Embed Size (px)
Citation preview

Retinol-deficient rats can convert apharmacological dose of astaxanthin to retinol:antioxidant potential of astaxanthin, lutein, and b-carotene
R.K. Sangeetha and V. Baskaran
Abstract: Retinol (ROH) and provitamin-A carotenoids are recommended to treat ROH deficiency. Xanthophyll carote-noids, being potent antioxidants, can modulate health disorders. We hypothesize that nonprovitamin-A carotenoids may yieldROH and suppress lipid peroxidation under ROH deficiency. This study aimed to (i) study the possible bioconversion of astax-anthin and lutein to ROH similar to b-carotene and (ii) determine the antioxidant potential of these carotenoids with referenceto Na+/K+-ATPase, antioxidant molecules, and lipid peroxidation (Lpx) induced by ROH deficiency in rats. ROH deficiencywas induced in rats (n = 5 per group) by feeding a diet devoid of ROH. Retinol-deficient (RD) rats were gavaged with astaxan-thin, lutein, b-carotene, or peanut oil alone (RD group) for 7 days. Results show that the RD group had lowered plasma ROHlevels (0.3 mmol/L), whereas ROH rose in astaxanthin and b-carotene groups (4.9 and 5.7 mmol/L, respectively), which wassupported by enhanced (69% and 70%) intestinal b-carotene 15,15’-monooxygenase activity. Astaxanthin, lutein, and b-carotene lowered Lpx by 45%, 41%, and 40% (plasma), respectively, and 59%, 64%, and 60% (liver), respectively, comparedwith the RD group. Lowered Na+/K+-ATPase and enhanced superoxide dismutase, catalase, and glutathione-S-transferase activ-ities support the lowered Lpx. To conclude, this report confirms that astaxanthin is converted into b-carotene and ROH inROH-deficient rats, and the antioxidant potential of carotenoids was in the order astaxanthin > lutein > b-carotene.
Key words: antioxidant enzymes, astaxanthin, b-carotene, lipid peroxidation, lutein, retinol, retinol deficiency.
Resume : Le retinol (ROH) et les carotenoıdes provitamine-A sont recommandes pour traiter la deficience en ROH. Lescarotenoıdes xanthophylles sont de puissants antioxydants qui peuvent moduler les affections. Nous emettons l’hypotheseque les carotenoıdes non provitamine-A pourraient fournir du ROH et supprimer la peroxydation lipidique lors d’une defi-cience en ROH. La presente etude a eu pour but 1) d’examiner la possible bioconversion de l’astaxanthine et de la luteineen ROH, au meme titre que le b-carotene, et 2) de determiner le potentiel antioxydant de ces carotenoıdes par rapport a laNa+/K+-ATPase, aux molecules antioxydantes et a la peroxydation lipidique (Lpx) induite par une deficience en ROH. Ona induit la deficience en ROH chez des rats (n = 5/groupe) par une diete sans ROH. On a gave les rats deficients en retinol(DR) soit avec de l’astaxanthine, de la luteine, du b-carotene ou de l’huile d’arachide seulement (groupe DR) pendant 7jours. Les resultats montrent que le groupe DR a eu un taux plus faible de ROH plasmatique (0,3 mmol/L), alors que cetaux a augmente (4,9 et 5,7 mmol/L) chez les groupes astaxanthine et b-carotene, ce qui a ete appuye par une augmentationde l’activite de la b-carotene 15,15’-monooxygenase intestinale (60, 70 %). L’astaxanthine, la luteine et le b-carotene ontdiminue la Lpx de 45, 41 et 40 % (plasma) et de 59, 64 et 60 % (foie) respectivement par rapport a ce qui a ete observechez le groupe DR. Une diminution de l’activite de la Na+/K+-ATPase et une augmentation des activites de la superoxydedismutase, de la catalase et de la glutathion-S-transferase accompagnent la diminution de Lpx. Cette etude confirme quel’astaxanthine est convertie en b-carotene et en ROH chez les rats deficients en ROH, le potentiel antioxydant des carote-noıdes se presentant dans l’ordre suivant : astaxanthine > luteine > b-carotene.
Mots-cles : enzymes antioxydantes, astaxanthine, b-carotene, peroxydation lipidique, luteine, retinol, deficience en retinol.
[Traduit par la Redaction]
Introduction
Globally, at least 254 million preschool children sufferfrom clinical and subclinical vitamin A (retinol) deficiency
(WHO 2000). It is reported that 25% of the 15 million blindpeople in the world are from India and one fifth of thesecases of blindness are caused by retinol (ROH) deficiency(Rajyalakshmi et al. 2001). Epidemiological studies have es-
Received 11 March 2010. Accepted 7 May 2010. Published on the NRC Research Press Web site at cjpp.nrc.ca on 24 September 2010.
Abbreviations: GST, glutathione-S-transferase; Lpx, lipid peroxides; MUFA, monounsaturated fatty acids; Pi, inorganic phosphate;PUFA, polyunsaturated fatty acids; RD, retinol deficient; ROH, retinol; SOD, superoxide dismutase; TMP, 1,1,3,3-tetramethoxypropane.
R.K. Sangeetha and V. Baskaran.1 Department of Biochemistry and Nutrition, Central Food Technological Research Institute, CSIR,Mysore, Karnataka 570 020, India.
1Corresponding author (e-mail: [email protected]).
977
Can. J. Physiol. Pharmacol. 88: 977–985 (2010) doi:10.1139/Y10-074 Published by NRC Research Press

tablished a correlation between vitamin A deficiency and in-creased risk of morbidity and mortality in children (West2000). It has also been suggested that ROH deficiency mayprovoke a general alteration in the physical characteristics ofcell membranes. ROH deficiency has a pro-oxidative effectand increases oxidative stress in rats (Kaul and Krishnakan-tha 1997; Sangeetha et al. 2009). Cellular and subcellularmembranes are susceptible to lipid oxidation owing to highconcentration of polyunsaturated fatty acids and close prox-imity to oxygen, transition metals, and peroxidases (Anzulo-vich et al. 2000). The provitamin-A carotenoids b-carotene,a-carotene, b-cryptoxanthin, and g-carotene are recom-mended to treat ROH deficiency (Olson 1994).
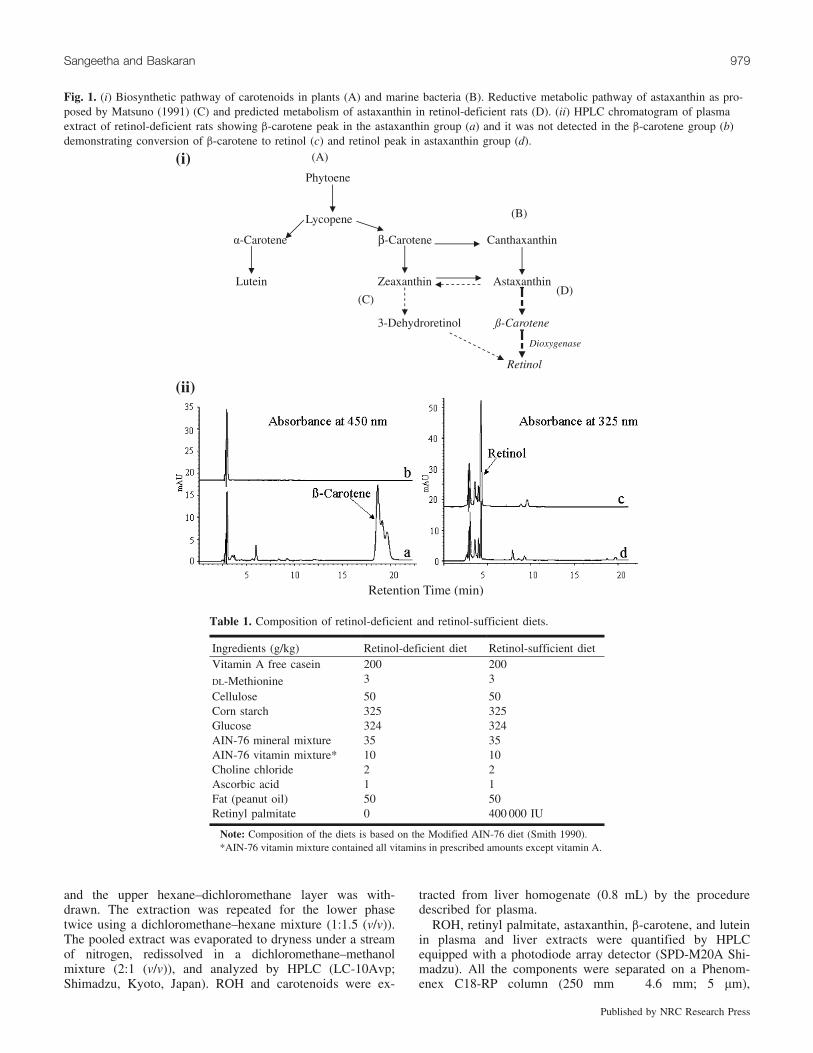
Earlier, Gross and Budowski (1966), Goswami and Barua(1986), and Moren et al. (2002) studied the possible biocon-version of nonprovitamin-A carotenoids, astaxanthin, can-thaxanthin, lutein, and zeaxanthin, in lower vertebrates.Matsuno (1991) and Grangaud et al. (1961) have reportedthe conversion of astaxanthin to retinol in a murine model.In plants, carotenoids have a common biosynthetic pathwayfrom the precursor phytoene and thereafter undergo elonga-tion and cyclization giving rise to a variety of secondarymetabolites (Fig. 1). Evidence indicates that carotenoidsmay protect against certain types of cancer (Kotake-Nara etal. 2001). Exploring the potential of nonprovitamin-A caro-tenoids in comparison with b-carotene towards amelioratingROH deficiency induced cellular alterations may advancetheir health benefits. However, studies of the role of xantho-phylls in modulating vitamin A deficiency disorders at thecellular level are scanty.
In view of the above, it is hypothesized that under retinol-deficient (RD) conditions, the nonprovitamin-A carotenoidsastaxanthin and lutein may convert to ROH and suppresslipid peroxidation. Hence, the aim of this study was to (i)examine the possible bioconversion of astaxanthin and luteinto ROH and the activity of b-carotene cleavage enzymemonooxygenase and (ii) compare the antioxidant potentialof astaxanthin, b-carotene, and lutein in suppressing lipidperoxidation caused by ROH deficiency and activity of anti-oxidant molecules in rats. This study may provide new in-sights into the possible role of nonprovitamin-A carotenoidsin amelioration of ROH deficiency induced biochemicalchanges.
Methods and materials
ChemicalsAll-trans retinol (95%), retinal (98%), retinyl palmitate,
DL-a-tocopherol, all-trans b-carotene (97%), astaxanthin, lu-tein, fatty acid standards, thiobarbituric acid, ouabain,1,1,3,3-tetramethoxypropane (TMP), cytochrome c, xanthineoxidase, and boron trifluoride–methanol solution were pur-chased from Sigma–Aldrich (St. Louis, Mo.). Xanthine andHPLC-grade acetonitrile, hexane, methanol, and dichlorome-thane were purchased from Sisco Research Lab (Mumbai,India). Hydrogen peroxide solution (50%) was procuredfrom Merck (Mumbai, India). Vitamin A free casein, vita-mins, minerals, cellulose, and methionine were purchasedfrom Hi-Media (Mumbai, India). Choline chloride was pur-chased from Loba Chem. Lab (Mumbai, India). Cornstarchand peanut oil were obtained from the local supermarket.
AnimalsThe animal experiment was conducted after clearance
from the Institutional Animal Ethics Committee (IAECNo. 126/08). Weanling male albino rats (OUTB-Wistar,IND-cft (2c)) weighing 35 ± 2 g were housed in individualstainless steel cages in the institute animal house facility atroom temperature (28 ± 2 8C). A 12 h light/dark cycle wasmaintained, and the rats had free access to food and waterad libitum.
Induction of ROH deficiency and intubation withastaxanthin, lutein, and b-carotene
Rats were divided into 5 groups (n = 5 per group). Groups1–4 were fed an RD diet for 8 weeks (Sangeetha et al. 2009)to induce ROH deficiency, while group 5 received a dietcontaining ROH (control). After induction of ROH defi-ciency, groups 1–4 were gavaged (0.5 mL) with an equalpharmacological concentration (0.25 mg�rat–1�day–1) of ei-ther astaxanthin, lutein, or b-carotene dispersed in peanutoil or peanut oil alone (RD group) for 7 days. Group 5 con-tinued on a diet with ROH. The composition of the RD diet(Smith 1990) and control diet is given in Table 1. ROH de-ficiency was confirmed by estimating the plasma ROH level(0.3 mmol/L) (Olson 1982). Feed intake and body mass gainof animals were recorded during the experimental run. Dur-ing the experimental run, rats received diet (RD diet) andwater ad libitum. At the termination of the experiment, ratsin each group were killed using ether anesthesia and bloodwas drawn from the heart into heparinized tubes. The liverwas sampled and washed with ice-cold saline.
Preparation of plasma, liver homogenate, andmicrosomes
All the samples were processed under a dim yellow lighton ice (<5 8C) to minimize isomerization and oxidation ofcarotenoids. The plasma was separated from the blood bycentrifugation (Remi India Ltd., Mumbai) at 1000g for15 min at 4 8C. Liver (1 g) was homogenized (Potter Elveh-jem homogenizer) at 4 8C in buffered saline for the ROHassay or in 120 mmol/L KCl and 30 mmol/L phosphate buf-fer (pH 7.2) for the lipid peroxidation assay. The suspensionwas centrifuged at 10 000g for 10 min at 4 8C to remove nu-clei and cell debris. The supernatant was used for the assayof the antioxidant enzymes (Gonzalez-Flecha et al. 1991).Liver microsomes (hereafter referred to as microsomes)were prepared according to Gutman and Katzper-Shamir(1971) from a 10% homogenate in 0.25 mol/L sucrose, 0.03mol/L histidine, and 0.001 mol/L EDTA (pH 7.4). Protein inplasma, liver homogenate, and microsomes was measured bythe method of Lowry et al. (1951).
HPLC analysis of retinol, retinyl palmitate, andcarotenoids
Astaxanthin, b-carotene, lutein, ROH, and retinyl palmi-tate were extracted from the plasma and liver homogenatesas per Raju and Baskaran (2009) and Lakshminarayana etal. (2008). Briefly, plasma (0.8 mL) was diluted with 3 mLof a dichloromethane–methanol mixture (1:2 (v/v)) contain-ing 2 mmol/L a-tocopherol and mixed for 1 min using avortex mixer, followed by the addition of 1.5 mL hexane.The mixture was centrifuged at 5000g for 3 min at 4 8C
978 Can. J. Physiol. Pharmacol. Vol. 88, 2010
Published by NRC Research Press
and the upper hexane–dichloromethane layer was with-drawn. The extraction was repeated for the lower phasetwice using a dichloromethane–hexane mixture (1:1.5 (v/v)).The pooled extract was evaporated to dryness under a streamof nitrogen, redissolved in a dichloromethane–methanolmixture (2:1 (v/v)), and analyzed by HPLC (LC-10Avp;Shimadzu, Kyoto, Japan). ROH and carotenoids were ex-
tracted from liver homogenate (0.8 mL) by the proceduredescribed for plasma.
ROH, retinyl palmitate, astaxanthin, b-carotene, and luteinin plasma and liver extracts were quantified by HPLCequipped with a photodiode array detector (SPD-M20A Shi-madzu). All the components were separated on a Phenom-enex C18-RP column (250 mm � 4.6 mm; 5 mm),
Fig. 1. (i) Biosynthetic pathway of carotenoids in plants (A) and marine bacteria (B). Reductive metabolic pathway of astaxanthin as pro-posed by Matsuno (1991) (C) and predicted metabolism of astaxanthin in retinol-deficient rats (D). (ii) HPLC chromatogram of plasmaextract of retinol-deficient rats showing b-carotene peak in the astaxanthin group (a) and it was not detected in the b-carotene group (b)demonstrating conversion of b-carotene to retinol (c) and retinol peak in astaxanthin group (d).
Table 1. Composition of retinol-deficient and retinol-sufficient diets.
Ingredients (g/kg) Retinol-deficient diet Retinol-sufficient dietVitamin A free casein 200 200
DL-Methionine 3 3
Cellulose 50 50Corn starch 325 325Glucose 324 324AIN-76 mineral mixture 35 35AIN-76 vitamin mixture* 10 10Choline chloride 2 2Ascorbic acid 1 1Fat (peanut oil) 50 50Retinyl palmitate 0 400 000 IU
Note: Composition of the diets is based on the Modified AIN-76 diet (Smith 1990).*AIN-76 vitamin mixture contained all vitamins in prescribed amounts except vitamin A.
Sangeetha and Baskaran 979
Published by NRC Research Press

isocratically eluting (1 mL/min) with an acetonitrile–dichloromethane–methanol mixture of 60:20:20 (by vol-ume) containing 0.1% ammonium acetate as the mobilephase. ROH and retinyl palmitate were monitored at325 nm, b-carotene and lutein were monitored at 450 nm,while astaxanthin was monitored at 478 nm using Shi-madzu Class-VP v. 6.14SP1 software. The peak identitieswere confirmed by their characteristic UV-vis spectra com-pared with authentic standards and quantified by comparingtheir peak areas.
Assay for b-carotene 15,15’-monooxygenase enzymeb-Carotene 15,15’-monooxygenase (E.C. 14.99.36) (previ-
ously known as b-carotene 15,15’-dioxygenase) activity wasmeasured as per Nagao et al. (1996). In brief, the upper por-tion of the small intestine of control and experimental ani-mals was rinsed with saline, and the mucosal layer(jejunum) was scraped off (15 cm) using a cover glass andhomogenized with 5 volumes (5 mL) of 50 mmol/LHEPES–KOH buffer (pH 7.4) containing 0.54 mol/L KCl,1 mmol/L EDTA, and 0.1 mmol/L dithiothreitrol (DTT).The homogenate was centrifuged at 10 000g for 30 min, thesupernatant was applied to a sephadex G-25 column (1.5 cm� 5.5 cm) equilibrated with 10 mmol/L HEPES–KOH buf-fer (pH 7.4) containing 0.1 mmol/L EDTA, 0.05 mol/L KCl,and 0.1 mmol/L DTT, and the elute was used as the enzymesource. The reaction mixture (0.4 mL) contained tricine–KOH buffer (pH 8.0), 15 mmol/L b-carotene, 0.5 mmol/LDTT, 0.1 mmol/L a-tocopherol, 0.15% Tween-40, and en-zyme (~1.5 mg protein) after preincubation at 37 8C for5 min. The reaction was initiated by adding 1.2 nmol(80 mL) of b-carotene solubilized in a-tocopherol and aque-ous Tween-40 and 0.32 mL of reaction mixture and incu-bated at 37 8C for 60 min in the presence of atmosphericoxygen under dim yellow light. The reaction was terminatedby addition of 0.1 mL 37% formaldehyde and further incu-bated at 37 8C for 10 min followed by addition of 0.5 mLacetonitrile and cooled on ice for 5 min. The insoluble mat-ter in the reaction mixture was removed by centrifugation at10 000g at 4 8C for 10 min. The supernatant (20 mL) wassubjected to HPLC analysis to determine the amount of reti-nal formed.
Lipid peroxidation assayThe level of lipid peroxides (Lpx) in plasma, liver homo-
genate, and microsomes was estimated using TBA as de-scribed previously (Ohkawa et al. 1979). Briefly, 20%acetic acid (1.5 mL), 8% sodium dodecyl sulphate(0.2 mL), and 0.8% thiobarbituric acid (1.5 mL) were added,in that order, to the sample, incubated for 1 h in a boilingwater bath, cooled at room temperature followed by the ad-dition of n-butanol (5 mL), and centrifuged at 3000g for15 min. The upper n-butanol phase containing thiobarbituricacid-reactive substances was read spectrofluorometrically(Hitachi, Japan) with an excitation at 515 nm and emissionat 553 nm. TMP was used as a standard to estimate the thi-obarbituric acid-reactive substances.
Assay for antioxidant enzymesActivity of catalase (CAT; E.C. 1.11.1.6) in plasma and
liver homogenates was determined by measuring the de-
crease in absorption at 240 nm using a spectrophotometer(Shimadzu-Japan, 1601) in a reaction mixture containingphosphate buffer (0.1 mmol/L, pH 7.0) and hydrogen perox-ide (8.8 mmol/L) according to Aebi (1984). One CAT unitis defined as the amount of enzyme required to decompose1 mmol/L of H2O2/min. Superoxide dismutase activity(SOD; E.C. 1.15.1.1) in plasma, liver homogenate, and mi-crosomes was measured by the inhibition of cytochrome creduction mediated via superoxide ions generated by thexanthine–xanthine oxidase system (Flohe and Otting 1984)and measured using a spectrophotometer at 550 nm. OneSOD unit is defined as the amount required to inhibit cyto-chrome c reduction by 50%. Glutathione-S-transferase(GST; E.C. 2.5.1.18) activity in liver homogenates and mi-crosomes was determined following the formation of conju-gate of reduced glutathione (GSH) and 1-chloro-dinitro-benzene at 340 nm in a reaction mixture containing GSH(20 mmol/L), 1-chloro-dinitro-benzene (20 mmol/L), andphosphate buffer (0.1 mmol/L, pH 6.5) (Gluthenberg et al.1985). GSH levels in plasma, liver, and microsomes weremeasured by monitoring the rate of 5-thio-2-nitrobenzoicacid formation at 412 nm (Owens and Belcher 1965).
Fatty acid analysisFatty acids methyl esters were prepared from plasma and
microsomes using boron trifluoride in methanol (Morrisonand Smith 1964) and analyzed by gas chromatography (Shi-madzu 14B, Shimadzu, Kyoto, Japan) fitted with a flameionization detector and a fused silica capillary column(25 m � 0.25 mm; Konik Tech, Barcelona, Spain). The in-jector, column, and detector temperatures were 220, 230,and 240 8C, respectively, with nitrogen as the carrier gas at1 mL/min. Individual fatty acids were identified by compar-ing their retention time with those of standards and quanti-fied using a Clarity Lite 420 integrator.
Na+/K+-ATPase assayNa+/K+-ATPase activity in microsomes was estimated by
the method of Kaplay (1978). Briefly, buffer compositionused for the assay was MgCl2 (3 mmol/L), KCl (14 mmol/L), NaCl (140 mmol/L), EDTA (0.2 mmol/L), and Tris–HCl (20 mmol/L, pH 7.4). Samples were run in 2 batches,one with the inhibitor ouabain (2 mmol/L) and one without.The sample blank contained no assay standard and micro-somes were also run simultaneously. The reaction wasstopped by the addition of trichloroacetic acid (10%). Liber-ated inorganic phosphate (Pi) was determined in aliquots(0.7 mL) of incubated mixtures by the addition of ascorbicacid – ammonium molybdate solution (0.3 mL) prepared ac-cording to the method of Ames (1966). Then, the reactionmixture was mixed well and incubated at 45 8C for 20 min.Extinction at 820 nm was measured by a UV–visible spec-trophotometer (Shimadzu 1601, Kyoto, Japan). Specific ac-tivity was expressed as mmol Pi�h–1�(mg protein)–1.
Statistical analysisThe experimental data obtained for different parameters
were subjected to analysis of variance (ANOVA). In casesof a significant difference, mean separation was accom-plished by Tukey’s test using Origin software (v. 5.1). The
980 Can. J. Physiol. Pharmacol. Vol. 88, 2010
Published by NRC Research Press
percent differences among groups were calculated and con-sidered significant at the level of p < 0.05.
Results
Induction of retinol deficiencyThe ROH level in the control group was 0.733 mmol/L,
whereas its level in the RD group was 0.3 mmol/L, indicat-ing ROH deficiency (Olson 1982). ROH deficiency resultedin lowered body mass (by 25%) compared with the controlwhile liver mass was unaffected. However, the ROH levelin the liver was depleted (p < 0.05) to 2 nmol/g as a resultof feeding a RD diet when compared with the control(65.18 mmol/g). Rats in the RD group did not exhibit anyabnormal morphological signs during the experimental run.
Plasma and liver levels of retinol, astaxanthin, b-carotene, and lutein
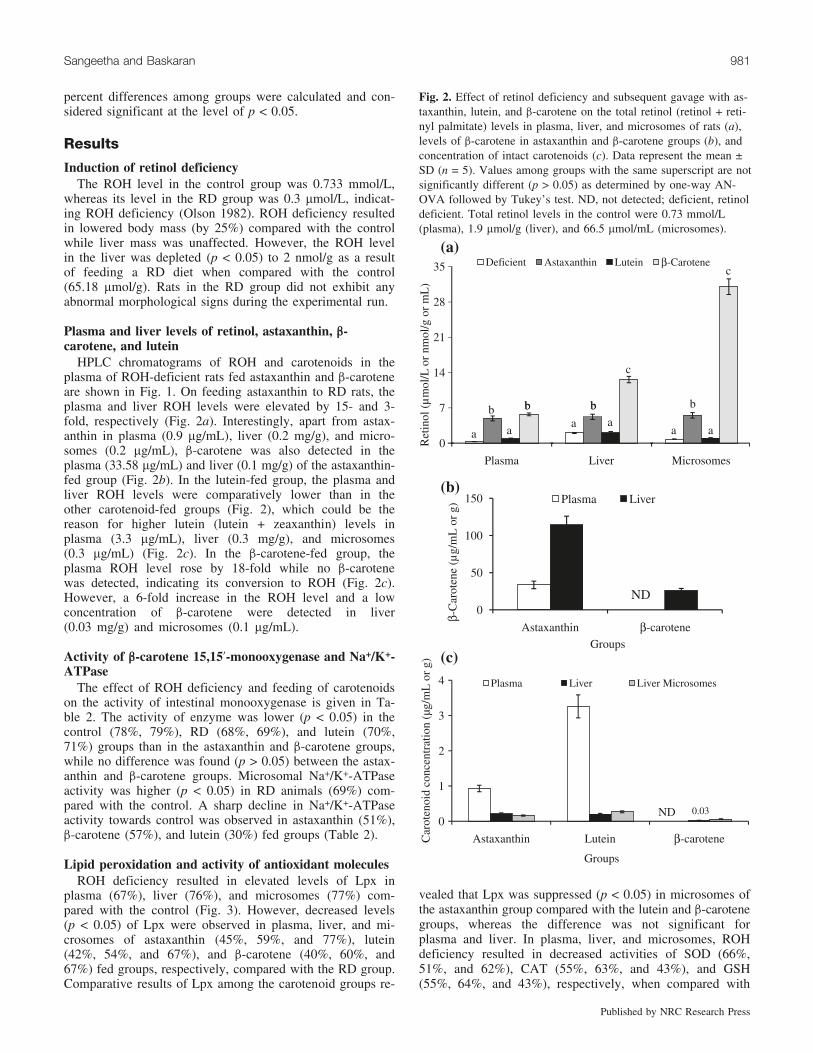
HPLC chromatograms of ROH and carotenoids in theplasma of ROH-deficient rats fed astaxanthin and b-caroteneare shown in Fig. 1. On feeding astaxanthin to RD rats, theplasma and liver ROH levels were elevated by 15- and 3-fold, respectively (Fig. 2a). Interestingly, apart from astax-anthin in plasma (0.9 mg/mL), liver (0.2 mg/g), and micro-somes (0.2 mg/mL), b-carotene was also detected in theplasma (33.58 mg/mL) and liver (0.1 mg/g) of the astaxanthin-fed group (Fig. 2b). In the lutein-fed group, the plasma andliver ROH levels were comparatively lower than in theother carotenoid-fed groups (Fig. 2), which could be thereason for higher lutein (lutein + zeaxanthin) levels inplasma (3.3 mg/mL), liver (0.3 mg/g), and microsomes(0.3 mg/mL) (Fig. 2c). In the b-carotene-fed group, theplasma ROH level rose by 18-fold while no b-carotenewas detected, indicating its conversion to ROH (Fig. 2c).However, a 6-fold increase in the ROH level and a lowconcentration of b-carotene were detected in liver(0.03 mg/g) and microsomes (0.1 mg/mL).
Activity of b-carotene 15,15’-monooxygenase and Na+/K+-ATPase
The effect of ROH deficiency and feeding of carotenoidson the activity of intestinal monooxygenase is given in Ta-ble 2. The activity of enzyme was lower (p < 0.05) in thecontrol (78%, 79%), RD (68%, 69%), and lutein (70%,71%) groups than in the astaxanthin and b-carotene groups,while no difference was found (p > 0.05) between the astax-anthin and b-carotene groups. Microsomal Na+/K+-ATPaseactivity was higher (p < 0.05) in RD animals (69%) com-pared with the control. A sharp decline in Na+/K+-ATPaseactivity towards control was observed in astaxanthin (51%),b-carotene (57%), and lutein (30%) fed groups (Table 2).
Lipid peroxidation and activity of antioxidant moleculesROH deficiency resulted in elevated levels of Lpx in
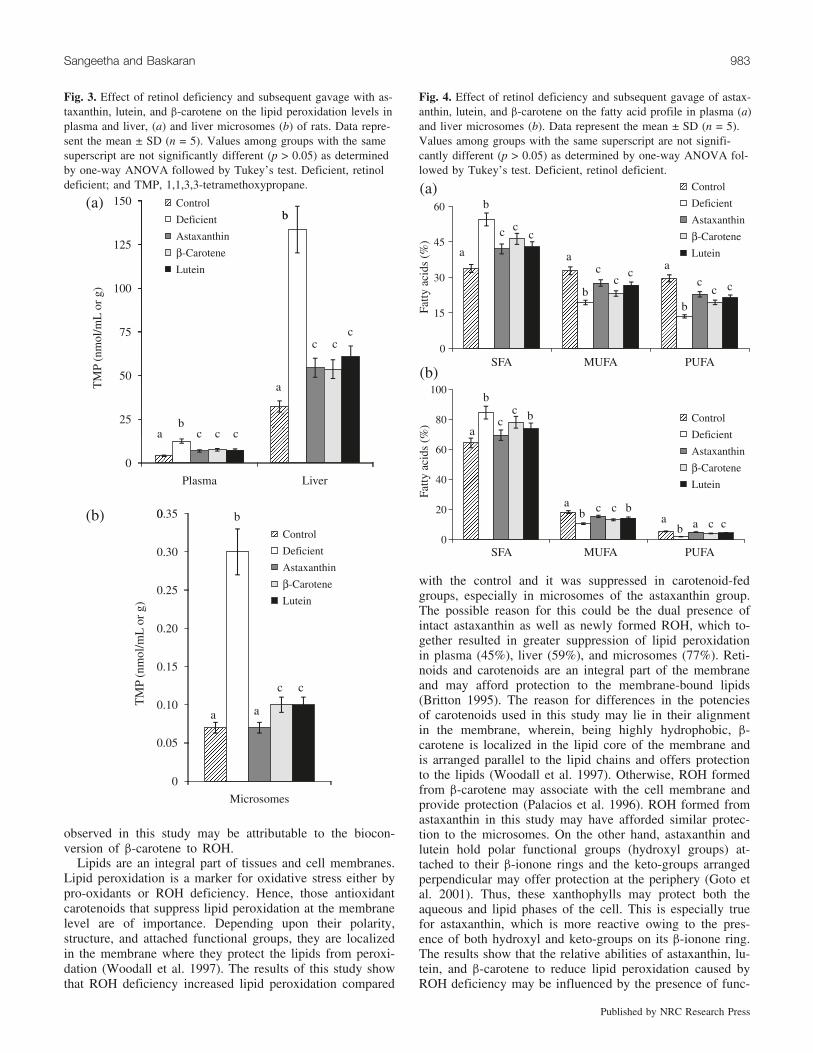
plasma (67%), liver (76%), and microsomes (77%) com-pared with the control (Fig. 3). However, decreased levels(p < 0.05) of Lpx were observed in plasma, liver, and mi-crosomes of astaxanthin (45%, 59%, and 77%), lutein(42%, 54%, and 67%), and b-carotene (40%, 60%, and67%) fed groups, respectively, compared with the RD group.Comparative results of Lpx among the carotenoid groups re-
vealed that Lpx was suppressed (p < 0.05) in microsomes ofthe astaxanthin group compared with the lutein and b-carotenegroups, whereas the difference was not significant forplasma and liver. In plasma, liver, and microsomes, ROHdeficiency resulted in decreased activities of SOD (66%,51%, and 62%), CAT (55%, 63%, and 43%), and GSH(55%, 64%, and 43%), respectively, when compared with
Fig. 2. Effect of retinol deficiency and subsequent gavage with as-taxanthin, lutein, and b-carotene on the total retinol (retinol + reti-nyl palmitate) levels in plasma, liver, and microsomes of rats (a),levels of b-carotene in astaxanthin and b-carotene groups (b), andconcentration of intact carotenoids (c). Data represent the mean ±SD (n = 5). Values among groups with the same superscript are notsignificantly different (p > 0.05) as determined by one-way AN-OVA followed by Tukey’s test. ND, not detected; deficient, retinoldeficient. Total retinol levels in the control were 0.73 mmol/L(plasma), 1.9 mmol/g (liver), and 66.5 mmol/mL (microsomes).
Sangeetha and Baskaran 981
Published by NRC Research Press
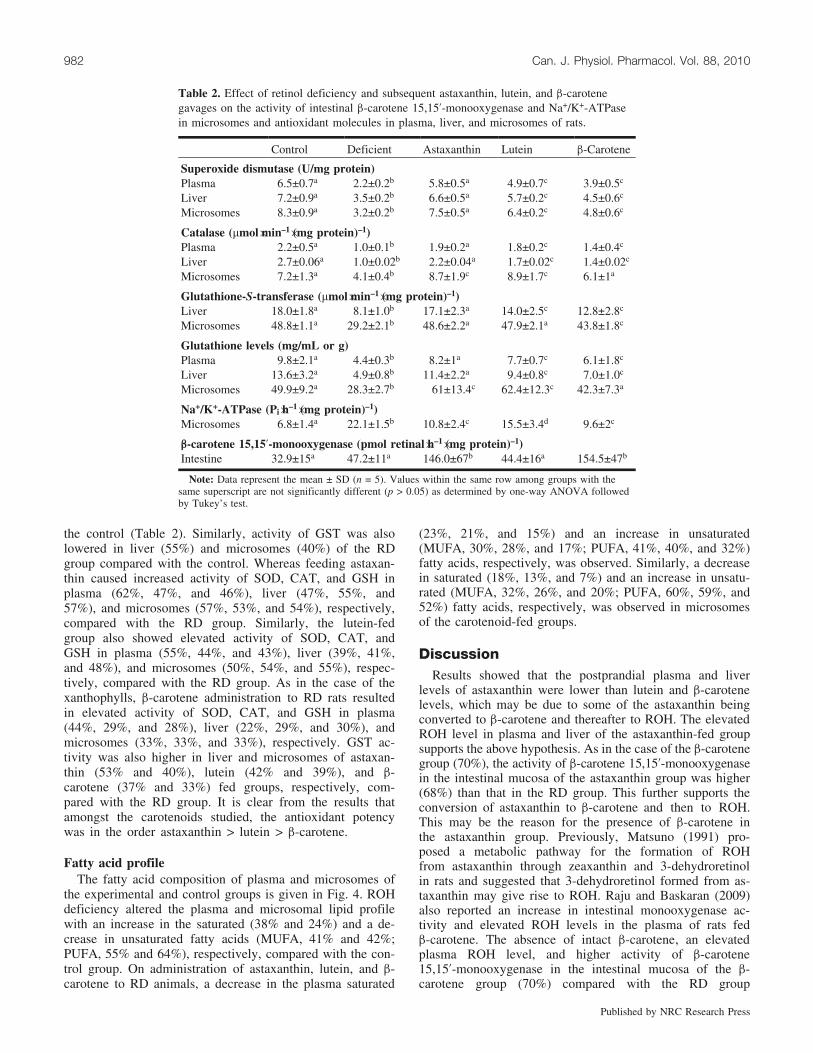
the control (Table 2). Similarly, activity of GST was alsolowered in liver (55%) and microsomes (40%) of the RDgroup compared with the control. Whereas feeding astaxan-thin caused increased activity of SOD, CAT, and GSH inplasma (62%, 47%, and 46%), liver (47%, 55%, and57%), and microsomes (57%, 53%, and 54%), respectively,compared with the RD group. Similarly, the lutein-fedgroup also showed elevated activity of SOD, CAT, andGSH in plasma (55%, 44%, and 43%), liver (39%, 41%,and 48%), and microsomes (50%, 54%, and 55%), respec-tively, compared with the RD group. As in the case of thexanthophylls, b-carotene administration to RD rats resultedin elevated activity of SOD, CAT, and GSH in plasma(44%, 29%, and 28%), liver (22%, 29%, and 30%), andmicrosomes (33%, 33%, and 33%), respectively. GST ac-tivity was also higher in liver and microsomes of astaxan-thin (53% and 40%), lutein (42% and 39%), and b-carotene (37% and 33%) fed groups, respectively, com-pared with the RD group. It is clear from the results thatamongst the carotenoids studied, the antioxidant potencywas in the order astaxanthin > lutein > b-carotene.
Fatty acid profileThe fatty acid composition of plasma and microsomes of
the experimental and control groups is given in Fig. 4. ROHdeficiency altered the plasma and microsomal lipid profilewith an increase in the saturated (38% and 24%) and a de-crease in unsaturated fatty acids (MUFA, 41% and 42%;PUFA, 55% and 64%), respectively, compared with the con-trol group. On administration of astaxanthin, lutein, and b-carotene to RD animals, a decrease in the plasma saturated
(23%, 21%, and 15%) and an increase in unsaturated(MUFA, 30%, 28%, and 17%; PUFA, 41%, 40%, and 32%)fatty acids, respectively, was observed. Similarly, a decreasein saturated (18%, 13%, and 7%) and an increase in unsatu-rated (MUFA, 32%, 26%, and 20%; PUFA, 60%, 59%, and52%) fatty acids, respectively, was observed in microsomesof the carotenoid-fed groups.
DiscussionResults showed that the postprandial plasma and liver
levels of astaxanthin were lower than lutein and b-carotenelevels, which may be due to some of the astaxanthin beingconverted to b-carotene and thereafter to ROH. The elevatedROH level in plasma and liver of the astaxanthin-fed groupsupports the above hypothesis. As in the case of the b-carotenegroup (70%), the activity of b-carotene 15,15’-monooxygenasein the intestinal mucosa of the astaxanthin group was higher(68%) than that in the RD group. This further supports theconversion of astaxanthin to b-carotene and then to ROH.This may be the reason for the presence of b-carotene inthe astaxanthin group. Previously, Matsuno (1991) pro-posed a metabolic pathway for the formation of ROHfrom astaxanthin through zeaxanthin and 3-dehydroretinolin rats and suggested that 3-dehydroretinol formed from as-taxanthin may give rise to ROH. Raju and Baskaran (2009)also reported an increase in intestinal monooxygenase ac-tivity and elevated ROH levels in the plasma of rats fedb-carotene. The absence of intact b-carotene, an elevatedplasma ROH level, and higher activity of b-carotene15,15’-monooxygenase in the intestinal mucosa of the b-carotene group (70%) compared with the RD group
Table 2. Effect of retinol deficiency and subsequent astaxanthin, lutein, and b-carotenegavages on the activity of intestinal b-carotene 15,15’-monooxygenase and Na+/K+-ATPasein microsomes and antioxidant molecules in plasma, liver, and microsomes of rats.
Control Deficient Astaxanthin Lutein b-Carotene
Superoxide dismutase (U/mg protein)Plasma 6.5±0.7a 2.2±0.2b 5.8±0.5a 4.9±0.7c 3.9±0.5c
Liver 7.2±0.9a 3.5±0.2b 6.6±0.5a 5.7±0.2c 4.5±0.6c
Microsomes 8.3±0.9a 3.2±0.2b 7.5±0.5a 6.4±0.2c 4.8±0.6c
Catalase (mmol�min–1�(mg protein)–1)Plasma 2.2±0.5a 1.0±0.1b 1.9±0.2a 1.8±0.2c 1.4±0.4c
Liver 2.7±0.06a 1.0±0.02b 2.2±0.04a 1.7±0.02c 1.4±0.02c
Microsomes 7.2±1.3a 4.1±0.4b 8.7±1.9c 8.9±1.7c 6.1±1a
Glutathione-S-transferase (mmol�min–1�(mg protein)–1)Liver 18.0±1.8a 8.1±1.0b 17.1±2.3a 14.0±2.5c 12.8±2.8c
Microsomes 48.8±1.1a 29.2±2.1b 48.6±2.2a 47.9±2.1a 43.8±1.8c
Glutathione levels (mg/mL or g)Plasma 9.8±2.1a 4.4±0.3b 8.2±1a 7.7±0.7c 6.1±1.8c
Liver 13.6±3.2a 4.9±0.8b 11.4±2.2a 9.4±0.8c 7.0±1.0c
Microsomes 49.9±9.2a 28.3±2.7b 61±13.4c 62.4±12.3c 42.3±7.3a
Na+/K+-ATPase (Pi�h–1�(mg protein)–1)Microsomes 6.8±1.4a 22.1±1.5b 10.8±2.4c 15.5±3.4d 9.6±2c
b-carotene 15,15’-monooxygenase (pmol retinal�h–1�(mg protein)–1)Intestine 32.9±15a 47.2±11a 146.0±67b 44.4±16a 154.5±47b
Note: Data represent the mean ± SD (n = 5). Values within the same row among groups with thesame superscript are not significantly different (p > 0.05) as determined by one-way ANOVA followedby Tukey’s test.
982 Can. J. Physiol. Pharmacol. Vol. 88, 2010
Published by NRC Research Press
observed in this study may be attributable to the biocon-version of b-carotene to ROH.
Lipids are an integral part of tissues and cell membranes.Lipid peroxidation is a marker for oxidative stress either bypro-oxidants or ROH deficiency. Hence, those antioxidantcarotenoids that suppress lipid peroxidation at the membranelevel are of importance. Depending upon their polarity,structure, and attached functional groups, they are localizedin the membrane where they protect the lipids from peroxi-dation (Woodall et al. 1997). The results of this study showthat ROH deficiency increased lipid peroxidation compared
with the control and it was suppressed in carotenoid-fedgroups, especially in microsomes of the astaxanthin group.The possible reason for this could be the dual presence ofintact astaxanthin as well as newly formed ROH, which to-gether resulted in greater suppression of lipid peroxidationin plasma (45%), liver (59%), and microsomes (77%). Reti-noids and carotenoids are an integral part of the membraneand may afford protection to the membrane-bound lipids(Britton 1995). The reason for differences in the potenciesof carotenoids used in this study may lie in their alignmentin the membrane, wherein, being highly hydrophobic, b-carotene is localized in the lipid core of the membrane andis arranged parallel to the lipid chains and offers protectionto the lipids (Woodall et al. 1997). Otherwise, ROH formedfrom b-carotene may associate with the cell membrane andprovide protection (Palacios et al. 1996). ROH formed fromastaxanthin in this study may have afforded similar protec-tion to the microsomes. On the other hand, astaxanthin andlutein hold polar functional groups (hydroxyl groups) at-tached to their b-ionone rings and the keto-groups arrangedperpendicular may offer protection at the periphery (Goto etal. 2001). Thus, these xanthophylls may protect both theaqueous and lipid phases of the cell. This is especially truefor astaxanthin, which is more reactive owing to the pres-ence of both hydroxyl and keto-groups on its b-ionone ring.The results show that the relative abilities of astaxanthin, lu-tein, and b-carotene to reduce lipid peroxidation caused byROH deficiency may be influenced by the presence of func-
Fig. 3. Effect of retinol deficiency and subsequent gavage with as-taxanthin, lutein, and b-carotene on the lipid peroxidation levels inplasma and liver, (a) and liver microsomes (b) of rats. Data repre-sent the mean ± SD (n = 5). Values among groups with the samesuperscript are not significantly different (p > 0.05) as determinedby one-way ANOVA followed by Tukey’s test. Deficient, retinoldeficient; and TMP, 1,1,3,3-tetramethoxypropane.
Fig. 4. Effect of retinol deficiency and subsequent gavage of astax-anthin, lutein, and b-carotene on the fatty acid profile in plasma (a)and liver microsomes (b). Data represent the mean ± SD (n = 5).Values among groups with the same superscript are not signifi-cantly different (p > 0.05) as determined by one-way ANOVA fol-lowed by Tukey’s test. Deficient, retinol deficient.
Sangeetha and Baskaran 983
Published by NRC Research Press

tional groups with increasing polarities, as well as by thenumber of double bonds. Although xanthophylls do not pos-sess provitamin-A activity, they have been reported to havesimilar or even greater antioxidant properties compared withb-carotene (McNulty et al. 2007).
The suppression of Lpx by the carotenoids is further cor-roborated by the activity of antioxidant molecules. Vitaminsand carotenoids are known to scavenge peroxides throughup-regulation of the activities of antioxidant enzymes (Fanget al. 2000). In this study, ROH deficiency was associatedwith decreased (p < 0.05) activities of SOD, catalase, GST,and GSH compared with the control group. Decreased Lpxin carotenoid-fed groups may be due to increased antioxi-dant enzyme activities. The order of antioxidant potency ofthe carotenoids was astaxanthin > lutein > b-carotene.
Maintenance of the membrane lipid profile is critical, aschanges due to ROH deficiency would result in its alteredstructure and function. With respect to the fatty acid profileof plasma and microsomes, increased saturated (38%) anddecreased unsaturated (MUFA, 41%; PUFA, 55%) fattyacid levels observed in plasma of the RD group in the cur-rent study were corrected upon administration of astaxan-thin, lutein, and b-carotene. In all 3 carotenoid groups, theplasma (23%, 21%, and 15%) and microsomes (18%, 13%,and 8%) saturated fatty acids were lowered while there wasan increase in the unsaturated fatty acids level in plasma(35%, 33%, and 22%) and microsomes (38%, 33%, and28%) as compared with the RD group. The results thus dem-onstrate that there was amelioration of the fatty acid profiletowards the control in plasma and microsomes of astaxan-thin, lutein, and b-carotene groups and the effectiveness wasin the same order. The alteration in saturated and unsatu-rated fatty acids in astaxanthin, lutein, and b-carotene groupscan be attributed to the fact that these carotenoids might in-hibit desaturase activity and thereby increase levels of unsa-turated fatty acids, suggesting that the metabolism of fattyacids is affected (Grolier et al. 1991).
ROH deficiency alters the membrane-bound enzyme Na+/K+-ATPase (Kaul and Krishnakantha 1997). Hence, it isconsidered a marker for vitamin A deficiency. Our resultsshowed increased Na+/K+-ATPase activity in the RD groupand a decrease (p < 0.05) in the astaxanthin, lutein, and b-carotene groups. The ameliorative effect exhibited by astax-anthin and b-carotene was evident by the lowered activity(51.1% and 56.6%), which was greater than the lutein group(29.9%). A significant reversal in the activity of Na+/K+-ATPase in the astaxanthin and b-carotene groups was prob-ably due to the rise in ROH levels, which was formed as aresult of b-carotene cleavage and may be by conversion ofastaxanthin to b-carotene and thereafter to ROH. On theother hand, lutein did not yield ROH but brought down theactivity of Na+/K+-ATPase and enhanced the activity of anti-oxidant molecules, which may be due to its potent antioxi-dative property. Lowered Lpx in this group may be due tothe above reason, resulting in protection of the membraneand its components.
ROH deficiency in rats resulted in depleted ROH levelsand activity of antioxidant enzymes, while Na+/K+-ATPaseactivity and lipid peroxide levels in plasma, liver, and mi-crosomes were elevated. Carotenoid gavage amelioratedthese effects. b-Carotene and ROH detected in the astaxan-
thin group suggests that astaxanthin may protect the cellmembrane from ROH deficiency, unlike lutein. This is fur-ther supported by the elevated monooxygenase activity inthe astaxanthin group. All 3 carotenoids lowered Lpx in-duced by ROH deficiency in the order astaxanthin > lutein >b-carotene. These results provide greater understanding ofthe role of nonprovitamin-A carotenoids in meeting ROH re-quirements and protecting against ROH deficiency inducedbiochemical changes. Therefore, extracts of microalgae Hae-matococcus sp. and seafood like salmon, trout, red seabream, shrimp, and lobster can be used as sources of astax-anthin for food applications. Since bioconversion of xantho-phylls to ROH was the objective of this study, othermetabolites that may have been formed have not been inves-tigated here. Although studies of expression of mRNAmodulated by retinoic acid on feeding nonprovitamin-A car-otenoids under ROH deficiency is warranted, this reportconfirms that astaxanthin is converted to b-carotene andROH under ROH deficiency in the murine model.
AcknowledgementsSangeetha Ravi Kumar acknowledges the University
Grants Commission, Government of India, New Delhi, forthe award of a Senior Research Fellowship. The authors ac-knowledge the help of Asha M in chromatographic analysesand Adrian Alan Bennett, Tennessee Department of Trans-portation, Nashville, Tennessee, for editing this manuscriptwith reference to English and grammar.
ReferencesAebi, H. 1984. Catalase in vitro. In Oxygen radicals in biological
systems. Methods in enzymology. Vol. 105. Edited by L. Parker.Academic Press, Orlando, Fla. pp. 121–126.
Ames, B.N. 1966. Assay of inorganic phosphate, total phosphateand phosphatases. In Complex carbohydrates. Methods in enzy-mology. Vol. 8. Edited by E.F. Neufeld and V. Ginsburg. Aca-demic Press, New York. pp. 115–118.
Anzulovich, A.C., Oliveros, B.L., Muþoz, E., MartUnez, L.D., andGimOnez, M.S. 2000. Nutritional vitamin A deficiency altersantioxidant defenses and modifies the liver histoarchitecture inrat. J. Trace Elem. Exp. Med. 13: 343–357. doi:10.1002/1520-670X(2000)13:4<343::AID-JTRA3>3.0.CO;2-J.
Britton, G. 1995. Structure and properties of carotenoids in relationto function. FASEB J. 9(15): 1551–1558. PMID:8529834.
Fang, Y.Z., Yang, S., and Wu, G. 2000. Free radicals, antioxidants,and nutrition. Nutrition, 18(10): 872–879. doi:10.1016/S0899-9007(02)00916-4.
Flohe, L., and Otting, F. 1984. Superoxide dismutase assays In Oxygenradicals in biological systems. Methods in enzymology. Vol. 105.Edited by L. Parker. Academic Press, Orlando, Fla. pp. 93–104.
Gluthenberg, C., Alin, P., and Mannervik, B. 1985. Glutathionetransferase from rat testis. In Glutamate, glutamine, glutathioneand related compounds. Methods in enzymology. Vol. 113. Edi-ted by A. Meister. Academic Press, Orlando, Fla. pp. 507–510.
Gonzalez-Flecha, B., Llesuy, S., and Boveris, A. 1991. Hydroper-oxide initiated chemiluminescence: an assay for oxidative stressin biopsies of liver, heart and muscle. Free Radic. Biol. Med.10(2): 93–100. doi:10.1016/0891-5849(91)90002-K.
Goswami, U.C., and Barua, A.C. 1986. Intestinal conversion of lu-tein into 3-dehydroretinol in freshwater fish, Heteropneustes fos-silis and Channa straitus. Indian J. Biochem. Biophys. 18: 88.PMID:7275161.
984 Can. J. Physiol. Pharmacol. Vol. 88, 2010
Published by NRC Research Press

Goto, S., Kogure, K., Abe, K., Kimata, Y., Kitahama, K., Yama-shita, E., and Terada, H. 2001. Efficient radical trapping at thesurface and inside the phospholipid membrane is responsible forhighly potent antiperoxidative activity of the carotenoid astax-anthin. Biochim. Biophys. Acta, 1512(2): 251–258. doi:10.1016/S0005-2736(01)00326-1. PMID:11406102.
Grangaud, R., Massonet, R., Conquy, T., and Ridolfo, J. 1961.Transformation of astaxanthine into vitamin A in albino rats:neoformation in vivo and in vitro. C.R. Hebd. Seance. Acad.Sci. 252: 1854–1856. PMID:14470960.
Grolier, P., Cisti, A., Daubeze, M., and Narbonne, J.F. 1991. Theinfluence of dietary vitamin A intake on microsomal membranefluidity and lipid composition. Nutr. Res. 11(6): 567–574.doi:10.1016/S0271-5317(05)80347-1.
Gross, J., and Budowski, P. 1966. Conversion of carotenoids intovitamins A1 and A2 in two species of freshwater fish. Biochem.J. 101(3): 747–754. PMID:16742455.
Gutman, Y., and Katzper-Shamir, Y. 1971. The effect of urea, so-dium and calcium on microsomal ATPase activity in differentparts of the kidney. Biochim. Biophys. Acta, 233(1): 133–136.doi:10.1016/0005-2736(71)90365-8. PMID:4252906.
Kaplay, S.S. 1978. Erythrocyte membrane Na+ and K+ activatedadenosine triphosphatase in PCM. J. Clin. Nutr. 31: 579–584.
Kaul, S., and Krishnakantha, T.P. 1997. Influence of retinol defi-ciency and curcumin/turmeric feeding on tissue microsomalmembrane lipid peroxidation and fatty acids in rats. Mol. Cell.Biochem. 175(1–2): 43–48. doi:10.1023/A:1006829010327.PMID:9350032.
Kotake-Nara, E., Kushiro, M., Zhang, H., Sugawara, T., Miyashita,K., and Nagao, A. 2001. Carotenoids affect proliferation of hu-man prostate cancer cells. J. Nutr. 131(12): 3303–3306. PMID:11739884.
Lakshminarayana, R., Aruna, G., Sangeetha, R.K., Bhaskar, N., Diva-kar, S., and Baskaran, V. 2008. Possible degradation/biotransfor-mation of lutein in vitro and in vivo: isolation and structuralelucidation of lutein metabolites by HPLC and LC-MS (atmo-spheric pressure chemical ionization). Free Radic. Biol. Med.45(7): 982–993. doi:10.1016/j.freeradbiomed.2008.06.011. PMID:18640265.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., and Randall, R.J. 1951.Protein measurement with the Folin phenol reagent. J. Biol.Chem. 193(1): 265–275. PMID:14907713.
Matsuno, T. 1991. Xanthophylls as precursors of retinoids. PureAppl. Chem. 63(1): 81–88. doi:10.1351/pac199163010081.
McNulty, H.P., Byun, J., Lockwood, S.F., Jacob, R.F., and Mason,R.P. 2007. Differential effects of carotenoids on lipid peroxida-tion due to membrane interactions: X-ray diffraction analysis.Biochim. Biophys. Acta, 1768(1): 167–174. doi:10.1016/j.bbamem.2006.09.010. PMID:17070769.
Moren, M., Naess, T., and Hamre, K. 2002. Conversion of b-caro-tene, canthaxanthin and astaxanthin to vitamin A in Atlantic ha-libut (Hippoglossus hippoglossus L.) juveniles. Fish Physiol.Biochem. 27(1/2): 71–80. doi:10.1023/B:FISH.0000021819.46235.12.
Morrison, W.R., and Smith, L.M. 1964. Preparation of fatty acidmethyl esters and dimethylacetals from boron fluoride-methanol.J. Lipid Res. 5: 600–608. PMID:14221106.
Nagao, A., During, A., Hoshino, C., Terao, J., and Olson, J.A.1996. Stoichiometric conversion of all trans-b-carotene to retinalby pig intestinal extract. Arch. Biochem. Biophys. 328(1): 57–63. doi:10.1006/abbi.1996.0142. PMID:8638938.
Ohkawa, H., Ohishi, N., and Yagi, K. 1979. Assay for lipid perox-ides in animal tissues by thiobarbituric acid reaction. Anal. Bio-chem. 95(2): 351–358. doi:10.1016/0003-2697(79)90738-3.PMID:36810.
Olson, J.A. 1982. New approaches to methods for assessment ofnutritional status of the individual. Am. J. Clin. Nutr. 36: 1166–1168.
Olson, J.A. 1994. Absorption, transport and metabolism of carote-noids in humans. Pure Appl. Chem. 66(5): 1011–1016. doi:10.1351/pac199466051011.
Owens, C.W.I., and Belcher, R.V. 1965. A colorimetric micro-method for the determination of glutathione. Biochem. J. 94:705–711. PMID:14340062.
Palacios, A., Piergiacomi, V.A., and Catala, A. 1996. Vitamin Asupplementation inhibits chemiluminescence and lipid peroxida-tion in isolated rat liver microsomes and mitochondria. Mol.Cell. Biochem. 154(1): 77–82. doi:10.1007/BF00248464. PMID:8717420.
Raju, M., and Baskaran, V. 2009. Bioefficacy of b-carotene is im-proved in rats after solubilized as equimolar dose of b-caroteneand lutein in phospholipid-mixed micelles. Nutr. Res. 29(8):588–595. doi:10.1016/j.nutres.2009.07.005. PMID:19761893.
Rajyalakshmi, P., Venkatalaxmi, K., Venkatalakshmamma, K.,Jyothsna, Y., Balachandramani Devi, K., and Suneetha, V.2001. Total carotenoid and beta-carotene contents of forestgreen leafy vegetables consumed by tribals of south India. PlantFoods Hum. Nutr. 56(3): 225–238. doi:10.1023/A:1011125232097. PMID:11442223.
Sangeetha, R.K., Bhaskar, N., and Baskaran, V. 2009. Comparativeeffects of beta-carotene and fucoxanthin on retinol deficiency in-duced oxidative stress in rats. Mol. Cell. Biochem. 331(1–2):59–67. doi:10.1007/s11010-009-0145-y. PMID:19421712.
Smith, J.E. 1990. Preparation of vitamin A-deficient rats and mice.In Retinoids part B: cell differentiation and clinical application.Methods in enzymology. Vol. 190. Edited by T. Kealy and L.Parker. Academic Press, San Diego, Calif. pp. 229–236.
West, C.E. 2000. Meeting requirements for vitamin A. Nutr. Rev.58(11): 341–345. doi:10.1111/j.1753-4887.2000.tb01831.x.PMID:11140905.
Woodall, A.A., Britton, G., and Jackson, M.J. 1997. Carotenoidsand protection of phospholipids in solution or in liposomesagainst oxidation by peroxyl radicals: relationship between caro-tenoid structure and protective ability. Biochim. Biophys. Acta,1335: 575–586.
World Health Organization. 2000. Vitamin A deficiency. Availablefrom http://www.who.int/vaccines-diseases [accessed 15 June2007].
Sangeetha and Baskaran 985
Published by NRC Research Press





























