Embed Size (px)
Citation preview

Eur Reeplr J 1991, 4, 860-368
Possible roles of the peripheral vagal nerve in histamineinduced bronchoconstriction in guinea-pigs
H. lnoue, H. Aizawa, N. Miyazaki, T. lkeda, N. Shigematsu
Possible roles of the peripheral vagal nerve in histamine-induced bronchoconstriction in guinea-pigs. H. Inoue, H. Aizawa, N. Miyazaki, T. Ikeda, N. Shigematsu. ABSTRACT: Although the Importance of the vagal nerve In the patbogenesls of bronchial asthma has been reported, Its precise contribution Is still not fully understood. To shed more light on this area, we evaluated the possible contribution of vagal reflex In histamine-Induced bronchoconstrldlon (BIB), and decided the site of action of histamine on the vagal nerve.
For this purpose, we studied the effects of the bilateral cervical vagotomy, hexamethonium (l mg·kg·1) or tetrodotoxin (0.5 mg•kg·1) on BIB (81J.i•kg'1, iv) In anaesthetized and mechanlcaUy-ventllated guinea-pigs. We also studied whether or not atropine (1 mg•kg'1) decreases HIB after vagotomy, Including either the treatment of hexamethonium or tetrodo· toxin. Airway responses were assessed by measurement of pulmonary resistance.
The following results were obtained; 1) the response to histamine was signlftcantly enhanced by the vagotomy, hexamethonium or tetrodotoxin; 2) propranolol Increased HIB, and HIB was further enhanced by the vagotomy In the animals treated with propra.nolol; 3) atropine significantly suppressed BIB afte.r the vagotomy, hexamethonium or tetrodotoxin.
These results suggest that the postganglionic vagal nerve plays an excltatory role In BIB through the release of acetylcholine from the nerve terminals. It Is also suggested that the vagal reflex mainly exhibits an Inhibitory role In the HIB of guinea-pigs, presumably by the action of the nonadrenergfc Inhibitory nervous system. Eur Respir J., 1991, 4, 860-866.
Research Institute for Disease of the Chest, Faculty of Medicine, Kyushu University, Fukuoka, Japan.
Correspondence: H. Aizawa, Research Institute for Disease of the Chest, Faculty of Medicine, Kyushu Unive~ity, Ma.idashl3-1-1, Higashi-ku, Fu.lruoka 812, Japan.
Keywords: Atropine; histamine-induced broncho· constriction; hexamethonium; tetrodotoxin; vagal nerve; vagotomy.
Received: June 29, 1990; accepted after revision April 17, 1991.
This study was supported by Grants No. 01570432 for General Scientific Research from the Ministry of Education, Science and Culture of Japanese Government.
The precise mechanisms involved in airway hyperresponsiveness are not yet fully understood. However, it has been considered that the vagal nerve plays an important role in the pathogenesis of airway hyperresponsiveness (1-5]. It has been suggested that an augmented vagal reflex may be involved in the development of the increased responsiveness of the airways, since the stimuli such as histamine and allergen to the vagal sensory nerve cause bronchoconstriction which can be inhibited by sectioning/cooling of the vagal nerves or by a cholinergic antagonist [6-9]. However, it was also reported that a vagotomy does not suppress the bronchoconstriction induced by histamine [10, 11], and therefore the possible role of vagal reflex is still controversial.
through the release of acetylcholine [13-15]. However, the mechanism of activation of the peripheral vagal nerve by histamine remains to be clarified.
Furthermore, it has been suggested that the vagal nerve may participate locally in the bronchoconstriction (12]. Namely, the stimulation of afferent vagal nerve endings may result in an "axon reflex" by releasing tachykinins which cause bronchoconstriction. It is also possible that histamine may directly stimulate the efferent vagal nerve terminals and cause bronchoconstriction
The first aim of this study was to evaluate the relative contributions of the vagal reflex involved in the airway response to histamine. The second was to determine the site of action of histamine on the peripheral vagal nerve. The third was to clarify the inhibitory mechanism involved in the vagal reflex.
Methods
Procedure
Guinea-pigs, of either sex, weighing 500-900 g, were anaesthetized with pentobarbital sodium (50 mg·kg-1, i.p.}, and ventilated with a respirator (Harvard Apparatus, Model no. 680) through a tracheal cannula at a constant tidal volume (7-8 ml·kg-1) and a rate of 60 breaths·min·1•
IUSTAMINE AND VAOAL NERVE 861
A catheter was inserted into a jugular vein for the administration of drugs, and another catheter was in· serted into a carotid artery for the measurement of blood pressure using an electric manometer (Nihon Kohden, LPU-0.1). A fluid-filled catheter was inserted into the oesophagus to measure the oesophageal pressure for an evaluation of pleural pressure. The oesophageal catheter was positioned at the point where the maximal amplitude of pleural pressure can be obtained. The bilateral cervical vagal nerves were exposed. Sham surgery was performed in the control group. The animals were placed supine in a body-plethysmograph. Plethysmograph airflow (V) was measured with a Fleisch pneumotachograph (Nihon Kohden, TV-1321') and a differential pressure transducer (Nihon Kohden, TP-602T). The tidal volume (VT) was obtained by electrical integration of the flow signal. Transpulmonary pressure (Ptp) was estimated from the difference between the oesophageal and airway pressure measured by a differential pressure transducer (Nihon Kohden, TP-603T). ~lmonary resistance (RL) was calculated from Ptp and V by the method of AMoUR and MEAD (16].
The airway responses, that were assessed by the changes of RL, to intravenous infusions of histamine were obtained as follows: the lung was inflated to three times that of the tidal volume and the baseline RL was meas· ured one minute after the inflation. The animals were then given either an intravenous infusion of saline or histamine and then the RL was measured.
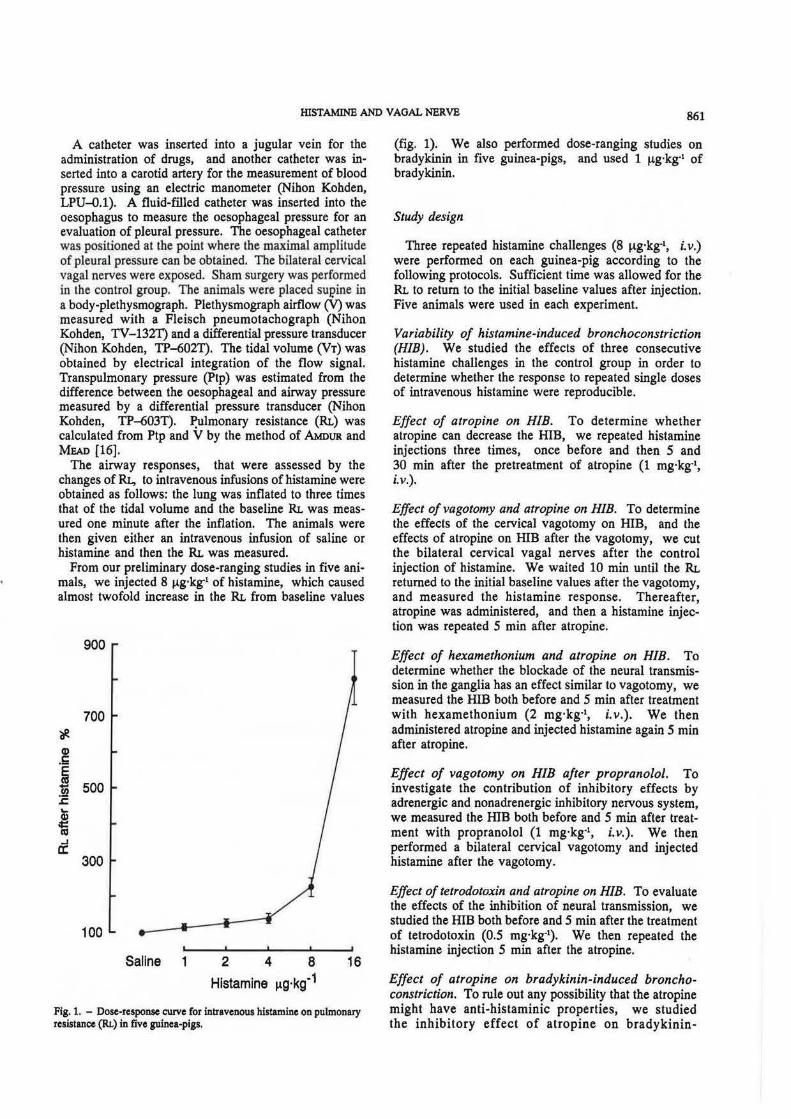
From our preliminary dose-ranging studies in five ani· mals, we injected 8 J,lg·kg-1 of histamine, which caused almost twofold increase in the RL from baseline values
900
700
* Q) c: -~ ]i 500 .s:: ... ~ ..J a:
300
100
Saline 2 4 6 16
Histamine J!Q·kg·1
Fig. 1. - Dose-response CIIIVe for intravenous histamine on pulmonary resistance (RL) in five guinea-pigs.
(fig. 1). We also performed dose-ranging studies on bradykinin in five guinea-pigs, and used 1 Jlg·kg·1 of bradykinin.
Study design
Three repeated histamine challenges (8 J.~.g·kg-1, i.v.) were performed on each guinea-pig according to the following protocols. Sufficient time was allowed for the RL to return to the initial baseline values after injection. Five animals were used in each experiment.
Variability of histamine-induced bronchoconstriction (HIB). We studied the effects of three consecutive histamine challenges in the control group in order to determine whether the response to repeated single doses of intravenous histamine were reproducible.
Effect of atropine on HJB. To determine whether atropine can decrease the HIB, we repeated histamine injections three times, once before and then 5 and 30 min after the pretreatment of atropine (1 mg·kg·t, i.v.).
Effect of vagotomy and atropine on HIB. To determine the effects of the cervical vagotomy on HIB, and the effects of atropine on IDB after the vagotomy, we cut the bilateral cervical vagal nerves after the control injection of histamine. We waited 10 min until the RL returned to the initial baseline values after the vagotomy, and measured the histamine response. Thereafter, atropine was administered, and then a histamine injection was repeated 5 min after atropine.
Effect of hexamethonium and atropine on HJB. To determine whether the blockade of the neural transmission in the ganglia has an effect similar to vagotomy, we measured the HIB both before and 5 min after treatment with hexamethonium (2 mg·kg·1, i. v.). We then administered atropine and injected histamine again 5 min after atropine.
Effect of vagotomy on HIB after propranolol. To investigate the contribution of inhibitory effects by adrenergic and nonadrenergic inhibitory nervous system, we measured the HIB both before and 5 min after treat· ment with propranolol (1 mg·kg-1, i. v.). We then performed a bilateral cervical vagotomy and injected histamine after the vagotomy.
Effect of tetrodotoxin and atropine on HJB. To evaluate the effects of the inhibition of neural transmission, we studied the HIB both before and 5 min after the treatment of tetrodotoxin (0.5 mg·kg-1
) . We then repeated the histamine injection 5 min after the atropine.
Effect of atropine on bradykinin-induced bronchoconstriction. To rule out any possibility that the atropine might have anti-histaminic properties, we studied the inhibitory effect of atropine on bradykinin-

862 H. INOUE ET AL.
induced bronchoconstriction (1 J,tg·kg·1, i. v.). We injected bradykinin before, and then 5 and 30 min after the atropine.
Time-matched control on HIB. It is not possible to exclude that the effect of first treatment, such as vagotomy, hexamethonium, tetrodotoxin or propranolol, could be time dependent, and thus contribute to changes ascribed to second treatment, such as atropine or vagotomy. We studied the HIB with or without the second treatment after the first treatment. In the experiment of time-matched control, we injected a vehicle or did sham surgery instead of the second treatment. Ten animals were used in each group. The values of RL are expressed as percentage of the maximal response after the first treatment.
Data analysis
The values of pulmonary resistance are expressed as the percentage of the initial baseline values, except in the time-matched control group. Data are expressed as arithmetic means and standard errors of the means in each experimental series. Differences between the means were determined using Student's t-test for paired samples. A p value <0.05 was considered to be statistically significant.
Drugs
The following drugs were used; pentobarbital sodium (Abbott Lab., North Chicago, IL), histamine diphosphate, atropine sulphate, propranolol hydrochloride, hexamethonium bromide and tetrodotoxin (Sigma Chemical, St. Louis, MO).
Results
Histamine injection increased Ptp, decreased V and markedly increased RL in the control guinea-pigs. The changes started at about 3 s after injection. The maximal change of the RL from the baseline values occurred at about 10 s after injection. These changes were transient and returned to baseline values within 3 min. Systemic blood pressure started to fall about 5 s after injection, and returned to the baseline within 3 min (fig. 2). A vehicle injection had no effect on the RI. or blood pressure.
Variability of HIB. In the control experiments, repeated histamine injections in the same animal resulted in virtually identical airway responses, as shown in table 1.
Effects of atropine on HIB. Atropine caused a significant suppression of airway responsiveness to intravenously administered histamine (p<O.Ol) (fig. 3). Thirty minutes after treatment with atropine, the response to
+ 0
1 ~ ] 111 11 111111111111111WIIIIllllll ll l lll lllllllll lll llllll ll llllllllllllllllllrrJIIIIIIIIIIIIlllllll~ Ptp "' :X: E u
..... V
, (/) e 1~]
01 ] :X: BP E
E
.....
] 'lP e
AL 6 "' :X:
6 0 2
M in
Fig. 2. - '!lace showing effeca of intravenous injection.of histamine (8 l'&·kg·1) on transpulmonary pressure (Ptp), air flow (V), systemic blood pressure (BP), and pulmonary resistance (Rt.) in a guinea-pig. Arrow indicates the time of injection.
Table 1. - Baseline and maximum change of AL for the 1st, 2nd, and 3rd histamine Injections obtained In five animals
Histamine injections 1st 2nd 3rd
Baseline; pre-blstamtne
Rt. c~O·mi·1·s %baseline
After histamine
Rt. cmHp·m1·1·s %baseline
0.61:0.054 0.60:t:0.057 0.60:t:0.057 100 98.4:t:1.6 97.8:t:1.6
l.lO:t:O.ll 178.8:t:8.0
1.08:t:O.ll 175.8:t7.1
1.14:t0.12 186.2:t9.4
Rt.: pulmonary resistance.
histamine returned to control values (p>O.l). The baseline RL values before, 5 min and 30 min after the atropine were not significantly different.
Effects of vagotomy and atropine on HIB. The surgical procedure for the bilateral vagotomy caused transient increases in RL, and the changes in RL returned spontaneously to initial baseline le~els. The vagotomy
HISTAMINE AND VAGAL NERVE 863
* (I) c 200 ·e ~ :E 100 .....
~ ...J a: 0
Control 5 min after 30 min after atropine atropine
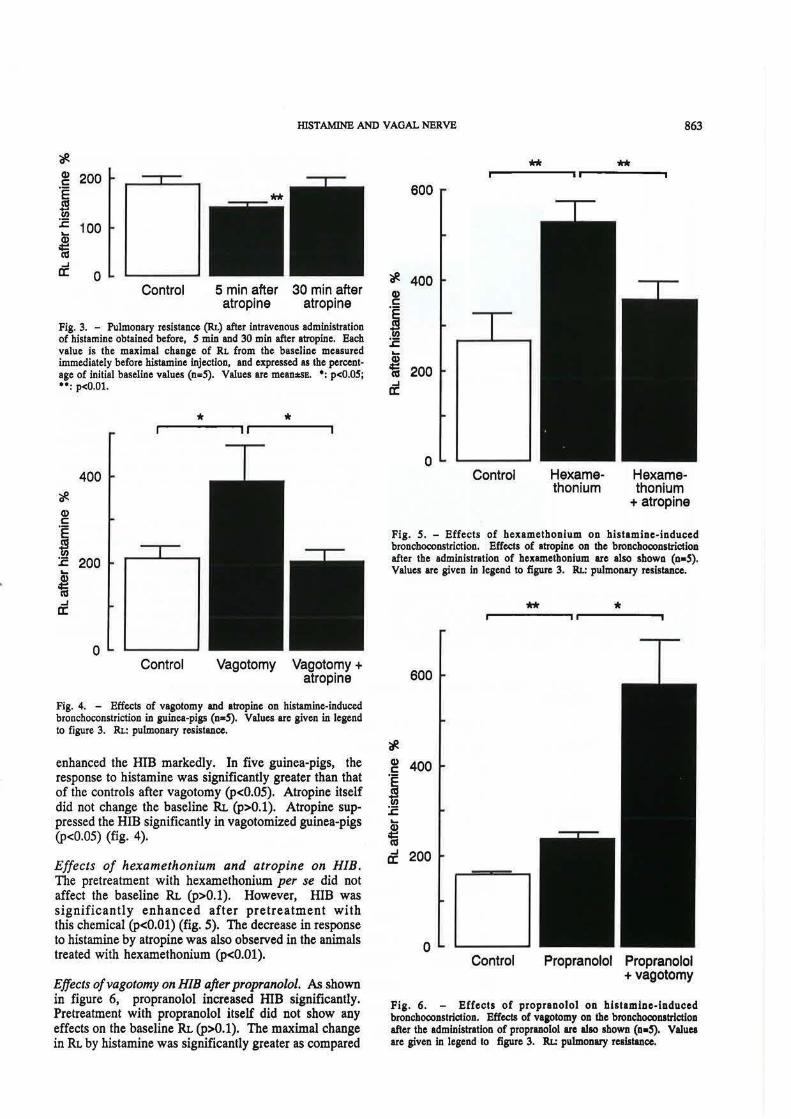
Fig. 3. - Pulmonary resistance (RI.) after intravenous administration of histamine obtained before, 5 min and 30 min after atropine. Bach value is the maximal change of RL from the baseline measured immediately before histamine injection, and expressed as the percentage of initial baseline values (n.,S). Values are meanzsl!. •: p<0.05; ••: p<O.Ol.
400
* (I)
i .£: 200 ..... (I)
li ..../ a:
0
*
I
Control Vagotomy Vagotomy+ atropine
Fig. 4. - Effects of vagotomy and atropine on histamine-induced bronchoconstriction in guinea-pigs (n•S). Values are given in legend to figure 3. RL: pulmonary resistance.
enhanced the HIB markedly. In five guinea-pigs, the response to histamine was significantly greater than that of the controls after vagotomy (p<0.05). Atropine itself did not change the baseline RL (p>O.l). Atropine suppressed the HIB significantly in vagotomized guinea-pigs (p<0.05) (fig. 4).
Effects of hexamethonium and atropine on HIB. The pretreatment with hexamethonium per se did not affect the baseline RL (p>O.l). However, HIB was significantly enhanced after pretreatment with this chemical (p<O.Ol) (fig. 5). The decrease in response to histamine by atropine was also observed in the animals treated with hexamethonium (p<O.Ol).
Effects of vagotomy on HIB after propranolol. As shown in figure 6, propranolol increased HIB significantly. Pretreatment with propranolol itself did not show any effects on the baseline RL (p>O.l). The maximal change in RL by histamine was significantly greater as compared
600
* 400 (I)
I :E .... (I)
li 200 ...J a:
0
**
I
Control Hexamethonium
**
Hexamethonium
+atropine
Fig. 5. - Effects of hexamethonium on histamine-induced bronchoconstriction. Effecta of atropine on the bronchoconstriction after the administration of hexamethonium are also shown (n•S). Values are given in legend to figure 3. Rt.: pulmonary resistance .
600
"#. (I) 400
I :c ....
~ cf 200
0
**
Control
*
Propranolol Propranolol +vagotomy
Fig. 6. - Effects of propranolol on histamine-Induced bronchoconstriction. Effecta of vaaotomy OD the broochoconslrictiOD after the administration of propranolol are also shown (o•S). Values are given in legend to figure 3. Rt.: pulmonary resistance.
864
400
~ Cl) c:
i I
:c 200
~ ....1 a:
0 Control
*
H. INOUB ET AL.
*
Tetrodotoxin Tetrodotoxin +atropine
to the control response after the application of propranolol (p<O.Ol). A bilateral vagotomy further increased changes in RL by histamine in the animals treated with propranolol (p<O.OS).
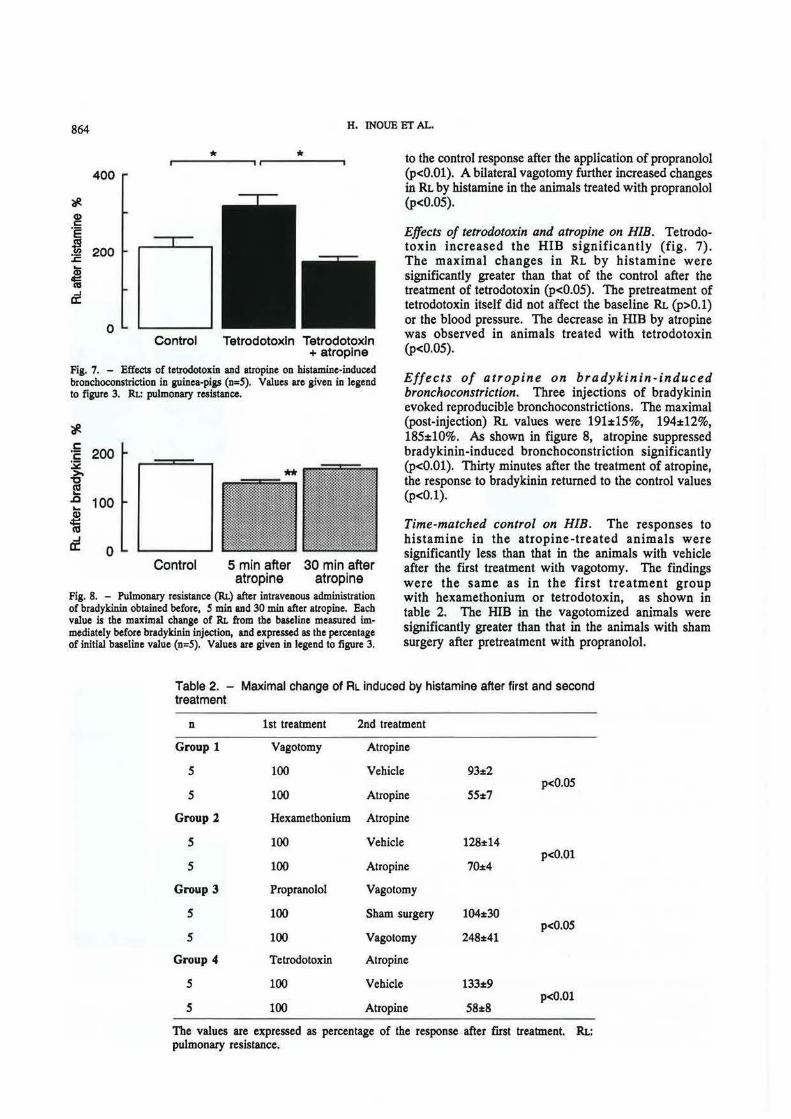
Effects of tetrodotoxin and atropine on HIB. Tetrodotoxin increased the HIB significantly (fig. 7). The maximal changes in RL by histamine were significantly greater than that of the control after the treatment of tetrodotoxin (p<0.05). The pretreatment of tetrodotoxin itself did not affect the baseline RL (p>O.l) or the blood pressure. The decrease in HIB by atropine was observed in animals treated with tetrodotoxin (p<0.05).
Fig. 7. - Effects of tetrodotoxin and atropine on histamine-induced bronchoconstriction in guinea-pigs (n=S). Values are given in legend to figure 3. RI.: pulmonary resistance.
Effects of atropine on bradykinin-induced bronchoconstriction. Three injections of bradykinin evoked reproducible bronchoconstrictions. The maximal (post-injection) RL values were 191:t15%, 194:t12%, 185:t10%. As shown in figure 8, atropine suppressed bradykinin-induced bronchoconstriction significantly (p<O.Ol). Thirty minutes after the treatment of atropine, the response to bradykinin returned to the control values (p<O.l) .
.,. c: '2 200 :52 ~ ~ .Q 100 .... Cl)
li ...J a: 0
Control 5 min after 30 min after atropine atropine
Fig. 8. - Pulmonary resistance (RI.) after intravenous administration of bradykinin obtained before, S min and 30 min after atropine. Each value is the maximal change of Rt. from the baseline measured immediately before bradykinin injection, and expressed as the percentage of initial baseline value (n=S). Values are given in legend to figure 3.
Time-matched control on HIB. The responses to histamine in the atropine-treated animals were significantly less than that in the animals with vehicle after the first treatment with vagotomy. The findings were the same as in the first treatment group with hexamethonium or tetrodotoxin, as shown in table 2. The HIB in the vagotomized animals were significantly greater than that in the animals with sham surgery after pretreatment with propranolol.
Table 2. - Maximal change of RL induced by histamine after first and second treatment
D 1st treatment 2nd treatment
Group 1 Vagotomy Atropine
5 100 Vehicle 93:2 p<0.05
5 100 Atropine 55:7
Group 2 Hexamethonium Atropine
5 100 Vehicle 128±14 p<O.Ol
5 100 Atropine 70:t4
Group 3 Propranolol Vagotomy
5 100 Sham surgery 104:30 p<0.05
5 100 Vagotomy 248:t41
Group 4 Tetrodotoxin Atropine
5 100 Vehicle 133±9
5 100 Atropine 58:t8 p<O.Ol
The values are expressed as percentage of the response after first treatment. RL: pulmonary resistance.

IDSTAMJNE AND VAGAL NERVE 865
Discussion
This study demonstrates that airway response to histamine was enhanced by the bilateral cervical vagotomy or the treatment with hexamethonium or tetrodotoxin. It is also demonstrated that atropine markedly suppressed the response to histamine even after the vagotomy. These results indicate that the vagal reflex may play an inhibitory role in the histamine-induced bronchoconstriction, and that the cholinergic bronchoconstriction may still remain even after the vagotomy.
It has been reported that bronchoconstriction induced by histamine may involve vagal reflex. The bronchoconstriction induced by histamine injection into bronchial arteries or inhalation was suppressed either by cooling or culling the cervical vagal nerves in dogs [6, 7, 9] and cats (17]. In awake guinea-pigs, it has also been reported that vagotomy attenuates HlB [8]. The importance of the vagal reflex was also suggested by the reduction in Hl.B by atropine in guinea-pigs (18, 19]. These results suggest that the vagal reflex might be an important factor for development of increased responsiveness of the airways.
In the present study, however, vagotomy did not attenuate but enhanced the HIB, and atropine did suppress HIB even after vagotomy. These findings raise the possibility that the "peripheral vagal nerve" may play a role in the enhancement of HIB, by releasing acetylcholine. The response after vagotomy consists of both the direct action to smooth muscle and the enhancement by the peripheral vagal nerve. Atropine eliminates the component of vagally mediated response. This possibility was supported by the results that the treatment with hexamethonium enhanced HIB, and that atropine also suppressed HIB after hexamethonium. Furthermore, atropine suppressed the bronchoconstriction induced by another bronchoconstrictor, bradykinin, as well as histamine. We consider this result rules out the possibility that the suppression of HIB by atropine may be due to an anti-histaminic activity of this agent.
Discrepancy in the present and previous fmdings on the effects of vagotomy is not fully understood, however, several methodological differences may partly account for it. We used anaesthetized animals under mechanical ventilation. It has been reported that there was less vagally mediated bronchoconstriction in ventilated than spontaneously breathing rabbits (20]. Anaesthesia has also been shown to suppress vagal reflexes (21]. We studied the effects of the cervical vagotomy on HIB using different anaesthesia, alphachloralose and urethan. The results of the experiment with chloralose and urethan were the same as that with pentobarbital (the data were not shown). We consider the difference of anaesthesia had little influence on the present findings.
The present findings are supported by several previous investigations. SHoRE et al. [9] reported that atropine attenuated HIB after bilateral vagotomy in dogs. Other previous studies reported that the bronchoconstrictor response to histamine was unaffected by either vagal
sectioning or cooling in either guinea-pigs [10, 11] or dogs (22--24].
Concerning the sites of action of histamine on the peripheral vagal nerve, there are two possibilities. Firstly, histamine may stimulate efferent vagal nerve terminals to release acetylcholine, since atropine suppressed HIB after tetrodotoxin in the present study. Treatment of tetrodotoxin completely blocked the neural transmission of vagal nerve, because electrical vagal stimulation (10 V, 50Hz, 1 ms, Ss) did not evoke any bronchoconstriction after the administration of tetrodotoxin (data not shown). These results support the possibility that histamine might stimulate efferent nerve terminals directly and release acetylcholine. Secondly, stimulation of afferent nerve terminals by histamine may cause a "local reflex" and release acetylcholine.
The augmentation of HIB after vagotomy suggests an inhibitory role of a "central vagal reflex", rather than an excitatory role. The adrenergic system may participate in this inhibitory effect, because propranolol enhanced the response to histamine. HIB was further enhanced by the vagotomy in animals treated with propranolol. This result suggests the contribution of the nonadrenergic inhibitory nervous system (NAIS) on the vagal reflex, since NAIS is involved in the vagal trunk [25, 26]. It has recently been reported that NAIS can be reflexly activated in humans [27] and cats [28]. A previous study from our laboratory demonstrated that hexamethonium induces airway hyperresponsiveness in cats treated with atropine and propranolol, suggesting that the blockade of NAIS causes hyperresponsiveness [25].
In conclusion, this study has demonstrated the possibility that the "peripheral vagal nerve", at a site close to an airway, has an excitatory effect independent of the "central vagal reflex" in HIB. It is also suggested that the "central vagal reflex" mainly exhibits an inhibitory role in guinea-pigs in HIB, presumably due to the action of NAIS.
Aclcnowl1dgements: The authors thank Drs Y. Ito and T. Hirose for reviewing the manuscript, and Miss H. Sawae for technical assistance.
References
1. Simonsson BG, Jacobs FM, Nadel JA. - Role of autonomic nervous system and the cough reflex in the increased responsiveness of airways in patients with obstructive airway disease. J Clin Invest, 1967, 46, 1812-1818. 2. Boushey HA, Holtzman MJ, Sheller JR, Nadel JA. -Bronchial hyperreactivity. Am Rev Respir Dis, 1980, 121, 389-413. 3. Richardson JB. - Nerve supply to the lungs. Am Rev Respir Dis, 1979, 119, 78~02. 4. Nadel JA, Bames PJ. - Autonomic regulation of the airways. Ann Rev Med, 1984, 35, 451-467. 5. Barns PJ. - Neural control of human airways in health and disease. Am Rev Respir Dis, 1986, 134, 1289-1314. 6. DeKock MA, Nadel JA, Zwi S, Colebatch HJH, Olsen CR. - New method for perfusing bronchial arteries: histamine bronchoconstriction and apnea. J Appl Physiol, 1966, 21, 185-194.

866 H. INOUE ET AL.
7. Gold WM, Kessler G-F, Yu DYC. - Role of vagus nerves in experimental asthma in allergic dogs. J Appl Physiol, 1972, 33, 719-725. 8. Mills JE, Widdicombe JG. - Role of the vagus nerves in anaphylaxis and histamine-induced bronchoconstrictions in guinea-pigs. Br J Pharmacol, 1970, 39, 724-731. 9. Shore SA, Bai TR, Wang CG, Martin JO. - Central and local cholinergic components of histamine-induced bronchoconstriction in dogs. J Appl Physiol, 1985, 58, 443-451. 10. Gordon T, Venugopalan CS, Amdur MO, Drazen JM. -Ozone-induced airway hyperreactivity in the guinea-pig. J Appl Physiol: Respirat Environ Exercise Physiol, 1984, 57, 1034-1038. 11. Mauser PJ, Edelman NH, Chapman RW. - Central nervous system control of airway tone in guinea-pigs: the role of histamine. J Appl Physiol, 1988, 65, 2024-2029. 12. Bames PJ. - Asthma as an axon reflex. Lancet, 1986, i, 242-245. 13. Inoue T, Ito Y. - Characteristics of neuro-effector transmission in the smooth muscle layer of dog bronchiole and modifications by autacoids. J Physiol, 1986, 370, 551-565. 14. ShoreS, Irvin CG, Shenkier T, Martin JG. - Mechanisms of histamine-induced contraction of canine airway smooth muscle. J Appl Physiol: Respirat Environ Exercise Physiol, 1983, 55, 22-26. 15. Drazen JM, Schneider MW. - Comparative responses of tracheal spirals and parenchyma! strips to histamine and carbachol in vitro. J Clin Invest, 1978, 61, 1441-1447. 16. Amdur MO, Mead J. - Mechanics of respiration in unanesthetized guinea-pigs. Am J Physiol, 1958, 192, 364-368. 17. Zimmermann I, Ulmer WT. - The role of vagus nerve on histamine- and acetylcholine-induced bronchoconstriction (experiments on cats). Res Exp Med, 1986, 186, 29-34. 18. Drazen JM, Austen KF. - Atropine modification of the pulmonary effects of chemical mediators in the guinea-pig. J Appl Physio~ 1975, 38, 834-838. 19. Douglas JS, Dennis MW, Ridgway P, Bouhuys A. -Airway constriction in guinea-pigs: interaction of histamine and autonomic drugs. J Pharmacal Exp Ther, 1973, 184, 169-179. 20. Karczewski W, Widdicombe JG. - The role of the vagus nerves in the respiratory and circulatory responses to intravenous histamine and phenyl diguanide in rabbits. J Physiol (Lond), 1969, 201, 271-291. 21. Holtzman MJ, Hahn HL, Sasak:i K, Skoogh B-E, Graf PD, Fabbri LM, Nadel JA - Selective effect of general anesthetics on reflex bronchoconstrictor response in dogs. J Appl Physiol: Respirat Environ Exercise Physiol, 1982, 53, 126-133. 22. Irvin CG, Dempsey JA. - Role of H1 and H, receptors in increased small airways resistance in the dog. Respir Physiol, 1978, 35, 161-176. 23. Loring SH, Drasen JM, Ingram RH Jr. - Canine pulmonary response to aerosol histamine: direct versus vagal effects. J Appl Physiol: Respirat Environ Exercise Physiol, 1977, 42, 946-952.
24. Snapper JR, Drazen JM, Loring SH, Braasch PS, Ingram RH Jr. - Vagal effects on histamine, carbachol, and prostaglandin F,. responsiveness in the dog. J Appl Physiol: Respirat Environ Exercise Physiol, 1979, 47, 13-16. 25. Aizawa H, Matsuzaki Y, lshibashi M, Domae M, Hirose T, Shigematsu N, Tanaka K. - A possible role of a nonadrenergic inhibitory nervous system in airway hyperreactivity. Respir Physiol, 1982, 50, 187-196. 26. Richardson JB. - Nerve supply to the lungs. Am Rev Respir Dis, 1979, 119, 785-802. 27. Michoud M-C, Amyot R, Jeanneret-Grosjean A, Couture J. - Reflex decrease of histamine-induced bronchoconstriction after laryngeal stimulation in humans. Am Rev Respir Dis, 1987, 136, 618-622. 28. Szarek JL, Gillespie MN, Altiere RJ, Diamond L. - Reflex activation of the nonadrenergic noncholinergic inhibitory nervous system in feline airways. Am Rev Respir Dis, 1986, 133, 1159-1162.
ROles possibles du nerf vague piriperique dan s la bronchoconstriction induite par / 'histamine chez le cobaye. H. Jnoue, H. Aizawa, N. Miyazaki, T. Jkeda, N. Shigematsu. RESUME: Quoique !'importance du nerf vague dans la pathogtnie de l'asthme bronchique ait 6te signalte, sa contribution exacte n'est pas encore compl!tement comprise. Afin d'~lairer davantage cette probltmatique, nous avons evalu6 la contribution possible d'un reflexe vagal dans la bronchoconstriction induite par !'histamine (HIB), et recherche le site d'action de l'bistamine sur le nerf vague.
Dans cette optique, nous avons etudie les effets de la vagotomie cervicale bilaterale, de !'hexamethonium (2 mg·kg) ou de la tetrodotoxine (0.5 mg·kg•) sur HIB (8 J.Lg·kg"', i. v.) chez des cobayes anesthesies et ventilts mecaniquement. Nous avons etudie egalement si l'atropine diminuait oui ou non HIB apr!s vagotomie (1 mg·kg-1) , en incluant le traitment, soit par !'hexamethonium soit par la tetrodotoxine. Les r6ponse des voies aeriennes ont ete apprecites par mesu.re de la resistance pulmonaire.
Les resultats suivants ont ett obtenus: 1) La reponse A !'histamine est fortement accentute par la vagotomie, !'hexamethonium ou la tetrodotoxine. 2) le propranolol a augmente HIB et HIB a ete accentute davantage par la vagotomie chez les animaux traites au propranolol. 3) !'atropine rtduit de fa~;on significative Hm apr!s vagotomie, hexamethonium ou tetrodotoxine.
Ces resultats sugg!rent que le nerf vague post-ganglionnaire joue un rllle excitateur dans HIB par la libtration d'acttylycholine A partir des terminaisons nerveuses. L'on sugg!re egalement que le reflexe vagal exerce principalement un role inhibiteur dans la HIB des cobayes, probablement par I' action du syst~me nerveux inhibiteur non adr6nergique. Eur Respir J., 1991, 4, 860-866.