Embed Size (px)
Citation preview

RRNNcclloorroopplláá
rreeppll
DD
Departament de Bioquímica iBiología Molecular
AA lliiggaassaass nnuucclleeaarreess yy ssttiiccaass:: ppoossiibbllee ppaappeell eenn llaa iiccaacciióónn ddee llooss vviirrooiiddeess
iieeggoo MMoolliinnaa SSeerrrraannoo VVaalleenncciiaa,, 22000088


CCoonnffííaa eenn eell ttiieemmppoo,, qquuee ssuueellee ddaarr dduullcceess ssaalliiddaass aa mmuucchhaass
aammaarrggaass ddiiffiiccuullttaaddeess..
MMiigguueell ddee CCeerrvvaanntteess


AA mmiiss ppaaddrreess yy hheerrmmaannooss


RRNNAA LLIIGGAASPPOOSSIIBBLLEE
Departament de Bioquímica i Biología Molecular
SAASS NNUUCCLLEEAARREESS YY CCLLOORROOPPLLÁÁSSTTIICCAASS:: PPAAPPEELL EENN LLAA RREEPPLLIICCAACCIIÓÓNN DDEE LLOOSS
VVIIRROOIIDDEESS
Memoria de Tesis Doctoral presentada por
DIEGO MOLINA SERRANO
para optar al grado de
DOCTOR EN BIOLOGÍA
Directores
José Antonio Daròs Arnau
Ricardo Flores Pedauyé
Valencia, 2008


Don Ricardo Flores Pedauyé, Doctor en Ciencias Químicas, Profesor de Investigación del Consejo Superior de Investigaciones Científicas, y Don José Antonio Daròs Arnau, Doctor en Ciencias Biológicas, Científico Titular, ambos pertenecientes al Instituto de Biología Molecular y Celular de Plantas (Universidad Politécnica de Valencia-Consejo Superior de Investigaciones Científicas) de Valencia, CERTIFICAN: que Don Diego Molina Serrano ha realizado bajo su dirección el trabajo que con el título ‘RNA ligasas nucleares y cloroplásticas: posible papel en la replicación de los viroides’ presenta para optar al grado de Doctor en Ciencias Biológicas. Para que así conste a los efectos oportunos, firman el presente certificado en Valencia, a 10 de Octubre de 2008.
Fdo. Ricardo Flores Pedauyé Fdo. José Antonio Daròs Arnau




Agradecimientos
Agradecimientos En primer lugar quisiera agradecer a mis directores la posibilidad de realizar esta
tesis en sus laboratorios. A José Antonio Daròs su dedicación incondicional a nuestro
proyecto, y a Ricardo Flores sus consejos e intentos de transmitirme su experiencia;
procuraré que no caigan en saco roto.
Parte de esta tesis no podría haberse realizado sin la colaboración de las
Doctoras Marisa Salvador y Loreto Suay, que aportaron su trabajo y esfuerzo, siendo
mis particulares Ariadnas en el laberinto de las Chlamydomonas.
Quiero recordar la ayuda de los Doctores Ismael Rodrigo y Susana Tárraga en
mis intentos de purificación enzimática por técnicas cromatográficas, que si bien tienen
poco reflejo en esta Tesis, no por ello fue una tarea menos ardua, ni merecen menos
reconocimiento.
Por supuesto no me puedo olvidar de la gente que trabaja en el IBMCP, MD
(Chispi) por ayudarme con mis tímidas aproximaciones al microscopio y sobre todo por
ser como es, Eugenio y Ana por su estupendo trabajo de secuenciación, la gente del
invernadero, especialmente Nacho, por cuidar de mis espinacas como si fueran suyas,
Assumpta por conseguirme esos artículos imposibles, Santiago y José por mantener el
material en perfecto funcionamiento, Rafa por procurar orden en la sala de isótopos,
Alexis y Ramón por ayudarme a mantener el ordenador en condiciones de trabajo, las
chicas de administración por hacerme el papeleo un poco más fácil, Juni y Estela por
tramitar mis pedidos, y a todos aquellos que en alguna ocasión me han ayudado con
consejos y/o material.
Que habría sido de mí sin la inestimable colaboración de mis compañeros del
laboratorio de Ricardo, que me han ayudado desde el principio hasta el final de esta
tesis. Selma, cuyos consejos tanto científicos como personales han sido tan valiosos
como nuestras conversaciones literarias, Emilio, que siempre ha tenido un minuto para
contestar a mis dudas y responder a mis bromas, Sonia, cuya incondicional ayuda ha
sido inestimable tantas y tantas veces, Alberto, con quien he compartido tantas penas y
alegrías de la Tesis, Douglas, porque sus dedicación y amistad han sido inestimables
I

Agradecimientos
para encontrar mi postdoc, Amparo, porque ha sido como una madre para mi, y el resto
de gente que ha pasado por el laboratorio, porque siempre me dieron buenos consejos.
Muchos de estos experimentos no habrían salido sin ellos. Gracias también a todo el
grupo de virus y viroides porque sus críticas constructivas ayudaron a mejorar este
trabajo, y a Toni especialmente, porque sin el mis cloroplastos no serían los mismos.
Mis más de cuatro años de tesis no se pueden recordar sin mencionar a
muchísima gente, y espero no dejarme a nadie en el tintero. A mis otros dos compañeros
fundadores del consejo, Jorge y Pedro, porque vuestra amistad no se expresa con
palabras. A J. Antonio (Kapi) y Pablo porque siempre intentásteis que viviésemos
juntos, y ahora no se como podré abandonaros, y a Barto porque es parte del alma de
ese piso. A Santi y Carlos (el alumno por siempre jamás) por nuestras fantásticas noche
de fútbol y sepias, a Patricia porque siempre pude confiar en ella, a Steph por compartir
conmigo los buenos y malos momentos, a Aurora por su cariño, a Mª Ángeles por su
compañerismo en el laboratorio y fuera de el, a Marina por ser durante tanto tiempo mi
amiga, a Vicent, Antoñete y Hugo porque las risas que nos hemos echado durante
nuestros cónclaves y E-mails del consejo no se pagan con dinero, a Alessandro por ser
tan auténtico y espontáneo, a Leti y Sandra porque siempre han tenido una palabra de
ánimo, a Carlos (amorsote), Silvia, Jorge Lozano y Teresa por tantos y tantos bailoteos.
A la gente que anima la hora de la comida y a todos aquellos que saludan por las
mañanas.
Como todo en la vida no es la Tesis, aunque a veces lo parezca, tampoco puedo
olvidar a Juan por ayudarme en mis inicios científicos y por tantos viajes maravillosos
juntos con sus correspondientes anécdotas, a Susana porque siempre he sentido su
apoyo desde Alicante, a Roque Bru por acogerme en su laboratorio cuando solo era un
polluelo y a Luis por seguir mis andanzas con interés desde Venezuela. A Juanjo (el
mofli) por tantos almuerzos vileros de nardo y “punxaetes”, y a toda la gente con la que
he compartido cervezas y alegrías. A Elisabetta, porque desde la distancia siempre
estuvo conmigo en los momentos más duros, y solo ella sabe cuan frecuentes fueron y
cuanto me ayudó.
A mis padres, porque sin ellos nunca lo habría conseguido, a mis hermanos, por
creer siempre en mí, y a mi abuela, porque todo lo que soy es por ella.
II



Resumen
Resumen Los viroides están constituidos por una molécula circular de RNA
monocatenario que no codifica proteínas. A pesar de su pequeño tamaño, entre 246-401
nt, son capaces de replicarse autónomamente, moverse sistémicamente, y en la mayoría
de los casos provocar enfermedades en sus plantas hospedadoras. Los viroides se
replican en el núcleo (familia Pospiviroidae) o en el cloroplasto (familia Avsunviroidae)
mediante un modelo de círculo rodante que comprende tres etapas: transcripción RNA-
RNA, corte y ligación. En el presente trabajo hemos profundizado en el estudio de la
tercera etapa del ciclo replicativo de estos agentes infecciosos, la circularización de los
RNAs monómericos, caracterizando una RNA ligasa nuclear y otra cloroplástica que
podrían catalizar la ligación de los miembros de la familia Pospiviroidae y
Avsunviroidae, respectivamente. Además, hemos desarrollado un sistema heterólogo
basado en la transformación del cloroplasto de Chlamydomonas reinhardti que resulta
interesante para estudiar in vivo el ciclo replicativo de los miembros de la familia
Avsunviroidae.
A partir de cloroplastos íntegros de espinaca, y mediante fraccionamiento de sus
proteínas por cromatografía en una columna de heparina, se ha detectado una actividad
que cataliza la circularización de los RNAs monoméricos lineales del viroide del
manchado solar del aguacate (Avocado sunblotch viroid, ASBVd) y del viroide latente
de la berenjena (Eggplant latent viroid, ELVd) resultantes del autocorte mediado por
ribozimas de cabeza de martillo. Sin embargo, no ha podido detectarse en las mismas
condiciones la circularización de los otros dos miembros de la familia Avsunviroidae, el
viroide del moteado clorótico del crisantemo (Chrysanthemum clorotic mottle viroid,
CChMVd) y el viroide del mosaico latente del melocotonero (Peach latent mosaic
viroid, PLMVd). Dicha actividad, la primera de esta clase identificada en cloroplastos,
reside en una enzima que no requiere ni NTPs ni Mg2+ como cofactores, a diferencia de
la mayoría de RNA ligasas conocidas. Experimentos de extensión de cebador indican
que el enlace formado por la RNA ligasa cloroplástica no es 3’,5’-fosfodiéster como
ocurre en las reacciones catalizadas por las RNA ligasas nucleares, sino un enlace
atípico que podría ser 2’,5’-fosfodiéster como el que generan las 2’,5’-RNA ligasas de
bacterias y arqueas.
La estrategia de RACE circular sobre molde de simple cadena puesta a punto en
el presente trabajo ha permitido identificar el mRNA de la tRNA ligasa de tomate a
V

Resumen
partir de la información proporcionada por etiquetas de secuencias expresadas
(expressed sequence tag, EST) de tomate con similitud de secuencia con los mRNAs de
las tRNA ligasas de plantas conocidas. El cDNA correspondiente a la tRNA ligasa de
tomate identificada fue clonado y expresado en la cepa de Escherichia coli Rosetta 2,
que corrige la preferencia codónica y facilita la traducción de proteínas heterólogas de
gran tamaño. La expresión en este sistema bacteriano de la tRNA ligasa de tomate y su
homóloga de Arabidopsis thaliana condujo a rendimientos desiguales, probablemente
debido a que ambas proteínas tienen tiempos de vida media distintos. Ambas enzimas, a
diferencia de sus homólogas de trigo y levadura, catalizan la ligación con otros NTPs
además de ATP, si bien el resto de propiedades bioquímicas ensayadas son muy
similares en todas ellas.
Finalmente, se ha transformado el cloroplasto del alga unicelular
Chlamydomonas reinhardtii para expresar heterólogamente RNAs diméricos de ambas
polaridades de diversos miembros de la familia Avsunviroidae. Estos RNAs, además de
expresarse correctamente en el cloroplasto de dicha alga, son procesados a sus formas
lineales y circulares en una proporción similar a la observada en extractos de tejidos
infectados con los mismos viroides. Por el contrario, el RNA dimérico del CEVd, un
viroide de replicación y localización nuclear, es transcrito pero no procesado. Sin
embargo, y a diferencia de lo que ocurre en sus hospedadores naturales, en el
cloroplasto de C. reinhardtii no se ha detectado transcripción RNA-RNA,
probablemente por la presumible ausencia de la RNA polimerasa cloroplástica
dependiente de DNA codificada en el núcleo.
VI



Abreviaturas
Abreviaturas ACNasa: anticodón nucleasa
ASBVd: viroide del manchado solar del aguacate
CCCVd: viroide del cadang-cadang del cocotero
CCR: región central conservada
CChMVd: viroide del moteado clorótico del crisantemo
CEVd: viroide de la exocortis de los cítricos
CPD y CPDasa: dominio y enzima ciclofosfodiesterasa, respectivamente
CpU: citidilil-2’,5’-uridilato (pCpU y CpUp: citidilil-2’,5’-uridilato fosforilado en 5’ y
3’, respectivamente)
DTT: ditiotreitol
EDTA: ácido etilendiaminotetraacético
ELVd: viroide latente de la berenjena
EST: marcador de secuencias expresadas
HDV: virus de la hepatitis delta
gRNA: RNA guía
HS: medio de alta concentración de sales para Chlamydomonas reinhardtii HSHA: medio de alta concentración de sales con acetato para Chlamydomonas
reinhardtii IPTG: isopropil-β-D-tiogalactopiranósido IR: región invertida LB: medio Luria-Bertani
NEP: RNA polimerasa cloroplástica codificada en el núcleo
nt: nucleótido
NTA: ácido nitriloacético
ORF: pauta de lectura abierta
PAGE: electroforesis en gel de poliacrilamida
pb: par de bases
PEP: RNA polimerasa cloroplástica codificada en el plastidio
PLMVd: viroide del mosaico latente del melocotonero
PNK: polinucleótido quinasa (dominio y enzima)
PSTVd: viroide del tubérculo fusiforme de la patata
PVP: polivinilpirrolidona
IX

Abreviaturas
RACE: amplificación rápida de los extremos de un cDNA
RLM-RACE: amplificación rápida de los extremos de un cDNA mediada por una RNA
ligasa
SDS: dodecil sulfato sódico
TCH: horquilla terminal conservada
TCR: región terminal conservada
TLC: cromatografía en capa fina
X-Gal: 5-bromo-4-cloro-3-indolil- beta-D-galactopiranósido
X



Índice
AGRADECIMIENTOS ................................................................................................. I
RESUMEN ................................................................................................................... V
ABREVIATURAS ...................................................................................................... IX
ÍNDICE ..................................................................................................................... XIII
1. INTRODUCCIÓN ..................................................................................................... 1
1.1. Los viroides .............................................................................................................. 3
1.1.1. Características generales ................................................................................ 3 1.1.2. Estructura y clasificación de los viroides ....................................................... 3 1.1.3. Síntomas causados por los viroides ................................................................ 9 1.1.4. Localización subcelular................................................................................... 9
1.1.5. Replicación de los viroides ......................................................................... 10
1.1.6. Enzimas y ribozimas implicadas en el ciclo replicativo de los viroides ...... 11
1.1.6.1. Iniciación y elongación ................................................................... 12
1.1.6.2. Corte ................................................................................................ 14
1.1.6.3. Ligación ......................................................................................... 16
1.2. Las RNA ligasas ................................................................................................... 18
1.2.1 Características generales y mecanismos de ligación .................................... 18
1.2.2. RNA ligasas víricas ..................................................................................... 22
1.2.2.1. T4.1 RNA ligasa ............................................................................ 22
1.2.2.2. T4.2 RNA ligasa ............................................................................ 23
1.2.2.3. Otras RNA ligasas víricas .............................................................. 24
1.2.3. RNA ligasas de bacterias y arqueas ............................................................ 25
1.2.3.1. 2’,5’-RNA ligasa ............................................................................ 25
1.2.3.2. Otras RNA ligasas de bacterias y arqueas ..................................... 26
1.2.4. RNA ligasas eucariotas ............................................................................... 27
1.2.4.1. tRNA ligasa .................................................................................... 27
1.2.4.1.1. Características generales y función biológica ................ 27
1.2.4.1.2. Mecanismo de ligación .................................................. 30
1.2.4.2. RNA ligasas mitocondriales .......................................................... 32
1.2.4.3. Otras RNA ligasas eucariotas ........................................................ 33
XIII

Índice
1.2.5. Comparación entre mecanismos .................................................................. 34
1.3. Chlamydomonas reinhardtii como organismo modelo ....................................... 37
1.3.1. Características generales .............................................................................. 37
1.3.2. El cloroplasto y su origen endosimbióntico ................................................. 39
1.3.3. El genoma de Chlamydomonas reinhardtii ................................................. 40
1.3.4. Regulación de la expresión génica .............................................................. 41
1.3.5. Transformación del cloroplasto ................................................................... 42
2. OBJETIVOS ............................................................................................................ 45
3. MATERIALES Y MÉTODOS ............................................................................... 49
3.1. Extracción de proteínas de cloroplastos de espinaca ........................................ 51
3.1.1. Obtención de cloroplastos intactos .............................................................. 51
3.1.2. Rotura de los cloroplastos y solubilización de las proteínas ....................... 51
3.1.3. Fraccionamiento de proteínas por cromatografía en columna
de heparina .................................................................................................. 52
3.2. Obtención de un extracto de germen de trigo con actividad RNA ligasa ....... 52
3.3. Síntesis in vitro de RNA viroidal ........................................................................ 53
3.4. Análisis de la actividad RNA ligasa .................................................................... 54
3.5. Fraccionamiento de RNAs en PAGE .................................................................. 55
3.6. Transferencia de RNAs a membranas de hibridación ...................................... 55
3.7. Análisis mediante hibridación northern ............................................................ 56
3.8. Extracción y fraccionamiento de mRNA de plantas ......................................... 56
3.9. Transcripción inversa (reverse transcription, RT) ............................................. 57
3.10. Reacción en cadena de la polimerasa (polymerase chain reaction, PCR) ...... 57
3.11. Amplificación rápida de los extremos de un cDNA (rapid amplification
of cDNA ends, RACE) ........................................................................................ 58
3.11.1. 3’-RACE ................................................................................................... 58
3.11.2. 5’-RACE ................................................................................................... 58
3.11.2.1. Adición al cDNA de una cola en 3’ con transferasa
terminal ..................................................................................... 58
XIV

Índice
3.11.2.2. Amplificación rápida de los extremos de un cDNA
mediada por RNA ligasa (RNA ligase-mediated
rapid amplification of cDNA ends, RLM-RACE) .................. 59
3.11.2.3. Selección y amplificación de mRNAs con estructura de
caperuza mediante un adaptador de DNA (Cap-finder) .......... 60
3.11.2.4. PCR con cebadores degenerados ............................................. 60
3.11.3. RACE circular .......................................................................................... 61
3.11.3.1. cDNA molde de doble cadena ................................................. 61
3.11.3.2. cDNA molde de simple cadena ................................................ 61
3.11.4. Análisis de los productos obtenidos en los experimentos de RACE ........ 62
3.12. Electroforesis en geles de agarosa .................................................................... 63
3.13. Modificaciones de los ácidos nucleicos ............................................................. 63
3.13.1. Fosforilación ............................................................................................ 63
3.13.2. Desfosforilación ...................................................................................... 63
3.13.2.1. Extremos 5’ y 3’ ....................................................................... 63
3.13.2.2. Extremos 2’,3’-fosfodiéster cíclico .......................................... 64
3.14. Extensión del cebador ........................................................................................ 64
3.15. Detección de enlaces anómalos en el RNA por cromatografía en capa
fina (thin layer cromatography, TLC) ............................................................... 65
3.16. Obtención de construcciones para expresar proteínas en E. coli
Rosetta 2 .............................................................................................................. 67
3.17. Expresión de proteínas en E. coli Rosetta 2 ..................................................... 67
3.17.1. Inducción ................................................................................................. 67
3.17.2. Extracción ................................................................................................ 68
3.17.3. Purificación ............................................................................................. 68
3.18. Electroforesis de proteínas en geles de poliacrilamida desnaturalizantes
(SDS-PAGE) ....................................................................................................... 69
3.19. Construcciones para expresar RNAs viroidales en el cloroplasto de
Chlamydomonas reinhardtii ............................................................................... 70
3.20. Transformación y crecimiento de cultivos de C. reinhardtii .......................... 71
3.21. Extracción y fraccionamiento de RNA viroidal de C. reinhardtii .................. 72
XV

Índice
3.22. Construcción de un árbol filogenético de las tRNA ligasas de plantas ......... 72
4. RESULTADOS ........................................................................................................ 75
4.1. Actividad RNA ligasa en cloroplastos aislados de espinaca ............................. 77
4.1.1. Obtención de una fracción proteica de cloroplastos de espinaca con
actividad RNA ligasa .................................................................................. 77
4.1.2. Efecto de la fuerza iónica en la actividad RNA ligasa cloroplástica ........... 78
4.1.3. Efecto de los ribonucleótidos trifosfato en la actividad RNA ligasa
cloroplástica ................................................................................................ 79
4.1.4. Inactivación de la actividad RNA ligasa cloroplástica por proteólisis y
desnaturalización térmica ........................................................................... 80
4.1.5. Cinética de las reacciones catalizadas por la RNA ligasa de espinaca y
de germen de trigo ...................................................................................... 81
4.1.6. Estudio del factor limitante de la reacción de ligación ................................ 83
4.1.7. Efecto del pH y del Mg2+ en la actividad RNA ligasa cloroplástica ........... 85
4.1.8. Extremos del RNA requeridos por la RNA ligasa cloroplástica ................. 86
4.1.9. Actividad de la RNA ligasa cloroplástica sobre los RNAs monoméricos
de la familia Avsunviroidae y autoligación de los mismos ......................... 88
4.1.10. Determinación del enlace generado por la RNA ligasa cloroplástica ....... 90
4.1.11. Purificación de la RNA ligasa cloroplástica y posible relación con
otras proteínas ............................................................................................ 95
4.2. Clonación, secuenciación y expresión de la tRNA ligasa de tomate
(Solanum lycopersicum) ........................................................................................ 97
4.2.1 Búsqueda de etiquetas de secuencias expresadas (expressed sequence
tags, ESTs) de tomate con similitud con los genes de las tRNA ligasas
de plantas ...................................................................................................... 97
4.2.2. Clonación del cDNA candidato de la tRNA ligasa de tomate ..................... 98
4.2.2.1. Empleo de técnicas de amplificación rápida de los extremos
de un cDNA (Rapid amplification of cDNA ends, RACE)
para determinar las regiones terminales 5’ y 3’ ............................. 98
4.2.2.2. Empleo de la técnica de RACE circular para determinar la
región 5’ del mRNA candidato a codificar la tRNA ligasa
de tomate ...................................................................................... 100
XVI

Índice
4.2.2.3. Amplificación, secuenciación y análisis del cDNA completo
de la presumible tRNA ligasa de tomate ..................................... 102
4.2.3. Clonación y expresión de los cDNAs que codifican las tRNA ligasas
de tomate y A. thaliana ............................................................................. 109
4.2.4. Caracterización bioquímica comparada de las tRNA ligasas de tomate
y A. thaliana .............................................................................................. 114
4.2.4.1. Efecto del pH ............................................................................... 114
4.2.4.2. Efecto de la temperatura .............................................................. 115
4.2.4.3. Requerimiento de ribonucleótidos trifosfato ............................... 116
4.3. Chlamydomonas reinhardtii: un sistema modelo para el estudio de
factores del hospedador implicados en la replicación de los viroides
cloroplásticos ..................................................................................................... 119
4.3.1. Transformación de C. reinhardtii y selección de líneas
homoplásmicas .......................................................................................... 119
4.3.2. Obtención de plásmidos para transformar C. reinhardtii con cDNAs
viroidales ................................................................................................... 121
4.3.3. Análisis del corte y la ligación de los RNAs viroidales en líneas
transplastómicas de C. reinhardtii ...................................................................... 122
4.3.3.1. ASBVd ......................................................................................... 125
4.3.3.2. CChMVd ...................................................................................... 125
4.3.3.3. ELVd ............................................................................................ 127
4.3.3.4. CEVd ........................................................................................... 128
4.3.4. Análisis de la replicación de los viroides por círculo rodante en las
C. reinhardtii transformadas ..................................................................... 128
5. DISCUSIÓN .......................................................................................................... 129
5.1. Una RNA ligasa cloroplástica capaz de catalizar la circularización de
los RNAs de ciertos miembros de la familia Avsunviroidae ........................... 134
5.2. La tRNA ligasa de tomate: un hospedador natural de viroides .................... 138
5.3. Los RNAs de la familia Avsunviroidae se procesan correctamente en el
cloroplasto de C. reinhardtii .............................................................................. 143
6. CONCLUSIONES ................................................................................................ 147
XVII

Índice
7. BIBLIOGRAFÍA ................................................................................................... 151
8. ANEJOS ................................................................................................................. 179
I. Cebadores ........................................................................................................... 181
II. Servidores Web ................................................................................................. 183
XVIII



11.. IINNTTRROODDUUCCCCIIÓÓNN


Introducción
1.1. Los viroides
1.1.1. Características generales
Los viroides son pequeños RNAs circulares desnudos de cadena simple con un
elevado contenido en estructura secundaria (Diener, 2001; Flores et al., 2004; Tabler y
Tsagris, 2004; Flores et al., 2005a; Daròs et al., 2006; Ding e Itaya, 2007). Hasta su
descubrimiento se había considerado que los sistemas genéticos más pequeños eran los
virus, que son capaces de manipular el aparato de traducción de las células que infectan
para dirigir la síntesis de proteínas necesarias para completar su ciclo infeccioso. La
caracterización del agente causal de la enfermedad del tubérculo fusiforme de la patata
(Potato spindle tuber viroid, PSTVd) (Diener, 1971), el primer viroide descrito, supuso
una sorpresa pues su genoma tiene un tamaño aproximadamente diez veces inferior al
genoma de los virus de RNA más sencillos conocidos. Los viroides no codifican
proteína alguna, como muestran los experimentos realizados tanto in vitro (Davies et
al., 1974; Hall et al., 1974) como in vivo (Semancik et al., 1977). A causa de su
simplicidad, los viroides dependen en mayor medida del hospedador que los virus, ya
que deben parasitar la maquinaria transcripcional de la célula para ser transcritos por
una de sus RNA polimerasas.
La simplicidad estructural y funcional de los viroides, junto con la actividad
catalítica que presentan algunos de ellos (ver más adelante), ha conducido a
considerarlos como fósiles moleculares derivados de los primeros replicones que
aparecieron en la Tierra, que se postula estaban constituidos por RNA porque es la
única biomolécula conocida capaz de almacenar información genética y expresar
actividad catalítica (Diener, 1989).
1.1.2. Estructura y clasificación de los viroides
Hasta el momento se han caracterizado biológica y molecularmente 29 especies
de viroides y numerosas variantes de secuencia de las mismas (Tabla 1.1), que se han
clasificado en dos familias: Pospiviroidae, cuya especie tipo es el PSTVd (Diener,
1971; Gross et al., 1978), y Avsunviroidae, cuya especie tipo es el viroide del manchado
solar del aguacate (Avocado sunblotch viroid, ASBVd) (Symons, 1981).
3
Introducción
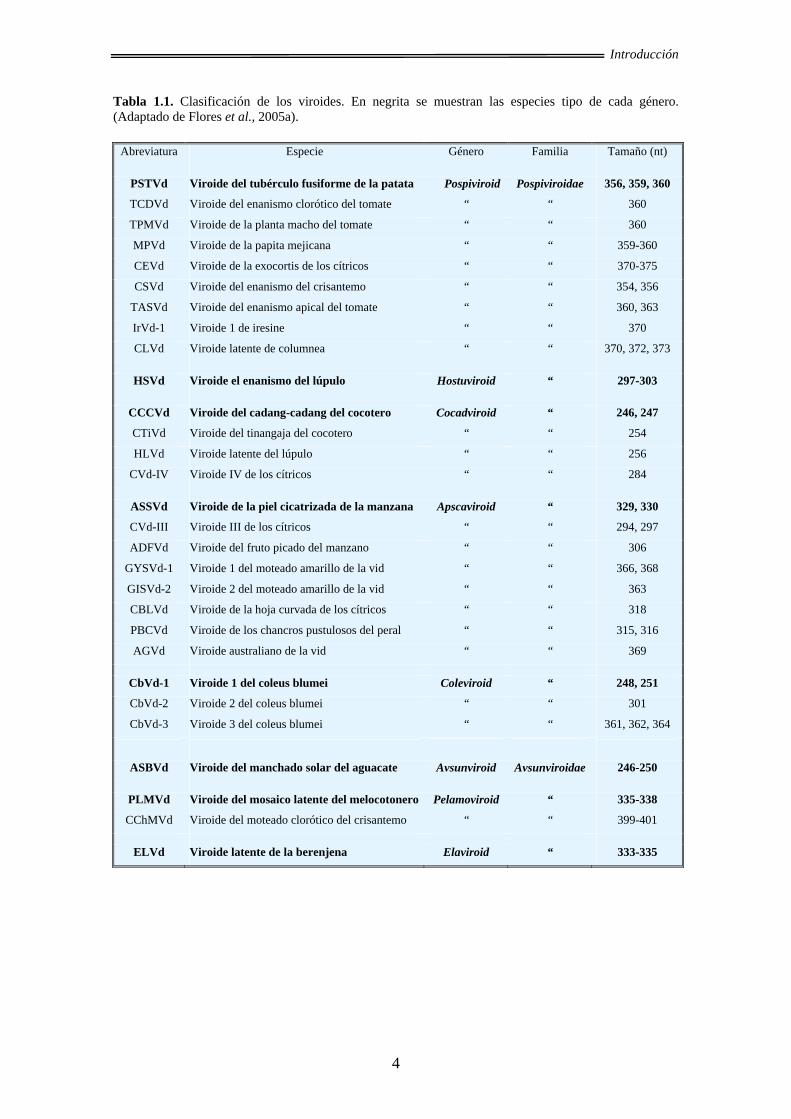
Tabla 1.1. Clasificación de los viroides. En negrita se muestran las especies tipo de cada género. (Adaptado de Flores et al., 2005a).
Abreviatura Especie Género Familia Tamaño (nt)
PSTVd Viroide del tubérculo fusiforme de la patata Pospiviroid Pospiviroidae 356, 359, 360
TCDVd Viroide del enanismo clorótico del tomate “ “ 360
TPMVd Viroide de la planta macho del tomate “ “ 360
MPVd Viroide de la papita mejicana “ “ 359-360
CEVd Viroide de la exocortis de los cítricos “ “ 370-375
CSVd Viroide del enanismo del crisantemo “ “ 354, 356
TASVd Viroide del enanismo apical del tomate “ “ 360, 363
IrVd-1 Viroide 1 de iresine “ “ 370
CLVd Viroide latente de columnea “ “ 370, 372, 373
HSVd Viroide el enanismo del lúpulo Hostuviroid “ 297-303
CCCVd Viroide del cadang-cadang del cocotero Cocadviroid “ 246, 247
CTiVd Viroide del tinangaja del cocotero “ “ 254
HLVd Viroide latente del lúpulo “ “ 256
CVd-IV Viroide IV de los cítricos “ “ 284
ASSVd Viroide de la piel cicatrizada de la manzana Apscaviroid “ 329, 330
CVd-III Viroide III de los cítricos “ “ 294, 297
ADFVd Viroide del fruto picado del manzano “ “ 306
GYSVd-1 Viroide 1 del moteado amarillo de la vid “ “ 366, 368
GISVd-2 Viroide 2 del moteado amarillo de la vid “ “ 363
CBLVd Viroide de la hoja curvada de los cítricos “ “ 318
PBCVd Viroide de los chancros pustulosos del peral “ “ 315, 316
AGVd Viroide australiano de la vid “ “ 369
CbVd-1 Viroide 1 del coleus blumei Coleviroid “ 248, 251
CbVd-2 Viroide 2 del coleus blumei “ “ 301
CbVd-3 Viroide 3 del coleus blumei “ “ 361, 362, 364
ASBVd Viroide del manchado solar del aguacate Avsunviroid Avsunviroidae 246-250
PLMVd Viroide del mosaico latente del melocotonero Pelamoviroid “ 335-338
CChMVd Viroide del moteado clorótico del crisantemo “ “ 399-401
ELVd Viroide latente de la berenjena Elaviroid “ 333-335
4

Introducción
Los viroides de la primera familia se caracterizan por poseer una región central
conservada (central conserved region, CCR), así como por su replicación y localización
nuclear. Los miembros de la segunda familia no poseen la CCR, y se replican y
localizan en el cloroplasto. Estos últimos además contienen en sus cadenas de ambas
polaridades ribozimas de cabeza de martillo, ausentes en los miembros de la familia
Pospiviroidae. Dichas ribozimas tienen un papel central en la replicación, como se verá
más adelante.
Los viroides nucleares adoptan una estructura en varilla con cinco dominios
(Keese y Symons, 1985). Dichos dominios son conocidos como central (C), patogénico
(P), variable (V) y terminal derecho (TR) e izquierdo (TL). El dominio central contiene
la CCR, mientras que el terminal izquierdo incluye a su vez una región terminal
conservada (terminal conserved region, TCR) o una horquilla terminal conservada
(terminal conserved hairpin, TCH) (Fig. 1.1). En un principio se asignó una función a
cada dominio; por ejemplo, el dominio P se asoció con los efectos patogénicos del
PSTVd y otros viroides relacionados. Sin embargo, la situación no es tan sencilla y la
expresión de síntomas parece estar controlada por determinantes discretos situados en
los dominios TL, P, V y TR (Sano et al., 1992).
El tipo de CCR, junto con la presencia o ausencia de los motivos TCR y TCH, se
utilizan como criterios en la clasificación de los miembros de la familia Pospiviroidae
(Flores et al., 2005b). La TCH está presente en los géneros Hostuviroid y Cocadviroid,
mientras que la TCR es característica a los géneros Pospiviroid y Apscaviroid, así como
en los dos miembros de mayor tamaño del género Coleviroid.
TCH TCR CCR
TL P C V TR Fig. 1.1. Esquema de la estructura característica de un miembro de la familia Pospiviroidae. La posición de los cinco dominios C (central), P (patogénico), V (variable), y TR y TL (terminal derecho e izquierdo, respectivamente) se indica en la parte inferior. Asimismo en la parte superior se muestran las regiones conservadas CCR, TCR y TCH, cuya presencia y secuencia se emplean como criterio taxonómico. Como CCR se muestra la del género Pospiviroid, siendo esta región diferente en los otros géneros. (Adaptado de Flores et al., 2004; Daròs et al., 2006).
5

Introducción
El elevado contenido en estructura secundaria de los RNAs viroidales es
consecuencia de la gran autocomplementariedad de sus secuencias. Mediante cálculos
termodinámicos con programas informáticos se ha determinado la estructura secundaria
de mínima energía libre que adoptan estos RNAs, en la que tramos apareados
intramolecularmente se alternan con pequeños bucles desapareados adquiriendo una
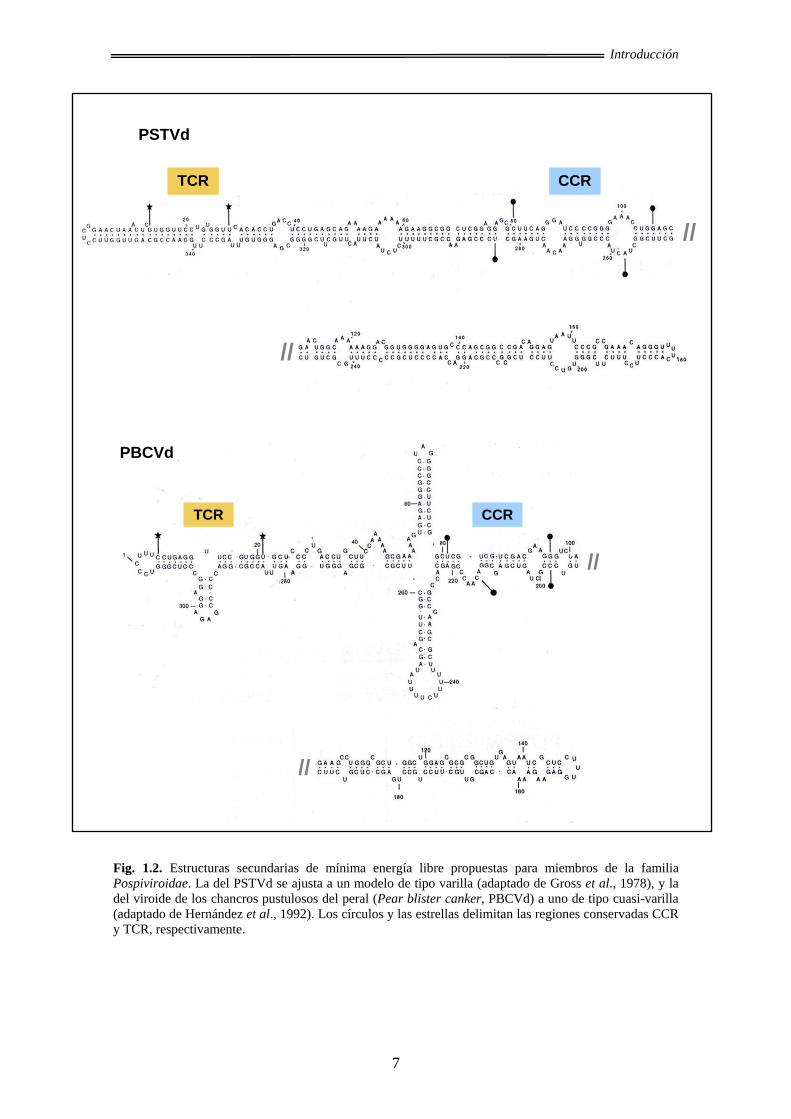
forma de varilla (Fig. 1.2). También se han descrito otras conformaciones posibles de
cuasi-varilla (Fig. 1.2) y ramificadas, estas últimas exclusivas de algunos miembros de
la familia Avsunviroidae (Fig. 1.3). Mediante microscopía electrónica se ha comprobado
que el PSTVd muestra una estructura de varilla, confirmando in vitro la predicción
termodinámica (Sogo et al., 1973; Sänger et al., 1976). Tanto la introducción de
deleciones artificiales en este viroide (Wassenegger et al., 1984) como la aparición de
repeticiones naturales en otras especies de la familia Pospiviroidae (Haseloff et al.,
1982; Semancik et al., 1994; Fadda et al., 2003b), conducen a variantes que preservan
la estructura de tipo varilla, indicando que ésta también es relevante in vivo.
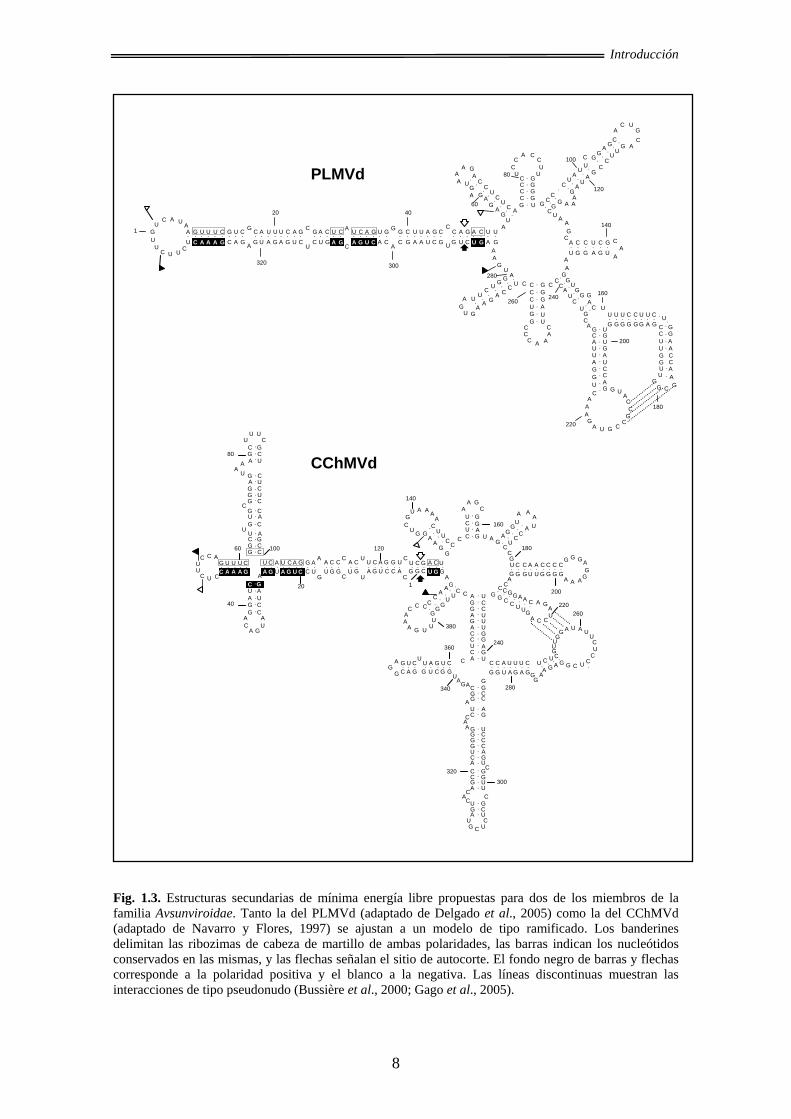
Con respecto a los miembros de la familia Avsunviroidae, el análisis de variantes
naturales de secuencia del viroide del mosaico latente del melocotonero (Peach latent
mosaic viroid, PLMVd) (Ambrós et al., 1998; Ambrós et al., 1999; Pelchat et al., 2000;
Malfitano et al., 2003 Rodio et al., 2006) y del viroide del moteado clorótico del
crisantemo (Chrysanthemum chlorotic mottle viroid, CChMVd) (Navarro y Flores,
1997; De la Peña et al., 1999; De la Peña y Flores, 2002) han permitido observar
diversas covariaciones y mutaciones compensatorias acordes con la adopción de una
estructura ramificada. Ambos RNAs viroidales son insolubles en 2 M LiCl, mientras
que los RNAs con la estructura de tipo varilla o cuasi-varilla característica de los
miembros de la familia Pospiviroidae y de otro miembro de la familia Avsunviroidae,
son solubles en estas condiciones (Navarro y Flores, 1997). Además, mediante
experimentos in vitro de cartografiado con nucleasas y de desplazamiento en gel
mediante unión de oligonucleótidos se ha detectado en el PLMVd una interacción de
tipo pseudonudo entre bucles (Bussière et al., 2000). Una interacción similar se predijo
para el CChMVd analizando las covariaciones presentes en variantes naturales del
mismo, que se confirmó in vitro con experimentos de retardo en gel e in vivo con
bioensayos de infectividad de RNAs con diversas mutaciones puntuales en dichos
bucles (Gago et al., 2005).
6
Introducción
PSTVd
TCR CCR
//
//
PBCVd
TCR CCR
//
//
Fig. 1.2. Estructuras secundarias de mínima energía libre propuestas para miembros de la familia Pospiviroidae. La del PSTVd se ajusta a un modelo de tipo varilla (adaptado de Gross et al., 1978), y la del viroide de los chancros pustulosos del peral (Pear blister canker, PBCVd) a uno de tipo cuasi-varilla (adaptado de Hernández et al., 1992). Los círculos y las estrellas delimitan las regiones conservadas CCR y TCR, respectivamente.
7
Introducción
8
Fig. 1.3. Estructuras secundarias de mínima energía libre propuestas para dos de los miembros de la familia Avsunviroidae. Tanto la del PLMVd (adaptado de Delgado et al., 2005) como la del CChMVd (adaptado de Navarro y Flores, 1997) se ajustan a un modelo de tipo ramificado. Los banderines delimitan las ribozimas de cabeza de martillo de ambas polaridades, las barras indican los nucleótidos conservados en las mismas, y las flechas señalan el sitio de autocorte. El fondo negro de barras y flechas corresponde a la polaridad positiva y el blanco a la negativa. Las líneas discontinuas muestran las interacciones de tipo pseudonudo (Bussière et al., 2000; Gago et al., 2005).
C
G
G C U G A
120
C. U
U. C. A G G U. C
U. C. G. A. UG
. . . . .
GA
160
180
200
240
380
GGC
ACCUGAU
G
C
C
U A G
CGU. C C. A. A. C. C C. C .
G G G A G
GAA A G GGGUUGGG
AC
G
GGCC
AG
UCCCAGU
CGGUUC
GCUC
UCGU
AGUC
ACA
GCCACUGGGGA
ACC
UA G
GCAGA
UGGCUGGACG
GA U
G. U. C. U A. G U. C. C
CCG.
AGGUC . .
..
. .
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
. .
1
UCCUUUGGAGUA
CUCCAGAGGA .
.
.
.
.
.
.
.
.
.
.
C. C. G. G. A. A. C. GUUCCGUU
GG
GU
UUGA
AA
C C CCC
AA
AG. G
UA A
AUA
CC
UC
. .
. .
CUCU
AA G
GGAG
.
.
.
. AGGU
CG
U A A AA
CU. U
. CC.
. .
140
60 100
U C C A
U . U U. C . U . C . A . U . C . A G. G . A . A
A. C. C. C
A. . . .
U
G G C U G U G
C G .
20
UC
GGUG
U C U G AA
G AAAC C U C
U
40
A U C C
A U
G A C A
G G A U C .
.
.
.
.
80
UA
A A
G G A G
G C
U U U
C
C
C
C C
U C C A
A G
G C U
U C
.
.
.
.
.
.
.
.
.
.
.
. .
.
320
340
360
280
C C A U U U CGAGAUGG. . . . . . . .
300
U C.
U. GA
AG
G
U
A G
CA
A. C
GUUG
G A A U U C U
C C U C G G
260
.
CU
A 220
G
CChMVd
G A
U . G
U . A
C . G
U C . G
G C A U A U
.
.
.
. U
G C G C
A
. .
U U A
U A .
C
C
U G C . G C . G C . G
C C . G C . G C . G U A
U G G . U
C C
C A A A C
C C
G . C
G . A
U G .
C G. G
A . U C .
G C A U G
C G A U U G U A A U
U . G
U . G
U . G
C . G
C . G
U
A A
A . U
C . G
C . G
U . A
C . G
G . U
C
A A
C G
A A U
G C
G C
U
A A G C . A
G C A
A U
U A
U . G
C G C
G C
A U
G U
G C A
A
C
C
U G
. . .
. .
. .
. .
. .
.
. .
.
.
.
.
.
.
.
.
.
240
.
100
120
140
160
200
260
C G . C G .
. U
180
U A . G
G C G
.
. G C
220
U A
G C
A A A G
C
A U G C C
U . C U
A G
A
A A
U G
G A
U .
G G . U
A U
A
U A
G C
A G
U G
G C . A
U . G C. G
U C A
A A
A G
A
.
. .
. .
C
60
. G C U U
C U C C
A C
80
280
.
G A .
G .
U . A
C . G
U
A . . C
G U
C U . A . G . C . A U C G
C . G
U . A
U . A
G . C
A
G G . C
40
300
G . U . U . U . C . A . U C A A
U . A
U
C
C
A A . U
G . C
C. A.
G U
G . C
A A C U
C
G U
C U U
U U
G . C
U . A
C . G A
G C . G U
A . U . A
U . G
C . G
U . A
C . G
U . A
C . A . G U
G . C
1
20
320
PLMVd

Introducción
1.1.3. Síntomas causados por los viroides
Las infecciones viroidales inducen síntomas muy parecidos a los provocados por
las virales. Las alteraciones más frecuentes causadas por los viroides incluyen
enanismo, epinastia, malformaciones, clorosis y necrosis en hojas, distorsiones y
decoloraciones en frutos, agrietamientos del tallo y la corteza y, en algunos casos, la
muerte de la planta. A diferencia de lo que ocurre con la mayoría de los virus, la
acumulación de los viroides y la expresión de los síntomas que inducen se ve favorecida
por elevadas temperaturas (30-33ºC) (Sänger y Ramm, 1975) y altas intensidades
lumínicas (Harris y Browning, 1980). Esta peculiaridad explica que la mayoría de
enfermedades viroidales se presente principalmente en zonas tropicales y subtropicales,
en climas continentales con veranos cálidos, y en cultivos de invernadero.
Mediante microscopía electrónica se han observado alteraciones citológicas
inducidas por viroides consistentes en malformaciones de la pared celular y
acumulación de depósitos electrodensos (Lawson y Hearon, 1971; Semancik y
Vanderwoude, 1976; Wahn et al., 1980), y en modificaciones de la estructura de los
cloroplastos (Hari 1980; Desjardins, 1987; Rodio et al., 2007). También se han
observado alteraciones bioquímicas en los niveles de acumulación de varias proteínas
del tipo PR (pathogenesis related), de ciertos metabolitos (poliaminas) (Conejero y
Semancik, 1977; Conejero et al., 1979) y de fitohormonas (etileno) (Bellés et al., 1989).
1.1.4. Localización subcelular
Esta cuestión se estudió inicialmente mediante centrifugación diferencial,
observándose que el PSTVd y otros viroides similares se acumulan en el núcleo
(Diener, 1971; Sänger, 1972; Semancik et al., 1976). Por lo que respecta a los miembros
de la familia Avsunviroidae, mediante esta misma técnica se observó que el ASBVd
muestra una acumulación cloroplástica (Mohamed y Thomas, 1980). Estudios
posteriores de hibridación in situ con sondas fluorescentes y microscopía láser confocal
de preparaciones de núcleos purificados y de secciones finas permitieron localizar en el
núcleo al PSTVd, al viroide del cadang-cadang del cocotero (Coconut cadang-cadang
viroid, CCCVd) y al viroide de la exocortis de los cítricos (Citrus exocortis viroid,
CEVd) (Harders et al., 1989; Bonfiglioli et al., 1996), mostrando el PSTVd y el CCCVd
una localización predominantemente nucleolar. Técnicas similares de hibridación in situ
y microscopía electrónica de transmisión han permitido localizar a dos miembros de la
9

Introducción
familia Avsunviroidae, ASBVd y PLMVd, en el cloroplasto (Lima et al., 1994; Bussière
et al., 1999), y más específicamente, en las membranas tilacoidales en el caso del
ASBVd (Bonfiglioli et al., 1994).
1.1.5. Replicación de los viroides
Los experimentos destinados a detectar un DNA homólogo del PSTVd o del
CEVd en plantas infectadas dieron resultados negativos (Branch y Dickson, 1980;
Zaitlin et al., 1980), mientras que sí se han identificado presumibles intermediarios
replicativos de RNA (Grill y Semancik, 1978). RNAs similares también se han
indentificado en viroides cloroplásticos como el ASBVd y PLMVd (Daròs et al., 1994;
Bussière et al., 1999 Delgado et al., 2005). De estos resultados se infiere que la
replicación de los viroides ocurre a través de un mecanismo con sólo intermediarios de
RNA.
La detección de RNAs multiméricos de polaridad negativa en plantas infectadas
por el PSTVd, junto con la naturaleza circular del molde inicial, condujo a proponer un
mecanismo de círculo rodante con dos variantes para la replicación de los viroides de
las dos familias (Branch y Robertson, 1984; Ishikawa et al., 1984; Hutchins et al.,
1985). En la variante asimétrica que siguen los miembros de la familia Pospiviroidae, el
RNA monomérico circular más abundante [al que se asigna arbitrariamente la polaridad
positiva (+)], actúa como molde para la síntesis de oligómeros de polaridad
complementaria, que a su vez sirven como molde para una segunda trascripción RNA-
RNA. Los multímeros de polaridad (+) resultantes de esta segunda trascripción son
cortados en fragmentos de longitud unitaria por una RNasa, para finalmente ser
circularizados por una RNA ligasa. Los miembros de la familia Avsunviroidae siguen la
variante simétrica, donde los multímeros de polaridad (-) resultantes de la trascripción
del RNA circular monomérico de polaridad (+) son procesados a formas circulares
monoméricas que sirven de molde para la segunda parte del ciclo de replicación,
simétrica a la primera. (Fig. 1.4). Una diferencia sustancial en la familia Avsunviroidae
es que el corte necesario para generar las formas de longitud unitaria ocurre
autocatalíticamente por la acción de las ribozimas de cabeza de martillo que pueden
formar los RNAs de ambas polaridades.
El soporte experimental que sostiene estas dos variantes proviene de estudios
encaminados a detectar el monómero circular de polaridad (-) en plantas infectadas.
10

Introducción
Estos estudios proporcionaron resultados negativos con el PSTVd, ya que de los
complejos replicativos sólo se recuperó el monómero circular (+) (Branch et al., 1988).
Por el contrario, los monómeros circulares de ambas polaridades se han detectado en los
casos del ASBVd y del PLMVd (Hutchins et al., 1985; Daròs et al., 1994; Bussière et
al., 1999; Navarro et al., 1999; Delgado et al., 2005). Estas observaciones, junto con la
capacidad de los RNAs de las dos polaridades de los cuatro miembros de la familia
Avsunviroidae de autocortarse in vitro a través de ribozimas de cabeza de martillo
(Hutchins et al., 1986; Hernández y Flores, 1992; Daròs et al., 1994; Navarro y Flores,
1997; Fadda et al., 2003a), sostienen que la variante que opera en esta familia es la
simétrica.
1.1.6. Enzimas y ribozimas implicadas en el ciclo replicativo de los viroides
La replicación de los viroides requiere tres actividades catalíticas: una RNA
polimerasa para generar los multímeros de ambas polaridades, una RNasa para cortar
estos multímeros a longitud unitaria, y una RNA ligasa para circularizar las formas
monoméricas lineales.
11
Fig. 1.4. Replicación de los viroides. Un mecanismo de círculo rodante de RNA con dos variantes distintas, asimétrica y simétrica, que siguen los miembros de las familias Pospiviroidae y Avsunviroidae respectivamente, permite explicar su replicación. Las líneas rojas y azules indican los RNAs de polaridad (+) y (-), respectivamente, y los triángulos los sitios de corte de las ribozimas de cabeza de martillo (Rz, en azul), y de factores del hospedador (FH, en rojo). Resultados recientes (Gas et al., 2007; Gas et al., 2008) muestran que los extremos de los RNAs monoméricos lineales de polaridad (+) generados en la variante asimétrica son de tipo 5’-P y 3’-OH, como los que producen las RNasas de tipo III. (Adaptado de Flores et al., 2004).
Rz
5’
+
5’3’
+
2’ P 3’
5’ OH 5’ OH 2’ P 3’
_
5’
3’
Rz
5’3’5’ 3’
FH FH
3’5’ P 3’ OH
Variante asimétrica (familia Pospiviroidae)
Variante simétrica (familia Avsunviroidae)

Introducción
Aunque el sitio de replicación y acumulación de los viroides no tiene que ser
necesariamente el mismo (podrían ser sintetizados en un compartimento celular y
posteriormente ser translocados a otro), la detección de una serie de intermediarios
replicativos del PSTVd en el núcleo (Spiesmacher et al., 1983) y del ASBVd en el
cloroplasto (Navarro et al., 2000), muestran que la replicación y acumulación ocurren
en el mismo compartimento.
1.1.6.1. Iniciación y elongación
La transcripción en el núcleo está catalizada por cuatro RNA polimerasas
dependientes de DNA que, además de por el tipo de RNA generado, se diferencian por
otras características bioquímicas y en particular por su sensibilidad a la toxina fúngica
α-amanitina (Roeder, 1976; Marzluff y Huang, 1984; Cox y Golberg, 1988). Mientras
que la RNA polimerasa II es inhibida por concentraciones nanomolares de este péptido,
se requieren concentraciones mil veces superiores del mismo para inhibir la RNA
polimerasa III, siendo la RNA polimerasa I insensible a dicha toxina. Experimentos in
vivo (Mühlbach y Sänger, 1979) e in vitro (Flores y Semancik, 1982; Schindler y
Mühlbach, 1992) han revelado que la replicación del PSTVd, del CEVd y del viroide
del enanismo del lúpulo (Hop stunt viroid, HSVd), todos ellos de la familia
Pospiviroidae (Tabla 1.1), es inhibida por concentraciones nanomolares de α-amanitina.
Por otra parte, ensayos de inmunoprecipitación empleando un anticuerpo monoclonal
contra el dominio conservado carboxilo terminal de la subunidad mayor de la RNA
polimerasa II, han mostrado que esta subunidad coprecipita con las cadenas del CEVd
de ambas polaridades presentes en preparaciones nucleares de tejido infectado capaces
de elongar in vitro RNAs de este viroide (Warrilow y Symons, 1999). Así pues, ambas
líneas experimentales implican a la RNA polimerasa II en la replicación de los viroides
nucleares. La RNA polimerasa IV descrita reciente en plantas tiene un papel en el
mecanismo de silenciamiento génico mediado por RNAs y, más concretamente, en la
síntesis de pequeños RNAs nucleares (Onodera et al., 2005; Herr et al., 2005; Kanno et
al., 2005), pero hasta la fecha no hay datos acerca de su sensibilidad a la α-amanitina ni
de su implicación en la replicación de los viroides.
La transcripción en el cloroplasto está catalizada por dos RNA polimerasas
dependientes de DNA: la polimerasa codificada en el genoma nuclear (nuclear encoded
polymerase, NEP) y la polimerasa codificada en el genoma plastídico (plastid encoded
12

Introducción
polymerase, PEP), esta última homóloga a la RNA polimerasa eubacteriana. Ambas
enzimas cloroplásticas pueden diferenciarse por su sensibilidad a la tagetitoxina, que
inhibe la PEP (Steinberg et al., 1990; Mathews y Durbin, 1990) pero no la NEP (Lukens
et al., 1987; Mathews y Durbin, 1990; Liere y Maliga, 1999). En experimentos con
preparaciones cloroplásticas de aguacate infectado por ASBVd se ha comprobado que la
adición de este inhibidor en un intervalo de 1 a 50 µM provoca una fuerte inhibición de
la transcripción de los RNAs cloroplásticos, mientras que la transcripción del ASBVd
solo se ve inhibida en torno a un 25% incluso a una concentración 100 µM de la toxina.
Este resultado sugiere la participación de la NEP en la replicación del ASBVd, aunque
no puede descartarse la existencia de una tercera polimerasa cloroplástica resistente a la
tagetitoxina (Navarro et al., 2000). En experimentos recientes realizados con
melocotoneros infectados con una variante de secuencia del PLMVd que provoca una
clorosis foliar muy intensa se ha observado que la PEP no es funcional en las regiones
albinas, mientras que los RNAs transcritos por la NEP no se ven afectados. En dichas
regiones, las formas circulares y lineales de ambas polaridades del PLMVd se acumulan
incluso en mayor medida que en las regiones adyacentes verdes de la hoja, por lo que
estos resultados in vivo sostienen que la RNA polimerasa implicada en la replicación del
PLMVd es la NEP (Rodio et al., 2007)
Resulta notable que a pesar de estar descritas y caracterizadas en plantas varias
RNA polimerasas dependientes de RNA (Schiebel et al., 1998; revisado en
Wassenegger y Krczal, 2006), todos los datos disponibles sostienen que los viroides son
reconocidos y transcritos por RNA polimerasas cuyo molde en condiciones fisiológicas
es DNA. Cómo los viroides son capaces de forzar este cambio de especificidad de
molde está por resolver, si bien observaciones recientes muestran que la RNA
polimerasa II posee una actividad dependiente de RNA que, aunque más lenta y menos
procesiva, podría ser la utilizada por los viroides nucleares para su replicación
(Lehmann, et al., 2007).
Diversos experimentos han permitido cartografiar el posible sitio de inicio de la
transcripción del RNA del PSTVd, si bien estos resultados deben interpretarse con
cautela pues se han obtenido in vitro, condiciones éstas que pueden no reproducir
fielmente lo que ocurre in vivo. En un primer lugar, el RNA monomérico circular (+)
purificado de plantas infectadas se transcribió in vitro con la RNA polimerasa II
purificada de tomate y germen de trigo (Tabler y Tsagris, 1990), y con extractos
nucleares de cultivos celulares de patata (Fels et al., 2001). Los RNAs de polaridad (-)
13

Introducción
así obtenidos se transcribieron inversamente y se amplificaron por PCR,
cartografiándose el sitio de inicio de la transcripción en tres posiciones diferentes
caracterizadas por formar parte de una horquilla. Experimentos más recientes de
extensión del cebador sobre el RNA (-) sintetizado de novo con extractos nucleares de
patata, han determinado otro sitio alternativo de inicio de la transcripción de las cadenas
de polaridad (-) del PSTVd en el bucle terminal izquierdo (Kolonko et al., 2006). En
este caso, las cadenas de polaridad (-) se purificaron por cromatografía de afinidad con
RNAs biotinilados de polaridad complementaria, evitando así la presencia de otros
RNAs contaminantes que condujeron a los resultados erróneos de los primeros
experimentos (Fels et al., 2001).
Con respecto al sitio de iniciación en la familia Avsunviroidae, conviene
destacar que el extremo 5’ de los transcritos primarios cloroplásticos, pero no el de los
RNAs resultantes de su procesamiento, tiene un grupo trifosfato libre que puede ser
específicamente marcado in vitro con [α−32P]GTP y guanililtransferasa. La aplicación
de este marcaje a los RNAs lineales monoméricos (+) y (-) del ASBVd aislados de
tejido de aguacate infectado, en combinación con ensayos de protección frente a RNasas
usando ribosondas específicas de este viroide, ha revelado que la transcripción de ambas
cadenas del ASBVd empieza con la secuencia UAAAA, que se localiza en los
respectivos bucles terminales derechos ricos en A+U de las estructuras secundarias de
cuasi-varilla predichas para los dos RNAs viroidales. Este resultado ha sido además
confirmado por experimentos de extensión de cebador (Navarro y Flores, 2000). Más
recientemente se han determinado los sitios de iniciación de la transcripción de cada una
de las cadenas del PLMVd en la proximidad del sitio de corte de la cadena de la otra
polaridad. Estos resultados se han obtenido por RT-PCR empleando la técnica de
amplificación rápida de los extremos de un cDNA mediada por RNA ligasa (RNA
ligase-mediated rapid amplification of cDNA ends, RLM-RACE), y por experimentos
de extensión del cebador (Delgado et al., 2005).
1.1.6.2. Corte
Una vez sintetizados los multímeros de ambas polaridades, bien a través de la
variante simétrica o la asimétrica, éstos deben ser cortados a formas monoméricas de
longitud unitaria, para lo que existe un mecanismo diferente en cada familia. Como ya
se ha comentado, los miembros de la familia Avsunviroidae emplean su capacidad de
14

Introducción
formar ribozimas de cabeza de martillo para cortarse autocatalíticamente y generar así el
RNA monomérico lineal. En los miembros de la familia Pospiviroidae se ha observado
que extractos nucleares de patata son capaces de procesar un RNA (+) del PSTVd de
longitud mayor que la unitaria a formas circulares monoméricas e infecciosas (Tsagris
et al., 1987). Sorprendentemente, la ribonucleasa fúngica T1 también es capaz de
catalizar in vitro las mismas reacciones de corte y ligación (Tabler et al., 1992),
sugiriendo que la especificidad de éstas debe estar determinada por conformaciones del
RNA viroidal.
Recientemente, en plantas de Arabidopsis thaliana transformadas con cDNAs
que expresan transcritos diméricos (+) de cinco especies representativas de la familia
Pospiviroidae, se ha observado el procesamiento correcto a las correspondientes formas
monoméricas circulares (+). Por el contrario, el trascrito dimérico (-) de uno de estos
viroides no fue procesado en las mismas condiciones, indicando que esta propiedad es
intrínseca de la polaridad, y que probablemente viene determinada por la adopción de
conformaciones específicas (Daròs y Flores, 2004). La observación de que en cultivos
celulares y en plantas infectadas con PSTVd las cadenas (+) y (-) se acumulan en el
nucleoplasma, mientras que las (+) se localizan en el nucleolo, sugiere que el
procesamiento de las últimas ocurre en este compartimento subnuclear (Qi y Ding,
2003), donde también se procesan los precursores de los rRNAs y tRNAs.
Diferentes resultados muestran la importancia de las ribozimas de cabeza de
martillo en el corte in vivo de los miembros de la familia Avsunviroidae. Primero, los
extremos 5’ de los RNAs monoméricos lineales del ASBVd, CChMVd y PLMVd
aislados de tejido infectado coinciden con los obtenidos en las reacciones de autocorte
in vitro de estos mismos RNAs (Marcos y Flores, 1992; Daròs et al., 1994; Navarro y
Flores, 1997; Navarro et al., 2000; Delgado et al., 2005). Segundo, los cambios
nucleotídicos detectados en las ribozimas de cabeza de martillo de diversas variantes de
secuencia de los viroides cloroplásticos no afectan a su estabilidad, bien porque se
localizan en los bucles, bien porque las mutaciones son covariaciones (Navarro y Flores,
1997; Ambrós et al., 1998; De la Peña et al., 1999; Fadda et al., 2003a). Tercero, existe
una correlación entre la infectividad de diferentes variantes del CChMVd y del PLMVd,
y su nivel de autocorte durante la transcripción in vitro (Ambrós et al., 1998; De la Peña
et al., 1999). Y cuarto, en el sitio de corte de un RNA satélite de tipo viroidal con
ribozimas de cabeza de martillo (estos RNAs son estructuralmente similares a los
viroides pero funcionalmente dependientes de un virus) se ha observado un enlace 2’-
15

Introducción
fosfomonoéster, 3’,5’-fosfodiéster (Kiberstis et al., 1985), la huella característica de la
ligación mediada por una tRNA ligasa, que emplea los mismos extremos que los
generados por el autocorte mediado por estas ribozimas (ver más adelante).
1.1.6.3. Ligación
No existe consenso acerca del mecanismo de ligación de los RNAs
monoméricos viroidales una vez han sido cortados, pues aunque se ha observado que la
RNasa T1 fúngica cataliza in vitro el corte y la ligación correcta de un RNA (+) del
PSTVd de longitud mayor que la unitaria (Tabler et al., 1992), es más probable que los
miembros de la familia Pospirivoidae sean circularizados por una RNA ligasa. De
forma paralela, los miembros de la familia Avsunviroidae también podrían ser ligados
por una enzima de esta clase, si bien no puede descartarse una ligación mediada por las
ribozimas de cabeza de martillo, o incluso una autoligación que genera enlaces atípicos
(ver más adelante).
Ensayos in vitro realizados con la tRNA ligasa de germen de trigo muestran que
esta enzima cataliza la circularización de las formas lineales monoméricas de PSTVd
aisladas de tejido infectado (Branch et al., 1982). Un resultado similar se obtuvo con
extractos de Chlamydomonas reinhardtii, en los que se detectó una actividad capaz de
catalizar la ligación tanto de los fragmentos resultantes del corte de los pre-tRNAs como
de las formas lineales monoméricas del PSTVd aisladas de tejido infectado. Esta
ligación ocurre mediante la formación de un enlace 3’,5’-fosfodiéster y de un grupo 2’-
fosfomonoéster (Kikuchi et al., 1982). Como ya se ha comentado, este grupo
característico de la ligación mediada por las tRNA ligasas también se ha detectado en el
sitio de ligación de un RNA satélite de tipo viroidal que se autocorta por ribozimas de
cabeza de martillo (Kiberstis et al., 1985). Además, experimentos realizados con A.
thaliana transformada con cDNAs diméricos de cinco especies representativas de la
familia Pospiviroidae muestran que esta planta modelo posee una enzima capaz de
catalizar in vivo la circularización de las formas monoméricas lineales de estos viroides
(Daròs y Flores, 2004). Estos datos sugieren que en la familia Pospiviroidae una tRNA
ligasa nuclear del hospedador media la circularización de los RNAs monoméricos
lineales procedentes del corte de los intermediarios multiméricos de la replicación.
16

Introdución
Mientras que ciertas ribozimas como la de tipo horquilla catalizan el autocorte y
la ligación generando un enlace 3’,5’-fosfodiéster en ciertos RNAs satélites de tipo
viroidal (Hampel y Tritz, 1989), no hay pruebas de que ocurra lo mismo con las
ribozimas de cabeza de martillo en la familia Avsunviroidae. En relación con estas
últimas ribozimas, las formas monoméricas lineales del RNA satélite del virus de las
manchas anulares del tabaco (satellite RNA of tobacco ringspot virus, sTRSV) pueden
formar dímeros en experimentos in vitro en ausencia de proteínas (Prody et al., 1986).
También se ha observado ligación in vitro en ribozimas de cabeza de martillo mínimas
(formadas únicamente por los nucleótidos que componen el centro activo y las hélices
que las flanquean) (Hertel et al., 1994), si bien en ambos casos el equilibrio está
desplazado unas 100 veces hacia el corte, de modo que menos del 1% del sustrato es
ligado. Más recientemente se ha caracterizado la ligación en ribozimas de cabeza de
martillo extendidas (que incluyen las interacciones terciaria entre bucles periféricos al
centro activo que son clave para la actividad catalítica) del sTRSV y de un transcrito del
gusano platelminto parásito Schistosoma mansoni. Los correspondientes resultados
muestran un aumento significativo de la ligación en ambos casos, hasta alcanzar valores
del 6 al 23% del RNA presente en la reacción (Nelson et al., 2005; Canny et al., 2007).
Esta situación no es en principio extrapolable a RNAs viroidales completos que, tras
autocortarse, pueden plegarse adquiriendo una conformación de mínima energía libre
que impide la formación de las ribozimas (Forster y Symons, 1987). Sin embargo es
posible que in vivo una chaperona ayude al RNA a adquirir el plegamiento adecuado
para que ocurra la ligación ribozimática.
Por otra parte se ha observado que en presencia de una alta concentración de
Mg+2 (100 mM), el RNA monomérico lineal del PLMVd resultante de autocorte es
capaz de autoligarse in vitro formando un enlace 2’,5’-fosfodiéster (Côté y Perreault,
1997), proponiéndose la existencia in vivo de moléculas circulares de este viroide con
dicho enlace (Côté et al., 2001). Sin embargo, no se conocen otros RNAs naturales con
enlaces 2’,5’-fosfodiéster que sirvan como moldes de trascripción, y además dicha
autoligación no parece ser ribozimática sino dirigida por la conformación de energía
mínima que adquiere el PLMVd tras su autocorte, ya que una reacción similar se ha
descrito incluso para el PSTVd que no contiene ribozimas de cabeza de martillo
(Baumstark et al., 1997).
Finalmente conviene resumir lo que se conoce respecto a la ligación del RNA
del virus delta de la hepatitis humana (hepatitis delta virus, HDV) (Sharmeen et al.,
17

Introdución
1989). Este es un virus defectivo que presenta un genoma de RNA circular
monocatenario como el de los viroides, y se replica por un mecanismo de círculo
rodante mediado por ribozimas de una clase especial. La autoligación del RNA del
HDV resultante de autocorte ocurre in vitro en ausencia (Wu y Lai, 1989) o en
presencia de un RNA complementario generando un enlace 2’,5’-fosfodiéster
(Sharmeen et al., 1989), aunque en condiciones lejos de las fisiológicas. También se ha
descrito que si bien el Mg+2 es necesario para el autocorte, la autoligación de dicho
RNA ocurre en ausencia de este catión (Wu y Lai, 1989), si bien otros resultados
indican que el Mg+2 es necesario para ambos procesos (Rosenstein y Been, 1990).
Además, la eficiencia de la autoligación es muy baja, y trabajos más recientes sugieren
que la ligación podría estar catalizada por la tRNA ligasa nuclear y generar un enlace
3’,5’-fosfodiéster (Lazinski y Taylor, 1995; Reid y Lazinski, 2000).
1.2. Las RNA ligasas
1.2.1 Características generales y mecanismos de ligación
Desde que se observó por primera vez una actividad de ligación del RNA en
células de E. coli infectadas por el bacteriófago T4 (Silber et al., 1972), se han
descubierto enzimas que catalizan esta reacción de formación de enlaces fosfodiéster en
organismos de los tres dominios filogenéticos: Bacteria, Arquea y Eucaria (revisado en
Shuman y Lima, 2004). Las RNA ligasas se clasifican en seis grupos (revisado en Arn y
Abelson, 1998): i) las del fago T4, una de las cuales repara las roturas que genera E. coli
en sus tRNAs como defensa frente a la infección por este virus, mientras que la otra
repara roturas del RNA de forma más genérica, ii) las que median la edición del mRNA
en kinetoplástidos, iii) la RNA ligasa de arqueas, que interviene en diversas reacciones
de procesamiento del RNA, iv) la tRNA ligasa de eucariotas que cataliza la ligación de
los precursores de los tRNAs una vez se ha eliminado el intrón del anticodón, v) una
RNA ligasa exclusiva de metazoos también implicada en la maduración de los pre-
tRNAs, y vi) la 2’,5’-RNA ligasa de bacterias y arqueas, la única que no requiere
ningún cofactor nucleotídico y de la que se desconoce su papel biológico.
Las reacciones catalizadas por estas enzimas difieren en función de los
requerimientos de NTPs, de los extremos ligados, y de la procedencia del grupo
fosforilo que participa en la génesis del enlace. De esta forma pueden diferenciarse
cuatro mecanismos diferentes catalizados por: i) las T4 RNA ligasas y RNA ligasas de
18

Introdución
kinetoplástidos, ii) las tRNA ligasas, iii) las RNA ligasas de animales y arqueas, y iv)
las 2’,5’-RNA ligasas (Fig 1.5).
T4 RNA ligasas/RNA ligasa de kinetoplástidos
Fig. 1.5. Comparación de los diferentes mecanismos de acción de las RNA ligasas. Ap, adenosina monofosfato; App, adenosina difosfato; Appp, adenosina trifosfato; ppi, pirofosfato; Gppp, guanosina trifosfato; Ø, no emplea Nppp. El grupo fosforilo que se incorpora en el enlace fosfoéster se marca con un asterisco. (Adaptado de Arn y Abelson, 1998).
Los dos primeros mecanismos son muy parecidos (Fig. 1.5), aunque la tRNA
ligasa cataliza una serie de pasos que en la ruta en la que participa la T4.1 RNA ligasa
son mediados previamente por la enzima T4 polinucleótido quinasa (polynucleotide
kinase, PNK) (Richardson 1965; Novogrodsky y Hurwitz, 1966a; Novogrodsky y
Ø Ø
2’
OH 3’ 5’
OH
P* APPP
5’3’
2’OH
AP + P*
tRNA ligasa
2’
RNA ligasa de animales/arqueas
3’ 5’
P*
HO
5’3’
2’OH
P*
APPP
Ø
? Ø
P* 3’
2’
5’
OH
HO
GPPP ?
Animales
Arqueas (Euryarqueotas)
Arqueas (Crenarqueotas)
2’-5’ RNA ligasa
3’
2’
5’
P*
HO OH3’
2’
5’
P*
APP AP +
APPP* APPP
3’
2’
5’
P
HO 5’3’
2’P
+ +
P*
19

Introdución
Hurwitz 1966b; Cameron y Uhlenbeck, 1977; Wang y Shuman, 2002; Wang et al.,
2002). En ambas reacciones el grupo fosforilo que forma parte del enlace generado
proviene de la fosforilación del extremo 5’-hidroxilo. Sin embargo, en el caso del fago
T4 es la PNK la que cataliza esta fosforilación, mientras que en eucariotas es la tRNA
ligasa, que contiene un dominio polinucleótido quinasa (ver más adelante), la que
cataliza la fosforilación del RNA antes de su ligación. La intervención de la PNK no
sería necesaria en kinetoplástidos, ya que los resultados sugieren que la endonucleasa
que media el corte del RNA genera directamente los extremos 3’-hidroxilo y 5’-
fosfomonoéster necesarios para la ligación (Sabatini y Hajduk, 1995).
Las RNA ligasas de T4 y de kinetoplástidos son las únicas que requieren
extremos 3’-hidroxilo 5’-fosfomonoéster, mientras que el resto catalizan la ligación de
extremos 2’,3’-fosfodiéster cíclico y 5’-hidroxilo. El mecanismo de las RNA ligasas de
animales y arqueas está menos estudiado que el de las T4 RNA ligasas y tRNA ligasas,
y difiere fundamentalmente en que el grupo fosforilo del nuevo enlace generado
proviene del extremo 3’ y no de la fosforilación de novo del extremo 5’. La reacción
catalizada por la 2’,5’-RNA ligasa ocurre en ausencia de cofactor nucleotídico, y al
igual que en el caso de las RNA ligasas de animales y arqueas, el grupo fosforilo del
nuevo enlace fosfodiéster proviene del extremo 2’,3’-fosfodiéster cíclico (revisado en
Arn y Abelson, 1998).
Las RNA ligasas dependientes de ATP, las DNA ligasas dependientes de ATP
(presentes en los tres dominios filogenéticos) y de NAD+ (detectadas en bacterias y
entomopoxvirus), y las transferasas dependientes de GTP que añaden la estructura de
caperuza al mRNA, componen la superfamilia de nucleotidil transferasas covalentes
(revisado en Shuman y Lima, 2004). Todas catalizan la transferencia de una adenina
monofosforilada al extremo 5’ de un ácido nucleico mediante la formación de un
intermediario enzima-(lisil-N)-NMP (fosforamidato, Fig. 1.6), en un paso conocido
como adenilación. La reacción de ligación de las mismas consiste en tres pasos: i)
adenilación de la enzima mediante la incorporación de un AMP a partir de un ATP y
liberación de un grupo pirofosfato, ii) transferencia del AMP al extremo 5’-
fosfomonoéster del RNA (extremo donador), y iii) ligación de la molécula mediante el
ataque del extremo 3’-hidroxilo (extremo aceptor) sobre el 5’-adenilado (extremo
donador), generando un enlace fosfodiéster con la consiguiente liberación del AMP
(Fig. 1.6A). Este mecanismo es compartido por los otros miembros de las superfamilia
de nucleotidil transferasas covalentes, con la particularidad de que el paso final de
20

Introdución
ligación no ocurre en las enzimas que catalizan la adición de la estructura de caperuza
(Fig. 1.6).
A) E + PPPA E-PA + PPi
E-PA + PRNA E · APPRNA RNAOH + E · APPRNA RNAPRNA + PA + E
B) E + PPPA E-PA + PPi
E-PA + PDNA E · APPDNA DNAOH + E · APPDNA DNAPDNA + PA + E
C) E + PPPG E-PG + PPi
E-PG + PPRNA GPPPRNA + E
Lys – N + – P – O – (Nucleósido)
O
O –
H––
H
–=
Fosforamidato
Lys – N + – P – O – (Nucleósido)
O
O –
H––
H
–=
Fosforamidato
Fig. 1.6. Mecanismo conservado de adenilación (ó guanilación) y transferencia nucleotídica a los extremos 5’ de los ácidos nucleicos. A) RNA ligasa, B) DNA ligasa, C) transferasa de GMP. El primer paso implica el ataque de la enzima (E) al grupo fosforilo α (P) del cofactor nucleotídico para formar un intermediario fosforamidato entre el NMP y la enzima (E-PA o E-PG). El segundo paso consiste en la transferencia del NMP (Ap o Gp) al extremo 5’-fosforilado del ácido nucleico. Finalmente, el extremo 3’-hidroxilo (rosa) ataca al extremo 5’-adenilado (azul) generando el enlace y liberando AMP (Ap). La estructura del intermediario fosforamidato se muestra en la parte inferior de la figura. (Adaptado de Shuman y Lima, 2004).
Esta superfamilia está definida por cinco motivos peptídicos que forman el
bolsillo de unión del nucleótido monofosforilado y proporcionan las cadenas laterales
necesarias para la catálisis. El más importante de los cinco motivos es el I, formado por
la secuencia KXXG (la segunda X es habitualmente D o G), conservada en todos los
miembros de la superfamilia. La lisina tiene un papel fundamental, pues incorpora el
nucleótido monofosforilado durante la adenilación. Los otro motivos, III, IIIa, IV y V
también desempeñan una función en la catálisis, como han puesto de manifiesto
diferentes análisis mutacionales (Sawaya et al., 2003; Wang et al., 2003; Yin et al.,
21

Introdución
2003; Wang y Shuman, 2005; Wang et al., 2006b). El estudio de las estructuras
cristalográficas de varios miembros de las tres familias (DNA y RNA ligasas y
transferasas de GMP) (Odell et al., 2000; Fabrega et al., 2003; Nandakumar et al.,
2006; El Omari et al., 2006), conduce a considerar su evolución a partir de un ancestro
común que posiblemente estuviera implicado en la reparación del RNA. Esta evolución
conllevaría la adquisición de dominios que otorgaran especificidad por DNA o RNA (y
dentro de este último, por mRNA o tRNA), y posteriormente la diferenciación del
dominio de transferencia del residuo nucleotídico para unir ATP, GTP y NAD+
(revisado en Shuman y Lima, 2004).
1.2.2. RNA ligasas víricas
1.2.2.1. T4.1 RNA ligasa
La primera de las RNA ligasas descritas, la T4.1 RNA ligasa (antes denominada
T4 RNA ligasa simplemente), fue descubierta por su capacidad para catalizar la unión
de los extremos 3’-hidroxilo y 5’-fosfomonoéster de cadenas artificiales de poli(A)
(Silber et al., 1972). Esta reacción es dependiente de Mg2+ y ATP, que se rompe
liberando AMP y pirofosfato en cantidades equimoleculares a los enlaces formados
(Sugino et al., 1977). La T4.1 RNA ligasa es una enzima con escasa especificidad de
sustrato, siendo incluso capaz de catalizar la unión entre RNA y DNA sin que se afecte
demasiado la velocidad de reacción, siempre que el DNA sea el sustrato donador
(Snopek et al., 1976; McCoy y Gumport, 1980). Sin embargo, los requerimientos para
el sustrato aceptor son mayores, catalizando la ligación de los aceptores
desoxirriboadenilados doscientas veces más lentamente que los riboadenilados, lo que
proporciona a la enzima su especificidad por el RNA. En cuanto al cofactor
nucleotídico, la enzima muestra una especificidad muy alta por NTPs frente a dNTPs, y
más concretamente por el ATP (Cranston et al., 1974). Esta RNA ligasa se ha detectado
en los fagos de la serie T-par, pero no en los de la serie T-impar ni en el fago λ (Silber et
al., 1972).
Mediante cromatografía de afinidad de ácidos nucleicos pudo aislarse una
proteína de 43 kDa asociada con la actividad T4.1 RNA ligasa (Snopek et al., 1976). El
gen codificante de esta enzima (el 63 del genoma de T4) fue identificado cartografiando
mutaciones que reducían dicha actividad (Snopek et al., 1977). La proteína de 375
aminoácidos codificada por este gen también desempeña un papel en la unión de las
22

Introdución
fibras de la cola al cuerpo del fago (Snopek et al., 1977). Ambas funciones se localizan
en dos sitios distintos, pues mutaciones que inactivan la actividad RNA ligasa no
afectan a la unión de las fibras de la cola al cuerpo del fago (Runnels et al., 1982).
Los análisis de secuencia y cristalográfico de esta enzima han puesto de
manifiesto la presencia de los cinco motivos peptídicos que caracterizan a la
superfamilia de las nucleotidil transferasas covalentes y de un sitio activo con dos iones
Mg+2 (El Omari et al., 2006). Mutaciones puntuales que afectan a diversos aminoácidos
de estos motivos conllevan una disminución o la anulación de la actividad catalítica
(Wang et al., 2003; Wang et al., 2006b).
La función biológica de dicha enzima fue estudiada en una cepa de E. coli
(CTr5x) resistente a la infección por cepas de T4 carentes de actividad PNK o RNA
ligasa. El fago expresa un péptido denominado Stp que inactiva el complejo de
modificación-restricción de la bacteria. Por su parte, la cepa CTr5x de E. coli posee la
enzima PrrC, que cataliza el corte del anticodón del tRNALys (anticodón nucleasa,
ACNasa), y que es inactiva en condiciones normales (Amitsur et al., 1992). Como
consecuencia de la infección por T4, PrrC se activa al alterarse el complejo de
modificación-restricción de la bacteria en presencia de la proteína Stp vírica, y media el
corte del tRNALys previniendo la síntesis de proteínas del invasor y del hospedador. Sin
embargo, esta ACNasa genera extremos 2’,3’-fosfodiéster cíclico y 5’-hidroxilo, que las
cepas víricas silvestres pueden transformar en extremos 3’-hidroxilo y 5’-fosmonoéster
por mediación de la T4 PNK, para que posteriormente la T4.1 RNA ligasa restaure el
tRNALys cortado. Este mecanismo regenera tRNAs funcionales que permiten progresar
a la infección vírica (Amitsur et al., 1987), por lo que el mecanismo sólo ha podido
estudiarse en E. coli infectada por cepas de T4 deficientes en la T4 PNK o la T4.1 RNA
ligasa. Sin embargo, en Neisseria se ha detectado un homólogo de PrrC, así como otras
ACNasas secretables en enterobacterias (colicina E5 y D) y hongos (zymocina)
(Kaufmann, 2000; Lu et al., 2005), que hacen pensar que se trata de un mecanismo de
defensa contra invasores bastante extendido y que, por añadidura, los mecanismos de
reparación del RNA mediados por RNA ligasas también deben ser frecuentes.
1.2.2.2. T4.2 RNA ligasa
Recientemente se ha descubierto una segunda RNA ligasa en T4 (T4.2) (Ho y
Shuman, 2002) con baja similitud de secuencia con la otra RNA ligasa de este fago
23

Introdución
(T4.1). Ambas enzimas sólo se asemejan por contener los motivos nucleotidil
tranferasa, guardando la T4.2 una mayor similitud con las ligasas que editan el RNA en
tripanosomas, con algunas RNA ligasas de arqueas, y con las RNA ligasas de
entomopoxvirus. La T4.2, que cataliza la unión de los mismos extremos que la T4.1 y
también requiere ATP y Mg+2 (o Mn+2), está codificada en el gen 24.1 del genoma de
T4 y formada por 334 aminoácidos (Ho y Shuman 2002). Al igual que en la T4.1, el
sustrato aceptor determina la especificidad de la enzima por el RNA, siendo esta
especificidad mayor en la T4.2 (Nandakumar y Shuman, 2004). Mediante experimentos
de deleción se ha comprobado que mientras que el reconocimiento del extremo 3’-
hidroxilo y el ataque sobre el RNA adenilado es mediado por aminoácidos del extremo
amino terminal en la T4.2 RNA ligasa, la actividad de transferencia del AMP
incorporado a la enzima al extremo 5’-fosfomonoéster del RNA reside en el extremo
carboxilo terminal (Ho et al., 2004). Esta enzima, además de catalizar la ligación entre
RNAs monocatenarios, sella roturas en RNAs de doble cadena y en híbridos
RNA:DNA, sugiriendo que podría mediar un mecanismo de reparación ancestral de
ácidos nucleicos (Nandakumar et al., 2004; Ho et al., 2004).
Un homólogo de la T4.2 RNA ligasa se ha encontrado en el fago KV40, donde
la enzima se encuentra predominantemente adenilada. La incubación de un RNA con
extremos 5’-fosfomonoéster y 3’-hidroxilo con una preparación purificada de esta
enzima adenilada genera moléculas circulares en ausencia de ATP. Al añadir ATP a la
reacción, el complejo RNA-proteína se desensambla sin llevar a cabo el último paso de
ligación, generando mayoritariamente RNA adenilado en vez de circularizado, de forma
similar a lo que ocurre con las trasferasas de GMP durante la adición de la estructura de
caperuza al mRNA. Como ya se ha comentado, estas últimas y las RNA ligasas
dependientes de ATP comparten un ancestro común, por lo que se presume que la
reacción catalizada por la enzima del fago KV40 sería un paso evolutivo intermedio
entre ambas funciones (Yin et al., 2004).
1.2.2.3. Otras RNA ligasas víricas
En baculovirus se ha descubierto recientemente un nuevo tipo de RNA ligasa
similar a la T4.1 que presenta una característica única entre las RNA ligasas no
eucarióticas: la presencia de las actividades polinucleótido quinasa y RNA ligasa en una
misma proteína y con la misma orientación (amino terminal-ligasa-quinasa-
24

Introdución
ciclofosfodiesterasa-carboxilo terminal) observada en las tRNA ligasas de levaduras y
plantas (ver más adelante) (Martins y Shuman, 2004a). Esta enzima proviene
posiblemente de la fusión de las enzimas T4 PNK y T4.1 RNA ligasa que en el fago T4
residen en proteínas distintas, y podría ser el antecesor de las tRNA ligasas presentes en
todos los eucariotas.
1.2.3. RNA ligasas de bacterias y arqueas
1.2.3.1. 2’,5’-RNA ligasa
Durante la maduración de los tRNAs de arqueas, levaduras y eucariotas
superiores, el pre-tRNA sufre la excisión de su intrón, generando dos mitades de RNA
(además del intrón escindido), que posteriormente son ligadas para formar el tRNA
maduro. En 1983 se detectó en diferentes bacterias, entre ellas E. coli, una actividad
capaz de mediar la ligación de las dos mitades de los tRNAs de la levadura común
(Saccharomyces cerevisiae) obtenidas por transcripción in vitro (Greer et al., 1983a).
Un análisis más exhaustivo de dicha actividad reveló que era específica para los
intermediarios del procesamiento de cuatro de los diez tRNAs de la levadura (Tyr, Lys2,
Trp y Phe), y que catalizaba más eficientemente la ligación de las dos mitades de estos
tRNAs si una vez sintetizados eran incubados con un extracto de levadura y purificados
antes de añadirlos a la reacción de ligación (Arn y Abelson, 1996). Además, la ligación
catalizada por esta enzima ocurre en ausencia de NTPs y NAD+, generando un enlace
2’,5’-fosfodiéster (Greer et al., 1983a). La naturaleza de este enlace se comprobó por su
resistencia a la digestión con las nucleasas P1 y RNasa T2, detectándose el
correspondiente dinucleótido mediante cromatografía en capa fina.
La reacción catalizada in vitro por la 2’,5’-RNA ligasa (de 19.9 kDa) tiene una
constante de equilibrio aparente cercana a la unidad, si bien favorable al corte de los
enlaces 2’,5’-fosfodiéster. La dirección del equilibrio in vivo dependerá de la
concentración efectiva de los sustratos disponibles, aunque como no se conocen
sustratos con enlace 2’,5’-fosfodiéster sobre los que pueda actuar, lo más probable es
que en condiciones fisiológicas funcione como una RNA ligasa (Arn y Abelson, 1996).
La enzima podría tener un papel relacionado con la ligación de los extremos producidos
por la ACNasa de E. coli en el tRNALys, si bien es cuestionable que esto ocurra in vivo
pues los fragmentos de tRNA se acumulan durante la infección con cepas del virus T4
mutados en el gen de la RNA ligasa T4.1 (Greer et al., 1983a). No obstante, la 2’,5’-
25

Introdución
RNA ligasa debe tener un papel en la célula, pues se han encontrado homólogos en α y
γ-proteobacterias, así como en bacterias verdes del azufre, en Gram positivas con bajo
contenido en G+C (Greer et al., 1983a) y en arqueas termófilas (Arn y Abelson, 1996;
Kato et al., 2003).
La 2’,5’-RNA ligasa cataliza la unión de extremos diferentes a los que requieren
las enzimas de T4 pero idénticos a los requeridos por las RNA ligasas eucarióticas, es
decir, 5’-hidroxilo y 2’,3’-fosfodiéster cíclico. Estos son los extremos que generan las
endonucleasas encargadas de la maduración de los tRNAs, unas enzimas presentes en
eucariotas y arqueas que son homólogas entre ellas (Peebles et al., 1983; Trotta et al.,
1997; Li et al., 1998; Paushkin et al., 2004; revisado en Abelson et al., 1998). De estas
observaciones se infiere que, conjuntamente con dichas endonucleasas, la 2’,5’-RNA
ligasa podría mediar la maduración de los RNAs. Por otro lado, su estructura
cristalográfica muestra similitud con la ciclofosfodiesterasa (CPD) de A. thaliana (Kato
et al., 2003), la enzima que procesa la ADP-ribosa 1”,2” fosfodiéster cíclico resultante
de eliminar el grupo 2’-fosforilo del enlace catalizado por las tRNA ligasas (Culver et
al., 1994; Genschik et al., 1997; Hofmann et al., 2000; ver más adelante). Las 2’,5’-
RNA ligasas, como las CPDs de A. thaliana levadura y vertebrados, y el dominio
ciclofosfodiesterasa de las tRNA ligasas (ver más adelante), pertenecen a la
superfamilia de fosfotransferasas 2H que catalizan la hidrólisis de enlaces fosfodiéster
cíclicos (Nasr y Filipowicz, 2000). Por lo tanto es factible que las 2’,5’-RNA ligasas
actúen alternativamente como CPD o RNA ligasas en función de los sustratos presentes
en la célula.
1.2.3.2. Otras RNA ligasas de bacterias y arqueas
En los dos reinos de arqueas, además de las enzimas homólogas a la 2’,5’-RNA
ligasa y la T4.2 RNA ligasa ya mencionadas, se ha detectado otra RNA ligasa que
participa en el procesamiento de los intrones de los tRNA en Desulfurococcus mobilis
(Crenarqueota) y Haloferax volcanii (Euryarqueota) (Kjems y Garrett, 1988; Gomes y
Gupta, 1997). La enzima de H. volcanii está implicada, además de en la ligación de las
dos mitades de los pre-tRNAs, en la circularización de los intrones eliminados (Salgia et
al., 2003). Sin embargo, existen importantes diferencias entre las RNA ligasas de ambas
arqueas. La enzima de D. mobilis cataliza la ligación de extremos 3’-fosfomonoéster y
5’-hidroxilo produciendo un enlace 3’,5’-fosfodiéster en ausencia de iones metálicos y
26

Introdución
empleando GTP (Kjems y Garrett, 1988), mientras que la de H. volcanii media la unión
de extremos 2’,3’-fosfodiéster y 5’-hidroxilo en ausencia de ATP, GTP y Mg+2, pero en
contraste con las 2’,5’-RNA ligasas genera un enlace 3’,5’-fosfodiéster (Gomes y
Gupta, 1997; Zofallova et al., 2000; Salgia et al., 2003). En ambas reacciones, a
diferencia de lo que ocurre con las tRNA ligasas y las RNA ligasas de T4, el grupo
fosforilo que se incorpora al enlace es el que se encuentra inicialmente en 3’ (Fig. 1.5),
a través de un mecanismo relacionado con el descrito para la RNA ligasa de animales
(Kjems y Garrett, 1988; Gomes y Gupta, 1997; ver más adelante).
Otra RNA ligasa que merece atención es la encontrada en Deinococcus
radiodurans, una bacteria capaz de soportar las condiciones extremas de radiación
existentes en centrales nucleares. Para hacer frente a dichas condiciones la bacteria ha
desarrollado un sistema de reparación de daños en el DNA muy eficiente que está
siendo estudiado exhaustivamente. Además, el gen que codifica esta proteína aumenta
su expresión in vivo durante la exposición a la radiación, mostrando que su función de
reparación del RNA es importante para la célula (Martins y Shuman, 2004b). La región
carboxilo terminal de la RNA ligasa de D. radiodurans tiene similitud estructural con la
T4.2 RNA ligasa, aunque no es capaz de catalizar la ligación de RNAs monocatenarios
sino que sólo sella roturas en RNAs bicatenarios o en híbridos RNA-DNA (Martins y
Shuman, 2004b). El reconocimiento de estas roturas viene determinado en el extremo
amino terminal por un dominio con un plegamiento de tipo unión de
oligonucleótido/oligosacárido (OB-fold) que no está presente en la T4.2 RNA ligasa
(Raymond y Shuman, 2007).
1.2.4. RNA ligasas eucarióticas
1.2.4.1. tRNA ligasa
1.2.4.1.1. Características generales y función biológica
En eucariotas, como en arqueas, las tRNA ligasas catalizan uno de los pasos
finales de la maduración de los pre-tRNAs (revisado en Hopper y Phizicky, 2003), por
lo que su función es esencial para el crecimiento celular (Phizicky et al., 1992). Tras ser
transcritos, los pre-tRNAs contienen un intrón justo en la secuencia del anticodón que
imposibilita su función (revisado en Hopper y Phizicky, 2003). Este intrón es eliminado
por una endonucleasa que genera extremos 5’-hidroxilo, 2’,3’-fosfodiéster cíclico
(Peebles et al., 1983; Trotta et al., 1997; revisado en Abelson et al., 1998), al igual que
27

Introdución
ocurre con la ACNasa de E. coli frente a la infección por T4 (Amitsur et al., 1987). De
esta forma se generan dos fragmentos cuya unión es catalizada por la tRNA ligasa a
través de tres actividades enzimáticas diferentes, ciclofosfodiesterasa, polinucleótido
quinasa y RNA ligasa, cada una radicadas en un domino proteico. Finalmente, una 2’-
fosfotransferasa transfiere el grupo 2’-fosforilo producto de la ligación a una molécula
de NAD+ para generar ADP-ribosa 1”,2” fosfodiéster cíclico y nicotinamida (McCraith
y Phizicky, 1991; Culver et al., 1993; Sawaya et al., 2005). Sobre esta última actúa la
CPD, dando lugar a ADP-ribosa 1”-fosfomonoéster (apartado 1.2.3.1; Culver et al.,
1994; Genschik et al., 1997; Nasr y Filipowicz, 2000; Hofmann et al., 2000).
tRNA endonucleasa
tRNA ligasa 2’-fosfotransferasa CPD PNK
Ligasa
Fig. 1.7. Mecanismo de maduración de los tRNAs. Durante este proceso el intrón es eliminado por una tRNA endonucleasa y, a continuación, la tRNA ligasa cataliza la unión de los extremos 5’-OH y 2’,3’>P a través de sus actividades ciclofosfodiesterasa (CPD), polinucleótido quinasa (PNK) y RNA ligasa. Finalmente el grupo fosforilo presente en 2’ es eliminado por una 2’ fosfotransferasa. En negro se indican las enzimas que intervienen en el mecanismo, y en rojo las diferentes actividades enzimáticas de la tRNA ligasa. (Adaptado de Sawaya et al., 2003).
En levaduras la tRNA ligasa también interviene en el procesamiento de
determinados mRNAs reguladores de la respuesta a proteínas no plegadas, una vía de
señalización intracelular que activa la transcripción de genes de chaperonas cuando en
el retículo endoplásmico se acumulan proteínas desplegadas (Sidrauski et al., 1996;
Gonzalez et al., 1999; revisado en Ron y Walter, 2007). Indicios adicionales en esta
dirección se han obtenido al comprobar que la mutación de un gen que codifica una de
las cuatro subunidades de la tRNA endonucleasa de humanos afecta tanto a la
maduración de los tRNAs como de los mRNAs (Paushkin et al., 2004).
Las tRNA ligasas eucarióticas más estudiadas son la de S. cerevisiae (Greer et
al., 1983b; Phizicky et al., 1986; Greer et al., 1987; Westaway et al., 1988; Xu et al.,
1990; Apostol et al., 1991; Beldford et al., 1993; Westaway et al., 1993; Sawaya et al.,
2003; Wang y Shuman, 2005; Wang et al., 2006a) y la de germen de trigo (Konarska et
28

Introdución
al., 1981; Konarska et al., 1982; Gegenheimer et al., 1983; Furneaux et al., 1983;
Schwartz et al., 1983; Pick y Hurwitz, 1986; Pick et al., 1986, Makino et al., 2006).
Ambas, así como sus homólogas de hongos (Sawaya et al., 2003) y plantas (Englert y
Beier, 2005), comparten muchas similitudes. Las dos median la unión de extremos
2’,3’-fosfodiéster cíclico y 5’-hidroxilo (Konarska et al., 1982, Greer et al., 1983b),
poseen la misma distribución de las actividades RNA ligasa, polinucleótido quinasa y
ciclofosfodiesterasa, que respectivamente catalizan la ligación propiamente dicha, la
fosforilación del extremo 5’ y la hidrólisis del enlace 2’,3’-fosfodiéster cíclico (Xu et
al., 1990; Apostol et al., 1991; Englert y Beier, 2005), y presentan un mecanismo de
reacción similar. Sin embargo, la tRNA ligasa de S. cerevisiae muestra una alta
especificidad de sustrato, catalizando 104 veces menos eficientemente la ligación de
oligonucleótidos no estructurados que los productos del procesamiento de los pre-tRNA
(Phizicky et al., 1986; Greer et al., 1987), mientras que la de germen de trigo (y de otras
plantas) cataliza con la misma eficiencia la ligación de productos del procesamiento de
los pre-tRNA y de oligonucleótidos artificiales, con el único requerimiento de que
posean extremos 5’-hidroxilo y 2’,3’-fosfodiéster cíclico (Schwartz et al., 1983). La
menor especificidad de las tRNA ligasas de plantas podría haber sido aprovechada por
los viroides para completar su ciclo replicativo, pues extractos de germen de trigo y del
alga Chlamydomonas reinhardtii circularizan eficientemente RNAs monoméricos
lineales de PSTVd aislados de tejido infectado (Branch et al., 1982; Kikuchi et al.,
1982). Dicha especificidad de la tRNA ligasa de levadura por su sustrato no está
determinada por el dominio ciclofosfodiesterasa, pues éste media eficientemente la
hidrólisis del extremo 2’,3’-fosfodiéster cíclico de otros RNAs distintos de los pre-
tRNAs (Greer et al., 1983b).
La secuencia aminoacídica de la RNA ligasa de S. cerevisiae se determinó
parcialmente a partir de preparaciones purificadas de la misma (Phizicky et al., 1986),
empleándose esta información para clonar el gen que la codifica (Westaway et al.,
1988). El gen homólogo de plantas no pudo ser identificado en búsquedas
bioinformáticas debido a su escasa similitud de secuencia con el de la tRNA ligasa de
levadura. Recientemente se purificó la enzima de germen de trigo, y a partir de la
secuencia parcial obtenida, se identificaron en las bases de datos los correspondientes
genes de A. thaliana y Oryza sativa (Englert y Beier, 2005). Mas tarde se han
caracterizado hasta tres isoformas de la tRNA ligasa de germen de trigo (Makino et al.,
2006).
29

Introducción
1.2.4.1.2. Mecanismo de ligación
Como ya se ha mencionado, las tRNA ligasas realizan su función mediante tres
actividades enzimáticas diferentes cada una localizada en un dominio proteico. En
primer lugar se produce la interacción entre el RNA y la enzima (Fig. 1.8, etapa 1), y a
continuación el dominio ciclofosfodiesterasa situado en la región carboxilo terminal de
la misma cataliza la hidrólisis del 2’,3’-fosfodiéster cíclico dejando el grupo fosforilo en
2’ (Fig. 1.8, etapa 2). Seguidamente se produce la fosforilación del 5’-hidroxilo
mediante la acción del dominio polinucleótido quinasa situado en la región central de la
proteína, conjuntamente con la adenilación del dominio RNA ligasa amino terminal
(Fig. 1.8, etapas 3 y 4). La fosforilación genera un extremo 5’-fosfomonoéster al que se
transfiere el AMP previamente incorporado en el dominio RNA ligasa (Fig. 1.8, etapa
5). La última etapa consiste en el ataque del extremo 3’-hidroxilo de la otra mitad del
tRNA al 5’-adenilado generando el grupo 2’-fosfomonoéster, 3’,5’-fosfodiéster y
liberando el AMP (Fig. 1.8, etapa 6).
Aunque es posible ensayar la ligación a partir de cada uno de los intermediarios
de las tres reacciones, la eficiencia no es igual en todos los casos mostrando así que se
trata de un mecanismo cooperativo (Pick y Hurwitz, 1986; Pick et al., 1986). Si bien la
actividad ciclofosfodiesterasa de la tRNA ligasa de germen de trigo (Fig. 1.8, etapa 2)
puede ensayarse en ausencia de ATP y del extremo 5’-hidroxilo, la eficiencia de la
reacción aumenta en presencia de este último (Pick et al., 1986). Sin embargo, la
actividad de ligación presenta eficiencia similar empleando extremos 3’ aceptores tanto
con estructura cíclica como abierta (Schwartz et al., 1983; Pick et al., 1986), mostrando
que la actividad ciclofosfodiesterasa está coordinada con la polinucleótido quinasa pero
no con la de ligación propiamente dicha. También se ha observado que la adenilación
del dominio ligasa (Fig. 1.8, etapa 3) es mayor en presencia de un 5’-hidroxilo que de
un 5’-fosfomonoéster, sugiriendo que está coordinada con la actividad polinucleótido
quinasa, y que la de adenilación de la enzima es anterior a la fosforilación del 5’-
hidroxilo (Fig. 1.8, etapa 3 y 4) (Pick et al., 1986). Finalmente, la ligación es
aproximadamente cinco veces mayor si el sustrato posee un extremo 5’-hidroxilo en vez
de un 5’-fosfomonoéster, indicando que la actividad de ligación está también
coordinada con la de fosforilación (Pick et al., 1986). Una diferencia entre las tRNA
ligasas de germen de trigo y levadura es que mientras en la primera la adenilación
aumenta en presencia del extremo 5’, en la segunda no varía. Esta observación indica
30

Introducción
que la enzima de plantas no se adenila si no hay sustrato para la fosforilación (5’-
hidroxilo), mientras que la de levaduras se halla adenilada independientemente de si hay
o no RNA en el medio (Greer et al., 1983b; Pick et al., 1986).
5’
RNA
E + HORNA>P E · HORNA>P
E · HORNA>P E · HORNAOHP
-E · HORNAOH + PPiPE · HORNAOH + PPPAP
-E · PRNAOH + PPAP
-E · HORNAOH + PPPAP
-E · PRNAOHP E · APPRNAOH
P
HORNA>P + 2 PPPAE
P2’
P3’ + PPi + PPA + PA
E · APPRNAOHP E + + PA
(1) Unión enzima-RNA
(2) Rotura del fosfodiéster cíclico
(3) Complejo enzima-AMP
(4) Fosforilación en 5’
(5) Adenilación del RNA
(6) Ligación
2’
PRNA
5’ 3’
P
PA
PAPA
PA
5’
RNA
E + HORNA>P E · HORNA>PE + HORNA>P E · HORNA>P
E · HORNA>P E · HORNAOHPE · HORNA>P E · HORNAOHPE · HORNAOHP
-E · HORNAOH + PPiPE · HORNAOH + PPPAP
-E · PRNAOH + PPAP
-E · HORNAOH + PPPAPE · HORNAOH + PPPAP
-E · PRNAOHP E · APPRNAOH
P
-E · PRNAOHP E · APPRNAOH
PE · APPRNAOHP
HORNA>P + 2 PPPAE
P2’
P3’ + PPi + PPA + PA
E · APPRNAOHPE · APPRNAOHPE · APPRNAOHP E + + PA
(1) Unión enzima-RNA
(2) Rotura del fosfodiéster cíclico
(3) Complejo enzima-AMP
(4) Fosforilación en 5’
(5) Adenilación del RNA
(6) Ligación
2’
PRNA
5’ 3’
P2’
PRNA
5’ 3’
P
PA
PAPA
PA
Fig. 1.8. Esquema del mecanismo de la tRNA ligasa de germen de trigo (E). 1) Formación del complejo RNA-enzima; 2) apertura por la actividad ciclofosfodiesterasa del grupo 2’,3’>P generando extremos 3’-OH y 2’-P; 3) reacción del dominio RNA ligasa con una molécula de ATP para formar el complejo AMP-enzima; 4) fosforilación del extremo 5’-OH del pre-tRNA mediada por la actividad polinucleótido quinasa localizada en el dominio central de la enzima; 5) adenilación del extremo 5’-P generando el intermediario de la reacción enzima-AMP-5’-P-RNA; 6) ataque del extremo 3’-OH sobre el enlace entre el AMP y el extremo 5’-P, generando un enlace 3’,5’-fosfodiéster nuevo y liberando el AMP y la enzima. El código de colores facilita la identificación de los grupos químicos participantes, donde el azul se corresponde al sustrato de la ligación, el rojo al residuo adenilato implicado en la adenilación y el verde al grupo fosforilo que finalmente se incorpora en el enlace. (Adaptado de Pick et al., 1986).
Experimentos realizados con estas tRNA ligasas muestran que la de S. cerevisiae
puede catalizar la ligación con los cuatro NTPs, si bien emplea preferentemente ATP y
UTP (Greer et al., 1983b), mientras que la enzima de germen de trigo sólo actúa en
presencia de ATP (Pick et al., 1986). En otros experimentos con la tRNA ligasa de
31

Introducción
levadura sólo se observó ligación con ATP, por lo que el resultado anterior podría
deberse a una contaminación con nucleótidos durante la extracción (Westaway et al.,
1993; Belford et al., 1993). También se comprobó en estos trabajos que la fosforilación
del sustrato es mucho más eficiente con GTP, y que la ligación también aumenta en
presencia de GTP y ATP en vez de ATP sólo. Sin embargo, este último es esencial ya
que sin él la reacción no progresa. Experimentos posteriores de mutación dirigida han
revelado que la tRNA ligasa de levadura posee dos sitios de unión, uno para ATP y otro
para GTP. Mutaciones en el sitio para el ATP no son letales, pero sí lo son en el sitio
para el GTP, mostrando que este último es el verdadero sustrato que requiere la enzima
para la fosforilación del extremo 5’-hidroxilo (Sawaya et al., 2003). Este sitio
alternativo para el GTP no se ha descrito en la tRNA ligasa de germen de trigo ni en
ninguna de sus homólogas de plantas.
1.2.4.2. RNA ligasas mitocondriales
Los kinetoplástidos, pertenecientes al filo Euglenozoa (protozoos ciliados) del
reino Protista, incluyen dos parásitos de humanos muy estudiados: Tripanosoma brucei
y Leishmania tarentolae. La mitocondria de estos organismos posee una actividad de
edición de los mRNAs mediante incorporación o eliminación de residuos de uridilato
especificada por un RNA guía (gRNA). El dúplex pre-mRNA-gRNA se asocia a un
complejo multiproteico con capacidad de añadir y eliminar uridilatos, y de catalizar la
ligación finalmente del mRNA editado, generando nuevos codones de iniciación,
cambiando la pauta de lectura o incluso generando así pautas nuevas (Schnaufer et al.,
2001; McManus et al., 2001). La reacción requiere ATP o dATP y produce un enlace
3’,5’-fosfodiéster mediante un mecanismo compartido con las T4 RNA ligasas (White y
Borst, 1987; Bakalara et al., 1989). Sin embargo, la endonucleasa que cataliza el corte
parece generar extremos 5’-fosfomonoéster y 3’-hidroxilo, a diferencia de las
endonucleasas que preceden a la reacción de la T4.1 RNA ligasa y tRNA ligasa que
generan extremos 5’-hidroxilo, 2’,3’-fosfodiéster cíclico (Sabatini y Hajduk, 1995).
Estas RNA ligasas poseen los dominios que caracterizan a la superfamilia de las
nucleotidil transferasas covalentes (Schnaufer et al., 2001; McManus et al., 2001), y
están relacionadas con la T4.2 RNA ligasa (Ho y Shuman, 2002).
32

Introducción
1.2.4.3. Otras RNA ligasas eucariotas
Además del mecanismo conservado de ligación de los tRNAs, también se ha
detectado in vitro una actividad de ligación de pre-tRNAs de levaduras en extractos de
células HeLa y en núcleos de oocitos de Xenopus laevis (Nishikura y De Robertis, 1981;
Filipowicz y Shatkin, 1983; Filipowicz et al., 1983; Perkins et al., 1985). Los extremos
ligados son igualmente 5’-hidroxilo y 2’,3’-fosfodiéster cíclico, pero el enlace generado
es 3’,5’-fosfodiéster (sin el 2’-fosfomonoéster residual resultante de la reacción de
ligación de los tRNA de plantas y levadura), donde el grupo fosforilo proviene del
fosfodiéster cíclico del extremo aceptor como en las RNA ligasas de arqueas y no de un
cofactor de tipo NTP como ocurre con las tRNA ligasas (Filipowicz y Shatkin, 1983,
revisado en Arn y Abelson, 1998). En los mismos extractos que contienen esta RNA
ligasa se detectó una actividad fosfato ciclasa 3’-terminal dependiente de ATP, cuya
función precede a la RNA ligasa cuando se emplea como sustrato un RNA con
extremos 3’-fosfomonoéster (Filipowicz et al., 1983). Dicha RNA ligasa tiene un
tamaño de 160 kDa (Perkins et al., 1985), y se la conoce como la RNA ligasa de
animales porque sólo se ha detectado en vertebrados.
Existen pruebas bioquímicas de la coexistencia en células HeLa de este
mecanismo con otro del tipo tRNA ligasa (incluyendo la 2’-fosfotransferasa) en el que
el grupo fosforilo del nuevo enlace es aportado exógenamente (Zillmann et al., 1991;
Zillmann et al., 1992; Hu et al., 2003). Sin embargo, las enzimas implicadas en la
ligación serían diferentes pues la fosforilación del extremo 5’-hidroxilo está catalizada
en humanos por la RNA quinasa hClp1, que a diferencia de la actividad quinasa de la
tRNA ligasa, es una enzima independiente aunque parece formar un complejo con la
tRNA endonucleasa como demuestran los experimentos de copurificación (Paushkin et
al., 2004; Weitzer y Martínez, 2007).
Queda por dilucidar cuáles son la CPD y la RNA ligasa que median el
mecanismo de tipo tRNA ligasa en humanos. A este respecto, la 2′,3′-nucleótido cíclico
3′-fosfodiesterasa (2′,3′-cyclic nucleotide 3′-phosphodiesterase, CNP), con un tamaño
de 46 kDa y homóloga al dominio ciclofosfodiesterasa de las tRNA ligasas, es la única
CPD descrita en animales. La función de esta enzima es desconocida, si bien su
ausencia parece estar implicada en la aparición tardía de procesos neurodegenerativos
(Lappe-Siefke et al., 2003). Ratones con las dos copias de la CNP mutadas crecen
normalmente (Lappe-Siefke et al., 2003), y lo mismo ocurre cuando se mutan las dos
33

Introducción
copias de la 2’-fosfotransferasa (Harding et al., 2008). Por tanto, estos resultados
sugirien que el mecanismo de ligación de tipo tRNA ligasa no es necesario en
vertebrados, probablemente porque la RNA ligasa de animales, que no requiere
actividades CPD ni 2’-fosfotransferasa, interviene en la maduración de los tRNAs y
probablemente también en la respuesta a proteínas no plegadas (Harding et al., 2008).
Por el contrario la levadura, que carece de una ruta alternativa a la de tipo tRNA ligasa,
sí es sensible a la mutación del gen de la 2’-fosfotransferasa (Spinelli et al., 1997;
Culver et al., 1997). Recientemente se ha descrito que la CNP puede ligar extremos 5’-
hidroxilo y 2’,3’-fosfodiéster cíclico al igual que la mayoría de las RNA ligasas
conocidas (Schwer et al., 2008), pero se desconoce si esta función es importante para la
célula.
1.2.5. Comparación entre mecanismos
A pesar de la distancia evolutiva que separa el mecanismo de maduración de los
pre-tRNAs y el de restricción/reparación de los tRNAs de E. coli infectada por el fago
T4, la ligación en ambos casos es muy parecida. En la figura 1.9 puede observarse que
la secuencia de reacciones es compartida en ambos casos excepto en lo que respecta a la
actividad fosfatasa de la T4 PNK, que tras abrir el 2’,3’-fosfodiéster cíclico deja
extremos 3’ y 2’-hidroxilo (Cameron y Uhlenbeck, 1977), mientras que el dominio
ciclofosfodiesterasa de la tRNA ligasa no tiene actividad fosfatasa y deja el grupo
fosforilo en 2’. Esta diferencia entre ambos mecanismos de ligación conlleva un paso
adicional en eucariotas para eliminar el 2’-fosfomonoéster catalizado por una 2’-
fosfotransferasa (Fig. 1.7 y 1.10), donde el grupo fosforilo del RNA ataca la unión entre
la nicotinamida y la ribosa del NAD+ generando nicotinamida libre y ADP ribosa 1”,2”-
fosfodiéster cíclico (McCraith y Phizicky, 1991; Culver et al., 1993; Sawaya et al.,
2005). Como se ha comentado, la función que realiza esta enzima en levadura es
esencial para la maduración de los tRNAs y el crecimiento, y si no está presente el
anticodón no es funcional y no hay crecimiento vegetativo (Spinelli et al., 1997; Culver
et al., 1997). Dicha enzima está evolutivamente muy conservada y se encuentra en
bacterias, arqueas, protozoos, levaduras, plantas y vertebrados (Spinelli et al., 1998), si
bien su presencia en bacterias y arqueas es un enigma, pues según el mecanismo de
ligación que catalizan sus RNA ligasas, la 2’-fosfotransferasa sería prescindible.
34

Introducción
Fig. 1.9. Esquema de las diferentes etapas del mecanismo de ligación de RNA catalizado por las enzimas
de levadura y plantas (izquierda) y del fago T4 (derecha). (Adaptado de Arn y Abelson, 1998).
Mediante experimentos de complementación in vitro se ha comprobado que la
T4 PNK y la T4.1 RNA ligasa son capaces conjuntamente de catalizar la unión de las
mitades de los pre-tRNAs generadas por la endonucleasa de levadura (Greer et al.,
1983b). En experimentos in vivo se ha observado que un mutante de S. cerevisiae con la
tRNA ligasa inactivada es capaz de crecer normalmente al ser transformado con los
genes de la PNK y RNA ligasa 1 del fago T4, prescindiendo de la actividad 2’-
fosfotransferasa. Además, la actividad conjunta de la PNK y la RNA ligasa 1 de T4 es
incluso capaz de sustituir a la tRNA ligasa en la respuesta a proteínas no plegadas
(Schwer et al., 2004). En estos mismos experimentos se observó que la T4.1 RNA
ligasa junto con los dominios ciclofosfodiesterasa y polinucleótido quinasa de la tRNA
APPP*
OH
PA
OH 3’
2’
5’ P*
P
T4 RNA ligasa
OH 3’
2’
5’ P*
OH
5’ 3’
2’ OH
P*
tRNA ligasa
3’
2’
5’ HO
P
OH 3’
2’
5’ P*
P
3’ OH
2’
5’ HO
P 2’
P
APPP*
APP
5’ 3’
2’ P
P*
EPA
E
PPi
APPP
AP
PA
OH 3’
2’
5’ P*
OH
AP
OH 3’
2’
5’ HO
APP
P 3’ 5’
HO
AP
T4 PNK
EPA PPi
E APPP
35

Introducción
ligasa de levadura restauran el crecimiento del mutante de S. cerevisiae pero, sin
embargo, el mismo mutante transformado con el gen de la PNK de T4 junto con el
fragmento que codifica el dominio RNA ligasa de levadura no crece. Esta situación
deriva de que el grupo fosforilo en 2’ producto de la acción sobre el sustrato de los
dominios ciclofosfodiesterasa y polinucleótido quinasa de levadura no interfiere en la
ligación catalizada por la T4.1 RNA ligasa. Por el contrario, el dominio RNA ligasa de
la enzima de levadura sí requiere este grupo fosforilo en 2’ para su función, que sin
embargo es eliminado del RNA cuando se trata con la PNK de T4. Este mecanismo
podría ser una adaptación para evitar la ligación accidental por la tRNA ligasa de RNAs
con extremos 3’ y 2’-hidroxilo, que son muy abundantes en la célula (Schwer et al.,
2004) (Fig. 1.10). También se ha comprobado que la expresión de una única copia de la
tRNA ligasa de A. thaliana bajo el control de un promotor constitutivo es suficiente
para restaurar el crecimiento de cepas de S. cerevisiae con el gen de su tRNA ligasa
mutado (Wang et al., 2006a).
Maduración de tRNAs
tRNA ligasa tRNA ligasa 2’-fosfotransferasaCPD ligasa
tRNA 5’-quinasa endonucleasa
Intrón
Restricción/reparación de tRNAs
PNK T4.1 RNA ligasa CPD/3’-fosfatasa PrrC
anticodonucleasa ligasa
5’-quinasa
Fig. 1.10. Comparación entre las funciones biológicas realizadas por las tRNA ligasas y la T4 RNA ligasa. En negro se indican las enzimas que intervienen en el mecanismo, en rojo, las diferentes actividades de dichas enzimas que actúan en cada momento y en el panel superior es azul el intrón que es eliminado durante la maduración de los tRNAs. (Adaptado de Schwer et al., 2004).
En la bacteria Clostridium thermocellum se ha descubierto recientemente una
polinucleótido quinasa-fosfatasa que parece estar implicada en un mecanismo de
reparación del RNA y, que al igual que la PNK de T4, genera extremos 3’ y 2’-
hidroxilo en el RNA mediante su actividad ciclofosfodiesterasa-fosfatasa (Martins y
36

Introducción
Shuman, 2005, Keppetipola y Shuman., 2007). Estos experimentos también muestran
que una simple mutación puntual provoca que, al abrirse el fosfodiéster cíclico, el grupo
fosforilo resultante no se hidrolice, sino que permanezca en 2’ como ocurre con el
dominio ciclofosfodiesterasa de las tRNA ligasas. De esta forma la enzima silvestre
genera extremos adecuados para las T4 RNA ligasas, mientras que la mutante (que no
tiene afectadas el resto de sus funciones) genera los extremos requeridos por las tRNA
ligasas eucarióticas. Esta peculiaridad observada en dicha polinucleótido quinasa-
fosfatasa muestra como podría haber ocurrido la evolución entre los mecanismos de
ligación mediados por la T4.1 y las tRNA ligasas (Keppetipola et al., 2007).
1.3. Chlamydomonas reinhardtii como organismo modelo
ada especificidad
de hosp
1.3.1. Características generales
ismo eucariótico unicelular fotosintético clasificado
taxonóm
género Chlamydomonas puede encontrarse en agua salada o dulce, en el suelo e
inclu
Los miembros de la familia Avsunviroidae muestran una elev
edador y, además, las especies que infectan no son habitualmente empleadas en
el laboratorio. Por esta razón se exploró la posibilidad de utilizar un sistema modelo
cloroplástico heterólogo para estudiar distintos aspectos de la biología molecular de los
miembros de esta familia en un contexto fisiológico. Entre los modelos clásicos para el
estudio del cloroplasto se encuentra el alga verde unicelular C. reinhardtii, que se eligió
para este trabajo por su facilidad para transformar sus tres genomas, que además están
completamente secuenciados.
C. reinhardtii es un organ
icamente dentro del orden Volvocales, que a su vez se encuadra en la división
Clorófitos (comúnmente conocidas como algas verdes) dentro del reino Planta, y que se
supone representan el ancestro de las actuales plantas superiores. Morfológicamente C.
reinhardtii es una célula con estructura polar y forma elipsoide de 10 µm de longitud y
3 µm de anchura, con dos flagelos anteriores, algunas mitocondrias y un único
cloroplasto basal que ocupa casi el 40% del volumen celular rodeando al núcleo (Harris,
1989).
El
so en la nieve. Se han descrito más de 500 especies del mismo, entre las que C.
reinhardtii es la más ampliamente empleada en el laboratorio. Además del crecer
fotosintéticamente, esta especie también es fácilmente cultivable en condiciones de
37

Introducción
oscuridad y heterotrofía empleando acetato como fuente única de carbono a través del
ciclo del glioxilato. En estas condiciones, y a diferencia de las plantas superiores, C.
reinhardtii es capaz de ensamblar correctamente su maquinaria fotosintética
permitiendo el estudio de mutantes de la misma (Harris, 1989). Por dicha razón se ha
empleado tradicionalmente este alga como organismo modelo para el estudio de la
fotosíntesis.
ig. 1.11. Reproducción sexual por meiósporas y distribución de los genomas en C. reinhardtii. Mientras las mutaciones nucleares se heredan siguiendo un patrón mendeliano, en cloroplastos y mitocondrias
ste organismo muestra un rápido crecimiento con divisiones cada 5-8 h y un
ciclo
Cariogamia
Meiosis
mt+ mt -
Tétrada
mt -mt+ mt+ mt -
Clor
Mitocondria
oplasto Núcleo
Fquese observa una herencia uniparental que depende de la polaridad de las células parentales. (Adaptado de Nickelsen y Kück, 2000).
E
biológico haplo-diploide. La gametogénesis puede ser inducida fácilmente en el
laboratorio mediante privación de nitrógeno en el medio de cultivo (Nickelsen y Kück,
2000). Los gametos son morfológicamente similares, y sólo se diferencian en el locus
genético (mt+ y mt-) que determina el tipo sexual (Ferris y Goodenough, 1994). Cuando
dos gametos opuestos se encuentran ocurre una fusión nuclear y cloroplástica, y el
38

Introducción
cigoto resultante se rodea de una pared celular impermeable que le protege de las
condiciones desfavorables. Cuando las condiciones nutricionales son adecuadas se
produce la meiosis, generando el cigoto cuatro células haploides (Fig. 1.11). Esta
característica resulta de gran utilidad al permitir el análisis genético y el seguimiento de
mutaciones heredables (Nickelsen y Kück, 2000).
1.3.2. El cloroplasto y su origen endosimbióntico
ma la
energ
ado que tanto el cloroplasto como la mitocondria
present
El cloroplasto, el orgánulo eucariótico responsable de la fotosíntesis, transfor
ía lumínica en ATP y poder reductor (NADPH) a través de los complejos
proteicos fotosintéticos de las membranas tilacoidales. Este proceso ocurre de forma
localizada en el estroma del cloroplasto, donde tienen lugar reacciones bioquímicas
como la fijación de carbono y producción de azúcares, la fijación de nitrógeno, azufre y
fósforo, así como la biosíntesis de ciertos aminoácidos, lípidos, almidón y pigmentos
(Bogorad, 2000; Zerges, 2000).
Está ampliamente acept
aban originariamente vida independiente como organismos procariotas (Martin y
Herrmann, 1998; Brinkman et al., 2002). Estos orgánulos eran eubacterias que
accedieron como endosimbiontes a una célula hospedadora, y gradualmente
evolucionaron hasta adquirir las características que actualmente presentan. Diversos
estudios han permitido establecer que las mitocondrias y las α-proteobacterias tienen un
ancestro común, así como los cloroplastos y las cianobacterias (revisado en Gray,
1993). También se ha establecido que el cloroplasto se incorporó a la célula eucariótica
posteriormente a la mitocondria, por lo que la distribución del primero dentro de la
división Eucaria es más restringida, pero existe la posibilidad de que ambos se
incorporaran tempranamente y luego el cloroplasto se perdiera en los linajes no
fotosintéticos (Andersson y Roger, 2002). Aunque durante la evolución del proceso
endosimbióntico la mayoría de los genes del endosimbionte fueron eliminados o
transferidos al núcleo (Martin y Herrmann, 1998), el genoma cloroplástico aún posee
muchas características comunes con el de bacterias, como se refleja en sus ribosomas
70S (Yamaguchi et al., 2003), y en las propiedades de sus RNAs mensajeros (Zurawski
et al., 1981). Sin embargo, debido al intercambio génico entre núcleo y cloroplasto a lo
largo de la evolución, no todos los mecanismos implicados en la expresión génica de
este orgánulo son estrictamente bacterianos (Yamaguchi et al., 2003). Se postula que
39

Introducción
parte del genoma endosimbióntico ancestral se mantiene en los cloroplastos porque, aún
siendo costoso para la célula, esa disposición ofrece la ventaja evolutiva de garantizar
una rápida respuesta frente al estrés oxidativo (Race et al., 1999).
1.3.3. El genoma de C. reinhardtii
s fotosintéticos, C. reinhardtii posee tres sistemas
genétic
as, y se caracteriza por ser
lineal
genoma cloroplástico muestra muchas características en común con el de
plant
mayoría de los genomas cloroplásticos presentan dos copias de dos grandes
regiones invertidas (inverted region, IR) que pueden variar de longitud como resultado
Al igual que otros eucariota
os independientes localizados en el núcleo, la mitocondria y el cloroplasto. El
genoma nuclear contiene 108 pares de bases (pb) distribuidos en 17 grupos de
ligamiento, es inusualmente rico en G+C (62%), y sus genes se heredan
mendelianamente durante la división meiótica (Harris, 1989). La transformación del
núcleo se lleva a cabo introduciendo el DNA foráneo en células carentes de pared
celular mediante agitación con bolas de vidrio (Kindle, 1990) o electroporación
(Shimogawara et al., 1998). En cualquier caso, la recombinación homóloga en el DNA
nuclear ocurre con una frecuencia muy baja y la inserción del DNA foráneo tiene lugar
al azar mediante recombinación heteróloga (Rochaix, 1995).
El genoma mitocondrial está presente en múltiples copi
y de pequeño tamaño (15.8 kpb) en comparación con los de otras algas verdes y
plantas superiores. Presenta regiones invertidas en los extremos y sólo codifica ocho
proteínas, tres tRNAs y dos rRNAs (Michaelis et al., 1990). Los gametos heredan el
DNA mitocondrial de la cepa mt- (Harris, 1989; Nickelsen y Kück, 2000). Este genoma
también puede ser transformado, aunque con baja eficiencia (Randolph-Anderson et al.,
1993).
El
as superiores, incluyendo la conservación de diversos genes. Es circular, de
aproximadamente 196 kpb (unas 34 veces mayor que el genoma mitocondrial), y
contiene alrededor de 80 genes en su mayoría implicados en la maquinaria fotosintética
y en la expresión génica del orgánulo (Nickelsen y Kück, 2000). Cada cloroplasto posee
aproximadamente 80 copias de este cromosoma, que se hereda monoparentalmente de
la cepa mt+ (Harris, 1989; Nickelsen y Kück, 2000). Tras la formación del cigoto, el
genoma cloroplástico de la cepa mt- es degradado por la acción de nucleasas (Kuroiwa,
1991).
La
40

Introducción
de un
s en operones, por lo general no ocurre así en C. reinhardtii,
aunqu
(Wendy I y II), que han
diver
Aunque la expresión génica nuclear está controlada principalmente a nivel
sugieren que la estabilización diferencial de
algun
a duplicación de, entre otros, los genes del rRNA. Entre los distintos taxones de
plantas superiores existen importantes variaciones en el tamaño del genoma
cloroplástico debidas a la longitud de estas regiones invertidas, sin que por ello se vea
afectada la complejidad genómica. En cambio, las variaciones de tamaño existentes
dentro del género Chlamydomonas se deben en gran medida a múltiples inserciones y
eliminaciones fuera de estas regiones invertidas (Palmer, 1985). La organización génica
dentro de las regiones invertidas es la típica de plantas superiores, excepto el gen que
codifica el 23S rRNA que contiene un intrón y el psbA que contiene cuatro intrones
(Maul et al., 2002).
Mientras que en plantas superiores la mayoría de los genes del cloroplasto
aparecen organizado
e puede observarse esta organización en las secuencias que codifican las proteínas
ribosomales (Sakamoto et al., 1994). Los genes que están dispuestos en operones sufren
un rápido procesamiento postranscripcional, pudiendo dar la impresión de ser
transcritos de forma independiente (Stern y Drager, 1998).
También es característica de C. reinhardtii la presencia de dos copias de un
elemento móvil de DNA de 2.4 kb denominado Wendy
gido por acumulación de mutaciones puntuales y por pequeñas inserciones y
eliminaciones. Estas dos copias alteran el orden de dos agrupamientos génicos en C.
reinhardtii que son contiguos en C. moewusii, sugiriendo que Wendy invadió C.
reinhardtii tras la separación de ambas especies (Fan et al., 1995). La abundancia de
repeticiones cortas dispersas en un bajo número de regiones codificantes, y una atípica y
exclusiva organización de los genes que codifican la RNA polimerasa, son también
característicos de la especie C. reinhardtii (Maul et al., 2002).
1.3.4. Regulación de la expresión génica
transcripcional, hay pruebas genéticas que
os RNAs podría ser esencial en la expresión de genes nucleares que codifican
proteínas cloroplásticas (Hahn et al., 1996). El control de la expresión génica
cloroplástica parece ocurrir fundamentalmente a nivel postranscripcional, actuando
sobre la estabilidad de los transcritos (Rochaix, 1996), y está regulado por los ciclos de
luz y oscuridad (Salvador et al., 1993a): la transcripción aumenta generalmente en los
41

Introducción
períodos de luz mientras que la estabilidad del RNA es mayor durante la fase oscura. El
análisis de mutaciones dirigidas en el cloroplasto revela que la degradación de los
RNAs no es un proceso al azar, sino que parece existir una regulación específica de
cada transcrito que viene determinada entre otros factores por las regiones 5’ y 3’ no
traducibles (Stern et al., 1991; Salvador et al., 1993b; revisado en Herrin y Nickelsen,
2004). Otros experimentos han mostrado que la maquinaria de degradación se ve
influida por el potencial redox del estroma (Salvador y Klein, 1999). Sin embargo, no
parece haber correlación entre la transcripción y la estabilidad de los mRNAs (Eberhard
et al., 2002; revisado en Herrin y Nickelsen, 2004).
1.3.5. Transformación del cloroplasto
La transformación genética del cloroplasto despierta un gran interés dentro de la
de características que resultan ventajosas en
comp
o wolframio con los que se bombardea el cultivo de C. reinhardtii mediante
acele
biotecnología de plantas por una serie
aración con la transformación nuclear. Entre estas ventajas están: i) la inserción
direccional del DNA foráneo por un fenómeno de recombinación homóloga, que
permite introducir en el lugar adecuado y con la orientación correcta el gen de interés,
ii) el procesamiento de mRNAs policistrónicos organizados en operones, facilitando de
esa forma la expresión fenotípica de caracteres multigénicos interesantes
agronómicamente, y iii) la herencia uniparental de este orgánulo, evitando la
transferencia vertical de los transgenes a poblaciones naturales. Estas características han
sido aprovechadas en diversos estudios para comprobar el enorme potencial del alga C.
reinhardtii como biofactoría para la expresión de proteínas heterólogas de interés
biotecnológico. Algunos ejemplos son la biosíntesis de polihidroxialcalonatos (PHAs),
una clase de biopolímeros biodegradables (Poirier et al., 1995; Heifetz, 2000),
anticuerpos monoclonales humanos (Mayfield et al., 2003), y antígenos bacterianos
como el TetC, que en forma de vacuna confiere resisténcia al tétanos (Tregoning et al.,
2004).
Para realizar la transformación cloroplástica, el DNA se adsorbe sobre proyectiles
de oro
ración en un flujo de helio. Los cultivos transformados carecen de la región de un
gen que codifica una proteína del aparato fotosintético, pudiendo seleccionarse los
transformantes porque restauran la actividad fotosintética mediante recombinación
homóloga (Boynton et al., 1988). También pueden seleccionarse los transformantes
42

Introducción
integrando durante la transformación marcadores heterólogos que confieran resistencia
a antibióticos (Goldschmidt-Clermont et al., 1991).
43


22.. OOBBJJEETTIIVVOOSS


Objetivos
En la presente Tesis Doctoral se ha profundizado en el estudio del
procesamiento de los RNAs viroidales durante su ciclo replicativo, y más
concretamente en la etapa final de ligación que ocurre en dicho ciclo.
Los objetivos específicos planteados han sido:
1) Detectar y caracterizar bioquímicamente una RNA ligasa cloroplástica que
pudiera estar implicada en la circularización de los miembros de la familia
Avsunviroidae
2) Identificar y expresar una tRNA ligasa nuclear de un hospedador natural de
miembros de la familia Pospiviroidae
3) Establecer un sistema heterólogo viroide-cloroplasto, basado en la
transformación del cloroplasto del organismo modelo Chlamydomonas
reinhardtii, que permita estudiar in vivo el ciclo replicativo de los miembros
de la familia Avsunviroidae
47


33.. MMAATTEERRIIAALLEESS YY MMÉÉTTOODDOOSS


Material y métodos
3.1. Extracción de proteínas de cloroplastos de espinaca
3.1.1. Obtención de cloroplastos intactos
El material de partida consistió en 25 g de hojas de espinaca (Spinacea oleracea
L.) cv. Matador cultivadas en cabina durante 1-2.5 meses, con un periodo de 12 h de luz
y 12 h de oscuridad, y una temperatura de 24ºC durante el día y 21ºC por la noche.
Alícuotas de 5 g de hojas cortadas en trozos de aproximadamente 1 cm2 se
homogeneizaron con un Omnimixer (Omni International) (3 pulsos de 5 s en la posición
nº 7) en 50 ml de tampón de aislamiento de cloroplastos [50 mM Tricina-NaOH pH 8.0,
0.6% polivinil pirrolidona (PVP), 5 mM ácido etilendiaminotetraacético (EDTA), 10
mM 2-mercaptoetanol y 0.3 M manitol] suplementado con 0.1% de albúmina de suero
bovino (BSA) y previamente enfriado a 4ºC. El homogeneizado se filtró a través de
cuatro capas de Miracloth (Calbiochem) y otra de nylon (20 µm) prensando
suavemente, y el filtrado se distribuyó en cuatro tubos, que se centrifugaron a 1000 x g
5 min con un rotor basculante. El sedimento de cada tubo se resuspendió en 1 ml del
mismo tampón empleando un pincel de pelos de oreja de camello, y se cargó sobre un
gradiente discontinuo formado por 10 ml de Percoll (Amersham) 40% y 10 ml de
Percoll 80% en tampón, que se centrifugó a 10000 x g durante 20 min con el mismo
rotor. Los cloroplastos intactos contenidos en la interfase entre ambas capas de Percoll
se recogieron con una pipeta con la punta recortada, y después de diluirlos en tampón se
centrifugaron de nuevo a 1000 x g 5 min (Navarro et al., 1999).
3.1.2. Rotura de los cloroplastos y solubilización de las proteínas
El sedimento de cloroplastos intactos se resuspendió en 4 ml de tampón de
extracción [100 mM Tris-HCl pH 8, 100 mM KCl, 10 mM MgCl2, 1% Nonidet P40 y
10 mM ditiotreitol (DTT) suplementado con una mezcla de inhibidores de proteasas
(incluídas serín, cisteín y metaloproteasas) (Complete, Roche)]. El extracto se agitó
durante 10 min para romper los cloroplastos y solubilizar su contenido, y a continuación
se centrifugó dos veces, primero a 10000 x g durante 15 min para eliminar las
membranas fragmentadas, y después a 100000 x g durante 2 h para eliminar el resto de
elementos insolubles de pequeño tamaño.
51

Material y métodos
3.1.3. Fraccionamiento de proteínas por cromatografía en columna de heparina
El sobrenadante de la ultracentrifugación se cargó en una columna de heparina
(Amersham) de 1 ml previamente equilibrada con 10 ml de tampón de extracción. La
columna se lavó con 10 ml de este mismo tampón y las proteínas se eluyeron con 10 ml
de un gradiente de KCl entre 0.1 y 1 M en tampón de extracción que se obtuvo
mediante un elaborador de gradientes. La cromatografía se llevó a cabo con un flujo de
0.5 ml/min, recogiéndose 20 fracciones.
3.2. Obtención de un extracto de germen de trigo con actividad RNA
ligasa
Aproximadamente 50 g de germen de trigo (Triticum aestivum L.) (Sigma) se
lavaron tres veces con una mezcla de 250 ml de tetracloruro de carbono y ciclohexano
en una proporción 5.5:1. El material recuperado se pesó, y se trituró en un mortero con
N2 líquido hasta obtener un polvo fino al que se le añadieron tres volúmenes de tampón
de extracción enfriado a 0ºC (10 mM Tris-acetato pH 7.6, 4 mM acetato magnésico, 50
mM acetato potásico, 1 mM DTT y la mezcla de inhibidores de proteasas Complete)
tomando como referencia para el volumen del tampón el peso del germen de trigo
lavado (p:v) (Konarska et al., 1981). Después de una centrifugación a 7800 x g durante
15 min para eliminar el material insoluble, el sobrenadante recuperado se sometió a una
nueva centrifugación a 100000 x g durante 2 h. Los ácidos nucleicos de la preparación
se eliminaron mediante precipitación con Polietilenimina P (Sigma) al 0.1%, agitando
con ese fin el extracto durante 15 min y centrifugando posteriormente a 7800 x g 10
min. Al sobrenadante recogido se le añadieron cantidades crecientes de sulfato amónico
hasta alcanzar saturaciones del 20%, 40%, 60% y 80% (Englard y Seifter, 1990). Tras
cada incremento de sulfato amónico el extracto se agitó durante 15 min y se centrifugó a
7800 x g 10 min. Los sedimentos obtenidos en cada paso del fraccionamiento se
resuspendieron en 2 ml de tampón de extracción, se centrifugaron a 12000 x g durante 5
min para eliminar el material no disuelto, y se introdujeron en un dializador con un
límite de masa molecular de 10000 kDa (Pierce). La diálisis se efectuó a lo largo de un
mínimo de 16 h frente a 1 l de tampón de extracción cambiando el tampón dos veces. El
volumen dializado se recogió, se centrifugó a 11000 x g durante 5 min, y se recuperó el
sobrenadante para ensayar su actividad. La muestra de sulfato amónico con mayor
actividad tRNA ligasa (apartados 3.4 a 3.7) se fraccionó cromatográficamente en una
52

Material y métodos
columna de heparina (apartado 3.1.3) y la actividad de ligación se ensayó nuevamente
(apartados 3.4 a 3.7), conservando a -20ºC la fracción más activa añadiéndole 2 g de
glicerol por cada ml de fracción. Todos los pasos de extracción se realizaron a 4ºC.
3.3. Síntesis in vitro de RNA viroidal
Los RNAs viroidales empleados durante este trabajo fueron el viroide del
manchado solar del aguacate (Avocado sunblotch viroid, ASBVd), el viroide del
moteado clorótico del crisantemo (Chrysanthemum chlorotic mottle, CChMVd), el
viroide latente de la berenjena (Eggplant latent viroid, ELVd) y el viroide latente del
melocotonero, que infectan naturalmente, como sus nombres indican, aguacate (Persea
americana Mill.), crisantemo (Dendranthema grandiflora Tzvelez), berenjena (Solanum
melongena L.) y melocotonero (Prunus persica L.), respectivamente. Los plásmidos
pBdASBVd/T3, pBdCChMVd/S, pBdELVd-2/S (apartado 3.19) y pBdPLMVd
(obtenido como se describe en Ambrós et al., 1998) se digirieron con las enzimas de
restricción EcoRI y XbaI (Fermentas) para obtener mediante transcripción in vitro los
RNAs monoméricos de la polaridad deseada. Los cortes con EcoRI (+) o XbaI (-) en
pBdASBVd/T3 y pBdPLMVd, y XbaI (+) o EcoRI (-) en pBdCChMVd/S y pBdELVd-
2/S permiten obtener las polaridades indicadas entre paréntesis. Alícuotas (1 µg) de
estos plásmidos abiertos se incubaron a 37ºC entre 30 min y 2 h en 20 µl de tampón de
transcripción (40 mM Tris-HCl pH 7.9, 6 mM MgCl2, 10 mM DTT, 10 mM NaCl y 2
mM espermidina) suplementado con 500 µM de cada uno de los NTPs, 20 U de
inhibidor de RNasas (Roche) y 10 U de RNA polimerasa (Fermentas) [T7 para
transcribir las polaridades (+) del ASBVd y PLMVd y (-) del CChMVd y ELVd, y T3
para las (-) del ASBVd y PLMVd y (+) del CChMVd y ELVd]. En el caso de
transcripciones marcadas radiactivamente la mezcla de reacción fue la misma, pero en
vez de emplear 500 µM de UTP se utilizaron 40 µCi de [α−32P]UTP (400 Ci/mmol).
Los RNAs viroidales transcritos son diméricos y contienen dos ribozimas de cabeza de
martillo que catalizan el autocorte ribozimático de dicho transcrito, generando RNAs de
diversos tamaños. La RNA polimerasa se inactivó añadiendo 20 µl de tampón de carga
desnaturalizante (10 mM Tris-HCl pH 8, 1 mM EDTA, 0.0025% azul de bromofenol
0.0025% cianol de xileno y 98% formamida) e incubando a 95ºC durante 90 s, y los
RNAs generados se fraccionaron mediante simple electroforesis en geles de
poliacrilamida (polyacrilamide gel electrophoresis, PAGE) desnaturalizante (apartado
53

Material y métodos
3.5). La región que contenía el RNA viroidal monomérico se aisló y se fragmentó en
pequeños cubos de aproximadamente 1 mm3. Tras añadir 500 µl de tampón de
extracción [100 mM Tris-HCl pH 9, 10 mM 2-mercaptoetanol, 10 mM EDTA y 1%
dodecil sulfato sódico (SDS)] y 100 µl de fenol:cloroformo (1:1) (saturando
previamente el fenol con agua y equilibrandolo a pH 8), la mezcla se agitó durante una
noche a temperatura ambiente. Después de centrifugar a 11000 x g 5 min, se recuperó la
fase acuosa, y a la fase orgánica se le añadieron 200 µl de tampón de extracción fresco,
agitando y centrifugando a 11000 x g 5 min. La segunda fase acuosa se combinó con la
primera y se reextrajeron con 200 µl de cloroformo, agitando y centrifugando a 11000 x
g 5 min. Los RNAs en la fase acuosa final se precipitaron con 1 volumen de isopropanol
y 0.1 volumen de 3 M acetato sódico pH 5.5, se recuperaron por centrifugación a 11000
x g 15 min y se resuspendieron en 100 µl de agua, estimándose su concentración por
absorbancia a 260 nm medida con un espectrofotómetro (Biophotometer, Eppendorf).
3.4. Análisis de la actividad RNA ligasa
Las RNA ligasas ensayadas fueron obtenidas tanto por clonación y expresión
heteróloga [T4.2 RNA ligasa, 2’,5’-RNA ligasa de Escherichia coli, tRNA ligasas de
tomate (Solanum lycopersicum L.) y de Arabidopsis (Arabidopsis thaliana (L.) Heynh)]
(apartados 3.16 y 3.17), como purificadas de extractos vegetales [RNA ligasa
cloroplástica de espinaca y tRNA ligasa de trigo] (apartados 3.1 y 3.2). Alícuotas (4 µl)
de diferentes preparaciones de estas RNA ligasas fueron ensayadas en 20 µl de tampón
de ligación [50 mM Tris-HCl pH 7.5, 5 mM DTT, 4 mM MgCl2 y 1 mM ATP (más 5
mM espermidina y 4 µg de RNAs 16S y 23S de E. coli en el caso de la 2’,5’-RNA
ligasa); adaptado de Kikuchi et al., 1982], empleando como sustrato de la ligación 100
ng de RNA monomérico lineal del ASBVd proveniente del autocorte in vitro de un
transcrito dimérico (apartado 3.3). La reacción se incubó a 30ºC durante 30 min (2 min
con la 2’,5’-RNA ligasa) y la enzima se inactivó añadiendo un volumen de tampón de
carga desnaturalizante (apartado 3.3) con 4 µg de RNAs 16S y 23S de E. coli (no
necesarios con la 2’,5’-RNA ligasa, pues se incorporaron con el tampón de reacción), y
calentando a 95ºC 90 s. El papel del RNA heterólogo es evitar tanto la unión de
proteínas del extracto con el sustrato que dificultan su separación y detección como la
degradación del mismo por RNasas contaminantes.
54

Material y métodos
3.5. Fraccionamiento de RNAs en PAGE
Las preparaciones de RNA se mezclaron con un volumen de tampón de carga
desnaturalizante (apartado 3.3) y se calentaron a 95ºC durante 90 s. Una vez enfriadas
en hielo se separaron mediante un gel desnaturalizante de poliacrilamida (5%
poliacrilamida, relación acrilamida:bisacrilamida 37.5:1) de 12 cm de largo, 14 cm de
ancho y 2 mm de grosor. El gel se preparó en tampón TBE (89 mM Tris-HCl, 89 mM
ácido bórico y 2 mM EDTA) con 8 M urea empleándose el mismo tampón sin urea para
los electrodos y aplicándose un voltaje de 200 V durante 3 h aproximadamente
(Sambrook y Russell, 2001). Una vez finalizada la electroforesis, el gel con los ácidos
nucleicos separados por tamaño se agitó durante 15 min en una solución preparada
disolviendo 20 µl de una solución 5 mg/ml de bromuro de etidio en 200 ml de agua, y se
lavó tres veces con agua antes de observar los resultados empleando un transiluminador
de luz ultravioleta (TM-20E, UVP).
Cuando fue necesario un mayor poder de resolución, los RNAs se mezclaron
con tampón de carga (50% glicerol, 10 mM EDTA pH 8, 0.025% azul de bromofenol
0.025% cianol de xileno) en una relación 9:1 (RNA:tampón) y se separaron primero en
un gel no desnaturalizante de poliacrilamida del mismo poro y dimensiones de los ya
descritos. La electroforesis se realizó en tampón TAE (40 mM Tris-acetato, 20 mM
acetato sódico y 1 mM EDTA, pH 7.2) en el gel y los electrodos, aplicando una
intensidad constante de 75 mA durante aproximadamente 90 min. Para revelar el RNA
se hizo una tinción con bromuro de etidio. A continuación, la región del gel que
contenía las formas circulares y lineales del viroide se cortó y se colocó sobre un
segundo gel desnaturalizante del mismo poro y dimensiones que el anterior. El tampón
de los electrodos fue TBE (0.25X) y el del gel el mismo con 8 M urea. Para la
separación de los RNAs se aplicó una intensidad constante de 25 mA durante
aproximadamente 120-150 min, y para revelarlos se tiñó el gel con bromuro de etidio.
Esta segunda electroforesis separa las moléculas circulares y lineales del viroide con
mayor resolución que una simple PAGE desnaturalizante (Schumacher et al., 1983;
Flores et al., 1985).
3.6. Transferencia de RNAs a membranas de hibridación
Para este fin se empleó la modalidad de transferencia semiseca (Amersham). El
gel, la membrana de nylon y el papel absorbente se equilibraron en tampón de
55

Material y métodos
electrotransferencia (25 mM Tris, 192 mM glicina) durante 10 min, se montaron con la
disposición cátodo - papel (3 capas) - gel - membrana - papel (3 capas) - ánodo, y se
aplicó una intensidad de 1 mA por cm2 de membrana durante 1 h. Una vez transferidos,
los RNAs se fijaron a la membrana irradiando con 0.12 J/cm2 de luz ultravioleta
(UVC500, Hoefer).
3.7. Análisis mediante hibridación northern
Para detectar los RNAs viroidales separados mediante doble o simple PAGE y
transferidos a membrana se realizó una prehibridación con 5 ml de solución de
hibridación [50% formamida, tampón SSC (5X) (SSC: 150 mM NaCl y 15 mM citrato
sódico, pH7), 0.1% Ficoll, 0.1% PVP, 1% SDS y 0.01% de DNA de esperma de
salmón] a 70ºC durante 1 h, sustituyéndose esta solución por 5 ml de la misma solución
atemperada a 70ºC. La ribosonda [entre 1x105 y 5x106 cpm, medidas en un contador de
centelleo, (Wallac 1410, Pharmacia)] se diluyó en 100 µl de solución de hibridación, se
desnaturalizó a 95ºC durante 90 s y se mezcló con la solución atemperada a 70ºC. La
hibridación se llevó a cabo a esta temperatura durante 12-14 h, transcurridas las cuales
la membrana se lavó durante 10 min con SSC (2X) y 0.1% SDS a temperatura ambiente
tres veces, y 15 min con SSC (0.1X) y 0.1% SDS a 55ºC una vez. La membrana seca se
reveló con una película autorradiográfica y en ocasiones se cuantificó su señal con un
analizador de imagen (Phosphorimager FLA 5100, Fujifilm).
3.8. Extracción y fraccionamiento de mRNAs de plantas
El material de partida (1 gramo de tejido) se trituró con nitrógeno líquido, se
homogeneizó con tres volúmenes de tampón de extracción (350 mM glicina, 50 mM
NaOH, 350 mM NaCl, 35 mM EDTA, 4% SDS y 14 mM 2-mercaptoetanol) y se
centrifugó a 11000 x g 5 min. El sobrenadante se mezcló con un volumen de
fenol:cloroformo (1:1) (apartado 3.3) y se centrifugó de nuevo a 11000 x g 5 min. Los
RNAs de la fase acuosa se fraccionaron añadiendo 0.25 volúmenes de 10 M LiCl e
incubando a 4ºC durante toda la noche, y el precipitado se recuperó por centrifugación a
11000 x g durante 15 min. El sedimento se resuspendió en 200 µl de agua y se volvió a
precipitar con 2.5 volúmenes de etanol 96% y 0.1 volumen 3 M acetato sódico pH 5.5
durante 2 h. Tras una centrifugación a 11000 x g 15 min, el sedimento se resuspendió en
200 µl de agua, y se determinó su absorbancia a 260 nm. El mRNA se purificó con un
56

Material y métodos
estuche de RNA poli(A) (Sigma) a partir de 500 µg del RNA insoluble en 2 M LiCl.
Este estuche contiene una suspensión de esferas de poliestireno con una cola de poli(dT)
incorporada que hibrida con la cola poli(A) de los mRNAs permitiendo separarlos por
centrifugación. De esta forma las esferas precipitan arrastrando el mRNA unido a las
mismas y separándolo del RNA soluble. En un último paso el RNA poli(A) se recuperó
elevando la temperatura para que el híbrido poli(A)/poli(dT) se desnaturalizara, se
precipitó con etanol y se resuspendió en 25 µl de agua.
3.9. Transcripción inversa (reverse transcription, RT)
El cebador (entre 25 y 50 ng, en función de su longitud) y el RNA molde [entre
10 y 200 ng de RNA poli(A) o entre 1 y 5 µg de RNA total (apartado 3.8)] se mezclaron
en un volumen de 6 µl, calentándose a 98ºC durante 90 s y enfriándose en hielo para
promover el anillamiento entre ambos. El volumen se ajustó a 10 µl en tampón de
transcripción inversa (50 mM Tris-HCl pH 8.3, 75 mM KCl, 3 mM MgCl2 y 2.5 mM
DTT) suplementado con 500 nM dNTPs, 20 U de inhibidor de RNasas y 100 U de
transcriptasa inversa Superscript III (Invitrogen). La mezcla de reacción se incubó a
55ºC 45 min, a 60ºC 10 min y a 65ºC 5 min (en un termociclador Geneamp PCR system
2400, Perkin Elmer). Finalmente, la enzima se inactivó a 70ºC durante 15 min y el RNA
se digirió con 0.5 U de RNasa H (Roche) a 37ºC durante 30 min para evitar
interferencias en reacciones posteriores.
3.10. Reacción en cadena de la polimerasa (polymerase chain reaction,
PCR)
Para esta reacción se emplearon DNA polimerasas termoestables distintas: Tth
(Biotools) de Thermus thermophylus, que no posee actividad correctora de errores, y la
mezcla contenida en el producto Expand high fidelity PCR system (Roche), formada por
Taq (muy procesiva) y Tgo (con actividad correctora de errores) que permite amplificar
productos grandes con una alta fidelidad. El DNA molde, y entre 250 y 500 ng (en
función de su longitud) de cada cebador, se mezclaron en un volumen final de 50 µl de
tampón de reacción [75 mM Tris-HCl pH 9, 50 mM KCl, 2 mM MgCl2 y 20 mM
(NH4)2SO4 para la enzima Tth, y el tampón recomendado por el proveedor para la
mezcla de enzimas Expand high fidelity PCR system] suplementado con 200 µM dNTPs
y 1-5 U de la DNA polimerasa. La reacción se incubó en un termociclador a 94ºC
57

Material y métodos
durante 2 min y luego se sometió a 30 ciclos calentando a 94ºC 40 s para desnaturalizar
el DNA, enfriando a 50-60ºC 30 s para anillar los cebadores y el DNA molde y
calentando a 72ºC durante 2-3 min para elongar las cadenas. La amplificación concluyó
con una extensión final a 72ºC durante 10 min. Cuando fue necesario una segunda PCR
para obtener mayor amplificación del DNA molde, 1 µl del producto de la primera
reacción se tomó como sustrato y otra PCR de 50 µl se preparó en las condiciones
descritas anteriormente.
3.11. Amplificación rápida de los extremos de un cDNA (rapid
amplifications of cDNA ends, RACE)
3.11.1. 3’-RACE
Los mRNAs (100-200 ng) se transcribieron inversamente (apartado 3.9)
empleando un cebador (25-50 ng) con una cola 3’ poli(dT) que hibrida con la cola
poli(A) de los mRNAs, y una secuencia 5’conocida que se incorpora durante la reacción
a los cDNAs generados (Sambrook y Russell, 2001). Estos cDNAs se amplificaron con
la DNA polimerasa termoestable Tth mediante dos PCRs sucesivas (apartado 3.10), la
segunda con cebadores (250-500 ng) más internos que los empleados en la primera. Un
cebador de cada pareja se diseñó complementario a la secuencia conocida del cDNA,
mientras que el otro estaba derivado de la secuencia introducida con el cebador
poli(dT).
3.11.2. 5’-RACE
3.11.2.1. Adición al cDNA de una cola en 3’ con transferasa terminal
El mRNA (100-200 ng) se transcribió inversamente (apartado 3.9) empleando un
cebador específico (25-50 ng), y el cDNA obtenido se precipitó con 2.5 volúmenes de
etanol y 0.1 volúmenes de 3 M acetato sódico pH 5.5 durante 2 h. El precipitado se
resuspendió en 20 µl de tampón de transferasa terminal (25 mM Tris-HCl pH 6.6, 200
mM cacodilato potásico y 0.25 mg/ml BSA) suplementado con 1.25 mM de CoCl2, 0.5
mM de dATP y 800 U de esta enzima (Roche), que cataliza la adición de una cola
poli(dA) en el extremo 3’ del cDNA (Frohman et al., 1988). La reacción se incubó a
37ºC 20 min y se calentó a 70ºC 5 min para inactivar la enzima. El producto obtenido se
precipitó con 2.5 volúmenes de etanol y 0.1 volúmenes de 3 M acetato sódico pH 5.5
58

Material y métodos
durante 2 h, y se resuspendió en 20 µl de agua. La amplificación se llevó a cabo con dos
PCRs sucesivas (apartado 3.10) empleando la DNA polimerasa termoestable Tth. En la
primera reacción, uno de los cebadores fue el mismo que en la RT, mientras que el otro
contenía una cola poli(dT) en 3’ que hibrida con la cola poli(dA) incorporada al cDNA
y una secuencia 5’conocida que se incorpora durante la reacción a los cDNAs
generados. Para la segunda PCR se diseñó un cebador específico interno al empleado en
la RT y otro derivado del cebador con la cola poli(dT). Ambas PCRs se realizaron con
250-500 ng de cada cebador (Schaefer, 1995).
3.11.2.2. Amplificación rápida de los extremos de un cDNA mediada por RNA
ligasa (RNA ligase-mediated rapid amplification of cDNA ends, RLM-RACE)
Los mRNAs (100-200 ng) se desfosforilaron incubando a 50ºC 30 min con 10 U
de fosfatasa alcalina intestinal bovina (Roche) en 50 µl de tampón de reacción (50 mM
Tris-HCl pH 8.5, 0.1 mM EDTA) con 5 U de inhibidor de RNasas. La enzima se
inactivó calentando a 95ºC 90 s, y una vez enfriada la reacción se añadieron
nuevamente 5 U de inhibidor de RNasas y 10 U de la enzima incubándose a 50ºC
durante otros 30 min. Esta reacción genera extremos 5’-hidroxilo excepto en aquellos
mRNAs con estructura de caperuza. El RNA se extrajo una vez con fenol:cloroformo
(apartado 3.3) y otra con cloroformo, y se precipitó con 2.5 volúmenes de etanol y 0.1
volúmenes de 3 M acetato sódico pH 5.5 durante 2 h (este tratamiento de
desproteinización se repitió tras cada reacción enzimática). Una vez recuperado el RNA
por centrifugación a 11000 x g 15 min, se trató con la pirofosfatasa ácida de tabaco
(tobacco acid pyrophosphatase, TAP, Epicentre) para eliminar la estructura de
caperuza, dejando extremos 5’-fosfomonoéster sólo en aquellos mRNAs con el extremo
5’ intacto (los que tenían la estructura de caperuza). La reacción se realizó en 10 µl de
tampón de TAP (50 mM acetato sódico pH 6.0, 1 mM EDTA, 0.1% β-mercaptoetanol y
0.1% Triton X-100) con 5 U de inhibidor de RNasas y 5 U de la enzima a 37ºC 60 min.
A los RNAs que poseían el extremo 5’-fosfomonoéster se les unió un adaptador de
RNA de secuencia conocida (250 ng) en una reacción (30 µl) en tampón de T4.1 RNA
ligasa (50 mM HEPES-NaOH pH 8.0, 10 mM MgCl2 y 10 mM DTT) suplementado con
1 mM ATP y 15 U de esta enzima (Epicentre), e incubando a 37ºC durante 60 min (Liu
y Gorovsky, 1993). El RNA así obtenido se empleó como molde para una RT (apartado
3.9) empleando un cebador específico (25-50 ng). Una alícuota de esta reacción se
59

Material y métodos
amplificó por PCR con este mismo cebador y otro complementario a la secuencia del
adaptador de RNA ligado al extremo 5’. Una segunda PCR se realizó con cebadores
más internos para intentar aumentar la concentración de los productos de amplificación.
Para las reacciones de PCR se emplearon 250-500 ng de cada cebador.
3.11.2.3. Selección y amplificación de mRNAs con estructura de caperuza mediante
un adaptador de DNA (Cap-finder)
La transcriptasa inversa del virus de la leucemia murina de Moloney y sus
derivadas a las que se les ha eliminado la actividad RNasa H (denominadas
comercialmente Superscript), pueden introducir hasta cuatro residuos C en el extremo
3’ del cDNA cuando alcanzan la estructura de caperuza del molde de mRNA (Schimdt
y Mueller, 1999; Schramm et al., 2000). Para la transcripción inversa se emplearon 100-
200 ng de mRNA, un cebador específico de secuencia (25-50 ng) y un adaptador de
DNA (50-100 ng) con una cola poli(dG) en 3’ que, al hibridar con la cola 3’ poli(dC)
del cDNA naciente, permite a la transcriptasa inversa continuar la elongación sobre la
secuencia del adaptador adyacente a las cuatro Gs. La reacción se llevó a cabo con
transcriptasa inversa Superscript III en un tampón adaptado para introducir hasta cuatro
Gs (50 mM Tris-HCl pH 8.3, 6 mM MgCl2, 2 mM MnCl2, 10 mM DTT, 0.2 mg/ml
BSA y 1 mM dNTPs) a una temperatura de 45ºC durante 60 min (Schramm et al.,
2000), seguida de una inactivación de la enzima a 70ºC 15 min y digestión del RNA con
0.5 U de RNasa H a 37ºC 30 min. El cDNA así obtenido se amplificó por PCR con un
cebador específico y con otro complementario a la secuencia introducida en 3’ derivada
del adaptador. Una segunda PCR se realizó con cebadores más internos, empleando en
ambas reacciones 250-500 ng de cada cebador.
3.11.2.4. PCR con cebadores degenerados
Los mRNAs se transcribieron inversamente (apartado 3.9) empleando un
cebador específico, y el cDNA obtenido se amplificó con este mismo cebador y otro
degenerado diseñado partir de una secuencia cercana al extremo 5’ de la región
codificante muy conservada en los supuestos genes homólogos. Una segunda PCR se
realizó con cebadores más internos (Sambrook y Russell, 2001).
60

Material y métodos
3.11.3. RACE circular
3.11.3.1. cDNA molde de doble cadena
El cDNA se generó con una RT (10 µl) (apartado 3.9) empleando un cebador
específico (25-50 ng) previamente fosforilado (apartado 3.13.1) y 100-200 ng de
mRNA. La segunda cadena de este cDNA se sintetizó siguiendo un protocolo descrito
por Okayama y Berg (1982). La reacción se llevó a cabo en 80 µl de tampón (20 mM
Tris-HCl, pH 7.5, 100 mM KCl, 10 mM (NH4)2SO4, 5 mM MgCl2, 150 µM β-NAD y
0.05 mg/ml BSA) suplementado con 250 µM dNTPs, 1 U de RNasa H, 25 U de DNA
polimerasa I (Fermentas) y 5 U de DNA ligasa E. coli (New England Biolabs), e
incubando a 16ºC durante 90 min. En esta mezcla, la RNasa H genera cortes en el
mRNA, y los fragmentos resultantes sirven como cebadores para la síntesis de la doble
cadena de cDNA por la DNA polimerasa I. Las mellas que quedan en la segunda cadena
de cDNA como consecuencia de los cortes que introduce la RNasa H en el mRNA son
selladas por la DNA ligasa de E. coli, dejando romos los extremos del cDNA de doble
cadena resultante mediante incubación a 16ºC 45 min con 10 U de T4 DNA polimerasa
(Fermentas). El producto de la reacción se extrajo una vez con fenol:cloroformo
(apartado 3.3) y otra con cloroformo, y el cDNA de la fase acuosa se precipitó con 2.5
volúmenes de etanol y 0.1 volúmenes de 3 M acetato sódico pH 5.5 durante 2 h. Tras
centrifugar a 11000 x g 15 min, el cDNA sedimentado se resuspendió en agua y se
autoligó en una reacción (50 µl) en tampón de T4 DNA ligasa (40 mM Tris-HCl pH 7.8,
10 mM MgCl2, 10 mM DTT y 500 µM ATP) con 5 U de esta enzima (Fermentas). La
reacción se incubó a 37ºC 2 h, y los productos resultantes se purificaron con un estuche
de minicolumnas (Zymo), empleándose a continuación como molde en dos PCR
consecutivas con dos parejas de cebadores específicos divergentes y la DNA polimerasa
termoestable Tth (apartado 3.10).
3.11.3.2. cDNA molde de simple cadena
Con un cebador específico (25-50 ng) previamente fosforilado (apartado 3.13.1)
y 100-200 ng de mRNA se llevó a cabo una RT en un volumen final de 10 µl (apartado
3.9). Una vez finalizada la reacción, el mRNA se digirió con RNasa H durante 60 min,
y el cDNA de simple cadena generado se autoligó con 5-25 U de T4.1 RNA ligasa,
aumentando el volumen de la reacción a 100 µl (para favorecer la circularización) con el
tampón apropiado para esta enzima (apartado 3. 11.2.2) y 1 mM ATP, e incubando a
61

Material y métodos
16ºC durante 16 h (Snopek et al., 1976; McCoy y Gumport, 1980). El producto
resultante se precipitó con 2.5 volúmenes de etanol y 0.1 volúmenes de 3 M acetato
sódico pH 5.5 durante 2 h, y tras centrifugar a 11000 x g 15 min, el sedimento se
resuspendió en 20 µl de agua. Empleando este cDNA circularizado se llevaron a cabo
dos PCR consecutivas con dos parejas de cebadores específicos divergentes y la DNA
polimerasa termoestable Tth (apartado 3.10). Esta técnica permite amplificar fragmentos
de 500-900 pares de bases. Para progresar en la determinación de las secuencias del
mRNA se emplearon los datos obtenidos en un primer experimento de RACE circular
para diseñar nuevos cebadores con los que se repitió el procedimiento hasta alcanzar el
extremo 5’ de dicho mRNA.
3.11.4. Análisis de los productos obtenidos en los experimentos de RACE
Los productos de la amplificación se separaron electroforéticamente en un gel de
agarosa (apartado 3.12) y, tras tinción con bromuro de etidio (apartado 3.5) se eluyó el
fragmento de interés con un estuche de minicolumnas. Dicho fragmento se introdujo en
el plásmido pTZ57R/T (Fermentas) ligándolo durante 16 h en una mezcla de reacción
(10 µl) con 25 pg de plásmido abierto, tampón de T4 DNA ligasa (apartado 3.11.3.1) y
5 U de esta enzima. Este plásmido está abierto por el sitio EcoRV y posee una T
protuberante en el extremo 3’ de cada cadena que puede hibridar con la A que algunas
DNA polimerasas termoestables introducen en el extremo 3’ del DNA amplificado. Esta
ligación entre extremos adhesivos facilita la incorporación del inserto en el vector. El
plásmido recombinante se purificó con el mismo estuche de minicolumnas y se
electroporó (en un aparato ECM399, BTX) a una suspensión de células competentes de
E. coli DH5α. Las células transformadas se crecieron en una placa con medio Luria-
Bertani (LB) sólido (1% triptona, 0.5% extracto de levadura, 1% NaCl, 1.5% agar)
suplementado con ampicilina (50 µg/ml) (LBA) y 5-bromo-4-cloro-3-indolil-beta-D-
galactopiranósido (X-Gal). Este último se disolvió previamente a una concentración de
50 mg/ml en dimetil formamida, y 30 µl de esta solución se distribuyeron
homogéneamente sobre la superficie de la placa antes de sembrar la suspensión
bacteriana. Tras crecer las células transformadas, las colonias blancas (el plásmido
pTZ57R/T permite selección por color) se aislaron y crecieron en medio LBA líquido
(1% triptona, 0.5% extracto de levadura, 1% NaCl, 50 µg/ml de ampicilina), y el DNA
plasmídico de las mismas se purificó empleando un estuche de minipreparaciones
62

Material y métodos
(Roche). La secuencia de los insertos en ambas orientaciones se obtuvo con un
secuenciador capilar automático (ABI 3100, Applied Biosystems).
3.12. Electroforesis en geles de agarosa
Para separar el DNA en función de su tamaño se añadió a las muestras tampón
de carga (apartado 3.5) y se depositaron sobre un gel de agarosa del 1% preparado en
tampón TAE (apartado 3.5) y con unas dimensiones de 8 cm de largo, 6 cm de ancho y
un grosor aproximado de 0.5 cm. El tampón de los electrodos fue también TAE,
aplicándose un voltaje de 75 V durante aproximadamente 1 h (Sambrook y Russell,
2001), y una vez finalizada la electroforesis, se tiñó el gel con bromuro de etidio para
ver la migración de los DNAs (apartado 3.5).
3.13. Modificaciones de los ácidos nucleicos
3.13.1. Fosforilación
El DNA o RNA sustrato (25-500 ng) se incubó a 37ºC entre 30 y 60 min en 50
µl de tampón de fosforilación (50 mM Tris-HCl pH 7.6, 10 mM MgCl2, 5 mM DTT,
100 µM espermidina y 100 µM EDTA) suplementado con 400 µM ATP y 10 U de T4
polinucleótido quinasa (PNK, Fermentas) (Richardson, 1965). La inactivación de la
enzima se realizó calentando la mezcla de reacción a 65ºC durante 3 min. Los ácidos
nucleicos se recuperaron por precipitación con 2.5 volumen de etanol y 0.1 volúmenes
de 3 M acetato sódico pH 5.5 (más 10 µM MgCl2 y 0.5 µl de glicógeno en el caso de
precipitar cebadores) y se resuspendieron en 10 µl de agua.
3.13.2. Desfosforilación
3.13.2.1. Extremos 5’ y 3’
El sustrato (entre 100 y 500 ng) y 1 U de fosfatasa alcalina de gamba (Roche) se
mezclaron en un volumen de 40 µl en tampón de desfosforilación (50 mM Tris-HCl pH
8.5 y 5 mM MgCl2), y se incubaron a 37ºC entre 10 y 60 min. La inactivación de la
enzima se realizó calentando la mezcla de reacción a 65ºC durante 15 min y los
productos de la reacción se precipitaron con 2.5 volumen de etanol y 0.1 volúmenes de
3 M acetato sódico pH 5.5, resuspendiéndose en 10 µl agua.
63

Material y métodos
3.13.2.2. Extremos 2’,3’-fosfodiéster cíclico
El sustrato (100-500 ng) y 10 U de T4 PNK se mezclaron en un volumen de 20
µl en tampón 100 mM Tris-HCl pH 6.5, 50 mM MgCl2 y 10 mM DTT, y se incubaron a
37ºC durante 1 h (Cameron y Uhlenbeck, 1977). La inactivación de la enzima se realizó
calentando la mezcla de reacción a 65ºC durante 3 min y los ácidos nucleicos se
precipitaron con 2.5 volumen de etanol y 0.1 volúmenes de 3 M acetato sódico pH 5.5,
resuspendiéndose en 10 µl de agua.
3.14. Extensión del cebador
Para la detección de enlaces anómalos en una molécula de RNA se diseñó un
cebador que hibridara con dicho RNA. Entre 1 y 2.5 µg de cebador (en función de la
longitud) se purificaron en una simple PAGE desnaturalizante (apartado 3.5) del 15 %
de poliacrilamida y 1 mm de grosor aplicándose un voltaje de 200 V durante 2.5 h
aproximadamente. La elución se hizo como se relata en el apartado 3.3, y el cebador se
precipitó durante toda la noche con 1 volumen de isopropanol, 0.1 volumen de 3 M
acetato sódico pH 5.5, 10 mM MgCl2 y 0.5 µg de glicógeno. Una vez resuspendido en
10-25 µl de agua, se estimó su concentración por absorbancia a 260 nm medida con un
espectrofotómetro. Entre 0.5 y 1 µg de cebador se fosforilaron en 10 µl de tampón de
fosforilación (apartado 3.13.1), con 20 µCi de [γ−32P]ATP (3000 Ci/mmol) y 10 U de
T4 PNK, incubando a 37ºC 30 min. El cebador fosforilado se precipitó con 2.5
volúmenes de etanol, 0.1 volúmenes de 3 M acetato sódico pH 5.5, 10 mM MgCl2 y 0.5
µg de glicógeno durante toda la noche, y se resuspendió en un volumen entre 5 y 20 µl
de agua. El cebador resuspendido (200-400 ng) y el sustrato (25-100 µg) se mezclaron
en un volumen de 10 µl de tampón de transcripción inversa (apartado 3.9) suplementado
con 500 nM de cada dNTP, 20 U de inhibidor de RNasas y 100 U de transcriptasa
inversa Superscript III. La mezcla se incubó a 55ºC 45 min, a 60ºC 10 min y a 65ºC 5
min, y la enzima se inactivó a 70ºC 15 min. Al finalizar la reacción se añadieron 20 µl
de tampón de carga desnaturalizante (apartado 3.3) y se desnaturalizó el cDNA obtenido
incubando a 95ºC durante 90 s. Una alícuota de 15 µl se fraccionó mediante simple
PAGE desnaturalizante (apartado 3.5) en un gel del 5% de poliacrilamida aplicando un
voltaje de 200 V durante aproximadamente 100 min. El gel se secó durante 2 h (en un
aparato MGD-5040, VWR) y se reveló con una película autorradiográfica (Sambrook y
Russell, 2001).
64

Material y métodos
3.15. Detección de enlaces anómalos en el RNA por cromatografía en
capa fina (thin layer chromatography, TLC)
El RNA monomérico lineal (+) del ASBVd se obtuvo disolviendo 10 µg del
plásmido pBdASBVd/T3 linealizado con EcoRI en 200 µl tampón de transcripción
suplementado con 500 µM de cada uno de los NTPs (excepto UTP), 100 µCi de [α-32P]UTP (400 Ci/mmol), 80 U de inhibidor de RNasas y 100 U de T7 RNA polimerasa,
e incubando a 37ºC durante 2 h (para más detalles ver apartado 3.3). La reacción se
dividió en tres partes, una de 10 µl que se reservó para purificar el RNA monomérico
lineal, y dos de 95 µl cuyo RNA se ligó alternativamente con la RNA ligasa
cloroplástica de espinaca o con la 2’,5’-RNA ligasa de E. coli. La reacción con la RNA
ligasa cloroplástica se llevó a cabo en un volumen final de 200 µl de tampón de ligación
con 40 µl de enzima e incubando a 30ºC 30 min, mientras que la reacción con la 2’,5’-
RNA ligasa se llevó a cabo en este mismo volumen final de tampón de ligación con 40
µl de enzima más 5 mM espermidina y 40 µg de RNAs 16S y 23S de E. coli, e
incubando a 30ºC 2 min (para más detalles ver apartado 3.4). Los productos resultantes
de las reacciones se precipitaron con 2.5 volúmenes de etanol, 0.1 volúmenes de 3 M
acetato sódico pH 5.5 y 7.5 µg de glicógeno en el caso de la muestra ensayada con la
2’,5’-RNA ligasa de E. coli, y 2.5 volúmenes de etanol, 0.1 volúmenes de 3 M acetato
sódico pH 5.5, 7.5 µg de glicógeno y 40 µg de RNAs 16S y 23S de E. coli en la muestra
ensayada con la RNA ligasa cloroplástica, resuspendiéndose los precipitados en 40 µl
de tampón de carga (apartado 3.5). Las muestras se fraccionaron mediante doble PAGE
(apartado 3.5), y los RNAs monoméricos circulares se eluyeron de la correspondiente
región del segundo gel (apartado 3.3). Paralelamente, la alícuota no tratada proveniente
de la transcripción inicial se mezcló con un volumen de tampón de carga
desnaturalizante y los RNAs se fraccionaron mediante simple PAGE desnaturalizante
(apartado 3.5), eluyéndose el RNA monomérico lineal (apartado 3.3). Los RNAs
eluidos se recuperaron por precipitación con 2.5 volúmenes de etanol, 0.1 volúmenes de
3 M acetato sódico pH 5.5, y 7.5 µg de glicógeno, y la radiactivivad del precipitado se
midió con un contador de centelleo (Wallac 1410, Pharmacia). La muestra con menor
emisión se resuspendió en 5 µl de agua, y a las otras dos se les añadió un volumen de
agua adecuado para que las tres tuvieran una radiactividad (cpm) por µl equivalente.
Alícuotas de 5 µl de las tres muestras se digirieron añadiendo 0.6 µl de tampón de
nucleasa S1 (400 mM acetato sódico pH 4.5, 300 mM NaCl y 2 mM ZnSO4) y 40 U de
65

Material y métodos
esta enzima (Fermentas), e incubando a 37ºC 30 min. Los nucleótidos y dinucleótidos
así obtenidos se extrajeron con 10 µl de cloroformo, agitando y centrifugando a 11000 x
g 5 min. Este paso se repitió una vez, y 5 µl de la fase acuosa se recuperaron para
separar su contenido por TLC.
Alícuotas (entre 1 y 5 µl) de las muestras resultantes de las digestiones
enzimáticas y los patrones correspondientes se aplicaron puntualmente a 2 cm del borde
de una capa de celulosa fijada a un soporte de cristal. La placa se introdujo en una
cubeta cromatográfica con aproximadamente 150 ml de eluyente (0.1 M fosfato sódico
pH 6.8, sulfato amónico y n-propanol en una proporción 100:60:2 v:p:v), y se dejó que
el mismo ascendiera por capilaridad hasta alcanzar el borde opuesto de la placa (de 12 a
15 h) (Silberklang et al., 1979). Una vez extraída de la cubeta, la placa se secó y los
productos separados se revelaron mediante iluminación UV cenital y aplicación de una
película autorradiográfica.
Alternativamente se obtuvo el RNA monomérico circular (+) del ASBVd en
ausencia de marcaje radiactivo, para lo cual se repitió el procedimiento anterior
empleando UTP no marcado. Los RNAs circulares y lineales purificados se digirieron
en 20 µl de tampón para RNasa ONE (10 mM Tris-HCl pH 7.5, 5 mM EDTA y 200
mM acetato sódico) con 10 U de esta enzima (Promega), e incubando a 37ºC 16 h. 5 U
de esta enzima se añadieron nuevamente, y se incubó la reacción a 37ºC 2 h. El agua de
la muestra se evaporó en un aparato de vacío (DNA 120, Savant), y el precipitado se
resuspendió en 20 µl de tampón de fosforilación (apartado 3.13.1), suplementado con 5
µCi de [γ-32P]ATP (3000 Ci/mmol) y 10 U de T4 PNK. La reacción se incubó a 37ºC 30
min, y el exceso de ATP marcado se agotó añadiendo glucosa hasta una concentración
final de 2 mM y 0.01 U de hexoquinasa (Sigma). La reacción se incubó a 37ºC 10 min,
y transcurrido este tiempo se añadieron 2.5 nmol de ATP, incubándose otros 10 min a la
misma temperatura. Este paso se repitió una vez, y los nucleótidos y dinucleótidos
producto de la digestión se extrajeron con 25 µl de cloroformo, agitando y
centrifugando a 11000 x g 5 min. Tras repetir de nuevo la extracción, la fase acuosa (20
µl) se recuperó, y se le añadió una solución de acetato amónico pH 5.3 hasta alcanzar
una concentración final de 75 mM, y 2 µg de nucleasa P1 (Sigma) previamente
disueltos en 75 mM acetato amónico pH 5.3. La reacción se incubó a 37ºC durante 3 h y
se aplicó en una TLC.
En esta ocasión fue necesaria una mayor capacidad de resolución de las
muestras, por lo que se llevó a cabo una separación bidimensional. Para este cometido,
66

Material y métodos
una alícuota de 4 µl se aplicó puntualmente a 2 cm del borde de una capa de celulosa
fijada a un soporte de cristal. La placa se introdujo en una cubeta cromatográfica con
150 ml de eluyente apolar (98% ácido isobutírico, 30% NH4OH y agua en una relación
volumétrica 66:1:33) y se dejó que el eluyente ascendiera por capilaridad hasta alcanzar
el borde superior de la placa (aproximadamente 12 h). La placa se secó un mínimo de 6
h en una campana de extracción de gases y se introdujo girada 90º en sentido contrario a
las agujas del reloj en una segunda cubeta cromatográfica con 150 ml de eluyente polar
(0.1 M fosfato sódico pH 6.8, sulfato amónico y n-propanol en una proporción 100:60:2
v:p:v), dejando que ascendiera por capilaridad (de 12 a 15 h) hasta alcanzar el borde
superior de la placa (Silberklang et al., 1979). Tras secarla, los productos separados se
revelaron mediante iluminación UV cenital y aplicación de una película
autorradiográfica.
3.16. Obtención de construcciones para expresar proteínas en E. coli
Rosetta 2
El cDNA que codifica las proteínas expresadas se obtuvo por RT (apartado 3.9)
y se amplificó por PCR (apartado 3.10) con cebadores específicos y la mezcla de DNA
polimerasas termoestables Expand high fidelity PCR system. Los cebadores se diseñaron
de forma que se eliminara el codón de parada de la secuencia y se incluyeran sitios de
restricción apropiados durante la PCR. El cDNA amplificado y digerido con las enzimas
de restricción adecuadas se introdujo en los plásmidos de expresión pET23a (+) o
pET23d (+) (Novagen) conservando la pauta de lectura y en fase con la secuencia del
vector que codifica seis histidinas a partir del último aminoácido de la proteína. Las
ligaciones inserto:vector se llevaron a cabo en una proporción molar 10:1 en 10 µl de
tampón de ligación de T4 DNA ligasa con 5 U de esta enzima e incubando a 16ºC toda
la noche.
3.17. Expresión de proteínas en E. coli Rosetta 2
3.17.1. Inducción
Las colonias de E. coli Rosetta 2 (Novagen) transformadas con las
construcciones a expresar se inocularon en un matraz de 500 ml de LB líquido con
ampicilina y cloranfenicol (50 y 34 µg/ml respectivamente). El cultivo se creció con
67

Material y métodos
agitación hasta alcanzar una densidad óptica aproximada de 0.6 a 600 nm, momento en
el que se añadió isopropil-β-D-tiogalactopiranósido (IPTG) hasta una concentración
final de 400 µM, incubándose a continuación el cultivo a 25ºC entre 2 y 3 h.
3.17.2. Extracción
Las células inducidas se recogieron mediante centrifugación a 9600 x g durante
15 min y se resuspendieron en 5 ml de agua con una mezcla concentrada (una pastilla
en 12.5 ml) de inhibidores de proteasas Complete. Para romper su membrana celular, las
células se congelaron a -20ºC durante 1 h, y una vez descongeladas se digirió el DNA
añadiendo 200 U de DNasa I (Fermentas) en tampón 50 mM Tris-HCl pH 7.5, 1%
Nonidet P40, 2.5 mM MgCl2 y 10 mM 2-mercaptoetanol, y agitando con rotación a 4ºC
durante 45 min. Para evitar interacciones inespecíficas entre las proteínas durante el
paso de purificación (apartado 3.17.3) se añadió NaCl hasta una concentración final de
500 mM. El volumen del extracto se ajustó a 10 ml con agua y tras una incubación con
rotación a 4ºC durante 15 min, se ultracentrifugó a 90000 x g durante 30 min.
3.17.3. Purificación
Las proteínas expresadas se purificaron mediante una cromatografía de afinidad
con iones metálicos inmovilizados. Para este cometido se empleó la resina Ni-NTA
agarosa (Qiagen), donde los átomos de Ni se complejan con ácido nitrilotriacético
(nitrilotriacetic acid, NTA), que ocupa cuatro de los seis enlaces de coordinación que
posee este metal (Porath et al., 1975; Sulkowski, 1985). A su vez, el NTA se encuentra
unido a agarosa, facilitando así la manipulación. El plásmido de expresión (apartado
3.16) permite la adición al extremo carboxilo terminal de la proteína de seis histidinas
(6xHis), que a un pH en torno a 8 se unen con gran afinidad a los dos enlaces de
coordinación libres del Ni. De esta forma, las proteínas que posean dicha cola 6xHis son
retenidas. Para eluirlas se empleó imidazol, un precursor de la histidina muy similar
estructuralmente que compite por el Ni. Un volumen de 500 µl de Ni-NTA agarosa se
equilibró con 1.25 ml de tampón de lavado (50 mM Tris-HCl pH 7.5, 500 mM NaCl,
1% NP40, 10 mM 2-mercaptoetanol y 20 mM imidazol), agitando durante 5 min y
centrifugando a 150 x g otros 5 min. Este paso se repitió una vez, y tras decantar el
tampón de lavado, se añadió el sobrenadante de la ultracentrifugación (apartado 3.17.2)
previamente suplementado con 20 mM imidazol pH 8 (ajustado con HCl). Esta
68

Material y métodos
concentración de imidazol evita las interacciones inespecíficas de las proteínas con el
Ni. Tras incubación a 4ºC durante 1 h, la mezcla se transfirió a una columna
cromatográfica, y tras percolar la fase móvil, la fase estacionaria se trató tres veces con
2.5 ml de tampón de lavado, y la proteína unida se eluyó en cinco fracciones de 250 µl
de tampón de elución (500 mM imidazol en 0.5X tampón de lavado).
3.18. Electroforesis de proteínas en geles de poliacrilamida
desnaturalizantes (SDS-PAGE)
Las preparaciones de proteínas fueron analizadas por electroforesis en geles de
poliacrilamida de porosidad variable (entre 7.5% y 15% de poliacrilamida, relación
acrilamida:bisacrilamida 37.5:1) en función del tamaño molecular de los componentes a
separar (Laemmli, 1970). El gel resolutivo, de 12 cm de largo, 14 de ancho y 1 mm de
grosor se preparó en tampón 375 mM Tris-HCl pH 8.8 y 0.05% SDS, mientras que para
apilar las proteínas se empleó un gel del 5% en tampón 125 mM Tris-HCl pH 6.8 y
0.05% SDS. El tampón de los electrodos consistió en 50 mM Tris, 384 mM glicina y
0.05% SDS. Las proteínas se desnaturalizaron añadiendo tampón de carga (300 mM
Tris-HCl pH 6.8; 10% SDS, 500 mM DTT, 60% glicerol y 0.05% azul de bromofenol)
en una relación 4:1 (proteína:tampón) e incubando a 95ºC durante 90 s, y a continuación
se separaron aplicando 200 V al gel durante aproximadamente 2.5 h.
Tras la electroforesis, el gel de poliacrilamida se fijó durante 1 h con una
solución de 10% ácido acético y 40% metanol. Tras retirar el fijador, el gel se lavó con
agua durante 10 min, repitiéndose este paso dos veces más. A continuación se añadió la
solución de azul de Coomassie coloidal, consistente en una parte de metanol y cuatro
partes de una solución de 10% sulfato amónico, 1% ácido fosfórico y 0.1% azul de
Coomassie G250 (dispersado previamente en agua) (Neuhoff et al., 1988). Tras 16 h, el
gel se lavó con agua atemperada a 45-55ºC dos o tres veces hasta alcanzar una relación
de contraste adecuada entre las bandas de proteína y el fondo. Todos los pasos se
llevaron a cabo con agitación orbital.
Cuando fue necesaria una mayor sensibilidad, el gel de poliacrilamida se fijó
durante 30 min con una solución de 10% ácido acético y 50% metanol, se rehidrató 15
min en 5% metanol y se lavó con agua durante 5 min, repitiendo este último paso dos
veces más. Una vez descartada el agua del último lavado, se añadió una solución de
0.02% Na2S2O3 y se incubó durante 60 s, retirándola rápidamente y lavando con agua
69

Material y métodos
durante 30 s tres veces. Tras este lavado, el gel se trató con una solución de 0.2%
AgNO3 durante 25 min, y tras tres lavados con agua de 60 s cada uno, se reveló con 3%
Na2CO3, 0.02% HCOH y 2% de la solución de Na2S2O3 preparada anteriormente
(0.02%). Una vez alcanzado el nivel de detección adecuado, se retiró esta última
solución y se añadió otra 1.4% EDTA para parar la reacción, manteniéndola 10 min y
lavando a continuación con agua (Blum et al., 1987). Todos los pasos se llevaron a cabo
con agitación orbital.
3.19. Construcciones para expresar RNAs viroidales en el cloroplasto
de Chlamydomonas reinhardtii
Para expresar en cloroplastos de Chlamydomonas (Chlamydomonas reinhardtii
Dang.) los RNAs del ASBVd, del CChMVd, del ELVd y del viroide de la exocortis de
los cítricos (Citrus exocortis viroid, CEVd) se generaron construcciones insertando el
correspondiente cDNA viroidal en el vector de transformación cloroplástico pCrc+157
(Suay et al., 2005) a través de tres pasos de clonación. En primer lugar se digirió con
enzimas de restricción adecuadas el cDNA dimérico viroidal contenido en plásmidos
derivados de pBluescript II KS (+) (Stratagene). Dichos cDNAs diméricos tienen una
orientación cabeza cola, donde la secuencia de ambos monómeros es consecutiva y con
la misma orientación. pBdASBVd contiene la secuencia dimérica del cDNA del
ASBVd, comenzando en la posición 154 y finalizando en la 153 para la polaridad (+), y
70/69 (comienzo y final) para la (-). pBdELVd-2 contiene la secuencia dimérica del
ELVd (variante 2) (210/209), pBdCChMVd la del CChMVd (295/294) y pBdCEVd la
del CEVd (40/39) [los números mostrados entre paréntesis indican la posición
inicial/final de la polaridad (+)]. Los cDNAs diméricos viroidales se digirieron con las
enzimas de restricción (todas suministradas por Fermentas) SpeI/SmaI (pBdASBVd),
BamHI/PstI (pBdCChMVd), ClaI/EcoRI (pBdELVd-2) y EcoRI/SpeI (pBdCEVd), y los
extremos del fragmento se hicieron romos con la nucleasa de la judía mungo [Vigna
radiata (L.) Wilczek] (New England Biolabs) (Kowalski et al., 1976; Kroeker et al.,
1976). El fragmento resultante se ligó en el sitio SmaI del plásmido pBluescript II KS
(+) con T4 DNA ligasa y, tras purificar los productos de la reacción con un estuche de
minicolumnas los plásmidos recombinantes se electroporaron a una suspensión de
células de E. coli DH5α competentes. Los DNAs plasmídicos de las colonias
transformadas se extrajeron con un estuche de minipreparaciones, y se seleccionaron
70

Material y métodos
mediante ensayos de restricción aquellos con el inserto en la orientación adecuada. Los
nuevos plásmidos se nombraron pBdASBVd/T3, pBdELVd-2/S, pBdCChMVd/S y
pBdCEVd/S. El segundo paso de clonación consistió en introducir el cDNA dimérico
viroidal extraído con las enzimas EcoRI y XbaI en el plásmido pBrbcL, un derivado de
pBluescript II KS (+) que contiene el promotor, la región 5’ no traducible (5’-UTR) y
los 70 primeros nucleótidos de la pauta abierta de lectura (open reading frame, ORF)
del gen rbcL de C. reinhardtii (entre -70 y +157, indicando +1 el inicio de la
transcripción). Las construcciones obtenidas se electroporaron a una suspensión de
células de E. coli DH5α competentes y mediante ensayos de restricción se seleccionaron
aquellas con el inserto adecuado. Finalmente el fragmento NheI-XbaI (que contenía el
cDNA dimérico viroidal precedido por la secuencia reguladora de rbcL) se transfirió a
pCrc+157. Este plásmido derivado de pUC8 también contiene la región reguladora de
rbcL, una copia del gen de la β-glucuronidasa (GUS), la región 3’ no traducible del gen
psaB y otras características necesarias para transformar C. reinhardtii. En pCrc+157, el
cDNA dimérico viroidal y la región reguladora de rbcL introducidos substituyen dicha
región reguladora del propio plásmido y el gen GUS. El cDNA dimérico viroidal se
subclona en pBrbcL porque pCrc+157 no posee ningún sitio de restricción adecuado
para incorporar la construcción directamente. En la construcción final, el cDNA
dimérico viroidal quedó precedido por el promotor, la región 5’ no traducible y el
comienzo de la pauta abierta de lectura del gen rbcL (entre -70 y +157), y seguido por la
región 3’ no traducible del gen psaB.
3.20. Transformación y crecimiento de cultivos de C. reinhardtii
La cepa mutante CC-373, que posee una deleción parcial del gen atpB que la
convierte en fotosintéticamente inactiva (obtenida del Chlamydomonas Genetics Center
en Duke University, NC, USA), se creció a temperatura ambiente en condiciones de
baja luminosidad (0.05 µmol/s·m2) en un medio con alta concentración de sales (high
salts, HS) suplementado con 2.5 g/l de acetato potásico (high salts acetate, HSHA)
(Sueoka, 1960). Para la transformación del mutante CC-373, se recogieron las células
cultivadas en 500 ml de medio HSHA durante dos días y se resuspendieron en medio
HSHA con 0.1% de agar. La suspensión se sembró sobre placas Petri de HSHA con
1.5% de agar, y se bombardeó con microproyectiles de oro de 0.6 µm (sobre los que se
había precipitado previamente las construcciones del DNA viroidal) empleando un
71

Material y métodos
aparato (PDS-1000/He, BioRad) con una presión de helio de 1550 psi (Boyton et al.,
1988). Las células se mantuvieron 24 h en oscuridad, y los transformantes estables se
seleccionaron creciéndolos durante dos semanas en placas de medio HS con 1.5% agar
en presencia de luz. A partir de las colonias seleccionadas, se establecieron cultivos
líquidos en medio HS que se crecieron hasta alcanzar una biomasa aproximada de entre
0.5 a 1 g de peso fresco por 50 ml de medio. Los cultivos líquidos se agitaron
continuamente mediante burbujeo de aire enriquecido con un 2% de CO2.
3.21. Extracción y fraccionamiento de RNA viroidal de C. reinhardtii
Las células contenidas en 50 ml de cultivo se centrifugaron a 500 x g durante 10
min, y el sedimento se resuspendió en 4 ml de tampón de extracción (0.1 M Tris-HCl
pH 9, 0.1 M NaCl, 10 mM EDTA y 0.1 M 2-mercaptoetanol). El resuspendido se
añadió a un tubo con un volumen de fenol:cloroformo (1:1) (apartado 3.3), y 0.2
volúmenes de 10% SDS, previamente mezclados y atemperados a 65ºC. La mezcla, tras
homogeneización por inversión, se incubó a 65ºC durante 15 min y se centrifugó a 7800
x g 5 min. La fase acuosa se transfirió a un nuevo tubo con un volumen de
fenol:cloroformo (1:1) (apartado 3.3), se agitó y se centrifugó a 7800 x g 5 min. Para
fraccionar los RNAs se empleó celulosa no iónica (CF11, Whatman), que en presencia
de una solución 35% etanol retiene específicamente los RNAs con un alto contenido en
estructura secundaria como los RNAs viroidales. El volumen de la fase acuosa
recuperada del paso anterior se ajustó a 5 ml con agua y se añadió etanol 96% (hasta
alcanzar una concentración final del 35%) y 0.2 g de celulosa, manteniéndose con
agitación continua durante 60 min a temperatura ambiente. La celulosa se recuperó
centrifugando a 1950 x g 5 min y se lavó tres veces con 3 ml de etanol al 35% en
tampón de extracción. Los RNAs se eluyeron de la celulosa lavándola tres veces con 1
ml de tampón de extracción sin etanol, se recuperaron por precipitación con 2.5
volumen de etanol y 0.1 volúmenes de 3 M acetato sódico pH 5.5, y se resuspendieron
en 200 µl de agua. Para detectar los RNAs viroidales mediante análisis northern se
emplearon 20 µl de cada muestra.
72

Material y métodos
3.22. Construcción de un árbol filogenético de las tRNA ligasas de
plantas
El modelo de sustitución al que mejor se acomodan estas secuencias se infirió
aplicando el programa ProtTest (http://darwin.uvigo.es/software/prottest_server.html).
A partir de estos datos, el árbol filogenético se construyó por el método del vecino más
próximo (neighbor-joining) utilizando el programa MEGA (Kumar et al., 2004), y su
fiabilidad se evaluó por el método de Bootstrap realizando diez mil pseudoréplicas.
73


44.. RREESSUULLTTAADDOOSS


Resultados
4.1. Actividad RNA ligasa en cloroplastos aislados de espinaca
Diversos experimentos con extractos de germen de trigo y Chlamydomonas
reinhardtii sugieren que la ligación de los RNAs monoméricos lineales del viroide del
tubérculo fusiforme de la patata (Potato spindle tuber viroid, PSTVd) puede estar
catalizada por una tRNA ligasa (Branch et al., 1982; Kikuchi et al., 1982). De forma
paralela a como una RNA ligasa nuclear podría catalizar la ligación in vivo de los
miembros de la familia Pospiviroidae, podría existir una enzima cloroplástica de esta
clase que mediara la circularización de los RNAs monoméricos lineales resultantes del
autocorte de los miembros de la familia Avsunviroidae. Para detectar esta actividad se
partió inicialmente de cloroplastos íntegros de aguacate, el hospedador natural del
viroide del manchado solar del aguacate (Avocado sunblotch viroid, ASBVd), la especie
tipo de esta familia. Sin embargo, el rendimiento fue muy bajo por lo que se decidió
emplear espinaca, una planta muy utilizada en estudios que conllevan la purificación y
caracterización bioquímica de enzimas cloroplásticas.
4.1.1. Obtención de una fracción proteíca de cloroplastos de espinaca con actividad
RNA ligasa
Partiendo de hojas de plantas de espinaca de aproximadamente 45 días de edad
cultivadas a 20ºC, se llevó a cabo una purificación de cloroplastos con una etapa final
de centrifugación en un gradiente discontinuo de Percoll. Los cloroplastos intactos se
recuperaron, se lisaron y las proteínas se fraccionaron en una columna de heparina con
un gradiente de 0.1 a 1 M de KCl.
La actividad RNA ligasa de las 20 fracciones recogidas se ensayó empleando
como sustrato RNA monomérico lineal de polaridad positiva (+) del ASBVd
proveniente del autocorte ribozimático in vitro de un transcrito dimérico. Los productos
de la reacción se separaron por simple PAGE desnaturalizante y se detectaron mediante
hibridación northern con una ribosonda específica, observándose una actividad de
ligación en las fracciones eluidas con una concentración salina entre 500 y 640 mM de
KCl, con un máximo en la fracción correspondiente a 550-600 mM (Fig. 4.1, carriles 8
a 10). Como control de ligación en éste y otros experimentos se empleó una preparación
de germen de trigo que contenía la tRNA ligasa nuclear obtenida por fraccionamiento
con sulfato de amonio y cromatografía en heparina (ver apartado 3.2 de Materiales y
Métodos) (Fig. 4.1, carril 18).
77

Resultados
Fig. 4.1. Fraccionamiento y detección de una actividad RNA ligasa en cloroplastos de espinaca. Análisis por simple PAGE desnaturalizante e hibridación northern de los productos generados por las fracciones de un extracto cloroplástico en columna de heparina con concentraciones crecientes de KCl (indicadas con un triángulo en la parte superior). Como sustrato se empleó el RNA monomérico lineal (+) del ASBVd proveniente de autocorte. (1) Control sin extracto; (2) sobrenadante del extracto cloroplástico centrifugado a 100000 x g; (3-17) fracciones de la cromatografía; (18) control con una preparación parcialmente purificada de germen de trigo con actividad tRNA ligasa. Las posiciones del RNA monomérico circular (c) y lineal (l) del ASBVd se indican a la derecha.
4.1.2. Efecto de la fuerza iónica en la actividad RNA ligasa cloroplástica
Como la distinta concentración de KCl en las fracciones eluidas podría afectar a
la actividad de ligación, una alícuota de la fracción de la columna de heparina con el
máximo de actividad RNA ligasa (Fig. 4.1, carril 9) se dializó frente a tampón de
extracción con 0.1% Nonidet P40. La actividad de la fracción dializada se ensayó con
concentraciones crecientes de KCl, detectándose dicha actividad en las reacciones que
contenían entre 30 y 250 mM de KCl (Fig. 4.2, carriles 2 a 7), aunque el óptimo se
observó entre 100 y 150 mM (Fig. 4.2, carriles 4 y 5). En el experimento del apartado
anterior (Fig. 4.1), la concentración de sal inicial de la fracción con mayor actividad
(aproximadamente 600 mM) se diluyó cinco veces en el volumen final de la reacción de
ligación, por lo que la concentración final fue de 120 mM. Este valor se encuentra
dentro del intervalo de concentración de KCl para el que la actividad RNA ligasa
cloroplástica es máxima (Fig. 4.2), por lo que las condiciones en las que se detectó
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
[KCl] = 235 - 910 mM
- c
- l
78

Resultados
dicha actividad en las fracciones de la cromatografía en heparina son óptimas en cuanto
a la concentración de esta sal.
[KCl] = 30 – 500
1 2 3 4 5 6 7 8 9 10 11 12 13
- c
- l
Fig. 4.2. Efecto de la concentración de KCl en la actividad RNA ligasa cloroplástica. Análisis por simple PAGE desnaturalizante e hibridación northern de los productos resultantes de la ligación catalizada por la fracción cloroplástica más activa de la columna de heparina. (1) Control sin proteína; (2-12) reacciones con concentraciones crecientes de KCl (30 a 500 mM, con intervalos de 50 mM excepto las dos primeras reacciones, con un intervalo de 20 mM entre ellas); (13) control con una preparación parcialmente purificada de germen de trigo con actividad tRNA ligasa. Las posiciones del RNA monomérico circular (c) y lineal (l) de ASBVd se indican a la derecha. Otros detalles como en la Fig. 4.1.
4.1.3. Efecto de los ribonucleótidos trifosfato en la actividad RNA ligasa
cloroplástica
Generalmente, las RNA ligasas eucarióticas conocidas utilizan dos moléculas de
ATP como cofactores en dos de los pasos de la reacción de ligación: la fosforilación del
extremo 5’-hidroxilo del RNA y la adenilación del nuevo extremo 5’-fosfomonoéster
previa a la formación del enlace. Aunque la mayoría de las RNA ligasas emplean ATP,
algunas como la de la levadura Saccharomyces cerevisiae pueden emplear GTP para la
fosforilación.
Para estudiar la dependencia de la actividad RNA ligasa cloroplástica de algún
NTP específico, se diseñó un experimento de ligación con ATP, CTP, GTP o UTP, a la
vez que se incluyó un control sin ningún NTP. De esta forma se observó que la ligación
del sustrato ocurría en ausencia de NTPs con la misma eficiencia que en presencia de
79
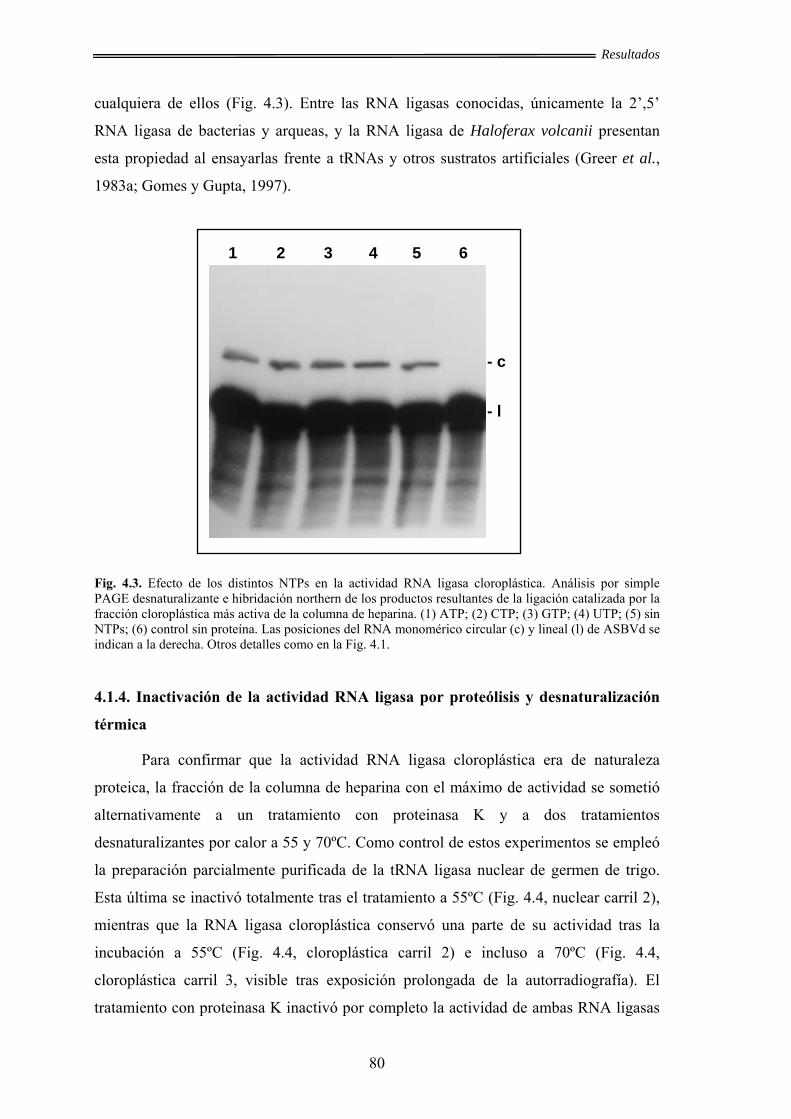
Resultados
cualquiera de ellos (Fig. 4.3). Entre las RNA ligasas conocidas, únicamente la 2’,5’
RNA ligasa de bacterias y arqueas, y la RNA ligasa de Haloferax volcanii presentan
esta propiedad al ensayarlas frente a tRNAs y otros sustratos artificiales (Greer et al.,
1983a; Gomes y Gupta, 1997).
1 2 3 4 5 6
- c
- l
Fig. 4.3. Efecto de los distintos NTPs en la actividad RNA ligasa cloroplástica. Análisis por simple PAGE desnaturalizante e hibridación northern de los productos resultantes de la ligación catalizada por la fracción cloroplástica más activa de la columna de heparina. (1) ATP; (2) CTP; (3) GTP; (4) UTP; (5) sin NTPs; (6) control sin proteína. Las posiciones del RNA monomérico circular (c) y lineal (l) de ASBVd se indican a la derecha. Otros detalles como en la Fig. 4.1.
4.1.4. Inactivación de la actividad RNA ligasa por proteólisis y desnaturalización
térmica
Para confirmar que la actividad RNA ligasa cloroplástica era de naturaleza
proteica, la fracción de la columna de heparina con el máximo de actividad se sometió
alternativamente a un tratamiento con proteinasa K y a dos tratamientos
desnaturalizantes por calor a 55 y 70ºC. Como control de estos experimentos se empleó
la preparación parcialmente purificada de la tRNA ligasa nuclear de germen de trigo.
Esta última se inactivó totalmente tras el tratamiento a 55ºC (Fig. 4.4, nuclear carril 2),
mientras que la RNA ligasa cloroplástica conservó una parte de su actividad tras la
incubación a 55ºC (Fig. 4.4, cloroplástica carril 2) e incluso a 70ºC (Fig. 4.4,
cloroplástica carril 3, visible tras exposición prolongada de la autorradiografía). El
tratamiento con proteinasa K inactivó por completo la actividad de ambas RNA ligasas
80

Resultados
(Fig. 4.4, carriles 4), mostrando que en los dos casos la reacción está mediada por una
proteína, aunque con una termoestabilidad significativamente mayor en la enzima
cloroplástica.
Cloroplástica Nuclear
C 1 2 3 4 1 2 3 4
- c
- l
Fig. 4.4. Inactivación de la actividad RNA ligasa por calor y proteólisis. Análisis por simple PAGE desnaturalizante e hibridación northern de los productos resultantes de la ligación catalizada por las RNA ligasas cloroplástica y nuclear contenidas en las fracciones más activas de la columna de heparina incubadas 30 min a dos temperaturas o 15 min en presencia de proteinasa K. (C) Control sin proteína; (1) sin pretratamiento; (2 a 4) pretratamiento a 55ºC, 70ºC y con proteinasa K, respectivamente. Las posiciones del RNA monomérico circular (c) y lineal (l) del ASBVd se indican a la derecha. Otros detalles como en la Fig. 4.1.
4.1.5. Cinética de las reacciones catalizadas por la RNA ligasa de espinaca y
germen de trigo
Para comparar las propiedades bioquímicas de la RNA ligasa cloroplástica de
espinaca frente a la previamente caracterizada tRNA ligasa nuclear de germen de trigo
se llevó a cabo un análisis cinético. Con ese fin se tomaron alícuotas de la reacción a
tiempos de entre 15 min y 4 h en presencia y ausencia de ATP. Mientras que la
actividad cloroplástica alcanzó el máximo de producto ligado a tiempos muy cortos, no
progresando la reacción transcurridos 30 min (Fig. 4.5A), con la preparación de germen
de trigo se observó aumento de la formación de producto hasta al menos 240 min (Fig
4.5B). Además, la cinética de la RNA ligasa cloroplástica, al contrario que la nuclear,
81

Resultados
fue idéntica en presencia y ausencia de ATP (Fig. 4.5A y B, comparar carriles 2 a 6 con
8 a 12), confirmando el resultado obtenido anteriormente (Fig. 4.3)
A + ATP Ø ATP
1 2 3 4 5 6 7 8 9 10 11 12
- c
- l
B + ATP Ø ATP 1 2 3 5 4 6 7 8 9 10 11 12
- c
- l
Fig. 4.5. Cinética de las RNA ligasas cloroplástica y nuclear. Análisis por simple PAGE desnaturalizante e hibridación northern de los productos resultantes de la ligación catalizada por las RNA ligasas cloroplástica (A) y nuclear (B) presentes en las fracciones más activas de la columna de heparina. La actividad enzimática se determinó a diferentes tiempos en presencia (1 a 6) y ausencia (7 a 12) de ATP. (1 y 7) Control sin enzima; (2 a 6 y 8 a 12) ligación a 15, 30, 60, 120 y 240 min respectivamente. Las posiciones del RNA monomérico circular (c) y lineal (l) del ASBVd se indican a la derecha de cada panel. Otros detalles como en la Fig. 4.1.
Las señales obtenidas en el análisis northern se cuantificaron mediante un
analizador de imagen, y los valores correspondientes al producto formado en presencia
de ATP se representaron gráficamente. De esta forma se observó que el producto en la
reacción catalizada por la enzima nuclear a lo largo del tiempo ensayado se acumuló
linealmente, mientras que en la reacción catalizada por la enzima cloroplástica se
alcanzó el máximo muy rápidamente (cuando sólo se había ligado un 2 % del sustrato)
(Fig. 4.6).
82

Resultados
0,16 0,14
RN
Ac/
RN
Ao
0,12 0,1
Cloroplástica0,08 Nuclear
0,06 0,04 0,02
0 0 20 40 60 80 100 120
Tiempo (min)
Fig. 4.6. Cinética de la actividad RNA ligasa catalizada por las RNA ligasas cloroplástica y nuclear. En el eje de abscisas se representa el tiempo de reacción, y en el de ordenadas la fracción del sustrato inicial (RNA0) convertida en producto (RNAC).
4.1.6. Estudio del factor limitante en la reacción de ligación
La falta de progreso de la reacción catalizada por la preparación cloroplástica de
RNA ligasa a tiempos superiores a 30 min podría deberse, entre otras causas, a la
inactivación de la enzima pocos minutos después del inicio de la reacción, o a que sólo
una pequeña parte del sustrato es ligable. Para discriminar entre estas alternativas se
llevó a cabo un experimento de ligación donde se duplicó la concentración del sustrato,
de la enzima o de ambos, y las señales obtenidas en el análisis northern se cuantificaron
mediante un analizador de imagen. En la reacción control, incubada durante 60 min, se
detectó una cantidad de producto acorde a la observada en experimentos previos (Fig.
4.7, carril 2). En una de las mezclas de reacción la concentración de sustrato se duplicó
transcurridos 30 min del inicio de la reacción, y se incubó otros 30 min, obteniéndose
aproximadamente el doble de producto que en la reacción control (Fig. 4.7, comparar
carriles 2 y 3). Por otra parte se duplicó la concentración de la enzima en dos mezclas
de reacción: en una de ellas transcurridos 30 min (Fig. 4.7, carril 4) y en la otra desde el
mismo inicio de la reacción (Fig. 4.7, carril 6), no detectándose en ninguno de los dos
casos diferencias en la cantidad de producto con respecto a la reacción control (Fig. 4.7,
comparar carriles 2, 4 y 6). Finalmente, al duplicar conjuntamente la concentración de
enzima y sustrato una vez transcurridos 30 min del inicio de la reacción e incubar otros
83
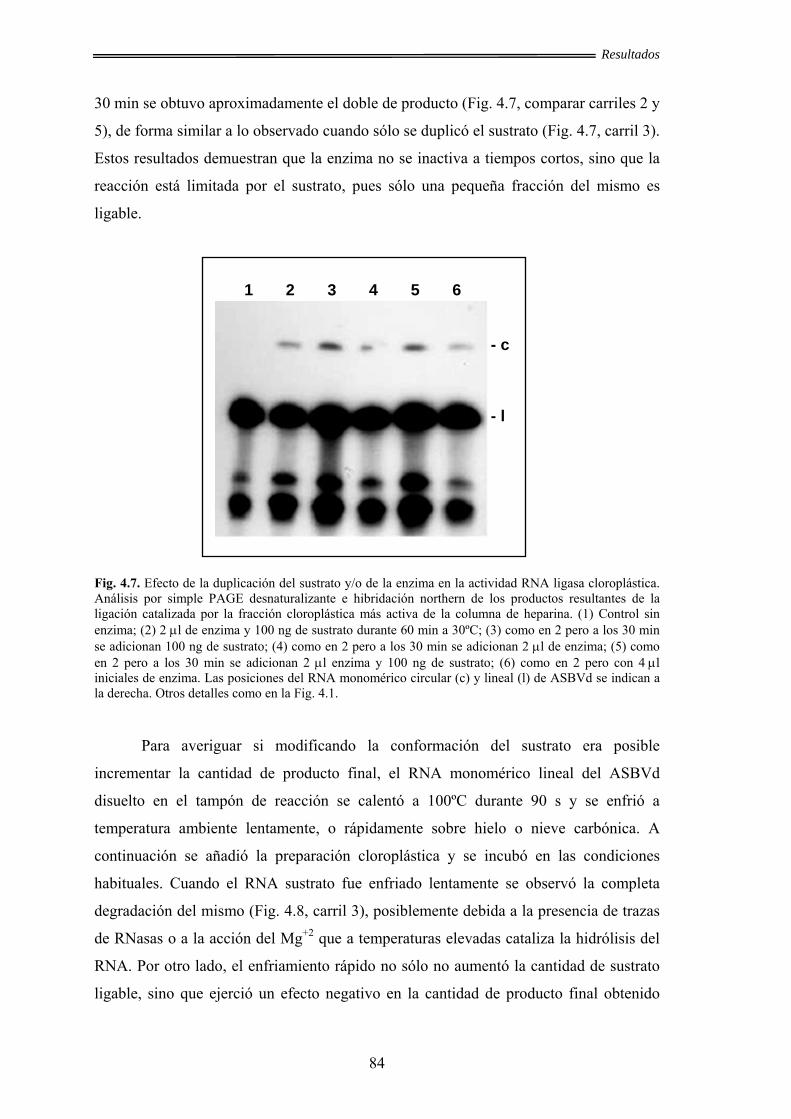
Resultados
30 min se obtuvo aproximadamente el doble de producto (Fig. 4.7, comparar carriles 2 y
5), de forma similar a lo observado cuando sólo se duplicó el sustrato (Fig. 4.7, carril 3).
Estos resultados demuestran que la enzima no se inactiva a tiempos cortos, sino que la
reacción está limitada por el sustrato, pues sólo una pequeña fracción del mismo es
ligable.
1 2 3 4 5 6
- c
- l
Fig. 4.7. Efecto de la duplicación del sustrato y/o de la enzima en la actividad RNA ligasa cloroplástica. Análisis por simple PAGE desnaturalizante e hibridación northern de los productos resultantes de la ligación catalizada por la fracción cloroplástica más activa de la columna de heparina. (1) Control sin enzima; (2) 2 µl de enzima y 100 ng de sustrato durante 60 min a 30ºC; (3) como en 2 pero a los 30 min se adicionan 100 ng de sustrato; (4) como en 2 pero a los 30 min se adicionan 2 µl de enzima; (5) como en 2 pero a los 30 min se adicionan 2 µl enzima y 100 ng de sustrato; (6) como en 2 pero con 4 µl iniciales de enzima. Las posiciones del RNA monomérico circular (c) y lineal (l) de ASBVd se indican a la derecha. Otros detalles como en la Fig. 4.1.
Para averiguar si modificando la conformación del sustrato era posible
incrementar la cantidad de producto final, el RNA monomérico lineal del ASBVd
disuelto en el tampón de reacción se calentó a 100ºC durante 90 s y se enfrió a
temperatura ambiente lentamente, o rápidamente sobre hielo o nieve carbónica. A
continuación se añadió la preparación cloroplástica y se incubó en las condiciones
habituales. Cuando el RNA sustrato fue enfriado lentamente se observó la completa
degradación del mismo (Fig. 4.8, carril 3), posiblemente debida a la presencia de trazas
de RNasas o a la acción del Mg+2 que a temperaturas elevadas cataliza la hidrólisis del
RNA. Por otro lado, el enfriamiento rápido no sólo no aumentó la cantidad de sustrato
ligable, sino que ejerció un efecto negativo en la cantidad de producto final obtenido
84

Resultados
(Fig. 4.8, comparar carriles 2, 4 y 5). De este experimento se infiere que las mejores
condiciones para obtener la conformación adecuada de ligación son aquellas en las que
se realiza la purificación del RNA, y que la desnaturalización y renaturalización del
sustrato no aumenta la fracción de moléculas ligables.
1 2 3 4 5
- c
- l
Fig. 4.8. Efecto de la conformación del sustrato en la actividad RNA ligasa cloroplástica. Análisis por simple PAGE desnaturalizante e hibridación northern de los productos resultantes de la ligación catalizada por la fracción cloroplástica más activa de la columna de heparina, empleando como sustrato el RNA del ASBVd incubado a 100ºC durante 90 s y sometido a diferentes tratamientos de enfriamiento para alterar su conformación. (1) Control sin enzima; (2) control sin tratamiento; (3) enfriamiento lento; (4) enfriamiento rápido en hielo; (5) enfriamiento rápido en nieve carbónica. Las posiciones del RNA monomérico circular (c) y lineal (l) del ASBVd se indican a la derecha. Otros detalles como en la Fig. 4.1.
4.1.7. Efecto del pH y del Mg2+ en la actividad RNA ligasa cloroplástica
Con el fin de conocer el pH óptimo de la RNA ligasa cloroplástica, se ensayó la
actividad catalítica de la fracción más activa de la columna de heparina en un intervalo
de pH entre 6 y 9. El óptimo se observó a pH 8 (valor similar al de las tRNA ligasas; ver
más adelante, apartado 4.2.4.1), si bien la actividad fue detectable en todo el intervalo
ensayado (Fig. 4.9, carriles 3 a 6).
Algunas RNA ligasas de bacterias y arqueas catalizan la reacción en ausencia de
Mg2+ (Greer et al., 1983a; Kjems y Garrett, 1988; Gomes y Gupta, 1997), en contraste
con lo que sucede con la mayoría de las enzimas de esta clase. Para ensayar la actividad
de la RNA ligasa cloroplástica en estas condiciones, el Mg2+ presente en la preparación
85

Resultados
cloroplástica se acomplejó con EDTA, y la reacción se llevó a cabo con tampón de
ligación sin este catión. La actividad detectada en dichas condiciones fue similar a la de
la reacción control (Fig. 4.9, comparar carriles 2 y 5), si bien se apreció una ligera
disminución en la cantidad de producto obtenido similar a la observada cuando la
ligación se ensayó a pHs distintos del óptimo.
1 2 3 4 5 6
- c
- l
Fig. 4.9. Efecto del pH y del Mg+2 en la actividad de la RNA ligasa cloroplástica. Análisis por simple PAGE desnaturalizante e hibridación northern de los productos resultantes de la ligación catalizada por la fracción cloroplástica más activa de la columna de heparina. (1) Control sin enzima; (2) ligación sin Mg2+ a pH 8; (3 a 6) ligación en presencia de Mg2+ a pH 6, 7, 8 y 9, respectivamente. Las posiciones del RNA monomérico circular (c) y lineal (l) del ASBVd se indican a la derecha. Otros detalles como en la Fig. 4.1.
4.1.8. Extremos del RNA requeridos por la RNA ligasa cloroplástica
El autocorte mediado por las ribozimas de cabeza de martillo genera RNAs con
extremos 5’-hidroxilo y 2’,3’-fosfodiéster cíclico, que son los reconocidos por las tRNA
ligasas de levadura y plantas de forma más eficiente. Para ensayar los requerimientos a
este respecto de la RNA ligasa cloroplástica de espinaca se empleó como sustrato RNA
monomérico lineal del ASBVd resultante del autocorte de un transcrito dimérico.
Previamente a su ensayo, este sustrato se sometió a: i) rotura química del grupo 2’,3’-
fosfodiéster cíclico por tratamiento ácido (que deja el grupo fosforilo equimolarmente
en 2’ y 3’), ii) desfosforilación del producto de la reacción anterior, tanto en 2’ como en
86

Resultados
3’, con fosfatasa alcalina de gamba, iii) fosforilación en 5’ con T4 polinucleótido
quinasa (PNK), y iv) otras combinaciones. Como control de las reacciones de
modificación del RNA se ensayó su ligación con T4.2 RNA ligasa, que sólo cataliza la
unión de extremos 5’-fosfomonoéster y 3’ y 2’-hidroxilo.
De este experimento sólo pudo concluirse que la RNA ligasa cloroplástica no es
activa frente a extremos 5’-fosfomonoéster, pues al analizar los controles con T4.2
RNA ligasa se observó que los tratamientos de modificación no generaron los extremos
pretendidos. Una posible interpretación es la ineficiencia de la reacción de rotura
química del fosfodiéster cíclico, y otra, probablemente más importante, la imposibilidad
de controlar durante las reacción de fosforilación con PNK su actividad
ciclofosfodiesterasa-fosfatasa. Si bien en ausencia de ATP esta enzima cataliza la
hidrólisis del extremo 2’,3’-fosfodiéster cíclico y elimina posteriormente el grupo
fosforilo, no puede ensayarse su actividad quinasa sin que opere simultáneamente la
actividad ciclofosfodiesterasa-fosfatasa, a pesar de que ambas actividades tienen un pH
óptimo distinto. Como resultado, se genera una mezcla de RNAs con diferentes
extremos que no pueden aislarse para ensayarlos por separado.
Con el fin de averiguar si la RNA ligasa cloroplástica catalizaba la unión de
extremos que no fuesen 2’,3’-fosfodiéster cíclico, el RNA monomérico del ASBVd
procedente de autocorte se trató previamente con tRNA ligasa de Arabidopsis thaliana
en ausencia de NTPs. En estas condiciones únicamente es activo el dominio
ciclofosfodiesterasa de la enzima, que a diferencia de la PNK no tiene actividad
fosfatasa, produciendo extremos 5’ y 3’-hidroxilo y 2’-fosfomonoéster. Al incubar este
sustrato con la preparación cloroplástica no se observó ligación ni en ausencia ni en
presencia de NTPs (Fig. 4.10, carriles 4 y 5), lo que sugiere que la RNA ligasa
cloroplástica es incapaz de ligar estos extremos. Aunque no hemos podido ensayar un
sustrato con extremos 3’-fosfomonoéster y 5’-hidroxilo, todo apunta a que el
mecanismo de la RNA ligasa cloroplástica procede a través de una transesterificación,
donde la rotura del grupo 2’,3’-fosfodiéster cíclico aporta la energía necesaria para la
formación del nuevo enlace, 3’-5’ o 2’-5’.
87

Resultados
1 2 3 4 5
- c
- l
Fig. 4.10. Ligación por la RNA ligasa cloroplástica de un sustrato con diferentes extremos 3’. Análisis por simple PAGE desnaturalizante e hibridación northern de los productos resultantes de la ligación catalizada por la fracción cloroplástica más activa de la columna de heparina en ausencia de NTPs (excepto cuando se indica lo contrario). Como sustrato se empleó RNA monomérico del ASBVd proveniente de autocorte o este mismo tratado con tRNA ligasa de A. thaliana para hidrolizar su extremo 2’,3’-fosfodiéster cíclico. (1) Control sin enzima con un sustrato 5’-OH; 2’,3’>P; (2) control sin enzima con un sustrato 5’-OH; 3’-OH, 2’-P; (3) ligación con un sustrato con extremos 5’-OH; 2’,3’>P; (4) ligación con sustrato con extremos 5’-OH; 3’-OH, 2’-P; (5) ligación de sustrato con extremos 5’-OH; 3’-OH, 2’-P en presencia de NTPs. Las posiciones del RNA monomérico circular (c) y lineal (l) del ASBVd se indican a la derecha. Otros detalles como en la Fig. 4.1.
4.1.9. Actividad de la RNA ligasa cloroplástica sobre los RNAs monoméricos de la
familia Avsunviroidae y autoligación de los mismos
Para conocer si la RNA ligasa cloroplástica era capaz de ligar otros RNAs
viroidales se obtuvieron RNAs monoméricos lineales procedentes de autocorte del
ASBVd, del viroide del mosaico latente del melocotonero (Peach latent mosaic viroid,
PLMVd) del viroide del moteado clorótico del crisantemo (Chrysanthemum chlorotic
mottle, CChMVd) y del viroide latente de la berenjena (Eggplant latent viroid, ELVd),
y se ensayó la ligación de los mismos en las condiciones habituales. Alternativamente,
se ha descrito en la bibliografía que el RNA monomérico PLMVd procedente de
autocorte ribozimático es capaz de ligarse in vitro en ausencia de proteínas, si bien esta
reacción, denominada autoligación, requiere alta concentración de Mg2+, y genera un
enlace 2’,5’-fosfodiéster (Côté y Perreault 1997). Con el fin de examinar esta actividad
paralelamente a la ligación catalizada por la RNA ligasa cloroplástica, los RNAs de
cuatro miembros de la familia Avsunviroidae se incubaron también en tampón de
autoligación (4 mM Tris-HCl pH 9, 100 mM MgCl2) a 16ºC durante 12 h.
88

Resultados
A ASBVd B C DPLMVd CChMVd ELVd
Fig. 4.11. Ligación y autoligación de los RNAs viroidales cloroplásticos. Análisis por simple PAGE desnaturalizante e hibridación northern de los productos de la ligación catalizada por la fracción cloroplástica más activa de la columna de heparina y de la autoligación a 100 mM Mg2+. Como sustrato se emplearon los RNAs monoméricos procedentes del autocorte ribozimático de los cuatro miembros de la familia Avsunviroidae. (1) Controles sin enzima (en tampón con 4 mM Mg2+); (2) controles sin enzima (en tampón 100 mM MgCl2); (3) reacciones con la RNA ligasa cloroplástica. Las posiciones de los RNA monoméricos circulares (c) y lineales (l) se indican a la derecha de cada panel. Otros detalles como en la Fig. 4.1.
A 100 mM de MgCl2 se observó autoligación de los RNAs del PLMVd y del
ELVd (Fig. 4.11B y 4.11D, carriles 2). Además este último también se autoligó, aunque
en menor medida, en el tampón que se empleó como control con una concentración de
Mg+2 de tan sólo 4 mM (Fig. 4.11D, carril 1). Sin embargo, los RNAs del ASBVd y
CChMVd no mostraron autoligación apreciable en alta concentración de Mg2+ (Fig.
4.11A y 4.11C, carriles 2). El análisis de la estructura primaria de los dos RNAs
viroidales que se autoligan a 100 mM Mg+2 muestra que el enlace se genera entre una C
y una U en el PLMVd (Fig. 4.12B), y entre una A y una G en el ELVd (Fig. 4.12D),
mientras que en los RNAs no autoligables en las condiciones ensayadas el enlace es
entre una C y una U en el ASBVd (Fig. 4.12A), y entre una C y una G en el CChMVd
(Fig. 4.12C). En cuanto a la estructura secundaria, la región donde se localiza el sitio de
unión en el ASBVd, PLMVd y CChMVd forma una doble hebra (Fig. 4.12A, 4.12B y
4.12C), si bien en el primero la G situada a continuación de la U que forma el enlace
está desapareada. En el ELVd la G del enlace también está formando parte de una
región con estructura secundaria, pero la A y los cuatro nucleótidos anteriores están
1 2 31 2 3
- c
- l
2 311 2 3
- l
- c
- l
- c
- l
89

Resultados
desapareados (Fig. 4.12D). Por tanto, no parece haber ninguna relación entre la
estructura primaria o secundaria de los RNAs viroidales y su capacidad de autoligación.
Con respecto a la actividad RNA ligasa cloroplástica, además del ASBVd,
también se mostró capaz de circularizar el ELVd incluso con mayor eficiencia (Fig.
4.11A y 4.11D, comparar carriles 3), pero no se detectó ligación del PLMVd o del
CChMVd (Fig. 4.11B y 4.11C, carriles 3). La ligación catalizada por la RNA ligasa
cloroplástica ocurre en los RNAs viroidales que tienen alguna base desapareada en el
sitio de unión (ELVd) o adyacente a el (ASBVd) (Fig. 4.12D y 4.12A,
respectivamente), mientras que la enzima no cataliza la ligación de aquellos RNAs en
los que el sitio de unión se encuentra en una zona de doble hélice (PLMVd y CChMVd)
(Fig. 4.12B y 4.12C, respectivamente). Además, los RNAs viroidales circularizables por
esta enzima tienen una estructura de cuasi-varilla (ASBVd y ELVd) mientras que los no
circularizables son muy ramificados, sugiriendo que la RNA ligasa cloroplástica
requiere determinadas conformaciones del RNA para su función, como también parece
inferirse del experimento previo de calentamiento y enfriamiento del ASBVd (Fig. 4.8).
A CB D
AG
UGU. .
CChMVd
GCU
CUCG A U
GGC
A GUC
14 396
30123
. . .....
PLMVd
UG C AGUCA.G U.U
G...C
U
CCG.
45
293
53
285
. .
ASBVd
C U.AC
.A A AG
U.G
U
A
GC.
184
58
Fig. 4.12. Esquema del sitio de procesamiento de los RNAs viroidales cloroplásticos. Las flechas indican el sitio donde ocurre el autocorte catalizado por las ribozimas de cabeza de martillo, y los números la posición que ocupan en el RNA viroidal el primer y último nucleótido de los fragmentos representados.
4.1.10. Determinación del enlace generado por la RNA ligasa cloroplástica
Con el fin de examinar si la reacción catalizada por la RNA ligasa cloroplástica
genera un enlace 3’,5’-fosfodiéster como el descrito para la mayoría de las RNA ligasas
conocidas (revisado en Arn y Abelson, 1998), o 2’,5’-fosfodiéster como el generado en
la autoligación del RNA del PLMVd (Côté y Perreault, 1997) y en la reacción
catalizada por las 2’,5’-RNA ligasas de bacterias y arqueas (Greer et al., 1983a), se
aplicaron dos metodologías para detectar enlaces anómalos. En una de ellas se trató el
191
52
. .
ELVd
GCC.
UG
AG
G.
AUU
193
5332
199
1AG
UGU. .
CChMVd
GCU
CUCG A U
GGC
A GUC
14 396
30123
. . .....
PLMVd
UG C AGUCA.G U.U
G...C
U
CCG.
45
293
53
285
. .
ASBVd
C U.AC
.A A AG
U.G
U
A
GC.
184
58
1
52
. .
ELVd
GCC.
UG
AG
G.
AUU
193
5332
199
1
19
90

Resultados
RNA ligado con RNasas y los productos resultantes se analizaron mediante
cromatografía en capa fina (thin layer cromatography, TLC) junto con patrones para
determinar la presencia de dinucleótidos con enlaces no digeribles por las RNasas. En la
otra se examinó la existencia en el RNA ligado de enlaces diferentes del 3’,5’-
fosfodiéster mediante experimentos de extensión del cebador.
La primera de estas metodologías, como ya se ha comentado, consistió en digerir
el RNA con RNasas y separar los productos generados mediante TLC. El enlace a que
da lugar la ligación del RNA monomérico lineal del ASBVd ocurre entre una C y una U
y, si el enlace que se forma es 2’,5’-fosfodiéster, resulta resistente a las RNasas,
acumulándose por tanto un dinucleótido CpU. Este dinucleótido puede detectarse por su
movilidad cromatográfica, que es diferente a la de los cuatro nucleótidos 5’-
monofosforilados. En un primer experimento el RNA monomérico lineal del ASBVd
procedente de autocorte, marcado internamente empleando [α−32P]UTP durante la
transcripción, se dividió en tres alícuotas: la primera no se sometió a tratamiento alguno,
mientras que la segunda se circularizó con la 2’,5’-RNA ligasa de Escherichia coli y la
tercera con la RNA ligasa cloroplástica de espinaca. Los productos resultantes se
separaron mediante doble PAGE, se eluyeron y se digirieron totalmente con nucleasa
S1, que genera nucleótidos monofosforilados en 5’. Esta reacción también provoca la
liberación del grupo fosforilo radiactivo de una fracción de los nucleótidos,
transformándolos en nucleósidos indetectables por autorradiografía. Si el sustrato está
ligado con un enlace 2’,5’-fosfodiéster la digestión genera además de pU, un pCpU,
ambos marcados. Tras la separación de los productos de la digestión con un eluyente
polar, el revelado autorradiográfico de la placa de celulosa mostró una mancha en la
posición correspondiente al patrón de pU en todas las muestras (Fig. 4.13, comparar
carriles 3 a 5 con 1), pero en ninguna de ellas se detectó la mancha correspondiente al
patrón de CpU (Fig. 4.13, comparar carriles 3 a 5 con 2). El grupo fosfato radiactivo
que, por su polaridad migra al extremo superior de la placa, se detectó en las tres
muestras (Fig. 4.13, carriles 3 a 5).
En la muestra resultante de la digestión del sustrato lineal (Fig. 4.13, carril 3) no
se observó ninguna mancha extra, como cabía esperar para un RNA con todos sus
enlaces 3’,5’-fosfodiéster. Sin embargo, en la muestra producto de la digestión del RNA
circularizado con la 2’,5’-RNA ligasa de E. coli se observó una débil mancha (Fig. 4.13,
carril 5) con movilidad cromatográfica ligeramente menor que la del patrón CpU (Fig
4.13, carril 2), y que podría corresponder a pCpU. En la muestra resultante de la
91

Resultados
digestión del sustrato circularizado con la RNA ligasa cloroplástica se detectó una
mancha (Fig. 4.13, carril 4) que, al igual que en la muestra ligada con la 2’,5’-RNA
ligasa de E. coli, presentó una movilidad cromatográfica ligeramente menor que la del
patrón CpU (Fig. 4.13, comparar carriles 4 y 5). Debido a que la movilidad del
presumible pCpU es diferente de la del patrón comercial CpU, se intentó fosforilar este
último con PNK y [γ-32P]ATP (ya que la forma fosforilada no se pudo obtener
comercialmente), pero las reacciones necesarias para este proceso generan múltiples
moléculas con diferente movilidad cromatográfica que no se pudieron identificar,
probablemente por la presencia de productos de degradación derivados del dinucleótido
y de trazas de otras moléculas presentes en los reactivos empleados en la fosforilación.
En consecuencia resultó imposible concluir a partir de estos experimentos que la RNA
ligasa cloroplástica genera un enlace 2’,5’-fosfodiéster, si bien se detectó la aparición de
una nueva especie marcada.
1 2 3 4 5P -
pU -
CpU -
Fig. 4.13. Detección de enlaces anómalos en los RNAs circularizados por la RNA ligasa cloroplástica. Análisis mediante TLC con un eluyente polar de los productos resultantes de digerir con la nucleasa S1 los RNAs circulares y lineales del ASBVd. (1) Patrón de pU; (2) patrón de CpU; (3, 4 y 5) productos de la digestión del RNA lineal del ASBVd, circularizado con la RNA ligasa cloroplástica y con la 2’,5’-RNA ligasa de E. coli, respectivamente. Las muestras se aplicaron en la parte inferior de la placa. La posición de los patrones pU, CpU y del grupo fosfato liberado por la digestión (P) se indica a la izquierda. Los círculos punteados delimitan la posición de los patrones no radiactivos.
92

Resultados
Empleando la misma técnica (TLC) para separar los productos de la digestión
del RNA circularizado, se aplicó una estrategia alternativa para intentar mejorar los
resultados anteriores. En esta ocasión los RNAs obtenidos de idéntica forma que en el
experimento anterior, pero en ausencia de precursor radiactivo, se digirieron con RNasa
ONE (Promega). Dicha enzima, a diferencia de la nucleasa S1 empleada previamente,
libera nucleóticos con el grupo fosforilo en 3’ (Np). Estos nucleótidos, y en su caso el
CpUp, se fosforilaron en 5’ con PNK y [γ-32P]ATP, agotándose el exceso de este último
con hexoquinasa y glucosa (dando lugar a glucosa-6-P marcada). Finalmente se eliminó
el grupo fosforilo en 3’ con nucleasa P1, generando nucleótidos marcados
radiactivamente en 5’ y, en su caso pCpU también marcado en 5’. Los productos
resultantes se separaron mediante TLC bidimensional, con un primer eluyente apolar y
un segundo polar, y la placa se reveló autorradiográficamente. Sin embargo, al igual que
sucedió en el experimento anterior cuando se fosforiló el dinucleótido CpU, se
detectaron múltiples manchas que no pudieron asignarse a patrones conocidos.
La segunda metodología empleada para examinar la naturaleza del enlace
generado por la RNA ligasa cloroplástica consistió en experimentos de extensión del
cebador utilizando como molde RNA circularizado. Durante la síntesis de cDNA, si
todos los enlaces del molde son 3’,5’-fosfodiéster la transcripción inversa no sufre
pausas (o son limitadas), mientras que si el molde está cerrado por un enlace atípico la
enzima no progresa y da lugar a un transcrito de longitud menor a la esperada. Para este
experimento el RNA monomérico lineal del ASBVd procedente de autocorte in vitro se
trató tanto con la 2’,5’-RNA ligasa de E. coli como con la RNA ligasa cloroplástica, y el
RNA circular obtenido se purificó mediante doble PAGE. Como control se empleó
RNA monómérico circular del ASBVd extraído de plantas infectadas y purificado de la
misma forma. La reacción se llevó a cabo con el cebador C1 previamente fosforilado
con PNK y [γ-32P]ATP, que hibrida con el molde a 116 nucleótidos (nt) (contados desde
su extremo 5’) del sitio de ligación del RNA viroidal. Los productos de la extensión del
cebador, separados por simple PAGE desnaturalizante y revelados
autorradiográficamente, deberían mostrar la señal correspondiente a un cDNA de 116 nt
si el RNA molde está cerrado por un enlace atípico.
93

Resultados
Fig. 4.14. Ensayo de extensión del cebador sobre RNAs viroidales circulares para detectar la presencia de enlaces anómalos. Análisis por simple PAGE desnaturalizante de los productos obtenidos por transcripción inversa con un cebador marcado radiactivamente en 5’ y distintos RNAs circulares de ASBVd. (M) Marcador de tamaño molecular; (1) molde circularizado con la RNA ligasa cloroplástica; (2) molde circularizado con la 2’,5’-RNA ligasa de E. coli; (3) molde circular proveniente de planta infectada. La posición de los marcadores (con su tamaño en nt) se indica a la izquierda; las bandas de menor intensidad entre 250 y 500 nt corresponden a tamaños de 300, 350, 400 y 450 nt.
Cuando en la reacción se empleó un molde circularizado con la 2’,5’-RNA
ligasa de E. coli se detectó un transcrito mayoritario de 116 nt (Fig. 4.14, carril 2),
mientras que la reacción con un molde de ASBVd circular extraído de planta infectada
no se observaron señales intensas correspondientes a cDNAs truncados (Fig. 4.14, carril
3). Este resultado muestra que el enlace que cierra el RNA viroidal in vivo es
presumiblemente 3’,5’-fosfodiéster, y las señales de baja intensidad detectadas
corresponden a paradas prematuras de la transcriptasa inversa que resultan del alto
grado de autocomplementariedad del RNA viroidal. Finalmente, la reacción con un
molde de RNA circularizado con la RNA ligasa cloroplástica generó un cDNA
predominante de 116 nt similar al resultante de emplear como molde el RNA
circularizado por la enzima de E. coli (Fig. 4.14, comparar carriles 1 y 2). Considerando
que el RNA sustrato de la reacción mediada por la RNA ligasa cloroplástica posee
extremos 5’-hidroxilo y 2’,3’-fosfodiéster cíclico, y que no requiere aporte de grupos
fosforilo externos, la ligación presumiblemente ocurre por transesterificación generando
un enlace fosfodiéster 3’-5’ o 2’-5’. Como los resultados de extensión del cebador
M 1 2 3nt
500 -
200 - 250 -
150 -
100 -
50 -
94

Resultados
muestran que el enlace no es 3’-5’, cabe inferir que probablemente sea 2’,5’-
fosfodiéster. Ésta y otras características bioquímicas, como son la ligación de sólo una
pequeña fracción del sustrato y el no requerimiento de NTPs y Mg+2 (Greer et al., 1983;
Arn y Abelson, 1996), establecen una estrecha relación entre la RNA ligasa
cloroplástica y las 2’,5’-RNA ligasas de bacterias y arqueas.
4.1.11. Purificación de la RNA ligasa cloroplástica y posible relación con otras
proteínas
Para este cometido se empleó una precipitación fraccionada con sulfato amónico
y distintas técnicas cromatográficas, entre ellas de exclusión molecular con Superdex 75
10/300 GL, de afinidad con heparina y de intercambio iónico con Sephadex SP25. Con
este procedimiento se detectó la actividad de ligación a lo largo de la purificación. Sin
embargo, tras analizar la preparación final mediante SDS-PAGE y teñir el gel con plata,
dicha actividad no pudo asociarse a ninguna banda visible, por lo que la RNA ligasa
cloroplástica de espinaca debe acumularse in vivo a muy baja concentración.
Durante el transcurso de estos experimentos se encontró una secuencia de
leguminosa (Medicago truncatula Gaertn.), con entrada Q1SQ96 en la base de datos
Uniprot/TrEMBL, para la que los programas Pfam e InterPro predicen un dominio
2’,5’-RNA ligasa/ciclofosfodiesterasa. El gen correspondiente se clonó introduciendo
en 3’ una secuencia codificante de seis histidinas carboxilo terminales, y se expresó en
células de E. coli Rosetta 2. La proteína (con un tamaño predicho de 21 kDa) se purificó
con una columna de Ni-NTA-agarosa que retiene específicamente a las proteínas que
contienen una cola de seis histidinas.
Aunque se obtuvo una proteína de aproximadamente 21 kDa con un buen
rendimiento (Fig. 4.15, carril E), no se detectó actividad enzimática en las mismas
condiciones empleadas con la RNA ligasa cloroplástica de espinaca. Este dominio 2’,5’-
RNA ligasa/ciclofosfodiesterasa no ha sido predicho en las proteínas homólogas de A.
thaliana ni levadura, lo que unido a los resultados negativos de ligación, sugiere que la
predicción in silico es errónea. Por lo tanto, para la identificación de la RNA ligasa
cloroplástica será necesario realizar una búsqueda más precisa de dominios 2’,5’-RNA
ligasa/ciclofosfodiesterasa en las secuencias anotadas en las bases de datos, o intentar la
secuenciación directa de un fragmento de la proteína una vez se disponga de mayores
cantidades de la misma.
95
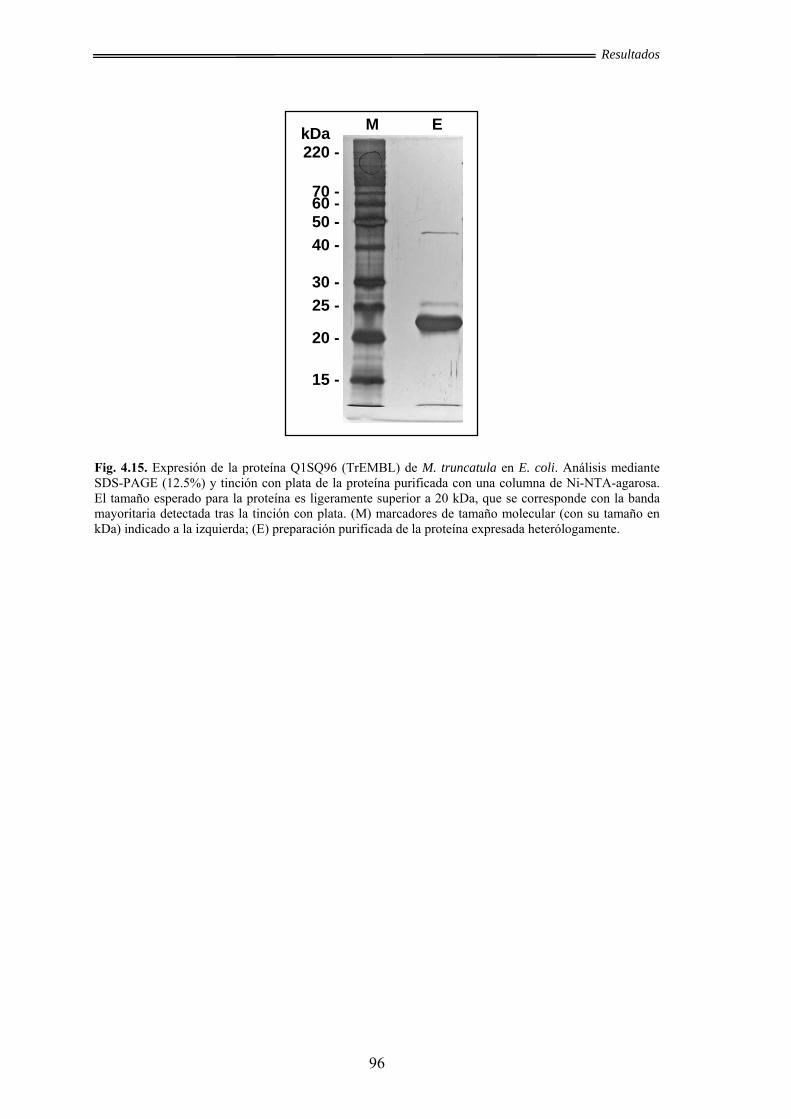
Resultados
Fig. 4.15. Expresión de la proteína Q1SQ96 (TrEMBL) de M. truncatula en E. coli. Análisis mediante SDS-PAGE (12.5%) y tinción con plata de la proteína purificada con una columna de Ni-NTA-agarosa. El tamaño esperado para la proteína es ligeramente superior a 20 kDa, que se corresponde con la banda mayoritaria detectada tras la tinción con plata. (M) marcadores de tamaño molecular (con su tamaño en kDa) indicado a la izquierda; (E) preparación purificada de la proteína expresada heterólogamente.
M E
220 -kDa
150 -160 -170 -
140 -
130 -125 -
120 -
115 -
96

Resultados
4.2. Clonación, secuenciación y expresión del gen de la tRNA ligasa de
tomate (Solanum lycopersicum)
Como ya se ha mencionado anteriormente, la ligación de los RNAs
monoméricos lineales del PSTVd con extractos de germen de trigo y C. reinhardtii
parece estar mediada por una tRNA ligasa (Branch et al., 1982: Kikuchi et al., 1982).
En plantas se han identificado y caracterizado los genes de las tRNA ligasas de arroz y
A. thaliana (Englert y Beier, 2005), así como de las tres isoformas de la tRNA ligasa
que catalizan la maduración de los tRNAs en trigo (Makino et al., 2006). Como ninguna
de estas especies es hospedador natural de viroides, en el presente trabajo se planteó el
objetivo de clonar el mRNA de la tRNA ligasa de una planta que sí lo fuera, y disponer
así de una herramienta que permitiera estudiar in vitro la ligación de los miembros de la
familia Pospiviroidae. Entre las posibles opciones se eligió el tomate porque es
infectado por el PSTVd, el miembro tipo de esta familia, y porque la secuenciación de
su genoma está bastante avanzada.
4.2.1. Búsqueda de etiquetas de secuencias expresadas (expressed sequence tags,
ESTs) de tomate con similitud a los genes de las tRNA ligasas de plantas
Partiendo de las secuencias de las tRNA ligasas ya caracterizadas de arroz, A.
thaliana y trigo, se buscaron las presumibles secuencias homólogas, tanto nucleotídicas
como aminoacídicas, en las bases de datos de solanáceas de Cornell University
(http://www.sgn.cornell.edu/solanaceae-project/index.pl) y del National Center for
Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.gov). Con este fin se
emplearon los algoritmos BLASTN (Altschul et al., 1990) y PSI-BLAST (Altschul et
al., 1997; Schaffer et al., 2001). El resultado más interesante de la búsqueda fue una
secuencia nucleotídica combinada de 679 pares de bases (pb), obtenida a partir de
cuatro ESTs solapantes de tomate, anotada como SGN-U223941. Esta secuencia mostró
una identidad del 70% o superior al compararla con la de los cDNAs de las tRNA
ligasas conocidas, por lo que se consideró una buena candidata a ser un fragmento del
gen homólogo de tomate.
97

Resultados
4.2.2. Clonación del cDNA candidato de la tRNA ligasa de tomate
4.2.2.1. Empleo de técnicas de amplificación rápida de los extremos de un cDNA
(Rapid amplification of cDNA ends, RACE) para determinar las regiones
terminales 5’ y 3’
La secuencia combinada de los cuatro ESTs depositada en la base de datos de
Cornell University (SGN-U223941) sirvió para diseñar cebadores que, mediante
técnicas de RACE, permitieran determinar las regiones terminales 5’ y 3’ del mRNA.
Para este cometido se utilizó RNA poliadenilado [poli(A)] purificado de hojas de
tomate. Los experimentos de 3’-RACE se llevaron a cabo empleando un cebador (C2),
con una cola poli(dT) en 3’ y una secuencia definida en 5’, con el que se transcribieron
inversamente los mRNAs (Sambrook y Rusell, 2001). Un fragmento comprendido entre
la secuencia conocida del cDNA de tomate y la introducida en 5’ por el cebador C2 se
amplificó por PCR con los cebadores C3 y C4, el primero derivado de SGN-U223941 y
el segundo complementario a C2. Para obtener un producto visible mediante
electroforesis en gel de agarosa y tinción con bromuro de etidio fue necesaria una
segunda amplificación con cebadores más internos (C5 y C6). Este producto se clonó en
el plásmido pTZ57R/T (abierto en el sitio EcoRV y con una T protuberante en el
extremo 3’ de cada cadena que puede hibridar con la A que algunas DNA polimerasas
termoestables introducen en el extremo 3’ del DNA amplificado) y se secuenció,
determinándose así los aproximadamente 1000 nt del extremo 3’ del mRNA candidato a
codificar la tRNA ligasa de tomate.
Para amplificar la región terminal 5’ de este mRNA se emplearon diversas
estrategias basadas en RT-PCR. La primera consistió en añadir mediante la enzima
transferasa terminal una cola poli(dA) en el extremo 3’ del cDNA generado por RT con
el cebador C7, derivado de la secuencia SGN-U223941 (Frohman et al., 1988). Para la
amplificación por PCR se empleó el mismo cebador que en la RT y otro con una cola
poli(dT) en 3’ (C2), llevándose a cabo una segunda PCR con los cebadores C8
(derivado de una posición interna de la secuencia SGN-U223941) y C4
(complementario a C2).
En segundo lugar se aplicó la metodología de amplificación rápida de los
extremos de un cDNA mediada por RNA ligasa (RNA ligase-mediated rapid
amplification of cDNA ends, RLM-RACE) para la que un adaptador de RNA (C9) de
secuencia conocida se unió a los mRNAs mediante la T4.1 RNA ligasa tras eliminar la
98

Resultados
estructura de caperuza de los mismos (Liu y Gorovsky, 1993). El cDNA obtenido por
RT con el cebador C7 se amplificó por PCR con este mismo cebador y otro (C10)
complementario a la secuencia del adaptador de RNA (C9) ligado al extremo 5’. Una
segunda PCR se llevó a cabo con cebadores más internos (C8 y C11), el primero
derivado de SGN-U223941 y el otro complementario de C9.
En tercer lugar, para aplicar la metodología denominada Cap-finder basada en la
capacidad de algunas transcriptasas inversas de añadir hasta cuatro Cs cuando alcanzan
la estructura de caperuza (Schmidt y Mueller, 1999; Schramm et al., 2000), la
transcripción inversa se efectuó con el cebador específico C7 en presencia de un
adaptador de DNA (C12) de secuencia conocida que contiene una cola poli(dG) en 3’
(Schramm et al., 2000). Durante la reacción, el adaptador hibrida con las Cs que la
transcriptasa inversa añade cuando se encuentra la estructura de caperuza, y esta misma
enzima continúa la reacción transcribiendo la secuencia del adaptador. El cDNA
obtenido se amplificó por PCR con el cebador C7 y con otro complementario a la
secuencia del adaptador introducida en 5’ (C4). Una segunda reacción de amplificación
se llevó a cabo con cebadores más internos (C6 y C8), el primero complementario a
C12 y el segundo derivado de la secuencia SGN-U223941.
Finalmente, un último intento de amplificación se acometió obteniendo el cDNA
con el cebador específico C7 y empleando para la PCR este mismo cebador y otro
degenerado (C14) diseñado a partir de una secuencia próxima al extremo 5’ de la región
codificante que está muy conservada en todas las tRNA ligasas conocidas de plantas.
Una segunda reacción de amplificación se llevó a cabo con el cebador específico C8 y
otro degenerado (C13), ambos más internos que los empleados en la primera reacción.
Con estas técnicas de RACE aplicadas a un molde lineal, o bien no se obtuvo
ningún producto detectable mediante tinción con bromuro de etidio tras separar los
productos de la PCR por electroforesis en un gel de agarosa, o bien la amplificación fue
inespecífica. Una posible explicación a estos resultados negativos sería que la
concentración del mRNA candidato es baja. Además, el fragmento que se pretendía
transcribir inversamente era bastante grande (aproximadamente 2500 nt), aumentando la
probabilidad de encontrar un elemento de estructura secundaria en el mRNA que
desacoplara la transcriptasa inversa (Schaefer, 1995). Por otra parte, la amplificación
inespecífica con estas técnicas es consecuencia de emplear un cebador derivado de una
secuencia introducida durante la transcripción reversa (o en pasos posteriores) del
mRNA. A pesar de realizarse sobre un cDNA transcrito con un cebador específico, se
99

Resultados
ha observado que este procedimiento disminuye la especificidad de la reacción con
respecto a las PCRs realizadas con los dos cebadores complementarios de dicha
secuencia, y esa es una de las razones del desarrollo de las técnicas de RACE basadas
en un molde circular (ver más adelante). Estas técnicas permiten emplear cebadores
específicos en ambas direcciones (Fu et al., 2004; Polidoros et al., 2006), por lo que se
procedió a ensayar las mismas.
4.2.2.2. Empleo de la técnica de RACE circular para determinar la región 5’ del
mRNA candidato a codificar la tRNA ligasa de tomate
La amplificación de la región terminal 5’ del presumible mRNA de la tRNA
ligasa de tomate se obtuvo con una técnica de RACE consistente en la circularización
del cDNA molde previa a la PCR (Fu et al., 2004). En un primer intento mediante
digestión suave con RNasa H se introdujeron una serie de cortes en el híbrido
mRNA:cDNA obtenido por transcripción inversa con el cebador C2 previamente
fosforilado. La segunda cadena de cDNA se sintetizó siguiendo el protocolo de
Okayama y Berg (1982), y el producto bicatenario se circularizó con T4 DNA ligasa. La
aplicación sobre este molde de dos PCRs consecutivas con dos pares de cebadores
divergentes derivados de los extremos de la secuencia SGN-U223941 (C3-C7 y C5-C8)
condujo a un producto de amplificación específico, que si bien era de tamaño inferior al
esperado (2500 pb aproximadamente), sirvió como punto de partida para adaptar esta
técnica de RACE circular a nuestro propósito.
En un siguiente intento se introdujo una modificación consistente en emplear la
T4.1 RNA ligasa para circularizar directamente el cDNA de simple cadena (Snopek et
al., 1976; McCoy y Gumport, 1980). En paralelo a esta modificación, se llevó a cabo un
experimento control siguiendo el protocolo para cDNA de doble cadena con el fin de
comparar los resultados obtenidos. En ambos casos, con cDNA molde de simple o doble
cadena, se efectuaron dos PCRs con dos pares de cebadores adyacentes y divergentes
(Fig. 4.16). Empleando las parejas de cebadores C3 y C7 primero, y C5 y C8 después,
se obtuvo amplificación con ambos moldes, siendo el fragmento generado mayor con el
cDNA de simple cadena, con la ventaja adicional de que el procedimiento resulta más
sencillo.
La técnica de RACE circular con cDNA molde de simple cadena permitió
obtener productos de amplificación de entre 500 y 900 pb Debido a que la secuencia del
100

Resultados
mRNA que se pretendía amplificar era de aproximadamente 2500 nt, fue necesario
reiterar la aplicación varias veces para alcanzar el extremo 5’ del mismo.
Después de cada experimento se comprobó que el fragmento amplificado
pertenecía al presumible mRNA de la tRNA ligasa de tomate clonándolo en el plásmido
pTZ57R/T, secuenciándolo y alineando esta secuencia con las de las tRNA ligasas
presuntamente homólogas. A partir de cada nueva región secuenciada se diseñaron
cebadores (C15-C27) que permitieron aplicar de nuevo la técnica hasta amplificar la
totalidad del mRNA.
5’-CAP 3’-OH mRNA
Fig. 4.16. Esquema del protocolo adaptado de RACE circular. A partir de un fragmento de secuencia interna de un mRNA es posible amplificar el resto con cebadores derivados de la misma tanto en dirección 5’ como 3’. La línea roja representa la secuencia conocida, y la negra y azul, las desconocidas. La flecha verde indica el cebador que genera el cDNA durante la RT, y las flechas azul turquesa y negra los cebadores divergentes derivados de la secuencia conocida empleados durante la PCR. La región en azul del mRNA puede amplificarse con un cebador con una cola poli(dT) en 3’ y cebadores derivados del fragmento conocido.
RT
3’-OH
PCR
Ligación
5’-P cDNA
3’-OH 5’-OH DNA
amplificado 5’-OH 3’-OH
101

Resultados
4.2.2.3. Amplificación, secuenciación y análisis del cDNA completo de la
presumible tRNA ligasa de tomate
Cuando el producto de amplificación obtenido acabó en el mismo nucleótido en
dos experimentos de RACE circular independientes, se alineó su secuencia con la de los
cDNAs de las tRNA ligasas de plantas conocidas empleando el algoritmo ClustalW
(http://www.ebi.ac.uk/clustalw/) (Chenna et al., 2003), y se localizó por comparación el
codón de iniciación más probable. Este es el primer codón AUG que se encuentra en la
secuencia considerando la fase en la que se traduce la proteína, mientras que el siguiente
codón AUG se encuentra inmerso en una región muy conservada en todas las tRNA
ligasas y cercana a la que codifica el motivo de adenilación de la enzima. Este codón
inicial más probable no se ajusta bien a las reglas de Kozak que determinan el sitio de
inicio de la traducción (Kozak, 1986; Lütcke et al., 1987; revisado en Fütterer y Hohn,
1996), si bien ocurre lo mismo con las tRNA ligasas de plantas descritas (Tabla 4.1).
Tabla 4.1. Reglas de Kozak para vertebrados y plantas, y su aplicación al sitio de inicio de la traducción de las tRNA ligasas de plantas. En azul se indican los nucleótidos que siguen la regla en las cuatro enzimas analizadas, en rojo los nucleótidos que cumplen la regla en alguna de las cuatro enzimas, y en verde los nucleótidos que son comunes a dos tRNA ligasas pero no se ajustan a la regla.
-5 -4 -3 -2 -1 +1 +2 +3 +4 +5
Vertebrados C C A/G C C A U G G - Plantas - A A C A A U G G C
Con
sens
o
Tomate C A C U C A U G C C
A. thaliana C G A A G A U G G A Arroz C U U A U A U G U C
tRN
A li
gasa
Trigo C U U A/G U A U G U C
Las reglas de Kozak adaptadas a plantas dan importancia a la presencia de una C
en posición +5 y una G en posición +4 (Lütcke et al., 1987). La primera puede
encontrarse en todas las secuencias de tRNA ligasas de plantas conocidas excepto en A.
thaliana, mientras que la segunda sólo se observa en esta última especie. Por otro lado,
hay una C en posición -5 no contemplada en las reglas adaptadas a plantas, que sin
embargo parece tener un papel importante pues es el único nucleótido conservado en la
región en torno al codón inicial en las cuatro especies (Tabla 4.1).
102

Resultados
En animales se ha observado que la eficiencia durante el inicio de la traducción
aumenta en un entorno subóptimo con respecto a las reglas de Kozak si puede formarse
una pequeña horquilla cercana a la posición +14 (Kozak, 1990). Este elemento de
estructura secundaria parece que actúa prolongando el tiempo que necesita el ribosoma
para localizar el codón de inicio, aumentando así la eficiencia del reconocimiento. Sin
embargo, el análisis con el programa Mfold (http://frontend.bioinfo.rpi.edu/zukerm/cgi-
bin/rna-index.cgi) de la región en torno al codón AUG inicial de los mRNAs que
codifican las tRNAs ligasas de plantas conocidas no predijo ninguna horquilla próxima
a la posición +14.
La región 5’ no traducible del presumible mRNA de la tRNA ligasa de tomate se
comparó con la de los mRNAs homólogos de trigo, arroz y A. thaliana, comprobándose
que su longitud se encuentra comprendida entre los valores máximos y mínimos de las
regiones equivalentes de las otras tRNA ligasas conocidas (Tabla 4.2). A este respecto
cabe destacar que la longitud de la región 5’ no traducible de la isoforma 1 de germen
de trigo es apreciablemente menor que la de las otras dos isoformas de este organismo y
sus homólogas de arroz y A. thaliana, por lo que es probable que esté incompleta.
Teniendo en cuenta estas observaciones, se asumió que la secuencia del mRNA
de la presumible tRNA ligasa de tomate era de longitud completa o casi completa.
Una vez conocidos los extremos, se diseñaron los cebadores C28 y C29 para
amplificar la secuencia codificante en su totalidad con una mezcla de DNA polimerasas
termoestables compuesta por Taq (muy procesiva) y Tgo (con actividad correctora de
errores). Esta mezcla permite amplificaciones eficientes y con una baja tasa de
mutación. La pauta abierta de lectura (open reading frame, ORF) completa, clonada en
el plásmido de expresión pET23a (+) (ver más adelante), se secuenció con siete
cebadores (T7, C8, C15, C16, C21, C27 y C28). Esta información, junto a la obtenida
previamente con los experimentos de RACE circular y 3’-RACE, permitió establecer la
secuencia completa del mRNA de la presunta tRNA ligasa de tomate (Fig. 4.17).
103

Resultados
Tabla 4.2. Regiones 5’ no traducibles de los mRNAs de las tRNA ligasas de plantas.
Especie Secuencia
Tomate GAGACCAUCUCCCUCACAAUCUCUCUUCUUCUUCUUCCUCAUAUUUCCCUUCUAGAAUCUUCUUUCGCUUUCAAUCUCGUUCCCUCACACACAGUACCUCGCUAUUCUCUUCACUC
A. thaliana GUGUGUUUUUUACUUUUCUGCAACUAAAUCGUGUUUCUUCCUAAGAACCCUA
AAUCUGAACUGUCUCAUCUUCUCCGAUAUCAGUGAGAUCAAUAAAUCAUCUUCUCACCCUAGACGACAGGGAAACUCGGGGAUUGAUUCGGAAAAACGAGUUCGUUUUUCUGCAAGAAAAAGAGAGAUCACGCUGAGCAGAAGUGGCAAGUAAAACCGAAG
Arroz GACCCAUUCUCCCCUCUCUCCGCCGAACCCUCCAGACUCCGCCGUGCCACGCACGGCGGCGCCGCCCCUCUAAACCCACCAACUCCGGCGCCGCCUCGCCCCCGCACUUAU
Trigo 1 UCCCCACCGACUUCCAUCCCCCGCCCCGGCGUCGUGCCCGGCACUUAU
Trigo 2 AAUAAUGCCAUUUGCAGGCGAGCCUCAAAAAUGGCCCGGACCAAGUCUUAGC
AUUAGCAGCAUUCCUGCCACCGCCGCUAAACCCUCCUGAACCCGCCGCGCGUAGGCGGCGCCGCUUUCCCCACCGACUUCCAUCCCCCGCCCCGGCGUCGUGGCCGGCACUUAU
Trigo 3 AAUAGUGCCAUUUUCAGGCGAGCCUCGGGCCGUCCCAAAUGGCCCGGCCCAAGUCUUAGCAUUAGCAUUCCUGCCACCGCCGCUAAACCCUCCUGAACCUGCCGUGCCUUCGGCGGCGCCACUUCCCCACCGACUUCUUCCAUCCCCCGCCCCGGCGUGGCCAGCACUUGU
El mRNA completo se tradujo in silico (http://www.bioinformatics.org/sms), y
la secuencia resultante se alineó con la de las tRNA ligasas de plantas conocidas (A.
thaliana, arroz y las tres isoformas de trigo, Fig. 4.18) empleando el algoritmo
ClustalW. En arroz y trigo, ambas monocotiledóneas, la secuencia en torno al codón
inicial de la traducción es idéntica, mientras que en tomate y A. thaliana, las dos
dicotiledóneas, dicha secuencia no se parece entre sí y tampoco se asemeja a la
compartida por trigo y arroz.
El análisis de la secuencia aminoacídica de la presumible tRNA ligasa de tomate
permitió localizar los residuos conservados característicos de los diferentes dominios
funcionales de las tRNA ligasas: adenililtransferasa/ligasa, polinucleótido quinasa y
ciclofosfodiesterasa (Fig. 4.18). La enzima comparte además una identidad mayor del
62% con las otras tRNA ligasas de plantas conocidas, a pesar de que la región amino
terminal de éstas es muy variable. La región carboxilo terminal también presenta mayor
variabilidad, aunque mucho menor que la de la región amino terminal.
104

Resultados
GAGACCATCTCCCTCACAATCTCTCTTCTTCTTCTTCCTCATATTTCCCTTCTAGAATCTTCTTTCGCTTTCAATCTCGTTCCCTCACACACAGTACCTCGCTATTCTCTTCACTCATGCCAAACAATCAGAGAAAGGGTGGTTATAAAGAAAAAAAATGGCAAGTGAGGAGTTCCAATAGGGTACCAGGTTCATCTTCAAATGTCGAACCTGCATCTCCTGCAACTACTGGAGCCATTGCTGACCGTCTAAGTTCCTTGAATATTACTGAAAGTGGTGCACAGTCTAGTGTTCCAGTTGCATCTCTTCAGTTTGGCAGCGTTGGATTAGCACCCCAATCACCTGTGCAGCATCAAAAAGTCATCTGGAAACCCAAGTCATATGGAACAGTGTCTGGAGCCCCAAAGATTGAAGCTGTAAAAACACCAAATGAACAAAAAAGTGCTCTTTTGAGCAAATTGTTTAAGGGTAGTTTATTGGAAAATTTTACTGTAGATAACTCCACATTCTCAAAAGCCCAAATAAGGGCCACTTTCTACCCAAAATTTGAGAATGAGAAATCAGATCAGGAGATTAGGACAAGGATGATAGAGATGGTCTCCAAAGGCTTGGCTACAGTGGAGGTCTCTCTTAAGCATTCTGGATCTCTTTTTATGTATGCTGGGCATAAGGGTGGAGCATATGCCAAGAATAGCTTTGGAAATATCTATACTGCCGTTGGTGTCTTTGTTCTTGGACGGATGCTTCGTGAGACATGGGGTACTCAAGCAAGTAAGAAGCAAGCAGAGTTCAATGAGTTTCTTGAGCGCAATCGCATGTGTATATCAATGGAGTTGGTCACGGCAGTGTTGGGGGACCACGGACAACGCCCACGAGATGATTATGCGGTTGTGACTGCAGTCACGGAGTTGGGAAGTGGAAAACCAAATTTCTATTCAACTCCCGATGTAATTGCTTTTTGCAGAGAATGGAGATTACCAACAAATCATATATGGCTGTTCTCAACAAGGAAATCAGTTACTTCCTTCTTTGCTGCGTTTGATGCACTCTGTGAGGAAGGTACAGCAACCTCTGTTTGCCAGGCTCTTGCTGAAGTTGCTGACATTTCTGTACCTGGATCAAAAGACCATATAAAAGTGCAGGGTGAAATTTTGGAGGGCCTCGTGGCACGCATTGTAAAACGTGAGAGCTCAGAGCATATGGAGCGGGTTCTAAGAGATTTTCCTCCTCCTCCATTAGAGGGTGAGGGTTTGGACTTGGGACCTACGCTGCGTGAGGTTTGTGCTGCAAATAGATCAGAAAAGCAGCAAATCAAGGCGCTTCTTCAGAGTGCTGGCACGGCTTTCTGCCCGAATTATTTGGACTGGTTTGGAGATGATGATTCTGGCTCACATTCAAGAAATGCTGATCGCTCTGTTGTGTCAAAGTTCTTACAGTCACATCCTGCTGATTTTTCTACAGGAAAATTACAGGAAATGGTTCGTTTGATGGGGGAAAAACGCTTTCCTGCTGCTTTCAAGTGTTACTATAACTTCCATAAAATTAATGATTTATCGAGTGACAACCTGCCTTTCAAAATGGTGATCCACGTACATAGTGATTCAGGCTTCCGCCGGTACCAGAAAGAGATGAGACACAAACCAGGACTGTGGCCTTTGTATCGAGGCTTTTTTGTTGACCTGGATTTATTCAAGGTCAATGAGAAGAAAACTGCCGAAATGGTAGGAAGTAGCAATCAAATGGTAAAAAATGAGGAGGAGGATAGCCGTTTAGCTGATGAAGATGCAAATTTGATGGTCAAGATGAAATTCCTTCCTTACAAGTTGAGAACTTTTTTGATCCGTAATGGCTTGTCGACTCTTTTCAAAGAAGGCCCTTCTGCGTATAAGGCTTATTACCTGAGGCAAATGAAAATATGGAATACTTCAGCAGCCAAGCAACGAGAACTCAGCAAGATGCTTGATGAATGGGCAGTATATATACGCAGAAAATATGGGAACAAACCATTGTCATCATCCACATACCTAAGTGAAGCTGAGCCCTTCCTTGAGCAATATGCAAAGTGTAGTCCACAAAATCAGGCGTTGATAGGATCTGCTGGAAATTTTGTCAAAGTTGAAGACTTCATGGCTATTGTTGAAGGAGAAGATGTAGAGGGCGATCTTGAGCC GACCAAAGATATTGCTCCTTCAAGCCCTAATATTTCCAGCAAAGACATGGTCGCAAAGAATGAGGGTCTCATCGTTTTTTTTCCAGGAATACCAGGTTGTGCTAAATCTGCACTTTGTAAGGAAATACTGAATGCTCCAGGAGGGCTTGAAGATGATCGGCCAATTCATAGTTTAATGGGTGATCTTATCAAAGGTAGATATTGGCAAAAAGTTGCTGATGAACGTCGAAGAAAACCTTACTCGATCATGCTTGCTGACAAGAATGCACCAAATGAGGAAGTATGGAAACAAATTGAGAACATGTGTCTAAGCACCAAAGCATCTGCTATTCCAGTTATACCTGATTCAGAAGGAACTGAAATCAATCCATTCTCTATTGATGCACTGGCGGTTTTTATATTCCGAGTACTTCAGCGTGTTAATCATCCGGGAAATCTTGACAAGTCATCTCCAAATGCTGGATATGTGATGTTAATGTTTTATCACCTTTATGATGGAAAGAGCCGTCAGGAGTTCGAGAGTGAACTTATTGAACGTTTTGGATCACTTGTCAGAATTCCTCTACTGAAACCTGAAAGGTCTCCTCTTCCGGATTCTGTGAGGTCTATTGTTGAGGAGGGAATCAATCTGTACAGGCTTCATACTAACAAACATGGAAGATTGGAGTCTACAAAAGGGACTTTTGTAAAAGAGTGGGTTAAATGGGAGAAGCAATTGAGGGATATTCTACATGGAAATGCGGACTATCTCAATTCAATACAGGTTCCATTTGAATTTGCTGTTAAAAAAGTCCTTGAACAACTGAAAGCTATTGCGAGGGGCGAATATGCAGCTCCTTCTTCTGAGAAGAGGAAGCTAGGATCCATTGTGTTTGCCGCTATCAGCCTGCCAGTTCCAGAAATTCTAGGTCTTCTGAATGATCTAGCCAAGAAAGATCTGAAGGTTGGTGATTTCTTGAAGGACAAAAGCCTGGAGAGCTGCATTCAGAAAGCCCATTTGACCCTGGCTCACAAGAGAAGTCATGGTGTCACTGCAGTTGCCAATTACGGTTCCTTTCTTCACCAAAACGTCCCAGTAGACGTGGCTGCTTTGTTGTTCTCCGATAAATTGGCTGCCCTAGAAGCTGAGCCTGGTTCTGTCGAAGGTGAAAAGGTCGATTCTAAAAACCCATGGCCCCACGTCACAATATGGACTGGGGCAGGAGCTACAGCCAAAGATGCCAATACACTACCACAGTTACTTTCTCAAGGGAAGGCTATCCGCATTGATATTAATCCACCCGTCACTATAACTGGCACTCTCGAATTCTTTTGAACTCTACATGTAATTCCTCATCTTGTGTGGATTTACCTAATGTAAACCCTACATGGGTTCTGAGTGAATATCATGTAATGACACTCCCGAGTGGTAGGAACAATTTTGTAGGATCGTGCGCTTTGGCTATATAGCAGTCTTGCAGTTGCAGGATAGAAATAGATTTAGAGAAAATGTAAAGATGTTGAGGCAATAAAGGTCTTAAGTCGTGGGCGTTTTGAAATCCCCGATGGCATTTCTTGAATAAAGCACACCATTTTCTACTCAAAAAAAAAAAAAA
Fig. 4.17. Secuencia completa del cDNA correspondiente al mRNA de la presumible tRNA ligasa de tomate obtenida por RT-PCR a partir de RNA poliadenilado de hojas de tomate. Las regiones 5’ y 3’ no traducidas aparecen en azul y la secuencia codificante en negro.
105

Resultados
106
Lig I
Lig IIIa
Lig IV
T. aestivum 1 MSLS----------LSLSHSPRRFLPLLLP----------APPFAMPPRRD--HGKQTQQ 38 T. aestivum 2 MSLS----------LSLSHSPRRFLPLLLP----------APPFAMPPRRD--HGKQTQQ 38 T. aestivum 3 MSLS----------LSLSHSPRRFLPLLLP----------APPFAMPPRRD--HGKQAHQ 38 O. sativa MSLSSTSTSSLLLLLSPRGNPRRLLPLLLPLRLSSSSSAAAAAAAMPPRRDRHHGKQPQQ 60 A. thaliana MDAPFESG------------------------------DSSATVVAEAVNNQFGGLSLKE 30 S. lycopersicum MPNNQRKGGYKEKKWQVRSSNRVPGSSSNV--------EPASPATTGAIADRLSSLNITE 52 . :.. . . : . . : T. aestivum 1 RWKPKAAPAPS----------------------SAAVERMALAPSTAG-AAQVWVPRGYA 75 T. aestivum 2 RWKPKATPTPAPAPAPAPS--------------SAAVERMSLAPSTAG-AAQVWVPRGYA 83 T. aestivum 3 RWKPKAAPAPAPAPAPAPS--------------SAAVERMALAPSAAG-AAQVWVPRGYA 83 O. sativa QWKPKATDPAAAAPAVVAAASSPASSTSSSADFAGGVERMTISPSPQGGASQVWVPRGYA 120 A. thaliana S--NTNAPVLP----------------------SQT-------TSNHRVQNLVWKPKSYG 59 S. lycopersicum SGAQSSVPVAS----------------------LQFGSVGLAPQSPVQHQKVIWKPKSYG 90 . . . * :* *:.*. T. aestivum 1 TSASS--------SSSS-AVTAAEQGGAGDKLSRLIKG--AAEFSVDNNTFTEAQIRATF 124 T. aestivum 2 TSASS--------SSSS-AVAAAEQGGASDKLSRLIKG--AAEFSVDNNTFTEAQIRATF 132 T. aestivum 3 TSASS--------SSSS-AVTAAEQGGAGDKLSRLIKG--AAEFSVDNNTFTEAQIRATF 132 O. sativa TSASS--------SSSSSSAAAAEQRIDAEKLSRVFKA--APNFEVDNNTFIQSQIRATF 170 A. thaliana TVSGSSSATEVGKTSAVSQIGSSGDTKVGLNLSKIFGGNLLEKFSVDKSTYCHAQIRATF 119 S. lycopersicum TVSG---------APKIEAVKTPNEQKSAL-LSKLFKGSLLENFTVDNSTFSKAQIRATF 140 * :. :. :. : . **::: . :* **:.*: .:****** T. aestivum 1 YPKFENEKSDQEKRTRMIEIVSQGLATIEVTQKHSGSLFMYAGHRGGAYAKNSFGNIFTA 184 T. aestivum 2 YPKFENEKSDQETRTRMIEIVSQGLATIEVTQKHSGSLFMYAGHRGGAYAKNSFGNIFTA 192 T. aestivum 3 YPKFENEKSDQETRTRMIEIVSQGLATIEVTQKHSGSLFMYAGHLGGAYAKNSFGNIFTA 192 O. sativa YPKFENEKSDQETRTRMIEMVSHGLATLEVTLKHSGSLFMYAGHHGGAYAKNSFGNIYTA 230 A. thaliana YPKFENEKTDQEIRTRMIEMVSKGLATLEVSLKHSGSLFMYAGHKGGAYAKNSFGNIYTA 179 S. lycopersicum YPKFENEKSDQEIRTRMIEMVSKGLATVEVSLKHSGSLFMYAGHKGGAYAKNSFGNIYTA 200 ********:*** ******:**:****:**: ************ ************:** T. aestivum 1 VGVFVLGRLFHEAWGGKAPKMQAEFNDFLEKNRICISMELVTAVLGDHGQRPKDDYAVVT 244 T. aestivum 2 VGVFVLGRLFREAWGSKAPKMQAEFNDFLEENRICISMELVTAVLGDHGQRPKDDYAVVT 252 T. aestivum 3 VGVFVLGRLFREAWGSKAPKMQAEFNDFLEKNRICISMELVTAVLGDHGQRPKDDYAVVT 252 O. sativa VGVFVLGRLFREAWGKEAPRMQEEFNVFLEKKCISISMELVTAVLGDHGQRPKDDYAVIT 290 A. thaliana VGVFVLSRMFREAWGTKAPKKEAEFNDFLEKNRMCISMELVTAVLGDHGQRPLDDYVVVT 239 S. lycopersicum VGVFVLGRMLRETWGTQASKKQAEFNEFLERNRMCISMELVTAVLGDHGQRPRDDYAVVT 260 ******.*:::*:** :*.: : *** ***.: :.***************** ***.*:* T. aestivum 1 AVTELGHGKPQFYSTPEVISFCRKWRLPTNHVWLFSTRKSATSFFAAYDALCEEGTATPV 304 T. aestivum 2 AVTELGHGKPQFYSTPEVISFCRKWRLPTNHVWLFSTRKSATSFFAAYDALCEEGTATPV 312 T. aestivum 3 AVTELGHGKPQFYSTPEVISFCRKWRLPTNHVWLFSTRKSATSFFAAYDALCEEGTATPV 312 O. sativa AVTELGHGKPKFYSTPEVIEFCRKWRLPTNHVWLFSTRKSASSFFAAYDALCEEGTATSV 350 A. thaliana AVTELGNGKPQFYSTSEIISFCRKWRLPTNHVWLFSTRKSVTSFFAAFDALCEEGIATSV 299 S. lycopersicum AVTELGSGKPNFYSTPDVIAFCREWRLPTNHIWLFSTRKSVTSFFAAFDALCEEGTATSV 320 ****** ***:****.::* ***:*******:********.:*****:******* **.* T. aestivum 1 CKALDEIADISVPGSKDHVMVQGEILEGLVARIVSRESSVQMEEVLRNFPIPSLDGGDSD 364 T. aestivum 2 CKALDEIADISVPGSKDHVMVQGEILEGLVARIVSRESSVQMEEVLRNFPIPSLDGGDSD 372 T. aestivum 3 CKALDEIADISVPGSKDHVMVQGEILEGLVARIVSRESSVQMEEVLRNFPIPSLDGGDSD 372 O. sativa CKALDEIADVAVPGSKDHVKVQGEILEGLVARIVSRESSVQIEEVLRNYPLPPLDGVGSD 410 A. thaliana CRALDEVADISVPASKDHVKVQGEILEGLVARIVSSQSSRDMENVLRDHPPPPCDGANLD 359 S. lycopersicum CQALAEVADISVPGSKDHIKVQGEILEGLVARIVKRESSEHMERVLRDFPPPPLEGEGLD 380 *:** *:**::**.****: **************. :** .:*.***:.* *. :* . * T. aestivum 1 LGPSLRDICAANRSDEKQQIKSLLENVGSSMCPDHRDWFGYSGLE-AQSRNADKSVVTHF 423 T. aestivum 2 LGPSLRDICAANRSDEKQQIKALLENVGSSMCPDHRDWFGYSGLE-PQSRNADKSVVTHF 431 T. aestivum 3 LGPSLRDICAANRSDEKQQIKALLENVGSSMCPDHRDWFGYSGLE-PQSRNADKSVVTHF 431 O. sativa LGPSLREICAANRSDEKQQIKALLENVGPSMCPDHSDWFGCSGLDDHQSPSANRSVVTKF 470 A. thaliana LGLSLREICAAHRSNEKQQMRALLRSVGPSFCPSDVEWFGDE----SHPKSADKSVITKF 415 S. lycopersicum LGPTLREVCAANRS-EKQQIKALLQSAGTAFCPNYLDWFGDDDSG-SHSRNADRSVVSKF 438 ** :**::***:** ****:::**...*.::**. :*** . :. .*::**:::*

Resultados
PNK
T. aestivum 1 LQAHPTDYATKKLQEMIGLMKRKNFSASFKSYWNYQKVDSLSNDNLCYKMVIHVYSDSVF 483 T. aestivum 2 LQAHPTDYATKKLQEMIGLMKRKNFSASFKSYWNYQKVDSLSNDNLCYKMVIHVYSDSVF 491 T. aestivum 3 LQAHPTDYATKKLQEMIGLMKRKNFSASFKSYWNYQKVDSLSNDNLCYKMVIHVYSDSVF 491 O. sativa LQAHPTDYTTKKLQEMIRVMKQRNFPAAFKCYWNYHKIDSLSNDSLYYKMVIHVLSDSVF 530 A. thaliana LQSQPADYSTSKLQEMVRLMKEKRLPAAFKCYHNFHRAEDISPDNLFYKLVVHVHSDSGF 475 S. lycopersicum LQSHPADFSTGKLQEMVRLMGEKRFPAAFKCYYNFHKINDLSSDNLPFKMVIHVHSDSGF 498 **::*:*::* *****: :* .:.:.*:**.* *::: :.:* *.* :*:*:** *** * T. aestivum 1 RRYQQEMRKNQELWPLYRGFFVDVNLFKANNKKAAELSKDSNTLLRNINGALDSSLSSKD 543 T. aestivum 2 RRYQQEMRKNQELWPLYRGFFVDVNLFKANNKKAAELSKDSNTLLRNINGALDSSLSSKD 551 T. aestivum 3 RRYQQEMRKNQELWPLYRGFFVDVNLFKANNKKAAELSKDSNTLLRNINGALDSSLSSKD 551 O. sativa RRYQQEMRRNQGLWPLYRGFFVDVNLFKVNNMKSSIPSEDIDTSLKNINGALDSNSSAKD 590 A. thaliana RRYHKEMRHMPSLWPLYRGFFVDINLFKSNKGRDLMALKSIDNASE------NDGRGEKD 529 S. lycopersicum RRYQKEMRHKPGLWPLYRGFFVDLDLFKVNEKKTAEMVGSSNQMVK------NEE--EDS 550 ***::***: ***********::*** *: : . : . :. .. T. aestivum 1 GLADEDSNLMVKLKFLTYKIRTFLIRNGLSTLFKDGPSAYRTYYLRQMKIWGTSASKQKE 603 T. aestivum 2 GLADEDSNLMVKLKFLTYKIRTFLIRNGLSTLFKDGPSAYKTYYLRQMKIWGTSASKQKE 611 T. aestivum 3 GLADEDSNLMVKLKFLTYKIRTFLIRNGLSTLFKDGPSAYRTYYLRQMKIWGTSASKQKE 611 O. sativa GLADEDSNLMVKLKFLTYKLRTFLIRNGLSTLFKDGPSAYKTYYLRQMKNWGTSASKQKE 650 A. thaliana GLADDDANLMIKMKFLTYKLRTFLIRNGLSILFKDGAAAYKTYYLRQMKIWGTSDGKQKE 589 S. lycopersicum RLADEDANLMVKMKFLPYKLRTFLIRNGLSTLFKEGPSAYKAYYLRQMKIWNTSAAKQRE 610 ***:*:***:*:***.**:********** ***:*.:**::******* *.** .**:* T. aestivum 1 LTKMLDEWAVYIRRKYQNKQLPSSTYLSEAEPFLEQYAKRSPANQALIGAAGDLVQTENF 663 T. aestivum 2 LTKMLDEWAVYIRRKYQNKQLPSSTYLTEAEPFLEQYAKRSPANQALIGAAGDLVQTENF 671 T. aestivum 3 LTKMLDEWAVYIRRKYQNKQLPSSTYLSEAEPFLEQYAKRSPANQALIGAAGDLVQTENF 671 O. sativa LSKLLDEWAVYIRRKYGNKPLSSSTYLSEAEPFLEQYAKRSPENQALIGAAGDLVQTENF 710 A. thaliana LCKMLDEWAAYIRRKCGNDQLSSSTYLSEAEPFLEQYAKRSPKNHILIGSAGNLVRTEDF 649 S. lycopersicum LSKMLDEWAVYIRRKYGNKPLSSSTYLSEAEPFLEQYAKCSPQNQALIGSAGNFVKVEDF 670 * *:*****.***** *. *.*****:*********** ** *: ***:**::*:.*:* T. aestivum 1 LAILDAQRDEEGDLQPERGTAPSSPTSTSLDVVSKTEGLIVFFPGIPGCAKSALCEQILN 723 T. aestivum 2 LAILDAQRDEEGDLQPERGTAPSSPTSTSLDVVSKTEGLIVFFPGIPGCAKSALCEQILN 731 T. aestivum 3 LAILDAQRDEEGDLQAERGTAPSSPTSTSLDVVSKTEGLIVFFPGIPGCAKSALCEQILN 731 O. sativa LAILEAKRDEEGDLQAERGTAPPSPTSTSLDVVPKAEGLIVFFPGIPGCAKSALCKEILT 770 A. thaliana LAIVDGDLDEEGDLVKKQGVTPATPEPAVKEAVQKDEGLIVFFPGIPGSAKSALCKELLN 709 S. lycopersicum MAIVEGE-DVEGDLEPTKDIAPSSPNISSKDMVAKNEGLIVFFPGIPGCAKSALCKEILN 729 :**::.. * **** :. :*.:* : : * * ************.******:::*. T. aestivum 1 TPGGLGDNRPLHSLMGDRTKGRYWQKVADERKKKPFRITLADKNAPNEEVWRQIEDMCGM 783 T. aestivum 2 TPGGLGDNRPLHSLMGDRTKGRYWQKVADERKKKPFRITLADKNAPNEEVWRQIEDMCGM 791 T. aestivum 3 TPGGLGDNRPLHSLMGDRTKGRYWQKVADERKKKPFRITLADKNAPNEEVWRQIEDMCGM 791 O. sativa TPGGLGDNRPLHSLMGDLIKGRYWQKVADERKKKPFRITLADKNAPNEEVWRQIEDMCRT 830 A. thaliana APGGFGDDRPVHTLMGDLVKGKYWPKVADERRKKPQSIMLADKNAPNEDVWRQIEDMCRR 769 S. lycopersicum APGGLEDDRPIHSLMGDLIKGRYWQKVADERRRKPYSIMLADKNAPNEEVWKQIENMCLS 789 :***: *:**:*:**** **:** ******::** * *********:**:***:** T. aestivum 1 TKAAAVPVIPDSEGTDSNPFSLEALAVFMFRVLQRVNHPGNLDKASPNAGYILLMFYNLY 843 T. aestivum 2 TKAAAVPVIPDSEGTDSNPFSLEALAVFMFRVLQRVNHPGNLDKASPNAGYILLMFYNLY 851 T. aestivum 3 TKAAAVPVIPDSEGTDSNPFSLEALAVFMFRVLQRVNHPGNLDKSSPNAGYILLMFYNLY 851 O. sativa TKAAAVPVIPDSEGTDSNPFSLDALAVFMFRVLQRDNHPGNLDKASPNAGYVLLMFYNLY 890 A. thaliana TRASAVPIVADSEGTDTNPYSLDALAVFMFRVLQRVNHPGKLDKESSNAGYVLLMFYHLY 829 S. lycopersicum TKASAIPVIPDSEGTEINPFSIDALAVFIFRVLQRVNHPGNLDKSSPNAGYVMLMFYHLY 849 *:*:*:*::.*****: **:*::*****:****** ****:*** *.****::****:** T. aestivum 1 DGKSRREFESELYERFGSLVKMPLLKPERAPLPGDVKTILDEGMSLFRLHQSRHGRAEPS 903 T. aestivum 2 DGKCRREFESELYERFGSLVKMPLLKPERAPLPGDVKTILDEGMSLFRLHQSRHGRVEPS 911 T. aestivum 3 DGKRRREFESELYERFGSLVKMPLLKPERAPLPGDVKTILDEGMSLFRLHQSRHGRVEPS 911 O. sativa DGKSRREFESELYERFGSLVKMPLLKPDRAPLPDEVKAILDEGISLFRLHQSRHGRAEPS 950 A. thaliana EGKNRNEFESELIERFGSLIKMPLLKSDRTPLPDPVKSVLEEGIDLFNLHSRRHGRLEST 889 S. lycopersicum DGKSRQEFESELIERFGSLVRIPLLKPERSPLPDSVRSIVEEGINLYRLHTNKHGRLEST 909 :** *.****** ******:::****.:*:***. *:::::**:.*:.** :*** *.:
107

Resultados
CPD
CPD
T. aestivum 1 KGSYAQEWAQWEKRLRVVLSRNANYLTSIQVPFDVAVKEVLEQLKAVAKGDVKTPDTAKR 963 T. aestivum 2 KGSYAQEWAQWEKRLRVVLSRNANYLTSIQVPFDVAVKEVLEQLKAVAKGDVKTPDTAKR 971 T. aestivum 3 KGSYAQEWAQWEKRLRVVLSRNANYLTSIQVPFDVAVKEVLEQLKAVAKGDVKTPDTAKR 971 O. sativa KGAYAKEWAQWEKRLRQVLFANKDYLNSIQVPFDFAVKEVLEQLKSVAKGDLKTPDTAKR 1010 A. thaliana KGTYAAEWTKWEKQLRDTLVANSEYLSSIQVPFESMVHQVREELKTIAKGDYKPPSSEKR 949 S. lycopersicum KGTFVKEWVKWEKQLRDILHGNADYLNSIQVPFEFAVKKVLEQLKAIARGEYAAPSSEKR 969 **::. **.:***:** * * :**.******: *::* *:**::*:*: .*.: ** T. aestivum 1 RFGNIVFAAVTVPQADILSLLRKLGENDGDVNNFLN--GIKVEDNLSKAHVTLAHKRAHG 1021 T. aestivum 2 RFGNIIFAAVTVPQADILSLLRKLGENDGDVNNFLN--GIKVEDNLSKAHVTLAHKRAHG 1029 T. aestivum 3 RFGNIVFAAVTVPQADILSLLRKLGQNDGDVNNFLN--GIKVEDNLSKAHVTLAHKRAHG 1029 O. sativa KFGNIVFAAVTLPPADILGALPKLAEDT-DANKFLN--NTKLADNLTKAHVTLAHKRAHG 1067 A. thaliana KHGSIVFAAINLPATQVHSLLEKLAAANPTMRSFLEGKKKSIQEKLERSHVTLAHKRSHG 1009 S. lycopersicum KLGSIVFAAISLPVPEILGLLNDLAKKDLKVGDFLK--DKSLESCIQKAHLTLAHKRSHG 1027 : *.*:***:.:* .:: . * .*. .**: .: . : ::*:******:** T. aestivum 1 VAAVASYGVYQNQEVPVSFNAFLYTDKMAALEAQLGTVNGEKIDSKNDWPHVTLWTAPGV 1081 T. aestivum 2 VAAVASYGVYQNQEVPVSFNAFLYTDKMAALEAQLGTVNGEKIDSKNDWPHVTLWTAPGV 1089 T. aestivum 3 VAAVASYGVYQNQEVPVSFNAFLYTDKMAALEAQLGTVNGEKIDSKNDWPHVTLWTAPGV 1089 O. sativa VAAVSSYGVYQNHQVPVIFNAFLFSDKMAALEVELGTVNGEKIASRNDWPHATLWTAPGV 1127 A. thaliana VATVASYSQHLNREVPVELTELIYNDKMAALTAHVGSVDGETVVSKNEWPHVTLWTAEGV 1069 S. lycopersicum VTAVANYGSFLHQNVPVDVAALLFSDKLAALEAEPGSVEGEKVDSKNPWPHVTIWTGAGA 1087 *::*:.*. . :::*** . :::.**:*** .. *:*:**.: *:* ***.*:**. *. T. aestivum 1 APKEANMLPQLFSSGQAKRVLIDPPITITGVLDFY 1116 T. aestivum 2 APKEANMLPQLFSAGQAKRVLIDPPITISGVLDFY 1124 T. aestivum 3 APKEANMLPQLFSSGQAKRVLIDPPITISGVLDFY 1124 O. sativa APKEANTLPQLVTEGKAKRVAIDPPITISGVLDFY 1162 A. thaliana TAKEANTLPQLYLEGKASRLVIDPPVSISGPLEFF 1104 S. lycopersicum TAKDANTLPQLLSQGKAIRIDINPPVTITGTLEFF 1122 :.*:** **** *:* *: *:**::*:* *:*:
Fig. 4.18. Secuencia aminoacídica de las tRNA ligasas conocidas de plantas. Las secuencias de T. aestivum (AB235905, AB235906, AB235907, AB235909 y AB235910), Oriza sativa (AK066389), A. thaliana (AC026875) y S. lycopersicum (pendiente de anotación en base de datos) se alinearon con la ayuda del programa ClustalW. Los motivos necesarios para la ligación (Lig I, IIIa y IV), fosforilación (PNK) e hidrólisis del grupo 2’,3’-fosfodiéster cíclico (CPD) se resaltan con un recuadro en gris, verde y azul, respectivamente.
Para estudiar las relaciones filogenéticas entre las tRNA ligasas conocidas de
plantas, las secuencias aminoacídicas se alinearon nuevamente con el programa
MUSCLE (Edgar, 2004), y el árbol se construyó por el método del vecino más próximo
(neighbor-joining) empleando el algoritmo MEGA (Kumar et al., 2004) (Fig. 4.19).
Este árbol mostró una mayor similitud entre trigo y arroz, un resultado esperable ya que
ambas son gramíneas, mientras que A. thaliana y tomate son crucíferas y solanáceas,
respectivamente.
108

Resultados
T.aestivum1
T.aestivum3
T.aestivum2
O.sativa
S.lycopersicum
A.thaliana
1 0 0
5 8
1 0 0
0.05
T. aestivum 1 T. aestivum 3 T. aestivum 2
O. sativa S. lycopersicum
A. thaliana
Fig. 4.19. Árbol filogenético de las tRNA ligasas de plantas conocidas. El árbol se construyó por el método del vecino más próximo con el programa MEGA, evaluando su fiabilidad con el ensayo bootstrap (10000 pseudoréplicas). Únicamente se muestran los valores superiores al 50%.
4.2.3. Clonación y expresión de los cDNAs que codifican las tRNA ligasas de
tomate y A. thaliana
El cDNA completo de la tRNA ligasa de tomate se obtuvo empleando un
cebador (C2) con una cola poli(dT) en 3’ y RNA poliadenilado extraído de hojas
jóvenes del cultivar Rutgers. La secuencia codificante de este cDNA se amplificó por
PCR con los cebadores C28 y C29, el primero de los cuales hibrida con la región que
contiene el codón ATG inicial y el segundo con la que contiene el codón del último
aminoácido de la proteína. Los cebadores se diseñaron con sitios de restricción NheI
(C28) y XhoI (C29) para introducir la secuencia en fase en el plásmido de expresión
pET23a (+), y adicionalmente C29 se diseñó de forma que el cDNA amplificado no
incluyera el codón de terminación. Los productos de la PCR se separaron por
electroforesis en gel de agarosa, y el cDNA del tamaño esperado se eluyó e insertó en el
plásmido de expresión empleando los sitios de restricción ya mencionados. De esta
forma se obtuvo un plásmido recombinante que al traducirse en E. coli añade a la
proteína una metionina y una alanina extra en la región amino terminal, junto con una
leucina y un ácido glutámico en la región carboxilo terminal. A continuación de dicha
región carboxilo terminal, el plásmido recombinante posee una corta secuencia que al
ser traducida incorpora una cola de seis histidinas. Estas histidinas sirven para la
purificación de las proteínas por afinidad con una resina de níquel (Porath et al., 1975;
Sulkowski, 1985).
El procedimiento empleado con la tRNA ligasa de A. thaliana fue distinto. El
RNA poliadenilado extraído de hojas del ecotipo Columbia-0 se transcribió
109

Resultados
inversamente empleando el cebador específico C31, y el cDNA resultante se amplificó
por PCR con los cebadores C30 y C31, que hibridan con las regiones 5’ y 3’ terminal de
la secuencia codificante. El plásmido pET23a (+) también se amplificó por PCR con los
cebadores divergentes C32 y C33, obteniéndose un plásmido lineal de extremos romos
(sin el sitio de clonación múltiple) que se ligó con la secuencia codificante de la tRNA
ligasa de A. thaliana. A diferencia de la construcción de la enzima de tomate, con este
procedimiento se obtuvo un plásmido recombinante que al traducirse en E. coli no
añade aminoácidos extra en los extremos amino o carboxilo terminales de la proteína
expresada, excepto la cola de seis histidinas en el extremo carboxilo terminal necesaria
para su purificación. Esta estrategia, sin embargo, es más complicada y laboriosa.
El código genético es universal, y por tanto compartido por bacterias, arqueas y
eucariotas (Hinegardner y Engelberg, 1963; Wong, 1976). Al ser además degenerado,
un mismo aminoácido puede estar codificado por varios codones, aunque no todos se
emplean con la misma frecuencia porque la concentración intracelular de algunos de los
tRNAs que reconocen los codones es baja. Este fenómeno, conocido como preferencia
codónica, no ocurre de igual forma en eucariotas, bacterias y arqueas (Grantham et al.,
1980), por lo que la traducción de un gen de plantas en un sistema bacteriano como E.
coli puede verse afectada porque algunos de los tRNAs frecuentes en eucariotas sean
escasos en bacterias (revisado en Kurland y Gallant, 1996). Esta posibilidad aumenta
con el tamaño de la proteína, motivo por el cual la expresión en un sistema bacteriano
de la tRNA ligasa de levadura (de un tamaño mayor de 90 kDa) ocurre con un
rendimiento muy bajo, recomendándose un sistema libre de células derivado de germen
de trigo para obtener proteínas de esta familia y otras de gran tamaño (Englert y Beier,
2004). Para corregir estos problemas durante la expresión de proteínas heterólogas, la
cepa comercial de E. coli Rosetta 2, posee un plásmido que codifica siete tRNAs
escasos en bacterias que son comunes en eucariotas, aumentando su concentración
intracelular y evitando que la traducción se atenúe por el déficit de alguno de ellos
(Brinkmann et al., 1989; revisado en Kane, 1995). Éste es el sistema que se empleó en
el presente trabajo para expresar con resultados satisfactorios las tRNA ligasas de A.
thaliana y tomate.
Las construcciones de tomate y A. thaliana se electroporaron en la cepa DH5α
de E. coli, y las colonias que incorporaron el plásmido con inserto se seleccionaron por
ausencia de color en presencia de X-Gal. Mediante ensayos de restricción adecuados se
comprobó qué construcciones eran correctas, y se transformaron con ellas la cepa de
110

Resultados
expresión Rosetta 2 de E. coli. Partiendo de diez colonias transformadas (cinco por cada
una de las dos construcciones), se inocularon diez cultivos líquidos de 50 ml que fueron
inducidos con isopropil-β-D-tiogalactopiranósido para expresar heterólogamente las
tRNA ligasas. Ambas enzimas se purificaron de los correspondientes extractos por
cromatografía de afinidad en columnas de Ni-NTA-agarosa, que retienen a las proteínas
por las histidinas de su extremo carboxilo terminal. La elución de ambas se efectuó con
un tampón que contenía 500 mM imidazol, y se recogieron cinco fracciones. La
actividad de ligación de las mismas se ensayó empleando como sustrato RNA
monomérico lineal (+) del ASBVd proveniente de autocorte, y la colonia de la que se
purificó la preparación más activa se inoculó a otro cultivo de mayor volumen (500 ml)
para obtener la enzima en cantidades superiores. Este procedimiento se aplicó a las dos
tRNA ligasas, detectándose en ambos casos la mayor actividad de ligación en la
segunda fracción de la columna.
Estos ensayos mostraron que la proteína de tomate expresada heterólogamente
posee actividad RNA ligasa, confirmándose así definitivamente que se trata de una
enzima homóloga a las tRNA ligasas de A. thaliana, arroz y trigo. Sin embargo, la
fracción de monómero circularizado en la reacción mediada por la tRNA ligasa de A.
thaliana fue apreciablemente mayor que el obtenido con la de tomate (Fig. 4.20,
comparar carriles E y 1 a 5). La fracción más activa de la enzima de A. thaliana ligó la
casi totalidad del sustrato en 30 min (Fig. 4.20, A. thaliana carril 2), mientras que la de
tomate dejó aproximadamente un 40% del sustrato por ligar en ese mismo tiempo (Fig.
4.20, tomate carril 2). Además, todas las fracciones de la preparación de tRNA ligasa de
A. thaliana mostraron actividad, mientras que sólo las dos primeras de la preparación de
la tRNA ligasa de tomate fueron activas.
111

Resultados
Tomate A. thaliana
Fig. 4.20. Actividad catalítica de las tRNA ligasas de A. thaliana y tomate expresadas en E. coli Rosetta 2. Análisis por simple PAGE desnaturalizante e hibridación northern de los productos resultantes de la reacción catalizada por los extractos y las fracciones de las tRNA ligasas de A. thaliana y tomate purificadas con columnas de Ni-NTA-agarosa. (C-) control sin extracto; E) extractos no fraccionados; (1 a 5) fracciones resultantes de purificar los extractos; (C+) control con una fracción de germen de trigo con actividad tRNA ligasa. Las posiciones del RNA monomérico circular (c) y lineal (l) del ASBVd se indican a la derecha, y las de los marcadores de RNA (con su tamaño en nt) a la izquierda. Otros detalles como en la Fig. 4.1.
Para determinar si la menor fracción de monómero circularizado en la reacción
mediada por la tRNA ligasa de tomate era consecuencia de una menor concentración de
la enzima en la preparación, las proteínas de los extractos y de las cinco fracciones
eluidas de la columna de Ni-NTA-agarosa se separaron por SDS-PAGE. La tRNA
ligasa completa tiene un tamaño en torno a 125 kDa, pero en un gel de poliacrilamida
desnaturalizante aparece como un doblete, con una banda en el lugar esperado y otra
con una masa molecular alrededor de 110 kDa, producto del procesamiento de la
proteína (Makino et al., 2006).
Mientras que el doblete se detectó mediante tinción con azul Coomassie coloidal
en las cinco fracciones de la preparación de la enzima de A. thaliana (Fig. 4.21, carriles
1 a 5), en las de tomate sólo se observó mediante tinción con plata en la segunda
fracción (datos no mostrados).
C- E 1 2 3 4 5 E 1 2 3 4 5 C+
- c
- l
1200 -
1300 -
1400 -
1600 - 1800 - 1000 -
nt
112

Resultados
Fig. 4.21. Proteínas presentes en los extractos y fracciones de las tRNA ligasas de A. thaliana y tomate expresadas en E. coli Rosetta 2. Separación por SDS-PAGE y tinción con azul Coomassie coloidal de las proteínas purificadas con columnas de Ni-NTA-agarosa y de los extractos iniciales. (M) marcadores de tamaño molecular; (E) extractos no fraccionados; (1 a 5) fracciones resultantes de purificar los extractos; (C) extracto de germen de trigo. La masa molecular de los marcadores (en kDa) se indica a la izquierda.
La secuenciación del plásmido permitió verificar la correcta inserción del cDNA
de la tRNA ligasa de tomate en el vector de expresión, descartándose asimismo errores
durante la purificación repitiendo el experimento varias veces. Sin embargo, la
concentración de proteína total eluida fue siempre menor en tomate que en A. thaliana.
La región amino terminal de una proteína tiene un papel relevante en el tiempo
de vida media de una proteína en la célula, tal y como demuestran diversos trabajos en
vertebrados, levadura y bacterias (Bachmair et al., 1986; Gonda et al., 1989; Tobias et
al., 1991, revisado en Varshavsky, 1996). Determinados residuos aminoacídicos actúan
como señales positivas o negativas de degradación, y la combinación de estos residuos
en la región amino terminal constituye una señal más o menos intensa, en función de los
aminoácidos que la componen y su posición, para la unión de los sistemas de
degradación. Dicha degradación está mediada por ubiquitina en eucariotas y ClpS en
bacterias (revisado en Mogk et al., 2007), y por tanto, el estudio de dicha región permite
predecir la estabilidad de las proteínas en ambos dominios taxonómicos.
100 - 190 - 180 - 170 -
140 -
kDa
150 -
160 -
120 -
160 - 220 -
M E 1 2 3 4 5 E 1 2 3 4 5 Tomate A. thaliana
C
113

Resultados
La alta variabilidad de la secuencia amino terminal de las tRNA ligasas sugiere
que esta región podría estar implicada en la regulación de la concentración de estas
enzimas en la célula. Para estimar in silico la estabilidad de las tRNA ligasas de tomate
y A. thaliana se empleó el algoritmo ProtParam del servidor ExPASy
(http://www.expasy.ch/tools/protparam.html), basado en los datos de Bachmair et al.,
(1986), Gonda et al., (1989) y Tobias et al., (1991). Este algoritmo proporciona un
índice que clasifica a las proteínas como estables si es inferior a 40, e inestables cuando
es superior. Los valores estimados para las tRNA ligasas de tomate y A. thaliana son de
41.3, y de 38.65 respectivamente, lo que sugiere que el tiempo de vida media de ambas
es diferente. Sin embargo, la pequeña variación en los valores de estabilidad calculados
para estas enzimas difícilmente puede explicar por sí sola un rendimiento tan dispar en
su purificación, por lo que es posible que algún otro factor desconocido este influyendo.
En cualquier caso, la cantidad de proteína purificada en ambos casos fue suficiente para
ensayar la actividad de estas enzimas en diferentes condiciones bioquímicas.
4.2.4. Caracterización bioquímica comparada de las tRNA ligasas de tomate y A.
thaliana
4.2.4.1. Efecto del pH
Para averiguar el pH óptimo de reacción de las dos tRNA ligasas, se ensayó la
fracción de la columna de Ni-NTA-agarosa con el máximo de actividad con tampones
ajustados a diferentes valores de pH (entre 6 y 9, siendo 7.5 el pH del tampón de
ligación que se había venido empleando). Más específicamente, los valores ensayados
fueron 6, 7, 7.5, 8 y 9, detectándose la ligación en todos los casos con tampón Tris-HCl
(excepto el caso de pH 6, que fue MES-KOH), suplementado con 5 mM DTT, 4 mM
MgCl2 y 1 mM ATP.
La máxima ligación para las dos enzimas se obtuvo a pH 7.5 (Fig. 4.22, carriles
3), siendo apenas apreciable la disminución de actividad a pH 7 (Fig. 4.22, carriles 2).
Sin embargo, a pH 8 la actividad se vio afectada significativamente (Fig. 4.22, carriles
4). A pHs ligeramente ácidos por debajo de 7 no se detectó actividad (Fig. 4.22, carriles
1), mientras que a pH 9 todavía se observó una actividad residual (Fig. 4.22, carriles 5).
114

Resultados
Tomate A. thaliana
Fig. 4.22. Efecto del pH sobre la actividad de las tRNA ligasas. Análisis por simple PAGE desnaturalizante e hibridación northern de los productos resultantes de la reacción catalizada por la fracción de las tRNA ligasas de A. thaliana y tomate más activa de la columna de Ni-NTA-agarosa. (1 a 5) Ligación a pH 6, 7, 7.5, 8 y 9, respectivamente; (C) control sin extracto. Las posiciones del RNA monomérico circular (c) y lineal (l) del ASBVd se indican a la derecha, y las de los marcadores de tamaño molecular (en nt) a la izquierda. Otros detalles como en la Fig. 4.1.
4.2.4.2. Efecto de la temperatura
Con el fin de determinar la temperatura óptima de las tRNA ligasas de tomate y
A. thaliana se llevó a cabo un experimento de ligación a temperaturas entre 15 y 37ºC
(con incrementos de 5ºC excepto en el último caso que fue de 7ºC). La temperatura
óptima de ligación en incubaciones de 30 min se observó en torno a los 30ºC (Fig. 4.23,
carriles 4), que es el valor que se usó para todos los experimentos de ligación de este
trabajo. La temperatura óptima es similar para ambas enzimas, si bien las diferencias a
25ºC y 30ºC fueron mínimas (Fig. 4.23, comparar carriles 3 y 4 respectivamente).
1 2 3 4 5 1 2 3 4 5 nt C
1800 - 1000 -
1600 -
1400 -
- c 1300 -
- l
1200 -
115
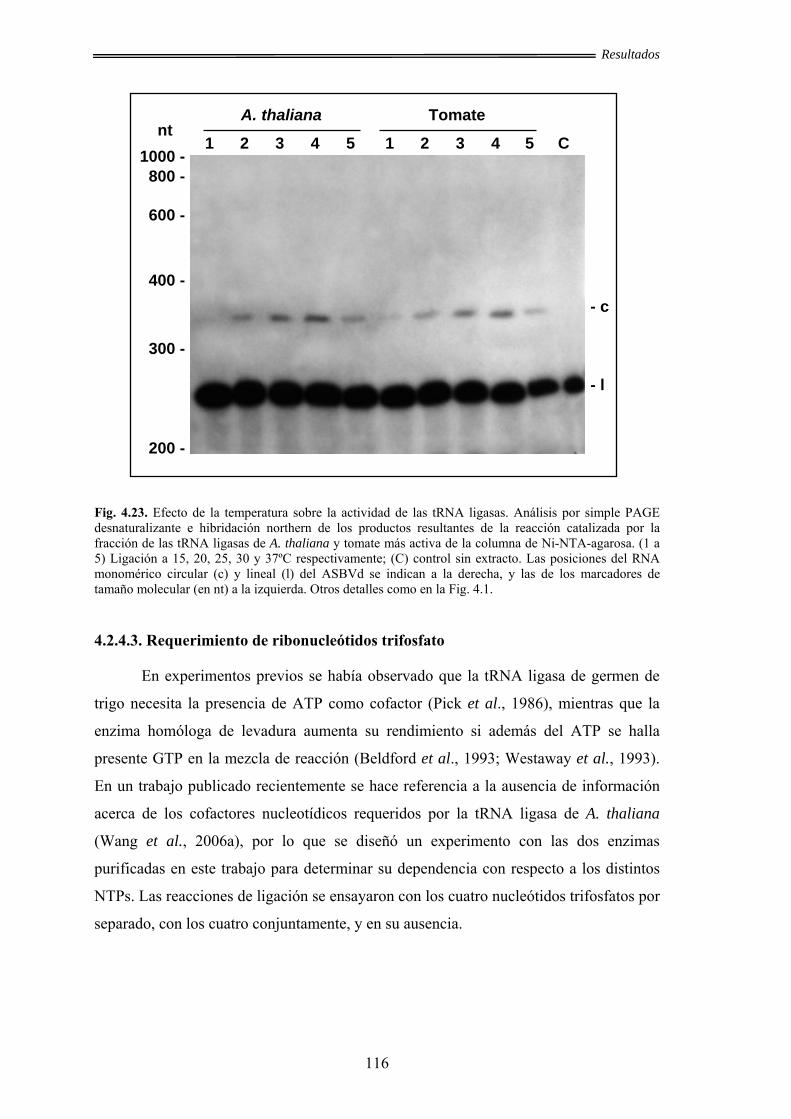
Resultados
Tomate A. thaliana nt
1 2 3 4 5 1 2 3 4 5 C
Fig. 4.23. Efecto de la temperatura sobre la actividad de las tRNA ligasas. Análisis por simple PAGE desnaturalizante e hibridación northern de los productos resultantes de la reacción catalizada por la fracción de las tRNA ligasas de A. thaliana y tomate más activa de la columna de Ni-NTA-agarosa. (1 a 5) Ligación a 15, 20, 25, 30 y 37ºC respectivamente; (C) control sin extracto. Las posiciones del RNA monomérico circular (c) y lineal (l) del ASBVd se indican a la derecha, y las de los marcadores de tamaño molecular (en nt) a la izquierda. Otros detalles como en la Fig. 4.1.
4.2.4.3. Requerimiento de ribonucleótidos trifosfato
En experimentos previos se había observado que la tRNA ligasa de germen de
trigo necesita la presencia de ATP como cofactor (Pick et al., 1986), mientras que la
enzima homóloga de levadura aumenta su rendimiento si además del ATP se halla
presente GTP en la mezcla de reacción (Beldford et al., 1993; Westaway et al., 1993).
En un trabajo publicado recientemente se hace referencia a la ausencia de información
acerca de los cofactores nucleotídicos requeridos por la tRNA ligasa de A. thaliana
(Wang et al., 2006a), por lo que se diseñó un experimento con las dos enzimas
purificadas en este trabajo para determinar su dependencia con respecto a los distintos
NTPs. Las reacciones de ligación se ensayaron con los cuatro nucleótidos trifosfatos por
separado, con los cuatro conjuntamente, y en su ausencia.
1800 - 1000 -
1600 -
1400 - - c
1300 -
- l
1200 -
116

Resultados
Tomate A. thaliana
Fig. 4.24. Efecto de los distintos NTPs en la actividad de las tRNA ligasas. Análisis por simple PAGE desnaturalizante e hibridación northern de los productos resultantes de la reacción catalizada por la fracción de las tRNA ligasas de A. thaliana y tomate más activa de la columna de Ni-NTA-agarosa. (1) ATP; (2) CTP; (3) GTP; (4) UTP, (5) los cuatro NTPs; (6) ningún NTP; (C) control sin extracto. Las posiciones del RNA monomérico circular (c) y lineal (l) del ASBVd se indican a la derecha, y las de los marcadores de tamaño molecular (en nt) a la izquierda. Otros detalles como en la Fig. 4.1.
A diferencia de lo descrito para la enzima de germen de trigo (Pick et al., 1986),
las tRNA ligasas de A. thaliana y tomate son capaces de circularizar el RNA
monomérico lineal del ASBVd en presencia de UTP (Fig. 4.24, carriles 4), aunque con
una eficiencia ligeramente menor que la observada en presencia de ATP (Fig. 4.24,
carriles 1). También se detectó ligación, aunque muy baja, en presencia de GTP (Fig.
4.24, carriles 3), no observándose actividad en presencia de CTP (Fig. 4.24, carriles 2).
En contraste con lo que ocurre con la tRNA ligasa de levadura, donde la adición de GTP
a la reacción con ATP incrementa la eficiencia con respecto a cuando sólo está presente
este último (Belford et al., 1993; Westaway et al., 1993), la actividad de las enzimas de
tomate y A. thaliana en presencia de ATP más cualquiera de los otros NTPs fue
indistinguible de la obtenida sólo con ATP.
1200 -
1300 -
1400 -
1600 - 1800 - 1000 -
nt 1 2 3 4 5 6 1 2 3 4 5 6 C
- c
- l
117


Resultados
4.3. Chlamydomonas reinhardtii: un sistema modelo para el estudio de
factores del hospedador implicados en la replicación de los viroides
cloroplásticos Como ya se ha comentado en la Introducción, los miembros de la familia
Avsunviroidae tienen una gama de hospedadores restringida esencialmente a las
especies vegetales que infectan de forma natural, ninguna de las cuales es un organismo
modelo de investigación en biología molecular. Este aspecto, junto con la dificultad de
transformar cloroplastos de plantas superiores, complica el análisis molecular de los
procesos que ocurren durante la interacción planta-viroide. Para solventar estos
problemas se consideró la posibilidad de emplear un sistema heterólogo basado en el
cloroplasto de C. reinhardti, un microorganismo unicelular modelo en estudios
bioquímicos y genéticos de este orgánulo, que presenta la ventaja adicional de que su
genoma está completamente secuenciado (Harris, 2001).
4.3.1. Transformación de C. reinhardtii y selección de líneas homoplásmicas
Las células empleadas, de la cepa CC-373, tienen una deleción en el gen
cloroplástico que codifica la subunidad β de la ATPasa (atpB), una enzima necesaria
para la fotosíntesis. Esta mutación impide el crecimiento autotrófico, pero no afecta a la
expresión del aparato fotosintético. Las células se transformaron mediante bombardeo
con proyectiles de oro recubiertos con el plásmido recombinante (Boyton et al., 1988).
El plásmido contiene, junto con el cDNA que codifica el RNA viroidal y secuencias
reguladoras adecuadas para su correcta expresión en el cloroplasto, una de las regiones
invertidas del cromosoma cloroplástico de C. reinhardtii y un fragmento del gen atpB
que incluye la región delecionada en la cepa CC-373. Tras el bombardeo, en aquellas
células que han incorporado el plásmido ocurre una recombinación homóloga del
mismo con el genoma cloroplástico, específicamente entre la región invertida y el gen
atpB, reparándose la copia truncada de este último (Fig. 4.25). En la recombinación se
incorpora permanentemente el cDNA viroidal y las secuencias reguladoras que lo
flanquean, y como consecuencia de la reparación del gen atpB, se restaura la capacidad
de crecimiento autotrófico permitiendo la selección de líneas transgénicas cuando se
retira la fuente de carbono del medio.
El cloroplasto posee múltiples copias de su único cromosoma, siendo muy baja
la probabilidad de que ocurra recombinación en más de una de ellas. Durante la
119

Resultados
duplicación celular, las copias del cromosoma que ha incorporado el cDNA viroidal no
se reparten de forma equivalente entre ambas células hijas. En un caso extremo una de
las células podría heredar todas las copias y la otra ninguna, por lo que no todas las
colonias transgénicas presentan el mismo nivel de expresión de RNA heterólogo. Esta
situación se denomina heteroplasmicidad, mientras que cuando la secuencia introducida
se encuentra en todas las copias del cromosoma se habla de homoplasmicidad. Así pues,
la expresión del transgén varía mucho entre clones.
A pCrc+157
Genoma cloroplástico
Fig. 4.25. Esquema de la transformación por recombinación homóloga del cloroplasto de C. reinhardtii. (A) El genoma cloroplástico posee dos regiones repetidas e invertidas (inverted region, IR) (en azul oscuro) y, en el caso de la cepa CC-373, el gen de la subunidad β de la ATPasa (atpB) truncado (en azul claro). (B) El plásmido pCrc+157 contiene el cDNA dimérico viroidal (naranja) flanqueado por la región 5’ no traducible (5’-UTR) y los 157 nt iniciales del gen de la subunidad grande de la ribulosa 1,5-bisfosfato carboxilasa oxigenasa (rbcL) (en verde claro), y por la 3’-UTR de una de las dos subunidades grandes del fotosistema I (psaB) (en verde oscuro), que otorgan estabilidad al RNA en el entorno cloroplástico. Este plásmido también incluye un fragmento de una de las dos IR y la parte del gen atpB delecionada en el genoma cloroplástico. Las regiones comunes entre el cromosoma cloroplástico y el plásmido (IR y atpB) recombinan homólogamente durante la transformación, incorporando el cDNA dimérico viroidal junto con sus secuencias reguladoras y restaurando la capacidad fotosintética de la cepa por reparación del gen atpB. El gráfico no está a escala.
atpB
IR atpB IR
5’-UTR 3’-UTRcDNA viroidal-cDNA viroidal
pCrc+157 + cDNA viroidal
pCrc+157 B
IR
rbcL psaB
120

Resultados
4.3.2. Obtención de plásmidos para transformar C. reinhardtii con cDNAs
viroidales
Para la expresión en el cloroplasto de los RNAs viroidales de polaridad (+), se
construyeron plásmidos que contenían cDNAs diméricos de viroides representativos de
los tres géneros de la familia Avsunviroidae, ASBVd, CChMVd y ELVd, así como de
otro representativo de la familia Pospiviroidae, el viroide de la exocortis de los cítricos
(CEVd). También se construyó un plásmido para expresar el RNA dimérico de
polaridad (-) del ASBVd (Fig. 4.26). La razón de emplear cDNAs diméricos viroidales
es que sus transcritos se asemejan a los intermediarios replicativos y, además son muy
infecciosos al inocularlos mecánicamente en sus hospedadores (Cress et al., 1983;
Navarro y Flores, 1997; Ambrós et al., 1998).
psaB
psaB
psaB
psaB
rbcL
333 nt 333 nt
399 nt 399 nt
247 nt 247 nt
247 nt 247 nt psaBASBVd(+) X52041
153 15455/56 153154 55/56
ASBVd(-) X52041
158 15770/69 158157 70/69
CChMVd(+) AJ878085
294 295399/1 294399/1295
CEVd(+) M34917G74∆370 nt 370 nt
39 40 3940
ELVd(+) AJ536613
209 210333/1 209333/1210
rbcL
rbcL
rbcL
rbcL psaB
psaB
psaB
psaB
rbcL
333 nt 333 nt
399 nt 399 nt
247 nt 247 nt
247 nt 247 nt psaBASBVd(+) X52041
153 154153 15455/56 153153154154 55/5655/56
ASBVd(-) X52041
158 15770/6970/69 158158157157 70/69
CChMVd(+) AJ878085
294 295399/1399/1 294294399/1295295
CEVd(+) M34917G74∆370 nt 370 nt
39 4039 40 39394040
ELVd(+) AJ536613
209 210209 210333/1333/1 209209333/1210210
rbcL
rbcL
rbcL
rbcL
Fig. 4.26. Esquema de los transcritos viroidales expresados en el cloroplasto de líneas de C. reinhardtii transplastómicas. Los RNA viroidales y sus polaridades [(+) y (-)] se indican a la izquierda, y los números de acceso de las variantes de secuencia empleadas a la derecha (el subíndice en la variante del CEVd indica una deleción con respecto a la depositada en la base de datos). Los rectángulos verde claro y oscuro representan la 5’-UTR y los 157 nt iniciales del gen de la subunidad grande de la ribulosa 1,5-bisfosfato carboxilasa oxigenasa (rbcL), y la 3’-UTR de una de las dos subunidades grandes del fotosistema I (psaB), respectivamente (no están a escala). Los rectángulos naranja corresponden a las unidades viroidales monoméricas (a escala). Las flechas muestran los sitios de autocorte de las ribozimas de cabeza de martillo, y las posiciones en las que empieza y acaba el transcrito viroidal y el tamaño de cada unidad monomérica se indican en la parte superior.
El cDNA dimérico viroidal se insertó a través de tres pasos de clonación en el
vector de transformación cloroplástico pCrc+157 (Suay et al., 2005), sustituyendo la
121

Resultados
ORF del gen de la β-glucuronidasa (GUS) (Fig. 4.27). En el plásmido recombinante,
dicho cDNA está precedido por el promotor, la región 5’ no traducible (5’-UTR) y el
comienzo de la ORF del gen rbcL (subunidad grande de la ribulosa 1,5-bisfosfato
carboxilasa oxigenasa, entre -70 y +157), y seguido de la 3’-UTR del gen psaB (una de
las dos subunidades grandes del fotosistema I). Las 5’ y 3’-UTR estabilizan al RNA que
flanquean retardando su degradación, mientras que el promotor asegura una expresión
abundante del mismo.
+157 +1 - 70
Fig. 4.27. Cassette de expresión del plásmido pCrc+157 a partir del que se generaron las construcciones para expresar los RNAs viroidales en el cloroplasto de C. reinhardtii. El cDNA dimérico viroidal sustituye la ORF del gen de la β-glucuronidasa (GUS) (en naranja), y se encuentra bajo el control de una región del cromosoma del cloroplasto (en rojo) que incluye el promotor de la subunidad grande de la ribulosa 1,5-bisfosfato carboxilasa oxigenasa (rbcL) (flecha negra). La 5’-UTR y los 67 nt iniciales de la ORF de rbcL (en verde claro), y la 3’-UTR de una de las dos subunidades grandes del fotosistema I (psaB) (en verde oscuro) otorgan estabilidad al RNA transcrito. NheI y XbaI delimitan los sitios de restricción empleados en el último paso de la construcción del plásmido. El fragmento del gen de la subunidad β de la ATPasa (atpB) permite la restauración de una copia completa del gen por recombinación homóloga con el genoma cloroplástico. Los números superiores indican el sitio de inicio de la transcripción, y los inferiores el tamaño de cada región. El gráfico no está a escala.
4.3.3. Análisis del corte y la ligación de los RNAs viroidales en líneas
transplastómicas de C. reinhardtii
Las células transformadas con las construcciones de los cDNAs viroidales se
cultivaron en medio HS sin acetato. De esta forma sólo pueden crecer autotróficamente
las células recombinantes con el gen atpB restaurado (Fig. 4.25). Un mínimo de diez
colonias recombinantes se inocularon en cultivos líquidos, de los que se extrajo su RNA
para examinar el nivel de expresión del dímero viroidal mediante simple PAGE
desnaturalizante e hibridación northern. Las colonias con una señal más intensa se
inocularon en un nuevo cultivo del que se purificó el RNA y analizó detalladamente
mediante simple PAGE desnaturalizante e hibridación Northern (Fig. 4.28). Los RNAs
viroidales que no se autocortan ni son procesados por la maquinaria enzimática del
cloroplasto de C. reinhardtii deberían generar una banda con el tamaño del transcrito
primario. En los que sí ocurre el procesamiento, debería observarse la aparición del
0.4 kb 0.23 kb
XbaI
1.9 kb
rbcL GUS psaB atpB2.9 kb
NheI
122

Resultados
RNA monomérico lineal producto del corte o autocorte del transcrito dimérico así como
del monomérico circular resultante de su ligación, el segundo retrasado con respecto al
primero por su estructura circular. Además, en estos últimos también debería observarse
en mayor o menor medida el transcrito primario no procesado.
A B
Fig. 4.28. Expresión y procesamiento en el cloroplasto de C. reinhardtii de los RNAs diméricos viroidales. Análisis mediante simple PAGE desnaturalizante e hibridación northern de líneas transplastómicas que expresan RNAs diméricos viroidales. Los RNAs se hibridaron con ribosondas para detectar ASBVd (+) (A), ASBVd (-) (B), CChMVd (+) (C), ELVd (+) (D), y CEVd (E). (1) Controles de aguacate infectado con ASBVd (A y B), crisantemo infectado con CChMVd (C), berenjena infectada con ELVd (D) y ginura infectada con CEVd (E); (2) controles de C. reinhardtii no transformada (A-E); (3) C. reinhardtii transformadas para expresar dímeros del ASBVd (+) (A y B), CChMVd (C), ELVd (D) y CEVd (E); (4) C. reinhardtii transformadas para expresar dímeros del ASBVd (-) (A y B). Las posiciones del RNA monomérico circular (c) y lineal (l) de cada viroide se indican a la derecha, y las de los marcadores de RNA (con su tamaño en nt) a la izquierda. Otros detalles como en la Fig. 4.1.
800 -600 -
400 -
300 -
200 -
nt 1 2 3 4
ASBVd (+) ASBVd (-)
1 2 3 4 nt
800 -600 -
400 -- c - c 300 -
- l - l 200 -
C D ECChMVd
800 -600 -
400 -
300 -
200 -
nt1 2 3
- c
- l
ELVd CEVd
1 2 3 1 2 3 nt nt
- c 800 - 800 -600 - 600 - - c
- l 400 - 400 -- l 300 -
300 -
200 - 200 -
123

Resultados
Las preparaciones de RNA de las líneas transplastómicas expresando los RNAs
diméricos (+) de los tres viroides cloroplásticos (ASBVd, CChMVd y ELVd) también
se examinaron por doble PAGE nativa/desnaturalizante e hibridación northern (Fig.
4.29). En esta técnica, las moléculas circulares comigran con las lineales en el primer
gel (nativo) permitiendo seleccionar los RNAs viroidales con el tamaño de interés, que
son separados en sus formas circulares y lineales en la segunda electroforesis
(desnaturalizante en condiciones de baja fuerza iónica). Así se resuelven mejor los
RNAs monoméricos circulares que en una simple PAGE desnaturalizante pueden
comigrar con otros RNAs lineales de mayor tamaño. A continuación se exponen los
resultados obtenidos para cada viroide.
1 2 3CChMVd
B
c -
l -
1 2 3ELVd
C
c -
l -
ASBVdA
1 2 3
c -
l -
1 2 31 2 3CChMVd
B
c -
l -
1 2 31 2 3ELVd
C
c -
l -
ASBVd1 2 31 2 3
c -
l -
A
Fig. 4.29. Expresión en el cloroplasto de C. reinhardtii de los RNAs diméricos viroidales. Análisis mediante doble PAGE nativa/desnaturalizante e hibridación northern de líneas transplastómicas que expresan RNAs viroidales. Las preparaciones se separaron con dos PAGEs consecutivas, la primera en condiciones no desnaturalizantes, para a continuación aplicar el segmento de gel nativo que contiene las formas lineales y circulares del viroide en una segunda PAGE desnaturalizante. Los RNAs viroidales se analizaron por hibridación northern con ribosondas para detectar ASBVd (+) (A), CChMVd (+) (B) y ELVd (+) (C). (1) Controles de aguacate infectado con ASBVd (A), crisantemo infectado con CChMVd (B) y berenjena infectada con ELVd (C); (2) controles de C. reinhardtii no transformada (A-C); (3) C. reinhardtii transformada para expresar dímeros del ASBVd (+) (A), CChMVd (B) y ELVd (C). Las posiciones de los RNA monoméricos circulares (c) y lineales (l) de cada viroide se indican a la izquierda de cada panel. Otros detalles como en la Fig. 4.1.
124

Resultados
4.3.3.1. ASBVd
En la línea de C. reinhardtii que expresa el RNA dimérico (+) del ASBVd se
observó por simple PAGE desnaturalizante el transcrito primario de alrededor de 800 nt
y, además, dos productos con las movilidades correspondientes al RNA monomérico
lineal (247 nt) y circular (por encima del marcador de 300 nt) (Fig. 4.28A, carril 3). Los
otros RNAs de diferentes tamaños observados resultan probablemente del autocorte
parcial del transcrito primario por una u otra de las ribozimas. También se detectaron
los RNAs monoméricos (+) lineales y circulares mediante doble PAGE
nativa/desnaturalizante (Fig. 4.29, carril 3). Así pues, el RNA dimérico (+) del ASBVd
se autocorta en C. reinhardtii de la forma predicha por las ribozimas de cabeza de
martillo, y una RNA ligasa cloroplástica o la propia ribozima es capaz de circularizar la
forma lineal. Por otra parte, analizando por simple PAGE desnaturalizante la línea de C.
reinhardtii transformada con la construcción correspondiente al ASBVd dimérico de
polaridad (-) se observaron resultados similares (Fig. 4.28B). Sin embargo, debido a una
expresión menos eficiente, se detectaron cantidades menores de los RNAs monoméricos
circular y lineal de esta polaridad, el último únicamente tras una exposición prolongada.
Es de destacar que el RNA monomérico circular de ambas polaridades del ASBVd se
acumula en mayor medida que el lineal en C. reinhardtii, al igual que en hojas de
aguacate infectado por este viroide (comparar los carriles 1 y 3 en Fig. 4.28A, carriles 1
y 4 en Fig. 4.28B, y carriles 1 y 3 en Fig. 4.29A). Este paralelismo refuerza la validez
de C. reinhardtii como sistema experimental.
4.3.3.2. CChMVd
El cDNA dimérico del CChMVd también se expresó en el cloroplasto de C.
reinhardtii, pero a diferencia del ASBVd, se procesó en sus formas lineales y circulares
con muy poca eficiencia. El RNA monomérico lineal del CChMVd (399 nt) se detectó
tanto por simple como por doble PAGE tras una exposición prolongada (Fig. 4.28C y
Fig. 4.29B, carril 3), mientras que la banda intensa menor de 800 nt que parece
corresponder con el RNA monomérico circular en la simple PAGE desnaturalizante,
mediante doble PAGE nativa/desnaturalizante se comprobó que realmente se trata de un
intermediario del autocorte del transcrito primario (Fig. 4.28C, y 4.29B, comparar
carriles 3). Estas bandas intensas de menor tamaño que el transcrito primario observadas
por simple PAGE desnaturalizante (Fig. 4.28, carril 3) muestran que el autocorte
125

Resultados
mediado por una u otra ribozima funciona eficientemente en el cloroplasto de C.
reinhardtii, pero que raramente ocurre de forma simultánea en ambos sitios. Por otra
parte, la señal correspondiente al RNA monomérico circular (visible tras una exposición
prolongada) es menor que la del RNA monomérico lineal (Fig. 4.29B, carril 3),
presentando una relación similar a la observada en plantas de crisantemo infectadas con
CChMVd.
Estudios previos han mostrado que el autocorte cotranscripcional de un RNA
dimérico del ASBVd (en ambas polaridades) ocurre preferentemente a partir de una
estructura ribozimática formada por nucleótidos de ambas unidades monoméricas
(estructura de doble cabeza de martillo) (Forster et al., 1988; Davies et al., 1991),
mientras que en el CChMVd la ribozima activa se pliega a partir de la secuencia
monomérica (estructura de simple cabeza de martillo). En consecuencia, es esperable
que esta última se forme con mayor facilidad que la del ASBVd y se autocorte mejor
(Navarro y Flores, 1997), en contraste con lo observado en C. reinhardtii.
Para examinar mejor esta cuestión, la construcción dimérica del CChMVd se
expresó in vitro bajo el control del promotor de la RNA polimerasa del fago T7. Con
este fin dicha construcción se amplificó por PCR a partir del plásmido de
transformación de C. reinhardtii con los cebadores C34 y C35, que hibridan con los
extremos de las regiones 5’ y 3’-UTR de rbcL y psaB, respectivamente, y se clonó en el
plásmido pTZ57R/T. Además se generaron dos construcciones similares, la primera
eliminando la mayor parte del sitio clonación múltiple del plásmido para que el RNA
obtenido fuera lo más parecido al transcrito generado en el cloroplasto, y la segunda con
el sitio de clonación múltiple del plásmido intacto para averiguar si una secuencia extra
en 3’ interfiere con el plegamiento catalíticamente activo de la ribozima. Mediante
separación de los productos de la transcripción in vitro por simple PAGE
desnaturalizante se observó que el autocorte de ambas ribozimas ocurre
simultáneamente con un rendimiento elevado (Fig. 4.30, carriles 1 y 2), aunque también
se detectaron en una proporción similar los intermediarios resultantes de la acción de
una u otra de las ribozimas. La presencia en el transcrito del fragmento con el sitio de
clonación múltiple del plásmido pTZ57R/T se reflejó en un aumento de 25 nt en el
producto del autocorte de menor tamaño, pero no se observaron diferencias apreciables
en la eficiencia del autocorte en las dos construcciones analizadas. Por tanto, la
presencia de una pequeña secuencia extra en 3’ no parece afectar al funcionamiento de
la ribozima. Como el plegamiento y autocorte de los RNAs viroidales ocurre
126
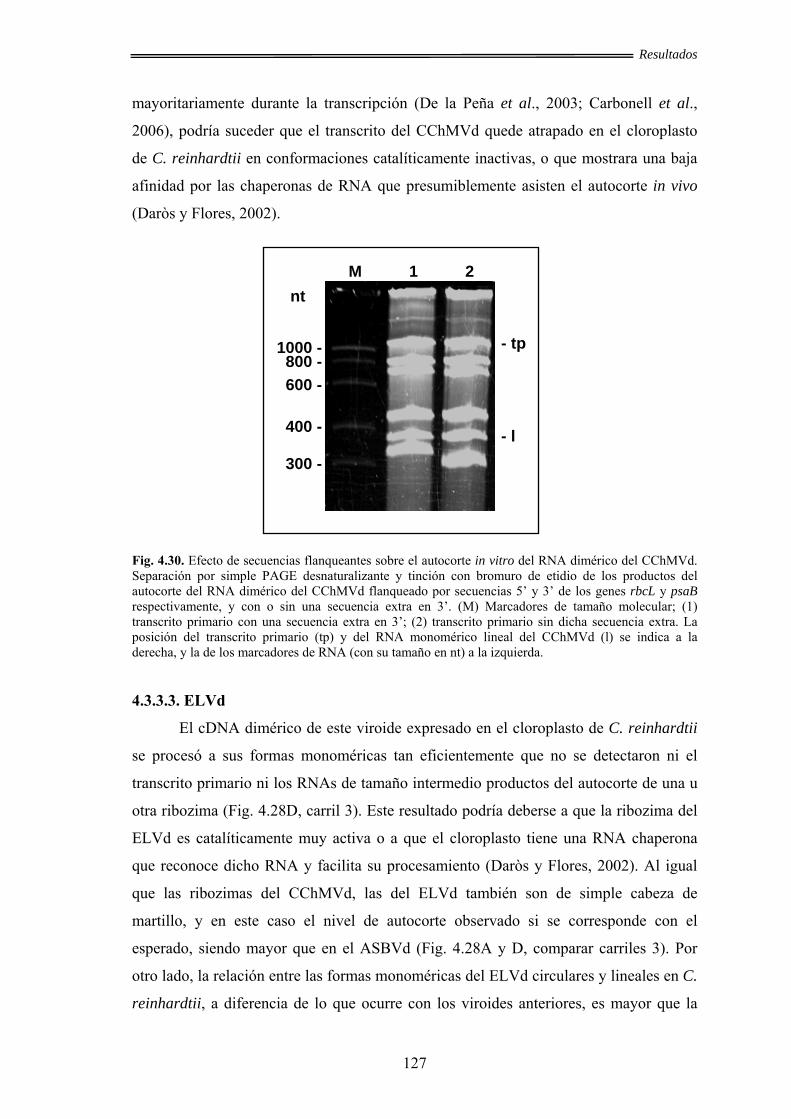
Resultados
mayoritariamente durante la transcripción (De la Peña et al., 2003; Carbonell et al.,
2006), podría suceder que el transcrito del CChMVd quede atrapado en el cloroplasto
de C. reinhardtii en conformaciones catalíticamente inactivas, o que mostrara una baja
afinidad por las chaperonas de RNA que presumiblemente asisten el autocorte in vivo
(Daròs y Flores, 2002).
Fig. 4.30. Efecto de secuencias flanqueantes sobre el autocorte in vitro del RNA dimérico del CChMVd. Separación por simple PAGE desnaturalizante y tinción con bromuro de etidio de los productos del autocorte del RNA dimérico del CChMVd flanqueado por secuencias 5’ y 3’ de los genes rbcL y psaB respectivamente, y con o sin una secuencia extra en 3’. (M) Marcadores de tamaño molecular; (1) transcrito primario con una secuencia extra en 3’; (2) transcrito primario sin dicha secuencia extra. La posición del transcrito primario (tp) y del RNA monomérico lineal del CChMVd (l) se indica a la derecha, y la de los marcadores de RNA (con su tamaño en nt) a la izquierda.
4.3.3.3. ELVd
El cDNA dimérico de este viroide expresado en el cloroplasto de C. reinhardtii
se procesó a sus formas monoméricas tan eficientemente que no se detectaron ni el
transcrito primario ni los RNAs de tamaño intermedio productos del autocorte de una u
otra ribozima (Fig. 4.28D, carril 3). Este resultado podría deberse a que la ribozima del
ELVd es catalíticamente muy activa o a que el cloroplasto tiene una RNA chaperona
que reconoce dicho RNA y facilita su procesamiento (Daròs y Flores, 2002). Al igual
que las ribozimas del CChMVd, las del ELVd también son de simple cabeza de
martillo, y en este caso el nivel de autocorte observado si se corresponde con el
esperado, siendo mayor que en el ASBVd (Fig. 4.28A y D, comparar carriles 3). Por
otro lado, la relación entre las formas monoméricas del ELVd circulares y lineales en C.
reinhardtii, a diferencia de lo que ocurre con los viroides anteriores, es mayor que la
1800 - 1600 -
1400 -
1300 -
M 1 2 nt
- tp 1000 -
- l
127

Resultados
observada en plantas de berenjena infectadas (Fig. 4.28D y 4.29C, comparar carriles 1 y
3), sugiriendo que no sólo el autocorte es muy eficiente, sino también la circularización.
4.3.3.4. CEVd
El análisis de las células de C. reinhardtii transformadas con el cDNA dimérico
del CEVd permitió detectar el transcrito primario expresado en el cloroplasto (Fig.
4.28E, carril 3), pero no ocurrió lo mismo con las formas lineales ni circulares. Este
resultado indica que dicho RNA dimérico viroidal no se procesa en el cloroplasto, algo
esperable pues carece de ribozimas de cabeza de martillo y en su contexto natural se
localiza en el núcleo. Experimentos realizados en líneas transgénicas de A. thaliana que
expresan construcciones diméricas de este mismo viroide en el núcleo muestran que el
transcrito sí es cortado y circularizado, y que sirve de molde para la transcripción RNA-
RNA que completa el ciclo replicativo (Gas et al., 2007).
4.3.4. Análisis de la replicación de los viroides por círculo rodante en las líneas
transformadas de C. reinhardtii
En las preparaciones de RNA de ambas polaridades del ASBVd procedentes de
las líneas de C. reinhardtii transplastómicas se intentó detectar la cadena de RNA
viroidal complementaria a la expresada para examinar la posible existencia de
transcripción RNA-RNA en el cloroplasto de dichas líneas. El análisis por hibridación
northern no detectó señal correspondiente a los RNA monoméricos circulares, lineales o
al transcrito primario cuando la preparación de RNAs viroidales de polaridad (+) se
hibridó con una sonda específica de la polaridad (-) (Fig. 4.28B, carril 3). El mismo
resultado negativo se obtuvo cuando la preparación de RNAs de polaridad (-) se hibridó
con una sonda específica de la polaridad (+) (Fig. 4.28A, carril 4). Las bandas más
tenues (visibles tras una exposición prolongada) que aparecen en la parte superior del
carril (Fig. 4.28A, carril 4) son probablemente debidas a hibridación inespecífica. Este
resultado indica que los cloroplastos de C. reinhardtii son incapaces de iniciar y/o
elongar el RNA viroidal a partir de un molde de RNA, y por lo tanto no puede
completarse la replicación.
128

55.. DDIISSCCUUSSIIÓÓNN


Discusión
Los viroides se replican en sus hospedadores a través de un mecanismo de
círculo rodante (Branch y Robertson, 1984; Ishikawa et al., 1984; Hutchins et al., 1985)
donde todos los intermediarios son de RNA (Grill y Semancik 1978). Este mecanismo
de replicación comprende tres etapas: transcripción, corte y ligación. En la variante
simétrica, que siguen los miembros de la familia Avsunviroidae (Daròs et al., 1994;
Bussière et al., 1999), los RNAs circulares de ambas polaridades sirven como moldes
para la síntesis de RNAs oligoméricos de polaridad complementaria que posteriormente
son procesados a sus formas monoméricas circulares. En la variante asimétrica,
característica de los miembros de la familia Pospiviroidae, únicamente opera un círculo
rodante en la síntesis de los RNAs multiméricos de polaridad (-), que sirven como
molde para la transcripción de los RNAs multiméricos de polaridad (+), los cuales son
procesados a las formas monoméricas circulares finales (Branch et al., 1988).
Desde el descubrimiento de los viroides, se han caracterizado alguno de los
factores que median su replicación. En relación con la primera etapa de la variante
asimétrica, se sabe que la RNA polimerasa II es la enzima implicada en la transcripción
de las cadenas de ambas polaridades (Mühlbach y Sänger, 1979; Flores y Semancik,
1982; Schindler y Mühlbach, 1992), y mediante diferentes experimentos in vitro
realizados con el viroide del tubérculo fusiforme de la patata (Potato spindle tuber
viroid, PSTVd) se ha determinado que dicha transcripción comienza en regiones
caracterizadas por formar un bucle (Tabler y Tsagris, 1990; Kolonko et al., 2006). En la
variante simétrica, los datos disponibles indican que una RNA polimerasa cloroplástica
dependiente de DNA codificada en el núcleo (NEP) transcribe los RNAs del viroide del
manchado solar del aguacate (Avocado sunblotch viroid, ASBVd) y del viroide del
mosaico latente del melocotonero (Peach latent mosaic viroid, PLMVd) comenzando la
transcripción en regiones similares en cada polaridad (Navarro et al., 2000; Navarro y
Flores, 2000; Delgado et al., 2005; Rodio et al., 2007). El corte de los multímeros en la
variante simétrica es mediado en ambas polaridades por una ribozima de cabeza de
martillo (Hutchins et al., 1986; Daròs et al., 1994), mientras que en la variante
asimétrica se ha propuesto que una endonucleasa cataliza esta etapa (Tsagris et al.,
1987). Sin embargo, se desconoce cómo ocurre en ambas variantes la última etapa de la
replicación, la ligación que conduce a los RNAs monoméricos circulares.
Al inicio de este trabajo no existían pruebas directas de que las ribozimas de
cabeza de martillo catalizaran la ligación, aunque otras ribozimas como la de tipo
horquilla son capaces de actuar como RNasas y RNA ligasas (Hampel y Tritz, 1989).
131

Discusión
Únicamente se había detectado ligación in vitro en el caso de ribozimas de cabeza de
martillo mínimas (que sólo contienen los nucleótidos que forman el centro catalítico),
aunque con un equilibrio desplazado unas 100 veces hacia el corte (Hertel et al., 1994).
Recientemente se ha observado ligación con dos ribozimas de cabeza de martillo
extendidas (que incluyen las interacciones terciaria entre bucles periféricos al centro
activo que son clave para la actividad catalítica): la del RNA satélite del virus de las
manchas anulares del tabaco (satellite RNA of tobacco ringspot virus, sTRSV) y la del
gusano platelminto parásito Schistosoma mansoni, donde se alcanzan valores de
ligación entre el 6 y el 23% (Nelson et al., 2005; Canny et al., 2007).
Por otra parte, en experimentos con RNAs monoméricos lineales del PLMVd
procedentes de autocorte, se ha observado autoligación in vitro en ausencia de proteínas
cuando el medio contiene 100 mM de MgCl2 (Côté y Perreault, 1997), generándose un
enlace 2’,5’-fosfodiéster. Se ha propuesto que dicho enlace también existe en moléculas
circulares de este mismo viroide aisladas de plantas infectadas (Côté et al., 2001),
aunque las pruebas a este respecto no son concluyentes. Es muy probable que esta
autoligación del PLMVd y la ligación descrita en las ribozimas de cabeza de martillo
mínimas in vitro no funcionen de la misma manera porque los RNAs viroidales con
ribozimas de cabeza de martillo, una vez autocortados, se pliegan adquiriendo una
conformación de mínima energía libre distinta a la catalíticamente activa (Forster y
Symons, 1987), mientras que las ribozimas mínimas y extendidas pueden adoptar
diversas conformaciones, una de las cuales es la catalíticamente activa (revisado en
Nelson y Uhlenbeck, 2006). Además, no se han descrito RNAs naturales con enlaces
2’,5’-fosfodiéster que sirvan como moldes de trascripción, por lo que no puede
desestimarse la implicación de una enzima en la circularización de los RNAs viroidales
o una ligación autocatalítica mediada por las ribozimas (asistidas por una RNA
chaperona) que generara un enlace 3’,5’-fosfodiéster.
Durante el trabajo descrito en esta memoria se ha detectado una RNA ligasa
cloroplástica que en principio podría mediar la circularización de los miembros de la
familia Avsunviroidae. Sin embargo, esta RNA ligasa presenta unas propiedades
atípicas en cuanto a sus requerimientos de NTPs y Mg+2, y los resultados
experimentales sugieren que el enlace generado sería de tipo 2’,5’-fosfodiéster, como el
descrito en la autoligación del PLMVd (Côté y Perreault, 1997). Asímismo, durante este
trabajo se ha observado mediante experimentos de extensión de cebador que en los
RNAs monoméricos circulares (+) del ASBVd aislados de plantas infectadas no se
132

Discusión
detectan enlaces atípicos (que como los 2’,5’-fosfodiéster, son un obstáculo para la
elongación catalizada por la transcriptasa inversa). Por lo tanto, estos datos suscitan
dudas acerca de si dicha RNA ligasa cloroplástica es la implicada en la circularización
de los RNAs de los miembros de la familia Avsunviroidae.
En el caso del PSTVd se ha observado que la nucleasa T1 fúngica es capaz de
catalizar el corte de RNAs de polaridad (+) de longitud mayor que la unitaria, así como
la circularización de los RNAs monoméricos resultantes (Tabler et al., 1992). Otros
trabajos muestran que una actividad del tipo tRNA ligasa presente en extractos de
Chlamydomonas reinhardtii y de germen de trigo cataliza eficientemente la
circularización de RNAs monoméricos lineales aislados de plantas infectadas por este
mismo viroide (Kikuchi et al., 1982; Branch et al., 1982). Estos últimos resultados con
extractos de plantas sustentan la hipótesis de que la circularización está mediada por una
RNA ligasa más que por una RNasa actuando en sentido inverso, razón por la que se
identificó y expresó durante el presente trabajo la tRNA ligasa de tomate, un
hospedador natural y experimental de diversos miembros de la familia Pospiviroidae
(Diener, 1979; Galindo et al., 1982; Kiefer et al., 1983). Esta enzima es la homóloga de
las tRNA ligasas previamente descritas en levadura (Greer et al., 1983b; Westaway et
al., 1988), germen de trigo (Konarska et al., 1981; Makino et al., 2006) Arabidopsis
thaliana y arroz (Englert y Beier, 2005). Los extremos ligados son del tipo 5’-hidroxilo,
2’,3’-fosfodiéster cíclico, que son los generados habitualmente por las endonucleasas
celulares incluyendo a las implicadas en la excisión de los intrones durante la
maduración de los pre-tRNAs previa a la ligación por las tRNA ligasas (Peebles et al.,
1983; Trotta et al., 1997). Sin embargo, otros resultados posteriores muestran que los
RNAs monoméricos lineales procedentes del procesamiento in vivo de miembros
representativos de la familia Pospiviroidae presentan predominantemente extremos 5’-
fosfomonoéster, 3’-hidroxilo, por lo que es posible la existencia de una segunda RNA
ligasa nuclear que catalizaría de este paso (Gas et al., 2007; Gas et al., 2008).
A pesar del progreso en la elucidación de los mecanismos implicados en la
replicación de los viroides cloroplásticos, todavía se desconocen muchos aspectos,
particularmente en lo que respecta a la interacción in vivo del RNA viroidal con los
factores del huésped que median en su replicación. Por esta razón se decidió explorar el
potencial de transformar el cloroplasto de C. reinhardtii para expresar RNAs diméricos
viroidales y estudiar su procesamiento. Dicha alga verde unicelular se eligió para el
presente trabajo por ser un organismo modelo cuyo genoma está secuenciado y para
133

Discusión
cuyo estudio se dispone de una batería de herramientas que incluyen la transformación
de sus tres genomas: nuclear, cloroplástico y mitocondrial (Harris, 2001).
5.1. Una RNA ligasa cloroplástica capaz de catalizar la circularización
de los RNAs de ciertos miembros de la familia Avsunviroidae
No se han descrito RNA ligasas en el cloroplasto, por lo que con el fin de
examinar si este orgánulo contiene alguna enzima capaz de catalizar la ligación de los
RNAs monoméricos lineales de la familia Avsunviroidae resultantes del autocorte
mediado por ribozimas de cabeza de martillo, se intentó purificar cloroplastos de hojas
jóvenes de aguacate, el hospedador natural del miembro tipo de esta familia, el ASBVd.
Sin embargo, el rendimiento de la purificación de cloroplastos de esta especie fue muy
bajo, por lo que se eligió espinaca como material de partida ya que es una planta modelo
para estudios bioquímicos centrados en este orgánulo. De esta forma pudo detectarse
una actividad RNA ligasa cloroplástica que presenta unas propiedades inusuales entre
las enzimas de esta clase conocidas en eucariotas. La reacción catalizada por dicha RNA
ligasa ocurre en ausencia de NTPs y Mg+2, el equilibrio de la reacción está desplazado
hacia las formas lineales y, además, el enlace generado no es del tipo 3’,5’-fosfodiéster
como en principio sería esperable.
Los resultados de este trabajo muestran que la enzima cloroplástica es capaz de
catalizar la formación de un enlace fosfodiéster sin aporte externo de energía (en forma
de NTPs), tal y como se ha descrito en algunas RNA ligasas de bacterias y arqueas
(Greer et al., 1983a; Gomes y Gupta, 1997). El resto de las RNA ligasas de eucariotas
(Konarska et al., 1981; Greer et al., 1983b; Filipowicz y Shatkin, 1983; White y Borst,
1987), de virus (Silber et al., 1972; Ho y Shuman, 2002) y una de las descritas en
arqueas (Kjems y Garrett, 1988), requieren la presencia de un NTP (generalmente ATP)
como fuente de energía para formar el nuevo enlace.
Comparando las cinéticas, la RNA ligasa cloroplástica alcanza el equilibrio de la
reacción a tiempos mucho más cortos que las tRNA ligasas nucleares, y además este
equilibrio está desplazado hacia las formas lineales del RNA viroidal. Una situación
similar ocurre con la 2’,5’-RNA ligasa de Escherichia coli, que tampoco requiere aporte
externo de energía en forma de NTPs, por lo que se consideró que esta enzima podría
funcionar in vivo como una RNasa en vez de como RNA ligasa. Sin embargo, este
último papel funcional no es factible ya que en E. coli no existen sustratos con dicho
134

Discusión
enlace atípico 2’,5’-fosfodiéster sobre los que pudiera actuar como RNasa (Arn y
Abelson, 1996). Con la RNA ligasa cloroplástica, una vez se ha alcanzado el equilibrio
(30 min en las condiciones ensayadas) la adición de sustrato fresco duplica la cantidad
de producto obtenido, indicando que la enzima no se inactiva tempranamente ya que se
obtiene la misma cantidad de producto que si se añadiese todo el sustrato desde el inicio
de la reacción. Otros resultados muestran que el sustrato ligable presenta extremos 5’-
hidroxilo y 2’,3’-fosfodiéster cíclico, los mismos requeridos por las tRNA ligasas
eucariotas (Konarska et al., 1982; Greer et al., 1983b). Por lo tanto, una
transesterificación aparece como el mecanismo más probable por el que ocurriría la
reacción a partir de estos extremos y sin aporte externo de energía. En esta reacción se
rompería el enlace 2’,3’-fosfodiéster cíclico y se formaría un nuevo enlace, 3’,5’ o
2’,5’-fosfodiéster, con un balance neto de energía esencialmente nulo. Mediante
experimentos de extensión de cebador hemos determinado que la reacción catalizada
por la RNA ligasa cloroplástica no genera un enlace 3’,5’-fosfodiéster como ocurre con
las tRNA ligasas de levadura y plantas (Greer et al., 1983b; Konarska et al., 1981) y la
RNA ligasa de animales (Filipowicz y Shatkin, 1983), lo que sugiere que la RNA ligasa
cloroplástica cataliza la circularización de los RNA viroidales generando un enlace
2’,5’-fosfodiéster, como sucede con las 2’,5’-RNA ligasas de bacterias y arqueas (Greer
et al., 1983a; Arn y Abelson, 1996). Puesto que todos los experimentos con 2’,5’-RNA
ligasas han sido realizados in vitro, no puede descartarse la posibilidad de que una
chaperona de RNA facilite la reacción in vivo, permitiendo al sustrato adquirir la
conformación necesaria para ligarse a través de un enlace 3’,5’-fosfodiéster típico. Por
otro lado, resultados no mostrados obtenidos en el presente trabajo indican que tanto la
2’,5’-RNA ligasa de E. coli como la RNA ligasa cloroplástica catalizan la ligación en
ausencia de Mg+2. Esta propiedad también relaciona ambas enzimas, pues el Mg+2 se
considera esencial en las reacciones mediadas por todas las RNA ligasas víricas y de
eucariotas, que pierden su actividad en ausencia de este catión (Silber et al., 1972; Greer
et al., 1983b; Perkins et al., 1985; Pick et al., 1986; White y Borst, 1987; Ho y Shuman,
2002).
Puesto que existen pruebas muy firmes de que el cloroplasto desciende de un
endosimibionte bacteriano (Bhattacharya y Medlin, 1998; Martin y Herrmann, 1998;
Brinkman et al., 2002; revisado en Gray, 1993), es posible que la RNA ligasa
cloroplástica sea homóloga de las 2’,5’-RNA ligasas caracterizadas en bacterias y
arqueas, aunque se desconoce cual sería su función. Los intrones bacterianos son de tipo
135

Discusión
I y II (autocatalíticos), excindiéndose y ligándose con un mecanismo ribozimático, por
lo que la intervención de la 2’,5’-RNA ligasa sería innecesaria. Sin embargo, la excisión
del intrón de los pre-tRNAs en arqueas es mediada por una endonucleasa homóloga a la
de eucariotas (Li et al., 1998; revisado en Abelson et al., 1998). De las cuatro RNA
ligasas caracterizadas o identificadas en arqueas (Kjems y Garrett, 1988; Arn y
Abelson, 1996; Gomes y Gupta, 1997; Ho y Shuman, 2002), la ligación de las dos
mitades de los pre-tRNAs en Haloferax volcanii está mediada por una RNA ligasa que
genera un enlace 3’,5’-fosfodiéster pero sin requerimientos de NTPs ni Mg+2 (Gomes y
Gupta, 1997; Zofallova et al., 2000; Salgia et al., 2003). Sin embargo, es posible que en
Termus thermophilus sea la 2’,5’-RNA ligasa la que catalice la unión de las dos mitades
de los pre-tRNAs (Kato et al., 2003).
La conservación de las 2’,5’-RNA ligasas en bacterias y arqueas, y posiblemente
en los cloroplastos de plantas superiores, indica que realizan una función importante
para la célula pudiendo utilizar esta actividad los viroides cloroplásticos para completar
su ciclo biológico. Por otra parte, no puede descartarse una ligación del RNA viroidal
de tipo ribozimático, pues esta reacción se ha descrito in vitro en ribozimas mínimas
(Hertel et al., 1994; Nelson et al., 2005; Canny et al., 2007). Sin embargo, este
mecanismo presenta problemas, pues la conformación de mínima energía libre
adquirida por los RNAs viroidales una vez cortados dificultaría la formación
nuevamente del centro catalítico de la ribozima (Forster y Symons, 1987), a diferencia
de lo que ocurre con las ribozimas mínimas (revisado en Nelson y Uhlenbeck, 2006). El
RNA viroidal podría ligarse ribozimáticamente mediante un reordenamiento estructural
hacia formas catalíticamente activas, pudiendo ocurrir este reordenamiento durante la
trascripción (Carbonell et al., 2006) y estar facilitado por una RNA chaperona
cloroplástica.
Alternativamente, en el PLMVd se ha descrito una autoligación in vitro a altas
concentraciones de Mg+2 y en ausencia de proteínas, en la que el enlace que cierra el
RNA viroidal es 2’,5’-fosfodiéster (Côté y Perreault, 1997). Este enlace se ha detectado
en RNAs viroidales aislados de plantas infectadas por el PLMVd (Côté et al., 2001). Sin
embargo, la autoligación y la ligación ribozimática descrita más arriba son
presumiblemente dos fenómenos distintos pues, como ya se ha comentado, la
conformación adoptada por el RNA viroidal plegado no es catalíticamente activa y es
improbable que la ribozima pueda formarse en estas condiciones sin la intervención de
un factor auxiliar. Además, el enlace 2’,5’-fosfodiéster detectado en RNAs circulares
136

Discusión
del PLMVd aislados de plantas infectadas (Côté y Perreault, 2001) pudiera ser
artefactual, ya que los resultados de extensión de cebador empleando las formas
circulares del ASBVd aisladas de tejido infectado que se detallan en el presente trabajo
sugieren que dicho enlace es 3’,5’-fosfodiéster. Por tanto, existen dudas de si la
autoligación descrita en el PLMVd ocurre realmente in vivo.
Los experimentos realizados en el presente trabajo con la RNA ligasa
cloroplástica muestran que en las condiciones ensayadas el RNA del PLMVd no es
circularizable por la enzima cloroplástica, mientras que el del ASBVd sí lo es. Cuando
se ensaya la autoligación (en tampón con 100 mM de MgCl2) ocurre lo contrario, pues
mientras que el RNA del PLMVd se circulariza de forma apreciable, el del ASBVd no.
Sólo el RNA del viroide latente de la berenjena (ELVd) es circularizable con la RNA
ligasa cloroplástica y mediante autoligación, mientras que el del viroide del moteado
clorótico del crisantemo (CChMVd) no lo es de ninguna de las dos maneras. Lo
esperable en un mecanismo no catalizado enzimáticamente sería que todos los RNAs
viroidales se autoligaran en mayor o menor medida, pues teóricamente todos pueden
adquirir la conformación necesaria para que ocurra esta reacción. Sin embargo, estos
resultados se interpretan mejor teniendo en cuenta que los dos RNAs viroidales
circularizables por la RNA ligasa cloroplástica, los del ASBVd y los del ELVd, adoptan
una estructura de cuasi-varilla, mientras que los del CChMVd y PLMVd adoptan una
estructura muy ramificada. Esta observación, junto con el resultado que muestra que
sólo una proporción del RNA total es circularizable in vitro por la RNA cloroplástica,
sugieren que determinadas conformaciones del RNA del ASBVd y ELVd acoplan
correctamente en el sitio catalítico, mientras que con el RNA del CChMVd y PLMVd
no ocurriría así.
Como ya se ha comentado, tanto si la ligación es ribozimática o enzimática,
otras proteínas podrían intervenir ayudando en el primer caso al RNA a adquirir una
conformación catalíticamente activa y en el segundo a que el enlace generado in vivo
sea 3’,5’-fosfodiéster, que es el que parece existir en el RNA circular del ASBVd
aislado de plantas infectadas.
137

Discusión
5.2. La tRNA ligasa de tomate: un hospedador natural de viroides
Como ya se ha comentado en esta discusión, existen indicios acerca de la posible
ligación de los RNAs viroidales nucleares por una tRNA ligasa (Branch et al., 1982;
Kikuchi et al., 1982). Esta enzima ha sido caracterizada en diferentes especies vegetales
como A. thaliana, trigo y arroz (Englert y Beier, 2005; Makino et al., 2006), si bien
ninguna de ellas es hospedador natural o experimental de viroides. Por dicha razón se
abordó en el presente trabajo la posibilidad de clonar la tRNA ligasa de tomate, que es
hospedador experimental del miembro tipo de la familia Pospiviroidae, el PSTVd
(Diener, 1979), y hospedador natural de otros miembros de esta familia como el viroide
de la planta macho del tomate (Tomato planta macho viroid, TPMVd), el viroide del
enanismo apical del tomate (Tomato apical stunt viroid, TASVd) y el viroide de la
exocortis de los cítricos (Citrus exocortis viroid, CEVd) (Galindo et al., 1982; Kiefer et
al., 1983; Mishra et al., 1991). La clonación y expresión de esta tRNA ligasa permitiría
explorar su implicación en la circularización de los viroides nucleares y, en tal caso,
determinar mediante estudios de mutación en el RNA viroidal las regiones del mismo
importantes para esta reacción.
La secuencia buscada se obtuvo partiendo de cuatro ESTs depositados en la base
de datos de la Universidad de Cornell (http://www.sgn.cornell.edu/solanaceae-
project/index.pl). Para obtener la secuencia completa del mRNA de la tRNA ligasa de
tomate se consideraron diversos protocolos de RACE (Zhang y Chiang, 1996; Fu et al.,
2004; Kuhn y Frank-Kamenetskii, 2005; Huang y Chen, 2006; Polidoros et al., 2006).
La variante de la técnica de RACE circular que emplea cDNA molde de simple cadena
puesta a punto en el presente trabajo tiene ventajas comparada con otras técnicas de
RACE, entre las que destaca su especificidad de amplificación debido al empleo de
cebadores específicos de la propia secuencia en ambas direcciones (Fu et al., 2004;
Polidoros et al., 2006). Por el contrario, las variantes de 5’ y 3’-RACE típicas consisten
en añadir una secuencia conocida en el extremo del cDNA lineal, que reduce la
especificidad porque todos los cDNAs presentes en la mezcla de reacción poseen esta
secuencia, facilitando así la amplificación de productos no específicos (Fu et al., 2004;
Polidoros et al., 2006). Otras ventajas de la variante de RACE circular aquí descrita es
su reproducibilidad y simplicidad con respecto a otras variantes de 5’-RACE (Liu y
Gorovsky, 1993; Schmidt y Mueller, 1999; Schramm et al., 2000; revisado en Schaefer,
1995) o de RACE circular (Fu et al., 2004; Kuhn y Frank-Kamenetskii, 2005; Huang y
138

Discusión
Chen, 2006). Además, los extremos 3’ y 5’ del cDNA pueden determinarse en un
mismo ensayo si la RT se realiza con un cebador poli[dT]. Por estas razones dicha
variante resulta de interés para amplificar mRNAs cuando sólo se conoce (o se presume
conservada) una secuencia incompleta del mismo. Esta variante, sin embargo, no está
exenta de aspectos negativos pues durante los experimentos realizados en el presente
trabajo no pudieron amplificarse secuencias mayores de 900 pb, por lo que se hizo
necesario reiterar el procedimiento varias veces. A pesar de todo, los resultados fueron
más reproducibles que las obtenidas con otras modalidades, y permitieron la obtención
del cDNA completo de la tRNA ligasa de tomate.
La secuencia nucleotídica completa de este cDNA, una vez traducida in silico, se
alineó con las secuencias aminoacídicas de las otras tRNA ligasas de plantas conocidas
mediante los programas ClustalW, Lalign y Multalin de la base de datos ExPASy
(http://us.expasy.org/tools/), mostrando una identidad aminoacídica mayor del 60% en
todos los casos. Esta identidad aún sería superior de no ser por la gran variabilidad de la
región amino terminal de las tRNA ligasas de plantas, que si se alinean de forma
independiente del resto de la proteína resulta imposible identificarlas como homólogas
con la excepción de las tres isoformas de trigo. Esta región amino terminal está mucho
más conservada en tRNA ligasas fúngicas como las de Saccharomyces cerevisiae,
Saccharomyces bayanus, Candida albicans, Neurospora crassa, Aspergillus nidulans, y
Schizosaccharomyces pombe (Sawaya et al., 2003). La diferencia de secuencia en las
tRNA ligasas de plantas conocidas podría regular su tiempo de vida media, porque la
región amino terminal de las proteínas determina en gran medida su estabilidad celular
(Bachmair et al., 1986; Gonda et al., 1989; Tobias et al., 1991), ya que sobre dicha
región actúan los sistemas de degradación a través de ubiquitina en eucariotas o de la
proteína ClpS en bacterias (revisado en Mogk et al., 2007). El alineamiento de las
secuencias aminoacídicas de las tRNA ligasas de trigo, arroz, A. thaliana y tomate
también permitió localizar los motivos catalíticos de los dominios ciclofosfodiesterasa,
polinucleótido quinasa y RNA ligasa propiamente dicha. A pesar de las diferencias
entre las secuencias de las tRNA ligasas de plantas y levaduras, estos motivos están
estrictamente conservados en ambos grupos taxonómicos, indicando que son esenciales
para las tres actividades que catalizan estas enzimas. Mediante experimentos de
mutación puntual en levaduras se ha observado que la actividad catalítica desaparece al
cambiar alguno de los aminoácidos de estos motivos (Sawaya et al., 2003; Wang y
Shuman, 2005).
139

Discusión
La introducción del cDNA de la tRNA ligasa de A. thaliana en la cepa Rosetta 2
de E. coli, que compensa la preferencia codónica de eucariotas, muestra que es posible
expresar con elevado rendimiento esta proteína en un sistema bacteriano. Experimentos
anteriores indicaban lo contrario, pues sólo se obtuvo un rendimiento alto al emplear un
sistema in vitro derivado de germen de trigo mientras que la expresión en E. coli
proporcionó un rendimiento muy bajo (Englert y Beier, 2004). El sistema in vitro de
germen de trigo fue empleado para expresar las tRNA ligasas de A. thaliana y arroz,
una vez fueron identificados sus potenciales mRNA, con el fin de comprobar su
actividad catalítica (Englert y Beier, 2005). El sistema in vivo tiene limitaciones, pues
algunas proteínas eucarióticas sufren diversas modificaciones postraduccionales (por
ejemplo, glicosilación) que no pueden ser aportadas por un sistema de expresión
bacteriano. Un segundo problema radica en el elevado poder reductor del citoplasma de
E. coli, que dificulta la correcta formación de los puentes disulfuro esenciales para el
correcto plegamiento y solubilidad de ciertas proteínas (revisado en Weickert et al.,
1996). Finalmente, el fenómeno de preferencia codónica, consistente en que bacterias y
eucariotas emplean con diferente frecuencia determinados codones (Grantham et al.,
1980), también contribuye a que la expresión en un sistema bacteriano de proteínas
grandes como las tRNA ligasas ocurra con un rendimiento bajo o insuficiente. Sin
embargo, la expresión en un sistema bacteriano de proteínas con pocos (o ningún)
puente disulfuro y que no necesiten modificaciones postraduccionales es de gran interés
porque el procedimiento es fácilmente escalable, económico y conduce a rendimientos
de hasta un 60-70% del total de proteína sintetizada por la célula (Kane, 1995).
La expresión en la cepa Rosetta 2 de E. coli del cDNA de la tRNA ligasa de
tomate y la posterior purificación y ensayo de la correspondiente proteína confirmó
definitivamente que se trataba de la tRNA ligasa buscada. Al analizar las proteínas
eluidas de la columna de Ni-NTA-agarosa mediante SDS-PAGE y tinción con plata se
observó que el rendimiento obtenido con la tRNA ligasa de tomate fue inferior al de su
homóloga de A. thaliana. Este menor rendimiento no es debido a errores de clonación,
pues la correcta inserción del cDNA de la tRNA ligasa de tomate en el vector de
expresión se comprobó secuenciando los extremos del mismo, descartándose también
errores experimentales en la purificación repitiendo el procedimiento varias veces.
Como ya se ha comentado, la diferencia más importante entre las secuencias
aminoacídicas de las tRNA ligasas de tomate y A. thaliana se localiza en la región
amino terminal. Mediante el uso del programa ProtParam de la base de datos ExPASy
140

Discusión
(http://us.expasy.org/tools/protparam.html) se observó que las enzimas de A. thaliana y
tomate tienen un índice de estabilidad teórico inferior y superior a 40, respectivamente,
que podría explicar las diferencias de rendimiento en su purificación. Sin embargo,
considerando que estas diferencias son pequeñas, no es posible descartar la influencia
de otros factores desconocidos.
Los experimentos bioquímicos realizados con las tRNA ligasas de A. thaliana y
tomate mostraron que ambas enzimas presentan valores similares de temperatura y pH
óptimo, y que dichos valores se asemejan a los descritos para la tRNA ligasa de germen
de trigo (Pick y Hurwitz, 1986; Pick et al., 1986). En estos trabajos previos se
comprobó que el ATP es el único NTP requerido para la ligación catalizada por la
enzima de germen de trigo, no habiendo sido estudiada esta característica bioquímica en
las tRNA ligasas de A. thaliana y arroz (Englert y Beier, 2005; Wang et al., 2006a). Los
resultados del presente trabajo muestran que las tRNA ligasas de tomate y A. thaliana
catalizan la ligación empleando no sólo ATP sino también UTP, si bien con un
rendimiento ligeramente inferior. También se detectó ligación con GTP, aunque con un
rendimiento muy pobre.
Experimentos similares realizados con la tRNA ligasa de levadura han revelado
que es catalíticamente activa en presencia de cualquiera de los cuatro NTPs, aunque con
preferencia por el ATP y el UTP (Greer et al., 1983b). Sin embargo, en trabajos
posteriores del mismo laboratorio se observó que la ligación sólo ocurre con ATP,
concluyéndose que los resultados previos (Greer et al., 1983b) fueron debidos a una
contaminación de la preparación con NTPs (Westaway et al., 1993; Belford et al.,
1993). Las tRNA ligasas emplean los NTPs en dos etapas diferentes, la fosforilación y
la adenilación, que están catalizadas por los dominios polinucleótido quinasa y RNA
ligasa, respectivamente. En experimentos realizados con la tRNA ligasa de levadura
(Westaway et al., 1993; Belford et al., 1993) se observó que el ATP es imprescindible
para la adenilación, y que si bien la fosforilación también ocurre con ATP, funciona
más eficientemente con cualquiera de los otros NTPs, fundamentalmente con GTP.
Westaway et al. (1993) también observaron que una tRNA ligasa de levadura a la que
se le había eliminado el módulo polinucleótido quinasa también catalizaba la ligación
con GTP, y en menor medida con UTP (a diferencia de lo observado con la enzima
completa). Además Beldford et al. (1993) mostraron que la tRNA ligasa de levadura
silvestre cataliza más eficientemente la ligación del sustrato si además de ATP está
presente en la reacción cualquiera de los otros tres NTPs (especialmente GTP). Ensayos
141

Discusión
de mutación posteriores confirmaron que dicha tRNA ligasa silvestre de levadura posee
dos sitios de unión diferentes para NTPs, uno para ATP y otro para GTP (Sawaya et al.,
2003).
Sin embargo, los experimentos de mutación puntual realizados con la tRNA
ligasa de A. thaliana no han permitido caracterizar el sitio de unión al GTP de esta
enzima (Wang et al., 2006). Además, en el presente trabajo se muestra que las enzimas
de tomate y A. thaliana catalizan por igual la ligación tanto con ATP como con una
combinación de los cuatro NTPs. Este resultado difiere del descrito para la tRNA ligasa
de levadura (Belford et al., 1993) y, conjuntamente con el observado por Wang et al.
(2006), sugiere que las tRNA ligasas de A. thaliana y tomate contienen un único sitio de
unión para NTPs. El UTP que estas dos tRNA ligasas pueden emplear para catalizar la
ligación del RNA es presumiblemente incorporado a través del sitio de unión para el
ATP, si bien son necesarios experimentos de mutación más detallados para esclarecer
este aspecto.
Una vez disponibles las secuencias del RNA del PSTVd (Gross et al., 1978) y
del cDNA de la tRNA ligasa de tomate, pueden acometerse diversos estudios
mutacionales que permitan analizar in vitro e in vivo qué motivos de secuencia y
estructura, tanto del RNA como de la enzima, están implicados en la reacción de
ligación. De esta forma podría conocerse un poco mejor cual es el mecanismo de
circularización que completa el ciclo replicativo de estos patógenos. Sin embargo,
aunque experimentos previos muestran que la tRNA ligasa de germen de trigo
circulariza in vitro los RNAs monoméricos lineales del PSTVd (Branch et al., 1982),
otros resultados recientes de nuestro laboratorio (Gas et al., 2007; Gas et al., 2008)
sugieren que los RNAs lineales de los viroides nucleares tienen extremos 5’-
fosfomonoéster y 3’-hidroxilo. Esta observación abre la puerta a la existencia de otra
RNA ligasa con propiedades diferentes, aunque la tRNA ligasa también podría catalizar
la circularización de RNAs viroidales con estos extremos siempre que el sustrato
contenga un grupo 2’-fosfomonoéster. En este supuesto sería necesaria la presencia en
la célula de una enzima con actividad quinasa en 2’ que no ha sido previamente
descrita. Experimentos adicionales permitirán resolver la cuestión de si la ligación es
catalizada por una nueva RNA ligasa o por la tRNA ligasa con la ayuda de una enzima
con actividad quinasa en 2’. En todo caso, la identificación y purificación de la tRNA
ligasa de tomate descrita en el presente trabajo será de utilidad para este cometido.
142

Discusión
5.3. Los RNAs de la familia Avsunviroidae se procesan correctamente
en el cloroplasto de C. reinhardtii
Los miembros de la familia Avsunviroidae son muy restrictivos en cuanto a sus
hospedadores, ninguno de los cuales es un organismo modelo. Además, la
transformación de cloroplastos de plantas superiores es complicada, por lo que el
sistema heterólogo basado en el cloroplasto de C. reinhardti descrito en el presente
trabajo resulta de interés para estudiar in vivo en condiciones de laboratorio dos de las
tres etapas de la replicación de los viroides cloroplásticos: el corte y la ligación. Las
construcciones de cDNAs viroidales diméricos del ASBVd, CChMVd, ELVd y CEVd
(todos cloroplásticos excepto este último, empleado como control) se transcribieron
correctamente en el cloroplasto de C. reinhardtii, detectándose la señal correspondiente
a los transcritos primarios y los productos de su procesamiento en experimentos de
hibridación northern. Los cDNAs viroidales empleados fueron diméricos porque los
transcritos obtenidos a partir de los mismos se asemejan a los intermediarios
replicativos de los RNAs viroidales in vivo, y porque los RNAs monoméricos generados
tras el procesamiento de las formas diméricas son muy infecciosos, como se ha
comprobado en experimentos de inoculación mecánica en sus hospedadores (Cress et
al., 1983; Navarro y Flores, 1997; Ambrós et al., 1998).
El corte de los RNAs oligoméricos en la familia Avsunviroidae es autocatalítico
y mediado por ribozimas de cabeza de martillo formadas por secuencias contenidas en
las cadenas de ambas polaridades (Hutchins et al., 1986; Prody et al., 1986; Flores et
al., 2001), como ponen de manifiesto una serie de trabajos in vitro e in vivo (Hernández
y Flores, 1992; Daròs et al., 1994; Navarro y Flores, 1997; Fadda et al., 2003a; Delgado
et al., 2005). Sin embargo, algunos aspectos críticos del funcionamiento de las
ribozimas de cabeza de martillo permanecen aún desconocidos, como queda ilustrado
por el reciente hallazgo de que interacciones entre regiones periféricas de esta clase de
ribozimas naturales incrementan notablemente su actividad catalítica a las bajas
concentraciones de magnesio existentes in vivo (Khvorova et al., 2003; De la Peña et
al., 2003; Martick y Scott, 2006). La expresión en cloroplastos de C. reinhardtii de los
RNAs diméricos del ASBVd (de ambas polaridades), del CChMVd y del ELVd
[polaridad (+)], muestra que el autocorte funciona en este contexto fisiológico
generando las formas monoméricas lineales. Mientras que este resultado es consistente
con los de experimentos previos in vitro (revisado en Flores et al., 2001), la eficiencia
143

Discusión
del autocorte depende de cada RNA viroidal.
Las ribozimas de cabeza de martillo pueden clasificarse en simples, si la
secuencias que forman el centro catalítico se encuentran incluidas en un RNA
monomérico, o dobles si dicho centro se forma a partir de RNAs diméricos (Hutchins et
al., 1986; Prody et al., 1986; Forster et al., 1988). Todos los viroides cloroplásticos, con
la excepción del ASBVd, contienen ribozimas de tipo simple que son
termodinámicamente más estables y se autocortan mejor in vitro. Sin embargo, los
resultados obtenidos en el presente trabajo muestran que no existe una relación entre la
eficiencia de la reacción de autocorte y que la misma esté mediada por ribozimas de
cabeza de martillo simples o dobles. Es presumible que in vivo dicha eficiencia venga
más condicionada por factores auxiliares del cloroplasto que por el tipo de ribozima. En
particular, debido a que el RNA se pliega y en su caso se autocorta durante la
transcripción (De la Peña et al., 2003; Mahen et al., 2005; Carbonell et al., 2006), el
bajo autocorte en C. reinhardtii de algunos RNA viroidales como el de polaridad (+) del
CChMVd pudiera resultar de quedar atrapados en conformaciones catalíticamente
inactivas (Foster y Symons, 1987), o de mostrar baja afinidad por chaperonas de RNA
cloroplásticas que presumiblemente asisten esta reacción in vivo. En este contexto es
pertinente resaltar que una proteína cloroplástica de aguacate que se une al ASBVd in
vivo se comporta como una chaperona de RNA que estimula el autocorte in vitro de un
RNA dimérico viroidal (Daròs y Flores 2002). Alternativamente, puesto que los RNAs
monoméricos provenientes de autocorte carecen de regiones 5’-UTR que como la del
gen rbcL otorgan estabilidad a los RNAs cloroplásticos (Suay et al., 2005), la estructura
secundaria de cada RNA viroidal podría condicionar diferencialmente la degradación de
el mismo.
Con respecto a la ligación de los RNAs monoméricos resultantes, ya se ha
comentado anteriormente que en el PLMVd puede ocurrir autocatalíticamente in vitro a
través de la formación de un enlace 2’,5’-fosfodiéster (Côté y Perreault, 1997). Sin
embargo, y a pesar de que se ha propuesto la existencia de este tipo de enlaces en RNAs
circulares del PLMVd aislados de tejido infectado (Côté et al., 2001), los resultados
obtenidos en el presente trabajo muestran que dicho enlace atípico no parece existir en
los RNAs circulares del ASBVd provenientes de plantas infectadas. Por lo tanto, la
autoligación descrita en el PLMVd posiblemente no esté actuando en el ASBVd, donde
es más probable que una RNA ligasa del hospedador catalicen la formación de enlaces
3’,5’-fosfodiéster. Dicha ligación enzimática podría estar catalizada en C. reinhardtii
144

Discusión
por la RNA ligasa detectada con propiedades similares a las tRNA ligasas si tiene
localización cloroplástica (Kikuchi et al., 1982), y en plantas por la RNA ligasa
cloroplástica descrita en el presente trabajo, en este segundo caso con la posible ayuda
de una chaperona que facilite la adquisición por el RNA viroidal de un plegamiento
adecuado. Alternativamente, las ribozimas de cabeza de martillo podrían funcionar a la
inversa, ya que esta ligación ha sido descrita en varias ribozimas de cabeza de martillo
mínimas o extendidas (Hertel et al., 1994; Nelson et al., 2005; Canny et al., 2007). En
este proceso también sería presumiblemente necesaria una chaperona para ayudar al
RNA a adquirir una conformación catalíticamente activa. En cualquier caso, la relación
entre los RNAs monoméricos circulares y lineales que se observa en los cloroplastos de
C. reinhardtii transformados es generalmente similar a la existente in planta y, por lo
tanto, este sistema transplastómico podría tener interés para identificar los factores del
hospedador que median esta reacción in vivo.
Para completar su ciclo replicativo los viroides cloroplásticos requieren que una
RNA polimerasa catalice la transcripción RNA-RNA (Hutchins et al., 1985; Daròs et
al., 1994). Experimentos previos han mostrado que en hospedadores naturales esta
enzima es muy probablemente del tipo NEP, pues altas concentraciones de tagetitoxina
que inhiben fuertemente la transcripción de genes cloroplásticos sólo disminuyen la
transcripción del ASBVd en un 25% (Navarro et al., 2000). Por otra parte, en
melocotoneros infectados con una variante de secuencia del PLMVd que provoca una
clorosis foliar muy intensa se ha observado que la PEP no es funcional en las regiones
albinas, mientras que los RNAs transcritos por la NEP no se ven afectados. En estas
regiones, la acumulación de las formas circulares y lineales de ambas polaridades del
PLMVd es incluso mayor que en las regiones adyacentes verdes. Estos resultados in
vivo sostienen que la RNA polimerasa implicada en la replicación del PLMVd también
es la NEP (Rodio et al., 2007).
La expresión en cloroplastos de C. reinhardtii de RNAs diméricos de una u otra
polaridad del ASBVd descrita en el presente trabajo no ha permitido detectar la
acumulación de las respectivas cadenas complementarias, sugiriendo que con este
viroide, y posiblemente con los otros miembros de la familia, no ocurre transcripción
RNA-RNA. Experimentos con rifampicina muestran que la transcripción cloroplástica
en C. reinhardtii es sensible a dicho antibiótico, un inhibidor de la PEP pero no de la
NEP (Eberhard et al., 2002). Este resultado, junto con los intentos infructuosos de
obtener mutantes de C. reinhardtii sin actividad PEP, sugiere que el cloroplasto de
145

Discusión
dicha alga carece de actividad NEP (Bohne et al., 2006), y explica la ausencia de
transcripción RNA-RNA en los cloroplastos de la misma transformados con RNA
viroidales. Este aspecto, lejos de ser un inconveniente, supone una ventaja para estudiar
de forma directa el efecto de mutaciones que afecten específicamente a las reacciones
de corte o ligación.
En resumen, el sistema experimental descrito en esta memoria consistente en la
expresión de RNAs viroidales en el cloroplasto de C. reinhardtii, puede facilitar el
estudio en un contexto similar al fisiológico de los motivos de RNA y de los factores
cloroplásticos implicados en la replicación viroidal. Este sistema, junto con la detección
y caracterización de la primera RNA ligasa cloroplástica y los últimos avances
relacionados con las ribozimas de cabeza de martillo funcionando como RNA ligasas,
podría aclarar la cuestión de si la ligación en los miembros de la familia Avsunviroidae
es enzimática o ribozimática. Desde otra perspectiva más biotecnológica, la expresión
de genes heterólogos en el cloroplasto es una alternativa experimental a la
transformación nuclear (Maliga, 2004), y los RNAs de la familia Avsunviroidae, los
únicos capaces de acceder y replicarse en este orgánulo, podrían servir como vectores
para este cometido. Así pues, el sistema heterólogo aquí descrito es una herramienta con
utilidad potencial para trabajos posteriores dirigidos a profundizar en la replicación de
los viroides y en posibles derivaciones biotecnológicas.
146

66.. CCOONNCCLLUUSSIIOONNEESS


Conclusiones
1) Se ha detectado una actividad de ligación de RNA en preparaciones de
cloroplastos de espinaca fraccionadas en columna de heparina. La
correspondiente RNA ligasa tiene unas características bioquímicas diferentes
de las tRNA ligasas descritas en eucariotas.
2) La RNA ligasa cloroplástica de espinaca es capaz de circularizar in vitro los
RNAs monoméricos lineales de polaridad positiva de dos miembros de la
familia Avsunviroidae, concretamente los que presentan una conformación
menos ramificada.
3) La ligación de los RNAs monoméricos viroidales está mediada por una
proteína, como demuestran los experimentos de digestión con proteinasa K, y
sucede en ausencia de NTPs y magnesio, generando un enlace diferente del
3’,5’-fosfodiéster fisiológico. Estas características bioquímicas establecen un
vínculo entre la RNA ligasa cloroplástica con las 2’,5’-RNA ligasas de
bacterias y arqueas.
4) Se ha puesto a punto una metodología de RACE circular con un molde de
simple cadena que permite caracterizar mRNAs completos a partir de una
secuencia interna conocida y que es más sencilla que otras metodologías
similares que emplean un molde de doble cadena.
5) Se ha clonado y caracterizado un homólogo de la tRNA ligasa de eucariotas en
tomate que tiene actividad con una gama de NTPs más amplia que las tRNAs
ligasas de germen de trigo y levadura. Esta propiedad es compartida por la
tRNA ligasa de A. thaliana homóloga.
6) Se ha expresado con un rendimiento elevado la tRNA ligasa de A. thaliana en
la cepa de E. coli Rosetta 2, que corrije la preferencia codónica, y que podría
sustituir satisfactoriamente a otros sistemas de expresión basados en extractos
celulares eucarióticos. Sin embargo, el rendimiento obtenido al expresar la
tRNA ligasa de tomate fue inferior, posiblemente porque su región amino
149

Conclusiones
terminal sea una diana mejor para los mecanismos de degradación celulares, si
bien no pueden descartarse la implicación de otros factores.
7) El sistema viroide-cloroplasto de Chlamydomonas reinhardtii permite la
expresión de RNAs viroidales diméricos en este orgánulo, así como su
correcto procesamiento. Además, la relación entre las formas lineales y
circulares de los RNAs monoméricos viroidales en el cloroplasto de este alga
se asemeja a la observada en plantas infectadas por estos mismos viroides en
tres de los cuatro casos analizados. En consecuencia, este sistema puede ser
muy interesante para el estudio de las interacciones planta-viroide en un
entorno similar al de sus hospedadores naturales, pero en un organismo
modelo de laboratorio que tiene su genoma secuenciado. Este último punto
puede facilitar mucho el análisis de los factores del hospedador relacionados
con el procesamiento de los viroides.
8) Al no producirse la transcripción RNA-RNA que completaría el ciclo
replicativo de los miembros de la familia Avsunviroidae, el sistema basado en
C. reinhardtii permite estudiar de forma directa el efecto de las mutaciones en
los transcritos viroidales primarios que afecten específicamente al corte o la
ligación.
150

77.. BBIIBBLLIIOOGGRRAAFFÍÍAA


Bibliografía
ABELSON, J., TROTTA, C.R. & LI, H. (1998). tRNA splicing. Journal of Biological Chemistry, 273, 12685-12688.
ALTSCHUL, S.F., GISH, W., MILLER, W., MYERS, E.W. & LIPMAN, D.J. (1990).
Basic local alignment search tool. Journal of Molecular Biology, 215, 403-410.
ALTSCHUL, S.F., MADDEN, T.L., SCHAFFER, A.A., ZHANG, J.H., ZHANG, Z., MILLER, W. & LIPMAN, D.J. (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Research, 25, 3389-3402.
AMBRÓS, S., HERNÁNDEZ, C., DESVIGNES, J.C. & FLORES, R. (1998). Genomic structure of three phenotypically different isolates of peach latent mosaic viroid: implications of the existence of constraints limiting the heterogeneity of viroid quasispecies. Journal of Virology, 72, 7397-7406.
AMBRÓS, S., HERNÁNDEZ, C. & FLORES, R. (1999). Rapid generation of genetic heterogeneity in progenies from individual cDNA clones of peach latent mosaic viroid in its natural host. Journal of General Virology, 80, 2239-2252.
AMITSUR, M., LEVITZ, R. & KAUFMANN, G. (1987). Bacteriophage-T4 anticodon
nuclease, polynucleotide kinase and RNA ligase reprocess the host lysine tRNA. EMBO Journal, 6, 2499-2503.
AMITSUR, M., MORAD, I., CHAPMANSHIMSHONI, D. & KAUFMANN, G. (1992). HSD restriction modification proteins partake in latent anticodon nuclease. EMBO Journal, 11, 3129-3134.
ANDERSSON, J.O. & ROGER, A.J. (2002). A cyanobacterial gene in nonphotosynthetic protists: an early chloroplast acquisition in eukaryotes? Current Biology, 12, 115-119.
APOSTOL, B.L., WESTAWAY, S.K., ABELSON, J. & GREER, C.L. (1991). Deletion
analysis of a multifunctional yeast tRNA ligase polypeptide: identification of essential and dispensable functional domains. Journal of Biological Chemistry, 266, 7445-7455.
ARN, E.A. & ABELSON, J.N. (1996). The 2'-5' RNA ligase of Escherichia coli: purification, cloning, and genomic disruption. Journal of Biological Chemistry, 271, 31145-31153.
ARN, E.A. & ABELSON, J. (1998). RNA ligases: function, mechanism and sequence conservation. In RNA Structure and Function (Simons, R. & Grunberg-Manago, M. ed.). Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 695-726.
BACHMAIR, A., FINLEY, D. & VARSHAVSKY, A. (1986). In vivo half-life of a protein is a function of its amino-terminal residue. Science, 234, 179-186.
153

Bibliografía
BAKALARA, N., SIMPSON, A.M. & SIMPSON, L. (1989). The Leishmania kinetoplast-mitochondrion contains terminal uridylyltransferase and RNA ligase activities. Journal of Biological Chemistry, 264, 18679-18686.
BAUMSTARK, T., SCHRODER, A.R.W. & RIESNER, D. (1997). Viroid processing:
switch from cleavage to ligation is driven by a change from a tetraloop to a loop E conformation. EMBO Journal, 16, 599-610.
BELFORD, H.G., WESTAWAY, S.K., ABELSON, J. & GREER, C.L. (1993). Multiple nucleotide cofactor use by yeast ligase in tRNA splicing: evidence for independent ATP-binding and GTP-binding sites. Journal of Biological Chemistry, 268, 2444-2450.
BELLÉS, J.M., GRANELL, A., DURAN-VILA, N. & CONEJERO, V. (1989). ACC synthesis as the activated step responsible for the rise of ethylene production accompanying citrus exocortis viroid infection in tomato plants. Journal of Phytopathology, 125, 198-208.
BHATTACHARYA, D. & MEDLIN, L. (1998). Algal phylogeny and the origin of land
plants. Plant Physiology, 116, 9-15.
BLUM, H., BEIER, H. & GROSS, H.J. (1987). Improved silver staining of plant-proteins, RNA and DNA in polyacrylamide gels. Electrophoresis, 8, 93-99.
BOGORAD, L. (2000). Engineering chloroplasts: an alternative site for foreign genes, proteins, reactions and products. Trends in Biotechnology, 18, 257-263.
BOHNE, A.V., IRIHIMOVITCH, V., WEIHE, A. & STERN, D.B. (2006). Chlamydomonas reinhardtii encodes a single sigma70-like factor which likely functions in chloroplast transcription. Current Genetics, 49, 333-340.
BONFIGLIOLI, R.G., MC FADDEN, G.I. & SYMONS, R.H. (1994). In situ hybridization localizes avocado sunblotch viroid on chloroplast thylakoid membranes and coconut cadang cadang viroid in the nucleus. Plant Journal, 6, 99-103.
BONFIGLIOLI, R.G., WEBB, D.R. & SYMONS, R.H. (1996). Tissue and intra-cellular distribution of coconut cadang cadang viroid and citrus exocortis viroid determined by in situ hybridization and confocal laser scanning and transmission electron microscopy. Plant Journal, 9, 457-465.
BOYNTON, J.E., GILLHAM, N.W., HARRIS, E.H., HOSLER, J.P., JOHNSON, A.M., JONES, A.R., RANDOLPH-ANDERSON, B.L., ROBERTSON, D., KLEIN, T.M., SHARK, K.B. & SANFORD, J.C. (1988). Chloroplast transformation in Chlamydomonas with high velocity microprojectiles. Science, 240, 1534-1538.
BRANCH, A.D., BENENFELD, B.J. & ROBERTSON, H.D. (1988). Evidence for a single rolling circle in the replication of potato spindle tuber viroid. Proceedings of the National Academy of Sciences of the United States of America, 85, 9128-9132.
154

Bibliografía
BRANCH, A.D. & DICKSON, E. (1980). Tomato DNA contains no detectable regions complementary to potato spindle tuber viroid as assayed by southern hybridization. Virology, 104, 10-26.
BRANCH, A.D. & ROBERTSON, H.D. (1984). A replication cycle for viroids and other small infectious RNAs. Science, 223, 450-455.
BRANCH, A.D., ROBERTSON, H.D., GREER, C., GEGENHEIMER, P., PEEBLES, C. & ABELSON, J. (1982). Cell-free circularization of viroid progeny RNA by an RNA ligase from wheat germ. Science, 217, 1147-1149.
BRINKMAN, F.S.L., BLANCHARD, J.L., CHERKASOV, A., AV-GAY, Y., RUNHAM, R.C., FERNANDEZ, R.C., FINLAY, B.B., OTTO, S.P., OUELLETTE, B.F.F., KEELING, P.J., ROSE, A.M., HANCOCK R.E.W. & JONES, S.J.M. (2002). Evidence that plant-like genes in chlamydia species reflect an ancestral relationship between chlamydiaceae, cyanobacteria, and the chloroplast. Genome Research, 12, 1159-1167.
BRINKMANN, U., MATTES, R.E. & BUCKEL, P. (1989). High-level expression of
recombinant genes in Escherichia coli is dependent on the availability of the dnaY gene product. Gene, 85, 109-114.
BUSSIÈRE, F., LEHOUX, J., THOMPSON, D.A., SKRZECZKOWSKI, L.J. &
PERREAULT, J.P. (1999). Subcellular localization and rolling circle replication of peach latent mosaic viroid: hallmarks of group A viroids. Journal of Virology, 73, 6353-6360.
BUSSIÈRE, F., OUELLET, J., CÔTÉ, F., LÉVESQUE, D. & PERREAULT, J.P.
(2000). Mapping in solution shows the peach latent mosaic viroid to possess a new pseudoknot in a complex, branched secondary structure. Journal of Virology, 74, 2647–2654.
CAMERON, V. & UHLENBECK, O.C. (1977). 3'-phosphatase activity in T4
polynucleotide kinase. Biochemistry, 16, 5120-5126.
CANNY, M.D., JUCKER, F.M. & PARDI, A. (2007). Efficient ligation of the Schistosoma hammerhead ribozyme. Biochemistry, 46, 3826-3834.
CARBONELL, A., DE LA PEÑA, M., FLORES, R. & GAGO, S. (2006). Effects of the trinucleotide preceding the self-cleavage site on eggplant latent viroid hammerheads: differences in co- and post-transcriptional self-cleavage may explain the lack of trinucleotide AUC in most natural hammerheads. Nucleic Acids Research, 34, 5613-5622.
CHENNA, R., SUGAWARA, H., KOIKE, T., LOPEZ, R., GIBSON, T.J., HIGGINS,
D.G. & THOMPSON, J.D. (2003). Multiple sequence alignment with the Clustal series of programs. Nucleic Acids Research, 31, 3497-3500.
CONEJERO, V., PICAZO, I. & SEGADO, P. (1979). Citrus exocortis viroid (CEV): protein alterations in different hosts following viroid infection. Virology, 97, 454-456.
155

Bibliografía
CONEJERO, V. & SEMANCIK, J.S. (1977). Exocortis viroid: alteration in proteins of Gynura aurantiaca accompanying viroid infection. Virology, 77, 221-232.
CÔTÉ, F., LÉVESQUE, D. & PERREAULT, J.P. (2001). Natural 2',5'-phosphodiester bonds found at the ligation sites of peach latent mosaic viroid. Journal of Virology, 75, 19-25.
CÔTÉ, F. & PERREAULT, J.P. (1997). Peach latent mosaic viroid is locked by a 2',5'-phosphodiester bond produced by in vitro self-ligation. Journal of Molecular Biology, 273, 533-543.
COX, K. H. & GOLBERG, R. B. (1988). Analysis of plant gene expression. In Plant Molecular Biology: A Practical Approach (C.H. Shaw, ed.). IRL Press, Oxford, 1-35.
CRANSTON, J.W., SILBER, R., MALATHI, V.G. & HURWITZ, J (1974). Studies on ribonucleic acid ligase: characterization of an adenosine triphosphate-inorganic pyrophosphate exchange reaction and demonstration of an enzyme-adenylate complex with T4 bacteriophage-induced enzyme. Journal of Biological Chemistry, 249, 7447-7456.
CRESS, D.E., KIEFER, M.C. & OWENS, R.A. (1983). Construction of infectious potato spindle tuber viroid cDNA clones. Nucleic Acids Research, 11, 6821-6835.
CULVER, G.M., CONSAUL, S.A., TYCOWSKI, K.T., FILIPOWICZ, W. & PHIZICKY, E.M. (1994). tRNA splicing in yeast and wheat germ: a cyclic phosphodiesterase implicated in the metabolism of ADP-ribose 1",2"-cyclic phosphate. Journal of Biological Chemistry, 269, 24928-24934.
CULVER, G.M., MCCRAITH, S.M., CONSAUL, S.A., STANFORD, D.R. &
PHIZICKY, E.M. (1997). A 2'-phosphotransferase implicated in tRNA splicing is essential in Saccharomyces cerevisiae. Journal of Biological Chemistry, 20, 13203-13210.
CULVER, G.M., MCCRAITH, S.M., ZILLMANN, M., KIERZEK, R., MICHAUD, N., LAREAU, R.D., TURNER, D.H. & PHIZICKY, E.M. (1993). A NAD derivative produced during transfer RNA splicing: ADP-ribose 1”-2” cyclic phosphate. Science, 261, 206-208.
DARÒS, J.A., ELENA, S.F. & FLORES, R (2006). Viroids: an Ariadne’s thread into the RNA labyrinth. EMBO Reports, 7, 593-598.
DARÒS, J.A. & FLORES, R. (2002). A chloroplast protein binds a viroid RNA in vivo and facilitates its hammerhead-mediated self-cleavage. EMBO Journal, 21, 749-759.
DARÒS, J.A. & FLORES, R. (2004). Arabidopsis thaliana has the enzymatic machinery for replicating representative viroid species of the family Pospiviroidae. Proceedings of the National Academy of Sciences of the United States of America, 101, 6792-6797.
156

Bibliografía
DARÒS, J.A., MARCOS, J.F., HERNÁNDEZ, C. & FLORES, R. (1994). Replication of avocado sunblotch viroid: evidence for a symmetrical pathway with two rolling circles and hammerhead ribozyme processing. Proceedings of the National Academy of Sciences of the United States of America, 91, 12813-12817.
DAVIES, J.W., KAESBERG, P. & DIENER, T.O. (1974). Potato spindle tuber viroid XII: an investigation of viroid RNA as a messenger for protein synthesis. Virology, 61, 281-286.
DAVIES, C., SHELDON, C.C. & SYMONS, R.H. (1991). Alternative hammerhead structures in the self-cleavage of avocado sunblotch viroid RNAs. Nucleic Acids Research, 19, 1893-1898.
DE LA PEÑA, M. & FLORES, R. (2002). Chrysanthemum chlorotic mottle viroid
RNA: dissection of the pathogenicity determinant and comparative fitness of symptomatic and non-symptomatic variants. Journal of Molecular Biology, 321, 411-421.
DE LA PEÑA, M., GAGO, S. & FLORES, R. (2003). Peripheral regions of natural
hammerhead ribozymes greatly increase their self-cleavage activity. EMBO Journal, 20, 5561-5570.
DE LA PEÑA, M., NAVARRO, B. & FLORES, R. (1999). Mapping the molecular determinant of pathogenicity in a hammerhead viroid: A tetraloop within the in vivo branched RNA conformation. Proceedings of the National Academy of Sciences of the United States of America, 96, 9960-9965.
DELGADO, S., MARTÍNEZ DE ALBA, A.E., HERNÁNDEZ, C. & FLORES, R. (2005). A short double-stranded RNA motif of peach latent mosaic viroid contains the initiation and the self-cleavage sites of both polarity strands. Journal of Virology, 79, 12934-12943.
DESJARDINS, P.R. (1987). Avocado sunblotch. In The Viroids (T.O. Diener, ed.). Plenum, New York, 299-313.
DIENER, T.O. (1971). Potato spindle tuber virus: plant virus with properties of a free nucleic acid III: subcellular location of PSTV-RNA and question of whether virions exist in extracts or in situ. Virology, 43, 75-98.
DIENER, T.O. (1979). Viroids and Viroid Diseases. Wiley-Interscience, New York.
DIENER, T.O. (1989). Circular RNAs: relics of precellular evolution? Proceedings of the National Academy of Sciences of the United States of America, 86, 9370-9374.
DIENER, T.O. (2001). The viroid: biological oddity or evolutionary fossil? Advances in Virus Research, 57, 137-184.
DING, B. & ITAYA, A. (2007). Viroid: a useful model for studying the basic principles of infection and RNA biology. Molecular Plant-Microbe Interactions, 20, 7-20.
157

Bibliografía
EBERHARD, S., DRAPIER, D. & WOLLMAN, F.A. (2002). Searching limiting steps in the expression of chloroplast-encoded proteins: relations between gene copy number, transcription, transcript abundance and translation rate in the chloroplast of Chlamydomonas reinhardtii. Plant Journal, 31, 149-160.
EDGAR, R.C. (2004). MUSCLE: multiple sequence alignment with high accuracy and
high throughput. Nucleic Acids Research, 32, 1792-1797. EL OMARI, K., REN, J., BIRD, L.E., BONA, M.K., KLARMANN, G., LEGRICE,
S.F.J. & STAMMERS, D.K. (2006). Molecular architecture and ligand recognition determinants for T4 RNA ligase. Journal of Biological Chemistry, 281, 1573-1579.
ENGLARD, S. & SEIFTER, S. (1990). Precipitation techniques. Methods in Enzymology, 182, 285-300.
ENGLERT, M. & BEIER, H. (2004). Efficient overexpression of functionally active
yeast tRNA ligase in the RTS 100 wheat germ lysate. Biochemica, 3, 27-29. ENGLERT, M. & BEIER, H. (2005). Plant tRNA ligases are multifunctional enzymes
that have diverged in sequence and substrate specificity from RNA ligases of other phylogenetic origins. Nucleic Acids Research, 33, 388-399.
FABREGA, C., SHEN, V., SHUMAN, S. & LIMA, C.D. (2003). Structure of an mRNA capping enzyme bound to the phosphorylated carboxy-terminal domain of RNA polymerase II. Molecular Cell, 11, 1549-1561.
FADDA, Z., DARÒS, J.A., FAGOAGA, C., FLORES, R. & DURAN-VILA, N. (2003a). Eggplant latent viroid, the candidate type species for a new genus within the family Avsunviroidae (hammerhead viroids). Journal of Virology, 77, 6528-6532.
FADDA, Z., DARÒS, J.A., FLORES, R. & DURAN-VILA, N. (2003b). Identification in eggplant of a variant of citrus exocortis viroid (CEVd) with a 96 nucleotide duplication in the right terminal region of the rod-like secondary structure. Virus Research, 97, 145-149.
FAN, W.H., WOELFLE, M.A. & MOSIG, G. (1995). Two copies of a DNA element,
'Wendy', in the chloroplast chromosome of Chlamydomonas reinhardtii between rearranged gene clusters. Plant Molecular Biology, 29, 63-80.
FELS, A., HU, K.H. & RIESNER, D. (2001) Transcription of potato spindle tuber viroid by RNA polymerase II starts predominantly at two specific sites. Nucleic Acids Research, 29, 4589-4597.
FERRIS, P.J. & GOODENOUGH, U.W. (1994). The mating-type locus of
Chlamydomonas reinhardtii contains highly rearranged DNA sequences. Cell, 76, 1135-1145.
FILIPOWICZ, W., KONARSKA, M., GROSS, H.J. & SHATKIN, A.J. (1983). RNA 3'-terminal phosphate cyclase activity and RNA ligation in HeLa cell extract. Nucleic Acids Research, 11, 1405-1418.
158

Bibliografía
FILIPOWICZ, W. & SHATKIN, A.J. (1983). Origin of splice junction phosphate in tRNAs processed by HeLa cell extract. Cell, 32, 547-557.
FLORES, R., DELGADO, S., GAS, M.E., CARBONELL, A., MOLINA, D., GAGO, S.
& DE LA PEÑA, M. (2004). Viroids: the minimal non-coding RNAs with autonomous replication. FEBS Letters, 567, 42-48.
FLORES, R., DURAN-VILA, N., PALLAS, V. & SEMANCIK, J.S. (1985). Detection
of viroid and viroid-like RNAs from grapevine. Journal of General Virology, 66, 2095-2102.
FLORES, R., HERNÁNDEZ, C., DE LA PEÑA, M., VERA, A. & DARÒS, J.A. (2001). Hammerhead ribozyme structure and function in plant RNA replication. Methods in Enzymology, 341, 540-552.
FLORES, R., HERNÁNDEZ, C., MARTÍNEZ DE ALBA, A.E., DARÒS, J.A. & DI
SERIO, F. (2005a). Viroids and viroid-host interactions. Annual Review of Phytopathology, 43, 117-139.
FLORES, R., RANDLES, J.W., OWENS, R.A., BAR-JOSEPH, M. & DIENER, T.O. (2005b). Viroids. In Virus Taxonomy, Classification and Nomenclature. VIIII Report of the International Committee on Taxonomy of Viruses (Fauquet, C.M, Mayo, M.A., Maniloff, J., Desselberger, U. & Ball, A.L. ed.). Elsevier/Academic press, London, 1145-59.
FLORES, R. & SEMANCIK, J.S. (1982). Properties of a cell-free system for synthesis
of citrus exocortis viroid. Proceedings of the National Academy of Sciences of the United States of America, 79, 6285-6288.
FORSTER, A.C., DAVIES, C., SHELDON, C.C., JEFFRIES, A.C. & SYMONS, R.H. (1988). Self-cleaving viroid and newt RNAs may only be active as dimers. Nature, 334, 265-267.
FORSTER, A.C. & SYMONS, R.H. (1987). Self-cleavage of plus and minus RNAs of a
virusoid and a structural model for the active sites. Cell, 49, 211-220. FROHMAN, M.A., DUSH, M.K. & MARTIN G.R. (1988). Rapid production of full-
length cDNAs from rare transcripts: amplification using a single gene-specific oligonucleotide primer. Proceedings of the National Academy of Sciences of the United States of America, 85, 8998-9002.
FU, G.K., WANG, J.T., YANG, J., AU-YOUNG, J. & STUVE, L.L. (2004). Circular
rapid amplification of cDNA ends for high-throughput extension cloning of partial genes. Genomics, 84, 205-210.
FURNEAUX, H., PICK, L. & HURWITZ, J. (1983). Isolation and characterization of
RNA ligase from wheat germ. Proceedings of the National Academy of Sciences of the United States of America, 80, 3933-3937.
FÜTTERER, J. & HOHN, T. (1996). Translation in plants: rules and exceptions. Plant Molecular Biology, 32, 159-189.
159

Bibliografía
GAGO, S., DE LA PEÑA, M. & FLORES, R. (2005). A kissing-loop interaction in a hammerhead viroid RNA critical for its in vitro folding and in vivo viability. RNA, 11, 1173-1183.
GALINDO, J., SMITH, D.R. & DIENER, T.O. (1982). Etiology of planta macho, a viroid disease of tomato. Phytopathology, 72, 49-54.
GAS, M.E., HERNÁNDEZ, C., FLORES, R. & DARÒS, J.A. (2007). Processing of nuclear viroids in vivo: an interplay between RNA conformations. PLoS Pathogens, 3, 1813-1826.
GAS, M.E., MOLINA-SERRANO, D., HERNÁNDEZ, C., FLORES, R. & DARÒS, J.A. (2008). Monomeric linear RNA of citrus exocortis viroid resulting from processing in vivo has 5'-phosphomonoester and 3'-hydroxyl termini: implications for the ribonuclease and RNA ligase involved in replication. Journal of Virology, 82, 10321-10325.
GEGENHEIMER, P., GABIUS, H.J., PEEBLES, C.L. & ABELSON, J. (1983). An RNA ligase from wheat germ which participates in transfer RNA splicing in vitro. Journal of Biological Chemistry, 258, 8365-8373.
GENSCHIK, P., HALL, J. & FILIPOWICZ, W. (1997). Cloning and characterization of the Arabidopsis cyclic phosphodiesterase which hydrolyzes ADP-ribose 1”,2”-cyclic phosphate and nucleoside 2’,3’-cyclic phosphates. Journal of Biological Chemistry, 272, 13211-13219.
GOLDSCHMIDT-CLERMONT, M., CHOQUET, Y., GIRARDBASCOU, J.,
MICHEL, F., SCHIRMERRAHIRE, M. & ROCHAIX, J.D. (1991). A small chloroplast RNA may be required for trans-splicing in Chlamydomonas reinhardtii. Cell, 65, 135-143.
GOMES, I. & GUPTA, R. (1997). RNA splicing ligase activity in the archaeon Haloferax volcanii. Biochemical and Biophysical Research Communications, 237, 588-594.
GONDA, D.K., BACHMAIR, A., WUNNING, I., TOBIAS, J.W., LANE, W.S. & VARSHAVSKY, A. J. (1989) Universality and structure of the N-end rule. Journal of Biological Chemistry, 264, 16700-16712.
GONZALEZ, T.N., SIDRAUSKI, C., DORFLER, S. & WALTER, P. (1999).
Mechanism of non-spliceosomal mRNA splicing in the unfolded protein response pathway. EMBO Journal, 11, 3119-3132.
GRANTHAM, R., GAUTIER, C., GOUY, M., MERCIER, R. & PAVÉ, A. (1980). Codon catalog usage and the genome hipotesis. Nucleic Acids Research, 8, r49-r62.
GRAY, M.W. (1993). Origin and evolution of organelle genomes. Current Opinion in Genetics and Development, 3, 884-890.
GREER, C.L., JAVOR, B. & ABELSON, J. (1983a). RNA ligase in bacteria: formation of a 2',5' linkage by an Escherichia coli extract. Cell, 33, 899-906.
160

Bibliografía
GREER, C.L., PEEBLES, C.L., GEGENHEIMER, P. & ABELSON, J. (1983b). Mechanism of action of a yeast RNA ligase in tRNA splicing. Cell, 32, 537-546.
GREER, C., SOLL, D. & WILLIS, I. (1987). Substrate recognition and identification of splice sites by the tRNA-splicing endonuclease and ligase from Saccharomyces cerevisiae. Molecular and Celular Biology, 7, 76-84.
GRILL, L.K. & SEMANCIK, J.S. (1978). RNA sequences complementary to citrus exocortis viroid in nucleic acid preparations from infected Gynura aurantiaca. Proceedings of the National Academy of Sciences of the United States of America, 75, 896-900.
GROSS, H.J., DOMDEY, H., LOSSOW, C., JANK, P., RABA, M., ALBERTY, H. & SÄNGER, H.L. (1978). Nucleotide sequence and secondary structure of potato spindle tuber viroid. Nature, 273, 203-208.
HAHN, D., BENNOUN, P. & KÜCK, U. (1996). Altered expressión of nuclear genes encoding chloroplast polypeptides in non-photosynthetic mutants of Chlamydomonas reinhardtii: evidence for post-transcriptional regulation. Molecular and General Genetics, 252, 362-370.
HALL, T.C., WEPPRICH, R.K., DAVIES, J.W., WEATHERS, L.G. & SEMANCIK, J.S. (1974). Functional distinctions between ribonucleic acids from citrus exocortis viroid and plant viruses: cell-free translation and aminoacylation reactions. Virology, 61, 486-492.
HAMPEL, A. & TRITZ, R. (1989). RNA catalytic properties of the minimum (-)sTRSV sequence. Biochemistry, 28, 4929-4933.
HARDERS, J., LUKACS, N., ROBERT-NICOUD, M., JOVIN, T.M. & RIESNER, D. (1989). Imaging of viroids in nuclei from tomato leaf tissue by in situ hybridization and confocal laser scanning microscopy. EMBO Journal, 8, 3941-3949.
HARDING, H.P., LACKEY, J.G., HSU, H-C., ZHANG, Y., DENG, J., XU, R.M., DAMHA, M.J. & RON, D. (2008) An intact unfolded protein response in Trpt1 knockout mice reveals phylogenic divergence in pathways for RNA ligation. RNA, 14, 225-232.
HARI, V. (1980). Ultrastructure of potato spindle tuber viroid-infected tomato leaf
tissue. Phytopathology, 70, 385-387. HARRIS, E.H. (1989). The Chlamydomonas Sourcebook: a Comprehensive Guide to
Biology and Laboratory Use. Academic Press, San Diego. HARRIS, E.H. (2001). Chlamydomonas as a model organism. Annual Review of Plant
Physiology and Plant Molecular Biology, 52, 363-406.
HARRIS, P.S. & BROWNING, I.A. (1980). The effects of temperature and light on the symptom expression and viroid concentration in tomato of a severe strain of potato spindle tuber viroid. Potato Research, 23, 85-93.
161

Bibliografía
HASELOFF, J., MOHAMED N.A. & SYMONS, R.H. (1982). Viroid RNAs of cadang-cadang disease of coconuts. Nature, 229, 316-321.
HEIFETZ, P.B. (2000). Genetic engineering of the chloroplast. Biochimie, 82, 655-666.
HERNÁNDEZ, C. & FLORES, R. (1992). Plus and minus RNAs of peach latent mosaic viroid self-cleave in vitro via hammerhead structure. Proceedings of the National Academy of Sciences of the United States of America, 89, 3711-3715.
HERNÁNDEZ, C., ELENA, S.F., MOYA, A. & FLORES, R. (1992). Pear blister canker viroid is a member of the apple scar skin subgroup (apscaviroids) and has also sequence homologies with viroids from other subgroups. Journal of General Virology, 73, 2503-2507.
HERR, A.J., JENSEN, M.B., DALMAY, T. & BAULCOMBE, D.C. (2005). RNA
polymerase IV directs silencing of endogenous DNA. Science, 308, 118-120.
HERRIN, D.L. & NICKELSEN, J. (2004). Chloroplast RNA processing and stability. Photosynthesis Research, 82, 301-314.
HERTEL, K.J., HERSCHLAG, D. & UHLENBECK, O.C., (1994) A kinetic and thermodynamic framework for the hammerhead ribozyme reaction. Biochemistry, 33, 3374-3385.
HINEGARDNER, R.T. & ENGELBERG, J. (1963). Rationale for a universal genetic
code. Science, 142, 1083-1085.
HO, C.K. & SHUMAN, S. (2002). Bacteriophage T4 RNA ligase 2 (gp24.1) exemplifies a family of RNA ligases found in all phylogenetic domains. Proceedings of the National Academy of Sciences of the United States of America, 99, 12709-12714.
HO, C.K., WANG, L.K., LIMA, C.D. & SHUMAN, S. (2004). Structure and mechanism of RNA ligase. Structure, 12, 327-339.
HOFMANN, A., ZDANOV, A., GENSCHIK, P., RUVINOV, S., FILIPOWICZ, W. & WLODAWER, A. (2000). Structure and mechanism of activity of the cyclic phosphodiesterase of Appr>p, a product of the tRNA splicing reaction. EMBO Journal, 19, 6207-6217.
HOPPER, A.K. & PHIZICKY, E.M. (2003). tRNA transfers to the limelight. Genes and Development, 17, 162-180.
HU, Q.D., LU, H., HUO, K., YING, K., LI, J., XIE, Y., MAO, Y. & LI, Y.Y. (2003). A human homolog of the yeast gene encoding tRNA 2’-phosphotransferase: cloning, characterization and complementation analysis. Cellular and Moleular Life Science, 60, 1725-1732.
HUANG, J-C. & CHEN, F. (2006). Simultaneous amplification of 5’ and 3’ cDNA ends based on template-switching effect and inverse PCR. Biotechniques, 40, 187-189.
162

Bibliografía
HUTCHINS, C.J., KEESE, P., VISVADER, J.E., RATHJEN, P.D., MCINNES, J.L. & SYMONS, R.H. (1985). Comparison of multimeric plus and minus forms of viroids and virusoids. Plant Molecular Biology, 4, 293-304.
HUTCHINS, C.J., RATHJEN, P.D., FORSTER, A.C. & SYMONS, R.H. (1986). Self-cleavage of plus and minus RNA transcripts of avocado sunblotch viroid. Nucleic Acids Research, 14, 3627-3640.
ISHIKAWA, M., MESHI, T., OHNO, T., OKADA, Y., SANO, T., UEDA, I. & SHIKATA, E. (1984). A revised replication cycle for viroids: the role of longer than unit length RNA in viroid replication. Molecular and General Genetics, 196, 421-428.
KANE, J.F. (1995). Effects of rare codon clusters on high-level expression of heterologous proteins in Escherichia coli. Current Opinion in Biotechnology, 6, 494-500.
KANNO, T., HUETTEL, B., METTE, M.F., AUFSATZ, W., JALIGOT, E.,
DAXINGER, L., KREIL, D.P., MATZKE, M. & MATZKE, A.J.M. (2005). Atypical RNA polymerase subunits required for RNA-directed DNA methylation. Nature Genetics, 37, 761-765.
KATO, M., SHIROUZU, M., TERADA, T., YAMAGUCHI, H., MURAYAMA, K.,
SAKAI, H., KURAMITSU, S. & YOKOYAMA, S. (2003). Crystal structure of the 2'-5' RNA ligase from Thermus thermophilus HB8. Journal of Molecular Biology, 329, 903-911.
KAUFMANN, G. (2000). Anticodon nucleases. Trends in Biochemical Sciences, 25, 70-74.
KEESE, P. & SYMONS, R.H. (1985). Domains in viroids: evidence of intermolecular RNA rearrangements and their contribution to viroid evolution. Proceedings of the National Academy of Sciences of the United States of America, 82, 4582-4586.
KEPPETIPOLA, N., NANDAKUMAR J. & SHUMAN, S. (2007). Reprogramming the tRNA-splicing activity of a bacterial RNA repair enzyme. Nucleic Acids Research, 35, 3624-3630.
KEPPETIPOLA, N. & SHUMAN, S. (2007). Characterization of the 2',3' cyclic phosphodiesterase activities of Clostridium thermocellum polynucleotide kinase-phosphatase and bacteriophage λ phosphatase. Nucleic Acids Research, 35, 7721-7732.
KHVOROVA, A., LESCOUTE, A., WESTHOF, E. & JAYASENA, S.D. (2003). Sequence elements outside the hammerhead ribozyme catalytic core enable intracellular activity. Nature Structural and Molecular Biology, 10, 708-712.
KIBERSTIS, P.A., HASELOFF, J. & ZIMMERN, D. (1985). 2' Phosphomonoester, 3'-5' phosphodiester bond at a unique site in a circular viral RNA. EMBO Journal, 4, 817-827.
163

Bibliografía
KIEFER, M.C., OWENS, R.A. & DIENER. T.O. (1983). Structural similarities between viroids and transposable genetic elements. Proceedings of the National Academy of Sciences of the United States of America, 80, 6234-6238.
KIKUCHI, Y., TYC, K., FILIPOWICZ, W., SÄNGER, H.L. & GROSS, H.J. (1982). Circularization of linear viroid RNA via 2'-phosphomonoester, 3',5'-phosphodiester bonds by a novel type of RNA ligase from wheat germ and Chlamydomonas. Nucleic Acids Research, 10, 7521-7529.
KINDLE, K.L. (1990). High-frequency nuclear transformation of Chlamydomonas reinhardtii. Proceedings of the National Academy of Sciences of the United States of America, 87, 1228-1232.
KJEMS, J. & GARRETT, R.A. (1988). Novel splicing mechanism for the ribosomal RNA intron in the archaebacterium Desulfurococcus mobilis. Cell, 54, 693-703.
KOLONKO, N., BANNACH, O., ASCHERMANN, K., HU, K.H., MOORS, M., SCHMITZ, M., STEGER, G. & RIESNER, D. (2006). Transcription of potato spindle tuber viroid by RNA polymerase II starts in the left terminal loop. Virology, 347, 392–404.
KONARSKA, M., FILIPOWICZ, W., DOMDEY, H. & GROSS, H.J. (1981).
Formation of a 2’-phosphomonoester, 3’,5’-phosphodiester linkage by a novel RNA ligase in wheat germ. Nature, 293, 112-116.
KONARSKA, M., FILIPOWICZ, W. & GROSS, H.J. (1982). RNA ligation via 2’-
phosphomonoester, 3’,5’-phosphodiester linkage: requirement of 2’.3’-cyclic phosphate termini and involvement of a 5’-hydroxyl polynucleotide kinase. Proceedings of the National Academy of Sciences of the United States of America, 79, 1474-1478.
KOWALSKI, D., KROEKER, W.D. & LASKOWSKI, M. (1976). Mung bean nuclease
I. Physical, chemical, and catalytic properties. Biochemistry, 15, 4457-4463.
KOZAK, M. (1986). Point mutations define a sequence flanking the AUG initiator codon that modulates translation by eukaryotic ribosomes. Cell, 44, 283-292.
KOZAK, M. (1990). Downstream secondary structure facilitates recognition of initiator
codons by eukaryotic ribosomes. Proceedings of the National Academy of Sciences of the United States of America, 87, 8301-8305.
KROEKER, W.D., KOWALSKI, D. & LASKOWSKI, M. (1976). Mung bean nuclease
I. Terminally directed hydrolysis of native DNA. Biochemistry, 15, 4463-4467.
KUHN, H. & FRANK-KAMENETSKII, M.D. (2005). Template-independent ligation of single-stranded DNA by T4 DNA ligase. FEBS Journal, 272, 5991-6000.
KUMAR, S., TAMURA, K. & NEI, M. (2004). MEGA 3: integrated software for molecular evolutionary genetics analysis and sequence alignment. Briefings in Bioinformatics, 5, 150-163.
164

Bibliografía
KURLAND, C. & GALLANT, J. (1996). Errors of heterologous protein expression. Current Opinion in Biotechnology, 7, 489-493.
KUROIWA, T. (1991). The replication, differentiation and inheritance of plastids with emphasis on the concept of organelle nuclei. International Review of Cytology, 128, 1-62.
LAEMMLI, U.K. (1970). Cleavage of structural proteins during assembly of head of bacteriophage T4. Nature, 227, 680-685.
LAPPE-SIEFKE, C., GOEBBELS, S, GRAVEL, M., NICKSCH, E., LEE, J., BRAUN, P.E., GRIFFITHS, I.R. & NAVE, C.A. (2003). Disruption of Cnp1 uncouples oligodendroglial functions in axonal support and myelination. Nature Genetics, 33, 366-374.
LAWSON, R.H. & HEARON, S.S. (1971). Ultrastructure of chrysanthemum stunt
virus-infected and stunt-free mistletoe Chrysanthemum. Phytopathology, 6, 653-656.
LAZINSKI, D.W. & TAYLOR, J.M (1995). Intracellular cleavage and ligation of hepatitis delta virus genomic RNA: regulation of ribozyme activity by cis-acting sequences and host factors. Journal of Virology, 69, 1190-1200.
LEHMANN, E., BRUECKNER, F. & CRAMER, P (2007). Molecular basis of RNA-
dependent RNA polymerase II activity. Nature, 450, 445-449. LI, H., TROTTA, C.R. & ABELSON, J. (1998). Crystal structure and evolution of a
transfer RNA splicing enzyme. Science, 280, 279-284.
LIERE, K. & MALIGA, P. (1999). In vitro characterization of the tobacco rpoB promoter reveals a core sequence motif conserved between phage-type plastid and plant mitochondrial promoters. EMBO Journal, 18, 249-257.
LIMA, M.I., FONSECA, M.E.N., FLORES, R. & KITAJIMA, E.W. (1994). Detection of avocado sunblotch viroid in chloroplasts of avocado leaves by in situ hybridization. Archives of Virology, 138, 385-390.
LIU, X. & GOROVSKY, M.A. (1993). Mapping the 5' and 3' ends of Tetrahymena thermophila mRNAs using RNA ligase mediated amplification of cDNA ends (RLM-RACE). Nucleic Acids Research, 21, 4954-4960.
LU, J., HUANG, B., ESBERG, A., JOHANSSON, M.J.O. & BYSTROM, A.S. (2005). The Kluyveromyces lactis gamma toxin targets tRNA anticodons. RNA, 11, 1648-1654.
LUKENS, J.H.N, MATHEWS, D.E. & DURBIN, R.D. (1987). Effect of tagetitoxin on the levels of ribulose 1,5-bisphosphate carboxylase, ribosomes, and RNA in plastids of wheat leaves. Plant Physiology, 84, 808-813.
LÜTCKE, H.A., CHOW, K.C., MICKEL, F.S., MOSS, K.A., KERN, H.F. & SCHEELE, G.A. (1987). Selection of AUG initiation codons differs in plants and animals. EMBO Journal, 6, 43-48.
165

Bibliografía
MAHEN, E.M., HARGER, J.W., CALDERON, E.M. & FEDOR, M.J. (2005). Kinetics and thermodynamics make different contributions to RNA folding in vitro and in yeast. Molecular Cell, 19, 27-37.
MAKINO, S., SAWASAKI, T., TOZAWA, Y., ENDO, Y. & TAKAI, K. (2006).
Covalent circularization of exogenous RNA during incubation with a wheat embryo cell extract. Biochemical and Biophysical Research Communications, 347, 1080-1087.
MALFITANO, M., DI SERIO, F., COVELLI, L., RAGOZZINO, A., HERNÁNDEZ, C. & FLORES, R. (2003). Peach latent mosaic viroid variants inducing peach calico contain a characteristic insertion that is responsible for this symptomatology. Virology, 313, 492-501.
MALIGA, P. (2004). Plastid transformation in higher plants. Annual Review of Plant
Biology, 55, 289-313.
MARCOS, J.F. & FLORES, R. (1992). Characterization of RNAs specific to avocado sunblotch viroid synthesized in vitro by a cell-free system from infected avocado leaves. Virology, 186, 481-488.
MARTICK, M. & SCOTT, W.G. (2006). Tertiary contacts distant from the active site prime a ribozyme for catalisis. Cell, 126, 309-320.
MARTIN, W. & HERRMANN, R.G. (1998). Gene transfer from organelles to the nucleus: how much, what happens, and why? Plant Physiology, 118, 9-17.
MARTINS, A. & SHUMAN, S. (2004a). Characterization of a baculovirus enzyme
with RNA ligase, polynucleotide 5'-kinase, and polynucleotide 3'-phosphatase activities. Journal of Biological Chemistry, 279, 18220-18231.
MARTINS, A. & SHUMAN, S. (2004b). An RNA ligase from Deinococcus radiodurans. Journal of Biological Chemistry, 279, 50654-50661.
MARTINS, A. & SHUMAN, S. (2005). An end-healing enzyme from Clostridium thermocellum with 5' kinase, 2',3' phosphatase, and adenylyltransferase activities. RNA, 11, 1271-1280.
MARZLUFF, W.F. & HUANG, R.C. (1984). Transcription of RNA in isolated nuclei. In Transcription and Translation: A Practical Approach (Hames, B.D. & Higgins, S.J., ed.). IRL Press, Oxford & Washington DC, 89-129.
MATHEWS, D.E. & DURBIN, R.D. (1990). Tagetitoxin inhibits RNA synthesis directed by RNA polymerases from chloroplasts and Escherichia coli. Journal of Biological Chemistry, 265, 493-498.
MAUL, J.E., LILLY, J.W., CUI, L., DEPAMPHILIS, C.W., MILLER, W., HARRIS, E.H. & STERN, D.B. (2002). The Chlamydomonas reinhardtii plastid chromosome: islands of genes in a sea of repeats. Plant Cell, 14, 2659-2679.
166

Bibliografía
MAYFIELD, S.P., FRANKLIN, S.E. & LERNER, R.A. (2003). Expression and assembly of a fully active antibody in algae. Proceedings of the National Academy of Sciences of the United States of America, 100, 438-442.
MCCOY, M.I.M. & GUMPORT, R.I. (1980). T4 ribonucleic acid ligase joins single-strand oligo (deoxyribonucleotides). Biochemistry, 19, 635-642.
MCCRAITH, S.M. & PHIZICKY, E.M. (1991). An enzyme from Saccharomyces cerevisiae uses NAD+ to transfer the splice junction 2'-phosphate from ligated tRNA to an acceptor molecule. Journal of Biological Chemistry, 266, 11986-11992.
MCMANUS, M.T., SHIMAMURA, M., GRAMS, J. & HAJDUK, S.L. (2001). Identification of candidate mitochondrial RNA editing ligases from Trypanosoma brucei. RNA, 7, 167-175.
MICHAELIS, G., VAHRENHOLZ, C. & PRATJE, E. (1990). Mitochondrial DNA of Chlamydomonas reinhardtii: the gene for apocytochrome-B and the complete functional map of the 15.8 Kb DNA. Molecular and General Genetics, 233, 211-216.
MISHRA, M.D., HAMMOND, R.W., OWENS, R.A., SMITH, D.R., & DIENER, T.O. (1991). Indian bunchy top disease of tomato plants is caused by a distinct strain of citrus exocortis viroid. Journal of General Virology, 72, 1781-1785.
MOGK, A., SCHMIDT, R. & BUKAU, B. (2007). The N-end rule pathway for regulated proteolysis: prokaryotic and eukaryotic strategies. Trends in Cell Biology, 17, 165-172.
MOHAMED, N.A. & THOMAS, W. (1980). Viroid-like properties of an RNA species
associated with the sunblotch disease of avocados. Journal of General Virology, 46, 157-167.
MÜHLBACH, H.P. & SÄNGER, H.L. (1979). Viroid replication is inhibited by alpha-amanitin. Nature, 278, 185-188.
NANDAKUMAR, J., HO, C.K., LIMA, C.D. & SHUMAN, S. (2004). RNA substrate specificity and structure-guided mutational analysis of bacteriophage T4 RNA ligase 2. Journal of Biological Chemistry, 279, 31337-31347.
NANDAKUMAR, J. & SHUMAN, S. (2004). How an RNA ligase discriminates RNA versus DNA damage. Molecular Cell, 16, 211-221.
NANDAKUMAR, J., SHUMAN, S. & LIMA, C.D. (2006). RNA ligase structures reveal the basis for RNA specificity and conformational changes that drive ligation forward. Cell, 127, 71-84.
NASR, F. & FILIPOWCZ, W. (2000). Characterization of the Saccharomyces cerevisiae cyclic nucleotide phosphodiesterase involved in the metbolism of ADP-ribose 1”,2”-cyclic phosphate. Nucleic Acids Research, 28, 1676-1683.
167

Bibliografía
NAVARRO, B. & FLORES, R. (1997). Chrysanthemum chlorotic mottle viroid: unusual structural properties of a subgroup of self-cleaving viroids with hammerhead ribozymes. Proceedings of the National Academy of Sciences of the United States of America, 94, 11262-11267.
NAVARRO, J.A., DARÒS, J.A. & FLORES, R. (1999). Complexes containing both polarity strands of avocado sunblotch viroid: identification in chloroplasts and characterization. Virology, 253, 77-85.
NAVARRO, J.A. & FLORES, R. (2000). Characterization of the initiation sites of both
polarity strands of a viroid RNA reveals a motif conserved in sequence and structure. EMBO Journal, 19, 2662-2670.
NAVARRO, J.A., VERA, A. & FLORES, R. (2000). A chloroplastic RNA polymerase resistant to tagetitoxin is involved in replication of avocado sunblotch viroid. Virology, 268, 218-225.
NELSON, J.A., SHEPOTINOVSKAYA, I. & UHLENBECK, O.C. (2005). Hammerheads derived from sTRSV show enhanced cleavage and ligation rate constants. Biochemistry, 44, 14577-14585
NELSON, J.A. & UHLENBECK, O.C. (2006). When to believe what you see.
Molecular Cell, 23, 447-450.
NEUHOFF, V. AROLD, N., TAUBE, D. & EHRHARDT, W. (1988). Improved staining of proteins in polyacrylamide gels including isoelectric focusing gels with clear background at nanogram sensitivity using Coomassie Brilliant Blue G-250 and R-250. Electrophoresis, 9, 255-262.
NICKELSEN, J. & KÜCK, U. (2000). The unicellular green alga Chlamydomonas reinhardtii as an experimental system to study chloroplast RNA metabolism. Naturwissenschaften, 87, 97-107.
NISHIKURA, K. & DEROBERTIS, E.M. (1981). RNA processing in micro-injected Xenopus oocytes: sequential addition of base modifications in a spliced transfer RNA. Journal of Molecular Biology, 145, 405-420.
NOVOGRODSKY, A. & HURWITZ, J. (1966a). The enzymatic phosphorylation of ribonucleic acid and deoxyribonucleic acid I. Phosphorylation at 5’-hydroxyl termini. Journal of Biological Chemistry, 241, 2923-2932.
NOVOGRODSKY, A. & HURWITZ, J. (1966b). The enzymatic phosphorylation of ribonucleic acid and deoxyribonucleic acid II. Further properties of the 5’-hydroxyl polynucleotide kinase. Journal of Biological Chemistry, 241, 2933-2943.
ODELL, M., SRISKANDA, V., SHUMAN, S. & NIKOLOV, D.B. (2000). Crystal structure of eukaryotic DNA ligase-adenylate illuminates the mechanism of nick sensing and strand joining. Molecular Cell, 6, 1183-1193.
OKAYAMA, H. & BERG, P. (1982). High-efficiency cloning of full-length cDNA. Molecular and Cellular Biology, 2, 161-170.
168

Bibliografía
ONODERA, Y., HAAG, J.R., REAM, T., NUNES, P.C., PONTES, O. & PIKAARD, C.S. (2005). Plant nuclear RNA polymerase IV mediates siRNA and DNA methylation-dependent heterochromatin formation. Cell, 120, 613-622.
PALMER, J.D. (1985). Comparative organization of chloroplast genomes. Annual Review of Genetics, 19, 325-354.
PAUSHKIN, S.V., PATEL, M., FURIA, B.S., PELTZ, S.W. & TROTTA, C.R. (2004). Identification of a human endonuclease complex reveals a link between tRNA splicing and pre-mRNA 3 ' end formation. Cell, 117, 311-321.
PEEBLES, C.L., GEGENHEIMER, P. & ABELSON, J. (1983). Precise excision of intervening sequences from precursor tRNAs by a membrane-associated yeast endonuclease. Cell, 32, 525-536.
PELCHAT, M., LEVESQUE, D., OUELLET, J., LAURENDEAU, S., LEVESQUE, S.,
LEHOUX, J., THOMPSON, D.A., EASTWELL, K.C., SKRZECZKOWSKI, L.J. & PERREAULT, J.P. (2000). Sequencing of peach latent mosaic viroid variants from nine North American peach cultivars shows that this RNA folds into a complex secondary structure. Virology, 271, 37-45.
PERKINS, K.K., FURNEAUX, H. & HURWITZ, J. (1985). Isolation and
characterization of an RNA ligase from HeLa cells. Proceedings of the National Academy of Sciences of the United States of America, 82, 684-688.
PHIZICKY, E.M., CONSAUL, S.A., NEHRKE, K.W. & ABELSON, J. (1992). Yeast
tRNA ligase mutants are non viable and accumulate tRNA splicing intermediates. Journal of Biological Chemistry, 267, 4577-4582.
PHIZICKY, E.M., SCHWARTZ, R.C. & ABELSON, J. (1986). Saccharomyces cerevisiae tRNA ligase: purification of the protein and isolation of the structural gene. Journal of Biological Chemistry, 261, 2978-2986.
PICK, L., FURNEAUX, H. & HURWITZ, J. (1986). Purification of wheat germ RNA ligase II: mechanism of action of wheat germ RNA ligase. Journal of Biological Chemistry, 261, 6694-6704.
PICK, L. & HURWITZ, J. (1986). Purification of wheat germ RNA ligase I: characterization of a ligase-associated 5'-hydroxyl polynucleotide kinase activity. Journal of Biological Chemistry, 261, 6684-6693.
POIRIER, Y., NAWRATH, C. & SOMERVILLE, C. (1995). Production of polyhydroxyalkanoates, a family of biodegradable plastics and elastomers, in bacteria and plants. Biotechnology, 13, 142-150.
POLIDOROS, A.N., PASENTSIS, K. & TSAFTARIS, A.S. (2006). Rolling circle amplification-RACE: a method for simultaneous isolation of 5’ and 3’ cDNA ends from amplified cDNA templates. Biotechniques, 41, 35-39.
169

Bibliografía
PORATH, J., CARLSSON, J., OLSSON, I. & BELFRAGE, G. (1975). Metal chelate affinity chromatography, a new approach to protein fractionation. Nature, 258, 598-599.
PRODY, G.A., BAKOS, J.T., BUZAYAN, J.M., SCHNEIDER, I.R. & BRUENING, G. (1986). Autolytic processing of dimeric plant virus satellite RNA. Science, 231, 1577-1580.
QI, Y.J. & DING, B. (2003). Differential subnuclear localization of RNA strands of opposite polarity derived from an autonomously replicating viroid. Plant Cell, 15, 2566-2577.
RACE, H., HERRMANN, R.G. & MARTIN, W. (1999). Why have organelles retained genomes? Trends in Genetics, 15, 364–370.
RANDOLPH-ANDERSON, B.L., BOYNTON, J.E., GILLHAM, N.W., HARRIS, E.H.,
JOHNSON, A.M., DORTHU, M.P. & MATAGNE, R.F. (1993). Further characterization of the respiratory deficient dum-1 mutation of Chlamydomonas reinhardtii and its use as a recipient for mitochondrial transformation. Molecular and General Genetics, 236, 235-244.
RAYMOND, A. & SHUMAN, S. (2007). Deinococcus radiodurans RNA ligase exemplifies a novel ligase clade with a distinctive N-terminal module that is important for 5’-PO4 nick sealing and ligase adenylylation but dispensable for phosphodiester formation at an adenylylated nick. Nucleic Acids Research, 35, 839-849.
REID, C.E. & LAZINSKI, D.W. (2000). A host-specific function is required for
ligation of a wide variety of ribozyme-processed RNAs. Proceedings of the National Academy of Sciences of the United States of America, 97, 424-429.
RICHARDSON, C.C. (1965). Phosphorylation of nucleic acid by an enzyme from T4
bacteriophage-infected Escherichia coli. Proceedings of the National Academy of Sciences of the United States of America, 54, 158-165.
ROCHAIX, J.D. (1995). Chlamydomonas reinhardtii as a photosintetic yeast. Annual review of genetics, 29, 209-230.
ROCHAIX, J.D. (1996). Post-transcriptional regulation of chloroplast gene expression in Chlamydomonas reinhardtii. Plant Molecular Biology, 32, 327-341.
RODIO, M.E., DELGADO, S., DE STRADIS, A., GÓMEZ, M.D., FLORES, R. & DI SERIO, F. (2007). A viroid RNA with a specific structural motif inhibits chloroplast development. Plant Cell, 19, 3610-3626.
RODIO, M.E., DELGADO, S., FLORES, R. & DI SERIO, F. (2006). Variants of Peach
latent mosaic viroid inducing peach calico: uneven distribution in infected plants and requirements of the insertion containing the pathogenicity determinant Journal of General Virology, 87, 231-240
170

Bibliografía
ROEDER, R.G. (1976). Eukaryotic nuclear RNA polymerases. In RNA Polymerases (Losick, R. & Chamberlin, M. ed.). Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 285-329.
RON, D. & WALTER, P. (2007). Signal integration in the endoplasmic reticulum unfolded protein response. Nature Reviews Molecular Cell Biology, 8, 519-529.
ROSENSTEIN, S.P & BEEN, M.D. (1990). Self-cleavage of hepatitis delta virus genomic strand RNA is enhanced under partially denaturing conditions. Biochemistry, 29, 8011-8016.
RUNNELS, J., SOLTIS, D., HEY, T. & SNYDER, L. (1982). Genetic and
physiological studies of the role of RNA ligase of bacteriofage T4. Journal of Molecular Biology, 154, 273-286.
SABATINI, R. & HAJDUK, S.L. (1995). RNA ligase and its involvement in guide RNA/mRNA chimera formation: evidence for a cleavage-ligation mechanism of Trypanosoma brucei messenger RNA editing. Journal of Biological Chemistry, 270, 7233-7240.
SAKAMOTO, W., STURM, N.R., KINDLE, K.L. & STERN, D.B. (1994). petD mRNA maturation in Chlamydomonas reinhardtii chloroplasts: role of 5' endonucleolytic processing. Molecular Cell Biology, 14, 6180-6186.
SALGIA, S. R., SINGH, S. K., GURHA, P. & GUPTA, R. (2003). Two reactions of Haloferax volcanii RNA splicing enzymes: joining of exons and circularization of introns. RNA, 9, 319-330.
SALVADOR, M.L. & KLEIN, U. (1999). The redox state regulates RNA degradation in the chloroplast of Chlamydomonas reinhardtii. Plant Physiology, 121, 1367-1374.
SALVADOR, M.L., KLEIN, U. & BOGORAD, L. (1993a). Light-regulated and endogenous fluctuations of chloroplast transcript levels in Chlamydomonas: regulation by transcription and RNA degradation. Plant Journal, 3, 213-219.
SALVADOR, M.L., KLEIN, U. & BOGORAD, L. (1993b). 5' sequences are important positive and negative determinants of the longevity of Chlamydomonas chloroplast gene transcripts. Proceedings of the National Academy of Sciences of the United States of America, 90, 1556-1560.
SAMBROOK, J. & RUSSELL, D.W. (2001). Molecular Cloning: A Laboratory Manual. 3ª ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
SÄNGER, H. L. (1972) An infectious and replicating RNA of low molecular weight: the agent of the exocortis disease of citrus. Advances in Biosciences, 8, 103-106.
SÄNGER, H.L., KLOTZ, G., RIESNER, D., GROSS, H.J. & KLEINSCHMIDT, A.K. (1976). Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proceedings of the National Academy of Sciences of the United States of America, 73, 3852-3856.
171

Bibliografía
SÄNGER, H.L. & RAMM, K. (1975). Radioactive labelling of viroid RNA. In Modification of the Information Content of Plant Cells (Markham, R., Davies, D. R., Hopwood, D. A., and Horne, R. W., ed.). North Holland, Amsterdam, 229-252.
SANO, T., CANDRESSE, T., HAMMOND, R.W., DIENER, T.O. & OWENS, R.A.
(1992). Identification of multiple structural domains regulating viroid pathogenicity. Proceedings of the National Academy of Sciences of the United States of America, 89, 10104-10108.
SAWAYA, R., SCHWER, B. & SHUMAN, S. (2003). Genetic and biochemical analysis of the functional domains of yeast tRNA ligase. Journal of Biological Chemistry, 278, 43928-43938.
SAWAYA, R., SCHWER, B. & SHUMAN, S. (2005). Structure-function analysis of the yeast NAD+-dependent tRNA 2'-phosphotransferase Tpt1. RNA, 11, 107-113.
SCHAEFER, B.C. (1995). Revolutions in rapid amplification of cDNA ends: new strategies for polymerase chain reaction cloning of full-lengh cDNA ends. Analytical Biochemistry, 227, 255-273.
SCHAFFER, A.A., ARAVIND, L., MADDEN, T.L., SHAVIRIN, S., SPOUGE, J.L., WOLF, Y.I., KOONIN, E.V. & ALTSCHUL, S.F. (2001). Improving the accuracy of PSI-BLAST protein database searches with composition-based statistics and other refinements. Nucleic Acids Research, 29, 2994-3005.
SCHIEBEL, W., PELISSIER, T., RIEDEL, L., THALMEIR, S., SCHIEBEL, R., KEMPE, D., LOTTSPEICH, F., SÄNGER, H.L. & WASSENEGGER, M. (1998). Isolation of an RNA-directed RNA polymerase-specific cDNA clone from tomato. Plant Cell, 10, 2087-2101.
SCHINDLER, I.M. & MÜHLBACH, H.P. (1992). Involvement of nuclear DNA-dependent RNA-polymerases in potato spindle tuber viroid replication: a reevaluation. Plant Science, 84, 221-229.
SCHMIDT, W.M. & MUELLER, M.W. (1999). CapSelect: a highly sensitive method for 5’ CAP-dependent enrichment of full-length cDNA in PCR-mediated analysis of mRNAs. Nucleic Acids Research, 27, e31.
SCHNAUFER, A., PANIGRAHI, A.K., PANICUCCI, B., IGO, P.P., WIRTZ, E., SALAVATI, R. & STUART, K. (2001). An RNA ligase essential for RNA editing and survival of the bloodstream form of Trypanosoma brucei. Science, 291, 2159-2162.
SCHRAMM, G., BRUCHHAUS, I. & ROEDER, T. (2000). A simple and reliable 5’-RACE approach. Nucleic Acids Research, 28, e96.
SCHUMACHER, J., RANDLES, J.W. & RIESNER, D. (1983). A two-dimensional electrophoretic technique for the detection of circular viroids and virusoids. Analytical Biochemistry, 135, 288-295.
172

Bibliografía
SCHWARTZ, R.C., GREER, C.L., GEGENHEIMER, P. & ABELSON, J. (1983). Enzymatic mechanism of an RNA ligase from wheat germ. Journal of Biological Chemistry, 258, 8374-8383.
SCHWER, B., ARONOVA, A., RAMIREZ, A., BRAUN, P. & SHUMAN, S. (2008). Mammalian 2',3' cyclic nucleotide phosphodiesterase (CNP) can function as a tRNA splicing enzyme in vivo. RNA, 14, 204 - 210.
SCHWER, B., SAWAYA, R., HO, C.K. & SHUMAN, S. (2004). Portability and fidelity of RNA-repair systems. Proceedings of the National Academy of Sciences of the United States of America, 101, 2788-2793.
SEMANCIK, J.S., CONEJERO, V. & GERHART, J. (1977). Citrus exocortis viroid: survey of protein synthesis in Xenopus laevis oocytes following addition of viroid RNA. Virology, 80, 218-221.
SEMANCIK, J.S., SZYCHOWSKI, J.A., RAKOWSKI, A.G. & SYMONS, R.H. (1994). A stable 463 nucleotide variant of citrus exocortis viroid produced by terminal repeats. Journal of General Virology, 75, 727-732.
SEMANCIK, J.S., TSURUDA, D., ZANER, L., GEELEN, J.L. & WEATHERS, J.G.
(1976). Exocortis disease: subcellular distribution of pathogenic (viroid) RNA. Virology, 69, 669-676.
SEMANCIK, J.S. & VANDERWOUDE, W.J. (1976). Exocortis viroid: cytopathic effects at plasma membrane in association with pathogenic RNA. Virology, 69, 719-726.
SHARMEEN, L., KUO, M.Y.P. & TAYLOR, J. (1989). Self-ligating RNA sequences on the antigenome of human hepatitis delta virus. Journal of Virology, 63, 1428-1430.
SHIMOGAWARA, K., FUJIWARA, S., GROSSMAN, A. & USUDA, H. (1998). High-efficiency transformation of Chlamydomonas reinhardtii by electroporation. Genetics. 148, 1821-1828.
SHUMAN, S. & LIMA, C.D. (2004). The polynucleotide ligase and RNA capping enzyme superfamily of covalent nucleotidyltransferases. Current Opinion in Structural Biology, 14, 757-764.
SIDRAUSKI, C., COX, J.S. & WALTER, P. (1996). tRNA ligase is required for regulated mRNA splicing in the unfolded protein response. Cell, 87, 405-413.
SILBER, R., MALATHI, V.G. & HURWITZ, J. (1972). Purification and properties of bacteriophage T4 induced RNA ligase. Proceedings of the National Academy of Sciences of the United States of America, 69, 3009-3013.
SILBERKLANG, M., GILLUM, A.M. & RAJBHANDARY, U.L. (1979). Use of in vitro 32P labelling in the sequence analysis of nonradioactive tRNAs. Methods in Enzymology, 59, 58-109.
173

Bibliografía
SNOPEK, T.J., SUGINO, A., AGARWAL, K.L. & COZZARELLI, N.R. (1976). Catalysis of DNA joining by bacteriophage T4 RNA ligase. Biochemical and Biophysical Research Communications, 68, 417-424.
SNOPEK, T.J., WOOD, W.B., CONLEY, M.P., CHEN, P. & COZZARELLI, N.R. (1977). Bacteriophage T4 RNA ligase is gene 63 product, protein that promotes tail fiber attachment to baseplate. Proceedings of the National Academy of Sciences of the United States of America, 74, 3355-3359.
SOGO, J.M., KOLLER, T. & DIENER, T.O. (1973). Potato spindle tuber viroid X: visualization and size determination by electron microscopy. Virology, 55, 70-80.
SPIESMACHER, E., MÜHLBACH, H.P., SCHNOLZER, M., HAAS, B. & SÄNGER, H.L. (1983). Oligomeric forms of potato spindle tuber viroid (PSTV) and of its complementary RNA are present in nuclei isolated from viroid-infected potato cells. Bioscience Reports, 3, 767-774.
SPINELLI, S.L., CONSAUL, S.A. & PHIZICKY, E.M. (1997). A conditional lethal yeast phosphotransferase (tpt1) mutant accumulates tRNAs with a 2'-phosphate and an undermodified base at the splice junction. RNA, 3, 1388-1400.
SPINELLI, S.L., MALIK, H.S., CONSAUL, S.A. & PHIZICKY, E.M. (1998). A functional homolog of a yeast tRNA splicing enzyme is conserved in higher eukaryotes and in Escherichia coli. Proceedings of the National Academy of Sciences of the United States of America, 95, 14136-14141.
STEINBERG, T.H., MATHEWS, D.E., DURBIN, R.D. & BURGESS, R.R. (1990). Tagetitoxin: a new inhibitor of eukaryotic transcription by RNA polymerase III. Journal of Biological Chemistry, 265, 499-505.
STERN, D.B. & DRAGER, R.G. (1998). Chloroplast RNA synthesis and processing. In The Molecular Biology of Chloroplast and Mitochondria in Chlamydomonas (Rochaix J.D., Goldschmidt-Clermont M., and Merchant S., ed.). Springer, Dordrecht, 165-181.
STERN, D.B., RADWANSKI, E.R. & KINDLEB, K.L. (1991). A 3' stem/loop structure of the Chlamydomonas chloroplast atpB gene regulates mRNA accumulation in vivo. Plant Cell, 3, 285-297.
SUAY, L., SALVADOR, M.L., ABESHA, E. & KLEIN, U. (2005). Specific roles of 5'
RNA secondary structures in stabilizing transcripts in chloroplasts. Nucleic Acids Research, 33, 4754-4761.
SUEOKA, N. (1960). Mitotic replication of deoxyribonucleic acid in Chlamydomonas reinhardtii. Proceedings of the National Academy of Sciences of the United States of America, 46, 83-91.
SUGINO, A., SNOPEK, T.J. & COZZARELLI, N.R. (1977). Bacteriophage T4 RNA ligase: reaction intermediates and interaction of substrates. Journal of Biological Chemistry, 252 , 1732-1738.
174

Bibliografía
SULKOWSKI, E. (1985). Purification of proteins by IMAC. Trends in Biotechnology, 3, 1-7.
SYMONS, R.H. (1981). Avocado sunblotch viroid: primary sequence and proposed secondary structure. Nucleic Acids Research, 23, 6527-6537.
TABLER, M. & TSAGRIS, M. (1990) Viroid replication mechanisms. In Recognition and Response in Plant-Virus Interactions, NATO ASI Series, Vol H41 (Fraser, R.S.S. ed.). Springer-Verlag, 185-205.
TABLER, M. & TSAGRIS, M. (2004). Viroids: petite RNA pathogens with
distinguished talents. Trends in Plant Science, 9, 339-348. TABLER, M., TZORTZAKAKI, S. & TSAGRIS, M. (1992). Processing of linear
longer-than-unit-length potato spindle tuber viroid RNAs into infectious monomeric circular molecules by a G-specific endoribonuclease. Virology, 190, 746-753.
TOBIAS, J.W., SHRADER, T.E., ROCAP, G. & VARSHAVSKY, A. (1991) The N-end rule in bacteria. Science, 254, 1374-1377.
TREGONING, J., MALIGA, P. DOUGAN, G. & NIXON, P.J. (2004). New advances in the production of edible plant vaccines: chloroplast expression of a tetanus vaccine antigen, TetC. Phytochemistry, 65, 989-994.
TROTTA, C.R., MIAO, F., ARN, E.A., STEVENS, S.W., HO, C.K., RAUHUT, R. & ABELSON, J.N. (1997). The yeast tRNA splicing endonuclease: a tetrameric enzyme with two active site subunits homologous to the archaeal tRNA endonucleases. Cell, 89, 849-858.
TSAGRIS, M., TABLER, M., MÜHLBACH, H.P. & SÄNGER, H.L. (1987). Linear oligomeric potato spindle tuber viroid (PSTV) RNAs are accurately processed in vitro to the monomeric circular viroid proper when incubated with a nuclear extract from healthy potato cells. EMBO Journal, 6, 2173-2183.
VARSHAVSKY, A. (1996). The N-end rule: Functions, mysteries, uses. Proceedings of the National Academy of Sciences of the United States of America, 93, 12142-12149.
WAHN, D., ROSENGERG DE GÓMEZ, R. & SÄNGER, H.L. (1980). Cytophathic effects at the plasma membrane in association with pathogenic RNA. Journal of General Virology, 49, 355-365.
WANG, L.K., HO, C.K., PEI, Y. & SHUMAN, S. (2003). Mutational analysis of
bacteriophage T4 RNA ligase 1: different functional groups are required for the nucleotidyl transfer and phosphodiester bond formation steps of the ligation reaction. Journal of Biological Chemistry, 278, 29454-29462.
WANG, L.K., LIMA, C.D. & SHUMAN, S. (2002). Structure and mechanism of T4 polynucleotide kinase: an RNA repair enzyme. EMBO Journal, 21, 3873-3880.
175

Bibliografía
WANG, L.K., SCHWER, B., ENGLERT, M., BEIER, H. & SHUMAN, S. (2006a). Structure-function analysis of the kinase-CPD domain of yeast tRNA ligase (Trl1) and requirements for complementation of tRNA splicing by a plant Trl1 homolog. Nucleic Acids Research, 34, 517-527.
WANG, L.K., SCHWER, B. & SHUMAN, S. (2006b). Structure-guided mutational analysis of T4 RNA ligase 1. RNA, 12, 2126-2134.
WANG, L.K. & SHUMAN, S. (2002). Mutational analysis defines the 5’-kinase and 3’-phosphatase active sites of T4 polynucleotide kinase. Nucleic Acids Research, 30, 1073-1080.
WANG, L.K. & SHUMAN, S. (2005). Structure-function analysis of yeast tRNA ligase. RNA, 11, 966-975.
WARRILOW, D. & SYMONS, R.H. (1999). Citrus exocortis viroid RNA is associated with the largest subunit of RNA polymerase II in tomato in vivo. Archives of Virology, 144, 2367-2375.
WASSENEGGER, M., HEIMES, S. & SÄNGER, H.L. (1984). An infectious viroid RNA replicon evolved from an in vitro-generated non-infectious viroid deletion mutant via a complementary deletion in vivo. EMBO Journal, 13, 6172-6177.
WASSENEGGER, M. & KRCZAL, G. (2006). Nomenclature and functions of RNA-
directed RNA polymerases. Trends in Plant Science, 11, 142-151. WEICKERT, M.J., DOHERTY, D.H., BEST, E.A. & OLINST, P.O. (1996).
Optimization of heterologous protein production in Escherichia coli. Current Opinion in Biotechnology, 7, 494-499.
WEITZER, S. & MARTINEZ, J. (2007). The human RNA kinase hClp1 is active on 3’
transfer RNA exons and short interfering RNAs. Nature, 447, 222-226. WESTAWAY, S.K., BELFORD, H.G., APOSTOL, B.L., ABELSON, J. & GREER,
C.L. (1993). Novel activity of a yeast ligase deletion polypeptide: evidence for GTP-dependent tRNA splicing. Journal of Biological Chemistry, 268, 2435-2443.
WESTAWAY, S.K., PHIZICKY, E.M. & ABELSON, J. (1988). Structure and function of the yeast tRNA ligase gene. Journal of Biological Chemistry, 263, 3171-3176.
WHITE, T.C. & BORST, P. (1987). RNA end-labeling and RNA ligase activities can produce a circular ribosomal RNA in whole cell-extracts from trypanosomes. Nucleic Acids Research, 15, 3275-3290.
WONG, J. T.F. (1976). The evolution of a universal genetic code. Proceedings of the National Academy of Sciences of the United States of America, 73, 2336-2340.
WU, H.N. & LAI, M.M. (1989). Reversible cleavage and ligation of hepatitis delta virus RNA. Science, 243, 652-654.
XU, Q., TEPLOW, D., LEE, T.D. & ABELSON, J. (1990). Domain structure in yeast tRNA ligase. Biochemistry, 26, 6132-6138.
176

Bibliografía
YAMAGUCHI, K., BELIGNI, M.V., PRIETO, S., HAYNES, P.A., MC DONALD, W.H., YATES III, J.R. & MAYFIELD, S.P. (2003). Proteomic characterization of the Chlamydomonas reinhardtii chloroplast ribosome: identification of proteins unique to the 70 s ribosome. Journal of Biological Chemistry, 278, 33774-33785.
YIN, S.M., HO, C.K., MILLER, E.S. & SHUMAN, S. (2004). Characterization of
bacteriophage KVP40 and T4 RNA ligase 2. Virology, 319, 141-151.
YIN, S.M., HO, C.K. & SHUMAN, S. (2003). Structure-function analysis of T4 RNA ligase 2. Journal of Biological Chemistry, 278, 17601-17608.
ZAITLIN, M., NIBLETT, C.L., DICKSON, E. & GOLDBERG, R.B. (1980). Tomato DNA contains no detectable regions complementary to potato spindle tuber viroid as assayed by solution and filter hybridization. Virology, 104, 1-9.
ZERGES, W. (2000). Translation in chloroplasts. Biochimie, 82, 583-601. ZHANG, X-H. & CHIANG, V.L. (1996). Single-stranded DNA ligation by T4 RNA
ligase for PCR cloning of 5’-noncoding fragments and coding sequence of a specific gene. Nucleic Acids Research, 24, 990-991.
ZILLMANN, M., GOROVSKY, M.A. & PHIZICKY, E.M. (1991). Conserved
mechanism of tRNA splicing in eukaryotes. Molecular and Cellular Biology, 11, 5410-5416.
ZILLMANN, M., GOROVSKY, M.A. & PHIZICKY, E.M. (1992). HeLa cells contain a 2'-phosphate-specific phosphotransferase similar to a yeast enzyme implicated in tRNA splicing. Journal of Biological Chemistry, 267, 10289-10294.
ZOFALLOVA, L., GUO, Y. & GUPTA, R. (2000). Junction phosphate is derived from
the precursor in the tRNA spliced by the archaeon Haloferax volcanii cell extract. RNA, 6, 1019-1030.
ZURAWSKI, G., PERROT, B., BOTTOMLEY, W. & WHITFELD, P.R. (1981). The
structure of the gene for the large subunit of ribulose 1,5-bisphosphate carboxylase from spinach chloroplast DNA. Nucleic Acids Research, 9, 3251-3270.
177


88.. AANNEEJJOOSS


Anejos
I. Cebadores Extensión del cebador sobre el RNA monomérico circular del ASBVd+
C1 => 5’- TGAAGAGACGAAGTGATCAAG - 3’
Experimentos de RACE (5’,3’ y circular)
C2 => 5’- CCAGTGAGCAGAGTGACGAGGACTCGAGCTCAAGCTTTTTTTTTTT
TTTTTTVN - 3’
C3 => 5’- ATCACCTTTATGATGGAAAGAGCCG - 3’
C4 => 5’- CCAGTGAGCAGAGTGACG - 3’
C5 => 5’- AGAGTGAACTTATTGAACGTTTTGG - 3’
C6 => 5’- GAGGACTCGAGCTCAAGC - 3’
C7 => 5’- TCCTGGAAAAAAAACGATGAGACCC - 3’
C8 => 5’- TGGAAATATTAGGGCTTGAAGGAGC - 3’
C9 => 5’- CGACUGGAGCACGAGGACACUGACAUGGACUGAAGGAGUAGAAA - 3’
C10 => 5’- TGGAGCACGAGGACACTGACATG - 3’
C11 => 5’- GACACTGACATGGATGAAGGAGTAG - 3’
C12 => 5’- CCAGTGAGCAGAGTGACGAGGACTCGAGCTCAAGCGGGG - 3’
C13 => 5’- TTYGGNAAYATHTAYACNGC - 3’
C14 => 5’- TTYTAYCCNAARTTYGARAAYG - 3’
C15 => 5’- TTCAGTAGAGGAATTCTGACAAGTG - 3’
C16 => 5’- CAAAGGCCACAGTCCTGGTTTGTGTC - 3’
C17 => 5’- GTGATCCACGTACATAGTGATTCAG - 3’
C18 => 5’- CTTCCGCCGGTACCAGAAAGACATG - 3’
C19 => 5’- AAGGCAGGTTGTCACTCGATAAATC - 3’
C20 => 5’- GGAAGTTATAGTAACACTTGAAAGC - 3’
C21 => 5’- GAGGAGGAGGAAAATCTCTTAGAAC - 3’
C22 => 5’- AAGTCCAGAGCCTCACCCTCTAATG - 3’
C23 => 5’- CTGCAAATAGATCAGAAAAGCAGC - 3’
C24 => 5’- GGACCTACGCTGCGTGAGGTTTGTG - 3’
C25 => 5’- CGTGCCAGCACTCTGAAGAAGCGCC - 3’
C26 => 5’- CCAGATGACTTTTCGATGCTGCAC - 3’
C27 => 5’- CAGACACTGTTCCATATGACTTGGG - 3’
181

Anejos
Amplificación de la tRNA ligasa de tomate completa
C28 => 5’- GGCGCGGC GCTAGC ATGCCAAACAATCAGAGAAAG - 3’
C29 => 5’- GCCGCGCC CTCGAG AAAGAATTCGAGAGTGCCAG - 3’
Amplificación de la tRNA ligasa de Arabidopsis thaliana completa
C30 => 5’- ATGGATGCTCCATTTGAATCCGGTGATAG - 3’
C31 => 5’- GAAAAACTCCAGAGGACCTGAGATTGACAC - 3’
Amplificación del plásmido pET23a(+) completo
C32 => 5’- ATGTATATCTCCTTCTTAAAGTTAAACAAAATTATTTC - 3’
C33 => 5’- CACCACCACCACCACCACTGAGATCCGGCTG - 3’
Amplificación de la construcción para expresar un RNA dimérico viroidal
en el cloroplasto de C. reinhardtii
C34 => 5’- AAATGTATTTAAAATTTTTCAAC - 3’
C35 => 5’- GCGGCCGCATTATAAAATCGGTTAGAC - 3’
Secuenciación
Directo => 5’- GTAAAACGACGGCCAGT - 3’
Reverso => 5’- CAGGAAACAGCTATGAC - 3’
Terminador T7 => 5’- TATGCTAGTTATTGCTCAG - 3’
182

Anejos
II. Servidores Web Bases de datos
1 => Universidad de Cornell: http://www.sgn.cornell.edu/solanaceae-project/index.pl
2 => NCBI: http://www.ncbi.nlm.nih.gov
3 => ExPASy: http://www.expasy.org/
Programas
4=> ClustalW: http://www.ebi.ac.uk/clustalw
5=> ProtParam: http://www.expasy.ch/tools/protparam.html
6=> Lalign: http://www.ch.embnet.org/software/LALIGN_form.html
7=> Multalin: http://www.toulouse.inra.fr/multalin/multalin.html
8=> Mfold: http://frontend.bioinfo.rpi.edu/zukerm/cgi-bin/rna-index.cgi
9=> MUSCLE: http://www.drive5.com/muscle
10=> ProtTest: http://darwin.uvigo.es/software/prottest_server.html
11=> MEGA: http://www.megasoftware.net
183




NNoo ddeesseessppeerreess,, nnii ssiiqquuiieerraa ppoorr eell hheecchhoo ddee qquuee nnoo ddeesseessppeerraass..
CCuuaannddoo ttooddoo ppaarreeccee tteerrmmiinnaaddoo,, ssuurrggeenn nnuueevvaass ffuueerrzzaass.. EEssttoo
ssiiggnniiffiiccaa qquuee vviivveess..
FFrraannzz KKaaffkkaa