Embed Size (px)
Citation preview

Seminar 5

Revision of pH

Weak acids
• are when:
• e.g. H2CO3, CH3COOH
𝛼 ≤ 0.05
OR
𝑐0
𝐾𝑎≥ 400

Weak acids
• only some small fraction of molecules in solution dissociates to anion and proton:
CH3COOH ⇌ CH3COO− + H+
𝐾𝑎 = 𝐶𝐻3𝐶𝑂𝑂− × [𝐻+]
[𝐶𝐻3𝐶𝑂𝑂𝐻]
𝐾𝑎 – acid dissociation constant
(I) initially:
(C) change:
𝑪𝟎
+𝒙
𝟎 𝟎
+𝒙 −𝒙
(E) quilibrium: 𝑪𝟎 − 𝒙 𝒙 𝒙
𝐾𝑎 = 𝑥 × 𝑥
𝐶0 − 𝑥

Ostwald’s dilution law
• is a relationship between the dissociation constant Kd and the degree of dissociation α of a weak electrolyte
𝐾𝑑 = 𝐴+ × [𝐵−]
[𝐴𝐵]=
𝛼2
1 − 𝛼× 𝑐0
𝐾𝑑 =𝛼2
1 − 𝛼× 𝑐0 ≈ 𝛼2 × 𝑐0
α =𝐾𝑑
𝑐0
𝐴+ = 𝐵− = 𝛼𝑐0 = 𝐾𝑑𝑐0

Practice Problems Calculate the pH of 0.2 mol/L acetic acid. (Ka = 1.74 × 10-5)
Answer:
𝐾𝑎 = 𝐶𝐻3𝐶𝑂𝑂− × [𝐻+]
[𝐶𝐻3𝐶𝑂𝑂𝐻]
1.74 × 10−5 = [𝐻+]2
[𝐶𝐻3𝐶𝑂𝑂𝐻]
𝐻+ = 1.74 × 10−5 × 0.2 = 1.86 × 10−3
𝑝𝐻 = − log (1. 86 × 10−3) = 2.73
𝛼 ≤ 0.05
OR 𝑐0
𝐾𝑎≥ 400

Practice Problems
Calculate the pH of 0.01 mol/L acetic acid. (Ka = 1.8 × 10-5)
Answer:
𝑝𝐻 =1
2 × 𝑝𝐾 −
1
2log [𝐴𝐻]
𝑝𝐾 = − log 1.8 × 10−5 = 4.7447
𝑝𝐻 =1
2 × 4.7447 −
1
2log0.01 = 2.372 − −1 = 3.372

Revision of Ion-Exchange
Chromatography

Ion-Exchange Chromatography (by charge)
• positively charged proteins (cationic proteins) can be separated on negatively charged columns
• negatively charged proteins (anionic proteins) can be separated on positively charged columns

more negative more positive

At a pH below their pI, proteins carry a net positive charge above their pI they carry a net negative charge
more negative protein => pI<pH
if pH is 6.5
more positive protein => pI>pH

• consider the following ion exchange chromatography test at pH 6.0
• a mixture of proteins were applied to the column in a low [NaCl] buffer
• proteins retained by the cation exchange column were eluted by gradually increasing the NaCl concentration within the buffer
• what can be concluded regarding the pI and relative charge of the protein(s) in each chromatogram peak?
Sample question

Glycolysis and metabolism of
pyruvate

• glycolysis comes from a merger of two Greek words
– Glykys = sweet
– Lysis = breakdown/ splitting
• it is also known as Embden-Meyerhof-Parnas pathway or EMP pathway
Glycolysis

• is the sequence of 10 enzyme-catalyzed reactions that converts glucose into pyruvate with simultaneous production on of ATP
• in this oxidative process, 1 mol of glucose is partially oxidised to 2 moles of pyruvate
• this major pathway of glucose metabolism occurs in the cytosol of all cell
• this unique pathway occurs aerobically as well as anaerobically and
doesn’t involve molecular oxygen
Glycolysis

• the glycolytic sequence of reactions differ from species to species only in the mechanism of its regulation and in the subsequent metabolic fate of the pyruvate formed
• in aerobic organisms, glycolysis is the prelude to Citric Acid Cycle and ETC
• glycolysis is the central pathway for glucose catabolism
Glycolysis

Glucose
Extracellular matrix and cell wall
polysachharide
Glycogen, Starch, Sucrose
Pyruvate
Ribose-5-phosphate
oxidation via pentose phosphate pathway
synthesis of structural polymers
storage
oxidation via glycolysis
Major pathways of glucose utilization


• glycolysis leads to breakdown of 6C glucose into two molecules of 3C pyruvate with the enzyme catalyzed reactions being bifurcated (categorized) into 2 phases:
– Phase 1: preparatory phase
– Phase 2: payoff phase
Two phases of glycolysis

• it consists of the first 5 steps of glycolysis in which the glucose is enzymatically phosphorylated by ATP to yield fructose-1,6-biphosphate
• this fructose-1,6-biphosphate is then split in half to yield 2 molecules of 3-carbon containing glyceraldehyde-3-phosphate (TP) and dihyroxyacteone phosphate (DHAP)
Preparatory phase

• the first phase results in cleavage of the hexose chain
• this cleavage requires an investment of 2 ATP molecules to activate the glucose molecule and prepare it for its cleavage into 3-carbon compound
Preparatory phase


• this phase constitutes the last 5 reactions of glycolysis
• this phase marks the release of ATP molecules during conversion of glyceraldehyde-3-phosphtae to 2 moles of pyruvate
• here 4 moles of ADP are phosphorylated to ATP Although 4 moles of ATP are formed, the net result is only 2 moles of ATP per mole of glucose oxidized
Payoff phase


Stepwise explanation of
glycolysis

STEP 1: PHOSPHORYLATION
• glucose is phosphorylated by ATP to form sugar phosphate
• this is an irreversible reaction and is catalyzed by hexokinase
• Thus the reaction can be represented as follows:
Glucose
Glucose-6-phosphate
Hexokinase ATP
ADP

STEP 2: ISOMERIZATION
• it is a reversible rearrangement of chemical structure of carbonyl oxygen from C1 to C2, forming a ketose from the aldose
• isomerization of the aldose glucose-6-phosphate gives the ketose fructose-6-phoshphate
Glucose-6-phosphate
Fructose-6-phosphate
Phosphoglucoisomerase

STEP 3: PHOPHORYLATION
• here the fructose-6-phosphate is phosphorylated by ATP to fructose-1,6-bisphosphate
• This is an irreversible reaction and is catalyzed by phosphofructokinase enzyme
Fructose-6-phosphate
Fructose-1,6-bisphosphate
ATP
ADP
Phosphofructokinase

STEP 4: BREAKDOWN
• this six carbon sugar is cleaved to produce two 3C molecules: glyceradldehyde-3-phosphate (TP or GAlP) and dihydroxyacetone phosphate (DHAP)
• this reaction is catalyzed by aldolase
Fructose-1,6-bisphosphate
Glyceraldehyde-3-phosphate
Dihydroxyacetone phosphate Triose phosphate
isomerase
Aldolase

STEP 5: ISOMERIZATION
• dihydroxyacetone phosphate is oxidized to form glyceraldehyde-3-phosphate
• this reaction is catalyzed by triose phosphate isomerase enzyme
Glyceraldehyde-3-phosphate
Dihydroxyacetone phosphate
triose phosphate isomerase
2
2

STEP 6: ADDING Pi
• 2 molecules of glyceraldehyde-3-phosphate are oxidized
• glyceraldehyde-3-phosphate dehydrogenase catalyzes the conversion of GAlP into 1,3-bisphosphoglycerate
Aldehyde Carboxylic acid
Carboxylic acid
Ortho-phosphate
Acyl-phosphate product

Resultant reaction
Glyceraldehyde-3-phosphate
1,3-bisphosphoglycerate
Glyceraldehyde-3-phosphate dehydrogenase
2NAD⁺ + 2Pi
2NADH + 2H⁺
2
2

STEP 7: PHOSPHORYLATION
• the transfer of high-energy phosphate group that was generated earlier to ADP, form ATP
• this phosphorylation is termed as substrate level phosphorylation as the phosphate donor is the substrate 1,3-bisphosphoglycerate
• the products of this reaction are 2 molecules of 3-phosphoglycerate (G3P)

1,3-bisphosphoglycerate
3-phosphoglycerate
Phosphoglycerate kinase
2 ADP
FIRST SUBSTRATE LEVEL PHOSPHORYLATION
2
2
2
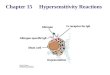
STEP 8: TRANSFER OF Pi
• the remaining phosphate-ester linkage in 3-phosphoglycerate, is moved from carbon 3 to carbon 2, because of relatively low free energy of hydrolysis, to form 2-phosphoglycerate
3-phosphoglycerate
2-phosphoglycerate
Phosphoglycerate mutase
2
2

STEP 9: DEHYDRATION OF G2P
• this is the second reaction in glycolysis where a high-energy phosphate compound is formed
• the 2-phosphoglycerate is dehydrated by the action of enolase to phosphoenolpyruvate (PEP) - this is a reversible reaction
• this compound is the phosphate ester of the enol tautomer of pyruvate

2-phosphoglycerate
Phosphoenol pyruvate
H₂O Enolase
2
2

STEP 10: PHOSPHORYLATION
• this last step is the irreversible transfer of high energy phosphoryl group from phosphoenolpuruvate to ADP
• this reaction is catalyzed by pyruvate kinase
• this is the second substrate level phosphorylation reaction in glycolysis which yields ATP
• this is a non-oxidative phosphorylation reaction

Phosphoenolpyruvate
Pyruvate
2
2
Pyruvate kinase
SECOND SUBSTRATE LEVEL
PHOSPHORYLATION
2ADP
2

OVERALL BALANCE OF GLYCOLYSIS
• the total input of all 10 reactions can be summarized as:
Glucose + 2ATP+ 2Pi+ 2NAD⁺+ 2H⁺+ 4ADP
2Pyruvate+ 2H⁺+ 4ATP+ 2H₂O+ 2NADH+ 2ADP

• glucose is oxidized to pyruvate
• NAD⁺ is reduced to NADH
• ADP is phosphorylated to ATP
Reactions involved in glycolysis
STEP NO. REACTION CONSUMPTION of ATP GAIN of ATP
1 glucose → glucose-6-phosphate - 1 ATP -
3 fructose-6-phosphate → fructose-1,6-biphosphate
- 1 ATP -
7 1,3-diphosphoglycerate → 3-phosphoglycerate
- 2 ATP
10 Phosphoenolpyruvate pyruvate - 2 ATP
TOTAL 2 4
Net gain: 4 – 2 = 2 ATP

Krebs cycle


Formation of acetyl-CoA

Formation of citric acid
Citrate synthase

Dehydration / Hydration
Aconitase

Decarboxylation
Isocitrate dehydrogenase

Oxidative decarboxylation
α-ketoglutarate dehydrogenase complex

succinyl CoA synthetase (succinate thiokinase)
Formation of succinate
the GTP formed is used for the decarboxylation of oxaloacetate to phosphoenolpyruvate in gluconeogenesis, and provides a regulatory link between citric acid cycle activity and the withdrawal of oxaloacetate for gluconeogenesis

Oxidation
succinate dehydrogenase

Hydration
fumarase (fumarate hydratase)

Dehydrogenation
malate dehydrogenase

TOTAL (for 2 cycles)
• 4 carbon dioxide molecules
• 6 NADH molecules
• 2 FADH2 molecules
• 2 ATP molecules
Summary of the Krebs Cycle

Gluconeogenesis

• gluconeogenesis occurs mainly in liver
• occurs to a more limited extent in kidney and small intestine under some conditions
• synthesis of glucose from pyruvate utilizes many of the same enzymes as glycolysis
• three glycolysis reactions have such a large negative DG that they are essentially irreversible
– hexokinase
– phosphofructokinase
– pyruvate kinase
Gluconeogenesis
these steps must be bypassed in gluconeogenesis
two of the bypass reactions involve simple hydrolysis reactions

• hexokinase or glucokinase (glycolysis) catalyzes:
glucose + ATP glucose-6-phosphate + ADP
• glucose-6-phosphatase (gluconeogenesis) catalyzes:
glucose-6-phosphate + H2O glucose + Pi
H O
OH
H
OHH
OH
CH2OH
H
OH
HH O
OH
H
OHH
OH
CH2OPO32
H
OH
HH2O
1
6
5
4
3 2
+ Pi
glucose-6-phosphate glucose
Glucose-6-phosphatase
Bypass 1

• glucose-6-phosphatase enzyme is embedded in the endoplasmic reticulum (ER) membrane in liver cells
• the catalytic site is found to be exposed to the ER lumen. Another subunit may function as a translocase, providing access of substrate to the active site
H O
OH
H
OHH
OH
CH2OH
H
OH
HH O
OH
H
OHH
OH
CH2OPO32
H
OH
HH2O
1
6
5
4
3 2
+ Pi
glucose-6-phosphate glucose
Glucose-6-phosphatase
Glucose-6-phosphatase

• phosphofructokinase (glycolysis) catalyzes:
fructose-6-P + ATP fructose-1,6-bisP + ADP
• fructose-1,6-bisphosphatase (gluconeogenesis) catalyzes:
fructose-1,6-bisP + H2O fructose-6-P + Pi
fructose-6-phosphate fructose-1,6-bisphosphate
Phosphofructokinase
CH2OPO32
OH
CH2OH
H
OH H
H HO
O
6
5
4 3
2
1 CH2OPO32
OH
CH2OPO32
H
OH H
H HO
O
6
5
4 3
2
1
ATP ADP
Pi H2O
Fructose-1,6-biosphosphatase
Bypass 2

• pyruvate kinase (last step of glycolysis) catalyzes:
phosphoenolpyruvate + ADP pyruvate + ATP
• for bypass of the pyruvate kinase reaction, cleavage of 2 ~P bonds is required
– DG for cleavage of one ~P bond of ATP is insufficient to drive synthesis of phosphoenolpyruvate (PEP)
– PEP has a higher negative DG of phosphate hydrolysis than ATP
Bypass 3

• bypass of pyruvate kinase needs 2 enzymes:
• pyruvate carboxylase (gluconeogenesis) catalyzes:
pyruvate + HCO3 + ATP oxaloacetate + ADP + Pi
• PEP carboxykinase (gluconeogenesis) catalyzes:
oxaloacetate + GTP PEP + GDP + CO2
C
C
CH2
O O
OPO32
C
C
CH3
O O
O
ATP ADP + Pi C
CH2
C
C
O
O O
OO
HCO3
GTP GDP
CO2
pyruvate oxaloacetate PEP
Pyruvate Carboxylase PEP Carboxykinase

• spontaneous decarboxylation contributes to spontaneity of the 2nd reaction
• cleavage of a second ~P bond of GTP also contributes to driving synthesis of PEP
C
C
CH2
O O
OPO32
C
C
CH3
O O
O
ATP ADP + Pi C
CH2
C
C
O
O O
OO
HCO3
GTP GDP
CO2
pyruvate oxaloacetate PEP
Pyruvate Carboxylase PEP Carboxykinase

• biotin has a 5C side chain whose terminal carboxyl is in amide linkage to the e-amino group of an enzyme lysine
• the biotin and lysine side chains form a long swinging arm that allows the biotin ring to swing back and forth between 2 active sites
pyruvate carboxylase uses biotin as prosthetic group
CHCH
H2C
S
CH
NH
C
HN
O
(CH2)4 C NH (CH2)4 CH
CO
NH
O
biotin
N subject to
carboxylation
lysine
residue
H3N+
C COO
CH2
CH2
CH2
CH2
NH3
H
lysine
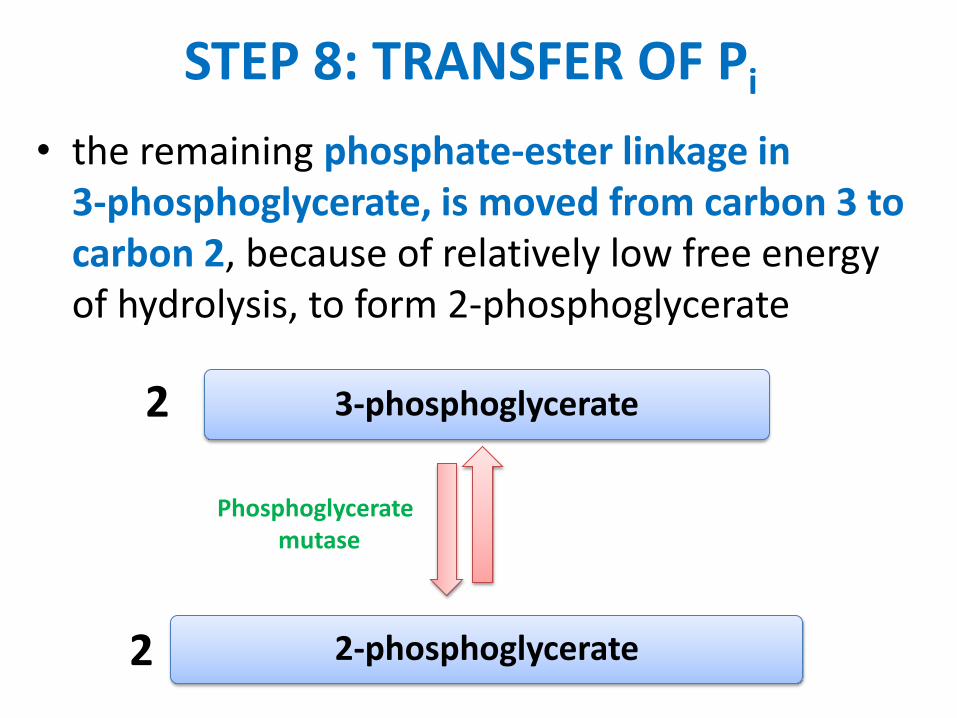
• biotin carboxylation is catalyzed at one active site of pyruvate carboxylase
• ATP reacts with HCO3 to yield carboxyphosphate
• the carboxyl is transferred from this ~P intermediate to N of a ureido group of the biotin ring. Overall:
biotin + ATP + HCO3 carboxybiotin + ADP + Pi
O P O
O
OH
C O
O
carboxyphosphate
CHCH
H2C
S
CH
NH
C
N
O
(CH2)4 C NH (CH2)4 CH
CO
NH
O
C
O
-O
carboxybiotin
lysine
residue

• at the other active site of pyruvate carboxylase the activated CO2 is transferred from biotin to pyruvate:
carboxybiotin + pyruvate
biotin + oxaloacetate
view an animation
CHCH
H2C
S
CH
NH
C
N
O
(CH2)4 C NH R
O
C
O
-OC
C
CH3
O O
O
C
CH2
C
C
O
O O
O
O
CHCH
H2C
S
CH
NH
C
HN
O
(CH2)4 C NH R
O
carboxybiotin
pyruvate
oxaloacetate
biotin

• when gluconeogenesis is active in liver, oxaloacetate is diverted to form glucose
• oxaloacetate depletion hinders acetyl CoA entry into Krebs Cycle
• the increase in acetyl CoA activates pyruvate carboxylase to make oxaloacetate
• pyruvate carboxylase (pyruvate oxaloactate)
is allosterically activated by acetyl CoA
• [Oxaloacetate] tends to be limiting for Krebs cycle
Glucose-6-phosphatase
glucose-6-P glucose
Gluconeogenesis Glycolysis
pyruvate
fatty acids
acetyl CoA ketone bodies
oxaloacetate citrate
Krebs Cycle

avidin
with bound biotin
• avidin, a protein in egg whites with a b barrel structure, tightly binds biotin
• excess consumption of raw eggs can cause nutritional deficiency of biotin
• the strong avidin-to-biotin affinity is used by biochemists as a specific "glue"
Avidin

• PEP carboxykinase catalyzes GTP-dependent oxaloacetate PEP (it is thought to proceed in 2 steps):
– oxaloacetate is first decarboxylated to yield a pyruvate enolate anion intermediate
– phosphate transfer from GTP then yields phosphoenolpyruvate (PEP)
C
C
CH2
O O
OPO32
C
CH2
C
C
O
O O
OO
CO2
C
C
CH2
O O
O
GTP GDP
oxaloacetate PEP
PEP Carboxykinase Reaction

in the bacterial enzyme, ATP is Pi donor instead of GTP
Mg++
pyruvate
Mn++
ATP
PEP Carboxykinase active site ligands PDB 1AQ2
• a metal ion such as Mn2+ is required for the PEP carboxykinase reaction, in addition to a Mg2+ ion that binds with the nucleotide substrate at the active site Mn2+ is thought to promote Pi transfer by interacting simultaneously with the enolate oxygen atom and an oxygen atom of the terminal phosphate of GTP or ATP

• the source of pyruvate and oxaloacetate for gluconeogenesis during fasting or carbohydrate starvation is mainly amino acid catabolism (some amino acids are catabolized to pyruvate, oxaloacetate, or precursors of these)
• muscle proteins may break down to supply amino acids. These are transported to liver where they are deaminated and converted to gluconeogenesis inputs
• glycerol, derived from hydrolysis of triacylglycerols in fat cells, is also a significant input to gluconeogenesis
The source of pyruvate and oxaloacetate

Glyceraldehyde-3-phosphate Dehydrogenase
Phosphoglycerate Kinase
Enolase
PEP Carboxykinase
glyceraldehyde-3-phosphate
NAD+ + Pi
NADH + H+
1,3-bisphosphoglycerate
ADP
ATP
3-phosphoglycerate
Phosphoglycerate Mutase
2-phosphoglycerate
H2O
phosphoenolpyruvate
CO2 + GDP
GTP
oxaloacetate
Pi + ADP
HCO3 + ATP
pyruvate
Pyruvate Carboxylase
Gluconeogenesis
Summary of gluconeogenesis pathway
• gluconeogenesis enzyme names in red
• glycolysis enzyme names in blue
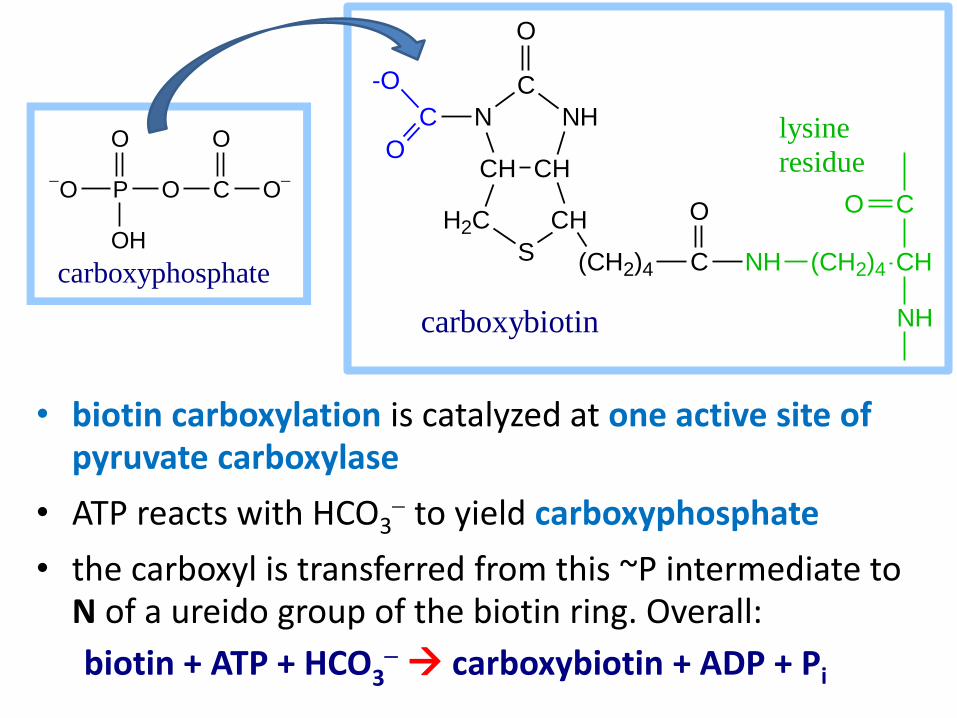
Glucose-6-phosphatase
Fructose-1,6-bisphosphatase
glucose Gluconeogenesis
Pi
H2O
glucose-6-phosphate
Phosphoglucose Isomerase
fructose-6-phosphate
Pi
H2O
fructose-1,6-bisphosphate
Aldolase
glyceraldehyde-3-phosphate + dihydroxyacetone-phosphate
Triosephosphate Isomerase (continued)

Glycolysis and Gluconeogenesis are both spontaneous. If both pathways were simultaneously active in a cell, it would constitute a "futile cycle" that would waste energy.
Glycolysis:
glucose + 2 NAD+ + 2 ADP + 2 Pi
2 pyruvate + 2 NADH + 2 ATP Gluconeogenesis:
2 pyruvate + 2 NADH + 4 ATP + 2 GTP
glucose + 2 NAD+ + 4 ADP + 2 GDP + 6 Pi
Questions:
1. Glycolysis yields how many ~P ?
2. Gluconeogenesis expends how many ~P ?
3. A futile cycle of both pathways would waste how many ~P per cycle ?
2
6
4

To prevent the waste of a futile cycle, Glycolysis and Gluconeogenesis are reciprocally regulated
Local Control includes reciprocal allosteric regulation by adenine nucleotides.
Phosphofructokinase (Glycolysis) is inhibited by ATP and stimulated by AMP.
Fructose-1,6-bisphosphatase (Gluconeogenesis) is inhibited by AMP.
fructose-6-phosphate fructose-1,6-bisphosphate
Phosphofructokinase
CH2OPO32
OH
CH2OH
H
OH H
H HO
O
6
5
4 3
2
1 CH2OPO32
OH
CH2OPO32
H
OH H
H HO
O
6
5
4 3
2
1
ATP ADP
Pi H2O
Fructose-1,6-biosphosphatase

The opposite effects of adenine nucleotides on
Phosphofructokinase (Glycolysis)
Fructose-1,6-bisphosphatase (Gluconeogenesis)
insures that when cellular ATP is high (AMP would then be low), glucose is not degraded to make ATP.
When ATP is high it is more useful to the cell to store glucose as glycogen.
When ATP is low (AMP would then be high), the cell does not expend energy in synthesizing glucose

Global Control in liver cells includes reciprocal effects of a cyclic AMP cascade, triggered by the hormone glucagon when blood glucose is low
Phosphorylation of enzymes and regulatory proteins in liver by Protein Kinase A (cAMP Dependent Protein Kinase) results in
inhibition of glycolysis
stimulation of gluconeogenesis,
making glucose available for release to the blood

Enzymes relevant to these pathways that are phosphorylated by Protein Kinase A include:
Pyruvate Kinase, a glycolysis enzyme that is inhibited when phosphorylated.
CREB (cAMP response element binding protein) which activates, through other factors, transcription of the gene for PEP Carboxykinase, leading to increased gluconeogenesis.
A bi-functional enzyme that makes and degrades an allosteric regulator, fructose-2,6-bisphosphate.

Reciprocal regulation by fructose-2,6-bisphosphate:
Fructose-2,6-bisphosphate stimulates Glycolysis.
Fructose-2,6-bisphosphate allosterically activates the Glycolysis enzyme Phosphofructokinase.
Fructose-2,6-bisphosphate also activates transcription of the gene for Glucokinase, the liver variant of Hexokinase that phosphorylates glucose to glucose-6-phosphate, the input to Glycolysis.
Fructose-2,6-bisphosphate allosterically inhibits the gluconeogenesis enzyme Fructose-1,6-bisphosphatase.

Recall that Phosphofructokinase, the rate-limiting step of Glycolysis, is allosterically inhibited by ATP.
At high concentration, ATP binds at a low-affinity regulatory site, promoting the tense conformation.
0
10
20
30
40
50
60
0 0.5 1 1.5 2
[Fructose-6-phosphate] mM
PF
K A
cti
vit
y
high [ATP]
low [ATP]
Sigmoidal dependence of reaction rate on [fructose-6-phosphate] is observed at high [ATP].
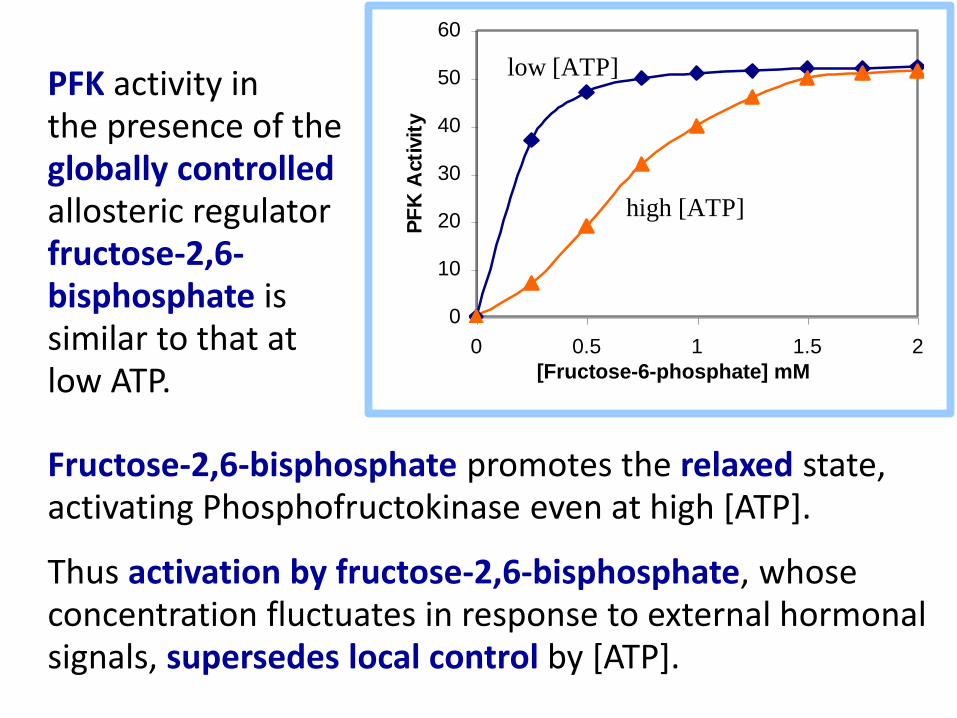
Fructose-2,6-bisphosphate promotes the relaxed state, activating Phosphofructokinase even at high [ATP].
Thus activation by fructose-2,6-bisphosphate, whose concentration fluctuates in response to external hormonal signals, supersedes local control by [ATP].
0
10
20
30
40
50
60
0 0.5 1 1.5 2
[Fructose-6-phosphate] mM
PF
K A
cti
vit
y
high [ATP]
low [ATP] PFK activity in the presence of the globally controlled allosteric regulator fructose-2,6-bisphosphate is similar to that at low ATP.

Summary of effects of glucagon-cAMP cascade in liver:
Gluconeogenesis is stimulated
Glycolysis is inhibited
Glycogen breakdown is stimulated
Glycogen synthesis is inhibited
Free glucose is formed for release to the blood
Glycogen Pyruvate
Gluconeogenesis
Glucose-1-P Glucose-6-P Glucose + Pi
Glucose-6-Pase
Glycolysis
Pathway
X
X

• the Cori Cycle operates during exercise
• for a brief burst of ATP utilization, muscle cells utilize ~P stored as phosphocreatine
• once phosphocreatine is exhausted, ATP is provided mainly by glycolysis, with the input coming from glycogen breakdown and from glucose uptake from the blood (Aerobic fat metabolism, discussed elsewhere, is more significant during a lengthy period of exertion such as a marathon run)
The Cori Cycle

Lactate produced from pyruvate passes via the blood to the liver, where it may be converted to glucose
The glucose may travel back to the muscle to fuel Glycolysis
Cori Cycle
Liver Blood Muscle
Glucose Glucose
2 NAD+ 2 NAD
+
2 NADH 2 NADH
6 ~P 2 ~P
2 Pyruvate 2 Pyruvate
2 NADH 2 NADH
2 NAD+ 2 NAD
+
2 Lactate 2 Lactate

The Cori cycle costs 6 ~P in liver for every 2 ~P made available in muscle. The net cost is 4 ~P.
Although costly in ~P bonds, the Cori Cycle allows the organism to accommodate to large fluctuations in energy needs of skeletal muscle between rest and exercise.
Cori Cycle
Liver Blood Muscle
Glucose Glucose
2 NAD+ 2 NAD
+
2 NADH 2 NADH
6 ~P 2 ~P
2 Pyruvate 2 Pyruvate
2 NADH 2 NADH
2 NAD+ 2 NAD
+
2 Lactate 2 Lactate

• the equivalent of the Cori Cycle also operates during cancer
• if blood vessel development does not keep pace with growth of a solid tumor, decreased O2 concentration within the tumor leads to activation of signal processes that result in a shift to anaerobic metabolism
The Cori Cycle

• energy dissipation by the Cori Cycle, which expends 6 ~P in liver for every 2 ~P produced via Glycolysis for utilization within the tumor, is thought to contribute to the weight loss that typically occurs in late-stage cancer even when food intake remains normal
Liver Blood Cancer Cell
Glucose Glucose
2 NAD+ 2 NAD
+
2 NADH 2 NADH
6 ~P 2 ~P
2 Pyruvate 2 Pyruvate
2 NADH 2 NADH
2 NAD+ 2 NAD
+
2 Lactate 2 Lactate