Embed Size (px)
Citation preview
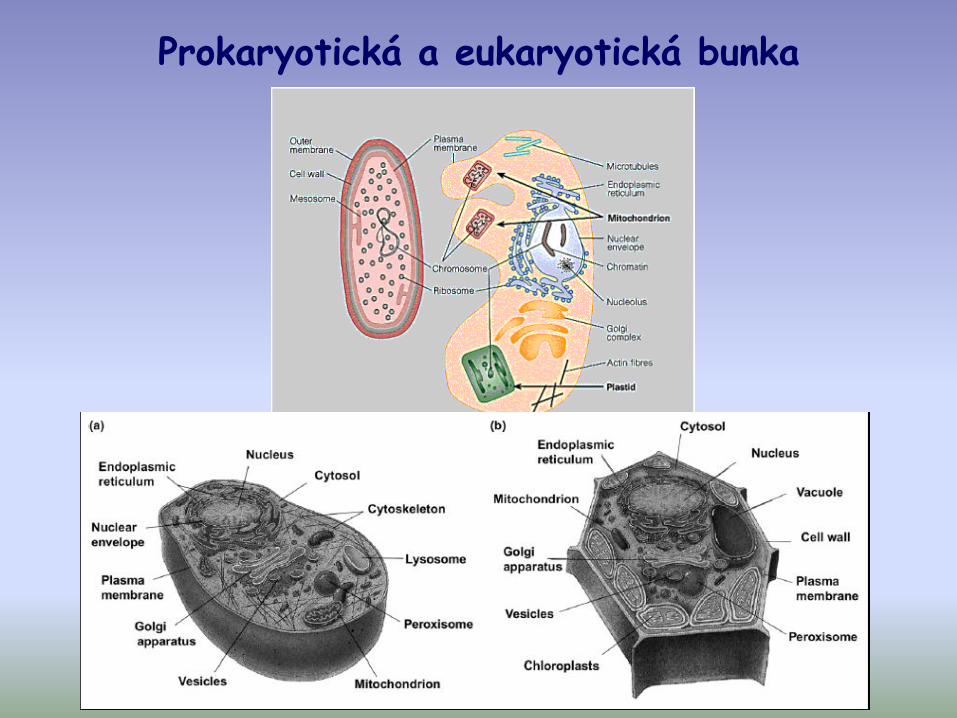
Prokaryotická a eukaryotická bunka
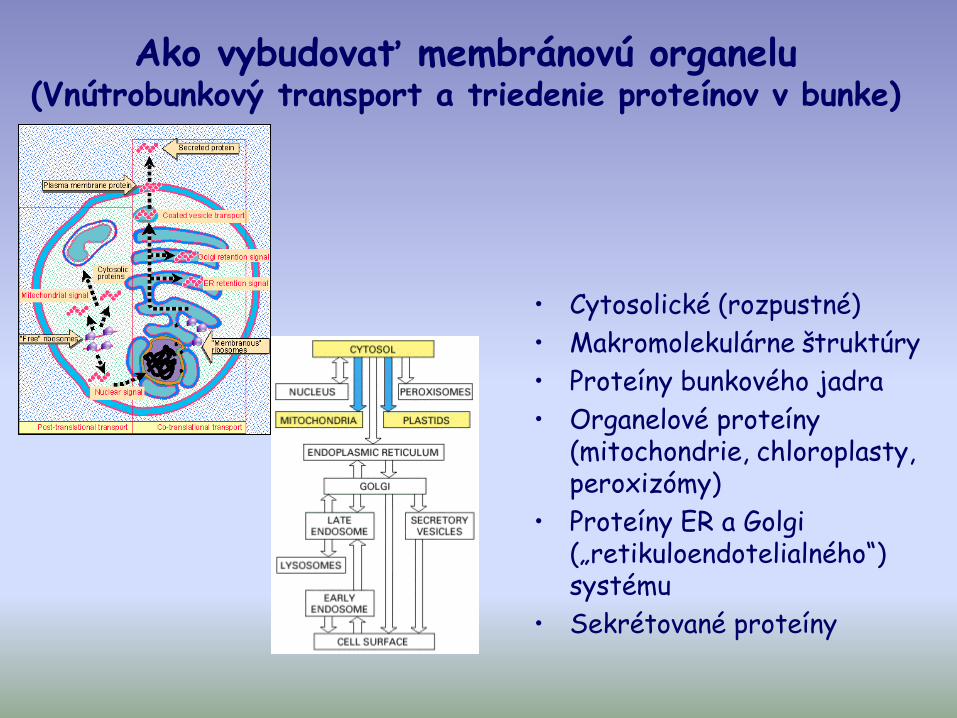
Ako vybudovať membránovú organelu(Vnútrobunkový transport a triedenie proteínov v bunke)
• Cytosolické (rozpustné)
• Makromolekulárne štruktúry
• Proteíny bunkového jadra
• Organelové proteíny (mitochondrie, chloroplasty, peroxizómy)
• Proteíny ER a Golgi („retikuloendotelialného“) systému
• Sekrétované proteíny
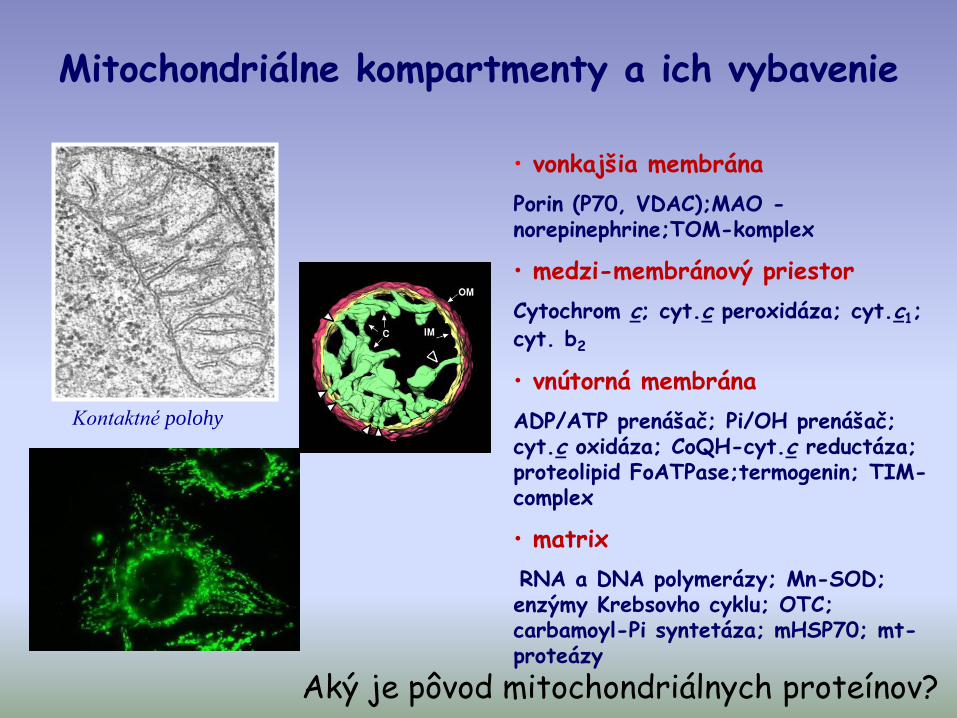
Mitochondriálne kompartmenty a ich vybavenie
• vonkajšia membrána
Porin (P70, VDAC);MAO -norepinephrine;TOM-komplex
• medzi-membránový priestor
Cytochrom c; cyt.c peroxidáza; cyt.c1; cyt. b2
• vnútorná membrána
ADP/ATP prenášač; Pi/OH prenášač; cyt.c oxidáza; CoQH-cyt.c reductáza; proteolipid FoATPase;termogenin; TIM-complex
• matrix
RNA a DNA polymerázy; Mn-SOD; enzýmy Krebsovho cyklu; OTC; carbamoyl-Pi syntetáza; mHSP70; mt-proteázy
Kontaktné polohy
Aký je pôvod mitochondriálnych proteínov?
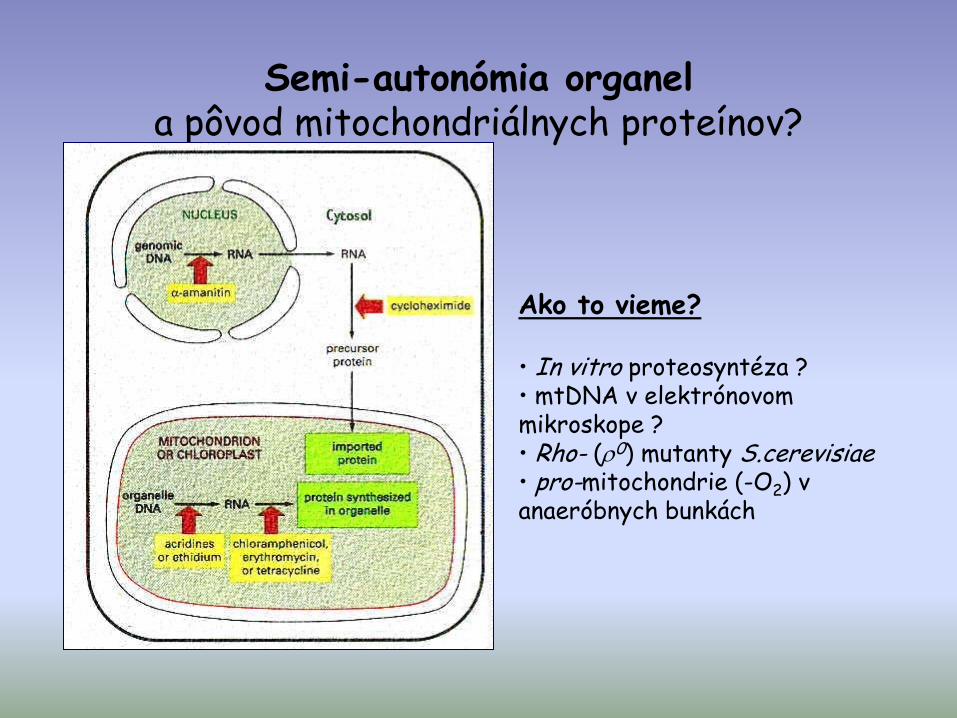
Semi-autonómia organel a pôvod mitochondriálnych proteínov?
Ako to vieme?
• In vitro proteosyntéza ?• mtDNA v elektrónovom mikroskope ?• Rho- (0) mutanty S.cerevisiae • pro-mitochondrie (-O2) v anaeróbnych bunkách
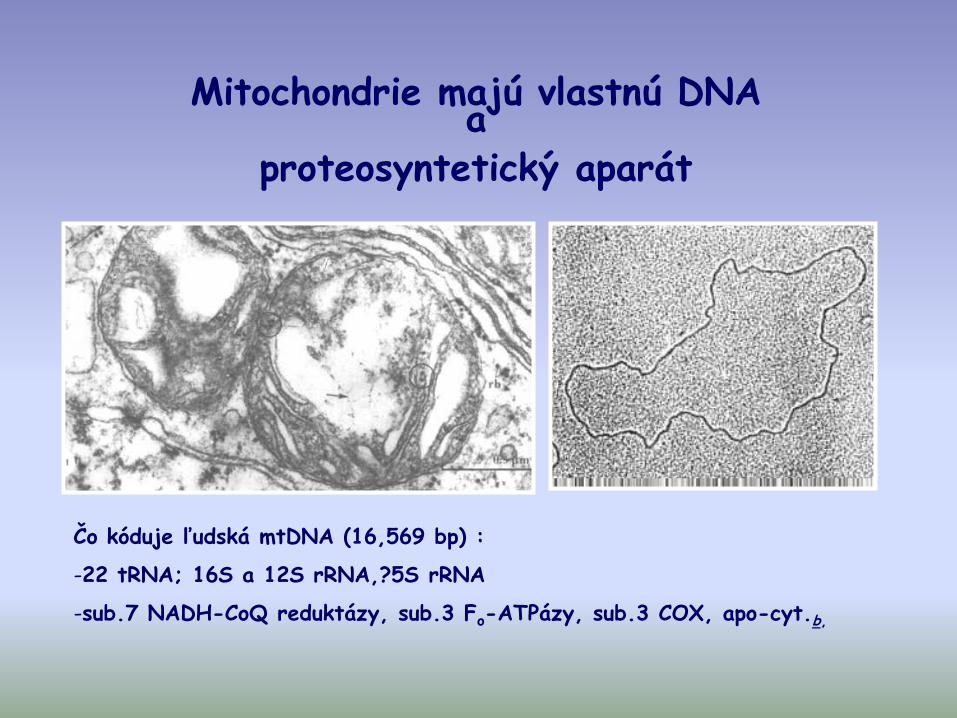
Mitochondrie majú vlastnú DNA a
proteosyntetický aparát
Čo kóduje ľudská mtDNA (16,569 bp) :
-22 tRNA; 16S a 12S rRNA,?5S rRNA
-sub.7 NADH-CoQ reduktázy, sub.3 Fo-ATPázy, sub.3 COX, apo-cyt.b,
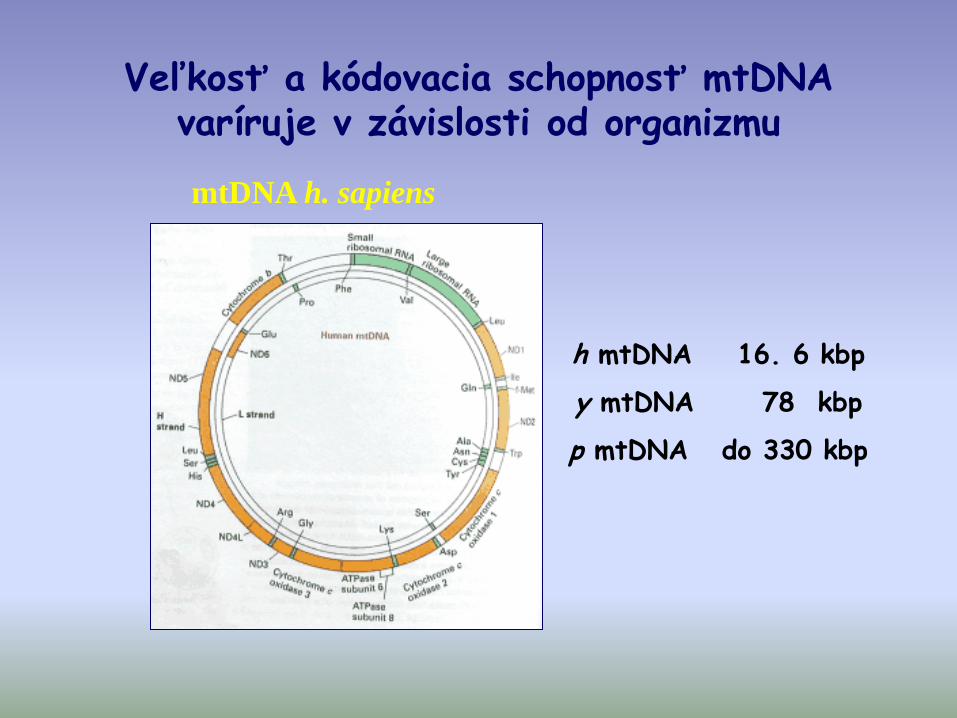
Veľkosť a kódovacia schopnosť mtDNA varíruje v závislosti od organizmu
mtDNA h. sapiens
h mtDNA 16. 6 kbp
y mtDNA 78 kbp
p mtDNA do 330 kbp
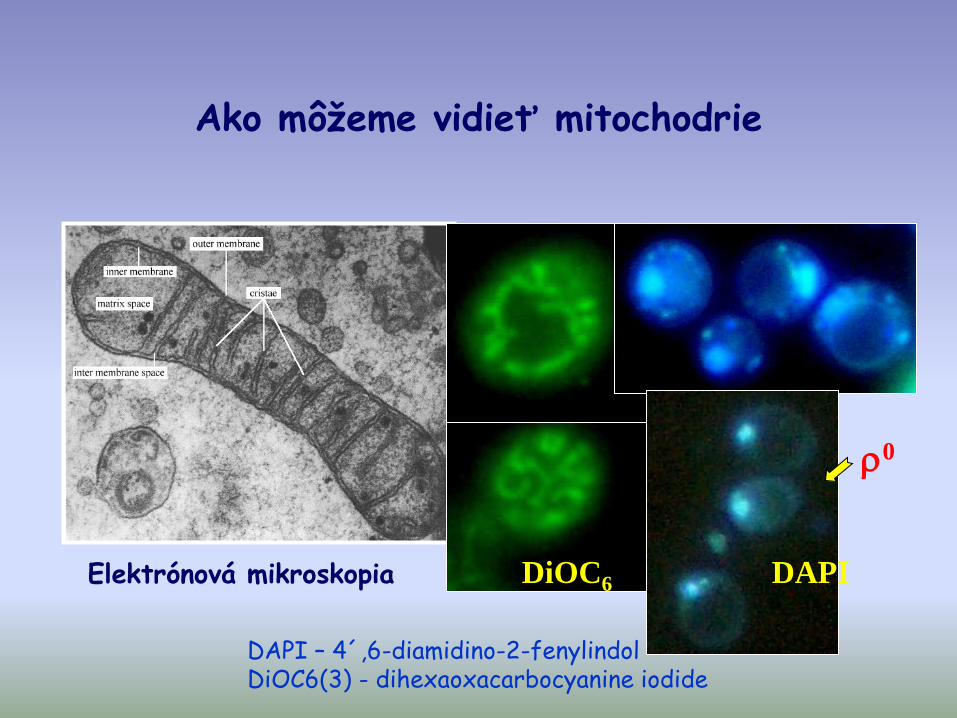
Ako môžeme vidieť mitochodrie
Elektrónová mikroskopia DiOC6 DAPI
0
DAPI – 4´,6-diamidino-2-fenylindolDiOC6(3) - dihexaoxacarbocyanine iodide
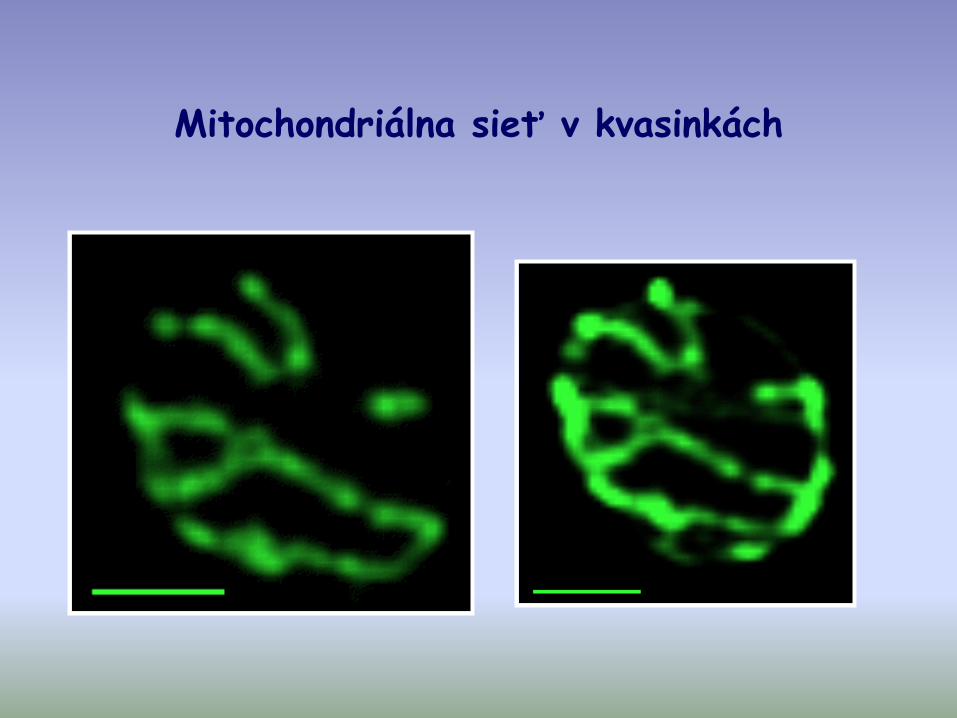
Mitochondriálna sieť v kvasinkách
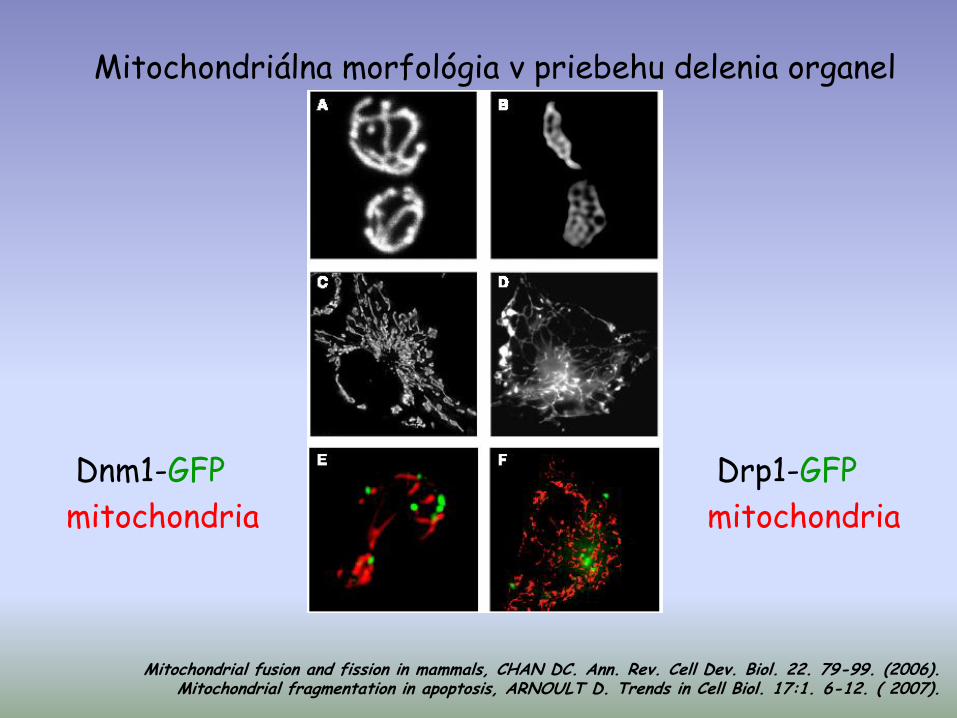
Mitochondriálna morfológia v priebehu delenia organel
Dnm1-GFP
mitochondria
Drp1-GFP
mitochondria
Mitochondrial fusion and fission in mammals, CHAN DC. Ann. Rev. Cell Dev. Biol. 22. 79-99. (2006). Mitochondrial fragmentation in apoptosis, ARNOULT D. Trends in Cell Biol. 17:1. 6-12. ( 2007).
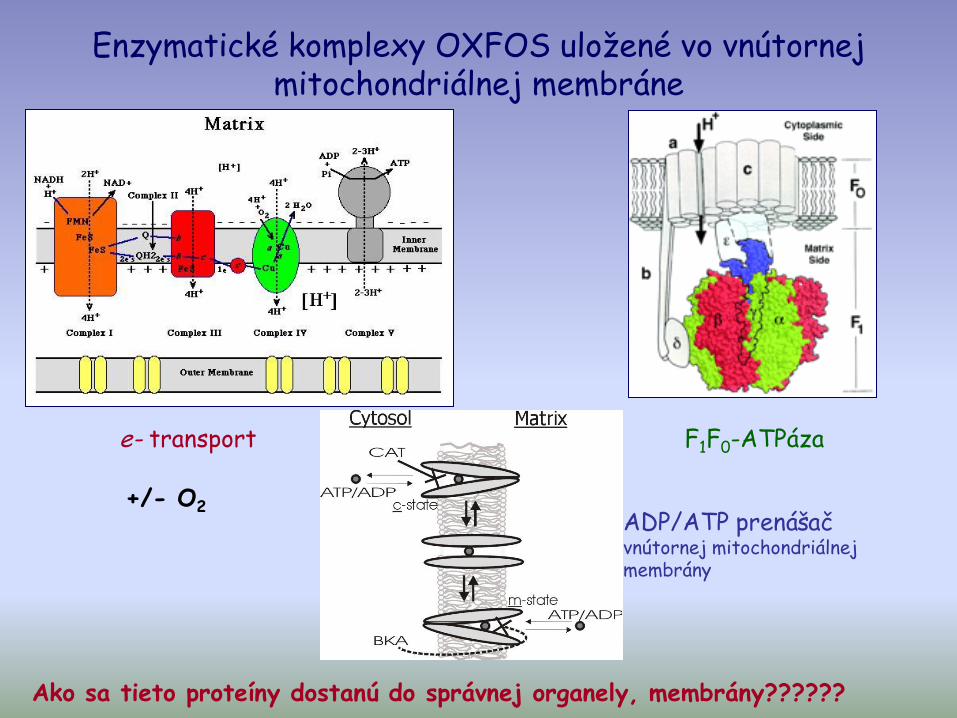
Enzymatické komplexy OXFOS uložené vo vnútornej mitochondriálnej membráne
ADP/ATP prenášač vnútornej mitochondriálnej membrány
F1F0-ATPázae- transport
+/- O2
Ako sa tieto proteíny dostanú do správnej organely, membrány??????
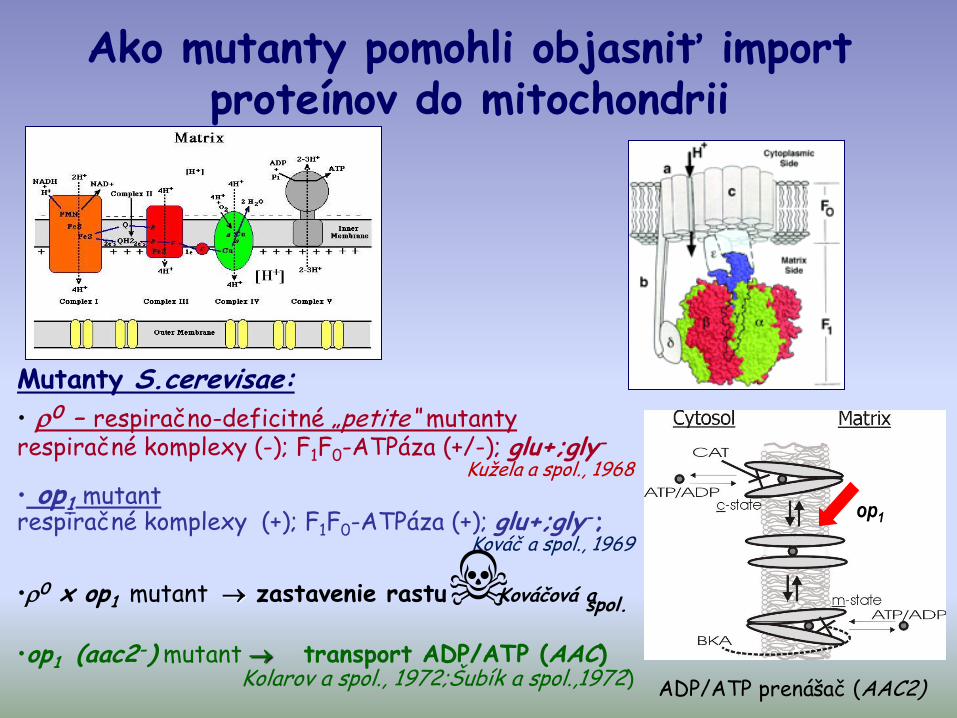
Ako mutanty pomohli objasniť import proteínov do mitochondrii
ADP/ATP prenášač (AAC2)
Mutanty S.cerevisae:
• 0 – respiračno-deficitné „petite“ mutantyrespiračné komplexy (-); F1F0-ATPáza (+/-); glu+;gly-
Kužela a spol., 1968
• op1 mutantrespiračné komplexy (+); F1F0-ATPáza (+); glu+;gly-;
Kováč a spol., 1969
•0 x op1 mutant zastavenie rastu Kováčová a spol.
•op1 (aac2-) mutant transport ADP/ATP (AAC)Kolarov a spol., 1972;Šubík a spol.,1972)
op1
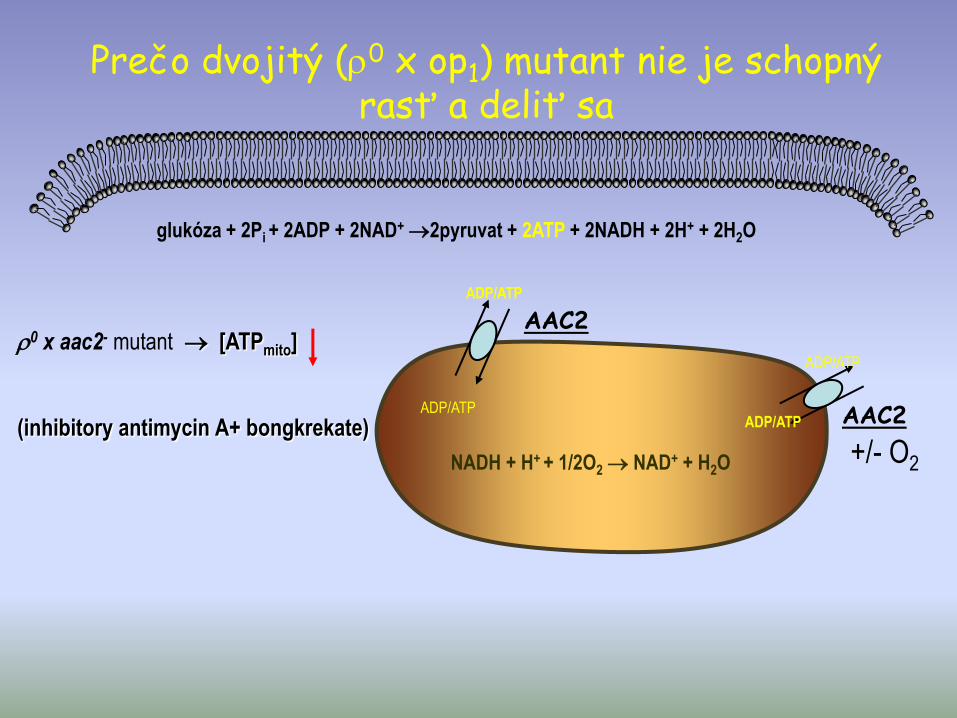
glukóza + 2Pi + 2ADP + 2NAD+ 2pyruvat + 2ATP + 2NADH + 2H+ + 2H2O
0 x aac2- mutant [ATPmito]
(inhibitory antimycin A+ bongkrekate)+/- O2
ADP/ATP
ADP/ATP
ADP/ATPADP/ATP
NADH + H+ + 1/2O2 NAD+ + H2O
AAC2
AAC2
Prečo dvojitý (0 x op1) mutant nie je schopný rasť a deliť sa

Záver:• Z neschopnosti rastu dvojitého mutanta (0 x
aac2-) vyplýva, že prítomnosť ATP vo vnútri mitochondrii je esenciálne pre život a delenie buniek.
Otázka: K čomu je potrebné ATP vo vnútri mitochondrii?
Experiment:
• k bunkám (0 x op1 ) mutanta a divého kmeňa (kontróla) pridať 35S-Met po čase pridať detergent antiserum mit.proteín.
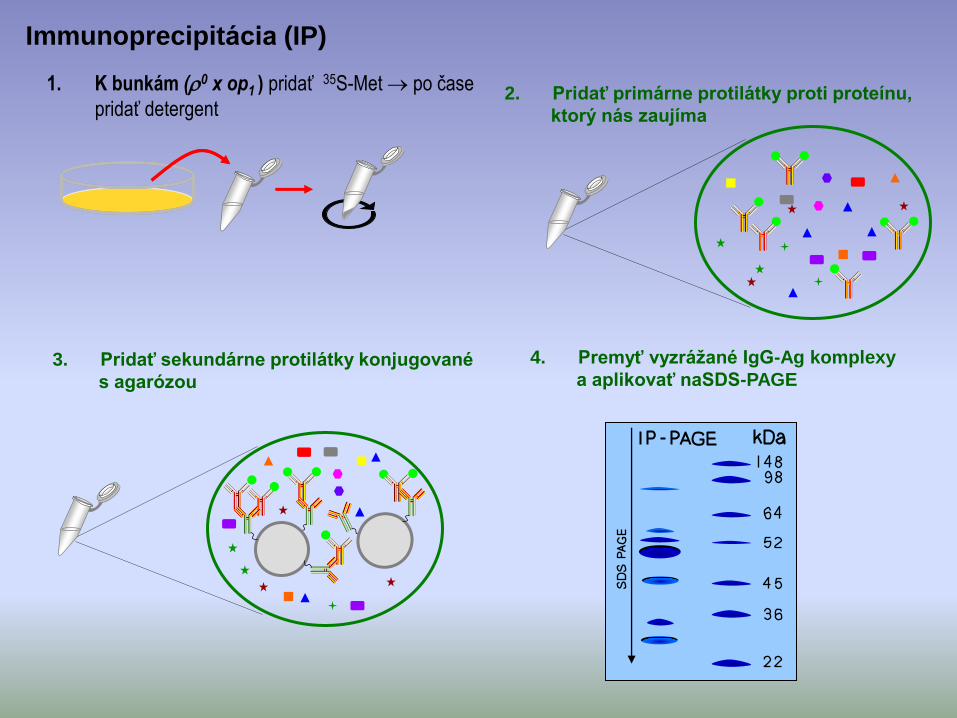
1. K bunkám (0 x op1 ) pridať 35S-Met po čase
pridať detergent
Immunoprecipitácia (IP)
2. Pridať primárne protilátky proti proteínu,
ktorý nás zaujíma
3. Pridať sekundárne protilátky konjugované
s agarózou
4. Premyť vyzrážané IgG-Ag komplexy
a aplikovať naSDS-PAGE
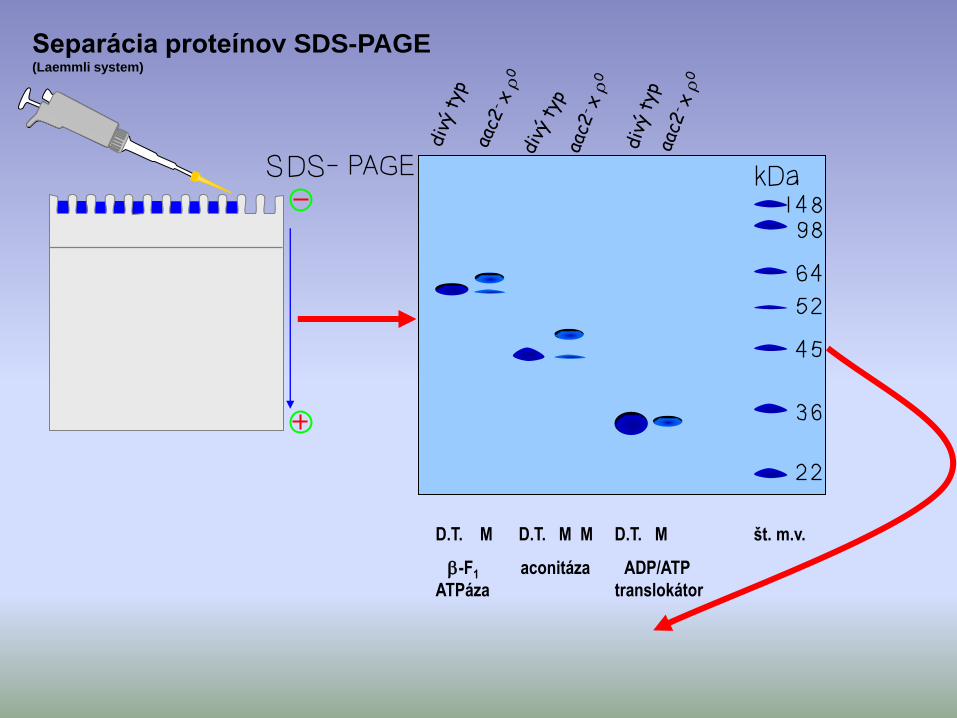
Separácia proteínov SDS-PAGE(Laemmli system)
D.T. M D.T. M M D.T. M št. m.v.
-F1 aconitáza ADP/ATP
ATPáza translokátor

Záver:
1) Intramitochondriálne ATP je esenciálne pre delenie eukaryotických buniek.
2) Mitochondriálne proteíny (kódované jadrovou DNA) sú syntetizované v cytoplazme ako prekurzory s väčšou mol.hmotnosťou.
3) Importované mito-prekurzory sú proteolitickým ošetrením (processing) premenené na dospelé formy.
4) Import mito-prekurzorov potrebuje intramitochondriálne ATP.
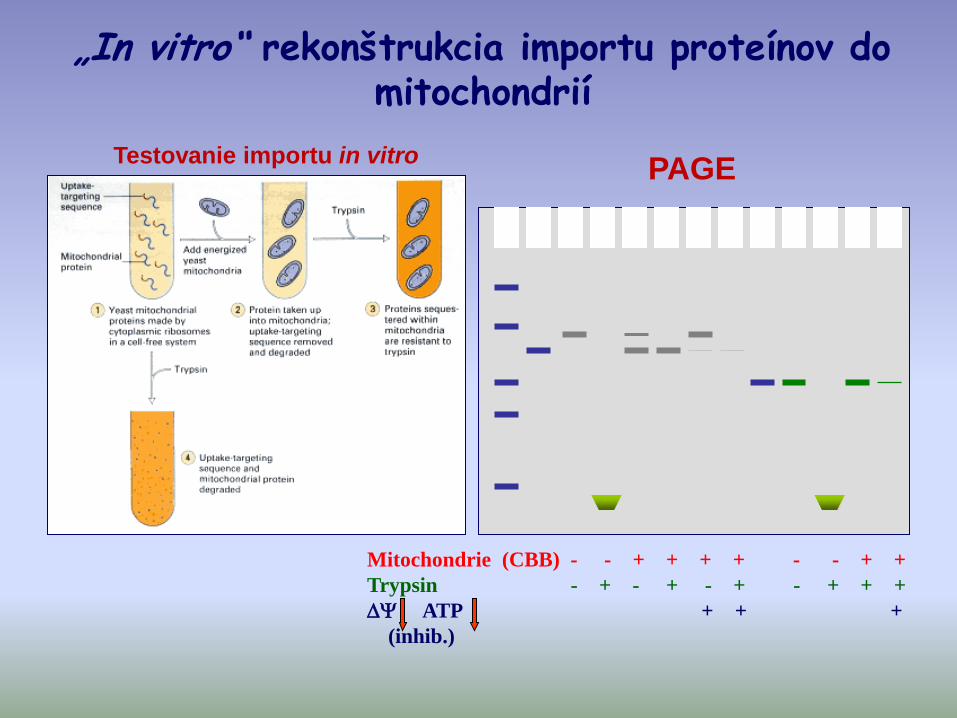
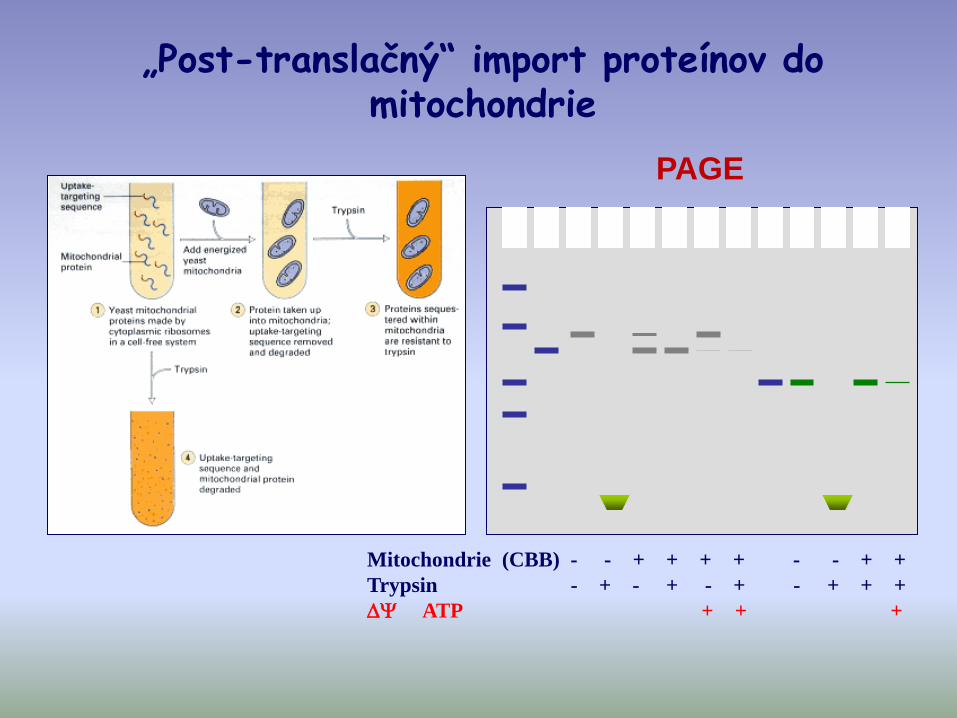
Mitochondrie (CBB) - - + + + + - - + +
Trypsin - + - + - + - + + +
ATP + + +
(inhib.)
„In vitro“ rekonštrukcia importu proteínov do mitochondrií
PAGETestovanie importu in vitro

Záver:
• Prekurzory sú najprv syntetizované v cytoplazme a potom „post-translačne“ sú importované do mitochondrie
• Okrem ATP je pre import potrebný
• Existujú mito-prekurzory, ktoré majú rovnakú mol. hmotnosť ako dospelá forma proteínu v mitochondriach.
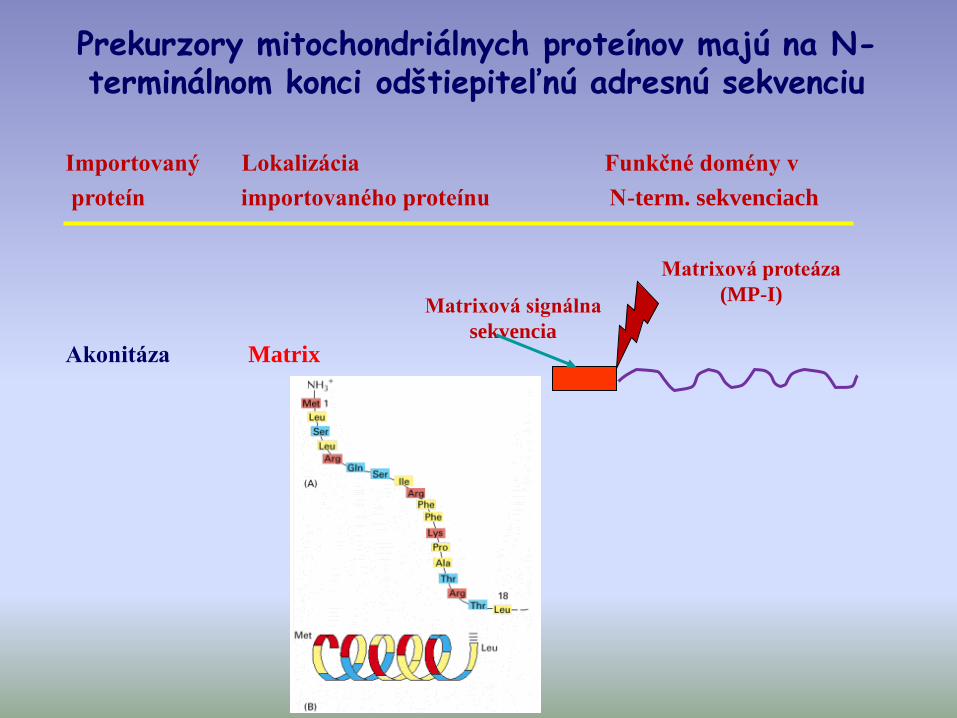
Importovaný Lokalizácia Funkčné domény v
proteín importovaného proteínu N-term. sekvenciach
Matrixová signálna
sekvencia
Matrixová proteáza
(MP-I)
Akonitáza Matrix
Prekurzory mitochondriálnych proteínov majú na N-terminálnom konci odštiepiteľnú adresnú sekvenciu

Vlastnosti mitochondriálnej adresnej sekvencie
• malá podobnosť medzi mito. adresnými sekvenciami
• vlastnosti vyplývajúce zo sekundárnej a terciárnej štruktúry adresnej sekvencie sú zodpovedné za rozpoznanie
? – Obsahuje adresná sekvencia všetku potrebnú informáciu pre lokalizáciu mitochondriálneho proteínu?
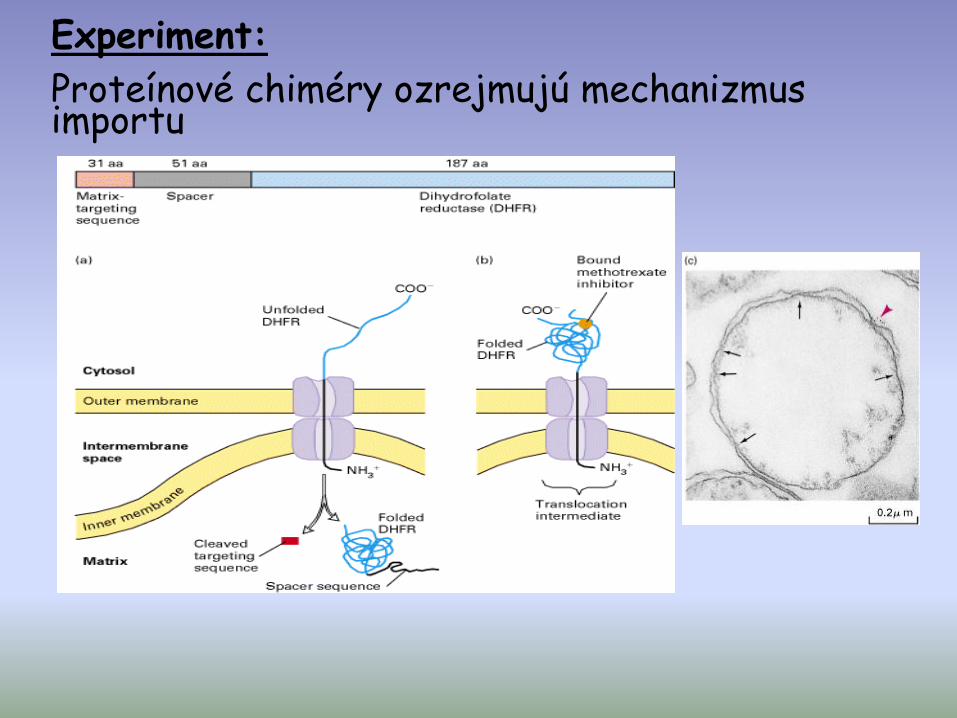
Experiment:
Proteínové chiméry ozrejmujú mechanizmus importu

Záver:
• Mito. adresná N-terminálna sekvencia je postačujúca pre import
• Importovaný peptid musí byť v rozvinutom (unfolded) stave
• Kontaktné polohy medzi OM a IM sú miestami pre import proteínov do mito.
!Tvorba stabilného translokačneho intermediatu!
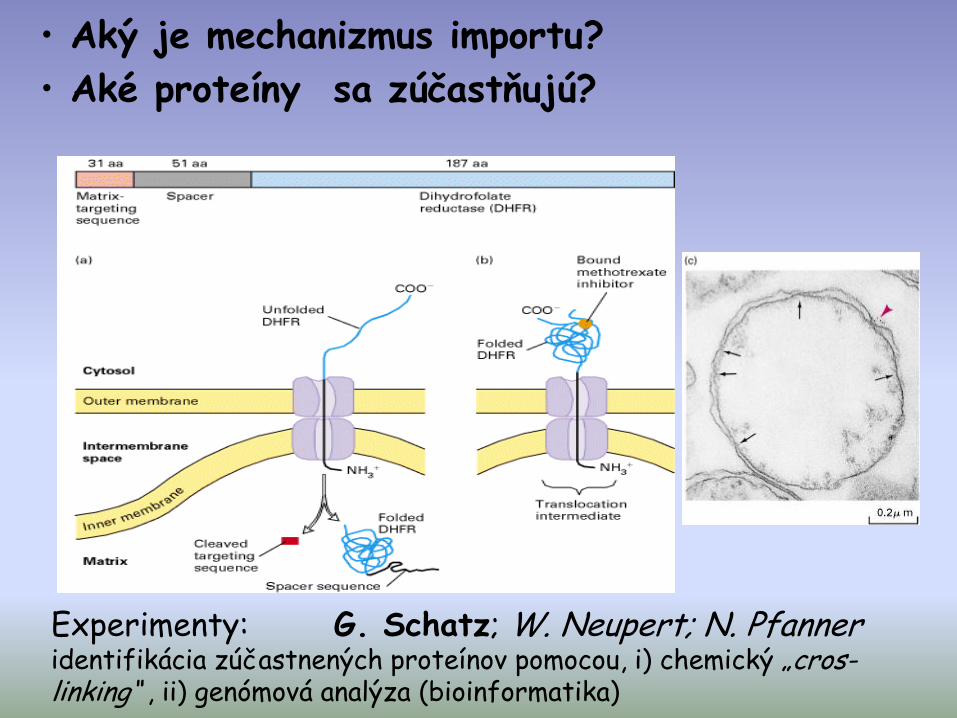
• Aký je mechanizmus importu?
• Aké proteíny sa zúčastňujú?
Experimenty: G. Schatz; W. Neupert; N. Pfanneridentifikácia zúčastnených proteínov pomocou, i) chemický „cros-linking“ , ii) genómová analýza (bioinformatika)

3 rôzne importné dráhy pre mitochondriálne proteíny syntetizované v cytosole
• import proteínov s N-terminálnou presekvenciou
• import proteínov s vnútornou signálnou sekvenciou
• proteíny s osobitou importnou dráhou
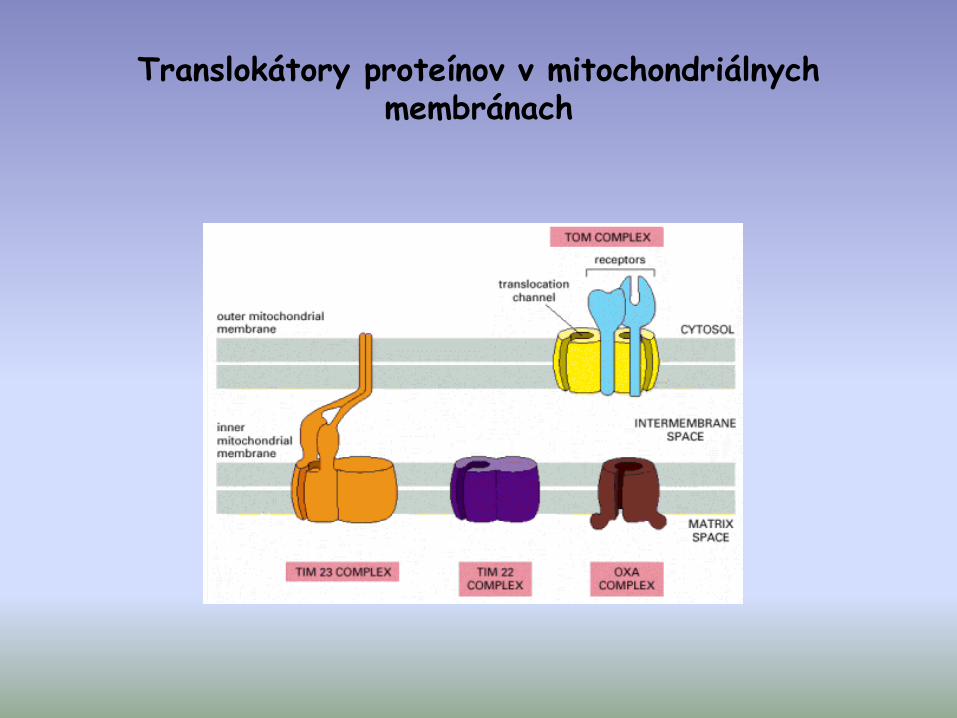
Translokátory proteínov v mitochondriálnych membránach
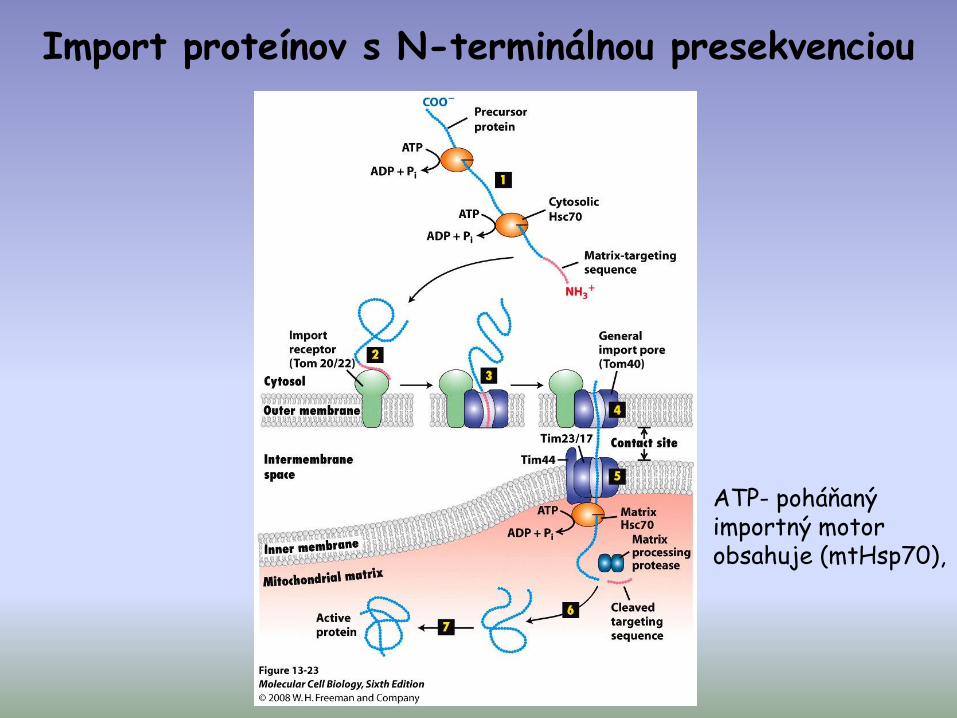
Import proteínov s N-terminálnou presekvenciou
ATP- poháňaný importný motor obsahuje (mtHsp70),
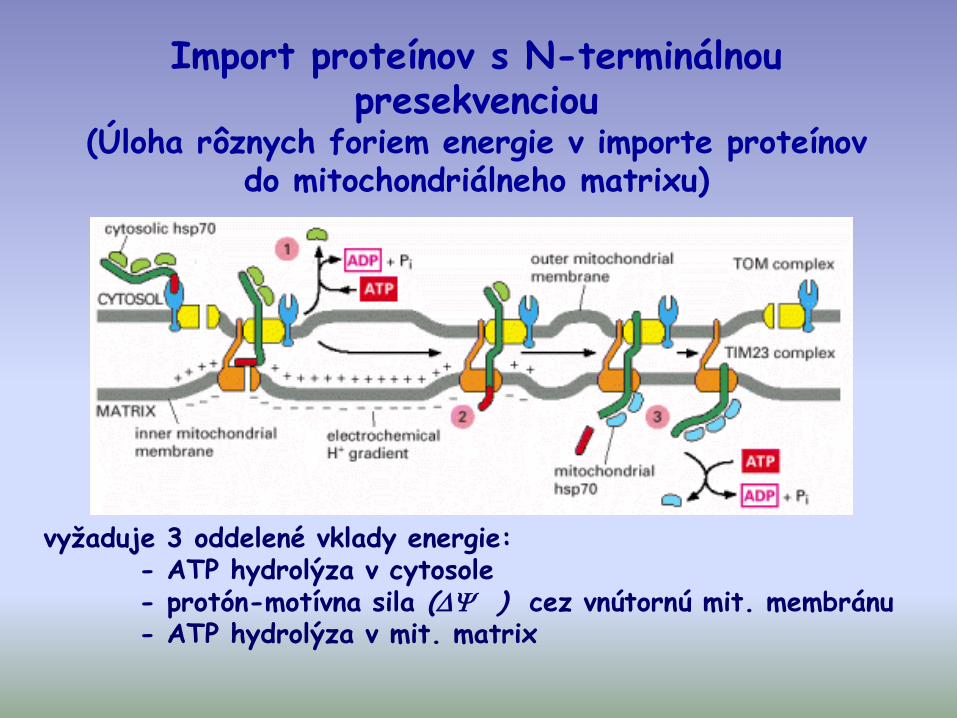
Import proteínov s N-terminálnou presekvenciou
(Úloha rôznych foriem energie v importe proteínov do mitochondriálneho matrixu)
vyžaduje 3 oddelené vklady energie:- ATP hydrolýza v cytosole- protón-motívna sila ( ) cez vnútornú mit. membránu- ATP hydrolýza v mit. matrix

Import proteínov bez N-terminálnej presekvencie
Sú to:
- niektoré proteíny importnej mašinérie vnútornej mitochondriálnej membrány (napr. Tim23)
- proteíny vnútornej mitochondriálnej membrány patriace do rodiny mitochondriálnych prenášačov (napr. ADP/ATP; Pi/OH-; citrát/malát, iné… )
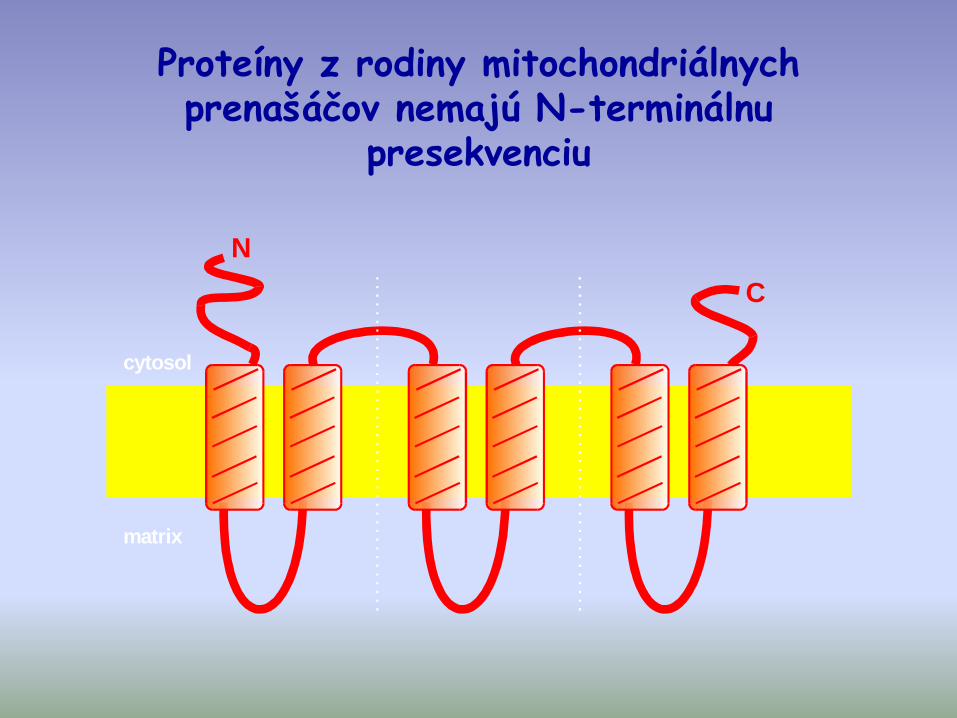
Proteíny z rodiny mitochondriálnych prenašáčov nemajú N-terminálnu
presekvenciu
cytosol
matrix
N
C
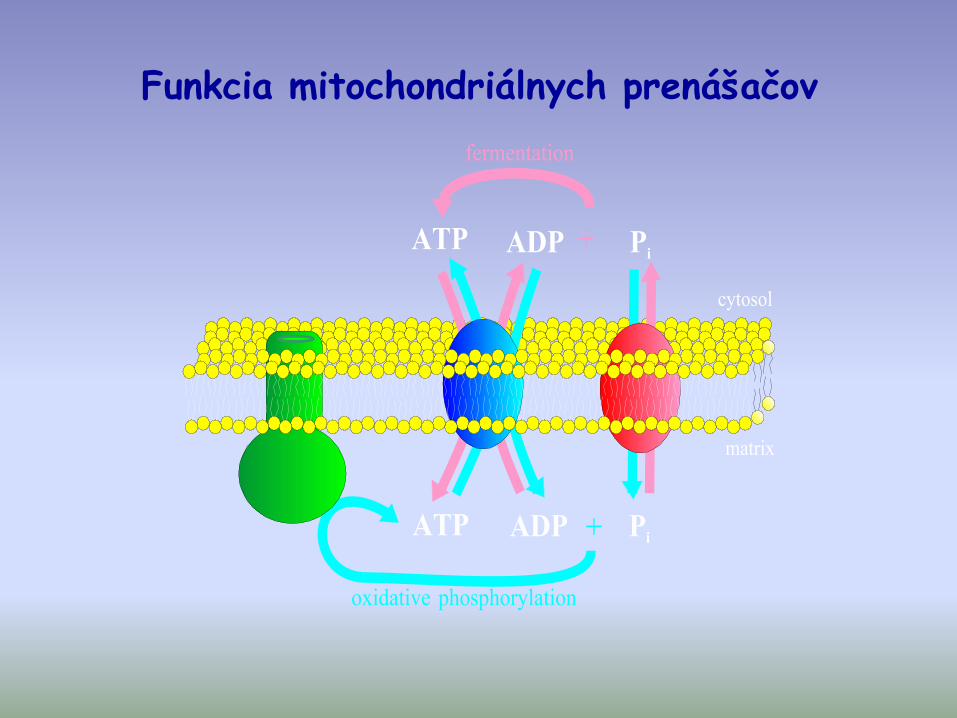
Funkcia mitochondriálnych prenášačov
Mitochondrie (CBB) - - + + + + - - + +
Trypsin - + - + - + - + + +
ATP + + +
„Post-translačný“ import proteínov do mitochondrie
PAGE
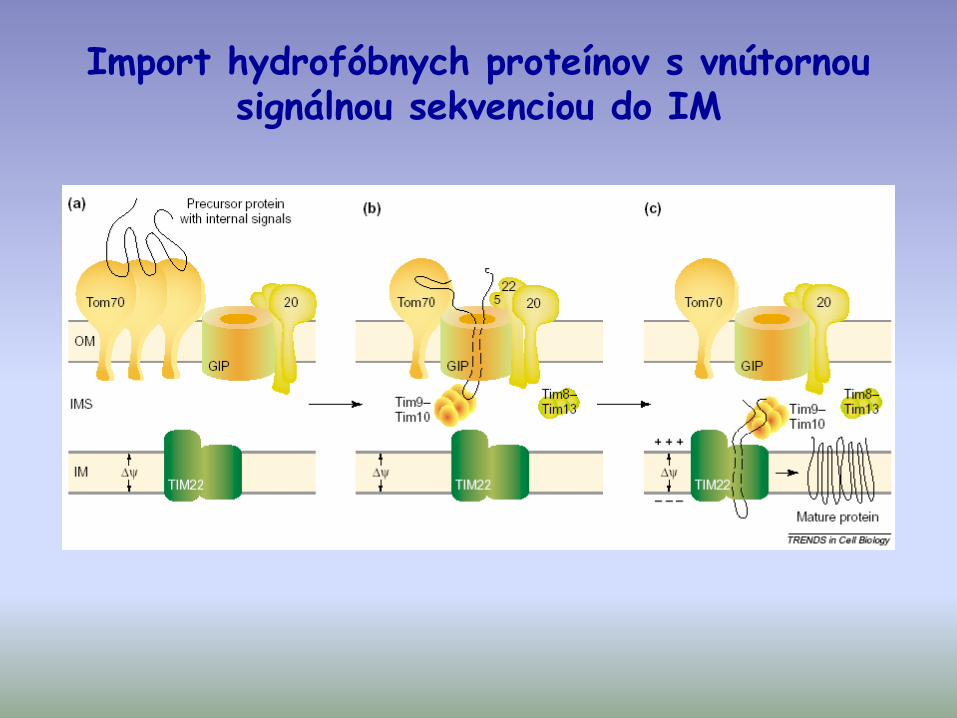
Import hydrofóbnych proteínov s vnútornou signálnou sekvenciou do IM
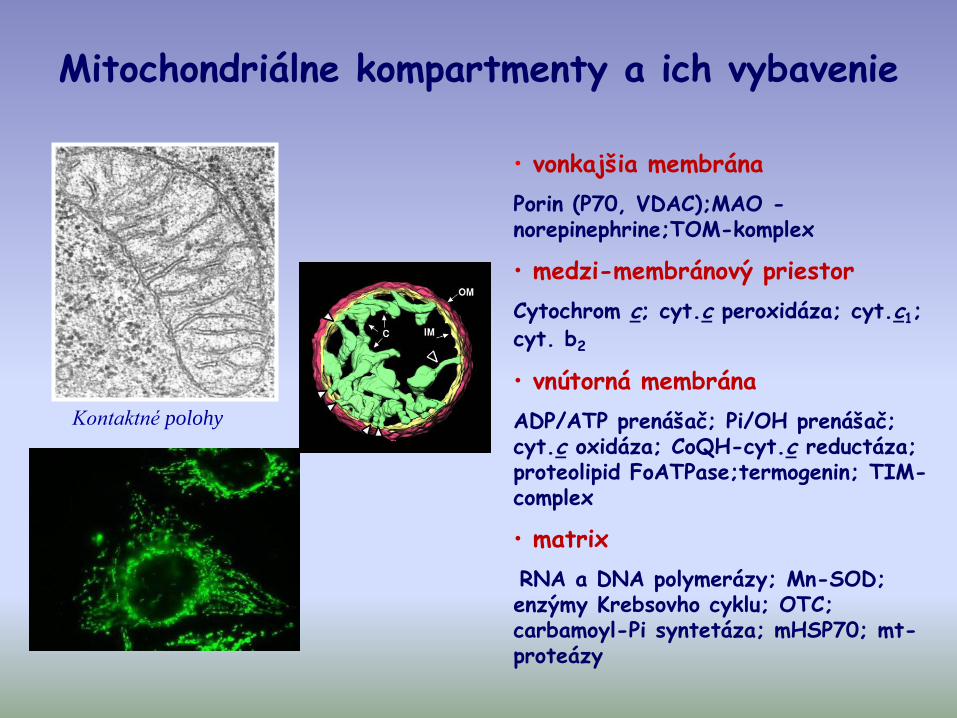
Mitochondriálne kompartmenty a ich vybavenie
• vonkajšia membrána
Porin (P70, VDAC);MAO -norepinephrine;TOM-komplex
• medzi-membránový priestor
Cytochrom c; cyt.c peroxidáza; cyt.c1; cyt. b2
• vnútorná membrána
ADP/ATP prenášač; Pi/OH prenášač; cyt.c oxidáza; CoQH-cyt.c reductáza; proteolipid FoATPase;termogenin; TIM-complex
• matrix
RNA a DNA polymerázy; Mn-SOD; enzýmy Krebsovho cyklu; OTC; carbamoyl-Pi syntetáza; mHSP70; mt-proteázy
Kontaktné polohy
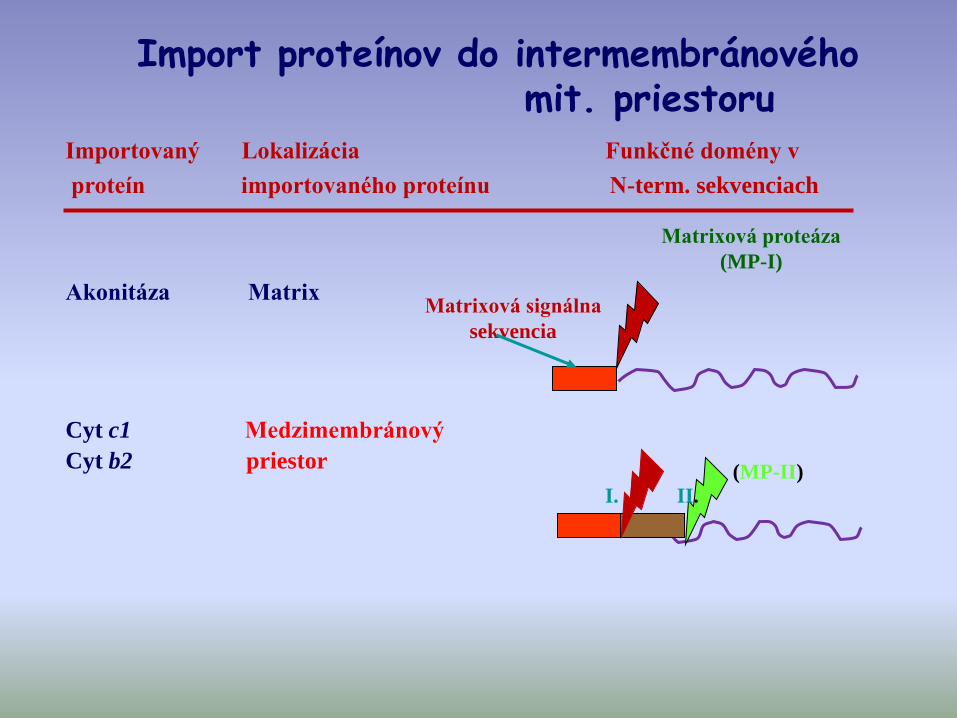
Importovaný Lokalizácia Funkčné domény v
proteín importovaného proteínu N-term. sekvenciach
Matrixová signálna
sekvencia
Matrixová proteáza
(MP-I)
I. II.
Akonitáza Matrix
Cyt c1 Medzimembránový
Cyt b2 priestor(MP-II)
Import proteínov do intermembránovéhomit. priestoru

Hierarchicky usporiadané sekvencie určujú lokalizáciu vo vnútri organel
• Obidve časti signálnejsekvencie majú odlišné zloženie.• N-terminálna časť sa podobá iným mit. signálnymsekvenciám. • Následných 19 AK je nepolárnych a vytvárajú neprerušený segment.
•Ani jedna sekvencia v maturovanom proteíne nie je prítomná.
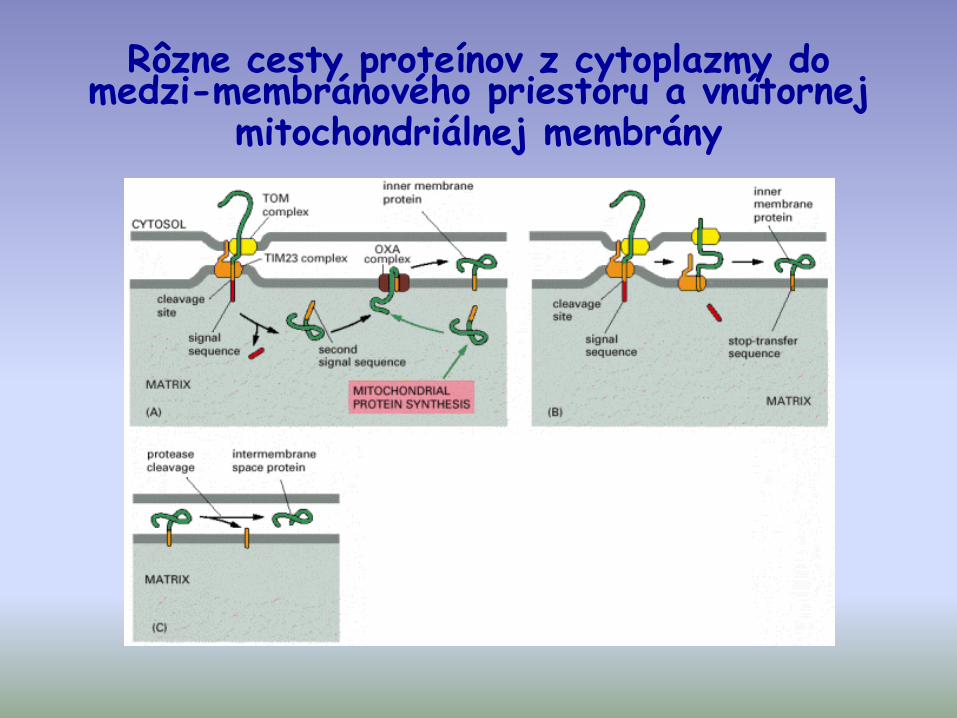
Rôzne cesty proteínov z cytoplazmy do medzi-membránového priestoru a vnútornej
mitochondriálnej membrány

Import cyt c do intermembránového priestoru -špecifická importná dráha
- bez N-terminálnej pre-sekvencie
- nevyžaduje prot. zložky Tim a Tom
- ireverzibilné umiestnenie v IMS je
zabezpečené zmenou konformácie
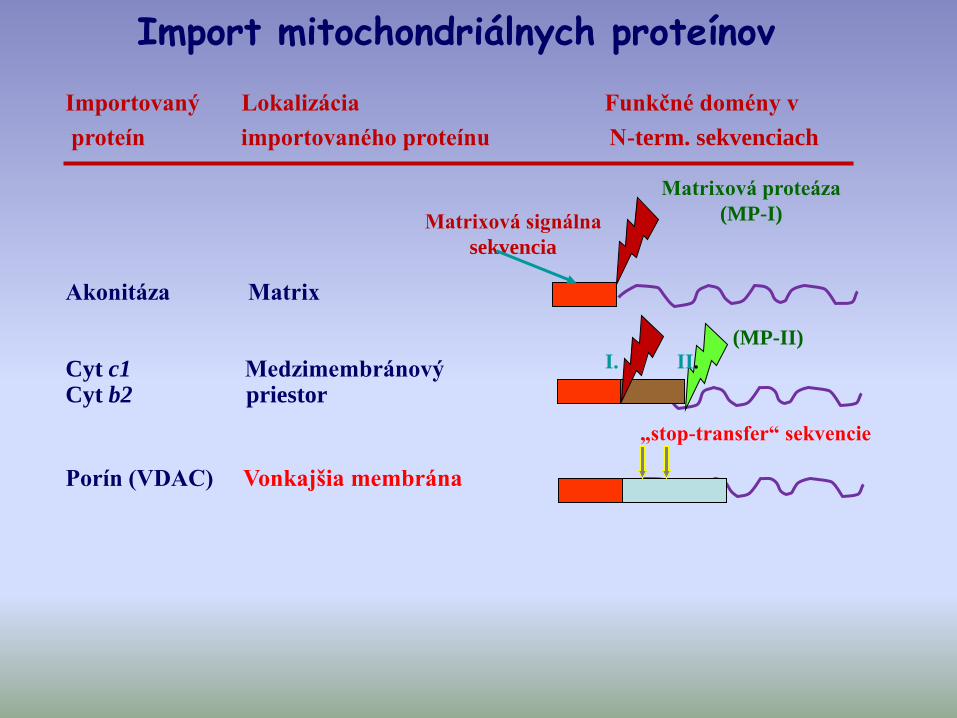
Importovaný Lokalizácia Funkčné domény v
proteín importovaného proteínu N-term. sekvenciach
Matrixová signálna
sekvencia
Matrixová proteáza
(MP-I)
I. II.
„stop-transfer“ sekvencie
Akonitáza Matrix
Cyt c1 Medzimembránový Cyt b2 priestor
Porín (VDAC) Vonkajšia membrána
(MP-II)
Import mitochondriálnych proteínov
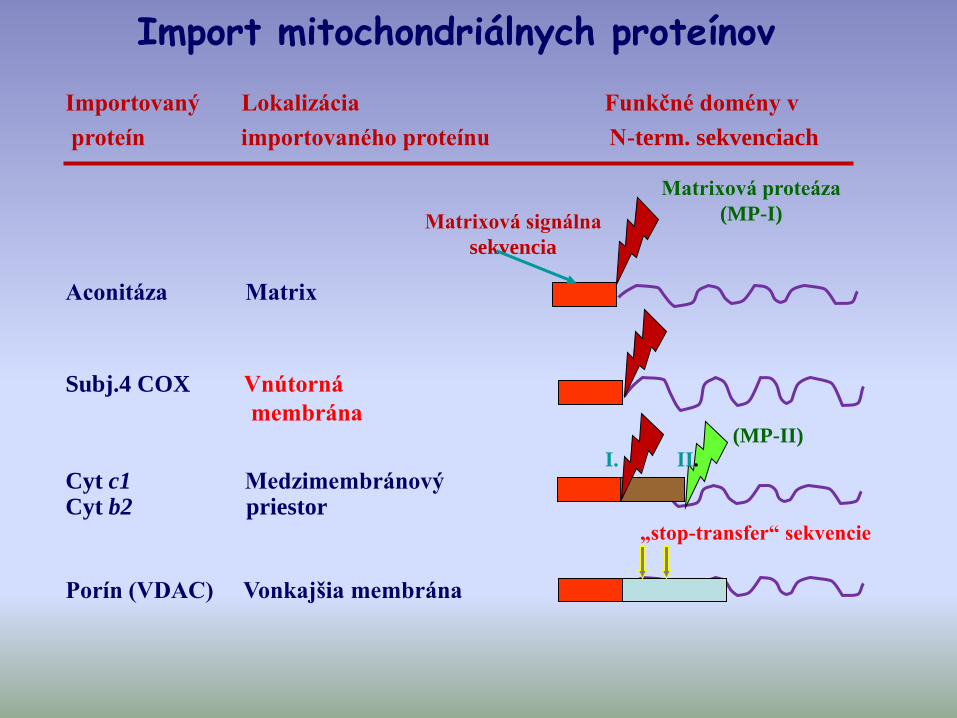
Importovaný Lokalizácia Funkčné domény v
proteín importovaného proteínu N-term. sekvenciach
Matrixová signálna
sekvencia
Matrixová proteáza
(MP-I)
I. II.
„stop-transfer“ sekvencie
Aconitáza Matrix
Subj.4 COX Vnútorná
membrána
Cyt c1 Medzimembránový Cyt b2 priestor
Porín (VDAC) Vonkajšia membrána
(MP-II)
Import mitochondriálnych proteínov

Metódy štúdia importu proteínov :
1. metódy in vitro
2. metódy in vivo
A. vyžadujúce dezintegráciu buniek:
i. stanovenie enzymatickej aktivity
ii. imunochemické metódy
B. nevyžadujúce dezintegráciu buniek:
i. využívajúce fúziu s reportérom (GFP)
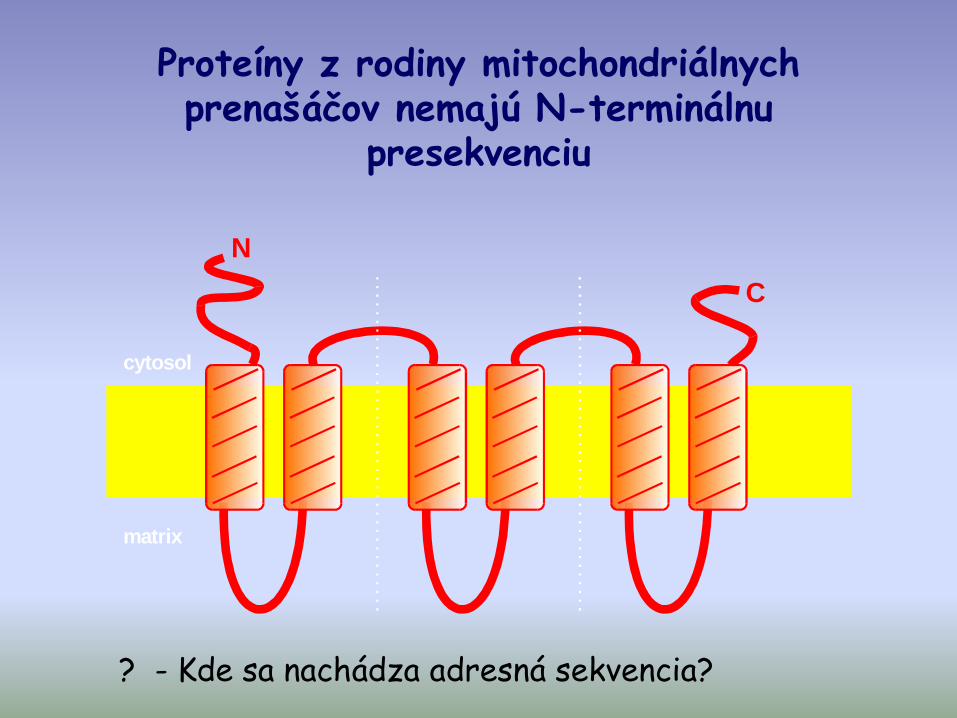
Proteíny z rodiny mitochondriálnych prenašáčov nemajú N-terminálnu
presekvenciu
cytosol
matrix
N
C
? - Kde sa nachádza adresná sekvencia?
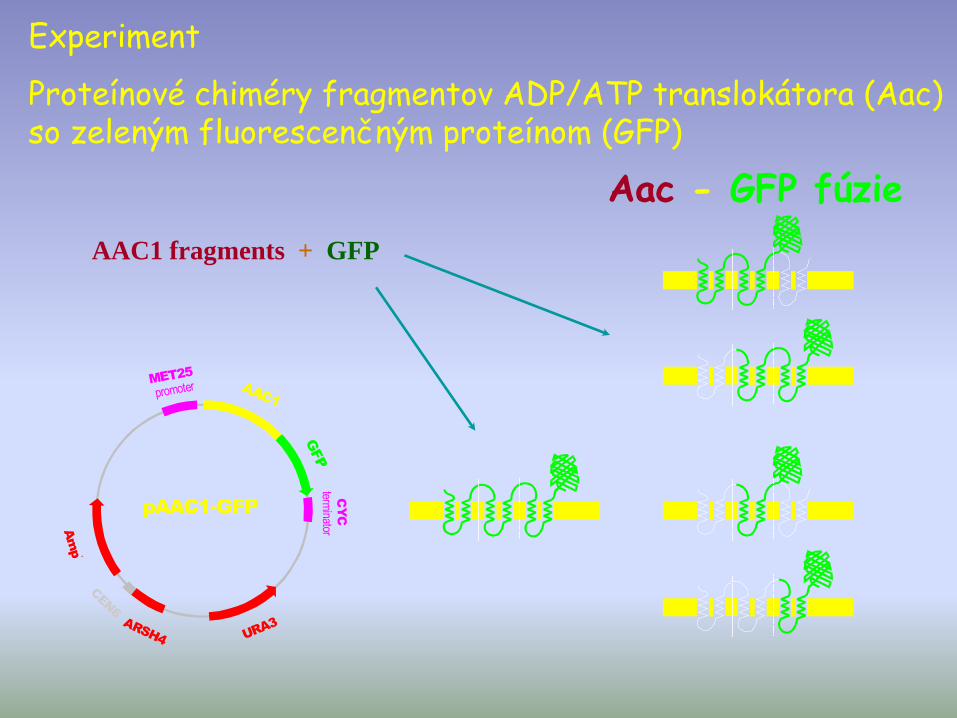
pAAC1-GFP
GFP
MET25
promoter
CYC
terminator
URA3
Amp
r
ARSH4
CEN6
AAC1
Aac - GFP fúzie
AAC1 fragments + GFP
Experiment
Proteínové chiméry fragmentov ADP/ATP translokátora (Aac) so zeleným fluorescenčným proteínom (GFP)
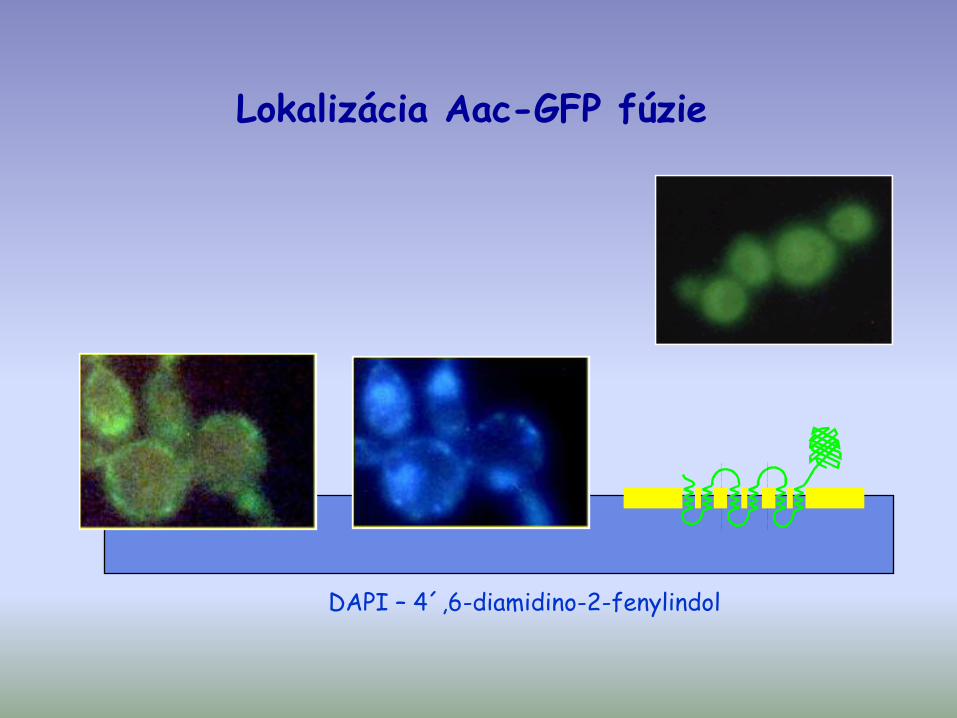
Lokalizácia Aac-GFP fúzie
DAPI – 4´,6-diamidino-2-fenylindol
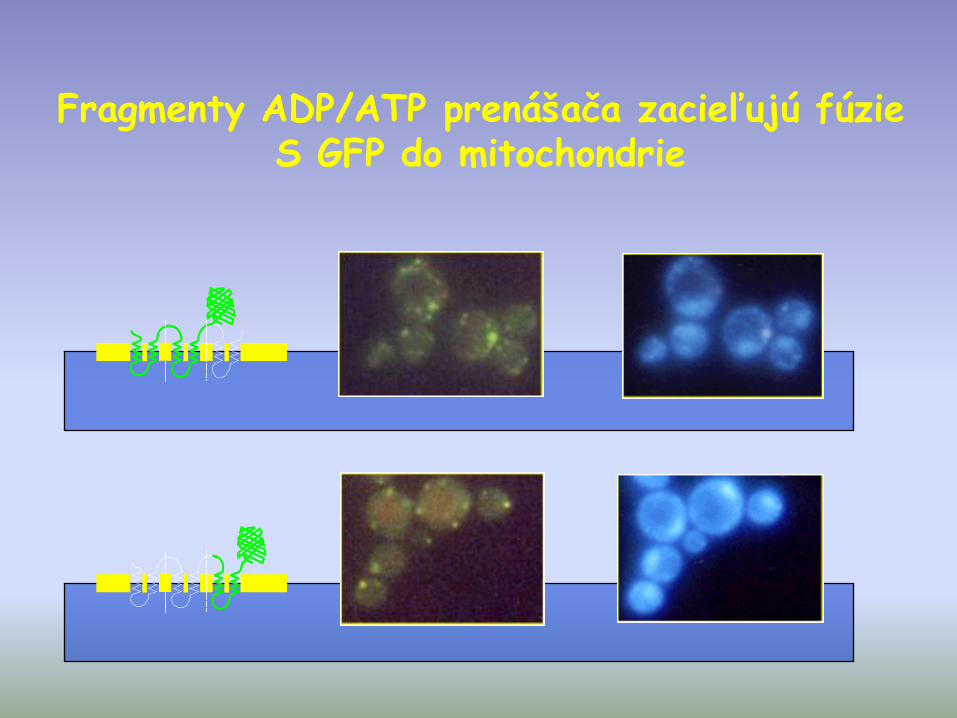
Fragmenty ADP/ATP prenášača zacieľujú fúzieS GFP do mitochondrie

• translokátory vnútornej mitochondriálnej membrány majú viacnásobnú adresnú sekvenciu, ktorá je reprezentovaná v každej transmembránovej slučke
• každá jedná transmembránova slučka obsahuje potrebnú informáciu pre lokalizáciu vo vnútornej mit. membráne.
Záver:
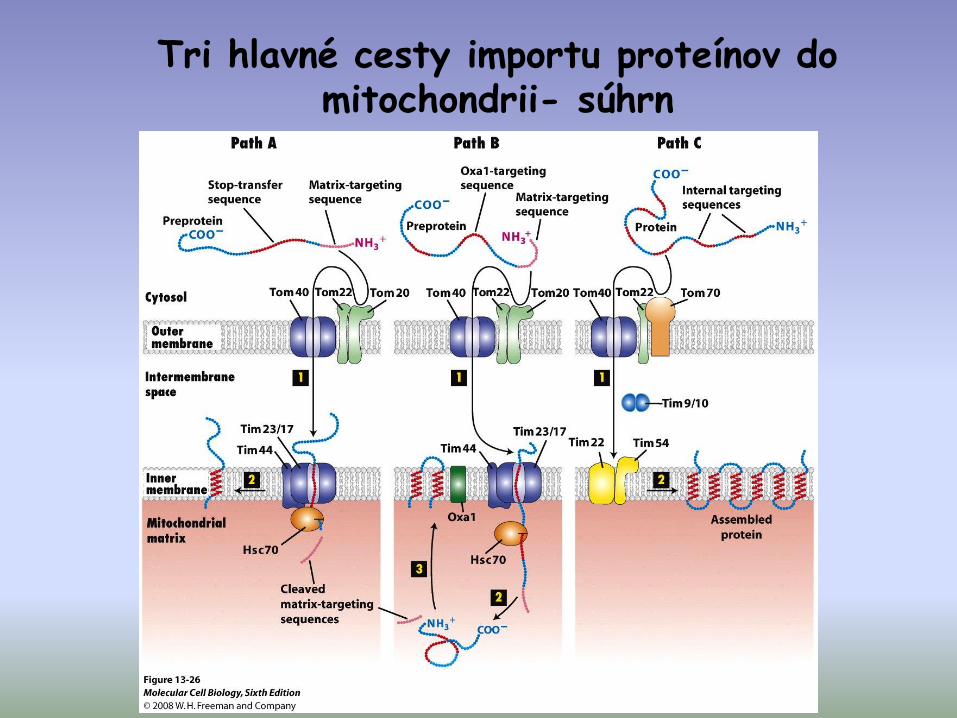
Tri hlavné cesty importu proteínov do mitochondrii- súhrn
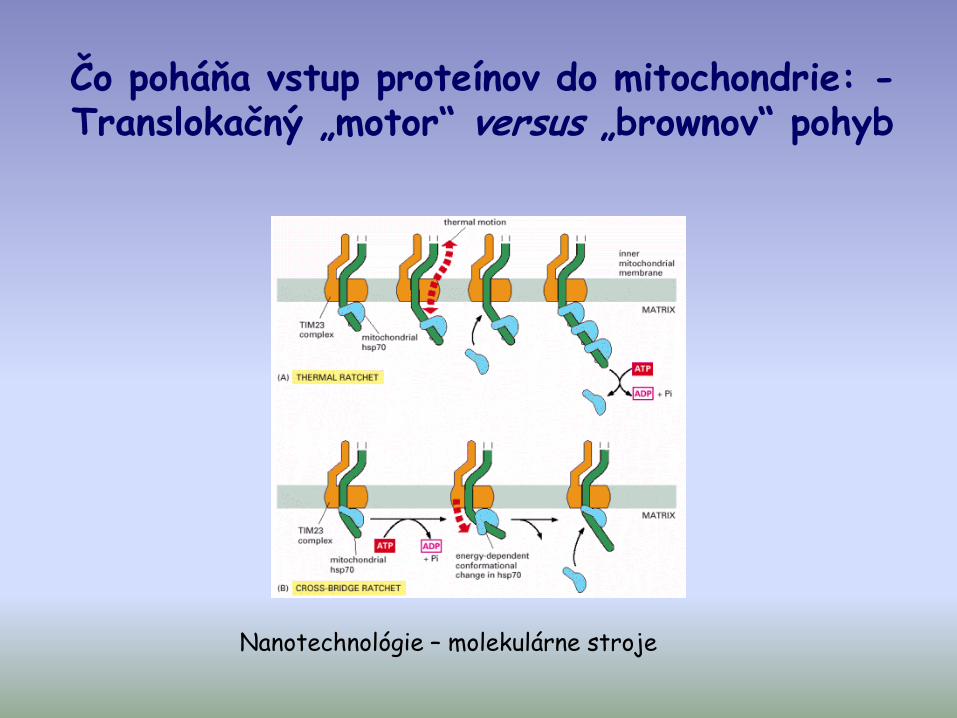
Čo poháňa vstup proteínov do mitochondrie: -Translokačný „motor“ versus „brownov“ pohyb
Nanotechnológie – molekulárne stroje
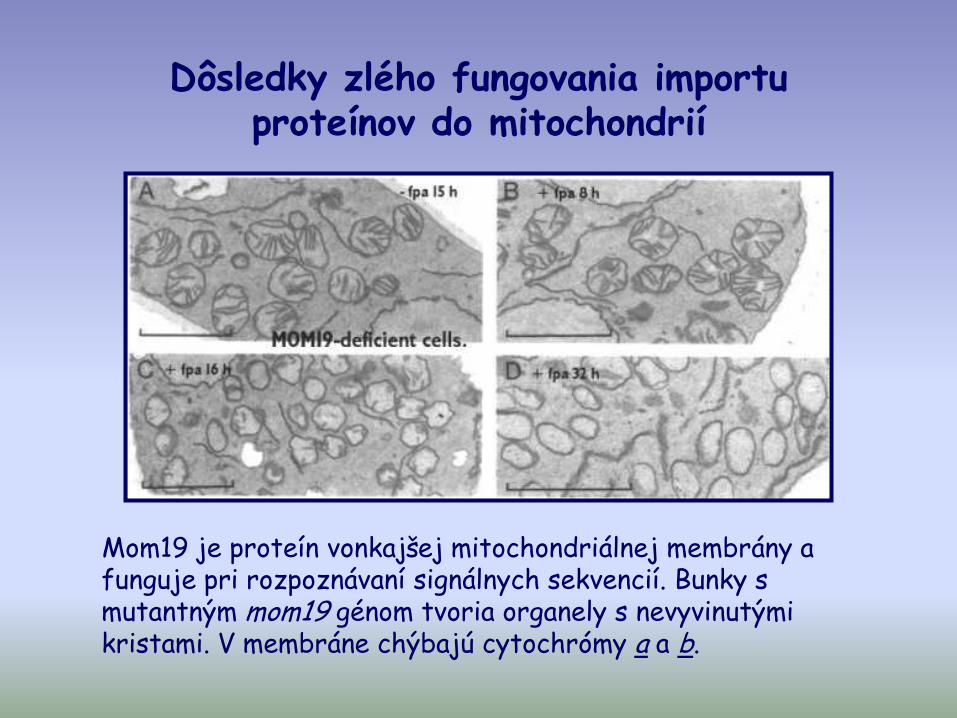
Dôsledky zlého fungovania importu proteínov do mitochondrií
Mom19 je proteín vonkajšej mitochondriálnej membrány a funguje pri rozpoznávaní signálnych sekvencií. Bunky s mutantným mom19 génom tvoria organely s nevyvinutými kristami. V membráne chýbajú cytochrómy a a b.




















