Embed Size (px)
Citation preview

Seleção de mutantes de Gluconacetobacter
diazotrophocus defectivos em produção de biofilme
Bianca Simões Gomes de Oliveira
Rio de Janeiro
2009

BIANCA SIMÕES GOMES DE OLIVEIRA
Aluna do Curso de BiotecnologiaMatrícula 0613800093
SELEÇÃO DE MUTANTES DE GLUCONACETOBACTER DIAZOTROPHICUS DEFECTIVOS EM PRODUÇÃO DE
BIOFILME
Trabalho de Conclusão de Curso, TCC,
apresentado ao Curso de Graduação em
Biotecnologia da UEZO como parte dos
requisitos para a obtenção do grau em
Biotecnólogo, sob a orientação da Profª
Drª Vânia Lúcia Muniz de Pádua.
Rio de Janeiro
Julho de 2009

SELEÇÃO DE MUTANTES DE GLUCONACETOBACTER
DIAZOTROPHICUS DEFECTIVOS EM PRODUÇÃO DE
BIOFILME
Elaborado por Bianca Simões Gomes de Oliveira
Aluna do Curso de Biotecnologia da UEZO
Este trabalho de Graduação foi analisado e aprovado com
Grau 9,5
Rio de Janeiro, 30 de julho de 2009.
___________________________________________
Drª Maria Luiza Gomes
___________________________________________
Drª Alaíde Barreto
__________________________________________
Drª Vânia Lúcia Muniz de Pádua, Orientadora e Presidente da Banca
RIO DE JANEIRO, RJ – BRASIL
JULHO DE 2009

RESUMO
A Gluconacetobacter diazotrophicus é uma bactéria diazotrófica endofítica que
coloniza o interior do tecido da cana-de-açúcar, promovendo fixação de nitrogênio e
aumento na produção de fitohormônios sem causar sintomas de doença, nas condições
naturais, as bactérias são encontradas em sua maioria associadas em forma de biofilme.
Biofilmes são associações de microorganismos e produtos extracelulares e representam
sistemas biológicos altamente organizados.
O objetivo do presente estudo foi a identificação de mutantes defectivos para genes
responsáveis pela formação do biofilme a partir de um banco cedido pela EMBRAPA-
Agrobiologia, a fim de identificar a provável função do gene no biofilme e entender a
possível participação na associação com a planta. O objetivo foi atingido utilizando a
técnica de sequenciamento de Sanger, com primers específicos para o transposon
possibilitando a identificação de mutantes defectivos em genes selecionados durante o
procedimento estabelecido para o crescimento da bactéria como biofilme.
Foram isolados 16 mutantes candidatos deficientes em produção de biofilme, sendo
que dez foram identificados pela técnica de sequenciamento, dentro das suas categorias
funcionais, enquanto os outros seis foram seqüenciados, porém não foi possível a
identificação.
Palavras-chave: biofilme, G. diazotrophicus, cana-de-açúcar

ABSTRACT
Gluconacetobacter diazotrophicus is a diazotrophic endophytic bacterium that
colonizes the tissue of sugar cane, promoting nitrogen fixation and increasing the
production of plant hormones without causing diseases. Biofilms are associations of
microorganisms and extracellular products and represent highly organized biological
systems.
The purpose of this study was the identification of mutant defects in genes
responsible for biofilm formation, from a bank assigned by EMBRAPA-Agrobiologia to
establish the putative function of the gene in biofilm and understand the possible
participation in association with the plant. The objective was achieved using the technique
of Sanger sequencing, with specific primers for the transposon allowing the identification
of mutants defects in genes selected during the procedure established for the growth of
bacteria in biofilm form.
16 candidates were screened, which 10 were identified by the technique of
sequencing DNA and classified into their functional categories.
Keywords: biofilm, G. diazotrophicus, sugar cane

SUMÁRIO
Resumo.................................................................................................................................iii
Abstract................................................................................................................................iv
1.INTRODUÇÃO.................................................................................................................1
1.1. Da cana – de – açúcar...............................................................................................1
1.2. Da bactéria.................................................................................................................1
1.3. Justificativa................................................................................................................4
2. OBJETIVOS.....................................................................................................................6
2.1. Objetivos gerais.........................................................................................................6
2.2. Objetivos específicos.................................................................................................6
3. METODOLOGIA............................................................................................................7
3.1. Pré - seleção dos mutantes defectivos na produção do biofilme...........................9
3.1.1. Meio de cultura utilizado..................................................................................9
3.1.2. O cristal violeta..................................................................................................9
3.1.3. A triagem dos mutantes..................................................................................10
3.2. Sequenciamento de DNA........................................................................................11
3.2.1. Extração de DNA.............................................................................................11
3.2.2. Quantificação de DNA....................................................................................12
3.2.3. Reação de sequenciamento.............................................................................12
3.2.4. Preparo da amostra para submeter á leitura do seqüenciador..................13
4. RESULTADOS E DISCUSSÃO...................................................................................15
4.1. Ensaio de biofilme...................................................................................................15
4.2. Sequenciamento.......................................................................................................18
5. CONCLUSÃO................................................................................................................22
6. REFERÊNCIAS BIBLIOGRÁFICAS.........................................................................23

1. INTRODUÇÃO
1.1) A CANA-DE-AÇÚCAR
O Brasil é o maior produtor de cana-de-açúcar do mundo, com os menores custos
de produção e, também, o maior exportador do produto. O país possui vantagens na
produção dados os diferenciais climáticos e as variedades de espécies (SOUZA, 2006).
A partir do processo de industrialização da cana-de-açúcar obtêm-se o açúcar,
como também o álcool, o vinhoto e o bagaço (http://infoener.iee.usp.br/scripts/biomassa
/brcana. asp).
Existem diversas utilizações para o álcool como: produção de bebidas alcoólicas,
aplicações na indústria química e farmacêutica, combustível veicular e a produção de
energia elétrica. Dentre elas, a utilização que mais se destaca no contexto atual é o álcool
combustível, que tem a vantagem de ser uma fonte de energia renovável e menos poluidora
que os derivados do petróleo, o que possibilitou o desenvolvimento de uma tecnologia
100% nacional, o Proálcool, um programa criado pelo governo brasileiro com o objetivo
de reduzir a importação de petróleo, assim como o impacto ambiental de sua utilização
(http://www.ambientebrasil.com.br/composer.php3base= ./energia/index.html&conteúdo
=./energia/etanol.html#futuro).
No Brasil, a safra da cana-de-açúcar ocorre no Centro-Sul, nos meses de setembro
a fevereiro e, no Norte/Nordeste, durante os meses de março a agosto. Em virtude disso, o
Brasil é o único país do mundo que produz açúcar e álcool ao longo de todo o ano
(SANTOS; EVANGELISTA, 2006).
1.2 ) A BACTÉRIA
Gluconacetobacter diazotrophicus é uma bactéria gram-negativa, tolerante á
ácido, aeróbica restrita, diazotrófica (fixadora de nitrogênio) e endofítica (coloniza o
interior do tecido), que possui crescimento e fixação de nitrogênio ótimos em temperatura
de 30° C e pH inicial entre 5,5 e 6,0 e coloniza o tecido vascular da cana-de-açúcar e de
outros vegetais sem causar sintomas de doença. A bactéria é raramente encontrada no solo

e não coloniza ervas daninhas ou outras plantas em campos de cana-de-açúcar
(DÖBEREINER et al.1988). Sua associação com a cana tem sido de grande interesse de
estudo devido á fixação de nitrogênio e produção de hormônios de crescimento como
auxinas e citocininas pela bactéria, que otimizam o desenvolvimento da planta, além de
exercer controle biológico contra patógenos. A bactéria secreta uma bacteriocina que inibe
o crescimento de um fitopatógeno da cana-de-açúcar - a Xanthomonas albilineans - e reduz
o pH do meio para abaixo de 3,0 o que inibe o crescimento de outro fitopatógeno da cana-
de-açúcar, o fungo Colletotrichum falcatum (MUTHUKUMARASAMY,REVATHI &
VADIVELU, 2000).
G. Diazotrophicus foi isolada pela primeira vez a partir de cana-de-açúcar em
1988 por Cavalcante e Döbereiner. Atualmente, encontra-se inclusa na classe das α-
Proteobactérias,ordem Rhodospirillales, família Acetobacteraceae e gênero
Gluconacetobacter(http://www.ncbi.nlm.nih.gov/sites/entrezcmd=Retrieve&db=nucleotide
&dopt=GenBank&RID=69K6RZBG016&log%24=nuclalign&blast_rank=2&list_uids
=161784420).
G.diazotrophicus é capaz de liberar nos tecidos da planta até 50% do nitrogênio
fixado (COJHO et al, 1993). Da família Acetobacteracea, o gênero Gluconacetobacter é o
único com espécies capazes fixar o nitrogênio atmosférico e pode ser encontrada no espaço
intercelular realizando associação endofítica com cana-de-açúcar, principalmente em áreas
onde utiliza-se pouco fertilizante nitrogenado. (GILLIS et al., 1989; REIS, OLIVARES &
DÖBEREINER, 1994; FUENTES - RAMÍREZ et al., 1999; FISHER & NEWTON, 2005).
G. Diazotrophicus também pode ser encontrada em associação com outros
vegetais como: batata doce (Ipomoea batatas) (DÖBEREINER, REIS & LAZARANI,
1988), café (Coffea arabica), nas gramíneas Eleusine coracana e Pennisetum purpureum,
ananás (Ananas comosus) (SANTOS, BUSTILLOS-CRISTALES & CABALERO-
MELLADO, 2001), cenoura (Daucus carota), rabanete (Raphanus sativus) e beterraba
(Beta vulgaris) (MUNÕZ-ROJAS, FUENTES-RAMÍREZ- & CABALLERO-MELLADO,
2005).
G.diazotrophicus é capaz de crescer e fixar nitrogênio em condições de sacarose
30% e pH baixo, aproximadamente 3,0. Além de ser capaz de fixar nitrogênio na presença
de nitrato e de oxigênio (CAVALCANTE E DÖBEREINER, 1988), o que é de grande
importância devido á sua utilização como insumo agrícola em cultivares tratados
anteriormente com fertilizantes á base de nitrato.

A cepa PAL 5 foi selecionada para ser decodificada pelo projeto RIOGENE
devido a análises anteriores que demonstraram por experimentos ser esta cepa a de maior
interesse econômico devido ao melhor desempenho na associação com a cana-de-açúcar
(MUÑOZ-ROJAS e CABALLERO-MELLADO , 2003).
O genoma completo de G. diazotrophicus possui aproximadamente 4.3 Mb, sendo
composto de um cromossoma circular, e 2 pequenos plasmídeos
(http://www.ncbi.nlm.nih.gov/sites/entrezDb=genome&Cmd=ShowDetailView&TermToS
earch=21712). Toda a seqüência genômica de G. diazotrophicus encontra-se depositada e
disponibilizada no banco de dados EMBL (European Molecular Biology Laboratory ) e
NCBI (National Center for Biotechnology Information) sob números de acesso
AM889285(plasmídeo pGDIPa15I), AM889286 (cromossomo) e AM889287 (plasmídeo
pGDIA01). A figura 1 ilustra a bactéria observada por microscopia eletrônica.
Figura 1. Microscopia eletrônica da bactéria G. diazotrophicushttp://www.cnpab.embrapa.br/educacao/baby/acetobac.html

1.3) JUSTIFICATIVA
Com o objetivo de aumentar o conhecimento científico acerca da associação da
Gluconacetobacter diazotrophicus PAL 5 com a cana-de-açúcar foi elaborado um projeto
para selecionar mutantes de interesse defectivos para formação de biofilme a fim de
possibilitar estudos e investigações posteriores que elucidem os mecanismos desta
interação. A proposta do presente estudo é considerada inovadora, visto que estudos
anteriores observam a formação de biofilme apenas para microorganismos patogênicos, o
que não é o caso da bactéria em questão.
Nas condições naturais, as bactérias são encontradas em sua grande maioria
associadas na forma de biofilme. Biofilmes são associações de microorganismos e seus
produtos extracelulares, principalmente carboidratos. Representam sistemas biológicos
altamente organizados, onde as bactérias estabelecem comunidades funcionais estruturadas
e coordenadas. A associação dos organismos em biofilmes constitui uma forma de
proteção ao seu desenvolvimento, aumentando a aquisição de nutrientes e facilitando a
comunicação celular, o que favorece relações simbióticas e auxilia na sobrevivência em
ambientes hostis (JEFFERSON, 2004). A resistência física conferida pelo biofilme é
essencial para a sobrevivência da bactéria em ambientes como, por exemplo, o xilema da
cana-de-açúcar, que tem o fluxo de seiva bruta muito intenso (TANG AND BOYER,
2008).
Além disso, a comunidade formada no biofilme é capaz de otimizar determinadas
estratégias que a bactéria possui, mas que em sua forma planctônica não teriam grandes
efeitos como a produção de toxinas, antibióticos, entre outros. A formação do biofilme
ocorre em três etapas, a aproximação e adesão das células, a colonização e posteriormente
a desagregação e migração para outros nichos (JEFFERSON, 2004), como pode ser
observado na figura 2.
Diante do reconhecimento de que a formação do biofilme é um aspecto
importante da maioria dos tipos de associações biológicas entre bactérias e seus
hospedeiros, foi desenvolvido este trabalho para a identificação de genes envolvidos na
formação do biofilme.

Figura 2: esquema de formação de biofilme
http://cqb.fc.ul.pt/biocorrosion/images/biofilm11.jpg
Aproximação e Adesão das células à superfície
Colonização: comunicação intercelular e secreção de polissacarídeos
Desagregação: migração para outros nichos, propagação

2. OBJETIVOS
2.1) OBJETIVOS GERAIS
Aumentar o conhecimento científico acerca da associação de Gluconacetobacter
diazotrophicus com planta hospedeira, através da identificação de genes envolvidos com o
estabelecimento do nicho bacteriano devido ao crescimento como biofilme.
2.2) OBJETIVOS ESPECÍFICOS
1. Estabelecer procedimento de triagem dos mutantes aleatórios relacionados à formação
de biofilme;
2. Selecionar as bactérias candidatas mutantes deficientes na formação de biofilme;
3. Identificar por sequenciamento de DNA o gene mutagenizado por inserção de
transposon;

3. METOLOLOGIA
O trabalho de identificação foi baseado em um banco de mutantes aleatórios
derivados de Gluconacetobacter diazotrophicus PAL 5, elaborado pela EMBRAPA
(Empresa Brasileira de Pesquisa Agropecuária ) Agrobiologia – Laboratório de Genética e
Bioquímica, depositado no banco de células ATCC sob nº 49037 e no banco de
microrganismos EMBRAPA-Agrobiologia sob nº BR11281; este banco contém
aproximadamente 7000 mutantes randômicos.
Os mutantes foram obtidos através do kit de mutagênese randômica EZ-Tn5TM
<KAN-2>Tnp TransposomeTM (Epicentre®).
O Transposoma EZ-Tn5 <KAN-2>Tnp contém o gene de resistência a canamicina
Tn903 (KanR) e é flanqueado por terminais de 19 pb que são normalmente reconhecidos
pela transposase EZ-Tn5 incluída no transposossoma introduzido em células bacterianas
através de eletroporação. A transposase é ativada por Mg2+ presente no ambiente
intracelular, resultando em inserções randômicas do transposon EZ-Tn5 <KAN-2> no
DNA genômico da bactéria eletroporada.
Para a produção dos mutantes, foram realizadas transformações através da
eletroporação, utilizando 1 µl do transposoma EZ-Tn5<KAN-2>Tnp (20 ng/µl) e 70 μl de
células eletrocompetentes de G. diazotrophicus PAL5 contidas em cubetas de
eletroporação e logo submetidas a 1,8 Kilovolts por 4,5-5 msegundos em um equipamento
Gene Pulser (Biorad®) visando a produção de poros na membrana e parede celular.
Imediatamente após o pulso, 350 μl de meio DYGS (DÖBEREINER et al., 1995)
foram adicionados á cubeta de eletroporação para recuperação das células transformadas,
posteriormente incubadas a 28°C por 2 horas sob leve agitação. As células foram
plaqueadas em meio DYGS sólido suplementado com 100 µg/mL de canamicina, e
incubadas a 28 °C durante 72h.
As colônias resistentes são os prováveis mutantes. Cerca de 7000 colônias foram
cultivadas em placas de 96 poços contendo 200 µl de meio DYGS suplementado com
glicerol a 25% e canamicina a 100 µg/mL por 72 horas, sendo em seguida estocados a -80
ºC.

A figura 3 demonstra a técnica utilizada para a elaboração dos mutantes seguida de
sequenciamento gênico para a identificação dos mesmos, procedimento utilizado na
elaboração do presente trabalho.
Figura 3: Esquema demonstrativo do processo de elaboração do banco de mutantes realizado pela
EMBRAPA - AGROBIOLOGIA, seguido do sequenciamento gênico realizado na presente pesquisa, a fim
de determinar os genes mutagenizados.
http://www.epibio.com/images/catalog/i_EZTn5_oriV_KAN2.gif

3.1) PRÉ - SELEÇÃO DOS MUTANTES DEFECTIVOS NA PRODUÇÃO DE
BIOFILME
Com o propósito de realizar a seleção de mutantes candidatos a serem defectivos
em produção de biofilme, foram realizados dois ensaios diferindo na presença e na
ausência de lã-de-vidro, usada como superfície para a adesão das bactérias formadoras de
biofilme. A utilização das duas estratégias fez-se necessária para a redução do número de
falsos positivos uma vez que uma técnica validaria a outra.
3.1.1) Meio de cultura utilizado
O meio de cultura LGIP foi preparado da seguinte maneira:
Açúcar cristal ..........................................................................100 g
K2HPO4 ..............sol. 10%.......................................................2 ml
KH2PO4 .............sol. 10% .......................................................6 ml
MgSO4.7H2O ......sol. 10% ......................................................2 ml
CaCl2.2H2O ........sol. 1% .........................................................2 ml
FeCl3.6H2O ........sol. 1% ........................................................1 ml
Na2MoO4.2H2O ..sol.0,1% ......................................................2 ml
Vitamina para meio de cultura ................................................1 ml
Azul de bromotimol solução a 0,5% em 0,2 N de KOH .........5 ml
O pH foi ajustado para 5,5 com solução de ácido acético a 10%. O volume final foi
completado para 1000 ml com água destilada.
3.1.2) O cristal violeta
O cristal violeta foi usado para corar o carboidrato proveniente da formação de
biofilme, funcionando dessa maneira como indicador da produção de biofilme, a partir da
leitura da coloração em espectrofotômetro com base no procedimento descrito por O'Toole
et al., (1999).

3.1.3) Triagem dos mutantes
Foram preparadas duas placas de noventa e seis poços, com capacidade para 2 mL
em cada poço, contendo 1,5 mL de meio LGIP. Após autoclavagem, foi feito o inóculo no
meio acima, suplementado com canamicina (50 microgramas por 50 mL de meio). As
bactérias inoculadas foram cultivadas sob agitação (250 rpm) a 28° C, durante sete dias.
Após o período de cultivo, a suspensão bacteriana foi centrifugada por 10 minutos
a 400 rpm, o sobrenadante de cada poço foi descartado e 100 microlitros de cristal violeta a
0,1% foram adicionados, homogeneizados em vórtex e mantidos em repouso por 10
minutos. A mistura foi centrifugada por 10 minutos a 4000 rpm e o sobrenadante
descartado. O material precipitado foi lavado duas vezes por 15 minutos com solução
salina 0,9% através de forte agitação em vórtex, seguida de centrifugação por 10 minutos a
4000 rpm. O sobrenadante foi retirado e o precipitado lavado com 1 mL de etanol 92,8%.
Uma alíquota de 200 microlitros foi transferida para a placa de Elisa, sendo que uma placa
somente com o etanol foi utilizada como branco. Procedeu-se com as leituras no
espectrofotômetro de placas utilizando o comprimento de onda de 560 nm. As leituras
foram organizadas em planilhas do programa Excel para o cálculo matemático da média
aritmética das leituras da placa, seguido da identificação das amostras com valores abaixo
da média da placa. As amostras identificadas por esse processo foram consideradas
candidatas a mutantes defectivos na formação de biofilme. Todo o procedimento foi
realizado em duplicata.
Um segundo procedimento de triagem baseado no uso de lã-de-vidro foi realizado
com os mesmos mutantes, seguindo o mesmo protocolo descrito acima com as seguintes
modificações:
- Antes das placas serem autoclavadas foi adicionada uma porção lã-de-vidro,
ocupando aproximadamente a terça parte de cada um dos poços.
- A retirada de sobrenadante foi feita com o auxílio de micropipeta, ao invés da
centrifugação.
Somente os mutantes que se comportaram como defectivos em formação de
biofilme nas quatro situações foram selecionados para o sequenciamento gênico.
O procedimento foi realizado a partir das placas do banco de mutantes nomeadas
MUT LIB 1 e MUT LIB 2 escolhidas aleatóriamente.

3. 2 ) SEQUENCIAMENTO DE DNA
O método utilizado foi baseado no processo descrito por Sanger em 1975. O
mutante foi identificado através do sequenciamento do DNA mutagenizado, o que foi
possibilitado pelo uso de primer homólogo à extremidade do transposon. Ferramentas
básicas de bioinformática foram utilizadas para analisar a sequência obtida e obter a
identificação precisa do gene contíguo à região do transposon anelada ao primer usado no
sequenciamento, fornecendo os dados para a identificação e as sugestões sobre as funções
dos genes.
Os mutantes selecionados foram inoculados em 10 mL de meio Dygs
suplementado por 10 microlitros de canamicina. As bactérias foram cultivadas sob
agitação (250 rpm) a 28° C por 72 horas, quando então foram coletadas para a extração de
DNA.
3.2.1) Extração de DNA:
Esta etapa foi realizada com o Kit Wizard genomic DNA purification (Promega)®.
A suspensão de células dos mutantes candidatos foi centrifugada por 5 minutos a
25° C e 7000 rpm e o sobrenadante foi descartado. O material precipitado no fundo de cada
tubo foi ressuspenso em 1 mL de solução salina (0,9%) e transferido para eppendorfs. O
material foi centrifugado por 3 minutos a 13000 rpm e o sobrenadante foi descartado.
Seiscentos microlitros de solução de lise foram adicionados e homogenizados
cuidadosamente, evitando a formação de bolhas para depois proceder a um banho seco a
80°C por 10 minutos. Em seguida, o material foi tratado com 3 microlitros de RNAse 37°C
e submetido ao banho-maria por 1 hora, as proteínas foram precipitadas pela adição de
200 microlitros de solução de precipitação. As amostras foram vigorosamente agitadas em
vórtex e incubadas por 5 minutos no gelo, sendo posteriormente centrifugadas por 5
minutos a 13000 rpm. O DNA foi precipitado após adição de 600 microlitros de
isopropanol e centrifugação por 10 minutos a 13000 rpm. O sobrenadante foi descartado e
o DNA precipitado ou pellet lavado com 500 microlitros de etanol 70%, seguido de
centrifugação a 13000 rpm por 2 minutos, sendo que o sobrenadante deve ser rapidamente

descartado. O pellet secou por aproximadamente 30 minutos à temperatura ambiente e foi
ressuspenso em 50 microlitros de água miliQ previamente autoclavada. O material foi
colocado na geladeira durante a noite (overnight).
3.2.2) Quantificação de DNA:
O DNA foi quantificado por eletroforese utilizando o programa Imagequant
(Molecular Dynamics®).
O gel de agarose (0,8%) foi preparado em TBE (Tris, Borato, EDTA) 0,5X
(Maniatis et al., 1982) adicionando-se 5 microlitros de brometo de etídio (10mg/ml).
As amostras foram adicionadas em microtubos previamente identificados: 6
microlitros de água miliq, 1 microlitro da amostra e como controle a mesma mistura, mas
contendo 1 microlitro de DNA de lambda com 100ng de DNA. Todo o conteúdo foi
aplicado no gel, que foi logo submetido á eletroforese por aproximadamente 25 minutos.
Uma foto do gel foi submetida a um programa de digitalização de imagens e o DNA foi
quantificado. O programa Imagequant permitiu a quantificação das amostras a partir da
comparação da obscuridade relativa ou da intensidade de bandas diferentes, considerando a
área e o volume da manchas das imagens digitalizadas do gel, Maiores detalhes são
encontrados em http://itc.virginia.edu/achs/documents/ACHS-104.pdf.
3.2.3) Reação de sequenciamento:
A reação foi realizada utilizando os reagentes do kit de sequenciamento da
Applied Biosystems. As amostras foram distribuídas em tubos identificados e foram
completadas com água miliq para 29 microlitros. Um microlitro de primer foi adicionado
em cada tubo, os tubos foram levados ao gelo e adicionou-se seis microlitros de tampão em
cada tubo, seguido de quatro microlitros de Big Dye terminator v1.1/v3.1. A mistura foi
toda para o fundo do tubo após uma breve centrifugação (spin).
As amostras foram submetidas ao termociclador no programa MUT SEQ
overnight. Programa MUT SEQ: desnaturação inicial a 95°C por 5 minutos, seguido por 60
ciclos compostos por uma desnaturação a 95 ºC durante 30 segundos, anelamento-
elongamento a 60°C por 4 minutos e um alongamento final a 60 °C durante 10 minutos.

Os primers utilizados nestas reações de sequenciamento são fornecidos pelo kit de
mutagênese randômica EZ-Tn5TM <KAN-2>Tnp TransposomeTM (Epicentre®), os quais
anelam-se nas extremidades do transposon EZ-Tn5 <KAN-2>, propiciando o
sequenciamento bidirecional do DNA genômico.
A descrição dos primers fornecidos pelo kit de mutagênese encontra-se na tabela
abaixo:
Nome Seqüência (5’→3’) Posição de anelamento no transposon EZ-Tn5 <KAN-2>*
KAN-2 Forward
KAN-2 Reverse
ACCTACAACAAAGCTCTCATCAACC
GCAATGTAACATCAGAGATTTTGAG
67 → 43
1127→1151
* Este transposon apresenta um tamanho de 1221 pb.
3.2.4) Preparo da amostra para submeter á leitura do seqüenciador capilar ABI3130
(Applied Biosystems®):
O DNA do sequenciamento foi precipitado após adição, em cada tubo, de 320
microlitros de isopropanol a 68%, misturados por inversão várias vezes e colocados em
temperatura ambiente por trinta minutos. Em seguida, foi realizada uma centrifugação por
quinze minutos a 1200 rpm e o sobrenadante foi descartado por inversão do tubo aberto,
cuidadosamente. O material foi lavado com cuidado adicionando quinhentos microlitros de
isopropanol 75% seguido de centrifugação a 1200 rpm por cinco minutos; em seguida o
sobrenadante foi aspirado com cautela e as amostras ficaram secando no escuro em
temperatura ambiente por uma hora a duas horas. Nesta etapa as amostras podem ser
guardadas no escuro, no freezer (-20oC) por até três dias;
Após este período, foram adicionados dez microlitros de formamida, o tubo foi
fechado, agitado vigorosamente, mantido por trinta minutos em temperatura ambiente e no
escuro, a amostra foi agitada novamente para em seguida ser desnaturada colocando-se a
95°C por cinco minutos e imediatamente depois no gelo. A amostra foi complemente
levada ao fundo por um spin.

O material das placas foi submetido ao seqüenciador capilar e as sequências
geradas foram comparadas com sequências depositadas no banco de dados NCBI, através
de alinhamentos utilizando a ferramenta BLAST (www.ncbi.nlm.nih.gov/BLAST/).
O NCBI foi fundado em 1988 como um recurso nacional para informações de
biologia molecular; é um banco público que cria bases de dados, realiza investigação em
biologia computacional, desenvolve software para análise de dados genoma e dissemina
informações científicas de diferentes áreas, principalmente a biomédica
(http://www.ncbi.nlm.nih.gov/).
A ferramenta BLAST compara sequências de nucleotídeos ou proteínas com
seqüências do banco de dados e calcula a sua significância estatística. O BLAST foi usado
para inferir funções e para ajudar a identificar os membros de famílias de genes
(http://blast.ncbi.nlm.nih.gov/Blast.cgi).

4. RESULTADOS E DISCUSSÃO
4.1) ENSAIO DE BIOFILME
Nesta etapa foi realizada uma pré-seleção de mutantes defectivos para formação
de biofilme, visando o posterior sequenciamento dos genes responsáveis pelo fenótipo
mutante. O ensaio foi realizado em duas condições, na presença e na ausência de lã-de-
vidro, sendo os procedimentos realizados em duplicata. A triagem resultou na seleção de
16 mutantes, sendo 8 da placa MUTLIB1(mutante 1 ao 8) e 8 da placa MUTLIB2 (mutante
9 ao 16). Os clones mutantes foram selecionados de acordo com as leituras de densidade
ótica (D.O) de cada um, em comparação com a média aritmética simples de leitura de
todos os mutantes da placa, sendo selecionados aqueles observados abaixo da média.
Embora o material tenha sido selecionado apenas quando reproduzido nas duas
situações, incluindo suas réplicas, esses dados devem ainda ser reproduzidos, apenas com a
bactéria selvagem (cepa PAL5), os mutantes selecionados e pelo menos cinco não
selecionados, com maiores volumes de cultura de modo a evitar erros de amostragem e no
volume da lã-de-vidro. Além disso, o uso de um protocolo adicional para quantificação
mais precisa da massa de biofilme é requisitado. O quadro 1 ilustra a leitura de densidade
ótica na presença de lã-de-vidro, e o quadro 2 na sua ausência.

COM LÃ -DE-VIDRO
MUTANTE MÉDIA D.O DA PLACA D.O DO MUTANTE1 1,5044 0,6506
Duplicata 1 1,3779 1,14352 1,5044 0,7703
Duplicata 2 1,3779 1,17323 1,5044 0,3434
Duplicata 3 1,3779 0,70054 1,5044 0,4009
Duplicata 4 1,3779 1,26395 1,5044 1,0085
Duplicata 5 1,3779 1,17796 1,5044 0,9357
Duplicata 6 1,3779 0,99467 1,5044 0,6367
Duplicata 7 1,3779 0,52598 1,5044 1,4852
Duplicata 8 1,3779 1,08279 0,3150 0,1859
Duplicata 9 0,1752 0,161710 0,3150 0,1598
Duplicata 10 0,1752 0,127111 0,3150 0,2829
Duplicata 11 0,1752 0,083612 0,3150 0,1939
Duplicata 12 0,1752 0,124413 0,3150 0,2039
Duplicata 13 0,1752 0,089914 0,3150 0,2606
Duplicata 14 0,1752 0,129815 0,3150 0,2157
Duplicata 15 0,1752 0,088816 0,3150 0,0822
Duplicata 16 0,1752 0,1343Quadro 1: Valores de densidade ótica verificados no experimento com a lã-de-vidro; média da D.O da placa e D.O do mutante em questão; Os mutantes 1 á 8 são provenientes da placa MUTLIB1, enquanto os mutantes 9 a 16 são provenientes da placa MUTLIB2.
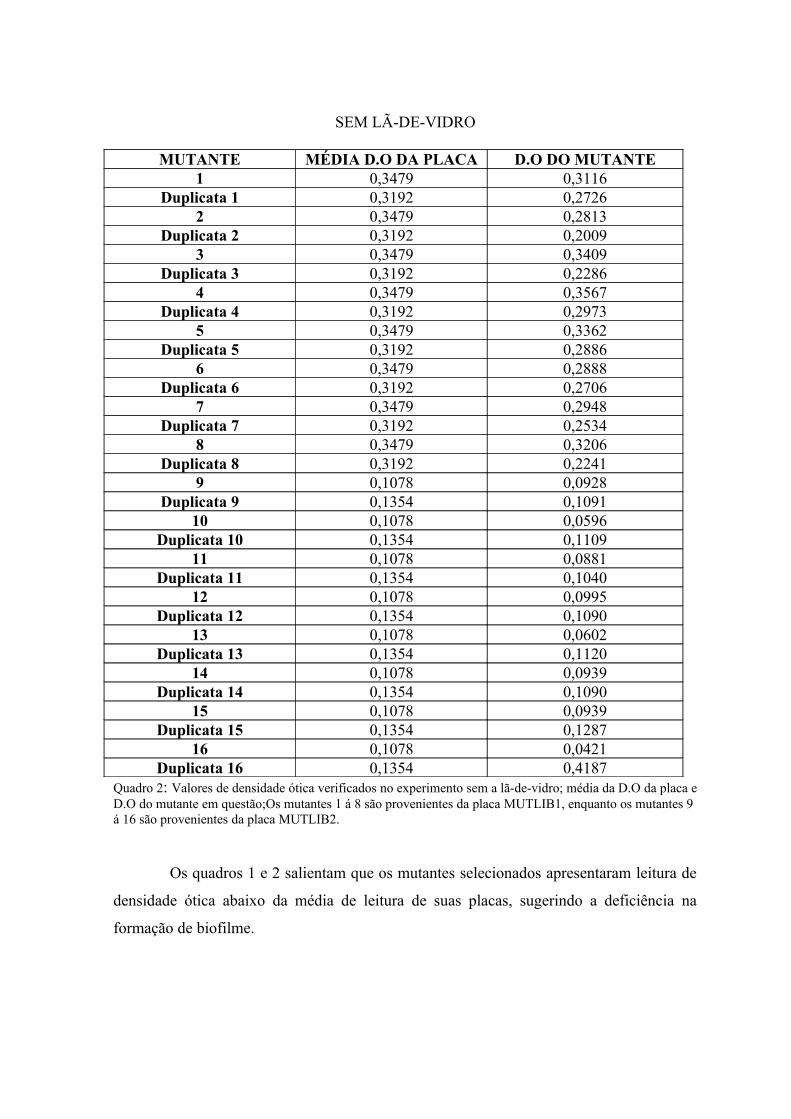
SEM LÃ-DE-VIDRO
MUTANTE MÉDIA D.O DA PLACA D.O DO MUTANTE1 0,3479 0,3116
Duplicata 1 0,3192 0,27262 0,3479 0,2813
Duplicata 2 0,3192 0,20093 0,3479 0,3409
Duplicata 3 0,3192 0,22864 0,3479 0,3567
Duplicata 4 0,3192 0,29735 0,3479 0,3362
Duplicata 5 0,3192 0,28866 0,3479 0,2888
Duplicata 6 0,3192 0,27067 0,3479 0,2948
Duplicata 7 0,3192 0,25348 0,3479 0,3206
Duplicata 8 0,3192 0,22419 0,1078 0,0928
Duplicata 9 0,1354 0,109110 0,1078 0,0596
Duplicata 10 0,1354 0,110911 0,1078 0,0881
Duplicata 11 0,1354 0,104012 0,1078 0,0995
Duplicata 12 0,1354 0,109013 0,1078 0,0602
Duplicata 13 0,1354 0,112014 0,1078 0,0939
Duplicata 14 0,1354 0,109015 0,1078 0,0939
Duplicata 15 0,1354 0,128716 0,1078 0,0421
Duplicata 16 0,1354 0,4187Quadro 2: Valores de densidade ótica verificados no experimento sem a lã-de-vidro; média da D.O da placa e D.O do mutante em questão;Os mutantes 1 á 8 são provenientes da placa MUTLIB1, enquanto os mutantes 9 á 16 são provenientes da placa MUTLIB2.
Os quadros 1 e 2 salientam que os mutantes selecionados apresentaram leitura de
densidade ótica abaixo da média de leitura de suas placas, sugerindo a deficiência na
formação de biofilme.

4.2) SEQUENCIAMENTO
Os dados dos resultados dos sequenciamentos encontram-se expostos no quadro 3,
que relaciona: a placa de origem dos mutantes, sua posição na placa, a identidade do
NCBI, o nome do mutante, e a posição aproximada da mutação no genoma.
PLACA MUTLIB
MUTANTE POSIÇÃO NA PLACA
IDENTIDADE NO NCBI
NOME POSIÇÃO NO GENOMA
1 1 A5 GDI_0278 diguanylate phosphodiesterase
2595736... 2596289
1 2 B10 GDI_0511 putative cell adhesion
508328...508564
1 3 D5 ------------------ -------------------- -------------1 4 E1 GDI_0692 LacI family
transcriptional regulator
703387... 703792
1 5 E2 ---------------- -------------------- -------------1 6 E6 GDI_1373 hypothetical protein 1376637... 13764881 7 G1 ------------------ ---------------------- -------------1 8 G6 ------------------ --------------------- -------------2 9 C5 GDI_1378 putative
ribonuclease PH1382134... 1382513
2 10 D4 GDI_1276 putative oxygen-independent coproporphyrinogen III oxidase
1273014...1273167
2 11 D9 GDI_2999 putative succinylglutamate desuccinylase
3086731... 3086789
2 12 E3 ------------------ -------------------- -------------2 13 E7 GDI_2780 transposase IS256 1542028... 5421192 14 F3 GDI_2402 Putative capsular
polysaccharide biosynth
----------------
2 15 H3 GDI_2414 putative inward rectifier potassium channel
2504733... 2504051
2 16 H12 ----------------- --------------------- -------------------Quadro 3: Quadro reunindo informações provenientes do sequenciamento gênico; os mutantes 3,5,7,8, 12 ,16 não apresentaram sequenciamento satisfatório, impossibilitando a identificação dos genes.
Os dados expostos no quadro 3 foram obtidos a partir de alinhamento da
sequencia resultante do sequenciamento com todas as sequencias depositadas nos bancos
de DNA públicos, utilizando a ferramenta BLASTn disponível no site

www.ncbi.nlm.nih.gov. A figura 4 exemplifica os resultados de alinhamento obtidos no
uso dessa ferramenta.
A localização da inserção do transposon que está sendo mostrada abaixo é baseada
no alinhamento do BLAST, mas há possibilidade do sítio de mutação estar em uma
posição mais 5’ ou 3’, porque nem todos os mutantes analisados resultaram em dados de
sequenciamento de boa qualidade nas duas direções, o que dificulta uma definição mais
precisa da localização. Definições precisas podem ser interessantes em alguns casos menos
freqüentes onde ocorre continuidade da expressão gênica dependendo da região da
inserção. Além disso, considerando que o genoma de organismos procariotos é enxuto,
com poucos espaços não utilizados entre os genes e abundância de operons, há
possibilidade de alteração da expressão de genes contíguos ao que recebeu a inserção, em
função do efeito de posição.
Conforme colocado acima, futuramente esses dados devem ser reproduzidos,
incluindo a bactéria cepa PAL5 e outros clones, com o propósito de evitar erros
experimentais. A anotação mais detalhada dos genes identificados está descrita no trabalho
de monografia de Costa (2009), aluna de Biotecnologia. Entretanto, vale a pena salientar
que a maioria dos genes identificados relaciona-se de forma bem consistente com a
formação de biofilme, porque são responsáveis por metabolismo de carboidrato, adesão
celular e sinalização.

Figura 4: Exemplo de resultado obtido pelo alinhamento utilizando a ferramenta BLASTn. Neste caso específico, o alinhamento foi feito com o gene diguanilato fosfodiesterase.

A figura 5 mostra a localização aproximada do gene diguanilato fosfodiesterase no
DNA genômico da bactéria, assim como a posição aproximada do transposon dentro do
gene. A figura ilustra como as sequências forward e reverse, que são aneladas na
extremidade do transposon Tn5, levam ao resultado de localização dos genes
mutagenizados bem como do inserto dentro no genoma.
Cromossomo
(Mb)0 3,
90,5
1,0
1,5
2,0
2,5
3,0
3,5
Tn 51221 pb
FR
2595736 2596289
2595719
2595097
Diguanilatofosfodiesterase2595083..2595856
CDS mutagenizado
pelo TransposonTn5
Alinhamento das sequências
geradas
SequênciaFowardATGGTTAATTTCGGGGTGGGACTCTAGAGCGACTGCAGGCTGCAAGCTTCAGGGTTGAGATGTGTATAAGAGACAGAACCAGGGAAACGAAGACGTCGATGCGGCGATTCACCAGATCGACGACGGGCTGAAAAAAACATCGTGAACGGAAACAGATCCTGATGGTCGTGGCAGGCCTGGCAGGGGGCGCGACTGGACGGTCCCTTTAAAAAAAAAAATGGGGGGTTTTTTTTGACGTGGATATTCTAATTCATGACTATCGCATGTCGGTCCCGGAAAGCAGCCCATATCCAGCCGCTTCGCTATCCGGGACGCCCCGCATGGCCAGTCGCGGTAAATCCGTGTAGCGTGTCCGCAACCGTGATTATCTTTCGCGGATCGGCATCGGGGAAATCGAAGCGCGGCCATGCACGACGTCCTCGTGGGGTTGATCCTGTTGTGTGTGGTCGTGATCTCCCCTTTGAATATCGGTGGTGGAGAAAAAAGCCTATCCCGCTGCCGCTGTGGCAGATCGTGCTGGGGGTGGTGGGGTCCGTGGCGGGTCTGCATGTTTTTTTTTAACCCCGACATGTTCATGCTGCTGTTCATTCCTCCACTGCTGTTCCCCGATGCGTTCCGCATGCCGACGCACGAATTCGGCCAGATGCGCGGCACATCSequênciaReverseCCCCCCCCCCCCGGCCAATCTCGATGATGGTTGAGATGTGTATAAGAGACAAGACCTGCCCTCGCGGCCCCGGGGGCGAGGGCGCGAGAGGGGCCTGGCGAGGTCACGGAGGAAAACCTGTATGCTTTAGAAAAGGGGGGCCGCACCCGGGCCATCCGGCTGGCGTCCCGGCTTGGGCTGGACCGCCGCCTGAGCATCAATTTCTTGCCCAACGCCGTCTATAACCCGCGCGCCTGCATCCAGTCTACTCTACAGGCCGCCCGCGAAAGCGATTTTCCCCTGGAAAACCTGACCTTCGAGATCCTCGAGACCGAACATCTGACCGAGACGAAGCACCTGCAGGACATCATCGGCGAATATCATCACCATGGCTTCAAGGTCGCCCTGGACGATTTCCCGACCGGACAGTCCGGGCTGATGCGGCTGGCCGACCCCAAGCCCGACATCATCAAGACCGACCGTGGCGGGGGACGGGATTGCGACCGGGACCAGGTGCGCCGCACGATCATGGCCGGCATCATACGAATTGCCACCGATCTGGGCGTCAAGGTCGTGATCGAGGGCGTGGAGCGTCTTGAGGAGGTGCGGGCCCTGCACGCCGTGGGCGGCCGCTTCTTCCAGGGGGTTCTTCTTCGCACGGCCGCTGTTCGAGCAGATCGCGCGGGAGGCGATATCCACTGGC
Representação gráfica dos resultados obtidos com sequenciamento de DNA Genômico purificado de mutantes Tn5 de Gluconacetobacter diazotrophicus PAL5
Figura 5: Exemplo de esquema da inserção do tranposon no genoma da bactéria.

5. CONCLUSÃO
Acredita-se que o biofilme seja um importante requisito para a produção de um
nicho bacteriano adequado e o estabelecimento de uma associação biológica eficiente entre
bactéria e o hospedeiro. Portanto, o estudo de mutantes relacionados ao biofilme deve
ajudar a aumentar o conhecimento científico acerca da associação de Gluconacetobacter
diazotrophicus com sua planta hospedeira, como a cana-de-açúcar.
Dezesseis bactérias G. diazotrophicus mutantes deficientes na formação de
biofilme foram selecionados neste trabalho. Com base no seqüenciamento de DNA
iniciado na sequência do transposon que foi inserido no genoma da bactéria, foi possível
descobrir a identidade do gene interrompido em dez dos dezesseis mutantes selecionados.
A caracterização detalhada de alguns dos mutantes selecionados, com relação ao
seu fenótipo durante o crescimento, sua ultraestrutura e interação com planta estão em
processo de investigação.

6. REFERÊNCIAS BIBLIOGRÁFICAS
CAVALCANTE, V.A. & DÖBEREINER, J. A new acid-tolerant nitrogen-fixing bacterium associated with sugarcane. Plant Soil. 108: 23-31, 1988.
COJHO, E. H. et al. Interactions of Acetobacter Diazotrophicus with an amylolytic yeast in nitrogen-free batch culture. FEMS Microbiol. Lett., v. 106, p. 341-346, 1993.
DÖBEREINER, J.; BALDANI, V.L.D.; BALDANI, J.I. Como isolar e identificar bactérias diazotróficas de plantas não-leguminosas. Brasília: Embrapa-SPI, 1995. 60p.
DÖBEREINER, J.; REIS, V.; LAZARINI, A. C. New N2 fixing bacteria in association with cereals and sugarcane. In: Bothe, H., de Briyn, F.J., Newton, W.E. Nitrogen Fixation.Stuttgart: Gustav Fisher, p. 717-722. 1988.
FISHER, K.; NEWTON, W. E. Nitrogenase proteins from Gluconacetobacter diazotrophicus, a sugarcane-colonizing bacterium. Biochim. Biophys. Acta., v. 1750, p. 154-165, 2005.
FUENTES-RAMÍREZ, L. E.; CABALLERO-MELLADO,J.;SEPÚLVEDA,J.; MARTÍNEZ -ROMERO, E. Colonization of sugarcane by Acetobacter Diazotrophicus is inhibited by high N-fertilization. FEMS Microbiol. Ecol., v. 29, p. 117-128, 1999.
GILDA ROSE SILVA DO AMARAL. Detecção de genes de secreção do tipo IV no genoma da bactéria Gluconacetobacter diazotrophicus e construção de mutantes em genes traG/virD4.2008.132 folhas.Mestrado.Ciências Biológicas.Genética.UFRJ.2008. Orientador: Dra. Ana Maria Abrantes Coelho.
GILDA ROSE SILVA DO AMARAL. Seqüenciamento do genoma da bactéria endofítica Gluconacetobacter diazotrophicus – Participação no consórcio RioGene.UFRJ.2005.59 folhas.
GILLIS, M; KERSTERS,K;HOSTE, B.; JANSSENS, D; R. M. KROPPENSTEDT, R.M.; STEPHAN, M. P.; TEIXEIRA, K.R.S ; J. DÖBEREINER,J.and J. DE LEY . Acetobacter Diazotrophicus sp. nov., a nitrogen-fixing acetic acid bacterium associated with sugarcane. Int. J. Syst. Bacteriol., v. 39, p. 361-364, 1989.
http://infoener.iee.usp.br/scripts/biomassa/br_cana.asp acessado em 20/07/2009 – 16:00
http://www.ambientebrasil.com.br/composer.php3?base=./energia/index.html&conteudo=./energia/etanol.html#futuro acessado em 16/07/2009 – 13:24
http://www.epibio.com/images/catalog/i_EZTn5_oriV_KAN2.gif acessado em 22/07/2009 - 22:30.

http://www.ncbi.nlm.nih.gov/sites/entrezcmd=Retrieve&db=nucleotide&dopt=GenBank&RID=69K6RZBG016&log%24=nuclalign&blast_rank=2&list_uids=161784420 acessado em 20/07/2009 – 17:03http://www.ncbi.nlm.nih.gov/sites/entrez Db=genome&Cmd=ShowDetailView&TermToSe arch=21712 acessado em 21/07/2009 -17:45
http://www.cnpab.embrapa.br/educacao/baby/acetobac.html - acessado em 22/07/2009 - 22:23
http://cqb.fc.ul.pt/biocorrosion/images/biofilm11.jpg acessado em 22/07/2009 - 22:29
JEFFERSON,KIMBERLY,K. What drives bacteria to produce a biofilm? FEMS Microbiology Letters 236 163–173, 2004.
MANIATIS T.; E.F.FRITISH; J. SAMBROOK. Molecular cloning: a laboratory manual, New York, Harbor, Vol. I-III, 1982.
MUNÕZ-ROJAS, J.; CABALLERO-M E L L A D O , J . Population Dynamics ofGluconacetobacter diazotrophicus in Sugarcane Cultivars and Its Effect on Plant Growth. Microb. Ecol., v. 46, p. 454-464, 2003.
MUNÕZ-ROJAS, J.; FUENTES-RAMÍREZ, L. E.; CABALLERO-MELLADO, J.Antagonism among Gluconacetobacter diazotrophicus strains in culture media and in endophytic association. FEMS Microbiol. Ecol. v. 54, p. 57-66, 2005.
MUTHUKUMARASAMY, R.;REVATHI, G. & VADIVELU, M. Antagonistic potential of N2-fixing Acetobacter diazotrophic against Colletotrichum falcatum Went: a causal organism of red-rot of sugarcane. Curr. Science. 78: 1063-1065, 2000.
REIS, V. M.; OLIVARES, F. L.; DÖBEREINER, J. Improved methodology for isolation ofAcetobacter Diazotrophicus and confirmation of its endophytic habitat. World J. Microbiol.Biotechnol., v. 10, p. 401-405, 1994.
SANTOS, J. A.; EVANGELISTA, F. R. Perspectivas do setor sucroalcooleironordestino. Fortaleza, Conjuntura Econômica, n. 8, jan.-mar. 2006
SANTOS, P. E. L.; BUSTILLOS-CRISTALES, R.; CABALLERO-MELLADO, J.Burkholderia, a genus rich in plant-associated nitrogen fixers with wide environmental and geographic distribution. Appl. Environ. Microbiol., v. 67, 2790-2798, 2001.
SOUZA, R. Panorama, oportunidades e desafios para o mercado mundialálcool automotivo. UFRJ. Rio Janeiro, 2006 (dissertação de mestrado).

TANG A-C AND BOYER J. S. Xylem tension affects growth-induced water potential and daily elongation of maize leaves. Journal of Experimental Botany 59(4):753-764, 2008.


























