Embed Size (px)
Citation preview

ELSEVIER Molecular and Cellular Endocrinology 109 (1995) 183-188
Sex- and tissue- specific regulation of 1 l/%hydroxysteroid dehydrogenase rnRNA
A.L. Albiston, R.E. Smith, Z.S. Krozowski* Baker Medical Research Institute, Melbourne, 3181, Australia
Received 1 December 1994; accepted 1 February 1995
Abstract
In the present study we have investigated the effect of hypophysectomy on llgHSD1 gene expression in a variety of tissues from male and female rats. Of the tissues analysed significant changes were observed only in the male and female livers, and in the female kidney. Hypophysectomy (hypox) increased hepatic message levels in the female 20-fold, and male levels rose 2-fold. Male renal llpHSD1 message levels remained constant following hypophysectomy, whereas female renal llpHSD1 mRNA levels increased approximately 2-fold. In intact animals hepatic ll/?HSDl mRNA levels are l&fold lower in the female than the male, while renal levels are l.Cfold higher in the male than the female. Hepatic 11/3HSDl levels in the female rat appear critically responsive to the pattern of growth hormone (GH) administration. Twice daily injections of GH had no significant effect on hepatic message levels in the hypox female rat, whereas constant infusions of GH, via an osmotic mini-pump produced a marked decrease. Renal 1 lpHSD1 mRNA levels were demonstrated to be under the regulation of estrogen, and ll@HSDl enzyme activity in both liver and kidney correlated with the observed changes in mRNA levels. 1 lgHSD1 gene expression is thus regulated in a tissue-specific and sexually dimorphic manner.
Keywords: 1 l/3-Hydroxysteroid dehydrogenase; Gene expression; Rat; Hypophysectomy; Growth hormone
1. Introduction
Classically the enzyme 1 1Shydroxysteroid dehydro- genase (11pHSD) was identified as catalysing the con- version of active glucocorticoids to their inactive 1 1-keto metabolites (Monder and Shackleton, 1984). The primary function of this enzyme was hypothesised to be a protec- tive mechanism preventing glucocorticoids from binding to the mineralocorticoid receptor (MR) (Stewart et al., 1987; Edwards et al., 1988; Funder et al., 1988). How- ever, 1 l/?HSD activity is not limited to aldosterone re- sponsive tissues, but is almost ubiquitous in its distribu- tion. This conundrum was resolved with the discovery that two distinct 11/3HSD enzymes exist (Mercer and Krozowski, 1992; Walker et al., 1992; Rusvai and Naray- Fejes-Toth, 1993). The ubiquitous 11/3HSD type I en- zyme (11/3HSDl) can be readily differentiated from the tissue specific 1lpHSD type II enzyme (11/3HSD2) on the basis of cofactor specificity (Mercer and Krozowski, 1992; Walker et al., 1992; Rusvai and Naray, 1993).
* Corresponding author.
The more recently identified enzyme, 1 lpHSD2, is the candidate most likely to confer specificity on the MR. To date it has been identified in high abundance in aldoster- one responsive tissues (Walker et al., 1992), appears to co-localise with the MR in the kidney (Mercer and Kro- zowski, 1992), has a high affinity for corticosterone (Rusvai and Naray, 1993) and is NAD specific (Mercer and Krozowski, 1992; Rusvai and Naray, 1993). In con- trast 1 lpHSD1 activity (Monder and Lakshmi, 1990) and gene expression (Mercer et al., 1993) is found in most tissues, its K,,, for corticosterone is at least lOO-fold lower than that of 11/3HSD2 and it is NADP dependent (Lakshmi and Monder, 1988). Unlike llBHSD2, 1 l/IHSDl clearly has both reductase and dehydrogenase activities (Lakshmi and Monder, 1985). Based on this knowledge the role of 1laHSDl is now proposed to be one of modulation of glucocorticoid action in target tis- sues (Moore et al., 1993). Recently, highly homologous human and ovine llfiHSD2 species have been cloned (Agarwal et al., 1994; Albiston et al., 1994). These stud- ies clearly show that the ll/lHSDl and ll/?HSD2 en- zymes are products of separate genes.
0303-7207/95/$09.50 0 1995 Elsevier Science Ireland Ltd. All rights reserved SSDI 0303-7207(95)03501-W

184 A.L. Albiston et al. I Molecular and Cellular Endocrinology IO9 (1995) 183-188
Surprisingly ll#?HSDl gene expression does not ap- pear to be modulated by adrenocorticoids (Krozowski et al., 1990), but it has long been known that 11/3HSD ac- tivity is regulated by sex hormones. Lax et al. showed that hepatic microsomal 1lgHSD activity is much higher in the male rat than in the female (Lax et al., 1978). Gonadectomy decreases the level of activity in males and the administration of testosterone to gonadectomised male and female rats results in a significant increase in the level of enzymatic activity. In contrast, the administration of oestradiol to gonadectomised rats of either sex resulted in a decrease in activity. However, the caveat on the sex hormone regulation of hepatic 1lgHSD activity was that the animal must be pituitary intact, suggesting that a hy- pophysial factor is the primary modulator (Lax et al., 1974, 1978). This phenomenon of sexually divergent regulation is not specific for 1 l/IHSD but encompasses a wide range of hepatic steroid metabolising enzymes, in- cluding 3/3HSD, the cytochrome P450 isozymes including testosterone 16a-hydroxylase, androstanediol disulphate 15fi-hydroxylase and Sa-reductase, all of which are thought to be under the regulation of a pituitary ‘feminising factor’ in the liver (for review see Gustafsson et al., 1983). In this study we have investigated the regu- lation of 11/3HSDl mRNA expression in both male and female rats in a range of mineralocorticoid and glucocor- ticoid target tissues, and attempted to elucidate which factors regulate the expression of the gene.
2. Materials and methods
2.1. Animal experiments All experiments were performed on adult male and
female Sprague-Dawley rats (13 weeks old) maintained in 12 h light and 12 h dark cycles with ad libitum access to rat chow and water. Hypophysectomised (hypox) rats also had access to 0.9% NaCl and 5% dextrose. All ani- mals were killed by decapitation and tissues were re- moved and immediately placed in liquid nitrogen prior to storage at -70°C until required. Hypox Sprague-Dawley rats were obtained from Monash University School of Anatomy, Melbourne, Australia. Animals were used 3 weeks after surgery. Only those animals gaining less than 5 g in weight per week were used. Ovine growth hormone was a generous gift of Bunge Scientific, Mel- bourne, Australia. Cortisone acetate, dihydrotestosterone and 17/3-oestradiol were obtained from Sigma Chemical Company, St Louis, MO, USA. Thyroxine sodium (T4) was from Wellcome (Sydney, Australia). Osmotic mini pumps, model 2002, were obtained from Alzet Corpora- tion, Palo Alto, CA, USA. All hormone and drug treat- ments were over a 4-day period.
2.2. Experiments In all experiments each group had an n = 4, tissue
samples were analysed individually and results are pre-
sented as x + SEM. Statistical significance was deter- mined by analysis of variance (ANOVA).
Experiment I. Hypophysectomised rats were injected (SC.) twice daily with a combination of ovine GH (75 puglanimal), T4 (2pg/lOO g), cortisone acetate (5O~g/lOO g) and 17/Ioestradiol (6pg/animal) for fe- males or dihydrotestosterone for males (lOO~g/animal).
Experiment 2. Ovariectomized animals were treated twice daily with S.C. injections of 17&oestradiol (6 puglanimal).
Experiment 3. Intact male and hypophysectomised female rats were treated with a continuous infusion of ovine growth hormone (4 pg/h) by an osmotic mini-pump.
2.3. Messenger RNA analysis Total RNA was extracted by the method of Chirgwin
et al. (1979), and solution hybridization nuclease protec- tion assay performed to determine 1 l/?HSDl mRNA lev- els, as previously described (Krozowski et al., 1992). Glyceraldehyde-3-phosphate dehydrogenase mRNA lev- els (GAP) were also determined and used as an internal control. mRNA abundance was calculated with a scan- ning densitometer (LKB Ultroscan XL Laser Densitome- ter) and normalised by dividing 1 l/IHSDl values by the corresponding GAP signal.
2.4. Assay of 1 IflSDl activity Tissues samples were homogenised and protein con-
centrations determined by the Bradford assay, with 2OOyg of protein incubated for 12 min at 37’C in 10 mM phosphate buffer (pH 7.4) containing lo-* M [3H]cortico- sterone with or without cofactor (1 n&I). The reaction was stopped and the steroids extracted by the addition of 3 ~01s. of ethyl acetate. Separation of steroids was by thin layer chromatography using silica gel plates in a chloro- form/ethanol (92:8) system.
3. Results
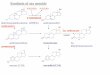
Message levels for 1 lpHSD1 were determined in liver, kidney, lung, heart, parotid, cerebellum, colon and hip- pocampus of intact, hypox and hypox plus hormone treated female rats, and in the liver, kidney, heart and lung of intact, hypox and hypox plus hormone treated male rats. Intact male rats were found to have 18-fold and 1 A-fold more 1 l#lHSDl mRNA in the liver and kidney, respectively, than intact female rats (Fig. 1). Of the above tissues only kidney and liver message levels were signifi- cantly altered by hypophysectomy. In hypox female rats 1 lPHSD1 mRNA levels rose 20- and 2-fold in the liver and kidney, respectively, compared with intact levels. Treatment of female hypox rats with a hormone cocktail restored renal 1 l#?HSDl mRNA to intact levels, but had no effect on the elevated hepatic levels. In contrast, in the

A.L. Albiston et al. I Molecular and Cellular Endocrinology IO9 (1995) 183-188 185
A
fl 1 lfi-HSDl
GAP
I Hypox +
Intact HYPOX Treated
1.0
E 0.8
9 9 0.6
= E 04 .
6 g 0.2
CA r r 0.0
C
I
0 INTACT 0 HYPOX
T w HYPOX TREATED
MALE FEMALE
B
fl 1 lfi-HSDl
GAP
I Hypox + Intact HYPOX Treated
I I 1 lf3-HSDl
I I
I 1.0
’ 0.8
9 s 0.6
E E 0.4
E; B 0.2
d ; 0.0
0 INTACT
D HYPOX n HYPOX TREATED
1 MALE FEMALE
Fig. 1. Effect of hypophysectomy and hormone replacement on 1 l#lHSDl mRNA levels in male and female adult rats. Sl nuclease protection analysis of 1 l/?HSDl and GAP mBNAs from (A) liver (B) kidney. Histogram of the above Sl protection assay (C) liver. +P c 0.01, ‘P < 0.001 versus intact male. *P < 0.001 versus intact female (D) kidney. +P < 0.01 versus intact female; *P < 0.01 versus hypox female.
male rats, renal 1 l#IHSDl mRNA levels were unaffected by hypophysectomy or the hormone cocktail while in the liver message levels increased 1.6-fold.
To further investigate sex-specific effects on 1 l/IHSDl gene expression, we studied the effect of oestrogens on renal and hepatic 1 1/3HSDl mRNA levels. As illustrated in Fig. 2 both hepatic and renal 1 lpHSD1 mRNA levels were significantly increased in ovariectomized (ovx) rats
9 1.5
E
% 3 1.0
f 0.5
6
$
-.- INTACT OVX OVX+ E2
compared with intact controls, with the magnitude of the increase similar to that observed in hypox rats. Treatment of ovx rats with estrogen returned renal message to intact levels, and decreased hepatic levels by 30%. The results of the hypox and the ovx experiments are consistent and suggest that estrogen modulates 11/3HSDl mRNA levels in the kidney independent of the pituitary gland. How- ever, in the liver estrogen did not affect 1 lgHSD1 mRNA
I3 +
r ” INTACT OVX OVX + E2
Fig. 2. Effect of ovariectomy and estrogen replacement on 1 lpHSD1 mRNA levels in (A) liver and (B) kidney. +P < 0.001 versus intact; *P < 0.01 versus ovx.

A.L. Albiston et al. I Molecular and Cellular Endocrinology 109 (1995) 183-188
* q MALE CONTROL q FEMALE CONTROL
ovx El ovx+E2
iJ0 CdFACTOR NADP NAD
Fig. 3. Changes in the levels of 11/3HSDl enzyme activity in the kidney following ovariectomy and estrogen replacement. Enzyme activities determined based on co-factor specificity: NADP+, ll/?HSDl and NAD+, 1 lr!?HSDZ. *P < 0.01, +P < 0.05 versus intact. ‘P < 0.05 versus
INTACT HYPOX HYPOX +GH
+
MALE FLLULE HYPOX HYPOX INTACT INTACT +OH
Fig. 4. Effect of a constant infusion of GH on hepatic llj3HSDl: (A) female mRNA levels. +P < 0.01 versus hypox. (B) Hepatic enzyme activity levels of intact mate, intact female, hypox female and hypox plus GH treated female. *P < 0.001 versus intact female, +P < 0.001 versus intact female. *P < 0.01 versus female hypox.
INTACT INTACT + GH
Fig. 5. Effect of a constant infusion of GH on male hepatic 11/3HSDl mRNA levels. +P < 0.01 versus control.
levels in the hypox rat, suggesting that the effects ob- served in ovariectomized rats were mediated via the pitui- tary. As shown in Fig. 3, changes in the level of renal 1 l/?HSDl (NADP dependent) enzyme activity follow the same pattern as observed for mRNA levels. It should be noted that the data in Fig. 3 are semi-quantitative due to the low initial substrate concentrations employed, and the fact that at the higher rates of corticosterone metabolism the percentage conversion of substrate is an under esti- mate of the true amount of enzyme activity present, due to limiting substrate conditions, and that the real amount of enzyme present is correspondingly higher, resulting in even larger differences between the groups. No signifi- cant differences in enzyme activity between sexes or treatments were observed when NAD+ was used as cofac- tor.
Since previous studies have shown that constant growth hormone (GH) infusion can modulate hepatic en- zymes in a sex specific manner (Gustafsson et al., 1983) we next sought to establish the role played by GH in modulating 1 l/lHSDl. Twice daily injection of GH to- gether with the other components of the cocktail, had no effect on 11/3HSDl message levels (see Fig. l), whereas administration of GH to female hypox rats by osmotic mini-pumps resulted in a significant decrease in levels of both mRNA and enzymatic activity compared with hypox rats (Figs. 4A,B). As in Fig. 3, the conditions of the assay have resulted in an underestimation of the true differences between experimental groups. Intact male rats treated in a similar manner showed a marked decrease in hepatic ll/?HSDl mRNA levels (Fig. 5) with renal message re- maining unaffected (results not shown).
4. Discussion
The present study clearly demonstrates a sexually dis- tinct and tissue specific pituitary-dependent regulation of ll/IHSDl gene expression. Of the eight tissues analysed in adult female rats (liver, kidney, lung, heart, parotid,

A.L. Albiston et al. I Molecular and Cellular Endocrinology 109 (1995) 183488 187
cerebellum colon and hippocampus) and four in males (liver, kidney, heart and lung) only the liver and kidney demonstrated significant changes in 11/3HSDl mRNA levels following hypophysectomy.
Male renal ll/IHSDl message levels remained con- stant following hypophysectomy, whereas female renal 1 l/?HSDl mRNA levels increased approximately 2-fold and returned to control levels after treatment with a hor- mone cocktail. The active hormone modulating female renal llj?HSDl message levels was subsequently shown to be estrogen since similarly elevated renal message levels were observed in ovx animals which returned to intact levels following estrogen administration. Previ- ously estrogen has been claimed to exert opposite effects on mRNA and 1 lj3HSDl activity in the female kidney, with message levels falling by 95% and activity increas- ing 70% (Low et al., 1993), however, in the present study estrogen halved both activity and mRNA levels. The dif- ferences observed between the two studies may reflect the differing doses used and the longer period (10 days) of hormone administration in the earlier study. For example, the effect of oestrogens on Sa-reductase activity has pre- viously been shown be dose dependent (Lax et al., 1979); in the present instance, since oestrogens have been shown to regulate the activity of another alcohol dehydrogenase by post-transcriptional mechanisms (Qulali and Crabb, 1992), it is possible that differing treatment regimes may differentially trigger one or both regulatory mechanisms.
The present study also shows that the mechanisms regulating 1 lgHSD1 and 11/3HSD2 enzyme activities appear to be different, with renal NAD dependent activity equivalent between the sexes and NADP activity mark- edly higher in the male kidney. The male kidney also ap- pears to possess higher levels of NAD than the female, since the addition of exogenous cofactor did not further increase enzyme activity in the male while female 1 lj3- HSD2 activity appear to increase.
A variety of genes have been shown to be tissue spe- cific in terms of their regulation by estrogen, of which retinol binding protein (Whitman et al., 1990) and orni- thine aminotransferase (Mueckler et al., 1984) are both modulated at the transcriptional level in the kidney, but not in the liver. An estrogen response element has not been identified in either the human or the rat 1 lj3HSDl genes (Tannin et al., 1991; Moisan et al., 1992), suggest- ing that the effect of the hormone may be indirect.
We have subsequently shown that at least one of the factors mediating the observed effects is the mode of GH secretion. These results reflect the differences between male and female patterns of hormone secretion. In the adult female rat a high pulse frequency results in a con- stant level of circulating GH; in contrast the male pitui- tary secrets GH episodically, resulting in periods where no GH is present in plasma. The present data showing the regulation of 1 l#IHSDl by estrogen in ovx, but not in hypox rats, are in agreement with previous studies on
enzyme:activity (Lax et al., 1978) and are consistent with the regulation of hepatic 1 l#IHSDl by GH. In the rat liver the two hormones have been shown to modulate dihy- drodiol dehydrogenase and 3-a-hydroxysteroid dehydro- genase activities independently of one another (Penning et al., 1992).
Previous studies have shown that the primary ‘feminising factor’ modulating hepatic steroidogenic en- zymes is the mode of GH release. Secretion of growth hormone by the pituitary is in part regulated by sex hor- mones, and administration of estrogen to male rats feminises the pattern of GH secretion, and in female rats testosterone masculinises it (Gustafsson et al., 1983) al- though how these steroids regulate the sex specific pattern of secretion remains unclear. The hypothalamic expres- sion of GH releasing factor and somatostatin are in part regulated by testosterone and estrogen (Hertz et al., 1989; Shirasu et al., 1990; Zeitler et al., 1990) but little is known about how these hormones modulate pituitary se- cretion patterns. The marked differences in hepatic levels between males and females, and the pituitary dependent nature of the phenomena, are also a feature of 38- hydroxysteroid dehydrogenase, and several hydroxyla- tion, sulphurylation and xenobiotic activities in the liver (Gustafsson et al., 1983; Naville et al., 1991). It is of in- terest to note that such enzymes are largely involved in the metabolism of sex hormones, whereas there is no evi- dence that 11/3HSDl metabolises these substrates.
A recent publication has indicated that 11/3HSDl may play a role in polycystic ovary disease (PCO) by indi- rectly modulating circulating androgen levels (Rodin et al., 1994). It has been suggested that increased levels of serum androgens in PC0 may be indirectly due to an in- crease in pituitary production of corticotropin, resulting from a lowering of circulating cortisol, a consequence of increased hepatic 1 lbHSD1 activity. A congenital defect in the reductase activity of 1 lj?HSD has also been shown to result in the appearance of secondary masculine fea- tures in women (Phillipou and Higgins, 1985) further suggesting a role for hepatic 1 laHSD1 in the modulation of androgen production.
The markedly lower levels of hepatic 1 lPHSD1 activ- ity in the female rat may result in higher levels of circulat- ing corticosterone than in the male providing the basis for a sex difference in glucocorticoid regulation of cortico- tropin (ACTH) production and thus adrenal androgen production. Studies by Trudel et al. (1991) demonstrated that in intact rats ACTH administration caused a marked increase in serum corticosterone levels with the amplitude of the response being markedly greater in the male com- pared to the female rat, suggesting a sex difference in adrenal cortical sensitivity to ACTH. Sex differences are also apparent in serum levels of both corticosterone and cortisol binding globulin (CBG) in the rat. Serum levels of CBG are 2.5-fold higher in the female than the male (Smith and Hammond, 1991; Mataradze et al., 1992) and

188 A.L. Albiston et al. I Molecular and Cellular Endocrinology 109 (1995) 183-188
are thought to be regulated by the sexually dimorphic mode of GH secretion (Mataradze et al., 1992) while cor- ticosterone levels were found to be approximately lo-fold higher in the female in one study (Trudel et al., 1991).
In conclusion the results presented in this paper show that the large differences in hepatic 1 l/?HSDl enzyme levels between male and female rats reflect comparable differences in message levels. The sex differences in gene expression are due to the different patterns of growth hormone secretion, and are specific to the liver.
Acknowledgments
The authors are in debt to Professor John Funder for helpful discussion throughout the course of the study and for critically reviewing the manuscript. This work was supported by a block grant from the NHMRC of Austra- lia.
References
Agarwal, A.K., Mune, T., Monder, C. and White, PC. (1994) J. Biol. Chem. 269,25952-25962.
Albiston, A.L., Obeyesekem, V.R., Smith, R.E. and Krozowski, Z.S. (1994) Mol. Cell. Endocrinol. 105, Rll-R17.
Chirgwin, J.M, Pryzbyla, A.E, MacDonald, R.J and Rutter, W.J. (1979) Biochemistry 18.5294-5299.
Edwards, C.R., Stewart, P.M., Burt, D., Brett, L., McIntyre, M.A., Sutanto, W.S., deKloet E,R. and Monder, C. (1988) Lancet 2,986 989.
Funder, J.W., Pearce, P.T., Smith, R. and Smith, AI. (1988) Science 242.583-585.
Gustafsson, J.A., Mode, A., Norstedt, G. and Skett, P. (1983) Annu. Rev. Physiol. 45,51-60.
Hertz, P., Silbermann, M., Even, L. and Hochberg, Z. (1989) Endocri- nology 125.581-585.
Krozowski, Z., Obeyesekere, V., Smith, R. and Mercer, W. (1992) J. Biol. Chem. 267.2569-2574.
Krozowski, Z., Stuchbery, S., White, P., Monder, C. and Funder, J.W. (1990) Endocrinology 127,3009-3013.
Lakshrni, V. and Monder, C. (1985) Endocrinology 116.552-560. Lakshmi, V. and Monder, C. (1988) Endocrinology 123.2390-2398. Lax, E.R., Ghraf, R. and Schriefers, H. (1978) Acta Endocrinol.
(Copenhagen) 89,352-358.
Lax, E.R., Hoff, H.G., Ghraf, R., Schroder, E. and Schriefers, H. (1974) Hoppe Seyler’s Z. Phys. Chem. 355, 1325-1331.
Lax, E.R., Kreuzfelder, E. and Schriefers, H. (1979) Hoppe Seyler’s Z. Phys. Chem. 360, 1807-1817.
Low, S.C., Assaad, S.N., Rajan, V., Chapman, K.E., Edwards, C. and Se&l, J.R. (1993) J. Endocrinol. 139.27-35.
Mataradze, G.D., Kurabekova, R.M. and Rozen, V.B. (1992) J. Endo- crinol. 132,235-240.
Mercer, W., Obeyesekere, V., Smith, R. and Krozowski, Z. (1993) Mol. Cell. Endocrinol. 92, 247-251.
Mercer, W.R. and Krozowski, Z.S. (1992) Endocrinology 130, 540- 543.
Moisan, M.P., Edwards, C. and Se&l, J.R. (1992) Mol. Cell. Endocri- nol. 6, 1082-1087.
Monder, C. and Lakshmi, V. (1990) Endocrinology 126,2435-2443. Monder, C. and Shackleton, C.H (1984) Steroids 44.383417. Moore, C., Mellon, S.H., Murai, J., Siiteri, P.K. and Miller, W.L.
(1993) Endocrinology 133,368-375. Mueckler, M.M,, Moran, S. and Pitot, H.C, (1984) J. Biol. Chem. 259,
2302-2305. Naville, D., Keeney, D.S., Jenkin, G., Murray, B.A., Head, J.R. and
Mason, I.J. (1991) Mol. Cell. Endocrinol. 5, 1090-1100. Penning, T.M., Isaacson, K. and Lyme, CR. (1992) Biochem. Pharma-
col. 43, 1148-1151. Phillipou, G. and Higgins, B.A. (1985) J. Steroid Biochem. 22, 435-
436. Qulali, M. and Crabb, D.W. (1992) Arch. Biochem. Biophys. 297,277-
284. Rodin, A., Thakkar, H., Taylor, N. and Clayton, R. (1994) N. Engl. J.
Med. 330,46&465. Rusvai, E. and Naray, F.T.A. (1993). J. Biol. Chem. 268, 10717-
10720. Shirasu, K., Stumpf, W.E. and Sar, M. (1990) Endocrinology 127, 344-
349. Smith, C.L. and Hammond, G.L. (1991) Endocrinology 128,983-988. Stewart, P.M. Wallace, A.M, Valentino, R., Burt, D., Schakleton, C.H
and Edwards, C.R.W. (1987) Lancet ii, 821-824. Tannin, G.M., Agarwal, A.K., Monder, C., New, M.I. and White, P.C.
(1991) J. Biol. Chem. 266. 16653-16658. Trudel, C:, Couet, J., Martel, C., Labrie, C. and Labrie, F. (1991) Endo-
crinology 129,2077-2084. Walker, B.R., Campbell, J.C., Williams, B.C. and Edwards, C.R.
(1992) Endocrinology 131,970-972. Whitman, M.M., Hamish, D.C., Soprano, K.J. and Soprano, D.R.
(1990) J. Lipid Res. 31, 1483-1490. Zeitler, P., Argente, J., Chowen, B.J., Clifton, D.K. and Steiner, R.A.
(1990) Endocrinology 127, 1362-1368.