Embed Size (px)
Citation preview

Biocutulysis and Biotrunsfomation, Vol. 19, pp. 4271142 Reprints available directly from the publisher Photocopying permitted by license only
0 2001 OPA (Overseas Publishers Association) N.V. Published by license under
the Hanvocd Academic Publishers imprint, part of Gordon and Breach Publishing
a member of the Taylor & Francis Group. All rights reserved.
PROTEINS ACCOMPANYING THE ESTROGEN RECEPTOR a AND p: A MODEL FOR STUDYING
PROTEIN HETERO-COMPLEXES
ELISABETH JISAa, KLAUS GRAUMANNb and L O I S JUNGBAUERa’*
aht i tUte of Applied Microbiology, University of Agricultural Sciences, Muthgasse 18, Vienna A-1190, Austria; bBiochemie GmbH, Kundl, Austria
(Received 6 April 2000; Revised 15 November 2000)
Estrogen receptor a forms a highly dynamic protein complex in its activated and inactivated state. The protein is complexed with heat shock proteins and other accompanying proteins. Upon ligand binding these proteins are shed off, the receptor dimerizes and forms a preinitiation complex with coactivators andlor corepressors. A plethora of proteins has been discovered, associated with this complex. These different proteins may either up- or down-regulate estrogen receptor-mediated transcription of target genes. Real time-biosensor technology is one approach to assessing these extremely dynamic protein complex formations.
Keywords: Estrogen; Immunophilins; Ligand Hetero-complexes
INTRODUCTION
Estrogen receptors belong to the family of steroid hormone receptors(SHR) (Evans, 1988). They play extremely important roles in growth and development, reproduction and hormone dependent cancers such as breast cancer. The receptors are associated with other proteins in their active, ligand-friendly state. Upon activation at least some of these proteins are shed off, the receptor dimerizes and is, if not already located there, transported into the nucleus to form an active transcription complex with other proteins. Functionality and fate of the receptor is highly determined by protein hetero-complexes (Graumann and
*Corresponding author. Fax: +43-136006-1249. E-mail: jungbaue@hpOl .boku.ac.at
421
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itat d
e G
iron
a on
11/
10/1
4Fo
r pe
rson
al u
se o
nly.

428 E. JISA et 01.
Jungbauer. 2000). Since the receptor itself is an extremely labile molecule it is very tedious to produce sufficient quantities for biochemical studies. The development of modem real-time biosensor techniques enabled the access to preliminary data on stability and thermodynamic properties of these hetero- complexes. In this review we describe the nature of the hetero-complex of unligandedinactive as well as ligandedactivated receptor and the methodology to measure these complexes using real-time biosensor technology.
Like any other members of the nuclear receptor superfamily estrogen receptor a and P (ERa and ERP) are composed of five different domains, which may differ in amino acid homology, but have similar function. The N-terminal A B domain is highly variable in sequence and length. It contains a ligand- independent transactivation function (AF- 1) which activates target gene transcription by directly interacting with components of the transcription machinery or with coactivators that mediate signaling to downstream proteins (Tora et al., 1989; Webb et al., 1998; Tremblay et al., 1999). The C domain comprises DNA binding function and dimerization sequences. Two zinc fingers are responsible for specific interaction between receptor molecules and estrogen response elements. The E domain is responsible for ligand binding, nuclear localization and ligand-dependent transactivation (AF-2) (Webster et al., 1988). The hinge domain between domains C and E contributes flexibility, and represents an anchor for corepressor proteins. The C-terminal F domain also contributes to transactivation capacity.
In 1996 a novel form of the estrogen receptor was discovered (Kuiper et al., 1996; Mosselman et al., 1996; Ogawa et al., 1998) and is referred to as ERP with the original form now known as ERa. ERP is able to partially substitute for the function of ERa in ERa knock out mice (Krege et al., 1998; Couse et al., 1999).
PROTEINS COMPLEXED WITH UNLIGANDED ERa AND ERP
As with many other intracellular proteins, SHR, such as ERa and ERP, form multi-protein complexes with members of the heat shock and immunophilin protein families (Pratt et al., 1996; F’ratt and Toft, 1997; Pratt, 1998). In their ligand free state, SHRs are found associated with hsp90 and the immunophilins FKBP52 or cyclophilin 40 (Cyp-40), as well as other proteins. Although it should be mentioned that not all of the proteins listed below have been detected in unliganded ERa or ERP hetero-complexes, most concepts seem to be true-at least for cytosolic ER.
Not surprisingly, one major role of these proteins is related to protein folding during receptor synthesis (Rutherford and Zuker, 1994). A core chaperone
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itat d
e G
iron
a on
11/
10/1
4Fo
r pe
rson
al u
se o
nly.

PROTEINS ACCOMPANYING THE ESTROGEN RECEPTOR a AND J3 429
complex formed of hsp90, p60 (Hop-heat shock protein organizing protein) and hsp70 binds nascent polypeptides (Chen et al., 1996). Other proteins like p23, hsp40 or p48 (Hip-heat shock interacting protein) are also involved (Johnson and Toft, 1995; Prapapanich et al., 1996; Kosano et al., 1998). In general, these protein hetero-complexes are highly dynamic, members are partly interchange- able. p60 or hsp70 have been detected in early stages of receptor complex formation while immunophilins replace them during a process called maturation. During this maturation process receptor molecules are not simply folded but transformed into a ligand-friendly state (Freeman et al., 1996).
Some aspects of receptor translocation from the cytosol to the nucleus and possible recycling are still rather unclear. Despite the high level of complexity due to the number of “players”, some light has been shed on hetero-complex formatiodcomposition and possible roles of individual complex members during the last years. Table I lists members of receptor hetero-complexes and their possible functions.
Hsp90 is one of the most abundantly expressed proteins in the cell and plays important roles in protein folding, translocation and also in cell signaling (Pratt, 1998). It forms the core of all known receptor hetero-complexes and is a pre- requisite for high affinity ligand binding. The stoichiometric ratio of hsp90 nuclear receptor molecule interaction appears to be two hsp90 molecules to each nuclear receptor molecule (Segnitz and Gehring, 1995). It is generally accepted that hsp90 is a nucleotide binding protein (Grenert et al., 1999). Graumann and Jungbauer (2000) have recently demonstrated that ATP changes the cooperativity of interaction between hsp90 and other relevant proteins, such as Hop or FKBP51 and FKBP52.
Hop is a member of early receptor hetero-complexes and, since hsp90 and hsp70 do not directly interact with each other, it forms a molecular bridge between these two chaperones. However, very recently Hop has been found to be
TABLE I Overview of biological functions of proteins accompanying unliganded SHRs
Protein Nucleotide Immuno suppression Cell-type Role in Role in Protein folding binding drug binding specijc action translocation transactiwation
hsp90 yes yes Yes ?
P60 FKBP51FKBP52 (yes) Yes Yes Yes cyp-40 Yes Yes ?
hsp40 yes (?I P23 1 ? ?
hsp70 yes yes
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itat d
e G
iron
a on
11/
10/1
4Fo
r pe
rson
al u
se o
nly.

330 E. JISA et nl.
not essential for the conversion of steroid receptors into hetero-complexes and a ligand binding state (Morishima et al., 2000).
Hsp70 itself has been found in early receptor complexes. Nucleotide binding modulates the interaction with substrate polypeptides. So far, its role in receptor hetero-complexes seems to be related to protein folding. However, as for hsp90, other functional proteins interact with the hetero-complex via hsp70. In this context, cofactors like BAG- 1 have been identified as modulators for chaperone activity (Kanelakis er al., 1999).
Immunophilins (FKBP.5 1, Fl(BP52, Cyp-40) possess peptidyl-prolyl isomer- ase function and therefore protein folding activity. They have also been linked to receptor shuttling in the cell (Pratt et al.. 1999). Another highly interesting feature of this class of proteins is that they are targets for immunosuppressant drugs such as rapamycin, FK506 or cyclosporin A. Therefore, these proteins form a crosslink to immunological function. Immunophilins, p60 (Hop) and hsp90 molecules interact via tetratricopeptide repeat (TPR) motifs with each other (Carrello et al., 1999). These TPR motifs are formed by a different number of rather distant amino acid patches.
More recently, p23 has been identified as a member of receptor hetero- complexes. Additionally this hsp90-binding protein has been linked to the ligand activation process (Kazlauskas el al., 1999) and DNA-receptor complex formation and therefore with transcription (Freeman et al., 2000). Hsp40 has been debated as an essential protein for receptor hetero-complex formation. A recent study with purified hsp90 and hsp70 showed that, as for Hop, hsp40 is not essential for hetero-complex formation. The multitude of proteins with different and rather diverse properties makes it clear that investigation of receptor hetero- complexes is a tool to learn more about functional cellular networks.
COFACTORS OF ER (Y AND f3
After activation by a hormone or hormone mimic the hetero-complex dissociates. Estrogen receptors stimulate gene transcription assembling into a stable preinitiation complex associated with basal transcription factors like TATA- binding protein (TBP) and TFIIB in order to serve as bridging proteins to RNA polymerase I1 (Horwitz er al., 1996; Smith et al., 1996 and references therein). For efficient hormone-controlled transcription a further class of proteins is required-the so-called coactivators and corepressors. Depending upon the cell type various coactivators and corepressors are present, the final composition of preinitiation complexes will depend on the composition of cofactors present in a given cell. A common characteristic for nuclear coactivator proteins is the
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itat d
e G
iron
a on
11/
10/1
4Fo
r pe
rson
al u
se o
nly.

PROTEINS ACCOMPANYING THE ESTROGEN RECEPTOR a AND p 43 1
signature sequence LXXLL mediating efficient and sufficient binding to liganded nuclear receptors (Heery et al., 1997). Prominent representatives of coactivators belong to the SRC-family consisting of SRC-1, TIF-2/GRIPl and RAC3/ACTR/pCIP/AIB- 1 (Leo and Chen, 2000).
Steroid receptor coactivator-1 (SRC-1) was shown to enhance transcription by ERa bound to ligands with mixed agonisthtagonist properties, but to inactivate ERa occupied by pure antiestrogens. Members of the SRC-family recruit additional coactivators such as p300/CBP in order to enhance transactivation (Onate et al., 1995; Horwitz et al., 1996; Leo and Chen, 2000). SRC-1 is a genuine coactivator for steroid receptor target gene expression acting by direct contact with receptor molecules (Onate et al., 1995). In contrast SRC-1 enhances transactivation by ERP both in the presence and absence of hormone suggesting distinct molecular mechanisms regulating the transcriptional activity of the two receptor subtypes. In vitro phosphorylation of AF-1 by MAP kinases leads to the recruitment of SRC-1 by ERP (Tremblay et al., 1997; Tremblay et al., 1999).
Another two proteins mediating the functions of activation function AF-2 are TIF-la and TIF-2. Like other coactivators TIF-la is able to enhance ER- dependent transcription in the presence of hormone by interacting with activation function AF-2. This interaction is selective for certain members of the nuclear receptor family including ER, TR, RAR, RXR and VDR (ThCnot et al., 1997). In the absence of hormone TIF-la interacts better with ERP independently of the presence of estrogen response element (ThCnot et al., 1997). TIF-2 could be characterized to act similar to TIF-1 exhibiting typical characteristics of nuclear receptor coactivators (Voegel et al., 1996).
A transcriptional coactivator for the C-terminal transactivation domain is glucocorticoid receptor interacting protein 1 (GRIPl), presumed to be the ortholog to human TIF-2. It binds to the ligand binding domain of steroid receptors in the presence but not in the absence of hormone, acting as a transcriptional coactivator for steroid receptors (Hong et al., 1997; Norris et al., 1998).
Receptor-associated coactivator 3 (RAC3) is highly related to SRC-1 and TIF- 2, enhancing ligand-dependent transcriptional activation. In addition it contains a transcriptional activation domain. (Li et al., 1997). RAC3 was found to activate transcription of several nuclear receptors, including the recently cloned estrogen receptor beta, exhibiting different alpha-helical LXXLL motifs of RAC3 for interaction (Leo et al., 2000).
Another member of the SRC-1 family is represented by the coactivator amplified in breast cancer-1 (AIB-1). AIB- 1 over-expression and amplification could be observed in a majority of ERa-positive breast and ovarian cancer cell lines. AIB-1 is able to interact with ERa in a ligand-dependent manner and to
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itat d
e G
iron
a on
11/
10/1
4Fo
r pe
rson
al u
se o
nly.

432 E. JISA et ul.
enhance target gene transcription (Anzick et al., 1997). AIB-1 is suspected of inducing estrogen-independent proliferation of breast and ovarian tumors when present in cell lines expressing mutated ERa binding AIB-1 or other members of the SRC-family hormone-independently. AIB-1 is able to interact with the ligand binding domain of a constitutively active ERa in vitro and to activate basal transcription when cotransfected (Eng et al., 1998).
In 1995 yet another coactivator RIP140 was characterized to interact with the AF-2 domain of ERor in virro. This interaction can be enhanced in the presence of estrogens but not of antiestrogens in vitro and in vivo (Cavailles et al., 1994; Cavailles et al., 1995). RIP140 was shown to block the stimulatory effect of TIF-2 on ER function suggesting a competitive mode of action of various coactivators. Further RIP140 may act as a negative regulatory factor in target gene transactivation. A remarkable observation was the fact that RIP140 can interact with the ligand binding domain of ERa in the presence of the pure antiestrogen ICI 164,384. This indicates a potential of the coactivator RIP140 to induce a switch of antagonistic effects into agonistic effects under certain conditions (Eng et al., 1998).
A novel nuclear receptor coactivator RAP250 has been shown to enhance the transcriptional activity of several nuclear receptors in a ligand-dependent or ligand-enhanced manner. It involves only one short LXXLL motif and exhibits an intnnsic activation domain (Caira et al., 2000).
p300KBP function as cointegrators for nuclear receptors and were found to exhibit histone acetyltransferase activity (Bannister and Kouzarides, 1996). In addition they associate with RNA polymerase I1 (Lee et al., 1999) suggesting a stimulatory role in target gene transcription by this enzyme. CBP was found to act synergistically with SRC-1 in ligand-dependent steroid receptor transactivation (Smith et al., 1996) and to interact with SRC-related proteins to augment nuclear receptor transactivation (Tremblay et al., 1997 and references therein). Kraus and Kadonaga, 1998 present a two stroke model for transcriptional activation by ERa and p300. (Kraus and Kadonaga, 1998).
A transcriptional coactivator p/CIP exists as a complex to CBP mediating activation of nuclear receptor target genes. NCoA-1, highly related to p/CIP is also specifically required for ligand-dependent transcriptional activation of nuclear receptors (Torchia et al., 1997).
Recently further coactivators have been discovered showing typical properties of nuclear hormone receptor coactivators, e.g. Tip60 (Brady et al., 1999), mZaclb acting as a coactivator and corepressor (Huang and Stallcup, 2000), ASC-2 (Lee et al.. 1999). Interestingly, an orphan nuclear receptor SHP (short heterodimer partner) has been shown to interact with estrogen receptors
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itat d
e G
iron
a on
11/
10/1
4Fo
r pe
rson
al u
se o
nly.

PROTEINS ACCOMPANYING THE ESTROGEN RECEPTOR a AND p 433
a and p and to inhibit their transactivation. S H P binds to the AF-2 domain of agonist-bound estrogen receptors via LXXLL-related motifs. SHP is suggested to exert its role as a negative coregulator via formation of ternary complexes with dimeric receptors on DNA (Johansson et al., 2000).
In contrast to factors activating target gene transcription, repression occurs upon association of unliganded estrogen receptors with negative coregulators such as nuclear receptor corepressor (N-CoR) and silencing mediator for RAR and TR (SMRT) (Lavinsky et al., 1998). N-CoR was found to exist complexed with a histone deacetylase, which is required for repression of N-CoR. Presumably a switch from a complex containing histone deacetylase to one containing histone acetylase induces a switch from target gene repression to activation (Heinzel et al., 1997).
ASSESSING PROTEIN HETERO-COMPLEX BY REAL-TIME BIOSENSOR TECHNOLOGY
The interaction of proteins and DNA can be assessed either in free solution or with one partner immobilized to a surface. An overview of methods is shown in Fig. 1. Depending on the type of assay the molecular assembly, the stoichiometry, the binding kinetics, affinity and thermodynamics (free energy and enthalpy) can be measured. Surface plasmon resonance can be used to quantify the complete picture of interaction of macromolecules except conformational changes and the size of the complex. To some extent the conformational change is reflected by the entropy change.
Both specificity of interaction and affinity can be assessed in real-time by biosensor systems based on SPR (O’Shannessy et al., 1993; O’Shannessy et al., 1994; Silin and Plant, 1997; Fivash et al., 1998). In Fig. 2 the interaction of hsp70 with p60, hsp90 is shown. After sample injection, analyte interacts with ligand immobilized to the sensor surface, which gives the association phase until a steady state is reached where association and dissociation rates are in equilibrium. As soon as the analyte injection is stopped and replaced by buffer flow, dissociation of analyte begins. Association and dissociation phase can be employed to derive kinetic constants. Assuming pseudo-first order kinetics,
A + B ~ A B kd
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itat d
e G
iron
a on
11/
10/1
4Fo
r pe
rson
al u
se o
nly.

434 E. JISA et al.
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itat d
e G
iron
a on
11/
10/1
4Fo
r pe
rson
al u
se o
nly.

PROTEINS ACCOMPANYING THE ESTROGEN RECEPTOR a AND p 435
the rate of complex formation during analyte injection is given by
where dRldt is the rate of change of the SPR signal, k, is the association, kd the dissociation rate constant, C is the analyte concentration, R,, the maximum analyte binding capacity in response units (RU), and R, is the signal at time t. The integral form of Eq. (2), expanded by Ri the background binding in response units (RU)
has been used to fit the response curve during the association phase. From these fits k, [M-ls-'], kd [s-l] and KA [M-'1 can be estimated.
The rate of dissociation can be expressed by
and the integrated form
can be used to fit response curves during the dissociation phase. Here Ro is the response at the beginning of the dissociation phase and R(t-03) is the response at infinite time accounting for the background signal. The dissociation constant can be estimated from this fit. At equilibrium, association and dissociation rates are equal and the equilibrium binding constant given by
ka KA = - k d
According to O'Shannessy et al., 1993, the value for kd obtained from first order approximation of a time-span of the dissociation phase can be inserted into the fitting equation for the adsorption phase (O'Shannessy and Winzor, 1996). The resulting values for k, and R,,, show higher confidence. Additionally, injection and analysis of different analyte concentrations can reveal deviations from first order kinetics and limitations due to mass transfer problems (Glaser, 1993). To avoid these problems highly reliable binding constants were
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itat d
e G
iron
a on
11/
10/1
4Fo
r pe
rson
al u
se o
nly.

336 E. JISA er nl.
determined by combining data sets from the maximal response at equilibrium (Req) for each injected protein concentration. Assuming a Langmuirian adsorption characteristic (Langmuir, 191 8), Re, is related to the equilibrium concentration C* by
KAC* 1 + K A C * '
Re, = Rmax (7)
C* corresponds to the injected analyte concentration (Co). Bound analyte is plotted against the ratio of bound (Req) and free analyte (C*) yielding the so- called Scatchard plot. The slope of the curve gives KA. The underlying hypothesis of this model is an interaction where the ligand binding sites do not interfere with each other during association and dissociation phases. When cooperative interaction takes place, the relationship between the equilibrium concentration bound to the solid phase (Req) and the analyte concentration can be described by the Hill-plot (Hill, 1910)
where n is the cooperativity coefficient. If n > 1.0. positive cooperativity is occurring. If 0 < n < 1. negative cooperativity is observed. For positive cooperativity, the Scatchard plot results in a characteristic curve-linear shape; for negative cooperativity the curve rapidly increases with lower bound analyte amounts (Re,).
Roos et al. (1998) have shown that the change in entropy (AS) and the change in enthalpy ( A H ) can be also assessed by real-time measurements AG, the change in free energy can be are expressed as
AG = A H - TAS. (9)
The most informative thermodynamic analysis uses kinetic data and applies transition state analysis of rate constants. following the Eyring theory (Roos et al., 1998). The kinetic rate constant of adsorption and desorption can be easily derived from the response curves of real-time biosensor measurements. k can be expressed as a function of AH* and AS*; the change of enthalpy and entropy in the transition state.
X~ AH* AS* +- k = -.e - ~
h RT R
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itat d
e G
iron
a on
11/
10/1
4Fo
r pe
rson
al u
se o
nly.

PROTEINS ACCOMPANYING THE ESTROGEN RECEPTOR Q AND p 437
k is the first order constant, kd (s-l), his the Planck’s constant (6.63 X Js), x is the Boltzmann’s constant (1.3806503 X JK-’) and Tis the temperature (K).
The asterisk indicates the transition state. From Eq. (10) a plot of ln2khlxT versus 1/T results in a straight line with a slope -AH*/R and an intercept of AS*/R.
kh AH* AS* In-=-- +-
XT R R
The free energy (AG), enthalpy (AH) and entropy (AS) changes at equilibrium can be derived from the transition state. The energies derived from the association rate constant are subtracted from the energies derived from the dissociation rate constant yielding into the free energy of the interaction.
Real-time biosensor measurements can be usually accomplished at different temperatures with a high accuracy. Therefore these experimental data are available. As pointed out in the previous two sections, a multitude of interactions is taking place with estrogen receptors. The experimental basis has been laid down to quantify these molecular events.
For a complete quantification of the interaction of macromolecules SPR-real time measurements (ka, k d , kA, AG, AH, AS), differential calorimetry (thermal stability) and electrophoretic mobilizing shift assay or analytical ultra centrifugation provide an ultimate view.
In Fig. 2 examples for measuring the interaction of the estrogen receptor and the respective accompanying proteins by SPR are shown. From these data the dynamics of complex formation could be derived. First a 6xhis-tagged p60 was pumped over a Ni-chelate chip, after continuing with running buffer hsp90, p23 and hsp70 were sequentially injected. The response signal increased with hsp90 and hsp70; not with p23 (Fig. 2A). This indicates that p23 does not take part in the interaction of p60 and hsp90.
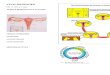
In Fig. 2B a similar experiment is shown with FKBP52, hsp70 and hsp90. From such analysis and painvise measurements of the reaction partners the size and composition of such a complex was developed (Fig. 3). Further detailed analysis was described by Graumann and Jungbauer, 2000.
Recently Myszka et al. (1998) have reported a simple two-compartment model for evaluation of real-time biosensor data to take the mass transport resistances
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itat d
e G
iron
a on
11/
10/1
4Fo
r pe
rson
al u
se o
nly.

438
23ooo ~
.
5 22Mx)- K m C
Y
c. - 2 21000-
g 2 2oooo:
a0 c
m
19ooo?,
E. JISA et a).
N a C L FKBP52 hspW
: I c 1 1
.
, 1 . 3 I . I . I , , . , , , , . I
into account. The model proposed by O’Shannessy and Winzor (1996), which has been discussed in detail, neglects mass transfer resistance of the migration of the solutes from the bulk liquid into the dextran coated layer. Simple algorithms are proposed by the manufacturers to check the mass transfer limitations. Knowing this fact the inherent influence of mass transfer properties on this kind of “interaction analysis” must be taken into account when measuring protein- protein interactions.
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itat d
e G
iron
a on
11/
10/1
4Fo
r pe
rson
al u
se o
nly.

PROTEINS ACCOMPANYING THE ESTROGEN RECEPTOR (Y AND p 439
FIGURE 3 according to Graumann and Jungbauer (2000).
Schematic presentation of hetero-complex associating human estrogen receptor (Y
References
Anzick, S.L., Kononen, J., Walker, R.L., Azorsa, D.O., Tanner, M.M., Guan, X.Y., Sauter, G., Kallioniemi, O.P., Trent, J.M. and Meltzer, P.S. (1997) ‘‘AIBl, a steroid receptor coactivator amplified in breast and ovarian cancer”, Science 277, 965-968.
Bannister, A.J. and Kouzarides, T. (1996) “The CBP co-activator is a histone acetyltransferase”, Nature 384,641-643.
Brady, M.E., Ozanne, D.M., Gaughan, L., Waite, I., Cook, S., Neal, D.E. and Robson, C.N. (1999) “Tip60 is a nuclear hormone receptor coactivator”, J. B i d . Chem. 274, 17599-17604.
Caira, F., Antonson, P., Pelto-Huikko, M., Treuter, E. and Gustafsson, J.A. (2000) “Cloning and characterization of RAP250, a novel nuclear receptor coactivator”, J. B i d . Chem. 275, 5308-5317.
Carrello, A., Ingley, E., Minchin, R.F., Tsai, S. and Ratajczak, T. (1999) “The common tetrabicopeptide repeat acceptor site for steroid receptor-associated immunophilins and hop is located in the dimerization domain of Hsp90”, J. B i d . Chem. 274,2682-2689.
Cavailles, V., Dauvois, S., Danielian, P.S. and Parker, M.G. (1994) “Interaction of proteins with transcriptionally active estrogen receptors”, Proc. Natl. Acad. Sci. USA 91, 10009- 10013.
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itat d
e G
iron
a on
11/
10/1
4Fo
r pe
rson
al u
se o
nly.

440 E. JISA era / .
Cavailles, V., Dauvois, S., CHorset, F.. Lopez, G.. Hoare, S., Kushner, P.J. and Parker, M.G. (1995) “Nuclear factor RIP140 modulates transcriptional activation by the estrogen receptor”, Embo. J. 14, 3741-3751.
Chen, S., Prapapanich, V., Rimerman, R.A., Honore, B. and Smith, D.F. (1996) “Interactions of p60, a mediator of progesterone receptor assembly, with heat shock proteins hsp90 and hsp70’, Mol. Endocrinol. 10,682-693.
Couse. J.F.. Hewitt, S.C.. Bunch, D.O., Sar, M., Walker, V.R., Davis, B.J. and Korach, K.S. (1999) “Postnatal sex reversal of the ovaries in mice lacking estrogen receptors alpha and beta”, Science 286,2328-2331.
Eng. F.C.. Barsalou, A,, Akutsu, N., Mercier, I., Zechel. C.. Mader, S. and White, J.H. (1998) “Different classes of coactivators recognize distinct but overlapping binding sites on the estrogen receptor ligand hinding domain”. J. B i d . Chem. 273. 28371 -28377.
Evans, R.M. (1988) “The steroid and thyroid hormone receptor superfamily”. Science 240, 889-895. Fivash, M.. Towler, E.M. and Fisher. R.J. (1998) “BIAcore for macromolecular interaction”, Curr:
Opin. BiotechnoL 9, 97 - 10 1. Freeman. B.C.. Toft, D.O. and Morimoto, R.I. (1996) “Molecular chaperone machines: chaperone
activities of the cyclophilin Cyp-40 and the steroid aporeceptor-associated protein p23”, Science 274, 1718-1720.
Freeman. B.C., Felts, S.J.. Toft, D.O. and Yamamoto, K.R. (20oO) “The p23 molecular chaperones act at a late step in intracellular receptor action to differentially affect ligand efficacies”, Genes Dev. 14, 422-434.
Glaser. R.W. ( 1993) “Antigen-antibody binding and mass transport by convection and diffusion to a surface: a two-dimensional computer model of binding and dissociation kinetics”, Anal. Biochern. 213, 152- 161.
Graumann, K. and Jungbauer, A. (2000) “Quantitative assessment of complex formation of nuclear- receptor accessory proteins”. Biochem. J . 345. 627-636.
Grenert, J.P.. Johnson. B.D. and Toft. D.O. (1999) “The importance of ATP binding and hydrolysis by hsp90 in formation and function of protein heterocomplexes”, J. B i d . Chern. 274, 17525- 17533.
Heery, D.M., Kalkhoven, E.. Hoare, S. and Parker, M.G. (1997) “A signature motif in transcriptional co-activators mediates binding to nuclear receptors”, Narrrre 387, 733-736.
Heiarel, T.. Lavinsky. R.M., Mullen. T.M.. Soderslrotii. M., Lahcrty, C.D., Torchia, J . , ’fang, W.M., Brard, G., Ngo, S.D.. Davie, J.R., Seto, E., Eisenman, R.N., Rose. D.W., Glass, C.K. and Rosenfeld. M.G. (1997) “A complex containing N-CoR, mSin3 and histone deacetylase mediates transcriptional repression”. Nature 387. 43-48.
Hill. A.V. (1910) “The possible effects of aggregation of the molecules of hemoglobin on its dissociation”. J. Physiol. (LondJ. iv-vii.
Hong, H.. Kohli. K.. Garabedian, M.J. and Stallcup. M.R. (1997) “GRIPI, a transcriptional coactivator for the AF-2 transactivation domain of steroid. thyroid, retinoid, and vitamin D receptors”, Mol . Cell Biol. 17. 2735-2744.
Horwitz, K.B.. Jackson. T.A.. Bain, D.L., Richer. J.K., Takimoto, G.S. and Tung, L. (1996) “Nuclear receptor coactivators and corepressors”. Mol. Eridocrinol. 10, 1167- 1177.
Huang, S.M. and Stallcup, M.R. (2000) “Mouse zacl, a transcriptional coactivator and repressor for nuclear receptors”. Mol. Cell B i d . 20. 1855-1867.
Johansson, L.. Bavner, A., Thomsen, J.S., Farnegardh. M.. Gustafsson, J.A. and Treuter, E. (2000) “The orphan nuclear receptor SHP utilizes conserved LXXLL-related motifs for interactions with ligand-activated estrogen receptors”, Mol. Cell Biol. 20, 1124- 1133.
Johnson, J.L. and Toft, D.O. (1995) “Binding of p23 and hsp90 during assembly with the progesterone receptor”, Mol. Endocrinol. 9. 670-678.
Kanelakis. K.C.. Morishima. Y., Dittmar, K.D., Galigniana. M.D., Takayama, S., Reed, J.C. andPratt, W.B. ( 1999) “Differential effects of the hsp70-binding protein BAG- 1 on glucocorticoid receptor folding by the hsp90-based chaperone machinery”, J. B i d . Chem. 274. 34134-34140.
Karlauskas, A.. Poellinger. L. and Pongatz. I . (1999) “Evidence that the co-chaperone p23 regulates ligand responsiveness of the dioxin (Aryl hydrocarbon) receptor”. J. Biol. Chem. 274, 1351 9- 13524.
Kosano, H.. Stensgard. B.. Charlesworth, M.C., McMahon. N. and Toft, D. (1998) “The assembly of progesterone receptor-hsp90 complexes using purified proteins”, J . B i d . Chem. 273, 32973 - 32979.
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itat d
e G
iron
a on
11/
10/1
4Fo
r pe
rson
al u
se o
nly.

PROTEINS ACCOMPANYING THE ESTROGEN RECEPTOR cx AND f3 441
Kraus, W.L. and Kadonaga, J.T. (1998) “p300 and estrogen receptor cooperatively activate transcription via differential enhancement of initiation and reinitiation”, Genes Dev. 12, 331-342.
Krege, J.H., Hodgin, J.B., Couse, J.F., Enmark, E., Warner, M., Mahler, J.F., Sar, M., Korach, K.S., Gustafsson, J.A. and Smithies, 0. (1998) “Generation and reproductive phenotypes of mice lacking estrogen receptor beta”, Proc. Natl. Acad. Sci. USA 95, 15677-15682.
Kuiper, G.G., Enmark, E., Pelto-Huikko, M., Nilsson, S. and Gustafsson, J.A. (1996) “Cloning of a novel receptor expressed in rat prostate and ovary”, Pmc. Natl. Acad. Sci. USA 93,5925-5930.
Langmuir, I. (1918) “The adsorption of gases on plane surfaces of glass, mica and platinum”, J. Am. Chem. SOC. 40, 1361-1403.
Lavinsky, R.M., Jepsen, K., Heinzel, T., Torchia, J., Mullen, T.M., Schiff, R., Del-Rio, A.L., Ricote, M., Ngo, S., Gemsch, J., Hilsenbeck, S.G., Osbome, C.K., Glass, C.K., Rosenfeld, M.G. and Rose, D.W. (1998) “Diverse signaling pathways modulate nuclear receptor recruitment of N- CoR and SMRT complexes”, Proc. Natl. Acad. Sci. USA 95, 2920-2925.
Lee, S.K., Anzick, S.L., Choi, J.E., Bubendorf, L., Guan, X.Y., Jung, Y.K., Kallioniemi, O.P., Kononen, J., Trent, J.M., Azorsa, D., Jhun, B.H., Cheong, J.H., Lee, Y.C., Meltzer, P.S. andLee, J.W. (1999) “A nuclear factor ASC-2, as a cancer-amplified transcriptional coactivator essential for ligand-dependent transactivation by nuclear receptors in vivo”, J. Biol. Chem. 274, 34283-34293.
Leo, C. and Chen, J.D. (2000) “The SRC family of nuclear receptor coactivators”, Gene 245, 1-1 1 . Leo, C., Li, H. and Chen, J.D. (2000) “Differential mechanisms of nuclear receptor regulation by
receptor-associated coactivator 3”, J. Biol. Chem. 275, 5976-5982. Li, H., Gomes, P.J. and Chen, J.D. (1997) “RAC3, a steroidnuclear receptor-associated coactivator
that is related to SRC-1 and TIFY, Pmc. Natl. Acad. Sci. USA 94, 8479-8484. Morishima, Y., Kanelakis, K., Silverstein, A., Dittmar, K., Estrada, L. and Pratt, W. (2000) “The hsp
organizer protein hop enhances the rate of but is not essential for glucocorticoid receptor folding by the multiprotein Hsp90-based chaperone system”, J. Biol. Chem. 275, 6894-6900.
Mosselman, S., Polman, J. and Dijkema, R. (1996) “ER beta: identification and characterization of a novel human estrogen receptor”, FEBS Lett. 392,49-53.
Myszka, D.G., He, X., Dembo, M., Morton, T.A. and Goldstein, B. (1998) “Extending the range of rate constants available from BIACORE: interpreting mass transport-influenced binding data”, Biophys. J. 75, 583-594.
Norris, J.D., Fan, D., Stallcup, M.R. and McDonnell, D.P. (1998) “Enhancement of estrogen receptor transcriptional activity by the coactivator GRIP-I highlights the role of activation function 2 in determining estrogen receptor pharmacology”, J. Biol. Chem. 273, 6679-6688.
Ogawa, S., Inoue, S., Watanabe, T., Hiroi, H., Orimo, A., Hosoi, T., Ouchi, Y. and Muramatsu, M. (1998) “The complete primary structure of human estrogen receptor beta (hER beta) and its heterodimerization with ER alpha in vivo and in vitro”, Biochem. Biophys. Res. Commun. 243, 122-126.
Onate, S.A., Tsai, S.Y., Tsai, M.J. and O’Malley, B.W. (1995) “Sequence and characterization of a coactivator for the steroid hormone receptor superfamily”, Science 270, 1354- 1357.
O’Shannessy, D.J. and Winzor, D.J. (1996) “Interpretation of deviations from pseudo-first-order kinetic behavior in the characterization of ligand binding by biosensor technology”, Anal. Biochem. 236, 275-283.
O’Shannessy, D.J., Brigham-Burke, M., Soneson, K.K., Hensley, P. and Brooks, I. (1993) “Determination of rate and equilibrium binding constants for macromolecular interactions using surface plasmon resonance: use of nonlinear least squares analysis methods”, Anal. Bzochem.
O’Shannessy, D.J., Brigham-Burke, M., Soneson, K.K., Hensley, P. and Brooks, I. (1994) “Determination of rate and equilibrium binding constants for macromolecular interactions by surface plasmon resonance”, Meth. Enzymol. 240, 323-349.
Prapapanich, V., Chen, S., Nair, S.C., Rimerman, R.A. and Smith, D.F. (1996) “Molecular cloning of human p48, a transient component of progesterone receptor complexes and an Hsp70-binding protein”, Mol. Endocrinol. 10, 420-43 1.
Pratt, W.B. (1998) “The hsp90-based chaperone system: involvement in signal transduction from a variety of hormone and growth factor receptors”, Proc. SOC. Exp. Biol. Med. 217, 420-434.
212,457-468.
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itat d
e G
iron
a on
11/
10/1
4Fo
r pe
rson
al u
se o
nly.

442 E. JISA et al.
Pratt. W.B. and Toft, D.O. (1997) “Steroid receptor interactions with heat shock protein and immunophilin chaperones”, Endoc: Reu. 18, 306-360.
Pratt. W.B.. Gehring, U. and Toft, D.O. f 1996) “Molecular chaperoning of steroid hormone receptors”, EXS 77, 79-95.
Pratt. W.B., Silverstein. A.M. and Galigniana, M.D. (1999) “A model for the cytoplasmic trafficking of signalling proteins involving the hsp90-binding immunophilins and p50cdc37”, Cell Signal. 11, 839-851.
Roos. H., Karlsson, R., Nilshans, H. and Persson. A. (1998) “Thermodynamic analysis of protein interactions with biosensor tehnology”, J. Mol. Recogn. 11, 204-210.
Rutherford. S.L. and Zuker, C.S. (1994) “Protein folding and the regulation of signaling pathways”, Cell79. 1129-1132.
Segnitz, B. and Gehring, U. (1995) “Subunit structure of the nonactivated human estrogen receptor”, Proc. Natl. Acad. Sci. USA 92. 2179-2183.
Silin, V. and Plant. A. ( 1997) “Biotechnological applications of surface plasmon resonance”, Tibtech
Smith, C.L.. Onate, S.A., Tsai. M.J. and O’Malley, B.W. (1996) “CREB binding protein acts synergistically with steroid receptor coactivator- 1 to enhance steroid receptor-dependent transcription”, Proc. Nurl. Acad. Sci. USA 93, 8884-8888.
Thenot. S.. Henriquet, C.. Rochefort. H. and CavaillGs, V. (1997) “Differential interaction of nuclear receptors with the putative human transcriptional coactivator hTIFl ”, J. Biol. Chem. 272,
Tora. L., White. J.. Brou. C.. Tasset, D., Webster, N., Scheer. E. and Chambon, P. (1989) “The human estrogen receptor has twao independent nonacidic transcriptional activation functions”, Cell 59, 477-487.
Torchia, J.. Rose, D.W.. Inostrora. J., Kamei, Y., Westin, S.. Glass, C.K. and Rosenfeld, M.G. (1997) “The transcriptional co-activator p/CIP binds CBP and mediates nuclear- receptor function”, Nature 387, 677-684.
Tremblay, G.B., Tremblay, A.. Copeland. N.G.. Gilbert, D.J.. Jenkins. N.A., Labrie, F. and Gigutre, V. (1997) “Cloning, chromosomal localization. and functional analysis of the murine estrogen receptor beta”. Mul. Endocrind. 11, 353-365.
Tremblay, A., Tremblay. G.B.. Labrie, F. and Giguere. V. (1999) “Ligand-independent recruitment of SRC-1 to estrogen receptor beta through phosphorylation of activation function AF-I”, MoZ. Cell 3, 513-519.
Voegel, J.J.. Heine, M.J., Zechel. C.. Chambon, P. and Gronemeyer. H. (1996) “TIF2, a 160kDa transcriptional mediator for the ligand-dependent activation function AF-2 of nuclear receptors”, Embo. J. 15, 3667-367s.
Webb. P., Nguyen. P.. Shinsako, J., Anderson, C.. Feng. W.. Nguyen, M.P., Chen, D., Huang, S.M., Subramanian. S., McKinemey, E., Katzenellenbogen. B.S., Stallcup, M.R. and Kushner, P.J. (1998) “Estrogen receptor activation function 1 works by binding p160 coactivator proteins”, Mol. Endocrinol. 12, 1605-1618.
Webster, N.J.. Green. S., Jin. J.R. and Chambon, P. (1988) “The hormone-binding domains of the estrogen and glucocorticoid receptors contain an inducible transcription activation function”, Cell 54. 199-207.
15. 353-359.
12062- 12068.
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
itat d
e G
iron
a on
11/
10/1
4Fo
r pe
rson
al u
se o
nly.