Embed Size (px)
Citation preview

RESPUESTAS FUNCIONALES DE CUATRO ESPECIES DE PLANTAS A LA
DISPONIBILIDAD DE NUTRIENTES DEL SUELO EN EL CAÑÓN DEL RÍO
CLARO
Laura Victoria Cano Arboleda
UNIVERSIDAD DE ANTIOQUIA
FACULTAD DE CIENCIAS EXACTAS Y NATURALES
INSTITUTO DE BIOLOGÍA
MEDELLÍN
2013

RESPUESTAS FUNCIONALES DE CUATRO ESPECIES DE PLANTAS A LA
DISPONIBILIDAD DE NUTRIENTES DEL SUELO EN EL CAÑÓN DEL RÍO
CLARO
LAURA VICTORIA CANO ARBOLEDA
Trabajo de grado para optar al título de Bióloga
Asesora
ESTELA QUINTERO VALLEJO, M.Sc.
Coasesora
CRISTINA LÓPEZ GALLEGO, Ph.D.
UNIVERSIDAD DE ANTIOQUIA
FACULTAD DE CIENCIAS EXACTAS Y NATURALES
INSTITUTO DE BIOLOGÍA
MEDELLÍN
2013

AGRADECIMIENTOS
A mi familia, quienes han sido incondicionales en todo, han creído en que la educación
es el mejor regalo que pueden darme y me han dado la oportunidad y todo el apoyo para
realizarme profesionalmente. Gracias papi, mami, Juan D y Juan J los amo y son lo más
importante que tengo. Gracias también a todas las tías, tíos, primas, primos que me
apoyaron de una manera u otra durante mi carrera.
A las mejores asesoras del mundo. Estelita, gracias por tu apoyo y por mostrarme el
maravilloso mundo de la ecología funcional, por compartirme tus conocimientos, tu
tiempo y toda la disposición para sacar a adelante este trabajo. A Cris, gracias por
compartirme tus conocimientos durante gran parte de la carrera, por tu apoyo y por tus
valiosos aportes a este trabajo. A Heri y a Oswald porque me convencieron de
emprender este trabajo, gracias por brindarme sus conocimientos y por el respaldo
durante este proceso. A los mejores ayudantes de campo que puedan existir: Laura
Toro, Yeison Montoya, David Taborda, Julian Zapata, Alejandro Ospina, Jose Orozco y
Don Omar, sin ustedes no hubiera sido posible llevar a cabo este trabajo, gracias por su
tiempo y esfuerzo en pro del trabajo, por apropiarse de el y por disfrutar conmigo todo
el proceso. Agradezco además a las organizaciones que financiaron este trabajo,
especialmente a la reserva natural Cañón del Río Claro “El refugio” por permitirme
realizar este estudio, por apoyarme con el alojamiento y por recibirme amablemente en
sus instalaciones. A The Explorers club por el soporte económico para la realización de
los análisis de suelo y a Idea Wild por su aporte con una balanza, un gps y una cámara
que complementaron muy bien mi trabajo de campo. A las niñas, Vale, Meli, Eli,
Yossi, Laura Toro gracias por el respaldo, por la valiosa amistad que construimos y por
todos los momentos de felicidad que perduraran en los recuerdos. A Val y a Jose,
gracias por todos los momentos compartidos hasta ahora, por el apoyo cuando más lo
necesité y por compartir conmigo mi felicidad y mis logros.
Gracias también a todos los profesores que tuve durante la carrera, personas admirables
a las que agradezco sus enseñanzas, especialmente a Dino Tuberquia, Cristina López,
Ricardo Callejas, Jhon Jairo Ramírez, Juan Parra, Aura Urrea y Vivian Páez.

TABLA DE CONTENIDO
1. RESÚMEN ................................................................................................................ 16
2. INTRODUCCIÓN ..................................................................................................... 17
2.1 Distribución de especies vegetales ........................................................................ 17
2.2 Hipótesis que explican la distribución de especies vegetales ................................ 19
2.3 Respuestas de las plantas a la disponibilidad de nutrientes en el suelo ................. 20
2.4 Rasgos funcionales y su relación con la disponibilidad de nutrientes en el suelo . 21
3. FORMULACIÓN DEL PROBLEMA ...................................................................... 24
4. HIPÓTESIS ............................................................................................................... 26
5. OBJETIVOS .............................................................................................................. 27
5.1 Objetivo general ..................................................................................................... 27
5.2 Objetivos específicos ............................................................................................. 28
6. JUSTIFICACIÓN ...................................................................................................... 29
7. MATERIALES Y MÉTODOS .................................................................................. 31
7.1 Descripción del área de estudio ............................................................................. 31
7.2 Distribución de especies y gradiente edáfico ......................................................... 33
7.3 Experimento de trasplante recíproco ..................................................................... 36
7.4 Análisis estadísticos ............................................................................................... 44
8. RESULTADOS ......................................................................................................... 47
8.1 Distribución de especies y gradiente edáfico ......................................................... 47
8.2 Análisis de suelo .................................................................................................... 48

8.3 Efecto de la disponibilidad de nutrientes en el suelo sobre el éxito de las
especies... .......................................................................................................................... 53
8.4 Variación de rasgos funcionales ............................................................................ 56
8.5 Relación entre los rasgos funcionales y el éxito de las especies. .......................... 88
9. DISCUSIÓN .............................................................................................................. 95
9.1 Patrón de distribución de las especies y gradiente de disponibilidad de
nutrientes… ....................................................................................................................... 95
9.2 Éxito de las especies respecto a la disponibilidad de nutrientes en el suelo .......... 97
9.3 Variación de los rasgos funcionales en relación a la disponibilidad de nutrientes en
el suelo ............................................................................................................................ 100
9.4 Relación entre los rasgos funcionales y el éxito de las especies ......................... 108
10. CONCLUSIONES ................................................................................................... 113
11. RECOMENDACIONES ......................................................................................... 115
REFERENCIAS BIBLIOGRÁFICAS ............................................................................... 116
ANEXOS ............................................................................................................................ 132

LISTA DE TABLAS
Tabla 1. Rasgos funcionales de las plantas relacionados con su respuesta a la
disponibilidad de nutrientes en el suelo, adaptado de Cornelissen et al. (2003) y Pérez-
Harguindeguy et al. (2013). .................................................................................................. 22
Tabla 2. Distribución de los valores de los rasgos funcionales en plantas indicadoras de dos
tipos de suelo. ....................................................................................................................... 27
Tabla 3. Descripción de la metodología para la medición de los rasgos funcionales
elegidos. ................................................................................................................................ 41
Tabla 4. Número de individuos de las especies focales encontrados en los transectos de
vegetación. ............................................................................................................................ 47
Tabla 5. Lista de las especies focales con su índice de especies indicadoras (IV) (resaltado)
y su valor p asociado y otras especies que mostraron IV altos y/o significativos. ............... 48
Tabla 6. Caracterización macroscópica de las muestras de suelo tomadas en los transectos
de vegetación. ....................................................................................................................... 49
Tabla 7. Promedios, desviación estándar y varianza de parámetros básicos de fertilidad y
elementos menores y clase textural para los sitios de suelo elaborado y roca caliza expuesta.
.............................................................................................................................................. 50
Tabla 8. Matriz de componentes principales. ...................................................................... 51

Tabla 9. Tasas de crecimiento relativo (RGR) e intervalo de confianza del 95% en
paréntesis, de plántulas de cada especie focal creciendo en ambos tipos de suelo, calculadas
en dos intervalos de cosecha. ................................................................................................ 54
Tabla 10. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en altura dependiendo del tipo de suelo, biomasa y tiempo para
cada especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de altura de la última cosecha entre tipos de suelo para
la especie C. brunnea............................................................................................................ 58
Tabla 11. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en SLA dependiendo del tipo de suelo, biomasa y tiempo para cada
especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de SLA de la última cosecha entre tipos de suelo para
la especie C. brunnea............................................................................................................ 60
Tabla 12. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en LMA dependiendo del tipo de suelo, biomasa y tiempo para
cada especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de LMA de la última cosecha entre tipos de suelo para
la especie C. brunnea............................................................................................................ 63
Tabla 13. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en área foliar dependiendo del tipo de suelo, biomasa y tiempo para
cada especie y sus interacciones correspondientes y valores del estadístico t y valores de

significancia del contraste de medias de área foliar de la última cosecha entre tipos de suelo
para la especie C. brunnea. ................................................................................................... 66
Tabla 14. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en LDMC dependiendo del tipo de suelo, biomasa y tiempo para
cada especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de LDMC de la última cosecha entre tipos de suelo
para la especie C. brunnea. ................................................................................................... 68
Tabla 15. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en resistencia foliar dependiendo del tipo de suelo, biomasa y
tiempo para cada especie y sus interacciones correspondientes y valores del estadístico t y
valores de significancia del contraste de medias de resistencia foliar de la última cosecha
entre tipos de suelo para la especie C. brunnea. ................................................................... 70
Tabla 16. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en SRL dependiendo del tipo de suelo, biomasa y tiempo para cada
especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de SRL de la última cosecha entre tipos de suelo para
la especie C. brunnea............................................................................................................ 73
Tabla 17. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en biomasa total dependiendo del tipo de suelo, biomasa y tiempo
para cada especie y sus interacciones correspondientes y valores del estadístico t y valores

de significancia del contraste de medias de biomasa total de la última cosecha entre tipos de
suelo para la especie C. brunnea. ......................................................................................... 75
Tabla 18. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en LMF dependiendo del tipo de suelo, biomasa y tiempo para cada
especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de LMF de la última cosecha entre tipos de suelo para
la especie C. brunnea............................................................................................................ 78
Tabla 19. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en SMF dependiendo del tipo de suelo, biomasa y tiempo para cada
especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de SMF de la última cosecha entre tipos de suelo para
la especie C. brunnea............................................................................................................ 80
Tabla 20. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en RMF dependiendo del tipo de suelo, biomasa y tiempo para cada
especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de RMF de la última cosecha entre tipos de suelo para
la especie C. brunnea............................................................................................................ 82
Tabla 21. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en DM dependiendo del tipo de suelo, biomasa y tiempo para cada
especie y sus interacciones correspondientes y valores del estadístico t y valores de

significancia del contraste de medias de DM de la última cosecha entre tipos de suelo para
la especie C. brunnea............................................................................................................ 85
Tabla 22. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en R:S dependiendo del tipo de suelo, biomasa y tiempo para cada
especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de R:S de la última cosecha entre tipos de suelo para
la especie C. brunnea............................................................................................................ 87
Tabla 23. Análisis de regresión múltiple, coeficientes B y valor de significancia para la
relación entre rasgos funcionales y éxito de las especies (biomasa) de las especies
estudiadas. ............................................................................................................................ 88
Tabla 24. Tendencias de la biomasa final y la tasa de crecimiento a través del tiempo de
plántulas de las especies focales creciendo en el sitio roca caliza expuesta y en el sitio suelo
elaborado. ............................................................................................................................. 98
Tabla 25. Principales tendencias de rasgos funcionales a través del tiempo de plántulas de
las especies focales creciendo en el sitio roca caliza expuesta y en el sitio suelo elaborado.
Los colores representan diferencias significativas, amarillo para plántulas creciendo en roca
caliza y verde para plántulas creciendo en el sitio suelo elaborado. .................................. 101
Tabla 26. Correlaciones significativas entre el éxito (biomasa total) de plántulas creciendo
en suelo elaborado y roca caliza expuesta y rasgos funcionales. El color amarillo resalta los
rasgos que tuvieron correlación significativa con biomasa en plántulas creciendo en roca
caliza y el verde en plántulas creciendo en sitio suelo elaborado. ..................................... 109

LISTA DE FIGURAS
Figura 1. Mapa de ubicación zona de estudio. .................................................................... 31
Figura 2. Mapa con la ubicación de los transectos trazados en suelos elaborados, suelos de
roca caliza expuesta y sitios de experimentación. SET1: Transecto 1 en suelo elaborado;
EXP SE: sitio de experimentación suelo elaborado; RCT1: transecto 1 en roca caliza
expuesta; RC EXP: sitio de experimentación roca caliza expuesta...................................... 34
Figura 3. Recolección de plántulas para su posterior siembra en los sitios de
experimentación. A: Clathrotropis brunnea; B: Caryodaphnopsis burgeri; C: Duguetia
colombiana y D: Peltogyne paniculata subsp. pubescens. ................................................... 37
Figura 4. Plántulas de A. C. burgeri, B. D. colombiana, C. Clathrotropis brunnea y D. P.
paniculata subsp. pubescens................................................................................................. 39
Figura 5. Establecimiento de las parcelas de experimentación en A y B: sitio suelo
elaborado y C y D: sitio roca caliza expuesta. ...................................................................... 40
Figura 6. Medición de área foliar y biomasa de tallo, raíz y hojas de las plántulas
cosechadas. ........................................................................................................................... 42
Figura 7. Diseño del penetrómetro ...................................................................................... 44
Figura 8. Gráfica del componente principal 1 versus el componente principal 2. .............. 52

Figura 9. Gráficas caja bigote de biomasa total medida en plántulas creciendo en ambos
sitios de experimentación. El rectángulo corresponde a los cuartiles Q1, Q2 y Q3, la barra
central a la media y los bigotes corresponden a los valores máximo y mínimo. A. C.
burgeri; B. D. colombiana; C. C. brunnea y D. P. paniculata subsp. pubescens. ............... 53
Figura 10. Gráficas que representan los cambios en la tasa de crecimiento relativo de
plántulas de las especies objeto de estudio entre sitios de experimentación. A. C.burgeri; B.
D. colombiana; C. C. brunnea y D. P. paniculata subsp. pubescens. .................................. 56
Figura 11. Tendencias de la Altura por especie y por tipo de suelo a lo largo del tiempo.
Los valores representan el promedio y las barras representan el error estándar. ................. 57
Figura 12. Tendencias del SLA por especie y por tipo de suelo a lo largo del tiempo. Los
valores representan el promedio y las barras representan el error estándar. ........................ 59
Figura 13. Tendencias del LMA por especie y por tipo de suelo a lo largo del tiempo. Los
valores representan el promedio y las barras representan el error estándar. ........................ 62
Figura 14. Tendencias del área foliar por especie y por tipo de suelo a lo largo del tiempo.
Los valores representan el promedio y las barras representan el error estándar. ................. 64
Figura 15. Tendencias de LDMC por especie y por tipo de suelo a lo largo del tiempo. Los
valores representan el promedio y las barras representan el error estándar. ........................ 67
Figura 16. Tendencias de resistencia foliar por especie y por tipo de suelo a lo largo del
tiempo. Los valores representan el promedio y las barras representan el error estándar ..... 69

Figura 17. Tendencias de SRL por especie y por tipo de suelo a lo largo del tiempo. Los
valores representan el promedio y las barras representan el error estándar ......................... 72
Figura 18. Tendencias de Biomasa total por especie y por tipo de suelo a lo largo del
tiempo. Los valores representan el promedio y las barras representan el error estándar ..... 74
Figura 19. Tendencias de LMF por especie y por tipo de suelo a lo largo del tiempo. Los
valores representan el promedio y las barras representan el error estándar ......................... 77
Figura 20. Tendencias de SMF por especie y por tipo de suelo a lo largo del tiempo. Los
valores representan el promedio y las barras representan el error estándar ......................... 79
Figura 21. Tendencias de RMF por especie y por tipo de suelo a lo largo del tiempo. Los
valores representan el promedio y las barras representan el error estándar ......................... 81
Figura 22. Tendencias de DM por especie y por tipo de suelo a lo largo del tiempo. Los
valores representan el promedio y las barras representan el error estándar ......................... 84
Figura 23. Tendencias de R:S por especie y por tipo de suelo a lo largo del tiempo. Los
valores representan el promedio y las barras representan el error estándar. ........................ 86
Figura 24. Gráficas de las correlaciones más significativas entre biomasa total y rasgos
funcionales medidos en plántulas creciendo en ambos tipos de suelo de la especie C.
burgeri y coeficientes de correlación asociados a las líneas de tendencia. .......................... 90

Figura 25. Gráficas de las correlaciones más significativas entre biomasa total y rasgos
funcionales medidos en plántulas creciendo en ambos tipos de suelo de la especie D.
colombiana y coeficientes de correlación asociados a las líneas de tendencia. ................... 91
Figura 26. Gráficas de las correlaciones más significativas entre biomasa total y rasgos
funcionales medidos en plántulas creciendo en el sitio suelo elaborado de la especie C.
brunnea y coeficientes de correlación asociados a las líneas de tendencia. ......................... 92
Figura 27. Gráficas de las correlaciones más significativas entre biomasa total y rasgos
funcionales medidos en plántulas de la especie P. paniculata subsp. pubescens creciendo en
el sitio suelo elaborado y coeficientes de correlación asociados a las líneas de tendencia. . 93
Figura 28. Gráficas de las correlaciones más significativas entre biomasa total y rasgos
funcionales medidos en plántulas de la especie P. paniculata subsp. pubescens creciendo en
el sitio roca caliza expuesta y coeficientes de correlación asociados a las líneas de
tendencia. .............................................................................................................................. 94

LISTA DE ANEXOS
Anexo 1. Lista de las especies registradas en los transectos de vegetación con su índice de
especies indicadoras (IV) y su valor de p asociado. Las especies que presentaron índices
significativos se resaltan con color rojo. ............................................................................ 132
Anexo 2. Caracterización macroscópica de las muestras de suelo tomadas en los transectos
trazados en el sitio de suelo elaborado y en el sitio roca caliza expuesta. .......................... 135
Anexo 3. Parámetros básicos de fertilidad para las muestras de suelo separadas según el
tipo de suelo al que corresponden....................................................................................... 138
Anexo 4. Clase textural y elementos menores de las muestras tomadas por transectos y por
tipo de suelo. ....................................................................................................................... 139
Anexo 5. Parámetros descriptivos de los valores de rasgos funcionales medidos en
plántulas de C. burgeri, C. brunnea, D. colombian y P. paniculata subsp. pubescens. .... 139

1. RESÚMEN El Cañón del Río Claro, localizado en el Departamento de Antioquia es una zona
constituida por formaciones de roca caliza, que presenta relieves escarpados y pendientes
pronunciadas. Esta geomorfología influencia los procesos de formación del suelo y
posiblemente determina la generación de un gradiente de nutrientes que según
observaciones preliminares en campo, podría determinar la distribución de las especies
vegetales en el cañón. El objetivo del presente trabajo fue responder si el patrón de
distribución de las especies Caryodaphnopsis burgeri y Duguetia colombiana (endémicas)
y Clathrotropis brunnea y Peltogyne paniculata subsp. pubescens (amenazadas), está
determinado por la disponibilidad de nutrientes en suelos de roca caliza expuesta y suelos
elaborados. También buscó determinar si la variación en rasgos funcionales relacionados
con la adquisición, uso y conservación de nutrientes, constituyen un posible mecanismo que
refleje las respuestas de las plantas a la disponibilidad de nutrientes. Se determinaron los
patrones de distribución por medio de transectos en ambos tipos de suelo. Adicionalmente
se midió la variación en rasgos funcionales y la supervivencia de estas especies en ambos
suelos, por medio de un experimento de transplante recíproco de plántulas. Sitios de roca
caliza expuesta presentan menor disponibilidad de nutrientes, donde las especies C.
brunnea y P. paniculata subsp. pubescens presentaron pobre desempeño. Por otro lado C.
burgeri y D. colombiana (indicadoras del sitio roca caliza expuesta) fueron exitosas tanto
en roca caliza expuesta como en suelo elaborado. Variación en rasgos relacionados con la
asignación de biomasa, el uso de los nutrientes como masa foliar por unidad de área foliar y
adquisitivos como la longitud específica de la raíz parecen ser una respuesta de las especies
estudiadas a la disponibilidad de nutrientes en este sitio.

2. INTRODUCCIÓN
2.1 Distribución de especies vegetales
El porqué de los patrones de distribución de las especies es una de las preguntas centrales
de la Ecología. Estos patrones son particularmente importantes en bosques tropicales, los
cuales poseen una alta diversidad y complejidad estructural (Gentry 1988; Condit et al.
2000; Barreto et al. 2010). Desde hace más de dos siglos geógrafos como Schimper
(1898) y Walter (1974) se cuestionaban sobre la distribución de las plantas, encontrando
patrones de distribución asociados a diferencias en atributos morfológicos. Asimismo,
Holdridge (1979) logró predecir las formas de vida predominantes en diferentes zonas
terrestres al estudiar los patrones climáticos y asociarlos con la distribución de las plantas.
En general, la distribución de las especies es el resultado de la interacción entre los
procesos biológicos, el conjunto de especies que cohabitan un espacio continuo y las
variables ambientales en diferentes escalas espaciales (Hall & Swaine 1981; Mateucci
1982). Por ejemplo, los patrones de distribución pueden estar influenciados por gradientes
latitudinales, altitudinales, climáticos, de precipitación e intercontinentales (Whittaker
1975; Gentry 1988). A escalas medianas, de tipo regional, la distribución de las especies
puede estar correlacionada con factores edáficos particulares como el estado hidrológico
del suelo, la disponibilidad de nutrientes y la textura del suelo (Swaine 1996; Davies et al.
1998; Clark et al. 1998; Palmiotto 2004; Andersen 2010) que a su vez están relacionados
con el material parental y la topografía del área. De ésta forma, la combinación de factores
edáficos y climáticos juega un papel importante al modificar las tasas de mineralización

del suelo y por lo tanto la disponibilidad de nutrientes en este (Brenes-Arguedas et al.
2008). Algunos estudios han mostrado que la distribución de ciertas especies arbóreas del
bosque tropical está a menudo relacionada con características edáficas de éste (Clark et al.
1999; Baltzer et al. 2005; John et al. 2007, Brenes-Arguedas 2008); y ciertos atributos de
las comunidades de plantas como la diversidad están correlacionados con la
disponibilidad de macro y micro nutrientes como fósforo, nitrógeno, magnesio, potasio,
calcio, sodio y aluminio en diferentes hábitats (Gentry 1988; Ashton & Hall 1992; Clark
1995; Poulsen et al. 2006; John 2007; Peña-Claros et al. 2012).
Así mismo, la composición florística de algunas regiones interactúa y modifica el suelo.
Las tasas de ciclaje de nutrientes pueden estar influenciadas por características
particulares de cada especie, tal como toma de nutrientes y liberación de estos a través de
la hojarasca, tasa de mineralización, entre otras. Las dinámicas de ciclaje de nutrientes
son, en parte, una función de la distribución de las especies y la abundancia en cualquier
sustrato (Gleason et al. 2010).
Aunque los gradientes mencionados anteriormente constituyen a menudo
discontinuidades conspicuas de las condiciones edáficas, que son características comunes
de los paisajes tropicales (Clarck et al. 1998), existe evidencia creciente de que gradientes
edáficos relativamente pequeños juegan un rol importante en la estructuración de paisajes
en los bosques húmedos tropicales (Tuomisto & Ruokolainen 1994). Es por esto que son
fundamentales los estudios que combinen el muestreo de vegetación con análisis de suelos
intensivos y manipulaciones experimentales. Estos estudios se requieren en todos los tipos
de bosques tropicales con el fin de encontrar una posible generalidad en los efectos de los

gradientes edáficos además de determinar los mecanismos por los cuales éstos operan y
cuáles son sus efectos sobre la estructura y función de los bosques (Clark et al. 1998).
2.2 Hipótesis que explican la distribución de especies vegetales
A escalas locales, como la del bosque tropical, la distribución de especies vegetales rara
vez es aleatoria, y en árboles tropicales suele ser generalmente agregada (Hubbell 1979;
Condit et al. 2000; Plotkin et al. 2000). Este nivel de agregación a menudo ha sido
explicado por medio de la diferenciación de nichos ecológicos (Ashton 1969; Connell,
1978; Tilman 1982; Leigh 1999) en los cuales las especies explotan los recursos
disponibles por medio de adaptaciones que determinan los sitios donde éstas son más
dominantes y abundantes con base en su capacidad de competencia (Hubbell & Foster
1983; Tilman & Pacala 1993). Por otro lado, una explicación alternativa para la
distribución agregada de las especies está dada por la teoría neutral de ensamblaje de las
comunidades que asume que no hay diferencias competitivas entre especies (cuyas
habilidades ecológicas se prevé son equivalentes) y que los patrones de distribución están
determinados por sus limitaciones en la dispersión (Hubbell 2001), así, por ejemplo, la
distribución de las especies es agregada a nivel poblacional porque los individuos en su
mayoría se reclutan cerca del parental (Bell 2001; Hubbell 2001). Una cuantificación
rigurosa de las relaciones entre las distribuciones de las especies y las características de
los hábitats, así como la identificación de las causas subyacentes a esos patrones se
requieren para determinar la contribución relativa de la especialización de hábitat al

mantenimiento de la diversidad en bosques tropicales (Hubbell & Foster 1983; Clark et al.
1999; Webb & Peart 2000).
2.3 Respuesta de las plantas a la disponibilidad de nutrientes en el suelo
Las especies vegetales difieren en la capacidad para soportar condiciones de baja
disponibilidad de recursos en el suelo. Por lo tanto especies que ocurren en ambientes
pobres en nutrientes presentan rasgos morfológicos y fisiológicos que incrementan la
eficiencia con la cual adquieren y usan los recursos más limitados dándoles ventajas en
crecimiento y sobrevivencia respecto a especies que no ocurren allí (Lambers & Dijkstra
1987). De esta manera las plantas pueden verse ante diferentes compromisos que
posiblemente se encuentren relacionados con la especialización y eficiencia en un tipo de
hábitat particular (Palmiotto et al. 2004; Baltzer et al. 2005). Por un lado la
especialización en un tipo de hábitat puede estar determinada por un compromiso entre el
crecimiento y la eficiencia con la cual la planta usa los recursos limitantes (Huante et al
1995; Aerts & Chapin 2000; Baltzer et al. 2005). En sitios donde los nutrientes son
limitantes, la eficiencia del uso del recurso puede incrementar por un aumento en la
longevidad foliar y una disminución de la concentración de nutrientes en la hoja,
incrementando así la ganancia de carbono por unidad de nutriente tomada (Huante et al
1995; Aerts & Chapin 2000). Por otro lado, la especialización puede darse por un
compromiso entre el crecimiento y la asignación de recursos para defensa en contra de
herbívoros. El nivel de inversión en defensa anti herbivoría puede incrementar a medida

que la tasa de crecimiento aumenta y la disponibilidad de recursos para remplazar el tejido
de la hoja perdido por herbivoría disminuye (Coley et al. 1985).
Ahora bien, estudiar las respuestas de las plantas respecto al gradiente de disponibilidad
de nutrientes puede contribuir al entendimiento del ensamblaje de la comunidad, ya que
algunas teorías sobre la coexistencia de plantas se basan en la habilidad diferencial de las
especies para explotar diferentes recursos como luz y nitrógeno (Tilman 1988). Esta
habilidad se determina por rasgos funcionales apropiados para establecerse en un sitio
determinado (Belyea & Lancaster 1999) después de sortear la influencia de filtros tanto
abióticos (Ackerly 2003) como bióticos (Cornwell & Ackerly 2009). Y esta habilidad
junto con el nivel de respuesta determina características de las plantas como especialistas
o generalistas (Valencia et al. 2004). Se ha hipotetizado que la especialización en un
recurso representa un compromiso respecto al desempeño en otros recursos y que las
especies generalistas presentan una habilidad competitiva reducida respecto a las especies
especialistas cuando se les compara en el ambiente del especialista (Futuyma & Moreno
1988; Baltzer et al. 2005).
2.4 Rasgos funcionales y su relación con la disponibilidad de nutrientes en el suelo
Los rasgos funcionales son caracteres fisiológicos, anatómicos y de historia de vida
relevantes ecológicamente para comprender y explicar la relación de una planta con su
entorno abiótico y biótico (Cornelissen et al. 2003; Pérez-Harguindeguy et al. 2013). La
ecología funcional se vale de la medición de rasgos funcionales para determinar los roles

o funciones que desempeñan las especies en el ecosistema al que pertenecen. La medición
de éstos rasgos constituye una aproximación importante para el entendimiento de patrones
de distribución de especies a lo largo de gradientes climáticos, disponibilidad de
nutrientes (tabla 1) y respuestas a la perturbación, (Chazdon & Fetcher 1984; Butler &
Chazdon 1998; Poorter et al. 2005; Lavorel et al. 2007).
Un ejemplo claro de rasgos funcionales que han mostrado variación según la
disponibilidad de nutrientes en el suelo son la tasa de crecimiento relativo (RGR) y la
asignación de biomasa (Chapin 1980; Hunt & Lloyd 1987; Korner & Renhardt 1987;
Shipley & Keddy 1988; Shipley & Peters 1990; Lambers & Poorter 1992). Esos estudios
han sugerido que especies de suelos de baja disponibilidad de nutrientes tienen tasas de
crecimiento relativo más bajas, mayor inversión en defensas químicas y mayor asignación
de biomasa para la raíz que plantas de hábitats con mayor disponibilidad de nutrientes
(Grime & Hunt 1975; Chapin 1980; Palmiotto et al. 2004).
Tabla 1. Rasgos funcionales de las plantas relacionados con su respuesta a la
disponibilidad de nutrientes en el suelo, adaptado de Cornelissen et al. (2003) y Pérez-
Harguindeguy et al. (2013).
Rasgo Correlaciones asociadas a los rasgos
Adquisición de nutrientes
Longitud específica
de la raíz
Tasa de toma de nutrientes y agua y tasa de crecimiento
relativo de las plántulas.
Cantidad de tejido de absorción usado por unidad de masa
invertida.

Composición química de la hoja
Concentración de
Nitrógeno y Fósforo
en la hoja
Disponibilidad de N y P en el suelo
Uso de nutrientes
Altura Habilidad competitiva e intercepción de luz.
Tasa de crecimiento
relativo (RGR)
Es un muy buen indicador de la estrategia de la planta
respecto a la productividad ante situaciones de estrés
ambiental y perturbaciones.
Área foliar
específica Tasa de crecimiento relativo o tasa fotosintética máxima.
Masa foliar por
unidad de área
foliar (LMA)
Relacionada con la densidad del tejido foliar, aumenta con
la longevidad de la hoja y con la inversión de carbono en
compuestos celulares secundarios como paredes
secundarias y lignina.
Área foliar
Medida del tamaño de la hoja relacionada, entre otros
factores, con la disponibilidad de nutrientes; el estrés por
nutrientes tiende a seleccionar hojas relativamente
pequeñas.
Conservación de nutrientes
Resistencia física de
las hojas (espesor
de la hoja)
Inversión relativa de carbono en la protección estructural
de los tejidos fotosintéticos.
Longevidad de las
hojas
Compromiso entre tasa de crecimiento y la defensa o
conservación de nutrientes por parte de la planta.
Contenido de
materia seca de la
hoja (LDMC)
Relacionado con la densidad promedio (masa fresca por
volumen fresco) de los tejidos foliares que a su vez está
influenciada por la disponibilidad de nutrientes en el suelo.
Se correlaciona negativamente con la tasa de crecimiento
relativo (RGR) y positivamente con la longevidad de la
hoja.
Asignación de biomasa
Biomasa total
individual Uso de los nutrientes en la producción de biomasa.

Fracción de
biomasa del tallo
(SMF)
Uso de los nutrientes en la producción de biomasa para el
tallo y a su vez para el sostenimiento de los ejes de la
planta.
Fracción de
biomasa de la raíz
Uso de los nutrientes en la producción de biomasa para la
raíz, relacionado con la adquisición de nutrientes.
Fracción de
biomasa de la hoja
(LMF)
Uso de los nutrientes en la producción de biomasa para la
hoja, relacionado con la adquisición y aprovechamiento de
la radiación solar.
Contenido de
materia seca (DM)
Relacionado con la cantidad de tejido estructural (fibras de
esclerénquima, paredes celulares lignificadas y otros
compuestos carbonados secundarios) que contiene la
planta. A mayor disponibilidad de nutrientes, mayor
inversión en tejidos estructurales.
Proporción raíz -
tallo
Compromiso entre la adquisición de nutrientes y el
crecimiento del tallo y la producción de hojas.
3. FORMULACIÓN DEL PROBLEMA
El Cañón del Río Claro se encuentra localizado en la vertiente sur oriental de la cordillera
Central, en el Sureste del departamento de Antioquia. Es una zona caracterizada por un
depósito de mármol de alta pureza con contenidos de Carbonato de Calcio (CaCO3)
superiores al 98 % y de óxidos inferiores al 0.52 %; la mineralogía está dominada por
calcita, cuarzo y moscovita (Restrepo 2011). El cañón presenta características típicas de
una región cárstica (Hernández y Vélez 1988) donde se dan procesos de carstificación
tropical (disolución de las rocas) (Cogollo 1986). En este sitio se hallan relieves muy
escarpados, abruptos y altos, los cuales conforman un relieve de tipo montañoso, donde las
laderas generalmente poseen pendientes muy fuertes, usualmente mayores a 40º. Las
características geomorfológicas del cañón están relacionadas con el hecho de que el suelo

en las zonas marmóreas es muy pequeño o está ausente, al igual que en zonas donde la roca
es en su mayoría cuarcitas, mientras que en zonas de esquistos está bien desarrollado
(Ocampo 2005). Así, la formación de relieves escarpados y pendientes pronunciadas junto
con los procesos de lixiviación de la roca por la lluvia estarían relacionados con un posible
gradiente de disponibilidad de nutrientes en el suelo, de menor a mayor disponibilidad
desde la cima hasta las bases de las colinas.
Las coberturas vegetales presentes en el Cañón del Río Claro corresponden, según la
clasificación de Holdridge (1979), a una transición entre bosque húmedo tropical (bh-T) y
bosque muy húmedo tropical (bmh-T), allí se ha observado que la distribución de las
especies Caryodaphnopsis burgeri, Clathrotropis brunnea, Duguetia colombiana, y
Peltogyne paniculata subsp. pubescens está relacionada con el tipo de hábitat. Por estudios
anteriores (Cogollo 1986) y observaciones en campo se ha encontrado que
Caryodaphnopsis burgeri y Duguetia colombiana se encuentran con mayor frecuencia en
las cimas de las colinas, las cuales se caracterizan por la presencia abundante de roca caliza
que en su mayoría se encuentra expuesta; mientras que las especies Clathrotropis brunnea
y Peltogyne paniculata subsp. pubescens, se encuentran con mayor frecuencia en las partes
bajas de las colinas donde el suelo es más elaborado pues contiene mayor cantidad de
materia orgánica y no se nota la presencia de roca caliza expuesta. Esto sugiere la
posibilidad de que un gradiente de disponibilidad de nutrientes en el suelo sea un factor que
esté determinando la distribución de éstas especies en el Cañón del Río Claro.
Por lo tanto, el presente trabajo pretende responder si el patrón de distribución
anteriormente mencionado está determinado por la disponibilidad de nutrientes en suelos

de roca caliza expuesta y suelos elaborados y si la variación en algunos rasgos funcionales
constituyen un posible mecanismo que refleje las respuestas de las plantas a la
disponibilidad de nutrientes en el suelo y les permita así un mejor desempeño en uno u otro
tipo de suelo.
4. HIPÓTESIS
Hipótesis 1: Si la distribución de plantas en el Cañón del Rio Claro responde a las
condiciones de nutrientes del suelo, entonces en suelos que contrastan en la disponibilidad
de sus nutrientes tales como suelos elaborados y suelos de roca caliza expuesta se
encontraran mayor abundancia y frecuencia de algunas especies que podrán ser indicadoras
de cada uno de los tipos de suelo.
Hipótesis 2: Si en el Cañón del Rio Claro se encuentran especies indicadoras de tipos de
suelos contrastantes, entonces, las especies indicadoras de cada uno de los dos tipos de
suelo serán más exitosas en su ambiente local.
Hipótesis 3: Si las especies de plantas que son indicadoras de suelos contrastantes en el
Cañón del Rio Claro maximizan su éxito en cada uno de sus ambientes a través de sus
rasgos funcionales, entonces plantas indicadoras de suelos elaborados y de suelos de roca
caliza expuesta presentaran valores diferentes en sus rasgos funcionales que posiblemente
estén relacionados con el éxito de las especies en uno u otro tipo de suelo. Las predicciones
de estos rasgos pueden ser observadas en la tabla 2.
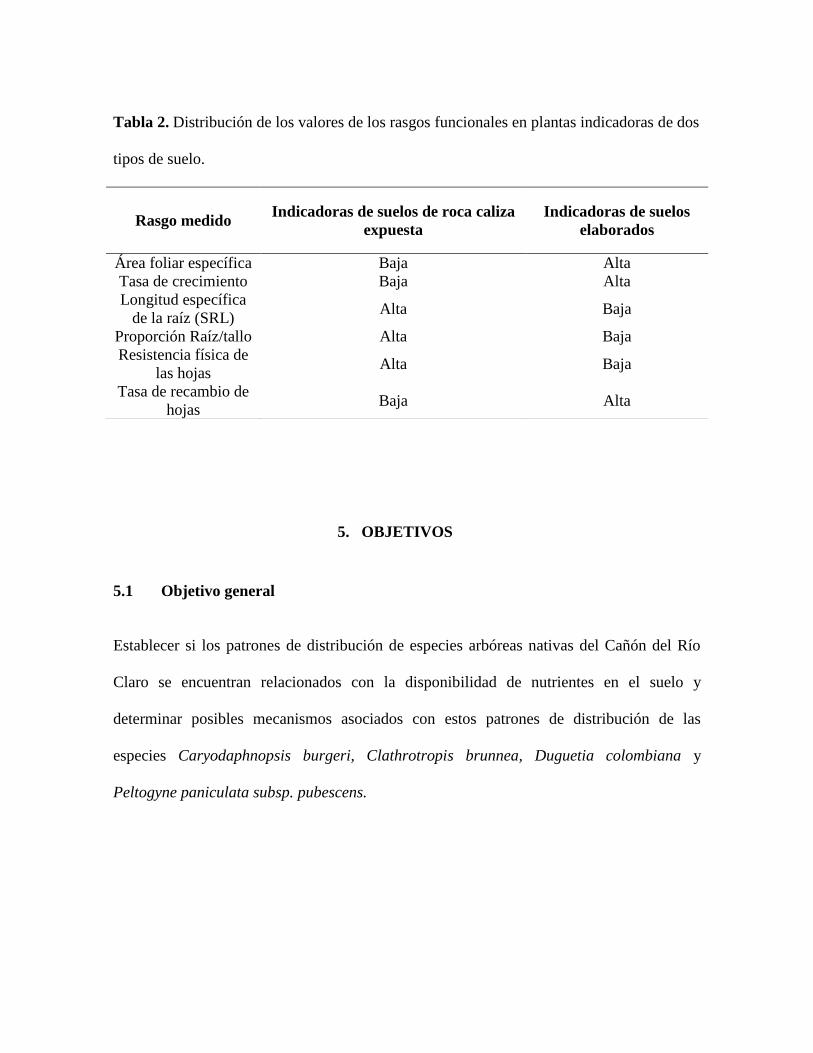
Tabla 2. Distribución de los valores de los rasgos funcionales en plantas indicadoras de dos
tipos de suelo.
5. OBJETIVOS
5.1 Objetivo general
Establecer si los patrones de distribución de especies arbóreas nativas del Cañón del Río
Claro se encuentran relacionados con la disponibilidad de nutrientes en el suelo y
determinar posibles mecanismos asociados con estos patrones de distribución de las
especies Caryodaphnopsis burgeri, Clathrotropis brunnea, Duguetia colombiana y
Peltogyne paniculata subsp. pubescens.
Rasgo medido Indicadoras de suelos de roca caliza
expuesta
Indicadoras de suelos
elaborados
Área foliar específica Baja Alta
Tasa de crecimiento Baja Alta
Longitud específica
de la raíz (SRL) Alta Baja
Proporción Raíz/tallo Alta Baja
Resistencia física de
las hojas Alta Baja
Tasa de recambio de
hojas Baja Alta

5.2 Objetivos específicos
Analizar la composición química de los suelos donde crecen las especies objeto de
estudio con el fin de caracterizar y verificar el posible gradiente de disponibilidad de
nutrientes.
Determinar si a partir de su abundancia y frecuencia las especies Caryodaphnopsis
burgeri, Clathrotropis brunnea, Duguetia colombiana y Peltogyne paniculata subsp.
pubescens pueden ser catalogadas como indicadoras de dos tipos de suelo contrastantes
como son roca caliza expuesta y elaborado.
Evaluar y comparar el éxito de las especies Caryodaphnopsis burgeri, Clathrotropis
brunnea, Duguetia colombiana y Peltogyne paniculata subsp. pubescens por medio de
la medición de biomasa de plántulas creciendo en suelos de roca caliza expuesta y
elaborado.
Establecer si las especies Caryodaphnopsis burgeri, Clathrotropis brunnea,
Duguetia colombiana y Peltogyne paniculata subsp. pubescens presentan variación en
algunos rasgos funcionales de plántulas creciendo en suelo elaborado y en suelo de
roca caliza expuesta y si éstos se encuentran relacionados con el éxito de estas especies
en su ambiente local.

6. JUSTIFICACIÓN
El cañón del Río Claro es una zona particular por su configuración fisiográfica e histórica,
lo que lo hace una región que presenta un alto grado de endemismo para especies vegetales.
Los ecosistemas con especies endémicas son de vital importancia para la conservación de la
biodiversidad, ya que constituyen sitios con condiciones únicas para que se presenten
procesos evolutivos que a su vez contribuyen al incremento de la diversidad regional.
Además de inventarios florísticos del Cañón del Rio Claro que han permitido conocer en
gran medida la composición florística de la zona y registrar algunas especies nuevas para la
ciencia, información detallada que asocie las características ambientales con los patrones de
distribución de especies vegetales en esta zona es escasa o inexistente. Por lo tanto, un
estudio que correlacione las características edáficas con las especies que habitan el lugar,
constituye una primera aproximación al entendimiento de éste particular ecosistema.
Adicionalmente, caracterizar los mecanismos por los cuales se dan los patrones de
distribución de las especies a escala del ecosistema constituye una aproximación al
entendimiento del grado de adaptación de las especies a los ambientes que habitan. Este
tipo de estudios contribuyen al conocimiento de la ecología y biogeografía de las especies
convirtiéndose en una herramienta fundamental que brinda mejores criterios para tomar
decisiones en cuanto a la conservación de estas especies, y restauración de los bosques
tropicales en general.
Al estudiar los rasgos funcionales de las especies presentes en el Cañón del Rio Claro, se
pretende conocer a fondo la ecología funcional de estas especies y por tanto tener una

mejor aproximación al entendimiento de la ecología funcional del bosque húmedo tropical.
Esta es una aproximación novedosa que permite avanzar en la investigación básica y
aplicada en el país. La ecología funcional se perfila como una herramienta que permite
comenzar a predecir procesos de sucesión y de respuesta de los organismos ante cambios
ambientales drásticos como el aumento de temperatura debido al cambio climático y ante
cambios en la disponibilidad de nutrientes en el suelo por efectos de erosión o fertilización.
En este caso, evaluar rasgos funcionales respecto a la disponibilidad de nutrientes permitirá
identificar algunas estrategias fisiológicas y anatómicas de adaptación en las plantas a
suelos infértiles, que a nivel de especie, es una información importante para la
implementación de programas de propagación, conservación y restauración enfocados a
especies de interés biológico, económico, cultural.
Finalmente esta investigación beneficiará a la comunidad científica en general, a la
Universidad de Antioquia y a las autoridades ambientales locales y regionales, ya que
aportará al conocimiento de algunos aspectos biológicos y ecológicos de las especies que
habitan el Cañón del Río Claro y en general del bosque húmedo tropical en Colombia.
Además se espera que los resultados de esta investigación puedan ser comparados con
estudios similares realizados en bosques húmedos tropicales de otras regiones.

7. MATERIALES Y MÉTODOS
7.1 Descripción del área de estudio
La zona de estudio comprende las coberturas boscosas presentes en la Reserva natural
Cañón del Río Claro “El Refugio” con una extensión de 250 Hectáreas y ubicada a los 5°
53’ N y 74 39’ W, en la vertiente sur oriental de la cordillera Central, en el Sureste del
departamento de Antioquia (Colombia), compartiendo límites entre los municipios de
Puerto Triunfo, San Luis y Sonsón, cerca del nacimiento del Río Claro, en el alto del tigre
y el alto de La osa (Sonsón) y a 57 kilómetros de la desembocadura en el río Magdalena y
en cercanías a la autopista Medellín-Bogotá. La ubicación del área de estudio se observa en
la figura 1.
Figura 1. Mapa de ubicación zona de estudio.

Las formaciones vegetales en el área de estudio corresponden, según la clasificación de
Holdridge (1982), a una zona de transición entre bosque húmedo tropical (bh-T) y bosque
muy húmedo tropical (bmh-T). El área presenta un rango de altitud que oscila entre 350 y
700 metros, una temperatura promedio de 24°C y una precipitación anual de 3000 a 5000
mm; estos factores y su interacción con otros como la topografía y los vientos, producen un
clima de transición entre húmedo tropical con lluvias todo el año y tropical monzónico con
tres meses de sequía en el año. El patrón de lluvias es bimodal, con dos épocas en el año
más húmedas (abril - mayo y septiembre – octubre) y otras dos épocas relativamente secas
(enero – febrero y junio – julio) (Cornare 1998).
Geológicamente el área presenta una composición de rocas metamórficas del paleozoico,
siendo estas principalmente ectinitas (formadas por micacitas de una y dos micas y
esquistos cloríticos) y formaciones calcáreas. En general, la geomorfología de la zona de
estudio está compuesta por mármoles, con características típicas de una región cárstica
(Hernández y Vélez 1988). El depósito de mármol es de alta pureza con contenidos de
CaCO3 superiores al 98% y de óxidos inferiores al 0.52% (Restrepo 2011). La deposición
de mármol confiere una topografía característica de colinas con lados verticales o muy
pendientes donde el río y las quebradas han socavado las bases de las colinas, dando lugar a
grandes cavernas (Hoyos & Hernández 1983; Cogollo 1986).
Las características geomorfológicas del cañón (relieves escarpados y pendientes
pronunciadas) junto con los procesos de lixiviación de la roca por la lluvia producen un
gradiente de disponibilidad de nutrientes en el suelo. Dado esto, es posible identificar dos
tipos de suelos contrastantes dentro del gradiente: suelo elaborado (mayor disponibilidad de

nutrientes), generalmente ubicados en las bases de las montañas y roca caliza expuesta
(menor disponibilidad de nutrientes) ubicados en las cimas de las montañas.
7.2 Distribución de especies y gradiente edáfico
7.2.1 Distribución de especies focales
Para determinar si la distribución de especies se encuentra relacionada con la disponibilidad
de nutrientes en los suelos, se establecieron 10 transectos distribuidos en la zona de estudio
procurando capturar la heterogeneidad de los suelos, es decir, 5 transectos en las cimas
donde se observara la roca caliza expuesta y 5 transectos en la base de las montañas donde
se observaran suelos elaborados (figura 2). Cada transecto consistió en la ubicación de una
línea recta de 50 m, la cual es el eje del transecto y a partir de éste, se midió un ancho de 2
m a cada lado para un ancho total de 4 m (parcelas tipo RAP). En cada transecto se
censaron todos los individuos (plántulas, juveniles y adultos) encontrados para las especies
Caryodaphnopsis burgeri, Clathrotropis brunnea, Duguetia colombiana y Peltogyne
paniculata subsp. pubescens. De igual manera se registraron todos los individuos de otras
especies con un DAP mayor a 5 cm que estuvieran dentro de los límites del transecto. En
ambos casos se tomaron muestras botánicas para su posterior identificación en el herbario
de la Universidad de Antioquia (HUA).

Figura 2. Mapa con la ubicación de los transectos trazados en suelos elaborados, suelos de
roca caliza expuesta y sitios de experimentación. SET1: Transecto 1 en suelo elaborado;
EXP SE: sitio de experimentación suelo elaborado; RCT1: transecto 1 en roca caliza
expuesta; RC EXP: sitio de experimentación roca caliza expuesta.
7.2.1.2 Colección y Análisis de suelos
Para caracterizar los suelos de cada uno de los hábitats se tomaron 5 muestras de suelo de
cada transecto, sobre la línea base comenzando a los 5m y a intervalos de 10 m. Para tomar
las muestras de suelo se removieron la hojarasca y el humus del suelo (primeros 5 cm). Se
tomó una muestra de 250 g a una profundidad máxima de 20 cm. Las muestras fueron
llevadas al laboratorio y allí se describieron en términos de estructura, textura y color,
basándose en la metodología propuesta por McGarry (2006) en su manual de campo y en

las cartas de color de suelos de Munsell (1994). Finalmente se tomó una alícuota de 50 g de
cada muestra y se hizo una muestra compuesta por cada transecto para su posterior análisis.
Cada muestra compuesta por transecto fue llevada al laboratorio de biogeoquímica de la
Universidad Nacional (Sede Medellín). En el laboratorio se analizaron las variables físicas
de textura, y las variables químicas de pH, contenido de materia orgánica, macronutrientes
(N, K, P) y micronutrientes (Al, Ca, Mg, K, Cu, Fe, Mn y Zn).
7.2.1.3 Categorización de especies
Con base en la información obtenida de los transectos sobre abundancia y frecuencia de las
especies, se calculó el índice de especies indicadoras (Drufene & Legendre 1997) que
permitió categorizar las especies como indicadoras de alguno de los dos tipos de suelo (roca
caliza expuesta y suelo elaborado) dependiendo de su abundancia y frecuencia en los dos
sitios.
El índice de especies indicadoras es un método numérico usado para determinar especies y
ensamblajes de especies que caracterizan sitios. La asociación entre las especies y un grupo
de sitios se basa en la abundancia relativa de las especies y su frecuencia de ocurrencia en
los diferentes sitios. Este índice es máximo cuando todos los individuos de una especie se
encuentran en un solo sitio, la fórmula para calcularlo es la siguiente:
Aij = Nindividuosij / Nindividuosi
Bij = Nsitiosij / Nsitiosj
INDVALij = Aij x Bij x 100

Donde i es especie y j es una agrupación de sitios. A es una medida de especificidad,
Nindividuosij es el número promedio de individuos de especies i en todos los sitios del
grupo j, mientras que Ninividuosi es la suma de los números promedio de individuos de
especies i en todos los grupos de sitios.
B es una medida de fidelidad, Nsitiosij es el número de sitios en el grupo j donde la especie
i está presente, mientras que Nsitiosj es el número total de sitios en ese grupo. Para probar
la significancia del índice se realiza un procedimiento de reasignación aleatoria de sitios
entre los grupos de sitios, usando permutaciones. La significancia se evalúa por la
diferencia entre el valor observado y el promedio de los valores obtenidos de las
permutaciones aleatorias, ponderado por la desviación estándar de los valores observados
aleatoriamente.
7.3 Experimento de trasplante recíproco
7.3.1 Recolección de plántulas y selección de sitios
Con el fin de determinar si especies indicadoras de cada tipo de suelo presentan mayor
éxito en su ambiente local y medir sus respuestas al tipo de suelo, se colectaron plántulas de
las especies objeto de estudio, evaluando que se encontraran en buen estado, procurando
que su altura fuera lo más homogénea posible (30 cm aproximadamente) y que provinieran
del mismo individuo parental o de parentales cercanos entre sí. Estas plántulas se
obtuvieron mediante recorridos en el área de estudio. Se extrajeron con cuidado de la tierra
y se envolvieron en papel periódico húmedo, luego se trasladaron en bolsas plásticas que
fueron selladas para evitar la desecación de las plántulas (Figura 3).

Figura 3. Recolección de plántulas para su posterior siembra en los sitios de
experimentación. A: Clathrotropis brunnea; B: Caryodaphnopsis burgeri; C: Duguetia
colombiana y D: Peltogyne paniculata subsp. pubescens.
A B
C D

Una vez colectadas las plántulas se establecieron parcelas de experimentación de trasplante
recíproco. Para esto se seleccionaron dos sitios, el primero ubicado en las cimas de las
colinas donde se observara roca caliza expuesta y el segundo ubicado en la parte baja de la
montaña donde se observara un suelo más elaborado.
7.3.2 Establecimiento del experimento in situ
En cada uno de los sitios se establecieron dos parcelas 2 x 3 m por cada una de las especies,
en cada una de las cuales se sembró la mitad de las plántulas colectadas para cada sitio. En
el sitio suelo elaborado se sembraron 43 plántulas de C. burgeri, 30 de P. paniculata subsp.
pubescens, 33 de C. brunnea y 20 de D. colombiana. En el sitio de roca caliza expuesta se
sembraron 45 plántulas de C. brugeri, 30 de P. paniculata subsp. pubescens, 36 de C.
brunnea y 13 de D. colombiana. Las plántulas fueron sembradas en bolsas plásticas con
suelo tomado del mismo sitio de siembra. Las bolsas plásticas fueron usadas para evitar
daños en las raíces en el momento de la cosecha (figura 4).
A B

Figura 4. Plántulas de A. C. burgeri, B. D. colombiana, C. Clathrotropis brunnea y D. P.
paniculata subsp. pubescens.
Una vez en las bolsas, las plántulas se enterraron en cada uno de los dos sitios donde se les
hizo un seguimiento continuo de su desarrollo (figura 5).
D C
A B

Figura 5. Establecimiento de las parcelas de experimentación en A y B: sitio suelo
elaborado y C y D: sitio roca caliza expuesta.
El experimento tuvo una duración de seis meses, contados a partir del momento de la
siembra. La fecha de la primera siembra fue el 30 de Octubre de 2012 y la última cosecha
se hizo el 5 de Junio de 2013. En la recolección de plántulas se tomaron plántulas
adicionales que funcionaron como reserva con el fin de remplazar plántulas muertas
durante los primeros 25 días después de la siembra (24 de Noviembre de 2012).
7.3.3 Cosecha y medición de rasgos funcionales
Con el fin de determinar las respuestas de las plántulas al tipo de suelo donde crecieron, se
midieron diferentes rasgos funcionales. Para esto se hicieron tres cosechas de plántulas
durante los seis meses de duración del experimento con el fin de estimar biomasa total en
cada etapa de crecimiento como una aproximación al crecimiento y cambios en los valores
de los rasgos funcionales durante el desarrollo. Cada 60 días aproximadamente se
cosecharon 10 plántulas por especie y se midieron los rasgos funcionales descritos en la
tabla 3.
C D

Tabla 3. Descripción de la metodología para la medición de los rasgos funcionales
elegidos.
Rasgo Cómo se mide
Adquisición de nutrientes
Longitud específica de la raíz
(SRL)
Proporción entre la longitud de la raíz y su masa,
expresada como m/g.
Uso de nutrientes
Altura Distancia entre el límite superior de las hojas y el nivel
del suelo, expresado en cm.
Tasa de crecimiento relativo
(RGR)
Ganancia de biomasa por unidad de biomasa
y tiempo. Las unidades son g/g.día
Área foliar específica (SLA) Área de una hoja fresca dividida por su masa seca,
expresada en mm2/mg.
Masa foliar por unidad de área
foliar (LMA) Es el inverso del área foliar específica: 1/SLA
Área foliar El área proyectada de una hoja individual expresada en
mm2.
Conservación de nutrientes
Resistencia física de las hojas
(espesor de la hoja)
Fuerza promedio necesaria para perforar una hoja
expresado en Newtons/cm2
Contenido de materia seca de la
hoja (LDMC)
Masa seca de la hoja (mg) dividida su masa fresca
saturada de agua (g). Se expresa en mg/g.
Asignación de biomasa
Biomasa total individual Suma de la biomasa de hojas, tallo y raíz
Fracción de biomasa del tallo
(SMF)
Relación de biomasa
de tallo y biomasa total de la planta. Se expresa en g
(tallo)/g (planta).
Fracción de biomasa de la raíz
(RMF)
Relación de biomasa
de raíz y biomasa total de la planta. Se expresa en g
(raíz)/ g(planta).
Fracción de biomasa de la hoja
(LMF)
Relación de biomasa
de hojas y la biomasa total de la planta. Se expresa en g
(hoja)/g (planta).
Contenido de materia seca (DM)
Relación de peso seco y
el peso fresco de la planta. Se expresa en g (peso
seco)/g (peso fresco).
Proporción raíz - tallo (R:S) Biomasa de la raíz sobre la biomasa de tallo y hojas.

El área foliar se calculó a partir de imágenes obtenidas con un escáner Canon MP280 y
analizadas con el programa “Image J” (figura 6). Para medir los pesos fresco y seco de las
hojas y raíces de cada plántula se usó una balanza digital Ohaus Adventurer. Para obtener el
peso seco (figura 6), las partes de cada planta se secaron previamente en un horno durante
48 horas a 70°C.
Figura 6. Medición de área foliar y biomasa de tallo, raíz y hojas de las plántulas
cosechadas.

La medición de la longitud de las raíces se hizo de acuerdo a la técnica propuesta por
Newman (1966), por la cual se usa una rejilla de cuadrados del mismo tamaño sumergida
en un recipiente con agua, se ubica la raíz sobre la rejilla y se cuenta el número de veces
que alguna parte de la raíz atraviesa alguna de las líneas que forma la cuadrícula.
Las tasas de crecimiento relativo por especie y por sitio se calcularon a través de un fichero
de Microsoft Excel 2000 desarrollado por Hunt et al. (2002). Cada tasa se calculó para dos
intervalos. La primera con pesos de plántulas cosechadas en la primera y en la segunda
cosecha y la segunda con pesos de plántulas cosechadas en la segunda y tercera cosecha.
El cálculo se hizo a partir de la siguiente fórmula:
RGR = (ln P2 –ln P1) / (t2–t1)
Dónde:
P2 y P1 son el peso de la plántula en los tiempos 2 y 1 (t2 y t1, respectivamente).
La fórmula asume que el crecimiento de las plántulas en los primeros estadios suele ser de
tipo exponencial. Para reducir errores se calcularon primero los logaritmos de los pesos
individuales de las plantas y luego, para cada tiempo, se calculó la media de esos
logaritmos como sugieren Hoffmann y Poorter (2002).
La resistencia de las hojas se midió usando un Penetrómetro, el cual determina el peso
necesario para romper una hoja extendida en una superficie, con la cabeza de un clavo de
diámetro conocido. El peso se determina adicionando agua a un recipiente de peso
conocido que se encuentra conectado al embolo de una jeringa que ha sido modificada con

un clavo invertido en su extremo. Una vez la hoja se rompe, se procede a determinar el
peso del agua adicionada. Con esta medida se determina la resistencia de la hoja (N/cm2)
(Rozendaal et al. 2006) (Figura 7).
Figura 7. Diseño del penetrómetro
7.4 Análisis estadísticos
Los tipos de pruebas que se realizaron se discriminan según la hipótesis a la que
correspondan.

Hipótesis 1
Con el fin de evidenciar las posibles diferencias en la disponibilidad de nutrientes de
los suelos se realizó un análisis de componentes principales con los resultados de los
análisis de suelo. El análisis de componentes principales se hizo en el programa IMB
SPSS statistics 20. Posteriormente se hizo un análisis de especies indicadoras usando el
método de Dufrede & Legendre (1997) por medio del programa PCORD versión 6
para determinar cuáles especies son indicadoras de suelo de roca caliza expuesta y
cuáles de suelo elaborado.
Hipótesis 2
Para evaluar el efecto de la disponibilidad de nutrientes del suelo o tipo de suelo
(factor) sobre el éxito de las especies (variable respuesta) se realizó una prueba t-
student por especie para el contraste de medias de la biomasa total de plántulas,
medida en la tercera cosecha. Se evaluó que los datos presentaran distribución normal
y homogeneidad de varianzas. Los datos que no cumplieron con estos requisitos fueron
analizados mediante la prueba no paramétrica Kruskal-Wallis, este fue el caso de la
especie D. colombiana. Ambas pruebas se hicieron con el programa IMB SPSS
statistics 20 con un nivel de confianza del 95%.
Hipótesis 3
Para determinar si hubo diferencias en los rasgos funcionales se llevó a cabo un
análisis de covarianza (ANCOVA) para cada especie cuyo factor fue tipo de suelo
(suelo) con dos niveles: suelo elaborado y de roca caliza expuesta y como covariables

el tiempo (días) y la biomasa total. Además se evaluaron las interacciones Sitio x
biomasa y sitio x tiempo. Las variables de respuesta fueron los rasgos funcionales. Para
la especie C. brunnea, no fue posible realizar el análisis de covarianza debido al bajo
número de plántulas cosechadas en los dos primeros periodos. Para evaluar el efecto
del tipo de suelo sobre los rasgos funcionales de esta especie, se aplicó una prueba t-
student para el contraste de las medias de los rasgos medidos durante la última
cosecha. Finalmente, para establecer si existe una relación entre los rasgos funcionales
y éxito de las especies se hizo una regresión múltiple con biomasa como variable
respuesta y rasgos funcionales como variables independientes. Se calcularon además
los coeficientes de correlación para cada línea de tendencia. Todos los análisis se
hicieron mediante el programa IMB SPSS statistics 20 con un nivel de confianza del
95%.

8. RESULTADOS
8.1 Distribución de especies y gradiente edáfico
8.1.1 Distribución de especies focales
De las especies objeto de estudio se encontraron en total 49 individuos entre plántulas,
juveniles y adultos distribuidos en suelo de roca caliza y suelo elaborado (tabla 4).
Tabla 4. Número de individuos de las especies focales encontrados en los transectos de
vegetación.
Especie N°
indiv
N° de indiv. en
sitio roca
caliza
N° de indiv. en
sitio suelo
elaborado
Caryodaphnopsis burgeri 7 7 0
Duguetia colombiana 19 19 0
Peltogyne paniculata subsp. pubescens 6 6 0
Clathrotropis brunnea 17 0 17
Total 49
Así mismo, se registraron 114 especies de plantas en los transectos trazados en ambos
sitios, 70 especies registradas en el sitio de suelo elaborado y 53 especies registradas en el
sitio de roca caliza expuesta. La lista completa de los individuos registrados se encuentra
en el anexo 1.
8.1.2 Categorización especies (Índice de especies indicadoras)
Con base en el índice de especies indicadoras (IV), las especies objeto de estudio C.
burgeri y D. colombiana son indicadoras del tipo de suelo roca caliza expuesta (tabla 5).
Por su parte las especies Clathrotropis brunnea y Peltogyne paniculata subsp. pubescens
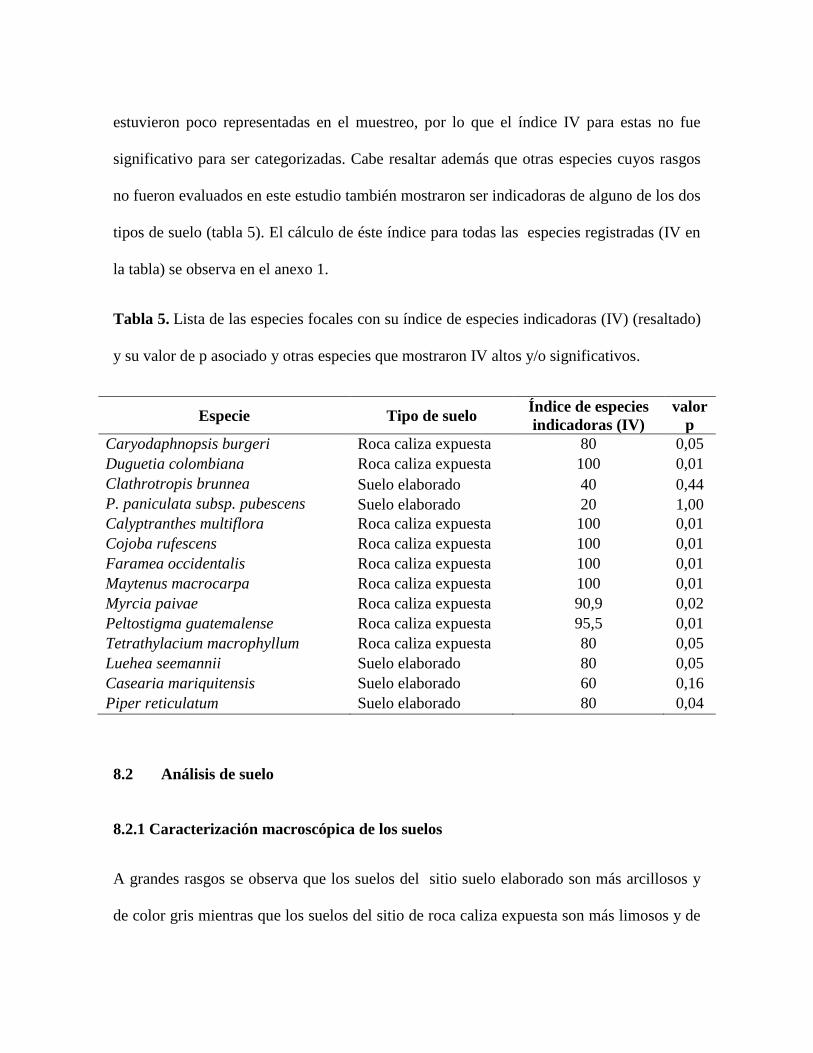
estuvieron poco representadas en el muestreo, por lo que el índice IV para estas no fue
significativo para ser categorizadas. Cabe resaltar además que otras especies cuyos rasgos
no fueron evaluados en este estudio también mostraron ser indicadoras de alguno de los dos
tipos de suelo (tabla 5). El cálculo de éste índice para todas las especies registradas (IV en
la tabla) se observa en el anexo 1.
Tabla 5. Lista de las especies focales con su índice de especies indicadoras (IV) (resaltado)
y su valor de p asociado y otras especies que mostraron IV altos y/o significativos.
Especie Tipo de suelo Índice de especies
indicadoras (IV)
valor
p
Caryodaphnopsis burgeri Roca caliza expuesta 80 0,05
Duguetia colombiana Roca caliza expuesta 100 0,01
Clathrotropis brunnea Suelo elaborado 40 0,44
P. paniculata subsp. pubescens Suelo elaborado 20 1,00
Calyptranthes multiflora Roca caliza expuesta 100 0,01
Cojoba rufescens Roca caliza expuesta 100 0,01
Faramea occidentalis Roca caliza expuesta 100 0,01
Maytenus macrocarpa Roca caliza expuesta 100 0,01
Myrcia paivae Roca caliza expuesta 90,9 0,02
Peltostigma guatemalense Roca caliza expuesta 95,5 0,01
Tetrathylacium macrophyllum Roca caliza expuesta 80 0,05
Luehea seemannii Suelo elaborado 80 0,05
Casearia mariquitensis Suelo elaborado 60 0,16
Piper reticulatum Suelo elaborado 80 0,04
8.2 Análisis de suelo
8.2.1 Caracterización macroscópica de los suelos
A grandes rasgos se observa que los suelos del sitio suelo elaborado son más arcillosos y
de color gris mientras que los suelos del sitio de roca caliza expuesta son más limosos y de
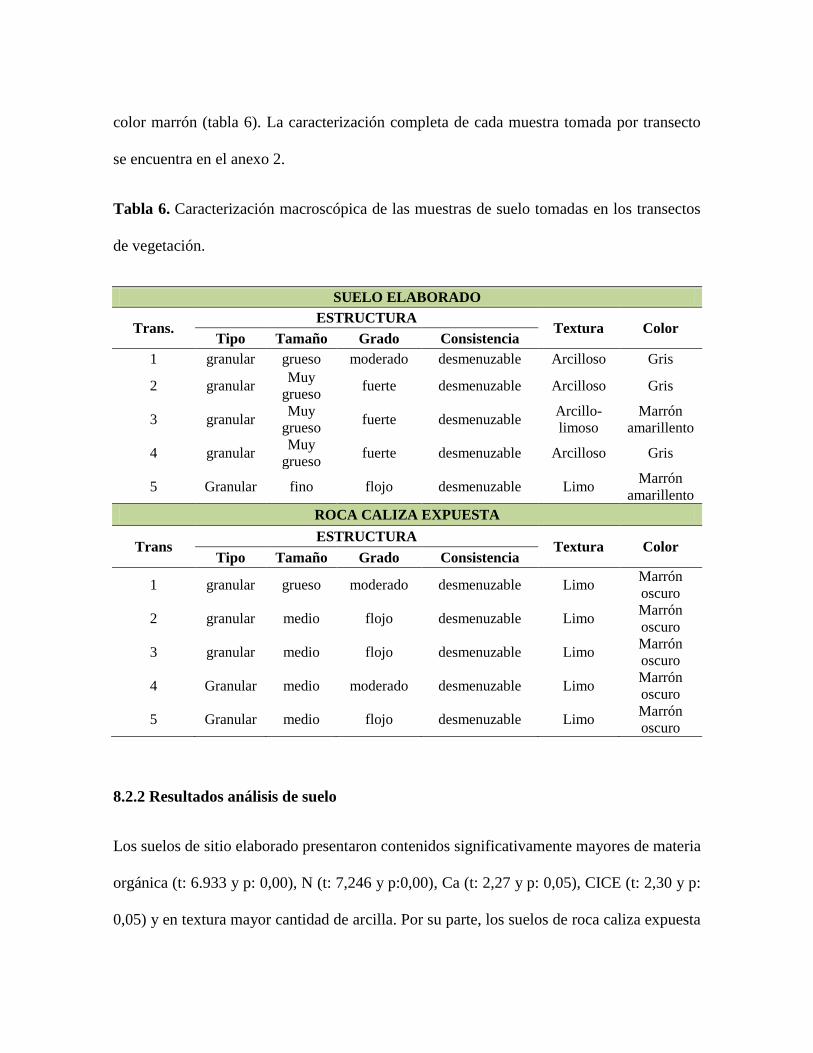
color marrón (tabla 6). La caracterización completa de cada muestra tomada por transecto
se encuentra en el anexo 2.
Tabla 6. Caracterización macroscópica de las muestras de suelo tomadas en los transectos
de vegetación.
SUELO ELABORADO
Trans. ESTRUCTURA
Textura Color Tipo Tamaño Grado Consistencia
1 granular grueso moderado desmenuzable Arcilloso Gris
2 granular Muy
grueso fuerte desmenuzable Arcilloso Gris
3 granular Muy
grueso fuerte desmenuzable
Arcillo-
limoso
Marrón
amarillento
4 granular Muy
grueso fuerte desmenuzable Arcilloso Gris
5 Granular fino flojo desmenuzable Limo Marrón
amarillento
ROCA CALIZA EXPUESTA
Trans ESTRUCTURA
Textura Color Tipo Tamaño Grado Consistencia
1 granular grueso moderado desmenuzable Limo Marrón
oscuro
2 granular medio flojo desmenuzable Limo Marrón
oscuro
3 granular medio flojo desmenuzable Limo Marrón
oscuro
4 Granular medio moderado desmenuzable Limo Marrón
oscuro
5 Granular medio flojo desmenuzable Limo Marrón
oscuro
8.2.2 Resultados análisis de suelo
Los suelos de sitio elaborado presentaron contenidos significativamente mayores de materia
orgánica (t: 6.933 y p: 0,00), N (t: 7,246 y p:0,00), Ca (t: 2,27 y p: 0,05), CICE (t: 2,30 y p:
0,05) y en textura mayor cantidad de arcilla. Por su parte, los suelos de roca caliza expuesta
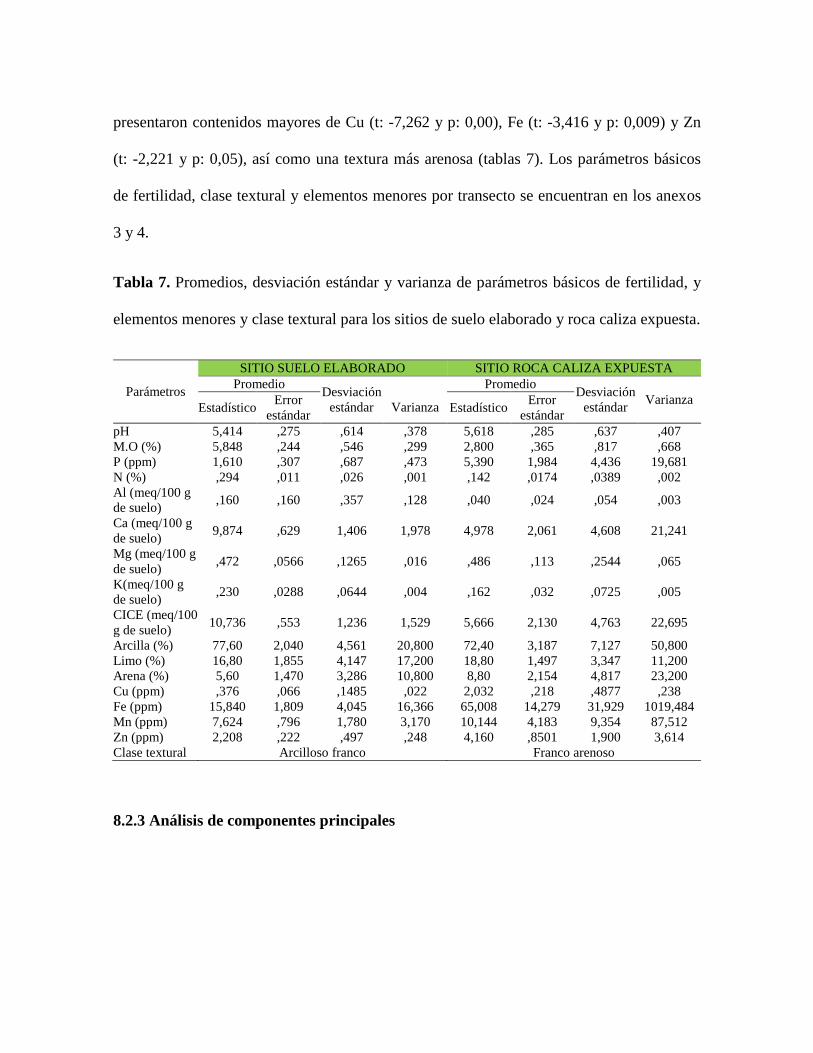
presentaron contenidos mayores de Cu (t: -7,262 y p: 0,00), Fe (t: -3,416 y p: 0,009) y Zn
(t: -2,221 y p: 0,05), así como una textura más arenosa (tablas 7). Los parámetros básicos
de fertilidad, clase textural y elementos menores por transecto se encuentran en los anexos
3 y 4.
Tabla 7. Promedios, desviación estándar y varianza de parámetros básicos de fertilidad, y
elementos menores y clase textural para los sitios de suelo elaborado y roca caliza expuesta.
Parámetros
SITIO SUELO ELABORADO SITIO ROCA CALIZA EXPUESTA
Promedio Desviación
estándar
Varianza
Promedio Desviación
estándar Varianza
Estadístico Error
estándar Estadístico
Error
estándar
pH 5,414 ,275 ,614 ,378 5,618 ,285 ,637 ,407
M.O (%) 5,848 ,244 ,546 ,299 2,800 ,365 ,817 ,668
P (ppm) 1,610 ,307 ,687 ,473 5,390 1,984 4,436 19,681
N (%) ,294 ,011 ,026 ,001 ,142 ,0174 ,0389 ,002
Al (meq/100 g
de suelo) ,160 ,160 ,357 ,128 ,040 ,024 ,054 ,003
Ca (meq/100 g
de suelo) 9,874 ,629 1,406 1,978 4,978 2,061 4,608 21,241
Mg (meq/100 g
de suelo) ,472 ,0566 ,1265 ,016 ,486 ,113 ,2544 ,065
K(meq/100 g
de suelo) ,230 ,0288 ,0644 ,004 ,162 ,032 ,0725 ,005
CICE (meq/100
g de suelo) 10,736 ,553 1,236 1,529 5,666 2,130 4,763 22,695
Arcilla (%) 77,60 2,040 4,561 20,800 72,40 3,187 7,127 50,800
Limo (%) 16,80 1,855 4,147 17,200 18,80 1,497 3,347 11,200
Arena (%) 5,60 1,470 3,286 10,800 8,80 2,154 4,817 23,200
Cu (ppm) ,376 ,066 ,1485 ,022 2,032 ,218 ,4877 ,238
Fe (ppm) 15,840 1,809 4,045 16,366 65,008 14,279 31,929 1019,484
Mn (ppm) 7,624 ,796 1,780 3,170 10,144 4,183 9,354 87,512
Zn (ppm) 2,208 ,222 ,497 ,248 4,160 ,8501 1,900 3,614
Clase textural Arcilloso franco Franco arenoso
8.2.3 Análisis de componentes principales
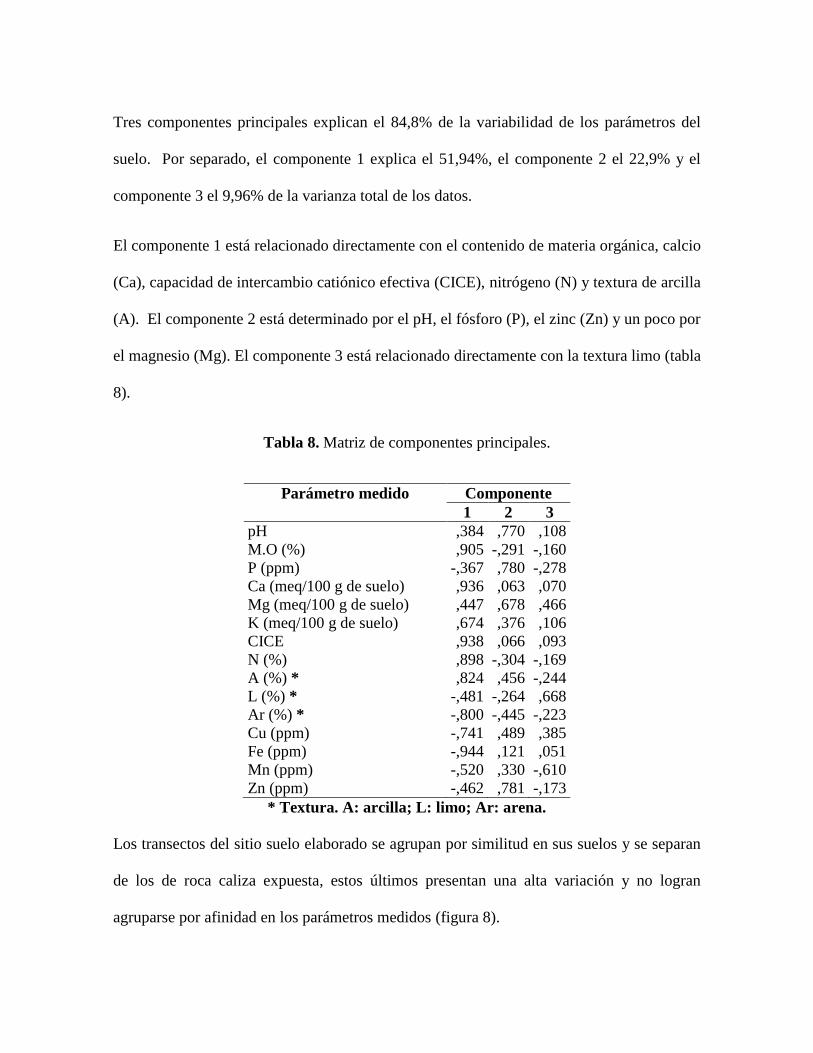
Tres componentes principales explican el 84,8% de la variabilidad de los parámetros del
suelo. Por separado, el componente 1 explica el 51,94%, el componente 2 el 22,9% y el
componente 3 el 9,96% de la varianza total de los datos.
El componente 1 está relacionado directamente con el contenido de materia orgánica, calcio
(Ca), capacidad de intercambio catiónico efectiva (CICE), nitrógeno (N) y textura de arcilla
(A). El componente 2 está determinado por el pH, el fósforo (P), el zinc (Zn) y un poco por
el magnesio (Mg). El componente 3 está relacionado directamente con la textura limo (tabla
8).
Tabla 8. Matriz de componentes principales.
Parámetro medido Componente
1 2 3
pH ,384 ,770 ,108
M.O (%) ,905 -,291 -,160
P (ppm) -,367 ,780 -,278
Ca (meq/100 g de suelo) ,936 ,063 ,070
Mg (meq/100 g de suelo) ,447 ,678 ,466
K (meq/100 g de suelo) ,674 ,376 ,106
CICE ,938 ,066 ,093
N (%) ,898 -,304 -,169
A (%) * ,824 ,456 -,244
L (%) * -,481 -,264 ,668
Ar (%) * -,800 -,445 -,223
Cu (ppm) -,741 ,489 ,385
Fe (ppm) -,944 ,121 ,051
Mn (ppm) -,520 ,330 -,610
Zn (ppm) -,462 ,781 -,173
* Textura. A: arcilla; L: limo; Ar: arena.
Los transectos del sitio suelo elaborado se agrupan por similitud en sus suelos y se separan
de los de roca caliza expuesta, estos últimos presentan una alta variación y no logran
agruparse por afinidad en los parámetros medidos (figura 8).
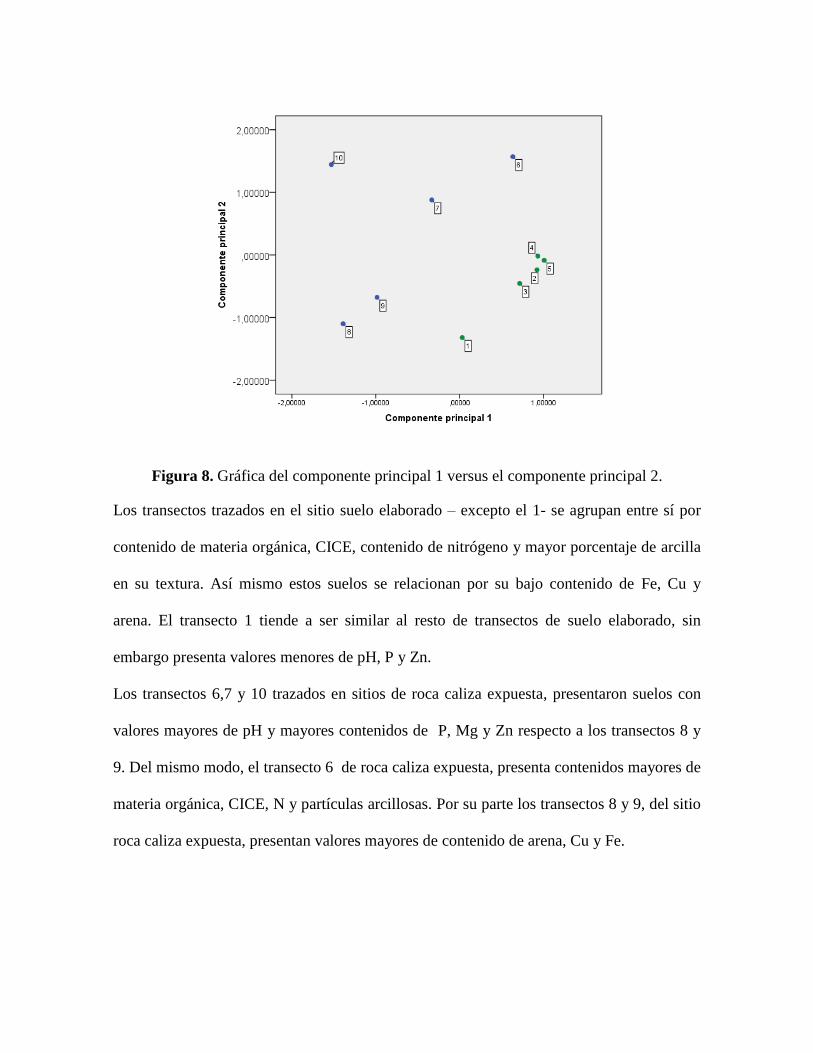
Figura 8. Gráfica del componente principal 1 versus el componente principal 2.
Los transectos trazados en el sitio suelo elaborado – excepto el 1- se agrupan entre sí por
contenido de materia orgánica, CICE, contenido de nitrógeno y mayor porcentaje de arcilla
en su textura. Así mismo estos suelos se relacionan por su bajo contenido de Fe, Cu y
arena. El transecto 1 tiende a ser similar al resto de transectos de suelo elaborado, sin
embargo presenta valores menores de pH, P y Zn.
Los transectos 6,7 y 10 trazados en sitios de roca caliza expuesta, presentaron suelos con
valores mayores de pH y mayores contenidos de P, Mg y Zn respecto a los transectos 8 y
9. Del mismo modo, el transecto 6 de roca caliza expuesta, presenta contenidos mayores de
materia orgánica, CICE, N y partículas arcillosas. Por su parte los transectos 8 y 9, del sitio
roca caliza expuesta, presentan valores mayores de contenido de arena, Cu y Fe.
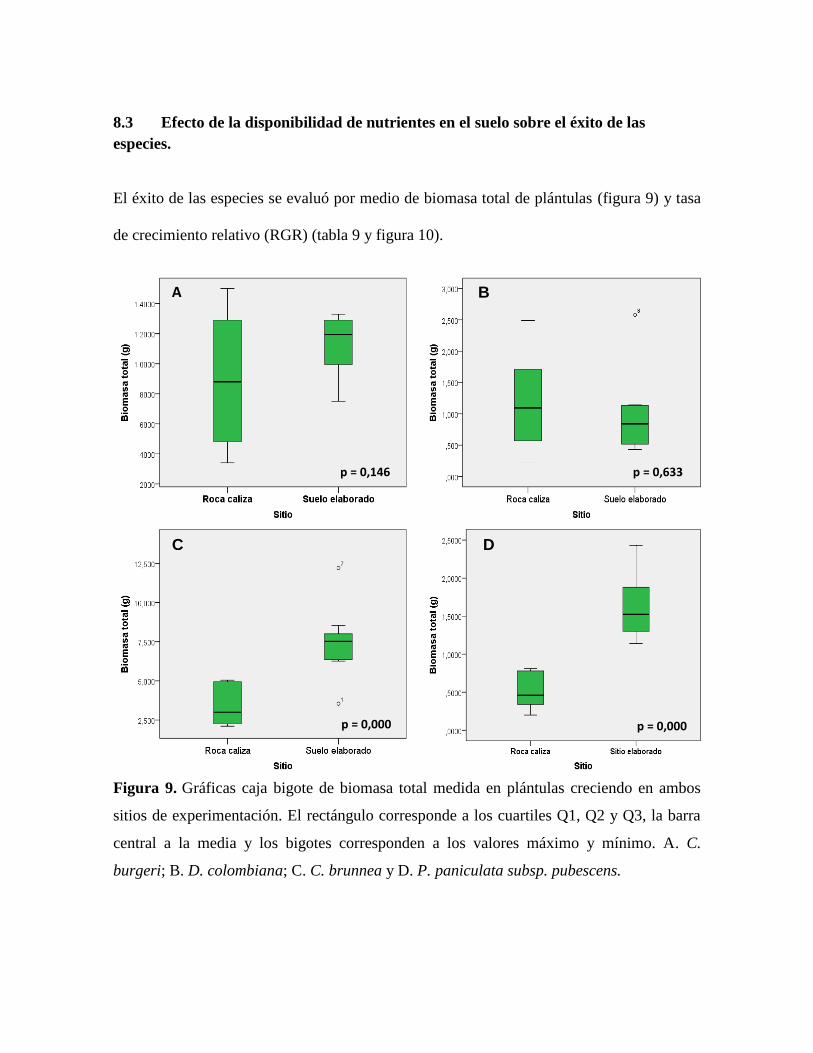
8.3 Efecto de la disponibilidad de nutrientes en el suelo sobre el éxito de las
especies.
El éxito de las especies se evaluó por medio de biomasa total de plántulas (figura 9) y tasa
de crecimiento relativo (RGR) (tabla 9 y figura 10).
Figura 9. Gráficas caja bigote de biomasa total medida en plántulas creciendo en ambos
sitios de experimentación. El rectángulo corresponde a los cuartiles Q1, Q2 y Q3, la barra
central a la media y los bigotes corresponden a los valores máximo y mínimo. A. C.
burgeri; B. D. colombiana; C. C. brunnea y D. P. paniculata subsp. pubescens.
A
p = 0,146
B
p = 0,633
C
p = 0,000
D
p = 0,000

Las especies de roca caliza expuesta (Caryodaphnopsis burgeri y Duguetia colombiana) no
presentaron diferencias significativas en los valores de biomasa total entre tipos de suelo,
aunque la biomasa fue variable en el sitio roca caliza expuesta para C. burgeri (figura 9A y
9B). Por otro lado, las especies del sitio suelo elaborado (Clathrotropis brunnea y
Peltogyne paniculata subsp. pubescens) fueron significativamente más exitosas en el sitio
suelo elaborado que en el sitio roca caliza expuesta (figura 9C y 9D)
En general se observó que las plántulas tuvieron un crecimiento lento con base en las tasas
de crecimiento registradas durante el tiempo de duración del experimento (tabla 9).
Tabla 9. Tasas de crecimiento relativo (RGR) e intervalo de confianza del 95% en
paréntesis, de plántulas de cada especie focal creciendo en ambos tipos de suelo, calculadas
en dos intervalos de cosecha.
Especie Tipo de suelo RGR (g/g.día)
Intervalo 1
RGR (g/g.día)
Intervalo 2
Caryodaphnopsis burgeri
Suelo
elaborado
-0,0036
(0,0089)
0,0034
(0,0083)
Roca caliza -0,0035 (0,011) 0,0033 (0,017)
Duguetia colombiana
Suelo
elaborado -0,017 (0,030) 0,0057 (0,032)
Roca caliza 0,0032 (0,034) 0,0034 (0,029)
Clathrotropis brunnea
Suelo
elaborado -0,0061 (0,018) 0,0082 (0,016)
Roca caliza 0,0017 (0,052) -0,0056
(0,037)
Peltogyne paniculata subsp.
pubescens
Suelo
elaborado 0,0033 (0,0082) 0,013 (0,010)
Roca caliza 0,0018 (0,019) -0,0013
(0,018)

Las plántulas de C. burgeri, aunque inicialmente mostraron una tasa de crecimiento
negativa, ésta tendió a incrementar similarmente en ambos sitios de experimentación
(figura 10A). Lo anterior se corresponde con el hecho de que las plántulas de esta especie
alcanzaron valores similares de biomasa en ambos sitios (figura 9A).
A
B
C
D
Figura 10. Gráficas que representan los cambios en la tasa de crecimiento relativo de
plántulas de las especies objeto de estudio entre sitios de experimentación. A. C.burgeri; B.
D. colombiana; C. C. brunnea y D. P. paniculata subsp. pubescens.

En contraste, para la especie D. colombiana la tasa de crecimiento aumentó en plántulas
creciendo en sitio roca caliza expuesta y disminuyó en plántulas creciendo en el sitio de
suelo elaborado. Vale la pena anotar que en esta especie las plántulas que crecieron en roca
caliza expuesta no tuvieron un crecimiento negativo, mientras que en el suelo elaborado, el
crecimiento se vio afectado inicialmente (figura 10B). Esto es similar a lo que se registró
para biomasa, los valores encontrados en plántulas creciendo en roca caliza expuesta fueron
mayores que los de plántulas creciendo en suelo elaborado (figura 9B).
8.4 Variación de rasgos funcionales
8.4.1 Descripción general rasgos funcionales
Los estadísticos descriptivos de los rasgos funcionales registrados para las plántulas de cada
especie se encuentran en el anexo 5.
8.4.2 Efecto del tipo de suelo sobre los rasgos funcionales de las plántulas
Altura
La altura de las plántulas cambió entre cosechas dependiendo del tipo de suelo donde
crecieron las plántulas (figura 11). Sin embargo, no hubo diferencias significativas entre los
tipos de suelo para ninguna especie mientras que el rasgo si mostró cambios a lo largo del
tiempo y a través del desarrollo (tabla 10).
La altura de las plántulas de C. burgeri varió poco entre tipo de suelo y cosechas, tendiendo
a aumentar un poco a través del tiempo (figura 11A). Sin embargo esta variación no fue
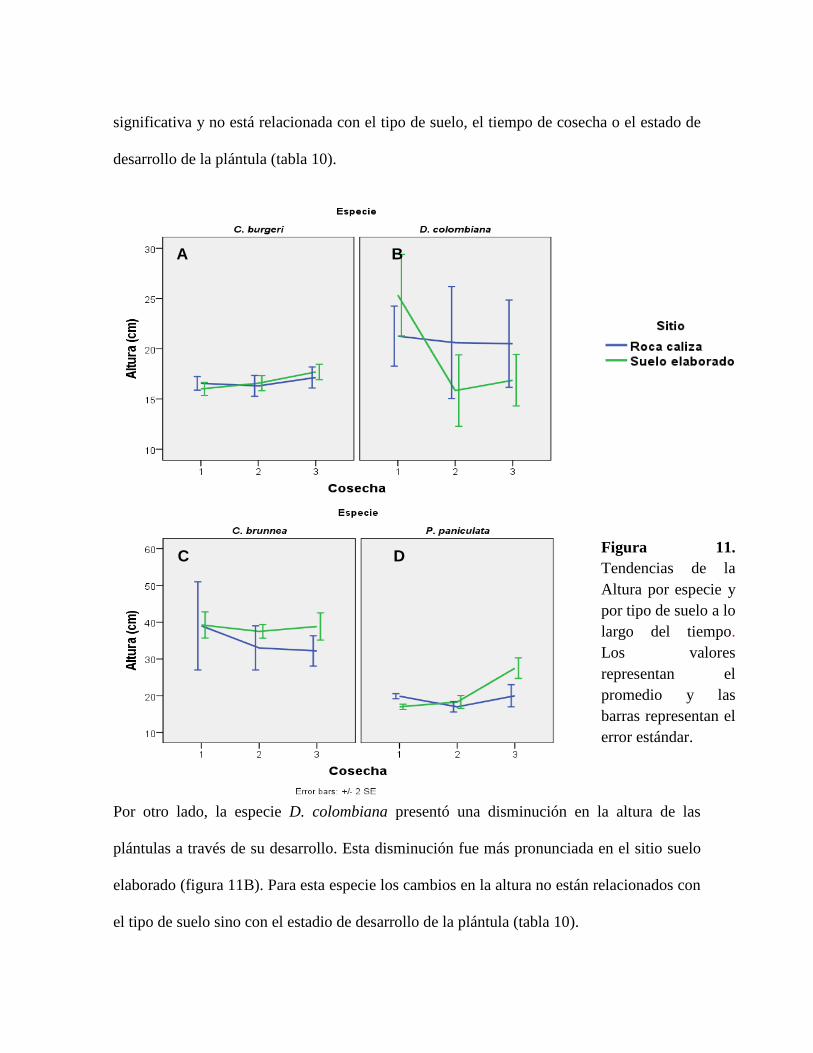
significativa y no está relacionada con el tipo de suelo, el tiempo de cosecha o el estado de
desarrollo de la plántula (tabla 10).
Figura 11.
Tendencias de la
Altura por especie y
por tipo de suelo a lo
largo del tiempo.
Los valores
representan el
promedio y las
barras representan el
error estándar.
Por otro lado, la especie D. colombiana presentó una disminución en la altura de las
plántulas a través de su desarrollo. Esta disminución fue más pronunciada en el sitio suelo
elaborado (figura 11B). Para esta especie los cambios en la altura no están relacionados con
el tipo de suelo sino con el estadio de desarrollo de la plántula (tabla 10).
B A
D C
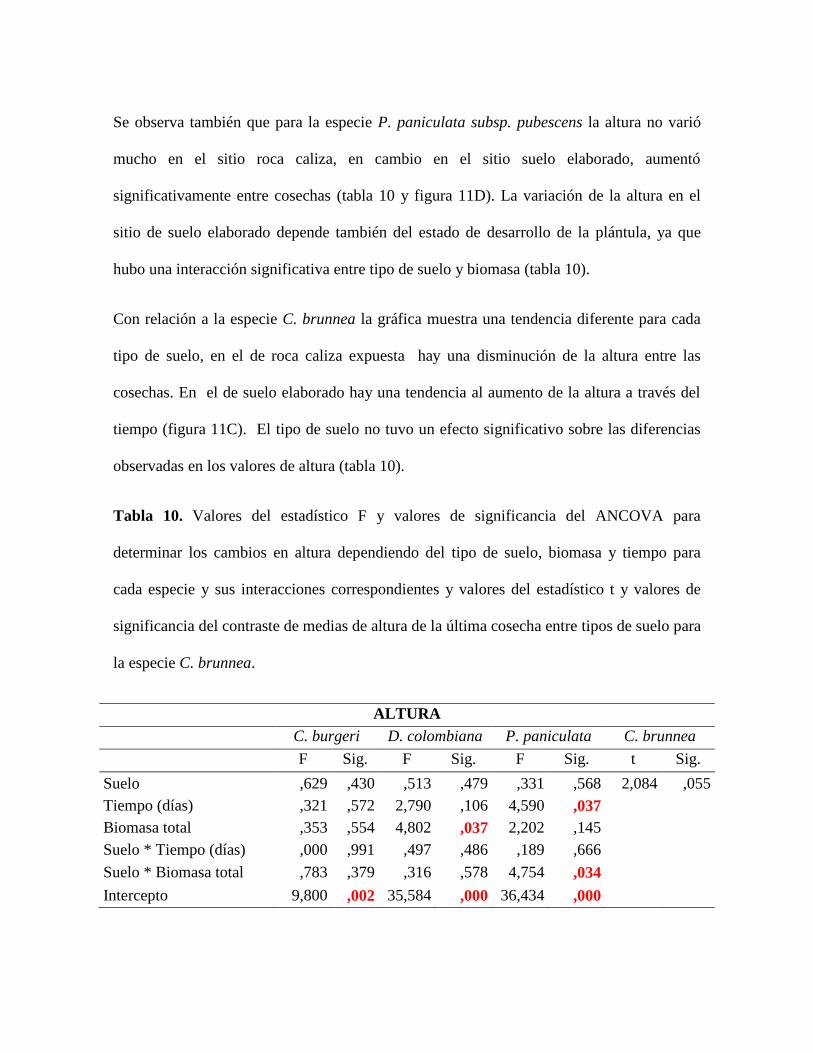
Se observa también que para la especie P. paniculata subsp. pubescens la altura no varió
mucho en el sitio roca caliza, en cambio en el sitio suelo elaborado, aumentó
significativamente entre cosechas (tabla 10 y figura 11D). La variación de la altura en el
sitio de suelo elaborado depende también del estado de desarrollo de la plántula, ya que
hubo una interacción significativa entre tipo de suelo y biomasa (tabla 10).
Con relación a la especie C. brunnea la gráfica muestra una tendencia diferente para cada
tipo de suelo, en el de roca caliza expuesta hay una disminución de la altura entre las
cosechas. En el de suelo elaborado hay una tendencia al aumento de la altura a través del
tiempo (figura 11C). El tipo de suelo no tuvo un efecto significativo sobre las diferencias
observadas en los valores de altura (tabla 10).
Tabla 10. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en altura dependiendo del tipo de suelo, biomasa y tiempo para
cada especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de altura de la última cosecha entre tipos de suelo para
la especie C. brunnea.
ALTURA
C. burgeri D. colombiana P. paniculata C. brunnea
F Sig. F Sig. F Sig. t Sig.
Suelo ,629 ,430 ,513 ,479 ,331 ,568 2,084 ,055
Tiempo (días) ,321 ,572 2,790 ,106 4,590 ,037
Biomasa total ,353 ,554 4,802 ,037 2,202 ,145
Suelo * Tiempo (días) ,000 ,991 ,497 ,486 ,189 ,666
Suelo * Biomasa total ,783 ,379 ,316 ,578 4,754 ,034
Intercepto 9,800 ,002 35,584 ,000 36,434 ,000
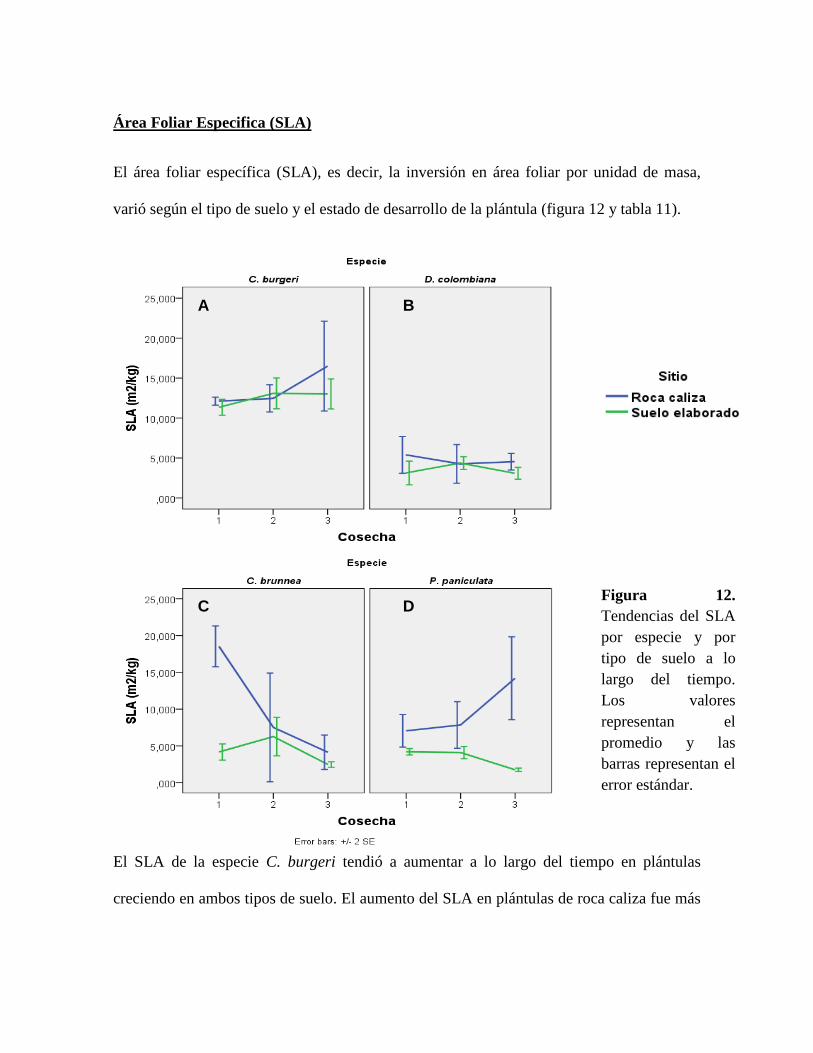
Área Foliar Especifica (SLA)
El área foliar específica (SLA), es decir, la inversión en área foliar por unidad de masa,
varió según el tipo de suelo y el estado de desarrollo de la plántula (figura 12 y tabla 11).
Figura 12.
Tendencias del SLA
por especie y por
tipo de suelo a lo
largo del tiempo.
Los valores
representan el
promedio y las
barras representan el
error estándar.
El SLA de la especie C. burgeri tendió a aumentar a lo largo del tiempo en plántulas
creciendo en ambos tipos de suelo. El aumento del SLA en plántulas de roca caliza fue más
B A
D C
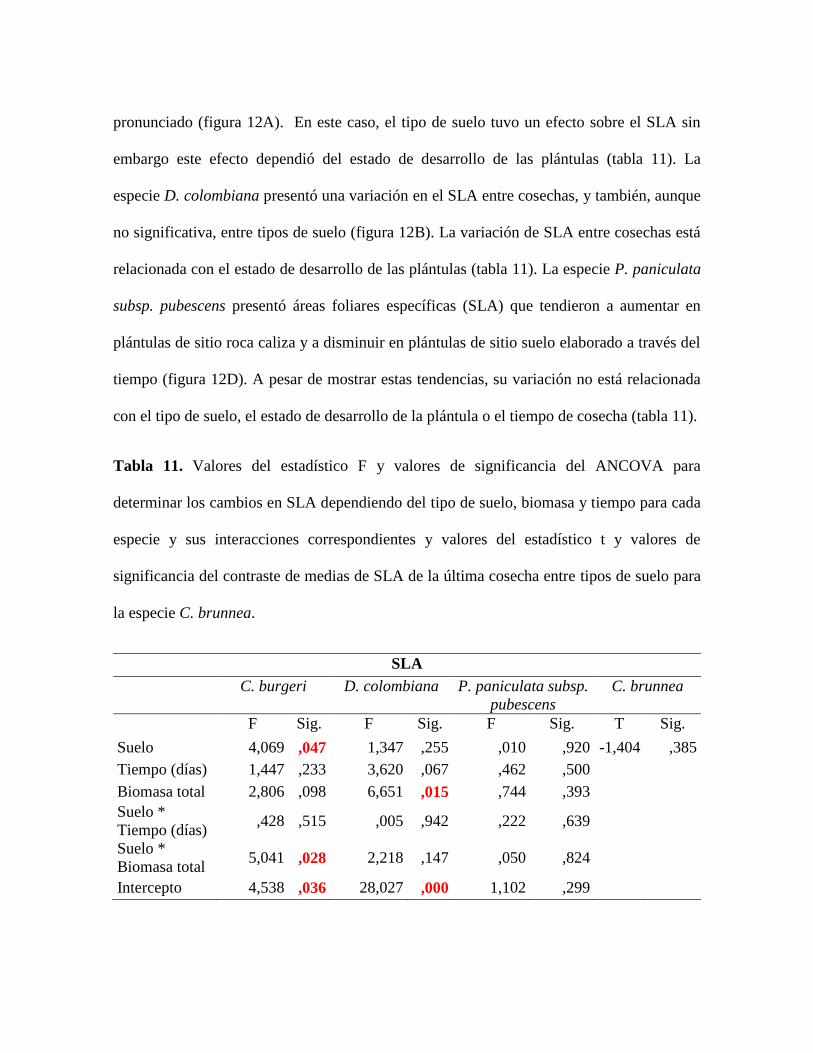
pronunciado (figura 12A). En este caso, el tipo de suelo tuvo un efecto sobre el SLA sin
embargo este efecto dependió del estado de desarrollo de las plántulas (tabla 11). La
especie D. colombiana presentó una variación en el SLA entre cosechas, y también, aunque
no significativa, entre tipos de suelo (figura 12B). La variación de SLA entre cosechas está
relacionada con el estado de desarrollo de las plántulas (tabla 11). La especie P. paniculata
subsp. pubescens presentó áreas foliares específicas (SLA) que tendieron a aumentar en
plántulas de sitio roca caliza y a disminuir en plántulas de sitio suelo elaborado a través del
tiempo (figura 12D). A pesar de mostrar estas tendencias, su variación no está relacionada
con el tipo de suelo, el estado de desarrollo de la plántula o el tiempo de cosecha (tabla 11).
Tabla 11. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en SLA dependiendo del tipo de suelo, biomasa y tiempo para cada
especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de SLA de la última cosecha entre tipos de suelo para
la especie C. brunnea.
SLA
C. burgeri D. colombiana P. paniculata subsp.
pubescens
C. brunnea
F Sig. F Sig. F Sig. T Sig.
Suelo 4,069 ,047 1,347 ,255 ,010 ,920 -1,404 ,385
Tiempo (días) 1,447 ,233 3,620 ,067 ,462 ,500
Biomasa total 2,806 ,098 6,651 ,015 ,744 ,393
Suelo *
Tiempo (días) ,428 ,515 ,005 ,942 ,222 ,639
Suelo *
Biomasa total 5,041 ,028 2,218 ,147 ,050 ,824
Intercepto 4,538 ,036 28,027 ,000 1,102 ,299
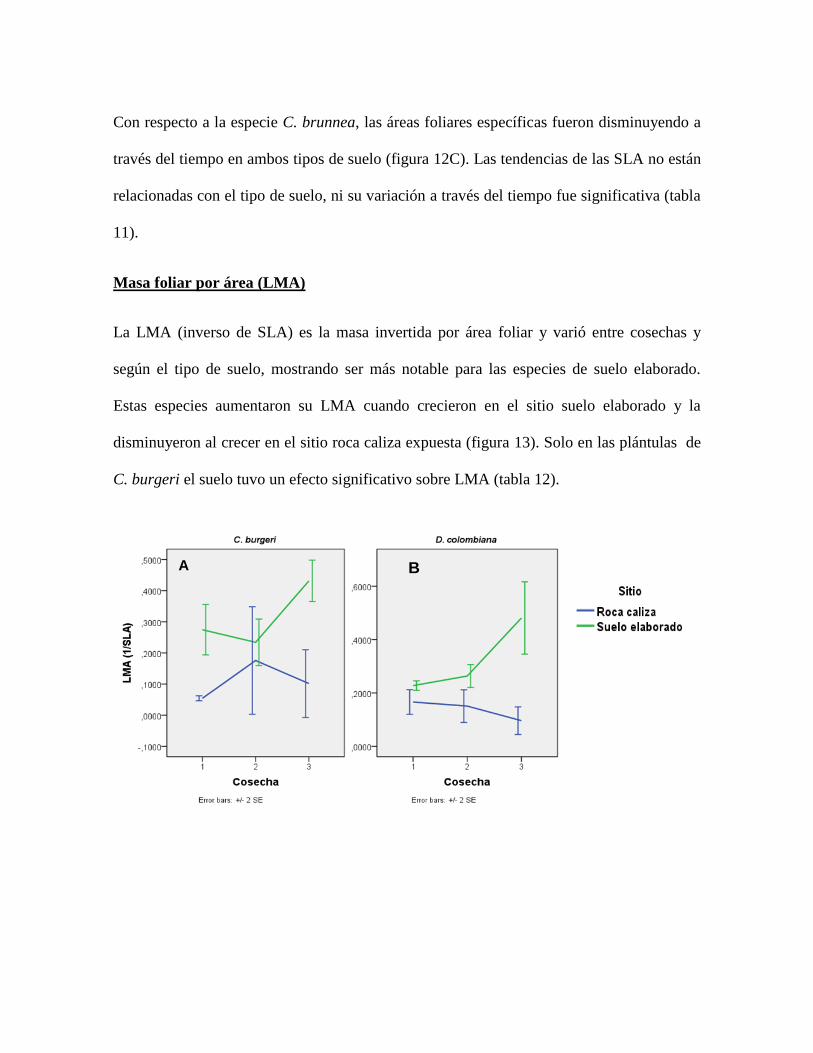
Con respecto a la especie C. brunnea, las áreas foliares específicas fueron disminuyendo a
través del tiempo en ambos tipos de suelo (figura 12C). Las tendencias de las SLA no están
relacionadas con el tipo de suelo, ni su variación a través del tiempo fue significativa (tabla
11).
Masa foliar por área (LMA)
La LMA (inverso de SLA) es la masa invertida por área foliar y varió entre cosechas y
según el tipo de suelo, mostrando ser más notable para las especies de suelo elaborado.
Estas especies aumentaron su LMA cuando crecieron en el sitio suelo elaborado y la
disminuyeron al crecer en el sitio roca caliza expuesta (figura 13). Solo en las plántulas de
C. burgeri el suelo tuvo un efecto significativo sobre LMA (tabla 12).
B A
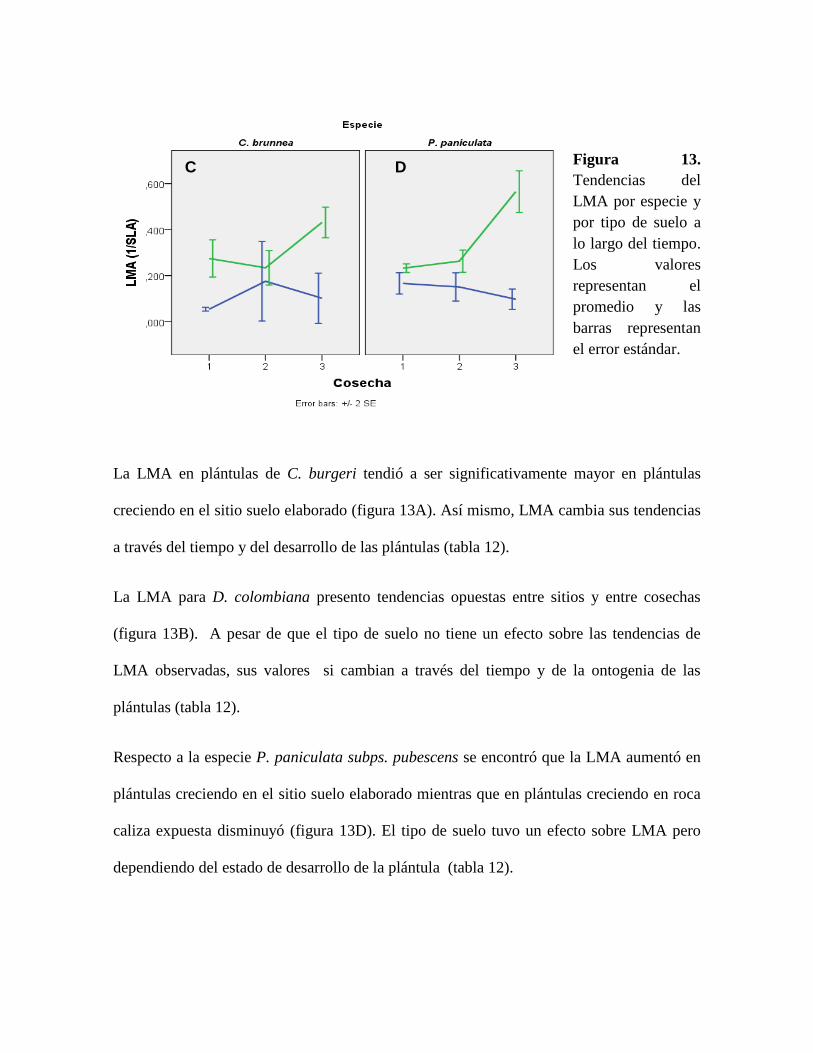
Figura 13.
Tendencias del
LMA por especie y
por tipo de suelo a
lo largo del tiempo.
Los valores
representan el
promedio y las
barras representan
el error estándar.
La LMA en plántulas de C. burgeri tendió a ser significativamente mayor en plántulas
creciendo en el sitio suelo elaborado (figura 13A). Así mismo, LMA cambia sus tendencias
a través del tiempo y del desarrollo de las plántulas (tabla 12).
La LMA para D. colombiana presento tendencias opuestas entre sitios y entre cosechas
(figura 13B). A pesar de que el tipo de suelo no tiene un efecto sobre las tendencias de
LMA observadas, sus valores si cambian a través del tiempo y de la ontogenia de las
plántulas (tabla 12).
Respecto a la especie P. paniculata subps. pubescens se encontró que la LMA aumentó en
plántulas creciendo en el sitio suelo elaborado mientras que en plántulas creciendo en roca
caliza expuesta disminuyó (figura 13D). El tipo de suelo tuvo un efecto sobre LMA pero
dependiendo del estado de desarrollo de la plántula (tabla 12).
D C
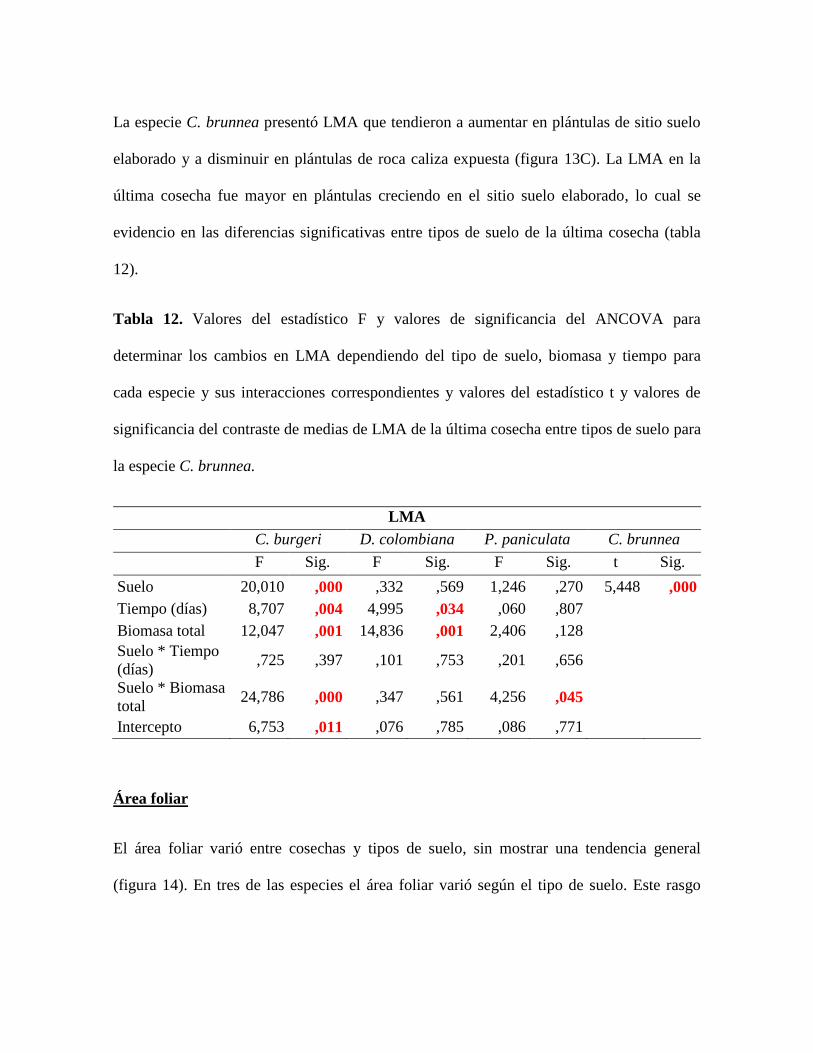
La especie C. brunnea presentó LMA que tendieron a aumentar en plántulas de sitio suelo
elaborado y a disminuir en plántulas de roca caliza expuesta (figura 13C). La LMA en la
última cosecha fue mayor en plántulas creciendo en el sitio suelo elaborado, lo cual se
evidencio en las diferencias significativas entre tipos de suelo de la última cosecha (tabla
12).
Tabla 12. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en LMA dependiendo del tipo de suelo, biomasa y tiempo para
cada especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de LMA de la última cosecha entre tipos de suelo para
la especie C. brunnea.
LMA
C. burgeri D. colombiana P. paniculata C. brunnea
F Sig. F Sig. F Sig. t Sig.
Suelo 20,010 ,000 ,332 ,569 1,246 ,270 5,448 ,000
Tiempo (días) 8,707 ,004 4,995 ,034 ,060 ,807
Biomasa total 12,047 ,001 14,836 ,001 2,406 ,128
Suelo * Tiempo
(días) ,725 ,397 ,101 ,753 ,201 ,656
Suelo * Biomasa
total 24,786 ,000 ,347 ,561 4,256 ,045
Intercepto 6,753 ,011 ,076 ,785 ,086 ,771
Área foliar
El área foliar varió entre cosechas y tipos de suelo, sin mostrar una tendencia general
(figura 14). En tres de las especies el área foliar varió según el tipo de suelo. Este rasgo

también depende del estado de desarrollo de las plántulas para algunas de las especies
focales (tabla 13).
Figura 14. Tendencias
del área foliar por
especie y por tipo de
suelo a lo largo del
tiempo. Los valores
representan el
promedio y las barras
representan el error
estándar.
Las áreas foliares de la especie C. burgeri fueron significativamente diferentes entre sitios
dependiendo del tiempo y del estado de desarrollo de la plántula (figura 14A y tabla 13).
B A
D C

Éstas fueron en principio similares y con el paso del tiempo presentaron tendencias
opuestas, alcanzando valores mayores en plántulas del sitio suelo elaborado (figura 14A).
La especie D. colombiana no presentó variaciones significativas en el área foliar entre tipos
de suelo más si entre cosechas (Figura 14B y tabla 13). En plántulas creciendo en el sitio
roca caliza se observó un tendencia al incremento en el área foliar mientras que en el sitio
de suelo elaborado se observó una disminución a través del tiempo.
En relación con áreas foliares de plántulas de C. brunnea se encontraron diferencias
significativas entre tipos de suelo (tabla 13). El área foliar presentó tendencias diferentes
según el tipo de suelo. . Mientras que en la roca caliza el área foliar siempre disminuyó, en
el sitio de suelo elaborado aumentó en la última cosecha (figura 14C).
Respecto a la especie P. paniculata subsp pubescens el área foliar de las plántulas fue
significativamente mayor en plántulas creciendo en el sitio suelo elaborado, respecto a las
que crecieron en el sitio roca caliza expuesta. Estas diferencias dependen además del estado
de desarrollo de las plántulas (figura 14D y tabla 13).
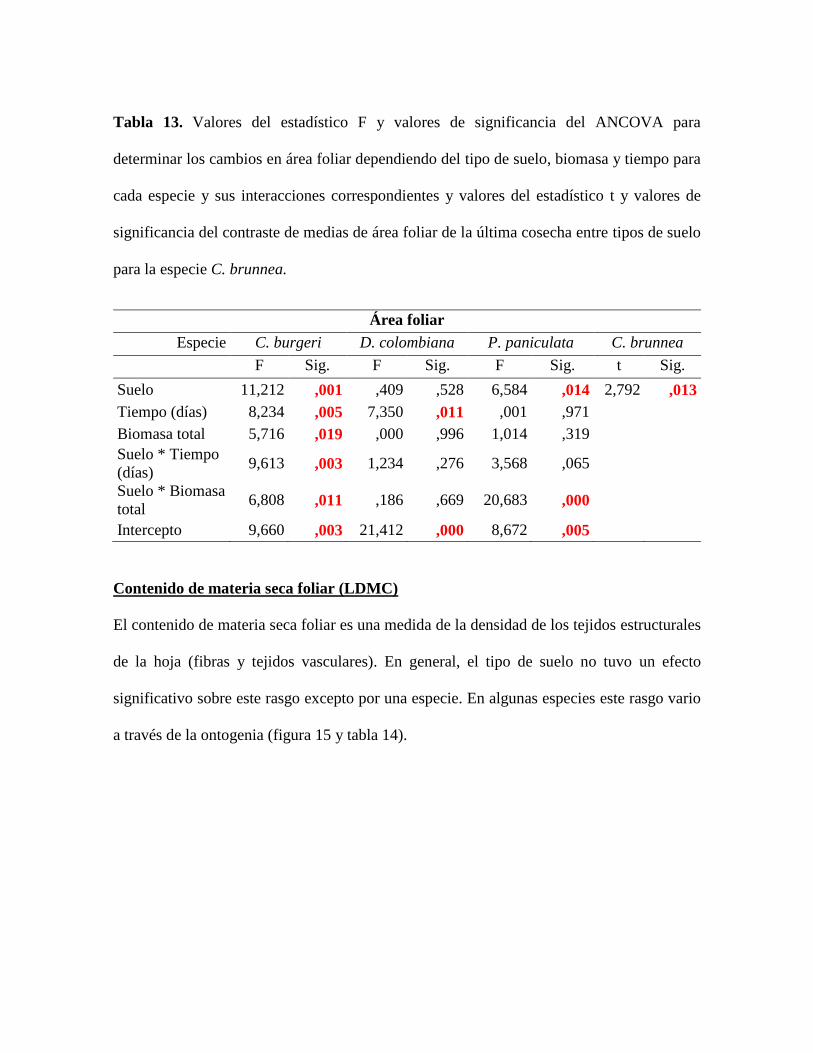
Tabla 13. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en área foliar dependiendo del tipo de suelo, biomasa y tiempo para
cada especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de área foliar de la última cosecha entre tipos de suelo
para la especie C. brunnea.
Área foliar
Especie C. burgeri D. colombiana P. paniculata C. brunnea
F Sig. F Sig. F Sig. t Sig.
Suelo 11,212 ,001 ,409 ,528 6,584 ,014 2,792 ,013
Tiempo (días) 8,234 ,005 7,350 ,011 ,001 ,971
Biomasa total 5,716 ,019 ,000 ,996 1,014 ,319
Suelo * Tiempo
(días) 9,613 ,003 1,234 ,276 3,568 ,065
Suelo * Biomasa
total 6,808 ,011 ,186 ,669 20,683 ,000
Intercepto 9,660 ,003 21,412 ,000 8,672 ,005
Contenido de materia seca foliar (LDMC)
El contenido de materia seca foliar es una medida de la densidad de los tejidos estructurales
de la hoja (fibras y tejidos vasculares). En general, el tipo de suelo no tuvo un efecto
significativo sobre este rasgo excepto por una especie. En algunas especies este rasgo vario
a través de la ontogenia (figura 15 y tabla 14).

Figura 15.
Tendencias de
LDMC por especie
y por tipo de suelo
a lo largo del
tiempo. Los valores
representan el
promedio y las
barras representan
el error estándar.
El rasgo funcional LDMC en las especies C. burgeri y D. colombiana presentó tendencias
diferentes y no estuvo influenciado por el tipo de suelo o por el estado de desarrollo de las
plántulas (figura 15 A, B y tabla 14).
Respecto al LDMC de la especie C. brunnea, este disminuyó entre cosechas en plántulas de
ambos tipos de suelo. Los LDMC de plántulas creciendo en el sitio roca caliza
B A
D C

disminuyeron más drásticamente hasta alcanzar valores significativamente menores que los
de plántulas creciendo en el sitio suelo elaborado (figura 15C y tabla 14).
La especie P. paniculata subsp. pubescens presentó un LDMC en plántulas creciendo en el
sitio roca caliza expuesta que disminuyó con su estado de desarrollo y de forma diferente
dependiendo del tipo de suelo (figura 15D y tabla 14).
Tabla 14. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en LDMC dependiendo del tipo de suelo, biomasa y tiempo para
cada especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de LDMC de la última cosecha entre tipos de suelo
para la especie C. brunnea.
LDMC
Especie C. burgeri D. colombiana P. paniculata C. brunnea
F Sig. F Sig. F Sig. T Sig.
Suelo 1,535 ,219 2,527 ,123 ,517 ,476 3,294 ,020
Tiempo (días) ,006 ,940 ,111 ,741 ,383 ,539
Biomasa total ,128 ,722 ,378 ,543 5,533 ,023
Suelo * Tiempo
(días) ,253 ,617 2,552 ,121 ,084 ,773
Suelo * Biomasa
total ,541 ,464 ,698 ,410 ,094 ,761
Intercepto 1,513 ,222 272,133 ,000 3,848 ,056
Resistencia foliar
La resistencia foliar está dada por la cantidad de tejido estructural de la hoja (fibras de
esclerénquima, células con paredes lignificadas, celulosa, etc.) y mostró en general ser

mayor en plántulas creciendo en el sitio roca caliza, aunque el tipo de suelo solo tuvo un
efecto significativo sobre la resistencia foliar de dos de las especies focales (figura 16 y
tabla 15).
Figura 16. Tendencias
de resistencia foliar
por especie y por tipo
de suelo a lo largo del
tiempo. Los valores
representan el
promedio y las barras
representan el error
estándar
La resistencia foliar de plántulas de C. burgeri fue similar entre tipos de suelo a través de
las cosechas y del desarrollo (figura 16A y tabla 15). En plántulas de la especie D.
B A
D C
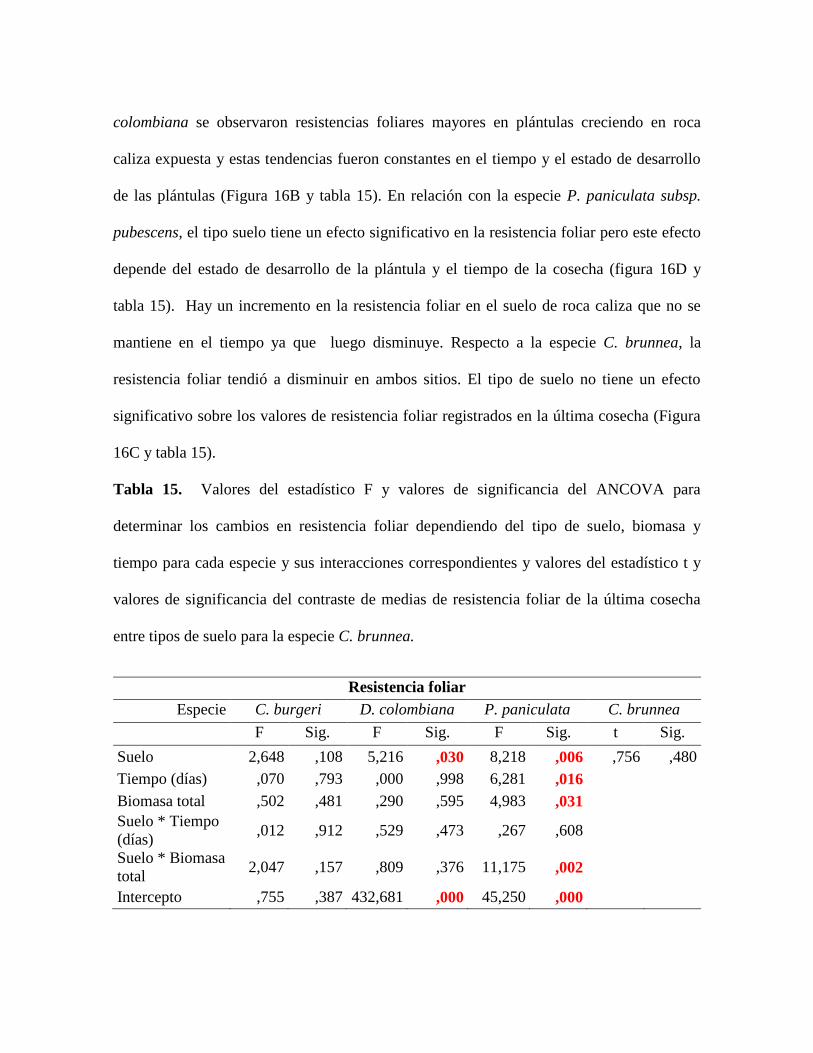
colombiana se observaron resistencias foliares mayores en plántulas creciendo en roca
caliza expuesta y estas tendencias fueron constantes en el tiempo y el estado de desarrollo
de las plántulas (Figura 16B y tabla 15). En relación con la especie P. paniculata subsp.
pubescens, el tipo suelo tiene un efecto significativo en la resistencia foliar pero este efecto
depende del estado de desarrollo de la plántula y el tiempo de la cosecha (figura 16D y
tabla 15). Hay un incremento en la resistencia foliar en el suelo de roca caliza que no se
mantiene en el tiempo ya que luego disminuye. Respecto a la especie C. brunnea, la
resistencia foliar tendió a disminuir en ambos sitios. El tipo de suelo no tiene un efecto
significativo sobre los valores de resistencia foliar registrados en la última cosecha (Figura
16C y tabla 15).
Tabla 15. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en resistencia foliar dependiendo del tipo de suelo, biomasa y
tiempo para cada especie y sus interacciones correspondientes y valores del estadístico t y
valores de significancia del contraste de medias de resistencia foliar de la última cosecha
entre tipos de suelo para la especie C. brunnea.
Resistencia foliar
Especie C. burgeri D. colombiana P. paniculata C. brunnea
F Sig. F Sig. F Sig. t Sig.
Suelo 2,648 ,108 5,216 ,030 8,218 ,006 ,756 ,480
Tiempo (días) ,070 ,793 ,000 ,998 6,281 ,016
Biomasa total ,502 ,481 ,290 ,595 4,983 ,031
Suelo * Tiempo
(días) ,012 ,912 ,529 ,473 ,267 ,608
Suelo * Biomasa
total 2,047 ,157 ,809 ,376 11,175 ,002
Intercepto ,755 ,387 432,681 ,000 45,250 ,000
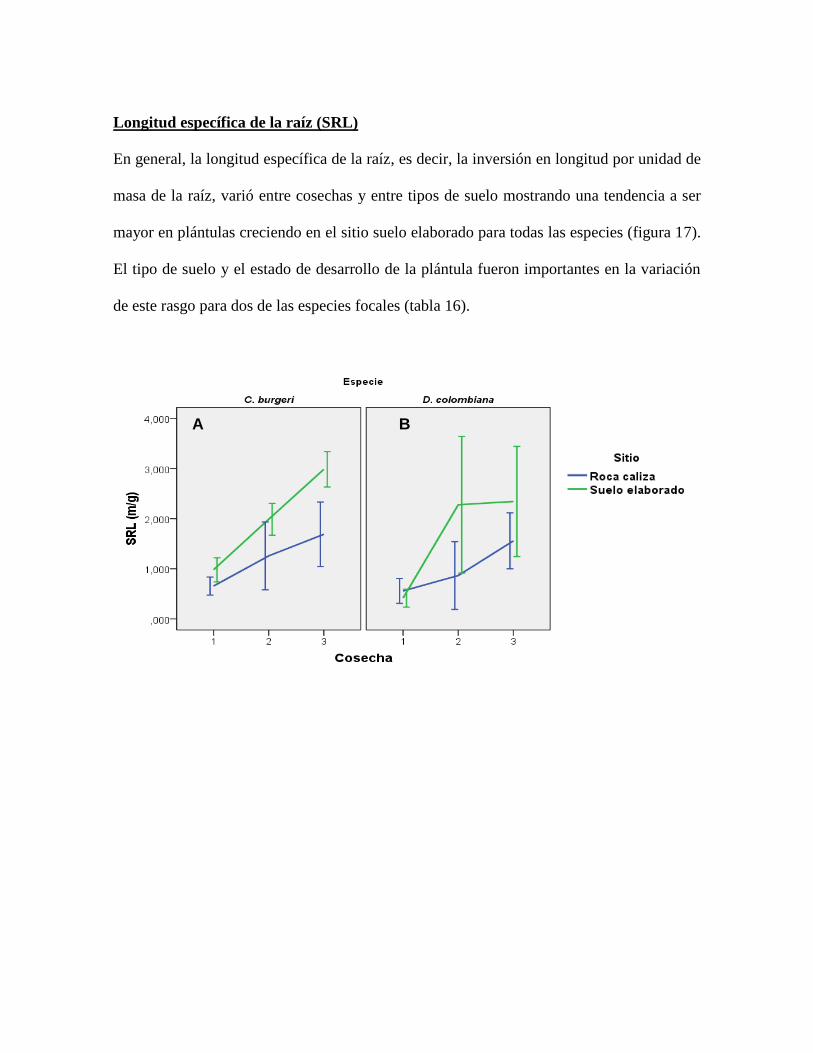
Longitud específica de la raíz (SRL)
En general, la longitud específica de la raíz, es decir, la inversión en longitud por unidad de
masa de la raíz, varió entre cosechas y entre tipos de suelo mostrando una tendencia a ser
mayor en plántulas creciendo en el sitio suelo elaborado para todas las especies (figura 17).
El tipo de suelo y el estado de desarrollo de la plántula fueron importantes en la variación
de este rasgo para dos de las especies focales (tabla 16).
B A

Figura 17. Tendencias de
SRL por especie y por tipo
de suelo a lo largo del
tiempo. Los valores
representan el promedio y
las barras representan el
error estándar
El tipo de suelo tuvo un efecto sobre SRL de la especie C. burgeri aunque este efecto fue
significativo solo en algunas cosechas (figura 17A y tabla 16). La longitud especifica de la
raíz en plántulas de D. colombiana no fue afectada por el tipo de suelo en el que las
plántulas crecieron, sin embargo mostró incrementos significativos a través del tiempo
(figura 17B y tabla 16).
En relación a la especie P. paniculata subps. pubescens se encontró que la SRL fue
significativamente diferente para los dos tipos de suelo (tabla 16); aumentó
progresivamente a través del tiempo en plántulas creciendo en el sitio suelo elaborado y lo
hizo más levemente en el sitio de roca caliza (figura 17D). De otro lado, los valores de SRL
de la especie C. brunnea se comportaron de manera similar inicialmente. Con el paso del
tiempo la SRL aumentó en plántulas del sitio suelo elaborado y disminuyó en plántulas de
D C
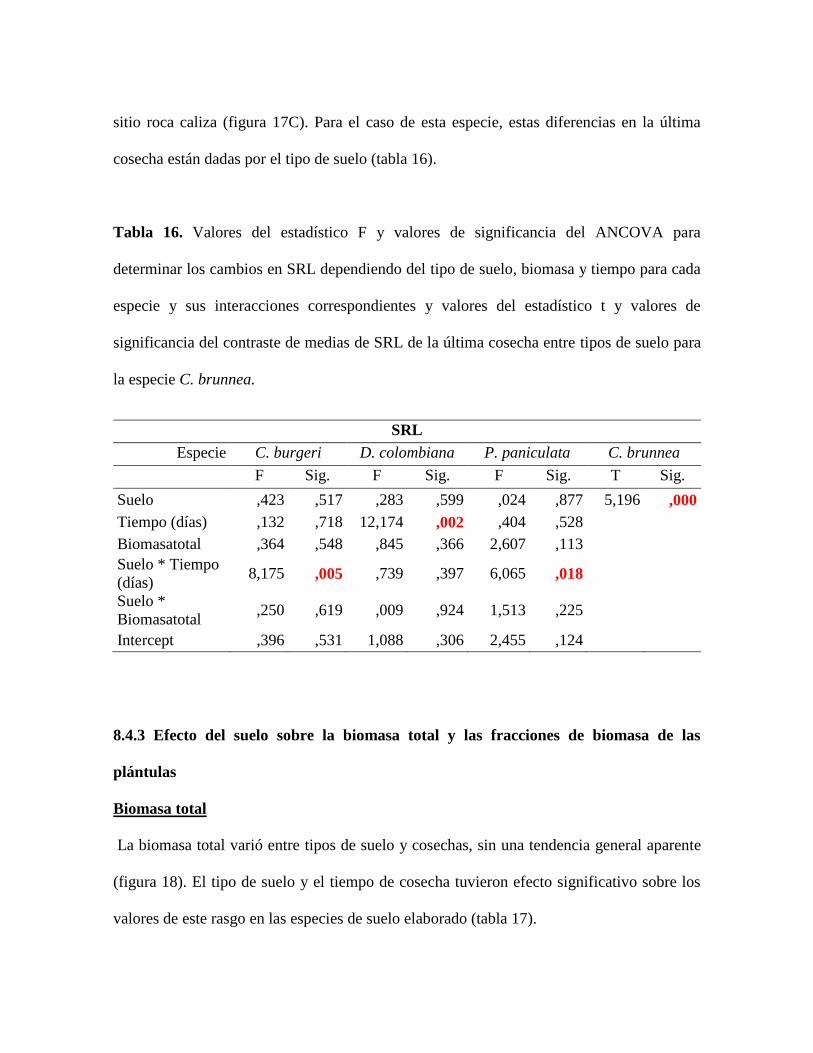
sitio roca caliza (figura 17C). Para el caso de esta especie, estas diferencias en la última
cosecha están dadas por el tipo de suelo (tabla 16).
Tabla 16. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en SRL dependiendo del tipo de suelo, biomasa y tiempo para cada
especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de SRL de la última cosecha entre tipos de suelo para
la especie C. brunnea.
SRL
Especie C. burgeri D. colombiana P. paniculata C. brunnea
F Sig. F Sig. F Sig. T Sig.
Suelo ,423 ,517 ,283 ,599 ,024 ,877 5,196 ,000
Tiempo (días) ,132 ,718 12,174 ,002 ,404 ,528
Biomasatotal ,364 ,548 ,845 ,366 2,607 ,113
Suelo * Tiempo
(días) 8,175 ,005 ,739 ,397 6,065 ,018
Suelo *
Biomasatotal ,250 ,619 ,009 ,924 1,513 ,225
Intercept ,396 ,531 1,088 ,306 2,455 ,124
8.4.3 Efecto del suelo sobre la biomasa total y las fracciones de biomasa de las
plántulas
Biomasa total
La biomasa total varió entre tipos de suelo y cosechas, sin una tendencia general aparente
(figura 18). El tipo de suelo y el tiempo de cosecha tuvieron efecto significativo sobre los
valores de este rasgo en las especies de suelo elaborado (tabla 17).

Figura 18. Tendencias
de Biomasa total por
especie y por tipo de
suelo a lo largo del
tiempo. Los valores
representan el
promedio y las barras
representan el error
estándar
El tipo de suelo tuvo un efecto sobre la biomasa total de plántulas de C. burgeri aunque este
efecto solo fue significativo en algunas cosechas (tabla 17) diluyéndose en la cosecha final
(figura 18A) y concordando con los resultados de biomasa total mostrados en la sección 8.3
(figura 9A). La especie D. colombiana mostró gran variación en la biomasa total entre
tipos de suelo y cosechas, con una tendencia opuesta en el primer estado de desarrollo
B A
D C

(figura 18B). Para esta especie el tipo de suelo no está determinando la variación de la
biomasa total (tabla 17) y esto se corresponde con lo encontrado en la sección 8.3 (figura
9B).
El tipo de suelo tuvo un efecto significativo en la biomasa total de P. paniculata subsp.
pubescens aunque este efecto estuvo determinado por el tiempo de cosecha y el estado de
desarrollo de las plántulas (tabla 17). La biomasa fue similar inicialmente en plántulas
creciendo en uno y otro sitio y posteriormente aumentó en plántulas creciendo en el sitio
suelo elaborado, mientras que la de plántulas creciendo en el sitio roca caliza disminuyó
(figura 18D).
Tabla 17. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en biomasa total dependiendo del tipo de suelo, biomasa y tiempo
para cada especie y sus interacciones correspondientes y valores del estadístico t y valores
de significancia del contraste de medias de biomasa total de la última cosecha entre tipos de
suelo para la especie C. brunnea.
Biomasa total
Especie C. burgeri D. colombiana P. paniculata C. brunnea
F Sig. F Sig. F Sig. T Sig.
Suelo 2,967 ,089 1,377 ,249 14,543 ,000 7,350 ,000
Tiempo (días) ,024 ,877 1,606 ,214 32,388 ,000
Suelo * Tiempo
(días) 4,571 ,036 1,601 ,215 41,372 ,000
Intercept 144,320 ,000 17,461 ,000 6,391 ,015
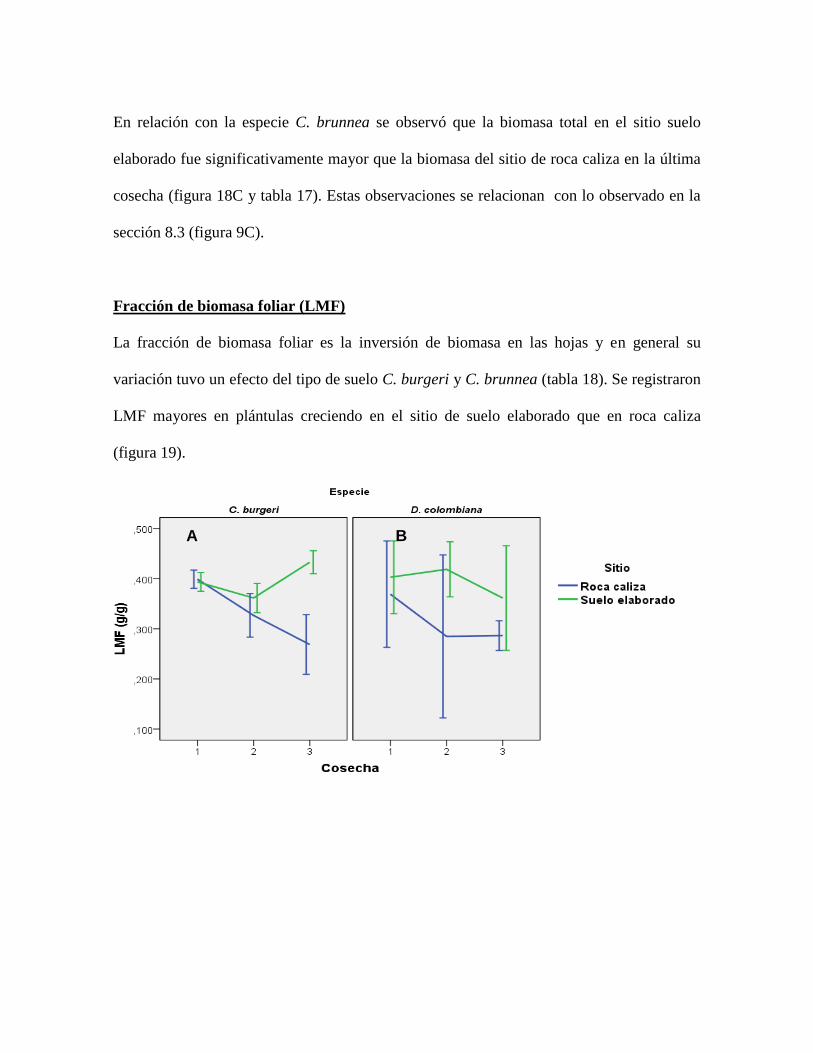
En relación con la especie C. brunnea se observó que la biomasa total en el sitio suelo
elaborado fue significativamente mayor que la biomasa del sitio de roca caliza en la última
cosecha (figura 18C y tabla 17). Estas observaciones se relacionan con lo observado en la
sección 8.3 (figura 9C).
Fracción de biomasa foliar (LMF)
La fracción de biomasa foliar es la inversión de biomasa en las hojas y en general su
variación tuvo un efecto del tipo de suelo C. burgeri y C. brunnea (tabla 18). Se registraron
LMF mayores en plántulas creciendo en el sitio de suelo elaborado que en roca caliza
(figura 19).
B A
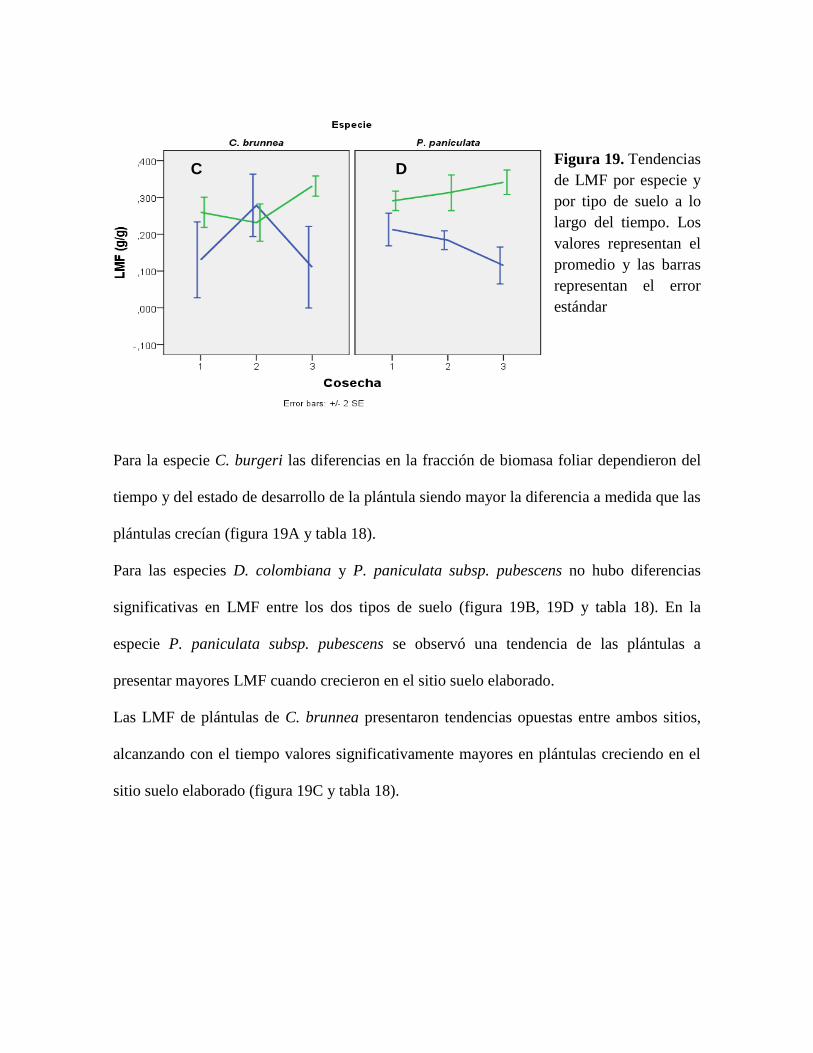
Figura 19. Tendencias
de LMF por especie y
por tipo de suelo a lo
largo del tiempo. Los
valores representan el
promedio y las barras
representan el error
estándar
Para la especie C. burgeri las diferencias en la fracción de biomasa foliar dependieron del
tiempo y del estado de desarrollo de la plántula siendo mayor la diferencia a medida que las
plántulas crecían (figura 19A y tabla 18).
Para las especies D. colombiana y P. paniculata subsp. pubescens no hubo diferencias
significativas en LMF entre los dos tipos de suelo (figura 19B, 19D y tabla 18). En la
especie P. paniculata subsp. pubescens se observó una tendencia de las plántulas a
presentar mayores LMF cuando crecieron en el sitio suelo elaborado.
Las LMF de plántulas de C. brunnea presentaron tendencias opuestas entre ambos sitios,
alcanzando con el tiempo valores significativamente mayores en plántulas creciendo en el
sitio suelo elaborado (figura 19C y tabla 18).
D C

Tabla 18. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en LMF dependiendo del tipo de suelo, biomasa y tiempo para cada
especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de LMF de la última cosecha entre tipos de suelo para
la especie C. brunnea.
LMF
Especie C. burgeri D. colombiana P. paniculata C. brunnea
F Sig. F Sig. F Sig. t Sig.
Suelo 5,156 ,026 ,352 ,558 ,300 ,587 3,856 ,010
Tiempo (días) 2,554 ,114 ,171 ,682 ,068 ,796
Biomasa total 6,665 ,012 ,401 ,531 ,392 ,534
Suelo * Tiempo
(días) 25,454 ,000 ,015 ,903 2,527 ,119
Suelo * Biomasa
total 14,020 ,000 ,036 ,852 ,223 ,639
Intercepto ,717 ,400 15,909 ,000 4,138 ,048
Fracción de biomasa del tallo (SMF)
El rasgo funcional SMF, es decir, la inversión de biomasa en el tallo, varió entre cosechas y
tipo de suelo en todas las especies, con tendencia a ser mayor en plántulas creciendo en el
sitio roca caliza expuesta (figura 20). El tipo de suelo tuvo un efecto significativo sobre la
variación de este rasgo para dos de las especies focales (tabla 19).

Figura 20. Tendencias
de SMF por especie y
por tipo de suelo a lo
largo del tiempo. Los
valores representan el
promedio y las barras
representan el error
estándar
La fracción de biomasa del tallo en plántulas de C. burgeri fue diferente dependiendo del
tipo de suelo. En el sitio de roca caliza hubo un aumento inicial de SMF seguido de una
leve disminución. En plántulas del sitio suelo elaborado la SMF disminuyo a través del
tiempo (figura 20A). La variación de SMF está afectada significativamente por el tipo de
suelo y cambia sus tendencias a través de la ontogenia de las plántulas (tabla 19).
B A
D C
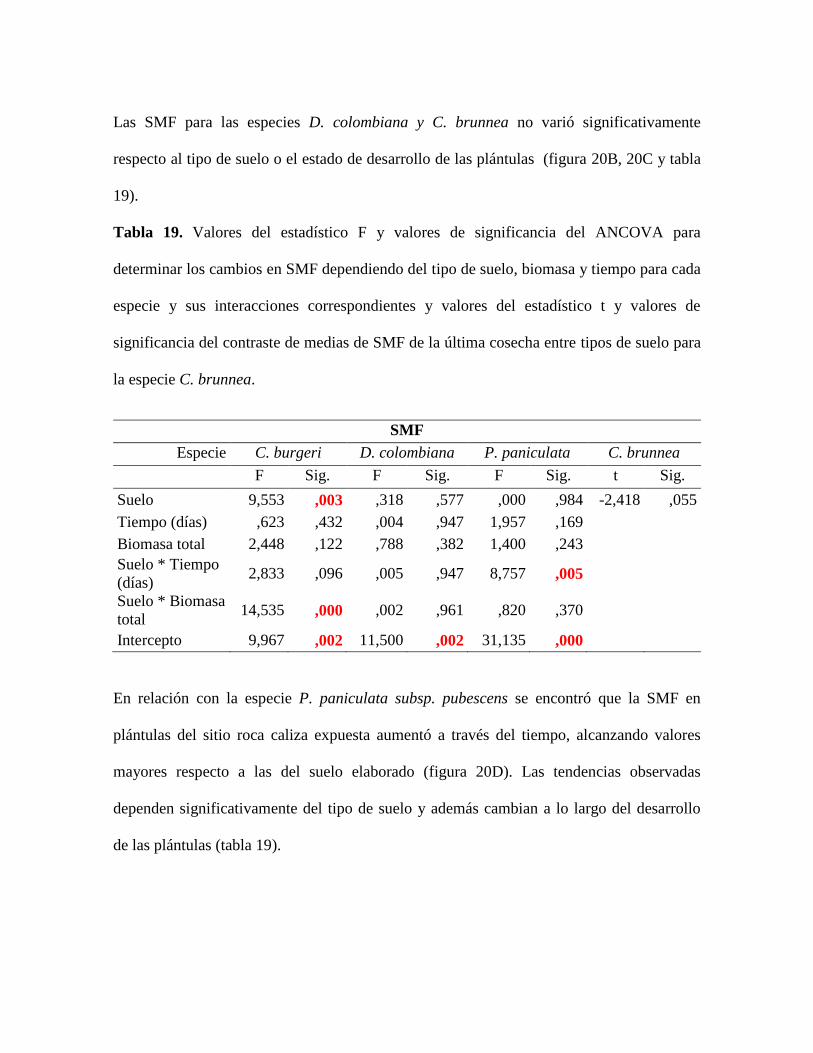
Las SMF para las especies D. colombiana y C. brunnea no varió significativamente
respecto al tipo de suelo o el estado de desarrollo de las plántulas (figura 20B, 20C y tabla
19).
Tabla 19. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en SMF dependiendo del tipo de suelo, biomasa y tiempo para cada
especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de SMF de la última cosecha entre tipos de suelo para
la especie C. brunnea.
SMF
Especie C. burgeri D. colombiana P. paniculata C. brunnea
F Sig. F Sig. F Sig. t Sig.
Suelo 9,553 ,003 ,318 ,577 ,000 ,984 -2,418 ,055
Tiempo (días) ,623 ,432 ,004 ,947 1,957 ,169
Biomasa total 2,448 ,122 ,788 ,382 1,400 ,243
Suelo * Tiempo
(días) 2,833 ,096 ,005 ,947 8,757 ,005
Suelo * Biomasa
total 14,535 ,000 ,002 ,961 ,820 ,370
Intercepto 9,967 ,002 11,500 ,002 31,135 ,000
En relación con la especie P. paniculata subsp. pubescens se encontró que la SMF en
plántulas del sitio roca caliza expuesta aumentó a través del tiempo, alcanzando valores
mayores respecto a las del suelo elaborado (figura 20D). Las tendencias observadas
dependen significativamente del tipo de suelo y además cambian a lo largo del desarrollo
de las plántulas (tabla 19).
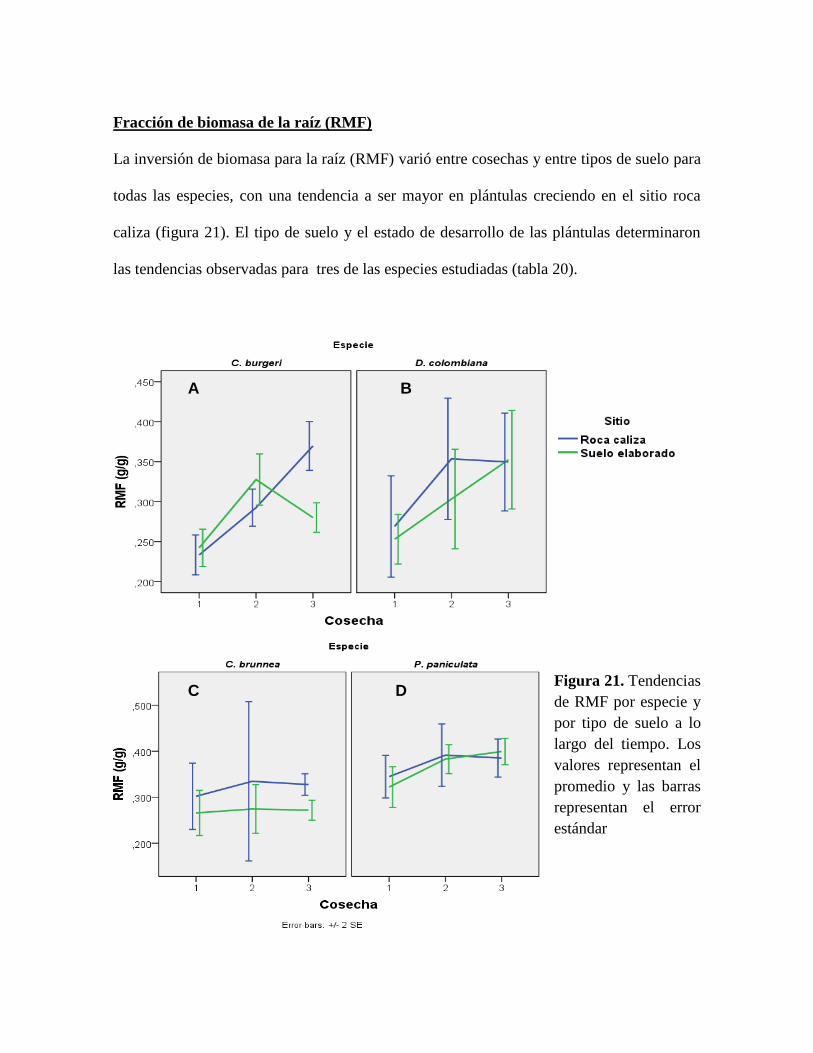
Fracción de biomasa de la raíz (RMF)
La inversión de biomasa para la raíz (RMF) varió entre cosechas y entre tipos de suelo para
todas las especies, con una tendencia a ser mayor en plántulas creciendo en el sitio roca
caliza (figura 21). El tipo de suelo y el estado de desarrollo de las plántulas determinaron
las tendencias observadas para tres de las especies estudiadas (tabla 20).
Figura 21. Tendencias
de RMF por especie y
por tipo de suelo a lo
largo del tiempo. Los
valores representan el
promedio y las barras
representan el error
estándar
B A
D C
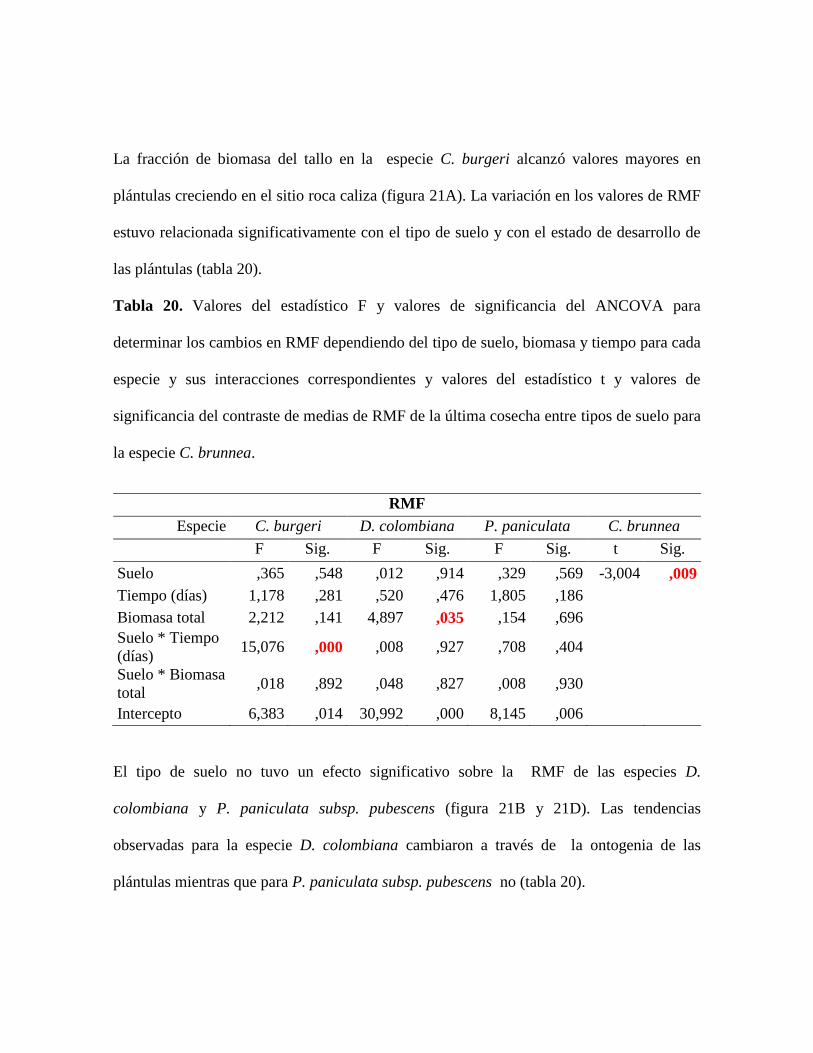
La fracción de biomasa del tallo en la especie C. burgeri alcanzó valores mayores en
plántulas creciendo en el sitio roca caliza (figura 21A). La variación en los valores de RMF
estuvo relacionada significativamente con el tipo de suelo y con el estado de desarrollo de
las plántulas (tabla 20).
Tabla 20. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en RMF dependiendo del tipo de suelo, biomasa y tiempo para cada
especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de RMF de la última cosecha entre tipos de suelo para
la especie C. brunnea.
RMF
Especie C. burgeri D. colombiana P. paniculata C. brunnea
F Sig. F Sig. F Sig. t Sig.
Suelo ,365 ,548 ,012 ,914 ,329 ,569 -3,004 ,009
Tiempo (días) 1,178 ,281 ,520 ,476 1,805 ,186
Biomasa total 2,212 ,141 4,897 ,035 ,154 ,696
Suelo * Tiempo
(días) 15,076 ,000 ,008 ,927 ,708 ,404
Suelo * Biomasa
total ,018 ,892 ,048 ,827 ,008 ,930
Intercepto 6,383 ,014 30,992 ,000 8,145 ,006
El tipo de suelo no tuvo un efecto significativo sobre la RMF de las especies D.
colombiana y P. paniculata subsp. pubescens (figura 21B y 21D). Las tendencias
observadas para la especie D. colombiana cambiaron a través de la ontogenia de las
plántulas mientras que para P. paniculata subsp. pubescens no (tabla 20).
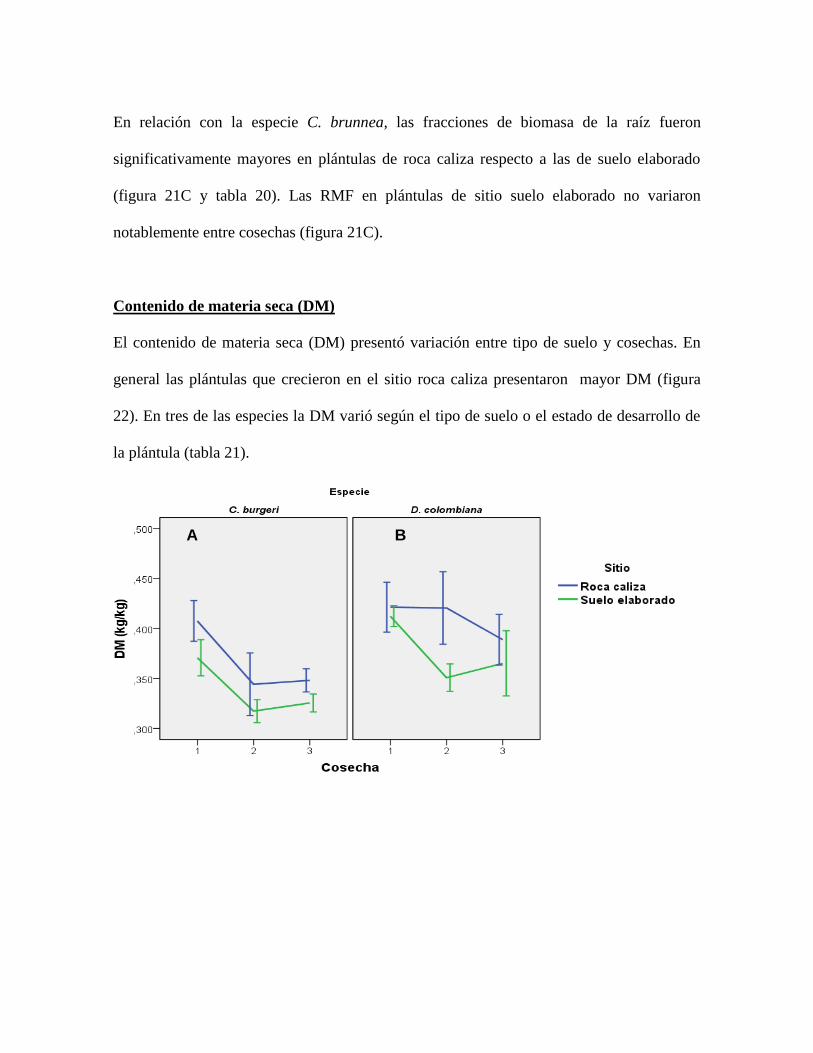
En relación con la especie C. brunnea, las fracciones de biomasa de la raíz fueron
significativamente mayores en plántulas de roca caliza respecto a las de suelo elaborado
(figura 21C y tabla 20). Las RMF en plántulas de sitio suelo elaborado no variaron
notablemente entre cosechas (figura 21C).
Contenido de materia seca (DM)
El contenido de materia seca (DM) presentó variación entre tipo de suelo y cosechas. En
general las plántulas que crecieron en el sitio roca caliza presentaron mayor DM (figura
22). En tres de las especies la DM varió según el tipo de suelo o el estado de desarrollo de
la plántula (tabla 21).
B A

Figura 22. Tendencias
de DM por especie y
por tipo de suelo a lo
largo del tiempo. Los
valores representan el
promedio y las barras
representan el error
estándar
En la especie C. burgeri se observó una tendencia de DM a disminuir y luego a aumentar
en ambos tipos de suelo a través del tiempo (figura 22A). Las diferencias que se observan
para este rasgo entre tipos de suelo y cosechas se corresponden con el hecho de que el tipo
de suelo tiene un efecto significativo sobre los valores de DM aunque también están
influenciados por el estado de desarrollo de las plántulas (tabla 21).
La DM en plántulas de D. colombiana y P. paniculta subsp. pubescens varió
significativamente según el tiempo de cosecha para la primera y dependiendo del estado de
desarrollo de las plántulas para la segunda (figura 22B, 22D y tabla 21)
Con relación a la DM registrada para la especie C. brunnea, se encontró que ésta tendió a
disminuir a través de las cosechas (figura 22C). A pesar de estas observaciones, esta
variación no fue significativa respecto al tipo de suelo o al estado de desarrollo de la
plántula (tabla 21).
D C
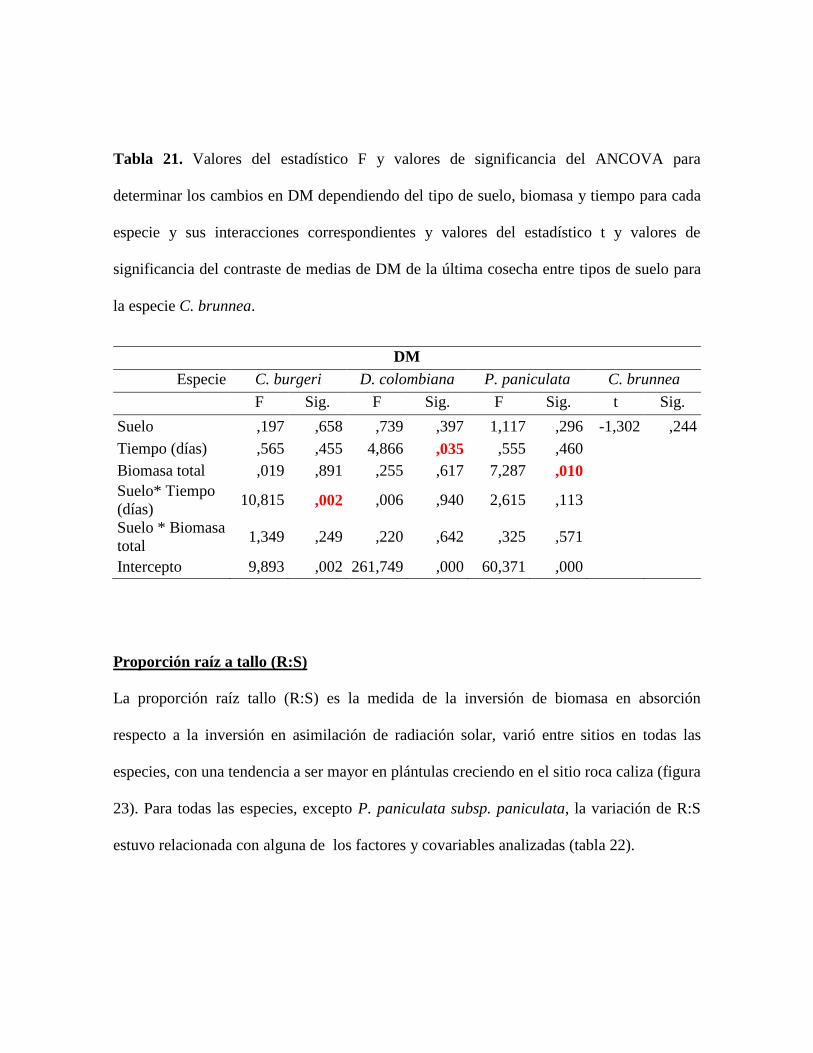
Tabla 21. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en DM dependiendo del tipo de suelo, biomasa y tiempo para cada
especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de DM de la última cosecha entre tipos de suelo para
la especie C. brunnea.
DM
Especie C. burgeri D. colombiana P. paniculata C. brunnea
F Sig. F Sig. F Sig. t Sig.
Suelo ,197 ,658 ,739 ,397 1,117 ,296 -1,302 ,244
Tiempo (días) ,565 ,455 4,866 ,035 ,555 ,460
Biomasa total ,019 ,891 ,255 ,617 7,287 ,010
Suelo* Tiempo
(días) 10,815 ,002 ,006 ,940 2,615 ,113
Suelo * Biomasa
total 1,349 ,249 ,220 ,642 ,325 ,571
Intercepto 9,893 ,002 261,749 ,000 60,371 ,000
Proporción raíz a tallo (R:S)
La proporción raíz tallo (R:S) es la medida de la inversión de biomasa en absorción
respecto a la inversión en asimilación de radiación solar, varió entre sitios en todas las
especies, con una tendencia a ser mayor en plántulas creciendo en el sitio roca caliza (figura
23). Para todas las especies, excepto P. paniculata subsp. paniculata, la variación de R:S
estuvo relacionada con alguna de los factores y covariables analizadas (tabla 22).
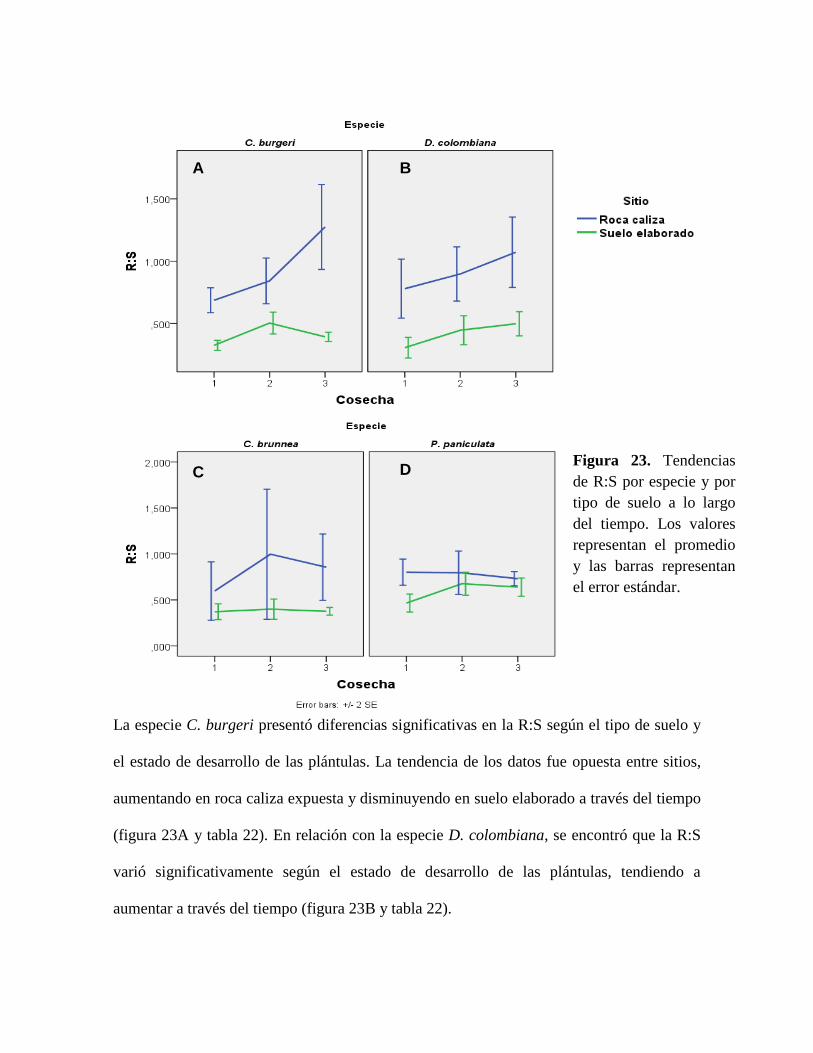
Figura 23. Tendencias
de R:S por especie y por
tipo de suelo a lo largo
del tiempo. Los valores
representan el promedio
y las barras representan
el error estándar.
La especie C. burgeri presentó diferencias significativas en la R:S según el tipo de suelo y
el estado de desarrollo de las plántulas. La tendencia de los datos fue opuesta entre sitios,
aumentando en roca caliza expuesta y disminuyendo en suelo elaborado a través del tiempo
(figura 23A y tabla 22). En relación con la especie D. colombiana, se encontró que la R:S
varió significativamente según el estado de desarrollo de las plántulas, tendiendo a
aumentar a través del tiempo (figura 23B y tabla 22).
B A
C D
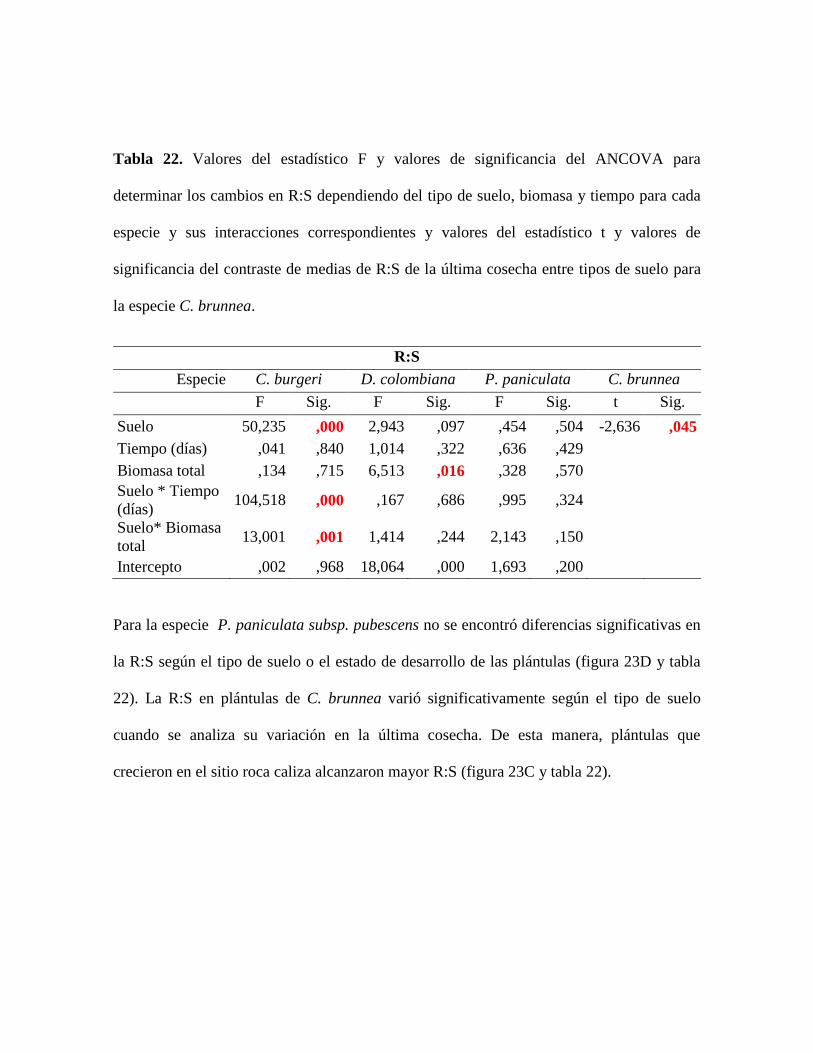
Tabla 22. Valores del estadístico F y valores de significancia del ANCOVA para
determinar los cambios en R:S dependiendo del tipo de suelo, biomasa y tiempo para cada
especie y sus interacciones correspondientes y valores del estadístico t y valores de
significancia del contraste de medias de R:S de la última cosecha entre tipos de suelo para
la especie C. brunnea.
R:S
Especie C. burgeri D. colombiana P. paniculata C. brunnea
F Sig. F Sig. F Sig. t Sig.
Suelo 50,235 ,000 2,943 ,097 ,454 ,504 -2,636 ,045
Tiempo (días) ,041 ,840 1,014 ,322 ,636 ,429
Biomasa total ,134 ,715 6,513 ,016 ,328 ,570
Suelo * Tiempo
(días) 104,518 ,000 ,167 ,686 ,995 ,324
Suelo* Biomasa
total 13,001 ,001 1,414 ,244 2,143 ,150
Intercepto ,002 ,968 18,064 ,000 1,693 ,200
Para la especie P. paniculata subsp. pubescens no se encontró diferencias significativas en
la R:S según el tipo de suelo o el estado de desarrollo de las plántulas (figura 23D y tabla
22). La R:S en plántulas de C. brunnea varió significativamente según el tipo de suelo
cuando se analiza su variación en la última cosecha. De esta manera, plántulas que
crecieron en el sitio roca caliza alcanzaron mayor R:S (figura 23C y tabla 22).

8.5 Relación entre los rasgos funcionales y el éxito de las especies.
Los análisis de regresión múltiple mostraron que algunos rasgos funcionales tienen un
impacto sobre el éxito de las especies, medido como biomasa total. Sin embargo solo
algunos rasgos tienen una relación directa con el aumento de biomasa (éxito) de las
plántulas creciendo en uno u otro tipo de suelo. LMA fue el rasgo de mayor importancia en
relación con el éxito de las plántulas, ya que presentó una relación significativa con la
biomasa de todas las especies (tabla 23).
Tabla 23. Análisis de regresión múltiple, coeficientes B y valor de significancia para la
relación entre rasgos funcionales y éxito de las especies (biomasa) de las especies
estudiadas.
Especie C. burger D. colombiana C. brunnea P. paniculata
Sitio Suelo elaborado Roca caliza Suelo
elaborado Roca caliza
Suelo
elaborado
Suelo
elaborado Roca caliza
Coef.B Valor
p
Coef
B
Valor
p
Coef
B
Valor
p
Coef
B
Valor
p CoefB
Valor
p
Coef
B
Valor
p
Coef
B
Valor
p
Constante 1,952 ,378 2,707 ,102 -,030 ,994 2,482 ,434 15,074 ,334 -,919 ,657 -,440 ,867
Altura -,010 ,435 ,000 ,961 -,009 ,800 ,030 ,255 -,022 ,495 ,007 ,332 ,010 ,196
SLA -,016 ,009 -,014 ,000 ,002 ,992 -,116 ,208 -,194 ,156 -,011 ,082 -,004 ,302
LMA ,171 ,884 6,869 ,000 4,039 ,002 3,677 ,028 18,70 ,000 2,113 ,000 2,993 ,000
Área
foliar
7,577E-
005 ,000 ,000 ,000 ,001 ,016 ,000 ,069 ,001 ,000 ,001 ,000 ,001 ,000
LDMC -,001 ,315 ,001 ,232 ,000 ,979 -,007 ,192 ,001 ,870 ,000 ,640 ,000 ,614
Resist.
foliar ,001 ,704 ,000 ,833 ,007 ,807 -,002 ,950 ,048 ,478 ,010 ,084 -,001 ,434
SRL ,036 ,131 -,007 ,623 -,158 ,309 ,020 ,508 -,223 ,433 -,060 ,020 ,001 ,962
LMF -1,53 ,488 -4,70 ,010 5,615 ,855 -4,10 ,091 -40,1 ,053 -1,14 ,617 -2,59 ,314
SMF -,093 ,964 -2,47 ,142 4,065 ,029 -2,29 ,571 -11,8 ,475 1,200 ,559 1,220 ,655
RMF -1,53 ,535 -1,74 ,281 -9,44 ,236 -21,4 ,445 -3,13 ,266 -1,77 ,456
DM ,368 ,718 -,484 ,244 -4,95 ,317 4,316 ,257 -7,02 ,284 -1,18 ,089 ,195 ,636
R:S ,521 ,361 -,038 ,579 5,370 ,101 -,173 ,792 5,109 ,572 1,806 ,053 ,663 ,046
En plántulas de C. burgeri creciendo en el sitio suelo elaborado, se encontró que los rasgos
SLA y tamaño foliar están relacionados directamente con el éxito de la especie en este
sitio, en el caso de SLA esta relación es negativa (tabla 23). El grado de relación entre los
rasgos y la biomasa fue débil (R2= 0,178 SLA y R
2= 0,313 área foliar) (figura 24 A y B).

Por otro lado, las plántulas que crecieron en el sitio roca caliza expuesta, presentaron una
relación significativa entre la biomasa total y los rasgos SLA, LMA, área foliar y LMF. La
relación de biomasa con SLA y LMF es negativa, mientras que con LMA y tamaño foliar
es positiva (tabla 23). Las asociaciones negativas presentaron relaciones con la biomasa
menos fuertes (R2 = 0,436 SLA y R
2 = 0,257 LMF) que las asociaciones positivas (R
2 =
0,738 LMA y R2 = 0,860 tamaño foliar) (figura 24 C, D, E y F).
SITIO SUELO
SITIO ROCA CALIZA EXPUESTA
A B
C D
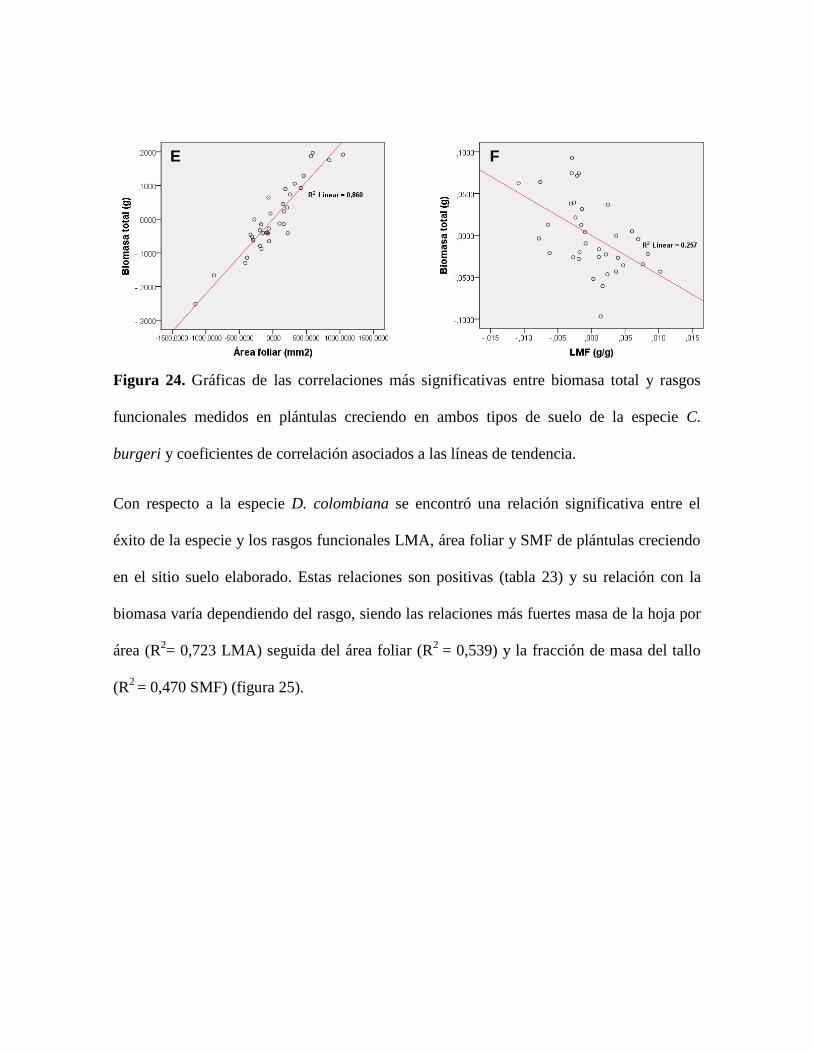
Figura 24. Gráficas de las correlaciones más significativas entre biomasa total y rasgos
funcionales medidos en plántulas creciendo en ambos tipos de suelo de la especie C.
burgeri y coeficientes de correlación asociados a las líneas de tendencia.
Con respecto a la especie D. colombiana se encontró una relación significativa entre el
éxito de la especie y los rasgos funcionales LMA, área foliar y SMF de plántulas creciendo
en el sitio suelo elaborado. Estas relaciones son positivas (tabla 23) y su relación con la
biomasa varía dependiendo del rasgo, siendo las relaciones más fuertes masa de la hoja por
área (R2= 0,723 LMA) seguida del área foliar (R
2 = 0,539) y la fracción de masa del tallo
(R2
= 0,470 SMF) (figura 25).
E F

SITIO SUELO ELABORADO
SITIO ROCA CALIZA
Figura 25. Gráficas de las correlaciones más significativas entre biomasa total y rasgos
funcionales medidos en plántulas creciendo en ambos tipos de suelo de la especie D.
colombiana y coeficientes de correlación asociados a las líneas de tendencia.
Por otra parte, el éxito de las plántulas de D. colombiana que crecieron en el sitio roca
caliza se relaciona únicamente con LMA. Esta relación es positiva (tabla 23) y
relativamente fuerte (R2=0,842) (figura 25 A, B y C).
A B
C D

Con relación a la especie C. brunnea, el éxito de las plántulas que crecieron en el sitio suelo
elaborado se relaciona significativamente con LMA, área foliar y LMF. La LMA y el área
foliar muestran una relación positiva (figura 26 A y B) mientras que LMF una relación
negativa (figura 26 C), aunque débil (R2
= 0.184), con la biomasa (tabla 23).
Figura 26. Gráficas de las correlaciones
más significativas entre biomasa total y
rasgos funcionales medidos en plántulas
creciendo en el sitio suelo elaborado de la
especie C. brunnea y coeficientes de
correlación asociados a las líneas de
tendencia.
Por lo que se refiere a la especie P. paniculata subsp. pubescens, se encontró que el éxito
de las plántulas que crecieron en el sitio suelo elaborado está relacionado de manera
significativa con LMA, área foliar y SRL (tabla 23). Los dos primeros presentan una
A B
C
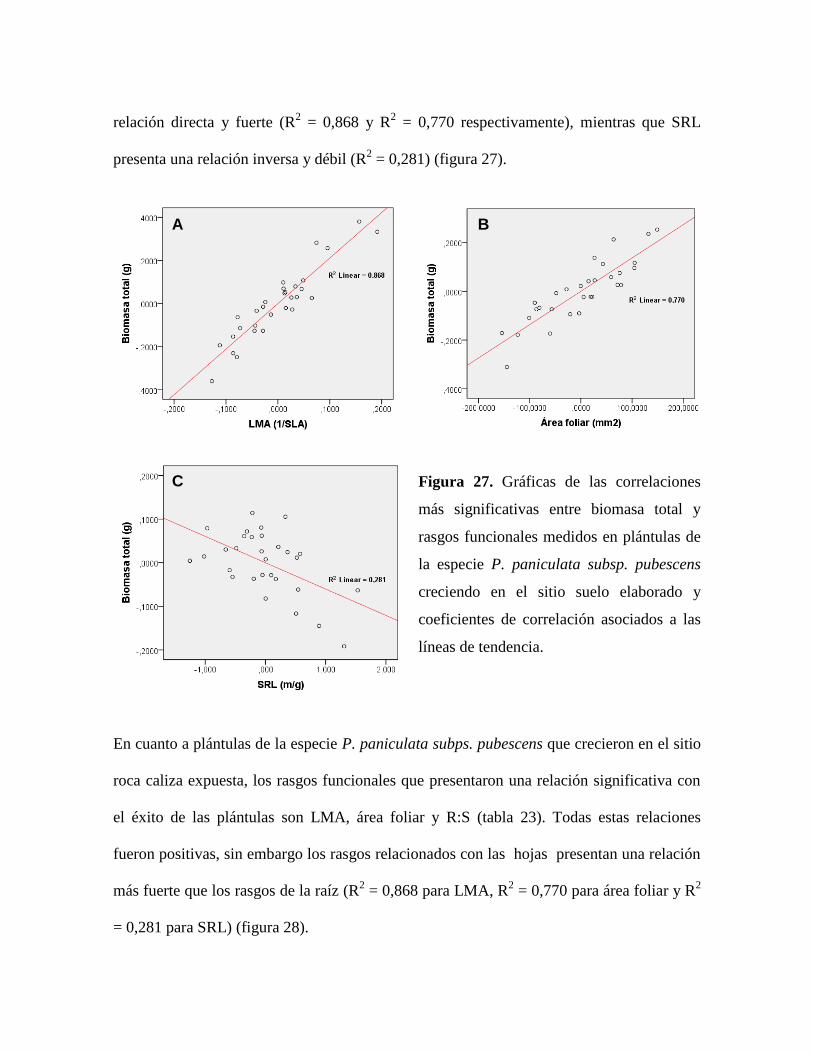
relación directa y fuerte (R2 = 0,868 y R
2 = 0,770 respectivamente), mientras que SRL
presenta una relación inversa y débil (R2 = 0,281) (figura 27).
Figura 27. Gráficas de las correlaciones
más significativas entre biomasa total y
rasgos funcionales medidos en plántulas de
la especie P. paniculata subsp. pubescens
creciendo en el sitio suelo elaborado y
coeficientes de correlación asociados a las
líneas de tendencia.
En cuanto a plántulas de la especie P. paniculata subps. pubescens que crecieron en el sitio
roca caliza expuesta, los rasgos funcionales que presentaron una relación significativa con
el éxito de las plántulas son LMA, área foliar y R:S (tabla 23). Todas estas relaciones
fueron positivas, sin embargo los rasgos relacionados con las hojas presentan una relación
más fuerte que los rasgos de la raíz (R2 = 0,868 para LMA, R
2 = 0,770 para área foliar y R
2
= 0,281 para SRL) (figura 28).
A B
C

Figura 28. Gráficas de las correlaciones
más significativas entre biomasa total y
rasgos funcionales medidos en plántulas
de la especie P. paniculata subsp.
pubescens creciendo en el sitio roca
caliza expuesta y coeficientes de
correlación asociados a las líneas de
tendencia.
A B
C

9. DISCUSIÓN
9.1 Patrón de distribución de las especies y gradiente de disponibilidad de
nutrientes
En el cañón del Río Claro existen diferencias en la disponibilidad de nutrientes en el suelo,
al menos entre los sitios denominados roca caliza expuesta y suelo elaborado. Se encontró
además que el patrón de distribución de las especies es agregado pues existen especies cuya
abundancia y frecuencia son mayores en roca caliza expuesta o en suelo elaborado. Esto es
similar a lo encontrado en la mayoría de los bosques tropicales (Hubbell 1979; Condit et al.
2000). A la luz de la teoría de diferenciación de nicho, ese patrón puede darse por un alto
grado de especialización en un tipo de hábitat (Ashton 1969; Connell 1978; Tilman 1982;
Clark et al. 1999; Hubbell 2001) o por limitaciones en la dispersión (Hubbell 2001). Se ha
encontrado que árboles y arbustos de bosques tropicales pueden presentar asociaciones con
hábitats particulares (Harms et al. 2001). El presente estudio sugiere que las especies
Caryodaphnopsis burgeri y Duguetia colombiana están asociadas a suelos de roca caliza
expuesta, confirmando lo encontrado en un estudio florístico y ecológico hecho en este
cañón (Cogollo 1986) y la información suministrada por pobladores locales. Del mismo
modo y basándose en las fuentes mencionadas anteriormente, se esperaba encontrar que las
especies Clathrotropis brunnea y Peltogyne paniculata subsp. pubescens estuvieran
asociadas a suelos elaborados en el cañón del Río Claro. Contrario a lo esperado, estas
especies no estuvieron bien representadas en el muestreo, posiblemente por el alto grado de
explotación a la que han sido sometidas durante las últimas décadas dado su característica

de maderables (Cárdenas & Salinas 2007). Las poblaciones se encuentran diezmadas ya
que la explotación ha venido en aumento significativo desde los años 80 con la apertura de
la autopista Medellín – Bogotá (Cogollo 1986). Un caso particular es que para la especie P.
paniculata subsp pubescens fueron encontrados 6 individuos, uno menos que para C.
burgeri (tabla 4). Sin embargo, la segunda resulto ser indicadora y la primera no. Esto es
posible dado que el índice de especies indicadoras tiene en cuenta la abundancia pero
también la frecuencia de los individuos (Dufrene & Legendre 1997). Para el caso de P.
paniculata subsp. pubescens los individuos encontrados se registraron solo en tres de los
diez transectos trazados.
La diferencia en disponibilidad de nutrientes de los suelos muestreados en relación con las
especies indicadoras halladas sugiere que el patrón de distribución de las especies puede
estar influenciado por factores edáficos. Esto se evidencia también por el hecho de que
además de las focales, otras especies registradas en los transectos son indicadoras de uno u
otro tipo de suelo (tabla 5). La tendencia de las especies focales a encontrarse
preferiblemente en uno u otro sitio puede deberse a que posiblemente estas presenten cierto
grado de especialización de hábitat. De hecho, la especialización de las especies como
consecuencia de gradientes de disponibilidad de nutrientes ha sido también reportada en
otras localidades (Swaine 1996; Clark et al. 1998, 1999; Svenning 2001; Lee et al. 2002;
Baltzer et al. 2005; John et al. 2007, Brenes-Arguedas 2008). Sin embargo, el patrón de
distribución de las especies puede estar influenciado por otros factores tanto bióticos como
abióticos (Hall & Swaine 1981; Mateucci 1982) y por diversos procesos como la limitación
en la dispersión (Hubbell 2001).

9.2 Éxito de las especies respecto a la disponibilidad de nutrientes en el suelo
En este estudio se evidenció que las especies estudiadas se encuentran preferiblemente en el
sitio roca caliza expuesta o suelo elaborado, lo que puede estar relacionado con posibles
adaptaciones de estas especies a la disponibilidad diferencial de nutrientes en el suelo. Esto
es sugerido por las diferencias significativas en el desempeño de plántulas entre dos tipos
de suelo con menor y mayor disponibilidad de nutrientes (tabla 24), similar a lo encontrado
en bosques tropicales de Malasia (Palmiotto et al. 2004). Las adaptaciones diferenciales de
las especies pueden estar relacionadas con una especialización de éstas, en este caso al tipo
de suelo. La especialización edáfica de las especies en bosques tropicales ha sido explicada
mediante la hipótesis del uso eficiente del recurso que indica que las especies presentan una
capacidad diferencial para resistir a condiciones de disponibilidad limitada de nutrientes
(Palmitto et al. 2004; Baltzer et al. 2005). Sin embargo, existen otras posibilidades, pues el
éxito diferencial de las especies entre diferentes hábitats también puede estar relacionado
con las adaptaciones que presenten las especies ante otros factores como la disponibilidad
de luz, las temperaturas, estrés por sequía, disturbios y presiones como la competencia intra
e interespecífica, (Denslow 1987; Hubbell 2001; Hoffmann et al. 2004), también con la
habilidad diferencial de las especies para responder ante esos filtros bióticos y abióticos
(Tilman 1988; Cornwell & Ackerly 2009); incluso con interacciones entre varios factores.
Por ejemplo, las plántulas en el bosque húmedo tropical pueden ser más sensibles a la luz
pero los nutrientes del suelo podrían jugar un papel importante en el crecimiento de éstas,
incluso en condiciones de sotobosque (Palow & Oberbauer 2009) y la distribución de

recursos puede estar asociada con interacciones entre disponibilidad de nutrientes, luz,
presiones por herbivoría y otras causas de mortalidad (Blundell & Peart 2001)
Tabla 24. Tendencias de la biomasa final y la tasa de crecimiento a través del tiempo de
plántulas de las especies focales creciendo en el sitio roca caliza expuesta y en el sitio suelo
elaborado.
ANCOVAS
Especie indicadora de creciendo en Biomasa final Tasa crecimiento
C Burgeri Roca caliza Roca caliza no cambia aumenta
C Burgeri Roca caliza Suelo elaborado no cambia aumenta
D Colombiana Roca caliza Roca caliza no cambia no cambia
D Colombiana Roca caliza Suelo elaborado no cambia aumenta
C brunnea Suelo elaborado Roca caliza menor disminuye
C brunnea Suelo elaborado Suelo elaborado mayor aumenta
P panicualata Suelo elaborado Roca caliza menor disminuye
P panicualata Suelo elaborado Suelo elaborado mayor aumenta
Relacionado tal vez con especialización en suelos elaborados, las especies C. brunnea y
P.paniculata subsp. pubescens, presentaron pobre desempeño cuando se les sometió a
condiciones de baja disponibilidad de nutrientes (roca caliza) (tabla 24), como se registró
para cuatro especies en bosques tropicales de Malasia (Palmiotto et al. 2004) y como se
espera en general para especies adaptadas a suelos ricos en nutrientes para las cuales la tasa
de crecimiento, la producción y asignación de biomasa son altamente afectadas por la
limitación de los recursos (Grime et al. 1986). Ambas especies en el sitio de roca caliza
expuesta, presentaron una tasa de mortalidad de plántulas significativa durante el
experimento, más alta en plántulas de C. brunnea (datos no mostrados, observación
personal). En el contexto de la hipótesis de eficiencia del uso del recurso, las especies

asociadas con un hábitat no serán capaces de aclimatarse lo suficiente para alcanzar la
misma eficiencia en el uso del recurso limitante como las especialistas asociadas con el
ambiente de nutrientes reducidos (Baltzer et al. 2005).
Las especies de roca caliza expuesta C. burgeri y D. colombiana no presentaron diferencias
significativas en su desempeño entre ambos tipos de suelo, como lo registraron Palmiotto y
colaboradores (2004). Sin embargo, la biomasa, en plántulas creciendo en roca caliza, fue
más variable (figura 9), esto puede estar relacionado con la heterogeneidad que se encontró
en la disponibilidad de nutrientes en los suelos de roca caliza expuesta. Es posible que si las
especies que ocurren en ambientes pobres en nutrientes han sido seleccionadas para rasgos
que promueven la eficiencia con la cual adquieren y usan los recursos limitantes (Lusk &
Matus 2000), las especies de roca caliza posiblemente se hayan especializado con
adaptaciones morfológicas o fisiológicas que les confieren ventajas en crecimiento y
sobrevivencia en el ambiente pobre en nutrientes (Lambers & Dijkstra 1987), en este caso,
el de roca caliza expuesta. La especialización en ciertos hábitats puede comprometer el
fitness en otros (Baltzer et al. 2005), sin embargo estas especies se comportaron de manera
similar en ambos tipos de suelos, alcanzado valores de biomasa parecidos y presentando
tasas de crecimiento que tendieron a incrementar en ambos tipos de suelo. Si se analiza esto
a partir de una posible especialización de estas especies en hábitats pobres en nutrientes,
cuando son sometidas a condiciones de mayor disponibilidad de nutrientes es posible que
no cambien su patrón de uso de nutrientes y crezcan a tasas similares ya que podrían estar
adaptadas a suelos de disponibilidad limitada de nutrientes. Esto es congruente con que
plantas que están restringidas a suelos infértiles, exhiben generalmente tasas de crecimiento

máximo menores y responden menos a la adición de nutrientes de lo que lo hacen plantas
de suelos más fértiles (Grime 1977; Chapin et al. 1986). Los resultados relacionados con el
éxito de las especies en roca caliza expuesta y en suelo elaborado evidencian que la
disponibilidad de nutrientes podría ser un filtro para el establecimiento de estas cuatro
especies y posiblemente de otras, en el cañón del Río Claro.
9.3 Variación de los rasgos funcionales en relación a la disponibilidad de
nutrientes en el suelo
En este estudio fue registrada una amplia variedad de valores de rasgos funcionales, tasa de
crecimiento relativo (RGR) y asignación de biomasa entre especies, que puede estar
relacionada con la disponibilidad de nutrientes del suelo en el cañón del Río Claro pero
también con otros factores bióticos y abióticos a los cuales están sometidas las especies.
Los rasgos funcionales que mostraron una variación significativa estadísticamente respecto
al suelo fueron principalmente los relacionados con la asignación de recursos en la planta,
es decir, biomasa total, fracción de biomasa foliar, fracción de biomasa del tallo, fracción
de biomasa de la raíz y la proporción raíz-tallo (tabla 25). Los rasgos masa por unidad de
área foliar (LMA), área foliar, resistencia foliar y longitud específica de la raíz (SRL)
también mostraron algún patrón respecto al tipo de suelo y durante el desarrollo de las
plántulas (tabla 25). Estas variaciones observadas incluyen respuestas en diferentes
aspectos involucrados en la relación planta – suelo. La SRL está relacionada con la
adquisición de los nutrientes; la tasa de crecimiento relativo, la LMA y el área foliar tienen
que ver con el uso que le da la planta a los nutrientes y la resistencia foliar está relacionada
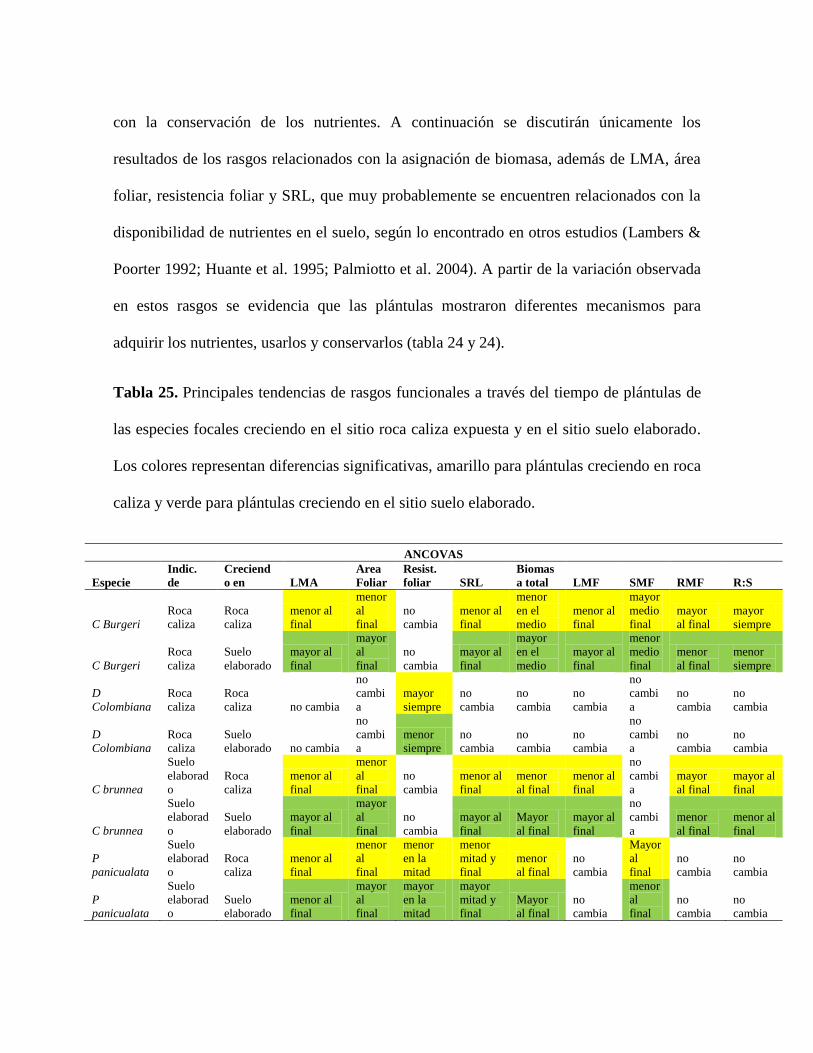
con la conservación de los nutrientes. A continuación se discutirán únicamente los
resultados de los rasgos relacionados con la asignación de biomasa, además de LMA, área
foliar, resistencia foliar y SRL, que muy probablemente se encuentren relacionados con la
disponibilidad de nutrientes en el suelo, según lo encontrado en otros estudios (Lambers &
Poorter 1992; Huante et al. 1995; Palmiotto et al. 2004). A partir de la variación observada
en estos rasgos se evidencia que las plántulas mostraron diferentes mecanismos para
adquirir los nutrientes, usarlos y conservarlos (tabla 24 y 24).
Tabla 25. Principales tendencias de rasgos funcionales a través del tiempo de plántulas de
las especies focales creciendo en el sitio roca caliza expuesta y en el sitio suelo elaborado.
Los colores representan diferencias significativas, amarillo para plántulas creciendo en roca
caliza y verde para plántulas creciendo en el sitio suelo elaborado.
ANCOVAS
Especie
Indic.
de
Creciend
o en LMA
Area
Foliar
Resist.
foliar SRL
Biomas
a total LMF SMF RMF R:S
C Burgeri
Roca
caliza
Roca
caliza
menor al
final
menor
al
final
no
cambia
menor al
final
menor
en el
medio
menor al
final
mayor
medio
final
mayor
al final
mayor
siempre
C Burgeri
Roca
caliza
Suelo
elaborado
mayor al
final
mayor
al
final
no
cambia
mayor al
final
mayor
en el
medio
mayor al
final
menor
medio
final
menor
al final
menor
siempre
D
Colombiana
Roca
caliza
Roca
caliza no cambia
no
cambi
a
mayor
siempre
no
cambia
no
cambia
no
cambia
no
cambi
a
no
cambia
no
cambia
D
Colombiana
Roca
caliza
Suelo
elaborado no cambia
no
cambi
a
menor
siempre
no
cambia
no
cambia
no
cambia
no
cambi
a
no
cambia
no
cambia
C brunnea
Suelo
elaborad
o
Roca
caliza
menor al
final
menor
al
final
no
cambia
menor al
final
menor
al final
menor al
final
no
cambi
a
mayor
al final
mayor al
final
C brunnea
Suelo
elaborad
o
Suelo
elaborado
mayor al
final
mayor
al
final
no
cambia
mayor al
final
Mayor
al final
mayor al
final
no
cambi
a
menor
al final
menor al
final
P
panicualata
Suelo
elaborad
o
Roca
caliza
menor al
final
menor
al
final
menor
en la
mitad
menor
mitad y
final
menor
al final
no
cambia
Mayor
al
final
no
cambia
no
cambia
P
panicualata
Suelo
elaborad
o
Suelo
elaborado
menor al
final
mayor
al
final
mayor
en la
mitad
mayor
mitad y
final
Mayor
al final
no
cambia
menor
al
final
no
cambia
no
cambia

Las especies C. burgeri y C. brunnea presentaron un patrón de asignación de biomasa
similar. Cuando las plántulas de ambas especies crecieron en el sitio suelo elaborado
tendieron a asignar más biomasa a las hojas y cuando crecieron en el sitio de roca caliza
expuesta asignaron más biomasa al tallo y a la raíz (tabla 25). Este es un patrón que se ha
encontrado con frecuencia en otros estudios en bosques templados (Grime et al. 1986;
Crick & Grime 1987; Campbell & Grime 1989; Shipley & Keddy 1988; Lambers &
Poorter 1992) y en bosques tropicales (Huante et al. 1995; Palmiotto et al. 2004). Estos
estudios han encontrado que las respuestas de las plantas a las variaciones en la
disponibilidad de nutrientes pueden involucrar cambios compensatorios en la asignación de
biomasa entre la raíz y el tallo (proporción raíz-tallo) con el fin de incrementar la
adquisición de recursos del suelo que están limitando el crecimiento (Bloom et al. 1985;
Chapin 1991). Esto es conocido como asignación funcional de biomasa (Poorter et al.
2011) por la cual especies creciendo en hábitats infértiles presentan rasgos que maximizan
la captura y el uso eficiente de los nutrientes, presentando por ejemplo mayor longitud
específica de la raíz y mayor fracción de biomasa para la raíz (Gleason et al. 2011;
Eissenstat 2000; Read et al. 2006). Especies de hábitats fértiles presentan rasgos que
maximizan la captura y el uso eficiente de la radiación solar aumentando por ejemplo la
fracción de biomasa foliar (Gleason et al. 2011).
El patrón observado en C. burgeri se complementa con que presentó un desempeño similar
en ambos suelos, posiblemente por las modificaciones que mostró en la asignación de
biomasa (tabla 25). Al parecer, a pesar de que mostró cambios en sus rasgos funcionales
correspondientes a lo esperado para los diferentes tipos de suelo, y de tener mayor

disponibilidad de nutrientes, las plántulas no presentaron un incremento significativo de su
biomasa en el sitio de suelo elaborado. Lo anterior sugiere poca plasticidad respecto a la
estrategia de conservación de nutrientes de esta especie. C. burgeri, en cuanto al uso de los
nutrientes, presentó áreas foliares mayores en plántulas que crecieron en el sitio de suelo
elaborado pero también mayor masa por unidad de área, es decir, las plántulas presentaron
hojas más grandes y de mayor grosor. Las plántulas posiblemente usaron este mecanismo
para aprovechar la radiación solar y para almacenar nutrientes, ya que éstos no estaban
siendo un factor limitante. Al parecer esta especie asigna una cantidad de biomasa
considerable por unidad de longitud a las raíces puesto que presenta valores bajos de
longitud específica de la raíz cuando creció en roca caliza expuesta, es decir, produjo raíces
más cortas pero más gruesas. Esto es similar a lo encontrado en plántulas de bosque seco
con baja disponibilidad de nutrientes, es posible que esta sea una estrategia para conservar
los nutrientes cuando son el recurso limitante (Markesteijn & Poorter 2009). Por el
contrario, en el sitio de suelo elaborado presentó raíces más largas pero más delgadas, esto
puede estar relacionado con que la textura que prima en los sitios de suelo elaborado es la
arcilla, esto hace que el suelo sea anóxico (Korning et al. 1994), una estrategia de la planta
podría ser extender las raíces para favorecer la aireación de las mismas. Adicionalmente,
bajo mayor disponibilidad de nutrientes, las raíces pueden proliferar para maximizar el área
de absorción (Hodge 2004), esto como una estrategia para la adquisición y búsqueda de
nutrientes (Reich et al. 1998; Hodge 2004) que requieren para mantener sus tasas de
crecimiento positivas (Markesteijn & Poorter 2009).

La especie C. brunnea, adaptada a suelos elaborados, presentó biomasas significativamente
mayores creciendo en su ambiente natural (tabla 24). Esto es congruente con lo encontrado
por Grime et al. (1986) según el cual especies adaptadas a suelos ricos en nutrientes
conservan un crecimiento rápido y alto rendimiento, mantenido por una alta tasa de
absorción de nutrientes cuando se encuentran en ambientes de recursos abundantes, sin
embargo la tasa de crecimiento y la producción y asignación de biomasa de esas especies
son altamente afectadas por la limitación de los recursos. Cuando los nutrientes son
escasos, estas especies reducen el rendimiento y la tasa de crecimiento e incrementan la
asignación de biomasa a la raíz como un mecanismo para adquirir recursos a través de
ajustes morfológicos (Grime et al. 1986). Aunque C. brunnea mostró algunos cambios en la
asignación de biomasa, al parecer no fueron suficientes para que tuviera éxito en el sitio de
roca caliza expuesta (tabla 24 y 25). Es posible además que las especies adaptadas al sitio
roca caliza presenten asociaciones micorricicas o con endosimbiontes fijadores de
nitrógeno que C. brunnea no posea. Plántulas que crecieron en el sitio suelo elaborado
presentaron áreas foliares mayores y mayor cantidad de masa por área, es decir más
densidad de tejido foliar, respecto a plántulas que crecieron en roca caliza donde hay menor
cantidad de elementos como N, fundamental en la formación de tejidos estructurales que le
confieren grosor a la hoja. De esta manera las plántulas que crecieron en el sitio de suelo
elaborado alcanzaron valores de biomasa total mayores. Posiblemente sus tasas de
crecimiento y producción fueron favorecidas y hubo mayor asignación de biomasa a las
hojas, dada la buena disponibilidad de nutrientes. Por otro lado, plántulas que crecieron en
el sitio roca caliza no pudieron tener éxito probablemente por la baja disponibilidad de
nitrógeno y materia orgánica, elementos esenciales de compuestos estructurales,

metabólicos y genéticos, fundamentales para el crecimiento de la planta (Taiz & Zeiger
2006). Ante esa situación posiblemente invirtieron mayor biomasa en la raíz en busca de
nutrientes y menos en área foliar y en masa foliar por área, ante la baja disponibilidad de
recursos para ello. A pesar de usar esta posible estrategia, no lograron obtener los nutrientes
necesarios para tener tasas de crecimiento y producción de biomasa positivas.
Por otro lado, el comportamiento de la longitud específica de la raíz fue contrario a lo
esperado, pues se ha encontrado que especies que crecen en hábitats de baja disponibilidad
de nutrientes generalmente tienen mayor longitud específica de la raíz (Lambers & Poorter
1992; Reich et al. 1998; Hodge 2004). En este caso, plántulas que crecieron en suelo
elaborado presentaron una longitud específica de la raíz mayor que las que crecieron en
roca caliza expuesta (tabla 25). Se ha registrado que especies que ocurren en suelos
infértiles, pueden presentar raíces gruesas y densas; este mecanismo está relacionado con
conservación de los nutrientes (Reich et a1. 1998; Markesteijn & Poorter 2009).
Adicionalmente se ha registrado que cuando las raíces encuentran una porción de suelo rico
en nutrientes, proliferan dentro de este, es decir, aumentando el número de raíces laterales o
la longitud de las ya formadas, maximizando el área de absorción (Hodge 2004). En
especies anuales de hábitats fértiles se ha registrado una alta longitud específica de la raíz,
que ha sido asociada con la adquisición y búsqueda de nutrientes (Reich et al. 1998; Hodge
2004), maximizando el área de intercambio con el suelo, permitiendo así un rápido acceso a
los recursos minerales. Es posible también que, como otras especies de bosque húmedo, las
especies focales demanden más nutrientes y agua, debido a su capacidad fotosintética
mayor y a altas tasas de crecimiento que son inherentes a ellas (Markesteijn & Poorter

2009). La especie D. colombiana fue un caso particular al analizar de qué manera respondió
al uso de los nutrientes respecto a la asignación de biomasa, pues no presento variaciones
significativas en casi ningún rasgo excepto en la resistencia foliar (tabla 25). Aun así,
mostró un desempeño similar en ambos tipos de suelo. Es posible que los mecanismos
asociados a su adaptación al sitio roca caliza sean funcionales para esta especie al verse
sometida a un ambiente de mayor disponibilidad de nutrientes o que los mecanismos para
adaptarse al sitio de suelo elaborado sean otros que no fueron medidos en este estudio. En
algunas investigaciones se ha encontrado que especies de suelos con baja disponibilidad de
nutrientes son menos plásticas en rasgos como área foliar específica y asignación de
biomasa a la raíz y muestran acumulación de nutrientes en el tejido vegetal y bajas tasas de
recambio de tejidos en conjunto con un sistema radicular longevo (Grime et al. 1986; Crick
& Grime 1987; Coomes & Grubb 2000; Palmiotto et al., 2004). Adicionalmente, se ha
predicho que en respuesta al enriquecimiento en nutrientes, especies adaptadas a suelos
infértiles usualmente muestran cambios más pequeños en sus tasas de crecimiento y
absorción (Chapin 1980; Shipley & Keddy 1988). Es posible que el caso de D. colombiana
se corresponda con lo encontrado en especies de suelos pobres en nutrientes, que
maximizan la toma de nutrientes más a través de una proporción raíz tallo constante y
posibles asociaciones con micorrizas que a través de una alta capacidad de absorción
radicular (Chapin 1980, 1988). Además, la distribución de recursos puede estar asociada
con interacciones entre disponibilidad de nutrientes, luz, herbívoros y otras causas de
mortalidad (Blundell & Peart 2001), que podrían estar influyendo sobre las respuestas de
esta especie. A pesar de mostrar poca variación en los rasgos asociados a la asignación de
biomasa, esta especie mostró diferencias significativas en la resistencia foliar, rasgo

asociado a la conservación de los nutrientes. Las plántulas que crecieron en el sitio de roca
caliza presentaron una resistencia foliar mayor, es decir, mayor densidad de tejidos
esclerenquimáticos y estructurales, esta puede ser una adaptación de esta especie a su
hábitat natural, pues se sabe que especies de suelos infértiles tienden a tener hojas con
mayor contenido de fibras esclerenquimáticas (Read et al. 2005) como una estrategia de
conservación de nutrientes o como una respuesta a presiones por ataques de herbívoros
(Coley & Barone, 1996; Schemske et al. 2009).
Con respecto a la especie P. paniculata subsp. pubescens se observó, que para los rasgos
relacionados con la asignación de biomasa, no presentó variaciones significativas (tabla
25). Esto puede estar relacionado con que las especies difieren en su capacidad para
responder a los cambios ambientales (Valencia et al. 2004) y que posiblemente sus
mecanismos de adaptación no hayan sido medidos en este estudio. Es posible que esta
especie no sea tan plástica en rasgos relacionados con asignación de biomasa pero si en
otros como área foliar específica, resistencia foliar, longitud específica de la raíz y área
foliar. Por ejemplo, similar a los observado para esta especie, cuando crecen bajo estrés por
nutrientes, especies asociadas con ambientes fértiles muestran un área foliar específica
(SLA) más alta que la de especies relacionadas con ambientes infértiles (Huante et al. 1995,
Pérez-Harguindeguy et al. 2013). En este caso, al parecer, el aumento de la fracción de
biomasa foliar se reflejó en un aumento del área foliar cuando creció en el sitio suelo
elaborado. De esta manera mostró el patrón esperado en plantas de suelos fértiles:
maximizó la captura y el uso eficiente de la radiación solar, que en su hábitat natural, es el
recurso limitante (Poorter et al. 2011; Gleason et al. 2011)

En todas las especies se observaron cambios en los rasgos funcionales a través del tiempo;
los requerimientos de nutrientes de las especies pueden cambiar durante su ciclo de vida y
pueden estar influenciadas por otros factores incluyendo micorrizas y asociaciones con
endosimbiontes fijadores de nitrógeno (Huante at al. 1993). En otros estudios se ha
encontrado que los rasgos pueden cambiar a medida que las plantas incrementan en tamaño,
por ejemplo, la tasa fotosintética decrece (Suzuki & Tsukahara 1987) y la asignación de
biomasa al tallo aumenta con la edad de la planta (Belford & Tennant 1990). Alteraciones
fisiológicas, morfológicas y de asignación de biomasa pueden ser el resultado de un
incremento en el tamaño de la planta, requiriendo una inversión extra en tejido de soporte
(Givnish 1986). Adicionalmente, cambios ambientales que generen una alteración de la
transpiración o del balance de carbono pueden influir sobre el crecimiento de las plántulas
(Poorter & Pothmann 1992)
9.4 Relación entre los rasgos funcionales y el éxito de las especies
El éxito de las especies en uno u otro tipo de suelo estuvo correlacionado
significativamente con rasgos foliares, principalmente masa foliar por unidad de área
(LMA) y área foliar (tabla 26). Para todas las especies el área foliar y la masa foliar por
unidad de área tuvieron un impacto sobre el éxito de las plántulas que crecieron en el sitio
roca caliza. Se ha encontrado que la acumulación de biomasa en plantas depende de varios
factores incluyendo, entre otros, número de hojas, área foliar, capacidad fotosintética de las
hojas, capacidad de acumular o usar eficientemente los nutrientes y la disponibilidad de
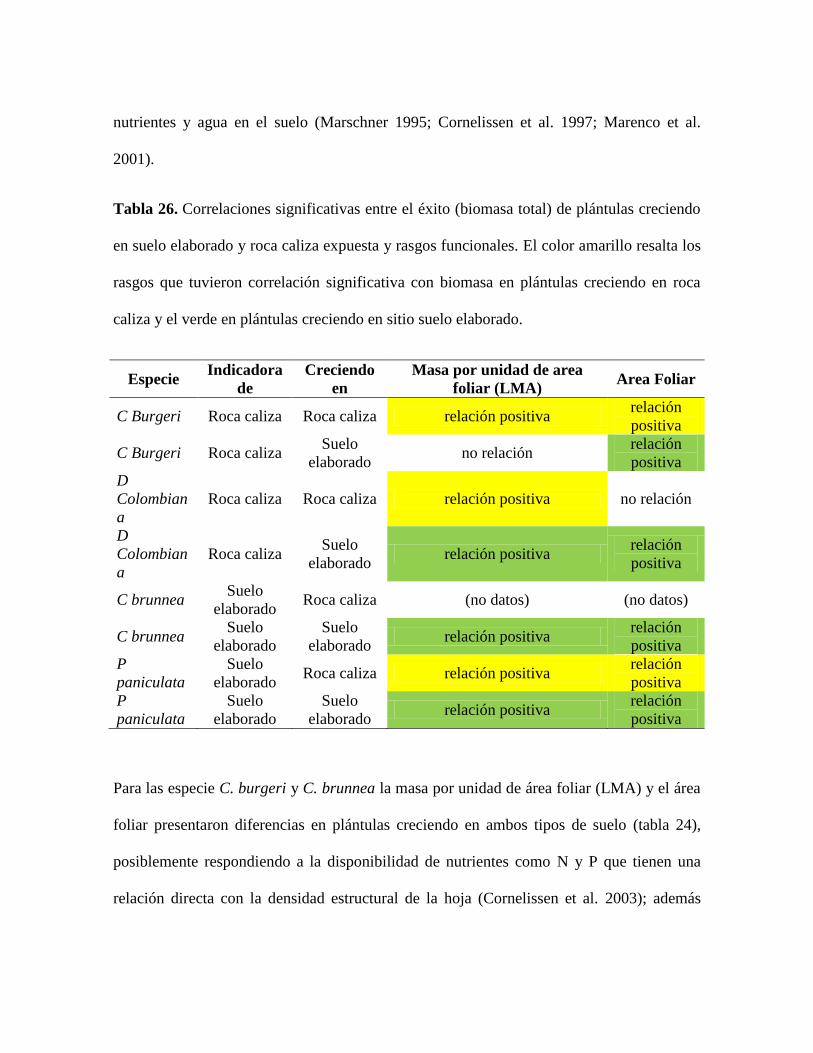
nutrientes y agua en el suelo (Marschner 1995; Cornelissen et al. 1997; Marenco et al.
2001).
Tabla 26. Correlaciones significativas entre el éxito (biomasa total) de plántulas creciendo
en suelo elaborado y roca caliza expuesta y rasgos funcionales. El color amarillo resalta los
rasgos que tuvieron correlación significativa con biomasa en plántulas creciendo en roca
caliza y el verde en plántulas creciendo en sitio suelo elaborado.
Especie Indicadora
de
Creciendo
en
Masa por unidad de area
foliar (LMA) Area Foliar
C Burgeri Roca caliza Roca caliza relación positiva relación
positiva
C Burgeri Roca caliza Suelo
elaborado no relación
relación
positiva
D
Colombian
a
Roca caliza Roca caliza relación positiva no relación
D
Colombian
a
Roca caliza Suelo
elaborado relación positiva
relación
positiva
C brunnea Suelo
elaborado Roca caliza (no datos) (no datos)
C brunnea Suelo
elaborado
Suelo
elaborado relación positiva
relación
positiva
P
paniculata
Suelo
elaborado Roca caliza relación positiva
relación
positiva
P
paniculata
Suelo
elaborado
Suelo
elaborado relación positiva
relación
positiva
Para las especie C. burgeri y C. brunnea la masa por unidad de área foliar (LMA) y el área
foliar presentaron diferencias en plántulas creciendo en ambos tipos de suelo (tabla 24),
posiblemente respondiendo a la disponibilidad de nutrientes como N y P que tienen una
relación directa con la densidad estructural de la hoja (Cornelissen et al. 2003); además

existe evidencia de que hay una variación coordinada de rasgos de la raíz y de la hoja
respecto a la disponibilidad de nutrientes en el suelo, por ejemplo la concentración de
nitrógeno y fósforo disponible en el suelo y acumulado en las hojas están relacionados
(Kerkhoff et al. 2006; Reich et al. 2008). De esta manera, el éxito de plántulas en suelos
pobres en nutrientes puede ser alcanzado por un incremento en la asignación de biomasa
para la raíz junto con rasgos foliares que les confieran eficiencia en la adquisición de luz,
como por ejemplo mayor área foliar para incrementar la adquisición de radiación solar
(Gleason et al. 2011). En el caso de las especies estudiadas, un conjunto de mecanismos
que incluyen la reasignación de biomasa y la plasticidad en rasgos foliares que responden
tanto a la adquisición de nutrientes como a la de radiación solar, están determinando el
éxito de las especies en uno u otro tipo de suelo. Para las especies de suelo elaborado que
no lograron tener éxito en el sitio de roca caliza, estos mecanismos no fueron suficientes y
su especialización en sitios ricos en nutrientes comprometió su fitness cuando crecieron
bajo poca disponibilidad de nutrientes. Un ejemplo de esto es que el estrés por un déficit en
nutrientes y el estrés por alta radiación tienden a seleccionar hojas de áreas relativamente
pequeñas (Pérez-Harguindeguy et al. 2013).
Los rasgos foliares están altamente influenciados por la disponibilidad de luz. Las plántulas
tienden a aumentar la intercepción de la luz produciendo hojas relativamente grandes y
delgadas con una baja masa por unidad de área foliar (LMA) (Evans and Poorter 2001).
Sin embargo, en muchas instancias, la asignación de biomasa a las hojas no es
particularmente sensible al aumento de la radiación y no es un factor importante respecto al
cambio en la tasa de crecimiento de la planta (Poorter & Nagel 2000). Es posible que en el

caso de las especies estudiadas, los rasgos foliares estén respondiendo también a la
disponibilidad de nutrientes, y que ésta sea una presión importante sobre la respuestas de
las plántulas.
Especies con baja LMA, como C brunnea creciendo en roca caliza, tienden a presentar
mayor contenido de células del mesófilo metabólicamente activas, más que células con
paredes gruesas y deposiciones de cutículas (Shipley et al. 2006). El aumento del grosor
del mesófilo implica un incremento en el número de cloroplastos y la cantidad de enzimas
fotosintéticas y de esta manera un aumento de la capacidad fotosintética de la hoja por
unidad de área foliar, lo que permite mayor productividad (Poorter & Nagel 2000). Para la
especie C. burgeri el mecanismo para tener éxito en el sitio roca caliza posiblemente
incluye producir hojas de área foliar pequeña y densidad de tejido alta ya que posiblemente
tengan mayor disponibilidad de radiación solar que en el sitio suelo elaborado (resultados
no mostrados), de esta manera pueden emplear una estrategia conservativa de los recursos,
que puede incluir mayor asignación de biomasa para la raíz y para la hoja en forma de
tejidos más densos, como respuesta a un déficit en la disponibilidad de nutrientes en el sitio
de roca caliza expuesta. Especies de sitios pobres en nutrientes generalmente tienen altos
valores de contenido de material seca foliar y altas concentraciones de paredes celulares y
compuestos secundarios, como una estrategia para conservar los recursos que puedan
adquirir (Coley et al. 1985; Reich 1998; Poorter & Garnier 1999). En el sitio de suelo
elaborado el mecanismo, ante menor disponibilidad de luz (resultados no mostrados), fue
aumentar el área foliar junto con el contenido de masa foliar, aprovechando los nutrientes

disponibles y como una posible estrategia para aprovechar la radiación solar, manteniendo
así sus tasas de crecimiento a pesar de crecer con menor disponibilidad de luz.
En el caso de C. brunnea, la variación del área foliar se corresponde con la disponibilidad
de luz en ambos sitios, siendo mayor en el sito de suelo elaborado, donde los nutrientes no
son el recurso limitante. Respecto a la masa por unidad de área foliar que fue menor en el
sitio roca caliza, es posible que debido al bajo desempeño de esta especie ante la baja
disponibilidad de nutrientes, debió reasignar los recursos a la raíz, impidiendo la
producción de hojas de mayor densidad estructural.
En el caso de D. colombiana la LMA tuvo un impacto sobre la biomasa total o éxito de las
plántulas que crecieron en el sitio suelo elaborado (tabla 26). Esta relación puede verse
reflejada en un aumento, aunque no significativo en LMA en suelo elaborado, acompañado
con un incremento en su tasa de crecimiento relativo. Una mayor inversión en tejido
fotosintético podría contribuir a un aumento de la capacidad fotosintética y así de la
productividad en términos de biomasa (Poorter & Nagel 2000). Esto también puede aplicar
para P. paniculata subsp. pubescens, para la cual, además de la LMA, el área foliar también
tuvo un impacto sobre el éxito de plántulas creciendo en suelo elaborado. Es posible que el
aumento del área foliar esté relacionado con la disponibilidad de luz en este sitio, pues, en
general, las hojas tienden a presentar mayor área ante una baja disponibilidad de luz (Pérez-
Harguindeguy et al. 2013). En este caso la especie estaría direccionando su biomasa al
aprovechamiento de la luz que es el recurso limitante, y ya que hay buena disponibilidad de
nutrientes en el sitio de suelo elaborado.

10. CONCLUSIONES
La variación en algunos rasgos funcionales medidos en este estudio, constituye un posible
mecanismo de respuesta de las plantas a la disponibilidad de nutrientes, que les puede
conferir un mejor desempeño en el sitio suelo elaborado o roca caliza expuesta en el cañón
del Río Claro.
Algunas especies responden más que otras a los cambios en la disponibilidad de nutrientes
del suelo en el cañón del Río Claro, de esta manera fue posible identificar dos estrategias.
La conservativa, identificada en Caryodaphnopsis burgeri y Duguetia colombiana, propia
de especies que conservan los nutrientes aumentando el grosor de tejidos estructurales y
que son menos plásticas en la asignación de biomasa. Por otro lado la adquisitiva,
identificada en Clathrotropis brunnea y Peltogyne paniculata subsp. pubescens, encontrada
en especies que usan los nutrientes disponibles para mantener sus tasas de crecimiento
positivas y que son más plásticas en la asignación de biomasa, respondiendo a la buena
disponibilidad de nutrientes en el suelo.
En el cañón del Río claro, donde existe una diferencia en la disponibilidad de nutrientes en
el suelo, los rasgos funcionales que mostraron cambios significativos en dos suelos
contrastantes fueron algunos asociados a la asignación de biomasa (fracción de biomasa de
la hoja, del tallo, de la raíz y proporción raíz-tallo), a la adquisición de nutrientes (longitud
específica de la raíz) y al uso de los nutrientes (masa por unidad de área foliar y área foliar).
Éstos posiblemente son los más relacionados con las adaptaciones de las plantas a la
disponibilidad de nutrientes del suelo en éste lugar.

La disponibilidad de nutrientes del suelo en el cañón del Río Claro puede influir sobre el
éxito de las especies vegetales en sitios de suelo elaborado o de roca caliza expuesta. El
éxito de las especies puede estar dado por diferencias en rasgos funcionales relacionados
con la adquisición, uso y conservación de los nutrientes. Especialmente, por adaptaciones a
nivel de la hoja, pues rasgos foliares relacionados con la disponibilidad de nutrientes y
adquisición de radiación solar, están correlacionadas significativamente con el éxito de las
especies en uno u otro tipo de suelo. El establecimiento de las especies en el cañón del Río
Claro parece estar mediado por la disponibilidad de nutrientes en el suelo, por lo tanto este
es un factor que tiene una influencia importante sobre los patrones de distribución espacial
de árboles en el cañón del Río Claro y de esta manera puede estar jugando un papel clave
en la estructuración de las comunidades vegetales de este bosque tropical.

11. RECOMENDACIONES
Otros rasgos funcionales como contenido foliar de nutrientes, densidad del tallo,
asociaciones de tipo micorriza, herbivoría, entre otro, pueden ser de importancia para
corroborar la posible relación entre la distribución de las especies y la disponibilidad de
nutrientes en el suelo, así como la variación de rasgos funcionales como posibles
mecanismos de respuesta al suelo y por lo tanto para la identificación de especies
especialistas de uno otro tipo de suelo en el cañón del Río Claro.
Es interesante y necesario llevar a cabo un experimento de mayor duración para tratar de
separar el efecto del estado de desarrollo de las plántulas del efecto de la disponibilidad de
nutrientes sobre la respuesta de las plantas, medida en este caso mediante algunos rasgos
funcionales.

REFERENCIAS BIBLIOGRÁFICAS
Ackerly D. 2003.Community assembly, niche conservatism, and adaptive evolution in
changing environments. International Journal of Plant Sciences, 164:165-184.
Aerts R & Chapin FS. 2000. The mineral nutrition of wild plants revisited: a re-evaluation
of processes and patterns. Advances in Ecological Research, 30:1–67.
Andersen KM, Corre MD, Turner BL, Dalling JW. 2010. Plant-soil associations in a lower
montane tropical forest: physiological acclimation and herbivore-mediated responses
to nitrogen addition. Functional Ecology, 24:1171–1180.
Ashton P S. 1969. Speciation among tropical forest trees: some deductions in the light of
recent evidence. Biological Journal of the Linnean Society, 1:155–196.
Ashton PS & Hall P. 1992. Comparisons of structure among mixed dipterocarp forests of
northwestern Borneo. Journal of Ecology, 80:459–481.
Baltzer JL, Thomas C, Nilus R, Burslem D. 2005. Edaphic specialization in Tropical Trees:
Physiological Correlates and Responses to reciprocal transplantation. Ecology, 86
(11):3063-3077.
Barreto J, Duque A, Cárdenas D, Moreno H. 2010. Variación florística de especies arbóreas
a escala local en un bosque de tierra firme en la Amazonia colombiana. Acta
Amazonica, 40 (1):179-188.

Bell, G. 2001. Ecology: neutral macroecology. Science, 293:2413–2418.
Belyea LR & Lancaster J. 1999.Assembly rules within a contingent ecology. Oikos,
86:402–416.
Bloom AJ, Chapin FS, Mooney HA. 1985. Resource limitation in plants – an economic
analogy. Annual Review of Ecology and Systematics, 16: 363–392.
Blundell AG, Peart DR. 2001. Growth strategies of a shade- tolerant tropical tree: the
interactive effects of canopy gaps and simulated herbivory. Journal of Ecology,
89:608–615.
Brenes-Arguedas T, Ríos M, Rivas-Torres G, Blundo C, Coley P & Kursar T. 2008. The
effect of soil on the growth performance of tropical species with contrasting
distributions. Oikos, 117:1453-1460.
Butler B J & Chazdon RL. 1998. Species richness, spatial variation, and abundance of the
soil seed bank of a secondary tropical rain forest. Biotropica, 30: 214-222.
Campbell BD & Grime JP. 1989. A comparative study of plants responsiveness to the
duration of episodes of mineral nutrient enrichment. New Phytologist, 112:261-267.
Cárdenas D & Salinas N. 2007. Libro Rojo de Plantas de Colombia. Vol. 4. Especies
Maderables Amenazadas de Colombia. Instituto Amazónico de Investigaciones
Científicas SINCHI. Bogotá. 232 p.

Chapin FS III. 1980. The mineral nutrition of wild plants. Annual Review Of Ecology And
Systematics, 11:233–260.
Chapin FS, Vitousek PM & Van Cleve K. 1986. The nature of nutrient limitation in plant
communities. American Naturalist, 127:48–58.
Chapin FS. 1991. Integrated responses of plants to stress. Bioscience, 41: 29 – 36.
Chazdon R & Fetcher N. 1984.Photosynthetic light environments in a lowland tropical rain
forest in Costa Rica. Journal of Ecology, 72: 553-564.
Clark, D. A., D. B. Clark, R. M. Sandoval, and M. V. Castro.1995. Edaphic and human
effects on landscape-scale distributions of tropical rain forest palms. Ecology
76:2581–2594
Clark D, Clark D & Read, J. 1998. Edaphic variation and the mesoscale distribution of tree
species in a neotropical rain forest. Journal of Ecology, 86:101–112.
Clark D.B, Palmer M.W. & Clark, D.A. 1999.Edaphic factors and the landscape-scale
distributions of tropical rain forest trees. Ecology, 80:2662–2675.
Cogollo A. 1986. Estudio Florístico y Ecológico en el Cañón del Río Claro en San Luis
(Antioquia). Actualidades Biológicas ed: Universidad De Antioquia, 45 (56): 35 – 40.
Coley, P.D., Bryant, J. & Chapin, F.S. 1985. Resource availability and plant antiherbivore
defense. Science, 230:895–899.

Coley PD & Barone JA. 1996. Herbivory and Plant Defenses in Tropical Forests. Annual
Review of Ecology and Systematics, 27(1):305–335.
Condit R, Ashton P, Baker P, Bunyavejchewin S, Gunatilleke S, Gunatilleke, N, Hubbell
S, Foster R, Itoh A, LaFrankie N, Lee H, Losos E, Manokaran T, Sukumar R
&Yamakura T. 2000. Spatial patterns in the distribution of tropical tree species.
Science, 288:1414-1418.
Connell, J.H. 1978. Diversity in tropical rain forest and coral reefs. Science, 199:1302-
1309.
Coomes DA & Grubb PJ. 2000. Impacts of root competition in forests and woodlands: A
theoretical framework and review of experiments. Ecological Monographs, 70:171–
207.
Cornare–Municipio de San Luis. 1998. Identificación del potencial ecoturístico del
municipio de San Luis. Proyecto Bosques Húmedos Tropicales, p. 295.
Cornelissen JHC, Thompson K. 1997. Functional leaf attributes predict litter decomposition
rate in herbaceous plants. New Phytologist, 35:109–114.
Cornelissen J, Lavorel S, Garnier E, Díaz S, Buchmann D, Gurvich D, Reich P, Steege H,
Morgan H, Van der Heijden M, .Pausas J, Poorter H. 2003. A handbook of protocols
for standardised and easy measurement of plant functional traits worldwide. Australian
Journal of Botany, 51(4), p.335.

Cornwell W.K. & Ackerly D.D. 2009. Community assembly and shifts in plant trait
distributions across an environmental gradient in coastal California. Ecological
Monographs, 79(1):109-126.
Crick JC & Grime JP. 1987. Morphological plasticity and mineral nutrient capture in two
herbaceous species of contrasted ecology. New Phytologist, 107:403–414.
Davies S.1998. Comparative ecology of 11 sympatric species of Macaranga in Borneo. The
Journal of Ecology, 86(4):662-673.
Denslow JS. 1987. Tropical rainforest gaps and tree species diversity. Annual Review of
Ecology and Systematics, 18: 431–451.
Dufrene M & Legendre P. 1997. Species assemblages and indicator species: the need for a
flexible asymmetrical approach. Ecological Monographs, 67(3): 345-366.
Eissenstat D. 2000. Root structure and function in an ecological context. New Phytologist,
148:353–354Futuyma D. J. & Moreno G. 1988. The evolution of ecological
specialization. Annual Review of Ecology and Systematics 19:207–233.
Evans JR & Poorter H. 2001. Photosynthetic acclimation of plants to growth irradiance :
the relative importance of specific leaf area and nitrogen partitioning in maximizing
carbon gain. Plant, Cell and Environment, 24:755–767.

Gentry A.H. 1988. Changes in Plant Community Diversity and Floristic Composition on
Environmental and Geographical Gradients. Annals of the Missouri Botanical Garden,
75(1)1-34.
Givnish TJ. 1986. Biomechanical constraints on crown geometry in forest herbs. In:
Givnish TJ. ed. On the economy of form and function. Cambridge, UK: Cambridge
University Press, pp 525–583.
Gleason S, Read J, Ares A & Metcalfe D. 2010. Species-soil associations, disturbance, and
nutrient cycling in an Australian tropical rainforest. Oecologia, 162(4):1047-1058.
Gleason SM, Read J & Ares A. 2011. Biomass allocation and phosphorus economics of
rain-forest seedlings: effects of fertilization and radiation on soil specialists and soil
generalists. Journal of Tropical Ecology, 27:147–161.
Grime J.P, Hunt R. 1975. Relative growth-rate: its range and adaptive significance in a
local flora. Journal of Ecology, 63:393-422.
Grime JP. 1977. Evidence for the existence of three primary strategies in plants and its
relevance to ecological and evolutionary theory. The American naturalist,
111(982):1169–1194.
Grime JP, Crick JC & Rincón JE. 1986. The ecological significance of plasticity. Plasticity
in Plants (eds D. H. Jennings & A. J. Trewavas). Cambridge, pp. 5-29.

Hall J.B. & Swaine M.D. 1981.Distribution and Ecology of Vascular Plants in a Tropical
Rain Forest. Dr W. Junk Publishers, The Netherlands.
Harms KE, Condit R & Hubbell SP. 2001. Habitat associations of trees and shrubs in a 50-
ha neotropical forest plot. Journal of Ecology, 89: 947–959.
Hernández B., Vélez H., 1988, Geomorfología de los mármoles de Río Claro
(Departamento de Antioquia). Trabajo Dirigido de Grado, Universidad Nacional,
Facultad de Minas, p. 171.
Hodge A. 2004. The plastic plant: root responses to heterogeneous supplies of nutrients.
New Phytologist, 162:9–24.
Hoffmann WA & Poorter H. 2002. Avoiding bias in calculations of relative growth rate.
Annals of Botany, 90:37-42.
Hoffmann WA, Orthen B & Franco A C. 2004. Constraints to seedling success of savanna
and forest trees across the savanna-forest boundary. Oecologia, 140(2):252–260.
Holdridge L. 1979. Ecología Basada en Zonas de Vida. Libros y Materiales Educativos,
No. 34. San José, Costa Rica: Instituto Interamericano de Ciencias Agrícolas.
Hoyos S & Hernández J. 1983. Estudio florístico de un bosque en el municipio de San
Luis. Actualidades biológicas, 12 (44):267-324.

Huante, P, Rincon E & Chapin III FS. 1995. Responses to Phosphorus of contrasting
successional tree-seedling species from the tropical deciduous forest of Mexico.
Functional Ecology, 9(5):760–766.
Hubbell S.P. & Foster R.B. 1983. Diversity of canopy trees in a neotropical forest and
implications for conservation. Tropical Rain Forest: Ecology and Management,
Blackwell Science, Oxford, UK, p. 25–41.
Hubbell SP.1979. Tree dispersion, abundance, and diversity in a tropical dry forest.Science
(New York, N.Y.), 203(4387), p.1299-1309.
Hubbell, S. 2001. The unified neutral theory of biodiversity and biogeography. Princeton
University Press, Princeton, NJ.
Hunt, R. & Lloyd, P.S. 1987. Growth and partitioning. New Phytologist 106 (Suppl), pp
235–249.
Hunt R, Causton D R, Shipley B & Askew A P. 2002. A modern tool for classical growth
analysis. Annals of Botany, 90:485-488.
John R. et al. 2007. Soil nutrients influence spatial distributions of tropical tree species.
Proceedings of the National Academy of Sciences, 104:864–869.
Kerkhoff AJ, Fagan WF, Elser JJ & Enquist BJ. 2006. Phylogenetic and growth form
variation in the scaling of nitrogen and phosphorus in the seed plants. The American
Naturalist, 168:E103–E122.

Lambers H, Korner Ch. & Renhardt U. 1987. Dry matter partitioning and root length/leaf
area ratios in herbaceous perennial plants with diverse altitudinal distribution.
Oecologia 74:411-418.
Kerkhoff AJ, Fagan WF, Elser JJ & Enquist BJ. 2006. Phylogenetic and growth form
variation in the scaling of nitrogen and phosphorus in the seed plants. The American
Naturalist, 168:E103–E122.
Korning J, Thomsen K, Dalsgaard K & Nornberg P. 1994. Characters of three udults and
their relevance to the composition and structure of virgin rain forest of amazonian
Ecuador. Geoderma, 63:145–164.
Lambers H, Dijkstra H. 1987. A physiological analysis of genetic variation in relative
growth rate: Can growth rate confer ecological advantage?. Disturbance in grasslands:
causes, effects and processes. The Netherlands. p. 237–252.
Lambers H, Poorter H. 1992. Inherent variation in growth rate between higher plants: a
earch for ecological causes and consequences. Advances in Ecological Research,
2:187–261.
Lavorel S, Díaz S, Cornelissen J, Garnier E, Harrison S, Mcintyre S & Pausas J. 2007.
Plant Functional Types : Are We Getting Any Closer to the Holy Grail ? Terrestrial
Ecosystems in a Changing World. Book Series Title: Global Change — The IGBP
Series. Springer Berlin Heidelberg. p. 149-164.

Lee HS, Davies SJ, LaFrankie JV, Tan S, Itoh A, Yamakura T, Ohkubo T & Ashton PS.
2002. Floristic and structural diversity of 52 hectares of mixed dipterocarp forest in
Lambir Hills National Park, Sarawak. Malaysia Journal of Tropical Forest Science,
14:379–400.
Leigh E.G. 1999. Tropical Forest Ecology: a View from Barro Colorado Island. Oxford
University Press, Oxford, UK.
Lusk CH & Matus F. 2000. Juvenile tree growth rates and species sorting on fine-scale soil
fertility gradients in a Chilean temperate rain forest. Journal of Biogeography,
27:1011–1020.
MacGarry D. A methodology of a visual soil - Field assessment tool to support, enhance
and contribute to the LADA program [Internet]. 2006. FAO. Queensland government,
Australia. Fecha de acceso: 07 de julio de 2013. Disponible en: <
ftp://ftp.fao.org/agl/agll/lada/vsfast_methodology.pdf >.
Marenco RA, Vieira G, Gonçalves JFC. 2003. Leaf micronutrient concentrations and
potential photosynthesis in Ochroma pyramidale established in a degraded land. Acta
Amazonica, 33:23–31.
Marschner H. 1995. Mineral Nutrition of Higher Plants, 2nd ed. Academic Press, New
York.

Markesteijn L & Poorter L. 2009. Seedling root morphology and biomass allocation of 62
tropical tree species in relation to drought- and shade-tolerance. Journal of Ecology,
97:311–325.
Matteucci SD, Colma A. 1982. Metodología para el estudio de la vegetación. Secretaria
General de la Organización de los Estados Americanos. Programa Regional de
Desarrollo Científico y Tecnológico, Washington D.C., USA. 168 pp.
Munsell soil color charts. 1994. Munsell Products, New York.
Newman E I. 1966. A method of estimating the total length of root in a sample. Journal of
Applied Ecology, 3:139-145.
Ocampo E. 2005. Caracterización petrográfica de los mármoles de Cementos Río Claro,
sector sur, con miras a estudiar su potencial de uso, especialmente en la industria
cementera y aumentar las reservas de materias primas de la empresa Cementos Río
Claro S.A. Tesis de Pregrado, Universidad Nacional de Colombia.
Palmiotto PA, Davies SJ, Vogt KA, Ashton MS, Vogt DJ& Ashton PS. 2004. Soil-related
habitat specialization in dipterocarp rain forest tree species in Borneo.Journal of
Ecology, 92:609–623.
Palow DT & Oberbauer SF. 2009. Soil type affects seedling shade response at low light for
two Inga species from Costa Rica. Plant soil, 319:25–35.

Peña-Claros M, Poorter L, Alarcón A, Blate G, Choque U, Fredericksen TS, Justiniano M.
2012. Soil Effects on Forest Structure and Diversity in a Moist and a Dry Tropical
Forest. Biotropica, 44(3): 276–283.
Pérez-Harguindeguy N. et al. 2013. New handbook for standardised measurement of plant
functional traits worldwide. Australian Journal of Botany, 61:167–234.
Plotkin J, Potts M Leslie N, Manokaran N, LaFrankie J &Ashton P. 2000. Species-area
curves, spatial aggregation, and habitat specialization in tropical forests. Journal of
Theoretical Biology, 207:81–99.
Poorter H & Pothmann P. 1992. Growth and carbon economy of a fast- growing and a
slow-growing grass species as dependent on ontogeny. New Phytologist, 120:159–
166.
Poorter H & Garnier E. 1999. Ecological significance of inherent variation in relative
growth rate and its components. Handbook of Functional Plant Ecology (eds F.I.
Pugnaire & F. Valladares), pp. 81–120. Marcel Dekker, New York.
Poorter L, Bongers F, Sterck F &Woll H. 2005. Beyond the regeneration phase:
differentiation of height-light trajectories among tropical tree species. Journal of
Ecology, 93: 256–267.

Poorter H, Niklas KJ, Reich PB, Oleksyn JP, Mommer L. 2012. Biomass allocation to
leaves, stems and roots : meta-analyses of interspecific variation and environmental
control. New Phytologist, 193:30–50.
Poorter H, Nagel O. 2000. The role of biomass allocation in the growth response of plants
to different levels of light, CO2, nutrients and water: a quantitative review. Australian
Journal of Plant Physiology, 27:595–607.
Poulsen A D, Tuomisto H, Balslev H (2006) Edaphic and floristic variation within a 1-ha
plot of lowland Amazonian rain forest. Biotropica 38:468–478
Read J, SansonGD, De Garine-Wichatitsky M & Jaffr´e T. 2006. Sclerophylly in two
contrasting tropical environments: low nutrients vs. low rainfall. American Journal of
Botany, 93:1601– 1614.
Reich PB, Tjoelker MG, Walters MB, Vanderklein DW & Bushena C. 1998. Close
association of relative growth rate, leaf and root morphology, seed mass and shade
tolerance in seedlings of nine boreal tree species grown in high and low light.
Functional Ecology, 12:327–338.
Reich PB, Tjoelker MG, Pregitze KS,Wright IJ, Oleksyn J & Machado JL. 2008. Scaling of
respiration to nitrogen in leaves, stems and roots of higher land plants. Ecology
Letters, 11:793–801.
Restrepo C. 2011. El sistema Karstico de La Danta. Dyna,169:239-246.

Rozendaal D. M., Hurtado V. H. & Poorter L. 2006. Plasticity in leaf traits of 38 tropical
tree species in response to light; relationships with light demand and adult stature.
Functional Ecology, 20(2):207-216.
Schemske DW, Mittelbach GG, Cornell HV, Sobel JM & Roy K. 2009. Is there a
latitudinal gradient in the importance of biotic interactions? Annual Review of
Ecology and Systematics, 40:245–269.
Schimper A. F. W. 1898. Plant geography on a physiological basis. G. Fischer, Jena,
Germany.
Shipley B & Keddy PA. 1988. The relationship between relative growth rate and sensitivity
to nutrient stress in twenty-eight species of emergent macrophytes. Journal of
Ecology, 76:1101-1110.
Shipley, B., Keddy, P.A, Moore, D.R.J &Lemky, K. 1989. Regeneration and establishment
strategies of emergent macrophytes. Journal of Ecology, 77:1093–1110.
Shipley, B. & Peters, R.H. 1990a. A test of the Tilman model of plant strategies: relative
growth rate and bio- mass partitioning. American Naturalist 136:139–153.
Shipley B. Lechoweicz MJ, Wright I & Reich PB. 2006. Fundamental trade- offs
generating the worldwide leaf economics spectrum. Ecology, 87:535–541.
Suzuki E & Tsukahara J. 1987. Age structure and regeneration of old growth Cryptomeria
japonica forests on Yakushima Island. Botanical Magazine, 100:223-241.

Svenning JC. 2001. On the role of microenvironmental heterogeneity in the ecology and
diversification of neotropical rain-forest palms (Arecaceae). Botanical Review, 67:1–
53.
Swaine MD. 1996. Rainfall and soil fertility as factors limiting forest species distributions
in Ghana. Journal of Ecology, 84: 419–428.
Taiz L & Zeiger E. 2006. Plant Physiology, Fourth Edition. Sinauer Associates.
Sunderland, MA. p 764.
Tilman D & Pacala S. 1993. The maintenance of species diversity in plant communities.
Species Diversity in Ecological Communities. University of Chicago Press, Chicago.
USA. p. 13-25.
Tilman D. 1982. Resources competition and community structure. Princeton University
Press, Princeton, New Jersey, USA.
Tilman D. 1988. Plant Strategies and the Dynamics and Structure of Plant Communities.
Princeton University Press, Princeton, NJ.
Tuomisto H. & Ruokolainen K. 1994. Distribution of Pteridophyta and Melastomataceae
along an edaphic gradient in an Amazonian rain forest. Journal of Vegetation Science,
5, p. 25-34.

Valencia R, Foster R, Villa G, Condit R, Svenning J, Hernández C, Romoleroux K, Losos
E, Magard E, Balslev H. 2004. Tree species distributions and local habitat variation in
the Amazon : large forest plot in eastern Ecuador. Journal of Ecology, 92:214-229.
Walter H. 1974.The vegetation of the earth. Gustav Fisher Verlag, Jena
Webb CO & Peart DR. 2000. Habitat associations of trees and seedlings in a Bornean rain
forest. Journal of Ecology, 88(3):464–478.
Whittaker RH 1975. Communities and ecosystems. Second edition. MacMillan, New York,
New York, USA.

ANEXOS
Anexo 1. Lista de las especies registradas en los transectos de vegetación con su índice de
especies indicadoras (IV) y su valor de p asociado. Las especies que presentaron índices
significativos se resaltan con color rojo.
Trans Especie
Tipo
de
suelo*
Valor del
indicador
observado
(IV)
IV de grupos
indicadores aleatorios
observados valor
p*
Promedio Desv.
estándar
1 Aegiphila truncata 1 20 20 0,28 1,00
2 Aiouea sp. 0 20 20 0,28 1,00
3 Albizia carbonaria 1 40 26,5 11,91 0,44
4 Allophylus edulis 0 20 20 0,28 1,00
5 Alseis sp. 0 60 32,2 14,71 0,16
6 Anaxagorea cf. brevipes 0 20 20 0,28 1,00
7 Aniba sp. 1 40 23,4 14,92 0,45
8 Aphelandra crenata 1 20 20 0,28 1,00
9 Ardisia sp. 0 40 23,1 14,88 0,44
10 Aspidosperma sp 2. 0 20 20 0,28 1,00
11 Brosimum guianense 0 20 20 0,28 1,00
12 Bunchosia argentea 0 20 20 0,28 1,00
13 Bunchosia
brevisurcularis
1 20 20 0,28 1,00
14 Calyptranthes multiflora 0 100 43,2 15,19 0,01
15 Calyptranthes sp. 0 40 26,2 12,44 0,45
16 Caryodaphnopsis
burgeri
0 80 36,4 15,05 0,05
17 Caryodaphnopsis
cogolloi
1 20 20 0,28 1,00
18 Caryodendron
orinocense
0 60 32,3 13,5 0,17
19 Casearia mariquitensis 1 60 32 12,96 0,16
20 Casearia pitumba 1 30 41,9 13,31 1,00
21 Cecropia sp 1 40 23,4 14,92 0,45
22 Chrysochlamys
dependens
1 20 20 0,28 1,00

23 Cinnamomum triplinerve 0 20 20 0,28 1,00
24 Clarisia racemosa 1 20 20 0,28 1,00
25 Clathrotropis brunnea 1 40 28,1 10,52 0,44
26 Coccoloba densifrons 0 40 23,4 14,92 0,45
27 Coccoloba sp. 0 20 20 0,28 1,00
28 Cojoba rufescens 0 100 42 13,2 0,01
29 Cordia alliodora 1 40 23,2 14,89 0,44
30 Cordia nodosa 1 20 20 0,28 1,00
31 Cynometra bauhiniifolia 0 20 20 0,28 1,00
32 Dendropanax arboreus 1 20 20 0,28 1,00
33 Desmopsis microcarpa 0 60 32,1 12,36 0,16
34 Duguetia colombiana 0 100 42,3 14,31 0,01
35 Dulacia candida 0 20 20 0,28 1,00
36 Ephedranthus
colombianus
0 20 20 0,28 1,00
37 Erythroxylum
plowmanianum
0 60 32,2 13,17 0,17
38 Faramea occidentalis 0 100 42,2 12,96 0,01
39 Ficus matiziana 0 20 20 0,28 1,00
40 Galipea panamensis 0 40 23,6 14,94 0,45
41 Gloeospermum
longifolium
0 20 20 0,28 1,00
42 Guapira costaricana 0 20 20 0,28 1,00
43 Guarea sp. 0 20 20 0,28 1,00
44 Guatteria cestrifolia 1 45 42,9 15,44 0,65
45 Gustavia romeroi S.A 1 20 20 0,28 1,00
46 Gustavia speciosa 0 60 32,2 13,79 0,17
47 Hamelia patens 1 20 20 0,28 1,00
48 Hasseltia floribunda 1 20 20 0,28 1,00
49 Herrania cf. purpurea . 1 20 20 0,28 1,00
50 Hirtella sp 0 40 25,3 13,27 0,45
51 Hura crepitans 1 40 25,2 13,26 0,45
52 Indet. 1 0 20 20 0,28 1,00
53 Inga marginata 1 26,7 32 12,26 1,00
54 Inga samanensis 1 20 20 0,28 1,00
55 Inga sp. 0 20 20 0,28 1,00
56 Jacaranda hesperia 1 60 32,1 13,51 0,16
57 Lonchocarpus cf. monilis 0 40 25 13,23 0,44
58 Luehea seemannii 1 80 37,2 14,79 0,05
59 Margaritaria nobilis 1 13,3 25,3 13,27 1,00
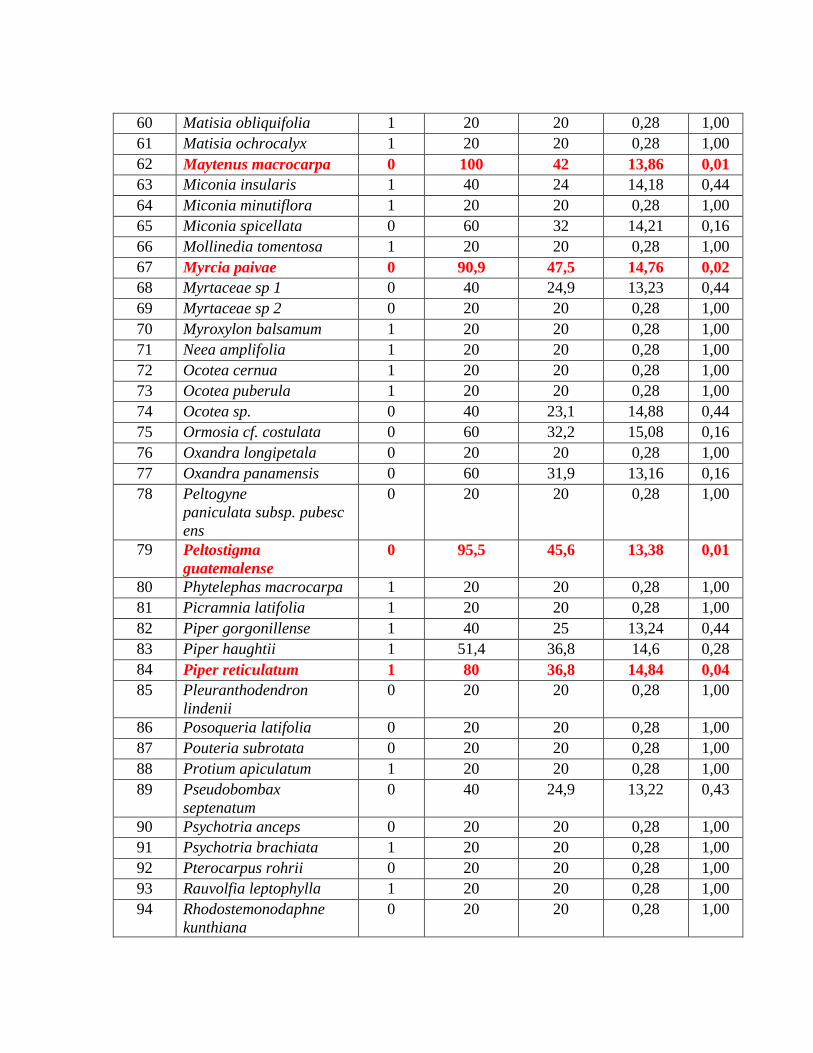
60 Matisia obliquifolia 1 20 20 0,28 1,00
61 Matisia ochrocalyx 1 20 20 0,28 1,00
62 Maytenus macrocarpa 0 100 42 13,86 0,01
63 Miconia insularis 1 40 24 14,18 0,44
64 Miconia minutiflora 1 20 20 0,28 1,00
65 Miconia spicellata 0 60 32 14,21 0,16
66 Mollinedia tomentosa 1 20 20 0,28 1,00
67 Myrcia paivae 0 90,9 47,5 14,76 0,02
68 Myrtaceae sp 1 0 40 24,9 13,23 0,44
69 Myrtaceae sp 2 0 20 20 0,28 1,00
70 Myroxylon balsamum 1 20 20 0,28 1,00
71 Neea amplifolia 1 20 20 0,28 1,00
72 Ocotea cernua 1 20 20 0,28 1,00
73 Ocotea puberula 1 20 20 0,28 1,00
74 Ocotea sp. 0 40 23,1 14,88 0,44
75 Ormosia cf. costulata 0 60 32,2 15,08 0,16
76 Oxandra longipetala 0 20 20 0,28 1,00
77 Oxandra panamensis 0 60 31,9 13,16 0,16
78 Peltogyne
paniculata subsp. pubesc
ens
0 20 20 0,28 1,00
79 Peltostigma
guatemalense
0 95,5 45,6 13,38 0,01
80 Phytelephas macrocarpa 1 20 20 0,28 1,00
81 Picramnia latifolia 1 20 20 0,28 1,00
82 Piper gorgonillense 1 40 25 13,24 0,44
83 Piper haughtii 1 51,4 36,8 14,6 0,28
84 Piper reticulatum 1 80 36,8 14,84 0,04
85 Pleuranthodendron
lindenii
0 20 20 0,28 1,00
86 Posoqueria latifolia 0 20 20 0,28 1,00
87 Pouteria subrotata 0 20 20 0,28 1,00
88 Protium apiculatum 1 20 20 0,28 1,00
89 Pseudobombax
septenatum
0 40 24,9 13,22 0,43
90 Psychotria anceps 0 20 20 0,28 1,00
91 Psychotria brachiata 1 20 20 0,28 1,00
92 Pterocarpus rohrii 0 20 20 0,28 1,00
93 Rauvolfia leptophylla 1 20 20 0,28 1,00
94 Rhodostemonodaphne
kunthiana
0 20 20 0,28 1,00
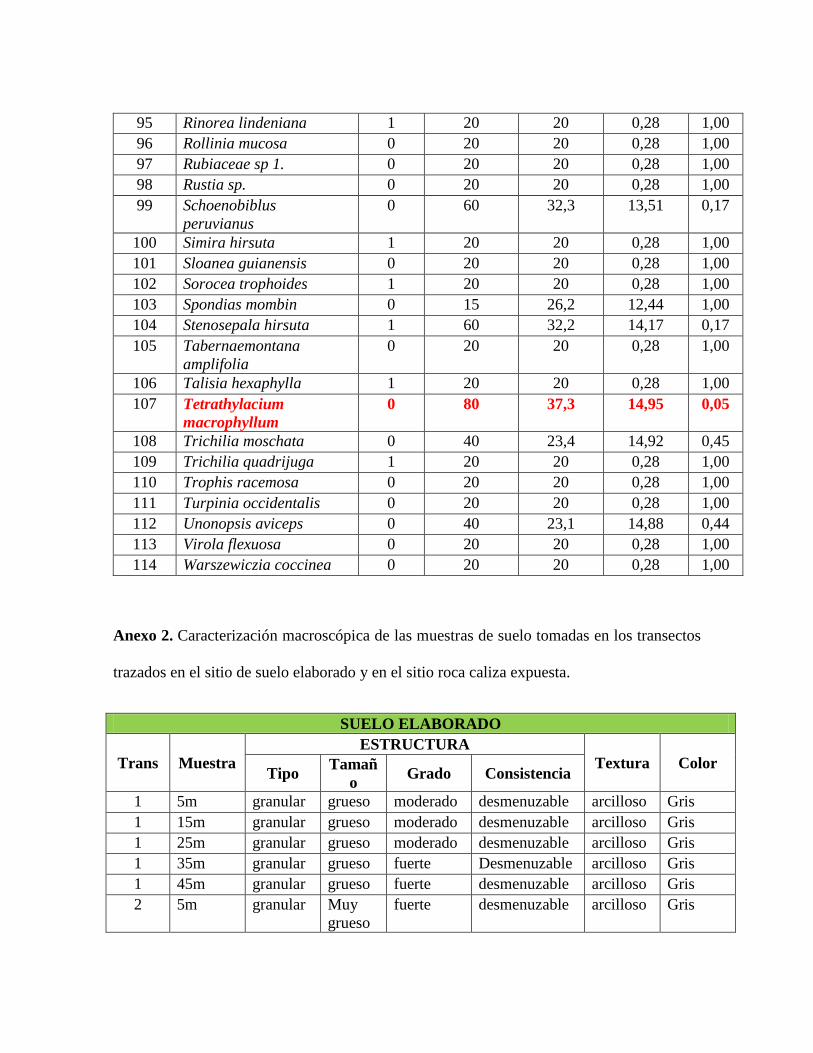
95 Rinorea lindeniana 1 20 20 0,28 1,00
96 Rollinia mucosa 0 20 20 0,28 1,00
97 Rubiaceae sp 1. 0 20 20 0,28 1,00
98 Rustia sp. 0 20 20 0,28 1,00
99 Schoenobiblus
peruvianus
0 60 32,3 13,51 0,17
100 Simira hirsuta 1 20 20 0,28 1,00
101 Sloanea guianensis 0 20 20 0,28 1,00
102 Sorocea trophoides 1 20 20 0,28 1,00
103 Spondias mombin 0 15 26,2 12,44 1,00
104 Stenosepala hirsuta 1 60 32,2 14,17 0,17
105 Tabernaemontana
amplifolia
0 20 20 0,28 1,00
106 Talisia hexaphylla 1 20 20 0,28 1,00
107 Tetrathylacium
macrophyllum
0 80 37,3 14,95 0,05
108 Trichilia moschata 0 40 23,4 14,92 0,45
109 Trichilia quadrijuga 1 20 20 0,28 1,00
110 Trophis racemosa 0 20 20 0,28 1,00
111 Turpinia occidentalis 0 20 20 0,28 1,00
112 Unonopsis aviceps 0 40 23,1 14,88 0,44
113 Virola flexuosa 0 20 20 0,28 1,00
114 Warszewiczia coccinea 0 20 20 0,28 1,00
Anexo 2. Caracterización macroscópica de las muestras de suelo tomadas en los transectos
trazados en el sitio de suelo elaborado y en el sitio roca caliza expuesta.
SUELO ELABORADO
Trans Muestra
ESTRUCTURA
Textura Color Tipo
Tamañ
o Grado Consistencia
1 5m granular grueso moderado desmenuzable arcilloso Gris
1 15m granular grueso moderado desmenuzable arcilloso Gris
1 25m granular grueso moderado desmenuzable arcilloso Gris
1 35m granular grueso fuerte Desmenuzable arcilloso Gris
1 45m granular grueso fuerte desmenuzable arcilloso Gris
2 5m granular Muy
grueso
fuerte desmenuzable arcilloso Gris
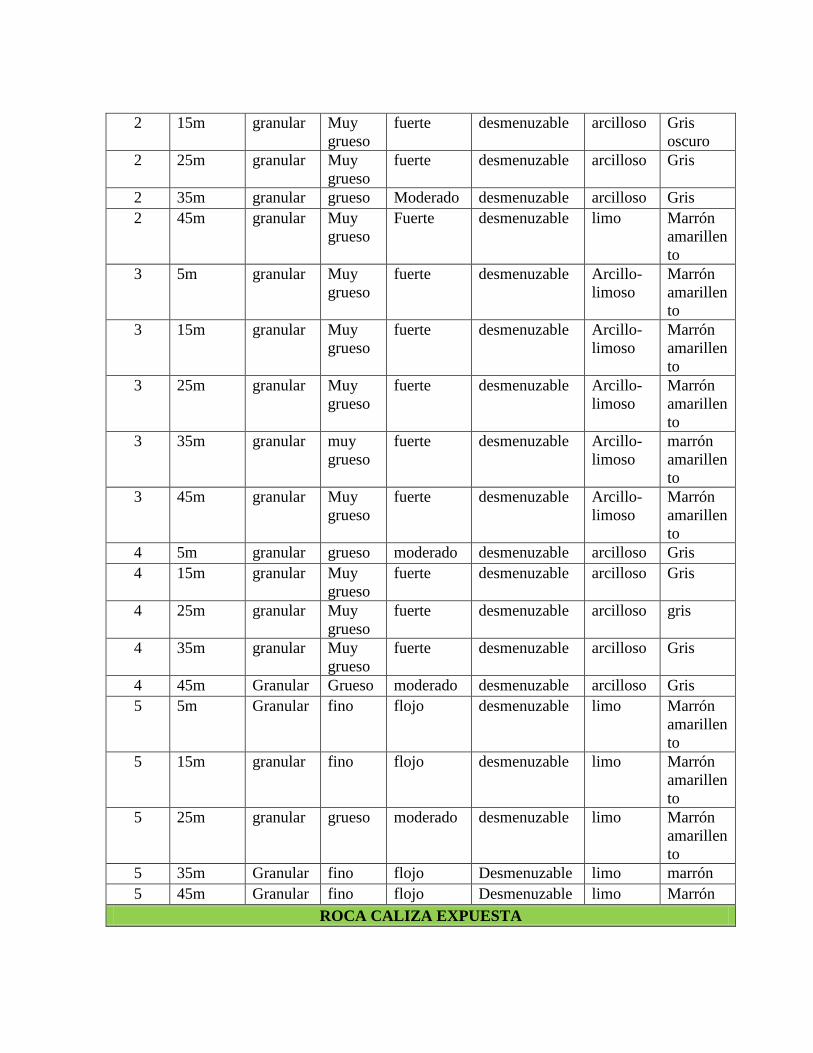
2 15m granular Muy
grueso
fuerte desmenuzable arcilloso Gris
oscuro
2 25m granular Muy
grueso
fuerte desmenuzable arcilloso Gris
2 35m granular grueso Moderado desmenuzable arcilloso Gris
2 45m granular Muy
grueso
Fuerte desmenuzable limo Marrón
amarillen
to
3 5m granular Muy
grueso
fuerte desmenuzable Arcillo-
limoso
Marrón
amarillen
to
3 15m granular Muy
grueso
fuerte desmenuzable Arcillo-
limoso
Marrón
amarillen
to
3 25m granular Muy
grueso
fuerte desmenuzable Arcillo-
limoso
Marrón
amarillen
to
3 35m granular muy
grueso
fuerte desmenuzable Arcillo-
limoso
marrón
amarillen
to
3 45m granular Muy
grueso
fuerte desmenuzable Arcillo-
limoso
Marrón
amarillen
to
4 5m granular grueso moderado desmenuzable arcilloso Gris
4 15m granular Muy
grueso
fuerte desmenuzable arcilloso Gris
4 25m granular Muy
grueso
fuerte desmenuzable arcilloso gris
4 35m granular Muy
grueso
fuerte desmenuzable arcilloso Gris
4 45m Granular Grueso moderado desmenuzable arcilloso Gris
5 5m Granular fino flojo desmenuzable limo Marrón
amarillen
to
5 15m granular fino flojo desmenuzable limo Marrón
amarillen
to
5 25m granular grueso moderado desmenuzable limo Marrón
amarillen
to
5 35m Granular fino flojo Desmenuzable limo marrón
5 45m Granular fino flojo Desmenuzable limo Marrón
ROCA CALIZA EXPUESTA
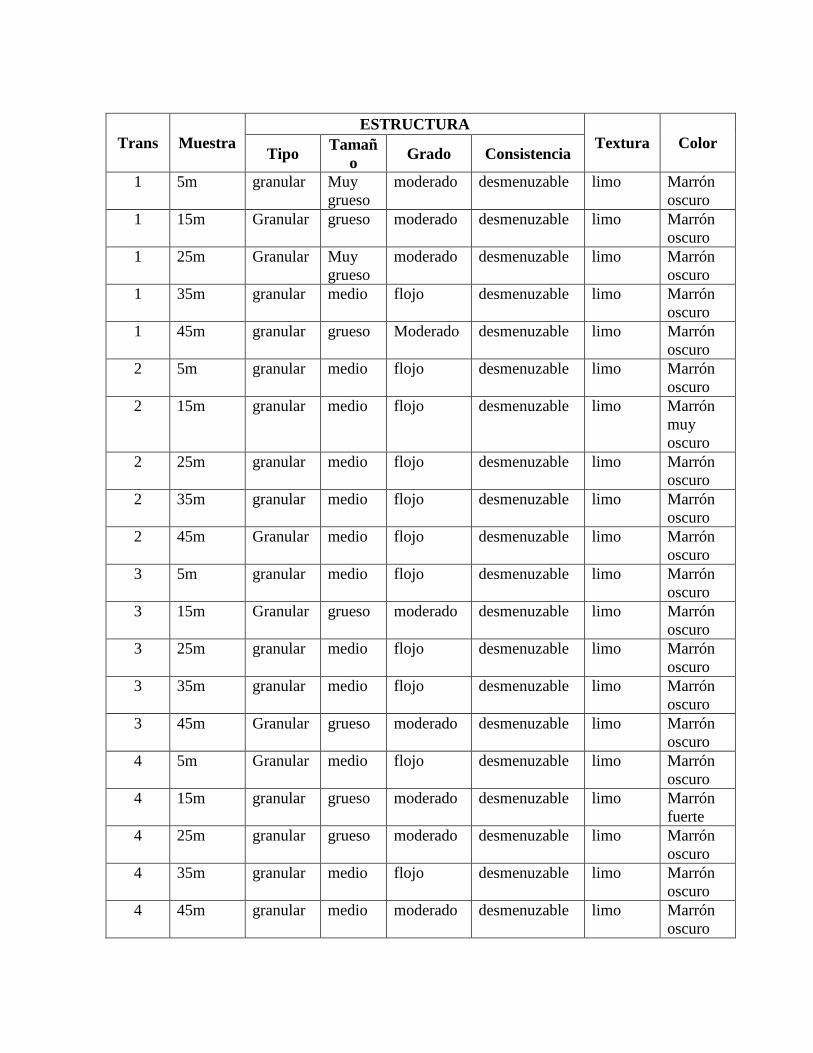
Trans Muestra
ESTRUCTURA
Textura Color Tipo
Tamañ
o Grado Consistencia
1 5m granular Muy
grueso
moderado desmenuzable limo Marrón
oscuro
1 15m Granular grueso moderado desmenuzable limo Marrón
oscuro
1 25m Granular Muy
grueso
moderado desmenuzable limo Marrón
oscuro
1 35m granular medio flojo desmenuzable limo Marrón
oscuro
1 45m granular grueso Moderado desmenuzable limo Marrón
oscuro
2 5m granular medio flojo desmenuzable limo Marrón
oscuro
2 15m granular medio flojo desmenuzable limo Marrón
muy
oscuro
2 25m granular medio flojo desmenuzable limo Marrón
oscuro
2 35m granular medio flojo desmenuzable limo Marrón
oscuro
2 45m Granular medio flojo desmenuzable limo Marrón
oscuro
3 5m granular medio flojo desmenuzable limo Marrón
oscuro
3 15m Granular grueso moderado desmenuzable limo Marrón
oscuro
3 25m granular medio flojo desmenuzable limo Marrón
oscuro
3 35m granular medio flojo desmenuzable limo Marrón
oscuro
3 45m Granular grueso moderado desmenuzable limo Marrón
oscuro
4 5m Granular medio flojo desmenuzable limo Marrón
oscuro
4 15m granular grueso moderado desmenuzable limo Marrón
fuerte
4 25m granular grueso moderado desmenuzable limo Marrón
oscuro
4 35m granular medio flojo desmenuzable limo Marrón
oscuro
4 45m granular medio moderado desmenuzable limo Marrón
oscuro

5 5m Granular Grueso moderado desmenuzable limo Marrón
oscuro
5 15m Granular medio flojo desmenuzable limo marrón
muy
oscuro
5 25m granular medio Flojo desmenuzable limo Marrón
oscuro
5 35m granular medio flojo desmenuzable limo Marrón
5 45m Granular medio flojo desmenuzable limo Marrón
oscuro
Anexo 3. Parámetros básicos de fertilidad para las muestras de suelo separadas según el
tipo de suelo al que corresponden.
Trans. Tipo de suelo pH M.O
(%)
P
(ppm)
N
(%)
Al Ca Mg K CICE
meq/100 g de suelo
1 Elaborado 4,32 5,31 1,19 0,27 0,8 8,01 0,35 0,16 9,32
2 Elaborado 5,67 6,15 1,61 0,31 - 11,60 0,41 0,21 12,22
3 Elaborado 5,60 6,57 2,80 0,33 - 9,58 0,43 0,19 10,20
4 Elaborado 5,72 5,31 1,26 0,27 - 9,29 0,49 0,32 10,10
5 Elaborado 5,76 5,90 1,19 0,29 - 10,89 0,68 0,27 11,84
6 Roca caliza
expuesta 6,66 4,05 8,89 0,20 -
12,97 0,70 0,19 13,86
7 Roca caliza
expuesta 5,60 3,12 2,31 0,16 -
4,58 0,80 0,26 5,64
8 Roca caliza
expuesta 5,22 2,53 2,24 0,13 0,1
3,31 0,25 0,09 3,75
9 Roca caliza
expuesta 5,00 2,36 2,10 0,12 0,1
2,56 0,25 0,09 3,00
10 Roca caliza
expuesta 5,61 1,94 11,41 0,10 -
1,47 0,43 0,18 2,08

Anexo 4. Clase textural y elementos menores de las muestras tomadas por transectos y por
tipo de suelo.
Trans
Tipo de suelo
Textura* Clase
Textural
**
Cu
Fe
Mn
Zn
A L Ar
% ppm
1 Elaborado 70 22 8 FA 0,64 15,36 10,4
4 2,16
2 Elaborado 80 16 4 AFR 0,32 11,00 8,04 2,88
3 Elaborado 78 12 10 FA 0,32 14,68 7,40 2,52
4 Elaborado 82 14 4 AFR 0,32 22,20 6,16 1,76
5 Elaborado 78 20 2 AFR 0,28 15,96 6,08 1,72
6 Roca caliza
expuesta 82 16 2 AFR 2,00 23,24 4,44 4,04
7 Roca caliza
expuesta 74 20 6 FA 2,40 47,08 5,84 5,28
8 Roca caliza
expuesta 62 24 14 FA 2,28 97,28 4,52 1,72
9 Roca caliza
expuesta 72 16 12 FA 1,20 61,48 9,44 3,12
10 Roca caliza
expuesta 72 18 10 FA 2,28 95,96
26,4
8 6,64
* A: arcilla; L: limo; Ar: arena.
** FA: franco arenoso; AFR: arcilloso franco.
Anexo 5. Parámetros descriptivos de los valores de rasgos funcionales medidos en
plántulas de C. burgeri, C. brunnea, D. colombiana y P. paniculata subsp. pubescens.
Caryodaphnopsis burgeri
COSECHA 1
Sitio N Promedio Desv. estándar Error estándar
promedio
Altura (cm) 1 16 15,88 1,218 0,305
2 9 16,41 1,018 0,339
SLA (m2/kg) 1 16 11,341 1,993 0,498
2 10 12,115 ,790 0,249
LMA (1/SLA) 1 16 0,091 0,019 0,005
2 10 0,082 0,005 0,002

Leaf size (mm2) 1 16 3635,443 807,203 201,800
2 10 4092,740 729,747 230,766
LDMC (mg/g) 1 16 290,992 26,359 6,589
2 8 313,902 9,169 3,241
Leaf toughness
(N/cm2)
1 16 51,391 4,424 1,106
2 10 56,625 2,516 0,795
SRL (m/g) 1 16 0,980 0,480 0,120
2 10 0,655 0,285 0,090
Biomasa total (g) 1 16 0,831 0,180 0,045
2 10 0,852 0,162 0,0514
LMF (g/g) 1 16 0,393 0,037 0,009
2 10 0,399 0,028 0,009
SMF (g/g) 1 16 0,354 0,042 0,010
2 10 0,346 0,037 0,011
RMF (g/g) 1 16 0,242 0,046 0,011
2 10 0,233 0,039 0,012
DM (kg/kg) 1 16 0,370 0,036 0,009
2 10 0,407 0,032 0,010
R:S 1 16 0,324 0,080 0,020
2 10 0,686 0,157 0,049
COSECHA 2
Sitio N Promedio Desviación
estándar
Error estándar
promedio
Altura (cm) 1 16 16,56 1,504 0,376
2 10 16,30 1,636 0,517
SLA (m2/kg) 1 14 13,083 3,623 0,968
2 8 12,453 2,411 0,852
LMA (1/SLA) 1 16 0,074 0,026 0,006
2 10 0,074 0,025 0,008
Leaf size (mm2) 1 16 4572,992 881,836 220,459
2 10 3158,298 736,556 232,919
LDMC (mg/g) 1 16 258,384 17,807 4,451
2 10 278,712 22,715 7,183
Leaf toughness
(N/cm2)
1 16 45,190 2,296 0,574
2 10 44,300 2,879 0,910
SRL (m/g) 1 16 1,989 0,6365 0,159
2 10 1,257 1,07 0,338
Biomasa total (g) 1 16 1,0 0E-7 0E-7
2 10 0,8 0,421 0,133
LMF (g/g) 1 16 0,361 0,058 0,014
2 10 0,326 0,068 0,021
SMF (g/g) 1 15 0,294 0,044 0,011
2 10 0,370 0,081 0,025
RMF (g/g) 1 16 0,327 0,064 0,016
2 10 0,292 0,036 0,011
DM (kg/kg) 1 16 0,317 0,023 0,005
2 10 0,344 0,049 0,015
R:S 1 16 0,503 0,174 0,043
2 10 0,841 0,289 0,091
COSECHA 3
Sitio N Promedio Desviación
estándar
Error estándar
promedio
Altura (cm) 1 16 17,69 1,537 ,384
2 15 17,13 2,031 ,524
SLA (m2/kg) 1 16 13,008 3,756 ,939
2 15 16,492 10,863 2,804
LMA (1/SLA) 1 15 ,079 ,022 ,006
2 15 ,086 ,045 ,011
Leaf size (mm2) 1 16 6343,104 2208,470 552,117
2 9 3049,845 255,222 85,074
LDMC (mg/g) 1 16 292,973 19,832 4,958
2 10 297,114 11,752 3,716
Leaf toughness
(N/cm2)
1 15 54,548 1,814 ,468
2 14 53,732 2,400 ,641
SRL (m/g) 1 15 2,986 ,682 ,176
2 15 1,689 1,247 ,322
Biomasa total (g) 1 16 1,116 ,196 ,049
2 15 ,932 ,407 ,105
LMF (g/g) 1 16 ,432 ,046 ,011
2 15 ,268 ,115 ,029
SMF (g/g) 1 16 ,277 ,038 ,009
2 15 ,355 ,147 ,037
RMF (g/g) 1 16 ,280 ,036 ,009
2 15 ,369 ,059 ,015
DM (kg/kg) 1 16 ,325 ,017 ,004
2 15 ,348 ,022 ,005
R:S 1 16 ,392 ,074 ,018
2 15 1,274 ,660 ,170
Duguetia colombiana
COSECHA 1
Sitio N Promedio Desviación
estándar
Error estándar
promedio
Altura (cm) 1 16 16,56 1,504 ,376
2 10 16,30 1,636 ,517
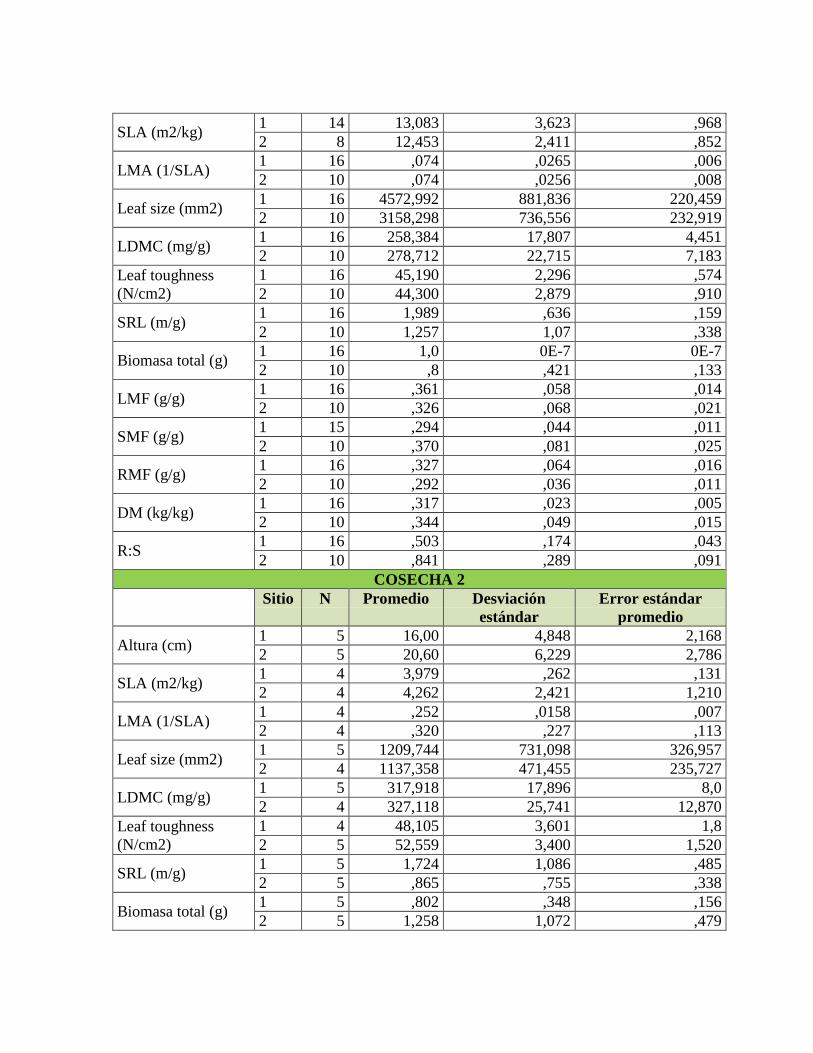
SLA (m2/kg) 1 14 13,083 3,623 ,968
2 8 12,453 2,411 ,852
LMA (1/SLA) 1 16 ,074 ,0265 ,006
2 10 ,074 ,0256 ,008
Leaf size (mm2) 1 16 4572,992 881,836 220,459
2 10 3158,298 736,556 232,919
LDMC (mg/g) 1 16 258,384 17,807 4,451
2 10 278,712 22,715 7,183
Leaf toughness
(N/cm2)
1 16 45,190 2,296 ,574
2 10 44,300 2,879 ,910
SRL (m/g) 1 16 1,989 ,636 ,159
2 10 1,257 1,07 ,338
Biomasa total (g) 1 16 1,0 0E-7 0E-7
2 10 ,8 ,421 ,133
LMF (g/g) 1 16 ,361 ,058 ,014
2 10 ,326 ,068 ,021
SMF (g/g) 1 15 ,294 ,044 ,011
2 10 ,370 ,081 ,025
RMF (g/g) 1 16 ,327 ,064 ,016
2 10 ,292 ,036 ,011
DM (kg/kg) 1 16 ,317 ,023 ,005
2 10 ,344 ,049 ,015
R:S 1 16 ,503 ,174 ,043
2 10 ,841 ,289 ,091
COSECHA 2
Sitio N Promedio Desviación
estándar
Error estándar
promedio
Altura (cm) 1 5 16,00 4,848 2,168
2 5 20,60 6,229 2,786
SLA (m2/kg) 1 4 3,979 ,262 ,131
2 4 4,262 2,421 1,210
LMA (1/SLA) 1 4 ,252 ,0158 ,007
2 4 ,320 ,227 ,113
Leaf size (mm2) 1 5 1209,744 731,098 326,957
2 4 1137,358 471,455 235,727
LDMC (mg/g) 1 5 317,918 17,896 8,0
2 4 327,118 25,741 12,870
Leaf toughness
(N/cm2)
1 4 48,105 3,601 1,8
2 5 52,559 3,400 1,520
SRL (m/g) 1 5 1,724 1,086 ,485
2 5 ,865 ,755 ,338
Biomasa total (g) 1 5 ,802 ,348 ,156
2 5 1,258 1,072 ,479
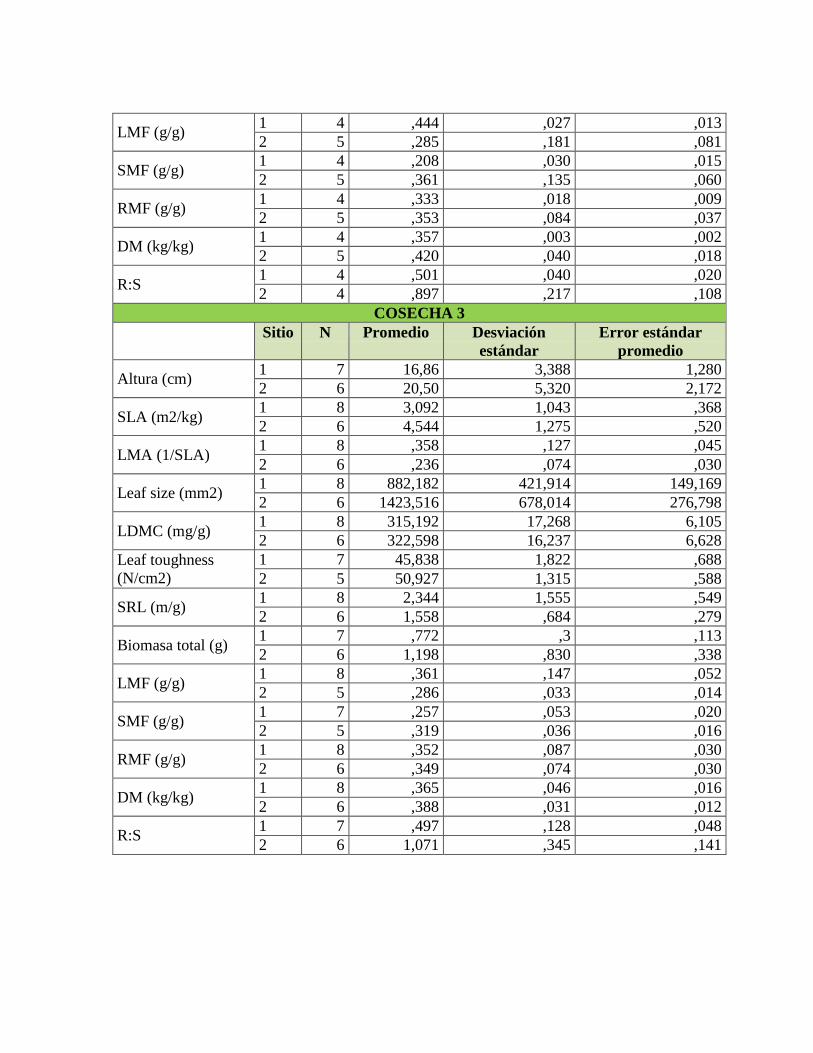
LMF (g/g) 1 4 ,444 ,027 ,013
2 5 ,285 ,181 ,081
SMF (g/g) 1 4 ,208 ,030 ,015
2 5 ,361 ,135 ,060
RMF (g/g) 1 4 ,333 ,018 ,009
2 5 ,353 ,084 ,037
DM (kg/kg) 1 4 ,357 ,003 ,002
2 5 ,420 ,040 ,018
R:S 1 4 ,501 ,040 ,020
2 4 ,897 ,217 ,108
COSECHA 3
Sitio N Promedio Desviación
estándar
Error estándar
promedio
Altura (cm) 1 7 16,86 3,388 1,280
2 6 20,50 5,320 2,172
SLA (m2/kg) 1 8 3,092 1,043 ,368
2 6 4,544 1,275 ,520
LMA (1/SLA) 1 8 ,358 ,127 ,045
2 6 ,236 ,074 ,030
Leaf size (mm2) 1 8 882,182 421,914 149,169
2 6 1423,516 678,014 276,798
LDMC (mg/g) 1 8 315,192 17,268 6,105
2 6 322,598 16,237 6,628
Leaf toughness
(N/cm2)
1 7 45,838 1,822 ,688
2 5 50,927 1,315 ,588
SRL (m/g) 1 8 2,344 1,555 ,549
2 6 1,558 ,684 ,279
Biomasa total (g) 1 7 ,772 ,3 ,113
2 6 1,198 ,830 ,338
LMF (g/g) 1 8 ,361 ,147 ,052
2 5 ,286 ,033 ,014
SMF (g/g) 1 7 ,257 ,053 ,020
2 5 ,319 ,036 ,016
RMF (g/g) 1 8 ,352 ,087 ,030
2 6 ,349 ,074 ,030
DM (kg/kg) 1 8 ,365 ,046 ,016
2 6 ,388 ,031 ,012
R:S 1 7 ,497 ,128 ,048
2 6 1,071 ,345 ,141
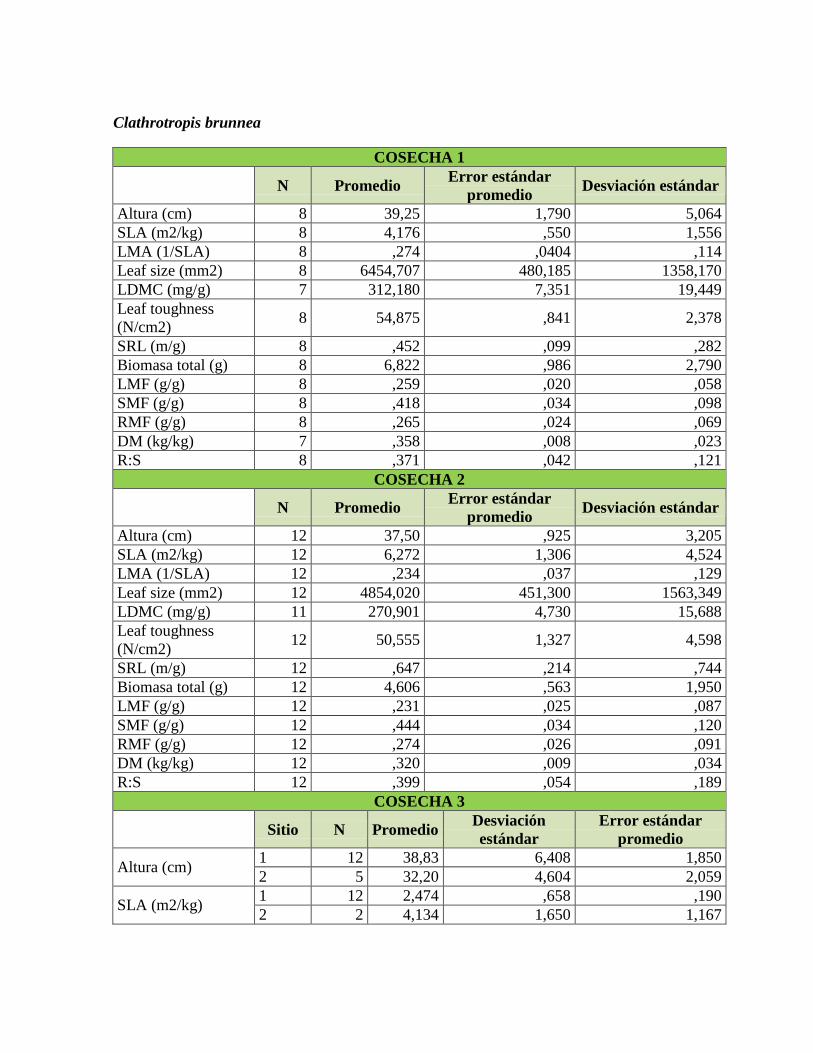
Clathrotropis brunnea
COSECHA 1
N Promedio
Error estándar
promedio Desviación estándar
Altura (cm) 8 39,25 1,790 5,064
SLA (m2/kg) 8 4,176 ,550 1,556
LMA (1/SLA) 8 ,274 ,0404 ,114
Leaf size (mm2) 8 6454,707 480,185 1358,170
LDMC (mg/g) 7 312,180 7,351 19,449
Leaf toughness
(N/cm2) 8 54,875 ,841 2,378
SRL (m/g) 8 ,452 ,099 ,282
Biomasa total (g) 8 6,822 ,986 2,790
LMF (g/g) 8 ,259 ,020 ,058
SMF (g/g) 8 ,418 ,034 ,098
RMF (g/g) 8 ,265 ,024 ,069
DM (kg/kg) 7 ,358 ,008 ,023
R:S 8 ,371 ,042 ,121
COSECHA 2
N Promedio
Error estándar
promedio Desviación estándar
Altura (cm) 12 37,50 ,925 3,205
SLA (m2/kg) 12 6,272 1,306 4,524
LMA (1/SLA) 12 ,234 ,037 ,129
Leaf size (mm2) 12 4854,020 451,300 1563,349
LDMC (mg/g) 11 270,901 4,730 15,688
Leaf toughness
(N/cm2) 12 50,555 1,327 4,598
SRL (m/g) 12 ,647 ,214 ,744
Biomasa total (g) 12 4,606 ,563 1,950
LMF (g/g) 12 ,231 ,025 ,087
SMF (g/g) 12 ,444 ,034 ,120
RMF (g/g) 12 ,274 ,026 ,091
DM (kg/kg) 12 ,320 ,009 ,034
R:S 12 ,399 ,054 ,189
COSECHA 3
Sitio N Promedio Desviación
estándar
Error estándar
promedio
Altura (cm) 1 12 38,83 6,408 1,850
2 5 32,20 4,604 2,059
SLA (m2/kg) 1 12 2,474 ,658 ,190
2 2 4,134 1,650 1,167
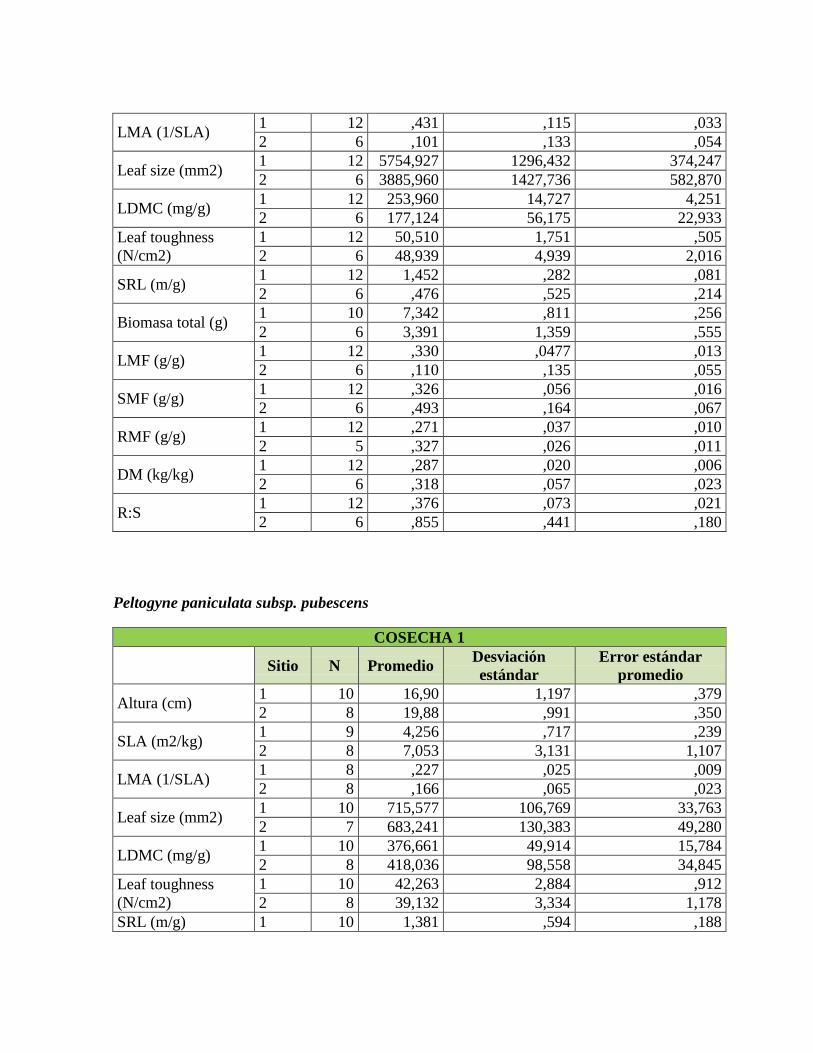
LMA (1/SLA) 1 12 ,431 ,115 ,033
2 6 ,101 ,133 ,054
Leaf size (mm2) 1 12 5754,927 1296,432 374,247
2 6 3885,960 1427,736 582,870
LDMC (mg/g) 1 12 253,960 14,727 4,251
2 6 177,124 56,175 22,933
Leaf toughness
(N/cm2)
1 12 50,510 1,751 ,505
2 6 48,939 4,939 2,016
SRL (m/g) 1 12 1,452 ,282 ,081
2 6 ,476 ,525 ,214
Biomasa total (g) 1 10 7,342 ,811 ,256
2 6 3,391 1,359 ,555
LMF (g/g) 1 12 ,330 ,0477 ,013
2 6 ,110 ,135 ,055
SMF (g/g) 1 12 ,326 ,056 ,016
2 6 ,493 ,164 ,067
RMF (g/g) 1 12 ,271 ,037 ,010
2 5 ,327 ,026 ,011
DM (kg/kg) 1 12 ,287 ,020 ,006
2 6 ,318 ,057 ,023
R:S 1 12 ,376 ,073 ,021
2 6 ,855 ,441 ,180
Peltogyne paniculata subsp. pubescens
COSECHA 1
Sitio N Promedio Desviación
estándar
Error estándar
promedio
Altura (cm) 1 10 16,90 1,197 ,379
2 8 19,88 ,991 ,350
SLA (m2/kg) 1 9 4,256 ,717 ,239
2 8 7,053 3,131 1,107
LMA (1/SLA) 1 8 ,227 ,025 ,009
2 8 ,166 ,065 ,023
Leaf size (mm2) 1 10 715,577 106,769 33,763
2 7 683,241 130,383 49,280
LDMC (mg/g) 1 10 376,661 49,914 15,784
2 8 418,036 98,558 34,845
Leaf toughness
(N/cm2)
1 10 42,263 2,884 ,912
2 8 39,132 3,334 1,178
SRL (m/g) 1 10 1,381 ,594 ,188
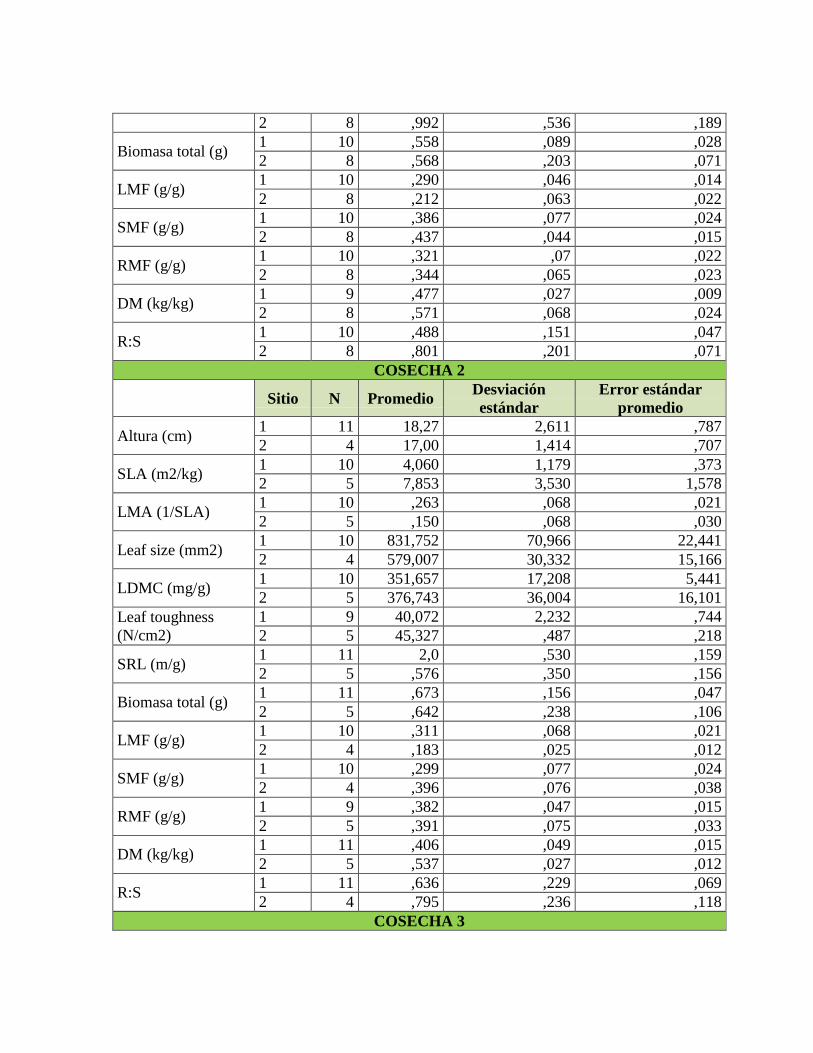
2 8 ,992 ,536 ,189
Biomasa total (g) 1 10 ,558 ,089 ,028
2 8 ,568 ,203 ,071
LMF (g/g) 1 10 ,290 ,046 ,014
2 8 ,212 ,063 ,022
SMF (g/g) 1 10 ,386 ,077 ,024
2 8 ,437 ,044 ,015
RMF (g/g) 1 10 ,321 ,07 ,022
2 8 ,344 ,065 ,023
DM (kg/kg) 1 9 ,477 ,027 ,009
2 8 ,571 ,068 ,024
R:S 1 10 ,488 ,151 ,047
2 8 ,801 ,201 ,071
COSECHA 2
Sitio N Promedio Desviación
estándar
Error estándar
promedio
Altura (cm) 1 11 18,27 2,611 ,787
2 4 17,00 1,414 ,707
SLA (m2/kg) 1 10 4,060 1,179 ,373
2 5 7,853 3,530 1,578
LMA (1/SLA) 1 10 ,263 ,068 ,021
2 5 ,150 ,068 ,030
Leaf size (mm2) 1 10 831,752 70,966 22,441
2 4 579,007 30,332 15,166
LDMC (mg/g) 1 10 351,657 17,208 5,441
2 5 376,743 36,004 16,101
Leaf toughness
(N/cm2)
1 9 40,072 2,232 ,744
2 5 45,327 ,487 ,218
SRL (m/g) 1 11 2,0 ,530 ,159
2 5 ,576 ,350 ,156
Biomasa total (g) 1 11 ,673 ,156 ,047
2 5 ,642 ,238 ,106
LMF (g/g) 1 10 ,311 ,068 ,021
2 4 ,183 ,025 ,012
SMF (g/g) 1 10 ,299 ,077 ,024
2 4 ,396 ,076 ,038
RMF (g/g) 1 9 ,382 ,047 ,015
2 5 ,391 ,075 ,033
DM (kg/kg) 1 11 ,406 ,049 ,015
2 5 ,537 ,027 ,012
R:S 1 11 ,636 ,229 ,069
2 4 ,795 ,236 ,118
COSECHA 3
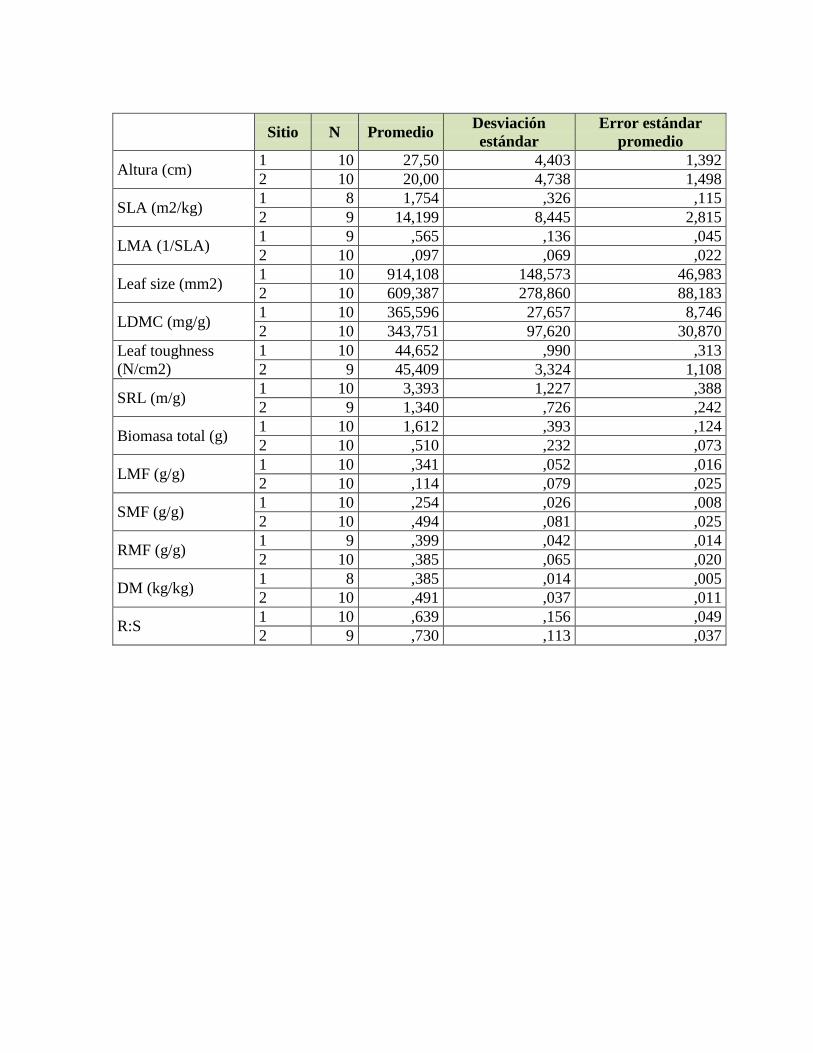
Sitio N Promedio Desviación
estándar
Error estándar
promedio
Altura (cm) 1 10 27,50 4,403 1,392
2 10 20,00 4,738 1,498
SLA (m2/kg) 1 8 1,754 ,326 ,115
2 9 14,199 8,445 2,815
LMA (1/SLA) 1 9 ,565 ,136 ,045
2 10 ,097 ,069 ,022
Leaf size (mm2) 1 10 914,108 148,573 46,983
2 10 609,387 278,860 88,183
LDMC (mg/g) 1 10 365,596 27,657 8,746
2 10 343,751 97,620 30,870
Leaf toughness
(N/cm2)
1 10 44,652 ,990 ,313
2 9 45,409 3,324 1,108
SRL (m/g) 1 10 3,393 1,227 ,388
2 9 1,340 ,726 ,242
Biomasa total (g) 1 10 1,612 ,393 ,124
2 10 ,510 ,232 ,073
LMF (g/g) 1 10 ,341 ,052 ,016
2 10 ,114 ,079 ,025
SMF (g/g) 1 10 ,254 ,026 ,008
2 10 ,494 ,081 ,025
RMF (g/g) 1 9 ,399 ,042 ,014
2 10 ,385 ,065 ,020
DM (kg/kg) 1 8 ,385 ,014 ,005
2 10 ,491 ,037 ,011
R:S 1 10 ,639 ,156 ,049
2 9 ,730 ,113 ,037





























