Embed Size (px)
Citation preview

Q34
Q3
Q4
The American Journal of Pathology, Vol. -, No. -, - 2013
1234567891011121314151617181920212223242526272829303132333435363738394041424344454647484950515253545556575859606162
ajp.amjpathol.org
6364656667686970717273747576777879808182
Slug/b-CatenineDependent Proinflammatory Phenotypein Hypoxic Breast Cancer Stem CellsGianluca Storci,*y Sara Bertoni,y Sabrina De Carolis,*y Alessio Papi,z Marina Nati,* Claudio Ceccarelli,* Chiara Pirazzini,*Paolo Garagnani,* Alberto Ferrarini,x Genny Buson,x Massimo Delledonne,x Michelangelo Fiorentino,{ Elisa Capizzi,{
Elisa Gruppioni,{ Mario Taffurelli,k Donatella Santini,k Claudio Franceschi,* Giuseppe Bandini,** Francesca Bonifazi,** andMassimiliano Bonafé*y
8384858687
From the Department of Experimental, Diagnostic and Specialty Medicine,* the Center for Applied Biomedical Research,y the Pathology Unit,{ AddariiInstitute of Oncology, and the Institute of Haematology “L & A Seragnoli,”** St. Orsola-Malpighi University Hospital, Bologna; the Departments ofBiological, Geological and Environmental Sciencesz and Biotechnologies,x Functional Genomics Center, University of Verona, Verona; and the Department ofMedical and Surgical Sciences,k University of Bologna, Bologna, Italy
888990
Accepted for publicationJuly 30, 2013.
2
C
P
h
919293949596979899
Address correspondence toMassimiliano Bonafé, M.D., orGianluca Storci, Ph.D., Depart-ment of Experimental, Diag-nostic and Specialty Medicine,University of Bologna,Bologna, Italy. E-mail:[email protected] [email protected].
opyright ª 2013 American Society for Inve
ublished by Elsevier Inc. All rights reserved
ttp://dx.doi.org/10.1016/j.ajpath.2013.07.020
100
FL
Cancer stem cell survival relies on the activation of inflammatory pathways, which is speculativelytriggered by cell autonomous mechanisms or by microenvironmental stimuli. Here, we observed thathypoxic bone marrow stromaederived transforming growth factor-b 1 promotes the growth of humanbreast cancer stem cells as mammospheres. The ensuing Slug-dependent serine 139 phosphorylation ofthe DNA damage sensor H2AX in breast cancer stem cells induces tumor necrosis factor-a and IL-8 mRNAs,whose stability is enhanced by cytoplasmic b-catenin. b-Catenin also up-regulates and binds miR-221,reducing the stability of the miR-221 targets Rad51 and ERa QmRNAs. Our data show that the Slug/b-cateninedependent activation of DNA damage signaling triggered by the hypoxic microenvironmentsustains the proinflammatory phenotype of breast cancer stem cells. (Am J Pathol 2013, -: 1e10;http://dx.doi.org/10.1016/j.ajpath.2013.07.020)
101
Supported by grant PRIN 2008KTRN38 Clinical, diagnostic and thera-peutic implications of studies on breast cancer stem cells (M.T. and M.B.)and by RFO funds ex 60%, Cornelia Pallotti and Roberto Pallotti Foun-dation Q30(M.B.).
102103104105106107108109110111112113114115116117118119120121122123
Cancer stem cells (CSCs) constitute a minor population ofimmunophenotypically identifiable cells endowed with hightumor-initiating capability in xenografts.1 Recent literatureshows that CSCs are endowed with a proinflammatoryphenotype.2e4 This feature is characterized by the over-activation of NF-kBeregulated proinflammatory cytokines[IL-6, tumor necrosis factor-a (TNF-a), and IL-8], whichare potent breast CSC growth factors.5e8 Notably, CSCsurvival and self-renewal rely on the activity of inflamma-tory pathways to a higher extent than in their normalcounterpart.2,9 Such an inflammatory addiction is currentlyinvestigated as a potential Achilles’ heel of CSCs.2
The CSC phenotype is promoted by exposure to lowoxygen tension.10,11 Hypoxic tissue regions, which fre-quently occur in the tumor mass,12 are currently regarded asthe niches for CSCs.13 Intriguingly, the bone marrow, aprivileged soil for metastatic breast cancer cell engraft-ment,14 harbors an oxygen-poor compartment enriched inhematopoietic stem cells niches.15 Conceivably, this mayconstitute a niche also for disseminating breast cancer cells.
stigative Pathology.
.
A 5.2.0 DTD � AJPA1476_proof �
Hypoxia promotes genetic instability and down-regulatesRad51, the major player of the homologous recombinationDNA repair pathway.16 Hypoxia also up-regulates the poly-comb 2 receptor complex component EZH2, an acknowl-edged Rad51 transcriptional repressor.17 Interestingly, EZH2overexpression enhances breast CSCs self-renewal,17 simi-larly to the knockdown of the genome integrity gatekeeperBrca1.18 Intriguingly, DNA-damaged cells cast inflamma-tory signals into the surrounding microenvironment.19e21
Hence, a close relationship between the activation of theDNA damage response and the proinflammatory status inCSCs can be hypothesized.
Here, we show that hypoxic bone marrow stroma (BMH)-derived transforming growth factor-b1 (TGF-b1) expandsbreast CSCs as mammospheres (MS). TGF-b1 has been
124
12 September 2013 � 2:47 am � EO: AJP13_0196

Q5
Q6
Q7
8
9
10
11
12
13
Storci et al
125126127128129130131132133134135136137138139140141142143144145146147148149150151152153154155156157158159160161162163164165166167168169170171172173174175176177178179180181182183184185186
187188189190191192193194195196197198199200201202203204205206207208209210211212213214215216217218219220221222223224225226227228229230231232233234235236237238239240241242243244245246247
previously described as autocrine breast CSCs’ promotingfactor22: here, we report that the peptide induces an autocrineloop of TNF-a, a previously identified CSC growthfactor.3,23,24 We describe the ensuing up-regulation of thetranscriptional repressor Slug, a master regulator of breastcancer and mammary gland stem cell self-renewal andsurvival.25e28 Slug engenders the serine 139 phosphorylationof the DNA damage sensor H2AX (g-H2AX),29 and theNF-kBemediated up-regulation of proinflammatory pep-tides IL-8 and TNF-a, in the absence of any detectablegenomic alteration. In MS, the Slug-dependent up-regulationof IL-8 and TNF-a expression also relies on the recentlycharacterized capability of b-catenin, a crucial modulator ofstem cell features,30 to bind and stabilize mRNAs.31 Onexposure to hypoxia, b-catenin interacts with the NF-kBtarget miR-221, which has been previously associated withbreast cancer cell aggressiveness.32,33 b-Catenin then affectsthe stability of two miR-221 target mRNAs: the DNA repairgene Rad5134 and estrogen receptor-a (ERa). These datadisclose that the Slug/b-catenin proinflammatory phenotypein CSCs is a consequence of the hypoxia-induced activationof DNA damage response.
Materials and Methods
Cell Lines and Chemicals
MCF7 cells were grown as adherent layer in RPMI 10% fetalbovine serum (Euroclone, Milan, Italy). MS were generatedfrom MCF7 cells in mammary epithelial growth medium(MEGM), supplemented with ad hoc aliquots of B27,epidermal growth factor, basic fibroblast growth factor,insulin, and hydrocortisone (Lonza, Basel, Switzerland).7
Briefly, 5 � 103 MCF7 cells were filtered through a 40-mmnylon mesh (Becton Dickinson, Franklin Lakes, NJ) andsuspended with 1 mL of complete MEGM in 1.5-cm2 lowattachment wells (Corning, Corning, NY). MS were scored atday 5. Normoxic cultures were kept at 37�C in a 5%CO2ehumidified atmosphere. Hypoxia (1% pO2, 5% pCO2,94% pN2) was obtained in an in vivo 300 hypoxia cabinet(Ruskinn Technology, Bridgend, UK). Recombinant humanTNF-a and TGF-b1 were purchased from Sigma-Aldrich(St. Louis, MO). Monoclonal neutralizing antibodies wereantieTNF-a antibody (clone 28401; R&D Systems, Min-neapolis, MN) and antieTGF-b1 antibody (clone 9016-2;Sigma-Aldrich).
MS Generation from Breast Cancer Tissues andIsolation of Fibroblasts from the Normal Bone Marrow
MS were generated in complete MEGM from human breastcarcinoma tissues that were characterized according to stan-dard diagnostic procedures3,7 (Supplemental Table S1). Theprocedure had been approved by the St. Orsola-MalpighiUniversity Hospital ethical committee (Prot n.75/2011).Immunohistochemistry was performed with antiehuman ER
2FLA 5.2.0 DTD � AJPA1476_proof �
(clone SP1), antiehuman progesterone receptor (PgR; clone1E2), and antiehuman epidermal growth factor receptor 2(HER2; Pathway clone 4B5) fromVentanaMedical Systems,Tucson, AZ. Immunostaining was performed on a VentanaBenchmark Ultra immunostainer and visualized using theUltraView DAB Universal Detection Kit Q. To assess self-renewal, MS were disaggregated at day 7 in 1� trypsin-EDTA (STEMCELL Technologies, Vancouver, BC,Canada), washed in complete MEGM, filtered throughouta 40-mm nylon mesh, and seeded to form higher (second,third, and fourth) generation MS. Fibroblasts were isolatedfrom bone marrow from one healthy donor. Adherentmononuclear cells were cultured in Dulbecco’s minimalessential medium Qand supplemented with 20% fetal bovineserum for 3 weeks. Second-passage fibroblasts were grown at75% confluence in hypoxic conditions, and culture super-natants were collected after 24 hours. Written informedconsent was obtained for primary cell isolation.
Generation of Slug and b-Catenin MCF7 Knockdownand of Slug Overexpressing MCF7 Clones
Stable Slug knockdown (shSlug) MCF7 cells has beenpreviously described.7 Stable knockdown of b-catenin Q
(shBeta) in MCF7 cells was obtained via the retroviraltransduction of pCtoGMB retroviral vector carrying humanb-cateninespecific 19-nt coding sequence (50-GAGCCTC-TATACCACCCAC-30 Q), as previously described.7 Slugoverexpressing (Babe-Slug) MCF7 clones were generatedfrom the BamHI/EcoRI full-length Slug cDNA, and thensubcloned into the pBabe-Puro retroviral vector (pBabe-Slug). Babe-Slug cells were selected in RPMI containing 1mg/mL puromycin and then seeded into 96-well plates forsingle-cell cloning. Three Babe-Slug MCF7 clones (A, B,C) were used for the study.
Transient Transfection of siRNA, PremiR, andAntagomiR
siRNA against H2AX-, SLUG Q-, and GC-matched scrambledcontrol (Life Technologies, Carlsbad, CA) were Qtransientlytransfected with Lipofectamine 2000 (Life Technologies) toadherent MCF7 cells (105 cells � 3 cm2 well), or withjetPEI (Polyplus, Illkirch, France) to MS, at a concentrationof 1 mg/well for 72 hours. Pre- and antagomiR-221 (LifeTechnologies) were transiently transfected to adherentMCF7cells (105 cells� 3 cm2 well) at a concentration of 100 pmol/well for 72 hours.
Luciferase Reporter Assay
NF-kB-Luc was provided by Dr. K.B. Marcu (Stony BrookUniversity, Stony Brook, NY); Slug-Luc reporter vectorwas provided by Dr. Togo Ikuta (Saitama Cancer Centre,Saitama, Japan); TOPFLASH reporter vector was providedby Dr. Rolf Kemler (Max Planck Institute, Heidelberg,
ajp.amjpathol.org - The American Journal of Pathology
248
12 September 2013 � 2:47 am � EO: AJP13_0196

14
DNA Damage Response and Proinflammatory Phenotype Q1
249250251252253254255256257258259260261262263264265266267268269270271272273274275276277278279280281282283284285286287288289290291292293294295296297298299300301302303304305306307308309310
311312313314315316317318319320321322323324325326327328329330331332333334335336337338339340341342343344345346347348349350351352353354355356357358359360361362363364365366367368369370371
Germany). Luciferase reporters were assessed using theDual-Luciferase Reporter Assay System and a GloMax 20/20 Luminometry System (Promega, Madison, WI).
RT-PCR and Real-Time PCR analysis
Total RNA was extracted from cells using TRIzol reagentaccording to the manufacturer’s instructions (Life Tech-nologies). RT-PCR primers were TNF-a: forward 50-GG-CCCAGGCAGTCAGATCAT-30 and reverse 50-GGGGC-TCTTGATGGCAGAGA-30 (310 bp); IL-8: forward 50-GCTTTCTGTGGAAGAGAGC-30, and reverse 50-GGCA-CAGTGGAACAAGGACT-30 (606 bp); Rad51: forward50GCCTGCTGGAGAGAGGA-30, and reverse 50-GGAA-GCTGGCAGGTGAC-30 (500 bp); ERa: forward 50-TGA-AAGGTGGGATACGAAAAGAC-30, and reverse 50-CAG-GATCTCTAGCCAGGCACAT-30 (410 bp); real-time pro-bes forGUS (b-glucuronidase, 4326320E), Slug (Hs00950344-m1), IL-8 (Hs00174103-m1), TNF-a (Hs00174128-m1), Brca1(Hs01556193-m1), Rad51 (Hs00153418-m1), ERa (Hs01046818-m1), TGF-b receptor 1 (TGFb-R1) (Hs100998133-m1), EZH2 (Hs001016789-m1), and miR-221 (TM000524)were obtained from Life Technologies.
Actinomycin D mRNA Stability Assay
A mRNA stability assay was performed exposing MCF7and shBeta MCF7 cells to actinomycin D at a concentrationof 100 ng/mL (Sigma-Aldrich) and assessed by RT-PCR atdifferent time points. The first time point was taken as thereference value for each time series.
ELISA Test
The EZH2 enzyme-linked immunosorbent assay (ELISA)test was purchased from Epigentek (Brooklyn, NY) andperformed following the manufacturer’s instructions.
Western Blot Analysis
Proteins were extracted with radioimmunoprecipitationassay buffer, with the exception of g-H2AX extraction,which was performed by adding HCl to the lysis buffer [10mmol/L HEPES (pH 7.9); 1.5 mmol/L MgCl2; 10 mmol/LKCl] to a final concentration of 0.2 mol/L. Sixty to 100 mgof proteins were assessed by Western blot analysis, usingthe following antibodies: anti-Rad51 (clone 3C10) andantiephospho-ser 139-H2AX (clone JBW301; Millipore,Billerica, MA); anti-Slug (clone L40C6; Cell SignalingTechnology, Danvers, MA); and antieb-catenin (E5) andanti-Actin (C4; Santa Cruz Biotechnology, Santa Cruz, CA).
mRNA Immunoprecipitation Assay
A mRNA immunoprecipitation assay was performed fol-lowing a previously published protocol.35 In brief, cultured
The American Journal of Pathology - ajp.amjpathol.orgFLA 5.2.0 DTD � AJPA1476_proof �
cells were suspended in lysis buffer [100 mmol/L KCl,5 mmol/L MgCl2, 10 mmol/L HEPES (pH 7.0), 0.5%Nonidet P-40] supplemented with RNase and proteaseinhibitors. Proteins were immunoprecipitated using proteinA beads (Santa Cruz Biotechnology) and either antieb-catenin (E5; Santa Cruz Biotechnology) or anti-HuR (Mo-lecular Probes, Eugene, OR), or normal IgG (sc-2025; SantaCruz Biotechnology) mouse antibodies in NT-2 buffer[50 mmol/L Tris (pH 7.4), 150 mmol/L NaCl, 1 mmol/LMgCl2, 0.05% Nonidet P-40], supplemented with RNaseinhibitor (40 U/mL), and dithiothreitol Q(1 mmol/L). Immu-noprecipitates were resuspended in TRIzol reagent (LifeTechnologies). mRNA was assessed by RT-PCR analysis(IL-8 and TNF-a) or real-time PCR (miR-221). Data wereexpressed as the fold increase of each mRNA level bound tomouse-specific antibody over the amount of the samemRNA bound to normal mouse IgG.
Microsatellite Instability
Microsatellite instability analysis was performed throughPCR reaction (Life Technologies, Foster City, CA), using theCC-MSI kit (AB Analitica, Padova, Italy), which allows theamplification of 10 markers (BAT25, BAT26, D2S123,D5S346, D17S250, NR21, NR24, BAT40, TGFb-RII, andD18S58) in two separate reactions. The fluorescent PCRproductswere analyzed inanABI3730XLDNAanalyzer usingthe GeneMapper software version 4.0 (Life Technologies).
In Situ Hybridization Analysis
Fluorescent in situ hybridization analyses for chromo-some 1 and 6 copy number variations were performedon paraformaldehyde-fixed/paraffin-embedded cell pelletsusing centromeric probes CEP1 (D1Z5 Spectrum OrangeProbe) and CEP6 (D6Z1 Spectrum Green Probe) fromAbbott Laboratories (Des Plaines, IL). Twenty nonover-lapping nuclei were counted at �1000 magnification andexamined using a Nikon Eclipse 80i microscope (NikonCorporation, Tokyo, Japan). The cutoff value for each probewas set equal to the mean � 3 SD of the control values.Dual-color silver/chromogenic in situ hybridization analysisfor HER2 gene status and chromosome 17 copy numbervariations was performed by INFORM HER2 Dual ISHDNA Probe Cocktail (Ventana Medical Systems/RocheDiagnostics, Mannheim, Germany). Twenty nonoverlappingnuclei were counted at �600 magnification.
Shotgun Whole-Genome Sequencing
DNA library preparation was carried out according to theIllumina TruSeq DNA Sample Prep protocol (Illumina, SanDiego, CA). A total amount of 2.5 mg of genomic DNA wassonicated to obtain 400-bp fragments, an end-repair stepwas carried out, and an A nucleotide was added to the 30
blunt ends before ligating multiple indexing adapters to the
3
372
12 September 2013 � 2:47 am � EO: AJP13_0196

Q15
Q16
Q17
½F1�½F1�
Figure 1 Hypoxic BMH supernatants enhance MS self-renewal via paracrine TGFb1/autocrine TNF-a crosstalk. A: Second- (II, n Z 3, samples S1, S3, S4),third- (III, n Z 1, sample S4), and fourth- (IV, n Z 1, sample S4) generation T-MS Q27assay on exposure to either 1% pO2 [hypoxic (Hyp)] or BMH supernatants.Normoxic (Nor) T-MS formation is shown as baseline. B: Real-time PCR analysis of TGF-b1 mRNA in Nor and Hyp BMH cells, and of TGFb-R1 expression in Nor/Hyp MCF7-MS. C: T-MS assay (sample S2) in response to hypoxia in presence/absence of 15 ng/mL TGF-b1. D: T-MS assay (sample S2) on BMH supernatants inpresence/absence of 1.5 mg/mL TGF-b1 neutralizing antibody (anti-Tb); Hypoxic T-MS formation is shown as baseline. E: Real-time PCR analysis of TNF-a mRNAlevel in T-MS (sample S3) on BMH supernatants; F: MCF7 MS assay on exposure to either BMH supernatants or 15 ng/mL TGF-b1, in the presence/absence of 1.5mg/mL TNF-a neutralizing antibody (anti-Ta); hypoxic T-MS formation is shown as baseline. Data are presented as means � SD. *P Z 0.05, **P < 0.01, and***P < 0.001 Q28.
Storci et al
373374375376377378379380381382383384385386387388389390391392393394395396397398399400401402403404405406407408409410411412413414415416417418419420421422423424425426427428429430431432433434
435436437438439440441442443444445446447448449450451452453454455456457458459460461462463464465466467468469470471472473474475476477478479480481482483484485486487488489490491492493494495
DNA fragments. The adapter-ligated DNA fragments weresize-selected from gel, and DNA fragments with adapters onboth ends were selectively enriched using 10 cycles of PCR.Libraries were pooled and sequenced using the IlluminaHiSeq 1000 sequencing system and applying standardIllumina protocols with the TruSeq SBS Kit v3-HS andTruSeq PE Cluster Kit v3-cBot-HS kits (lllumina). Cellploidy was evaluated as the ratio between the number ofreads of each sample over the reference genome, usinga window of 1 � 105 bases.
Bioinformatics and Statistics
Bioinformatics analysis on AU-rich (adenylate uridylateerich) elementecontaining mRNA was performed on theonline database AREsite (http://rna.tbi.univie.ac.at/cgi-bin/AREsite.cgi, last accessed). miRNA structure was esti-mated by the mfold Web Server (The RNA Institute, http://mfold.rna.albany.edu, last accessed). Statistical analysiswas performed using SPSS software (IBM, Armonk, NewYork). P values referring to t-test are reported.
Results
Hypoxic Bone Marrow Stromal Cells Enhance MSFormation via TGF-b1eInduced Autocrine TNF-a Loop
Hypoxia triggers stem cell features and pathways in can-cer cells.6,11,13 To reproduce in vitro the hypoxic niche
4FLA 5.2.0 DTD � AJPA1476_proof �
environment, we generated hypoxic MS in presence/absenceof hypoxic BMH supernatants. We observed that BMHconferred an increase in MS formation, compared to hyp-oxia alone (Figure 1A). To gain an insight into thisphenomenon, we availed ourselves of our recent observationthat breast cancereassociated mammary gland fibroblaststrigger MS formation via TGF-b1 secretion.3 We detectedincreased expression of TGF-b1 mRNA in hypoxia-exposedBMH fibroblasts and of TGFb-RI mRNA in hypoxia-exposed MS (Figure 1B). These data prompted us to testthe hypothesis that TGF-b1 signaling is enhanced in thehypoxic stromal niche. We found that TGF-b1 consistentlyincreased hypoxic MS generation (Figure 1C), and thatTGF-b1 neutralizing antibody hampered the BMH super-natanteinduced MS formation (Figure 1D). In keeping withthese results, we observed that hypoxic BMH supernatantstriggered TNF-a expression in MS (Figure 1E). Finally, thecapability of BMH supernatants and TGF-b1 to increase MSformation was tapered by TNF-a neutralizing antibody(Figure 1F). These data show that hypoxic BMH induceCSC’s autocrine TNF-a loop via TGF-b1.
Slug Overexpression Blunts Rad51 Expression andInduces H2AX Serine 139 in Absence of DNA Damage
Slug is a hypoxia-induced regulator of mammary gland stemcells.24 We observed that hypoxic TGF-b1 and BMHsupernatants engendered Slug up-regulation in MS, and thatBMH supernatanteinduced MS formation was drastically
ajp.amjpathol.org - The American Journal of Pathology
496
12 September 2013 � 2:47 am � EO: AJP13_0196
Q18
½F2�½F2�
19
½F3�½F3�
DNA Damage Response and Proinflammatory Phenotype
497498499500501502503504505506507508509510511512513514515516517518519520521522523524525526527528529530531532533534535536537538539540541542543544545546547548549550551552553554555556557558
559560561562563564565566567568569570571572573574575576577578579580581582583584
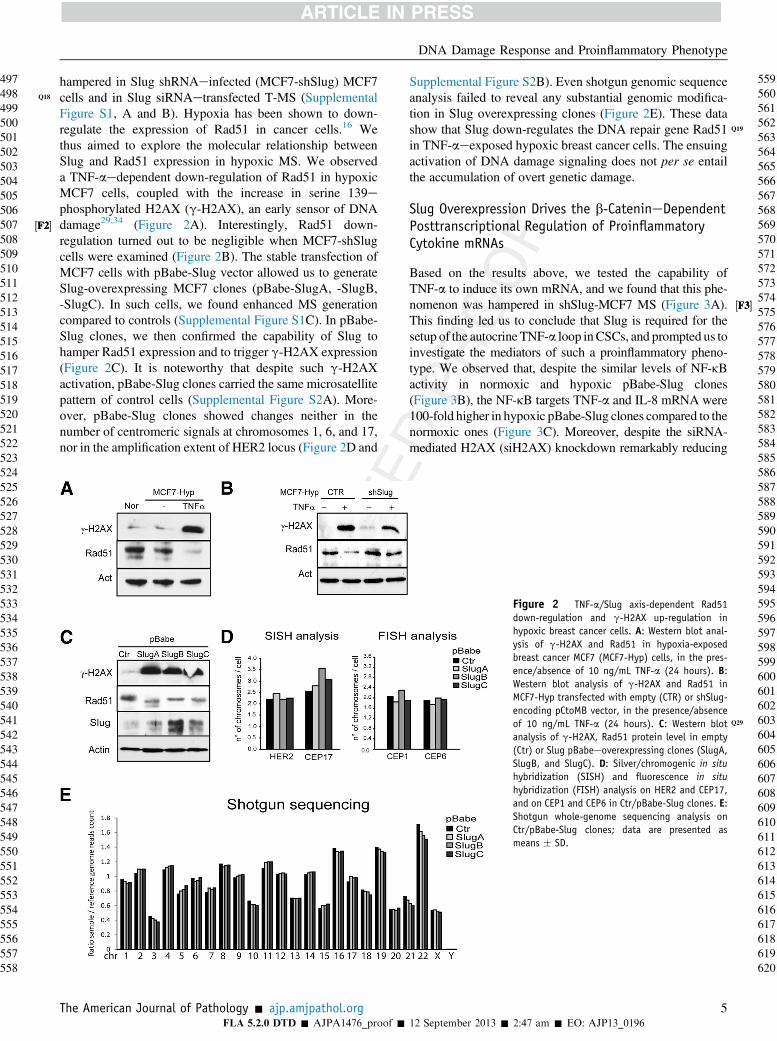
hampered in Slug shRNAeinfected (MCF7-shSlug) MCF7cells and in Slug siRNAetransfected T-MS (SupplementalFigure S1, A and B). Hypoxia has been shown to down-regulate the expression of Rad51 in cancer cells.16 Wethus aimed to explore the molecular relationship betweenSlug and Rad51 expression in hypoxic MS. We observeda TNF-aedependent down-regulation of Rad51 in hypoxicMCF7 cells, coupled with the increase in serine 139ephosphorylated H2AX (g-H2AX), an early sensor of DNAdamage29,34 (Figure 2A). Interestingly, Rad51 down-regulation turned out to be negligible when MCF7-shSlugcells were examined (Figure 2B). The stable transfection ofMCF7 cells with pBabe-Slug vector allowed us to generateSlug-overexpressing MCF7 clones (pBabe-SlugA, -SlugB,-SlugC). In such cells, we found enhanced MS generationcompared to controls (Supplemental Figure S1C). In pBabe-Slug clones, we then confirmed the capability of Slug tohamper Rad51 expression and to trigger g-H2AX expression(Figure 2C). It is noteworthy that despite such g-H2AXactivation, pBabe-Slug clones carried the same microsatellitepattern of control cells (Supplemental Figure S2A). More-over, pBabe-Slug clones showed changes neither in thenumber of centromeric signals at chromosomes 1, 6, and 17,nor in the amplification extent of HER2 locus (Figure 2D and
The American Journal of Pathology - ajp.amjpathol.orgFLA 5.2.0 DTD � AJPA1476_proof �
Supplemental Figure S2B). Even shotgun genomic sequenceanalysis failed to reveal any substantial genomic modifica-tion in Slug overexpressing clones (Figure 2E). These datashow that Slug down-regulates the DNA repair gene Rad51 Q
in TNF-aeexposed hypoxic breast cancer cells. The ensuingactivation of DNA damage signaling does not per se entailthe accumulation of overt genetic damage.
Slug Overexpression Drives the b-CatenineDependentPosttranscriptional Regulation of ProinflammatoryCytokine mRNAs
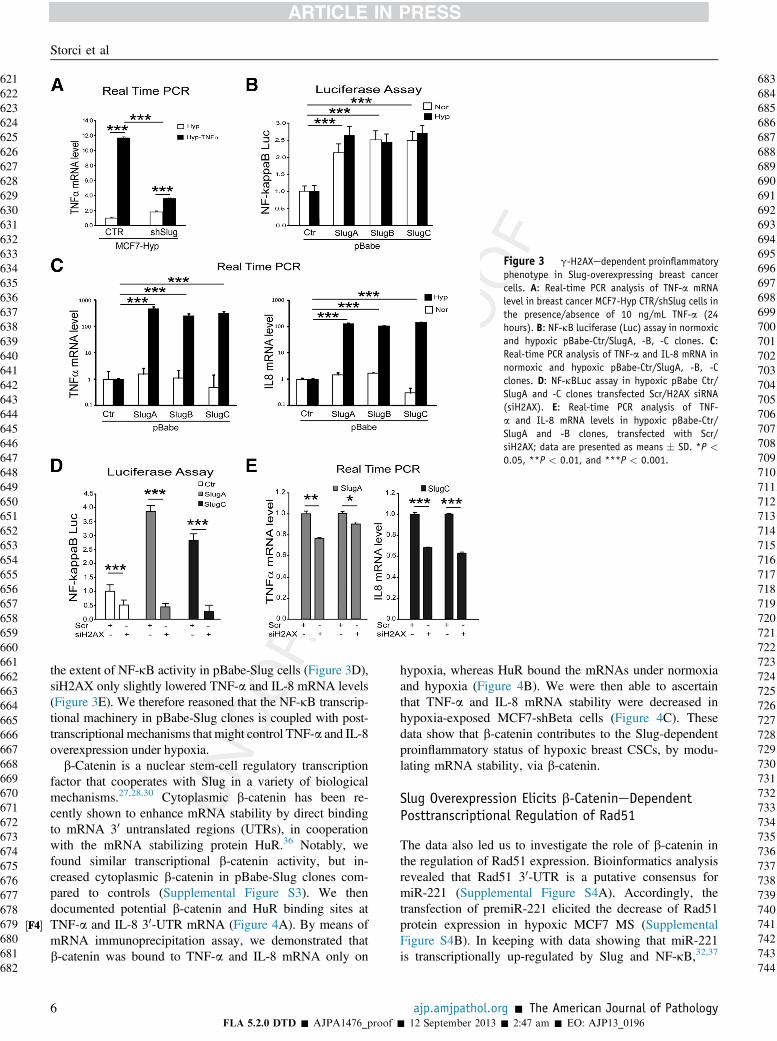
Based on the results above, we tested the capability ofTNF-a to induce its own mRNA, and we found that this phe-nomenon was hampered in shSlug-MCF7 MS (Figure 3A).This finding led us to conclude that Slug is required for thesetup of the autocrine TNF-a loop inCSCs, and prompted us toinvestigate the mediators of such a proinflammatory pheno-type. We observed that, despite the similar levels of NF-kBactivity in normoxic and hypoxic pBabe-Slug clones(Figure 3B), the NF-kB targets TNF-a and IL-8 mRNA were100-fold higher in hypoxic pBabe-Slug clones compared to thenormoxic ones (Figure 3C). Moreover, despite the siRNA-mediated H2AX (siH2AX) knockdown remarkably reducing
Figure 2 TNF-a/Slug axis-dependent Rad51down-regulation and g-H2AX up-regulation inhypoxic breast cancer cells. A: Western blot anal-ysis of g-H2AX and Rad51 in hypoxia-exposedbreast cancer MCF7 (MCF7-Hyp) cells, in the pres-ence/absence of 10 ng/mL TNF-a (24 hours). B:Western blot analysis of g-H2AX and Rad51 inMCF7-Hyp transfected with empty (CTR) or shSlug-encoding pCtoMB vector, in the presence/absenceof 10 ng/mL TNF-a (24 hours) Q29. C: Western blotanalysis of g-H2AX, Rad51 protein level in empty(Ctr) or Slug pBabeeoverexpressing clones (SlugA,SlugB, and SlugC). D: Silver/chromogenic in situhybridization (SISH) and fluorescence in situhybridization (FISH) analysis on HER2 and CEP17,and on CEP1 and CEP6 in Ctr/pBabe-Slug clones. E:Shotgun whole-genome sequencing analysis onCtr/pBabe-Slug clones; data are presented asmeans � SD.
5
585586587588589590591592593594595596597598599600601602603604605606607608609610611612613614615616617618619620
12 September 2013 � 2:47 am � EO: AJP13_0196
½F4�½F4�
Figure 3 g-H2AXedependent proinflammatoryphenotype in Slug-overexpressing breast cancercells. A: Real-time PCR analysis of TNF-a mRNAlevel in breast cancer MCF7-Hyp CTR/shSlug cells inthe presence/absence of 10 ng/mL TNF-a (24hours). B: NF-kB luciferase (Luc) assay in normoxicand hypoxic pBabe-Ctr/SlugA, -B, -C clones. C:Real-time PCR analysis of TNF-a and IL-8 mRNA innormoxic and hypoxic pBabe-Ctr/SlugA, -B, -Cclones. D: NF-kBLuc assay in hypoxic pBabe Ctr/SlugA and -C clones transfected Scr/H2AX siRNA(siH2AX). E: Real-time PCR analysis of TNF-a and IL-8 mRNA levels in hypoxic pBabe-Ctr/SlugA and -B clones, transfected with Scr/siH2AX; data are presented as means � SD. *P <
0.05, **P < 0.01, and ***P < 0.001.
Storci et al
621622623624625626627628629630631632633634635636637638639640641642643644645646647648649650651652653654655656657658659660661662663664665666667668669670671672673674675676677678679680681682
683684685686687688689690691692693694695696697698699700701702703704705706707708709710711712713714715716717718719720721722723724725726727728729730731732733734735736737738739740741742743
the extent of NF-kB activity in pBabe-Slug cells (Figure 3D),siH2AX only slightly lowered TNF-a and IL-8 mRNA levels(Figure 3E). We therefore reasoned that the NF-kB transcrip-tional machinery in pBabe-Slug clones is coupled with post-transcriptional mechanisms that might control TNF-a and IL-8overexpression under hypoxia.
b-Catenin is a nuclear stem-cell regulatory transcriptionfactor that cooperates with Slug in a variety of biologicalmechanisms.27,28,30 Cytoplasmic b-catenin has been re-cently shown to enhance mRNA stability by direct bindingto mRNA 30 untranslated regions (UTRs), in cooperationwith the mRNA stabilizing protein HuR.36 Notably, wefound similar transcriptional b-catenin activity, but in-creased cytoplasmic b-catenin in pBabe-Slug clones com-pared to controls (Supplemental Figure S3). We thendocumented potential b-catenin and HuR binding sites atTNF-a and IL-8 30-UTR mRNA (Figure 4A). By means ofmRNA immunoprecipitation assay, we demonstrated thatb-catenin was bound to TNF-a and IL-8 mRNA only on
6FLA 5.2.0 DTD � AJPA1476_proof �
hypoxia, whereas HuR bound the mRNAs under normoxiaand hypoxia (Figure 4B). We were then able to ascertainthat TNF-a and IL-8 mRNA stability were decreased inhypoxia-exposed MCF7-shBeta cells (Figure 4C). Thesedata show that b-catenin contributes to the Slug-dependentproinflammatory status of hypoxic breast CSCs, by modu-lating mRNA stability, via b-catenin.
Slug Overexpression Elicits b-CatenineDependentPosttranscriptional Regulation of Rad51
The data also led us to investigate the role of b-catenin inthe regulation of Rad51 expression. Bioinformatics analysisrevealed that Rad51 30-UTR is a putative consensus formiR-221 (Supplemental Figure S4A). Accordingly, thetransfection of premiR-221 elicited the decrease of Rad51protein expression in hypoxic MCF7 MS (SupplementalFigure S4B). In keeping with data showing that miR-221is transcriptionally up-regulated by Slug and NF-kB,32,37
ajp.amjpathol.org - The American Journal of Pathology
744
12 September 2013 � 2:47 am � EO: AJP13_0196

½F5�½F5�
Q20
Figure 4 b-Catenin binds and stabilizes TNF-a and IL-8 mRNAs. A:Scheme showing TNF-a and IL-8 mRNAs. b-Catenin and HuR binding sitesare highlighted. B: TNF-a and IL-8 mRNA immunoprecipitation assay inbreast cancer MCF7 Nor/Hyp cells with mouse antieb-catenin, anti-HuR,and IgG antibodies. C: TNF-a and IL-8 mRNA stability assay in MCF7-HypCTR/shBeta cells; data are presented as means � SD. Hyp, hypoxia; Nor,normoxia. ***P < 0.001.
Figure 5 The b-catenin/miR-221 interplay posttranscriptionally down-regulates Rad51 expression. A: Real-time PCR analysis of miR-221 inhypoxic pBabe-Ctr/SlugB, -C clones, transfected with Scr/siH2AX. B: mRNAimmunoprecipitation (IP) assay of miR-221 level in breast cancer MCF7Nor/Hyp cells with mouse IgG and anti-b-catenin antibodies. C: Rad51mRNA stability assay in MCF7-Hyp CTR/shBeta cells. D: WB analysis ofb-catenin and Rad51 in MCF7-Hyp CTR/shBeta cells; data are presented asmeans � SD. Hyp, hypoxia; Nor, normoxia. **P < 0.01, ***P < 0.001.
DNA Damage Response and Proinflammatory Phenotype
745746747748749750751752753754755756757758759760761762763764765766767768769770771772773774775776777778779780781782783784785786787788789790791792793794795796797798799800801802803804805806
807808809810811812813814815816817818819820821822823824825826827828829830831832833834835836837838839840841842843844845846847848849850851852853854855856857858859860861862863864865866867
we observed that miR-221 was down-regulated in siH2AX-transfected pBabe-Slug clones (Figure 5A), as well as inshSlug and shBeta MCF7 cells (Supplemental Figure S4C).Owing to the presence of a 30-ACUUU-50 sequence in themature miR-221 that may potentially bind b-catenin(Supplemental Figure S4D), we tested the hypothesis thatb-catenin might directly bind miR-221 and regulate Rad51expression. We found that on hypoxia, but at a very limitedextent under normoxia, miR-221 was immunoprecipitated bythe b-catenin specific antibody (Figure 5B). We then wereable to show that Rad51 mRNA stability and protein levelwere increased in hypoxic MCF7-shBeta cells (Figure 5, Cand D). Similar data were obtained for ERa mRNA, which
The American Journal of Pathology - ajp.amjpathol.orgFLA 5.2.0 DTD � AJPA1476_proof �
was stabilized in hypoxic MCF7-shBeta cells (SupplementalFigure S5A). Our data also confirmed that ERa is a targetof miR-221, because ERa expression was increased byantiemiR-221 and decreased by premiR-221 administration(Asiedu et al38) (Supplemental Figure S5B). These datahighlight the role of b-catenin/miR-221 circuitry in geneexpression profile of Slug overexpressing cells.
Discussion
This paper started with the observation that hypoxic BMHengender a TNF-aedriven Slug/b-catenin axis, which pro-motes the self-renewal of breast CSCs. In particular,hypoxia elicits TGF-b1 expression in stromal cells andTGFb-R1 in MS, thus facilitating the activation of thepathway, which is expected to be less active (or even latent)in the absence of hypoxia. It is noteworthy that we haverecently found that tumor-associated, but not normal, breastfibroblasts enhance MS formation via a similar paracrine/autocrine TGF-b1/TNF-a interplay.3 TGF-b1 has beendescribed as a potent breast CSC autocrine growth factor22
In turn, the growth-promoting activity of TNF-a on humanMS has been previously reported.23,24 Intriguingly, the twomediators elicit overt stem cell features when coadminis-tered to breast cancer cells.39 These data support the notion
7
868
12 September 2013 � 2:47 am � EO: AJP13_0196

Q21
Q22
Q23
24
25
½F6�½F6�
Figure 6 Scheme showing the sustenance of the CSC proinflammatoryphenotype in a hypoxic environment. In the hypoxic niche, Slug triggersg-H2AX activation, which engenders NF-kBemediated cytokine up-regulation. Slug also induces b-catenin to bind cytoplasmic cytokinemRNAs, thus promoting their stabilization. Moreover, b-catenin bindsmiR221, and this phenomenon associates with enhanced mRNA degrada-tion of miR221 targets, eg, Rad51and ERa. Other potential adenylate uri-dylate (AU)erich or miR-221 targets of such a mechanism, such as EZH2and Brca1, are indicated.
Storci et al
869870871872873874875876877878879880881882883884885886887888889890891892893894895896897898899900901902903904905906907908909910911912913914915916917918919920921922923924925926927928929930
931932933934935936937938939940941942943944945946947948949950951952953954955956957958959960961962963964965966967968969970971972973974975976977978979980981982983984985986987988989990991
that such different environmental stimuli converge on theinduction of the breast CSC phenotype.
Slug is a crucial regulator of mammary gland stem cellfunctioning.28 Published data show that Slug is a hypoxia-induced gene, and that it is a target of the TGF-b1 andTNF-a pathways.7,40 Accordingly, we here demonstrate thatSlug up-regulation follows the microenvironmental stimulithat pertain to the CSCs niche, ie, hypoxia and hypoxicstromal cellederived peptides. Our data suggest that similarclues may be provided by the cancer-associated stroma3 andby the stromal cells to which CSCs disseminate and seed.
We then shed light on major observations that have beenperformed in the recent past. The former is the proin-flammatory phenotype of breast CSCs,3 the latter is theassociation between the down-regulation of DNA repairgenes and the enhancement of breast CSCs features.17
As far as the CSCs proinflammatory phenotype is con-cerned, we find that Slug expression up-regulates the level ofthe DNA damage sensor g-H2AX.34 According to the notionthat the activation of the DNA damage response pathwayengenders the proinflammatory response,19,20 we show thatthe knockdown of the DNA damage sensor g-H2AX bluntsNF-kB activity and the expression of inflammatory media-tors. These data suggest that the proinflammatory phenotypeof CSCs originates from the activation of the DNA damagesensing pathway. Consequently, the accrual of DNA damagecan be expected to occur in such cells. On the contrary,although the onset of minor genomic alterations cannot beexcluded, we did not detect any overt DNA damage in Slugoverexpressing cells. In fact, g-H2AX activation might occurindependently from its activity as a DNA damage sensor.41,42
In particular, g-H2AX activation promotes angiogenesis andstem cell pool maintenance, two processes engendered byhypoxia.11,41,42 Therefore, we can conclude that the proin-flammatory phenotype of breast CSCs, which is not per seassociated with the onset of overt genomic abnormalities,relies on the activation of hypoxic DNA damage response. Inregard to this issue, we show that hypoxia also enhances themRNA stability of proinflammatory peptides via b-catenin.b-Catenin has long been recognized as a master controllerof mammary gland stem cell homeostasis via its nucleartranscriptional activity.11,30 However, cytoplasmic b-cateninbinds and stabilizes various cytoplasmic mRNA,31 includingIl-6 (D’Uva, unpublished data). In accordance with thenotion that posttranscriptional mechanisms gain importanceon hypoxia,44 the b-cateninedependent stabilization ofmRNAs becomes overt in hypoxia-exposed cells. Note-worthy, b-catenin triggers the inflammation-dependent ag-gressive behavior in hepatocellular carcinoma, suggestingthat the results presented here may extend to other tumortypes’ where proinflammatory peptides may act as CSCsgrowth factors.5e8,44
With reference to the relationship between CSCs and thedown-regulation of DNA repair genes, we convey that Slugdown-regulates Rad51 expression. Slug is a transcriptionalrepressor that hinders the expression of the genome
8FLA 5.2.0 DTD � AJPA1476_proof �
gatekeeper Brca1.45 We show that the b-catenin/miR-221interaction in Slug overexpressing cells regulates Rad51mRNA stability. Notably, Rad51 down-regulation has beenpreviously associated with increased breast cancer cell MS-forming capability.17 Such knockdown was achievedthrough the overexpression of a hypoxia-inducible repressorof Rad51, ie, the polycomb receptor complex componentEZH2.17 We found that that EZH2 expression is increasedin Slug overexpressing clones, as well as that EZH2 mRNAstability and protein level are reduced in b-catenin knock-down cells (Supplemental Figure S5, CeE). Following ouradditional observation that Brca1 mRNA half-life is reducedby b-catenin (Supplemental Figure S5F), we speculate thathypoxia operates on the CSCs’ phenotype, not only byeliciting growth-promoting proinflammatory factors (in-cluding EZH2),46 but also by hampering the expression ofgenes (including Brca1 and Rad51) Qthat potentially restrainthe onset of the CSCs phenotype. Because we show thatanother miR-221 target (ERa) is down-regulated by miR-221/b-catenin circuitry, the complex b-cateninedependentreshape of gene expression is expected to generate a profilethat is closely similar to the triple-negative/basal-like breastcancer.47 In such ERa-negative tumors, the overexpressionof a stem cellelike gene expression profile is accompaniedby the down-regulation of Rad51 and the up-regulation ofEZh2 Q, miR-221, and gH2AX.48e50 In conclusion, these datasupport the notion that the CSCs’ proinflammatory pheno-type is sustained by a Slug/b-cateninedriven molecularmachinery (Figure 6) that operates at both the transcriptional
ajp.amjpathol.org - The American Journal of Pathology
992
12 September 2013 � 2:47 am � EO: AJP13_0196

DNA Damage Response and Proinflammatory Phenotype
9939949959969979989991000100110021003100410051006100710081009101010111012101310141015101610171018101910201021102210231024102510261027102810291030103110321033103410351036103710381039104010411042104310441045104610471048104910501051105210531054
1055105610571058105910601061
and posttranscriptional levels to modulate mRNA stability inthe hypoxic environment.
Uncited Reference
43.
106210631064106510661067106810691070107110721073107410751076107710781079108010811082Acknowledgments
We thank Fondazione Carisbo and Fondazione del Montefor supporting the Center for Applied Biomedical Research(C.R.B.A.), St. Orsola-Malpighi University Hospital,Bologna, Italy.
G.S. and M.B. conceived and designed experiments;G.S., S.D.C., S.B., A.P., M.N., P.G., C.P., A.F., G.B., M.F.,E.C., E.G., and M.D. performed the experiments; M.T.,G.B., F.B., C.C., and D.S. provided material; M.B., G.S.,M.D., M.F., and C.F. analyzed data; and M.B. and G.S.wrote the manuscript.
Supplemental Data
Supplemental material for this article can be found athttp://dx.doi.org/10.1016/j.ajpath.2013.07.020.
108310841085108610871088108910901091109210931094109510961097109810991100110111021103110411051106110711081109111011111112111311141115
References
1. Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ,Clarke MF: Prospective identification of tumorigenic breast cancercells. Proc Natl Acad Sci U S A 2003, 100:3983e3988
2. Konopleva MY, Jordan CT: Leukemia stem cells and microenvi-ronment: biology and therapeutic targeting. J Clin Oncol 2011, 29:591e599
3. Papi A, Storci G, Guarnieri T, De Carolis S, Bertoni S, Avenia N,Sanguinetti A, Sidoni A, Santini D, Ceccarelli C, Taffurelli M,Orlandi M, Bonafé M: Peroxisome proliferator activated receptor-a/hypoxia inducible factor-1a interplay sustains carbonic anhydraseIX and apolipoprotein E expression in breast cancer stem cells. PLoSOne 2013, 8:e54968
4. Rajasekhar V, Studer L, Gerald W, Socci ND, Scher HI: Tumor-initiating stem-like cells in human prostate cancer exhibit increasedNF-kB signaling. Nat Commun 2011, 2:162
5. Korkaya H, Liu S, Wicha MS: Breast cancer stem cells, cytokinenetworks, and the tumor microenvironment. J Clin Invest 2011, 121:3804e3809
6. Sansone P, Storci G, Tavolari S, Guarnieri T, Giovannini C,Taffurelli M, Ceccarelli C, Santini D, Paterini P, Marcu KB,Chieco P, Bonafè M: IL-6 triggers malignant features in mammo-spheres from human ductal breast carcinoma and normal mammarygland. J Clin Invest 2007, 117:3988e4002
7. Storci G, Sansone P, Mari S, D’Uva G, Tavolari S, Guarnieri T,Taffurelli M, Ceccarelli C, Santini D, Chieco P, Marcu KB,Bonafè M: TNFalpha up-regulates SLUG via the NF-kappaB/HIF1alpha axis, which imparts breast cancer cells witha stem cell-like phenotype. J Cell Physiol 2010, 225:682e691
8. Korkaya H, Kim GI, Davis A, Malik F, Henry NL, Ithimakin S,Quraishi AA, Tawakkol N, D’Angelo R, Paulson AK, Chung S,Luther T, Paholak HJ, Liu S, Hassan KA, Zen Q, Clouthier SG,Wicha MS: Activation of an IL6 inflammatory loop mediates
The American Journal of Pathology - ajp.amjpathol.orgFLA 5.2.0 DTD � AJPA1476_proof �
trastuzumab resistance in HER2þ breast cancer by expanding thecancer stem cell population. Mol Cell 2012, 47:570e584
9. Prasad S, Ravindran J, Aggarwal BB: NF-kappaB and cancer: howintimate is this relationship. Mol Cell Biochem 2010, 336:25e37
10. Lee KE, Simon MC: From stem cells to cancer stem cells: HIF takesthe stage. Curr Opin Cell Biol 2012, 24:232e235
11. Harrison H, Rogerson L, Gregson HJ, Brennan KR, Clarke RB,Landberg G: Contrasting hypoxic effects on breast cancer stem cellhierarchy is dependent onER-a status. Cancer Res 2013, 73:1420e1433
12. Semenza GL: Hypoxia-inducible factors: mediators of cancerprogression and targets for cancer therapy. Trends Pharmacol Sci2012, 33:207e214
13. Seidel S, Garvalov BK, Wirta V, von Stechow L, Schänzer A,Meletis K, Wolter M, Sommerlad D, Henze AT, Nistér M,Reifenberger G, Lundeberg J, Frisén J, Acker T: A hypoxic nicheregulates glioblastoma stem cells through hypoxia inducible factor 2alpha. Brain 2010, 133:983e995
14. Hartkopf AD, Banys M, Krawczyk N, Staebler A, Becker S,Hoffmann J, Hahn M, Wallwiener M, Fehm T: Bone marrow versussentinel lymph node involvement in breast cancer: a comparison ofearly hematogenous and early lymphatic tumor spread. Breast CancerRes Treat 2012, 131:501e508
15. Lévesque JP, Helwani FM, Winkler IG: The endosteal ‘osteoblastic’niche and its role in hematopoietic stem cell homing and mobiliza-tion. Leukemia 2010, 24:1979e1992
16. Bindra RS, Glazer PM: Repression of RAD51 gene expression byE2F4/p130 complexes in hypoxia. Oncogene 2007, 26:2048e2057
17. Chang CJ, Yang JY, Xia W, Chen CT, Xie X, Chao CH,Woodward WA, Hsu JM, Hortobagyi GN, Hung MC: EZH2promotes expansion of breast tumor initiating cells through activationof RAF1-b-catenin signaling. Cancer Cell 2011, 19:86e100
18. Liu S, Ginestier C, Charafe-Jauffret E, Foco H, Kleer CG,Merajver SD, Dontu G, Wicha MS: BRCA1 regulates humanmammary stem/progenitor cell fate. Proc Natl Acad Sci U S A 2008,105:1680e1685
19. Rodier F, Coppé JP, Patil CK, Hoeijmakers WA, Muñoz DP,Raza SR, Freund A, Campeau E, Davalos AR, Campisi J: PersistentDNA damage signalling triggers senescence-associated inflammatorycytokine secretion. Nat Cell Biol 2009, 11:973e979
20. McCool KW, Miyamoto S: DNA damage-dependent NF-kB activa-tion: NEMO turns nuclear signaling inside out. Immunol Rev 2012,246:311e326
21. Niu J, Shi Y, Tan G, Yang CH, Fan M, Pfeffer LM, Wu ZH: DNAdamage induces NF-kB-dependent microRNA-221 up-regulation andpromotes breast cancer cell invasion. J Biol Chem 2012, 287:21783e21795
22. Scheel C, Eaton EN, Li SH, Chaffer CL, Reinhardt F, Kah KJ,Bell G, Guo W, Rubin J, Richardson AL, Weinberg RA: Paracrineand autocrine signals induce and maintain mesenchymal and stem cellstates in the breast. Cell 2011, 145:926e940
23. Bhat-Nakshatri P, Appaiah H, Ballas C, Pick-Franke P, Goulet R Jr.,Badve S, Srour EF, Nakshatri H: SLUG/SNAI2 and tumor necrosisfactor generate breast cells with CD44þ/CD24- phenotype. BMCCancer 2010, 10:411
24. Storci G, Sansone P, Trere D, Tavolari S, Taffurelli M, Ceccarelli C,Guarnieri T, Paterini P, Pariali M, Montanaro L, Santini D, Chieco P,Bonafé M: The basal-like breast carcinoma phenotype is regulated bySLUG gene expression. J Pathol 2008, 214:25e37
25. Pérez-Caro M, Bermejo-Rodríguez C, González-Herrero I, Sánchez-Beato M, Piris MA, Sánchez-García I: Transcriptomal profiling of thecellular response to DNA damage mediated by Slug (Snai2). Br JCancer 2008, 98:480e488
26. Ikuta T, Kawajiri K: Zinc finger transcription factor Slug is a noveltarget gene of aryl hydrocarbon receptor. Exp Cell Res 2006, 312:3585e3594
27. Proia TA, Keller PJ, Gupta PB, Klebba I, Jones AD, Sedic M,Gilmore H, Tung N, Naber SP, Schnitt S, Lander ES, Kuperwasser C:
9
1116
12 September 2013 � 2:47 am � EO: AJP13_0196

26
Storci et al
11171118111911201121112211231124112511261127112811291130113111321133113411351136113711381139114011411142114311441145114611471148114911501151115211531154115511561157115811591160116111621163116411651166116711681169117011711172117311741175117611771178
11791180118111821183118411851186118711881189119011911192119311941195119611971198119912001201120212031204120512061207120812091210121112121213121412151216121712181219122012211222
Genetic predisposition directs breast cancer phenotype by dictatingprogenitor cell fate. Cell Stem Cell 2011, 8:149e163
28. GuoW, Keckesova Z, Donaher JL, Shibue T, Tischler V, Reinhardt F,Itzkovitz S, Noske A, Zürrer-Härdi U, Bell G, TamWL, Mani SA, vanOudenaarden A, Weinberg RA: Slug and Sox9 cooperatively deter-mine the mammary stem cell state. Cell 2012, 148:1015e1028
29. Rogakou EP, Pilch DR, Orr AH, Ivanova VS, Bonner WM: DNAdouble stranded breaks induce histone H2AX phosphorylation onserine 139. J Biol Chem 1998, 273:5858e5868
30. Korkaya H, Paulson A, Charafe-Jauffret E, Ginestier C, Brown M,Dutcher J, Clouthier SG, Wicha MS: Regulation of mammarystem/progenitor cells by PTEN/Akt/beta-catenin signaling. PLoS Biol2009, 7:e1000121
31. Kim I, Kwak H, Lee HK, Hyun S, Jeong S: b-Catenin recognizesa specific RNA motif in the cyclooxygenase-2 mRNA 30-UTR andinteracts with HuR in colon cancer cells. Nucleic Acids Res 2012, 40:6863e6872
32. Galardi S, Mercatelli N, Farace MG, Ciafrè SA: NF-kB and c-Juninduce the expression of the oncogenic miR-221 and miR-222 inprostate carcinoma and glioblastoma cells. Nucleic Acids Res 2011,39:3892e3902
33. Stinson S, Lackner MR, Adai AT, Yu N, Kim HJ, O’Brien C,Spoerke J, Jhunjhunwala S, Boyd Z, Januario T, Newman RJ, Yue P,Bourgon R, Modrusan Z, Stern HM, Warming S, de Sauvage FJ,Amler L, Yeh RF, Dornan D: miR-221/222 targeting of tricho-rhinophalangeal 1 (TRPS1) promotes epithelial-to-mesenchymaltransition in breast cancer. Sci Signal 2011, 4(186):pt5
34. Krejci L, Altmannova V, Spirek M, Zhao X: Homologous recombi-nation and its regulation. Nucleic Acids Res 2012, 40:5795e5818
35. Tenenbaum SA, Lager PJ, Carson CC, Keene JD: Ribonomics: iden-tifying mRNA subsets in mRNP complexes using antibodies to RNA-binding proteins and genomic arrays. Methods 2002, 26:191e198
36. Heinonen M, Hemmes A, Salmenkivi K, Abdelmohsen K, Vilén ST,Laakso M, Leidenius M, Salo T, Hautaniemi S, Gorospe M,Heikkilä P, Haglund C, Ristimäki A: Role of RNA binding proteinHuR in ductal carcinoma in situ of the breast. J Pathol 2011, 224:529e539
37. Lambertini E, Lolli A, Vezzali F, Penolazzi L, Gambari R, Piva R:Correlation between Slug transcription factor and miR-221 in MDA-MB-231 breast cancer cells. BMC Cancer 2012, 12:445
38. Rao X, Di Leva G, Li M, Fang F, Devlin C, Hartman-Frey C,Burow ME, Ivan M, Croce CM, Nephew KP: MicroRNA-221/222confers breast cancer fulvestrant resistance by regulating multiplesignaling pathways. Oncogene 2011, 30:1082e1097
39. Asiedu MK, Ingle JN, Behrens MD, Radisky DC, Knutson KL:TGFbeta/TNFalpha-mediated epithelial-mesenchymal transitiongenerates breast cancer stem cells with a claudin-low phenotype.Cancer Res 2011, 71:4707e4719
10FLA 5.2.0 DTD � AJPA1476_proof �
40. Yin X, Wolford CC, Chang YS, McConoughey SJ, Ramsey SA,Aderem A, Hai T: ATF3, an adaptive-response gene, enhances TGF{beta} signaling and cancer-initiating cell features in breast cancercells. J Cell Sci 2010, 123:3558e3565
41. Andäng M, Hjerling-Leffler J, Moliner A, Lundgren TK, Castelo-Branco G, Nanou E, Pozas E, Bryja V, Halliez S, Nishimaru H,Wilbertz J, Arenas E, Koltzenburg M, Charnay P, El Manira A,Ibañez CF, Ernfors P: H2AX-dependent GABA(A) receptor regula-tion of stem cell proliferation. Nature 2008, 451:460e464
42. Economopoulou M, Langer HF, Celeste A, Orlova VV, Choi EY,Ma M, Vassilopoulos A, Callen E, Deng C, Bassing CH, Boehm M,Nussenzweig A, Chavakis T: Histone H2AX is integral to hypoxia-driven neovascularization. Nat Med 2009, 15:553e558
43. Gorospe M, Tominaga K, Wu X, Fahling M, Ivan M: Post-tran-scriptional control of the hypoxic response by RNA-binding proteinsand microRNAs. Front Mol Neurosci 2011, 4:7 Q
44. Anson M, Crain-Denoyelle AM, Baud V, Chereau F, Gougelet A,Terris B, Yamagoe S, Colnot S, Viguier M, Perret C, Couty JP:Oncogenic b-catenin triggers an inflammatory response that deter-mines the aggressiveness of hepatocellular carcinoma in mice. J ClinInvest 2012, 122:586e599
45. Wu ZQ, Li XY, Hu CY, Ford M, Kleer CG, Weiss SJ: Canonical Wntsignaling regulates Slug activity and links epithelial-mesenchymaltransition with epigenetic Breast Cancer 1, Early Onset (BRCA1)repression. Proc Natl Acad Sci U S A 2012, 109:16654e16659
46. Lee ST, Li Z, Wu Z, Aau M, Guan P, Karuturi RK, Liou YC, Yu Q:Context-specific regulation of NF-kB target gene expression byEZH2 in breast cancers. Mol Cell 2011, 43:798e810
47. Penault-Llorca F, Viale G: Pathological and molecular diagnosis oftriple-negative breast cancer: a clinical perspective. Ann Oncol 2012,6:vi19-22
48. Scully R, Chen J, Ochs RL, Keegan K, Hoekstra M, Feunteun J,Livingston DM: Dynamic changes of BRCA1 subnuclear locationand phosphorylation state are initiated by DNA damage. Cell 1997,90:425e435
49. Nagelkerke A, Van Kuijk SJ, Sweep FC, Nagtegaal ID,Hoogerbrugge N,Martens JW, TimmermansMA, van Laarhoven HW,Bussink J, Span PN: Constitutive expression of g-H2AX has prog-nostic relevance in triple negative breast cancer. Radiother Oncol 2011,101:39e45
50. Barbano R, Copetti M, Perrone G, Pazienza V, Muscarella LA,Balsamo T, Storlazzi CT, Ripoli M, Rinaldi M, Valori VM,Latiano TP, Maiello E, Stanziale P, Carella M, Mangia A,Pellegrini F, Bisceglia M, Muda AO, Altomare V, Murgo R,Fazio VM, Parrella P: High RAD51 mRNA expression characterizeestrogen receptor-positive/progesterone receptor-negative breastcancer and is associated with patient’s outcome. Int J Cancer 2011,129:536e545
ajp.amjpathol.org - The American Journal of Pathology
122312241225122612271228122912301231123212331234123512361237123812391240
12 September 2013 � 2:47 am � EO: AJP13_0196

Q31
Q32
Q33
12411242124312441245124612471248124912501251125212531254125512561257125812591260126112621263
12641265126612671268126912701271127212731274127512761277127812791280128112821283128412851286
Supplemental Figure S1 Slug expression promotes MS formation in breast cancer cells. A: Real-time PCR analysis of Slug expression in MS exposed tohypoxic (Hyp) TGF-b1 and hypoxic BMH supernatants; Normoxic (Nor) Slug mRNA level in is reported as reference. B: MS formation assay in CTR/shSlug-MCF7cells in the presence of BMH supernatants and T-MS assay in scr/siSlug transfected primary breast cancer cells (sample S1), on exposure to BMH. C: MS assay inpBabe-Ctr/SlugA, -B, and -C clones. *P < 0.05, **P < 0.01, and ***P < 0.001.
Supplemental Figure S2 Absence of genomic alterations in Slug overexpressing clones. A: Microsatellite analysis in pBabe-Ctr/SlugA, -B, and -C clones.B: Dual-color silver/chromogenic in situ hybridization (SISH) for HER2 gene (black) and chromosome 17 (red, magnification �1000 SISH) and fluorescence insitu hybridization (FISH) analysis with centromeric probes for chromosome 1 (orange) and chromosome 6 (green, magnification �400), in pBabe-Ctr/SlugA,-B, and -C clones.
Supplemental Figure S3 b-Cateninedependent posttranscriptional regulation of TNF-a and IL-8 mRNA. A: TOPFLASH activity in normoxic/hypoxicpBabe-Ctr/SlugA, -B, and -C clones. B: Immunofluorescence analysis of b-catenin in pBabe-Ctr/SlugA, -B, and -C clones. Ctr, control; Hyp, hypoxia; Nor,normoxia. Scale bar Z 20 mm.
Supplemental Figure S4 b-Catenin/miR-221edependent regulation of Rad51. A: Schematic representation of Rad51 mRNA 30-UTR with miR-221binding sequence. B: Western blot analysis of Rad51 in premiR CT/221 transfected breast cancer MCF7 cells. C: Real-time PCR analysis of Rad51 mRNAlevel in CTR/shSlug/shBeta MCF7 cells. D: Schematic representation of miR-221 sequence and secondary structure according to mfold software. **P < 0.01.
Supplemental Figure S5 b-Catenin posttranscriptionally regulates ERa, EZH2, and BRCA1 expression. A: ERa mRNA stability assay in CTR/shBeta MCF7-Hyp cells. B: Real-time PCR analysis of ERa mRNA level in MCF7 cells transfected with pre-/anti-miR CT/221. C: Real-time PCR analysis of EZH2 in normoxic(Nor) and hypoxic (Hyp) pBabe Ctr/SlugA and -B clones. D: EZH2 mRNA stability assay in MCF7-Hyp Ctr/shBeta cells. E: EZH2 ELISA test in MCF7 CTR/shBetacells exposed to 10 ng/mL TNF-a (24 hours) and 15 ng/mL TGF-b1 (24 hours). F: Brca1 mRNA stability assay in CTR/shBeta MCF7-Hyp cells; data are presentedas means � SD. CTR, control. **P < 0.01, ***P < 0.001.
FLA 5.2.0 DTD � AJPA1476_proof � 12 September 2013 � 2:47 am � EO: AJP13_0196





























