Embed Size (px)
Citation preview

菌物学报 [email protected] 15 May 2013, 32(3): 485-517 Http://journals.im.ac.cn Mycosystema ISSN1672-6472 CN11-5180/Q © 2013 IMCAS, all rights reserved.
*Corresponding author. E-mail: [email protected] Received: 18-12-2012, accepted: 13-03-2013
Species delimitation for Neonectria coccinea group including the causal agents of beech bark disease in Asia, Europe, and North America Yuuri HIROOKA1* Amy Y. ROSSMAN2 Wen-Ying ZHUANG3 Catalina SALGADO-SALAZAR4 Priscila CHAVERRI4
1Forestry & Forest Products Research Institute, Department of Forest Microbiology, 1 Matsunosato, Tukuba, Ibaraki, 305-8687, JAPAN 2Systematic Mycology and Microbiology Laboratory, United States Department of Agriculture, Agricultural Research Service, 10300
Beltsville Ave., Beltsville, Maryland 20705, USA 3State Key Laboratory of Mycology, Institute of Microbiology, Chinese Academy of Sciences, Beijing 100101, P.R. China 4University of Maryland, Department of Plant Science and Landscape Architecture, 2112 Plant Sciences Building, College Park, Maryland
20742, USA
Abstract: Neonectria coccinea has historically been known as a serious pathogen of beech trees in North America and
Europe. Phylogenetic relationships of this species and its relatives have been determined. Neonectria faginata, previously
referred to as Neo. coccinea var. faginata has been delimited from Neo. coccinea sensu stricto and Neo. punicea. Five
additional lineages supported by relatively high phylogenetic values are allied with Neo. coccinea and its relatives. Until
now, these lineages have not been recognized as species because no significant morphological differences among them were
found. Although each phylogenetic species appeared to be unique in host specificity and geography, relatively few specimens
of these species existed. We re-evaluated these species using newly obtained specimens and isolates from Asia, Europe, and
North America. Based on sequences of multiple loci, specifically act, ITS, LSU, rpb1, tef1 and tub, our results indicate that
these isolates represent four phylogenetic species, Neo. coccinea, Neo. faginata, Neo. microconidia, and Neo. punicea. In our
phylogeny, each species is well supported by high BI PP, MP BP, and ML BP values. The single-locus analyses of ITS and
LSU did not correlate with our species recognition criteria. Based on morphology, phylogeny, host specificity and
geographical traits, these four species are conclusively delimited. Each of them is re-described and illustrated. Epitype
specimens for Sphaeria coccinea and Neonectria coccinea var. faginata are designated. Because no type specimen of Neo.
punicea was found, this name is neotypified. Within Neo. coccinea, Neo. microconidia, and Neo. punicea, some sublineages
were supported by high phylogenetic values. In the future these may warrant recognition as distinct species.
Key words: invasive pathogens, molecular systematics, Neonectria coccinea, Neonectria faginata, Neonectria microconidia,
Neonectria punicea, species concept, typification

ISSN1672-6472 CN11-5180/Q Mycosystema May 15, 2013 Vol.32 No.3
http://journals.im.ac.cn/jwxtcn
486
含亚洲、欧洲、北美洲山毛榉病害病原菌的 Neonectria coccinea 复合群的物种概念 Yuuri HIROOKA1* Amy Y. ROSSMAN2 Wen-Ying ZHUANG3 Catalina SALGADO-SALAZAR4 Priscila CHAVERRI4 1Forestry & Forest Products Research Institute, Department of Forest Microbiology, 1 Matsunosato, Tukuba,
Ibaraki, 305-8687, JAPAN [日本] 2Systematic Mycology and Microbiology Laboratory, United States Department of Agriculture, Agricultural
Research Service, 10300 Beltsville Ave., Beltsville, Maryland 20705, USA [美国] 3State Key Laboratory of Mycology, Institute of Microbiology, Chinese Academy of Sciences, Beijing 100101,
P.R. China [中国] 4University of Maryland, Department of Plant Science and Landscape Architecture, 2112 Plant Sciences
Building, College Park, Maryland 20742, USA [美国]
摘 要:对包含亚洲、欧洲、北美洲山毛榉病害病原菌在内的 Neonectria coccinea 复合群中物种之间的系统演化关系
以及物种概念进行了探讨。Neonectria faginata(曾处理为 Neo. coccinea var. faginata)与狭义的 Neo. coccinea 和 Neo.
punicea 有明显区别。系统树显示 5 个分支与 Neo. coccinea 群的关系较为接近,由于缺乏显著的形态差异,它们始终
没有在物种的等级上区分。根据形态学、系统发育、寄主专化性和地理因素,采用多基因 act、ITS、LSU、rpb1、tef1
和 tub 序列分析的方法,对亚洲、欧洲、北美洲的相关材料进行了研究,结果表明,它们代表 4 个独立的物种,即
Neo. coccinea,Neo. faginata,Neo. microconidia 和 Neo. punicea,对每个种进行了详尽的描述和图示。并为 Sphaeria
coccinea 和 Neonectria coccinea var. faginata 指定了附加模式。
关键词:入侵性病原菌,分子系统学,猩红新丛赤壳,Neonectria faginata,小孢新丛赤壳,Neonectria punicea,物种
概念,选模式
INTRODUCTION The beech bark disease (BBD) constitutes one
of the greatest global threats to beech forests. This disease occurs following the interaction between the beech scale insect, Cryptococcus fagisuga Lind. and one of the three ascomycetous fungi, Neonectria coccinea (Pers.) Rossman & Samuels, Neo. faginata (M.L. Lohman, A.M.J. Watson & Ayers) Castl. & Rossman, and Neo. ditissima (Tul. & C. Tul.) Samuels & Rossman. The disease was first documented in Germany in 1849 (McIntosh 1849).
The beech scale insect was introduced into Nova Scotia through seedling beech trees brought from Europe in 1890 (Ehrlich 1934; Hewitt 1914). Recent molecular studies indicate that the causal agents of BBD were introduced from Europe, yet it is unclear if the beech scale insect and BBD fungal pathogens were introduced into North America at the same time (Mahoney et al. 1999; Gwiazdowski et al. 2006). The disease has now rapidly spread across the northeastern United States and southeastern Canada (Houston 1994; Griffin et al. 2003) with

Yuuri HIROOKA et al. / Species delimitation for Neonectria coccinea group including the causal agents of beech bark disease in Asia, …
Mycosystema
487
80%–90% mortality rates of mature beech (Houston 1994).
The BBD pathogenic fungi are Neonectria coccinea on Fagus in Europe, Neo. faginata on Fagus in North America, and Neo. ditissima on hardwoods in North America and Europe, rarely Asia. These fungi have red, globose, slightly roughened perithecia, 1-septate ascospores, and cylindrocarpon-like anamorphs (Rossman et al. 1999). They are morphologically similar and often misidentified because they are distinguished by only small differences in ascospore size. Castlebury et al. (2006) conducted taxonomic studies of these fungi based on morphological observation and combined phylogenetic analyses of rpb2, tef1, and tub gene regions. They clarified the differences between Neonectria coccinea, Neo. ditissima, and Neo. faginata (=Neo. coccinea var. faginata), newly recognized at the species rank. The phylogenetic tree of Castlebury et al. (2006) also showed that potentially five distinct species exist that are sister to Neo. coccinea. Although each phylogenetic species seemed to be segregated by host specificity and geography, no significant morphological differences among them were found due to the limited number of samples.
Neonectria coccinea was originally described as Sphaeria coccinea Pers.: Fr. by Persoon (1800). After Fries (1823) sanctioned Sphaeria coccinea, he transferred this species to Nectria (Fries 1849). Because Persoon (1800) and Fries (1823, 1849) did not typify any specimens, Booth (1959) found several Persoon specimens of S. coccinea and designated one of them as the lectotype. Since the 19th century, the broadly conceived genus Nectria has been narrowed. Most recently the genus Nectria was determined to include fungi having red perithecia without bright yellow scurf and
tubercularia-like anamorph (Hirooka et al. 2011, 2012). Previously considered to be a synonym of Nectria, Rossman et al. (1999) resurrected the genus Neonectria Wollenw. (Wollenweber 1916), typified by Neo. ramulariae Wollenw., for nectria-like species having a cylindrocarpon-like anamorph. Many nectrioid species having cylindrocarpon-like anamorphs were included in the genus Neonectria. Chaverri et al. (2011) divided Neonectria into five genera: Campylocarpon Halleen, Schroers & Crous, Ilyonectria P. Chaverri & C. Salgado, Neonectria Wollenw., Rugonectria P. Chaverri & Samuels, and Thelonectria P. Chaverri & C. Salgado. These genera were defined based on a multiple locus phylogeny and morphological observations.
Neonectria coccinea with its anamorph Cylindrocarpon candidum (Link: Fr.) Wollenw. has been considered a common species distributed widely in the northern hemisphere, frequently reported on twigs and wood of beech trees as well as on other deciduous trees, dung, and soil (Domsch et al. 2007). The basionym of the anamorphic fungus, Fusidium candidum Link, was described by Link (1809) and sanctioned by Fries (1832). Wollenweber (1928) transferred Fusidium candidum to Cylindrocarpon. Booth (1966) first introduced the teleomorph-anamorph connection as states of one species, and recent studies have confirmed the connection. The recent study by Castlebury et al. (2006) restricted this species to Fagus in Europe.
Using combined sequences of several loci, a number of new species within Neonectria sensu lato were recently described from China (Luo & Zhuang 2010; Zhao et al. 2011). Neonectria confusa J. Luo & W.Y. Zhuang was placed in the Neo. coccinea group while Neo. microconidia J. Luo, P. Zhao & W.Y. Zhuang was sister to the Neo. coccinea clade. Although Luo & Zhuang (2010) and Zhao et al.

ISSN1672-6472 CN11-5180/Q Mycosystema May 15, 2013 Vol.32 No.3
http://journals.im.ac.cn/jwxtcn
488
(2011) recognized Neo. confusa and Neo. microconidia as distinct species, the variation between them was relatively low with subtle morphological differences from other species in the Neo. coccinea group. A taxonomic reappraisal of the Neo. coccinea group is needed.
To determine the species circumscription within the Neo. coccinea group, detailed morphological and molecular phylogenetic analyses were undertaken. Many isolates from freshly collected and herbarium specimens around the northern hemisphere, particularly Asia, Europe, and North America, were used to define the species in the Neo. coccinea group. In order to identify the most suitable locus for recognition of cryptic species in this group, the multi-locus genealogies as well as a single locus approach were compared. Each species is described and illustrated, and a key is provided.
1 MATERIALS AND METHODS Specimens and isolates
Specimens and isolates of the Neo. coccinea group preserved in herbaria and culture collections were obtained and examined in this study. Fresh specimens of the teleomorphs were collected and sent by authors or collaborators. These specimens are deposited in the U.S. National Fungus Collections (BPI), Beltsville, Maryland, United States. Single ascospore or conidial cultures were established as described earlier (Hirooka et al. 2011, 2012). Newly isolated cultures are preserved at the CBS Fungal Biodiversity Centre (CBS, Utrecht, Netherlands), and/or GenBank, National Institute of Agrobiological Sciences (NIAS, Tsukuba, Ibaraki, Japan). Morphology
Methods of morphological observation are described by Hirooka et al. (2011, 2012). To observe sporulating structures, the cultures were grown on a
low nutrient agar (SNA; Nirenberg 1976). Cultures on SNA were incubated at 25℃ with alternating 12h/12h fluorescent light/darkness for 2–3 wks. Images were captured with a Nikon DXM1200 digital camera. Some composite images were made with Helicon Focus version 4.21.5 Pro (Helicon Soft, www.heliconfocus.com). Color terminology used for perithecia, ascospores, conidia, and top and reverse colony, were based on Kornerup & Wanscher (1978). Statistical analysis
Measurements of morphological characters such as length and width were made using Scion Image software beta version 4.0.2 (Scion Corporation, Frederick, Maryland, USA) and are based on up to 50 measurements for structures in each isolate and specimen. For morphological structures, descriptive statistics (minimum, mean, median, maximum and standard deviation) were computed. All computations were performed using Systat 10 (Systat Software, San José, California, USA). Only isolates for which all data existed were included in the analysis. Ranges are reported as mean values ± one standard deviation; the number of items measured is given in parentheses together with maximum and minimum. Growth trial
Disks of 5mm diam. were cut from the edge of young colonies and placed in the center of PDA plates, then incubated at temperatures from 15℃ to 35℃ at 5℃ intervals in complete darkness. Diameters of the colonies on three plates for each isolate at each temperature were measured daily for ten days. This trial was replicated three times for each isolate. DNA extraction, PCR, and sequencing
In total, 75 cultures of Neo. coccinea group and three isolates of Neonectria hederae (C. Booth)

Yuuri HIROOKA et al. / Species delimitation for Neonectria coccinea group including the causal agents of beech bark disease in Asia, …
Mycosystema
489
Castl. & Rossman (Table 1) were grown in Difco™ potato dextrose broth in 6cm diam. Petri plates for about one to two weeks at 20℃. Mycelial mats were obtained in a laminar flow hood and dried with clean, absorbent paper towels. DNA was extracted with Ultra Clean™ Plant DNA Isolation Kit (MO BIO Laboratories Inc., Solana Beach, California, USA).
For each isolate, six loci were sequenced, namely α-actin (act) (Carbone & Kohn 1999), β-tubulin (tub) (O’Donnell & Cigelnik 1997), RNA polymerase II subunit one (rpb1) (Castlebury et al. 2004), the internal transcribed spacer (ITS) (White et al. 1990), large subunit nuclear ribosomal DNA (LSU) (Vilgalys n.d.), and translation elongation factor 1-α (tef1) (Carbone & Kohn 1999; Rehner 2001). The primers and PCR protocol information are listed in Tables 2. PCR products were purified with ExoSAP-IT® (USB Corporation, Cleveland, Ohio, USA) following the manufacturer’s instructions. Clean PCR products were sequenced at the DNA Sequencing Facility (Center for Agricultural Biotechnology, University of Maryland, College Park, Maryland, USA) and at MCLAB (Molecular Cloning Laboratories, San Francisco, California, USA). Sequences were assembled and edited with Sequencher 4.9 (Gene Codes, Madison, Wisconsin, USA). Sequences are deposited in GenBank (Table 1). Phylogenetic analyses
Sequences of the six loci were aligned with MAFFT version 6 (Katoh 2008) and the alignment was visually improved with Mesquite version 2.74 (Maddison & Maddison 2010). Phylogenetic analyses with aligned sequences were performed using the Bayesian (BI), Maximum likelihood (ML) and Maximum parsimony (MP) analyses. Three isolates of Neo. hederae, CBS 714.97, A.R. 4523
and IMI 058770a, were used as the outgroup for inferring inter- and intra-specific relationships.
JMODELTEST (Posada 2008) was used to calculate the models of nucleotide substitutions for each gene/partition for the ML and BI analyses. The number of substitution schemes was set to 11, base frequencies +F, rate variation +I and +G, and the base tree for likelihood calculations was set to ‘ML optimized’. Eighty-eight models were compared. After calculation of likelihood scores, the models were selected according to the Akaike information criterion (AIC) (Posada & Buckley 2004). After jMODELTEST, likelihood settings for trees were set to each gene (Table 2).
BI analysis was done using MrBayes 3.1.2 (Huelsenbeck et al. 2001, 2002). BI data were partitioned by locus and the parameters of the nucleotide substitution models for each partition were set as described (Table 2). For these analyses, two independent analyses of two parallel runs and four chains were carried out for 10,000,000 generations using MrBayes. Analyses were initiated from a random tree and trees sampled every 100th generation. The first 20% of the resulting trees were eliminated (=‘burn in’). A consensus tree (‘sumt’ option) and posterior probabilities (PP) were calculated in MrBayes.
For the ML and bootstrap analyses (BP), GARLI version 0.96 (Zwickl 2006) was computed through the Grid computing (Cummings & Huskamp 2005) and The Lattice Project (Bazinet & Cummings 2008), which includes clusters and desktops in one integrated network (Myers et al. 2008). In GARLI, the starting tree was made by stepwise-addition and the number of runs or search replicates was set to 50. 2000 ML BP replicates were done in GARLI, with the starting tree chosen randomly.

ISSN1672-6472 CN11-5180/Q Mycosystema May 15, 2013 Vol.32 No.3
http://journals.im.ac.cn/jwxtcn
490
Tabl
e 1
Isol
ates
and
acc
essio
n nu
mbe
rs u
sed
in th
e ph
ylog
enet
ic a
naly
ses
Gen
Ban
k ac
cess
ion
No.
Sp
ecie
s Is
olat
e N
o.
Her
bariu
m N
o. S
ubst
rate
/Hos
t C
ount
ry
act
ITS
LSU
rp
b1
tef1
tu
b
Neo
nect
ria
cocc
inea
A
.R. 3
696=
CBS
1195
34
BPI
871
112
Fagu
s syl
vatic
a Fr
ance
K
C66
0420
K
C66
0501
K
C66
0619
K
C66
0670
K
C66
0436
K
C66
0723
Neo
. coc
cine
a A
.R. 3
694=
CBS
1189
14
BPI
870
938
Fagu
s syl
vatic
a Fr
ance
K
C66
0419
K
C66
0500
K
C66
0607
K
C66
0669
K
C66
0435
K
C66
0725
Neo
. coc
cine
a C
BS
1191
58=G
.J.S.
98-
114
BPI
748
295
Fagu
s sp.
G
erm
any
KC
6604
22
KC
6605
21
KC
6606
20
KC
6606
72
JF26
8734
* K
C66
0727
Neo
. coc
cine
a M
AFF
241
561=
TPP-
h430
B
PI 8
8194
2 Fa
gus c
rena
ta
Japa
n K
C66
0426
K
C66
0548
K
C66
0552
K
C66
0674
K
C66
0493
K
C66
0729
Neo
. coc
cine
a A
.R. 3
708=
CBS
1189
16
BPI
870
939
Fagu
s syl
vatic
a R
oman
ia
KC
6604
25
KC
6605
05
KC
6606
01
KC
6606
64
KC
6604
42
KC
6607
19
Neo
. coc
cine
a A
.R. 3
707=
CBS
1189
15
BPI
870
937
Fagu
s syl
vatic
a R
oman
ia
KC
6604
16
- K
C66
0578
K
C66
0666
K
C66
0441
K
C66
0718
Neo
. coc
cine
a G.
J.S. 9
2-33
=CBS
134
254
BPI
802
648
Fagu
s sp.
Sc
otla
nd
KC
6604
18
- K
C66
0581
K
C66
0673
K
C66
0467
K
C66
0722
Neo
. coc
cine
a A
.R. 3
700=
CBS
1191
56
BPI
870
940
Fagu
s syl
vatic
a Sl
ovak
ia
KC
6604
21
- K
C66
0579
K
C66
0671
K
C66
0437
K
C66
0726
Neo
. coc
cine
a A
.R. 3
705=
CBS
1191
50
BPI
870
941
Fagu
s syl
vatic
a Sl
ovak
ia
KC
6604
17
KC
6605
04
KC
6605
77
KC
6606
65
KC
6604
40
KC
6607
17
Neo
. coc
cine
a A
.R. 3
691=
CBS
1195
23
BPI
871
113
Fagu
s syl
vatic
a Sl
ovak
ia
KC
6604
23
KC
6604
99
KC
6606
09
KC
6606
68
KC
6604
34
KC
6607
21
Neo
. coc
cine
a A
.R. 3
712=
CBS
1195
22
BPI
871
115
Fagu
s syl
vatic
a Sl
ovak
ia
KC
6604
24
KC
6605
06
KC
6605
93
KC
6606
67
KC
6604
44
KC
6607
24
Neo
. coc
cine
a A
.R. 3
701
BPI
871
114
Fagu
s syl
vatic
a Sl
ovak
ia
KC
6604
15
KC
6605
02
KC
6605
91
KC
6606
63
KC
6604
38
KC
6607
16
Neo
. coc
cine
a A
.R. 3
687=
CBS
1342
53
- Fa
gus s
ylva
tica
Slov
akia
-
- -
- -
KC
6607
20
Neo
. fag
inat
a=
Cyl
indr
ocar
pon
fagi
natu
m C
BS
217.
67=A
TCC
1654
7=IM
I
1057
38
Fa
gus g
rand
ifolia
Can
ada
KC
6604
12
HQ
8403
85*
HQ
8403
82*
HQ
8403
93*
JF26
8746
* JF
2687
30*
Neo
. fag
inat
a A
.R. 4
307=
CBS
1342
46
BPI
878
329
Fagu
s gra
ndifo
liaU
nite
d St
ates
KC
6604
09
KC
6605
19
KC
6606
00
KC
6606
53
KC
6604
57
KC
6607
43
Neo
. fag
inat
a A
.R. 3
677=
CB
S 11
9154
-
Fagu
s gra
ndifo
liaU
nite
d St
ates
- -
- -
- K
C66
0742
Neo
. fag
inat
a A
.R. 3
682=
CB
S 11
9155
-
Fagu
s gra
ndifo
liaU
n it e
d St
ates
- -
- -
- K
C66
0733
Neo
. fag
inat
a N
CF
346=
AR
368
0=C
BS
1189
83
BPI
880
528
Fagu
s sp.
U
nite
d St
ates
KC
6604
07
KC
6604
98
KC
6605
71
KC
6606
55
KC
6604
33
KC
6607
41
Neo
. fag
inat
a A
.R. 4
130=
CBS
1191
61
BPI
871
095
Fagu
s gra
ndifo
liaU
nite
d St
ates
KC
6604
04
KC
6605
09
KC
6605
57
KC
6606
61
KC
6604
47
KC
6607
36
Neo
. fag
inat
a A
.R. 4
152=
CBS
1189
17
BPI
870
943
Fagu
s gra
ndifo
liaU
nite
d St
ates
KC
6604
08
KC
6605
13
KC
6605
62
KC
6606
58
KC
6604
51
KC
6607
35
Neo
. fag
inat
a A
.R. 4
149=
CBS
1195
19
BPI
871
123
Fagu
s syl
vatic
a U
nite
d St
ates
- K
C66
0511
K
C66
0559
K
C66
0651
K
C66
0449
K
C66
0730
Neo
. fag
inat
a A
.R. 4
197=
CBS
1189
18
BPI
870
945
Fagu
s gra
ndifo
liaU
nite
d St
ates
- K
C66
0518
K
C66
0560
K
C66
0657
K
C66
0456
D
Q78
9856
*
Neo
. fag
inat
a A
.R. 4
167=
CBS
1191
99
BPI
871
125
Fagu
s syl
vatic
a U
nite
d St
ates
KC
6604
06
KC
6605
17
KC
6605
55
KC
6606
52
KC
6604
55
KC
6607
40

Yuuri HIROOKA et al. / Species delimitation for Neonectria coccinea group including the causal agents of beech bark disease in Asia, …
Mycosystema
491
Ta
ble
1 co
ntin
ued
Neo
. fag
inat
a A
.R. 4
153=
CBS
1191
63
BPI
871
097
Fagu
s gra
ndifo
liaU
nite
d St
ates
KC
6604
05
KC
6605
14
KC
6605
54
KC
6606
56
KC
6604
52
KC
6607
32
Neo
. fag
inat
a A
.R. 4
151=
CBS
1191
62
BPI
871
096
Fagu
s gra
ndifo
liaU
nite
d St
ates
KC
6604
10
KC
6605
12
KC
6605
72
KC
6606
59
KC
6604
50
KC
6607
37
Neo
. fag
inat
a A
.R. 4
148=
CBS
1195
24
BPI
871
122
Fagu
s syl
vatic
a U
nite
d St
ates
KC
6604
13
KC
6605
10
KC
6606
13
KC
6606
62
KC
6604
48
KC
6607
31
Neo
. fag
inat
a A
.R. 4
097=
CBS
1189
38
BPI
870
942
Fagu
s gra
ndifo
liaU
nite
d St
ates
KC
6604
11
KC
6605
08
KC
6606
14
KC
6606
54
KC
6604
46
KC
6607
39
Neo
. fag
inat
a A
.R. 4
166=
CBS
1191
98
BPI
871
124
Fagu
s syl
vatic
a U
nite
d St
ates
KC
6604
14
KC
6605
16
KC
6605
51
KC
6606
60
KC
6604
54
KC
6607
38
Neo
. hed
erae
A
.R. 4
523=
CBS
1251
75
BPI
878
947
Hed
era
helix
Fr
ance
K
C66
0428
K
C66
0520
K
C66
0615
K
C66
0691
K
C66
0459
K
C66
0760
Neo
. hed
erae
C
BS
714.
97
BPI
871
043
(drie
d cu
lture
)
Hed
era
helix
N
ethe
rland
s K
C66
0429
-
KC
6606
16
KC
6606
93
KC
6604
61
DQ
7898
78*
Neo
. hed
erae
IM
I 058
770a
B
PI 8
7104
4
(drie
d cu
lture
)
Hed
era
helix
U
nite
d
Kin
gdom
KC
6604
30
AJ0
0925
4*K
C66
0617
K
C66
0692
D
Q78
9752
*D
Q78
9895
*
Neo
. mic
roco
nidi
a H
MA
S 98
294
W.Y
. Zhu
ang
and
Y. N
ong
5639
Twig
s C
hina
K
C66
0402
K
C66
0530
K
C66
0587
K
C66
0680
JF
2687
39*
JF26
8724
*
Neo
. mic
roco
nidi
a M
AFF
241
530=
TPP-
h260
B
PI 8
8209
8 B
ark
of d
ead
woo
dJa
pan
KC
6603
89
- K
C66
0618
K
C66
0678
K
C66
0478
K
C66
0757
Neo
. mic
roco
nidi
a M
AFF
241
556=
TPP-
h385
B
PI 8
8193
1 B
ark
of d
ead
woo
dJa
pan
KC
6603
98
KC
6605
45
KC
6605
85
KC
6606
86
KC
6604
89
KC
6607
58
Neo
. mic
roco
nidi
a M
AFF
241
560=
TPP-
h427
B
PI 8
8194
1 Tw
igs
Japa
n K
C66
0391
K
C66
0547
K
C66
0588
K
C66
0677
K
C66
0492
K
C66
0748
Neo
. mic
roco
nidi
a M
AFF
241
558=
TPP-
h390
B
PI 8
8193
3 C
eras
us
Jam
asak
ura
Japa
n K
C66
0390
K
C66
0546
K
C66
0580
K
C66
0681
K
C66
0490
K
C66
0752
Neo
. mic
roco
nidi
a M
AFF
241
506=
TPP-
h120
-
Stum
p Ja
pan
KC
6603
92
- K
C66
0583
K
C66
0684
K
C66
0472
K
C66
0746
Neo
. mic
roco
nidi
a M
AFF
241
570=
TPP-
h521
B
PI 8
8195
6 Fa
gus c
rena
ta
Japa
n K
C66
0395
K
C66
0549
K
C66
0608
K
C66
0683
K
C66
0494
K
C66
0751
Neo
. mic
roco
nidi
a M
AFF
241
559=
TPP-
h391
B
PI 8
8193
4 B
ark
of d
ead
woo
d
Japa
n K
C66
0394
-
KC
6605
84
KC
6606
76
KC
6604
91
KC
6607
53
Neo
. mic
roco
nidi
a M
AFF
241
555=
TPP-
h378
B
PI 8
8193
0 B
ark
of d
ead
woo
dJa
pan
KC
6603
99
- K
C66
0590
K
C66
0679
K
C66
0488
K
C66
0750
Neo
. mic
roco
nidi
a M
AFF
241
516=
TPP-
h176
B
PI 8
8216
3 Tw
igs
Japa
n K
C66
0393
-
KC
6606
21
KC
6606
85
KC
6604
75
KC
6607
49
Neo
. mic
roco
nidi
a M
AFF
241
518=
TPP-
h190
B
PI 8
8216
5 Tw
igs
Japa
n K
C66
0396
K
C66
0534
K
C66
0582
K
C66
0687
K
C66
0476
K
C66
0756
Neo
. mic
roco
nidi
a M
AFF
241
572=
TPP-
h542
B
PI 8
8195
8 Vi
tis c
oign
etia
e Ja
pan
KC
6604
01
KC
6605
50
KC
6605
76
KC
6606
89
KC
6604
95
KC
6607
55
Neo
. mic
roco
nidi
a M
AFF
241
493=
TPP-
h64
BPI
882
137
Wei
gela
cora
eens
is
Japa
n K
C66
0388
-
- K
C66
0675
K
C66
0471
K
C66
0745

ISSN1672-6472 CN11-5180/Q Mycosystema May 15, 2013 Vol.32 No.3
http://journals.im.ac.cn/jwxtcn
492
Ta
ble
1 co
ntin
ued
Neo
. mic
roco
nidi
a M
AFF
241
522=
TPP-
h227
-
Twig
s Ja
pan
KC
6604
27
KC
6605
35
KC
6605
96
KC
6606
90
KC
6604
77
KC
6607
59
Neo
. mic
roco
nidi
a M
AFF
241
514=
TPP-
h171
B
PI 8
8216
1 B
ark
of d
ead
woo
dJa
pan
KC
6604
00
KC
6605
33
KC
6605
97
KC
6606
88
KC
6604
74
KC
6607
54
Neo
. mic
roco
nidi
a M
AFF
241
552=
TPP-
h341
B
PI 8
8211
9 St
em
Japa
n K
C66
0397
K
C66
0544
K
C66
0586
K
C66
0682
K
C66
0487
K
C66
0747
Neo
. pun
icea
A
.R. 3
102=
CBS
1197
24
BPI
871
063
Fran
gula
aln
us
Aus
tria
KC
6603
86
KC
6604
96
KC
6605
68
KC
6606
49
KC
6604
31
DQ
7898
24*
Neo
. pun
icea
A
.R. 4
155=
CBS
1195
27
BPI
871
062
Rham
nus f
alla
x A
ustri
a K
C66
0403
K
C66
0515
K
C66
0575
K
C66
0647
K
C66
0453
D
Q78
9854
*
Neo
. pun
icea
H
MA
S 99
197
Y. N
ong,
W.Y
.
Zhua
ng 5
694
Twig
s of
a
dico
tyle
don
tree
Chi
na
KC
6603
67
KC
6605
31
KC
6605
58
KC
6606
42
JF26
8735
* K
C66
0714
Neo
. pun
icea
G.
J.S. 9
8-13
3=C
BS
1342
55
BPI
748
311
Har
dwoo
d tre
e Fr
ance
K
C66
0364
K
C66
0529
K
C66
0598
K
C66
0624
K
C66
0469
K
C66
0701
Neo
. pun
icea
A
.R. 4
522=
CBS
1342
47
BPI
878
875
Fran
gula
aln
us
Fran
ce
KC
6603
85
- K
C66
0567
K
C66
0648
K
C66
0458
K
C66
0715
Neo
. pun
icea
C
BS
125.
24=I
MI
1138
80=M
UC
L 98
08
BPI
871
038
Bar
k G
erm
any
KC
6603
80
- K
C66
0573
K
C66
0643
D
Q78
9719
*D
Q78
9862
*
Neo
. pun
icea
C
BS
208.
30
BPI
8710
35=C
entr
aalb
urea
u vo
or
Schi
mm
elcu
ltu
res H
erba
rium
H-1
1427
Ulm
us sp
. G
erm
any
KC
6603
82
- K
C66
0611
K
C66
0645
K
C66
0460
D
Q78
9867
*
Neo
. pun
icea
C
BS
242.
29
Rh
amnu
s sp.
G
erm
any
KC
6603
87
KC
6605
22
KC
6605
65
KC
6606
50
DQ
7897
30*
DQ
7898
73*
Neo
. pun
icea
M
AFF
241
548=
TPP-
h328
B
PI 8
8211
5 Tw
igs
Japa
n K
C66
0372
K
C66
0541
K
C66
0569
K
C66
0637
K
C66
0484
K
C66
0713
Neo
. pun
icea
M
AFF
241
546=
TPP-
h326
B
PI 8
8211
3 Pr
unus
×ye
doen
sis
Japa
n K
C66
0371
K
C66
0539
K
C66
0604
K
C66
0636
K
C66
0482
K
C66
0709
Neo
. pun
icea
M
AFF
241
550=
TPP-
h330
B
PI 8
8211
7 Tw
igs
Japa
n K
C66
0374
K
C66
0543
K
C66
0570
K
C66
0639
K
C66
0486
K
C66
0708
Neo
. pun
icea
M
AFF
241
513=
TPP-
h166
B
PI 8
8216
0 B
ark
of d
ead
woo
d
Japa
n K
C66
0368
K
C66
0532
K
C66
0594
K
C66
0632
K
C66
0473
K
C66
0705
Neo
. pun
icea
M
AFF
241
540=
TPP-
h296
-
Twig
s Ja
pan
KC
6603
70
KC
6605
37
KC
6605
95
KC
6606
34
KC
6604
80
KC
6607
12
Neo
. pun
icea
M
AFF
241
547=
TPP-
h327
B
PI 8
8211
4 B
ark
Japa
n K
C66
0375
K
C66
0540
K
C66
0561
K
C66
0641
K
C66
0483
K
C66
0710
Neo
. pun
icea
M
AFF
241
541=
TPP-
h297
B
PI 8
8210
7 Tw
igs
Japa
n K
C66
0379
K
C66
0538
K
C66
0610
K
C66
0635
K
C66
0481
K
C66
0706
Neo
. pun
icea
M
AFF
241
537=
TPP-
h290
B
PI 8
8210
4 Q
uerc
us c
risp
ula
Japa
n K
C66
0369
K
C66
0536
K
C66
0606
K
C66
0633
K
C66
0479
K
C66
0707

Yuuri HIROOKA et al. / Species delimitation for Neonectria coccinea group including the causal agents of beech bark disease in Asia, …
Mycosystema
493
Ta
ble
1 co
ntin
ued
Neo
. pun
icea
M
AFF
241
549=
TPP-
h329
B
PI 8
8211
6 Tw
igs
Japa
n K
C66
0373
K
C66
0542
K
C66
0599
K
C66
0638
K
C66
0485
K
C66
0711
Neo
. pun
icea
G.
J.S. 9
2-32
=CBS
119
530
BPI
802
647
Acer
mac
roph
yllu
m
Scot
land
K
C66
0378
K
C66
0527
K
C66
0564
K
C66
0631
K
C66
0466
K
C66
0699
Neo
. pun
icea
A
.R. 3
713=
CBS
1195
31
BPI
871
118
Fagu
s syl
vatic
a Sl
ovak
ia
KC
6603
76
KC
6605
07
KC
6605
53
KC
6606
30
KC
6604
45
KC
6606
98
Neo
. pun
icea
A
.R. 3
454=
CBS
1195
25
BPI
871
116
Fagu
s syl
vatic
a Sl
ovak
ia
KC
6603
60
KC
6604
97
KC
6605
92
KC
6606
22
KC
6604
32
KC
6606
96
Neo
. pun
icea
A
.R. 3
711=
CB
S 11
9533
B
PI 8
7111
7 Fa
gus s
ylva
tica
Slov
akia
K
C66
0362
-
KC
6605
66
KC
6606
23
KC
6604
43
KC
6606
97
Neo
. pun
icea
A
.R. 3
704=
CBS
1195
32
- Fa
gus s
ylva
tica
Slov
akia
K
C66
0361
K
C66
0503
K
C66
0563
K
C66
0628
K
C66
0439
K
C66
0728
Neo
. pun
icea
G.
J.S. 9
0-29
=CBS
119
529
BPI
110
7108
Fa
gus s
ylva
tica
Switz
erla
nd
KC
6603
83
KC
6605
26
KC
6605
74
KC
6606
44
KC
6604
65
KC
6606
94
Neo
. pun
icea
G.
J.S. 9
3-35
=CBS
119
528
BPI
802
504
Acer
sp.
Uni
ted
Stat
esK
C66
0384
K
C66
0528
K
C66
0602
K
C66
0640
K
C66
0468
D
Q78
9888
*
Neo
. pun
icea
A
.R. 3
675=
CBS
1191
53
- Fa
gus g
rand
ifolia
Uni
ted
Stat
es-
- -
- -
KC
6607
44
Neo
. pun
icea
A
.R. 4
096=
CBS
1192
31
BPI
870
944
Fagu
s gra
ndifo
liaU
nite
d St
ates
- -
- -
- K
C66
0734
Neo
. pun
icea
J.F
. 110
17=A
.R. 4
831=
CBS
1342
51
- Ac
er sp
. Fr
ance
K
C66
0381
-
KC
6606
12
KC
6606
46
- K
C66
0695
Neo
. pun
icea
C
.L.L
. 100
40=A
.R. 4
828=
CBS
1342
48
- C
asta
nea
sp.
Fran
ce
KC
6603
66
KC
6605
23
KC
6605
56
KC
6606
25
KC
6604
62
KC
6607
03
Neo
. pun
icea
C
.L.L
. 100
45=A
.R. 4
830=
CBS
1342
50
- Sa
lix sp
. Fr
ance
K
C66
0377
K
C66
0525
K
C66
0605
K
C66
0629
K
C66
0464
K
C66
0704
Neo
. pun
icea
J.F
. 110
23=A
.R. 4
832=
CBS
1342
52
- Sa
lix sp
. Fr
ance
K
C66
0365
-
KC
6605
89
KC
6606
27
KC
6604
70
KC
6607
02
Neo
. pun
icea
C
.L.L
. 100
42=A
.R. 4
829=
CBS
1342
49
- Ro
bini
a sp
. Fr
ance
K
C66
0363
K
C66
0524
K
C66
0603
K
C66
0626
K
C66
0463
K
C66
0700
Not
e: A
.R.:
Am
y Y.
Ros
sman
, USD
A-A
RS
MD
USA
; AT
CC
: A
mer
ican
Typ
e Cu
lture
Col
lect
ion,
Man
assa
s, VA
, USA
; B
PI:
U.S
. Nat
iona
l Fu
ngus
Col
lect
ions
USD
A-A
RS
MD
USA
; C
BS:
Cen
traal
bure
au v
oor
Schi
mm
elcu
lture
s, U
trech
t, Th
e N
ethe
rland
s; C
.L.L
.: Ch
ristia
n Le
chat
, Asc
ofra
nce,
Vill
iers
en
Bois
, Fra
nce.
; G.J
.S.:
Gar
y J.
Sam
uels,
USD
A-A
RS
MD
USA
; J.F
.: Ja
cque
s
Four
nier
, Asc
ofra
nce,
Vill
iers
en
Boi
s, Fr
ance
; MA
FF: M
AFF
Gen
eban
k, N
atio
nal I
nstit
ute
of A
grob
iolo
gica
l Sci
ence
s, Ib
arak
i, Ja
pan;
TU
A-T
PP-h
: Yuu
ri H
irook
a, T
orop
ical
Pla
nt P
lote
ctio
n La
b
Her
bariu
m, T
okyo
Uni
vers
ity o
f A
gric
ultu
re, T
okyo
Jap
an; T
PP-h
: Yuu
ri H
irook
a, T
orop
ical
Pla
nt P
lote
ctio
n La
b Cu
lture
Col
lect
ion,
Tok
yo U
nive
rsity
of
Agr
icul
ture
, Tok
yo J
apan
; Y.H
.: Yu
uri
Hiro
oka,
USD
A-A
RS
MD
USA
. * Sequ
ence
s obt
aine
d fro
m G
enB
ank.

ISSN1672-6472 CN11-5180/Q Mycosystema May 15, 2013 Vol.32 No.3
http://journals.im.ac.cn/jwxtcn
494
Table 2 Parameters used and statistical values resulting from the different phylogenetic analyses of individual datasets for members
of the Neonectria coccinea group
Locus Act ITS LSU Rpb1 Tef1 Tub Combined
Number of
characters
Included sites 663 449 788 650 440 535 3525
Phylogenetically
informative sites (%)
41 (6%) 22 (5%) 11 (1%) 72 (11%) 64 (15%) 64 (12%) 274 (8%)
Tree length 68 47 17 97 135 102 514
Consistency Index (CI) 0.912 0.766 0.941 0.825 0.837 0.863 0.768
Retention Index (RI) 0.978 0.893 0.990 0.968 0.960 0.973 0.942
Rescaled consistency
(RC)
0.892 0.684 0.931 0.798 0.803 0.840 0.724
MP
Homoplasy Index (HI) 0.088 0.234 0.059 0.175 0.163 0.137 0.232
ML and BI Nucleotide substitution
models
TrN+I TPM2uf+I+G TIM3 TrNef+I K80+I TIM1+I
Primers used (reference) Tact1, Tact2
(Samuels et al.
2006)
ITS5, ITS4
(White et al.
1990)
LR5, LROR
(Vilgalys
n.d.)
crpb1a, rpb1c
(Castlebury
et al. 2004)
tef1-728,
tef1-1567
(Carbone &
Kohn 1999;
Rehner 2001)
Btub-T1,
Btub-T2
(O’Donnell
& Cigelnik
1997)
PCR
PCR protocol: annealing
temp. & cycles
65℃, 30s, 15×
48℃, 30s, 30×
53℃, 1min,
35×
53℃, 1min,
35×
50℃, 2min,
40×
66℃, 55s, 9×
56℃, 55s,
35×
55℃, 30s,
35×
MP analyses were performed with PAUP* version 4.0 b10 (Swofford 2002), using 1,000 replicates of heuristic search with random addition of sequences and subsequent TBR branch swapping. Gaps (insertion/deletions) were treated as missing data. Bootstrap analysis was performed with 1,000 replicates, and a 70 % majority rule consensus tree was constructed in PAUP*. The three analyses for each locus, act, tub, rpb1, ITS, LSU, and tef1, were also done and produced each tree to identify the most suitable locus in Neo. coccinea group.
2 RESULTS Phylogenetic analyses
Because the BI, MP and ML trees showed
almost identical topologies in each dataset, the Bayesian trees were taken as representatives (Figs. 1−7). The number of taxa, substitution models and other statistical values resulting from the different analyses are presented in Table 2.
The combined BI, MP, and ML analyses of six loci using 75 isolates of the Neonectria coccinea group resolved four distinct species (Fig. 7). The first major clade (clade A) occurring only in Asia is here determined as Neo. microconidia. This species was supported by moderate values (BI PP 0.73, MP BP 77%, ML BP 74%). In the Neo. microconidia clade, MAFF 241514 and MAFF 241572 formed a clade A-1 supported by high BI PP, MP BP, and ML
Yuuri HIROOKA et al. / Species delimitation for Neonectria coccinea group including the causal agents of beech bark disease in Asia, …
Mycosystema
495
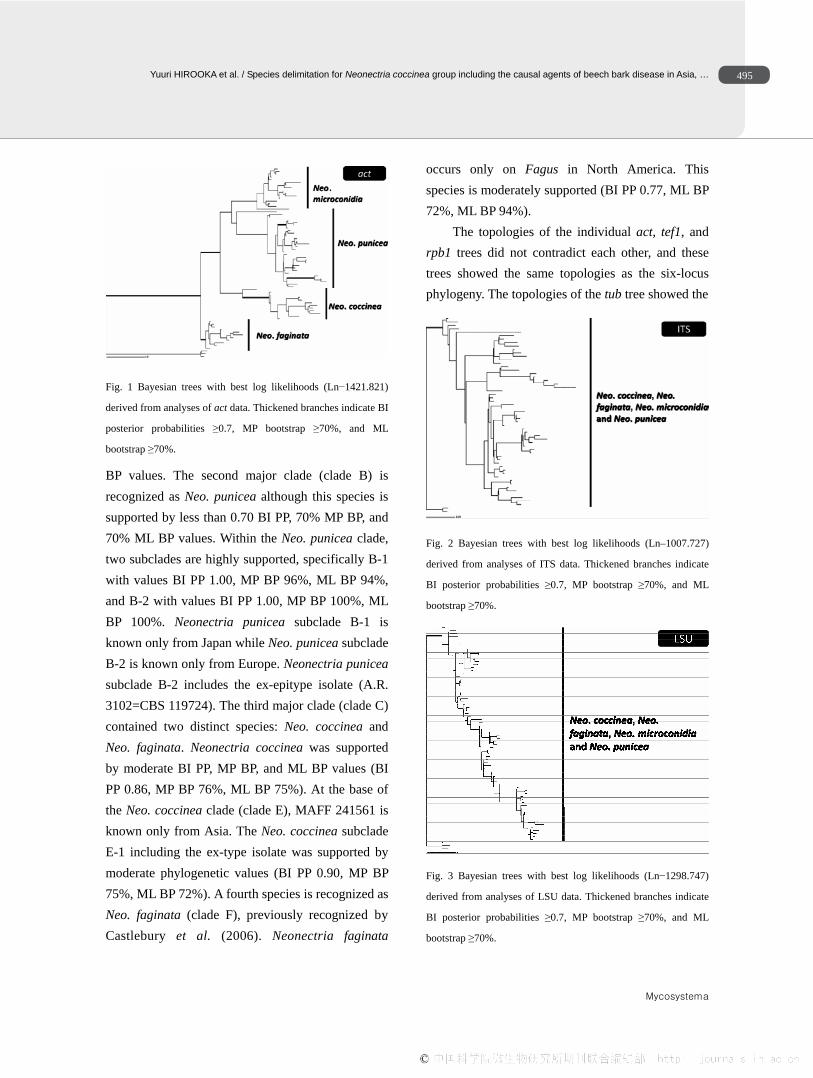
Fig. 1 Bayesian trees with best log likelihoods (Ln−1421.821)
derived from analyses of act data. Thickened branches indicate BI
posterior probabilities ≥0.7, MP bootstrap ≥70%, and ML
bootstrap ≥70%.
BP values. The second major clade (clade B) is recognized as Neo. punicea although this species is supported by less than 0.70 BI PP, 70% MP BP, and 70% ML BP values. Within the Neo. punicea clade, two subclades are highly supported, specifically B-1 with values BI PP 1.00, MP BP 96%, ML BP 94%, and B-2 with values BI PP 1.00, MP BP 100%, ML BP 100%. Neonectria punicea subclade B-1 is known only from Japan while Neo. punicea subclade B-2 is known only from Europe. Neonectria punicea subclade B-2 includes the ex-epitype isolate (A.R. 3102=CBS 119724). The third major clade (clade C) contained two distinct species: Neo. coccinea and Neo. faginata. Neonectria coccinea was supported by moderate BI PP, MP BP, and ML BP values (BI PP 0.86, MP BP 76%, ML BP 75%). At the base of the Neo. coccinea clade (clade E), MAFF 241561 is known only from Asia. The Neo. coccinea subclade E-1 including the ex-type isolate was supported by moderate phylogenetic values (BI PP 0.90, MP BP 75%, ML BP 72%). A fourth species is recognized as Neo. faginata (clade F), previously recognized by Castlebury et al. (2006). Neonectria faginata
occurs only on Fagus in North America. This species is moderately supported (BI PP 0.77, ML BP 72%, ML BP 94%).
The topologies of the individual act, tef1, and rpb1 trees did not contradict each other, and these trees showed the same topologies as the six-locus phylogeny. The topologies of the tub tree showed the
Fig. 2 Bayesian trees with best log likelihoods (Ln–1007.727)
derived from analyses of ITS data. Thickened branches indicate
BI posterior probabilities ≥0.7, MP bootstrap ≥70%, and ML
bootstrap ≥70%.
Fig. 3 Bayesian trees with best log likelihoods (Ln−1298.747)
derived from analyses of LSU data. Thickened branches indicate
BI posterior probabilities ≥0.7, MP bootstrap ≥70%, and ML
bootstrap ≥70%.

ISSN1672-6472 CN11-5180/Q Mycosystema May 15, 2013 Vol.32 No.3
http://journals.im.ac.cn/jwxtcn
496
almost similar topologies with the six-locus phylogeny as well as the other protein-coded dataset phylogeny. The topologies of the ITS and LSU trees conflict with the six-locus phylogeny. Among these single locus trees, tef1 provides the best resolution and thus is the most phylogenetically informative.
Fig. 4 Bayesian trees with best log likelihoods (Ln–1545.111)
derived from analyses of rpb1 data. Thickened branches indicate
BI posterior probabilities ≥0.7, MP bootstrap ≥70%, and ML
bootstrap ≥70%.
Fig. 5 Bayesian trees with best log likelihoods (Ln−1548.664)
derived from analyses of tef1 data. Thickened branches indicate
BI posterior probabilities ≥0.7, MP bootstrap ≥70%, and ML
bootstrap ≥70%.
Fig. 6 Bayesian trees with best log likelihoods (Ln–1458.84)
derived from analyses of tub data. Thickened branches indicate BI
posterior probabilities ≥0.7, MP bootstrap ≥70%, and ML
bootstrap ≥70%.
Morphological, colony growth and temperature analyses
The four species in the Neo. coccinea group are difficult to distinguish based solely on teleomorph morphology. Although perithecial characters, such as color, surface, and wall cell structure, are generally reliable for identifying the species or genera of other nectrioid fungi, this is not true for the segregate species of the Neo. coccinea group. The perithecial apex of Neo. coccinea, Neo. faginata, and Neo. punicea tends to be papillate and thus seems to be useful in distinguishing species but this characteristic varies within species. The perithecial wall surface of species in the Neo. coccinea group is slightly roughened. The perithecial walls are about the same thickness and cell walls form similar tissue of textura globulosa to textura angularis; thus, perithecial wall structure is not useful in distinguishing these species. According to Castlebury et al. (2006), the asci of Neo. coccinea are narrowly clavate with an apical ring while those

Yuuri HIROOKA et al. / Species delimitation for Neonectria coccinea group including the causal agents of beech bark disease in Asia, …
Mycosystema
497
Fig. 7 Bayesian tree with best log likelihood (Ln−8382.847) derived from analysis of combined data set (act, ITS, LSU, rpb1, tef1 and tub).
Values at branches indicate Bayesian posterior probabilities (BI PP)/Maximum Likelihood bootstrap (ML BP)/Maximum parsimony
bootstrap (MP BP). BI posterior probabilities ≤0.7, MP bootstrap ≤70%, and ML bootstrap ≤70% indicated by - . Thickened branches
indicate BI posterior probabilities ≥0.7, MP bootstrap ≥70%, and ML bootstrap ≥70%. The culture collection number, geography and host are
included after each taxon name. “EX” indicates ex-type cultures.

ISSN1672-6472 CN11-5180/Q Mycosystema May 15, 2013 Vol.32 No.3
http://journals.im.ac.cn/jwxtcn
498
of Neo. faginata are cylindrical without an apical ring and with uniseriate ascospores. In our study, we observed that all species of Neo. coccinea group have uniseriate or occasionally obliquely biseriate ascospores in asci with an apical ring. These morphological characters were not useful in distinguishing species. Although size, surface structure, and septation of ascospores are considered to be important in distinguishing these species, we found no significant differences among the four species of the Neo. coccinea group. We analyzed 95% confidence intervals of length/width ratios of ascospores on natural substrate (Fig. 8). Although length of ascospores in Neo. faginata is statistically shorter than Neo. coccinea, no differences of the length were found among Neo. faginata, Neo. microconidia and Neo. punicea. We were not able to examine the anamorphic states of species in the Neo. coccinea group in nature because we did not find many specimens with the anamorph fructification.
As for the anamorph in culture, colony colors are indistinguishable among species. Colonies are white to umber with ochraceous to sienna reverse. Although the optimal temperature for mycelial growth on PDA for each species is 20℃, the mycelial growth rates at 30℃ for 7days on PDA were different: Neo. punicea and Neo. faginata are less than 15mm diameter while Neo. microconidia and Neo. coccinea are greater than 15mm diameter. The morphology of conidiophores, conidiogenous cells, chlamydospores, and microconidia are similar for the four species of the N. coccinea group; however, these species can be distinguished based on the morphology of the macroconidia. On SNA, Neo. microconidia and Neo. faginata produce up to 8-septate macroconidia while Neo. punicea and Neo. coccinea produce up to 7-septate macroconidia. Size of conidia is also reliable; only Neo. faginata forms
macroconidia that are more than 80μm long when 5-septate while the other species produce macroconidia that are less than 80μm long when 5-septate.
Overall, Neo. faginata is easily identified because it is known only on species of Fagus in North America and produces macroconidia more than 80μm long when 5-septate. Neonectria coccinea is distinguishable based on host specificity occurring only on Fagus in Europe (except for one anomalous isolate from Japan) and the fast mycelial grow at 30℃ for 7d on PDA. Neonectria microconidia and Neo. punicea are similar because of the broad host range and the same size of macroconidia; however, the number of septa in the macroconidia and mycelial growth rate are different. In addition, Neonectria microconidia is known only from Asia. TAXONOMY
Based on our morphological and molecular analyses, the Neonectria coccinea group is re-determined as four distinct species, each of which is described and illustrated below. A key to these four species is provided.
Neonectria coccinea (Pers.: Fr.) Rossman & Samuels, in Rossman, Samuels, Rogerson & Lowen, Stud. Mycol. 42: 158. 1999. Fig. 9
≡Sphaeria coccinea Pers.: Fr., Persoon, Icon. & Descr. Fung. 2: 47. 1800: Fries, Syst. Mycol. 2: 412. 1823.
≡Nectria coccinea (Pers.: Fr.) Fr., Summa Veg. Scand. 2: 388. 1849.
=Fusidium candidum Link, Observationes I, Mag. Ges. Naturf. Freunde Berlin 3: 6. 1809.
≡Cylindrocarpon candidum (Link) Wollenw., Fus. Autogr. Del., ed. 2, no. 655. 1926.
=Sphaerostilbe caespitosa Fuckel, Jahrb. Nassauischen Vereubs Naturk. 27-28: 33. 1873.

Yuuri HIROOKA et al. / Species delimitation for Neonectria coccinea group including the causal agents of beech bark disease in Asia, …
Mycosystema
499
Fig. 8 Graphs of 95% confidence intervals of length to width ratios of ascospores and conidia.
≡Neonectria caespitosa (Fuckel) Wollenw., Angew. Bot. 8: 192. 1926.
=Fusidium fractum Sacc. & Cav., N. Giorn. Bot. ital. 7: 308. 1900.
≡Cylindrocarpon fractum (Sacc. & Cav.) Wollenw., Fus. Autogr. Del., ed. 1, no. 655. 1924.
Teleomorph on natural substrata: Mycelium sometimes visible around perithecia and on host. Stromata up to 0.3mm high and 8mm diam., erumpent through epidermis, red to bay, KOH+ dark red, LA+ yellow, pseudoparenchymatous, cells forming textura angularis to textura prismatica with cells oriented more or less vertically; cells 3–10μm diam., with 1–1.5μm thick walls, intergrading with ascomatal wall. Perithecia superficial on well-developed, erumpent stroma, solitary or
caespitose up to 38 on a stroma, subglobose to pyriform, 218–367μm high and 148–399μm diam. (n=21), red to sienna, often cupulate upon drying or collapsing by lateral pinching, papillate, apical region sometimes slightly darker, KOH+ purple, LA+ yellow, surface with smooth to rough. Perithecial surface cells forming textura globulosa or textura angularis, with walls pigmented, ca. 1.5μm thick. Perithecial wall ca. 17–49μm thick, of two distinct regions: outer region ca. 22–42μm thick, intergrading with stroma, cells forming textura globulosa or textura angularis, walls pigmented, ca. 1.5μm thick; inner region ca. 7–21μm thick, of elongated, thin-walled, hyaline cells, forming textura prismatica. Asci unitunicate, (58–)69–97(–124)× (7.8–)8.9–12.9(–15.9)μm (n=44), cylindrical to

ISSN1672-6472 CN11-5180/Q Mycosystema May 15, 2013 Vol.32 No.3
http://journals.im.ac.cn/jwxtcn
500
narrowly clavate, often with an apical ring at apex, stipitate, 8-spored, ascospores occasionally obliquely biseriate near apex. Ascospores ellipsoidal to fusiform with narrowly rounded ends, straight or slightly curved, hyaline, finely spinulose, 1 septate, (9.1–) 11.8–14.6(–19.3)×(3.7–)4.9–6.3(–7.6)μm (n=309).
Anamorph in culture: Optimum temperature for growth on PDA 20℃, colonies 39–56mm (average 45mm) diam. at 20℃ after 7d. Colony surface on PDA, radial, abundant cottony with aerial mycelium, sometimes wavy, white to saffron or umber, saffron to ochraceous sporodochial conidial masses producing after one week; reverse ochraceous to sienna. Sporulation on SNA from lateral phialidic pegs on submerged or commonly on aerial hyphae, 2.0–7.5μm long, 1.5–3.0μm wide at base. Conidiophores occasionally developing on aerial hyphae, unbranched, sometimes verticillate, 1–3-branched, becoming loosely to moderately densely branched, 6.5–124μm long, 2.1–3.0μm wide at base. Conidiogenous cells monophialidic, cylindrical and slightly tapering toward tip or narrowly flask-shaped with widest point in middle, 4–36μm long, 1.5–3.0μm wide at base. Conidia formed by monophialides on submerged or aerial hyphae, formed abundantly on slimy heads, ellipsoidal, oblong to long cylindrical, hyaline, smooth, straight or slightly curved with round at both ends, 0–7-septate; 0-septate: (3.4–)6.0–9.8 (–12.5)×(2.4–)2.7–3.9(–4.6)μm (n=120), 1-septate: 10.0–22.6(–43.5)×(3.6–)3.9–5.1(–5.6)μm (n=52), 2-septate: (22.5–)22.7–29.5(–33.1)×(4.1–)4.4–5.0μm (n=24), 3-septate: (28.1–)32.1–43.5(–52.2)×4.4–6.0 (–7.2)μm (n=48), 4-septate: (41.1–)44.5–62.1 (–62.6)×(5.4–)5.7–6.4(–6.6)μm (n=38), 5-septate: (52.2–)58.9–72.1(–80.4)×(4.2–)5.5–67.1(–8.0)μm (n=114), 6-septate: (70.0–)73.6–84.2(–85.7)×(5.3–) 5.3–6.3(–6.4)μm (n=18), 7-septate: (72.9–)88.4–
100.8(–111.0)×(5.1–)5.4–6.2(–6.5)μm (n=17). Chlamydospores intercalary in hyphae, globose to subglobose, sometimes ellipsoidal, smooth, hyaline. Perithecia not produced in culture.
Distribution: Asia (Japan) and Europe (France, Germany, Romania, Scotland, Slovakia).
Habitat: On Fagus crenata, F. sylvatica and Fagus sp.
Lectotype of Neonectria coccinea designated by Booth (1959). Herb. Lugd. Bat. No. 910, 270-56 on bark of Fagus, isolectotype BPI 738862.
Epitype of Neonectria coccinea designated here. Germany Saarland, Naturpark Saar-Hundsruck, between Leisel and Schollen, alt. 600m. 49°40'0''N 7°10'0''E, on hardwood, Fagus sp., Oct. 13, 1998, coll. G.J. Samuels, H.J. Schroers (BPI 748295; ex-epitype culture CBS 119158=G.J.S. 98-114).
Additional type specimens: A holotype of Sphaeria caespitosa exists at G, isotype K, and BR, according to Booth (1966) and Seifert (1985).
Additional specimens and isolates examined: France, on Fagus sylvatica, 7 Sep. 2001, coll. A. Kunca (BPI 871112; culture A.R. 3696=CBS 119534); on Fagus sylvatica, 7 Sep. 2001, coll. A. Kunca (BPI 870938; culture A.R. 3694=CBS 118914). Japan, Kanagawa Prefecture, Ashigarakami-gun, on Fagus crenata, 17 Apr. 2005, coll. H. Masuya, Y. Hirooka (BPI 881942; culture MAFF 241561=TPP-h430). Romania, Brasov, Busteni, on Fagus sylvatica, 26 Sep. 2000, coll. A. Kunca (BPI 870939; culture A.R. 3708=CBS 118916); Brasov, Busteni, on Fagus sylvatica, 26 Sep. 2000, coll. A. Kunca (BPI 870937; culture A.R. 3707=CBS 118915). Scotland, Cowal Peninsula, Argyll Forest Park. N. End of Loch Goil, alt. 25–250m, on Fagus sp., 12 Apr. 1992, coll. G.J. Samuels, D. Brayford (BPI 802648; culture G.J.S. 92-33=CBS 134254). Slovakia, Kacin, Male Karpety,

Yuuri HIROOKA et al. / Species delimitation for Neonectria coccinea group including the causal agents of beech bark disease in Asia, …
Mycosystema
501
Fig. 9 Neonectria coccinea. A, B: Perithecia in the natural environment; C: Median section of perithecium; D: Median section of perithecial
apex and wall; E: Ascus apices; F: Asci; G: Ascospores in optical section; H: Ascospores in surface view; I: Colony of A.R. 3700 after 7d at
20℃ on PDA; J: Colony of A.R. 3691 after 7d at 20℃ on PDA; K: Colony of A.R. 3707 after 7d at 20℃ on PDA; L: Colony of G.J.S.
92-33after 7d at 20℃ on PDA; M, N, Q–S: Conidiophores on SNA; O, P: Lateral phialidic pegs; T–V: Micro- and macroconidia on SNA.
Bar: A=3mm; B=300µm; C, M=100µm; D=30µm; E–H, N=20µm; I–L=30mm; O–V=10µm.

ISSN1672-6472 CN11-5180/Q Mycosystema May 15, 2013 Vol.32 No.3
http://journals.im.ac.cn/jwxtcn
502
on Fagus sylvatica, 14 Apr. 2000, coll. A. Kunca (BPI 870940; culture A.R. 3700=CBS 119156); Muranky planina, Pod Vel’kow stozkou, on Fagus sylvatica, 26 Oct. 2000, coll. A. Kunca (BPI 870941; culture A.R. 3705=CBS 119150); Bansky Studenec 5, Stiavnicke vrchy, on Fagus sylvatica, 10 Jun. 2001, coll. A. Kunca (BPI 871113; culture A.R. 3691=CBS 119523); Bansky Studenec 1, on Fagus sylvatica, 14 Mar. 2001, coll. A. Kunca (A.R. 3687=CBS 134253), Vel’ke Rovne, Javornicky, on Fagus sylvatica, 11 Jul. 2001, coll. A. Kunca (BPI 871115; culture A.R. 3712=CBS 119522); Nova Bystrica, Kysucke vrchy, on Fagus sylvatica, 20 Nov. 2000, coll. A. Kunca (BPI 871114; A.R. 3701).
Notes: Neonectria coccinea is here determined to include specimens collected on Fagus spp. from Europe and one isolate from Japan. To distinguish this species from others in the Neo. coccinea group, cultural characters are required. Neonectria coccinea is similar to Neo. microconidia in having 5-septate conidia that are 50–70μm long on SNA and a growth rate in culture of more than 15mm at 30℃ for 7days on PDA. In addition, septation of macroconidia in Neo. coccinea ranges from 1–7-septate while in Neo. microconidia the range is 1–8-septate. Also, Neo. coccinea is known only from Fagus while Neo. microconidia occurs on Fagus and other hosts. Surprisingly our phylogenetic analyses suggest a sister-group relationship between Neo. coccinea and MAFF 241561 collected from Japan (Fig. 7). However, based on our observations of the morphological characters in the natural environment and culture, MAFF 241561 completely matches Neo. coccinea. Although there are obvious geographical differences, we tentatively regard MAFF 241561 as Neo. coccinea.
Castlebury et al. (2006) mentioned that BPI 748295 and the derived CBS 119158 as a
representative isolate of N. coccinea but an epitype was not designated. Since this species is economically important, an epitype of the basionym Sphaeria coccinea is needed to distinguish Neo. coccinea from other species in the Neo. coccinea group. The specimen (BPI 748295, ex-epitype culture CBS 119158) is herein designate as epitype of S. coccinea. This specimen was collected on Fagus sp. from Germany, most likely the type locality of Sphaeria coccinea collected by Persoon (1800) because Persoon moved to Göttingen for botanical research and earned a doctorate from the Academy of Natural Sciences in Erlangen in 1777 (de Zeeuw 1939; Petersen 1977).
Neonectria faginata (M.L. Lohman, A.M.J. Watson & Ayers) Castl. & Rossman, in Castlebury, Rossman & Hyten, Can. J. Bot. 84: 1425. 2006. Fig. 10
≡Neonectria coccinea var. faginata M.L. Lohman, A.M.J. Watson & Ayers, Lloydia 6: 100. 1943.
=Cylindrocarpon faginatum C. Booth, Mycol. Pap. 104: 1. 1966.
Teleomorph on natural substrata: Mycelium sometimes visible around perithecia and on host. Stromata up to 0.3mm high and 8mm diam., erumpent through epidermis, red to bay, KOH+ dark red, LA+ yellow, pseudoparenchymatous, cells forming textura angularis to textura prismatica with cells oriented more or less vertically; cells 3–9μm diam., with 1–2μm thick walls, intergrading with ascomatal wall. Perithecia superficial on well-developed, erumpent stroma, solitary or caespitose up to 50 on a stroma, subglobose to pyriform, 195–399μm high and 156–302μm diam. (n=45), red to bay, often cupulate upon drying or collapsing by lateral pinching, papillate, apical region sometimes slightly darker, KOH+ purple,

Yuuri HIROOKA et al. / Species delimitation for Neonectria coccinea group including the causal agents of beech bark disease in Asia, …
Mycosystema
503
LA+ yellow, surface with smooth to rough. Perithecial surface cells forming textura globulosa or textura angularis, with walls pigmented, ca. 1.5μm thick. Perithecial wall ca. 31–50μm thick, of two distinct regions: outer region ca. 17–42μm thick, intergrading with stroma, cells forming textura globulosa or textura angularis, walls pigmented, ca. 1.5μm thick; inner region ca. 13–20μm thick, of elongated, thin-walled, hyaline cells, forming textura prismatica. Asci unitunicate, (65.8–)69.4–96.4 (–124.5)×(7.0–)8.9–11.9(–13.7)μm (n=47), cylindrical to narrowly clavate, often with an apical ring at apex, stipitate, 8-spored, ascospores occasionally obliquely biseriate near apex. Ascospores ellipsoidal to fusiform with narrowly rounded ends, straight or slightly curved, hyaline, finely spinulose, 1-septate, (8.5–)10.3–12.3(–14.3)×(4.1–)4.4–6.2 (–8.0)μm (n=249).
Anamorph in culture: Optimum temperature for growth on PDA 20℃, colonies 34–40mm (average 36mm) diam. at 20℃ after 7d. Colony surface on PDA, radial, abundant cottony with aerial mycelium, sometimes wavy, ochraceous, saffron to ochraceous sporodochial conidial masses producing after one week; reverse ochraceous. Sporulation on SNA from lateral phialidic pegs on submerged or commonly on aerial hyphae, 2.0–6.8μm long, 1.5–2.5μm wide at base. Conidiophores occasionally developing on aerial hyphae, unbranched, sometimes verticillate, 1–3-branched, becoming loosely to moderately densely branched, 6.5–87μm long, 2.1–3.0μm wide at base. Conidiogenous cells, monophialidic, cylindrical and slightly tapering toward tip or narrowly flask-shaped with widest point in middle, 8–29μm long, 1.5–2.5μm wide at base. Conidia formed by monophialides on submerged or aerial hyphae, formed abundantly on slimy heads, ellipsoidal, oblong to long cylindrical, hyaline,
smooth, straight or slightly curved with round at both ends, 0–8-septate; 0-septate: (2.5–)7.5–11.9 (–13.2)×(2.8–)3.3–4.5(–4.8)μm (n=140), 1-septate: (13.2–)13.4–16.6(–19.0)×(3.6–)3.7–4.7(–5.0)μm (n=40), 2-septate: (13.7–)15.5–25.9(–30.3)×(3.1–) 4.0–5.1(–5.8)μm (n=25), 3-septate: (20.4–)35.8– 50.5(–55.6)×(4.5–)4.8–6.7(–7.0)μm (n=21), 4-septate: (44.4–)59.1–67.2(–73.4)×(4.4–)4.8–6.2(–6.8)μm (n=22), 5-septate: (75.9–)76.7–92.3(–97.1)×(5.6–) 5.8–6.8(–7.0)μm (n=50), 6-septate: (78.2–)79.5– 92.1(–104.5)×(5.1–)5.9–7.2(–8.0)μm (n=14), 7-septate: (93.1–)94.1–102.3(–102.7)×5.0–6.6(–7.7)μm (n=35), 8-septate: (95.5–)97.3–109.0(–114.1)×(5.1–)5.9–7.5 (–8.1)μm (n=20). Chlamydospores intercalary in hyphae, globose to subglobose, sometimes ellipsoidal, smooth, hyaline. Perithecia not produced in culture.
Distribution: North America (Canada, United States).
Habitat: On Fagus grandifolia, F. sylvatica and Fagus sp.
Holotype of Neonectria faginata: United States, Maine, Meddybemps, on Fagus grandifolia, 4 Oct. 1933, coll. T.T. Ayers (BPI 551558).
Epitype of Neonectria faginata designated here: United States, Maine, on Fagus grandifolia, Apr. 2006, coll. M. Kassen (BPI 878329; ex-epitype culture A.R. 4307=CBS 134246).
Ex-holotype culture of Cylindrocarpon faginatum: Canada, New Brunswick York County, Rocky Brook, on Fagus grandifolia (CBS 217.67=ATCC 16547=IMI 105738).
Specimens and isolates examined: United States, New Hampshire, Grafton Co., Hubbard Brook Experimental Forest, on Fagus grandifolia, 1998, coll. E.M. Mahoney (culture A.R. 3677=CBS 119154); Maine, Penobscot Co., Penobscot Experimental Forest, on Fagus grandifolia, 1998,

ISSN1672-6472 CN11-5180/Q Mycosystema May 15, 2013 Vol.32 No.3
http://journals.im.ac.cn/jwxtcn
504
Fig. 10 Neonectria faginata. A–C: Perithecia in the natural environment; D: Median section of perithecium; E: Median section of perithecial
apex and wall; F: Asci; G: Ascospores in optical section; H: Ascospores in surface view; I: Colony of A.R. 4167 after 7d at 20℃ on PDA; J:
Colony of A.R. 4307 after 7d at 20℃ on PDA; K–O, Q–U: Conidiophores on SNA; M: Lateral phialidic pegs and conidiophores on SNA; P,
V: Micro- and macroconidia on SNA; W: Chlamydospores on SNA. Bar: A=3mm; B, C=300µm; D=100µm; E, F, K=30µm; G, H,
L–W=10µm; I, J=30mm.

Yuuri HIROOKA et al. / Species delimitation for Neonectria coccinea group including the causal agents of beech bark disease in Asia, …
Mycosystema
505
coll. E.M. Mahoney (culture A.R. 3682=CBS 119155); West Virginia, on Fagus sp., coll. E.M. Mahoney (BPI 880528; culture NCF 346=AR 3680=CBS 118983); West Virginia. Burner Mt. BBD plot tree 4, on Fagus grandifolia, Oct. 2004, coll. M. MacKenzie (BPI 871095; culture A.R. 4130=CBS 119161; West Virginia, Burner Mt. BBD plot tree 8, on Fagus grandifolia, Mar. 03 2005, coll. M. MacKenzie (BPI 870943; culture A.R. 4152=CBS 118917; North Carolina, Blue Ridge Parkway National Park, Richland Balsam, on Fagus sylvatica, Mar. 25 2004, coll. M. MacKenzie (BPI 871123; culture A.R. 4149=CBS 119519); Michigan, Luce Co., N of Bass Lake on County Road 421. 46.5°N 85.78°W, on Fagus grandifolia, Jun. 29 2005, coll. G. Joseph O’Brien (BPI 870945; culture A.R. 4197=CBS 118918); Pennsylvania, McKean Co., Bradford Rd., on Fagus sylvatica, May 12 2005, coll. M. MacKenzie (BPI 871125; culture A.R. 4167=CBS 119199); West Virginia, Burner Mt. BBD plot, tree gamma, on Fagus grandifolia, Mar. 31 2005, coll. M. MacKenzie (BPI 871097; culture A.R. 4153=CBS 119163; West Virginia, Burner Mt. BBD plot, tree beta, on Fagus grandifolia, Mar. 31 2004, coll. M. MacKenzie (BPI 871096; culture A.R. 4151=CBS 119162; North Carolina, Great Smoky Mountains National Park. Newfound Gap, on Fagus sylvatica, Dec. 03 2003, coll. B. Jones (BPI 871122; culture A.R. 4148=CBS 119524); Pennsylvania, Allegheny National Forest, tree 53, on Fagus grandifolia, Oct. 07 2004, coll. M. MacKenzie (BPI 870942; culture A.R. 4097=CBS 118938; West Virginia, Monogahela National Forest. Cheat Mt. Road, on Fagus sylvatica, May 04 2005, coll. M. MacKenzie (BPI 871124; culture A.R. 4166=CBS 119198).
Notes: Neonectria faginata was raised to the rank of species by Castlebury et al. (2006) based on
morphology, geography and phylogeny. Among the four species in the Neo. coccinea group, the morphology of Neo. faginata is relatively distinctive. The five-septate macroconidia of Neo. faginata are longer than the other species i.e., 80–90μm long in Neo. faginata vs. 50–70μm long in other species of Neo. coccinea group. Neonectria faginata is known only on Fagus spp. in North America. According to Castlebury et al. (2006), only Fagus grandifolia was recorded as the host of Neo. faginata. Four specimens on Fagus sylvatica are reported here.
The holotype specimen of Neo. coccinea var. faginata is preserved in BPI, but it lacks a living ex-type culture. Therefore, we epitypify this name with BPI 878329 (ex-epitype culture A.R. 4307=CBS 134246) collected from the same host and state in the United States as the holotype.
Neonectria microconidia J. Luo, P. Zhao & W.Y. Zhuang, in Zhao, Luo, Zhuang, Liu & Wu, Sci. China Life Sci. 54: 671. 2011. Fig. 11
Teleomorph on natural substrata: Mycelium sometimes visible around perithecia and on host. Stromata up to 0.4mm high and 12mm diam., erumpent through epidermis, red to bay, KOH+ dark red, LA+ yellow, pseudoparenchymatous, cells forming textura angularis to textura prismatica with cells oriented more or less vertically; cells 3–12μm diam., with 1–2μm thick walls, intergrading with ascomatal wall. Perithecia superficial on well-developed, erumpent stroma, solitary or caespitose up to 300 on a stroma, subglobose to pyriform, 203–380μm high and 175–357μm diam. (n=30), red to sienna, often cupulate upon drying or collapsing by lateral pinching, sometimes papillate, apical region sometimes slightly darker, KOH+ purple, LA+ yellow, surface with smooth to rough. Perithecial surface cells forming textura globulosa

ISSN1672-6472 CN11-5180/Q Mycosystema May 15, 2013 Vol.32 No.3
http://journals.im.ac.cn/jwxtcn
506
or textura angularis, with walls pigmented, ca. 1.5μm thick. Perithecial wall ca. 30–44μm thick, of two distinct regions: outer region ca. 20–40μm thick, intergrading with stroma, cells forming textura globulosa or textura angularis, walls pigmented, ca. 1.5μm thick; inner region ca. 10–18μm thick, of elongated, thin-walled, hyaline cells, forming textura prismatica. Asci unitunicate, (58–)64–94 (–102)×(6.3–)8.0–10.6(–11.1)μm (n=35), cylindrical to narrowly clavate, often with an apical ring at apex, stipitate, 8-spored, ascospores occasionally obliquely biseriate near apex. Ascospores ellipsoidal to fusiform with narrowly rounded ends, straight or slightly curved, hyaline, finely spinulose, 1-septate, (8.9–)10.6–15.8(–23.1)×(3.8–)4.5–6.5(–9.4)μm (n=192).
Anamorph in culture: Optimum temperature for growth on PDA 20℃, colonies 34–56mm (average 47mm) diam at 20℃ after 7d. Colony surface on PDA, radial, abundant cottony with aerial mycelium, sometimes wavy, white to ochraceous, saffron to ochraceous sporodochial conidial masses producing after one week; reverse ochraceous to rust. Sporulation on SNA from lateral phialidic pegs on submerged or aerial hyphae common, 2.0–7.2μm long, 1.5–2.5μm wide at base. Conidiophores occasionally developing on aerial hyphae, unbranched, sometimes verticillate, 1–3-branched, becoming loosely to moderately densely branched, 6.5–110μm long, 2.1–3.0μm wide at base. Conidiogenous cells, monophialidic, cylindrical and slightly tapering toward tip or narrowly flask-shaped with widest point in middle, 6–38μm long, 1.5–2.5μm wide at base. Conidia formed by monophialides on submerged or aerial hyphae, formed abundantly on slimy heads, ellipsoidal, oblong to long cylindrical, hyaline, smooth, straight or slightly curved with round at both ends,
0–8-septate; 0-septate: (3.1–)5.5–8.9(–12.3)×(1.7–) 2.1–3.3(–4.9)μm (n=288), 1-septate: (9.8–)12.2–19.2 (–31.0)×(2.5–)3.5–4.5(–5.6)μm (n=144), 2-septate: (12.2–)16.7–28.5(–34.8)×(3.4–)4.2–5.4(–5.6)μm (n=37), 3-septate: (24.6–)31.5–43.9(–49.7)×(4.0–) 4.8–6.2(–7.1)μm (n=84), 4-septate: (39.9–)44.5– 53.3(–57.7)×(4.4–)4.8–6.2(–6.8)μm (n=42), 5-septate: (47.4–)55.7–69.7(–76.4)×(4.0–)5.5–6.9(–7.6)μm (n=95), 6-septate: (63.9–)69.3–88.7(–101.9)×(5.0–) 5.9–7.5(–8.2)μm (n=54), 7-septate: (71.5–)80.3– 98.3(–118.2)×(5.1–)6.2–7.6(–8.5)μm (n=51), 8-septate: (79.1–)85.2–101.7(–120.0)×(5.2–)6.6–7.2(–8.0)μm (n=17). Chlamydospores intercalary in hyphae, globose to subglobose, sometimes ellipsoidal, smooth, hyaline. Perithecia not produced in culture.
Distribution: Asia (China, Japan). Habitat: On dead woody substrata, known from
Cerasus jamasakura, Fagus crenata, Vitis coignetiae and Weigela coraeensis.
Holotype of Neonectria microconidia: China, Hubei, Wufeng, 800m alt., on twigs, 13 Sep. 2004, coll. W.Y. Zhuang, Y. Nong (5639) (HMAS 98294).
Specimens and isolates examined: Japan, Fukuoka Prefecture, Oazanatuki, Tagawa city, on bark of dead wood, Apr. 02 2004, coll. Y. Hirooka (BPI 882098; culture MAFF 241530=TPP-h260); Kanagawa Prefecture, Yamakawa-cho, Ashigarakami-gun, on bark of dead wood, Oct. 31 2004, coll. Y. Hirooka (BPI 881931; culture MAFF 241556=TPP-h385); Kanagawa Prefecture, Kiyokawa-mura, Aigo-gun, on twigs, Apr. 16 2005, coll. Y. Hirooka (BPI 881941; culture MAFF 241560=TPP-h427); Kanagawa Prefecture, Yamakawa-cho, Ashigarakami-gun, on dead wood of Cerasus jamasakura, Oct. 31 2004, coll. Y. Hirooka (BPI 881933, culture MAFF 241558=TPP-h390), Kanagawa Prefecuutre, Atugi city, on stump, May 03 2003, coll. Y. Hirooka (culture MAFF

Yuuri HIROOKA et al. / Species delimitation for Neonectria coccinea group including the causal agents of beech bark disease in Asia, …
Mycosystema
507
241506=TPP-h120); Kanagawa Prefecture, Yamakitagawayose, Ashigarakami-gun, on Fagus crenata, Jul. 03 2005, coll. Y. Hirooka (BPI 881956; culture MAFF 241570=TPP-h521); Kanagawa Prefecture, Yamakawa-cho, Ashigarakami-gun, Genbu river, on bark of dead wood, Oct. 31 2004, coll. Y. Hirooka (BPI 881934, culture MAFF 241559=TPP-h391); Kanagawa Prefecture. Yamakawa-cho, Ashigarakami-gun, on bark of dead wood, Oct. 31 2004, coll. Y. Hirooka (BPI 881930; culture MAFF 241555=TPP-h378); Kochi Prefecture, Tosakitakaido, Tosa-cho, on twigs, Aug. 04 2003, coll. Y. Hirooka (BPI 882163; culture MAFF 241516=TPP-h176); Kochi Prefecture, Shirono, Kitagawa-mura, on twigs, Aug. 05 2003, coll. Y. Hirooka (BPI 882165; culture MAFF 241518=TPP-h190); Nagano Prefecture, Sugadaira, Ueda city, on twigs of Vitis coignetiae, Sep. 01 2006, coll. Y. Hirooka (BPI 881958; culture MAFF 241572=TPP-h542); Saitama Prefecture, Nakaizu, Tagata-gun, on twigs of Weigela coraeensis, Oct. 14 2002, coll. Y. Hirooka (BPI 882137; culture MAFF 241493=TPP-h64); Tokyo, Okutama-gun, on twigs, Nov. 20 2003, coll. Y. Hirooka (culture MAFF 241522=TPP-h227); Yamagata Prefecture, Akakura, mogami-cho, on bark of dead wood, Jul. 26 2003, coll. T. Tokiwa (BPI 882161; culture MAFF 241514=TPP-h171); Yamanashi Prefecture, Yamato-son, Higashiyamanashi-gun, Yamato-son, on stem, Sep 26, 2009, coll. T. Tokiwa, Y. Hirooka (BPI 882119; culture MAFF 241552=TPP-h341).
Notes: Neonectria microconidia was recently described from China (Zhao et al. 2011). Based on our phylogenetic analyses, the type isolate of this species clustered with many Japanese isolates. This clade is supported by more than 0.70 MI PP, 70% MPBP and 70% ML BP values (clade A in Fig. 7). Within the Neo. microconidia clade, subclade A-1 is
supported by high BIPP, MPBP and MLBP values. Although subclade A-1 includes only Japanese isolates MAFF 241514 and MAFF 241572, no morphological differences were found from the other isolates of Neo. microconidia. Thus subclade A-1 is not considered to be distinct from Neo. microconidia.
Neonectria microconidia is known only from Asia although Neo. punicea and one isolate of N. coccinea are also known from Asia. Neonectria microconidia can be distinguished from the other species of the Neo. coccina group using the morphological characters of the anamorph. Neonectria microconidia has 5-septate macroconidia that are 50–70μm long, conidia varying from 1–8-septate, a colony growth rate of more than 15mm in diameter after 7d at 30℃ and a wide host range.
Neonectria punicea (J.C. Schmidt: Fr.) Castl. & Rossman, in Castlebury, Rossman & Hyten, Can. J. Bot. 84: 1425. 2006. Fig. 12
≡Sphaeria punicea J.C. Schmidt, in Kunze & Schmidt, Mykologische Hefte (Leipzig) 1: 61. 1817.
=Fusarium album Sacc., Michelia 2: 132. 1880. ≡Cylindrocarpon album (Sacc.) Wollenw.,
Fusaria autographica delineate 1: no. 473. 1916. =Neonectria confusa J. Luo & W.Y. Zhuang,
Mycologia 102: 143. 2010. Teleomorph on natural substrata: Mycelium
sometimes visible around perithecia and on host. Stromata up to 0.4mm high and 10mm diam., erumpent through epidermis, scarlet to bay, KOH+ dark red, LA+ yellow, pseudoparenchymatous, cells forming textura angularis to textura prismatica with cells oriented more or less vertically; cells 4–14μm diam., with 1–1.5μm thick walls, intergrading with ascomatal wall. Perithecia superficial on well-

ISSN1672-6472 CN11-5180/Q Mycosystema May 15, 2013 Vol.32 No.3
http://journals.im.ac.cn/jwxtcn
508
Fig. 11 Neonectria microconidia. A–C: Perithecia in the natural environment; D: Median section of perithecium; E: Median section of
perithecial wall; F, G: Asci; H: Ascospores in optical section; I: Ascospores in surface view; J: Colony of MAFF 241516 after 7d at 20℃ on
PDA; K: Colony of MAFF 241493 after 7d at 20℃ on PDA; L: Colony of MAFF 241530 after 7d at 20℃ on PDA; M: Colony of MAFF
241559 after 7d at 20℃ on PDA; N, P–W: Conidiophores on SNA; O: Lateral phialidic pegs on SNA; X: Micro- and macroconidia on SNA.
Bar: A=3mm; B, C=300µm; D=100µm; E, U, V=30µm; F, G=20µm; H, I=5µm; J–M=30mm; N–T, W, X=10µm.

Yuuri HIROOKA et al. / Species delimitation for Neonectria coccinea group including the causal agents of beech bark disease in Asia, …
Mycosystema
509
developed, erumpent stroma, solitary or caespitose up to 40 on a stroma, subglobose to pyriform, 211–364μm high and 160–338μm diam. (n=50), red to sienna, often cupulate upon drying, papillate, apical region slightly darker, KOH+ purple, LA+ yellow, surface with smooth to rough. Perithecial surface cells forming textura globulosa or textura angularis, with walls pigmented, ca. 1.5μm thick. Perithecial wall ca. 17–46μm thick, of two distinct regions: outer region ca. 16–34μm thick, intergrading with stroma, cells forming textura globulosa or textura angularis, walls pigmented, ca. 1.5μm thick; inner region ca. 6–15μm thick, of elongated, thin-walled, hyaline cells, forming textura prismatica. Asci unitunicate, (48–)69–93(–109)× (6.1–)8.1–11.3(–14)μm (n=100), cylindrical to narrowly clavate, often with an apical ring at apex, stipitate, 8-spored, ascospores occasionally obliquely biseriate near apex. Ascospores ellipsoidal to fusiform, sometimes long fusiform, with narrowly rounded ends, straight or slightly curved, hyaline, finely spinulose, 1-septate, (8.3–)11.0–14.4(–18.5)× (2.9–)4.2–5.8(–7.6)μm (n=675).
Anamorph in culture: Optimum temperature for growth on PDA 20℃, colonies 31–54mm (average 41mm) diam at 20℃ after 7d. Colony surface on PDA, radial, abundant cottony with aerial mycelium, sometimes wavy, white to saffron or ochraceous, saffron to ochraceous sporodochial conidial masses producing after one week; reverse ochraceous to sienna. Sporulation on SNA from lateral phialidic pegs on submerged or aerial hyphae common, 2.0–7.5μm long, 1.5–3.0μm wide at base. Conidiophores occasionally developing on aerial hyphae, unbranched, sometimes verticillate, 1–3-branched, becoming loosely to moderately densely branched, 6.5–98μm long, 2.1–3.5μm wide at base. Conidiogenous cells, monophialidic,
cylindrical and slightly tapering toward tip or narrowly flask-shaped with widest point in middle, 4–24μm long, 2.5–3.5μm wide at base. Conidia formed by monophialides on submerged or aerial hyphae, formed abundantly on slimy heads, ellipsoidal, oblong to long cylindrical, hyaline, smooth, straight or slightly curved with round at both ends, 0–7-septate: 0-septate: (3.2–)6.1–9.7 (–18.3)×(1.5–)2.3–3.7(–6.1)μm (n=505), 1-septate: (8.6–)11.8–20.6(–27.7)×(2.3–)3.6–5.0(–6.6)μm (n=241), 2-septate: (20.3–)22.7–28.7(–31.2)×(4.0–) 4.3–6.1(–7.0)μm (n=50), 3-septate: (25–)31.4–42.2 (–52.5)×(3.5–)4.5–6.1(–8.0)μm (n=233), 4-septate: (37.7–)43.0–54.6(–64.9)×(4.3–)4.9–6.1(–7.2)μm (n=94), 5-septate: (40.9–)51.6–67.9(–79.9)×(4.0–) 5.2–6.6(–8.0)μm (n=199), 6-septate: (66.3–)68.6– 79.2(–84.7)× (4.7–)5.2–7.0(–7.8)μm (n=25), 7-septate: (80.7–)81.9–102.1(–103.8)×(5.0–)5.2–6.8(–6.9)μm (n=21). Chlamydospores intercalary in hyphae, globose to subglobose, sometimes ellipsoidal, smooth, hyaline. Perithecia not produced in culture.
Distribution: Asia (China, Japan); Europe (Austria, France, Germany, Scotland, Slovakia, Switzerland); North America (United States).
Habitat: On dead woody substrata including Acer macrophyllum, Acer sp., Frangula alnus, Fagus grandifolia, F. sylvatica, Prunus×yedoensis, Quercus crispula, Rhamnus fallax, Rhamnus sp., and Ulmus sp.
Neotype of Sphaeria punicea designated here: Austria, Kaernten, St. Margareten i. Ros., "Tumpfi". Grid square 9452/4, on Frangula alnus, 26 Sep. 1999, W. Jaklitsch (BPI 871063; ex-neotype culture A.R. 3102=CBS 119724).
Ex-type of Neonectria confuse: China, Hubei, Wufeng, 1,200m, on twigs of a dicotyledonous tree, 15 Sep. 2004, coll. Y. Nong, W.Y. Zhuang (Y. Nong, W.Y. Zhuang 5694; HMAS 99197).

ISSN1672-6472 CN11-5180/Q Mycosystema May 15, 2013 Vol.32 No.3
http://journals.im.ac.cn/jwxtcn
510
Specimens and isolates examined: Austria, Kaernten, Bad Vellach, Vellacher Kotschna. MTB 8653/1, on Rhamnus fallax, 17 Aug. 2004, coll. W. Jaklitsch (BPI 871062; culture A.R. 4155=CBS 119527). France, Pyrenees Atlantiques, Isle de Sauveterre de Bearn, alt. 100m, on hardwood tree, 25 Oct. 1998, coll. G.J. Samuels, F. Candoussau (BPI 748311; culture G.J.S. 98-133=CBS 134255); Foret du St. Sauvant, on dead twigs of Frangula alnus, 18 Apr. 2008, coll. A. Rossman (BPI 878875; culture A.R. 4522=CBS 134247); Village de Combret, 48 St. Germain du Teil, on Castanea, 19 Feb. 2011, coll. C. Lechat C.L.L. 10040 (A.R. 4828=CBS 134248); St. Maixent, on Robinia, 14 Feb. 2011, coll. C. Lechat C.L.L. 10042 (A.R. 4829=CBS 134249); Lauastric, on Salix, 12 Feb. 2011, coll. C. Lechat C.L.L. 10045 (A.R. 4830=CBS 134250); Las Muros, 09 Rimont, on Acer, 14 Mar 2011, coll. J. Fournier J.F. 11017 (A.R. 4831=CBS 134251); La Maille, 09 Rimont, on Salix, 25 May 2011, on Salix, 25 Mar 2011, coll. J. Fournier J.F. 11023 (A.R. 4832=CBS 134252). Germany, on bark, Mar. 1924, coll. H.W. Wollenweber (BPI 871038; culture CBS 125.24=IMI 113880=MUCL 9808); on Ulmus sp., Jul. 1930, coll. H.W. Wollenweber (BPI 871035=Centraalbureau voor Schimmelcultures Herbarium H-11427; culture CBS 208.30); former West-Germany, on bark of Rhamnus sp., Jul. 1929, coll. H.W. Wollenweber (culture CBS 242.29). Japan, Gifu Prefecture, Mizunashi city, Kashimo-mura, on twigs, 31 Aug. 2004, coll. Y. Hirooka (BPI 882115; culture MAFF 241548= TPP-h328); Niigata Prefecture Nakauonuma-gun, Tuna-cho, Akiyamago, on twigs of Prunus× yedoensis, 30 Aug. 2004, coll. Y. Hirooka (BPI 882113; culture MAFF 241546=TPP-h326); Gifu Prefecture, Mizunashi city, Kashimo-mura, on twigs,
31 Aug. 2004, coll. Y. Hirooka (BPI 882117; culture MAFF 241550=TPP-h330); Tokyo, Okutama-gun, on bark of dead wood, 12 Jul. 2003, coll. T. Tokiwa (BPI 882160; culture MAFF 241513=TPP-h166); Miyagi Prefecture, Taihaku-ku, akiu-cho, Akiuootaki, on twigs, 4 Aug. 2004, coll. Y. Hirooka (culture MAFF 241540=TPP-h296); Niigata Prefecture, Nakauonuma-gun, Tuna-cho, on bark, 31 Aug. 2004, coll. Y. Hirooka (BPI 882114; culture MAFF 241547=TPP-h327); Miyagi Prefecture, Taihaku-ku, akiu-cho, Akiuootaki, on twigs, 4 Aug. 2004, coll. Y. Hirooka (BPI 882107; culture MAFF 241541=TPP-h297); Miyagi Prefecture, Taihaku-ku, akiu-cho, Akiuootaki, on Quercus crispula, 4 Aug. 2004, coll. Y. Hirooka (BPI 882104; culture MAFF 241537=TPP-h290); Gifu Prefecture, Mizunashi city, Kashimo-mura, on twigs, 31 Aug. 2004, coll. Y. Hirooka (BPI 882116; culture MAFF 241549=TPP-h329). Scotland, Cowal Peninsula, Argyll Forest Park. N. End of Holy Loch, alt. 25–250m, on dead bark of Acer macrophyllum, 13 Apr. 1992, coll. G.J. Samuels, D. Brayford (BPI 802647; culture G.J.S. 92-32=CBS 119530). Slovakia, Volvec, Volovske vrchy, on Fagus sylvatica, 29 Oct. 1999, coll. A. Kunca (BPI 871118; culture A.R. 3713=CBS 119531); Stagiar, on Fagus sylvatica, 1999, coll. I. Mihal, A. Kunca (BPI 871116; A.R. 3454=CBS 119525); Stagiar, Kremnicke vrchy, on Fagus sylvatica, 16 Jul. 1999, coll. A. Kunca (BPI 871117; A.R. 3711=CBS 119533); Poruba-N6, on Fagus sylvatica, coll. A. Kunca (culture A.R. 3704=CBS 119532). Switzerland, Grisons (Graubunden), Canton, Malan Vicinity, alt. ca. 600m, on Fagus sylvatica, 5 Sep. 1990, coll. K.F. Rodrigues (BPI 1107108; culture G.J.S. 90-29=CBS 119529). United States, New York, Ulster Co., Catskill Mts. from Oliverea at end

Yuuri HIROOKA et al. / Species delimitation for Neonectria coccinea group including the causal agents of beech bark disease in Asia, …
Mycosystema
511
Fig. 12 Neonectria punicea. A–C: Perithecia in the natural environment; D, E: Median section of perithecia; F: Asci; G: Ascospores in
surface view; H: Colony of MAFF 241513 after 7d at 20℃ on PDA; I: Colony of MAFF 241549 after 7d at 20℃ on PDA; J: Colony of
CBS 125.24 after 7d at 20℃ on PDA; K, M–U: Conidiophores on SNA; L: Lateral phialidic pegs on SNA; V, W: Micro- and macroconidia
on SNA; X: Chlamydospores on SNA. Bar: A=3mm; B–D=300µm; E, K, R=100µm; F=20µm; G, L–N=5µm; H–J=30mm; O–Q,
S–X=10µm.

ISSN1672-6472 CN11-5180/Q Mycosystema May 15, 2013 Vol.32 No.3
http://journals.im.ac.cn/jwxtcn
512
of McKinley Hollow Road to Summit of Balsam Mt., alt. 550–1,098m. 42°4'0''N 74°22'0''W, on Acer sp., 7 May 1993, coll. W.R. Buck (BPI 802504; culture G.J.S. 93-35=CBS 119528); New York, Schuyler Co., Arnot Forest, on Fagus grandifolia,1988, coll. E.M. Mahoney (culture A.R. 3675=CBS 119153); Pennsylvania, Allegheny National Forest. Tree 6, on Fagus grandifolia, 7 Oct. 2004, coll. M. MacKenzie (BPI 870944; culture A.R. 4096=CBS 119231).
Notes: The basionym of Neonectria punicea, Sphaeria punicea, was described from Germany in 1817 (Kunze & Schmidt 1817), and the type specimens should be preserved in B. Despite intensively searching in several herbaria including B around Europe, this type specimen could not be located. Because no illustrations exist in the original paper (Kunze & Schmidt 1817), the name is herein
neotypified. The one isolate from Germany on Rhamnus sp., the original host of Sphaeria punicea, did not have a teleomorph specimen. A specimen with culture from Austria on Frangula alnus (previously known as Rhamnus frangula; Rhamnaceae) is herein designated the neotype of Sphaeria punicea (BPI 871063; ex-neotype culture A.R. 3102=CBS 119724).
Morphologically, Neo. punicea could be difficult to identify because it has a wide host range and is known in Asia, Europe, and North America. Based on our phylogenetic data, at least two cryptic species are embedded within Neo. punicea. These appear as subclades B-1 and B-2 each supported by high MI PP, MPBP and ML BP values. Although these specimens and isolates of these subclades were examined carefully, we could not find useful characters to distinguish them.
Key to species in the Neonectria coccinea GROUP
1. Mycelial growth >15mm at 30℃ for 7d on PDA············································································ 2
1. Mycelial growth <15mm at 30℃ for 7d on PDA············································································ 3
2. Macroconidia 1−7-septate; on Fagus; in Europe and Asia (one isolate from Japan) ····Neo. coccinea
2. Macroconidia 1−8-septate; known only from Asia (China and Japan)··················Neo. microconidia
3. Macroconidia 1−7-septate, 5-septate macroconidia 50–70µm long on SNA; known from diverse
woody hosts in Asia (China and Japan), Europe, and North America ··························· Neo. punicea
3. Macroconidia 1−8-septate, 5-septate macroconidia 80–90µm long on SNA; known only on Fagus
in North America·········································································································Neo. faginata
3 DISCUSSION Species delimitation
By combining a morphological species concept (John & Maggs 1997; Kirk et al. 2008) with genealogical concordance phylogenetic species recognition (Taylor et al. 2000), we are able to define four species in the Neonectria coccinea group. Similarly this approach has been used to divide the Nectria cinnabarina complex into four
species (Hirooka et al. 2011). In the N. cinnabarina species complex anamorphic states in nature showed the most useful characters for defining species. In the Neo. coccinea group these characters were rarely found.
Our phenotypic and genotypic analyses of the Neo. coccinea group delimited four previously known species, each of which is described and illustrated. Based on our analyses as well as the

Yuuri HIROOKA et al. / Species delimitation for Neonectria coccinea group including the causal agents of beech bark disease in Asia, …
Mycosystema
513
phylogenetic tree in Zhao et al. (2011), Neo. coccinea, Neo. faginata, Neo. microconidia and Neo. punicea form a monophyletic group within the genus Neonectria sensu Chaverri (Chaverri et al. 2011). Of the six loci used, two nuclear ribosomal DNA regions, ITS and LSU datasets, did not fully distinguish these species, but sequence data from the four protein-coding genes, act, tef1, tub, and rpb1, strongly supported them. In the tub dataset one isolate of Neo. punicea, A.R. 4155=CBS 119527, was not nested within the main Neo. punicea clade due to transitions of a few base pairs. However, this isolate falls in the Neo. punicea clade in the phylogeny produced by the other three protein-coding genes as well as in the combined dataset. This phylogenetic separation resulting from a few base pair positions also occurred in the Grosmannia serpens complex (Duong et al. 2012). Among the protein-coding genes used in this study, the tef1 dataset provided the most variable regions and thus was the most informative. Sequence data of the ITS, the official barcoding gene for fungi (Seifert 2009; Schoch et al. 2012), did not reliably resolve relationships within the Neo. coccinea group.
In our study, Neo. faginata and Neo. coccinea were segregated based on phylogenetic and morphological data as mentioned in Castlebury et al. (2006). Within Neo. punicea many distinct clades are included, although only two of them are phylogenetically supported by more than 0.70 MI PP, 70% MP BP and 70% ML BP values. The ex-neotype culture of the Neo. puniciea designated herein as A.R. 3102 (CBS 119724) is included in Neo. punicea subclade B-2 together with CBS 242.29 and A.R. 4522 (=CBS 134247) supported by high phylogenetic values. These three isolates could be regarded as Neo. punicea sensu stricto; the other isolates of Neo. punicea could be taxonomically
segregated from the sensu stricto isolates. Likewise, subclade B-1 of Neo. punicea consisting of isolates only from Japan is supported by high MI PP, MP BP and ML BP values. However, because no morphological differences could be observed, we do not recognize these two clades as distinct species. Similar subclades exist in Neo. coccinea and Neo. microconidia, but again no morphological characters exist on which to base species delimitations. Geography
The Neonectria coccinea group is known from throughout the world especially in the Northern Hemisphere (Castlebury et al. 2006). Within each species, geography is more or less useful. Neonectria microconidia is known only in Asia, Neo. faginata only in North America, and Nectria coccinea without MAFF 241561 only from Europe. Neonectria punicea is widespread in Asia, Europe and North America. These four species have been reported in tropical regions and the Southern Hemisphere (Doidge 1950); however, none of these reports could be confirmed. Host
Known only on Fagus, Neonectria coccinea and Neo. faginata are recognized as the causal agents of beech bark disease in Europe and North America, respectively. On the other hand Neo. punicea and Neo. microconidia are collected from a wide range of woody shrubs and trees including Fagus. Although we have not examined the pathogenicity of Neo. microconidia and Neo. punicea, they could be plant pathogenic. Most specimens used in this study were collected on newly killed branches or trunks. These fungi may exist as endophytes and then sporulate when the substrate becomes weakened (Chapela & Boddy 1988, Sieber 2007).
Knowledge of the geographic distribution and

ISSN1672-6472 CN11-5180/Q Mycosystema May 15, 2013 Vol.32 No.3
http://journals.im.ac.cn/jwxtcn
514
host association of species in the Neo. coccinea group has been limited primarily to North America and Europe. In our study, many samples of Neo. coccinea group from Asia, especially Japan, were included; most of these are not species of Neo. coccinea and Neo. faginata, cause of BBD. Surprisingly, MAFF 241561 from Fagus crenata in Kanagawa, Japan, appears to be basal to the rest of the isolates of Neo. coccinea on Fagus in Europe (subclade E-1). Subclade E-1 including the ex-type culture (CBS 119158) was supported by moderate to high MI PP, MP BP and ML BP values (BI PP 0.90, MP BP 75%, ML BP 72%) values. We propose two hypotheses about the origin of MAFF 241561. First MAFF 241561 may have been introduced from Europe to Japan and thus derived from European Neo. coccinea by a mutation. This is suggested by the high number of shared single nucleotide polymorphisms, the sister relationship between MAFF 241561 and the E-1 subclade indicates a recent speciation event. The host of MAFF 241561 is Japanese beech, Fagus crenata, a species that is sister to the European beech, Fagus sylvatica (Denk 2003). Another hypothesis is that MAFF 241561 may have originated in Japan and simply was not detected until now. To know the origin of the Japanese isolate, more collections are needed not only from Japan, but also from other Asian countries. The potential threat of BBD in Japan
Non-indigenous fungal pathogens can be highly disastrous to hosts that are closely related to the original host species because of host-pathogen co-adaptation (Parker & Gilbert 2004). The current forest inventory data in the United State suggest that BBD has invaded areas with relatively high densities of beech. BBD have degraded American beech trees (Fagus grandifolia) in the eastern deciduous forests of North America and are now spreading in
Canadian forests (Houston 1994; O’Brien et al. 2001; MacKenzie & Iskra 2005).
In our study, MAFF 241561 is identified for the first time as Neo. coccinea in Japan based on phenotypic and genotypic data. This fungus may already have spread around Japan and will cause serious damage to Japanese beech trees sooner or later as has occurred in North America. So far, the disease caused by this fungus has not been detected in Japan, possibly because the beech bark scale (Cryptococcus fagisuga) has not colonized Japan (Gwiazdowski et al. 2006). The scale insect makes feeding wounds in the bark that enables infection by Neonectria spp. In order to determine and prevent the possible threat of BBD, Japanese beech trees should be tested for pathogenicity using MAFF241561. A careful watch for the beech bark scale insect as well as other isolates of Neo. coccinea in Japan should be continued.
Acknowledgements: The authors thank Dr. Gary J.
Samuels (USDA-ARS, Beltsville, Maryland, USA) for
using the many specimens and isolates that he collected as
well as providing fruitful comments during the course of
this study. We also thank the many mycologists who
shared their fresh specimens and/or cultures with us,
specifically Matt Kassen, Walter Jaklitsch, Christian
Lechat, Martin MacKenzie, Hayato Masuya, and
Toshiyuki Tokiwa. We express sincere thanks to Sato
Toyozo, Takayuki Aoki, and Keisuke Tomioka (NIAS
Genebank, National Institute of Agrobiological Sciences,
Tukuba, Ibaraki, Japan), and Keiko T. Natuaki (Tokyo
University of Agriculture) for quickly depositing and
sending Japanese cultures. We express sincere thanks to
the curators and staff of the BPI and G. This study was
supported in part by the United States National Science
Foundation (NSF) PEET grant DEB-0731510
‘Monographic Studies in the Nectriaceae, Hypocreales:

Yuuri HIROOKA et al. / Species delimitation for Neonectria coccinea group including the causal agents of beech bark disease in Asia, …
Mycosystema
515
Nectria, Cosmospora, and Neonectria’ to University of
Maryland (PIs: P. Chaverri, A.Y. Rossman, G.J. Samuels).
[REFERENCES] Bazinet AL, Cummings MP, 2008. The Lattice project: a Grid
research and production environment combining multiple
Grid computing models. In: Weber WHW (ed.) Distributed
& Grid computing-science made transparent for everyone
principles, applications and supporting communities tectum.
Marburg, 2-13
Booth C, 1959. Studies of Pyrenomycetes: IV. Nectria (Part I).
Mycological Papers, 73: 1-115
Booth C, 1966. The genus Cylindrocarpon. Mycological Papers,
104: 1-56
Carbone I, Kohn LM, 1999. A method for designing primer sets
for speciation studies in filamentous ascomycetes.
Mycologia, 91: 553-556
Castlebury LA, Rossman AY, Sung GH, Hyten AS, Spatafora JW,
2004. Multigene phylogeny reveals new lineage for
Stachybotrys chartarum, the indoor air fungus. Mycological
Research, 108: 864-872
Castlebury LA, Rossman AY, Hyten AS, 2006. Phylogenetic
relationships of Neonectria/Cylindrocarpon on Fagus in
North America. Canadian Journal of Botany, 84(9):
1417-1433
Chapela IH, Boddy L, 1988. Fungal colonization of attached
beech branches. II. Spatial and temporal organization of
communities arising from latent invaders in bark and
functional sapwood under different moisture regimes. New
Phytologist, 110: 47-57
Chaverri P, Salgado C, Hirooka Y, Rossman AY, Samuels GJ,
2011. Delimitation of Neonectria and Cylindrocarpon
(Nectriaceae, Hypocreales, Ascomycota) and related genera
with Cylindrocarpon-like anamorphs. Studies in Mycology,
68: 57-78
Cummings MP, Huskamp JC, 2005. Grid computing. Educause
Review, 40: 116-117
de Zeeuw R, 1939. Notes on the life of Persoon. Mycologia,
31(3): 369-370
Denk T, 2003. Phylogeny of Fagus L. (Fagaceae) based on
morphological data. Plant Systematics and Evolution, 240:
55-81
Doidge EM, 1950. The South African fungi and lichens to the end
of 1945. Bothalia, 5: 1-1094
Domsch KH, Gams W, Anderson TH, 2007. Compendium of soil
fungi, 2nd taxonomically revised edition by W. Gams.
Eching, Germany, IHW-Verlag. 1-322
Duong TA, De Beer ZW, Wingfield BD, Wingfield MJ, 2012.
Phylogeny and taxonomy of species in the Grosmannia
serpens complex. Mycologia, 104(3): 715-732
Ehrlich J, 1934. The beech bark disease: a Nectria disease of
Fagus, following Cryptococcus fagi (Baer.). Canadian
Journal of Research, 10(6): 593-692
Fries EM, 1823. Systema Mycologicum.Vol. 2(2). Lund, Sweden.
Fries EM, 1832. Systema Mycologicum.Vol. 3(2). Sumptibus
Ernesti Mauritii, Greifswald.
Fries EM, 1849. Summa Vegetabilium Scan dinaviae II. Upsaliae.
Unknown publisher. 1-387
Griffin JM, Lovett GM, Arthur MA, Weathers KC, 2003. The
distribution and severity of beech bark disease in the Catskill
Mountains, N.Y. Canadian Journal of Forest Research, 33:
1754-1760
Gwiazdowski RA, Van Driesche RG, Desnoyers A, Lyon S, Wu
SA, Kamata N, Normark BB, 2006. Possible geographic
origin of beech scale, Cryptococcus fagisuga (Hemiptera:
Eriococcidae), an invasive pest in North America. Biological
Control, 39: 9-18
Hewitt CG, 1914. Note on the occurrence of the felted beech
coccus Cryptococcus fagi (Baerens.) Dougl. in Nova Scotia.
The Canadian Entomologist, 46: 15-16
Hirooka Y, Rossman AY, Chaverri P, 2011. A morphological and
phylogenetic revision of the Nectria cinnabarina species
complex. Studies in Mycology, 68: 35-56
Hirooka Y, Rossman AY, Samuels GJ, Lechat C, Chaverri P,
2012. A monograph of Allantonectria, Nectria, and
Pleonectria (Nectriaceae, Hypocreales, Ascomycota) and

ISSN1672-6472 CN11-5180/Q Mycosystema May 15, 2013 Vol.32 No.3
http://journals.im.ac.cn/jwxtcn
516
their pycnidial, sporodochial, and synnematous anamorphs.
Studies in Mycology, 71: 1-210
Houston DR, 1994. Major new tree disease epidemics: beech bark
disease. Annual Review of Phytopathology, 32: 75-87
Huelsenbeck JP, Ronquist F, Nielsen ES, Bollback JP, 2001.
Bayesian inference of phylogeny and its impact on
evolutionary biology. Science, 294: 2310-2314
Huelsenbeck JP, Larget B, Miller RE, Ronquist F, 2002. Potential
applications and pitfalls of Bayesian inference of phylogeny.
Systematic Biology, 51: 673-688
John DM, Maggs CA, 1997. Species problems in eukaryotic
algae: a modern perspective. Claridge MF, Dawah HA,
Wilson MR (eds.) In: Species: The Units of Biodiversity.
Chapman & Hall, London. 83-107
Katoh T, 2008. Recent developments in the MAFFT multiple
sequence alignment program. Briefings in Bioinformatics, 9:
286-298
Kirk PM, Cannon PF, Minter DW, Stalpers JA, 2008. Dictionary
of the fungi. 10th edition. CAB International, Wallingford.
1-771
Kornerup A, Wanscher JH, 1978. Methuen Handbook of Colour.
3rd edition. Methuen, London. 1-252
Kunze G, Schmidt JC, 1817. Mykologische Hefte 1: i-xvi, 1-109,
2 plates. Germany, Leipzig; G. Voss'sche Buchhandlung.
Link JHF, 1809. Magazin Gesellsehafl naturforschung Freunde,
Berlin 3: 8
Luo J, Zhuang WY, 2010. Three new species of Neonectria
(Nectriaceae, Hypocreales) with notes on their phylogenetic
positions. Mycologia, 102: 142-152
MacKenzie M, Iskra AJ, 2005. The first report of beech bark
disease in Ohio comes nineteen years after the first report of
the initiating scale. Plant Disease, 89: 203
Maddison WP, Maddison DR, 2010. Mesquite: a modular system
for evolutionary analysis. Version 2.73 http://
mesquiteproject.org. Accessed October 2012
Mahoney EM, Milgroon MG, Sinclair WA, Houston DR, 1999.
Origin, genetic diversity, and population structure of Nectria
coccinea var. faginata in North America. Mycologia, 91:
583-592
McIntosh C, 1849. Disease in beech trees. North British
Agriculturist and Journal of Horticulture, 1: 409
Myers DS, Bazinet AL, Cummings MP, 2008. Expanding the
reach of Grid computing: combining Globus- and
BOINC-based systems. In: Talbi EG, Zomaya A (ed.) Grids
for Bioinformatics and Computational Biology. John Wiley
& Sons, New York. 71-85
Nirenberg HI, 1976. Untersuchungen über die Morphologische
und biologische differenzierung in der Fusarium-Sektion
Liseola. Mitteilungen aus der Biologischen Bundesanstalt
für Land und Forstwirtschaft, Berlin-Dahlem, 169: 1-117
O’Brien JG, Ostry ME, Mielke ME, Mech R, Heyd RL,
McCullough DG, 2001. First report of beech bark disease in
Michigan. Plant Disease, 85: 921
O’Donnell K, Cigelnik E, 1997. Two divergent intragenomic
rDNA ITS2 types within a monophyletic lineage of the
fungus Fusarium are nonorthologous. Molecular
Phylogenetics and Evolution, 7: 103-116
Parker IM, Gilbert GS, 2004. The evolutionary ecology of novel
plant-pathogen interactions. Annual Review of Ecology,
Evolution, and Systematics, 35: 675-700
Persoon CH, 1800. Icones et Descriptiones Fungorum Minus
Cognitorum. Vol. 2. Leipzig.
Petersen RH, 1977. Some brief reflections on C.H. Persoon. Kew
Bulletin, 31(3): 695-698
Posada D, 2008. jModelTest: Phylogenetic Model Averaging
Averaging. http://darwin.uvigo.es/
Posada D, Buckley TR, 2004. Model selection and model
averaging in phylogenetics: advantages of the AIC and
Bayesian approaches over likelihood ratio tests. Systematic
Biology, 53: 793-808
Rehner SA, 2001. Primers for elongation factor 1-a (EF1-a).
www.nacse.org/yfaaberg/aftol/EF1primer.pdf
Rossman AY, Samuels GJ, Rogerson CT, Lowen R, 1999. Genera
of Bionectriaceae, Hypocreaceae and Nectriaceae
(Hypocreales, Ascomycetes). Studies in Mycology, 42: 1-248
Schoch CL, Seifert KA, Huhndorf S, Robert V, Spouge JL, et al.,

Yuuri HIROOKA et al. / Species delimitation for Neonectria coccinea group including the causal agents of beech bark disease in Asia, …
Mycosystema
517
2012. Nuclear ribosomal internal transcribed spacer (ITS)
region as a universal DNA barcode marker for Fungi.
Proceedings of the National Academy of Sciences, 109:
6241-6246
Seifert KA, 1985. A monograph of Stilbella and some allied
Hyphomycetes. Studies in Mycology, 27: 1-235
Seifert KA, 2009. Progress towards DNA barcoding of fungi.
Molecular Ecology Resources, 9: 83-89
Sieber TN, 2007. Endophytic fungi in forest trees: are they
mutualists? Fungal Biology Review, 21: 75-89
Swofford DL, 2002. PAUP*. Phylogenetic analysis using
parsimony (*and other methods), ver 4.10. Sinauer
Associates, Sunderland
Taylor JW, Jacobson DJ, Kroken S, Kasuga T, Geiser DM,
Hibbett DS, Fisher MC, 2000. Phylogenetic species
recognition and species concepts in fungi. Fungal Genetics
and Biology, 31: 21-32
Vilgalys R. n.d. Conserved primer sequences for PCR
amplification and sequencing from nuclear ribosomal RNA.
http://www.biology.duke.edu/fungi/mycolab/primers.htm
Vilgalys Lab, Durham, NC
White TJ, Bruns T, Lee S, Taylor J, 1990. Amplification and
direct sequencing of fungal ribosomal RNA genes for
phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White
TJ (eds.) PCR protocols: a guide to methods and
applications. Academic Press, San Diego, California, USA.
315-322
Wollenweber HW, 1916. Fusaria autographice delineate.
Published by the author, Berlin. 1100 plates
Wollenweber HW, 1928. Über Fructformen der krebserregenden
Nectriaceen. Z. Parasitenk. (Berlin), 1: 138-173
Zhao P, Luo J, Zhuang WY, Liu XZ, Wu B, 2011. Practice
towards DNA barcoding of the nectriaceous fungi. Fungal
Diversity, 46: 183-191
Zwickl DJ, 2006. Genetic algorithm approaches for the
phylogenetic analysis of large biological sequence datasets
under the maximum likelihood criterion. PhD dissertation,
The University of Texas at Austin, Austin. TX. 1-125





























