Embed Size (px)
Citation preview

Aus der Medizinischen Klinik 1 des Marienhospitals Herne - Universitätsklinik -
der Ruhr-Universität Bochum Kommisarischer Direktor : Prof. Dr. med. Voigtmann
_______________________________________________ Reaktive Sauerstoffradikale bei Patienten mit obstruktivem Schlaf-Apnoe-
Syndrom und Regulation der Radikalenbildung durch Angiotensin Rezeptoren
Inaugural-Dissertation zur
Erlangung des Doktorgrades der Medizin einer
Hohen Medizinischen Fakultät der Ruhr-Universität Bochum
vorgelegt von Susanne Stemmler
aus Würzburg 2001

II
Abstract
Stemmler
Susanne
Reaktive Sauerstoffradikale bei Patienten mit obstruktivem Schlaf-Apnoe-
Syndrom und Regulation der Radikalenbildung durch Angiotensin
Rezeptoren
Reaktive Sauerstoffradikale (ROS) spielen eine wesentliche Rolle bei der
intrazellulären Signaltransduktion. Hohe ROS-Konzentrationen führen aber zu
einer Schädigung von Proteinen, Enzymen oder Transkriptionsfaktoren und
damit zu einer Störung der regulären Zellfunktion bei einer Reihe von
Erkrankungen. Patienten mit obstruktivem Schlaf-Apnoe-Syndrom (OSAS)
haben häufig eine arterielle Hypertonie und eine erhöhte kardiovaskuläre
Mortalität. In der Untersuchung sollte jetzt geprüft werden, ob Veränderungen
von intrazellulären ROS bei Patienten mit OSAS vorliegen und welchen Einfluß
Angiotensin AT-2-Rezeptoren bei der Bildung der ROS haben.
Die ROS-Konzentration wurden fluoreszenzspektrophotometrisch in den
mononukleären Leukozyten bestimmt. Innerhalb von 30 Minuten kam es zu
einem spontanen Anstieg von intrazellulären ROS in den mononukleären
Leukozyten von Patienten mit OSAS um 2.8±0.4 (MW ± SEM, n=13) und von
den gesunden Kontrollpersonen um 3.9±0.6 (n=18). Die Stimulation der Zellen
mit Phorbol-myristat-acetat (PMA) führte zu einem Anstieg von intrazellulären
ROS bei Patienten mit OSAS um 10.4±1.6 und bei den gesunden
Kontrollpersonen um 13.8±1.5. Die Vorgabe des Singlet-oxygen-scavengers
Natriumazid, des Peroxynitrit-scavengers Ebselen und des ACE-Hemmers
Captopril führte zu einer signifikanten Verminderung der PMA-stimulierten
intrazellulären ROS-Bildung (jeweils p<0.01). Die Angiotensin AT-2-Rezeptor-
agonisten und der Angiotensin AT-1-Rezeptorantagonist führten ebenfalls zu
einer signifikanten Verminderung der PMA-stimulierten intrazellulären ROS-
Bildung (jeweils p<0.05), der Angiotensin AT-1-Rezeptoragonist und der
Angiotensin AT-2-Rezeptorantagonist zeigten jedoch keine Wirkung.
Die Arbeit zeigt, daß Patienten mit obstruktivem Schlaf-Apnoe-Syndrom keine
signifikanten Veränderungen von intrazellulären reaktiven Sauerstoffradikalen
aufweisen. Weiterhin scheint eine Regulation der Bildung von intrazellulären
reaktiven Sauerstoffradikalen über den Angiotensin AT-2-Rezeptor zu erfolgen.

III
.
.
Dekan : Professor Dr. med. G. Muhr
Referent : PD Dr. med. M. Tepel
Koreferent : Frau Professor Dr. med. M. Schläfke
Tag der mündlichen Prüfung : 14.01.2003

IV
Meiner Familie

V
Inhaltsverzeichnis
1. Einleitung 1
1.1. Schlaf-Apnoe Syndrom 1
1.2. Sauerstoffradikale und Calcium 3
1.3. Prinzip der Fluoreszenz und Durchführung der Messung 5
1.4. Fragestellung der Arbeit 8
2. Materialen und Methoden 9
2.1. Patienten und Kontrollpersonen 10
2.2. Blutentnahme, Präparation von mononukleären Leukozyten und
Versuchsablauf 12
2.2.1. Fluoreszenzmessung von reaktiven Sauerstoffradikalen 13
2.2.2. Fluoreszenzmessung von Calcium 13
2.3. Statistik 15
3. Ergebnisse 16
3.1. Reaktive Sauerstoffradikale bei Patienten mit obstruktivem Schlaf-
Apnoe-Syndrom und bei gesunden Kontrollpersonen 16
3.2. Wirkung von Scavengern und ACE-Hemmern auf die Bildung
reaktiver Sauerstoffradikale 24
3.3. Wirkung am Angiotensinrezeptor 27
3.4. Konzentrationsmessung von Calcium und von reaktive Sauerstoff-
radikalen unter Zugabe verschiedener Sauerstoffradikalstimulatoren
(PMA und Thapsigargin) 32
4. Diskussion 34
5. Zusammenfassung 39
6. Literaturverzeichnis 41
7. Danksagung 48
8. Lebenslauf 49

VI
Abbildungsverzeichnis
Abb.1 : Schematische Darstellung der Entstehung von reaktiven
Sauerstoffradikalen (ROS)
Abb.2 : Entstehung der Fluoreszenz/Emission
Abb.3 : Aufbau eines Fluorometers
Abb.4 : Präparation der mononukleären Leukozyten
Abb.5 : Fluoreszenz-Spektrum des radikalsensitiven Farbstoffes DCF-DA
Abb.6 : Konzentrationsmessung von Natriumazid
Abb.7 : Konzentrationsmessung von Captopril
Abb.8 : Konzentrationsanstieg von ROS über 60 Minuten
Abb.9 : spontane reaktive Sauerstoffradikalbildung von OSAS-Patienten im
Vergleich zu einer gesunden Kontrollgruppe
Abb.10 : PMA induzierte ROS-Bildung von OSAS-Patienten im Vergleich zu
einer gesunden Kontrollgruppe
Abb.11 : Wirkung von verschiedenen Substanzen/Scavengern auf die
ROS-Bildung bei Patienten mit OSAS
Abb.12 : Mechanismus der ROS-Bildung über die Angiotensin AT-1 und
AT-2-Rezeptoren
Abb.13 : Wirkung eines Stickstoffmonoxiddonors (SNAP) auf die ROS-
Entwicklung unter dem Einfluß verschiedener Radikalenstimulatoren
Abb.14 : Mechanismus der ROS-Bildung/Calciumionenkonzentration
Tabellenverzeichnis
Tabelle1 : klinische und biochemische Daten von Patienten mit OSAS
und gesunden Kontrollpersonen

1
1. Einleitung
1.1. Schlaf-Apnoe-Syndrom
Die Prävalenz des obstruktiven Schlaf-Apnoe-Syndroms (OSAS) liegt bei ca.
1-2% der Bevölkerung. Hauptsächlich sind Männer zwischen dem 40-60
Lebensjahr betroffen. Bei Patienten mit obstruktivem Schlaf-Apnoe-Syndrom
kommt es zu schlafbezogenen Atempausen, die länger als 10 Sekunden
andauern. Der Schlafapnoe-Index (AHI) gibt die Anzahl der Atempausen pro
Stunde an. Dabei sind Werte über 10/h pathologisch.
Zur Diagnosefindung ist die Anamnese (starkes Schnarchen, Kopfschmerzen,
Abgeschlagenheit, Tagesmüdigkeit mit Einschlafneigung und Leistungsknick)
einschließlich der Partnerbefragung von großer Bedeutung. Meßtechnisch wird
als erstes ein Screening mittels ambulantem Schlafmonitoring durchgeführt und
bei Hinweisen auf ein obstruktives Schlaf-Apnoe-Syndrom wird noch zusätzlich
eine polysomnographische Langzeitmessung im Schlaflabor herangezogen.
Während der Atempausen steigt der Streßfaktor bei den Patienten an. Dadurch
werden vermehrt Hormone, z.B. Katecholamine, ausgeschüttet, welche zu einer
Erhöhung des Energieumsatzes führen. Bei Oxidationsreaktionen, z.B. bei der
Gewinnung von ATP, fallen häufig reaktive Sauerstoffradikale (ROS) an. Freie
Radikale verursachen Schäden an biologischen Strukturen, so z.B. bei Lipiden,
Proteinen, Kohlenhydraten oder der DNS. Bei den Lipiden kann es durch die
Einwirkung der reaktiven Sauerstoffradikale zu Peroxidationen ungesättigter
Fettsäuren in Zellmembranen kommen. Bei den Proteinen führt dies zu einer
oxidativen Denaturierung, bei den Kohlenhydraten kommt es zu einer Spaltung
(Depolymerisation) von Polysaccharieden und bei der DNS führt dies zu
Hydroxylierungen von Basen, Strangabbrüchen und Quervernetzungen.
Daher werden verschiedene klinische Zustandsbilder mit freien Sauerstoff-
radikalen als zusätzliches mitverursachendes Agens in Zusammenhang
gebracht, wie z.B. bei Arthritiden, Herzkreislauf-Krankheiten (Herzinfarkt),
Lungenödem, Schock sowie Diabetes mellitus.
Das Risiko für Patienten mit obstruktivem Schlaf-Apnoe-Syndrom diese
Krankheiten zu bekommen, könnte bei vermehrter reaktiver Sauerstoffradikalen-
Bildung erhöht sein. Es ist klinisch jedoch schwer dies abzugrenzen, da die

2
Patienten meist noch zusätzliche Risikofaktoren für z.B. Herzinfarkt (Diabetes
mellitus, Adipositas, Hypertonie, Rauchen) aufweisen.
Durch die Gewinnung von mononukleären Leukozyten aus dem Blut von
Patienten, kann selektiv in vitro die Sauerstoffradikalenfreisetzung stimuliert und
mittels fluoreszenzspektrophotometrischer Messung bestimmt werden. In einem
zweiten Arbeitsgang kann man unter Zugabe verschiedener Substanzen (z.B.
von Scavengern oder bestimmten Medikamenten) versuchen, die gebildeten
Sauerstoffradikale zu vermindern.
In anderen Studien wurde bereits gezeigt, daß biologische Scavenger vor
Radikalen schützen. So wurde z.B. von Ballmer (Ballmer et al., 1988)
beschrieben, daß Allopurinol durch Blockierung der Xanthinoxidasereaktion die
Bildung von Superoxidradikalanionen (O2-) und von Wasserstoffperoxid (H2O2)
verhindert. Im Tierversuch wurden die postischämisch entstandenen Radikale
vermindert.
Aber auch der Körper selbst verfügt über einige Enzyme, die die reaktiven
Sauerstoffradikalen vermindern können, wie die Superoxiddismutase (SOD), die
Catalase und die Glutathionperoxidase. Treten öfters hypoxische Phasen auf,
kann dies laut Duan (Duan et al., 1998) zu einer Toleranzentwicklung des
Körpers gegenüber diesen Phasen führen, wobei die reaktiven Sauerstoff-
radikale (ROS) und ihre spezifischen Scavengerenzyme eine Rolle spielen.
Die Durchführung von fluoreszenzspektrophotometischen Bestimmungen
unterschiedlichster Art wird seit vielen Jahren angewandt. Es können direkt
fluoreszierende Substanzen aber auch mit einem Fluoreszenzfarbstoff
angefärbte Substanzen gemessen werden. So wendeten bereits Orie (Orie et al.,
1999), Nofer (Nofer et al., 1999) und Neusser (Neusser et al., 1999) die Methode
der Fluoreszenzspektrophotometrie für ihre Studien an.

3
1.2. Sauerstoffradikale und Calcium
Unter " Radikal " versteht man eine atomare oder molekulare Einheit, die ein
oder mehrere ungepaarte Elektronen auf der äußeren Hülle besitzen. Radikale
können andere Moleküle durch Entzug eines Elektrons oxidativ schädigen. So
kann z.B. die DNA verändert werden.
Reaktive Sauerstoffradikale mit potentiell schädigender Wirkung sind das
Superoxidradikal Anion (O2-), das Hydroxylradikal (OH⋅), das Wasserstoffperoxid-
molekül (H2O2), verschiedene Superoxidradikale (RO⋅, R = Fettsäure) und der
Singulettsauerstoff (1/2 O2).
Abbildung 1: Schematische Darstellung der Entstehung freier Sauerstoffradikale
und des Wirkortes der Enzyme, welche eine "Scavenger" Funktion haben, von
Pfaffendorf aus " Sauerstoffradikale und Arteriosklerose "
Sauerstoffradikale können im Körper große Schäden auslösen, so verursachen
sie eine Depolymerisation von Bindegewebe, Denaturierung von Enzymen,
Schädigung von Zellmembranen mit Gefäßpermeabilitätsveränderungen und
nachfolgender Zellschädigung (Man kennt diesen Vorgang auch als "ranzig-
werden" von Fetten und Ölen). Ist die DNS mitbetroffen, so geht die Zelle
zugrunde oder aber es treten Strangabbrüche mit der Gefahr der fehlerhaften
Reparatur und der Begünstigung von Mutationen auf. Dieser Mechanismus ist für

4
die krebserregende, aber auch für die therapeutisch genutzte zellzerstörende
Wirkung energiereicher Strahlung und einiger Zytostatika verantwortlich. So
bilden z.B. die Zytostatika Bleomycin und Doxorubicin in vitro und in vivo spontan
Radikale. Durch die energiereiche Strahlung entstehen z.B. durch homolytische
Spaltung von Wasser ein Hydrogen- und ein Hydroxydradikal.
Andererseits nutzt der Körper aber auch die freien Radikale, so z.B. zur Abwehr
von Krankheitserregern. Makrophagen bilden während der Phagozytose große
Mengen an Superoxidradikal Anionen. Die Bedeutung für das menschliche
Leben wurde erst in den letzten Jahren im vollen Umfang erfaßt.
Die intrazelluläre Calciumregulation spielt eine große Rolle bei dem Ablauf
entscheidender zellulärer Prozesse, wie bei verschiedenen Metabolismen, der
Proliferation, der Proteinbiosynthese sowie bei exokrinen und endokrinen Funk-
tionen. Im Zytoplasma liegt eine Calciumionenkonzentration von ca. 0,01-
0,1µmol/l vor. Dies ist eine um mehrere Größenordnungen niedrigere Konzen-
tration als im Plasma, wo die Calciumionen etwa zur Hälfte an Proteinen
gebunden und zur anderen Hälfte in freier Form vorliegen. Durch primär-aktiven
Transport und sekundär-aktiven Gegentransport werden die Calciumionen aus
der Zelle ausgeschleußt. Der Einstrom erfolgt zum einen passiv (durch das
Konzentrationsgefälle) und zum anderen aktiv durch z.B. Aktionspotentiale oder
über second messengers. Durch die äußeren Reize werden die Calciumkanäle
vorübergehend geöffnet, die Calciumionen binden an bestimmte Proteine und
lösen so in der Zelle eine Reaktion aus.

5
1.3. Prinzip der Fluoreszenz und Durchführung der Messung
Manche Moleküle können Licht einer bestimmten Wellenlänge absorbieren.
Dabei wird ein Außenelektron eines bestimmten Atoms in ein höheres
Energieniveau angehoben. Kehrt dieses Elektron wieder auf sein Ausgangs-
niveau zurück, gibt es einen Teil der Energie in Wärme und einen Teil in Licht
längerer Wellenlänge (geringere Frequenz) und somit geringerer Energie ab
(Emission/Fluoreszenz). Dies geschieht nach außerordentlich kurzer Zeit.
Abbildung 2: Entstehung der Fluoreszenzstrahlung / Emission, von Kaiser und
Henning aus physikalischer Chemie
A-B : Ein Elektron absorbiert Energie und geht damit vom Zustand A nach B
über
B-C : Die aufgenommene Energie wird schnell wieder abgegeben. Dies
geschieht teilweise als Wärme
C-D : und teilweise als energieärmeres Licht
Die Fluoreszenz wird mit bestimmten Meßinstrumenten (z.B. Fluoreszenz-
Spektrophotometer) nachgehalten. Je mehr Moleküle eines fluoreszierenden
Stoffes in einer Probe vorhanden sind, um so mehr Licht wird absorbiert und um
so höher ist daraufhin die Emission/Fluoreszenz. Die Intensität der Fluoreszenz
kann dann umgerechnet werden und damit die Konzentration der jeweiligen
Stoffe ermittelt werden.
Das Licht, mit dem die Fluoreszenz angeregt wird, heißt Primärstrahlung, das
durch die Fluoreszenz erzeugte Licht nennt man Sekundärstrahlung.

6
Die Fluoreszenzspektrophotometrie ist eine Methode, mit der unter anderem
Blutbestanteile und deren Reaktion auf bestimmte Reize in definierter
Versuchsanordnung untersucht werden können. Zur Aussendung der Primär-
strahlung wird hier eine Xenon-Lampe benutzt. Die von der Xenon-Lampe
ausgehende Strahlung tritt durch einen Filter (Exzitationsfilter), der nur für
Strahlung einer definierten Wellenlänge durchlässig ist, und trifft dann als
sogenannte Exzitationsstrahlung auf die Probe. Diese regt die Moleküle in der
Probe in einen höheren Energiezustand an, der jedoch nicht stabil ist. Die
angeregten Moleküle geben ihre zusätzliche Energie ab, teilweise in Wärme und
teilweise indem sie Strahlung aussenden, und gelangen somit in ihr
Ausgangsniveau zurück (Fluoreszenz). Diese Strahlung gelangt durch einen
Emissionsfilter, der wiederum nur eine bestimmte Wellenlänge passieren läßt.
Diese trifft dann auf einen Detektor, der die Strahlung registriert.

7
Abbildung 3: Aufbau eines Fluorometers, von K. Dörner aus Klinische Chemie.
Das Licht einer Xenon-Lampe tritt durch einen Exzitationsfilter, der nur für
Strahlung einer definierten Wellenlänge durchlässig ist, und trifft dann als
sogenannte Exzitationsstrahlung auf die Probe. Dies regt die Moleküle in der
Probe in einen höheren Energiezustand an, der jedoch nicht stabil ist. Die
angeregten Moleküle geben ihre zusätzliche Energie ab, teilweise in Wärme und
teilweise indem sie Strahlung aussenden, und gelangen somit in ihr
Ausgangsniveau zurück (Fluoreszenz). Diese Strahlung gelangt durch einen
Emissionsfilter, der wiederum nur eine bestimmte Wellenlänge passieren läßt.
Diese trifft dann auf einen Detektor, der die Strahlung registriert.

8
1.4. Fragestellung der Arbeit
Patienten mit obstruktivem Schlaf-Apnoe-Syndrom weisen vermehrte, längere
nächtliche Atempausen auf. Dabei stellt sich die Frage: Kommt es durch diesen
Sachverhalt zu einer vermehrten oxidativen Schädigung des Gewebes durch
erhöhte reaktive Sauerstoffradikalenbildung?
In der Arbeit wurden folgende Fragestellungen bearbeitet:
1. Bestehen Veränderungen der Bildung von reaktiven Sauerstoffradikalen
(ROS) in mononukleären Leukozyten bei Patienten mit obstruktiver Schlaf-
Apnoe-Syndrom im Vergleich zu gesunden Kontrollpersonen?
2. Wie verändern ROS-Scavenger oder Angiotensin Converting Enzym
Inhibitoren die Bildung von reaktiven Sauerstoffradikalen in mononu-
kleären Leukozyten?
3. Über welchen Mechanismus führen Angiotensin Converting Enzym
Inhibitoren zu einer Hemmung der ROS-Bildung in mononukleären
Leukozyten?
4. Besteht ein Zusammenhang zwischen der Bildung von reaktiven
Sauerstoffradikalen und der Steigerung der intrazellulären freien
Calciumionenkonzentration?

9
2. Materialien und Methoden
Die reaktive Sauerstoffradikalenbildung (ROS) wurde fluoreszenzspektrophoto-
metisch mit dem Fluoreszenz-Spektrometer F 2000 in den mononukleären
Leukozyten von Patienten mit obstruktivem Schlaf-Apnoe-Syndrom (OSAS) und
von gesunden Kontrollpersonen gemessen. Die Untersuchungen wurde bei 13
Patienten mit OSAS und bei 18 gesunden Kontrollpersonen durchgeführt.
Desweiteren wurden Messungen zur Frage der Angriffspunkte der reaktiven
Sauerstoffradikale an der Zelloberfläche (z.B. am Angiotensin Subtyp AT-1- und
AT-2 Rezeptor), zur Reduktion der intrazellulären ROS-Konzentration sowie zur
Bestimmung der intrazellulären Calciumionenkonzentration vorgenommen.
Als intrazelluläre Farbstoffe wurde 2.7.-Dichlorofluorescein-diacetat (DCF-DA)
zur reaktiven Sauerstoffradikalenmessung und 1-[2-(5-Carboxyoxazol-2-yl-6-
aminobenzofuran-5-oxyl]-2´amino-5´-methylphenoxy-ethan-N,N,N´,N´-tetraessig-
säure (Fura-2) zur Calciumionenkonzentrationsmessung verwendet. Zur
Sauerstoffradikalenstimulation wurde entweder Phorbol-myristat-acetat (PMA)
oder Thapsigargin zu der jeweiligen Probe hinzugegeben. Gemessen wurde bei
einer Emissionswellenlänge von 534nm und einer Exzitationswellenlänge von
488nm über einen Zeitraum von 60 Sekunden nach 0, 30 und 60 Minuten
Einwirkzeit der zugegebenen Substanzen (ROS-Bildung) sowie bei einer
Emissionswellenlänge von 510nm und einer Exzitationswellenlänge von 340nm
und 380nm über einen Zeitraum von 400 Sekunden nach 2 Minuten Einwirkzeit
der zugegebenen Substanzen. Gleichzeitig wurde bei jeder Messung eine
Eichung mit Ionomycin und EGTA durchgeführt (Calciumionenkonzentration).

10
2.1. Patienten und Kontrollpersonen
Es wurden insgesamt 13 Patienten im Alter von 32 bis 68 Jahren mit gesichertem
obstruktivem Schlaf-Apnoe-Syndrom untersucht. Durch eine Polysomnographie-
untersuchung im Schlaflabor wurde die Diagnose bestätigt. Zur klinischen und
biochemischen Charakterisierung der Patienten wurden verschiedene Daten z.B.
Lebensalter, Blutdruck/arterielle Hypertonie, Puls, einige Laborwerte und der
Bodymass-Index (BMI) gemessen, statistisch ausgewertet und gegen die Daten
der gesunden Kontrollpersonen in einer Tabelle eingetragen (Tabelle1). Die
Daten sind Durchschnittswerte der einzelnen Parameter für das jeweilige
Kollektiv.
Als Kontrollgruppe wurden 18 gesunde Personen im Alter von 24 bis 85 Jahren
untersucht. Auch bei diesen Personen wurden klinische und biochemische Daten
erhoben, die in Tabelle1 zusammengefaßt sind.

11
Tabelle1: Klinische und biochemische Daten von Patienten mit obstruktivem
Schlaf-Apnoe-Syndrom im Vergleich zu gesunden Kontrollpersonen
Patienten mit OSAS Gesunde Kontrollperson
Alter (Jahre) 53,8±10,0 51,1±20,6
Anzahl 13 18
Bodymass-Index (kg/m2) 33,2±5,0 23,2±3,7
Puls (Schläge/Minute) 75±9,3 85±22,8
Blutdruck
Systolisch (mmHg) * 128±17,6 130±16,9
Diastolisch (mmHg) * 76±9,2 75±10,4
Leukozyten (g/l) 5,7±1,0 9,4±3,7
Hämoglobin (g/dl) 15,4±0,9 13,5±2,1
Thrombozyten (g/l) 222±37,0 263±66,3
Kreatinin (mg/dl) 0,84±0,1 1,91±1,7
Harnstoff (mg/dl) 37±4,3 61±32,8
Calcium (mmol/l) 2,30±0,1 2,39±0,2
Kalium (mmol/l) 4,1±0,4 4,23±0,5
Natrium (mmol/l) 141±2,5 138±3,0
Hypertonie (%) 53,8 16,7
Koronare Herzkrankheit (%) 23,1 11
Diabetes mellitus (%) 8,0 33
Raucher (%) 23,1 11
* bei arterieller Hypertonie mit Antihypertonika eingestellt

12
2.2. Blutentnahme, Präparation von mononukleären Leukozyten und
Versuchsablauf
Es wurden jeweils 15-20ml Vollblut bei Patienten mit obstruktivem Schlaf-Apnoe-
Syndrom (OSAS) sowie bei gesunden Kontrollpersonen entnommen und zur
Gerinnungshemmung mit 1ml Heparin (Heparin-Natrium von Merck, Darmstadt)
versetzt. Zur Präparation der mononukleären Leukozyten wurde das Vollblut 15
Minuten bei 2000U/min mit der Zentrifuge von Heraeus (Heraeus Holding GmbH)
zentrifugiert , das überstehende Plasma verworfen und die Zellen 1:1 mit HBSS-
Lösung (Hund´s balanced salt sodium mit: Natriumchlorid, 7,94g/L,
Kaliumchlorid, 0,40g/L, Kaliumdihydrogenphosphat, 0,06g/L, D-Glucose, 1,0g/L,
Calciumchlorid, 0,147g/L, Hepes, 2,383g/L, Natriumdihydrogenphosphat, 0,06g/L
und Magnesiumchlorid, 0,203g/L bei einem pH-Wert von 7,40) vermischt. Nun
wurde jeweils 3ml Histopaque (Sigma-Aldrich Company, Taufbeuren) in einem
Reagenzglas vorgelegt, 5ml der Blut/HBSS-Mischung darübergeschichtet und
erneut 15 Minuten bei 2000U/min. zentrifugiert. Dadurch wurden die Leukozyten
von den übrigen Blutzellen getrennt und konnten mittels einer Pipette isoliert
werden.
Blutprobe
↓ +Heparin
↓ +Zentrifugation
Blutzellen
↓ +HBSS 1:1
↓ +Histopaque
↓ +Zentrifugation
mononukleäre Leukozyten
Abbildung 4: Präparation der mononukleären Leukozyten

13
2.2.1. Fluoreszenzmessung von reaktiven Sauerstoffradikalen
Nach dem Waschen der mononukleären Leukozyten wurden diese mit 2ml
HBSS-Lösung aufsuspendiert, mit 20µl DCF-DA (2.7.-Dichlorofluorescein-
diacetat) gefärbt und für 20 Minuten bei 37°C im Wasserbad inkubiert. Nach
erneuter Zentrifugation bei 2000U/min für 5 Minuten und Isolation der
mononukleären Leukozyten wurden diese mit 12ml HBSS versetzt, jeweils 1ml
der Suspension in eine 10x10mm Quarzglasküvette von der Firma Helma
gegeben, die zumessende Substanz zugegeben und nach 0, 30 und 60 Minuten
wurde die Probe fluoreszenzspektrophotometrisch (Fluoreszenz-Spektrometer F
2000 Hitachi Ltd. Tokyo, Japan) in einem Intervall von 2 Hz über 60 Sekunden
bei einer Exzitationswellenlänge von 488nm und einer Emissionswellenlänge von
534nm gemessen.
2.2.2. Fluoreszenzmessung von Calcium
Bei dieser Messung wurden die mononukleären Leukozyten nach dem Waschen
mit 2ml HBSS-Lösung aufsuspendiert und mit 20µl des Farbstoffes Fura-2 ( 1-[2-
(5-Carboxyoxazol-2-yl-6-aminobenzofuran-5-oxyl]-2´amino-5´-methylphenoxy-
ethan-N,N,N´,N´-tetraessigsäure ) und 10µl Pluronic versetzt und für 45 Minuten
bei 37°C im Wasserbad inkubiert. Nach der Zentrifugation für 5 Minuten bei
2000U/min und der Isolation der mononukleären Leukozyten wurde das
Reagenzglas mit 12ml HBSS aufgefüllt, je 1ml dieser Suspension in eine
10x10mm Quarzglasküvette gegeben und über 400 Sekunden mit einem Intervall
von 0,5 Sekunden bei den beiden wechselden Exzitationswellenlänge von 340
und 380nm und einer Emissionswellenlänge von 510nm gemessen.
Die jeweilige Substanzzugabe erfolgte 2 Minuten vor Meßbeginn. 50 Sekunden
nach dem Start der Messung wurden 10µl Phorbol-myristat-acetat (PMA), 150
Sekunden später, zur Eichung der Messung, 10µl Ionomycin und weitere 100
Sekunden danach 50µl EGTA hinzugegeben. Auch hier wurde das Fluoreszenz-
Spektrometer F2000 Hitachi Ltd. verwendet.
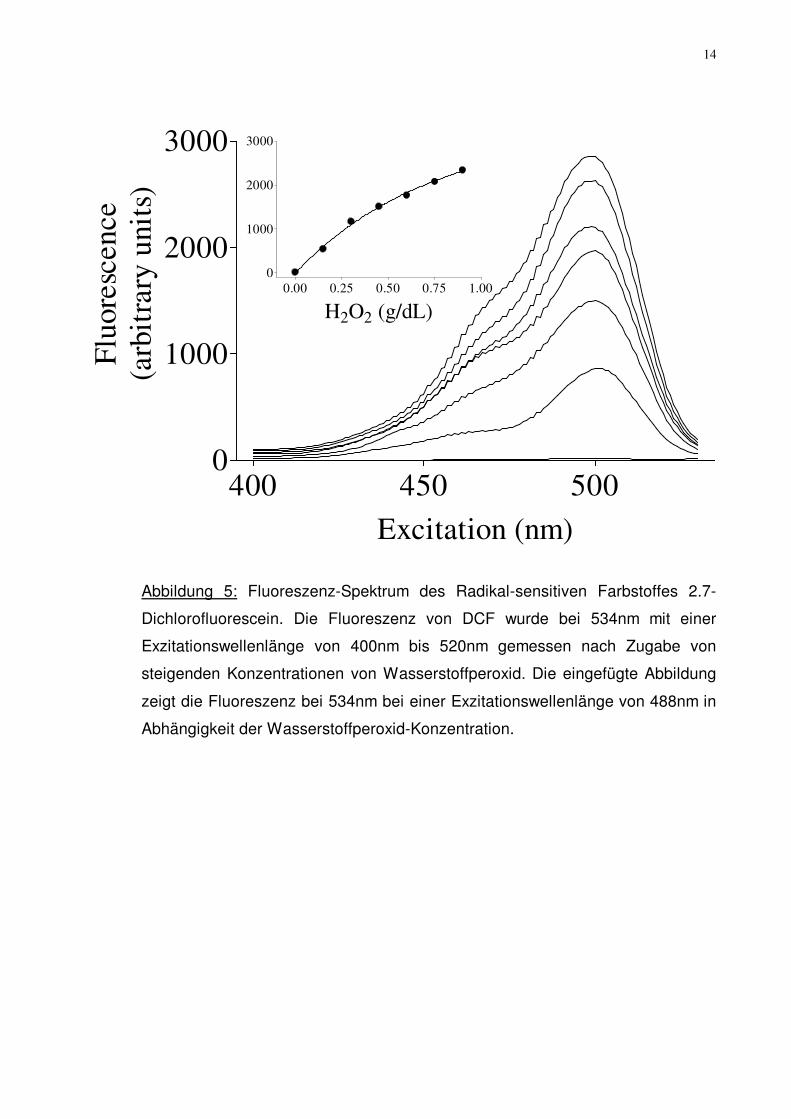
14
Abbildung 5: Fluoreszenz-Spektrum des Radikal-sensitiven Farbstoffes 2.7-
Dichlorofluorescein. Die Fluoreszenz von DCF wurde bei 534nm mit einer
Exzitationswellenlänge von 400nm bis 520nm gemessen nach Zugabe von
steigenden Konzentrationen von Wasserstoffperoxid. Die eingefügte Abbildung
zeigt die Fluoreszenz bei 534nm bei einer Exzitationswellenlänge von 488nm in
Abhängigkeit der Wasserstoffperoxid-Konzentration.
400 450 5000
1000
2000
3000
Excitation (nm)
Fluo
resc
ence
(arb
itrar
y un
its)
0.00 0.25 0.50 0.75 1.000
1000
2000
3000
H2O2 (g/dL)

15
2.3. Statistik
Zum Vergleich der Ergebnisse wurde der U-Test von Wilcoxon, Mann und
Whitney benutzt. Dieser Test für unverbundene Stichproben ist verteilungs-
unabhängig. Man bringt die (m+n) Stichprobenwerte in einer gemeinsamen
aufsteigenden Rangfolge. Die Summe der Rangzahlen von Stichprobe 1 sei R1,
die von Stichprobe 2 sei R2.
U1=mxn+0,5xmx(m+1)-R1
U2=mxn+0,5xnx(n+1)-R2
U1+U2=mxn
Die Nullhypothese wird verworfen, wenn U als der kleinere Wert von U1 und U2
kleiner oder gleich dem tabellierten kritischen Wert U (m,n;a) ist.
Die statistische Auswertung erfolgte mit der Computer Software Graph Pad
Prism Version 3,0 (Graph Pad, San Diego, USA).
Die Daten sind dargestellt als Mittelwerte ± Standardfehler des Mittelwertes
(SEM).

16
3. Ergebnisse
Zum einen wurde in dieser Arbeit untersucht, ob ein Unterschied in der reaktiven
Sauerstoffradikalenbildung (ROS) von Patienten mit obstruktivem Schlaf-Apnoe-
Syndrom (OSAS) und einer gesunden Kontrollgruppe besteht. Zum anderen
wurden verschiedene Substanzen getestet, die die Sauerstoffradikalenbildung
hemmen oder fördern könnten, um so einen möglichen Reaktionsmechanismus
und/oder -weg herauszufinden (insbesondere den Reaktionsmechanismus über
den Angiotensin Subtyp AT-1- und AT-2-Rezeptor) und als dritte Untersuchung
wurde der Calciumgehalt in den mononukleären Leukozyten gemessen.
Die Ergebnisse dieser Untersuchungen wurden in mehreren Abbildungen
gegliedert. Bei den einzelnen Messungen wurde die gemessene Fluoreszenz
meist zu einem Kontrollwert in Relation gesetzt (entweder zu den mit Phorbol-
myristat-acetat (PMA) stimulierten mononukleären Leukozyten oder zu der
spontanen reaktiven Sauerstoffradikalen(ROS)-Entwicklung [ohne Stimulation]).
Dabei wurde der Kontrollwert als 100% gewertet und die jeweiligen Ergebnisse
im Verhältnis dazu gesehen.
3.1. Reaktive Sauerstoffradikale bei Patienten mit obstruktivem Schlaf-
Apnoe-Syndrom und bei gesunden Kontrollpersonen
Um die benötigte Konzentration zur maximalen Hemmung der reaktiven
Sauerstoffradikalenbildung (ROS) in mononukleären Leukozyten herauszufinden,
wurden den Proben unterschiedliche Konzentrationen der jeweiligen Substanz,
hier als Beispiel Captopril und Natriumazid (NaN3), zugegeben. Es wurden
jeweils eine Kontrollmessung ohne Substanzzugabe und 6 Messungen mit
steigender Konzentration unter Stimulation mit dem Radikalenbildner Phorbol-
myristat-acetat (PMA) durchgeführt.
Abbildung 6 zeigt die Natriumazidmessung (n=6). Es wurden Endkonzen-
trationen von 0,01 bis 1,0µmol/L eingesetzt. Die maximale Hemmung der
reaktiven Sauerstoffradikalenbildung (ROS) nach Stimulierung mit 10µmol/L
Phorbol-myristat-acetat (PMA) wurde bei einer Endkonzentration von ca.
0,1µmol/L erreicht. Die PMA induzierte ROS-Bildung ließ sich auf 29,3%±3,1%
nach 60 Minuten vermindern.

17
Abbildung 7 zeigt die Messung mit Captopril (n=6). Es wurden Endkonzentra-
tionen von 0,1 bis 10,0µmol/L eingesetzt. Die maximale Hemmung der ROS-
Bildung nach Stimulierung mit 10µmol/L PMA wurde bei einer Endkonzentration
von ca. 5,0µmol/L erreicht. Die PMA induzierte ROS-Bildung ließ sich auf
65,4%±4,6% nach 60 Minuten vermindern.
Um zu zeigen, das die gemessene reaktive Sauerstoffradikalen-Bildung in den
mononukleären Leukozyten geschieht, erfolgten Leermessungen. Dabei wurde
einerseits HBSS und andererseits Plasma mit dem Farbstoff Dichlorofluorescein-
diacetat (DCF-DA) versetzt und die reaktive Sauerstoffradikalen-Bildung (ROS)
über 1 Stunde kontrolliert. Es konnte weder bei der HBSS- noch bei der
Plasmamessung ein Anstieg der reaktiven Sauerstoffradikale (ROS) festgestellt
werden. Daher ist davon auszugehen, daß der ROS-Anstieg in den Proben in
den mononukleären Leukozyten stattfindet.

18
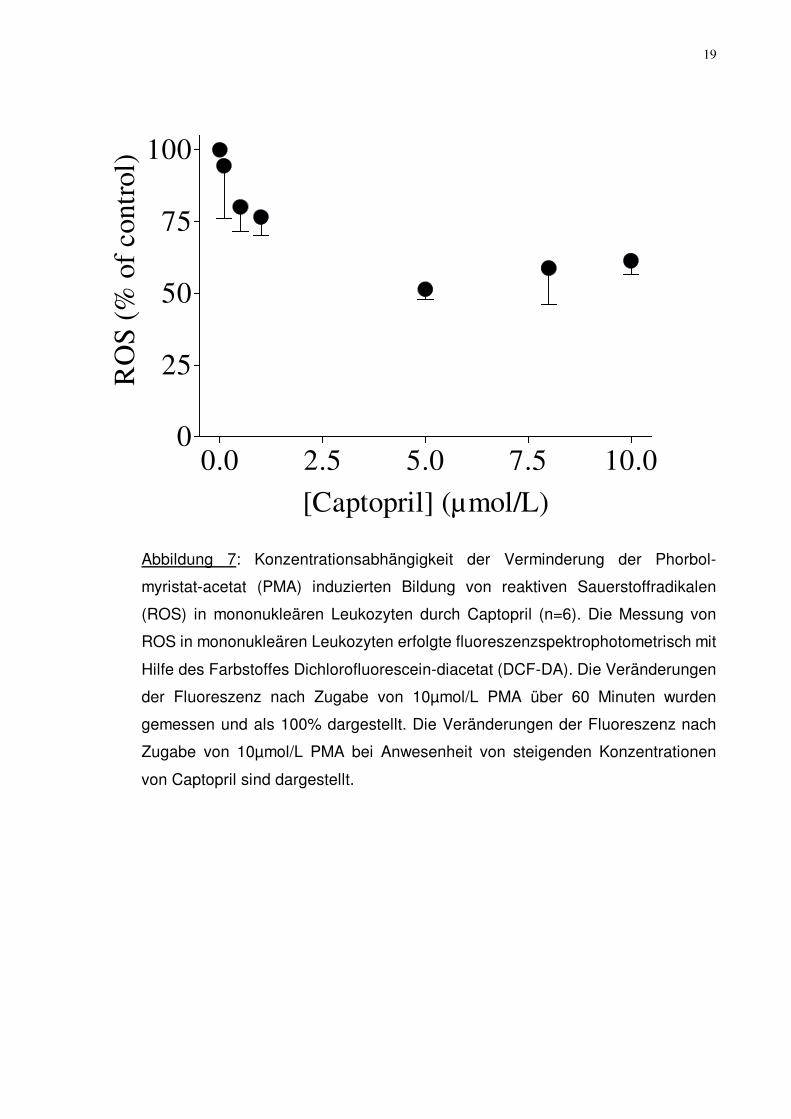
Abbildung 6: Konzentrationsabhängigikeit der Verminderung der Phorbol-
myristat-acetat (PMA) induzierten Bildung von reaktiven Sauerstoffradikalen
(ROS) in mononukleären Leukozyten durch Natriumazid (n=6). Die Messung von
ROS in mononukleären Leukozyten erfolgte fluoreszenzspektrophotometrisch mit
Hilfe des Farbstoffes Dichlorofluorescein-diacetat (DCF-DA). Die Veränderungen
der Fluoreszenz nach Zugabe von 10 µmol/L PMA über 60 Minuten wurden
gemessen und als 100% dargestellt. Die Veränderungen der Fluoreszenz nach
Zugabe von 10 µmol/L PMA bei Anwesenheit von steigenden Konzentrationen
von Natriumazid (NaN3) sind dargestellt.
0.00 0.25 0.50 0.75 1.000
25
50
75
100
[NaN3] (µmol/L)
RO
S (%
of c
ontr
ol)
19
Abbildung 7: Konzentrationsabhängigkeit der Verminderung der Phorbol-
myristat-acetat (PMA) induzierten Bildung von reaktiven Sauerstoffradikalen
(ROS) in mononukleären Leukozyten durch Captopril (n=6). Die Messung von
ROS in mononukleären Leukozyten erfolgte fluoreszenzspektrophotometrisch mit
Hilfe des Farbstoffes Dichlorofluorescein-diacetat (DCF-DA). Die Veränderungen
der Fluoreszenz nach Zugabe von 10µmol/L PMA über 60 Minuten wurden
gemessen und als 100% dargestellt. Die Veränderungen der Fluoreszenz nach
Zugabe von 10µmol/L PMA bei Anwesenheit von steigenden Konzentrationen
von Captopril sind dargestellt.
0.0 2.5 5.0 7.5 10.00
25
50
75
100
[Captopril] (µmol/L)
RO
S (%
of c
ontr
ol)

20
Die fluoreszenzspektrophotometrische Messung von Patienten mit obstruktivem
Schlaf-Apnoe-Syndrom und von den gesunden Kontrollpersonen wurde jeweils
über 60 Minuten beobachtet. Dabei wurde jede Probe 3 Mal (nach 0, 30, 60
Minuten) für je 60 Sekunden in das Fluoreszenzspektrophotophotometer
eingebracht und die gemessene Fluoreszenz gemittelt. Dabei konnte ein
kontinuierlicher Anstieg der reaktiven Sauerstoffradikale (ROS) über 60 Minuten
nachgewiesen werden.
In Abbildung 8 ist die kontinuierliche ROS-Entstehung wahlweise für die
Substanzen Phorbol-myristat-acetat (PMA), Captopril plus PMA und Natriumazid
(NaN3) plus PMA dargestellt (n=13). Es wurde jeweils eine Endkonzentration von
10µmol/L PMA und Captopril sowie von 1µmol/L Natriumazid eingesetzt.
In Abbildung 9 und 10 ist die reaktive Sauerstoffradikalenbildung in den
mononukleären Leukozyten von Patienten mit obstruktivem Schlaf-Apnoe-
Syndrom (n=13) gegen die der gesunden Kontrollpersonen (n=18) im direkten
Vergleich aufgetragen. Dabei zeigt Abbildung 9 die spontane und Abbildung 10
die mit Phorbol-myristat-acetat (PMA) induzierte Radikalbildung. Es erfolgte ein
spontaner Anstieg der intrazellulären ROS-Konzentration in den mononukleären
Leukozyten nach 30 Minuten von 2,8±0,4 bei den Patienten mit OSAS sowie von
3,9±0,6 bei den gesunden Kontrollpersonen. Nach Zugabe von 10ml PMA zeigte
sich ein Anstieg der intrazellulären ROS-Konzentration nach 30 Minuten von
10,4±1,6 bei den Patienten mit OSAS sowie von 13,8±1,5 bei den gesunden
Kontrollpersonen. In keiner der beiden Meßreihen konnte ein signifikanter
Unterschied zwischen den beiden Gruppen gezeigt werden.
21
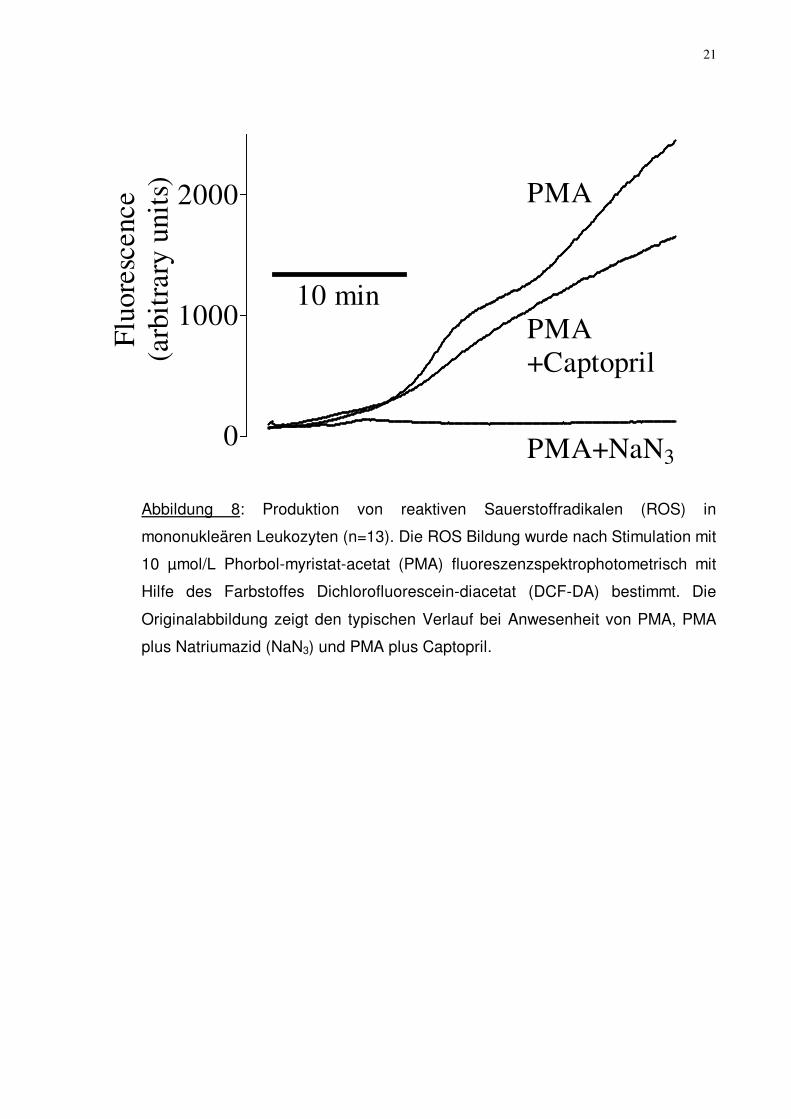
Abbildung 8: Produktion von reaktiven Sauerstoffradikalen (ROS) in
mononukleären Leukozyten (n=13). Die ROS Bildung wurde nach Stimulation mit
10 µmol/L Phorbol-myristat-acetat (PMA) fluoreszenzspektrophotometrisch mit
Hilfe des Farbstoffes Dichlorofluorescein-diacetat (DCF-DA) bestimmt. Die
Originalabbildung zeigt den typischen Verlauf bei Anwesenheit von PMA, PMA
plus Natriumazid (NaN3) und PMA plus Captopril.
0
1000
2000
10 min
Fluo
resc
ence
(arb
itrar
y un
its) PMA
PMA+Captopril
PMA+NaN3

22
Abbildung 9: Produktion von reaktiven Sauerstoffradikalen (ROS) in
mononukleären Leukozyten. Es wurde die spontane ROS Entwicklung (ohne
Stimulation) über einen Zeitraum von 60 Minuten fluoreszenzspektrophoto-
metrisch mit Hilfe des Farbstoffes Dichlorofluorescein-diacetat (DCF-DA)
bestimmt. Die Abbildung zeigt den Vergleich der ROS-Bildung von Patienten mit
obstruktivem Schlaf-Apnoe-Syndrom (OSAS, n=13) zu einer gesunden
Kontrollgruppe (Control, n=18). Der Ausgangswert der ROS-Konzentration ist mit
100% angesetzt.
Control OSAS0
250
500R
OS-
incr
ease
(%)
23
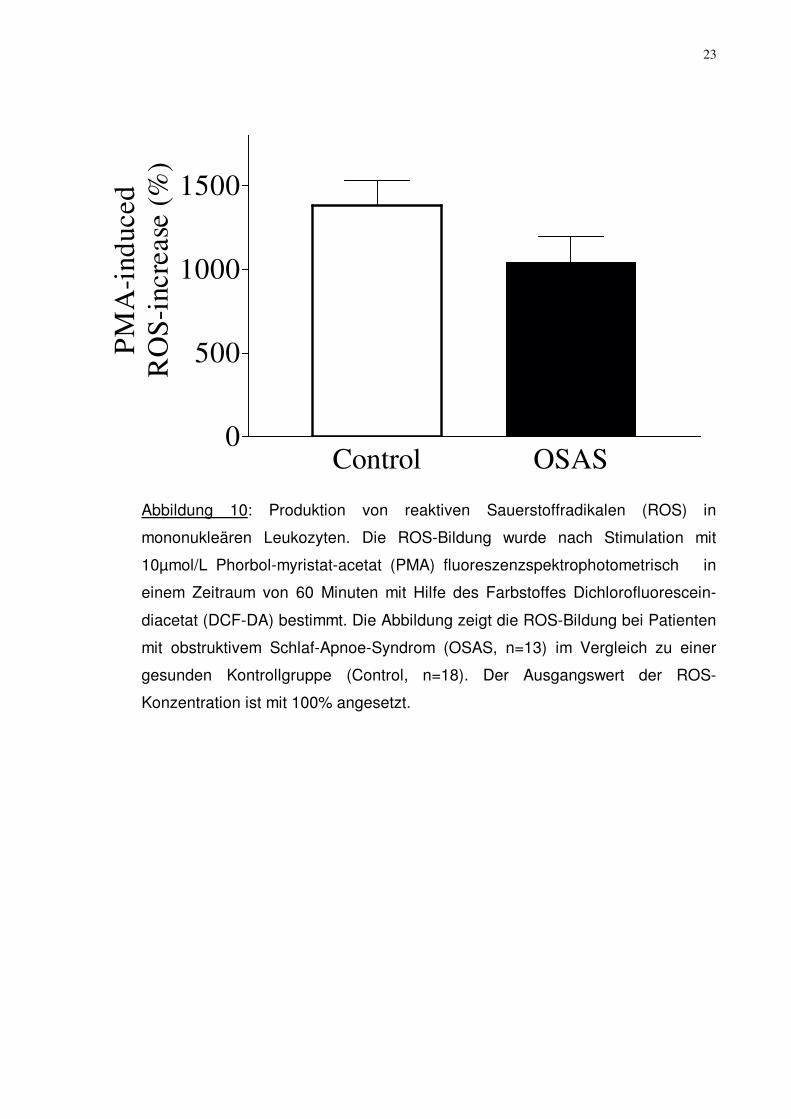
Abbildung 10: Produktion von reaktiven Sauerstoffradikalen (ROS) in
mononukleären Leukozyten. Die ROS-Bildung wurde nach Stimulation mit
10µmol/L Phorbol-myristat-acetat (PMA) fluoreszenzspektrophotometrisch in
einem Zeitraum von 60 Minuten mit Hilfe des Farbstoffes Dichlorofluorescein-
diacetat (DCF-DA) bestimmt. Die Abbildung zeigt die ROS-Bildung bei Patienten
mit obstruktivem Schlaf-Apnoe-Syndrom (OSAS, n=13) im Vergleich zu einer
gesunden Kontrollgruppe (Control, n=18). Der Ausgangswert der ROS-
Konzentration ist mit 100% angesetzt.
Control OSAS0
500
1000
1500
PMA
-ind
uced
RO
S-in
crea
se (%
)

24
3.2. Wirkung von Scavengern und ACE-Hemmern auf die Bildung
reaktiver Sauerstoffradikale
Zur Überprüfung der gefundenen Werte sowie zur fraglichen Unterscheidung von
Patienten mit obstruktivem Schlaf-Apnoe-Syndrom gegenüber den gesunden
Kontrollpersonen in der Bildung von unterschiedlichen reaktiven
Sauerstoffradikalen wurden einige Substanzen den Proben zugesetzt, die
bekanntermaßen die Bildung bestimmter Sauerstoffradikalengruppen
unterdrücken.
Die Superoxiddismutase (SOD) führt zu einer verminderten Konzentration des
Superoxidradikal Anions (O2-), die Catalase führt zu einer verminderten
Konzentration des Wasserstoffperoxidmoleküls (H2O2), Natriumazid (NaN3) führt
zu einer verminderten Konzentration von Singulettsauerstoff (1/2 O2) und
Ebselen führt zu einer verminderten Konzentration von Peroxinitrit (ONOO-).
Ro-32 (2-[8 [(Dimethylamino)methyl]-6,7,8,9-tetrahydropyrido[1,2-a]indol-3-yl)-3-
(methyl-1H-indol-3yl)maleimide, HCl) wurde als Hemmstoff der Protein Kinase C
und Tyrphostin als Hemmstoff der Tyrphostin Kinase eingesetzt.
Außerdem wurde bei dem Vergleich von Patienten mit obstruktivem Schlaf-
Apnoe-Syndrom und von gesunden Kontrollpersonen noch die als Medikamente
verwendeten Angiotensin Converting Enzym Hemmer (ACE-Hemmer) Captopril
und Ramiprilat sowie der Angiotensin AT-1-Rezeptorantagonist Losartan (mit
Exp3174 als pharmakologisch aktivem Metaboliten) mit untersucht.
In Abbildung 11 und teilweise in Abbildung 12 sind die jeweiligen Gruppen nach
Zugabe einiger Scavenger-Substanzen und einiger Medikamente dargestellt.
Hier konnte sowohl in der Gruppe der Patienten mit obstruktivem Schlaf-Apnoe-
Syndrom (OSAS), Abbildung 11 (n=13), als auch in der Gruppe der gesunden
Kontrollpersonen, Abbildung 12 (n=6-18), eine Verringerung der reaktiven
Sauerstoffradikale (ROS) 60 Minuten nach Zugabe von Dimethylsulfoxid
(DMSO), Natriumazid (NaN3) , Ebselen, des Protein Kinase Inhibitors Tyrphostin,
des Protein Kinase C Inhibitors Ro-32, der ACE-Hemmer Captopril und
Ramiprilat und des Angiotensin AT-1-Rezeptorantagonisten Exp3174

25
nachgewiesen werden. Da Exp3174 kaum wasserlöslich ist, wurde DMSO als
Lösungsmittel verwendet und das Ergebnis in Relation zu DMSO gewertet.
Bei den Patienten mit obstruktivem Schlaf-Apnoe-Syndrom (OSAS) zeigte sich
eine Verminderung der reaktiven Sauerstoffradikalenkonzentration (ROS) nach
Zugabe von DMSO auf 48,3%±3,0%, von NaN3 auf 29,3%±3,1%, von Ebselen
auf 54,1%±3,7%, von Tyrphostin auf 60,8%±4,5%, von Ro-32 auf 26,4%±4,1%,
von Captopril auf 65,4%±4,6%, von Ramiprilat auf 78,5%±5,4% und von
Exp3174 auf 53,0%±10,6% (28,7%±10,6%) in Bezug auf den Kontrollwert
(jeweils p< 0,02).
Bei den gesunden Kontrollpersonen ging die ROS-Konzentration nach Zugabe
von DMSO auf 49,2%±4,5%, von NaN3 auf 25,1%±2,3%, von Ebselen auf
55,0%±4,3%, von Tyrphostin auf 59,2%±5,8%, von Ro-32 auf 20,3%±3,1%, von
Captopril auf 67,2%±5,6%, von Ramiprilat auf 71,7%±3,6% und von Exp3174 auf
63,0%±5,3% (31,0%±5,3%) in Bezug auf den Kontrollwert zurück (jeweils
p<0.02).
Im direkten Vergleich der beiden Gruppen konnte kein signifikanter Unterschied
festgestellt werden.
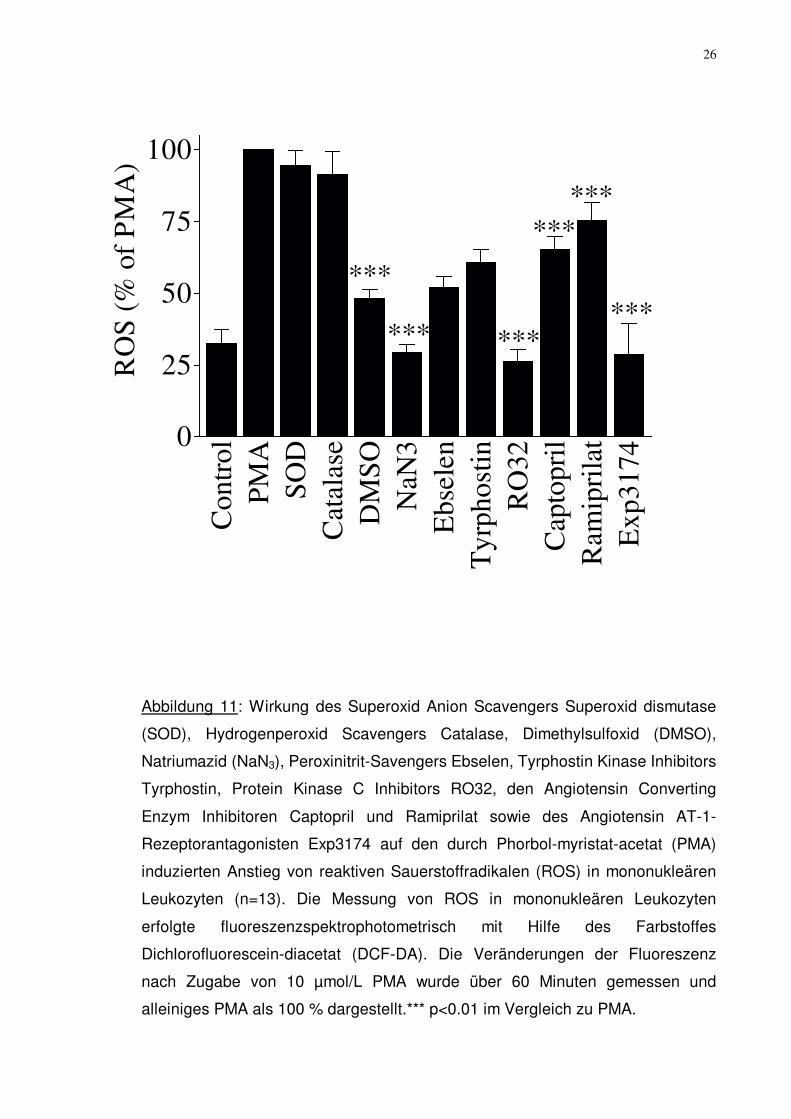
26
Abbildung 11: Wirkung des Superoxid Anion Scavengers Superoxid dismutase
(SOD), Hydrogenperoxid Scavengers Catalase, Dimethylsulfoxid (DMSO),
Natriumazid (NaN3), Peroxinitrit-Savengers Ebselen, Tyrphostin Kinase Inhibitors
Tyrphostin, Protein Kinase C Inhibitors RO32, den Angiotensin Converting
Enzym Inhibitoren Captopril und Ramiprilat sowie des Angiotensin AT-1-
Rezeptorantagonisten Exp3174 auf den durch Phorbol-myristat-acetat (PMA)
induzierten Anstieg von reaktiven Sauerstoffradikalen (ROS) in mononukleären
Leukozyten (n=13). Die Messung von ROS in mononukleären Leukozyten
erfolgte fluoreszenzspektrophotometrisch mit Hilfe des Farbstoffes
Dichlorofluorescein-diacetat (DCF-DA). Die Veränderungen der Fluoreszenz
nach Zugabe von 10 µmol/L PMA wurde über 60 Minuten gemessen und
alleiniges PMA als 100 % dargestellt.*** p<0.01 im Vergleich zu PMA.
Con
trol
PMA
SOD
Cat
alas
eD
MSO
NaN
3E
bsel
enT
yrph
ostin
RO
32C
apto
pril
Ram
ipril
atE
xp31
74
0
25
50
75
100
***
***
*** ******
***
RO
S (%
of P
MA
)

27
3.3. Wirkung am Angiotensinrezeptor
In einer weiteren Meßreihe wurde der Wirkungsmechanismus der reaktiven
Sauerstoffradikalenbildung und -hemmung in den mononukleären Leukozyten,
insbesondere die Wirkung an den Angiotensinrezeptoren (Subtyp AT-1/AT-2),
genauer untersucht. Dabei wurde das Blut gesunder Kontrollpersonen
verwendet. Da in der vorherigen Meßreihe bereits die ACE-Hemmer Captopril
und Ramiprilat und der Angiotensin AT-1-Rezeptorantagonist Exp3174
eingesetzt wurden, kamen nun noch die Angiotensin AT-2-Rezeptoragonisten
CGP42112 und Pyrrolidin-dithiocarbamat Ammonium (PDTC), der Angiotensin
AT-1-Rezeptoragonist Angiotensin l, der Angiotensin AT-1- und AT-2-
Rezeptoragonist Angiotensin ll, der Angiotensin AT-2-Rezeptorantagonist
PD123319, der Calciumantagonist Verapamil sowie ein weiterer ACE-Hemmer,
Lisinopril, hinzu. Wie in Abbildung 12 dargestellt, ließ sich hierbei zusätzlich zu
den obengenannten Substanzen, eine signifikante Verminderung der reaktiven
Sauerstoffradikalenkonzentration (ROS) bei Stimulation mit Phorbol-myristat-
acetat (PMA) bei dem Calciumantagonisten Verapamil auf 70,4%±4,5%, bei dem
Angiotensin AT-1- und AT-2-Rezeptoragonisten Angiotensin ll auf 82,6%±3,7%
und bei dem Angiotensin AT-2-Rezeptoragonisten CGP42112 auf 75,9%±3,0%
feststellen (jeweils p=0,03).
Lediglich der Angiotensin AT-1-Rezeptoragonist Angiotensin l führte zu einer
signifikanten Steigerung der ROS-Bildung auf 107,4%±1,2%.
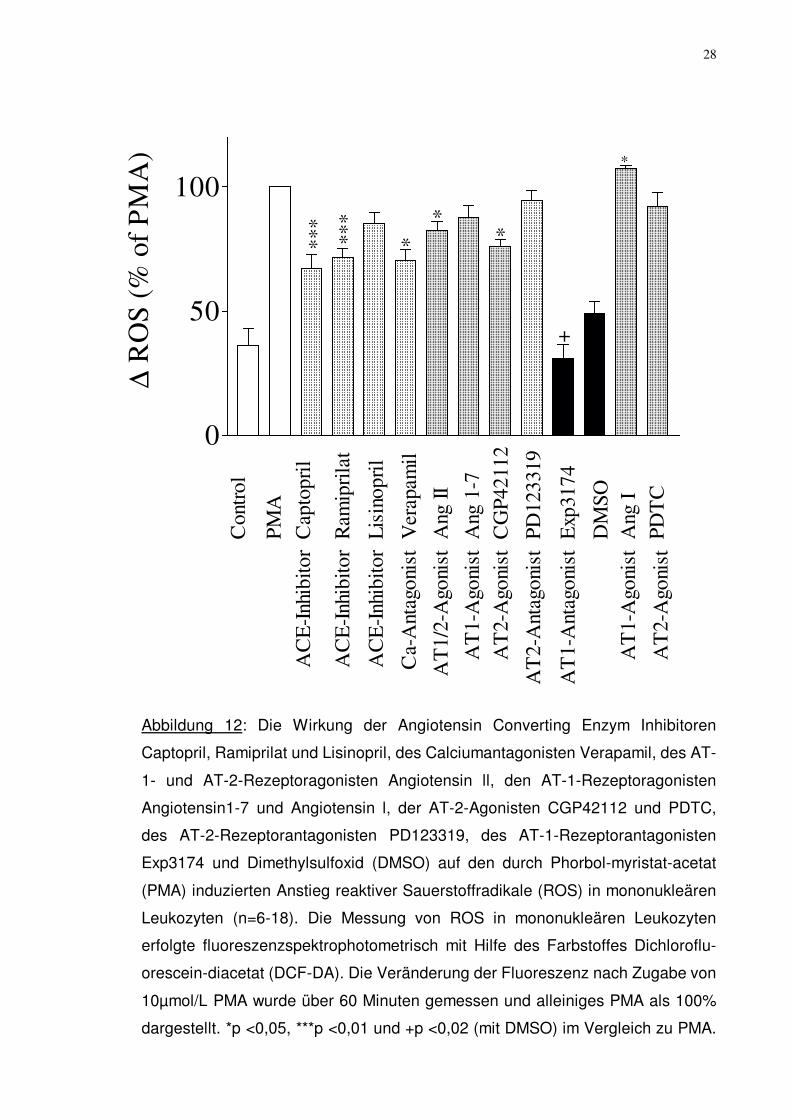
28
Abbildung 12: Die Wirkung der Angiotensin Converting Enzym Inhibitoren
Captopril, Ramiprilat und Lisinopril, des Calciumantagonisten Verapamil, des AT-
1- und AT-2-Rezeptoragonisten Angiotensin ll, den AT-1-Rezeptoragonisten
Angiotensin1-7 und Angiotensin l, der AT-2-Agonisten CGP42112 und PDTC,
des AT-2-Rezeptorantagonisten PD123319, des AT-1-Rezeptorantagonisten
Exp3174 und Dimethylsulfoxid (DMSO) auf den durch Phorbol-myristat-acetat
(PMA) induzierten Anstieg reaktiver Sauerstoffradikale (ROS) in mononukleären
Leukozyten (n=6-18). Die Messung von ROS in mononukleären Leukozyten
erfolgte fluoreszenzspektrophotometrisch mit Hilfe des Farbstoffes Dichloroflu-
orescein-diacetat (DCF-DA). Die Veränderung der Fluoreszenz nach Zugabe von
10µmol/L PMA wurde über 60 Minuten gemessen und alleiniges PMA als 100%
dargestellt. *p <0,05, ***p <0,01 und +p <0,02 (mit DMSO) im Vergleich zu PMA.
0
50
100*
Con
trol
PMA
Cap
topr
il
Ram
ipril
at
Lisi
nopr
il
Ver
apam
il
Ang
IIA
ng 1
-7C
GP4
2112
PD12
3319
Exp3
174
DM
SO
***
***
*
*
*
+
AC
E-In
hibi
tor
AT
1/2-
Ago
nist
AC
E-In
hibi
tor
AC
E-In
hibi
tor
AT
1-A
goni
stA
T2-
Ago
nist
AT
2-A
ntag
onis
t
AT
1-A
ntag
onis
t
Ang
IPD
TC
AT
1-A
goni
st
Ca-
Ant
agon
ist
AT
2-A
goni
st
∆ R
OS
(% o
f PM
A)

29
Wie in bereits erwähnt, erfolgte eine Abnahme der durch Phorbol-myristat-acetat
(PMA) induzierten reaktiven Sauerstoffradikalenbildung (ROS) nach Zugabe des
Angiotensin AT-2-Rezeptoragonisten CGP42112, des Angiotensin AT-1 und AT-
2-Rezeptoragonisten Angiotensin ll sowie des Calciumantagonisten Verapamil.
Dies legt die Vermutung nahe, daß die Regulierung der ROS-Entstehung bzw.
Hemmung über die AT-1- und/oder AT-2-Rezeptoren erfolgt.
Daraufhin wurden Messungen mit dem Angiotensin AT-1- und AT-2-Rezeptor-
agonisten Angiotensin ll ohne Zugabe von Phorbol-myristat-acetat (PMA)
durchgeführt (n=13). Dabei wurden steigende Endkonzentrationen von 1,0 bis
10000 µmol/L des AT-1- und AT-2-Rezeptoragonisten eingesetzt. Die alleinige
Zugabe von Angiotensin ll führte zu keiner signifikanten Änderung der ROS-
Produktion in den mononukleären Leukozyten.
Da Angiotensin ll eine AT-1- und eine AT-2-rezeptoragonistische Wirkung besitzt,
wurde bei einigen Messungen der Probe außer Angiotensin ll noch PD123319,
ein Angiotensin AT-2-Rezeptorantagonist, hinzugefügt, um die AT-2-
Rezeptorwirkung auszuschalten und die alleinige AT-1-Rezeptorwirkung des
Angiotensin ll zu testen (n=13). Wie in Abbildung 13 zu sehen ist, wurden hierbei
sowohl Messungen mit Stimulation der mononukleären Leukozyten durch
Phorbol-myristat-acetat (PMA) als auch ohne Zugabe von PMA durchgeführt. Bei
dieser Meßreihe konnte aber keine signifikante Änderung der Konzentration der
reaktiven Sauerstoffradikale (ROS) ermittelt werden. Es zeigte sich eine
Verminderung der ROS-Konzentration nach Zugabe von PD123319, Angiotensin
ll und PMA auf 90,8%+8,5%.
Des weiteren wurde die Wirkung von S-Nitroso-N-acetylpenicillinamin (SNAP)
auf die mononukleären Leukozyten bezüglich der reaktiven Sauerstoffradikale
(ROS) untersucht (n=7). SNAP ist ein Stickstoffmonoxid (NO) Donor. NO ist ein
relativ labiles Molekül, welches auch radikale Eigenschaften besitzt (daher kann
man das NO-Molekül nicht direkt einsetzen). In der Studie von Muriel (Muriel et
al., 2000) setzte dieser SNAP ein, um das kurzlebige NO-Molekül direkt am
gewünschten Wirkungsort zu erzeugen.

30
Die Untersuchung erfolgte zum einen mit Stimulation von Phorbol-myristat-acetat
(PMA) und zum anderen ohne Stimulation. Da SNAP nicht ausreichend in
Wasser löslich ist, wurde Dimethylsulfoxid (DMSO) als Lösungsmittel verwendet.
Dabei zeigte sich bei beiden Meßreihen eine signifikante Erhöhung der ROS-
Freisetzung gegenüber dem Kontrollwert.
Bei der ersten Meßreihe (ohne Zugabe von PMA) stieg die ROS-Konzentration
auf 102,7%±15,2% gegenüber 50,46%±11% nur mit dem Lösungsmittel DMSO.
Die zweite Meßreihe (mit Zugabe von PMA) ergab eine Erhöhung der ROS-
Konzentration auf 176,4%±33,9% gegenüber 67,6%±8,9% nur mit dem
Lösungsmittel DMSO.
Die angegebenen Werte sind in Abbildung 13 graphisch dargestellt.

31
Abbildung 13: Die Wirkung des Angiotensin AT-1-und AT-2-Rezeptoragonisten
Angiotensin ll in Gegenwart des Angiotensin AT-2-Rezeptorantagonisten
PD123319 (n=13) und des Stickstoff(2)oxid Donors S-Nitroso-N-acetylpenicillin-
amin (SNAP, n=7) auf die Konzentration reaktiver Sauerstoffradikale (ROS) in
mononukleären Leukozyten ohne und mit Induktion durch Phorbol-myristat-
acetat (PMA). Die Messung von ROS in den mononukleären Leukozyten wurde
fluoreszenzspektrophotometrisch mit Hilfe des Farbstoffes Dichlorofluoroscein-
diacetat (DCF-DA) bestimmt. Die Veränderung der Fluoreszenz nach Zugabe
von 10µmol/L PMA wurde über 60 Minuten gemessen und alleiniges PMA als
100% dargestellt.
** p < 0,01 im Vergleich zum Kontrollwert.
cont
rol
PMA
PD+A
ng
PD+A
ng+P
MA
DM
SO
SNA
P
PMA
+DM
SO
PMA
+SN
AP0
50
100
150
200
250
**
**
∆ R
OS
% o
f PM
A

32
3.4. Calciummessung und reaktive Sauerstoffradikalenmessung unter
Zugabe verschiedener Sauerstoffradikalenstimulatoren (PMA und
Thapsigargin)
Bei dieser Meßreihe wurde die reaktive Sauerstoffradikalenbildung durch zwei
verschiedene Stimulatoren ausgelöst. Zum einen durch das bis jetzt verwendete
Phorbol-myristat-acetat (PMA) zum anderen durch Thapsigargin. Hierbei sollte
geprüft werden, ob unterschiedliche Konzentrationen reaktiver Sauerstoffradikale
(ROS) entstehen. Außerdem wurde die Calciumionenkonzentration nach Zugabe
der verschiedenen Sauerstoffradikalenbildner gemessen.
Die Messung mit Thapsigargin erfolgte nach derselben Vorschrift wie die
Messung mit Phorbol-myristat-acetat (PMA). Auch hierbei wurden die ACE-
Hemmer Captopril und Ramiprilat, der Angiotensin AT-1-Rezeptorantagonist
EXP3174 und Dimethylsulfoxid (DMSO) jeweils mit einer Endkonzentration von
10µmol/L eingesetzt. Wie bereits in Abbildung 11 beschrieben zeigte sich eine
signifikante Verminderung der PMA induzierten reaktiven Sauerstoffradikalen
(ROS)-Konzentration bei den ACE-Hemmern wie auch bei dem Angiotensin AT-
1-Rezeptorantagonisten. Bei den Messungen mit dem Sauerstoffradikalen-
stimulator Thapsigargin konnte, wie in Abbildung 14 (n=6) zu sehen ist, nur eine
signifikante Verminderung der ROS-Bildung nach Zugabe des Angiotensin AT-1-
Rezeptorantagonisten Exp3174 auf 50,3%±7,4% nachgewiesen werden. Bei den
anderen Messungen zeigten sich keine signifikanten Veränderungen in der ROS-
Konzentration.
Die intrazelluläre Calciumionenkonzentrationsmessung wurde auch mit den
obengenannten Substanzen in gleicher Konzentration durchgeführt. Dabei
änderte sich die Calciumionenkonzentration signifikant nach Zugabe des
Angiotensin AT-1-Rezeptorantagonisten Exp3174 sowohl bei den durch PMA als
auch bei den durch Thapsigargin stimulierten mononukleären Leukozyten. Bei
der Messung mit PMA stieg die Calciumionenkonzentration auf 120,6%±49,4%
an und bei der Messung mit Thapsigargin fiel die Calciumionenkonzentration auf
34,6%±3,6% ab.
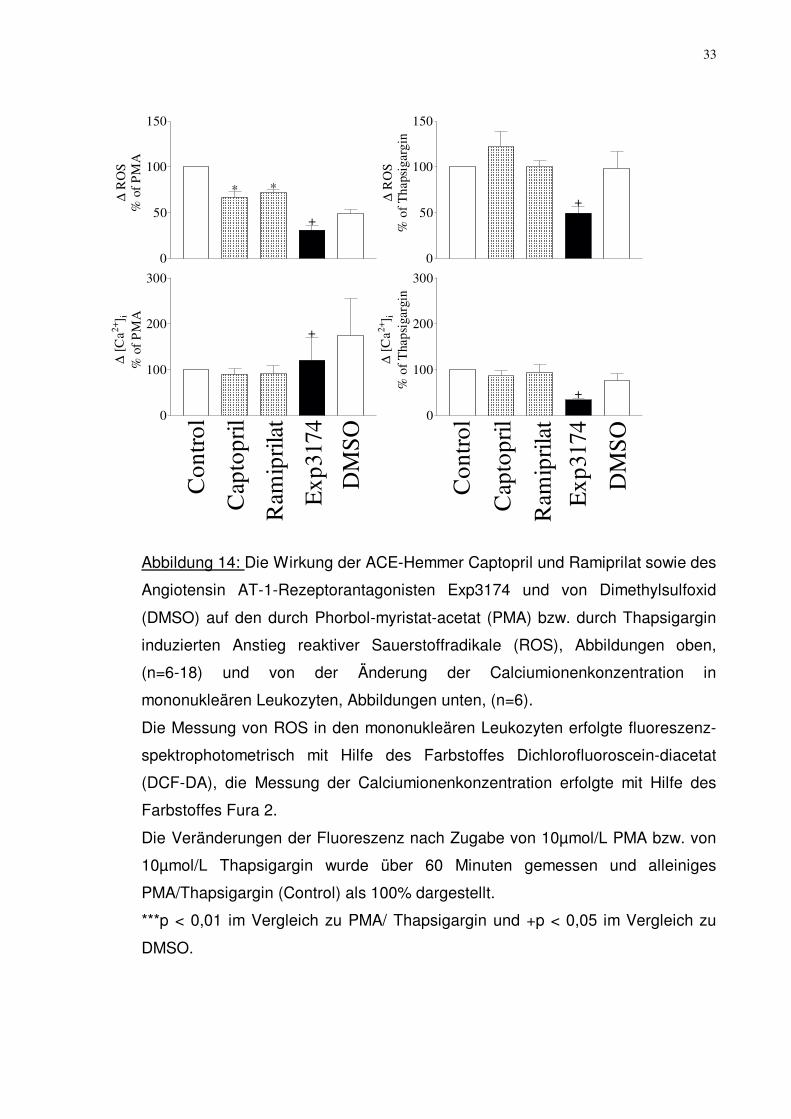
33
Abbildung 14: Die Wirkung der ACE-Hemmer Captopril und Ramiprilat sowie des
Angiotensin AT-1-Rezeptorantagonisten Exp3174 und von Dimethylsulfoxid
(DMSO) auf den durch Phorbol-myristat-acetat (PMA) bzw. durch Thapsigargin
induzierten Anstieg reaktiver Sauerstoffradikale (ROS), Abbildungen oben,
(n=6-18) und von der Änderung der Calciumionenkonzentration in
mononukleären Leukozyten, Abbildungen unten, (n=6).
Die Messung von ROS in den mononukleären Leukozyten erfolgte fluoreszenz-
spektrophotometrisch mit Hilfe des Farbstoffes Dichlorofluoroscein-diacetat
(DCF-DA), die Messung der Calciumionenkonzentration erfolgte mit Hilfe des
Farbstoffes Fura 2.
Die Veränderungen der Fluoreszenz nach Zugabe von 10µmol/L PMA bzw. von
10µmol/L Thapsigargin wurde über 60 Minuten gemessen und alleiniges
PMA/Thapsigargin (Control) als 100% dargestellt.
***p < 0,01 im Vergleich zu PMA/ Thapsigargin und +p < 0,05 im Vergleich zu
DMSO.
0
50
100
150
+
* *
∆ R
OS
% o
f PM
A
0
100
200
300
+
∆ [C
a2+] i
% o
f PM
A
0
50
100
150
∆ R
OS
% o
f Tha
psig
argi
n
+
0
100
200
300
∆ [C
a2+] i
% o
f Tha
psig
argi
n+
Con
trol
Cap
topr
il
Ram
ipril
at
Exp3
174
DM
SO
Con
trol
Cap
topr
il
Ram
ipril
atEx
p317
4
DM
SO

34
4. Diskussion
In dieser Arbeit wurde die Bildung reaktiver Sauerstoffradikale (ROS) in
mononukleären Leukozyten bei Patienten mit obstruktivem Schlaf-Apnoe-
Syndrom (OSAS) und im Vergleich dazu bei einer gesunden Kontrollgruppe
untersucht. Ferner wurden in dieser Arbeit mögliche Reaktionsmechanismen und
-wege der reaktiven Sauerstoffradikalen-(ROS)-bildung sowie -hemmung im
menschlichen Körper diskutiert. Besondere Aufmerksamkeit galt dabei dem
Angiotensin Subtyp AT-1- und AT-2-Rezeptor. Desweiteren wurde auch eine
mögliche Veränderung der Calciumkonzentration in den mononukleären
Leukozyten untersucht.
In geringer Konzentration sind reaktive Sauerstoffradikale (ROS) Nebenprodukte
von physiologischen metabolischen Prozessen in den Zellen, Orie (Orie et al.,
1999). Auch spielen sie bei der intrazellulären Signaltransduktion eine Rolle. Bei
höheren Konzentrationen werden die ROS jedoch für die Entstehung einer Reihe
von Erkrankung mitverantwortlich gemacht.
Bereits 1999 wurde in einer Untersuchung von Orie (Orie et al., 1999) die ROS-
Bildung bei Patienten mit arterieller Hypertonie sowie mit Diabetes mellitus
untersucht. Dabei konnte gezeigt werden, daß bei Patienten mit Diabetes
mellitus (Typ 2) sowohl die spontane ROS-Entwicklung als auch die durch
Phorbol-myristat-acetat (PMA) stimulierten Werte gegenüber einer gesunden
Kontrollgruppe signifikant erhöht waren. Dies gilt auch für Patienten mit einer
Niereninsuffizienz, wie in einer Untersuchung von Tepel (Tepel et al., 2000)
gezeigt werden konnte. In der jetzigen Arbeit wurden dann Patienten mit
obstruktivem Schlaf-Apnoe-Syndrom (OSAS) untersucht. Bei Patienten mit
OSAS ist eine erhöhte Inzidenz an Herz-Kreislauferkrankungen bekannt. Dies
konnte auch in dieser Studie im Verhältnis zu den gesunden Kontrollpersonen
gezeigt werden. Dabei fiel eine erhöhte Inzidenz bei der Hypertonie und bei der
koronaren Herzkrankheit (KHK) / Infarkten auf. Besonders deutlich ist der
Bodymass-Index (BMI) bei den Patienten mit OSAS mit durchschnittlich
33,2±5,0kg/m² gegenüber der gesunden Kontrollgruppe mit durchschnittlich
23,2±3,7kg/m² erhöht.

35
Bei der spontanen ROS-Bildung und unter Stimulation mit Phorbol-myristat-
acetat (PMA) konnte kein signifikanter Unterschied im Vergleich zwischen den
Patienten mit OSAS und der gesunden Kontrollgruppe nachgewiesen werden.
Nach Zugabe einiger Medikamente, den ACE-Hemmern Captopril bzw.
Ramiprilat und dem Angiotensin AT-1-Rezeptorantagonisten Exp3174
(Losartan), zu den mit Phorbol-myristat-acetat (PMA) stimulierten mononukleären
Leukozyten konnte in Bezug auf den PMA induzierten Kontrollwert eine
signifikante Verminderung der ROS-Bildung nachgewiesen werden. Es konnten
jedoch keine Unterschiede zwischen den beiden Untersuchungsgruppen
festgestellt werden.
In einer Veröffentlichung von Inagami (Inagami et al., 1998) wurden Nierenzellen
unter anderem auf Subtypen des Angiotensinrezeptors untersucht. Dabei wurden
2 Subtypen, der AT-1-Rezeptor und der AT-2-Rezeptor, unterschieden, die eine
antagonistische Wirkung zueinander zeigten. Es konnte bewiesen werden, daß
die Stimulation von AT-1-Rezeptoren und AT-2-Rezeptoren unterschiedliche
Wirkung auf den Blutdruck von Mäusen haben (über AT-1-Rezeptoren wird der
Blutdruck gesteigert und über AT-2-Rezeptoren wird der Blutdruck gesenkt),
sowie auf die Diurese und Natriurese in Rattennieren (der Angiotensin AT-2-
Rezeptoragonist CGP42112 führt zu einer Supression, der Angiotensin AT-2-
Rezeptorantagonist PD123319 führt zu einer Stimulation und der Angiotensin
AT-1-Rezeptorantagonist Losartan (Exp 3174) führt nur zu einer gesteigerten
Natriurese).
In der vorliegenden Arbeit konnte bereits eine Hemmung der durch Phorbol-
myristat-acetat (PMA) induzierten reaktiven Sauerstoffradikalenbildung (ROS)
durch einen Angiotensin AT-1-Rezeptorantagonisten, Exp3174 (Losartan), sowie
durch zwei verschiedene ACE-Hemmer, Captopril und Ramiprilat, (die die
Bildung von Angiotensin ll verhindern) gezeigt werden. Daraufhin aufbauend
sollte untersucht werden, ob eine unterschiedliche ROS-Bildung durch
Stimulierung oder Blockierung der AT-1- und/oder der AT-2-Rezeptoren
stattfindet. Es wurden zusätzlich mehrere andere angiotensinrezeptorwirksame
Substanzen, ein weiterer ACE-Hemmer und ein Calciumantagonist auf ihre
Wirkung in Bezug zur ROS-Konzentration in den mononukleären Leukozyten

36
getestet. Dabei zeigte sich nach allen Untersuchungen eine signifikante
Verminderung der PMA induzierten ROS-Bildung bei den ACE-Hemmern
Captopril und Ramiprilat, bei dem Calciumantagonisten Verapamil, bei dem
Angiotensin AT-2-Rezeptoragonisten CGP42112, bei dem Angiotensin AT-1- und
AT-2-Rezeptoragonisten Angiotensin ll und bei dem Angiotensin AT-1-
Rezeptorantagonisten EXP 3174 sowie eine vermehrte ROS-Bildung bei dem
Angiotensin AT-1-Rezeptoragonisten Angiotensin l.
Eine alleinige Gabe von Angiotensin ll (AT-1-und AT-2-Rezeptoragonist), ohne
PMA Stimulation, in ansteigender Konzentration sowie mit dem Angiotensin AT-
2-Rezeptorantagonisten PD 123319 in Kombination (alleinige AT-1-
Rezeptorwirkung) brachte dagegen keine signifikante Änderung der ROS-
Konzentration gegenüber dem Kontrollwert.
In einer weiteren Meßreihe erfolgte eine reaktive Sauerstoffradikalenmessung
(ROS) in den mononukleären Leukozyten mit dem Radikalenstimulator
Thapsigargin im Vergleich zu Phorbol-myristat-acetat (PMA) sowie eine Messung
der intrazelluläre Calciumionenkonzentration (je eine Meßreihe mit PMA und eine
mit Thapsigargin). In einem Artikel von Tepel (Tepel et al, 1999) wurde die
Aktivität des Na+/H+-Gegentransportsystems bei Patienten mit obstruktivem
Schlaf-Apnoe-Syndrom (OSAS) im Vergleich zu einer gesunden Kontrollgruppe
und der nächtliche Verlauf (bei den Patienten) gemessen. Bei den Patienten mit
OSAS kommt es zu Hypoxiezuständen mit Hypercapnie während der
Atempausen. In den Zellen kommt es zu einer anaeroben Stoffwechsellage
wobei Milchsäure entsteht. So fallen laufend Protonen in den Zellen an. Dadurch
wird der intrazelluläre pH-Wert verändert und bestimmte intracelluläre und
extracelluläre Puffersysteme beeinflußt. Durch das Na+/H+-Gegentransport-
system werden H+-Ionen aus der Zelle transportiert und somit der pH-Wert
reguliert.
Bei Patienten mit OSAS zeigte sich eine signifikant höhere Aktivität des Na+/H+-
Gegentransportsystems. Während der ganzen Nacht war die Aktivität
gleichermaßen hoch. Daher stellt sich die Frage, ob dadurch das Gleich-
gewichtssystem anderer Ionen, z.B. der Calciumionen, bei Patienten mit OSAS
verschoben ist.

37
In anderen Studien wurde bereits gezeigt, daß reaktive Sauerstoffmetabolite den
durch Calcium übertragenen Zellmetabolismus und spezifische membran-
gebundene Regulationsschritte in der Calciumsignalkaskade beeinträchtigen.
Thapsigargin erhöht die cytoplasmatische Calciumionenkonzentration laut
Thastrup (Tharsup et al., 1999) indem es die Wiederaufnahme der Calciumionen
in die zellulären Speicher hemmt. Des weiteren beschreiben Nofer (Nofer et al.,
1999) in einer Studie, daß das Na+/Ca²+-Gegentransportsystem in menschlichen
Lymphozyten durch Thapsigargin induziert wird. Dieses System spielt eine große
Rolle in der Regulation des Gleichgewichtssystems der Elektrolyte in den Zellen.
Takeyama (Takeyama et al., 1993) zeigen in einer Arbeit, daß die oxidative
Schädigung in Mitochondrien mit der Öffnung von nicht-spezifischen
Calciumkanälen zusammenhängt.
Auch in dieser Meßreihe wurden den Proben verschiedene Medikamente, die
ACE-Hemmer Captopril und Ramiprilat und der Angiotensin AT-1-Rezeptor-
antagonist Exp3174, hinzugesetzt. Bei der thapsigargininduzierten ROS-Bildung
konnte eine signifikante Minderung der ROS-Konzentration nach Zugabe des
Angiotensin AT-1-Rezeptorantagonisten Exp3174 festgestellt werden. Bei der
Messung der intrazellulären Calciumionenkonzentration zeigte sich auch bei dem
Angiotensin AT-1-Rezeptorantagonisten Exp3174 eine signifikante Änderung
nach Stimulation mit PMA sowie mit Thapsigargin.
Im Rahmen der hier durchgeführten Untersuchung konnte mit Hilfe der
fluoreszenzspektrophotometrischen Messungen gezeigt werden, daß die
Konzentration der reaktiven Sauerstoffradikale (ROS) in mononukleären
Leukozyten durch Zugabe verschiedener Medikamente signifikant gemindert
werden kann.
Andererseits konnte eine unterschiedliche Konzentration der ROS-Bildung weder
spontan noch nach Stimulation mit Phorbol-myristat-acetat (PMA) bei Patienten
mit obstruktivem Schlaf-Apnoe-Syndrom (OSAS) gegenüber einer gesunden
Kontrollgruppe nachgewiesen werden.

38
Zusammenfassend, mit den Ergebnissen anderer Arbeitsgruppen und Befunden
in der Literatur, läßt sich für einige Erkrankungen, z.B. Diabetes mellitus und
Niereninsuffizienz, eine Erhöhung der reaktiven Sauerstoffradikalen (ROS) in den
mononukleären Leukozyten feststellen. Jedoch gilt dies nicht für Patienten mit
obstruktivem Schlaf-Apnoe-Syndrom.
Die Arbeit zeigt, daß Patienten mit obstruktivem Schlaf-Apnoe-Syndrom keine
signifikanten Veränderungen von intrazellulären reaktiven Sauerstoffradikalen
gegenüber einer gesunden Kontrollgruppe aufweisen. Weiterhin wurden
Hinweise dafür gefunden, daß eine Regulation der Bildung von intrazellulären
reaktiven Sauerstoffradikalen über den Angiotensin Subtyp AT-2-Rezeptor
erfolgt.

39
5. Zusammenfassung
Reaktive Sauerstoffradikale bei Patienten mit obstruktivem Schlaf-Apnoe-
Syndrom und Regulation der Radikalenbildung durch Angiotensin
Rezeptoren
Reaktive Sauerstoffradikale (ROS) spielen eine wesentliche Rolle bei der
intrazellulären Signaltransduktion. Hohe ROS-Konzentrationen führen dagegen
zu einer Schädigung von intrazellulären Proteinen, Enzymen oder
Transkriptionsfaktoren und damit zu einer Störung der regulären Zellfunktion bei
einer Reihe von Erkrankungen wie Entzündungen, Arteriosklerose, arterieller
Hypertonie, kardialen und vaskulären Erkrankungen oder Neoplasien. Patienten
mit obstruktivem Schlaf-Apnoe-Syndrom (OSAS) haben häufig eine arterielle
Hypertonie und eine erhöhte kardiovaskuläre Mortalität. In der vorliegenden
Untersuchung sollte daher geprüft werden, ob Veränderungen von intrazellulären
ROS bei Patienten mit obstruktivem Schlaf-Apnoe-Syndrom vorliegen und
welchen Einfluß Angiotensin AT-2-Rezeptoren bei der Bildung der
Sauerstoffradikale haben.
Die ROS-Konzentration in den mononukleären Leukozyten wurde bei 13
Patienten mit obstruktivem Schlaf-Apnoe-Syndrom und bei 18 altersgleichen
gesunden Kontrollpersonen fluoreszenzspektrophotometrisch mit Hilfe des
intrazellulären Farbstoffs 2',7'-Dichlorofluorescein-diacetat (DCF-DA) bestimmt.
Die Messungen erfolgten bei 534nm mit einer Exzitationswellenlänge von
488nm. Innerhalb von 30 Minuten kam es zu einem spontanen Anstieg von
intrazellulären ROS in mononukleären Leukozyten von Patienten mit
obstruktivem Schlaf-Apnoe-Syndrom um 2.8±0.4 (MW ± SEM, n=13) und von
gesunden Kontrollpersonen um 3.9±0.6 (n=18). Die Stimulation der Zellen mit
Phorbol-myristat-acetat (PMA) führte zu einem Anstieg von intrazellulären ROS
in mononukleären Leukozyten von Patienten mit obstruktivem Schlaf-Apnoe-
Syndrom um 10.4±1.6 und von gesunden Kontrollpersonen um 13.8±1.5. Sowohl
der spontane Anstieg von ROS als auch die Stimulation der Zellen mit PMA
zeigte keinen signifikanten Unterschied zwischen den beiden Gruppen.

40
Der Zusatz des Singlet-oxygen-scavengers Natriumazid bzw. des Peroxynitrite-
scavengers Ebselen ergaben eine signifikante Verminderung der PMA-
stimulierten intrazellulären ROS-Bildung auf 29%±3% und auf 52%±4% des
Kontrollwertes (jeweils p<0.01). Die Vorgabe des Angiotensin Converting Enzym
Inhibitors Captopril führte zu einer signifikanten Verminderung der PMA-
stimulierten intrazellulären ROS-Bildung auf 65%±5% des Kontrollwertes
(p<0.01). Der Angiotensin AT-1- und AT-2-Rezeptoragonist Angiotensin II, der
spezifische Angiotensin AT-1-Rezeptorantagonist Exp3174 und der Angiotensin
AT-2-Rezeptoragonist CGP42112 verursachten ebenfalls eine signifikante
Verminderung der PMA-stimulierten intrazellulären ROS-Bildung (jeweils
p<0.05). Dagegen hatten der Angiotensin AT-1-Rezeptoragonist Angiotensin 1-7
und der spezifische Angiotensin AT-2-Rezeptorantagonist PD123319 keine
Wirkung auf die PMA-stimulierte ROS-Bildung. Dies deutet darauf hin, daß die
intrazelluläre ROS-Bildung durch eine AT-2-Rezeptor Aktivierung blockiert
werden kann.
Die Arbeit zeigt, daß Patienten mit obstruktivem Schlaf-Apnoe-Syndrom keine
signifikanten Veränderungen von intrazellulären reaktiven Sauerstoffradikalen
gegenüber einer gesunden Kontrollgruppe aufweisen. Weiterhin wurden
Hinweise dafür gefunden, daß eine Regulation der Bildung von intrazellulären
reaktiven Sauerstoffradikalen über den Angiotensin Subtyp AT-2-Rezeptor
erfolgt.

41
6. Literaturverzeichnis
1.) Ames BN, Dietary carcinogens and anticarcinogens, Science 221, 1256-
1264, (1983)
2.) Aruoma IO, Halliwell B, Dizdaroglu M, Iron ion-dependent modification of
bases in DNA by the superoxide radical-generating system
hypoxanthine/xanthine oxidase, J. Biol. Chem. 264, 13024-13028, (1989)
3.) Balasubramanyam M, Rohowsky-Kochan C, Reeves JP, Gardner JP,
Na+/Ca2+ exchange-mediated calcium entry in human lymphocytes, J.
Clin. Invest. 94, 2002-2008, (1994)
4.) Ballmer PE, Reinhard WH, Sauerstoffradikale -Leben oder Tod?-, Schweiz
med. Wschr. 118, 1195-1198, (1988)
5.) Breimer LH, Ionizing radiation-induced mutagenesis, Br. J. Cancer 57, 8-18,
(1988)
6.) Bulkley GB, The role of oxygen free radicals in human disease processes,
Surgery 94, 407-411, (1983)
7.) Casale AS, Bulkley GB, Bulkley BH, Flaherty JT, Gott VL, Gardner TJ,
Oxygen free-radicals scavengers protect the arrested, globally ischemic
heart upon reperfusion, Surg. Forum 34, 313-316, (1983)
8.) Christ DD, Human plasma protein binding of the angiotensin 2 receptor
antagonist losartan potassium (DuP753/MK954) and its pharmacologically
active metabolite Exp3174, J. Clin. Pharmacol. 35, 515-520, (1995)
9.) Cross EC, Halliwell B, Borish ET, Pryor WA, Ames BN, Saul RL, McCord
JM, Harmann D, Oxygen radicals and human disease, Ann. Intern. Med.
107, 526-545, (1987)

42
10.) Dijkhorst-Oei LT, Stroes ESG, Koomans HA, Rabelink TJ, Acute
simultaneous stimulation of nitric oxide and oxygen radicals by angiotensin
ll in vivo, J. Cardiovasc. Pharmacol. 33, 420-424, (1998)
11.) Dörner K, Klinische Chemie, Enke Verlag Stuttgart, ISBN 3432 973128,
(1992)
12.) Duan C-L, Yan F-S, Song X-Y, Lu G-W, Changes of superoxide dismutase,
glutathione perioxidase and lipid peroxides in the brain of mice
preconditioned by hypoxia, Biol. Signals Recept. 8, 256-260, (1998)
13.) Epstein FH, Oxygen-derived free radicals in postischemic injury, New Engl.
J. Med. 312, 159-163, (1985)
14.) Geeraerts MM, Ronveaux-Dupal MF, Lemasters JJ, Herman B, Cytosolic
free calcium and proteolysis in lethal oxidative injury in endothelial cells,
Am. J. Physiol. 261, C889-C896, (1991)
15.) Gluckman TL, Grossman JE, Folts JD, Kruse-Elliott KT, Regulation of
leukocyte funktion by nitric oxide donors: The effect of S-Nitroso-Thiol
complexes, J. Tox. Env. Health Part A 61, 9-26, (2000)
16.) Hla KM, Young TB, Bidwell T, Palta M, Skatrud JB, Dempsy J, Sleep
apnoea and hypertension. A population-based study, Ann. Intern. Med.
120, 382-388, (1994)
17.) Igarashi M, Ohyama A, Ohbayashi K, Kozaki S, Komiya Y, The mechanism
of the neurotransmitter release in growth cones, J. Neurosci. Res. 60, 743-
753, (2000)
18.) Inagami T, Kambayashi Y, Ichiki T, Tszuzuki S, Eguchi S., Yamakawa T,
Angiotensin rezeptors: Molecular biology and signalling, Clin. Exp.
Pharmacol. Physiol. 26, 544-549, (1999)

43
19.) Ishizaki H, Ohtawa M, Inhibitory effect of the nonpeptide angiotensin ll
receptor antagonist losartan and its active metabolite, E-3174, on cAMP
phosphodiesterase: additional action of the antagonists, Biochem.
Pharmacol. 48, 201-204, (1994)
20.) Kaiser R, Henning I, Physikalische Chemie, Verlag Gehlen Bad Homburg
vor der Höhe, Gehlenbuch 704, (1986)
21.) Keiser JA, Bjork FA, Hodges JC, Taylor DG Jr., Renal hemodynamic and
excretory response to PD123319 and lorsartan, nonpeptide AT1 and AT2
subtype-specific angiotensin2 ligands, J. Pharmacol. Exp. Ther. 263,
1154-1160, (1992)
22.) Klinke R, Silbernagl S, Lehrbuch der Physiologie, Georg Thieme Verlag
Stuttgart New York, (1994)
23.) Lazo JS, Sebti SM, Fildermann AE, Metabolism of bleomycin and
bleomycin-like compounds. In: Powis G, Prough R, editors. Metabolism
and action of anti-cancer drugs. London: Taylor and Francis, 194-210,
(1987)
24.) Livne A, Veitch R, Grinstein S, Balfe JW, Marquez-Julio A, Rothstein A,
Increased platelet Na+/H+ exchange rates in essential hypertension:
application of a novel test, Lancet, i: 533-536, (1987)
25.) Lo M, Liu KL, Lanteim P, Sassard J, Subtype 2 of angiotensin2 receptors
controls pressure-natriuresis in rats, J. Clin. Invest. 95, 394-397, (1995)
26.) Maitra SR, Sayeed MM, Effect of alpha-adrenergic blockage on cellular
Ca²+ mobilization during endotoxic shock, Circ. Shock. 37, 317-322, (1992)
27.) McCord JM, The superoxide free radical: its biochemistry and patho-
physiology, Surgery 94, 412-414, (1983)

44
28.) Muriel P, Sandoval G, Nitric oxide and peroxynitrite anion modulate liver
plasma membrane fluidity and Na+/K+-ATPase activity, Nitric Oxide: Biol.
Chem. 4, 333-342, (2000)
29.) Mutschler E, Arzneimittelwirkungen, wissenschaftliche Verlagsgesellschaft
mbh Stuttgart, ISBN 3-8047-1377-7, (1996)
30.) Narkiewicz K, Somers VK, The sympathetic nervous system and
obstructive sleep apnoea: implications for hypertension, J. Hypertens. 15,
1613-1619, (1997)
31.) Neusser M, Golinski P, Zhu Z, Zidek W, Tepel M, Thapsigargin-insensitive
Calcium pools in vascular smooth muscle cells, Clin. Exp. Hypertens.,
21(4), 395-405, (1999)
32.) Nofer J-R, Pulawski E, Junker R, Seedorf U, Assmann G, Zidek W, Tepel
M, Na+/Ca²+ exchange inhibitors modulate thapsigargin-inducted Calcium
and Natrium influx in human lymphozytes, Int. J. Clin. Lab. Res. 29, 89-92,
(1999)
33.) Nofer J-R, Tepel M, Walter M, Seedorf U, Assmann G,Zidek W,
Phosphatidylcholine-specific phospholipase C regulates thapsigargin-
induced Calcium influx in human lymphoctes, J. Biol. Chem. 272, 32861-
32868, (1997)
34.) Orie NN, Zidek W, Tepel M, Reaktive oxygen Species in Essential
hypertension and non-insulin-depended diabetes mellitus, Am. J.
Hypertens. 12, 1169-1174, (1999)
35.) Orie NN, Zidek W, Tepel M, Increased intracellular generation of reactive
oxygen species in mononuclear leukocytes from patients with diabetes
mellitus type 2, Exp. Clin. Endocrinol. Diab. 108, 175-180, (2000)

45
36.) Orie NN, Tepel M, Zidek W, Tyrosine and calcium/calmodulin kinases are
common signaling components in the generation of reactive oxygen
species in human lymphocytes, Life Sciences 20, 2135-2142, (1999)
37.) Orie NN, Zidek W, Tepel M, Chemoattractant- and mitogen-induced
generation of reactive oxygen species in human lymphocytes: The role of
calcium, Exp. Physiol. 84, 515-520, (1999)
38.) Partinen M, Jamieson A, Guilleminault C, C. Long-term outcome for
obstructive sleep apnoea syndrome patients: mortality, Chest. 94, 1200-
1204, (1988)
39.) Pfaffendorf M, Sauerstoffradikale und Arteriosklerose, Med. Mo. Pharm. 4,
102-113, (1994)
40.) Rasmussen H, Barrett P, Calcium messenger system: An integrated view,
Physiol. Rev. 64, 938-984, (1984 )
41.) Rose S, Sayeed M, Superoxide radical scavenging prevents cellular calcium
dysregulation during intraabdominal sepsis, Shock 7, 263-268, (1997)
42.) Schettler G, Greten H, Innere Medizin, Thieme Verlag Stuttgart, ISBN
3-13-444309-0, (1998)
43) Schulze-Osthoff K, Wesselborg S, Sauerstoffradikale als sekundäre
Botenstoffe der Genexpression, Dtsch. Med. Wschr. 121, 1301-1302,
(1996)
44.) Shiragi HM, Carey RM, Immunohistochemical localisation of rat
angiotensin2 AT1 receptor, J. Clin. Invest. 100, 264-269, (1997)
45.) Shiragi HM, Carey RM, The subtype 2 (AT2) angiotensin receptor regulates
renal cyclic guanosine in conscious rats, J. Clin. Invest. 97, 1978-1982,
(1996)

46
46.) Siffert W, Düsing R, Sodium-proton exchange and primary hypertension.
An update. Hypertension 26, 649-655, (1995)
47.) Silbernagl S, Despopoulos A, Taschenatlas der Physiologie, Thieme Verlag
Stuttgart, 4. Auflage ISBN 3135677044, (1991)
48.) Silomon M, Pizanis A, Rose S, Oxyradical-mediated hepatocellular Ca²+
alternations during hemorragic shock and resuscitation, Shock 11, 193-
198, (1999)
49.) Takejama N, Matsuo N, Tanaka T, Oxidative damage to mitochondria is
mediated by the Ca²+-dependent innermembrane permeability transition,
Biochem. J. 294, 719-725, (1993)
50.) Tepel M, Echelmeyer M, Orie NN, Zidek W, Increase intracellular reactive
oxygen species in patients with end-stage chronic renal failure: Effect of
hemodialysis, Kidney Int. 58, 867-872, (2000)
51.) Tepel M, Heidenreich S, Zhu Z, Zidek W, Cellular protective action of
angiotensin converting enzyme inhibitors, Exp. Clin. Endocrinol. Diabetes
105, 61-63, (1997)
52.) Tepel M, Heidenreich S, Zidek W, Transgenic hypertensive rats show a
reduced angiotensin 2 induced (Ca²+) response in glomerular mesangial
cells, Life Sci. 62, 69-76, (1998)
53.) Tepel M, Sanner B, van der Giet M, Zidek W, Increased sodium-proton
antiporter activity in patients with obstructive sleep apnoea, J. Sleep. Res.
9, 285-291, (2000)
54.) Tepel M, Schlotmann R, Barenbrock M, Kisters K, Klaus T, Spieker C,
Walter M, Meyer C, Bretzel RG, Zidek W, Lymphocytic Na+/H+ exchange
increases after an oral glucose challenge, Circ. Res. 77, 1024-1029,
(1995)

47
55.) Thastrup O, Role of Ca-ATP-ase in regulation of cellular signaling as
studied with the selective microsomal Ca-ATP-ase inhibitor, thapsigargin,
Agents Actions 26, 207, (1990)
56.) Van der Vliet A, Bast A, Effect of oxidative stress on receptors and signal
transmission, Chem. Biol. Interactions 85, 95-116, (1992)
57.) Weigel G, Griesmacher A, Toma CD, Baecker C, Heinzl H, Mueller MM,
Endothelial eikosanoid metabolism and signaltransduction during
exposure to oxygen radicals injury, Thromb. Res. 87, 363-375, (1997)
58.) Zhu Z, Tepel M, Neusser M, Zidek W, Mechanism of Angiotensin-
Converting Enzym Inhibitors on Agonist-Induced Ca²+-Influx, J. Vasc. Res.
31, 265-270, (1994)

48
7. Danksagung
Der experimentelle Teil dieser Arbeit fand im Zeitraum von August 1998 bis April
2000 im nephrologischen Labor der medizinischen Klinik 1 der Universitätsklinik
Marienhospital in Herne statt. Ich danke Herrn Professor Dr. med. W. Zidek für
die Möglichkeit, dieses Thema im nephrologischen Labor zu bearbeiten.
Besonderer Dank gilt meinem Doktorvater, Herrn PD Dr. med. M.Tepel, der stets
für mich ansprechbar war und mir mit fachlicher Kompetenz zu Rate stand.
Ebenso möchte ich mich bei der Station 6b bedanken, die mir bei der Auswahl
der richtigen Patienten half, bei allen gesunden Freiwilligen, sowie bei allen Mit-
Doktoranden, daß die Koordination im Labor so gut funktioniert hat und die
Hilfsbereitschaft sehr groß war.
Auch meiner Familie, meinen Eltern und Geschwistern, sowie meinem Freund
möchte ich danken, daß sie mir beim Schreiben dieser Arbeit mit Rat und Tat zur
Seite standen.
Außerdem möchte ich mich bei allen bedanken, die mich durch ihre Hilfsbereit-
schaft unterstützt haben.

49
8. Lebenslauf
Persönliche Daten
Name : Susanne Stemmler Geburtsdatum /-ort : 07.06.1972 in Würzburg Familienstand : Ledig Anschrift : Buschweg 21 51519 Odenthal Schulbildung
1978 - 1982 katholische Grundschule in Voiswinkel 1982 - 1988 Städtische-Realschule Bergisch-Gladbach mit
Fachoberschulreife 1991 - 1994 Dietrich-Bonhoeffer-Gymnasium Bergisch-Gladbach
mit allgemeiner Hochschulreife Berufsausbildung :
1988-1991 Ausbildung bei der Bayer AG Leverkusen zur Chemielaborantin
Studium :
1994 - 2000 1996 1997 1999 2000 1999 - 2000
Medizinstudium an der Ruhr-Universität Bochum Physikum 1.Staatsexamen 2.Staatsexamen 3.Staatsexamen Praktisches Jahr im Marienhospital in Herne Wahlfach: Gynäkologie
1998 - 2001 Doktorarbeit: Im nephrologischen Labor des Marienhospitals in Herne unter der Leitung von PD Dr. med. M.Tepel.
Nebentätigkeiten :
1996 - 2000 Aushilfe im Pflegedienst im Klinikum Leverkusen Beruf : 2000 - z.Zt.
AiP im Bereich Gynäkologie und Geburtshilfe im Marienhospital in Herne










![3. Fluoreszenz- und UV/VIS-spektroskopische ......49 3. Fluoreszenz- und UV/VIS-spektroskopische Untersuchungen der dargestellten Imidazo[4,5-c]carbazole, Carbazole und Phenazine 3.1](https://img.pdfslide.tips/doc/110x75/60b7520dfe96d2539945bd31/3-fluoreszenz-und-uvvis-spektroskopische-49-3-fluoreszenz-und-uvvis-spektroskopische.jpg)




![Untersuchung der Fluoreszenz- und ... · veröffentlichter Artikel berichtet von einem vielversprechenden Schritt in diese Richtung.[1] Ein internationales Forscherteam schildert](https://img.pdfslide.tips/doc/110x75/5e1e3b5c61721640f50677d0/untersuchung-der-fluoreszenz-und-verffentlichter-artikel-berichtet-von-einem.jpg)













