Embed Size (px)
Citation preview

Sistemul imunitar (S.I.) – caracterizare generală, proprietăţi
Sistemul imunitar este un organ difuz, sui generis, de aprox. 900 g la om, adesea ignorat
datorită dispersiei sale în organism, ca celule şi molecule libere în umorile organismului şi fixe, în
ţesuturi şi organe limfoide, care alcătuiesc sistemul limfoid.
Numeroase date din patologia S.I. demonstrează că este un organ vital, fără de care
organismul nu poate supravieţui, în condiţiile în care acesta este supus în permanenţă unor agresiuni
(virusuri, bacterii, microfungi, paraziţi). Deşi numărul celulelor (~1012 limfocite) şi moleculelor
( ~1020 molecule de anticorpi) sistemului imunitar este imens, acesta nu reflectă potenţialul real de
apărare, deoarece după stimularea antigenică are loc o amplificare enormă a unor subpopulaţii sau
clone de limfocite şi sinteza unui număr enorm de molecule de anticorpi specifici, la care se adaugă
şi potenţialul de refacere şi mobilizare; de ex., un om produce zilnic aprox. 10 miliarde de limfocite.
Celulele şi moleculele S.I. au capacitatea de circulare şi recirculare prin reţeaua vaselor de sânge şi
limfatice, astfel că «patrulează » şi asigură detectarea substanţelor străine, mobilizându-se pentru
anihilarea acestora.
Teoretic, capacitatea de recunoaştere este imensă, respectiv ~1030 molecule diferite structural
care formează «repertoriul» sistemului imunitar, reprezentat de unităţi de recunoaştere şi cuplare a
antigenului, codificate genetic ca receptori celulari şi paratopi (situsurile combinative ale
anticorpilor). Repertoriul real este mai mic (107 – 1011, după diferiţi autori), oricum este imens, cu
mult mai mare decât numărul proteinelor din organism. Acest repertoriu dă organismelor
posibilitatea de a recunoaşte nu numai toate antigenele naturale, dar chiar orice altă substanţă
artificială, sintetizată până în prezent sau care ar putea fi sintetizată în viitor. Combinarea specifică
dintre Ag şi Ac a fost adesea descrisă ca o reacţie de tip lacăt-cheie, în care cheia este Ac-ul, iar
regiunea complementară acesteia, respectiv lacătul este Ag-ul (sau mai precis, un epitop din
structura acestuia); în aceşti termeni, ar însemna că repertoriul receptorilor de pe limfocitele T şi al
moleculelor de Ig formează un set imens de chei pregătite dinainte, între care există o cheie potrivită
pentru orice lacăt teoretic posibil. Explicatia: capacitatea genelor care codifica Ig-le de a se combina
şi de a suferi mutaţii somatice în cursul proliferării şi diferenţierii dependente de Ag a limfocitelor B,
care vor avea o informaţie genetică (pentru regiunile variabile ale moleculelor de recunoaştere: Ac
liberi şi receptori), diferită de cea a celulelor-stem de origine.
1 1

Proprietăţile sistemului imunitar.
Organizarea S.I. în plan biologic este teleonomică, în sensul că este perfect organizat pentru
a-şi îndeplini funcţia sa esenţială: diferenţierea self-ului de non-self. Sistemul imunitar are
următoarele particularităţi:
1. Are o structură în reţea, ai cărei constituenţi sunt situaţi mai ales la zonele de contact cu
substanţele străine, la porţile de intrare în organism. În această reţea, celulele sistemului
imunitar îşi îndeplinesc rolul fiziologic prin interacţiuni cu alte celule şi mecanisme
fiziologice, inclusiv cu cele nespecifice, neadaptative, acestea reprezentând prima linie de
apărare (Fig. 1), pe când cele specifice, adaptative, fac parte din linia a II-a de aparare, care
acţionează după o perioadă de latenţă.
Fig. 1. Reprezentarea schematică a primei linii de apărare a organismului (după Roitt, 1991).
2. Recepţionează informaţii din mediul extern, le recunoaşte şi reacţionează faţă de
acestea, ocupând din acest punct de vedere, locul al II-lea după sistemul nervos. Sistemul
imunitar apare ca un ,,organ de simţ,, pentru stimuli (de natură chimică - macromolecule)
care nu sunt recunoscuţi de sistemul senzorial clasic.
3. Celulele specifice ale sistemului imunitar poartă pe suprafaţa lor un număr mare de
receptori membranari. De ex., o clonă de limfocite are receptori pentru Ag, cu situsuri de
combinare identice, care diferă de receptorii altor clone; rezultă că o clonă diferă de alta şi
prin natura antigenului pe care îl poate recunoaşte şi de care poate fi activată.
4. Capacitatea de a răspunde la toate antigenele pe care virtual le poate întâlni în natură,
datorată repertoriului imens de subpopulaţii sau clone diferite, fiecare purtând receptori
pentru anumite antigene, fiind deci imunocompetente. După pătrunderea în organism, un
antigen interacţionează, teoretic, numai cu o clonă limfocitară, deci cu un număr mic de
2 2

celule. Din acest punct de vedere există o redundanţă, care este benefică, pentru că deşi
numărul celulelor imunocompetente este mult mai mare, nu sunt utilizate decât câteva la un
moment dat, astfel ca apărarea organismului se realizează cu mare economie de mijloace.
După stimularea realizată prin contactul cu antigenul, clona respectivă este activată şi intră
într-un proces de proliferare intensă şi de diferenţiere funcţională, celulele devenind
efectoare, angajate (engl. commited), Ag-reactive. La baza răspunsului imun se află selecţia
clonală, care determină marea eficienţă a acestui răspuns, derivată din capacitatea de
amplificare a efectelor sale, consecutivă stimulării antigenice (fig. 2).
5. Activitatea de supraveghere a sistemului imunitar este facilitată de capacitatea de circulare
şi recirculare a limfocitelor, din sânge în organele limfoide secundare (ganglioni limfatici,
splină, amigdale, plăci Peyer) care reprezintă locul de întâlnire dintre limfocite şi Ag, şi
invers.
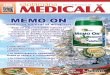
6. Memoria imunologică, netransmisibilă ereditar, determină un răspuns imun mai rapid, mai
intens, mai durabil, la a doua întâlnire şi urmatoarele, cu acelaşi antigen = răspuns
anamnestic sau fără uitare (gr. ana-mnesis = amintirea celor din trecut sau an-a-mnesis=ne-
ne-uitare) (fig. 3), pe această proprietate bazându-se procedeul imunoprofilaxiei prin
vaccinare.
CEL.
STEM
Proliferare
Maturare
LT LB Repertoriul de celule imunocompetente,
înainte de contactul cu Ag OOOOOOOOOOOOOOO = Clone de LT şi LB
Ag stimulare SELECŢIA CLONALĂ antigenică
Proliferare selectivă Diferenţiere functională Clona de limfociteAg-specifice
Fig. 2. Schema procesului de selecţie clonală a limfocitelor imunocompetente, consecutiv stimulării antigenice.
3 3
Fig. 3. Dinamica răspunsului imun primar şi secundar.Legenda: ______ Nivelul total al Ig-lor
----------- Titrul IgM _ο_ο_ο_ Titrul IgG
7. Sistemul imunitar al vertebratelor poate fi asimilat celorlalte sisteme şi poate fi
considerat integrat acestora, sistemului nervos în primul rând, ceea ce explică scăderea
capacităţii de apărare a sistemului imunitar, sub influenţa stresului prelungit. De asemenea,
orice disfuncţie a sistemului imunitar, perturbă funcţiile celorlalte sisteme. În medicină se
foloseşte expresia de «status imunologic», investigat şi în cazul unor boli ale altor sisteme,
deoarece este dovedit faptul că multe afecţiuni au o componentă imună.
De altfel, există numeroase analogii între sistemul imunitar şi sistemul nervos: complexitate
funcţională şi structură în reţea (~ creier mobil), capacitatea de a răspunde diferenţiat la o varietate
de stimuli, o serie de dichotomii şi dualităţi: ambele sisteme primesc şi transmit semnale, utilizează
semnale excitatorii şi inhibitorii, prezenţa unor organe limfoide primare şi secundare (similar celor
nervoase: centrale şi periferice).
Răspunsul imun este o reacţie adaptativă a sistemului imunitar; spre deosebire de mecanismele
de rezistenţă naturală sau imunitate înnăscută, răspunsul imun are caracter adaptativ, fiind îndreptat
spre o anumită substanţă non-self şi realizat prin mobilizarea anumitor celule preprogramate, care
«aşteaptă» să fie activate de un anumit antigen specific.
COMPONENTA UMORALĂ SPECIFICĂ A IMUNITĂŢII
Anticorpii (Ac). Bazele moleculare ale răspunsului imun
Imunoglobulinele = glicoproteine a căror sinteză este indusă de prezenţa în organism a unor
substanţe straine sau antigenice, care au functie de anticorpi şi rol determinant în reacţiile de
imunitate umorală.
4 4
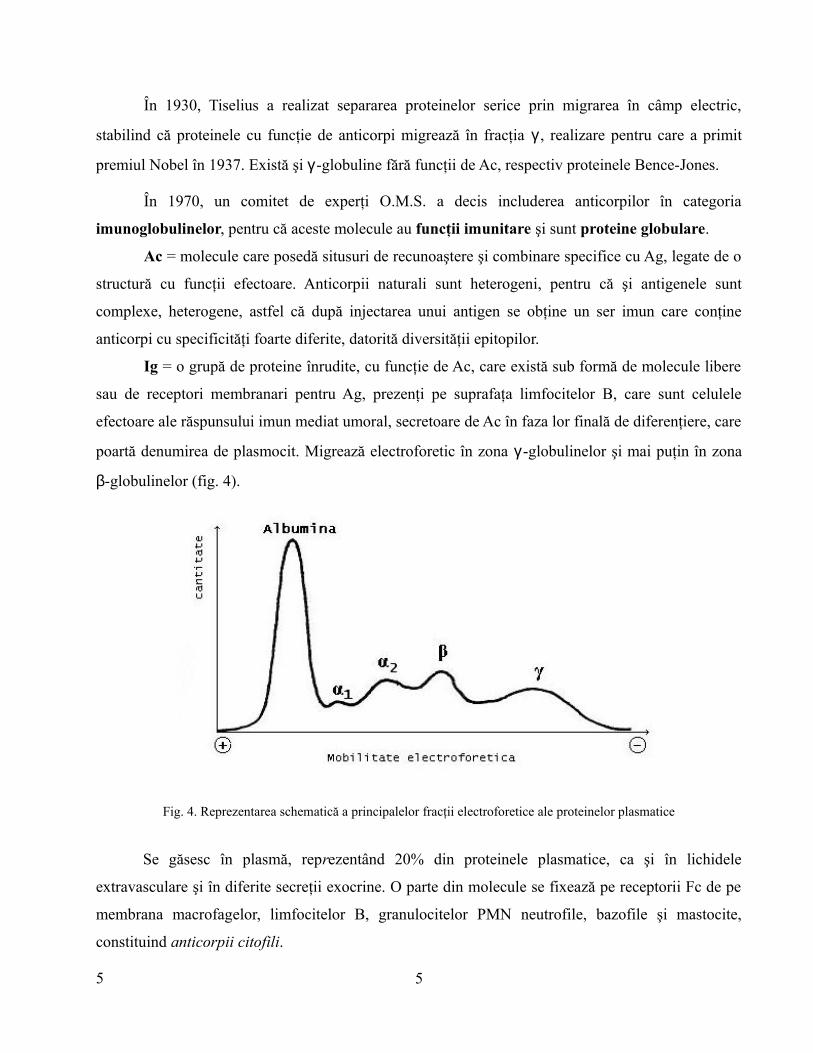
În 1930, Tiselius a realizat separarea proteinelor serice prin migrarea în câmp electric,
stabilind că proteinele cu funcţie de anticorpi migrează în fracţia γ , realizare pentru care a primit
premiul Nobel în 1937. Există şi γ -globuline fără funcţii de Ac, respectiv proteinele Bence-Jones.
În 1970, un comitet de experţi O.M.S. a decis includerea anticorpilor în categoria
imunoglobulinelor, pentru că aceste molecule au funcţii imunitare şi sunt proteine globulare.
Ac = molecule care posedă situsuri de recunoaştere şi combinare specifice cu Ag, legate de o
structură cu funcţii efectoare. Anticorpii naturali sunt heterogeni, pentru că şi antigenele sunt
complexe, heterogene, astfel că după injectarea unui antigen se obţine un ser imun care conţine
anticorpi cu specificităţi foarte diferite, datorită diversităţii epitopilor.
Ig = o grupă de proteine înrudite, cu funcţie de Ac, care există sub formă de molecule libere
sau de receptori membranari pentru Ag, prezenţi pe suprafaţa limfocitelor B, care sunt celulele
efectoare ale răspunsului imun mediat umoral, secretoare de Ac în faza lor finală de diferenţiere, care
poartă denumirea de plasmocit. Migrează electroforetic în zona γ -globulinelor şi mai puţin în zona
β-globulinelor (fig. 4).
Fig. 4. Reprezentarea schematică a principalelor fracţii electroforetice ale proteinelor plasmatice
Se găsesc în plasmă, reprezentând 20% din proteinele plasmatice, ca şi în lichidele
extravasculare şi în diferite secreţii exocrine. O parte din molecule se fixează pe receptorii Fc de pe
membrana macrofagelor, limfocitelor B, granulocitelor PMN neutrofile, bazofile şi mastocite,
constituind anticorpii citofili.
5 5

În afară de moleculele de Ig (Ac liberi), în organism există şi alte molecule de recunoaştere:
receptorii celulari de pe LT (TCR), receptorii imunoglobulinici de pe LB – cu rol de receptori pentru
Ag, moleculele CMH cls. I şi II etc. Toate aceste molecule alcătuiesc superfamilia Ig-lor, toate având
în structura lor 1 – n fragmente sau domenii polipeptidice, identice cu domeniul, ca unitate de bază a
moleculelor de Ig.
Structura imunoglobulinelor
Anticorpii constituie cele mai heterogene proteine cunoscute, dar în ciuda diversităţii lor,
unitatea de structură moleculară este caracteristică, tipică. Diversitatea se manifestă mai ales la
nivelul unor situsuri cu structură chimică diferită de la Ac la Ac, care determină specificitatea de
reacţie cu Ag-ul care le-a determinat sinteza. Structura imunoglobulinelor a fost descifrată de către
R. Porter şi G. Edelman, descoperire pentru care în 1972 au fost distinşi cu premiul Nobel.
Modelul general de structură al monomerilor de Ig a fost descifrat pe IgG, care reprezintă ~
80% din cantitatea totală de Ig. Autorii au scindat prin proteoliză enzimatică limitată moleculele de
Ig în fragmente şi lanţuri, care au fost studiate biochimic şi funcţional.
De ex., prin hidroliză enzimatică cu papaină s-au obţinut 3 fragmente (fig. 5):
Papaina
1) IgG 2 fragmente Fab (fragment antigen binding)
1 fragment Fc (fragment crystalisable; cytotropic, complement binding);
care cristalizeaza prin păstrare la 4ºC
Fig. 5. Scindarea enzimatică a moleculei de IgG sub acţiunea papainei.
Fab – g.m. 45kDa; situat la capatul NH2 terminal al lanturilor polipeptidice;
- leaga Ag, dar nu formeaza aglutinate si precipitate vizibile;
- monovalent;
6 6

Fc - g.m. 50kDa; situat la capatul COOH terminal al lanturilor polipeptidice;- leaga complementul;- se poate lega citofil la receptorii pentru Fc de pe membrana unor celule;
Sub acţiunea pepsinei se obţine un singur fragment (fig. 6):
Fig. 6. Scindarea enzimatică a moleculei de IgG sub acţiunea pepsinei.
Pepsina2) IgG F(ab’)2 – g.m. 100 kDa (fara Fc)
- fragment bivalent, care leagă Ag şi participă în calitate de Ac complet la reacţiile de aglutinare şi precipitare;
Reducerea –S-S-3) IgG 4 lanţuri polipeptidice: - 2 lanţuri grele –H (engl. heavy)- g.m. 50kDa Alchilare (2-mercaptoetanol + uree) - 2 lanţuri uşoare- L (engl. light)- g.m. 25kDa
- niciunul dintre lanţuri nu poate lega singur Ag, fiind necesară legarea lor 2 câte 2 (L+H);- cele 4 lanţuri sunt unite între ele prin punţi disulfidice –S-S-, formate la nivelul resturilor de
cisteină; intra- şi intercatenare;- legaturile –S-S- sunt esenţiale pentru asigurarea structurii tridimensionale a moleculei şi pentru
funcţionalitatea sa;- lanţurile L sunt comune pentru toate clasele de Ig; din punct de vedere antigenic sunt însă de tip
k sau λ;- în moleculele de Ig ambele lanţuri L sunt de acelaşi tip; 65% dintre moleculele de Ig umane
conţin lanţuri Lk; la şoarece- 95% - Lk.Lanţurile H conţin 450 – 600 AA; regiunea CH are 3 - 4 domenii alcătuite din 100 –110 AA:
CH1, CH2, CH3, (CH4). Ig din clasele G, A, D au 3 domenii C, iar clasele M şi E au 4 domenii
C.Domeniile se pot nota şi astfel Cγ1, Cγ2, Cγ3 –pentru IgG ş.a.m.d. Lanţurile H – se deosebesc prin
proprietăţile antigenice, conferind caracterul de “clasă” al Ig-lor :
- clasa IgG – lanţ γ - clasa IgM – lanţ µ - clasa IgA – lanţ α - clasa IgD – lanţ δ - clasa IgE - lanţ ε
7 7

Lanţurile L conţin 214 AA, au o g.m. de 25 kDa, se compun dintr-un domeniu variabil VL şi
unul constant CL, fiecare de 107 resturi de AA. Întotdeauna se ataşează Vk+Ck şi Vλ+Cλ.
Analiza structurală şi chimică a lanţurilor moleculelor de Ig s-a realizat folosind populaţii moleculare
omogene, de la bolnavi cu mielom multiplu (plasmocitom)- stare patologică caracterizată printr-o sinteză
crescută de molecule de Ig normale – uşor de purificat şi analizat (~ 95%). In alte stări patologice se
sintetizează în exces molecule patologice de Ig; de ex. în mielomul micromolecular sinteza de lanţuri L în
exces, conduce la dimerizarea şi eliminarea lor prin urină (proteine Bence - Jones); în boala lanţurilor grele
lipsesc AA care formează domeniul CH1.
Analiza cristalografică în raze X a Ig-lor a relevat faptul că lanţurile polipeptidice nu sunt
secvenţe lineare, ci pliate, formând regiuni globulare compacte numite domenii, spaţial distincte, cu
o structură tridimensională foarte asemănătoare (fig. 7).
Fig. 7. Structura pe domenii a moleculei de IgG, generate prin formarea de legături disulfidice intracatenare (http:www.medicine.uiowa.edu+martin.lab).
La bifurcaţia moleculei în Y se află regiunea „balama”, compusă din 15 AA şi 2 legături -S-
S-, care asigură legătura între cele 2 perechi de lanţuri şi simetria moleculei. Regiunea este sensibilă
la acţiunea enzimelor, datorită resturilor de prolină, care împiedică formarea structurii α-helicoidale.
Este regiunea care uneşte Fab şi Fc, rezultând un complex molecular unic.
Regiunea balama este formată dintr-un octopeptid ciclic, rigid, care acţionează ca un pivot al
părţii flexibile a regiunii, enzimele proteolitice (papaina, tripsina) clivând molecula de Ig înaintea
pivotului. IgA este mai puţin sensibilă la atacul enzimatic, datorită bogăţiei de HC din vecinătate. La
8 8

clasele IgM şi IgE această regiune lipseşte, fiind prezent un domeniu suplimentar, care este
considerat precursorul său evolutiv.
Regiunea ,,balama,, are un rol important şi în flexibilitatea şi funcţionalitatea moleculei, în
sensul că permite fragmentelor Fab orientarea în spaţiu, rotirea, desfacerea, apropierea braţelor în
funcţie de spaţierea epitopilor, asigurând eficienţa de legare a acestora.
Prin regiunea „balama”, se trasmit din fragmentele Fab (de la situsurile combinative), în
fragmentul Fc efectele complexării moleculei de Ig cu Ag, având deci şi un rol de transductor de
semnale informaţionale. În absenţa acestei regiuni, moleculele de Ig îşi pierd funcţia, astfel că prin
legarea directă a Fab de CH2 rezultă molecule rigide în forma literei T, cu funcţii alterate (aceste
molecule sunt incapabile să lege C1q, să interacţioneze cu receptorii Fc de pe membrana leucocitelor
PMNN, a limfocitelor B, au o slabă afinitate pentru cei de pe membrana macrofagelor, au o
capacitate redusă de legare de sinciţiotrofoblaştii placentari, dar îşi păstrează capacitatea de legare a
proteinei A stafilococice - SpA).
Situsurile combinative ale anticorpilor (S.C.) sau paratopii sunt situaţi la extremităţile NH2-
terminale ale fragmentelor Fab. Reprezintă locul prin care molecula de Ig recunoaşte şi leagă specific Ag.
Paratopul este alcătuit din regiunile variabile VH+VL, fiind diferit ca formă, dimensiuni. O moleculă de
IgG (monomer) are două situsuri combinative (paratopi). Situsul combinativ conţine 3 regiuni hipervariabile,
notate CDR (regiuni determinante de complementaritate), care sunt inserate între 4 segmente notate Fr (engl.
frame work = regiuni cadru), conservate în cursul evoluţiei. În aceste zone este prezent glicocolul, care
contribuie la invariabilitatea lor. Prin plierea lanţurilor H şi L, zonele hipervariabile de pe lanţul H se apropie
de cele de pe lanţul L, astfel încât formează o unitate funcţională, respectiv situsul combinativ sau paratopul.
Prin cercetări de radiocristalografie s-a demonstrat că suportul molecular al reacţiei Ag-Ac este reprezentat de
interacţia dintre D.A., cu anumiţi aminoacizi din regiunile hipervariabile ale moleculei de Ig. Ceilalţi
aminoacizi aparţin domeniilor variabile, deşi nu sunt implicaţi direct în legarea antigenului, îndeplinesc un rol
foarte important, funcţionând ca un schelet pentru plierea catenelor regiunilor variabile şi pentru păstrarea
integrităţii S.C. Secvenţa aminoacizilor în CDR este variabilă, numărul specificităţilor rezultate fiind
destul de mare pentru a corespunde tuturor epitopilor existenţi în natură.
Fragmentul Fc este alcătuit din domeniile constante CH2 şi CH3 ale lanţurilor H, unite prin
punţi disulfidice (un domeniu suplimentar CH4 este prezent la Ig M şi IgE). La un pH acid (2,5)
regiunea dintre domeniile CH2 şi CH3 devine sensibilă la clivajul enzimatic, iar la pH neutru
redevine rezistentă. Dacă prin hidroliza enzimatică cu papaină a moleculei de IgG se obţin 2
fragmente Fab şi un fragment Fc, prin acţiunea prelungită a acestei enzime, pe lângă fragmentele
9 9

Fab se obţine şi un Fragment Fc’ sau pFc (pepsic). Aşa s-a demonstrat că domeniul CH2 leagă
complementul şi este responsabil de rata catabolismului întregii molecule.
Moleculele de Ig reprezintă sediul unei dualităţi funcţionale: fragmentele Fab leagă specific
antigenul, datorită variabilităţii lor, pe când fragmentele Fc, caracterizate prin constanţă, conferă
moleculelor de Ig posibilitatea de a prezenta antigenele fixate de Fab, sistemelor celulare şi umorale
de distrugere şi eliminare.
Glucidele din structura Ig-lor (glicoproteine)
Toate clasele de Ig au o componentă glucidică (oligozaharidică) localizată în regiunea
constantă CH2, sub forma unor catene laterale simple sau complexe care reprezintă 3 – 13% din
greutatea moleculelor. Sunt secvenţe foarte heterogene, reprezentate de glucozamină (GlcN),
glucoză, galactozamină (GalN), acid sialic, fucoză, manoză, care se adaugă secvenţial pe parcursul
sintezei Ig-lor. Legarea începe înainte ca sinteza să fie completă şi se încheie la secreţia Ig-lor (la
trecerea din RER în aparatul Golgi, sub acţiunea transglucozidazelor membranare nespecifice).
Prezenţa lor este corelată, se pare, cu procesul de transport transmembranar şi de secreţie a Ig-lor.
Glucidele din structura glicoproteinelor, în general, conferă acestora următoarele proprietăţi:
- cresc solubilitatea proteinelor;- induc şi menţin lanţurile peptidice într-o conformaţie tridimensională funcţională;- protejează proteinele faţă de atacul enzimatic;- scad imunogenitatea;- controlează timpul de înjumătăţire al glicoproteinelor solubile şi celulare.Aşa cum s-a arătat, analiza cristalografică în raze X a Ig-lor a demonstrat că lanţurile
polipeptidice nu sunt lineare ci pliate, ca regiuni globulare, compacte, numite domenii, spaţial
distincte, cu structură 3-dimensională asemănătoare şi rezistente la proteoliză. Domeniul se înscrie
într-un cilindru cu diametrul de 3,5 nm şi este format din 60-70 aminoacizi, fiind stabilizat de o
punte –S-S- situată în regiunea centrală a unei unităţi de omologie formată din 110 aminoacizi. Toate
domeniile analizate au prezentat un model unic de pliere antiparalelă a lanţurilor polipeptidice, numit
β-pliere, pliere–Ig (engl. Ig-folding).
Domeniile fiecărui lanţ nu sunt independente de cele din lanţul opus, ci formează perechi de
unităţi de omologie care se constituie în unităţi structurale numite module (de ex., VL şi VH izolate
au slabă capacitate de a lega Ag, dar VL + VH formează situsul combinativ = paratop, cu mare
afinitate pentru Ag. Au fost descrise 2 tipuri de interacţiuni între domenii adiacente:
10 10

Heterogenitatea Ig-lorIg sunt foarte heterogene, în privinţa încărcăturii electrice şi a migrarii electroforetice,
specificităţii de Ac, caracterului antigenic etc. Heterogenitatea antigenică se relevă după inocularea
la alte organisme în care se comportă ca Ag, determinând sinteza de Ac anti-Ig. Această enormă
diversitate a Ig-lor este generată de secvenţa aminoacizilor de la nivelul diferitelor domenii, care
determină diferenţe antigenice şi existenţa următoarelor specificităţi:
1) Specificitatea sau heterogenitatea izotipică;
2) ,, ,, alotipică;
3) ,, ,, idiotipică.
1) Heterogenitatea izotipică a imunoglobulinelor
Reprezintă caracterul antigenic al moleculelor de Ig caracteristic tuturor indivizilor unei specii.
Pe baza acestei specificităţi, în cadrul aceleiaşi specii sunt mai multe clase şi subclase de molecule
de Ig, care diferă prin proprietăţile lor antigenice şi funcţionale. La mamifere se cunosc 5 clase de Ig
( fig. 8), unele diferenţiindu-se în subclase.
Fig. 8. Structura celor cinci clase de Ig (sferele indică localizarea determinanţilor antigenici izotipici ai lanţurilor H) .
De ex., clasa IgG are 4 subclase, lanţurile H ale acestora fiind notate cu γ1 – γ4. IgA şi IgM au
câte două subclase. La variantele antigenice izotipice contribuie şi frecvenţa răspândirii lanţurilor
uşoare k şi λ. Diferenţele antigenice ale lanţurilor H sunt semnificative, gradul de omologie fiind de
40%. Acesti determinanţi antigenici localizaţi în domeniile constante sunt comuni indivizilor unei
specii şi se manifestă ca antigene după inocularea într-un organism aparţinând altei specii. De ex.,
dacă se inoculează IgG umane la iepure, se obţine un antiser cu Ac anti-IgG umane, care va precipita
toate subclasele de IgG umane sau, un antiser preparat pe iepure faţă de lanţurile λ umane, va
11 11

reacţiona şi va precipita Ig-le umane din orice clasă care conţin lanţuri Lλ. Clasele şi subclasele de Ig
au proprietăţi structurale, antigenice, dar şi funcţionale diferite:
Clasa IgG – Ig-le din această clasă constituie tipul reprezentativ de Ig, fiind cel mai
bine cunoscute Ig; reprezintă 70-75% din totalul Ig-lor serice, astfel că sunt uşor de obţinut în stare
pură, de studiat biochimic şi funcţional. Formula moleculară: γ 2k2 sau γ 2λ2. Constanta de
sedimentare este de 7S, g.m. 150 kDa, concentraţia serică variind între 800– 2000 mg%. Moleculele
de IgG sunt termorezistente, nefiind denaturate la 75ºC, timp de 30min; au un mic procent de glucide
(2 –3%), un timp de înjumătăţire (T 1/2) lung, de 21-23 zile (mai redus la IgG3). O concentraţie mare
de IgG este prezentă în infecţiile cronice.
Moleculele IgG reprezintă anticorpii răspunsului imun secundar, fiind sintetizaţi după stimularea
antigenică secundară sau în cursul răspunsului primar, după comutarea clasei. Sunt anticorpii tipic
precipitanţi, fiind principalii Ac cu rol în neutralizarea toxinelor bacteriene şi a virusurilor. Mediază
citotoxicitatea anticorpo-dependentă, ca şi fagocitoza opsonică (imună), după interacţiunea cu
receptorii Fc de pe suprafaţa fagocitelor. IgG leagă C1q şi activează sistemul complement pe calea
clasică (cu excepţia subclasei IgG4), leagă proteina A stafilococică (SpA), reacţionează cu factorii
reumatoizi - FR (autoanticorpi IgM sau IgG, anti IgG proprii, modificate conformaţional, datorită
unui defect de glicozilare, care se produce în cursul infecţiilor streptococice cronice şi al
complicaţiilor acestora). De asemenea, se fixează de celule şi ţesuturi cu receptori Fc, inclusiv de
sinciţiotrofoblaştii placentari, fiind singurii Ac care pot traversa placenta, asigurând imunitatea
dobândită pasiv a nou-născutului. Nivelul seric al IgG la nou-născut este de 110% faţă de cel matern,
IgG fiind pompate activ în organismul fetal. Ac materni persistă până la vârsta de 3-6 luni, sinteza
proprie de IgG începe la 3 luni, iar nivelul normal este atins la 3 - 6 ani (fig. 9). Functiile IgG sunt
prezentate schematic in fig. 10. Filogenetic, IgG au apărut la peştii dipnoi.
În cadrul acestei clase există 4 subclase (fig. 11), prezente la toţi indivizii (este izotip).
12 12
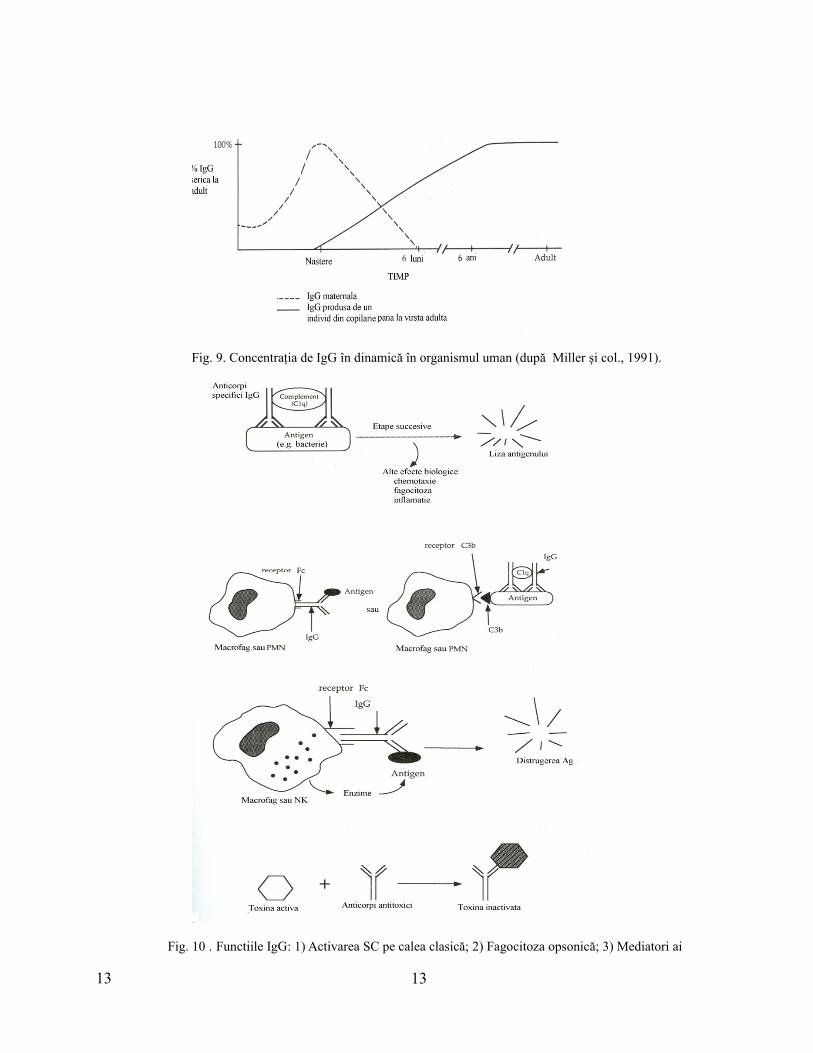
Fig. 9. Concentraţia de IgG în dinamică în organismul uman (după Miller şi col., 1991).
Fig. 10 . Functiile IgG: 1) Activarea SC pe calea clasică; 2) Fagocitoza opsonică; 3) Mediatori ai
13 13

citotoxicităţii celulare anticorpodependente (ADCC); 4) Neutralizarea toxinelor (reacţia de seroneutralizare) (după Miller şi colab., 1991).
Fig. 11. Structura moleculelor de IgG, aparţinând celor 4 subclase.
Clasa IgM. Moleculele de IgM au cea mai mare greutate moleculară - 950 kDa, 19S. Reprezintă
3 – 10% din cantitatea totală de Ig (concentraţie serică cuprinsă între 60 – 280 mg%). Lanţul µ are
576 de aminoacizi şi 5 domenii, dintre care 4 constante. IgM constituie anticorpii răspunsului imun
primar, care apar după primul contact cu Ag. Anticorpii IgM au T ½ = 10 zile, sunt termorezistenţi şi
au un conţinut de 12% glucide.
Structural sunt molecule pentamerice, libere în circulaţie sau molecule monomere fixate pe
membrana limfocitelor B. Pentamerul este alcătuit din 5 unităţi identice, legate între ele prin legături
disulfidice şi prezintă 10 situsuri identice de combinare cu Ag, dintre care sunt active 5, maximum 6,
datorită unui efect de mascare sterică. IgM pentameră are în structura sa şi un alt component, esenţial
pentru polimerizare, un lanţ care reuneşte monomerii la nivelul domeniului CH4, motiv pentru care a
fost numit lanţul J (engl. joining chain = lanţ de unire), deci formula moleculară este (µ2L2)5-J.
Ansamblul are o mare flexibilitate, molecula având formă de «stea» sau «păianjen», cu diametrul de
30 nm, formă ce se modifică în funcţie de relaţia cu Ag. Datorită volumului mare al moleculei, este
numită şi β2 – macroglobulina (fig. 12) şi prezentă mai ales în sistemul circulator şi în mică măsură
în ţesuturi (în lichidul interstiţial).
Moleculele de IgM au o mare capacitate de aglutinare, fiind Ac tipic aglutinanţi (capacitate de
1000x mai mare decât IgG), de precipitare şi mai ales de activare a sistemului complement pe calea
clasică (de 100x mai mare decât IgG). Pentru legarea componentului C1q şi activarea C pe calea
clasică, ce conduce la citoliză, condiţia obligatorie este ca 2 molecule de Ig foarte apropiate să lege
acest component, concomitent, la nivelul fragmentelor Fc. IgM fiind o moleculă pentameră, este
14 14

suficientă o singură moleculă pentru activarea C, deoarece condiţia obligatorie se realizează de la
sine, respectiv legarea C1q de 2 fragmente Fc. La IgM situsul de activare a C este situat la nivelul
domeniilor CH3 – CH4 şi devine accesibil (ca şi în cazul IgG) în urma modificării conformaţiei
sterice, după reacţia cu Ag, lanţul J nefiind implicat în acest proces.
Fig. 12. Structura moleculei pentamerice de IgM.
Anticorpii din clasa IgM au rol în apărarea faţă de antigene cu determinanţi antigenici repetitivi,
faţă de Ag complexe, activând citoliza celulelor străine mediată de complement, mecanism de
apărare activ în bacteriemii.
În dezvoltarea ontogenetică, primii Ac sintetizaţi aparţin acestei clase. În serul nou-născutului Ig-
le dominanante sunt deci IgM. Chiar şi la această vârstă, un nivel crescut de IgM sugerează existenţa
unor infecţii cu virusurile rubeolic sau citomegalic, cu bacteriile Treponema pallidum, Listeria
monocytogenes sau protozoarul Toxoplasma gondii, agenţi infecţioşi care se pot transmite
transplacentar. Atinge nivelul seric normal la vârsta de 1 an.
IgM sunt Ig-le caracteristică răspunsului imun primar, apărând la 3 zile după inocularea Ag-lui,
nivelul maxim este atins între 6 şi 10 zile, după care sunt înlocuite de IgG (prin procesul de comutare
a clasei, sinteza de IgM fiind inhibata printr-un proces de feed-back negativ, datorat afinităţii mai
mari a IgG pentru Ag, care competiţionează pentru acesta cu IgM), după aprox. 10 zile de la
inoculare (continuă şi sinteza de IgM, dar este mai redusă). Aglutininele α şi β din sânge, care au
capacitatea de a detemina aglutinarea Ag-lor de grup sanguin de pe hematiile histoincompatibile,
sunt Ac din clasa IgM. Filogenetic, forma monomeră a apărut la vertebratele inferioare, iar forma
polimerică, cu lanţul J – la rechini.
La adulţi, în serul bolnavilor cu artrită reumatoidă, apar Ig specifice numite factori reumatoizi
(FR), IgM (sau IgG) anti – IgG proprii, modificate conformaţional.
15 15

Spre deosebire de alte clase de Ig, sinteza IgM este mai puţin influenţată de acţiunea unor factori
imunodepresori, totuşi în sindroame de maladsobţie şi infecţiile cronice scade concentraţia de IgM
serică, fiind crescută în infecţiile acute, în general. O concentraţie crescută se înregistrează în artrite
reumatoide şi hepatite acute. Sunt Ac foarte eficienţi în bacteriemii, legarea toxinelor.
Monomeri de IgM se găsesc pe membrana limfocitelor B (IgMm), lanţurile grele ale acestora
având o regiune transmembranară hidrofobă şi una intracitoplasmatică; moleculele de IgMm au rol
de receptori pentru Ag (BCR = B cell receptor). Monomeri de IgM se găsesc şi în ser, în concentraţii
foarte mici in mod normal (care cresc însă într-o serie de boli, cum ar fi macroglobulinemia Waldenström,
lupusul eritematos diseminat –LED; in maladia Waldenström creşterea nivelului seric al IgM monoclonale,
însoţită de scăderea IgG şi IgA şi tendinţa acestor molecule de a forma polimeri, conduc la instalarea unui
sindrom de hipervâscozitate a sângelui).
Clasa IgA. Unitatea de bază este monomerul cu formula α2k2 sau α2λ2. Se prezintă şi în
formă dimerizată. Monomerul are 7S, g.m. 160 kDa şi o structură asemănătoare cu cea a IgG.
Dimerul se formează cu ajutorul unui lanţ de unire J. IgA sunt prezente atât în ser, cât şi în secreţii.
Clasa IgA este mai bogată în hidraţi de carbon comparativ cu IgG, legaţi în special de regiunea
«balama» (7-11%). Anticorpii IgA au T ½ = 6 zile şi sunt termorezistenţi.
Există două subclase de IgA: IgA1 şi IgA2 (fig. 13a). Subclasa IgA1 este prezentă în
proporţie de 90% în ser (80% fiind monomeră). Subclasa IgA2 are o structură originală, lanţurile L
fiind legate între ele prin legături covalente, iar de lanţurile H prin interacţiuni necovalente. Este
predominantă în secreţii, in forma dimerizata (fig. 13b).
IgA serică are o concentraţie relativ mare (90 – 450 mg%), reprezentând cca 15 -20% din
totalul Ig-lor aflate în circulaţie. Deşi în cantitate mare în ser, IgA serice au o activitate slabă de Ac.
Nivelul seric normal este atins la vârsta de 14 ani.
a) b)
Fig. 13. a) Structura monomerilor de IgA, aparţinând celor 2 subclase ; b) structura dimerului de sIgA (Kerr, 1990).
16 16
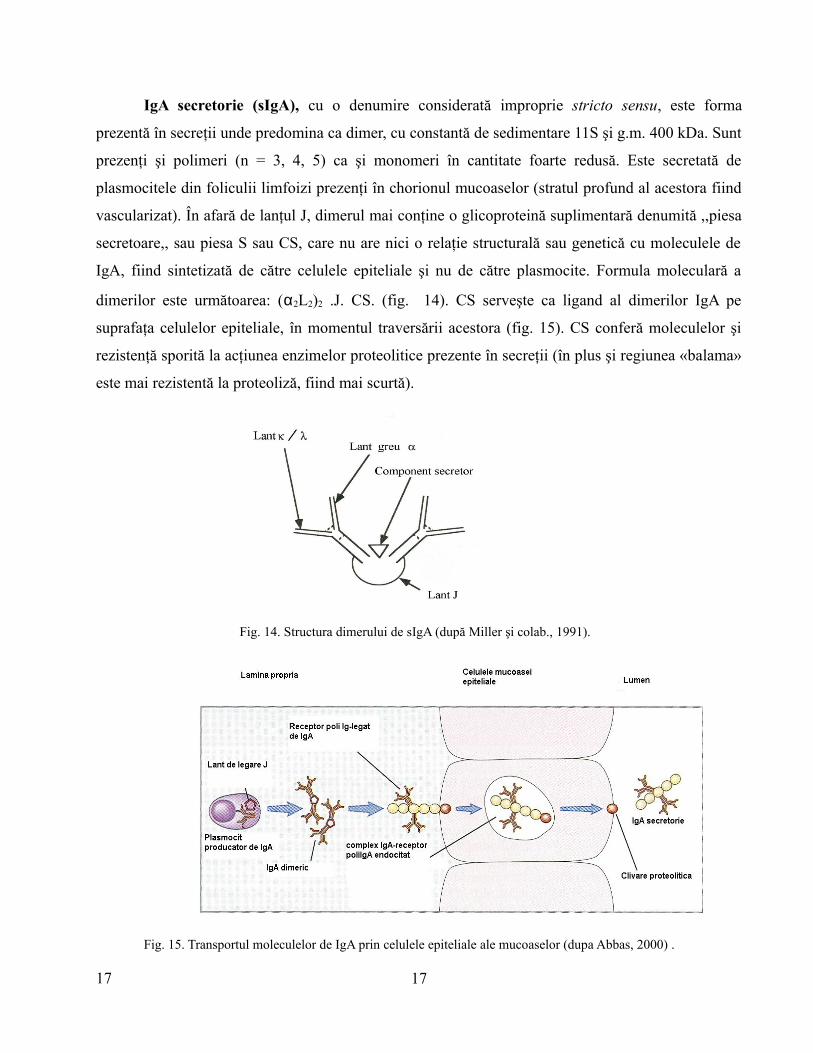
IgA secretorie (sIgA), cu o denumire considerată improprie stricto sensu, este forma
prezentă în secreţii unde predomina ca dimer, cu constantă de sedimentare 11S şi g.m. 400 kDa. Sunt
prezenţi şi polimeri (n = 3, 4, 5) ca şi monomeri în cantitate foarte redusă. Este secretată de
plasmocitele din foliculii limfoizi prezenţi în chorionul mucoaselor (stratul profund al acestora fiind
vascularizat). În afară de lanţul J, dimerul mai conţine o glicoproteină suplimentară denumită ,,piesa
secretoare,, sau piesa S sau CS, care nu are nici o relaţie structurală sau genetică cu moleculele de
IgA, fiind sintetizată de către celulele epiteliale şi nu de către plasmocite. Formula moleculară a
dimerilor este următoarea: (α2L2)2 .J. CS. (fig. 14). CS serveşte ca ligand al dimerilor IgA pe
suprafaţa celulelor epiteliale, în momentul traversării acestora (fig. 15). CS conferă moleculelor şi
rezistenţă sporită la acţiunea enzimelor proteolitice prezente în secreţii (în plus şi regiunea «balama»
este mai rezistentă la proteoliză, fiind mai scurtă).
Fig. 14. Structura dimerului de sIgA (după Miller şi colab., 1991).
Fig. 15. Transportul moleculelor de IgA prin celulele epiteliale ale mucoaselor (dupa Abbas, 2000) .
17 17

Funcţiile IgA.
Moleculele de IgA serice nu activează complementul pe calea clasică, nu favorizează fagocitoza
opsonică, nu aglutinează, nu precipită Ag-le. Rolul lor este de a îndepărta Ag-le provenite din
alimente sau Ag solubile ale bacteriilor din microbiota normală pătrunse în circulaţie, care astfel vor
fi eliminate rapid, împiedicând accesul acestora la celulele sistemului imunitar şi devierea
răspunsului imun de la funcţia sa normală de îndepărtare a non-self-ului. În alte lichide biologice ale
corpului, cum ar fi LCR, lichidul pleural, amniotic, sinovial, ca şi în ser - predomină IgG, raportul
IgG : IgAserică, fiind în jur de 5:1.
Moleculele de IgA secretorii sunt prezente în toate tipurile de secreţii (secreţia nazală, bronşică,
suc gastric, intestinal, secreţie biliară pancreatică, secreţia vaginală, lichidul prostatic, în lichidul
lacrimal, în salivă, în lapte şi colostru), dar în concentraţii diferite. Copilul la naştere are un nivel
seric redus de IgA, dar primeşte sIgA de la mamă, sinteza proprie începând după 30 de zile de la
naştere. De aceea, copiii alimentaţi natural sunt mai puţin receptivi la infecţii intestinale şi
respiratorii, comparativ cu cei alimentaţi artificial.
Moleculele de sIgA au un rol important în apărarea locală antivirală şi antibacteriană,
determinând imobilizarea şi agregarea agenţilor infecţioşi. Mecanismul de acţiune constă în
inhibarea aderenţei agenţilor infecţioşi la nivelul receptorilor specifici prezenţi pe suprafaţa celulelor
epiteliale şi a colonizării consecutive a mucoaselor, fenomenul fiind denumit excludere imună. S-a
demonstrat că nivelurile crescute de sIgA cu specificitate faţă de microorganismele cariogene au un
rol important in generarea rezistenţei la carii. Se pare că sIgA reglează cantitativ şi calitativ
(componenţa în specii) microbiota normală a mucoaselor. Dezvoltarea în filogenie a unui sistem
imunitar specializat al mucoaselor, apare foarte avantajoasă, luând în considerare suprafeţele
extensive ale corpului şi particularităţile epiteliilor acoperite de membrane mucoase vulnerabile. IgA
secretoare, component umoral specific al acestui sistem acţionează prin câteva mecanisme cum ar fi:
excluderea imuna (sIgA previne aderarea microorganismelor şi a proteinelor străine la suprafeţe
mucoase), urmată de clearance-ul complexelor imune, prin eliminarea lor de către curentul de
mucus.
Microorganismele patogene (bacterii, levuri) care colonizează mucoasele prezintă, în general,
proprietatea de a elabora enzime extracelulare cu ajutorul cărora contracarează efectul protector al
sIgA. În general, sinteza IgA - proteazelor este asociata la bacteriile respective cu prezenta capsulei
18 18

(efect anti-opsonizant), fapt care le confera avantaje deosebite în colonizarea mucoaselor. Pentru a
inhiba activitatea proteazelor, organismul-gazdă produce în cursul răspunsului imun anticorpi (IgA şi
IgG) anti-protează, rezistenţi la acţiunea enzimelor.
Indivizii cu deficit de IgA (1: 700 de subiecţi) sunt sensibili la infecţii ale mucoaselor
respiratorii, intestinale, ale conjunctivei etc., dar predispuşi şi la boli autoimune. In serul aceastor
indivizi nivelul IgG şi IgM se menţine în limite normale. În caz de mieloame de tip IgA predomină
forma polimerică de IgA (Kerr, 1990). Filogenetic, IgA este prezentă pentru prima dată la păsări
(Păunescu şi colab., 1999).
Clasa IgD – a fost descoperită în anul 1965 în serul unui bolnav cu mielom de tip IgD, nivelul
fiziologic în ser fiind foarte scăzut (cca 40mg/l), ceea ce reprezintă aprox. 0,2 – 1% din cantitatea
totală de Ig. Are o g.m. de 185kDa, 7S şi formula moleculară: δ2k2 sau δ2λ2. Lanţul δ are 3 domenii
constante şi o regiune «balama» foarte lungă (65 de aminoacizi), responsabilă de marea sensibilitate
a anticorpilor din această clasă la proteoliză enzimatică şi temperatură (la 56ºC, timp de 30 min).
Lanţurile δ sunt puternic glicozilate (9 - 14% hidraţi de carbon). Catabolismul acestor molecule este
extrem de rapid, T ½ = 3 zile. IgD există în organism sub două forme, şi anume ca molecule solubile,
circulante în plasmă şi ca receptor pentru Ag pe membrana limfocitelor B. Aceste molecule au un
fragment hidrofob la extremitatea carboxiterminală, prin care molecula se fixează la membrana LB,
datorită căruia au şi o g.m. mai mare decât a moleculelor libere.
Rolul biologic, în special al moleculelor serice, este de importanţă minoră. Astfel, IgD nu
fixează şi nu activează C, nu traversează placenta, nu provoacă degranularea bazofilelor şi a
mastocitelor, nu se fixează citofil pe monocite sau limfocite. Se pare că moleculele IgD ar avea un
rol important numai la nivel celular, ca receptor pentru Ag pe membrana LB, în asociere cu IgMm
monomere, cu aceeaşi specificitate. Se pare că intervin şi în diferenţierea limfocitelor B ca şi în
instalarea memoriei imunologice. Concentraţia serică la om creşte de la naştere până la 15 ani, după
care rămâne la nivel constant toată viaţa, în mod normal. Concentraţia IgD creşte în mod fiziologic
în sarcină şi mai ales în timpul naşterii şi patologic în unele infecţii cronice la copii, în astmul
bronşic, în pneumonii virale, în boala reumatoidă, L.E.D., leucemia limfatică cronică, mielomul de
tip IgD şi boala Hodgkin.
Clasa IgE – a fost descoperită în anul 1966. Notaţia E, derivă de la tipul de lanţ greu -ε, dar şi de
la „eritem” (roşeaţă), pentru că reprezintă aşa-numitele molecule de Ac reaginici sau Ac
19 19

sensibilizanţi ai pielii (citotropi). Formula moleculară este ε2k2 sau ε2λ2, g.m. 190 kDa, 8S, T ½ = 2
zile şi sunt molecule termosensibile (sunt denaturate la 56ºC, timp de 30 min). Prin încălzire IgE îşi
pierde capacitatea reaginică, nu însă şi funcţia de Ac, de recunoaştere specifică a Ag-lor. Lanţurile ε
au 5 domenii (g.m. 75 kDa) şi conţin un procent mare de hidraţi de carbon (12%). Concentraţia
serică este redusă, de aprox. 250 ng/ml (0,004% din totalul Ig-lor), dar crescută în colostru (sute de
ng), scăzând apoi în laptele matern, rămânând la valori crescute numai la mamele cu alergii. Nu se
transmit transplacentar, dar copilul o primeşte prin colostru, deşi sinteza proprie începe încă din
perioada intrauterină. Nivelul seric normal este atins la 14 ani.
IgE nu activează C, dar se leagă citofil de receptorii Fcε de pe membrana mastocitelor (bazofile
tisulare, prezente şi la nivelul mucoaselor şi a ţesutului conjunctiv), inducând degranularea acestora
şi eliberarea de amine vasoactive. IgE are un rol important şi în cursul inflamaţiei. În momentul în
care în organism pătrunde un alergen care este recunoscut şi reacţionează specific cu Ac IgE fixaţi
citofil, se transmit semnale în celulă, care determină degranularea mastocitelor şi eliberarea imediată
a unor cantităţi mari de amine vasoactive, care sunt mediatori farmacologic activi preformaţi de tipul
histaminei, serotoninei, care vor determina vasodilataţie, contracţia muşchilor netezi, creşterea
permeabilităţii vaselor sanguine. Sunt eliberaţi şi factori chemotactici pentru eozinofile, neutrofile,
proteaze ce afectează membrana endoteliilor şi TNF –α cu efect inflamator. Se sintetizează şi sunt
eliberaţi şi mediatori nou formaţi, de tipul prostaglandinelor care determină vasodilataţie şi
agregarea trombocitelor, de tipul leucotrienelor, cu rol în creşterea permeabilităţii vaselor sanguine şi
secreţia de mucus, generând simptomatologia tipică a reacţiilor de hipersensibilitate de tip I sau de
tip imediat (reacţii alergice). Simptomatologia depinde şi de situsul anatomic interesat. Un organism
sensibilizat faţă de un anumit antigen (alergen), răspunde exagerat, de unde denumirea de reacţie de
hipersensibilitate, de tip imediat pentru că se produce în câteva minute după contactul cu alergenul,
deoarece IgE sunt deja sintetizate şi legate citofil (fig. 16).
Ca şi celelalte molecule de Ig, moleculele de IgE sunt secretate de către plasmocite, aflate
sub controlul limfocitelor Th şi Ts, acţiunea acestui ultim tip celular tinzând să limiteze durata şi
intensitatea răspunsului imun în general, inclusiv prin IgE. De regulă răspunsul cu Ac IgE este
precoce, fiind sintetizaţi imediat după IgM şi înainte de IgG, producţia putându-se prelungi însă luni
şi chiar ani de zile, ceea ce explică durata mare a manifestărilor alergice.
20 20

Fig. 16. Fixarea citofilă a IgE la receptorii Fcε de pe suprafaţa bazofilelor şi mecanismul unei reacţii alergice, declanşată de pătrunderea unui alergen în organism şi degranularea consecutivă a bazofilelor.
COMPONENTA UMORALĂ NESPECIFICĂ
Sistemul complement (SC) şi reacţiile Ag-Ac cu participarea complementului
SC reprezintă un complex de proteine serice care interacţionează pentru a forma un sistem
efector, capabil să detemine liza unor celule străine şi / sau o serie de activităţi biologice importante,
cu rol esenţial în desfăşurarea răspunsului imun.
Acţiunea sa a fost descoperită de către Pfeiffer (1895), în cursul studiilor privitoare la
bacterioliza imună a vibrionului holeric, studii care au demonstrat că activitatea anticorpilor depinde
de prezenţa unui factor termolabil, prezent în serul sanguin proaspăt, numit de Buchner (1899)
alexina (gr. alexsein = a respinge, a apăra) şi de către Bordet (1900) – complement. Această
denumire exprima ideea, infirmată ulterior că SC ajută anticorpii să-şi exercite funcţiile efectoare de
apărare. Această concepţie iniţială a fost restructurată, inversând relaţia dintre anticorpi şi C, în
sensul că Ag, reprezentate de celule invadatoare, sunt atacate efectiv de complement, în timp ce Ac
au doar funcţia de recunoaştere şi identificare a acestora şi de activare a SC, a cărui funcţie
esenţială este citoliza celulelor străine. Complementul nu este o substanţă unică, ci un sistem
alcătuit din ~ 26 de componente proteice, organizate în 9 grupe, care se găsesc în ser în stare
inactivă, dar pot fi activate enzimatic pe două căi: clasică şi alternativă.
Proteinele SC reprezintă 10 – 15% din globulinele serice normale, a căror concentraţie nu
este influenţată de stimularea antigenică (imunizare), nici de gradul de activare a celulelor S.I., ci de
starea de nutriţie (mai ales de aportul proteic) şi de factorii genetici (de specia animalului considerat
21 21

– de ex., serul de cobai are cea mai mare concentraţie de complement). Sunt proteine termolabile
(inactivate la 56 o C, timp de 10 min.) sintetizate mai ales de către hepatocite, dar şi de către
macrofage, monocite; C1q este sintetizat exclusiv de către epiteliul gastrointestinal şi urogenital.
Unele componente au rol efector, altele au rol de reglare. Indiferent de calea de activare, primul
component odată activat, declanşează o reacţie în lanţ sau în “cascadă”, inducând activarea
următorului component ş.a.m.d.
Nomenclatură. Proteinele SC circulante, inactive sunt desemnate prin litera C, urmată de un
indice numeric de la 1 – 9, care corespunde ordinii în care proteina respectivă intră în reacţie: C1 -
C4 – C2 – C3 – C5 – C6 – C7 – C8 – C9 (numerotarea componentelor izolate şi identificate s-a făcut
înainte de stabilirea ordinii intrării lor în reacţie); alte componente sunt desemnate prin simboluri (I,
B, D, H) sau denumiri obişnuite – properdina.
Fragmentele rezultate din clivarea enzimatică a anumitor componente sunt notate cu litere
mici, adăugate la denumirea componentului normal. De ex., componentul C1 poate fi clivat în trei
subunităţi: C1q, C1r, C1s, care în mod normal sunt menţinute împreună prin legături necovalente şi
în prezenţa ionilor de Ca++. Componentele cu activitate enzimatică sau cele active biologic, sunt
desemnate print-o bară adăugată deasupra denumirii componentului inactiv; de ex.: C1s, C3b,
C5b67.
Activarea componentelor SC inactive poate fi realizată pe două căi:
1) Calea clasică – activată de formarea complexelor imune Ag-Ac (fig. 17);
2) Calea alternativă – activată de substanţe polimerice: LPS din structura peretelui celular al
bacteriilor Gram negative, polizaharide capsulare bacteriene şi fungice, Ig polimerizate.
Fig. 17. Activarea componentului C1 pe calea clasică, consecutiv reacţiei Ag-Ac (dupa legarea Ac-lor de Ag expuse pe membrana celulelor straine).
22 22

Din punct de vedere funcţional, moleculele SC se împart în 3 grupe: 2 realizează cascada de
reacţii care vor activa C3 (pe una din cele două căi); odată activat, C3 activează a 3-a grupă care
formează “complexul de atac al membranei” (C5 – C9).
Activarea precursorilor inactivi ai SC se face într-o succesiune riguroasă, prin clivare
proteolitică limitată, în general în două fragmente active:
a) fragmentul major - cu 2 situsuri, unul pentru ataşarea la membrana celulară şi un situs activ enzimatic,
necesar clivării următorului component, astfel că se creează un efect de “cascadă enzimatică”; o parte din
moleculele activate se leagă de membranele celulare (C5-C9) şi devin activatoare pentru secvenţele
ulterioare, restul sunt eliberate în circulaţie şi rapid degradate, inactivate; astfel efectele nocive ale SC
sunt limitate la Ag declanşator al răspunsului, iar celulele organismului-gazdă sunt protejate;
b) fragmentul minor – de regulă nu este implicat în citoliză; rămâne în circulaţie şi exercită efecte biologice
importante: chemotaxia, imunoaderenta, creşterea permeabilităţii vasculare si anafilaxia.
Activarea se realizează pe cele 2 căi:
1) calea clasică - mai rapidă şi mai eficientă – expresia unor mecanisme specifice;
2) calea alternativă – mai lentă şi mai puţin eficientă – o cale primitivă, nespecifică, cu rol
major în apărarea antibacteriană la vertebratele inferioare şi cu rol important la vertebratele
superioare, mai ales la începutul unei infecţii, înainte de sinteza Ac-lor specifici.
Indiferent de calea de activare a componentelor SC, odata activate acestea contribuie la
functia esenţială a SC, care este citoliza celulelor străine, mecanismul leziunilor induse de SC fiind
de tip osmotic; componentele asa-numitului “complex de atac al membranei” (C5 – C9) atasate
secvential la membrana celulei-straine creeaza în aceasta orificii suficient de mari pentru a putea
permite trecerea masiva a apei şi electroliţilor (fiind hidrofile în regiunea centrală), dar prea mici
pentru proteine şi acizi nucleici, ceea ce în final determină explozia celulei sau citoliza.
COMPONENTA CELULARĂ NESPECIFICĂ
A SISTEMULUI IMUNITAR
Răspunsul imun celular (RIC) este implicat în eliminarea a două categorii de celule:
singenice (self) şi alogenice (străine, transplantate). Celulele proprii pot deveni antigenice prin uzură,
infectare cu virusuri şi bacterii parazite intracelular, malignizare.
Componenta celulară a sistemului imunitar este reprezentată de celule nespecifice = fagocite
(PMN şi macrofage, numite şi celule accesorii ale S.I.) şi celule specifice: limfocite T şi B, ca şi
23 23

celule limfoide non- T, non– B = NK, K (nespecifice). Aceste elemente celulare sunt prezente atât în
organele limfoide, cât şi în sângele periferic (Fig. 18, A) şi pot fi evidenţiate pe frotiul de sânge (fig.
18, B).
Fig. 18, A. Liniile celulare derivate din celula stem pluripotentă.
24 24

Fig. 18, B. Elementele figurate ale sângelui, între care şi celulele sistemului imunitar, observabile pe frotiul de sânge; 1) monocit; 2) trombocite; 3) PMN neutrofil segmentat; 4) PMN bazofil;
5) limfocit; 6) PMN eozinofil; 7) hematie; 8) monocit; 9) PMN neutrofil nesegmentat.
* Valorile normale ale formulei leucocitare la adult: neutrofile nesegmentate- 1- 4%, neutrofile segmentate - 60-70%, eozinofile – 1 -4 %, bazofile –0 -1%, limfocite – 25 -30%, monocite – 4 -8%, plasmocite - 0- 0,5%; de menţionat că la copilul mic proporţiile de PMNN şi limfocite sunt inversate (Ţiţeica şi col., 1984).
Sistemul fagocitar si fagocitoza.
Fenomenul de fagocitoză a fost descoperit în 1882 de către Metchnikoff, care a dat
numele generic de “fagocite” = celule care mănâncă, celulelor libere şi fixe, care au capacitatea
de a îngloba în citoplasmă particulele fine cu care vin în contact şi de a le distruge prin digestie
intracelulară, ori de câte ori natura chimică a acestora le face accesibile enzimelor fagocitului.
Fagocitoza este foarte răspândită pe scara animală, funcţia de apărare a acestui proces derivând
din funcţia de nutriţie, pe care o are la protozoare şi metazoarele primitive. În afară de aceasta,
fagocitele sunt implicate în metamorfoza nevertebratelor şi a vertebratelor inferioare, realizând
resorbţia celulelor distruse. La organismele superioare sunt implicate în procesul de remaniere a
oaselor şi joacă rolul de ,,măturător,, prin aceea că înglobează şi digeră celulele devenite inutile
(uzate), pe cele cu viaţă scurtă şi pe cele moarte sub acţiunea unui agent patogen. Astfel
fagocitele contribuie la eliminarea unor resturi nefolositoare şi la reintroducerea în economia
generală a organismului a unor constituienţi chimici. Fagocitele se grupează în două categorii
majore:
25
9.
25

1) Fagocite ,,profesioniste,,- adaptate prin definiţie să ingere particule străine în
stadiul matur şi purtătoare de receptori specifici pentru opsonine: C3b şi IgG şi capabile de
opsonofagocitoză (fagocitoză imună):
- Sistemul fagocitar mononuclear = S.F.M. (monocite şi macrofage);
- ,, ,, polimorfonuclear = S.F.P.M.N. (neutrofile, eozinofile).
2) Fagocite ,,neprofesioniste,, - exercită această funcţie facultativ, ingerând cantităţi
mici de particule străine şi fără intervenţia receptorilor pentru opsonine; sunt reprezentate de
celule reticulare dendritice din organele limfoide secundare, fibroblaşti şi celule endoteliale.
Celulele dendritice sunt răspândite în toate ţesuturile corpului cu excepţia creierului şi
prezente în număr mare în piele şi în cortexul nodulilor limfatici, fiind o populaţie de celule
monocitare (similare macrofagelor); cele din piele poartă denumirea de celule Langerhans,
fiind celule ce prezintă prelungiri citoplasmatice filamentoase, lungi şi fără proprietăţi
fagocitare.
COMPONENTA CELULARA SPECIFICA A S.I.
Sistemul limfoid
Limfocitele sunt celulele efectoare esenţiale, specifice ale răspunsului imun, fiind
generate, stocate şi “educate” la nivelul ţesuturilor şi organelor limfoide specializate, structuri
cunoscute ca “sistem limfoid”, substratul morfologic şi funcţional al S.I., care asigură
imunostazia organismului.
Principala funcţie biologică a celulelor S.I. şi în special a celulelor limfoide
imunocompetente este recunoaşterea specifică a Ag-lor şi dezvoltarea de reacţii specifice, a
răspunsului imun prin care Ag este anihilat şi eliminat.
Ţesutul limfoid este format dintr-un reticul fibrilar în ale cărui “ochiuri” se găsesc celule libere.
Reticulul este alcătuit din fibre reticulare, celule reticulare sau dendritice şi macrofage fixe.
Celulele libere – limfocite (majoritatea) în diferite stadii de diferenţiere şi maturare, macrofage.
Organele limfoide, după modul de apariţie în embriogeneză şi după funcţiile lor, se împart în două
grupe: primare şi secundare.
La vertebratele superioare:
26 26

- organele limfoide primare apar iniţial în evoluţia filogenetică şi în embriogeneză:
- timusul – pentru LT;- bursa lui Fabricius – pentru LB, la păsări;- măduva osoasă (echivalentul bursal) – la mamifere.- organele limfoide secundare- ganglionii limfatici, splina, plăcile Peyer, amigdalele, apendicele
vermiform, vegetaţiile adenoide, celulele limfoide izolate din piele şi mucoase, limfocitele libere din
sânge şi limfă.
Organele limfoide primare şi secundare au caracteristici care le deosebesc fundamental. În
cele primare limfocitele suferă transformări care conduc la maturarea lor functională, iar cele
secundare sunt populate cu limfocite instruite în cele primare, la acest nivel având loc majoritatea
reacţiilor imunitare. Celulele care pot realiza potenţial un răspuns imun faţă de un Ag, se numesc
imunocompetente, iar cele care au fost stimulate şi reacţionează faţă de un anumit Ag, se numesc
celule angajate, Ag-reactive.
Funcţia limfopoietică a organelor limfoide primare este condiţionată de popularea lor cu celule tinere,
imature, progenitoare, derivate din celulele suşă, matcă sau stem, de la nivelul măduvei osoase. Organele
limfoide secundare sunt populate cu celule limfoide educate în cele primare.
Ontogenia şi rolul diferitelor subpopulaţii de limfocite T
Limfocitele T se dezvoltă in 3 stadii succesive (fig. 19):
1) stadiul pretimic (în măduva osoasă); 2) stadiul intratimic (LT sunt „educate” la acest nivel şi îşi dobândesc imunocompetenţa),
aceste prime 2 stadii fiind independente de Ag;3) stadiul posttimic, la nivelul organelor limfoide secundare, după migrarea limfocitelor
(educate în timus) şi întâlnirea cu Ag, diferenţierea se face sub influenţa Ag-lui.
Fig. 19. Etapele diferentierii (ontogenia LT).
27 27

Caracterizarea generală a principalelor subpopulaţii funcţionale de LT
Studiul corelativ al particularităţilor fenotipice/funcţionale ale LT periferice a dus la
împărţirea lor, în funcţie de exprimarea Ag-lor CD4 şi CD8, în 2 subpopulaţii celulare, care sunt
din punct devedere funcţional mutual exclusive.
Limfocitele Th. Limfocitele care prezintă markerul membranar CD4 sau T4, se notează
ca celule CD4+ sau T4+ şi aşa sunt desemnate limfocitele acestei subpopulaţii, înainte de
întâlnirea cu antigenul specific. Celulele latente activate de către Ag specific prezentat de către
celulele prezentatoare de antigen (CPA) în asociere cu molecule CMH cls. II, proliferează
(expansiunea clonală) şi se diferenţiază la celule efectoare numite LT ajutătoare = Th (engl.
helper).
Funcţiile Th– stimulează LB să prolifereze şi să se diferenţieze la plasmocite, secretoare de Ac;
- „ Tc–să acţioneze asupra celulelor-ţintă: celule tumorale, infectate cu virusuri
- „ Ts – să-şi exercite funcţiile de modulare a răspunsului imun;
- „ activitatea celulelor NK;
- interacţionează cu macrofagele (CPA) în cursul răspunsului imun;
- stimulate, produc IL-2 – limfokină care activează alte subpopulaţii celulare.
Rezultă rolul central al limfocitelor Th în declanşarea răspunsului imun mediat celular şi
umoral, majoritatea antigenelor naturale fiind timodependente şi necesitând interacţiunea Th -
LB, pentru sinteza de Ac.
Limfocitele Tc/s
Tc = celule de tip efector, care determină liza celulelor străine: infectate cu virusuri,
tumorale, grefe incompatibile.
Ts = celule rol în supresia răspunsului imun, contribuie la inducerea toleranţei faţă de
self, desăvârşesc educaţia LT din timus; în caz de deficit al acestor celule, se produc boli de tip
autoimun. Deoarece toate organismele conţin aceleaşi sisteme embrionare de gene pentru Ig şi
TCR şi deci au acelaşi potenţial de răspuns imun, este evident că toleranţa faţă de self trebuie
dobândită – în timus- unde are loc educaţia limfocitelor pentru a discrimina self-ul de non-self),
proprietate-cheie a sistemului imunitar (S. Tonegawa - premiul Nobel 1987, pentru conceptul de
educaţie a limfocitelor, ca şi pentru elucidarea structurii TCR).
Celulele Tc/s = T8 = CD8+ recunosc moleculele CMH I, în asociere cu Ag străin =
recunoaştere asociată, dublă, cu restricţie genetică.
28 28

Concluzii. LT recunosc Ag expuse pe suprafaţa celulelor, produc limfokine (IL, IFN,
factori de creştere şi de diferenţiere a diferitelor celule imunitare), induc sinteza şi exprimarea
receptorilor pentru limfokine, dezvoltă funcţii citotoxice. LT mature au o importanţă
fundamentală: activitate antivirală, antifungică, antitumorală şi de respingere a grefelor– reacţii
ale imunităţii mediate celular.
Limfocitele B îşi au originea în celulele stem din măduva osoasă. Etapa iniţială de
diferenţiere, independentă de Ag trece prin stadiile de celule B medulare, celule pre-B, celule B
mature imunocompetente. În etapa a II-a, ca urmare a întâlnirii cu Ag, LB mature sunt activate şi
suferă un proces de proliferare intensă, de expansiune clonală, proces asociat cu o diferenţiere
dependentă de Ag (stimulată de Th şi reglată de Ts), la celule terminale = plasmocite secretoare
de Ac (Fig. 20 a, b), şi LB de memorie, cu viaţă lungă. Fiecare LB matur, în repaus (înainte de
întâlnirea cu Ag) sintetizează cantităţi mici dintr-un tip de Ig caracteristice, care rămân legate de
membrana LB (Ig M, IgD), ca molecule-receptor pentru Ag (BCR), asemănătoare unor antene de
detectare a Ag-lor. Fiecare clona de LB este programată genetic să producă un anumit tip de Ig şi
capabile să recunoască un anumit Ag. În repaus, LB au receptori de mică afinitate pentru IL-2,
dar după stimulare exprimă receptori de mare afinitate pentru IL-2 care determină proliferarea
LB şi secreţia de IgM.
Etapa de diferenţiere independentă de Ag. La nivelul măduvei, din celulele stem
derivă precursorii LB, dintre care supravieţuiesc numai cei care îşi rearanjează eficient linia
germinală şi care se acumulează în spaţiile sinusoidale, aproape de sinusul central, de unde trec
în circulaţie ca celule mature, virgine. Majoritatea mor în măduvă (95% la oaie), fiind rapid
fagocitate, precursorii fiind supuşi aici primei selecţii la care rezistă doar cele care sintetizează
intracitoplasmatic lanţul µ, apoi L şi δ şi care în final exprimă receptorii IgM şi IgD.
În circulaţie LB sunt supuse la a II-a selecţie, majoritatea având viaţă scurtă, fiind
omorâte şi depozitate în splină (în pulpa roşie). Celulele care rezistă ajung în ariile B-dependente
din ganglioni şi splină unde trăiesc 6-8 săptămăni. Dacă nu întâlnesc Ag corespunzător mor, fiind
înlocuite cu altele. Dacă întâlnesc Ag-ul urmează a III-a selecţie, şi etapa de diferenţiere
dependentă de Ag în care au loc hipermutaţii somatice, cu evoluţie spre plasmocite şi LB de
memorie.
29 29

Fig. 20: a) Etapele diferentierii (ontogenia) LB (dupa Zarnea, 1990). b) Reprezentarea schematică a unui plasmocit.
Plasmocitele secretă Ac liberi în circulaţie, solubili, identici ca specificitate cu IgM
(BCR=receptorul pentru Ag) (teoretic, pentru că practic afinitatea Ac secretaţi pentru Ag
corespunzător este mai mare). Plasmocitele au o durată de viaţă de 2-3 zile. Celulele care
evoluează spre plasmocit încep să-şi dezvolte maşinăria de sinteză a Ac-lor: îşi pierd receptorii
de membrană, se dezvoltă reticulul endoplasmic, aparatul Golgi, creşte numărul poliribosomilor,
ceea ce demonstrează că funcţia esenţială a LB este cea de sinteză şi secreţie de Ig. La început
sintetizează IgM, apoi se comută clasa la IgG. Capacitatea de sinteză este foarte mare: 10.000
molecule de Ig/sec. După formare, au loc reacţii Ag-Ac, care conduc la neutralizarea şi
eliminarea Ag-lor şi a celulelor care le poartă (prin fagocitoză opsonică sau activarea SC).
Au fost evidenţiate şi LB prezentatoare de Ag (CPA) şi LB supresoare, ceea ce înseamnă că LB
intervin în procese de cooperare şi reglare ale răspunsului imun. Distribuţia LB în organele limfoide este
diferită şi opusă celei LT. Antigenele care stimulează LB sunt reprezentate de diferite molecule din
alimente, de MO ale microbiotei normale sau invadante.
30 30












![Untitled-1 [] · 2017-02-13 · imunitar, eliminarea toxinelor, curata colonul, protejarea celulelor impotriva stresului oxidativ. Maca Hraneste sistemul endocrin, contribuie in mod](https://img.pdfslide.tips/doc/110x75/5e445891fc83c80b8279d3e8/untitled-1-2017-02-13-imunitar-eliminarea-toxinelor-curata-colonul-protejarea.jpg)