Embed Size (px)
Citation preview

Proteus morganii törzsek szerológiai keresztkapcsolatai
PhD értekezés
írta Dr. Péterfi Zoltán
Doktori iskola vezetője: Prof. Dr. Szolcsányi János Programvezető: Prof. Dr. Emődy Levente Témavezető: Dr. Kocsis Béla
PÉCSI TUDOMÁNYEGYETEM, ÁLTALÁNOS ORVOSTUDOMÁNYI KAR ORVOSI MIKROBIOLÓGIAI ÉS IMMUNITÁSTANI INTÉZET
2007

Proteus morganii törzsek szerológiai keresztkapcsolatai
2
TARTALOMJEGYZÉK
TARTALOMJEGYZÉK......................................................................................................................................... 2 RÖVIDÍTÉSJEGYZÉK .......................................................................................................................................... 4 1. BEVEZETÉS ...................................................................................................................................................... 5 1.1. A keresztkapcsolatok vizsgálataiban szereplő baktériumok ............................................................................ 6 1.1.1. Proteus (Morganella) morganii .................................................................................................................... 6 1.1.2 Escherichia coli.............................................................................................................................................. 7 1.1.3. Salmonella..................................................................................................................................................... 7 1.1.4. Shigella ......................................................................................................................................................... 8 1.2 A Gram-negatív bakteriális sejtfal szerkezete................................................................................................... 8 1.2.1. Citoplazma membrán: ................................................................................................................................... 9 1.2.2. Peptidoglikán réteg: .................................................................................................................................... 10 1.2.3. Periplazmatikus tér:..................................................................................................................................... 10 1.2.4. Külső membrán:.......................................................................................................................................... 10 1.3. A lipopoliszacharid ........................................................................................................................................ 11 1.3.1. Történeti áttekintés:..................................................................................................................................... 11 1.3.2. A lipopoliszacharid szerkezete.................................................................................................................... 12 1.3.3. A lipopoliszacharid genetikája .................................................................................................................... 15 1.3.4. Biológiai hatás – hatásmechanizmus........................................................................................................... 15 1.4. Külső membrán fehérjék (OMP).................................................................................................................... 17 1.5. A keresztkapcsolatok ..................................................................................................................................... 18 2. CÉLKITŰZÉSEK ............................................................................................................................................. 21 3. ANYAG ÉS MÓDSZER................................................................................................................................... 22 3.1. Felhasznált baktériumok: ............................................................................................................................... 22 3.2. Baktériumtenyésztés: ..................................................................................................................................... 22 3.3. A lipopoliszacharid kivonása: ........................................................................................................................ 23 3.4. Külső membrán fehérjék (OMP) kivonása:.................................................................................................... 23 3.5. Oszlopkromatográfia:..................................................................................................................................... 24 3.6. Poliklonális immunsavók előállítása nyúlban: ............................................................................................... 24 3.7. Poliakrilamid gélelektroforézis (PAGE): ....................................................................................................... 25 3.8. Csőagglutináció:............................................................................................................................................. 26 3.9. Indirekt hemagglutináció: .............................................................................................................................. 26 3.10. Dot blot: ....................................................................................................................................................... 27 3.11. Immunoblot:................................................................................................................................................. 27 3.12. ELISA: ......................................................................................................................................................... 28 3.13. Kapilláris elektroforézis (CE): ..................................................................................................................... 29 3.14. Lab-on-a-Chip (microchip) technológia:...................................................................................................... 29 3.15. Vékonyréteg kromatográfia (TLC): ............................................................................................................. 30 3.16. Gázkromatográfia, tömegspektrometria (GC-MS):...................................................................................... 30

Proteus morganii törzsek szerológiai keresztkapcsolatai
3
3.17. Nukleáris mag rezonancia (NMR): .............................................................................................................. 30 3.18. Növekedésgátlási vizsgálat: ......................................................................................................................... 31 3.19. Opszonizált baktériumok makrofág inváziója:............................................................................................. 31 3.20. Genetikai vizsgálatok:.................................................................................................................................. 32 3.21. Statisztikai analízis....................................................................................................................................... 32 4. EREDMÉNYEK ............................................................................................................................................... 33 4.1. Metodikai fejlesztés az LPS ELISA módszer érzékenységének növelésére .................................................. 33 4.2. Az újonnan beállított módszer klinikai használhatóságának kipróbálása....................................................... 37 4.3. A keresztreakcióban résztvevő antigének azonosítása és ezek tisztított formában való kinyerése ................ 39 4.4. A korábban leírt szerológiai keresztkapcsolatok megerősítése, pontosítása érzékenyebb módszerek
használatával ......................................................................................................................................................... 41 4.5. A hőstabil (LPS) antigén, cukor szerkezetének feltárása és összehasonlítása................................................ 44 4.6. Genetikai hasonlóság detektálása a három vizsgált törzs rfb gén clusterében ............................................... 50 4.7. A hőlabilis (fehérje) antigén identifikálása, ................................................................................................... 52 4.8. A keresztreagáló antigének kiváltják-e a keresztprotektívitást okozó ellenanyagok termelését?................... 57 4.9. A további kutatási lehetőségek feltárása ........................................................................................................ 59 5. MEGBESZÉLÉS .............................................................................................................................................. 60 6. AZ ÚJ EREDMÉNYEK ÖSSZEFOGLALÁSA............................................................................................... 67 7. IRODALOMJEGYZÉK.................................................................................................................................... 69

Proteus morganii törzsek szerológiai keresztkapcsolatai
4
RÖVIDÍTÉSJEGYZÉK
BSA bovine serum albumin
CE capillary electrophoresis
Col colitóz
DC desoxycholate
DIC diffuse inravascular coagulation
DMSO dimethyl-sulfoxid
DPS degradált poliszacharid
ELISA enzyme linked immunosorbent assay
FID flame ionization detector
Gal galaktóz
GC gas chromatography
Glu glukóz
HUS hemolytic uremic syndrome
IL interleukin
KDO 2-keto-3-deoxyoktonsav
LBP lipopolysaccharide binding protein
LIF laser induced fluorescence
LOS lipooligoszacharid
LPS lipopoliszacharid
GluA N-acetil-glukózamin
NMR nukleáris magrezonancia
OD optikai denzitás
OMP outer membrane protein
PAF thrombocyta aktiváló faktor
PAGE polyacrylamide gel electrophoresis
PBS phosphat buffered saline
PCP phenol-chlorophorm-petrolether
SDS sodium dodecyl sulfate
TLC thin layer chromatography
TLR toll-like receptor
TNF tumor necrosis factor

Proteus morganii törzsek szerológiai keresztkapcsolatai
5
1. BEVEZETÉS
A szervezetet veszélyeztető, behatoló kórokozókkal szemben fajlagos antitestek
termelődnek, amelyek képesek a pathogen kórokozóhoz kapcsolódni és azok eliminációjában
részt venni. A kórokozó felismerése nem mindig specifikus. Sok esetben a hasonló vagy
azonos antigént hordozó mikroorganizmusok is immunreakciót adnak ezen ellenanyagokkal.
Ilyen esetekben szerológiai keresztreakcióról beszélhetünk, mivel az egyik, antigénként
viselkedő mikroba ellen termelt antitest kapcsolódik egy másik mikroba felszínén levő
antigénnel is.
Mind az irodalomban, mind a klinikai vagy mikrobiológiai gyakorlatban gyakran
szembesülünk különböző pathogén és apathogén mikroorganizmusok közötti
keresztreakciókkal. Ezek egy része hasznos, mert az ismert keresztkapcsolatokra alapozva,
diagnosztikus céllal használhatjuk fel a hasonló antigént hordozó törzset. Itt elsősorban olyan
mikroorganizmusokról van szó, amelyek nehezen vagy egyáltalán nem tenyészthetők, illetve
tenyésztésük csak megfelelő biztonsági fokozattal rendelkező laboratóriumokban lehetséges
[Kondakova 2003]. Ilyen elven alapul pl. a Weil-Felix reakció, amikor a nehezen tenyészthető és
magas biztonsági fokozatot igénylő Rickettsiák (R. prowazekii) kimutatására a proteusok
bizonyos O antigénjeit (OX18, OX2, OXK) használják.
Más esetekben viszont a keresztreakció zavaró hatással lehet, mivel szerológiai
módszerrel nem tudjuk meghatározni a kórokozó kilétét, ahhoz más, esetleg bonyolultabb
módszerek szükségesek, növelve a diagnosztika költségeit. A kórokozó pontos azonosítása
fontos lehet az alkalmazott terápia vagy az epidemiológiai nyomonkövetés szempontjából.
A keresztreakcióban résztvevő antigének ismerete segítséget nyújthat mind az
identifikálásában, mind pedig az érzékenyebb diagnosztikai módszerek kidolgozásában. Ezen
kívül a tiszta antigének kinyerésével, az azok szintézisét meghatározó genetikai háttér és
immunológiai sajátságok ismeretével hatékony védőoltásokat dolgozhatunk ki, akár több
kórokozóval szemben is. „Oltóanyagként” adott esetben akár a nem kórokozó baktériumot is
használhatjuk, amennyiben ezek megfelelő minőségű, úgynevezett protektív antitesttermelést
kiváltó hasonló vagy megfelelő antigént tartalmaznak [Aucken 1991].
Korábban a Proteus morganii törzsekkel végzett szerológiai vizsgálatok bizonyították,
hogy egyes szerocsoportok nagyfokú keresztreakciót mutatnak a genuson belüli más
szerocsoportokkal vagy a Proteus genuson kívüli egyéb Gram-negatív baktériumokkal is.
Évtizedekkel ezelőtt a Pécsi Orvostudományi Egyetem Mikrobiológiai Intézetében Prof.

Proteus morganii törzsek szerológiai keresztkapcsolatai
6
Rauss Károly, Ralovich Béla és Vörös Sándor úttörő munkát folytattak egyes
keresztkapcsolatok jellegének tisztázásában [Ralovich 1967, Rauss 1967]. Ezek a munkálatok
azonban csak az akkor rendelkezésre álló alap immunológiai módszereken, főként
csőagglutinációs vizsgálatokon alapultak, amelyről tudjuk, hogy nem eléggé érzékeny
módszer. Ezen vizsgálatokkal mind lipopoliszacharid (LPS) (hőstabil), mind kapszid fehérje
természetű (hőlabilis) keresztreakció igazolódott a P. morganii O34 (8662/64)-E. coli O111
között [Ralovich 1967, Rauss 1967]. A P. morganii törzsek ritkán okoznak súlyos
gastrointestinális tüneteket, míg az E. coli, Salmonella, Shigella törzsek által okozott
gastroenteritis csecsemőkben vagy idősekben akár halálos kimenetelű is lehet. Más esetekben
súlyos mellékhatások jelentkezhetnek, mint pl. hemolitikus urémiás szindróma (HUS) [Beutin
2004].
Ezen ismereteket felhasználva érdemesnek tartottuk a keresztkapcsolatok
kialakításában résztvevő antigéneket, azok kémiai szerkezeti és immunológiai tulajdonságait a
jelenleg rendelkezésre álló módszerek felhasználásával tanulmányozni. Ezen vizsgálatoktól
elsősorban alapkutatási eredményeket és módszertani jellegű fejlesztéseket várunk, amelyeket
egyrészt további antigén keresztkapcsolatok tanulmányozásában alkalmazhatnak, másrészt
vakcinológiai fejlesztésekben lehetnek felhasználhatók.
1.1. A keresztkapcsolatok vizsgálataiban szereplő baktériumok
1.1.1. Proteus (Morganella) morganii
A Proteus (Morganella) morganii-t 1906-ban Morgan izolálta először csecsemők hasmenéses
székletéből. Egészséges felnőttek székletében 0,5-1%, kórházban ápolt csecsemők és
gyermekek székletében mintegy 10%-ban fordul elő. Enteropathogén szerepét nem erősítették
meg. Elhúzódó húgyúti fertőzésekben és sebgennyedésekből azonban gyakran lehet
identifikálni [Lányi 1980].
Alaki sajátságok: Gram szerint festve, Gram-negatív, peritrich csillós, tok nélküli, 1-3 μm
hosszú pálca.
Tenyésztés: Könnyen tenyészthető egyszerű agar táptalajon is. Közepes nagyságú, sima
felszínű telepeket alkot. Szélükből olykor kesztyűujjszerű nyúlványok indulnak. Az egész
lemezre terjedően, azaz rajzással csak lágy táptalajon, szobahőmérsékleten növekszik. Véres
agaron, a hemolizin termelő törzsek a telepek körül hemolitikus udvart képeznek.
Dezoxycholát (DC) tartalmú táptalajon üvegszerű vagy opálosan áttetsző telepek alatt a
táptalaj sárga lesz.

Proteus morganii törzsek szerológiai keresztkapcsolatai
7
Biokémiai és szerológiai tulajdonságok: Legállandóbb tulajdonsága az intenzív ureáz
képzés, indol termelés, valamint az a tény, hogy a mannitot, laktózt, szacharózt nem bontja és
képtelen H2S képzésre. A glukózt kevés gázképződés közben hasítja. A gelatint nem
folyósítja [Rauss 1959]. Szerológiai sémájuk kialakításában döntő szerepet játszott intézetünk,
Rauss és Vörös révén [Rauss 1959]. Trehalóz fermentálás alapján két alfajt különítenek el: a
nem fermentáló Proteus morganii subsp. morganiit és a fermenentáló Proteus morganii
subsp. sibioniit [Janda 1996].
1.1.2 Escherichia coli
Az Enterobacteriaceae család leggyakrabban izolált képviselője, és ugyanakkor a normál
bélflóra tagja. Környezeti jelenléte ételben, vízben faecalis szennyezettségre utal, így
közegészségügyi indikátor. Az enteralis pathogen csoportba tartozó törzsek súlyos
hasmenéses, mások húgyúti és egyéb extraintestinális megbetegedéseket okozhatnak.
Alaki sajátságok: Gram-negatív, csillós, rövid pálca. Az extraintestinális kórokozók
többsége poliszacharid tokkal is rendelkezik.
Tenyésztés: Tápanyagok tekintetében igénytelen, könnyen tenyészthető egyszerű táptalajon
is. Közepes nagyságú, sima felszínű, színtelen telepeket alkot. Diagnosztikai célra laktózt és
indikátort tartalmazó táptalajon tenyésztik.
Biokémiai és szerológiai tulajdonságok: Laktóz bontás, indol termelés és metilvörös próba
pozitív. A szénhidrátok széles skáláját sav és gázképződés közben fermentálja. Antigén
szerkezetileg heterogén: eddig mintegy 170 O antigént, 100-nál több K antigént, 55 H
antigént és 15-20 F antigént különítettek el [Holmes 1994, Brooks 1998].
1.1.3. Salmonella
A genus tagjai igen elterjedtek a természetben, mint a legkülönbözőbb állatok
bélcsatornájában élősködő pathogének. Köztük vannak emberhez adaptált szerovariánsok (S.
Typhi, S. Paratyphi A, B, C), egy meghatározott állatfajhoz nagy fokban adaptálódott
szerovariánsok (pl. S. Choleraesuis), valamint széles gazdaspecificitással rendelkezők (S.
Enteritidis, S. Typhimurium). Az utóbbi két csoport tagjai emberben elsősorban
gastroenteritist okoznak.
Alaki sajátságok: Peritrich csillós, tok nélküli (kivételt képez a S. Typhi, S. Paratyphi és S.
Dublin Vi-antigénje), Gram-negatív, 2-3x0,4-0,6 μm hosszú pálca. Egyes törzsek fimbriával
is rendelkeznek.

Proteus morganii törzsek szerológiai keresztkapcsolatai
8
Tenyésztés: Könnyen tenyészthető egyszerű táptalajon is, de a vizsgálati anyagból szelektív
(szelenit, brillantzöld, bizmut szulfit tartalmú) táptalajról izolálják őket.
Biokémiai és szerológiai tulajdonságok: Számos szénhidrátot sav és rendszerint
gázképződéssel fermentálnak. A laktózt azonban csak a III. subgenus tagjai bontják. Intenzív
H2S termelők, ureáz negatívak. Antigénszerkezetük O és H antigének által meghatározott.
Egyes törzsek rendelkeznek K antigénnel is, amit Vi antigénnek nevezünk. Eddig több mint
2200 szerotípus ismert, melyet a Kauffmann-White szerocsoportokba (A, B, C, stb) soroltak.
A törzsek döntő többsége kétfázisú H antigén képzésére képes [Holmes 1994, Brooks 1998].
1.1.4. Shigella
A genusba tartozó subgenusok, a S. dysenteriae, S. flexneri, S. boydii, S. sonnei a bakteriális
vérhast okozzák. A külvilág hatásaira sokkal érzékenyebbek, mint a korábban említettek. A
shigellák szigorúan emberhez adaptálódott pathogének.
Alaki sajátságok: Gram-negatív, csillótlan pálcák. Fiatal tenyészetekben coccobacilláris
formák jelenhetnek meg.
Tenyésztés: Igényesebb, fakultatív anaerob baktérium azonban jól kinő egyszerű agaron is.
Telepei apró, áttetsző, sima felszínűek. Diagnosztikai célra szelektív (DC, XLD) táptalajokat
használnak.
Biokémiai tulajdonságok: Biokémiailag relatíve inaktívak: kevés cukrot fermentálnak, azt is
gázképződés nélkül. Laktóz negatívak (a S. sonnei késői laktóz pozitív), ureázt, H2S-t nem
termelnek. Antigénszerkezeti alapon a S. dysenteriae subgenusba 11, a S. flexneri subgenusba
12-14, a S. boydii subgenusba 17, míg a S. sonnei subgenusba 1 szerotípus tartozik. A S.
sonnei esetében fázisról beszélünk: I. fázis a vad törzs, II. fázis, a virulencia plazmidot
vesztett mutáns ”R” tözs [Cooper 1957, Rauss 1961, Pál 1999]. Indukált mutációval további ”R”
mutánsok származtathatók (lásd 1.3.2 fejezetet).
1.2 A Gram-negatív bakteriális sejtfal szerkezete
A Gram-negatív baktériumokat a külvilág felől a sejtburok határolja (1. ábra). Ez a
burok nemcsak elválasztja a sejtet környezetétől, hanem össze is köti azzal: egyrészt rajta
keresztül mindkét irányban transzport folyamatok zajlanak, másrészt, mint külső felületnek
meghatározó szerepe van a baktériumsejtnek a gazdasejtek felszínéhez való kapcsolódásában,
megbetegítő képességében, az immunológiai folyamatok elindításában. Kémiai összetétele és
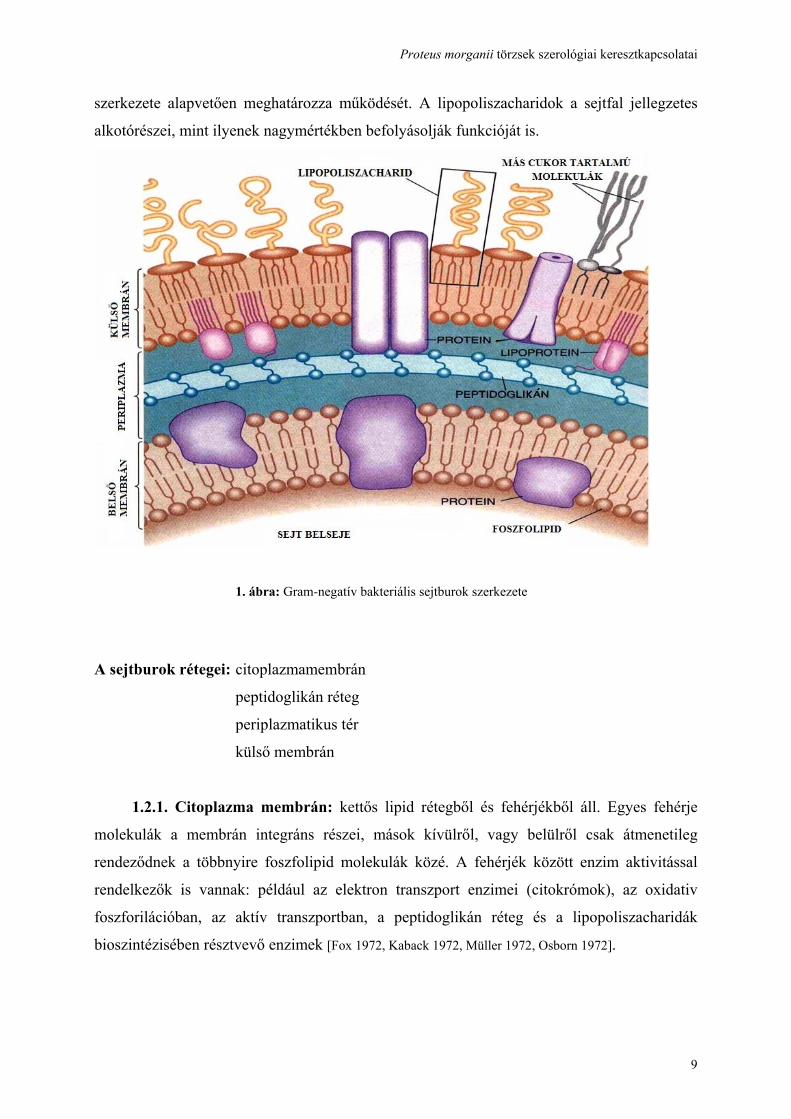
Proteus morganii törzsek szerológiai keresztkapcsolatai
9
szerkezete alapvetően meghatározza működését. A lipopoliszacharidok a sejtfal jellegzetes
alkotórészei, mint ilyenek nagymértékben befolyásolják funkcióját is.
A sejtburok rétegei: citoplazmamembrán
peptidoglikán réteg
periplazmatikus tér
külső membrán
1.2.1. Citoplazma membrán: kettős lipid rétegből és fehérjékből áll. Egyes fehérje
molekulák a membrán integráns részei, mások kívülről, vagy belülről csak átmenetileg
rendeződnek a többnyire foszfolipid molekulák közé. A fehérjék között enzim aktivitással
rendelkezők is vannak: például az elektron transzport enzimei (citokrómok), az oxidativ
foszforilációban, az aktív transzportban, a peptidoglikán réteg és a lipopoliszacharidák
bioszintézisében résztvevő enzimek [Fox 1972, Kaback 1972, Müller 1972, Osborn 1972].
1. ábra: Gram-negatív bakteriális sejtburok szerkezete

Proteus morganii törzsek szerológiai keresztkapcsolatai
10
1.2.2. Peptidoglikán réteg: N-acetil-glukózaminból és N-acetil-muraminsavból álló
poliszacharid láncokból és azokat összekötő peptidekből épül fel. Vastagsága a Gram-negatív
baktériumokban lényegesen szerényebb, mint a Gram-pozitívakban és mivel kevesebb a
kereszt kötés, relatíve nyitottabb is. Ez adja a baktérium stabil, rugalmas vázát. Fontos szerepe
van a sejt növekedésében és osztódásában. Több antibiotikum (penicillinek, cefalosporinok)
támadáspontja, egyesek azonban hatásukat igazán csak a Gram-pozitívakban fejtik ki, mert
külső membrán hiányában könnyen érik el ezt a réteget. (vancomycin) [Lugtenberg 1977].
1.2.3. Periplazmatikus tér: a citoplazma membrán és a külső membrán között
helyezkedik el. Részben szabad, részben kötött speciális lipoproteineket és ionokat tartalmaz.
Hidrolitikus, antibakteriális vegyületeket, detoxifikáló enzimaktivitású fehérjék mellett,
aminosav és aminocukor kötő proteineket is kimutattak. Feltételezések szerint ez a térség a
lipopoliszacharid szintézisében is szerepet játszik [Mühlradt 1977, Stock 1977].
1.2.4. Külső membrán: elektronmikroszkópos vizsgálattal identikusnak bizonyult más
biológiai membránokkal. Ez a Gram-negatív sejtburok legjellegzetesebb alkotó eleme [Nikaido
1979]. A lipid kettősréteg kialakításában a lipopoliszacharidok, foszfolipidek, valamint
fehérjék (major, minor protein, porin) vesznek részt. A kétértékű ionok a membrán
stabilitásáért felelősek (Ca2+, Mg2+). A foszfolipidek a belső, a lipopoliszacharidák a külső
oldalon elhelyezkedve aszimmetrikussá teszik a membránt [Funahara 1980]. A lipoproteineknek
fontos szerepük van a külső membrán peptidoglikán réteghez való kötésében. A peptidoglikán
réteg sérülése esetén megváltozik a külső membrán penetrabilitása, ezért a peptidoglikán réteg
a külső membrán bázisának tekinthető [Costerton 1974].
Ez a felépítés sajátságos tulajdonságot kölcsönöz a külső membránnak: barrier
funkciója révén hidrofób és nagy molekulákat nem enged át, ugyanakkor a tápanyagok
könnyedén átdiffundálnak rajta. A külső membrán nagymértékben befolyásolja az
antibiotikumok bejutását a baktériumba, így egyes antibiotikumok (lincomycin, clindamycin)
számára átjárhatatlan. Hidrofil külső felszíne ellenállóvá teszi a baktériumot az
emésztőnedvekkel, fagocitózissal, komplement rendszerrel szemben. Ezek a tulajdonságok
meghatározó szerepet játszanak a baktérium megbetegítő képességének kialakításában [Doyle
1989]. A membránt felépítő molekulák általában jó antigének és immunmodulánsként is
hasznosíthatók [Appelmelk 1987].
A lipopoliszacharid jelenléte alapvetően meghatározza a membrán tulajdonságait. Az
LPS-t érintő mutációk, illetve vegyszeres kezelése (pl. EDTA) a külső membrán szerkezetét

Proteus morganii törzsek szerológiai keresztkapcsolatai
11
megbontják és károsítják annak működését. Még nem ismert olyan Gram-negatív baktérium,
amely lipopoliszacharid hiányában képes lenne szaporodásra.
A Gram-negatív baktériumok sejtburkát egyes esetekben poliszacharidákból vagy
proteinből álló tok, vagy szerkezet nélküli nyák veszi körül. A sejtburkon csillók vagy
fimbriák hatolhatnak keresztül, módosítva a sejtfelület tulajdonságait.
1.3. A lipopoliszacharid
1.3.1. Történeti áttekintés: A lipopoliszacharidok kutatását a XIX. század hetvenes
éveinek végén kezdték el röviddel azután, hogy Koch megállapította: minden fertőző betegség
egy-egy meghatározott mikrobától ered. Kimutatták, hogy a baktériumok gyakran toxinokat
termelnek. Az elsőként elkülönített bakteriális méreganyagok az exotoxinok voltak.
Akkoriban a hőérzékenység volt a legfőbb ismertetőjelük: mivel ezek fehérjék, hő hatására
elveszítik megbetegítő képességüket. 1892-ben Pfeiffer felfedezte, hogy a kolera kórokozója,
a Vibrio cholerae rendelkezik egy hőérzékeny és egy hőstabil toxinnal is. Ez utóbbit nem
választja ki környezetébe és csak akkor szabadul fel, ha a baktérium szétesik. Ezért
endotoxinnak nevezte el. 1894-ben Centanni, Pfeiffertől függetlenül a Salmonella Typhiben
talált egy hőstabil toxint. Mivel a nyulakban lázkeltő volt, a pirotoxin nevet adta [Westphal
1983, Westphal 1985, Rietschel 1992].
Boivin, Morgan és Goebel (1933) arra a következtetésre jutottak, hogy az összes
Gram-negatív baktérium rendelkezik hőstabil toxinnal, ami poliszacharidból, lipidekből és
fehérjékből áll.
Shear (1943) szintén megállapította, hogy az általa izolált daganatpusztító és toxikus
anyag poliszacharidokból és lipidből áll. Ő a lipopoliszacharid (LPS) elnevezést használta.
Westphal, Lüderitz és Bister (1952) kidolgozott egy eljárást, amellyel a baktériumból nagy
mennyiségű és igen tiszta kivonatot tudtak nyerni. Ezek után egyértelművé vált, hogy a
mérgező alkotórész mindig csak poliszacharidból, lipidből valamint foszforsavból áll és a
korábbi vizsgálatoknál talált fehérje alkotórész nem szükséges a toxikus hatás létrejöttéhez
[Westphal 1952, Westphal 1985]. Westphal és Lüderitz annak kimutatásában is szerepet játszott,
hogy endotoxint a Gram-negatív baktériumok mindig, míg más élőlények egyáltalán nem
termelnek. A pirotoxin megjelölés ma már nem használatos, az endotoxin és lipopoliszacharid
között a szaknyelv már nem tesz különbséget, bár az endotoxin inkább biológiai, a
lipopoliszacharid inkább biokémiai megközelítést sugall.

Proteus morganii törzsek szerológiai keresztkapcsolatai
12
1.3.2. A lipopoliszacharid szerkezete
Az LPS molekulát bakteriológiai, szerológiai, kémiai, genetikai és biológiai
tulajdonságai alapján több szerkezeti egységre oszthatjuk: [Lüderitz 1983, Seydel 2000].
O-specifikus oldallánc
oligoszacharid mag (core)
Lipid-A
Az egyes részek kovalens kötéssel kapcsolódnak egymáshoz. Ez az alapséma elsősorban a
megbetegítő képességgel is rendelkező Gram-negatív, pálca alakú bélbaktérium törzsekre
jellemző. Ettől eltérő szerkezetű endotoxinokat is találunk az élővilágban, pl. Neisseria
meningitidis, Haemophilus influenzae, Chlamydia stb. törzseinél úgynevezett
lipooligoszacharid (LOS) fordul elő.
1. O-specifikus oldallánc: Ez a molekula a bakteriális sejtburok külvilág felé eső
része, amely egy ismétlődő egységekből összetevődő és hidrofil tulajdonságokkal rendelkező
polimer. Az alegységek 2-8 különböző, néha azonos cukormolekulából glikozid kötéssel
jönnek létre. A polimerizáltság foka igen változó (20-40), még egy adott törzs esetén is
heterogenitás észlelhető [Jann 1984, Nowotny 1990]. Az ismétlődő egységek monoszacharidjai
között sokféle és sokszor ritka cukrok is, pl. dideoxihexózok, aminocukrok, aminouronsavak
stb. találhatók. Ezért az O-specifikus oldallánc az endotoxin legváltozékonyabb része, de ez
felel a fajlagos immunitásért is, ezáltal meghatározva a baktérium szerológiai viselkedését.
Tekintettel a környezetünkben előforduló szerocsoportok nagy számára, melyet viszonylag
kevés monoszacharidból felépülő oligoszacharid határoz meg, elkerülhetetlen, hogy
egymástól rendszertanilag eltérő baktériumoknál, azonos vagy hasonló felépítésű és
szerológiai viselkedésű O oldallánccal rendelkező LPS-ek szintetizálódjanak. Ezek a hasonló,
vagy azonos endotoxin részleteket hordozó baktériumok a szerológiai vizsgálatok során
egymással keresztreakciót adnak.
Az O-specifikus oldallánc szintézisét meghatározó gének csoportját rfb- nek nevezzük.
Ha pl. deleciós mutáció következik be, akkor előfordulhat, hogy az O-oldallánc nem
szintetizálódik. Az ilyen lipopoliszacharidot hordozó baktériumokat R mutánsnak nevezték el,
mivel szemben a vad törzs sima (S-smooth) telepeivel, ezek rücskös (R-rough) telepeket
képeznek és elvesztik eredeti megbetegítő képességüket. [Kauffmann 1966, Lányi 1966/67, Lányi
1970, Seydel 2000, Raetz 2002].

Proteus morganii törzsek szerológiai keresztkapcsolatai
13
2. Oligoszacharid mag (core): A core régió belső magra (lipidhez kapcsolódó részre)
és külső magra (az O-specifikus lánchoz kötődő részre) osztható. Ez a régió már nem mutat
olyan nagy változékonyságot, mint az O-oldallánc. A szervezetre sem gyakorol olyan
erőteljes hatást. Bár, ha az O-lánc hiányzik az endotoxinból ez is beindíthatja az ellenanyag
termelést.
Több oligoszacharid mag típus ismert: Salmonella spp.-nél egyféle (Ra) mag található
minden szerotípusnál, míg Shigella, E. coli esetében ötfélét (R1-R4, K12) írtak le. Proteus és
Serratia esetén a mag szerkezete még nem teljesen ismert [Lebrecht 1990, Nowotny 1990]. A
strukturális különbségek a külső magban lelhetők fel.
Külső mag: különböző hexózokból áll: D-glukóz (Glu), D-galaktóz (Gal), N-acetil D-
glukózamin (GluA). Mindegyik piranóz formában és általában anomer kombinációban
van. Receptorként szolgál specifikus bakteriofágoknak, ugyanakkor R mutánsoknál a
mag ezen része felel a szerológiai specificitásért is. Ezen túlmenően valószínű, hogy
az aktivált T sejt lectinszerű receptorához a galaktóz reziduum segítségével köti az
LPS-t vagy baktériumot. Ezért feltételezik, hogy szerepe van a T helper sejtek
differenciálódásában és a baktériumok elölését kiváltó folyamatokban [Nowotny 1990].
Belső mag: az oligoszacharid mag izgalmasabb része, minthogy ritkán előforduló
cukormolekulákat, mint heptózt, gyakran L-glicero-D-manno-heptózt, ritkábban D-
glicero-D-manno-heptózt, valamint 2-keto-3-deoxioktonsavat (KDO) tartalmaz. A
KDO savlabilis ketozidos kötéssel α-2-6’ kapcsolódik a Lipid-A-hoz. Nagyon
jellegzetes és nélkülözhetetlennek tűnő alkotórésze a lipopoliszacharidának [Osborn
1963, Under 1981]. Nyolc szénatomot tartalmaz és néhány növényt kivéve, csak a
baktériumokban fordul elő. Minden endotoxin tartalmaz legalább egy KDO vagy
KDO-szerű molekulát. Szintézisének gátlása a Gram-negatív baktérium pusztulásához
vezet, ezért kitűnő célpont lehet antimikróbás szerek számára, azonban figyelembe
kell venni, hogy a kórokozók és a normál flóra között nem tesz különbséget. A KDO-
régió nagy affinitást mutat a bivalens kationok iránt. A magas Ca2+, Mg2+ koncetráció
a bakteriális felszínen esszenciálisnak tűnik a külső membrán strukturális és
funkcionális integritásában [Nowotny 1990]. A KDO-nak továbbá szerepe lehet a
hőlabilis exotoxinok megkötésében is, ami magyarázza a Vibrio cholerae által termelt
enterotoxin erősebb aktivitását is az ETEC hasonló molekulájával szemben, ugyanis
míg a V. cholerae csak egy KDO-val rendelkezik az E. coli esetében 2-3 KDO is
fellelhető [Horstman 2004]. 2-3 KDO-ból álló oligoszacharidához kapcsolódik a heptóz-

Proteus morganii törzsek szerológiai keresztkapcsolatai
14
régió. Úgy tűnik, hogy az endotoxin cukor régiói közül a belső mag szerkezete a
legkonzervatívabb [Seydel 2000, Raetz 2002].
3. Lipid-A: A lipopoliszacharid legbelső, egyben a legkevésbé variábilis része. Azt
már Westphal és Lüderitz is kimutatta, hogy a Lipid-A főként cukorból (glukózamin),
foszfátcsoportból és hosszú szénláncú zsírsavakból áll. 1983-ban Zähringer, Seydel és
Rietschel feltárta az E. coli Lipid-A szerkezetét, ami azonosnak bizonyult a Takayama által
leírt Salmonella Typhimurium Lipid-A szerkezetével.
A molekula vázát két D-glukózamin képezi, szokatlan módon (β 1-6 kötésben)
kapcsolódva egymáshoz. Az egyik glukózamin első szénatomja egy oxigén atomon keresztül
a másik glukózamin hatodik szénatomjához kapcsolódik, miközben az oxigén a glukózaminok
síkja fölé emelkedik (béta konfiguráció). Általában a béta-konfigurációban kötődő
glukózamin párokban az oxigén az egyes és négyes, esetleg az egyes és hármas szénatomot
kapcsolja össze egymással. Az egyes és hatos szénatom közti kapcsolatot az endotoxin Lipid-
A-ján kívül sehol sem találtak. Néhány esetben, a 2-amino-glukóz, 3-amino-glukóz
helyettesíti. Erre a vázra, amid és észter kötéssel hosszú szénláncú zsírsavak kötődnek: β-
hidroxi-mirisztinsav (C16), laurinsav (C12), mirisztinsav (C14), palmitinsav (C16). A
zsírsavak biztosítják a lipopoliszacharid hidrofobicitását és lipofil jellegét, mely lehetővé teszi
a membrán lipid kettős rétegébe való beágyazódását.
Hosszú évek kutatásainak eredményeként a Lipid-A-t tekintik a lipopoliszacharid
endotoxikus tulajdonságáért felelős molekula részének [Rietschel 1987, Nowotny 1990, Rietschel
1993]. Ezt Watson és Kim (1963) kutatásai megerősítették, ugyanis a Lipid-A-t és KDO régiót
tartalmazó endotoxin ugyanolyan toxikusnak és lázkeltőnek bizonyult, mint a teljes
poliszacharidot is tartalmazó molekula. A Lipid-A kulcsszerepe csak akkor vált egyértelművé,
amikor Siba, Kuszomoto és munkatársai szintetizálták az E. coli teljes Lipid-A-ját. A
mesterséges változat éppen úgy viselkedett, mint a természetes eredetű Lipid-A, azonban a
teljes értékű biológiai hatás kifejeződéséhez szükség van a lipopoliszacharid molekula minden
összetevőjére, ugyanis a térszerkezet kialakulása elősegíti az endotoxin kölcsönhatását a
szervezet sejtjeivel [Rietschel 1984, Galanos 1986, Rietschel 1987, Brade 1988, Rietschel 1991, Seydel
2000, Raetz 2002].

Proteus morganii törzsek szerológiai keresztkapcsolatai
15
1.3.3. A lipopoliszacharid genetikája
A bioszintézis genetikai hátterének nagy része már ismert. Jobban ismertek a mag és az
O-specifikus oldallánc bioszintézisében szerepet játszó génclusterek, amelyek közül a
legfontosabbak és azok funkciói a következők:
rfa - a mag bioszintézisét
rfb - az O-oldallánc alegységek szintézisét
rfc - az O-lánc alegységek polimerizálását
rfa-L és rfb-T -a mag és az O-specifikus oldallánc összekapcsolódását
rfe - az O-lánc bioszintézisét irányítja.
A különböző pontmutációk részfunkció változásokat hozhatnak létre, amelyek hozzájárulnak
az endotoxin heterogenitásának kialakulásához [Nowotny 1980].
Alaposabban tanulmányozott az rfb géncluster, amely az LPS-k biovariabilitásáért is
felelős. A vizsgálatunk tárgyát képező baktériumok O-oldallánca egy egyedi, a természetben
ritkán előforduló cukor molekulát, egy 3,6-dideoxihexózt, a colitózt (3,6-dideoxi-galaktóz) is
tartalmazza. Ezt a cukormolekulát eddigi ismereteink alapján csak néhány baktérium
lipopoliszacharidja tartalmazza (E. coli O55, O111, O157, S. enterica O30, O35, O50, Y.
enterocolitica O9, Vibrio cholerae O1). A colitóz szintézisért felelős gének és az rfb cluster
felépítése a következő képet mutatja:
Ahol a galF - glukóz-1-foszfát uridililtranszferáz, a wbdH - glycosil transzferáz, a gmd -
GDP-D-mannóz dehidratáz, a manC - mannóz-1-foszfát guanililtranszferáz, a manB -
foszfomannomutáz, wzx - O egység flippáz, a wzy - O antigén polimeráz, a wbdL - glycosil
transzferáz, a wbdM - glycosil transzferáz, a gnd - 6-foszfogluconát dehidrogenáz enzimek
szintézisét kódolja [Wang 1998, Bastin 1995, Wang 2000].
1.3.4. Biológiai hatás – hatásmechanizmus
Az első leírások az endotoxin lázkeltő és daganatölő hatásáról számoltak be [Snell
1975]. Ma tudjuk, hogy az endotoxinok csak akkor fejtik ki hatásukat, ha felszabadulnak a
baktériumsejt felszínéről. Ez akkor történik meg, ha a baktérium elpusztul vagy szaporodni
kezd [Bratková 1988].
wzxgalF wbdH gmd gmm wbdI manC manB wbdL wbdKwbdJ wbdMwzy gnd

Proteus morganii törzsek szerológiai keresztkapcsolatai
16
Hatásai sokrétűek, amelyeket nem közvetlenül a fertőzött sejt elpusztításával fejt ki. A
lázat sem közvetlenül a hipotalamusz hőszabályozó sejtjeire hatva hozza létre, hanem
közvetítő anyag segítségével (IL-1, TNF, stb.)
Mergenhangen és munkatársai (1969) bizonyították, hogy az endotoxinok főként a
makrofágokon keresztül hatnak. A makrofágokkal való kapcsolat után, az különféle
molekulákat választ ki, amelyek vagy függetlenül egymástól vagy szinergista módon
beindítják, illetve felerősítik a specifikus és nem specifikus immunválaszt [Nikaido 1979]. Az
így elindított cytokin kaszkád komoly orvosi problémát is jelenthet.
A mediátorok alacsony szintje esetén azok jótékony hatása érvényesül, többek
között az enyhe láz, az immunrendszer általános serkentése segíti a szervezetet a fertőzések
leküzdésében, valamint a daganatos sejtek burjánzásának gátlásában. Egyes kutatók szerint
fontos szerepe van az immunrendszer kialakulásában is [Raetz 2002].
Nagy mennyiségű, a szervezetet elárasztó endotoxin hatására, a káros hatások
érvényesülnek. Így magas láz mellett leukopenia, hipotenzió, sokk jelentkezik a szervek
rossz keringésével és elégtelen oxigénellátásával. A helyzetet súlyosbítja a kialakuló acidózis,
a komplement rendszer és a thrombotikus rendszer aktiválódásával kialakuló DIC, amely
gyakran halálos kimenetelű [Senior 1987, Martich 1993]. A Gram-negatív szepszis erélyes
baktericid antibiotikumos kezelése is sokszor magas keringő endotoxin szintet eredményez,
mivel az, az elölt lizált baktériumokból felszabadul. Ezért ilyen esetekben megfontolandó az
antibiotikumok steroid profilaxis melletti alkalmazása. A kis dózisú steroid kedvezően
befolyásolja a sepsis kimenetelét [Nowotny 1980, Waage 1991, Annane 2002].
Az LPS hatására felszabaduló mediátorok három csoportba oszthatók: fehérjék,
szabad gyökök és zsírok.
A fehérjék közül a legfontosabb a TNFα, IL-1, IL-6, IL-8: lázat, nagy adagban
irreverzibilis sokkot, halált okozhatnak. Kedvező tulajdonságuk, hogy a fertőzés helyére
csalják a fehérvérsejteket és a TNFα a daganatsejtek pusztítását is elősegíti. Az IL-6 mint
fontos sepsis marker már a klinikai gyakorlatban is egyre elfogadottabbá válik [Xiao 2006].
Szabad gyökök közül az oxigén szabad gyök (O-2), hidrogénperoxid (H2O2),
nitrogénmonoxid (NO), szintén hozzájárul a betolakodó mikroorganizmusok pusztításához.
Lipid eredetű mediátorok közül pedig a PGE2, TXA, PAF lehet fontos.
Egyre több megfigyelés bizonyítja, hogy az endotoxin a vérkeringésben lipid kötő
fehérjéhez (LBP) majd ez a komplex a makrofágok CD-14 receptorához tud kötődni.
Morrison és munkatársai (1992) olyan molekulákat azonosítottak a makrofágok felszínén,
amelyek közvetlenül kapcsolódhatnak az endotoxin Lipid-A-jához. Lehetséges, hogy a CD-14

Proteus morganii törzsek szerológiai keresztkapcsolatai
17
molekula mellett ezekhez a receptorokhoz is kapcsolódik az endotoxin [Morrison 1992]. A
másik lehetőség, az endotoxinnak a TLR 4 receptorokhoz való kötődése, mely később
elindítja a cytokin és eicosanoid termelést [Raetz 2002, Raetz 2006]. A receptorhoz való
kapcsolódás után aktiválódik a foszfolipáz C és megemelkedik a vízoldékony foszfoinozitol
4,5 bisfoszfát (PIP2), inozitol trifoszfát (IP3), valamint a zsíroldékony diacilglicerol (DAG)
szintje. Az IP3 mobilizálja az intracelluláris Ca2+-t az endoplazmatikus retikulum raktáraiból,
míg a DAG aktiválja a proteinkináz C-t. Ennek az eredménye különböző fehérjék
foszforilációja. A foszforiláció mértékének megváltozása zavart eredményez a glikogenolízis
és a glükoneogenézis folyamatában létrehozva az endotoxikózist és sepsist [Nowotny 1990].
A Lipid-A kevésbé vagy egyáltalán nem toxikus töredékei úgy tűnik, hogy
meggátolják a Lipid-A által kiváltott mediátor termelést, feltehetően úgy, hogy elfoglalják a
szabad receptorokat, így az endotoxin nem tud kötődni. A Lipid-A részletei vagy módosított
változatai a szervezet általános védekező képességét is fokozhatják [Raetz 2002]. Ez fontos
lehet, ha a beteg természetes védelmi mechanizmusai fertőzés vagy daganatbetegség
következtében károsodtak.
Az endotoxin szerkezetét megbontó gyógyszerek segíthetnek a Gram-negatív
baktériumok elpusztításában. Mivel a KDO minden Gram-negatív baktérium elengedhetetlen
része, analógjai az összes Gram-negatív baktériumra gátló hatással vannak. Az ilyen analógok
a jövőben nagy hatású antibakteriális szerek lennének, mivel még idáig egyetlen Gram-
negatív baktériumról sem tudunk, amely endotoxin hiányában képes lenne megfelelően
szaporodni [Nowotny 1990, Seydel 2000, Raetz 2002, Horstman 2004, Raetz 2006].
1.4. Külső membrán fehérjék (OMP)
Amint a belső membrán, a külső membrán is tartalmaz különböző fehérjéket, melyek
meghatározzák a membrán tulajdonságait, különböző funkcióit. Ilyenek a nagy mennyiségben
előforduló major, a kis mennyiségben előforduló minor fehérjék, és porinok [Vaara 1984,
Nishimura 1996]. Ezek közül a legismertebbek a porinok, melyek pórusokat formálnak
különböző anyagok transportját téve ezzel lehetővé.
Egy részüknek nincs specifikus funkciójuk és képesek különböző kis mólsúlyú hidrofil
jellegű molekulák áteresztésére (OmpC, OmpD, PhoE, OmpF stb.). Az egyes anyagok között
a fiziko-kémiai tulajdonságok, mint a méret, vízoldékonyság, töltés alapján tesznek
különbséget.

Proteus morganii törzsek szerológiai keresztkapcsolatai
18
A másik csoport tagjai specifikusak és csak egy-egy fajta molekula áteresztésére
képesek. Például a LamB protein maltóz és maltodextrin, a Tsx protein a nukleozid, FhuA
protein a ferrichrom, a BtuB protein a B12 vitamin, míg FepA protein a Fe3+ transzportban
játszanak szerepet. Ezen túlmenően ezek a fehérjék receptorként is szolgálnak különböző
bakteriofágok (λ, T1, T5, T6) és colicinek (colicin B, D, E, K) számára. Mint felületi
antigének fontos szerepet játszanak a szervezet immunológiai válaszának kialakításában,
valamint egyes sejtekhez való kötődésében [Bassford 1977, Datta 1977, Pugsley 1978, Yu 1982]. Más
külső membrán fehérjék, mint az E. coli OmpA membránfehérjéje a konjugációban betöltött
szerepükről ismertek.
A következő csoportnak a peptidoglikán réteg, külső membrán közti kapcsolat
kialakításában van strukturális funkciója. Ilyen a murein lipoprotein. A külső membránban
helyet kapó enzimek, mint a foszfolipáz A1 és számos proteáz a baktériumok inváziós
tulajdonságait is meghatározzák [Vaara 1984].
A külső membrán fehérjék vizsgálatára több módszer is rendelkezésünkre áll, ezek
közül talán a legismertebbek a fágok segítségével történő tipizálás, a fehérjék szétválasztását
célzó elektroforézises (gél, kapilláris, microchip) és az immunológiai metodikák [Kustos 1998/a,
Grubor 2005, Schaller 2006].
1.5. A keresztkapcsolatok
Számos fertőző betegség etiológiai diagnózisa a genetikai módszerek széleskörű
elterjedése ellenére, továbbra is immunológiai módszerek felhasználásán, a specifikus
antitestek kimutatásán alapul. Ilyenek az intracelluláris kórokozók, nehezen vagy egyáltalán
nem tenyészthető baktériumok (Mycoplasma, Leptospira, Treponema) vagy vírusok által
okozott kórképek, valamint mindazon betegségek, amelyek kórokozóinak tenyésztése magas
biztonsági fokozatú laboratóriumi felszerelést igényel. Azonban a szerológiai eredményeket
óvatosan kell értékelnünk, mert keresztreakció mindig előfordulhat. A vizsgálatunk tárgyát
képező baktériumok könnyen tenyészhetők és a diagnosztikus célú antitest-meghatározás
jelentősége elenyésző. Szerológiai vizsgálatot rutinszerűen csak a tipizáláshoz használnak.
Ezen törzsek keresztreakcióinak vizsgálatára mégis szükség van, mivel ezáltal egy-egy
bakteriális antigén szerkezetét jobban megközelíthetjük [Maurin 1997].
Az immunkémiai tanulmányok a keresztreakciók molekuláris szinten történő
megismerésében és a kémiai alapon történő klasszifikáció kidolgozásában nyújtanak
segítséget. A keresztkapcsolatok létrejöttében legtöbbször többféle antigén is részt vesz

Proteus morganii törzsek szerológiai keresztkapcsolatai
19
(fehérje, LPS) és ezek pontos meghatározása, a bakteriális fertőzőképességben játszott
szerepük feltárása nem könnyű feladat [Literacka 1999].
Különböző tanulmányokra alapozva elmondhatjuk, hogy a nagyfokú immunológiai
keresztreakció korrelál a genetikai homológiával. Tehát a szerológiai vizsgálatok egyes
baktériumok genetikájáról is felvilágosítást nyújthatnak. Ez pedig fontos támpontot jelent a
molekuláris genetikai vizsgálatok elkezdésében [Shu 1999].
A keresztreakciók, mint minden humorális immunológiai reakció antigén-antitest
reakción alapulnak. Az antitestek nagyfokú specificitással rendelkeznek, csak bizonyos
epitópok felismerésére képesek.
A keresztreakciók vizsgálati eredményeit az immunglobulinok típusai
befolyásolhatják. Az IgM, pentamer molekula, nagyobb partikulák összekötésével
agglutinációs (látható) reakciót hozhat létre, de kevésbé specifikus egy-egy meghatározott
epitópra. Az IgG már specifikusabb, érettebb, de agglutináció kialakítására kisebb mértékben
képes. Pontosabb, érzékenyebb szerológiai vizsgálatok kivitelezésére használhatók. Az IgA
szerepe a nyálkahártya védelemben kulcsfontosságú, szerológiai vizsgálatokra speciális
esetben használják.
Természetes körülmények között egy-egy baktérium ellen poliklonális immunsavó
termelődik. Előnye, hogy hatékony, sok epitóp ellen termelt antitest van jelen. Hátránya, hogy
nem tudjuk az egyes epitópokat azonosítani. Ezzel szemben a monoklonális ellenanyagot csak
laboratóriumi körülmények között lehet előállítani (kivétel a monoclonalis immunglobulint
termelő daganatos megbetegedések), egy meghatározott epitópot ismer fel és az, ha fedésben,
takarásban van nem használható a keresztreakció vizsgálatára (pl. ”S” baktérium esetén a
KDO régió elleni antitest).
A bakteriológiában, elsősorban a Gram-negatív baktériumoknál, gyakran találkozunk
LPS szintű keresztkapcsolatokkal. Gyakran a keresztreagáló antitestek azonos vagy hasonló
szerkezetű oligoszacharid fragmentumot ismernek fel (diszacharidtól tetraszacharid
hosszúságig). Pl. az LPS core régiójában előforduló L-glicero-D-mannoheptóz vagy D-
glicero-D-mannoheptóz bizonyos esetekben immunológiailag viselkedhet úgy, mint a mannóz
[Literacka 1999, Grubor 2006].
Világszerte erőfeszítések folynak a keresztreagáló nem pathogén baktériumok
védőoltásokra való felhasználásának kiaknázására [Aucken 1991, Ansaruzzaman 1999, Wang
1999]. A nagyszámú szerotípust felvonultató baktériumok esetében ez kudarcra ítélt
vállalkozás, mivel az antigének nagy száma miatt minden antigénnel szemben jó
immunválaszt kiváltó vakcina nehezen elképzelhető. A keresztreagáló antigének kiszűrésével

Proteus morganii törzsek szerológiai keresztkapcsolatai
20
lényegesen szűkíthetjük a védőoltáshoz szükséges antigének számát. Az antigéneket kódoló
gének ismeretével és a modern molekuláris genetika segítségével egy olyan pluripotens, nem
pathogén baktérium is előállítható, amely tartalmazhatja számos szerotípus elleni védelem
kiváltásában fontos antigéneket.
Az azonos antigénszerkezettel rendelkező baktériumok más-más betegségek
kiváltásában játszhatnak szerepet. Pl. esetünkben a Proteus morganii gastrointestiális
megbetegedést nem vagy nagyon ritkán okoz, de a Salmonella Adelaide O35, E. coli O111
súlyos hasmenést idézhetnek elő. Felvetődik a kérdés, hogy a fenti pathogen baktériumok
elleni immunitás kialakításához - első lépésben modellkísérletekben - a P. morganii-t
használjuk.
A klinikumban ellenkező esettel is találkozhatunk, mikor a klinikai tünetek, kémiai
laborparaméterek alapján nem tudunk differenciálni két egészen eltérő kórokozó között. A
leptospirosis és hantavírus fertőzés lehet erre egy jó példa. Az egyik baktérium, míg a másik
vírus. Gyakran együtt betegítik meg az egyént, mivel azonos epidemiológiai előfordulással,
reservoirral rendelkeznek, azonos tüneteket, és laboratóriumi eltéréseket okoznak. Kezelésük
azonban eltérő, ezért ilyen esetben a szerológiai vizsgálatok eredményére kell támaszkodnunk
[Nemes 2000, Péterfi 2000/b].
A keresztreagáló antigének kémiai szerkezetének, vagyis az antigént felépítő
komponenseknek a meghatározása segíthet a keresztreakciók mechanizmusainak
megítélésében is. Vizsgálatainkat is ezen megközelítésre alapoztuk és állítottuk fel
célkitűzéseinket.

Proteus morganii törzsek szerológiai keresztkapcsolatai
21
2. CÉLKITŰZÉSEK
Egyes Gram-negatív baktériumok közötti szerológiai keresztreakciót és az O antigének
kémiai hasonlóságát már többen leírták, azonban ezidáig csupán három esetben bizonyították,
hogy az O antigén két rendszertanilag távol álló baktériumtörzs esetében egyforma, éspedig
E. coli O111 és S. enterica O35; E. coli O55 és S. enterica O50, valamint E. coli O157 és S.
enterica O30 eseteiben [Jann 1984, Jann 1987, Wang 2000]. A P. morganii O34 (8662/64) LPS
kémiai szerkezetéről és annak az E. coli O111 LPS szerkezetéhez való viszonyáról még idáig
nem volt információnk. Irodalmi adatokból ismert, hogy a külső membrán fehérjék és az LPS
nemcsak mint strukturális együttes állnak szoros kapcsolatban egymással, hanem a hasonló
LPS-t hordozó baktériumok külső membránfehérjéje is hasonló lehet [DeMaria 1988].
A cukor és fehérje antigének közösen határozzák meg egy-egy baktérium szerológiai
tulajdonságait, különböző bakteriofágokkal szemben észlelt érzékenységét. Éppen ezért a
keresztkapcsolatok vizsgálatánál figyelembe kell vennünk ezen interakciót is, vagyis a
szerkezetek ismeretén túl az egymásra való hatásukat is.
Prof. Rauss Károly, Ralovich Béla és Vörös Sándor ugyan már korábban leírtak
keresztkapcsolatokat az általunk vizsgált baktériumok között [Ralovich 1967, Rauss 1967], de
ezek az eredmények csak csőagglutináción alapultak, a keresztreakció háttere nem került
felderítésre és elmaradt az antigének szerkezetének meghatározása is.
A fenti kérdések tisztázására irányuló munkánkat a következőképpen csoportosíthatjuk: 1. Metodikai fejlesztés az LPS ELISA érzékenységének és fajlagosságának növelésére,
hogy alkalmas legyen finom antigén különbségek detektálására is. 2. A továbbfejlesztett módszer klinikai használhatóságának kipróbálása. 3. A korábban leírt szerológiai keresztkapcsolatok megerősítése, pontosítása
érzékenyebb módszerek használatával és az álpozitív eredmények kiszűrésével. 4. A keresztreakcióban résztvevő antigének tisztított formában való kinyerése és
azonosítása. 5. A hőstabil (LPS) antigén cukor szerkezetének feltárása és összehasonlítása. 6. Genetikai hasonlóság detektálása a három vizsgált törzs rfb gén clusterében 7. A keresztreakcióban résztvevő hőlabilis (fehérje) antigén identifikálása, szerepének
meghatározása. 8. Megvizsgáljuk, hogy a keresztreagáló antigének képesek-e keresztvédelem
kiváltására és a keresztreakciók használhatók-e a vakcinológia területén. 9. A további tudományos, kísérleti munka során használható lehetőségek feltárása.

Proteus morganii törzsek szerológiai keresztkapcsolatai
22
3. ANYAG ÉS MÓDSZER
3.1. Felhasznált baktériumok:
Proteus morganii O34 (8662/64) O1, O9 és O43, Escherichia coli O21, O22, O33, O61, O68
[Bettelheim 1972] és O111, Salmonella Urbana O30, Salmonella Adelaide O35, Yersinia
enterocolitica O9, Shigella sonnei: S. sonnei fázis I és mutánsok S. sonnei fázis II [Rauss 1961],
S. sonnei Re 4350 [Kocsis 1984], Bacteroides fragilis.
3.2. Baktériumtenyésztés:
A megfelelő mennyiségű lipopoliszacharid kivonat elkészítéséhez nagy mennyiségű
baktériumra volt szükségünk. A baktériumtenyésztést Mueller-Hinton táptalajban végeztük a
Braun Melsungen (Germany) cég által gyártott Biostat U 30 típusú fermentorban (pH 7,2;
37°C).
Az előzőleg egy órán keresztül 121°C-on sterilizált táptalajt, két liter,
rázótermosztátban (New Brunswick Psycrotherm, USA) termelt előtenyészettel oltottuk be. A
tenyészetet 37°C-on 25 l/perc levegő befújása mellett 200/perc fordulattal kevertük. A pH-t a
fermentor folyamatos ellenőrzés és 10 n NaOH korrigálás segítségével állandó értéken
tartotta. A habzás fékezésére automatikus adagolással szilikon olajat alkalmaztunk. A
termelést tíz óra inkubáció után a logaritmikus fázis végén állítottuk le. A növekedést
lemezöntéses csíraszámlálással, illetve 600 nm hullámhosszon Perkin Elmer (USA)
fotométerrel ellenőriztük. A nagy térfogatú tenyészetet 4°C-ra hűtöttük, 100 ml koncentrált
formalin hozzáadásával elöltük és átfolyós centrifugán (Zuglói Gépgyár, FS-45,
Magyarország) perisztaltikus pumpa segítségével centrifugáltuk. Az így nyert, kb. 700 g
nedves baktériumot acetonnal szárítottuk. Az intracelluláris vizet is eltávolítva finom
szürkésfehér port kaptunk. Az LPS kivonása az S baktériumokból fenol-vizes módszerrel
[Westphal 1952], míg az R baktériumokból fenol-kloform-petroléteres eljárással [Galanos 1969]
történt. A Bacteroides fragilis LPS RIBI Immunochem Research Inc., (Hamilton, Montana,
USA) adománya volt. A Lipid-A-t és DPS-t az LPS 1%-os ecetsavas (100°C, 90 min.)
hidrolízise után nyertük.
A külső membrán kivonathoz a baktériumokat rázótermosztátban (New Brunswick
Psycrotherm, USA) 2000 ml Mueller-Hinton táptalajon tenyésztettük (24 h, 37°C, 100 rpm).
A baktériumok további felhasználásra a tenyészet centrifugálása (5000 g, 15 min., 4°C) és
kétszeri fiziológiás sóoldatos mosása után váltak alkalmassá.

Proteus morganii törzsek szerológiai keresztkapcsolatai
23
3.3. A lipopoliszacharid kivonása:
3.3.1. Westphal-féle fenol-vizes kivonó eljárás.
A beszárított baktériumot desztillált vízben szuszpendáltuk (1 g baktérium: 20 ml desztillált
víz) és homogenizáltuk. Az elegyet 68°C-ra melegítettük és azonos hőmérsékletű 90%-os
fenollal (1:1/v:v) kevertük össze. A keveréket 5 perc után 4°C-ra hűtöttük és 15 percig
Janetzki K-70-es centrifugán ülepítettük (fordulatszám 2000 rpm.). A felülúszó vizes fázis
tartalmazta a szükséges endotoxint, amit leszívtunk. Az eljárást még kétszer megismételtük.
Az összegyűjtött vizes fázisokat három napig folyamatosan csapvízben, majd újabb három
napig 12 óránként cserélt desztillált vízben dializáltuk. A dializálható anyagoktól megtisztított
kivonatot Büchi-féle rotációs vákuum bepárlón kb. 3 g/100 ml-re koncentráltuk. A
koncentrátumot ultracentrifugán (Janetzki VAC 602 Germany) 100.000 g-vel, 4 órán át
centrifugáltuk. Az üledéket desztillált vízben szuszpendáltuk és az eljárást még kétszer
megismételtük. Liofilezés után rendelkezésünkre állt a nagy tisztaságú, por alakú, fehérje és
nukleinsav mentes endotoxin preparátum [Westphal 1952].
3.3.2. R és abszolút R törzsek esetén a Galanos féle fenol-kloroform-petroléteres
(PCP) kivonó eljárást alkalmaztuk. Az acetonnal szárított baktériumot
fenol/kloroform/petroléter (2:5:8/v:v:v) keverékével kezeltük, majd centrifugáltuk. A
felülúszót vákuumban koncentráltuk. Háromszori kivonás után a koncentrált fenolos fázisból
cseppenként adagolt vízzel csaptuk ki az LPS-t. A fenol nyomokat éteres mosással
távolítottuk el. Az üledéket desztillált vízben vettük fel, 105.000 g-vel 4,5 órát
ultracentrifugáltuk. Az LPS-t tartalmazó üledéket liofileztük [Galanos 1969].
3.4. Külső membrán fehérjék (OMP) kivonása:
A baktériumokat 37°C-on folyékony táptalajban fermentorban tenyésztettük, amely a
tenyészetet 24 óráig logaritmikus fázisban tartotta. Ezután a baktériumokat 4°C-on Janetzki K
70-es centrifugán 15 percig, 2000 rpm-el (5000 g; 4°C) centrifugáltuk. Az üledéket fiziológiás
sóoldatban szuszpendáltuk és a táptalaj alkotórészek kimosása céljából a centrifugálást
megismételtük. Ezt követően lizozimes kezelést alkalmaztunk. Az üledéket 4°C-on, állandó
kevergetés mellett 25 ml, 0,75 M szacharóz és 7,5 pH-jú, 10 mM TRIS HCl oldatban
szuszpendáltuk. Mikor a szuszpenzió homogénné vált, változatlan hőmérsékleten 2 percig,
250 μl (10 μg/ml) lizozimmel inkubáltuk, majd kb. 10 perc alatt lassan, 50 ml, 1,5 mM
EDTA-t (pH 7,5) adtunk hozzá. Ezt követte a sejtek ultrahangos feltárása jeges fürdőben, 4x2
percig 500 W-on. Az intakt sejteket 4°C-on történő 15 perces centrifugálással távolítottuk el.

Proteus morganii törzsek szerológiai keresztkapcsolatai
24
A külső membrán fehérjék kinyerésére 1 órás ultracentrifugálást alkalmaztunk 100.000 g-n
(Dupont-Sorval, USA). Az üledéket, mely a külsőmembrán fehérjéket is tartalmazta grádiens
ultracentrifugálással (24.000 rpm, 4°C, 21 óra) (Beckman Spinco Division, Palo Alto, CA,
USA) választottuk el.
A frakciókat külön-külön automata pipettával leszívtuk és deionizált vízben oldottuk,
majd újabb 1 órás ultracentrifugálással (Dupont-Sorval, USA) a cukor grádiens maradványát
is eltávolítottuk. Az üledéket deionizált vízben oldottuk és további használatig –20°C-on
fagyasztva tároltuk [Osborn 1972].
3.5. Oszlopkromatográfia:
Az LPS Lipid-A és poliszacharid részre való bontásához enyhe savas hidrolysist alkalmaztunk
1%-os ecetsav segítségével (100°C, 90 min). Az így nyert csapadékot centrifugálás
segítségével távolítottuk el. A felülúszót, amely a DPS-t tartalmazta további tisztításnak
vetettük alá 2,5x80 cm hosszú Sephadex G-50 (Sigma Chemicals, St. Louis, MO, USA)
oszlopon. A szétválasztáshoz pyridin-ecetsav puffert (4 ml pyridin és 10 ml ecetsav 1000 ml
vízben) használtunk. Az 5 ml frakciók cukortartalmát koncentrált kénsav és fenol segítségével
(100 µl minta, 100 µl fenol, 800 µl cc. H2SO4) határoztuk meg. A photometrálás segítségével
elkészített optikai denzitás görbék mutatták a tiszta frakciókat tartalmazó csövek pozícióját.
Az így nyert oldatot Büchi-féle rotációs vákuum bepárló segítségével koncentráltuk, többszöri
desztillált vizes oldás és bepárlás segítségével a nyomokban jelenlévő ecetsavat és pyridint is
eltávolítottuk. [Osborn 1972, Vinogradov 2002].
3.6. Poliklonális immunsavók előállítása nyúlban:
Az immunizáláshoz használt baktériumot 37°C állandó hőmérsékleten, egy éjszakán keresztül
tenyésztettük. Mosás után fiziológiás sóoldatban szuszpendáltuk, hogy a koncentrációja 108
cfu/ml legyen. A baktérium szuszpenziót 100°C fokon, egy órát főztük.
Új-zélandi nyulakat (kb. 3 kg) intravénásan a következő séma szerint oltottuk: 0,2 ml
(0. nap), 0,4 ml (5. nap), 0,8 ml (10. nap), 1,2 ml (15. nap), 1,6 ml (20. nap), 1,8 ml (25. nap).
Az utolsó injekció előtt próbavért vettünk és annak titerét csőagglutinációval határoztuk meg.
Az 1:3200, 1:6400 hígítást tartottuk elfogadhatónak. Ha nem volt ilyen magas a titer, akkor
tovább immunizáltunk. Az utolsó injekció után kb. 14 nappal, a nyulakat elvéreztettük. A vért
steril üvegedénybe gyűjtöttük. A kapott vért egy órán át 30°C-on, majd további kb. 10 órán át
4°C-on tartottuk, hogy a véralvadás teljes legyen. Ezután Janetzki K-70-es centrifugán 4°C–

Proteus morganii törzsek szerológiai keresztkapcsolatai
25
on, 15 percig, 2000-es fordulattal centrifugáltuk. A savót leszívtuk és Seitz szűrőn sterilre
szűrtük, majd steril műanyag edényekben, –20°C-on tároltuk [Clausen 1991, Péterfi 2000].
A külső membrán proteinek elleni immunsavót szintén nyulakban termeltük. A
kipreparált külső membrán fehérjéket SDS-PAGE segítségével választottuk szét, ezt követően
azokat nitrocellulóz membránra blottoltuk. A nitrocellulóz membránra átvitt fehérjét 0,02%-
os Ponceau S (Merck, Darmstadt, Germany) segítségével tettük láthatóvá. Ezt követően a
vizsgálat szempontjából kérdéses sávokat kivágtuk, DMSO-ban oldottuk és komplett Freund
adjuváns egyenlő arányú keverékével immunizálást végeztünk. Az oltóanyagot egyidőben
alkalmaztuk mind subcutan, mind intraperitonealisan, mind intramuscularisan (mindkét oldali
comb), egy-egy régióba egyszerre 1 ml keveréket juttatva. Az emlékeztető oltás 2 hét múlva,
majd havonta követte az előző oltást, melyben DMSO-ban oldott fehérje tartalmú
nitrocellulóz csíkok és inkomplett Freund adjuváns egyenlő arányú keverékét alkalmaztuk
[Norrander 2000, Toller 2004]. Az immunizálás végén az immunsavót a korábban ismertetett
módon nyertük ki és tároltuk.
3.7. Poliakrilamid gélelektroforézis (PAGE):
A gélelektroforézis BioRad Mini-Protean II (BioRad Laboratories, Hercules, Ca, USA)
készülékkel Laemmli discontinuus rendszerben történt. A szeparáló gél 12%-os, a stacking
gél 5%-os volt. A gélre minden anyagból (bromophenol blue-val festett) 5-10 μg mennyiséget
vittünk fel. Anyagunkat 200 V feszültség alatt a 0,053 M Tris-HCl (pH=8,3), 0,052 M glicin
és 0,1% SDS elektród puffer segítségével futtattuk, míg a frontvonal a gél alját elérte.
3.7.1. A fehérjék Coomassie Brilliant Blue festése: A fehérje mintát tartalmazó
géleket Coomassie Brilliant Blue R250 (0,125% Coomassie Brillinat Blue R-250, 50%
methanol, 10% ecetsav) segítségével festettük meg. A quantitatív analízist denzitométer (Bio-
Capt, East Port, France) segítségével, míg a molekulasúlyt, molekulasúly standard (Low
molecular standard kit, Pharmacia Biotech, Uppsala, Sweden) használatával értékeltük: α
lactalbumin (14400 Da), trypsin inhibitor (20100 Da), carbonic anhydrase (30000 Da),
ovalbumin (43000 Da), BSA (67000 Da) [Laemmli 1970].
3.7.2. LPS minták előhívása ezüstözéssel: Az LPS-t tartalmazó géleket a módosított
ezüstöző eljárással festettük meg. A friss géleket egy éjszakán keresztül 200 ml 25% propan-
2-olt, 7% ecetsavat tartalmazó oldatban fixáltuk. Ezt követően perjódsavas oxidáció történt.
Gyakori csere mellett, a több mint 4 órás desztillált vizes mosás után 15 percig
ammóniumtartalmú ezüstnitrátos (1,4 ml cc. ammónia, 21 ml 0,36% NaOH, 19,4% AgNO3
100 ml desztillált vízben) festést alkalmaztunk. A közel 40 perces mosás után az előhívás a

Proteus morganii törzsek szerológiai keresztkapcsolatai
26
frissen elkészített 200 ml 0,005% citromsavas, 0,019% formaldehides oldatban történt, melyet
a reakció leállítása céljából többszöri desztillált vizes mosás követett [Hancock 1988, Nelson
1990].
3.8. Csőagglutináció:
Widal csövekben létrejövő antigén-antitest kapcsolódás során kialakult agglutinátum
vizsgálatára alkalmas. A fiziológiás konyhasóoldatban végzett 1:100 hígítású immunsavóból
10 Widal csőben felező hígítást készítettünk (1 ml minden csőben), majd 25 μl főzött (100°C,
60 perc) baktérium szuszpenziót (A600=1.2) (UV-VIS photometer Perkin-Elmer, Germany)
adtunk. Az eredményt agglutinoscop segítségével 37°C-on történő 24 órás inkubáció után
olvastuk le.
3.9. Indirekt hemagglutináció:
A Gram-negatív baktériumok lipopoliszacharid antigénjei ellen keletkezett ellenanyagok
kimutatására igen jól bevált, egyszerű és gyors módszer a passzív hemagglutináció. A
módszer hátránya, hogy többnyire IgM tipusú antitestet mutat ki és érzékenysége elmarad az
ELISA érzékenységétől.
A vizsgálat lényege az, hogy a szuszpendált endotoxin kivonatot a vörösvérsejtek
felületére adszorbeáljuk (szenzitizáljuk a vörösvérsejtet), és így az új antigén specificitást
nyert vörösvérsejtek a homológ ellenanyagot tartalmazó savóban jellegzetes, könnyen
leolvasható reakciót, hemagglutinációt mutatnak. A passzív hemagglutinációhoz a
legalkalmasabb a birka, de használható más állatfaj és ember vére (O; Rh negatív) is. A friss
defibrinált vérből a plazmát legalább háromszori mosással el kell távolítani. A friss, mosott
vörösvérsejt szuszpenziók Alsever konzerváló oldatban 3-4 hónapig használhatók. Az Alsever
oldat összetétele: glukóz 20,5 g; NaCl 4,2 g; desztillált víz ad 1000 ml; pH-ját 0,15 M-os
citromsav oldattal 6,1-re állítjuk, és szűréssel sterilizáljuk. A friss vér nem optimális a
hemagglutinációra, a konzervált vörösvérsejtek használatát is ajánlatos néhány nappal az
elkészítés után kezdeni [Lányi 1980].
Vörösvérsejt szenzitizálása: Mosott vagy konzervált vörösvérsejt szuszpenzióból
centrifugálással (2000 rpm, 5 perc) nyert üledék 0,5 ml-éhez adtuk az előzőleg 2 ml
fiziológiás konyhasóban szuszpendált 3 mg LPS-t és vízfürdőben egy órán át 37°C-on
inkubáltuk. Ezután a nem adszorbeálódott, fölöslegben maradt antigént háromszori mosással
(centrifugálás 2000-rpm-el, 5 percig) eltávolítottuk. Az üledékből kb. 2-5% -os szuszpenziót

Proteus morganii törzsek szerológiai keresztkapcsolatai
27
készítettünk. A vizsgálandó savóból Takátsy lemezen fiziológiás konyhasóval kettős léptékű
tovafutó hígítást végeztünk a következőképpen:
50 μl fiziológiás konyhasó minden furatba
50 μl immunsavó az első vályúlatba és összekeverés után
50 μl-t tovább vittünk kihagyva az utolsó furatot, amit savómentes kontrollnak
használtunk (felező hígítás). A következő sor első furatába összesen 100 μl fiziológiás
konyhasót tettünk, ugyancsak 50 μl savóval kevertük és az előzőhöz hasonlóan elkészítettük a
hígítási sort. (harmadoló, majd köztes felező hígítás). A hígítás elkészítése után minden
vályúlatba 50 μl kb. 2%-os szenzitizált vörösvérsejt szuszpenziót tettünk és
szobahőmérsékleten inkubáltuk. Az eredmény már 10 óra után leolvasható volt.
Negatív kontroll céljából megvizsgáltuk a szenzitizált vér reakcióját fiziológiás
konyhasóban is, mivel a spontán agglutináló szenzibilizátum nem használható.
3.10. Dot blot:
A Magyar és munkatársai által leírt eljárást alkalmaztuk [Magyar 1991], amelyet a
következőképpen módosítottunk. 108 cfu/ml baktérium tenyészet 10 μl szuszpenzióját
csepegtettük a nitrocellulóz membránra (Sigma Chemicals, St. Louis, MO, USA), majd a
cseppet beszárítottunk. A membránokat szobahőmérsékleten 20 percig blokkoló oldattal
blokkoltuk (1% kazein, 0,05% Tween 20, 0,025 M Tris és 0,192 M glicin desztillált vízben
oldva és a pH 8,3-ra állítva). Blokkolás után a membránokat 1:1000 hígítású poliklonális
nyúlsavóval, majd peroxidázzal konjugált anti-nyúl IgG-vel további 20-20 percig inkubáltuk.
A nem-kötő ellenanyagokat kétszeri NaCl-os blokkoló oldattal, majd PBS-el mostuk 5-5
percig. A reakciót 4-kloronaftol-peroxidáz szubsztrát hozzáadásával tettük láthatóvá. A
reakció erősségét denzitometriás (Bio-Capt, East Port, France) mérés után egy 256-s
szürkeskála alapján analizáltuk
3.11. Immunoblot:
A módszer lényege, hogy előzetes gél elektroforézises szétválasztás után az antigének (fehéje
vagy poliszacharid) egy nitrocellulóz membránra kerülnek, ahol immunológiai sajátosságaikat
felhasználva célzottan vizsgálhatók, tehetők láthatóvá.
A poliakrilamid gélelektroforézis után a géleket 30 percig blottoló pufferben áztattuk,
majd 350 mA-n, a BioRad Mini Trans-Blot Electrophoretic Transfer Cell (BioRad
Laboratories, Hercules, CA, USA) segítségével 4°C-on, 60 perc alatt, nitrocellulóz
membránra helyeztük (Sigma Chemicals, St. Louis, MO, USA) (pórusnagyság 0,22 μm). A

Proteus morganii törzsek szerológiai keresztkapcsolatai
28
transfert a Bio-Rad használati utasításának megfelelően végeztük. A megszárított
membránokat a korábban ismertetett eljárással kezeltük [Magyar 1991].
3.12. ELISA:
Először az Engvall és Perlmann által leírt (1972), később a Takahashi és munkatársai
által módosított eljárást (1992) használtuk ELISA módszerünk alapjául, amelyet a
következőképpen fejlesztettünk tovább:
Lemez előérzékenyítése: 100 μl, 0,01 M, 7,2 pH-jú foszfát pufferben (PBS) oldott 10 μg/ml
koncentrációjú poly-L-lysint (260.000 mólsúly) (Sigma Chemicals, St. Louis, MO, USA)
helyeztünk a polisztirén lemez vályulataiba (Nunc Immunoplate, Intermed, Dánia), amit
szobahőmérsékleten, egy éjszakán keresztül inkubáltunk.
LPS lemezhez kötése: 100 μl különböző koncentrációjú LPS oldatot adtunk a poly-L-lysinnel
előérzékenyített lemezhez, amit ezután 1 órán át 37°C-on inkubáltunk. A lemezt 4x mostuk
0,05% Tween 20-t tartalmazó PBS-el (T-PBS). A rendszer beállítása után a lemez
érzékenyítésére 1 μg/ml LPS-t használtunk.
Nem specifikus kötőhelyek blokkolása: BSA-t, kazeint, kecske, juh, sertés, bovin, tengeri
malac, ló savókat használtunk blokkoláshoz (Sigma Chemicals, St. Louis, MO, USA). A
blokkolást 200 μl különböző hígítású kazein és BSA oldatokkal, valamint különböző
állatfajok savóival 30 percig 37°C-on végeztük, amit a lemez 4x T-PBS mosása követett. A
módszer beállítása után a nem specifikus helyek blokkolására már csak 1:20 hígítású
kecskesavót alkalmaztunk.
Immunsavó: 100 μl korábban kipróbált optimális hígítású savót adtunk minden lyukba, a
lemezeket 1 órán keresztül 37°C-on inkubáltuk, majd 4x mostuk T-PBS-el.
Konjugátum: 100 μl, 1:500 hígítású peroxidázzal konjugált anti-nyúl IgG (kecske savó)
(Sigma Chemicals, St. Louis, MO, USA) adtunk minden lyukba. 1 óra 37 °C fokos inkubálás
után 4x mostuk T-PBS-el.
Szubsztrát: minden lyukba 100 μl 0,01% o-fenilén diamin peroxidáz szubsztrát (Sigma
Chemicals, St. Louis, MO, USA) és 0,03% H2O2 keverékét adtunk. A reakciót 10 perccel
később 4N kénsavval leállítottuk. Az extinkciót 492 nm-n mértük le Titertek Uniscan
fotométerrel (Flow Laboratories, Helsinki, Finland)
Minőség ellenőrzés és értékelés: A fotométert levegőre nulláztuk. A puffer optikai denzitása
(OD) 0,000-0,100 közötti értéket kellett, hogy adjon. Ha nagyobb volt, mint 0,100 akkor a
nem megfelelő mosás miatt kísérleteinket megismételtük. Ezután a fotométert a negatív

Proteus morganii törzsek szerológiai keresztkapcsolatai
29
kontrollra nulláztuk és olvastuk le az eredményeket. A pozitív és negatív (IgG mentes)
kontroll közti különbség nagyobb volt, mint 0,800 [Péterfi 2000].
Vizsgálatok humán savókkal: A humán eredetű szérumok vizsgálatához további
módosításokra, rendszerbeállításokra volt szükségünk. Immunsavó helyett humán szérumot
használtunk, majd detektáláshoz peroxidázzal konjugált anti-human immunglobulin keveréket
(IgG, IgM és IgA), valamint IgM (Sigma Chemicals, St. Louis, MO, USA) tartalmú savót
alkalmaztunk. Egyebekben a módszer megegyezett a korábban ismertetettekkel.
A reakciók kiértékelése: Pozitívnak akkor értékeltük a kapott eredményeket, ha azok
meghaladták a 0,3 optikai denzitást. Ez az érték a negatív kontrollok háromszoros standard
deviációjának felel meg. Titeremelkedés kritériuma a negatív kontroll legalább kétszeres
standard deviációjának megfelelő optikai denzitás növekedés volt [Péterfi 2006].
3.13. Kapilláris elektroforézis (CE):
Mintáinkat a CE-SDS kit futtató pufferét használva a Biofocus 3000 (BioRad Laboratories,
Hercules, CA, USA) kapilláris elektroforetizáló rendszeren futtattuk. A vizsgálathoz 24 cm,
50 μm átmérőjű szilikonos kapillárist alkalmaztunk. Mintapufferünk összetétele megegyezett
a szokványos poliakrilamid gélelektroforésisnél használt mintapufferel (a glicerol és
brómfenolkék kivételével). A minta felvitele hidrodinamikai módszerrel történt (40-50 psi x
s). A használt feszültség 15 kV, 19 μA, a hőmérséklet 20°C volt, míg a detektálás 220 nm
hullámhosszon történt. A molekulasúly meghatározására a Pharmacia molekulasúly
kalibrációs standard görbéit, belső standardnak benzoesavat használtunk. Az egyes csúcsok
alatti területeket és azok arányát Biofocus Integrator program segítségével határoztuk meg
[Kustos 1998a, 1998b].
3.14. Lab-on-a-Chip (microchip) technológia:
A chip alapú elválasztást a kereskedelmi forgalomban is kapható Agilent 2100 Bioanalyzer
system segítségével végeztük. A vizsgálatokhoz Protein 200 Plus LabChip Kits chipet
használtunk. Ezek a chipek 11 minta vizsgálatára alkalmasak és egyetlen elválasztó csatornát
használnak. A chipek előkészítése és kezelése a Protein 200 LabChip Kit használati
utasításnak megfelelően történt. Detektálásra laser indukált fluorescenciát (LIF) használtunk.
Az eljárás, mint szűrőrendszert használja a microcapillárisokban lévő futtató puffert. A
fehérje festéket már a szűrőmátrix tartalmazza, ezért az SDS-protein komplex még a chipben
történő futása alatt gyorsan megfestődik, kevesebb, mint 100 ms a szeparáló csatornába
történő lépéstől számítva. A fluorescens háttér csökkentése céljából SDS hígítás került a

Proteus morganii törzsek szerológiai keresztkapcsolatai
30
kapilláris oszlop után beiktatásra. Az adatok kezelése, tárolása és értékelése Protein 200 assay
software segítségével történt. A molekulasúly meghatározáshoz pedig a LabChip Kit Protein
200 molekulasúly standardját használtuk, mely így 6-210 kDa közötti molekulasúly
meghatározását tette lehetővé. A fehérje koncentrációjának meghatározása pontkalibrációval
történt, amikor a rendszer a myosin marker csúcsot a vizsgált fehérje csúcsával hasonlította
össze [www.agilent.com/chem/labonachip, Péterfi közlésre elfogadva].
3.15. Vékonyréteg kromatográfia (TLC):
A minták futtatása felszálló irányban Shandon-féle kromatográfiás edényben történt. A
monoszacharidokat az LPS-ből 100 °C-on és 14 órán át tartó egy mólos kénsavas hidrolízis
során nyertük. A semlegesítést BaOH-al végeztük. Az oldatot vákuumban bekoncentráltuk,
majd kis térfogat desztillált vízben reszuszpendáltuk. Mintáinkat Cellulose DC Plastikfolia (E.
Merk, Darmstadt, Germany) vékonyrétegre (futtató közeg) csepegtettük és beszárítás után a
következő eleggyel futtattuk (mozgó fázis): n-butanol-piridin-víz (6:4:3), valamint piridin-
etilacetát-víz-ecetsav (5:5:3:1). Standardként ribóz, mannóz, galaktóz, glukóz, heptóz és
glukózamin keverékét használtuk. Az előhívás Trevelyan és munkatársai által kifejlesztett
ezüstözési módszerrel történt [Trevelyan 1950].
3.16. Gázkromatográfia, tömegspektrometria (GC-MS):
A gázkromatogram készítéséhez a vizsgálandó anyagot először illékonnyá kell tenni. Első
lépésben a monoszacharidokat Na-bórhidriddel (13 mg Na-bórhidrid 1 ml desztillált vízben)
redukáltuk. A semlegesítés 200 μl koncentrált ecetsav hozzáadásával történt. A bekoncentrált
anyagot 10x2 ml desztillált methanollal szárazra pároltuk, majd vízmentes környezetben
piridin jelenlétében 14 órán keresztül 100°C-on ecetsavanhidrid segítségével acetiláltuk, majd
1 ml kloroformban oldottuk és Agilent GC-MSD gázkromatográffal (Agilent technologies,
USA) 30 m, 251 µm átmérőjő Agilent DB-225 HP-1MS (Agilent technologies, USA)
kapilláris oszlopon analizáltuk. A vivőgáz 15 ml/perc sebességgel 21 psi nyomáson
áramoltatott nagy tisztaságú hélium volt. Az injector hőmérséklete 220°C, az oszlopé 180°C,
a lángionizációs detektoré (FID) szintén 220°C. Belső standardként mesoinozitolt használtunk
[Sawardeker 1965, www.agilent.com].
3.17. Nukleáris mag rezonancia (NMR):
A 13C-NMR spektrumot Varian UNITYINOVA 400 WB (100 MHz for 13C) spectrometer
segítségével készítettük. A méréseket 303K hőmérsékleten és 8 mg/ml D2O oldatban

Proteus morganii törzsek szerológiai keresztkapcsolatai
31
végeztük. Belső standardnak acetont használtunk (δC = 31.07 ppm). A mérések alatt 46000
indukciós bomló jelt gyűjtöttünk (FID). A méréseket 64000 adalék pontértékekkel, 30°
(2.5μs) pulzussal és 3s relaxaciós idővel futtattuk. A Fourier transformáció előtt 3 Hz soros
bővítést alkalmazva a FID jeleket exponenciálisan megsokszoroztuk [Gupta 1995, Péterfi közlésre
benyújtva].
3.18. Növekedésgátlási vizsgálat:
A párhuzamosan vizsgált felező savóhígításokat 1 ml táptalajt tartalmazó Widal csövekben
végeztük. A P. morganii O34 (8622/64), E. coli O111 és S. Adelaide O35 ellen termelt
immunsavó hígítása után minden tesztcsőbe 10 μl standardizált komplementet (Institute
Virion Ltd., Switzerland) és 5 μl baktérium (A600 = 0.6) (UV-VIS photometer Perkin-Elmer,
Germany) szuszpenziót adtunk. A táptalajok abszorpcióját 24 óra után 37°C-on történő
inkubálás után olvastuk le. Mindezek után egy gátlási indexet (I.I.) számoltunk ki minden
egyes vizsgált törzs esetében, ahol az alacsony gátlási index nagyfokú növekedésgátlást
jelentett. (I.I. = a növekedésében gátolt baktérium szuszpenzió abszorpciója/ a nem gátolt
baktérium szuszpenzió abszorpciója x 100)
3.19. Opszonizált baktériumok makrofág inváziója:
Az eredetileg Mills (1994), valamint Murray (2006) által leírt módszer módosításával
végeztük a vizsgálatokat. A log fázisban lévő baktérium tenyészetből (A600 = 0,5) 10 μl-t
adtunk a monolayer RAW264.7 makrofág tenyészethez. Az opsonizációhoz a baktérium
tenyészetet homológ, vagy heterológ immunsavó 1:1000 hígításával kevertük. Negatív
kontroll céljából normál nyúlszérumot használtunk. A szérumban lévő ismeretlen mennyiségű
komplement inaktiválását 30 percig 56°C-ra történő melegítésével végeztük. A baktériumok
és makrofágok közötti kapcsolat kialakulását 5 perces centrifugálással (650g) segítettük. A
harminc perces inkubálás után az extracellulárisan maradt baktériumokat 80 μg/ml gentamicin
oldattal elöltük (90 perces inkubálással). Minden vizsgált baktériumtörzsünk gentamicin
érzékeny volt. A makrofágok által bekebelezett baktériumokat 1% Triton X-100 (Sigma
Chemicals, St. Louis, MO, USA) segítségével történt sejtlízis után szilárd táptalajon
szélesztettük, 24 órás tenyésztés után néhány kolóniát polytrop táptalajon identifikáltunk,
hogy ellenőrizzük azonosságát, majd a telepeket leszámoltuk és a telepszámokat
összehasonlítottuk az opsonizáció nélküli eredményekkel.

Proteus morganii törzsek szerológiai keresztkapcsolatai
32
3.20. Genetikai vizsgálatok:
Az E. coli O111 és S. Adelaide O35 LPS szintézisét irányító enzimek, az O antigén gén
clustere jól ismert, a génszekvencia megtalálható és letölthető a GeneBank
(www.ncbi.nlm.nih.gov/Genbank/) adatbázisából (AF078736, AF285969). A primereket a P.
morganii O34 (8662/64) megfelelő génjének szintetizálásához a vizsgált törzsek manB
(foszfomannomutáz) szekvenciáit használva választottuk ki. A DNS extrakciója 2 percig,
93°C-on történt. A PCR reakciót a következő ciklusok 40-szeri ismétlésével végeztünk. A
94°C-on, 1 percig tartó denaturációt, 90 sec.-ig, 50°C-os megtapadás, majd 72°C-on történő
szintézis követte. A szekvencia analízis a Wang és munkatársai (2000) által leírtaknak
megfelelően történt.
3.21. Statisztikai analízis elkészítéséhez az Origin 7.5 grafikai és statisztikai programot
használtuk (OriginLab Corporation, Northampton MO, USA).
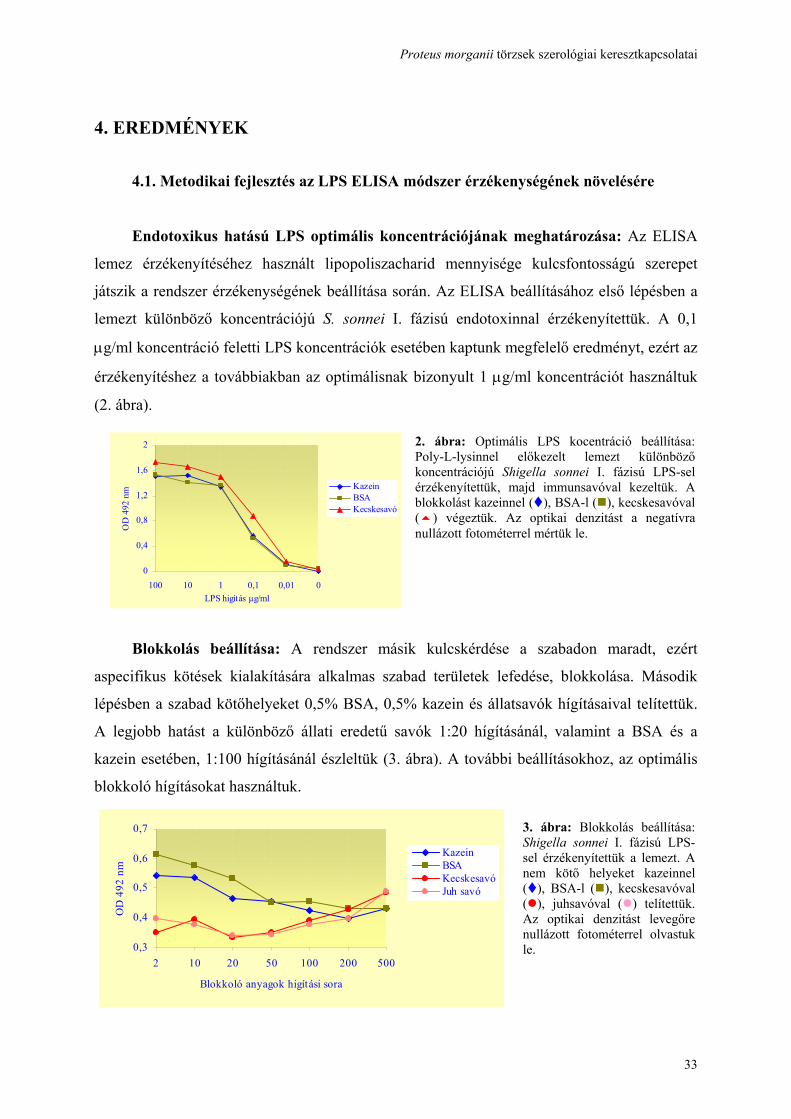
Proteus morganii törzsek szerológiai keresztkapcsolatai
33
4. EREDMÉNYEK
4.1. Metodikai fejlesztés az LPS ELISA módszer érzékenységének növelésére
Endotoxikus hatású LPS optimális koncentrációjának meghatározása: Az ELISA
lemez érzékenyítéséhez használt lipopoliszacharid mennyisége kulcsfontosságú szerepet
játszik a rendszer érzékenységének beállítása során. Az ELISA beállításához első lépésben a
lemezt különböző koncentrációjú S. sonnei I. fázisú endotoxinnal érzékenyítettük. A 0,1
μg/ml koncentráció feletti LPS koncentrációk esetében kaptunk megfelelő eredményt, ezért az
érzékenyítéshez a továbbiakban az optimálisnak bizonyult 1 μg/ml koncentrációt használtuk
(2. ábra).
Blokkolás beállítása: A rendszer másik kulcskérdése a szabadon maradt, ezért
aspecifikus kötések kialakítására alkalmas szabad területek lefedése, blokkolása. Második
lépésben a szabad kötőhelyeket 0,5% BSA, 0,5% kazein és állatsavók hígításaival telítettük.
A legjobb hatást a különböző állati eredetű savók 1:20 hígításánál, valamint a BSA és a
kazein esetében, 1:100 hígításánál észleltük (3. ábra). A további beállításokhoz, az optimális
blokkoló hígításokat használtuk.
0,3
0,4
0,5
0,6
0,7
2 10 20 50 100 200 500
Blokkoló anyagok higítási sora
OD
492
nm
KazeinBSAKecskesavóJuh savó
0
0,4
0,8
1,2
1,6
2
100 10 1 0,1 0,01 0LPS higítás µg/ml
OD
492
nm Kazein
BSAKecskesavó
2. ábra: Optimális LPS kocentráció beállítása: Poly-L-lysinnel előkezelt lemezt különböző koncentrációjú Shigella sonnei I. fázisú LPS-sel érzékenyítettük, majd immunsavóval kezeltük. A blokkolást kazeinnel ( ), BSA-l ( ), kecskesavóval ( ) végeztük. Az optikai denzitást a negatívra nullázott fotométerrel mértük le.
3. ábra: Blokkolás beállítása: Shigella sonnei I. fázisú LPS-sel érzékenyítettük a lemezt. A nem kötő helyeket kazeinnel ( ), BSA-l ( ), kecskesavóval ( ), juhsavóval ( ) telítettük. Az optikai denzitást levegőre nullázott fotométerrel olvastuk le.

Proteus morganii törzsek szerológiai keresztkapcsolatai
34
A blokkoló anyagok összehasonlítása: A kis mennyiségű antigén, valamint a
minimális különbséget hordozó antigének jó és megbízható kimutatására, elkülönítésére a
pozitív és negatív kontroll értéke közötti nagy különbség és a széles mérési tartomány a
legmegfelelőbb. Ennek egyik feltétele, hogy a nem specifikus kötődések, és a háttér a lehető
legkisebb színreakciót adja. Ilyen megfontolásból kerestünk olyan anyagot, amely eleget tud
tenni a fenti feltételeknek. Vizsgálataink során BSA, kazein, kecske, juh, sertés, marha,
tengerimalac, ló savókat teszteltünk és hasonlítottunk össze. Negatív kontroll esetén
statisztikailag szignifikánsan a legkisebb OD-t (p<0,001) a kecskesavóval történt
blokkoláskor kaptuk. Pozitív kontroll esetében a legjobb eredményt szintén a kecskesavó
szolgáltatta, a különbség itt is szignifikáns volt (p<0,001). Az egyes állatsavók hatásai között
nem észleltünk szignifikáns különbséget. Ezzel szemben a hagyományosan használt kazein és
BSA a vizsgálatok során egyaránt elégtelennek bizonyultak. A kecskesavót alkalmaztuk a
további vizsgálatokban azon megfontolás alapján is, hogy ezzel minimálisra csökkentsük az
interakciókat, ugyanis a konjugátum kecskében termelt nyúl ellenes IgG volt.
A megbízhatóság vizsgálata: A megbízhatóság és ismételhetőség bizonyítására 12
különböző kísérletben 6-6 párhuzamos meghatározást végeztünk. Az „intra-assay” változások
elhanyagolható mértékűek voltak (a pozitív kontroll szórása SD=0,033, míg a negatív kontroll
szórása SD=0,023). Ugyanakkor az „inter-assay” szórások sem mutattak lényeges
különbséget (pozitív kontroll szórása SD=0,088, míg a negatív kontrollé SD=0,097). Mindkét
esetben a korábban már ismert szignifikáns különbségek változatlanul detektálhatók voltak a
kecskesavóval illetve a hagyományos BSA-val vagy kazeinnel történt blokkolások között.
Mivel nem ismert, hogy a szérum egyes fehérje komponensei milyen mértékben
játszanak szerepet a szabadon maradt felület lefedésében, 20 különböző egyedből származó
kecskesavót is összehasonlítottunk. Ezáltal ki lehetett szűrni az egyedi fehérje összetételű
savók használatából eredő esetleges hibákat is. Az egyes mérések közt nem találtunk
szignifikáns eltérést, a szórás is a korábban regisztrált tartományban maradt (pozitív kontroll
esetén SD=0,080, a negatív kontrollé SD=0,044 ).
Kompetitív blokkolás: Választ kerestünk arra a kérdésre is, hogy miért kapunk jobb
eredményeket a kecskesavó használatával, mint a hagyományos blokkoló anyagokkal? Jobb
fedést biztosítanak, vagy pedig kevesebb az aspecifikus kötődés? Ismert, hogy az LPS nagyon
könnyen alkot komplexet BSA-val. Ennek a kérdésnek a megválaszolására kompetitív
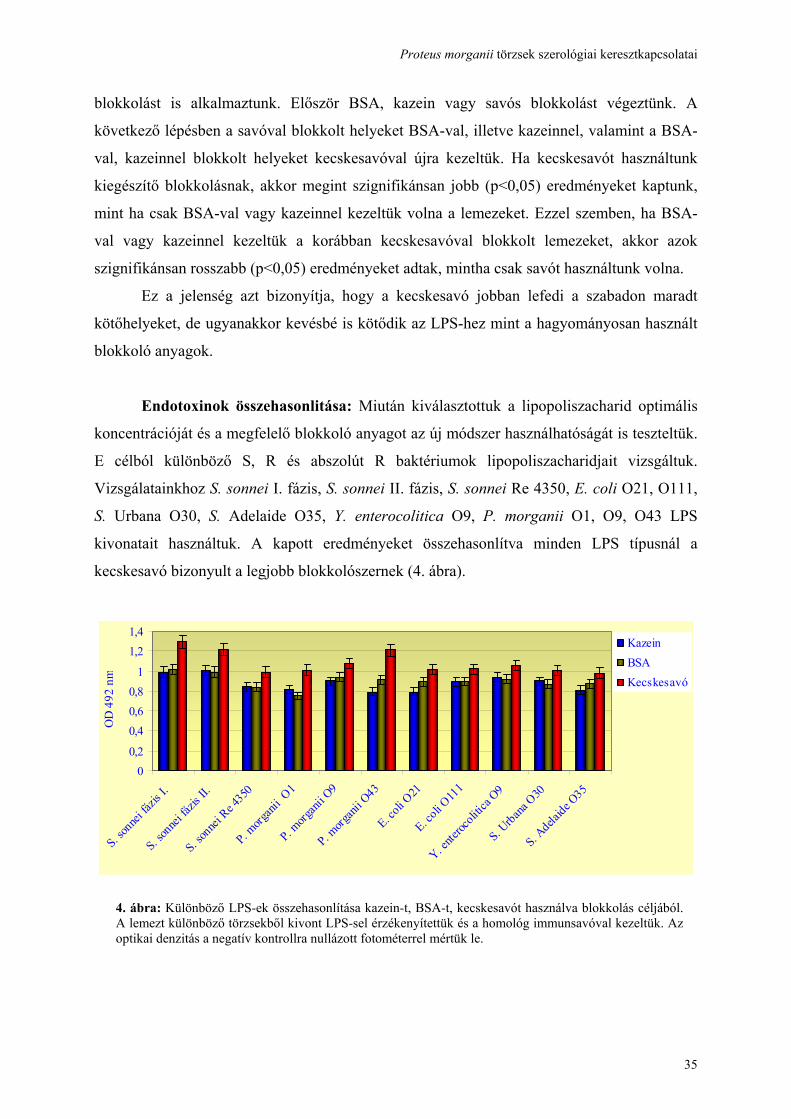
Proteus morganii törzsek szerológiai keresztkapcsolatai
35
blokkolást is alkalmaztunk. Először BSA, kazein vagy savós blokkolást végeztünk. A
következő lépésben a savóval blokkolt helyeket BSA-val, illetve kazeinnel, valamint a BSA-
val, kazeinnel blokkolt helyeket kecskesavóval újra kezeltük. Ha kecskesavót használtunk
kiegészítő blokkolásnak, akkor megint szignifikánsan jobb (p<0,05) eredményeket kaptunk,
mint ha csak BSA-val vagy kazeinnel kezeltük volna a lemezeket. Ezzel szemben, ha BSA-
val vagy kazeinnel kezeltük a korábban kecskesavóval blokkolt lemezeket, akkor azok
szignifikánsan rosszabb (p<0,05) eredményeket adtak, mintha csak savót használtunk volna.
Ez a jelenség azt bizonyítja, hogy a kecskesavó jobban lefedi a szabadon maradt
kötőhelyeket, de ugyanakkor kevésbé is kötődik az LPS-hez mint a hagyományosan használt
blokkoló anyagok.
Endotoxinok összehasonlitása: Miután kiválasztottuk a lipopoliszacharid optimális
koncentrációját és a megfelelő blokkoló anyagot az új módszer használhatóságát is teszteltük.
E célból különböző S, R és abszolút R baktériumok lipopoliszacharidjait vizsgáltuk.
Vizsgálatainkhoz S. sonnei I. fázis, S. sonnei II. fázis, S. sonnei Re 4350, E. coli O21, O111,
S. Urbana O30, S. Adelaide O35, Y. enterocolitica O9, P. morganii O1, O9, O43 LPS
kivonatait használtuk. A kapott eredményeket összehasonlítva minden LPS típusnál a
kecskesavó bizonyult a legjobb blokkolószernek (4. ábra).
00,2
0,40,60,8
1
1,21,4
S. sonnei
fázis
I.
S. sonnei
fázis
II.
S. sonnei
Re 4350
P. morg
anii O
1
P. morg
anii O
9
P. morg
anii O
43
E. coli O
21
E. coli O
111
Y. ente
rocoliti
ca O9
S. Urbana
O30
S. Adela
ide O35
OD
492
nm
KazeinBSAKecskesavó
4. ábra: Különböző LPS-ek összehasonlítása kazein-t, BSA-t, kecskesavót használva blokkolás céljából. A lemezt különböző törzsekből kivont LPS-sel érzékenyítettük és a homológ immunsavóval kezeltük. Az optikai denzitás a negatív kontrollra nullázott fotométerrel mértük le.
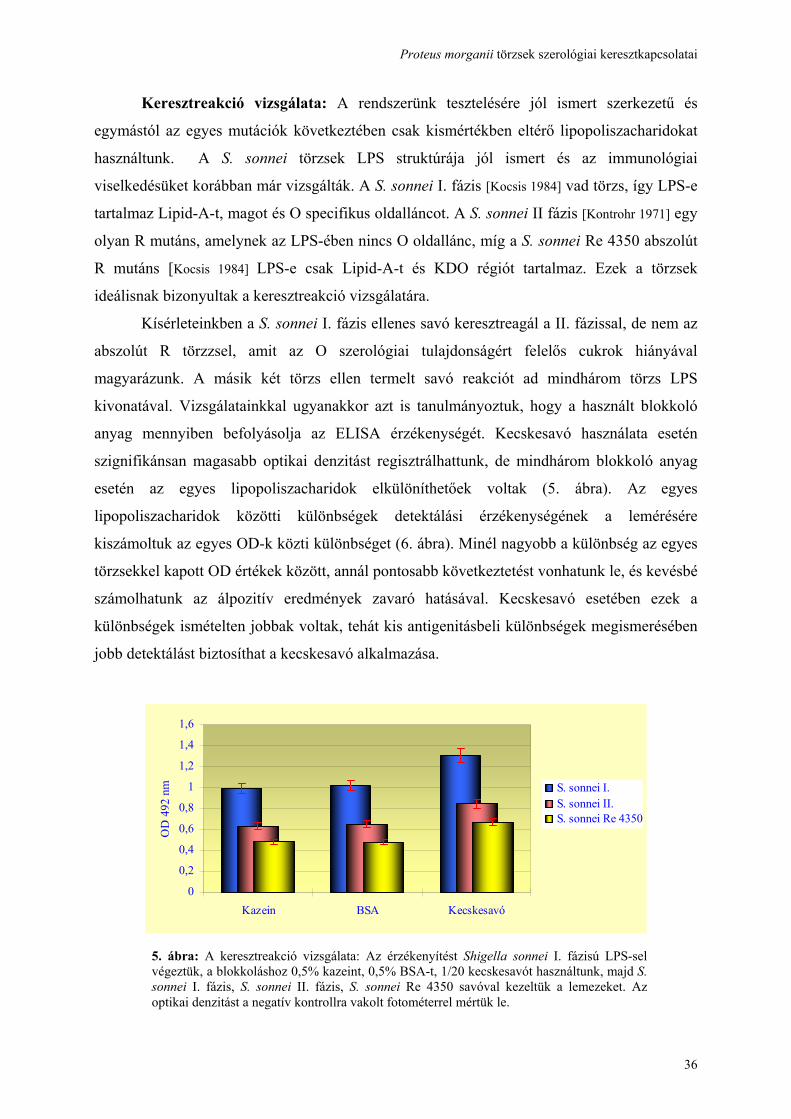
Proteus morganii törzsek szerológiai keresztkapcsolatai
36
Keresztreakció vizsgálata: A rendszerünk tesztelésére jól ismert szerkezetű és
egymástól az egyes mutációk következtében csak kismértékben eltérő lipopoliszacharidokat
használtunk. A S. sonnei törzsek LPS struktúrája jól ismert és az immunológiai
viselkedésüket korábban már vizsgálták. A S. sonnei I. fázis [Kocsis 1984] vad törzs, így LPS-e
tartalmaz Lipid-A-t, magot és O specifikus oldalláncot. A S. sonnei II fázis [Kontrohr 1971] egy
olyan R mutáns, amelynek az LPS-ében nincs O oldallánc, míg a S. sonnei Re 4350 abszolút
R mutáns [Kocsis 1984] LPS-e csak Lipid-A-t és KDO régiót tartalmaz. Ezek a törzsek
ideálisnak bizonyultak a keresztreakció vizsgálatára.
Kísérleteinkben a S. sonnei I. fázis ellenes savó keresztreagál a II. fázissal, de nem az
abszolút R törzzsel, amit az O szerológiai tulajdonságért felelős cukrok hiányával
magyarázunk. A másik két törzs ellen termelt savó reakciót ad mindhárom törzs LPS
kivonatával. Vizsgálatainkkal ugyanakkor azt is tanulmányoztuk, hogy a használt blokkoló
anyag mennyiben befolyásolja az ELISA érzékenységét. Kecskesavó használata esetén
szignifikánsan magasabb optikai denzitást regisztrálhattunk, de mindhárom blokkoló anyag
esetén az egyes lipopoliszacharidok elkülöníthetőek voltak (5. ábra). Az egyes
lipopoliszacharidok közötti különbségek detektálási érzékenységének a lemérésére
kiszámoltuk az egyes OD-k közti különbséget (6. ábra). Minél nagyobb a különbség az egyes
törzsekkel kapott OD értékek között, annál pontosabb következtetést vonhatunk le, és kevésbé
számolhatunk az álpozitív eredmények zavaró hatásával. Kecskesavó esetében ezek a
különbségek ismételten jobbak voltak, tehát kis antigenitásbeli különbségek megismerésében
jobb detektálást biztosíthat a kecskesavó alkalmazása.
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
Kazein BSA Kecskesavó
OD
492
nm S. sonnei I.
S. sonnei II.S. sonnei Re 4350
5. ábra: A keresztreakció vizsgálata: Az érzékenyítést Shigella sonnei I. fázisú LPS-sel végeztük, a blokkoláshoz 0,5% kazeint, 0,5% BSA-t, 1/20 kecskesavót használtunk, majd S. sonnei I. fázis, S. sonnei II. fázis, S. sonnei Re 4350 savóval kezeltük a lemezeket. Az optikai denzitást a negatív kontrollra vakolt fotométerrel mértük le.

Proteus morganii törzsek szerológiai keresztkapcsolatai
37
4.2. Az újonnan beállított módszer klinikai használhatóságának kipróbálása
A rendszer beállítása után megvizsgáltuk, hogy az hogyan működik a klinikai
gyakorlatban, mennyire használható klinikai minták vizsgálatára. Appendicitis miatt műtéten
átesett betegek savóit használtuk erre a célra.
A 47 operált egyén közül 9-nél az eltávolított appendix szövettani elváltozást nem
mutatott, vagyis az appendicitis diagnózisa szövettanilag nem igazolódott. Ezeket a
személyeket használtuk negatív kontroll céljából. A betegek közül 12 esetben simplex, 18
esetben phlegmonosus (gangrenas) és 8 beteg esetében pedig perforalt appendixet találtak.
Ugyanezeknek a savóknak egy korábbi vizsgálatában, melyben haemagglutinációs módszert
alkalmaztak [Antal 1998], a különböző, normál bakteriális flórához tartozó baktériumok
lipopoliszacharidjai ellen emelkedő haemagglutinációs titert lehetett regisztrálni, azonban az
alacsony titerek miatt ezek értékelése nehézségekbe ütközött. ELISA módszerünk
segítségével, amely a gyulladás súlyosságával párhuzamosan emelkedő ellenanyag titereket
mutatott, egyértelmű szoros összefüggést találtunk a szerológiai viselkedés és a szövettani kép
között. A negatív kontroll és appendicitisben szenvedő betegek savói között a különbség
szignifikáns volt (E. coli p<0,001, B. fragilis p<0,005). Néhány reprezentatív eredményt a 1.
táblázat és 7. ábra mutat be. A gyulladás különböző fokozatai között a különbség nem volt
szignifikáns.
6. ábra: Keresztreakció vizsgálata: A módszer megegyezik az előző ábrán alkalmazottal. OD különbségeket számoltunk a S. sonnei I. fázis és S. sonnei Re 4350 ( ), S. sonnei I. fázis és S. sonnei II. fázis ( ) és S. sonnei II.fázis és S. sonnei Re 4350 ( ) között.
0
0,10,2
0,3
0,40,5
0,6
0,70,8
0,9
Kazein BSA Kecskesavó
OD
kül
önbs
égek
S. sonnei I.- S. sonnei Re 4350S. sonnei I.- S. sonnei II.S. sonnei II.- S. sonnei Re 4350
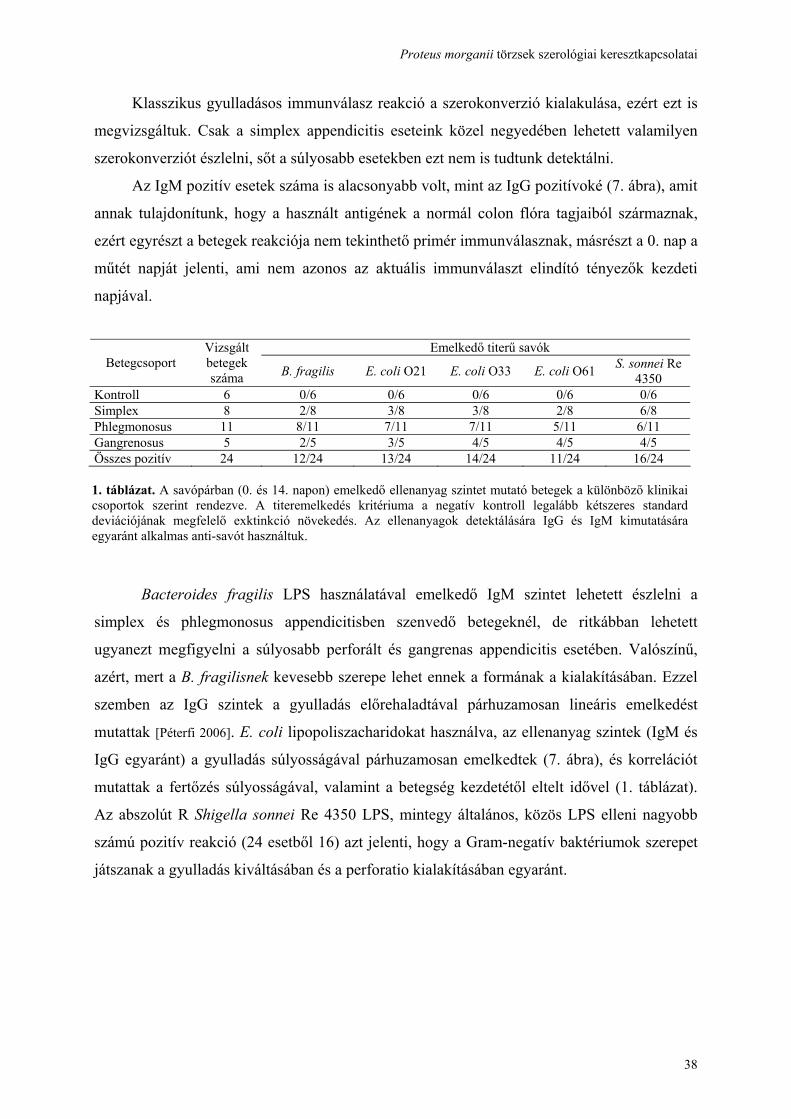
Proteus morganii törzsek szerológiai keresztkapcsolatai
38
Klasszikus gyulladásos immunválasz reakció a szerokonverzió kialakulása, ezért ezt is
megvizsgáltuk. Csak a simplex appendicitis eseteink közel negyedében lehetett valamilyen
szerokonverziót észlelni, sőt a súlyosabb esetekben ezt nem is tudtunk detektálni.
Az IgM pozitív esetek száma is alacsonyabb volt, mint az IgG pozitívoké (7. ábra), amit
annak tulajdonítunk, hogy a használt antigének a normál colon flóra tagjaiból származnak,
ezért egyrészt a betegek reakciója nem tekinthető primér immunválasznak, másrészt a 0. nap a
műtét napját jelenti, ami nem azonos az aktuális immunválaszt elindító tényezők kezdeti
napjával.
Bacteroides fragilis LPS használatával emelkedő IgM szintet lehetett észlelni a
simplex és phlegmonosus appendicitisben szenvedő betegeknél, de ritkábban lehetett
ugyanezt megfigyelni a súlyosabb perforált és gangrenas appendicitis esetében. Valószínű,
azért, mert a B. fragilisnek kevesebb szerepe lehet ennek a formának a kialakításában. Ezzel
szemben az IgG szintek a gyulladás előrehaladtával párhuzamosan lineáris emelkedést
mutattak [Péterfi 2006]. E. coli lipopoliszacharidokat használva, az ellenanyag szintek (IgM és
IgG egyaránt) a gyulladás súlyosságával párhuzamosan emelkedtek (7. ábra), és korrelációt
mutattak a fertőzés súlyosságával, valamint a betegség kezdetétől eltelt idővel (1. táblázat).
Az abszolút R Shigella sonnei Re 4350 LPS, mintegy általános, közös LPS elleni nagyobb
számú pozitív reakció (24 esetből 16) azt jelenti, hogy a Gram-negatív baktériumok szerepet
játszanak a gyulladás kiváltásában és a perforatio kialakításában egyaránt.
Emelkedő titerű savók Betegcsoport
Vizsgált betegek száma B. fragilis E. coli O21 E. coli O33 E. coli O61 S. sonnei Re
4350 Kontroll 6 0/6 0/6 0/6 0/6 0/6 Simplex 8 2/8 3/8 3/8 2/8 6/8 Phlegmonosus 11 8/11 7/11 7/11 5/11 6/11 Gangrenosus 5 2/5 3/5 4/5 4/5 4/5 Összes pozitív 24 12/24 13/24 14/24 11/24 16/24
1. táblázat. A savópárban (0. és 14. napon) emelkedő ellenanyag szintet mutató betegek a különböző klinikai csoportok szerint rendezve. A titeremelkedés kritériuma a negatív kontroll legalább kétszeres standard deviációjának megfelelő exktinkció növekedés. Az ellenanyagok detektálására IgG és IgM kimutatására egyaránt alkalmas anti-savót használtuk.
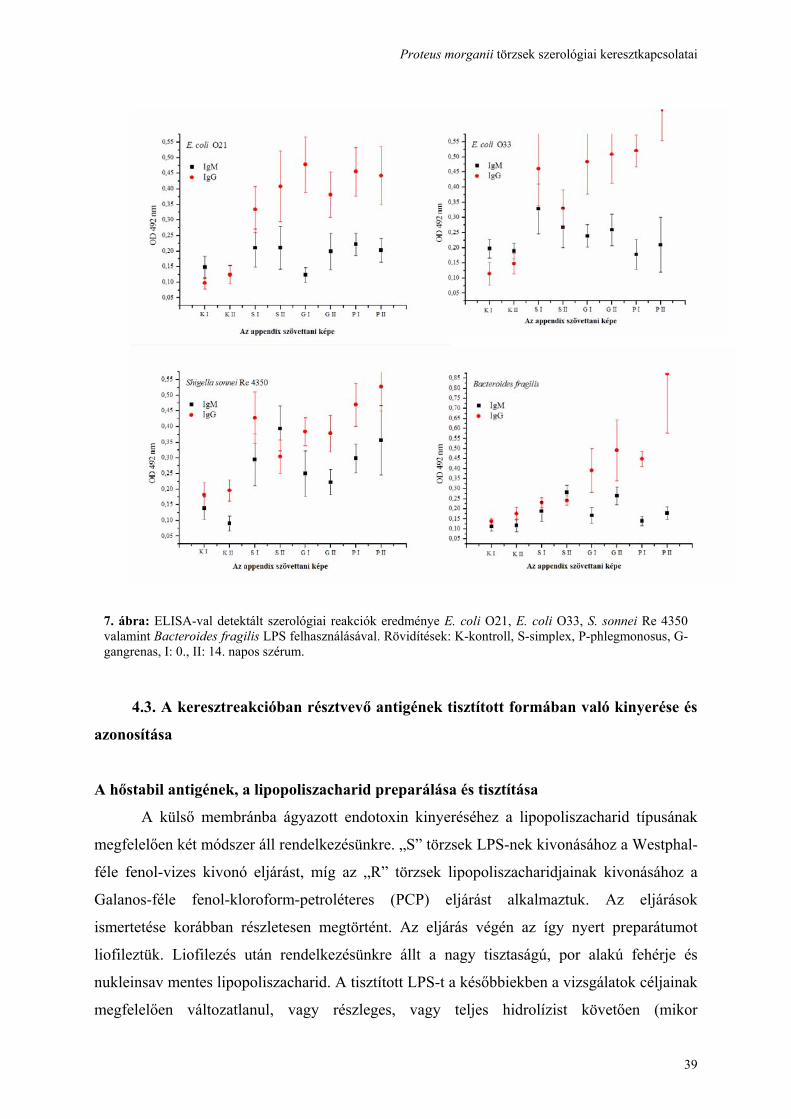
Proteus morganii törzsek szerológiai keresztkapcsolatai
39
4.3. A keresztreakcióban résztvevő antigének tisztított formában való kinyerése és
azonosítása
A hőstabil antigének, a lipopoliszacharid preparálása és tisztítása
A külső membránba ágyazott endotoxin kinyeréséhez a lipopoliszacharid típusának
megfelelően két módszer áll rendelkezésünkre. „S” törzsek LPS-nek kivonásához a Westphal-
féle fenol-vizes kivonó eljárást, míg az „R” törzsek lipopoliszacharidjainak kivonásához a
Galanos-féle fenol-kloroform-petroléteres (PCP) eljárást alkalmaztuk. Az eljárások
ismertetése korábban részletesen megtörtént. Az eljárás végén az így nyert preparátumot
liofileztük. Liofilezés után rendelkezésünkre állt a nagy tisztaságú, por alakú fehérje és
nukleinsav mentes lipopoliszacharid. A tisztított LPS-t a későbbiekben a vizsgálatok céljainak
megfelelően változatlanul, vagy részleges, vagy teljes hidrolízist követően (mikor
7. ábra: ELISA-val detektált szerológiai reakciók eredménye E. coli O21, E. coli O33, S. sonnei Re 4350 valamint Bacteroides fragilis LPS felhasználásával. Rövidítések: K-kontroll, S-simplex, P-phlegmonosus, G-gangrenas, I: 0., II: 14. napos szérum.
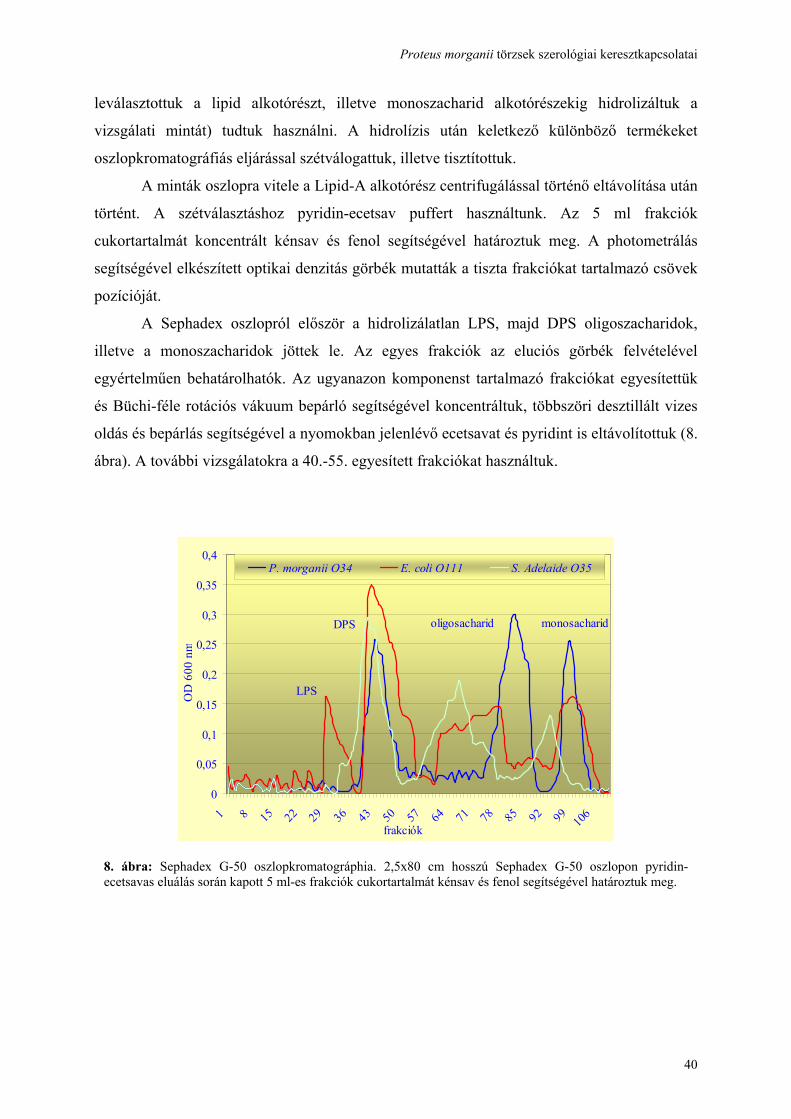
Proteus morganii törzsek szerológiai keresztkapcsolatai
40
leválasztottuk a lipid alkotórészt, illetve monoszacharid alkotórészekig hidrolizáltuk a
vizsgálati mintát) tudtuk használni. A hidrolízis után keletkező különböző termékeket
oszlopkromatográfiás eljárással szétválogattuk, illetve tisztítottuk.
A minták oszlopra vitele a Lipid-A alkotórész centrifugálással történő eltávolítása után
történt. A szétválasztáshoz pyridin-ecetsav puffert használtunk. Az 5 ml frakciók
cukortartalmát koncentrált kénsav és fenol segítségével határoztuk meg. A photometrálás
segítségével elkészített optikai denzitás görbék mutatták a tiszta frakciókat tartalmazó csövek
pozícióját.
A Sephadex oszlopról először a hidrolizálatlan LPS, majd DPS oligoszacharidok,
illetve a monoszacharidok jöttek le. Az egyes frakciók az eluciós görbék felvételével
egyértelműen behatárolhatók. Az ugyanazon komponenst tartalmazó frakciókat egyesítettük
és Büchi-féle rotációs vákuum bepárló segítségével koncentráltuk, többszöri desztillált vizes
oldás és bepárlás segítségével a nyomokban jelenlévő ecetsavat és pyridint is eltávolítottuk (8.
ábra). A további vizsgálatokra a 40.-55. egyesített frakciókat használtuk.
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
1 8 15 22 29 36 43 50 57 64 71 78 85 92 99 106
frakciók
OD
600
nm
P. morganii O34 E. coli O111 S. Adelaide O35
LPS
DPS oligosacharid monosacharid
8. ábra: Sephadex G-50 oszlopkromatográphia. 2,5x80 cm hosszú Sephadex G-50 oszlopon pyridin-ecetsavas eluálás során kapott 5 ml-es frakciók cukortartalmát kénsav és fenol segítségével határoztuk meg.

Proteus morganii törzsek szerológiai keresztkapcsolatai
41
A hőlabilis, fehérje természetű antigének preparálása, tisztítása:
A külső membrán fehérjék kivonásához a korábban
ismertetett protokoll (Anyag és módszer: 3.4.) szerint történt. A
grádiens ultracentrifugálás után (9. ábra) a különböző frakciókat
deionizált vízben oldva –20°C-on fagyasztva tároltuk. A külső
membránt tartalmazó frakció fehérje összetevőit jellemeztük. A
36, 41 kDa nagyságú major fehérje mindhárom vizsgált törzsben
megtalálható volt.
4.4. A korábban leírt szerológiai keresztkapcsolatok megerősítése, pontosítása
érzékenyebb módszerek használatával
4.4.1. Reakciók teljes baktériumokon:
4.4.1.1. Csőagglutináció: Csőagglutinációval reprodukálni tudtuk a Rauss és Vörös
által elért eredményeket [Rauss 1967]. Nem találtunk lényeges eltérést az egyes baktérium
törzsek között (2. táblázat). Az E. coli-val szemben kapott magasabb agglutinációs titerek
feltehetően az E. coli sejtek felületén nagyobb számban megjelenő antigéndeterminánsoknak
köszönhetők.
Agglutinációs titer Baktérium törzsek P. morganii O34 E. coli O111:B4 S. Adelaide O35
P. morganii O34 3200 3200 1600 E. coli O111:B4 12800 12800 6400 S. Adelaide O35 3200 3200 3200
2. táblázat. P. morganii O34, E. coli O111 és S. Adelaide O35 csőagglutinációs vizsgálata. Minden immunsavó felező hígítása után 25 μl (A600=1.2), 100 C fokon elölt baktérium szuszpenziót használtunk. Az eredményt 24 órás 37 C fokos inkubálás után olvastuk le.
9. ábra: A külső membrán fehérjék grádiens ultracentrifugálás után
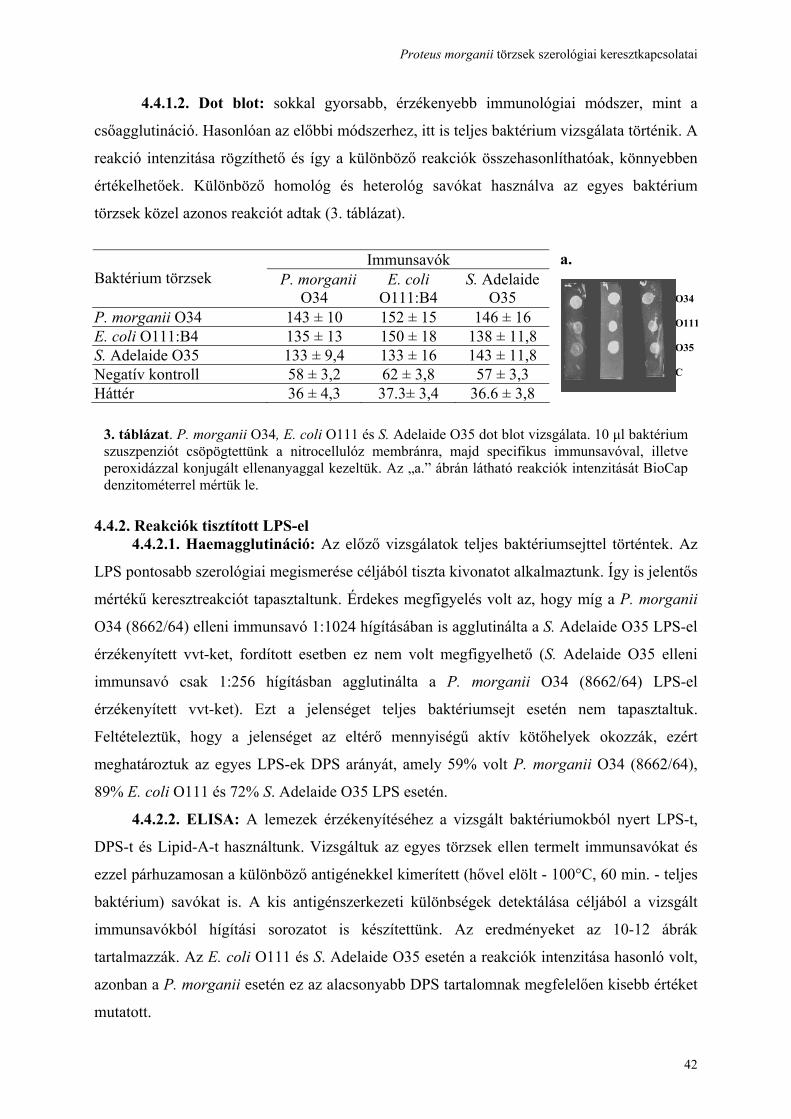
Proteus morganii törzsek szerológiai keresztkapcsolatai
42
O34 O111 O35 C
4.4.1.2. Dot blot: sokkal gyorsabb, érzékenyebb immunológiai módszer, mint a
csőagglutináció. Hasonlóan az előbbi módszerhez, itt is teljes baktérium vizsgálata történik. A
reakció intenzitása rögzíthető és így a különböző reakciók összehasonlíthatóak, könnyebben
értékelhetőek. Különböző homológ és heterológ savókat használva az egyes baktérium
törzsek közel azonos reakciót adtak (3. táblázat).
a.
4.4.2. Reakciók tisztított LPS-el
4.4.2.1. Haemagglutináció: Az előző vizsgálatok teljes baktériumsejttel történtek. Az
LPS pontosabb szerológiai megismerése céljából tiszta kivonatot alkalmaztunk. Így is jelentős
mértékű keresztreakciót tapasztaltunk. Érdekes megfigyelés volt az, hogy míg a P. morganii
O34 (8662/64) elleni immunsavó 1:1024 hígításában is agglutinálta a S. Adelaide O35 LPS-el
érzékenyített vvt-ket, fordított esetben ez nem volt megfigyelhető (S. Adelaide O35 elleni
immunsavó csak 1:256 hígításban agglutinálta a P. morganii O34 (8662/64) LPS-el
érzékenyített vvt-ket). Ezt a jelenséget teljes baktériumsejt esetén nem tapasztaltuk.
Feltételeztük, hogy a jelenséget az eltérő mennyiségű aktív kötőhelyek okozzák, ezért
meghatároztuk az egyes LPS-ek DPS arányát, amely 59% volt P. morganii O34 (8662/64),
89% E. coli O111 és 72% S. Adelaide O35 LPS esetén.
4.4.2.2. ELISA: A lemezek érzékenyítéséhez a vizsgált baktériumokból nyert LPS-t,
DPS-t és Lipid-A-t használtunk. Vizsgáltuk az egyes törzsek ellen termelt immunsavókat és
ezzel párhuzamosan a különböző antigénekkel kimerített (hővel elölt - 100°C, 60 min. - teljes
baktérium) savókat is. A kis antigénszerkezeti különbségek detektálása céljából a vizsgált
immunsavókból hígítási sorozatot is készítettünk. Az eredményeket az 10-12 ábrák
tartalmazzák. Az E. coli O111 és S. Adelaide O35 esetén a reakciók intenzitása hasonló volt,
azonban a P. morganii esetén ez az alacsonyabb DPS tartalomnak megfelelően kisebb értéket
mutatott.
Immunsavók Baktérium törzsek P. morganii
O34 E. coli
O111:B4 S. Adelaide
O35 P. morganii O34 143 ± 10 152 ± 15 146 ± 16 E. coli O111:B4 135 ± 13 150 ± 18 138 ± 11,8 S. Adelaide O35 133 ± 9,4 133 ± 16 143 ± 11,8 Negatív kontroll 58 ± 3,2 62 ± 3,8 57 ± 3,3 Háttér 36 ± 4,3 37.3± 3,4 36.6 ± 3,8
3. táblázat. P. morganii O34, E. coli O111 és S. Adelaide O35 dot blot vizsgálata. 10 μl baktérium szuszpenziót csöpögtettünk a nitrocellulóz membránra, majd specifikus immunsavóval, illetve peroxidázzal konjugált ellenanyaggal kezeltük. Az „a.” ábrán látható reakciók intenzitását BioCap denzitométerrel mértük le.
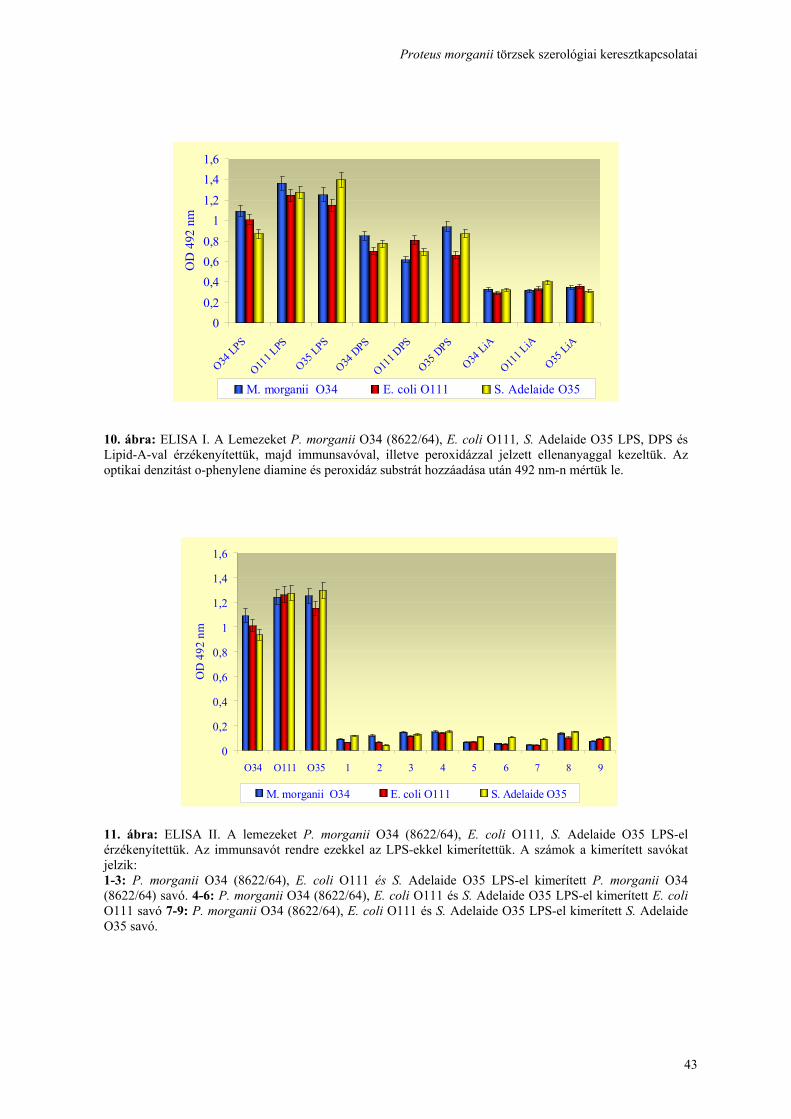
Proteus morganii törzsek szerológiai keresztkapcsolatai
43
00,20,40,60,8
11,21,41,6
O34 LPS
O111 L
PS
O35 LPS
O34 D
PS
O111 D
PS
O35 D
PS
O34 LiA
O111 L
iA
O35 LiA
OD
492
nm
M. morganii O34 E. coli O111 S. Adelaide O35
10. ábra: ELISA I. A Lemezeket P. morganii O34 (8622/64), E. coli O111, S. Adelaide O35 LPS, DPS és Lipid-A-val érzékenyítettük, majd immunsavóval, illetve peroxidázzal jelzett ellenanyaggal kezeltük. Az optikai denzitást o-phenylene diamine és peroxidáz substrát hozzáadása után 492 nm-n mértük le.
11. ábra: ELISA II. A lemezeket P. morganii O34 (8622/64), E. coli O111, S. Adelaide O35 LPS-el érzékenyítettük. Az immunsavót rendre ezekkel az LPS-ekkel kimerítettük. A számok a kimerített savókat jelzik: 1-3: P. morganii O34 (8622/64), E. coli O111 és S. Adelaide O35 LPS-el kimerített P. morganii O34 (8622/64) savó. 4-6: P. morganii O34 (8622/64), E. coli O111 és S. Adelaide O35 LPS-el kimerített E. coli O111 savó 7-9: P. morganii O34 (8622/64), E. coli O111 és S. Adelaide O35 LPS-el kimerített S. Adelaide O35 savó.
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
O34 O111 O35 1 2 3 4 5 6 7 8 9
OD
492
nm
M. morganii O34 E. coli O111 S. Adelaide O35
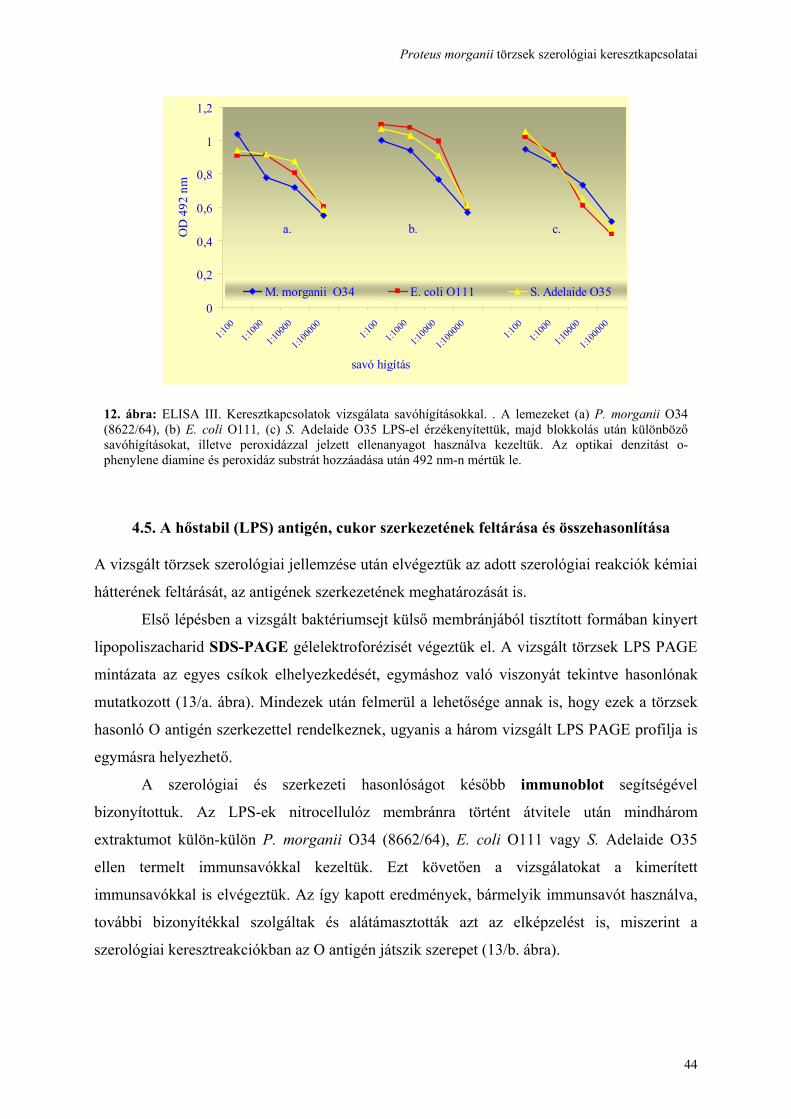
Proteus morganii törzsek szerológiai keresztkapcsolatai
44
a. b. c.
0
0,2
0,4
0,6
0,8
1
1,2
1:100
1:100
0
1:100
00
1:100
000
1:100
1:100
0
1:100
00
1:100
000
1:100
1:100
0
1:100
00
1:100
000
savó hígítás
OD
492
nm
M. morganii O34 E. coli O111 S. Adelaide O35
4.5. A hőstabil (LPS) antigén, cukor szerkezetének feltárása és összehasonlítása
A vizsgált törzsek szerológiai jellemzése után elvégeztük az adott szerológiai reakciók kémiai
hátterének feltárását, az antigének szerkezetének meghatározását is.
Első lépésben a vizsgált baktériumsejt külső membránjából tisztított formában kinyert
lipopoliszacharid SDS-PAGE gélelektroforézisét végeztük el. A vizsgált törzsek LPS PAGE
mintázata az egyes csíkok elhelyezkedését, egymáshoz való viszonyát tekintve hasonlónak
mutatkozott (13/a. ábra). Mindezek után felmerül a lehetősége annak is, hogy ezek a törzsek
hasonló O antigén szerkezettel rendelkeznek, ugyanis a három vizsgált LPS PAGE profilja is
egymásra helyezhető.
A szerológiai és szerkezeti hasonlóságot később immunoblot segítségével
bizonyítottuk. Az LPS-ek nitrocellulóz membránra történt átvitele után mindhárom
extraktumot külön-külön P. morganii O34 (8662/64), E. coli O111 vagy S. Adelaide O35
ellen termelt immunsavókkal kezeltük. Ezt követően a vizsgálatokat a kimerített
immunsavókkal is elvégeztük. Az így kapott eredmények, bármelyik immunsavót használva,
további bizonyítékkal szolgáltak és alátámasztották azt az elképzelést is, miszerint a
szerológiai keresztreakciókban az O antigén játszik szerepet (13/b. ábra).
12. ábra: ELISA III. Keresztkapcsolatok vizsgálata savóhígításokkal. . A lemezeket (a) P. morganii O34 (8622/64), (b) E. coli O111, (c) S. Adelaide O35 LPS-el érzékenyítettük, majd blokkolás után különböző savóhígításokat, illetve peroxidázzal jelzett ellenanyagot használva kezeltük. Az optikai denzitást o-phenylene diamine és peroxidáz substrát hozzáadása után 492 nm-n mértük le.
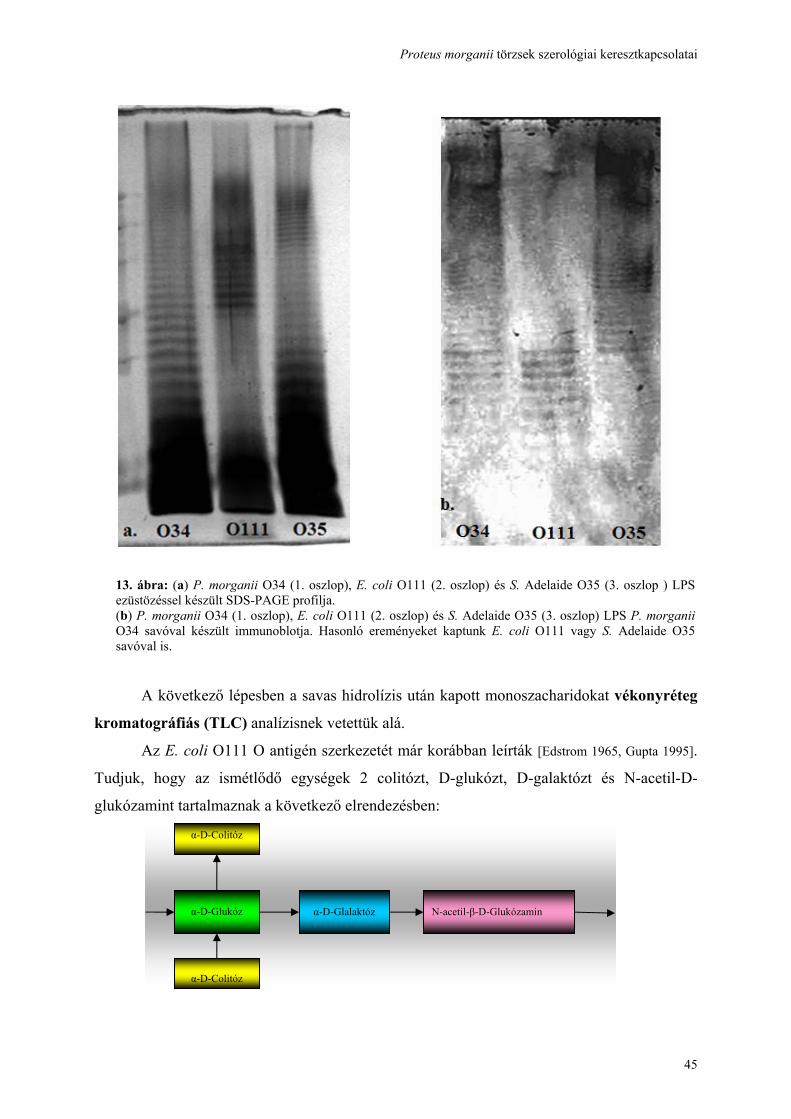
Proteus morganii törzsek szerológiai keresztkapcsolatai
45
A következő lépesben a savas hidrolízis után kapott monoszacharidokat vékonyréteg
kromatográfiás (TLC) analízisnek vetettük alá.
Az E. coli O111 O antigén szerkezetét már korábban leírták [Edstrom 1965, Gupta 1995].
Tudjuk, hogy az ismétlődő egységek 2 colitózt, D-glukózt, D-galaktózt és N-acetil-D-
glukózamint tartalmaznak a következő elrendezésben:
α-D-Colitóz
α-D-Colitóz
α-D-Glukóz
α-D-Glalaktóz
N-acetil-β-D-Glukózamin
13. ábra: (a) P. morganii O34 (1. oszlop), E. coli O111 (2. oszlop) és S. Adelaide O35 (3. oszlop ) LPS ezüstözéssel készült SDS-PAGE profilja. (b) P. morganii O34 (1. oszlop), E. coli O111 (2. oszlop) és S. Adelaide O35 (3. oszlop) LPS P. morganii O34 savóval készült immunoblotja. Hasonló ereményeket kaptunk E. coli O111 vagy S. Adelaide O35 savóval is.

Proteus morganii törzsek szerológiai keresztkapcsolatai
46
A vékonyréteg kromatográfián a fenti monoszacharidok megjelenését, detektálását
vártuk, mindhárom törzs esetén, amennyiben egyforma a három vizsgált baktérium O-
oldallánca. A vékonyréteg kromatográfia során a neutrális cukrok között két foltot találtunk,
melyek a standardként használt galaktóz és glukóz vonalában mutatkoztak és a retenciós idő
is megfelelt a két neutrális cukor retenciós idejének. A colitóz (3,6 dideoxy-L-galactose),
mely biztos összetevője az E. coli O111 O antigénnek, megfelelő mennyiségű autentikus
colitóz hiányában nem volt meghatározható, de a glukóznak megfelelő foltok sokkal
nagyobbak voltak, nagyobb mennyiségű anyagot mutattak, mint azt mólsúlyuk miatt vártuk.
Feltehető, hogy a colitóz a glukózzal fut együtt. Ezt a colitóz szerkezetével tudjuk
magyarázni, ugyanis az a galaktóztól abban különbözik, hogy a 3 és 6 pozícióban hiányzik a
hydroxil gyök. Ez olyan változásokat eredményezhet, hogy a kromatográfiás eljárás során
gyorsabban vándorol, mint a galaktóz, de a glukózzal egyszerre, és így a két cukor egymásra
vetül. Mivel ez a természetben ritkán előforduló cukor molekula a kereskedelmi forgalomban
nem kapható, azt standardként használni nem tudtuk. Elméletünket ily módon alátámasztani
sem lehet. Azonban, hogyha meghatározzuk a glukóznak és galaktóznak megfelelő foltok
területét és intenzitását visszakapjuk a glukóz (colitózzal együtt) és galaktóz 3:1 arányát,
vagyis az ismétlődő egységekben ismert cukorarányt (14. ábra). Mivel a colitózt nem sikerült
azonosítanunk további vizsgálatokra volt szükségünk.
A tömegspektrométerrel összekötött gázkromatográfia alkalmas lehet az összetevő
monoszacharidok pontos identifikálására, az NMR vizsgálat segítségével pedig akár az egyes
összetevők egymáshoz viszonyított helyzetét, a pontos szerkezetet és felépítést is meg lehet
határozni.
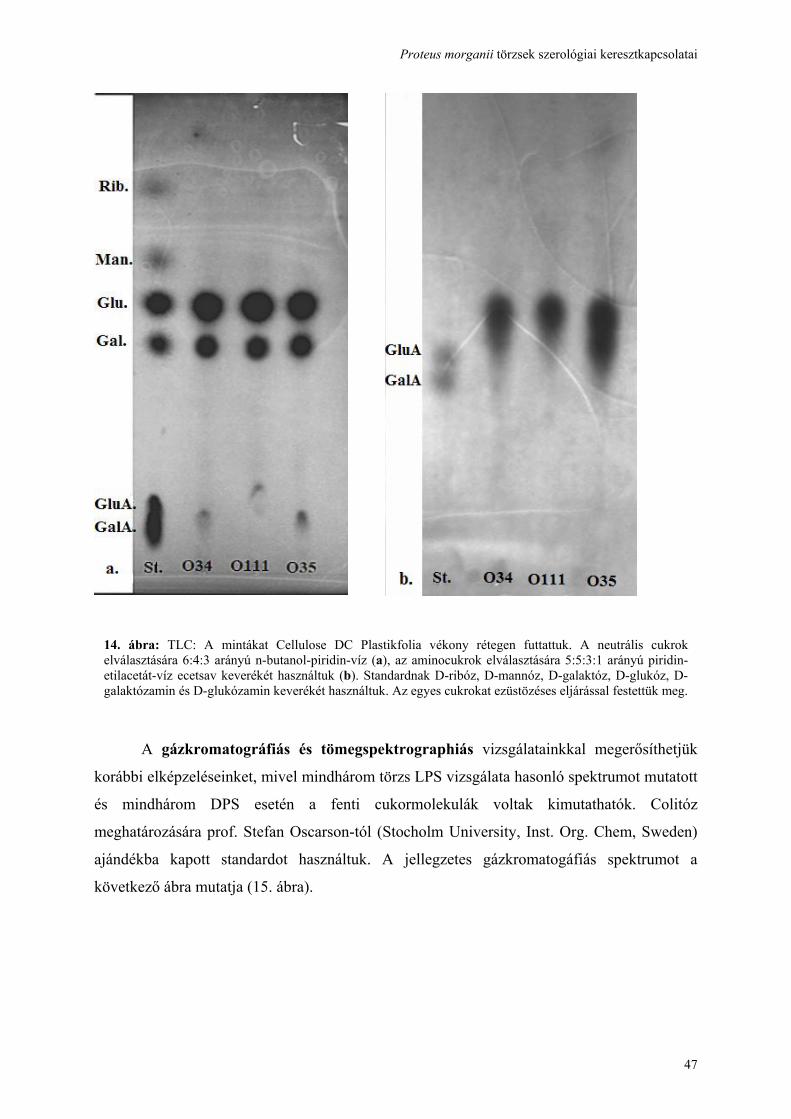
Proteus morganii törzsek szerológiai keresztkapcsolatai
47
A gázkromatográfiás és tömegspektrographiás vizsgálatainkkal megerősíthetjük
korábbi elképzeléseinket, mivel mindhárom törzs LPS vizsgálata hasonló spektrumot mutatott
és mindhárom DPS esetén a fenti cukormolekulák voltak kimutathatók. Colitóz
meghatározására prof. Stefan Oscarson-tól (Stocholm University, Inst. Org. Chem, Sweden)
ajándékba kapott standardot használtuk. A jellegzetes gázkromatogáfiás spektrumot a
következő ábra mutatja (15. ábra).
14. ábra: TLC: A mintákat Cellulose DC Plastikfolia vékony rétegen futtattuk. A neutrális cukrok elválasztására 6:4:3 arányú n-butanol-piridin-víz (a), az aminocukrok elválasztására 5:5:3:1 arányú piridin-etilacetát-víz ecetsav keverékét használtuk (b). Standardnak D-ribóz, D-mannóz, D-galaktóz, D-glukóz, D-galaktózamin és D-glukózamin keverékét használtuk. Az egyes cukrokat ezüstözéses eljárással festettük meg.
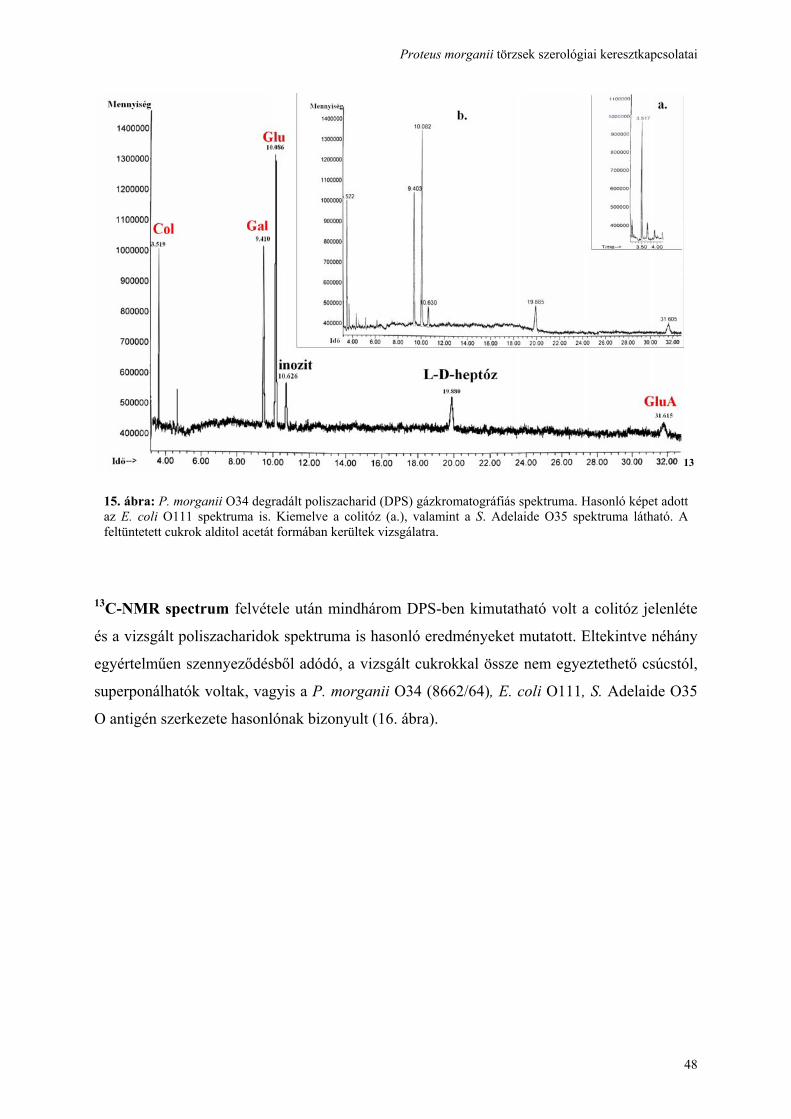
Proteus morganii törzsek szerológiai keresztkapcsolatai
48
13
13C-NMR spectrum felvétele után mindhárom DPS-ben kimutatható volt a colitóz jelenléte
és a vizsgált poliszacharidok spektruma is hasonló eredményeket mutatott. Eltekintve néhány
egyértelműen szennyeződésből adódó, a vizsgált cukrokkal össze nem egyeztethető csúcstól,
superponálhatók voltak, vagyis a P. morganii O34 (8662/64), E. coli O111, S. Adelaide O35
O antigén szerkezete hasonlónak bizonyult (16. ábra).
15. ábra: P. morganii O34 degradált poliszacharid (DPS) gázkromatográfiás spektruma. Hasonló képet adott az E. coli O111 spektruma is. Kiemelve a colitóz (a.), valamint a S. Adelaide O35 spektruma látható. A feltüntetett cukrok alditol acetát formában kerültek vizsgálatra.
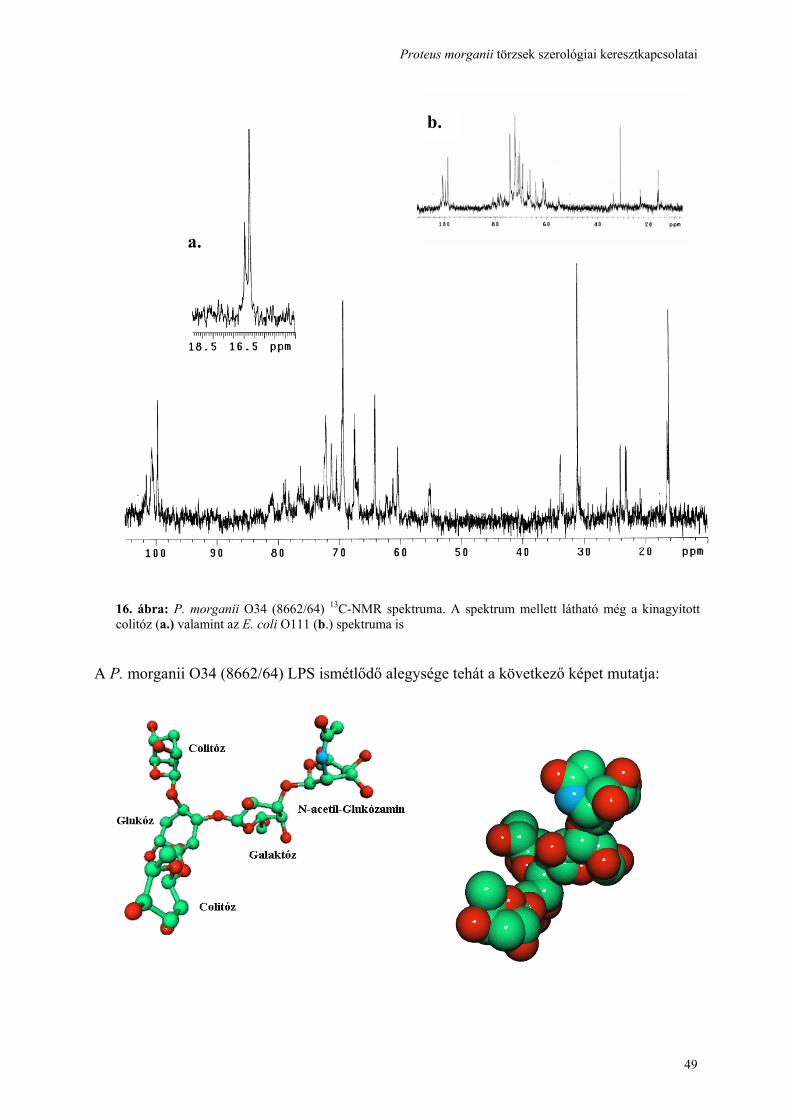
Proteus morganii törzsek szerológiai keresztkapcsolatai
49
A P. morganii O34 (8662/64) LPS ismétlődő alegysége tehát a következő képet mutatja:
a.
b.
16. ábra: P. morganii O34 (8662/64) 13C-NMR spektruma. A spektrum mellett látható még a kinagyított colitóz (a.) valamint az E. coli O111 (b.) spektruma is

Proteus morganii törzsek szerológiai keresztkapcsolatai
50
4.6. Genetikai hasonlóság detektálása a három vizsgált törzs rfb gén clusterében
Hasonló, vagy esetleg egyforma LPS szerkezet hátterében feltételezzük, hogy a
lipopoliszacharid felépítését és szintézisét irányító gének is hasonlóságot mutatnak. Bár idő és
megfelelő technikai háttér hiányában nem szándékoztunk a P. morgani O34 LPS
génszerkezetét feltárni és a bázissorrendet meghatározni, próba vizsgálatként az O antigén
gén cluster colitóz szintéziséért felelős gének egyikének, nevezetesen a manB DNS
szekvenciáját sikerült meghatároznunk, a primerek összeállításában a korábban ismert E. coli
O111 megfelelő génjét használva mintaként. A szintetizált génszakasz szekvenciasorrendjét a
génbankba helyeztük, mely EF051576 szám alatt érhető el. Az így nyert DNS szintetizátum és
szekvencia vizsgálata 70%-os megegyezést mutatott a P.
morganii O34 (8662/64) és S. Adelaide O35, valamint 69%-os
megegyezést mutatott a P. morganii O34 (8662/64) és E. coli
O111 DNS szekvenciái között. Az E. coli O111 és S. Adelaide
O35 DNS korábban tárgyalt génszakasza viszont 84,4%-os
homológiát mutat. Bár a P. morganii O34 teljes O antigénjének
szintézisét irányító gének szerkezete teljességgel még nem
ismert, feltételezzük, hogy az további homológiát mutathat az E.
coli O111 és S. Adelaide O35 O antigén szerkezetével (17.-20.
ábrák).
17. ábra: PCR vizsgálat után készült DNS szekvencia agaróz gélelektroforézises képe.
18. ábra: A P. morganii O34 O antigén manB génjének szekvenciája
* 10 * 40 * 60 AGCAGCAATTCTAGCCCAATAAGCGAAGATACTGGACTTAAAGAAATACAAAAACTAGCT * 80 * 100 * 120 GAGTCAAATAATTTTCTTATCGTGGGTACTGAAAAAAAAGGTGTGGTATATAGAAAAGTA * 140 * 160 * 180 TCCACCTTAGATAGCTATATTGACCACCTAATGTCTTACATAACACCGACTTCAATCAAA * 200 * 220 * 240 CCTTTAAAATTAAATGTTAATAGTGGAAACGGTGTAGCAGGTCATGTTATCAATGAATTA * 260 * 280 * 300 GAAAAAAGGG TTATTCATCTTAACATTCCCGTTACATTTGTAAAAATCAATAATATTCCT * 320 * 340 * 360 GATGGTTCATTCCCTAATGGAATTCCCAATCCTTTGCTACCAGAAAATCGTCAAGATACC * 380 * 400 * 413 ATTGATGCTGTTTTAAAATATCACGCTGATATTGGTATTGCATTTGATGGTGA
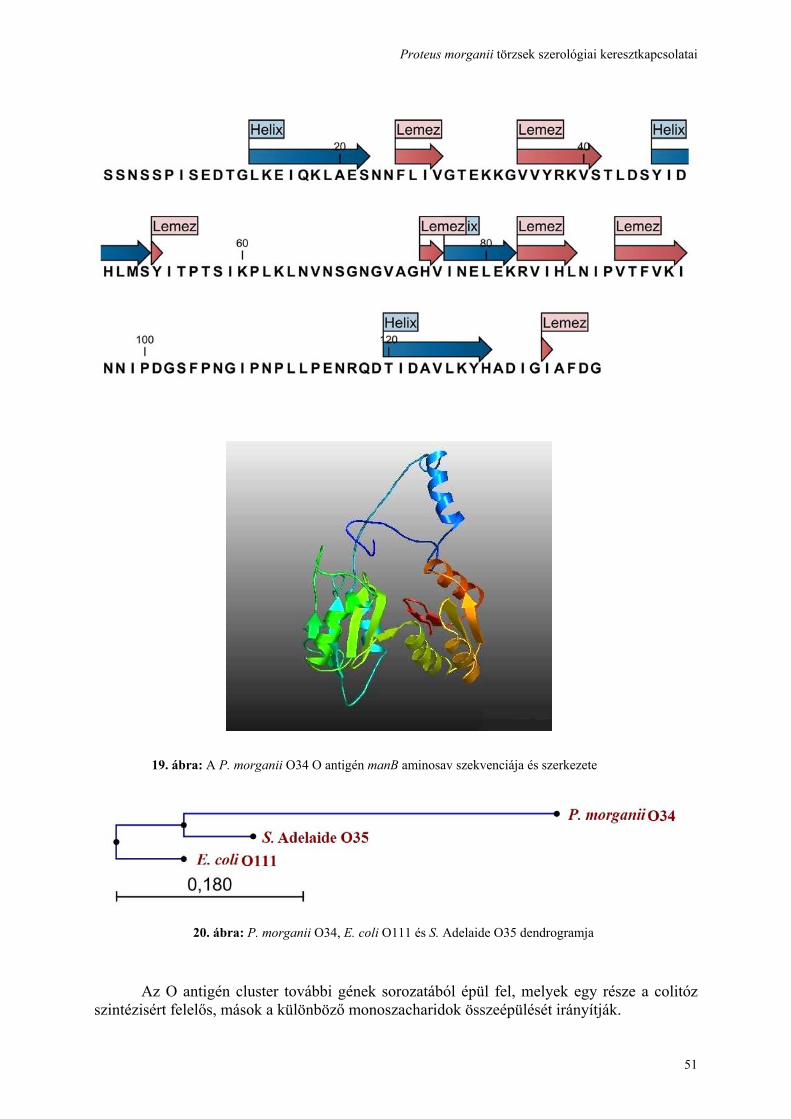
Proteus morganii törzsek szerológiai keresztkapcsolatai
51
Az O antigén cluster további gének sorozatából épül fel, melyek egy része a colitóz
szintézisért felelős, mások a különböző monoszacharidok összeépülését irányítják.
19. ábra: A P. morganii O34 O antigén manB aminosav szekvenciája és szerkezete
20. ábra: P. morganii O34, E. coli O111 és S. Adelaide O35 dendrogramja

Proteus morganii törzsek szerológiai keresztkapcsolatai
52
4.7. A hőlabilis (fehérje) antigén identifikálása, szerepének meghatározása és az
LPS-el való kapcsolatának tisztázása
4.7.1. A külső membrán fehérjék kapilláris elektroforézises profilja
Első lépésben a vizsgálatunk tárgyát képező három baktérium külső membrán fehérjéit
akartuk jellemezni, melyet mind a konvencionális kapilláris elektroforesis, mind a chip
technológia segítségével elvégeztünk. A kapilláris elektroforézis során készült OMP képet a
21. ábra mutatja. Minden vizsgálatot 10 percen belül végeztünk el. A belső standardként
használt benzoesav a második percben vált láthatóvá. A vizsgált baktériumok külső membrán
fehérjéinek kapilláris elektroforézises vizsgálata során az első csúcsokat az 5,2 percben
lehetett regisztrálni.
Mindhárom kapilláris elektroforetikus OMP profilt a 35-50 kDa. közötti csúcsok dominálták,
melyek jól korreláltak a mikrochip alapú elektroferogrammokkal. A P. morganii O34
(8662/64) két jellegzetes csúcsot adott (21/b. ábra). Az E. coli O111 több karakterisztikus
csúcsot mutatott, de a 36, 40, 45 kDa. fehérjék domináló jellege itt is kimutatható (21/a. ábra),
míg a S. Adelaide O35 esetén a több jellegzetes csúcs mellett a közös domináló és a 39 kDa
nagyságú fehérje is kimutatható (21/c. ábra).
4.7.2. Az OMP chip alapú elektroforesise
A chip alapú electroferogramokat az 22. ábrán mutatjuk be. A chip alapú vizsgálatok
esetében az elektroforetikus mérés mindössze 45 másodpercig tartott. Az alacsony és magas
mólsúlyú standardok körülbelül 20.-43. másodperc között jelentek meg. Minden mérést
három alkalommal ismételtünk meg. A mikrochip alapú elektrophoretikus képen mindhárom
törzs esetében a 35-50 kDa molekulasúlynak megfelelően találtuk a domináló fehérje
csúcsokat.
Az irodalmi adatoknak megfelelően a Gram-negatív baktériumok ebben a
molekulasúly tartományban tartalmazzák a jellegzetes és domináló külső membrán fehérjéket.
A P. morganii O34 (8662/64) membránprofilja két major külső membrán fehérjét tartalmaz a
36 és 41 kDa molekulasúly tartományban (22/b ábra). Ugyanebben a régióban az E. coli O111
membránprofilja három különböző fehérjét mutat a 36, 41 és 45 kDa-nak megfelelően,
azonban a 36 kDa fehérje mennyisége sokkal nagyobb a másik kettőhöz viszonyítva (22/a
ábra). A S. Adelaide O35 külsőmembrán fehérje képét a 39 kDa nagyságú fehérje dominálja.
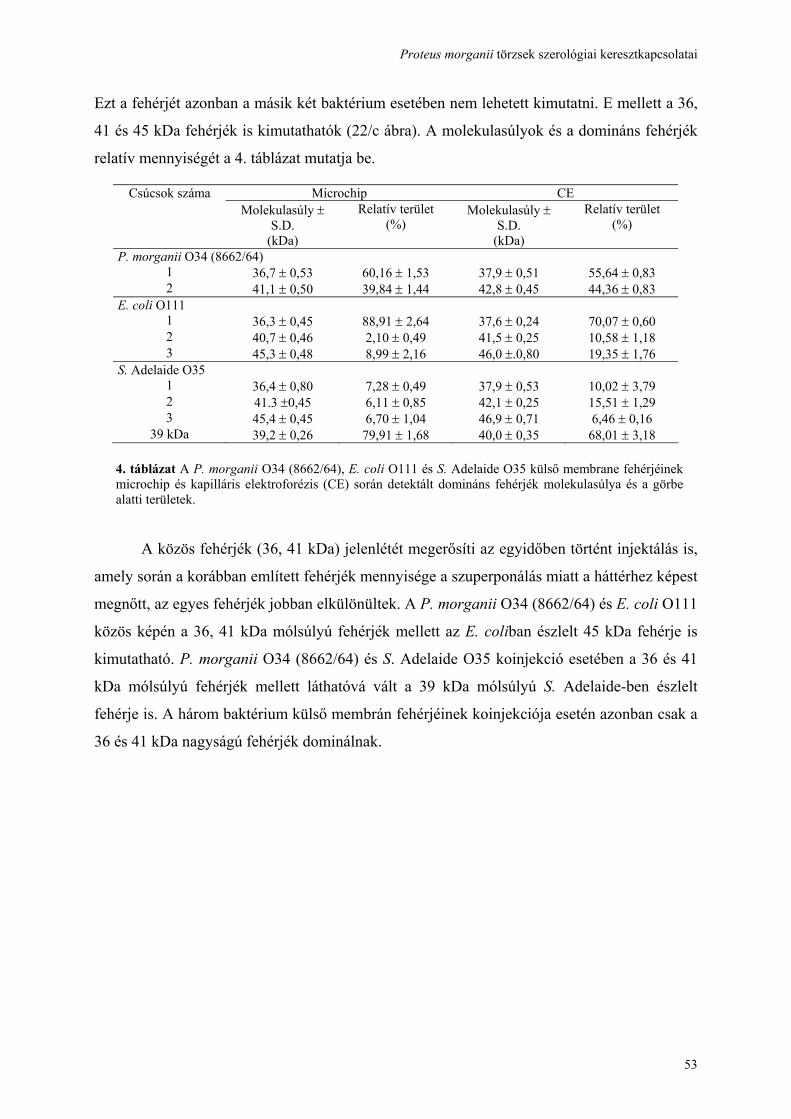
Proteus morganii törzsek szerológiai keresztkapcsolatai
53
4. táblázat A P. morganii O34 (8662/64), E. coli O111 és S. Adelaide O35 külső membrane fehérjéinek microchip és kapilláris elektroforézis (CE) során detektált domináns fehérjék molekulasúlya és a görbe alatti területek.
Ezt a fehérjét azonban a másik két baktérium esetében nem lehetett kimutatni. E mellett a 36,
41 és 45 kDa fehérjék is kimutathatók (22/c ábra). A molekulasúlyok és a domináns fehérjék
relatív mennyiségét a 4. táblázat mutatja be.
A közös fehérjék (36, 41 kDa) jelenlétét megerősíti az egyidőben történt injektálás is,
amely során a korábban említett fehérjék mennyisége a szuperponálás miatt a háttérhez képest
megnőtt, az egyes fehérjék jobban elkülönültek. A P. morganii O34 (8662/64) és E. coli O111
közös képén a 36, 41 kDa mólsúlyú fehérjék mellett az E. coliban észlelt 45 kDa fehérje is
kimutatható. P. morganii O34 (8662/64) és S. Adelaide O35 koinjekció esetében a 36 és 41
kDa mólsúlyú fehérjék mellett láthatóvá vált a 39 kDa mólsúlyú S. Adelaide-ben észlelt
fehérje is. A három baktérium külső membrán fehérjéinek koinjekciója esetén azonban csak a
36 és 41 kDa nagyságú fehérjék dominálnak.
Csúcsok száma Microchip CE Molekulasúly ±
S.D. (kDa)
Relatív terület (%)
Molekulasúly ± S.D.
(kDa)
Relatív terület (%)
P. morganii O34 (8662/64) 1 36,7 ± 0,53 60,16 ± 1,53 37,9 ± 0,51 55,64 ± 0,83 2 41,1 ± 0,50 39,84 ± 1,44 42,8 ± 0,45 44,36 ± 0,83
E. coli O111 1 36,3 ± 0,45 88,91 ± 2,64 37,6 ± 0,24 70,07 ± 0,60 2 40,7 ± 0,46 2,10 ± 0,49 41,5 ± 0,25 10,58 ± 1,18 3 45,3 ± 0,48 8,99 ± 2,16 46,0 ±.0,80 19,35 ± 1,76
S. Adelaide O35 1 36,4 ± 0,80 7,28 ± 0,49 37,9 ± 0,53 10,02 ± 3,79 2 41.3 ±0,45 6,11 ± 0,85 42,1 ± 0,25 15,51 ± 1,29 3 45,4 ± 0,45 6,70 ± 1,04 46,9 ± 0,71 6,46 ± 0,16
39 kDa 39,2 ± 0,26 79,91 ± 1,68 40,0 ± 0,35 68,01 ± 3,18
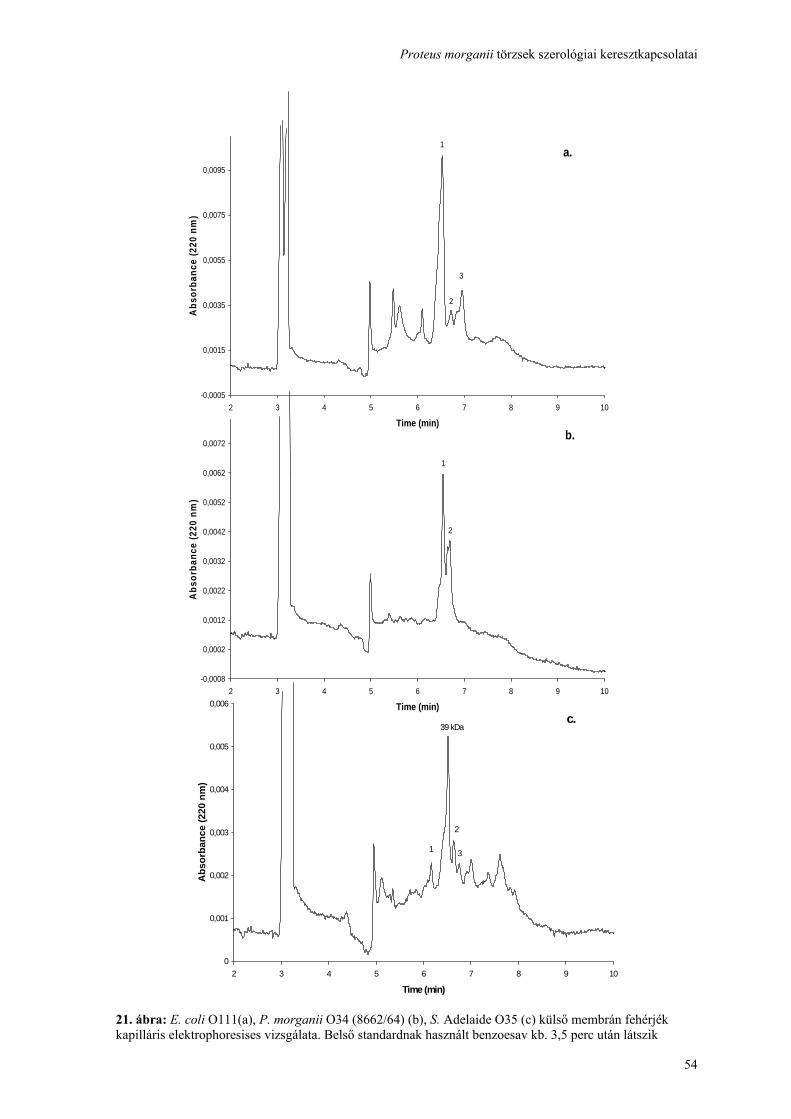
Proteus morganii törzsek szerológiai keresztkapcsolatai
54
21. ábra: E. coli O111(a), P. morganii O34 (8662/64) (b), S. Adelaide O35 (c) külső membrán fehérjék kapilláris elektrophoresises vizsgálata. Belső standardnak használt benzoesav kb. 3,5 perc után látszik
-0,0008
0,0002
0,0012
0,0022
0,0032
0,0042
0,0052
0,0062
0,0072
2 3 4 5 6 7 8 9 10
Time (min)
Abs
orba
nce
(220
nm
)
1
2
b.
-0,0005
0,0015
0,0035
0,0055
0,0075
0,0095
2 3 4 5 6 7 8 9 10
Time (min)
Abs
orba
nce
(220
nm
)
1
2
3
a.
0
0,001
0,002
0,003
0,004
0,005
0,006
2 3 4 5 6 7 8 9 10
Time (min)
Abs
orba
nce
(220
nm
)
1
2
3
39 kDac.
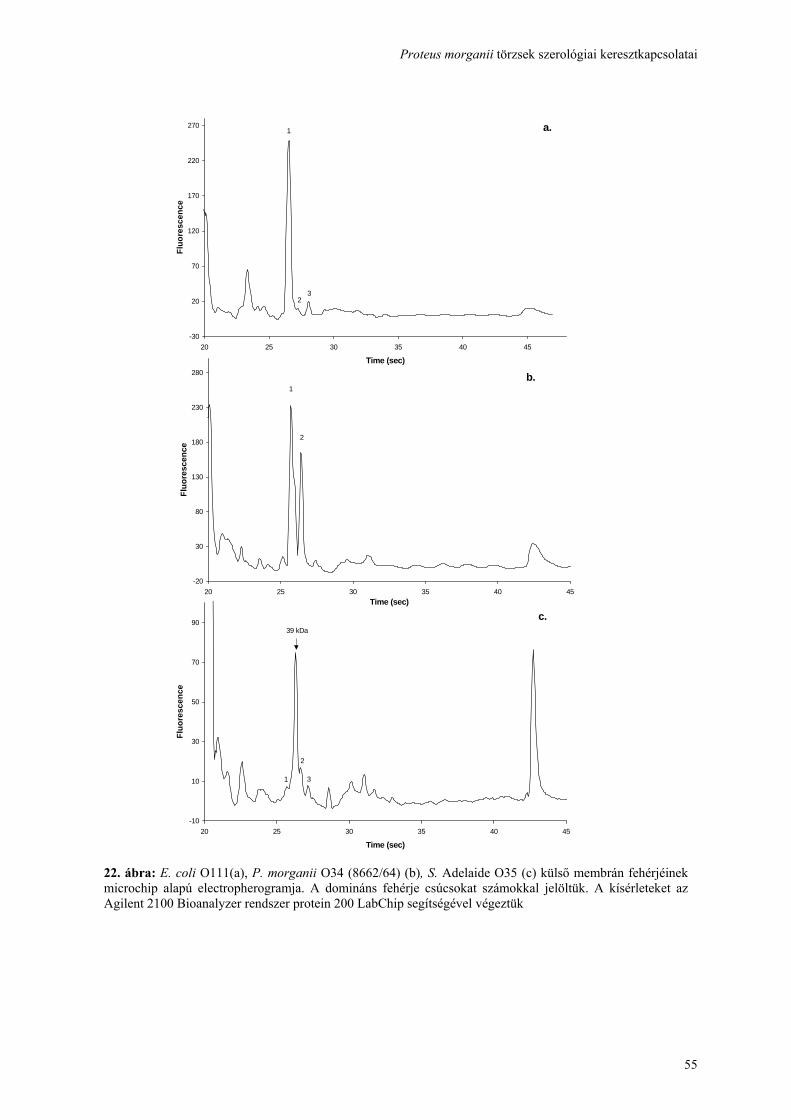
Proteus morganii törzsek szerológiai keresztkapcsolatai
55
-30
20
70
120
170
220
270
20 25 30 35 40 45
Time (sec)
Fluo
resc
ence
a. 1
23
-20
30
80
130
180
230
280
20 25 30 35 40 45Time (sec)
Fluo
resc
ence
1
2
b.
-10
10
30
50
70
90
20 25 30 35 40 45
Time (sec)
Fluo
resc
ence
1
2
3
39 kDa
c.
22. ábra: E. coli O111(a), P. morganii O34 (8662/64) (b), S. Adelaide O35 (c) külső membrán fehérjéinek microchip alapú electropherogramja. A domináns fehérje csúcsokat számokkal jelöltük. A kísérleteket az Agilent 2100 Bioanalyzer rendszer protein 200 LabChip segítségével végeztük
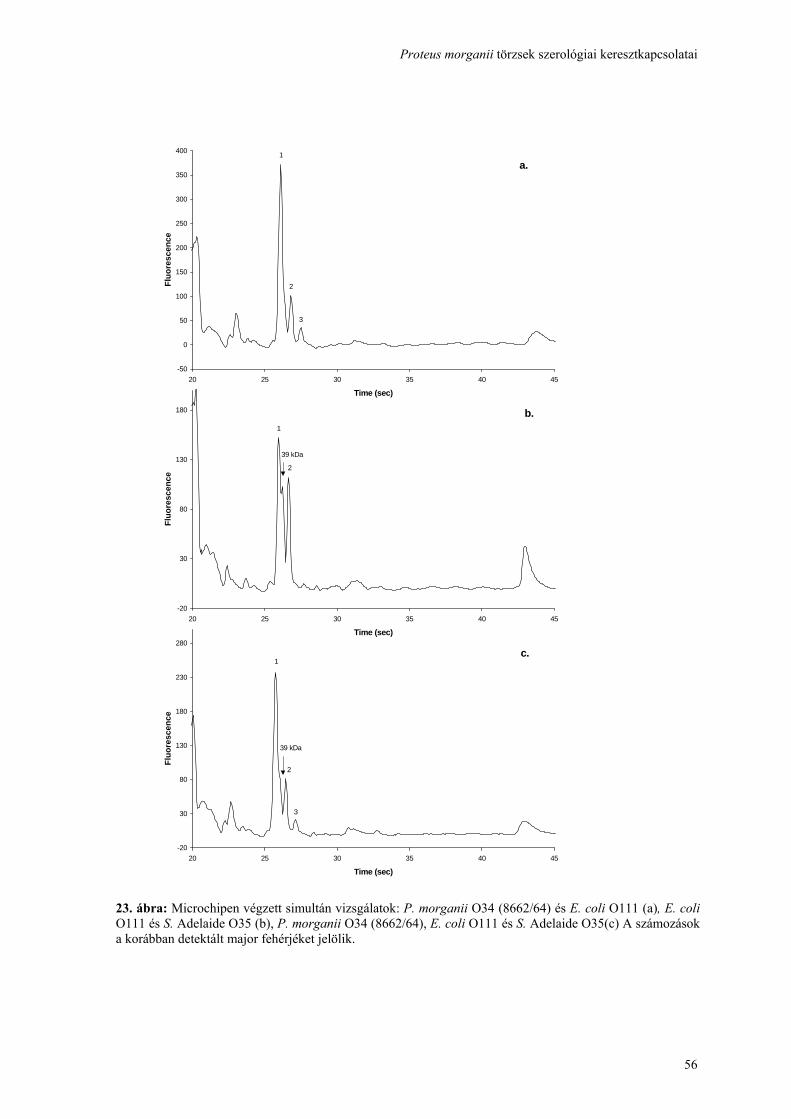
Proteus morganii törzsek szerológiai keresztkapcsolatai
56
23. ábra: Microchipen végzett simultán vizsgálatok: P. morganii O34 (8662/64) és E. coli O111 (a), E. coli O111 és S. Adelaide O35 (b), P. morganii O34 (8662/64), E. coli O111 és S. Adelaide O35(c) A számozások a korábban detektált major fehérjéket jelölik.
-50
0
50
100
150
200
250
300
350
400
20 25 30 35 40 45
Time (sec)
Fluo
resc
ence
1
2
3
a.
-20
30
80
130
180
20 25 30 35 40 45
Time (sec)
Fluo
resc
ence
b. 1
2
39 kDa
-20
30
80
130
180
230
280
20 25 30 35 40 45
Time (sec)
Fluo
resc
ence
1
2
3
39 kDa
c.
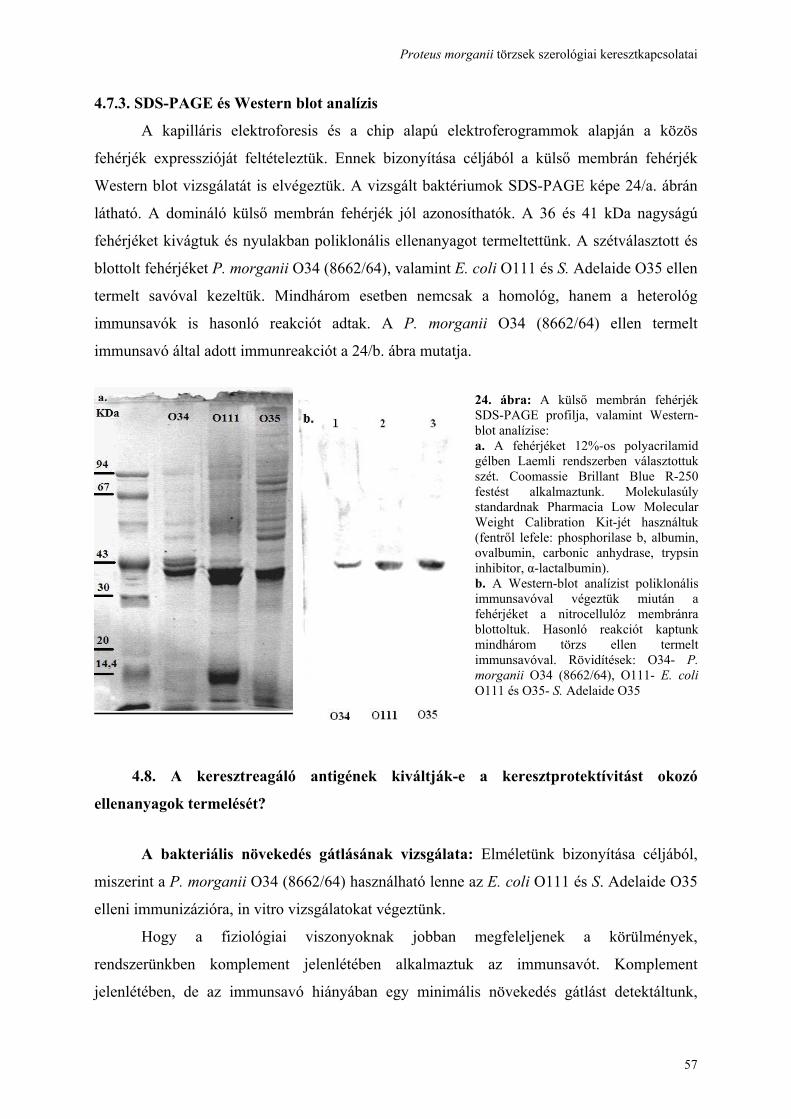
Proteus morganii törzsek szerológiai keresztkapcsolatai
57
24. ábra: A külső membrán fehérjék SDS-PAGE profilja, valamint Western-blot analízise: a. A fehérjéket 12%-os polyacrilamid gélben Laemli rendszerben választottuk szét. Coomassie Brillant Blue R-250 festést alkalmaztunk. Molekulasúly standardnak Pharmacia Low Molecular Weight Calibration Kit-jét használtuk (fentről lefele: phosphorilase b, albumin, ovalbumin, carbonic anhydrase, trypsin inhibitor, α-lactalbumin). b. A Western-blot analízist poliklonális immunsavóval végeztük miután a fehérjéket a nitrocellulóz membránra blottoltuk. Hasonló reakciót kaptunk mindhárom törzs ellen termelt immunsavóval. Rövidítések: O34- P. morganii O34 (8662/64), O111- E. coli O111 és O35- S. Adelaide O35
4.7.3. SDS-PAGE és Western blot analízis
A kapilláris elektroforesis és a chip alapú elektroferogrammok alapján a közös
fehérjék expresszióját feltételeztük. Ennek bizonyítása céljából a külső membrán fehérjék
Western blot vizsgálatát is elvégeztük. A vizsgált baktériumok SDS-PAGE képe 24/a. ábrán
látható. A domináló külső membrán fehérjék jól azonosíthatók. A 36 és 41 kDa nagyságú
fehérjéket kivágtuk és nyulakban poliklonális ellenanyagot termeltettünk. A szétválasztott és
blottolt fehérjéket P. morganii O34 (8662/64), valamint E. coli O111 és S. Adelaide O35 ellen
termelt savóval kezeltük. Mindhárom esetben nemcsak a homológ, hanem a heterológ
immunsavók is hasonló reakciót adtak. A P. morganii O34 (8662/64) ellen termelt
immunsavó által adott immunreakciót a 24/b. ábra mutatja.
4.8. A keresztreagáló antigének kiváltják-e a keresztprotektívitást okozó
ellenanyagok termelését?
A bakteriális növekedés gátlásának vizsgálata: Elméletünk bizonyítása céljából,
miszerint a P. morganii O34 (8662/64) használható lenne az E. coli O111 és S. Adelaide O35
elleni immunizázióra, in vitro vizsgálatokat végeztünk.
Hogy a fiziológiai viszonyoknak jobban megfeleljenek a körülmények,
rendszerünkben komplement jelenlétében alkalmaztuk az immunsavót. Komplement
jelenlétében, de az immunsavó hiányában egy minimális növekedés gátlást detektáltunk,
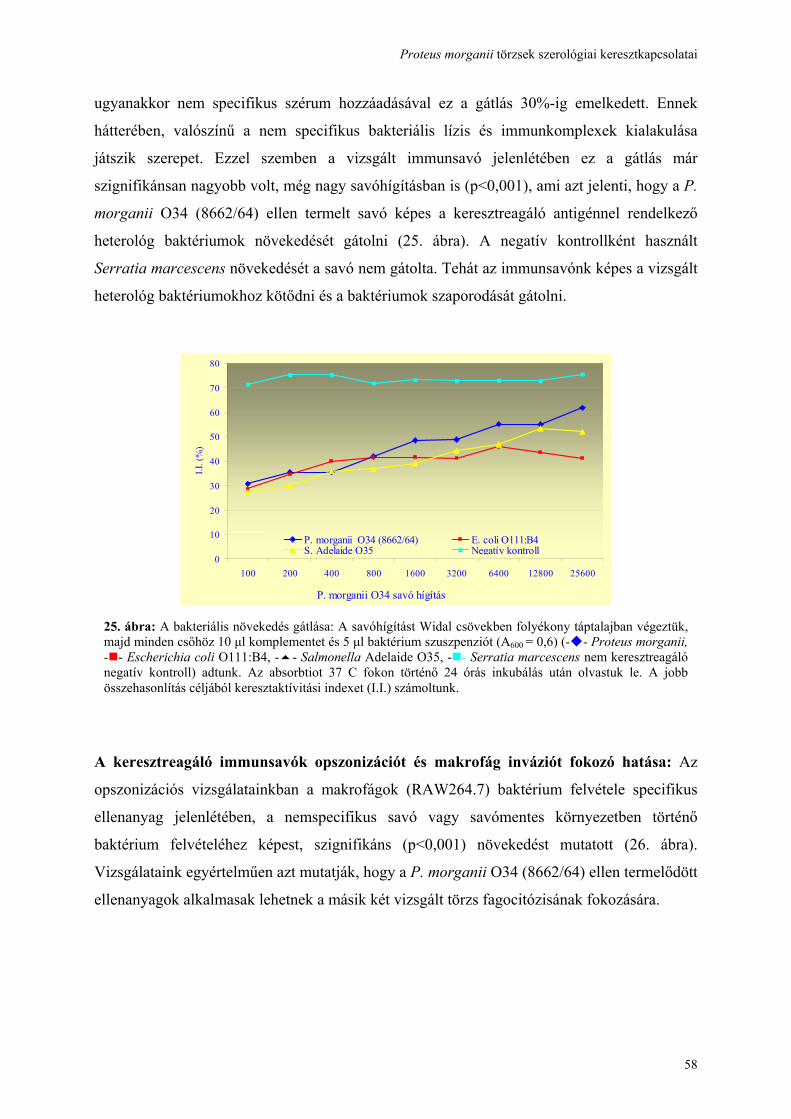
Proteus morganii törzsek szerológiai keresztkapcsolatai
58
ugyanakkor nem specifikus szérum hozzáadásával ez a gátlás 30%-ig emelkedett. Ennek
hátterében, valószínű a nem specifikus bakteriális lízis és immunkomplexek kialakulása
játszik szerepet. Ezzel szemben a vizsgált immunsavó jelenlétében ez a gátlás már
szignifikánsan nagyobb volt, még nagy savóhígításban is (p<0,001), ami azt jelenti, hogy a P.
morganii O34 (8662/64) ellen termelt savó képes a keresztreagáló antigénnel rendelkező
heterológ baktériumok növekedését gátolni (25. ábra). A negatív kontrollként használt
Serratia marcescens növekedését a savó nem gátolta. Tehát az immunsavónk képes a vizsgált
heterológ baktériumokhoz kötődni és a baktériumok szaporodását gátolni.
0
10
20
30
40
50
60
70
80
100 200 400 800 1600 3200 6400 12800 25600
P. morganii O34 savó hígítás
I.I. (
%)
P. morganii O34 (8662/64) E. coli O111:B4S. Adelaide O35 Negatív kontroll
A keresztreagáló immunsavók opszonizációt és makrofág inváziót fokozó hatása: Az
opszonizációs vizsgálatainkban a makrofágok (RAW264.7) baktérium felvétele specifikus
ellenanyag jelenlétében, a nemspecifikus savó vagy savómentes környezetben történő
baktérium felvételéhez képest, szignifikáns (p<0,001) növekedést mutatott (26. ábra).
Vizsgálataink egyértelműen azt mutatják, hogy a P. morganii O34 (8662/64) ellen termelődött
ellenanyagok alkalmasak lehetnek a másik két vizsgált törzs fagocitózisának fokozására.
25. ábra: A bakteriális növekedés gátlása: A savóhígítást Widal csövekben folyékony táptalajban végeztük, majd minden csőhöz 10 μl komplementet és 5 μl baktérium szuszpenziót (A600 = 0,6) (- - Proteus morganii, - - Escherichia coli O111:B4, - - Salmonella Adelaide O35, - - Serratia marcescens nem keresztreagáló negatív kontroll) adtunk. Az absorbtiot 37 C fokon történő 24 órás inkubálás után olvastuk le. A jobb összehasonlítás céljából keresztaktívitási indexet (I.I.) számoltunk.

Proteus morganii törzsek szerológiai keresztkapcsolatai
59
a. b. c.
0
20
40
60
80
100
120
savó n
élkül
norm
.O34
O111
O35
savó n
élkül
norm
.O34
O111
O35
savó né
lkül
norm
.O34
O111
O35
nyúlsavók
CFU
4.9. A további kutatási lehetőségek feltárása
A fentiekben bemutatott eredmények a már eddig is ismert szempontok mellett felhívják a
figyelmet azokra a további lehetőségekre, amelyeket a bakteriális antigének közötti
keresztkapcsolatok vizsgálata nyújthat. Rámutatnak arra, hogy ezek a kapcsolatok nem csak a
diagnosztika területén hasznosíthatók, hanem keresztprotektívitás kiváltása révén lehetőséget
nyújthatnak vakcinológiai alkalmazásra.
A korábban tárgyalt törzsek mellett még további hasonló keresztkapcsolatok kerültek
előtérbe, melyek előzetes vizsgálata után azok alaposabb ellemzése, megismerése a későbbi
kutatások tárgyát képezheti. A múltbéli a vizsgálatok eredményei elsősorban szerológián
alapszanak.
Az általunk eddig végzett szerológiai (csőagglutináció, haemagglutináció, dot blot,
ELISA, növekedés gátlás), valamint szerkezeti vizsgálatokon alapuló eredmények arra
utalnak, hogy a P. morganii O43:12 (Louvain) - Y. enterocolitica O9 - E. coli O157:H7 - S.
Urbana O30 - Brucella abortus, valamint a P. morganii O9 (352/56) - E. coli O21;K20 - S.
Inverness O38 - Salmonella O44 - Citrobacter freundi O14, illetve a P. morganii O29
(1594) - E. coli O112ac;K68 - S. dysenteriae 2 törzsek között szerológiai hasonlóság van.
Ezeknek a kapcsolatoknak a további vizsgálata, finom szerkezeti elemzése ugyan meghaladja
ezen disszertáció kereteit, de további kutatásaink központi célkitűzései közé tartozna.
26. ábra: Opsonizáció és invázió vizsgálata: RAW264.7 makrofág sejtkultúrához inaktivált normál nyúlszérummal, vagy immunsavóval illetve savó nélküli P. morganii O34 (8662/64) (a.), E. coli O111 (b.), S. Adelaide O35 (c.) baktérium szuszpenziót adtunk. Az internalizált baktériumok számát a makrofágok lízise után a telepképzők mennyiségének meg-határozásával állapítottuk meg. Rövidítések: norm – normál nyúlszérum; O34 - P. morganii O34 és immunszérum; O111 - E. coli O111 és immunszérum; O35 - S. Adelaide O35 és immunszérum keveréke.

Proteus morganii törzsek szerológiai keresztkapcsolatai
60
5. MEGBESZÉLÉS
A különböző szerológiai vizsgálatokra, antigének, illetve antitestek kimutatására,
keresztreakciók bizonyítására, ma már egyik leggyakrabban használt módszer az ELISA,
amely érzékeny, reprodukálható, viszonylag gyors és olcsó. A fehérje természetű antigének
esetében, mivel azok jó antigének, könnyen kivitelezhető és kipróbált rendszerek állnak
rendelkezésünkre. Nem fehérje természetű antigének esetén, mint az LPS több probléma is
felmerül: a rossz kötődés a lemezhez, az antitestek nem specifikus kötődése a lemez falához
vagy a blokkolásra használt anyaghoz.
Az első probléma a lemez poly-L-lysines előkezelésével megoldódott [Engvall 1972].
Eredeti leírás szerint a reakció láthatóvá tételéhez peroxidázzal konjugált Protein A-t
használtak, de a pozitív reakciók esetében kapott legnagyobb exktinkció nem volt elég nagy
(saját kísérleteinkben 0,751). Ha blokkolás nélkül használtuk a peroxidázzal konjugált anti-
IgG-t, a háttér optikai denzitása túl magas volt (kísérleteinkben 0,946).
A nem specifikus kötőhelyeket éppen ezért blokkolni kell, erre a célra általában
valamilyen, a rendszer szempontjából semleges hatású blokkoló anyagot használnak.
Legelterjedtebb a 0,5% BSA vagy 0,5% kazein [Freudenberg 1989, Trautmann 1996]. BSA vagy
kazein 1:100 hígítását használva (legjobb hígítás), a kapott eredményeink a korábbiakhoz
képest javulnak. Sajnálatos módon ezek a fehérjék aspecifikusan kötődnek az LPS-hez is,
csökkentve az antigén-antitest reakcióban résztvevő epitópok számát, több álpozitív
eredményt produkálnak, és ezáltal befolyásolják a rendszer érzékenységét is. Ezen ismeretek
birtokában született az ötlet, hogy keressünk olyan blokkoló anyagot, ami kevésbé
befolyásolja az adott reakciókat. Különböző állatokból származó savókat kipróbálva, azok
hatékonyabbnak bizonyultak, mint a hagyományosan használt blokkoló anyagok.
A kecskesavóval történő blokkolás esetében sokkal kisebb mértékű az aspecifikus
kötődés és az eredmények is ennek megfelelően jobbak, mintha a BSA vagy kazein bármely
hígítását alkalmaztuk volna. A kecskesavó használata esetén a különböző vizsgálat
sorozatokban minden esetben szignifikánsan jobb eredményeket adtak (p<0,001). Azonban
megmondani, hogy a savó mely komponense felelős a hatékonyabb blokkolásért nem tudtuk,
de több egyedből származó savó összehasonlítása azt mutatja, hogy a különböző eredetű
kecskesavók között nincs lényeges különbség. Bizonyítandó a kecskesavó jobb hatékonyságát
azon túl, hogy összehasonlítottuk a különböző blokkoló anyagokat, kereszt-blokkolásokat is
végeztünk, azaz a kazein vagy BSA blokkolás után kecskesavót használtunk és fordítva. Ha a

Proteus morganii törzsek szerológiai keresztkapcsolatai
61
kecskesavót másodszori blokkolásra alkalmaztuk az eredmények szignifikánsan javultak, ami
a savó jobb befedő tulajdonságának tulajdonítható. Fordított esetben, amikor a BSA vagy
kazein volt a második blokkoló anyag az eredmények szignifikánsan romlottak, mert ezek a
fehérjék elfedik a LPS reakcióképes helyeit.
Vizsgálataink azt bizonyítják, hogy blokkolásra a kecskesavó használata jobb, nem
csak az LPS molekula, mint egész, hanem a kis antigenitásbeli eltérést mutató antigének
kimutatásában is, hiszen a S. sonnei O oldallácot és core-t érintő csak 1-1 cukormolekula
eltérést mutató mutánsait egyértelműen azonosítani tudtuk. Ezért alkalmasabbnak bizonyult
az LPS keresztreakciók analízisére is. Ez a megállapítás érvényes általában minden fajta LPS-
re és hidrolízis utáni származékaira, így S, R, abszolút R törzsek lipopoliszacharidja, DPS-e és
Lipid-A-ja esetén is. Ez a módszer kiválóan alkalmas különböző rokonsági fokú antigének
kimutatására, akár hígítási sorozat nélkül is, és elég érzékenynek bizonyul az antigének
közötti kismértékű eltérések vizsgálatára is. Ugyanakkor a reakciók ismétlése során nem
tapasztalunk szignifikáns eltérést, ami szintén a rendszer stabilitását, megbízhatóságát
bizonyítja. Tapasztalataink megerősítésére klinikai mintákból származó savókat használtunk.
Így nagyszámú és változatos betegpopulációból származó minták közötti különbségek
megfelelő támpontot mutatnak a megbízhatóság tesztelésére.
Célunk elérésére lehetőség kínálkozott appendicitises betegekből származó
histológiailag, tehát a gyulladás mértékében is eltérő, így várhatóan különböző anti-LPS titerű
szérumok felhasználásával.
Az acut appendicitis kialakításában egyaránt fontos szerepe van az obstrukciónak és
a fertőzésnek is [Pieper 1982/a, Pieper 1982/b]. A gyulladt féregnyúlvány tenyésztése során
gyakran aerob (E. coli) és anaerob (B. fragilis) baktériumokat egyaránt sikerül kitenyészteni.
Ezen baktériumok szerepe azonban a gyulladás kialakításában mindezidáig még nem
tisztázott. A szerológiai vizsgálatok a gyulladás kiváltásában játszott szerepüket erősítik
[Pieper 1982].
A vizsgálatok elkezdése előtt azonban a baktériumok, szerotípusok nagy száma miatt
tisztázni kellene mi a gyulladás kiváltó oka. Ez sajnos a legtöbb esetben lehetetlen, ezért a
szerológiai módszerekhez olyan, az egyébként normál bélflórához tartozó baktérium törzseket
lehet használni, amelyek hasonló körülmények között viszonylag gyakrabban izolálhatók. Az
ELISA [Lindberg 1979] a történések követésére releváns szerológiai módszernek bizonyulhat.
A normál bélflóra E. coli szerotípusai szerteágazóak és folyamatosan változnak [Burkitt
1971, Leigh 1974, Ehlag 1986]. Az immunválasz detektálására használt szerotípusokat úgy
válogattuk, hogy azok a normál bélflórát reprezentálják. ELISA vizsgálatainkhoz E. coli O21,

Proteus morganii törzsek szerológiai keresztkapcsolatai
62
O22, O33, O61, O68 [Bettelheim 1972] és B. fragilist használtunk. Mivel a bél normál flóráját
többnyire Gram-negatív baktériumok alkotják, ezért un. „általános” antigénként az abszolút R
mutáns S. sonnei Re4350-t választottuk. Ezzel az antigénnel az O szerotípustól függetlenül
detektálhatók a legkülönbözőbb Gram-negatív baktériumok által indukált ellenanyagok is,
mivel a Lipid-A és a belső core konzervatív régiót (KDO) tartalmazó lipooligoszacharid
(LOS) keresztreagál a többi lipopoliszachariddal is.
Vizsgálataink során az E. coli és B. fragilis-el szemben termelődött ellenanyagok titere
a betegség súlyosságával párhuzamosan emelkedett. A negatív kontroll és appendicitisben
szenvedő betegek savói között a különbség szignifikáns volt (p<0,001). Amíg magas E. coli
ellenes IgG szintet az esetek 50-93%-ban lehetett detektálni, addig emelkedett IgM szintet az
esetek 5-60%-ban észleltünk. Mindezek után úgy gondoljuk, hogy a gyulladás kiváltásában
nem csak egy baktériumtörzs vagy species játszik szerepet, hanem több, a normál flórához
tartozó E. coli és B. fragilis törzs szinergén hatásáról beszélhetünk. Intra-abdominalis
sepsisben ilyen szinergén hatást E. coli és B. fragilis között, kísérletes körülmények mellett,
korábban Onderdonk és munkatársai is leírtak [Onderdonk 1976].
Feltételezésünk szerint a B. fragilis és E. coli közötti szinergizmus elsősorban a
betegség, a gyulladás megjelenésében játszik fontos szerepet, de a későbbi súlyosabb
kórformák és a perforatio kialakulásában már az E. coli szerepe kiemelkedő, míg az anaerob
B. fragilis szerepe elhanyagolhatóbb, ugyanis kevesebb esetben találtunk magasabb B. fragilis
elleni IgM szintet. Lehetséges, hogy az endotoxinok az appendix falának sérülése után
kerülnek a keringésbe és aktiválják az immunrendszert, mivel a gyulladt bélfal sokkal
áteresztőbb a baktériumok és bakteriális termékek tekintetében, mint a normál, ép bélfal.
Az a tény, hogy a magasabb ellenanyag szinteket elsősorban a súlyosabb gyulladásos
folyamatot mutató phlegmonosus és gangrenás formákban észleltünk, azt mutatja, hogy itt
súlyosabb a szervkárosodás és valószínű hosszabb idő is eltelt a betegség inicializálódása és a
mintavétel között. Azokra az esetekre, amikor nem detektálható ellenanyagválasz, két
lehetséges magyarázatot adhatunk, vagy a szervezet immunválasz készsége csökkent, zavart,
vagy pedig nem a megfelelő antigént illetve ellenanyagot használtuk az immunreakció
lemérésére.
Mindezek után úgy gondoljuk, hogy módosított ELISA tesztünk a fenti célokra jól
alkalmazható és hasonló vizsgálatok elvégzéséhez a módszert ajánlani tudjuk. Ismeretlen
lipopoliszacharid detektálására pedig az abszolút R mutáns, mint „általános” antigén szintén
jól használható, bár kevésbé specifikus. Amennyiben IgG vagy IgA és IgM egyidejű
detektálása szükséges, teszteléshez kiváló az említett immunglobulin keveréke ellen termelt

Proteus morganii törzsek szerológiai keresztkapcsolatai
63
anti-savó konjugátum alkalmazása, mivel az így kapott eredmények jól korrelálnak a
gyulladás súlyosságával [Péterfi 2006].
Az ELISA módszer beállítása és klinikai tesztelése után kezdtük el a vizsgálataink
alapját képező keresztreakciók tanulmányozását.
Vizsgálatainkat először immunológiai módszerekkel kezdtük, majd kémiai, genetikai
illetve preparatív eljárásokkal folytattuk.
Az LPS három fő alkotórészből épül össze: a lipid (Lipid-A) részével kapcsolódik a
külső membránhoz, és felel az LPS endotoxicitásáért, az oligoszacharid core régió és az O
specifikus (O-antigén) poliszacharid rész, mely a baktérium immunspecificitását határozza
meg. Az endotoxin ellen termelt ellenanyagokat általában csőagglutinációs módszerrel a
Widal reakció felhasználásával, passzív haemagglutinációval vagy ELISA-val mutatják ki [Coughlin 1987].
A Widal reakció nem érzékeny és az eredményeket nem specifikus reakciók, mint pl. a
spontán agglutináció, gyakran zavarják. Mind a passzív haemagglutináció, mind a
csőagglutináció hátránya, hogy elsősorban IgM detektálására alkalmas (kb. 10 vagy 100x
érzékenyebb IgM, mint IgG kimutatására) [Carlsson 1972]. Az IgG sokkal specifikusabban
kapcsolódik bizonyos epitópokhoz, így kevesebb nem specifikus reakció keletkezik.
Immunizálás után hosszabb ideig magas szinten jelen van a keringésben, így tovább biztosít
védelmet a betolakodó baktériumokkal szemben. A korábban csőagglutináció segítségével
leírt keresztkapcsolatok bizonyítására ezért elsősorban IgG detektálására alkalmas
módszereket, dot blot, ELISA és Western blot, használtunk.
Eredményeink azt mutatják, hogy a P. morganii O34 (8662/64) hasonló LPS-t hordoz, mint
az E. coli O111 valamint a S. Adelaide O35. Csőagglutinációs vizsgálatokkal sikerült
reprodukálnunk a Rauss és Vörös által leírt agglutinációs eredményeket. Az alaposabb
vizsgálatokra érzékenyebb módszereket alkalmaztunk. A haemagglutinációnál észlelt
agglutinációs titerkülönbségeket az egyes LPS-ek eltérő Lipid-A és DPS arányával
magyarázhatjuk, ami P. morganii O34 (8662/64) esetében 59%, E. coli O111 89%, míg a S.
Adelaide O35 LPS hidrolizátuma 72% DPS-t mutatott. ELISA-val nem tudtunk különbséget
kimutatni a vizsgált három LPS között. Kereszt kimerítések után továbbra sem lehetett eltérést
detektálni a három LPS között. A Lipid-A zavaró hatásának kiszűrése céljából a
vizsgálatainkat DPS felhasználásával is elvégeztük. Az alacsonyabb mért optikai denzitás a
konzervatív core régió keresztkapcsolatokban való elenyésző szerepét támasztja alá, de a
Lipid-A-nak is van némi szerepe. Ez nem meglepő, ha figyelembe vesszük, hogy ennek
felépítésében a természetben viszonylag kisszámú zsírsav vesz részt. A hasonló

Proteus morganii törzsek szerológiai keresztkapcsolatai
64
keresztreakciók csak szerológiai jellemzése nem elégséges, mivel gyakran a keresztreagáló
ellenanyagok nemcsak egyforma, hanem hasonló oligoszacharid fragmenteket ismernek fel.
Az O oldallánc kémiai jellemzését vékonyréteg kromatográfiás vizsgálatokkal
indítottuk. Az LPS hidrolízisével előállított monoszacharidok vizsgálata során a vizsgált
törzsek hasonló arányban tartalmazták az egyforma monoszacharidokat. A kromatográfiás
vizsgálatok azonban nem adtak információt a colitóz jelenlétéről, csak sejtéseink és a
mólarányok figyelembe vétele után feltételeztük, hogy mindhárom lipopoliszacharid
ismétlődő egységeiben jelen vannak. Nehézséget jelentett az a tény, hogy az említett cukor
kereskedelmi forgalomban nem kapható, így standardként használni nem tudtuk és a retenciós
faktora is ismeretlen.
Kémiai szerkezete alapján feltételezzük, hogy rendszerünkben a kiinduló vegyületénél
(galaktóz) gyorsabban fut, mivel kevesebb hydroxil csoportja van és ezért a glukózzal egy
magasságban, azzal együtt lehet láthatóvá tenni. Ezt látszik alátámasztani az egyes foltok
nagyságának összehasonlítása is. A colitóz meghatározásához további vizsgálatokra volt
szükségünk, így a tömegspektrométerhez kötött gázkromatográf segítségével a TLC-nél
finomabb elkülönítést, és az egyes monoszacharidok pontos meghatározását lehetett elérni.
A rendelkezésünkre álló kis mennyiségű colitóz gázkromatográfiás és
tömegspektrometriás meghatározása után lehetőségünk nyílt azt a vizsgált baktériumok LPS
hidrolizátumában is identifikálni. Mindhárom törzs (P. morganii O34 (8662/64, E. coli O111
és S. Adelaide O35) lipopoliszacharidja elemzésre került, és mindannyian tartalmazták a
colitózt is. Ugyanakkor a többi cukorösszetevők gázkromatográfiás spektruma is
superponálható volt, ami a hasonló minőségű cukrok hordozását bizonyítja.
Igazából pontos szerkezeti meghatározást mégis a fenti lipopoliszacharidok DPS-einek
NMR vizsgálata hozott. A három endotoxin esetén hasonló spektrumot kaptunk, a colitóz
egyértelműen kimutatható mindhárom endotoxinban. Mindez azt bizonyítja, hogy a P.
morganii O34 (8662/64) O oldallánca ugyanolyan ismétlődő egységekből épül fel, mint a
korábban már megismert E. coli O111 és S. Adelaide O35 lipopoliszacharidja, vagyis két
colitóz (3,6-dideoxy-L-galaktóz), mely glukóz molekulán keresztül kapcsolódik a galaktózhoz
és az N acetyl-galaktózaminhoz.
A genetikai vizsgálatok eredményei, bár korántsem mondhatóak teljesnek, mivel a
teljes gén cluster szekvencia analízise nem történt meg, azt mutatják, hogy ezek a gének a
vizsgált baktériumokban bizonyos fokú rokonságot, konzerváltságot mutatnak.
Néhány esetben a keresztreakciókat nem a lipopoliszacharidok, hanem a külső
membrán fehérjék adják [Literacka 1999]. Vizsgálataink bizonyították, hogy a keresztreakciók

Proteus morganii törzsek szerológiai keresztkapcsolatai
65
egy részéért az egyforma oligoszacharid fragmentek tehetők felelőssé, de néhány esetben már
korábban is leírták, hogy szoros összefüggés figyelhető meg a különböző szerotípusú E. coli
törzsek LPS és külső membrán fehérjéi között [DeMaria 1988, Orskov 1990, Nishimura 1996]. Ezért
megvizsgáltuk a külső membrán fehérjéket is.
A Gram-negatív baktériumok nagyon változatos antigén szerkezettel rendelkeznek. A
felszínükön megjelenő fehérje vagy szénhidrát alapú anyagok határozzák meg a szerológiai
viselkedésüket, ezek akár csak minimális megváltozása már eltérő ellenanyag választ
eredményez.
A fent ismertetett módon kipreparált külső membrán fehérjéket elektroforesises
módszerekkel vizsgáltuk. A kapilláris elektroforesises és chip alapú vizsgálatokkal a 35-50
kDa nagyságú régióban találtunk néhány domináns fehérjét. Az OMP expressiós képe az
Enterobacteriaceae családban nagyfokú konzerváltságot mutat [Molly 2001] és a major
fehérjék elsősorban ebben a molekulasúly tartományban mutathatók ki [Vaara 1984, Bassford
1977], azonban a különböző specieseknek megfelelően ezek a fehérjék egyediek és lényeges
eltérést mutatnak.
Mindhárom törzs elektroferogramján jól látható a 36 és 41 KDa nagyságú fehérje
jelenléte. Az egyes törzseknél további egyedi major fehérjék jelenléte is kimutatható (pl. a S.
Adelaide O35 esetében egy 39 kDa nagyságú fehérje), amely azonban nem játszik szerepet a
keresztreakció kialakításában. Habár a S. Adelaide O35 elektroforetikus képén a 36 és 41 kDa
fehérjék mennyisége jóval kevesebb, de jelenlétük elégséges egy keresztkapcsolat
kialakításához. A fehérjék azonosságát a különböző törzsek együttes injektálásával is
bizonyítottuk, amikor az injektálás után a többi fehérjéhez képest a két közös fehérje
mennyisége szignifikánsan megnövekedett.
Eredményeink további megerősítésére Western blot technikát alkalmaztunk,
amelyhez a korábban ismertetésre került eljárással, a 36 és 41 kDa fehérje ellen termelt
poliklonális immunsavót használtunk. A P. morganii O34 (8662/64) ellen termelt
immunsavó, hasonlóan az E. coli O111 és S. Adelaide O35 ellen termelt immunsavókhoz,
csak az említett fehérjékkel adott keresztreakciót.
Mindezek ismeretében feltételezzük, hogy a P. morganii O34 (8662/64), E. coli O111
és Salmonella Adelaide O35 keresztreakcióinak kialakításában nemcsak az egyforma
ismétlődő alegységeket tartalmazó lipopoliszacharidok játszanak szerepet, hanem a külső
membrán fehérjék is.
Az antigén rokonságot elvileg ki lehet használni a súlyosabb megbetegedést okozó
baktériumok elleni harcban. Az LPS protektív antigén, mivel mind a poliklonális, mind a

Proteus morganii törzsek szerológiai keresztkapcsolatai
66
monoklonális ellenanyagok komplement jelenlétében baktericid hatásúak és segítik az
opsonizációt, valamint kísérletes körülmények között egerekben és kutyákban az LPS ellen
termelődött ellenanyagok kivédték a fertőzést. [Coughlin 1987, Gupta 1995].
Ezen ismeretek alapján határoztuk el egy olyan vakcina lehetőségének a vizsgálatát,
amely keresztprotektívitást biztosító O specifikus ellenanyagok termelésén alapul.
Elképzelésünkkel nem vagyunk egyedül, ugyanis az Enterobacteriaceae családba tartozó
baktériumokkal történő oltóanyagok kidolgozásán már évek óta több kutatócsoport is
foglalkozik, és hoztak is némi eredményt [Rauss 1972, DeMaria 1988, Schwartzer 1988, Baumgartner
1991, Gupta 1995, Wang 1999, Nagy 2004, Nagy 2006].
Az LPS önmagában nem használható védőoltásként toxicitása miatt, a DPS pedig
nem elég immunogén, valószínűleg alacsony mólsúlya miatt [Gupta 1995]. Az E. coli O111 és
Salmonella Adelaide O35 a különböző virulencia faktorok kombinációjával hozzák létre az
eltérő klinikai képet, így azok szerepével is számolnunk kell [Donnenberg 1989, Whittam 1993].
Az egészséges önkéntesek hővel elölt, az Enterobacteriaceae családba tartozó
baktériumokkal történő oltása általában csak IgM immunválaszt vált ki és az újraoltás nem
eredményez booster hatást. [DeMaria 1988, Schwartzer 1988, Baumgartner 1991]. Néhány oralis
vakcina biztonságosságát és immunogenitását Rauss és munkatársai is vizsgálták
újszülötteken. A vakcina 1687 vizsgálatba bevont alany esetében 41%-os hatékonyságúnak
bizonyult [Rauss 1972]. Egerek E. coli O111 hidrazonizált LPS-tetanus toxoid intramusculáris
bevitele után erőteljesebb IgG immunválsszal reagáltak, mint más konjugált vakcinák
esetében [Gupta 1995]. Egerek E. coli O111 O antigént expresszáló attenuált Salmonella
enterica sv. Typhimurium aroA törzsével történt intraperitonealis oltásra jó IgG
immunválaszt adtak, míg oralis alkalmazás esetén az IgG és IgA immunválasz is erőteljes volt
az E. coli O111 ellen [Wang 1999].
In vitro kísérleteink eredményei, amelyek a növekedés gátlását, valamint az
opszonizáció kialakulását célozták, támogatják elképzeléseinket. Figyelembe kell venni azt is,
hogy az LPS önmagában nem alkalmas T sejtes válasz, valamint memória sejtek kialakítására.
A jó memória és T sejtes immunitás kiváltásához konjugálni kell valamilyen jó
immunogenitást is mutató fehérjével, vagy teljes baktériumot kell használni az
immunizáláshoz. Bár a vakcinológia ma már az egyre kevesebb fölösleges antigént tartalmazó
oltóanyagok fejlesztése felé mozdult el (kevesebb teher az oltandó szervezetnek), oralis
alkalmazás esetén még napjainkban is megfontolandó a teljes baktériumokkal történő
immunizálás.

Proteus morganii törzsek szerológiai keresztkapcsolatai
67
Az O specifikus poliszacharid protektív antigén, tehát elméletileg a per os
alkalmazott keresztreagáló baktérium alkalmazása védelmet biztosíthatna a
keresztantigenitású kórokozókkal szemben. A P. morganii-hez hasonlóan nem
enteropathogén más baktériumok, amelyek jelentős kórokozókkal mutatnak antigén
rokonságot, hasonló kísérletek alapjául szolgálhatnak, majd perspektívikusan heterológ
vakcinatörzsként jöhetnek szóba.
6. AZ ÚJ EREDMÉNYEK ÖSSZEFOGLALÁSA
A Gram-negatív baktériumok között gyakran előforduló szerológiai kereszkapcsolatok
vizsgálatára ma már számos olyan módszer áll rendelkezésünkre, amely az immunológiai
jellegű technikákon túl genetikai, molekuláris biológiai, szerkezeti meghatározásokon
keresztül próbálja feltárni ezen reakciók jellegét.
1-2. A leggyakrabban használt immunológiai módszerek egyike az ELISA. A
rendelkezésünkre álló, kipróbált rendszerek érzékenysége nem felelt meg az elvárásoknak,
mert a blokkolásra használt BSA aspecifikus kötődése az LPS-hez a reakció
alkalmazhatóságát korlátozta. A probléma kiküszöbölése érdekében módszertani fejlesztést
végeztünk. Az optimálisnak talált blokkoló anyag (kecskesavó) használatával szignifikánsan
jobb eredményeket értünk el, a módszer alkalmassá vált kis mértékű antigénszerkezetbeli
különbségek megbízható detektálására is. Módszerünket különböző rendszerekben, így a S.
sonnei mutánsok elkülönítésén túl, klinikai minták vizsgálatával is teszteltük.
3. A korábban csak csőagglutinációs módszerrel megállapított P. morganii O34
(8662/64), E. coli O111 és S. Adelaide O35 közötti keresztkapcsolatokat újabb, érzékenyebb
immunológiai módszerek széles skálájának felhasználásával bizonyítottuk, és
megerősítettük.
4.-5. Az immunológiai reakciókban résztvevő hőstabil, lipopoliszacharid antigéneket
kipreparáltuk, tisztítottuk és kémiai módszerek segítségével azok szerkezetét is
meghatároztuk. Korábban három esetben bizonyították, hogy két rendszertanilag egymástól
távol álló baktérium egyforma O antigént hordoz. Vizsgálataink után ezt a sort egy újabb
taggal bővíthetjük, ugyanis eredményeink azt bizonyítják, hogy a P. morganii O34 (8662/64),
E. coli O111 és S. Adelaide O35 lipopoliszacharidjainak ismétlődő egységei azonosak.
Mindhárom baktérium tartalmaz egy, a természetben ritkán előfoduló cukormolekulát, a 3,6-
dideoxy-L-galaktózt, közismert nevén colitózt, amit mind gázkromatográfiával és
tömegspektrográfiával, mind NMR segítségével ki tudtunk mutatni.

Proteus morganii törzsek szerológiai keresztkapcsolatai
68
6. A vizsgált baktériumok genetikai rokonságának bizonyítására a P. morganii O34
(8662/64) colitóz szintéziséért felelős géncsoport egyik génjének (manB) szekvenálását is
elvégeztük. A kérdéses gén nagyfokú hasonlóságot mutatott E. coli O111 és S. Adelaide O35
megfelelő génjével. A szintetizált génszakasz szekvencia sorrendjét a génbankba helyeztük,
ott EF051576 szám alatt érhető el.
7. Ismert, hogy a hasonló LPS-t hordozó baktériumok külső membrán fehérjéi
is hasonlóak, ezért megvizsgáltuk, mely fehérjék játszanak szerepet a keresztreakció
kialakításában. Egy 36 és egy 41 kDa nagyságú fehérje mindhárom törzsben kimutatható volt,
és immunblot reakció alapján szerológiailag is hasonlónak bizonyultak.
8. Hasonló vagy egyforma antigének hordozása esetén jogosan merül fel a kérdés,
vajon ezek a közös antigének (fehérje vagy lipopoliszacharid molekulák) alkalmasak
lehetnek-e keresztimmunitás kiváltására. Ennek bizonyítására elvégzett növekedés gátlási és
opszonizációs vizsgálatok megerősítették, hogy az egyik törzs ellen termelődött ellenanyagok
komplement jelenlétében baktericid hatásúak a keresztreagáló törzsekkel szemben, emellett
opszonizáló hatásukkal önmagukban is elősegítik azok makrofágok általi bekebelezését.
9. A bevezetett és eredményesen használt immunkémiai módszerek további
lehetőségeket biztosítanak a Gram-negatív baktériumok közötti szerológiai
keresztkapcsolatok további, szélesebbkörű vizsgálatára.

Proteus morganii törzsek szerológiai keresztkapcsolatai
69
7. IRODALOMJEGYZÉK
1. Annane D., Sebille V., Charpentier C., Bollaert P.E., Francois B., Korach J.M., Capellier G.,
Cohen Y., Azoulay E., Troche G., Chaumet-Riffaut P., Bellissant E.: Effect of treatment with low
doses of hydrocortisone and fludrocortisone on mortality in patients with septic shock. JAMA. 2002;
288:862-871.
2. Ansaruzzaman M., Shimada T., Bhuiyan N.A., Nahar S., Alam K., Islam M.S. Albert J.: Cross
reaction between a strain of Vibrio mimicus and V. cholerae O139 Bengal. J. Med. Microbiol. 1999;
48:873-877.
3. Antal A., Kocsis B., Jeges S., Hideg Gy., Tornoczky E.: Serologic examinations in acute appendicits.
Acta Chirurg. Hung. 1998; 37(1-2):39-44.
4. Appelmelk B.J.: Antibodies to the lipopolysaccharides core region and their protective role in Gram-
negative sepsis, Academic thesis Amsterdam 1987;
5. Aucken H.M., Pitt T.L.: Serological relationships of the O antigens of Klebsiella pneumoniae O5,
Escherichia coli O8 and a new serotype of Serratia marcescens. FEMS Microbiol. Letters. 1991; 80:93-
98
6. Bassford P.J., Dietrich D.L., Schnaitman C.L., Reeves P.: Outer membrane proteins of Escherichia
coli VI. Protein Alteration in bacteriophage-resistant mutants. J. Bact. 1977; 131 (2):608-622
7. Bastin D.A., Reeves P.R.: Sequence and analysis of the O antigen gene (rfb) cluster of Escherichia coli
O111. Gene. 1995; 16;164(1):17-23.
8. Baumgartner J.D., Heumann D., Calandra T., Glause M.P.: Antibodies to lipopolysaccharide after
immunization of humans with rough mutant Escherichia coli. J. Infect. Dis. 1991, 163:769-772
9. Bettelheim K.A., Faiers M., Shooter R.A.: Serotypes of Escherichia coli in normal stools. The Lancet
1972; 2(7789):1224-1226.
10. Beutin L., Krause G., Zimmermann S., Kaulfuss S., Gleier K.: Characterization of Shiga toxin-
producing Escherichia coli strains isolated from human patients in Germany over a 3-year period. J.
Clin. Microbiol. 2004; 42(3):1099-108
11. Boivin A., Mesrobeanu I., Mesrobeanu L.: Extraction d'un complex toxique et antigenique a partir du
bacille d'aertrycke. Compt. Rend. Soc. Biol. 1933; 114:307-310
12. Brade H., Brade L., Rietschel E.Th.: Structure activity relationships of bacterial lipopolysaccharide
(endotoxins). Current and future aspects. Zbl. Bakt. Hyg. A. 1988; 286:151-179.
13. Bratková, G. , Majtán, V.: Properties and Participation of "Free" Endotoxin in the Toxic Activity of
Morganella morganii. Folia Microbiol. 1988; 33:323-328.
14. Brooks, G. F., Butel, J. S., Morse, S. A.: Medical Microbiology. Appleton & Lange, 1998; 218-231
15. Burkitt D.P.: The aetiology of appendicitis. Brit. J. Surg. 1971; 58:695-9.
16. Carlsson H.E., Lindberg A.A., Hammaström S.: Titration of antibodies to Salmonella O antigens by
Enzyme-Linked Immunosorbent Assay. Inf. Immun., 1972; 6(5):703-708

Proteus morganii törzsek szerológiai keresztkapcsolatai
70
17. Clausen J.: Immunochemical techniques for the identification and estimation of macromolecules.
Burdon, R.H., Knippenberg, P. H. (Eds) Laboratory techniques in biochemistry and molecular biology.
vol. 1. part. 3., Elsevier, Amsterdam 1991
18. Cooper M.L., Keller H.M., Walters E.W.: Microscopic characteristics of colonies of Shigella flexneri
2a and 2b and their relation to antigenic composition, mouse virulence and immunogenicity, J.
Immunol. 1957; 78:160-171
19. Costerton, J.W., Ingram, J.M., Cseng, K.J.: Structure and function of the cell envelope of Gram-
negative bacteria. Bact. Rew. 1974; 38:87-110.
20. Coughlin R.T., Bogard W.C. Jr.: Immunoprotective murine monoclonal antibodies specific antibodies
for outer core polysaccharide and for the O-antigen of Escherichia coli O111:B4 lipopolysaccharide
(LPS). J. Immunol. 1987; 139:557-561
21. Datta D.B., Arden B., Henning U.: Major proteins of the Escherichia coli outer cell envelope
membranes as bacteriophage receptors. J. Bact. 1977; 131(3): 821-829
22. DeMaria A., Johns M.A., Berbeerich H., McCabe W.R.:. Immunization with rough mutants of
Salmonella minnesota: initial studies in human subjects. J. Infect. Dis. 1988; 158: 301-311
23. Donnenberg M.S., Donahue-Rolfe A., Keusch G.T.: Epithelial cell invasion: an overlooked property
of enteropathogenic Escherichia coli (EPEC) associated with the EPEC adherence factor. J. Infect. Dis.
1989; 160:452-459
24. Doyle R.J., Sonnenfeld E.M.: Properties of the Cell Surfaces of Pathogenic Bacteria. Int. Rew. of
Cytology. 1989; 118: 33-92.
25. Edstrom R.D, Heath E.E.: Isolation of colitose-containing oligosaccharides from the cell wall
lipopolysaccharide of Escherichia coli. Biochem. Biophys. Res. Commun. 1965; 21:638-643
26. Ehlag K.M., Alwan M.H., Al-Adnani M.S., Sherif R.A..: Bacteroides fragilis is a silent pathogen in
acute appendicitis. J. Med. Microbiol. 1986; 21:245-249.
27. Engvall E., Perlmann P.: Enzyme linked immunosorbent assay, ELISA. 3. Quantitation of specific
antibodies by enzyme-labeled anti-immunglobulin in antigen-coated tubes. J. Immunol. 1972;
109:129-35.
28. Fox C.F.: The structure of cell membrane. Sci. Amer. 1972; 226(2):30-38.
29. Freudenberg M.A., Fomsgaard A., Mitov I., Galanos C.: ELISA for antibodies to lipid A,
lipopolysaccharides and other hydrophobic agents. Infection 1989; 17:56/322-62/328.
30. Funahara Y., Nikaido H.: Asymmetric localizacion of Lipopolysaccharides on the Outer Membrane of
Salmonella typhimurium. J. Bacteriol. 1980; 141:1463-1454.
31. Galanos C., Lüderitz O., Westphal O.: A new method for the extraction of R lipopolysaccharides.
Eur. J. Biochem. 1969; 9:245-69.
32. Galanos C., Rietschel E.Th.: Bacterial endotoxins. Infection. 1986; 4:155.
33. Grubor N.M., Armstrong D.W., Jankowiak R.: Flow-through partial-filling affinity capillary
electrophoresis using a crossreactive antibody for enantiomeric separations. Electrophoresis 2006;
27:1078–1083
34. Gupta R.K., Egan W., Bryla D.A., Robbins J.B., Szu S.C.: Comparative immunogenicity of
conjugates composed of Escherichia coli O111 O-specific polysaccharide, prepared by treatment with

Proteus morganii törzsek szerológiai keresztkapcsolatai
71
acetic acid or hydrazine, bound to tetanus toxoid by two synthetic schemes. Infect. Immun. 1995;
63(8):2805-2810
35. Hancock I.C. and Poxton I.R.: Bacterial cell surface techniques. Wiley, Chichester, U.K. 1988
36. Holmes B., Gross J.R., Old D.C.: Principles of bacteriology, virology and immunity. Vol. 2.
Systematic bacteriology. B. C. Decker Inc., ed. Parker, T. M., Collier L.,H., 1994; 416-427, 456-468,
470-493
37. Horstman A.L., Bauman S.J., Kuehn M.J.: Lipopolysaccharide 3-Deoxy-D-manno-octulosonic Acid
(Kdo) Core Determines Bacterial Association of Secreted Toxins. J. Biol. Chem., 2004; 279(9):8070-
8075.
38. Janda M.J., Abbott S.L., Khashe S., Robin T.: Biochemical Investigation of Biogroups and
Subspecies of Morganella morganii. J. Clin. Microbiol. 1996; 34:108-113.
39. Jann K., Jann B.:. Polysaccharide antigens of E. coli. Rev. Infect. Dis. 1987; 9:S517-S526
40. Jann K., Jann B.:. Structure and Biosynthesis of O antigens. In: Handbook of Endotoxin, Vol 1 1984;
pp.138-186
41. Kaback H.R.: Transport across isolated bacterial cytoplasmic membranes. Biochem. Biophis. Acta.
1972; 256:367-416.
42. Kauffmann F.: The bacteriology of Enterobacteriaceae. Munsgaard Copenhagen 1966.
43. Kocsis B., Kontrohr T. Isolation of adenosine 5’-Diphosphate-L-glycero-D-mannoheptose, the
assumed substrate of heptose transferase(s), from Salmonella minnesota R595 and Shigella sonnei Re
mutants. J. Biol. Chem 1984; 259:11858-60.
44. Kondakova A.N., Vinogradov E., Lindner B., Knirel Y.A., Amano K.: Structural studies on the
lipopolysaccharide core of Proteus OX strains used in Weil-Felix test: a mass spectrometric approach.
Carbohydr. Res. 2003; 338(23):2697-709.
45. Kontrohr T.: The identification of 2-amino-2 deoxy-L-altruronic acid as a constituent of Shigella
sonnei phase I. LPS. Carbohydr. Res. 1977; 58:498-500.
46. Kustos I., Kocsis B., Kerepesi I., Kilár F.: Changes in outer membrane protein profile of bacteria after
meropenem-induced postantibiotic effect studied by capillary electrophoresis. Electrophoresis 1998;
19:2324-2330
47. Kustos I., Kocsis B., Kerepesi I., Kilár F.: Protein profile characterization of bacterial lysates by
capillary electrophoresis. Electrophoresis 1998; 19:2317-2323
48. Lüderitz O., Freudenberg M.A., Galanos C., Lehmann V., Rietschel E.Th., Shaw D.H.:
Lipopolysaccharides of Gram-negative bacteria. Cur. Top. in Membranes and Transport. 1983; 17:79-
151.
49. Laemmli U.K.: Cleavage of structural proteins during the assembly of the head of bacteriophage T4.
Nature (London) 1970; 227:680-685
50. Lányi B.: Járványügyi és klinikai bakteriológia. Módszertani útmutató. Országos Közegészségtani
Intézet Budapest 1980
51. Lányi B.: Serological Properties of Psedomonas aeruginosa I. Acta microbiol. Acad. Sci. Hung.
1966/67; 13(4): 295-318.

Proteus morganii törzsek szerológiai keresztkapcsolatai
72
52. Lányi B.: Serological Properties of Psedomonas aeruginosa II. Acta microbiol. Acad. Sci. Hung. 1970;
17(1): 35-48.
53. Lebrecht R.J., Pietrasik K.D., Mayer H.: Terminal and Chain-linked Residues of D-Galacturonic
Acid: Characteristic constituents of the R core Regions of Proteaceae and Serratia marcescens. System.
Appl. Microbiol. 1990; 13:214-219.
54. Leigh D.A., Simmons K., Norman E.: Bacterial flora of the appendix fossa in appendicitis and
postoperative wound infection. J. Clin. Pathol. 1974; 27:997-1000
55. Lindberg A.A., Weintraub A., Nord C.E.: The Humoral Antibody Response to Bacteroides fragilis
Infections in Humans. Scand. J. Infect. Dis. Suppl. 1979; 19:46-51
56. Literacka E., Perepelov A.V., Senchenkova S.N., Zatonsky G.V., Shashkov A.S., Knirel Y.A.,
Kaca W.: Structures of the O-specific polysaccharides and serological cross-reactivity of the
lipopolysaccharides of Proteus mirabilis O24, and O29. FEBS Lett. 1999; 456(2):227-231
57. Lugtenberg B., Bronstein H., N. van Slem, Peters R.: Peptidoglycan-associated outer membrane
proteins in Gram-negative bacteria. Biochem. Biophys. Acta. 1977; 465:571-578.
58. Magyar T., Rimler R.B.: Detection and enumeration of toxin-producing Pasteurella multocida with a
colony-blot assay. J. Clin. Microbiol. 1991; 29(7):1328-1332.
59. Martich G.D., Bonjoukos A.J., Suffredini A.F.: Response of Man to Endotoxin. Immunbiol. 1993;
187:403-416.
60. Maurin M., Eb F., Etienne J., Raoult D.: Serological cross-reactions between Bartonella and
Chlamydia species: Implication for diagnosis. J. Clin. Micobiol. 1997; 35(9):2283-2287
61. Mergenhagen S.E., Snyderman R., Gewurz H., Shin H.S.: Significance of complement to the
mechanism of action of endotoxin. Curr. Top. Microbiol Immunol. 1969; 50:37-77.
62. Mills S.D. and Finlay B.: Comparison of Salmonella typhi and Salmonella typhimurium invasion,
intracellular growth and localization in cultured human epithelial cells. Microbiol. Pathog. 1994;
17:409-423
63. Molloy M.P., Phadke N.D., Maddock J.R., Andrews P.C.: Two-dimensional electrophoresis and
peptide mass fingerprinting of bacterial outer membrane proteins. Electrophoresis 2001; 22:1686-1696
64. Morrison D.C., Ryan J.L.: Bacterial Endotoxic Lipopolysaccharides: Vol. I. Cellular Biology and.
Molecular Biochemistry: Vol. II. Immunopharmacology and Pathophysiology. CRC Press 1992.
65. Murray G.L., Attridge S.R., Morona R.: Altering the length of the lipopolysaccharide O antigen has
an impact on the interaction of Salmonella enterica serovar Typhimurium with macrophages and
complement. J. Bact. 2006; 188(7):2735-2739
66. Mühlradt P., Menzel J., Goletzki J., Speth V.: Outer membrane of Salmonella typhimurium and
Escherichia coli. J. Biol.Chem. 1977; 252:7850-7861.
67. Müller A., Hinckley A., Rothfield L.: Studies of phospholipid-requiring bacterial enzymes. III.
Purification and Properties of uridine diphosphate glucose: Lipopolysaccharide glucosyl transferase I. J.
Biol. Chem. 1972; 247:2614-2622.
68. Nagy G., Dobrindt U., Hacker J., Emody L.: Oral immunization with an rfaH mutant elicits
protection against salmonellosis in mice. Infect. Immun. 2004; 72(7):4297-301.

Proteus morganii törzsek szerológiai keresztkapcsolatai
73
69. Nagy G., Danino V., Dobrindt U., Pallen M., Chaudhuri R., Emody L., Hinton J.C., Hacker J.:
Down-regulation of key virulence factors makes the Salmonella enterica serovar Typhimurium rfaH
mutant a promising live-attenuated vaccine candidate. Infect. Immun. 2006; 74(10):5914-25.
70. Nelson D., Neill W., Poxton I.R.: A comparison of immunoblotting, flow cytometry and ELISA to
monitor the binding of anti-lipopolysaccharide monoclonal antibodies. J. Immunol. Methods. 1990;
133:227-233.
71. Nemes Zs., Péterfi Z.: Együttes leptospira- és hantavírus-fertőzés ugyanabban a betegben. Orv. Hetil
2000, 141(10):499-502
72. Nikaido H., Nakae T.: The outer membrane of Gram-negative bacteria. Adv. Microbiol. Physyol. 1979;
20:163-240.
73. Nishimura L.S., Ferreira L.C.S., Pacheco A.B.F., Guth B.E.C.: Relationship between outer
membrane protein and lipopolysaccharide profiles and serotypes of enterotoxigenic Escherichia coli
isolated in Brazil. FEMS Microbiol. Letters. 1996; 143:253-258
74. Norrander J.M., deCathelineau A.M., Brown J.A., Porter M.E., Linck R.W.: The Rib43a protein is
associated with forming the specialized protofilament ribbons of flagellar microtubules in
Chlamydomonas. Mol. Biol. Cell. 2000; 11(1):201-15.
75. Nowotny A., Nowotny M.A., Behling U.H.: The Neglected problem of Endotoxin heterogenity in:
“Bacterial Endotoxins and host response” Agarwal, M. K. (ed). Elsevier North Holland B.V. 1980.
76. Nowotny A., Spitzer J.J., Ziegler E.J.: Cellular and Molecular Aspects of Endotoxin Reactions in:
Endotoxin Research Series Vol. I. Nowotny (ed).-Molecular structure of bacterial endotoxin in relation
to bioactivity- Elsevier Sci. Publ.1990.
77. Onderdonk A.B., Bartlett J.G., Louie T.J., Sullivian-Seigler N. and Gorbach S.L.: Microbial
synergy in experimental intra-abdominal sepsis. Infect. Immun. 1976; 13:22-26
78. Orskov F., Whittam T.S., Cravioto A., Orskov I.,: Clonal relationships among classic
enteropathogenic Escherichia coli (EPEC) belong to different O groups. J. Infect. Dis. 1990; 162:76-81
79. Osborn M.J., Gander J.E., Parisi E., Carson J.: Mechanism of assembly of the outer membrane of
Salmonella typhimurium. Site of synthesis of lipopolysaccharide. J. Biol. Chem. 1972; 247:3973-3986.
80. Osborn M.J.: Studies on the gram-negative cell wall, I. Evidence for the role of 2-keto-3-
deoxyoctonate in the lipopolysaccharide of Salmonella typhimurium. Biochemistry. 1963; 50:499-506
81. Pál T: Shigella. Czirók É. (főszerkesztő) Klinikai és járványügyi bakteriológia, Melánia kiadó 1999;
366-372
82. Péterfi Z., Kocsis B.: Comparison of blocking agents for an ELISA for LPS. J. Immunoassay 2000;
21(4):341-4
83. Péterfi Z., Kovács K., Antal A., Kocsis B.: Anti-lipopolysaccharide antibodies in acute appendicitis
detected by ELISA. APMIS. 2006; 114(4):265-269.
84. Péterfi Z., Kustos I., Kilár F., Kocsis B.: Capillary electrophoretic characterization of cross-reacting
Gram-negative bacterial (Proteus morganii O34, Escherichia coli O111, Salmonella Adelaide O35)
outer membrane proteins. közlésre elfogadva (J. Chromatography A)
85. Péterfi Z., Nemes Zs.: Leptospirosis és hantavírus fertőzés. Orvostudományi Értesítő 2000; 73:172-
175

Proteus morganii törzsek szerológiai keresztkapcsolatai
74
86. Péterfi Z., Ősz E., Reuter G., Kilár A., Kilár F., Kocsis B.: Structure and serologic properties of O-
specific polysaccharide from Proteus morganii O34 (8662/64) possessing cross-reactivity with
Escherichia coli O111 and Salmonella Adelaide O35, bírálat alatt (J. Chromatography B)
87. Pieper R., Kager L., Tiedefeldt U.: Obstruction of appendix vermiformis causing acute appendicitis.
Acta Chir. Scand. 1982; 148:63-72
88. Pieper R., Kager L., Weintraub A., Lindberg A.A., Nord C.E.: The role of Bacteroides fragilis in
the pathogenesis of acute appendicitis. Acta Chirurg Scand 1982; 148:39-44.
89. Pugsely A.P., Schnaitman C.A.: Outer membrane proteins of Escherichia coli VII. Evidence that
bacteriophage-directed protein 2 function as a pore. J. Bact. 1978; 133(3):1181-1189
90. Raetz C.R.H., Garrett T.A., Reynolds C.M., Shaw W.A., Moore J.D., Smith D.C. Jr, Ribeiro A.A.,
Murphy R.C., Ulevitch R.J., Fearns C., Reichart D., Glass C.K., Benner C., Subramaniam S.,
Harkewicz R., Bowers-Gentry R.C., Buczynski M.W., Cooper J.A., Deems R.A., Dennis E.A.:
Kdo2-Lipid A of Escherichia coli, a defined endotoxin that activates macrophages via TLR-4. J. Lipid.
Res. 2006; 47(5):1097-111
91. Raetz C.R.H., Whitfield C.: Lipopolysaccharide endotoxins. Annu. Rev. Biochem 2002; 71:635-700
92. Ralovich B., Vörös S.: Morganella morganii antigenically related to Escherichia coli O111: K58 (B4).
Acta. Microbiol. Acad. Sci. Hung. 1967; 14:189-194.
93. Rauss K., Ketyi I., Matusovits E., Szendrei L., Vertenyi A. and Varbiro B.: Specific oral prevention
of infantile enteritis. 3. Experiments with corpuscular vaccine. Acta. Microbiol. Acad. Sci. Hung. 1972,
19:19-28.
94. Rauss K., Kétyi I., Vertényi A., Vörös S.: Studies on the nature of phase variation of Shigella sonnei.
Acta. Microbiol. Hung. 1961; 8:53-63.
95. Rauss K., Vörös S.: Antigenic relationships between Morganella morganii and different genera of
enterobacteriacea. Acta. Microbiol. Acad. Sci. Hung. 1967; 14:199-204
96. Rauss K., Vörös S.: The biochemical and serological properties of Proteus morganii. Acta. Microbiol.
Acad. Sci. Hung. 1959; 6:233-248
97. Rietschel E.T., Kirikae T., Schade F.U., Ulmer A.J., Holst O., Brade H., Schmidt G., Mamat U.,
Grimmecke H.D., Kusumoto S.: The chemical structure of bacterial endotoxin in relation to
bioactivity. Immunobiology. 1993; 187(3-5):169-90.
98. Rietschel E.Th., Brade H.: Bacterial endotoxins. Sci. Amer. 1992; 267:26-33.
99. Rietschel E.Th., Seydel U., Zähringer U., Schade U.F., Brade L., Loppnow H., Feist W., Wang
M.H., Ulmer A.J., Flad H.D.: Bacterial endotoxin: molecular relationships between structure and
activity. Infect. Dis. Clin. North Am. 1991; 5(4):753-79.
100. Rietschel E.Th., Wollenweber H.W., Brade H., Zähringer B.H., Giesbrecht P.: Structure and
conformation of the Lipid - A component of lipopolysaccharides in: ”Handbook of Endotoxin” Vol. I.
Chemistry of Endotoxin. Rietschel, E. Th. (ed). Elsevier Sci. Publ. B. V. 1984; 187-220.
101. Rietschel E.Th., Zähringer U., Brade H.: Bacterial Endotoxins: relation of chemical structure to
biological activity. Immunopharmacology of Inf. Diseases Vaccine Adjuvants and Modulators of Non
Specific Resistance. Ala R. Liss. Inc. 1987; 75-89.

Proteus morganii törzsek szerológiai keresztkapcsolatai
75
102. Sawardeker, J.S., Sloneker J.H., Jeanes A.:. Quantitative determination of monosaccharides as their
alditol acetates by gas liquid chromatography, Anal. Chem. 1965; 37:1602-1604
103. Schaller A., Troller R., Molina D., Gallati S., Aebi C., Stutzmann Meier P.: Rapid typing of
Moraxella catarrhalis subpopulations based on outer membrane proteins using mass spectrometry.
Proteomics 2006, 6:172–180
104. Schwartzer T.A., Alcid D.V., Numsuwan V., Gocke D.J.: Characterization of the human antibody
response to an Escherichia coli O111:B4 and J5 vaccine. J. Infect. Dis. 1988; 158:1135-1136
105. Senior B.W.: The typing of Morganella morganii by bacteriocin production and sensitivity. J. Med.
Microbiol. 1987; 23:33-39.
106. Seydel U., Schromm A.B., Blunck R., Brandenburg K.: Chemical structure, molecular conformation,
and bioactivity of endotoxins. Chem. Immunol. 2000; 74:5-24.
107. Shu Q., Bird S.H., Gill H.S., Rowe J.B.: Immunological cross-reactivity between the vaccine and
other isolates of Streptococcus bovis and Lactobacillus. FEMS Immunol. Med. Microbiol. 1999;
26:153-158.
108. Snell E.S.: Gram-negative bacterial endotoxin and the pathogenesis of fever. Prog. Drug. Res. 1975;
19:402-11.
109. Stock J.B., Ranch B., Roseman S.: Periplasmic space in Salmonella typhimurium and Escherichia
coli. J. Biol. Chem. 1977; 252:7850-7861.
110. Takahashi K., Fukada M., Kawai M. Yokochi T.: Detection of lipopolysaccharide (LPS) and
identification of its serotype by an enzyme-linked immunosorbent assay (ELISA) using poly-L-lysine.
J. Immunol. Methods 1992; 153:67-71.
111. Toller G.L., Horvath J.E., Schally A.V., Halmos G., Varga J.L., Groot K., Chism D., Zarandi M.:
Development of a polyclonal antiserum for the detection of the isoforms of the receptors for human
growth hormone-releasing hormone on tumors. Proc. Natl. Acad. Sci. USA. 2004; 101(42):15160-
15165.
112. Trautmann M., Cross A.S., Reich G., Held H., Podschun R., Marre. R.: Evaluation of a competitive
ELISA method for the determination of Klebsiella O antigens. J. Med. Microbiol. 1996; 44:44-51.
113. Trevelyan W.E., Procter D.P. and Harrison J.S.: Detection of sugars on paper chromatogramms.
Nature 1950; 166:444-445.
114. Under F.M.: The chemistry and biological significance of 3-deoxi-D-manno-2 octulosonic acid
(KDO). Adv. In Carbohyd. Chem. Biochem. 1981; 38:324-388
115. Vaara, M., Nikaido H.: Molecular organization of bacterial outer membrane, Handbook of endotoxin,
Vol. 1. Chemistry of endotoxin. Elsevier, Amsterdam, ed. Rietchel E. Th. 1984; 1-45.
116. Vinogradov, E.:. Structure of the O-specific polysaccharide chain of the lipopolysaccharide of
Bordetella hinzii. Carbohydrate Research 2002; 337: 961-963
117. Waage A., Brandtzaeg P., Espevik T., Halstensen A.: Current understanding of pathogenesis of
Gram-negative shock. Infect. Disease Clin. North Amer. 1991; 5(4):781-791.
118. Wang L., Curd H., Qu W. and Reeves P.R.: Sequencing of Escherichia coli O111 O-antigen gene
cluster and identification of O111-specific genes. J. Clin. Microbiol. 1998; 36(11): 3182-3187

Proteus morganii törzsek szerológiai keresztkapcsolatai
76
119. Wang L., Curd H., Reeves P. R.:. Immunization of mice with live oral vaccine based on a Salmonella
enterica (sv Typhimurium) aroA strain expressing the Escherichia coli O111 O antigen. Microb.
Pathog. 1999; 27(1):55-9
120. Wang L., Reeves P.R.: The Escherichia coli O111 and Salmonella enterica O35 Gene Clusters: Gene
Clusters Encoding the Same Colitose-Containing O Antigen Are Highly Conserved. J. Bact. 2000;
182(18):5256-5261
121. Watson D.W., Kim Y.B.: Modification of host responses to bacterial endotoxins. I. Specificity of
pyrogenic tolerance and the role of hypersensitivity in pyrogenicity, lethality, and skin reactivity. J.
Exp. Med. 1963; 118:425-46.
122. Westphal O., Jann K., Himmelspach K.: Chemistry and Immunochemistry of Bacterial
Lipopolysaccharides as Cell Wall Antigens and Endotoxins. Prog. Allergy. 1983; 33:9-39.
123. Westphal O., Lüderitz O., Galanos C., Mayer H., Rietschel E.Th.: The story of bacterial endotoxin,
3-rd International Conference on Immunpharmacology, Firenze 1985.
124. Westphal O., Lüderitz O., Bister F.: Über die Extraktion von Bakterien mit Phenol/Wasser. Z.
Naturforsch. Teil B Anorg. Chem. Org. Chem. Biochem. Biophys. Biol. 1952; 7:148-55.
125. Whittam T.S., Wolfe M.L., Wachsmuth I.K., Orskov F., Orskov I., Wilson R.W.: Clonal
relationships among Escherichia coli srains that cause haemorrhagic colitis and infantile diarrhea.
Infect. Immun. 1993; 63:1619-1629
126. www.agilent.com/
127. www.agilent.com/chem/labonachip
128. www.ncbi.nlm.nih.gov/Genbank/
129. Xiao H., Siddiqui J., Remick D.G.: Mechanisms of mortality in early and late sepsis. Infect. Immun.
2006; 74(9):5227-35.
130. Yu F., Mizushima S.: Roles of lipopolysaccharide and outer membrane protein OmpC of Escherichia
coli K-12 in the receptor function for bacteriophage T4. J. Bact. 1982; 151(2):718-722

Proteus morganii törzsek szerológiai keresztkapcsolatai
77
Közlemények és előadások jegyzéke
Közlemények: 1. Péterfi Z, Kocsis B: Comparison of blocking agents for an ELISA for LPS, J. Immunoassay 2000, 21(4):
341-354 IF: 1,286 2. Péterfi Z, Kocsis B: Hogyan válasszunk blokkoló anyagot az endotoxin-ELISA érzékenységének
növeléséhez? Orvostudományi Értesítő 2000, 73:166-171 3. Péterfi Z, Kustos I, Kocsis B: Szerologiai keresztkapcsolatok vizsgálata a Proteus morganii - Escherichia
coli – Salmonella Adelaide törzsek között. Orvostudományi Értesítő 2003/2; 76: 142-150 4. Péterfi Z, Kovács K, Antal A, Kocsis B: Anti-LPS antibodies in acute appendicitis dectected by ELISA.
APMIS 2006; 114(4):265-9. IF: 2,127 5. Péterfi Z, Kustos I, Kilár F, Kocsis B: Microfluid chip analysis of outer membrane proteins responsible for
serological cross reaction between three Gram-negative bacteria: Proteus morganii O34, Escherichia coli O111 and Salmonella Adelaide O35 J. Chromatography A, közlésre elfogadva IF: 3,096
6. Péterfi Z, Ősz E, Reuter G, Kilár F, Kocsis B: Structural and serologic properties of O-specific polysaccharide from Proteus morganii O34 (8662/64) possessing cross-reactivity with Escherichia coli O111 and Salmonella Adelaide O35. J Chromatography B, közlésre elküldve IF: 2,391
7. Nemes Zs, Kiss G, Madarassi PE, Péterfi Z, Ferenczi E., Bakonyi T, Ternák G: Nosocomial transmission of Dengue. Emerging Infect Dis. 2004; 10(10):1880-1 IF: 5,643
8. Nemes Zs, Péterfi Z: A Campylobacter jejuni enteritis, mint a dysenteria syndroma képviselője, Medicus Universalis XXXI/1, 1998, 13-15
9. Nemes Zs, Péterfi Z: Kullancs által közvetített újabb betegség - a humán ehrlichiózis, Praxis 1998; 7(11):56-58
10. Nemes Zs, Péterfi Z: Együttes leptospira- és hantavírus-fertőzés ugyanabban a betegben, Orv. Hetil 2000, 141 (10): 499-502
11. Péterfi Z, Nemes Zs: Leptospirosis és hantavírus fertőzés. Orvostudományi Értesítő 2000, 73:172-175 12. Péterfi Z: Ketolidok - Megoldás a rezisztens légúti kórokozók kezelésében? Th. Antimicrobialis 2002;
8:21-27 13. Péterfi Z: Az osteomyelitis korszerű kezelése Th. Antimicrobialis 2003; 9:9-20 14. Péterfi Z: Toxoplasmosis Th. Antimicrobialis 2003; 11:3-12 15. Péterfi Z: Régi vagy új betegség? Listeriosis, Therapia Antimicrobialis 2003; 12:19-24 16. Péterfi Z: Tularemia biológiai fegyverként is használt kullancsbetegeség, Th. Antimicrob. 2003; 13:42-50 17. Péterfi Z: Emerging fertőző betegségek Orvostudományi Értesítő 2004/1; 77(1):52-62 18. Péterfi Z: A staphylococcusok mikrobiológiai tulajdonságai, rezisztencia változásai, Infekció &
Infekciókontroll 2004; 1:5-13. 19. Péterfi Z: Legionellosis és más atípusos pneumoniák, Infekció & Infekciókontroll 2005; 2(1):13-27 20. Péterfi Z: Újabban megismert hepatitis vírusok (HGV, TTV, SEN-V), Infekció & Infekciókontroll 2005;
2(2):143-152 21. Péterfi Z: Bőrtünetekkel járó zoonózisok, Infekció & Infekciókontroll 2005;2(4):325-336 22. Nemes Zs, Péterfi Z: Necrotizáló fascitist okozó Streptococcus toxicus shock syndroma. Infekció &
Infekciókontroll 2005; 2(4):276-282 23. Péterfi Z: Az influenzavírusok, Infekció & Infekciókontroll 2005; 2(5):377-384 24. Péterfi Z: Legionellosis és más atípusos pneumoniák, Magyar Orvos 2006; 15(5):18-25 25. Péterfi Z: Növekvő fenyegetés, bomba az élelmiszerben? A salmonellosisról. Infekció & Infekciókontroll.
Megjelenés alatt

Proteus morganii törzsek szerológiai keresztkapcsolatai
78
Könyvfejezetek: Betegség enciklopédia Springer Tudományos Kiadó, Budapest 26. Péterfi Z: Anthrax, Betegség enciklopédia 2002, 591-592 27. Péterfi Z: Oesophagitis infectiosa, Betegség enciklopédia 2002, 765-766 28. Péterfi Z: Chancroid, Betegség enciklopédia 2002, 621-622 29. Péterfi Z: Cryptococcosis, Betegség enciklopédia 2002, 632-633 30. Péterfi Z: Csontokba és izületekbe ültetett protézisek infekciói: Betegség enciklopédia, 2002, 633-634 31. Péterfi Z: Donovanosis, Betegség enciklopédia 2002, 645-646 32. Péterfi Z: Escherichia coli okozta infectiók, Betegség enciklopédia 2002, 674-675 33. Péterfi Z: Diphyllobotriasis, Betegség enciklopédia 2002, 644-645 34. Péterfi Z: Emerging fertőző betegségek, Betegség enciklopédia 2002, 656-659 35. Péterfi Z: Ételmérgezések, Betegség enciklopédia 2002, 676-679 36. Péterfi Z: Leptospirosis, Betegség enciklopédia 2002, 733-735 37. Péterfi Z: Paratyphus, Betegség enciklopédia 2002, 776-777 38. Péterfi Z: Parotitis epidemica, Betegség enciklopédia 2002, 778 39. Péterfi Z: Pneumocistis carinii fertőzés, Betegség enciklopédia 2002, 789-790 40. Péterfi Z: Protozoonok által okozott egyéb megbetegedések (Giardiasis, Trichomoniasis), Betegség
enciklopédia 2002, 799-800 41. Péterfi Z: Secunder és tertier peritonitisek, Betegség enciklopédia 2002, 819-821 42. Péterfi Z: Toxocariasis, Betegség enciklopédia 2002, 847-848 43. Péterfi Z: Urethritis férfiakban, Betegség enciklopédia 2002, 864-865 44. Péterfi Z: Urethritis nőkben, Betegség enciklopédia 2002, 866 Absztraktok: 1. Péterfi Z, Mazák I, Kocsis B, Vörös S: Immunochemical and serological analysis of Proteus morganii
strains, Acta Microbiol. Hung. 1999. 46: 131 2. Péterfi Z, Kocsis B: Optimization of ELISA test used for detection of serological cross-reaction between
lipopolysaccharides, Acta Microbiol. Hung. 2000. 47:215 3. Kocsis B, Kustos I, Péterfi Z: Isolation and characterization of a Shigella sonnei absolute rough mutant,
Acta Microbiol. Hung. 2000. 47:217 4. Péterfi Z, Kovács K, Antal A, Kocsis B: Serological examinations in acute appendicitis Acta Microbiol.
Hung. 2004. 51:156 5. Péterfi Z, Kustos I, Kocsis B: Serological cross reaction examination between Proteus morganii -
Escherichia coli – Salmonella Adelaide Acta Microbiol. Hung. 2003. 50:231 6. Kilár F; Bui A; Péterfi Z; Kustos I; Kocsis B.: Bacterial Endotoxins. A complex study by capillary
electrophoresis with laser induced fluorescence, Rome, Italy, ITP, 2004. - L 25
7. Péterfi Z, Nemes Zs: A májcirrhosisos betegeink infekcióinak klinikai tünetei, megoszlása, Infektológia és Klinikai Mikrobiológia 1998. október – V/1:S22
8. Nemes Zs, Szabó J, Péterfi Z: Epidemiológiai és klinikai érdekességek Q lázas betegeinkben, Infektológia és Klinikai Mikrobiológia 1999. Október-VI:21
9. Nemes Zs, Péterfi Z, Ferenczi E: Zoonozisok együttes előfordulása betegeinkben Infektológia és Klinikai Mikrobiológia 2000. Október- VII/1:S30
10. Kiss G, Péterfi Z, Madarassi PE: Pneumoniák etiológiája, kórlefolyása, kezelése a Baranya Megyei Kórház Infektológiai Osztály 5 éves beteganyagában, Infektológia és Klinikai Mikrobiológia 2001
11. Péterfi Z, Madarassi PE, Molnár L, Nemes Zs: Chlamydia pneumoniae infekció kapcsán felismert autoimmun betegség esete Infektológia és Klinikai Mikrobiológia 2002 IX/1: S16
12. Kiss G, Péterfi Z:, Szabó J, Ternák G: Entamoeba histolytica szerológiai vizsgálatok a székletminta amoeba pozitivitása esetén Infektológia és Klinikai Mikrobiológia 2002 IX/1: S14
13. Nemes Zs., Péterfi Z, Takács M: Az újonnan felfedezett hepatitis vírusok előfordulása májbetegeinkben. Infektológia és Klinikai Mikrobiológia 2006; 13(1):S9

Proteus morganii törzsek szerológiai keresztkapcsolatai
79
Előadások és poszter bemutatások: 1. Péterfi Z, Mazák I, Kocsis B, Vörös B: A Morganella morganii törzsek szerológiai vizsgálata. előadás a
POTE TDK konferencián, Pécs, 1997 2. Péterfi Z, Mazák I, Kocsis B, Vörös B: A Morganella morganii törzsek szerológiai vizsgálata. poszter,
Korányi Frigyes Tudományos Fórum, Budapest, 1997 3. Péterfi Z, Mazák I, Kocsis B, Vörös S: A Proteus morganii törzsek immunkémiai és szerológiai vizsgálata.
előadás, Magyar Mikrobiológiai Társaság Nagygyülése, Miskolc, 1998. aug. 4. Péterfi Z, Kocsis B: Optimization of ELISA test used for detection of serological cross-reaction between
lipopolysaccharides. poszter, 13th International Congress of the Hungarian Society for Microbiology. Budapest. 1999. aug. 30
5. Kocsis B, Kustos I, Péterfi Z: Isolation and characterization of a Shigella sonnei absolute rough mutant. előadás, 13th International Congress of the Hungarian Society for Microbiology. Budapest. 1999. aug. 30
6. Péterfi Z, Nemes Zs: Két zoonozis egyidejű előfordulása ugyanabban a betegben. előadás, MIT tudományos ülése 1999. jún.16.
7. Péterfi Z, Kocsis B: Hogyan válasszunk blokkoló anyagot az endotoxin-ELISA érzékenységének növeléséhez? előadás EME Orvostudományi és Gyógyszerészeti Szakosztály X. Tudományos Ülésszaka, Székelyudvarhely, Románia 2000. május 11-13.
8. Péterfi Z, Nemes Zs: Leptospirosis és hantavírus fertőzés. előadás EME Orvostudományi és Gyógyszerészeti Szakosztály X. Tudományos Ülésszaka, Székelyudvarhely, Románia 2000. Május 11-13.
9. Péterfi Z, Kustos I, Kocsis B: Szerologiai keresztkapcsolatok vizsgálata a Proteus morganii - Escherichia coli – Salmonella Adelaide törzsek között, poszter Magyar Mikrobiológiai Társaság Nagygyülése, Balatonfüred, 2001
10. Péterfi Z, Kovács K, Antal A, Kocsis B: Acut appendicitis szerológiai vizsgálata előadás Magyar Mikrobiológiai Társaság Nagygyülése, Balatonfüred, 2002
11. Péterfi Z, Kustos I, Kocsis B: Szerologiai keresztkapcsolatok vizsgálata a Proteus morganii - Escherichia coli – Salmonella Adelaide törzsek között, előadás EME Orvostudományi és Gyógyszerészeti Szakosztály XIII. Tudományos Ülésszaka, Sepsiszentgyörgy, Románia, 2003. Május 29-31
12. Bui A, Péterfi Z, Kustos I, Kocsis B, Kilár F: Analyzation of 8-aminopyrene-1,3,6-trisulfonic acid-labelled hydrolized endotoxins by capillary electrophoresis, poszter 5th International symposium and course, Teaching and learning of „analytical and bioanalytical monitoring methods” 2004 31 May-07 June Sofia, Bulgaria
13. Bui A, Péterfi Z, Kustos I, Kocsis B, Kilár F: Optimization of experimental conditions in the analysis of 8-aminopyrene-1,3,6-trisulfonic acid-labelled hydrolyzed endotoxins by capillary electrophoresis poszter 14th International Symposium on Capillary Electroseparation Techniques, Rome, Italy, September 12-15, 2004
14. Kilár F, Bui A, Péterfi Z, Kustos I, Kocsis B: Bacterial endotoxins. A complex study by capillary electrophoresis with laser induced fluorescence előadás 14th International Symposium on Capillary Electroseparation Techniques, Rome, Italy, September 12-15, 2004
15. Nemes Zs, Péterfi Z: A bakteriális fertőzések előfordulásának gyakorisága és elemzése májcirrhosisos betegeinkben. poszter, Magyar Gasztroenterológiai Társaság 40. Nagygyűlése, Balatonaliga, 1998. Május
16. Péterfi Z: A bakteriális fertőzések előfordulása és azok elemzése májcirrhozisos betegeken. előadás Pécsi Tudományos Hétvége, Pécs 1998 jún.
17. Péterfi Z, Nemes Zs: A májcirrhosisos betegeink infekcióinak klinikai tünetei, megoszlása. poszter, Magyar Infektológiai Társaság éves kongresszusa, Szeged 1998 okt.
18. Nemes Zs, Szabó J, Péterfi Z: Lehet, hogy a foltos alopecia a Q láz ritka szövődménye? poszter Új és veszélyes fertőző betegségek I. hazai kogresszusa 1999. Szept 17-18
19. Nemes Zs, Szabó J, Péterfi Z: Epidemiológiai és klinikai érdekességek Q lázas betegeinkben. poszter, Magyar Infektológiai Társaság éves kongresszusa, Debrecen 1999. Okt.
20. Nemes Zs, Péterfi Z, Ferenczi E: Zoonozisok együttes előfordulása betegeinkben poszter Magyar Infektológiai Társaság éves kongresszusa, Budapest 2000. Okt. 12-14
21. Kiss G, Péterfi Z, Madarassi PE: Pneumoniák etiológiája, kórlefolyása, kezelése a Baranya Megyei Kórház Infektológiai Osztály 5 éves beteganyagában poszter Magyar Infektológiai Társaság éves kongresszusa, Szombathely 2001
22. Nemes Zs, Péterfi Z, Madarassi PE, Ternák G: Afrikából behurcolt rickettsiosis esetei előadás Magyar Infektológiai Társaság tudományos ülése 2001
23. Péterfi Z, Madarassi PE, Molnár L, Nemes Zs: Chlamydia pneumoniae infekció kapcsán felismert autoimmun betegség esete poszter Magyar Infektológiai Társaság éves kongresszusa, Szekszárd 2002

Proteus morganii törzsek szerológiai keresztkapcsolatai
80
24. Kiss G, Péterfi Z, Szabó J, Ternák G: Entamoeba histolytica szerológiai vizsgálatok a székletminta amoeba pozitivitása esetén poszter Magyar Infektológiai Társaság éves kongresszusa, Szekszárd 2002
25. Péterfi Z: Emerging fertőző betegségek előadás EME Orvostudományi és Gyógyszerészeti Szakosztály XIII. Tudományos Ülésszaka, Sepsiszentgyörgy, Románia, 2003. május 29-31.
26. Péterfi Z: Evidence based medicine (AIDS, hasmenéses megbetegedések) előadás Szekszárd továbbképző tanfolyam, 2003. november
27. Péterfi Z: Gastrointestinális infekciók, előadás Pécs, Korszerű Antiinfektív Terápia kongresszus, 2004. április 23-24.
28. Péterfi Z, Madarassi PE: Nosocomialis dengue lázas megbetegedés előadás Magyar Infektológiai Társaság tudomáyos ülése 2004-06-10
29. Péterfi Z: Gastrointestinális infekciók, előadás, Pécs Szakvizsga előkészítő tanfolyam, 2004 30. Péterfi Z: Streptococcus pneumoniae fertőzések klinikuma és terápiája, előadás Pécs, A Magyar
Infekciókontroll egyesület és az ÁNTSZ Baranya Megyei Intézetének kórházhigiénés rendezvénye 2004 31. Péterfi Z: Központi idegrendszeri infekciók, kullancsencephalitis, előadás Pécs, Infektológiai továbbképzés
2004. márc. 19 32. Péterfi Z: Központi idegrendszeri infekciók, kullancsencephalitis, előadás Pécs, Szakvizsga továbbképzés
2005. október 33. Péterfi Z: Gastrointestinális infekciók, előadás, Pécs, Szakvizsga továbbképzés, 2005. október 34. Péterfi Z: Központi idegrendszeri infekciók, kullancsencephalitis, előadás Tatabánya, Infektológiai
továbbképzés 2005. október 35. Péterfi Z: Gastrointestinális infekciók, előadás, Tatabánya, Infektológiai továbbképzés, 2005. október 36. Péterfi Z: Új fertőző betegségek, Pécs. előadás Családszakorvos továbbképzés, 2005 december 37. Péterfi Z: Neuroinfekciók, Pécs Infekció, infekciókontroll, trópusi betegségek, migráció továbbképző
előadás 2005. december 38. Nemes Zs, Péterfi Z: Ribavirin okozta bőrtünetekkel járó eseteink tanulságai, előadás Bükkfürdő 2006.
február 39. Péterfi Z: A hasmenések differenciál diagnosztikája az infektológus szemszögéből, előadás Pécs 2006.
március 40. Péterfi Z, Nemes Zs, Ternák G: Afrikából behurcolt rickettsiosis esetei, előadás Pécs, PTE Orvostudományi
és Egészségtudományi Szakosztálya, 2006. február 27. 41. Péterfi Z: Varicella járvány és az infektológia jelenlegi helyzete, előadás, Pécs, Háziorvosi továbbképzés,
2006. május 2. 42. Nemes Zs., Péterfi Z, Takács M: Az újonnan felfedezett hepatitis vírusok előfordulása májbetegeinkben.
előadás. 2006. október 6. Magyar Infektológiai és Klinikai Mikrobiológiai Társaság 34. Kongresszusa

Proteus morganii törzsek szerológiai keresztkapcsolatai
81
Független idézettség: Péterfi Z, Kocsis B: Comparison of blocking agents for an ELISA for LPS, J. Immunoassay 2000, 21(4): 341-354
1. Rüdiger Bräuming: Analyse von Protein-Protein-Wechselwirkungen mittels Phage Display, PhD disszertáció. Ruprech-Karls Univ, Heidelberg, disszertáció, 2001, 113
2. Coille I: Thermodynamic and kinetic characterisation of antibody / hapten pairs and optimisation of an immunoassay of fluorescence in homogeneous phase. PhD. Disszertáció, Eberhard-Karls Univ., Tübingen. 2001, 96, 98, 120
3. Ochoa R, Martinez JC, Ferriol X, Gracia AM, Estrada E, Blanco R, Licea T, Sotolongo F: Coating of ELISA plates with vaccine antigens, VacciMonitor 2001; 10(4):14-17
4. Catherine A. Young: An immunolocalization study of the life stages of Kudoa thyrsites in Atlantic salmon (Salmo salar). Bachelor of Science degree at Malaspina University-College, Nanaimo, British Columbia, 2002
5. Zheng Y, Rundell A: Biosensor immunosurface engineering inspired by B-cell membrane-bound antibodies: modeling and analysis of multivalent antigen capture by immobilized antibodies. IEEE Trans Nanobioscience. 2003 Mar;2(1):14-25
6. Huang T, Geng T, Sturgis J, Li HB, Gomez R, Bashir R, Bhunia AK, Robinson JP, Ladisch MR: Lysozyme for capture of microorganisms on protein biochips. Enzyme and Microbial Technology 2003; 33(7):958–966
7. Huang TT, Sturgis J, Gomez R, Geng T, Bashir R, Bhunia AK, Robinson JP, Ladisch MR.: Composite surface for blocking bacterial adsorption on protein biochips. Biotechnol Bioeng. 2003; 81(5):618-24.
8. Jin Z, Jin S, Ren C, Xiao Z, Huang X: Chinese J Preventive Med., 2003; 37(4):271-273 9. De Smedt A, Vanderlinden E, Demanet C, De Waele M, Goossens A, Noppen M.: Characterisation of
pleural inflammation occurring after primary spontaneous pneumothorax. Eur Respir J. 2004; 23(6):896-900.
10. Wessolowski A: Amphipathic Hexapeptides - Interaction with Membranes., PhD. Disszertáció. Free Univ. Berlin, 2004
Nemes Zs, Kiss G, Madarassi PE, Péterfi Z, Ferenczi E., Bakonyi T, Ternák G: Nosocomial transmission of Dengue. Emerging Infect Dis. 2004; 10(10):1880-1881
11. Beekmann SE, Henderson DK: Protection of healthcare workers from bloodborne pathogens; Current Opinion in Infect. Dis. 2005; 18(4):331-336
12. Chen LH, Wilson ME: Nosocomial dengue by mucocutaneous transmission: Emerg. Dis. 2005: 11(5):775-775
13. Chen LH, Wilson ME: Non-Vector Transmission of Dengue and Other Mosquito-Borne Flaviviruses. Dengue Bulletin 2005; 29:18-31
Péterfi Z: A staphylococcusok mikrobiológiai tulajdonságai, rezisztencia változásai, Infekció & Infekciókontroll 2004; 1:5-13
14. Rozgonyi Ferenc: Klinikai, járóbeteg-szakorvosi és háziorvosi microbiologiai gyorsdiagnostica, 2. bővített kiadás, 2006
Péterfi Z: Legionellosis és más atípusos pneumoniák, Infekció & Infekciókontroll 2005; 2(1):13-27
15. Rozgonyi Ferenc: Klinikai, járóbeteg-szakorvosi és háziorvosi microbiologiai gyorsdiagnostica, 2. bővített kiadás, 2006
Nemes Zs, Péterfi Z: Necrotizáló fascitist okozó Streptococcus toxicus shock syndroma. Infekció & Infekciókontroll 2005; 2(4):276-282
16. Rozgonyi Ferenc: Klinikai, járóbeteg-szakorvosi és háziorvosi microbiologiai gyorsdiagnostica, 2. bővített kiadás, 2006

Proteus morganii törzsek szerológiai keresztkapcsolatai
82
Nemes Zs, Péterfi Z: Együttes leptospira- és hantavírus-fertőzés ugyanabban a betegben, Orv. Hetil 2000, 141 (10): 499-502
17. Castillo C, Sanhueza L: Sindrome pulmonar por hantavirus. Medwave. 2002; 2(10) 18. Lietaert, L. Dans le cadre d'une etude de la fievre hemorragique avec syndrome renal, mise en place de
piegeages de micromammiferes en lorraine et dans les ardennes. A study of the haemorrhagic fever with renal syndrome; small mammals trapping in lorraine and french ardennes. Fievre hemorragique/ piegeage/ epidemiologie/ syndrome renal/ campagnol roussatre. These de doctorat veterinaire (Ecole nationale vétérinaire de Nantes. France) 2002; 103 p
19. Lienard E.: Contribution à l'étude des carnivores sauvages comme sentinelles de la fièvre hémorragique avec syndrôme rénal. Thèse, Docteur vétérinaire, Ecole nationale vétérinaire de Nantes. France 2003; 145 p.





![D]o] >µl] - SZPZ...Enterobacter, Citrobacter, Morganella morganii, Serratia, Proteus spp t hiter nastanek odpornosti ob zdravljenju s tretjo generacijo cefalosporinov W ] }ov]lµ](https://img.pdfslide.tips/doc/110x75/5e510267a496ad4dc61630b7/do-l-enterobacter-citrobacter-morganella-morganii-serratia-proteus.jpg)























