Embed Size (px)
Citation preview

hon p.1 [100%]
61
富山大学大学院医学薬学研究部(〒9300194 富山市杉
谷 2630)e-mail: hayashi9@pha.u-toyama.ac.jp本総説は,平成 18 年度宮田記念学術論文賞の受賞を記
念して記述したものである.
61YAKUGAKU ZASSHI 128(1) 61―79 (2008) 2008 The Pharmaceutical Society of Japan
―Reviews―
天然物の抗ウイルス活性評価と応用に関する研究
林 利 光
Studies on Evaluation of Natural Products for Antiviral EŠects and Their Applications
Toshimitsu HAYASHI
Graduate School of Medicine and Pharmaceutical Sciences for Research, University of Toyama,2630 Sugitani, Toyama City 9300194, Japan
(Received August 23, 2007)
In the search for novel antiviral molecules from natural products, we have discovered various antiviral moleculeswith characteristic mechanisms of action. Scopadulciol (SDC), isolated from the tropical medicinal plant Scoparia dul-cis L., showed stimulatory eŠects on the antiviral potency of acyclovir (ACV) or ganciclovir (GCV). This eŠect of SDCwas exerted via the activation of viral thymidine kinase (HSV-1 TK) and, as a result, an increase in the cellular concen-tration of the active form of ACV/GCV, i.e., the triphosphate of ACV or GCV. On the basis of these experimentalresults, cancer gene therapy using the HSV-1 tk gene and ACV/GCV together with SDC was found to be eŠective in sup-pressing the growth of cancer cells in animals. Acidic polysaccharides such as calcium spirulan (Ca-SP) from Spirulinaplatensis, nosto‰an from Nostoc ‰agelliforme, and a fucoidan from the sporophyll of Undaria pinnatiˆda (mekabufucoidan) were also found to be potent inhibitors against several enveloped viruses. Their antiviral potency was depen-dent on molecular weight and content of the sulfate or carboxyl group as well as counterion species chelating with sulfategroups, indicating the importance of the three-dimensional structure of the molecules. In addition, unlike dextran sul-fate, Ca-SP was shown to target not only viral absorption/penetration stages but also some replication stages of progenyviruses after penetration into cells. When mekabu fucoidan or nosto‰an was administered with oseltamivir phosphate,their synergistic antiviral eŠects on in‰uenza A virus were conˆrmed in vitro as well as in vivo.
Key words―antiviral eŠect; Scoparia dulcis; scopadulciol; calcium spirulan; nosto‰an; mekabu fucoidan
1. はじめに
後天性免疫不全症候群(エイズ:AIDS)は,
1981 年の夏に世界で初めて発見された感染症で,2
年後にウイルス(human immunode‹ciency virus,
HIV)がその病原体であることが明らかにされ
た.1,2)それから既に 25 年あまりが経過したが,こ
の間に,エイズの診断方法が確立されるとともに,
HIV の子孫ウイルスの複製に関与する特定の分子
を標的にした種々の抗 HIV 薬の開発が行われ,臨
床応用されるようになった.すなわち,これまでに,
HIV に特有の逆転写酵素やプロテアーゼなどに対
する阻害剤3,4)や,エンベロープを持つウイルスが
宿主細胞に吸着・侵入する際に関与するケモカイン
(CCR5)などのコレセプターへの結合阻害剤5)など
が開発され,臨床利用されている.現在,エイズ
は,もはや「死の病」ではないとされているが,そ
れは多剤併用療法(HAAT)6,7)をきちんと生涯に渡
って受けることが可能ならばという条件付きであ
る.サハラ砂漠以南のアフリカ,インド,東南アジ
アや南米では,相変わらず新たな HIV 感染者が発
生している.わが国においても感染者が増加してお
り,かならずしも楽観できない状況である.HIV
感染症が難治性である最大の理由は,HIV のゲノ
ムが宿主のゲノムに組み込まれてしまう点である.
現在,実施されている治療法には,いずれも,組み
込まれた HIV ゲノムを追い出し,消滅させる効果
はない.そのため,HIV 感染者は,抗ウイルス薬
を生涯服用しなければならなず,過重な経済的負担
や副作用の出現,耐性ウイルスの発生などの重大な
問題に悩まされている.加えて,今世紀になってか
ら,重症急性呼吸器症候群(SARS)や鳥インフル

hon p.2 [100%]
62
Fig. 1. Structures of Diterpenes Isolated from S. dulcis and Aphidicolin
62 Vol. 128 (2008)
エンザなどの高病原性ウイルス感染症が発生し新た
な問題となっている.
一方,この間に,医療技術の進歩や衛生環境の改
善などを背景にして高齢化社会が到来するととも
に,生活様式の欧米化に伴ったがん患者や種々の生
活習慣病患者の増加や,臓器移植者の増加がみられ
る.このような人達においては,生体の防御力であ
る免疫力が低下しており,感染症に罹り易く,しか
も重症化する傾向が生まれている.それゆえ,現
在,様々な感染症に対する新たな治療戦略の構築へ
の期待が高まっている.
筆者らは,古来から人々の健康の維持・増進や疾
病の予防・症状の改善に用いられてきた薬用植物や
食用藻類などのウイルス感染症に対する有用性の評
価とそれらの応用について検討してきたので,本稿
ではこれらの研究の主な成果について概説する.
2. 熱帯性薬用植物 Scoparia dulcis L. 由来新規
抗ウイルス活性ジテルペンに関する研究
2-1. Scoparia dulcis L. 由来新規ジテルペン類の
単離と抗ウイルス活性評価 Scoparia dulcis L.
は,南米,アジア及びアフリカ大陸の熱帯及び亜熱
帯地域に広く自生するゴマノハグサ科の多年生草本
で,いずれの地域でも,民間薬として用いられてい
る.筆者らは,1985 年に開始された国際協力事業
団の支援事業「南米パラグァイ共和国の先住民族グ
ァラニーインディオの伝承薬の調査研究」に参加す
る機会を得たが,S. dulcis は,その際に実施した生
物活性試験の対象とした薬草の一種である.本薬草
の成分検索の結果,二環性ジテルペン(scoparic
acid A, scoparic acid B, scoparic acid C, scopadiol)
とともに,カビの代謝産物で,DNA polymerase a
阻害薬として市販されている aphidicolin と類似し
た構造を有する四環性ジテルペン類[scopadulin,
scopadulcic acid A (SDA), scopadulcic acid B
(SDB), scopadulciol (SDC)]が単離された(Fig.
1).810)これらのジテルペン類について,宿主細胞
に対する 50%増殖阻害濃度(CC50)と単純ヘルペ
スウイルス 1 型(HSV-1)に対する 50%増殖阻害
濃度(IC50)をそれぞれ求め,両者の比(選択指数,
SI, CC50/IC50)を算出して抗ウイルス活性を評価し
たところ,SDB と SDC に活性が認められた.両者
の作用機序を検討するために,DNA polymerase に
対する阻害作用を調べた結果,SDB は aphidicolin
と同じように HSV-1 由来 DNA polymerase 阻害作
用を示した.また,これらの阻害強度は,宿主細胞
(HeLa cells)由来 DNA polymerase に対する作用
より顕著であった.一方,SDC にはこのような
DNA polymerase 阻害作用は認められなかったが,
HSV-1 を SDC の存在下で 6 時間処理後,残存する
ウイルスの感染力を調べたところ,濃度依存的に感

hon p.3 [100%]
63
Table 1. EŠects of SDB on Experimental HSV-1 Corneal Infection
Sample Dose(mg/kg/day)
Duration oftreatment(days)
Survivors/total
Mean lag phase ofhamster that developed
facial lesion (days±S.D.)
Mean survival timeof hamster that
died (days±S.D.)
Time intervalof death(days)
H2O 0 7 0/5 4.0±0.71 7.6±0.55 78SDB 20 7 0/5 5.4±1.34 8.2±0.84 79
SDB 100 7 2/5 5.2±3.29 9.3±1.53 811
H2O 0 5 0/9 3.4±0.88 6.7±0.71 68
SDB 100 5 2/9 4.6±0.50a) 8.4±1.51b) 710
Hamsters were infected with 3×104 PFU of HSV-1 HF strain and treated perorally twice per day. a) Statistically diŠerent from the H2O-treated animals (p<0.01, Student's t-test). b) Statistically diŠerent from the H2O-treated animals (p<0.05, Student's t-test).
63No. 1
染力が減弱した.これらの実験結果から SDC の抗
ウイルス作用はウイルスの不活化によることが示唆
された.10)
次に,SDB の in vivo での抗 HSV-1 作用を確認
するために,ゴールデンハムスターを用いて評価し
た.11)すなわち,ゴールデンハムスターの右眼の角
膜に HSV-1 (3×104 PFU/10 ml)を接種後,10 mg/
kg 又は 50 mg/kg の SDB を,それぞれ 7 日間又は
5 日間,1 日 2 回経口投与し,炎症の発生までの日
数と生存日数について対照群(蒸留水投与群)と比
較した.Table 1 に示したように,SDB (10 mg/
kg)を 7 日間経口投与した場合は,対照群に比べ
て炎症の発生が遅延するとともに,生存日数の延長
が確認された.また,50 mg/kgの SDB を 1日 2 回,
5 日間経口投与した場合は,統計的に有意な炎症発
生の遅延と生存日数の延長が認められた.なお,
SDB 投与群の動物には,生存期間中に活動や食欲
の変化は観察されなかった.また,生存した動物の
炎症は,ウイルス感染 2 週間後に治癒し始めた.
2-2. SDC と抗ヘルペスウイルス薬との相乗作用
SDC は SDB のカルボキシル基がヒドロキシルメチ
ル基と置換した構造を持つが,その抗 HSV-1 活性
発現の一端はウイルスの不活化作用によると推察さ
れた.このような作用は,臨床利用されている抗ヘ
ルペスウイルス薬 acyclovir (ACV)の作用機序と
は明らかに異なる.ちなみに,ACV は,HSV-1 感
染細胞内では,ウイルス由来チミジンリン酸化酵素
(TK)によってリン酸化されて一リン酸化体
(ACV-MP)となり,さらに ACV-MP は宿主細胞
由来のリン酸化酵素によりリン酸化されて二リン酸
化体(ACV-DP),三リン酸化体(ACV-TP)にま
で代謝される(Fig. 2).このように細胞内で生じ
た ACV-TP が活性本体として作用することが知ら
れている.12)そこで,HSV-1 増殖に及ぼす SDC と
ACV の相乗作用の有無を調べた.すなわち,
HSV-1 感染 HeLa 細胞を種々の濃度の SDC と
ACV を含む培地で培養し,それぞれの組み合わせ
における抗 HSV-1 作用を評価した.その結果,
0.050.1 mM SDC と 0.410 mM ACV の組み合わせ
間で相乗作用が認められた.13)なお,SDC の細胞
毒性は,100 mM 以下の ACV の併存下でも増強さ
れなかった.このような相乗作用の発現メカニズム
を解明するために,SDC と ACV 併用投与時にお
け る 細 胞 内 の deoxyribonucleoside triphosphate
pools (dNTP)を分析したところ,Table 2 に示し
たような結果が得られた.すなわち,SDC と ACV
を併用投与した場合,ACV 単独投与の場合に比べ
て dCTP, dTTP, dATP, dGTP のいずれの三リン酸
化体の含量も有意に増大した.また,この場合の細
胞内の ACV の三リン酸化体(ACV-TP)の濃度は
ACV 単独投与の場合と比べて 4 倍以上に増加した.
ACV によるウイルス DNA 合成阻害作用は感染細
胞内で生成される ACV-TP の量に依存していると
考えられるので,SDC は,ACV-TP 量を増加させ
ることによってその HSV-1 DNA polymerase との
結合を促進し,結果的に ACV の抗 HSV-1 活性を
増強させたと推察される.すなわち,HSV-1 感染
細胞内における dNTPs 含量の増大は,ACV-TP の
DNA polymerase 阻害作用によって dNTP pools か
ら DNA への合成が抑制された結果であると考えら
れる.このことは, SDC 単独投与の場合には
dNTPs 含量が増えなかったことからも支持され
る.ところで,SDC による ACV-TP 含量増大のメ
カニズムについては,1 つの可能性として TK の活

hon p.4 [100%]
64
Fig. 2. Enzymatic Phosphorylation of ACV and GCV to Their Mono-, Di-, and Triphosphate Derivatives which Contribute to An-tiviral Therapies
Table 2. EŠects of SDC and ACV on Intracellular Deoxyribonucleoside Triphosphate Pools
SDC(mM)
ACV(mM)
pmole per 106 cells
dCTP dTTP dATP dGTP ACV-TP
0 0 10.0±0.6 551±46 9.6±2.5 9.4±1.1 ―
0.1 0 10.1±1.7 464±44b) 8.7±2.2 6.4±2.0 ―
0 100 19.4±5.9a) 659±123a) 50±16a) 24.2±2.7d) (100±16)
0.1 100 46.0±19.2a) 1184±266c) 92±22d) 31.3±4.8d) (443±81)d)
Cells were infected with 10 PFU of HSV-1 per cell. After 1 h, the indicated concentrations of SDC and ACV were added. After an additional incubation of 8 h,the pool sizes were analysed by HPLC. Data were expressed as the mean±standard deviations of quadruplicate experiments. a) Signiˆcant increase comparedwith untreated control on the basis of the Student's t-test (p<0.05). b) Signiˆcant decrease compared with untreated control on the basis of the Student's t-test(p<0.05). c) Signiˆcant increase compared with untreated control on the basis of the Student's t-test (p<0.01). d) Signiˆcant increase compared with untreat-ed control on the basis of the Student's t-test (p<0.005).
64 Vol. 128 (2008)
性化が考えられる.そこで,ACV のリン酸化に関
与している 2 種類のリン酸化酵素,すなわち,
HSV-TK と細胞内 TK の酵素活性に及ぼす SDC の
影響を調べるため,TK 非発現 HeLa 細胞(HeLa-
TK-)と HSV-tk 発現 HeLa 細胞(HeLa-TK+)に
おける TK 活性を比較したところ,SDC は HeLa-
TK- 細胞の TK 活性には影響を与えなかったが,
HeLa-TK+ 細胞における TK 活性を有意に上昇させ
た.
以上のように,SDC は,従来報告されてきた抗
ウイルス活性物質には見い出されたことがない特異
な作用を示すことが明らかになった.本研究結果
は,現在臨床利用されている抗ウイルス薬の副作用
や耐性ウイルスの出現などの問題点を克服する上で
有用な知見となるであろう.
2-3. SDC のがん遺伝子治療への応用の試み
がん遺伝子治療法として,自殺遺伝子と呼ばれてい
る HSV-TK 遺伝子(HSV-tk)を用いる方法がある
が,これは,HSV-tk をがん細胞に導入後,ACV
又は ganciclovir (GCV)を投与してがん細胞の増
殖を抑制しようとするものである.14)先述のように,
SDC は HSV-TK 活性化作用を持つので,このがん
遺伝子治療に応用できると思われた.そこで,株化
された種々のがん細胞に HSV-tk を導入し,ACV
又は GCV の細胞毒性が SDC と併用することによ
り増強されるか否かを検討した.15,16)

hon p.5 [100%]
65
Table 3. Cell Growth Inhibition of ACV and GCV against Human Cancer Cells Expressing HSV-TK in the Ab-sence or Presence of 0.04 mM SDC
Cell lineCytotoxicity (IC50, mM)a)
ACV ACV+SDC GCV GCV+SDC
HEp-2 1947±328b) 2120±191 760±62 677±76HEp-2-HSTK 119±11 8.8±0.70c) 6.5±2.1 0.64±0.24d)
EJ-1 1917±150 1963±214 423±16 453±44
EJ-1-HSTK 185±38 23±5.1d) 49±6.3 1.2±0.16c)
HeLa 229 3147±434 3350±656 490±26 488±49HeLa 229-HSTK 166±9.1 10.9±3.0c) 3.4±0.44 0.15±0.015c)
YKG-1 2193±156 1873±367 497±85 610±61
TKG-1-HSTK 83±5.1 4.0±1.9c) 1.6±0.42 0.32±0.165c)
a) 50% inhibitory concentration (IC50) of ACV or GCV for cell inhibition. b) Each value is the mean±S.D. of triplicate assays.c) p<0.01 vs. prodrug alone. d) p<0.001 vs. prodrug alone.
Fig. 3. In vivo eŠects of SDC on Antitumor Actions of ACVin HSV-TK-expressing Tumors
SDC (1 mg/day) and ACV (4 mg/day) were orally administrated threetimes a day (every 8 h). Bar: mean±S.D. from ten determinations. +++p<0.001 vs. no drug control, #p<0.05 and ##p<0.01 vs. ACV monotherapy
65No. 1
まず,ヒトの株化がん細胞である larynx carcino-
ma (HEp-2), bladder caricinoma (EJ-1), cervical
carcinoma (HeLa 229),及び glioblastoma (YKG-1)
の増殖に対する ACV 及び GCV の影響を調べたと
ころ,顕著な阻害作用は認められなかった.一方,
HSV-tk を導入したそれぞれのがん細胞の増殖に対
しては,著しい阻害活性の上昇が観察された.すな
わち,HSV-tk を導入したがん細胞に対する ACV
及び GCV の細胞毒性(IC50)は,野生株に対する
細胞毒性に比べて,それぞれ 1026 倍及び 9311 倍
増強された(Table 3).SDC (0.04 mM)の単独投
与は,野生株及び TK 発現細胞の両者に毒性を示さ
なかった.また,SDC を ACV 又は GCV と併用し
た場合でも,野生株に対する細胞毒性は増大しなか
った.ところが,HSV-tk を導入したがん細胞に対
する細胞毒性は,SDC を ACV と併用した場合,
ACV 単独の場合に比べて 821 倍も強くなった
(Table 3).SDC を GCV と併用した場合も同様の
傾向が認められた(GCV 単独の場合の 541 倍の増
強).したがって,細胞株の種類の違いによりその
程度に差があるものの,いずれの TK 発現がん細胞
株に対しても,SDC は,ACV や GCV の細胞毒性
を顕著に増強することが確認された.
次に, in vivo での効果を確認するために,
HeLa-TK+ 細胞をヌードマウスの皮下に接種し,
腫瘍組織の増殖に及ぼす SDC と ACV 又は GCV と
の併用効果を検討した.SDC を単独で経口投与(1
mg/day, 1 日 3 回)した場合の腫瘍組織量は,対照
群と比較して有意差は認められなかったが,ACV
と併用した場合の腫瘍組織量は,対照群のみならず
ACV 単独投与群と比べて有意に抑制された(Fig.
3).なお,被検試料を腹腔内投与した場合も同様の
結果が得られた.また,SDC (1 mg/day)を GCV
(2.5 mg/day)と併用して腹腔内投与した場合の腫
瘍組織の増殖も,対照群及び GCV 単独投与群と比
べて有意に抑制された.ところで,現実的には,腫
瘍組織のすべての細胞に TK を発現させることは困
難である.そこで,HeLa-TK+ 細胞と HeLa-TK-
細胞を 1:9 に混合したものを皮下に接種した場合

hon p.6 [100%]
66
Fig. 4. In vivo Bystander EŠects of ACV on Tumors Inducedby Mixtures of 90% HSV-TK- and 10% HSV-TK+ HeLaCells in the Absence or Presence of SDC
SDC (1 mg/day) and ACV (4 mg/day) were orally administrated threetimes a day (every 8 h). Bar: mean±S.D. from ten determinations. +p<0.05, ++p<0.01 and +++p<0.001 vs. no drug control, #p<0.05, ##p<0.01and ###p<0.001 vs. ACV monotherapy.
66 Vol. 128 (2008)
に,同様の効果が得られるか否かを検討した.
Figure 4 に示したように,ACV 単独投与の場合
は,腫瘍組織の増殖が有意に抑制され,SDC と併
用することにより,さらに強い増殖抑制効果が認め
られた.GCV の場合も同様に SDC との併用によ
る腫瘍組織増殖抑制効果が確認された.なお,一部
の細胞に対する効果が周囲の細胞にも影響が及ぶ現
象は bystander eŠect17) と呼ばれているが,今回こ
のような bystander eŠect が認められたことは,
SDC を ACV/GCV によるがんの遺伝子治療に応用
することが可能であることを示している.
2-4. SDB 及び SDC に認められたその他の生物
活性 S. dulcis は,熱帯及び亜熱帯地域で民間薬
として使用され,その適応症は,消化,健胃,腹
痛,鎮咳,解熱,肺炎,気管支炎,喉頭炎,肝臓障
害,麻疹,糖尿病,高血圧など多岐に渡っている.
そこで,本植物から単離された SDB や SDC につ
いて,種々の生物活性を評価したところ,プロトン
ポンプ抑制作用18,19)や骨吸収抑制作用20)などが新た
に見い出され,それぞれ胃潰瘍や骨粗鬆症などに対
して有用である可能性が示唆された.
SDB や SDC の構造が高等植物由来成分としては
珍しい炭素骨格を持つことや,本稿で紹介したよう
な多彩な生物活性を示したことから,国内外の合成
化学者の注目を集め,化学合成が試みられた結果,
カリフォルニア大学の Overman ら21,22)やエール大
学の Ziegler ら23)らによって相ついで全合成が達成
された.
3. 食用藻類由来新規抗ウイルス活性糖鎖分子に
関する研究
藻類は光合成能を有しているが,その多くは,陸
上の高等植物とは異なる環境下に生育していること
から,藻類特有の代謝経路を持ち,産生される成分
も多様である.これまで,海藻由来の硫酸化多糖類
は,抗ウイルス活性を示すことが報告されている
が,その多くは,硫酸化多糖のポリアニオンとして
の性質に基づくウイルスの宿主細胞への吸着・侵入
段階が作用標的であると報告されている.24,25)
3-1. スピルリナ Spirulina platensis 由来抗ウイ
ルス活性分子に関する研究
3-1-1. スピルリナ由来抗 HSV-1 活性硫酸化多糖
の分離と構造 スピルリナは螺旋状の藍藻で,強
アルカリ性(pH 10 程度)の塩湖に自生する微細藻
類の一種である.本藻体は良質のタンパク質のほ
か,糖質,脂質,カロテノイドやビタミン類を豊富
に含んでいることから,新しい食糧源として注目さ
れている.また,その抽出物は,抗腫瘍作用,血清
コレステロール低下作用,血圧低下作用,抗体産生
促進作用などの多彩な生物活性を示すことが報告さ
れている.2628)一方,スピルリナは,人工培養によ
る大量生産が可能となり,現在,健康食品や機能性
食品としても利用されている.
スピルリナの凍結乾燥粉末の熱水抽出エキスから
調製した透析内液(SP-H)について,抗 HSV-1 活
性を指標にしながら活性成分の分離・精製を行っ
た.すなわち,SP-H の trichloroacetic acid 可溶部
について,陰イオン交換樹脂(Toyopearl DEAE
650M)カラムクロマトグラフィーとゲルろ過
(Sepharose 6B, Toyopearl HW-65)を組み合わせて
分画したところ,HPLC 分析において単一ピーク
として検出され,セルロースアセテート膜電気泳動
においても単一バンドとして泳動される硫酸化多糖
(Na-SP)が得られた.29) Na-SP の見掛けの分子量
は 7.44×104 で,主な構成単糖は rhamnose, 3-O-
methylrhamnose (acofriose)及びウロン酸であっ

hon p.7 [100%]
67
Table 4. Anti-HSV Activity of Metal Cation Exchanged Derivatives of Na-SP
Sample CytotoxicityCC50 (mg/ml)
Antiviral activity (IC50 (mg/ml)) Selectivity index (CC50/IC50)
A B A B
Na-SP 6000 0.46 4.8 13043 1250
Ca-SP >10000 0.74 4.7 >14000 >2100K-SP 7700 0.88 3.5 8750 2200
Ag-SP 7.0 1.6 3.2 4.4 2.2
Mg-SP 310 1.7 4.3 182 72
Mn-SP 630 1.8 5.0 350 126Co-SP 100 1.1 3.1 91 32
Ni-SP 260 2.8 5.2 93 50
Cu-SP 28 0.23 3.3 122 8.5
Zn-SP 320 3.3 5.3 97 60Cd-SP 2.2 1.0 1.1 2.2 2.0
Pb-SP 3200 1.3 1.8 2462 1778
Fe(II)-SP 3200 2.9 8.4 110 381
Fe(III)-SP 2200 >10 >10 <220 <220Al-SP 6200 2.6 8.5 2384 729
Cr(III)-SP 1100 >100 >100 <11 <11
A: Sample was added to the medium during viral infection and throughout the incubation, B: Sample was added to the medium immediately after viral infec-tion.
67No. 1
た.また,重水素化 methyl iodide (CD3I)を用い
て Na-SP を完全メチル化し,加水分解産物の GC-
MS 分析や部分加水分解産物の ESI-MS 分析を行っ
たところ,Na-SP の構造は,rhamnose と glucuron-
ic acid 又は galacturonic acid が交互に結合した糖
鎖のコア部分に,rhamnose と acofriose が交互に結
合した糖鎖のラムナン部分が結合したもので,さら
に,ラムナン部分に硫酸基が結合していると推察さ
れた(Fig. 5).30)従来,ラムナン骨格を有する多糖
は,緑藻ヒトエグサ Monostroma nitidum やヒロハ
ノヒトエグサ M. latissimum 由来ラムナン硫酸など
の単離例があるが,他の報告例は少ない.また,
Na-SP はその糖鎖中に,親水性の硫酸基と疎水性
のメチル基が存在することから,両親媒的性質を持
っていると推察され,この特徴的構造が多様な生物
活性の発現に関与していると考えられる.
3-1-2. Na-SP の Na+ と他の金属イオンとの置
換による抗ウイルス活性の変化 あらかじめ
Na+ 以外の金属塩溶液で平衡化しておいた陽イオ
ン交換樹脂(Dowex 50 W×8)を充したカラム
に Na-SP 溶液を付したのち,蒸留水で溶出させる
と他の金属イオンに置換した多糖体が得られる.様
々な金属イオンとの置換体について,抗 HSV-1 活
性を評価し,Table 4 にその結果を示した.31) Ca2+
塩の活性は上昇したのに対し,K+ 塩の活性はやや
低下した.他の置換体の選択的活性はいずれも顕著
に低下したが,その主たる要因は細胞毒性の上昇で
あると考えられた.
3-1-3. Na-SP の抗 HSV-1 作用のメカニズム
Na-SP の Na+ を Ca2+ と置換して得られた Ca-
SP が最も強い抗 HSV-1 活性を示したので,Ca-SP
の抗ウイルス活性スペクトルを調べたところ,本多
糖体は,エンベロープを有するウイルスに対して顕
著な増殖抑制作用を示したが,poliovirus や cox-
sackievirus などのエンベロープを持たないウイルス
に対しては全く活性を示さなかった(Table 5).32)
また,ウイルス感染前から培養液中に試料が存在し
た場合の方が感染と同時に試料を添加した場合より
抗 HSV-1 活性が強かったことから,Ca-SP の作用
標的段階はウイルスの宿主細胞への吸着・侵入段階
であると推測された.この点を確認するために,
time-of-addition 実験を行った.Table 6 に示したよ
うに,ウイルス感染中に Ca-SP が存在した場合に
顕著な増殖抑制作用が認められたが,感染 2 時間後
に添加した場合にも抗 HSV-1 活性が認められた.
このような作用は,Na-SP にも認められた.これ
らの結果は,Ca-SP や Na-SP は,ウイルスの吸
着・侵入段階を標的とするだけでなく,侵入後の子

hon p.8 [100%]
68
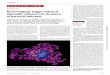
Fig. 5. Proposed Linkage Mode of Component Sugars of Na-SP
Fig. 6. Confocal Laser Fluorescence microscopy of HSV-1-infected Cells Treated with FI-SPFI-SP was added to the medium of HSV-1-infected cells and incubated for 3 h. The cells were ˆxed with 3% paraformaldehyde and treated with rabbit anti-
HSV-1 serum and goat Cy3-conjugated anti-rabbit IgG.
68 Vol. 128 (2008)
孫ウイルスの複製段階をも標的とすることを示唆し
ている.
そこで,まず,4°C で HSV-1 を宿主細胞に吸着
させたのち,種々の濃度の Ca-SP の存在下で培養
温度を 37°C に上昇させ,1 又は 6 時間後に宿主細
胞内へ侵入したウイルス量を調べたところ,Ca-SP
は濃度依存的にウイルスの細胞内への侵入を抑制し
た.32)また,HIV-1 感染における細胞間の融合に及

hon p.9 [100%]
69
Table 5. Inhibitory EŠect of Ca-SP on the Replication of DiŠerent Viruses
Virus Host cell Cytotoxicity(CC50, mg/ml)a)
Antiviral activity(ID50, mg/ml)b)
Selectivity index(CC50/IC50)
Ac) Bd) A B
HSV-1 HeLa 7900 16.5 0.92 479 8587
HCMV HEL 4800 41 8.3 117 578
Measelesvirus Vero 6300 39 17 162 371
Mumsvirus Vero 6300 92 23 68 274IFV MDCK 5400 230 9.4 23 574
HIV-1 MT-4 2900 11.4 2.3 254 1261
Poliovirus Vero 6300 2300 2200 2.7 2.9
Coxackievirus Vero 6300 2600 1850 2.4 3.4
a) Concentration required to reduce cell growth by 50%, mean value for two experiments. b) Concentration required to reduce virus replication by 50%, meanfor two experiments. c) Ca-SP was added to the medium immediately after viral infection. d) Ca-SP was added to the medium 3 h before viral infection.
Table 6. EŠect of Time-of-addition of Ca-SP on HSV-1 Replication
Time of additionAntiviral activity(IC50, mg/ml)3 h before
infectionDuring
infection01 h
postinfection12 h
postinfection224 h
postinfection
+a) - - - - >200b)
- + - - - 0.97±0.10- - + - - >200- - - + - >200- - - - + 24.5±4.6+ + - - - 0.92±0.20- + + - - 1.2±0.15- - + + - >200+ + + + + 0.95±0.19- + + + + 0.83±0.12- - + + + 13.2±2.2
a) HeLa cells were treated in the absence (-) or presence (+) of diŠerent concentrations of Ca-SP during the period indicated. b) Each value is the mean±S.D. of triplicate assays.
69No. 1
ぼす Ca-SP の効果を検討するために,Molt-4 細胞
と Molt-4 /HTLV-IIIB 細胞との共培養系を用い
て,惹起される巨細胞の形成に対する作用を調べ
た.その結果,Ca-SP は 25 mg/ml 以上の濃度で巨
細胞形成を完全に抑制することが分かった(Table
7).一方,Ca-SP から金属イオンを除去した場合
(H-SP)や脱硫酸化した場合(desulfated SP)の抗
HSV-1 活性や HIV-1 の巨細胞形成抑制作用は顕著
に低下又は消失した.なお,H-SP や desulfated SP
は 5 mg/ml 以下の低濃度では,逆に巨細胞形成を促
進した.ちなみに,このような巨細胞形成促進作用
は,25 mg/ml 以上の濃度で顕著な巨細胞抑制作用
を示した Na-SP や dextran sulfate (DS)にも認め
られた.これらの実験結果は,Na+ や Ca2+ などの
特定の金属イオンと硫酸基との結合により形成され
る糖鎖分子の立体構造が活性発現に重要な役割を果
たしていることを示唆している.
次に,HSV-1 感染細胞において,硫酸化多糖分
子が実際に細胞内へ取り込まれるか否かを明らかに
するために,Na-SP の蛍光標識体(FI-SP)を調製
して検討した.FI-SP を HSV-1 感染 6 時間後に添
加し,3 時間培養後,固定した細胞をさらに抗
HSV-1 血清と Cy3 標識 2 次抗体を用いて HSV-1 感
染細胞を検出するという二重染色法で処理した.そ
のときの FI-SP の局在部位を共焦点レーザー蛍光
顕微鏡で観察した.Figure 6 に示したように,FI-
SP はウイルス感染 6 時間後に明らかに細胞内に局
在することが認められたのに対し,非感染細胞内に

hon p.10 [100%]
70
Table 7. EŠect of Ca-SP, Na-SP, H-SP and Desulfated SP on Syncytium Formation by Cocultivation of Motl-4 and Motl-4/HTLV-IIIB a)
SamplePercent inhibition of syncytium formation 50% inhibitory
concentration(mg/ml)0.2 mg/ml 1 mg/ml 5 mg/ml 25 mg/ml 100 mg/ml
Ca-SP 6.3±1.2 14±3.9 27±6.7 99±1.2 100 7.3
Na-SP -8±5.0 -7.3±2.9 10±2.2 95±2.9 99±1.2 10.5
H-SP -8.7±1.7 -9.7±6.0 -1.3±0.45 13±3.4 27±8.1 >100
Desulfated SP -12±4.9 -13±5.4 -12±3.5 9.3±4.0 NDb) >25DSc) -13±5.4 -8.6±1.2 3.8±0.64 74±9.9 98±1.6 14.2
a) Sample was added at the time of cocultivation of Motl-4 cells with the counterpart. Each value is the mean±S.D. of triplicate assays. b) ND: not determinedbecause of its high cytotoxicity. c) DS: dextran sulfate.
Fig. 7. In vivo EŠects of Na-SP and ACV on the Development of Facial Lesions after Corneal Inoculation of HSV-1 to BALB/c MiceOral treatment was started just after inoculation and consisted of 2 daily applications.
70 Vol. 128 (2008)
は観察されなかった.このことは,FI-SP がウイル
ス感染細胞に選択的に取り込まれたことを示してい
る.なお,FI-SP は抗 HSV-1 活性を保持している
ことが確認された.
3-1-4. Na-SP の in vivo で の 抗 HSV-1 活 性
以上のように,Na-SP は HSV-1 の細胞への吸
着・侵入段階を標的とするのみならず,自らも
HSV-1 の感染細胞内へ取り込まれ,細胞内で起こ
る子孫ウイルスの複製に関与している生体分子をも
標的とすることが示唆された.そこで,次に,マウ
スを用いて Na-SP の in vivo での抗 HSV-1 作用を
調べた.すなわち,BALB/c マウスの右眼を擦過後,
HSV-1 を感染させ,Na-SP 水溶液を 1 日 2 回経口
投与した.HSV-1 による病変の進行状況は,5 段階
(0:無症状,1:マウスの顔面右側のみの軽度の炎
症,2:マウス顔面右側のみの中程度の炎症,3:マ
ウス顔面右側から左側に及ぶ中程度の炎症,4:マ
ウス顔面全面に及ぶ重度の炎症)で判定し,スコア
化して評価した.対照として抗ヘルペス薬として臨
床で用いられている ACV を用いたが,Na-SP は
ACV 程顕著ではないものの,用量依存的に病変の
進行を抑制した(Fig. 7).
3-1-5. Na-SP の低分子量化の抗 HSV-1 活性に
及ぼす効果 硫酸化多糖の抗ウイルス活性は分子
量の大きさにより変化するとの報告があることから,
Na-SP の低分子量化による抗 HSV-1 活性の変化を
調べた.すなわち,Na-SP を 0.1 M 酢酸緩衝液に溶
解後,H2O2 を添加してラジカル反応を行い,2, 4,
6, 8, 12 及び 24 時間後に一定量の反応液を採取して
低分子量化体を得た.抗 HSV-1 活性は,低分子量
化に伴って減少したが,中でも反応時間が 12 時間
と 24 時間の場合の反応産物[LW-SP (12)と LW-

hon p.11 [100%]
71
Table 8. Anti-HSV-1 Activity of Na-SP and Its Depolymeraized Derivatives (LW-P)
Sample Mr CC50
(mg/ml)
IC50 (mg/ml) Selectivity index (CC50/IC50)
A B A B
Na-SP 210000 6000 0.46 4.8 13000 1300
LW-SP(2)a) 37000 5600 0.5 5.7 11000 980
LW-SP(4) 22600 5000 0.62 7.0 8100 710LW-SP(6) 18500 4800 1.3 16 3700 300
LW-SP(12) 12700 5200 2.1 28 2500 190
LW-SP(24) 10700 4900 5.4 >100 910 <49
a) Numbers in parentheses indicate reaction time (h) with H2O2. A: Sample was added to the medium during viral infection and throughout the incubation,B: Sample was added to the medium immediately after viral infection.
Table 9. Anti-HSV-1 Activity of Oversulfated Spirulan (OS-SPs) and Partially Desulfated Spirulan (PDS-SPs)
Sample Sulfate(%)
Cytotoxicity(CC50, mg/ml)
Anti-HSV-1 activity(IC50, mg/ml)
Selectivity index(CC50/IC50)
A B A B
Na-SP 17.5 7100 0.63 3.1 13000 2300
OS-SP-1 21.1 4400 0.64 0.69 6900 6400
OS-SP-2 23.0 2500 0.46 0.75 5400 3300
OS-SP-3 25.6 3100 0.61 0.64 5100 4800OS-SP-4 27.8 3900 0.74 0.84 5300 4600
OS-SP-5 28.1 3300 0.84 0.76 3900 4300
PDS-SP-1 17.1 6500 1.7 4.1 4200 1600
PDS-SP-2 15.9 5300 1.7 3.8 3500 1400PDS-SP-3 14.4 4500 1.8 12 2500 390
PDS-SP-4 13.8 4500 2.6 19 1800 240
PDS-SP-5 12.3 5100 3.2 20 1600 260
PDS-SP-6 10.7 5100 5.9 34 880 150PDS-SP-7 8.6 3800 20 150 200 26
PDS-SP-8 6.8 3700 49 710 7.9 5.4
PDS-SP-9 5.5 3900 >1000 >1000 <5 <5
PDS-SP-10 2.5 4900 >1000 >1000 <5 <5DS-SP n.d. 4900 >1000 >1000 <5 <5
A: Sample was added to the medium during viral infection and throughout the incubation thereafter, B: Sample was added to the medium immediately afterviral infection, n.d: not detected.
71No. 1
SP (24)]の間に顕著な差が認められた(Table
8).33)このことは,抗 HSV-1 活性発現には,1 万程
度の分子量が必要であることを示している.なお,
両者の構成成分を分析したところ,LW-SP (24)
の硫酸基,uronic acid,及び acofriose の含量が
LW-SP (12)のそれらと比較して少なかったのに
対し,rhamnose の含量には差がなかった.
3-1-6. Na-SP の部分脱硫酸化及び過硫酸化の抗
HSV-1 活性に及ぼす効果 Na-SP の硫酸基を完
全に除去した場合,抗 HSV-1 活性が消失した.そ
こで,Na-SP を部分的に脱硫酸化した場合と,硫
酸基が結合していない水酸基を新たに硫酸化して多
糖分子内の硫酸基の割合を増やした場合の抗
HSV-1 活性への影響を調べた.結果を Table 9 に示
したが,硫酸基の含量が 8.6%まで減少するにつれ
て抗 HSV-1 活性も低下し,さらに減少させると活
性は消失した.33)一方,Na-SP を過硫酸化して得ら
れた OS-SP の場合,細胞毒性が強くなったが,濃
度依存的ではなく頭打ち傾向であった.また,Na-
SP の場合,ウイルス感染と同時に試料を添加した
ときの子孫ウイルスの増殖に対する阻害活性は,ウ
イルス感染直後に添加したときに比べてかなり強か
ったが,OS-SPs の場合,試料の添加時期の違いに
よる顕著な差は認められなかった.34)そこで,OS-

hon p.12 [100%]
72
Table 10. Time-of-addition Experiments of Na-SP and OS-Ps
TreatmentNa-SP OS-SP-2 OS-SP-5
IC50a) SI b) IC50 SI IC50 SI
During infection >200 >200 >200During infection and for 24 h thereafter 0.57 12456 0.87 2874 0.90 3667
024 h postinfection 2.9 2448 0.92 2717 1.3 2538
224 h postinfection 13 546 11 227 8.7 379
424 h postinfection 25 284 14 179 15 220624 h postinfection 50 142 27 93 32 103
824 h postinfection 82 87 38 66 37 89
a) The 50% inhibitory concentration (mg/ml). b) SI: Selectivity index.
72 Vol. 128 (2008)
SP-2 と OS-SP-5 について time-of-addition 実験を
行い,Na-SP の場合と比較した.これらの試料を
ウイルス感染中及びそれ以後も培地中に添加した場
合の IC50 値は OS-SPs と Na-SP の間に差はない
が,感染後に添加した場合は,OS-SPs の IC50 値は
Na-SP に比べて多少小さくなった(Table 10).特
に注目すべき点は,感染 8 時間後に試料を添加した
場合でも OS-SPs にウイルスの増殖阻害効果が認め
られたことである.これらの実験結果は,OS-SPs
が感染細胞への吸着・侵入後に起こる子孫ウイルス
の複製段階に対してより強く作用することを示唆し
ているが,その詳細な作用機序については不明であ
る.
3-2. メカブ由来新規抗ウイルス活性分子に関す
る研究
3-2-1. メカブ由来抗ウイルス活性糖鎖分子の分
離と構造 メカブは褐藻コンブ目チガイソ科に属
するワカメ Undaria pinnatiˆda の葉状体の下方部
分の胞子葉のことで,ヌメリのある食材として利用
されている.本藻体の乾燥破砕物を EtOH で処理
して脱脂後,残渣を 0.15N HCl で抽出した.抽出
液を中和後,4 倍量の EtOH を加えて得られた沈澱
物について,陰イオン交換樹脂(Toyopearl DEAE
650M, Q-Sepharose FF)やゲルろ過(Sephacryl
S-300 HR)を組み合わせて抗 HSV-1 活性画分を分
離・精製したところ,見掛けの分子量が 9000 の多
糖体が得られた.本多糖体には,fucose と glactose
が 1:1.1 の割合で含まれるほか,uronic acid が
1.9%,硫黄が 10.4%含まれていたことから,本多
糖体は fucoidan (FU)であると推察された.また,
硫酸化度から,10 糖残基当たり 7 個の硫酸基が結
合していると算出された.さらに,FU 及びその脱
硫酸化体のメチル化分析において極めて多様な結合
様式が観察されたことから,FU は,複雑な枝分れ
構造を持ち,硫酸基は fucose 残基の 2 位及び
galactose 残基の 3 位又は 6 位に結合していると推
測された(Table 11).35)
3-2-2. Mekabu fucoidan の 抗 ウ イ ル ス 活 性
FU について,抗ウイルス活性を調べたところ,
エンベロープを有する HSV-1, HSV-2, human
cytomegalovirus (HCMV)及び in‰uenza A virus
(IFV)に対しては顕著な増殖阻害活性を示したが,
エンベロープを持たない poliovirus や coxsackie vi-
rus に対しては無効であった.また,ウイルスの感
染と同時に FU を添加した場合の方が感染直後に添
加した場合より大きい選択指数を示したことから,
本糖鎖分子はウイルスの宿主細胞への吸着・侵入段
階を阻害すると推察された.
3-2-3. Mekabu fucoidan の A 型インフルエンザ
ウイルスに対する作用 最近,高病原性鳥インフ
ルエンザが世界各地で発生し,重大な社会問題とな
り,近い将来における新型インフルエンザの世界的
流行が懸念されている.わが国においても,厚生労
働省が IFV 治療薬である oseltamivir の備蓄を関連
諸機関に要請した.本薬剤は,インフルエンザウイ
ルスの増殖に関与している neuraminidase を特異的
に阻害する化学合成薬であるが,頻用による副作用
や耐性ウイルスの出現などの新たな問題が発生して
いる.36)特に,oseltamivir 服用者に観察された異常
行動との関連性を廻って種々の議論がなされた結
果,慎重投与が添付文書に記載されるようになっ
た.そこで,FU の抗 IFV 活性を oseltamivir のそ

hon p.13 [100%]
73
Table 11. Results of Methylation Analysis of Mekabu Fucoidan and Its Desulfated Derivative
Identiˆcation Deducedlinkage
Primary massfragments (m/z)
Fucoidan(mol%)
Desulfatedfucoidan (mol%)
2,3,4-Me3-Fuc a) t-Fuc 117, 131, 161, 175 7.8 11.6
2,4-Me2-Fuc 3-Fuc 117, 131, 233, 247 9.1 24.7
2,3-Me2-Fuc 4-Fuc 117, 161, 203, 247 4.2 7.8
4-Me-Fuc 2,3-Fuc 131, 261 11.3 5.53-Me-Fuc 2,4-Fuc 189, 203 4.9 2.4
2,3,4,6-Me4-Gal t-Gal 45, 117, 161, 205 6.0 12.8
2,4,6-Me3-Gal 3-Gal 45, 117, 161, 233, 277 7.3 6.8
2,3,6-Me3-Gal 4-Gal 45, 117, 161, 233, 277 6.3 10.12,3,4-Me3-Gal 6-Gal 117, 161, 189, 233 14.3 15.0
2,6-Me2-Gal 3,4-Gal 45, 117, 305 4.6
2,4-Me2-Gal 3,6-Gal 117, 189, 233, 305 15.4 1.0
2,3-Me2-Gal 4,6-Gal 117, 11, 261, 305 8.8 2.4
a) 2,3,4-Me3-Fuc means 1,5-di-O-acetyl-2,3,4-tri-O-methylfucitol.
Fig. 8. Time-of-addition Experiments Comparing the Antiviral Activity of FU to That of OseltamivirM-FC (100 mg/ml) or oseltamivir (5 mM) was added to cells infected with IFV at the indicated time points. Data are presented as the mean of duplicate assays.
73No. 1
れと比較するとともに,oseltamivir との併用効果
も調べた.37)
まず,FU について,time-of-addition 実験を行
い,oseltamivir と比較した.Figure 8 に示したよう
に,FU と oseltamivir のそれぞれをウイルス感染と
同時に添加後,24 時間培養して,子孫ウイルス量
を測定したところ,両者とも顕著な抑制効果を示し
た.また,感染中にのみ試料が存在するようにした
場合は,FU 投与群は対照群の 40%までウイルス産
生を抑えたが,それ以外の時間帯に添加した場合の
FU の効果は,oseltamivir と比較して極めて弱いも
のであった.
3-2-4. Mekabu fucoidan の免疫系に及ぼす作用
生体にウイルスが感染すると自然免疫系と特異免
疫系が応答する.自然免疫系を担っているマクロフ
ァージは,感染したウイルスを排除するための貪食
能を保有している.そこで,マクロファージの貪食
能について,latex beads の取り込み量を測定して
評価したところ,FU は亢進作用を示すことが認め
られた.また,natural killer (NK)細胞も自然免
疫系を担う細胞の一種であるが,加齢,ストレス,
外科手術,放射線療法や化学療法などにより惹起さ

hon p.14 [100%]
74
Fig. 9. Synergistic EŠect of FU and Oseltamivir on IFVReplication In vitro
FU(250 mg/ml) was combined with oseltamivir (0.5200 nM) againstIFV. The drug interaction was evaluated by isobologram.
74 Vol. 128 (2008)
れる免疫力の低下状態からの回復に寄与している.
抗がん剤投与により免疫力が低下することが知られ
ているので 5-‰uorouracil を用いて実験を行った.
すなわち,BALB/c マウスを 5-‰uorouracil で処理
すると,NK 活性は低下したが,このときに FU を
経口投与すると,NK 活性は正常値にまで回復し
た.ところが,NK 活性が正常に保たれている 5-
‰uorouracil 非処理群では,FU 投与による NK 活性
の上昇はみられなかった.さらに,BALAB/c マウ
スの脾臓から採取した B 細胞を FU の存在下で培
養し,抗体産生細胞への形質転換(blast formation)
に及ぼす効果を調べたところ,有意の促進効果が認
められた.
3-2-5. Mekabu fucoidan と oseltamiviv との併用
効果 FU は明らかに oseltamivir と異なる作用機
序を持つことが判明したので,両者の併用効果を検
討した.最初に,培養細胞を用いる評価系において,
FU (250 mg/ml)と oseltamivir (0.5200 nM)の種
々の濃度の組み合わせによる抗 IFV 活性の変動を
調べた.Figure 9 はその結果を示したものであり,
両者は明らかに相乗作用を示すことが分かった.な
お,いずれの組み合わせの場合にも細胞毒性の上昇
は認められなかった.
そこで,次に,in vivo での効果を検討した.す
なわち,1×105 PFU の IFV (A/NWS/33)を含む
ウイルス液を BALB/c マウスの鼻腔内に接種し,
FU (5 mg)と oseltamivir (0.1 mg)をそれぞれ単
独又は両者を併用して,1 日 2 回,7 日間経口投与
した.Figure 10 に示したように,FU 単独投与の
場合の肺のウイルス量には対照群と比べて顕著な減
少は認められなかったが,oseltamivir 単独投与群
及び併用投与群には顕著なウイルス量の減少が認め
られた.また,併用投与群の場合,oseltamivir 単
独投与群と比較した場合でも,肺のウイルス量に有
意な減少が観察された.一方,気管支洗浄液中の中
和抗体価を調べたところ,FU 単独投与群では,対
照群と比べて有意な中和抗体価の上昇が認められ
た.これに対し,oseltamivir 投与群での 14 日目と
21 日目の中和抗体価は,対照群に比べて低い値で
あった(Fig. 11).
IFV は,気道粘膜で増殖することから,気道での
分泌型 IgA が感染防御に大きな役割を果すとされ
ている.そこで,IFV 感染後の気管支洗浄液中のウ
イルス特異的 IgA 抗体量を ELISA で分析した.
Figure 12 に示したように,FU 投与群では感染 21
日後の IgA 量は有意に増加したが,対照群,osel-
tamivir 投与群,及び oseltamivir と FU 併用投与群
の IgA 量には相互に有意差は認められなかった.
以上のように,生体内でのウイルス増殖に及ぼす
FU は,抑制作用の点では oseltamivir に比べてかな
り弱いが,中和抗体産生能増強作用の点では極めて
強いといえる.したがって,両者のそれぞれの特性
を活かすことにより,高病原性ウイルス感染症の問
題にも対処することが可能であると思われる.
3-3. 髪菜 Nostoc ‰agelliforme 由来新規抗ウイル
ス活性分子に関する研究
3-3-1. 髪菜由来新規抗ウイルス活性糖鎖分子の
分離と構造解析 Nostoc ‰agelliforme は,中国
の内陸部やモンゴルの砂漠地域に自生する陸生藻
(藍藻)で,黒色の毛髪に似た形状をしている.本
藻体の乾燥品を短く切断したものを熱水抽出後,80
% EtOH で処理して得られた沈澱部を水に対して
透析した.透析内液について,抗 HSV-1 活性を指
標にしながら,陰イオン交換カラム(Toyopearl
DEAE 650M)やゲルろ過(Sepharose 6B, Toyo-
pearl HW-55F)を組み合わせて含有成分を分離・

hon p.15 [100%]
75
Fig. 10. EŠects of Peroral Treatment with FU (5 mg/day) and Oseltamivir (0.1 mg/day) Alone or in Combinations on Daily VirusTiters in the Lung of Mice Infected with IFV
Treatment of mice (n=3) was twice a day beginning just after virus inoculation. Signiˆcant levels were determined using Student's t-test versus control groups,p<0.05, p<0.01,p<0.001. Signiˆcant levels were determined using Student's t-test versus oseltamivir alone treated groups, ##p<0.01, ###p<0.001.
Fig. 11. EŠects of Peroral Treatment with FU and Oseltamivir Alone or in Combinations on the Production of Neutralizing An-tibody in the Mucosa of Mice Infected with IFV
Treatment was done as described in Fig. 10. p<0.05: signiˆcant increase as compared with control groups, #p<0.05: signiˆcant reduction as compared withcontrol groups.
75No. 1
精製したところ,抗 HSV-1 活性分子として,分子
量的にも荷電的にも均一な酸性多糖体(見掛けの分
子量 2.11×105, nosto‰an と命名)が単離された.
本多糖体を完全加水分解して構成単糖を分析したと
ころ,D-glucose, D-galactose, D-xylose, D-mannose
がそれぞれ 42.8, 20.7, 29.9, 6.6 mol%の割合で検出
された.また,これらとは別に 13.3%の uronic
acid も含まれていることが明らかになったが,
nosto‰an を NaBH4 で還元したところ,D-glucose
の含量が顕著に増加したことから,この uronic
acid は D-glucuronic acid であることが判明した.
一方,構成単糖の結合様式を明らかにするために

hon p.16 [100%]
76
Fig 12. EŠects of Peroral Treatment with FU and Oseltamivir Alone or in Combinations on the Production of IFV-speciˆc IgA in theMucosa of Mice Infected with IFV
Treatment was done as described in Fig. 10. p<0.05: signiˆcant increase as compared with control groups.
Table 12. Results of Methylation Analysis of Nosto‰an and Its Reduced Product
Identiˆcation Deducedlinkage
Primary massfragments (m/z)
Nosto‰an(mol%)
Reducedproduct (mol%)
2,3,4,6-Me4-Glc a) t-Glc 45, 118, 205, 206 6.4 13.5
2,3,4,6-Me4-Gal t-Gal 45, 118, 205, 206 2.0 1.3
2,3,4,6-Me4-Man t-Man 45, 118, 205, 206 4.3 2.92,3,6-Me3-Glc 4-Glc 45, 118, 162, 233, 277 18.2 17.7
2,3,6-Me3-Gal 4-Gal 45, 118, 162, 233, 277 22.2 16.7
2,3-Me2-Xyl 4-Xyl 118, 162, 189, 233 16.8 19.4
2,6-Me2-Gal 3,4-Gal 45, 118, 305 4.3 3.52-Me-Xyl 3,4-Xyl 118, 261 5.8 4.9
2,4-Me2-Glc 3,6-Glc 118, 189, 234, 305 2.3 2.2
2,3-Me2-Glc 4,6-Glc 118, 162, 261, 305 17.7 18.0
a) 2,3,4,6-Me4-Glc means 1,5-di-O-acetyl-2,3,4,6-tetra-O-glucitol.
76 Vol. 128 (2008)
nosto‰an と還元体のメチル化分析を行った.Table
12 にその結果を示したが,1,4-linked glucose, 1,4-
linked galactose, 1,4-linked xylose 及び 1,4,6-linked
glucose が約 1:1:1:1 の割合で存在し,少量の
D-mannose は,非還元末端位に結合していること
が示唆された.また,uronic acid 還元化物のメチ
ル化分析の結果を nosto‰an の場合と比較すると,
glucuronic acid は非還元末端位に結合していること
が示唆された.さらに,nosto‰an の部分加水分解
により得られたオリゴ糖の各種 NMR スペクトルを
解析した結果,b-D-Glcp-(1→4)-D-Xylp 及び b-D-
GlcAp-(1→6)-b-D-Glcp-(1→4)-D-Galp のような糖
鎖配列の存在が示唆された.なお,従来報告された
藻類由来酸性多糖の多くは硫酸化多糖であったが,
本多糖体は分子内に硫酸基やタンパク質を含有して
いないことが分かった.
3-3-2. Nosto‰an の抗ウイルス活性 Nosto‰an
は強い抗 HSV-1 活性を示したので,他のウイルス
に対する増殖阻害作用をも調べた結果,nosto‰an
は Na-SP や FU と同様,エンベロープを有するウ
イルスに対してのみ増殖阻害活性を示した.また,
time-of-addition 実験において,nosto‰an をウイル
ス感染と同時に添加した場合にのみ抗 HSV-1 活性
が認められたことから,ウイルスの宿主細胞への吸

hon p.17 [100%]
77
Fig. 13. EŠect of Combination of Nosto‰an and Oseltamiviron Virus Production in the Lung of Mice
Virus titers in the lung were estimated at day 3 postinfection. Signiˆcantlevels were determined using Student's t-test versus control groups, p<0.05, p<0.005, p<0.001. Signiˆcant levels were determined usingStudent's t-test versus oseltamivir-treated groups, #p<0.05, ##p<0.005,###p<0.001.
77No. 1
着段階及び侵入段階に対する作用をそれぞれ調べた
ところ,本物質は吸着段階を選択的に阻害すること
が分かった.このような結果は硫酸化多糖の場合と
著しく異なる点である.なお,IFV に対して同様の
実験を行い,HSV-1 の場合と同じ結果を得た.す
なわち,nosto‰an はエンベロープを有するウイル
スが宿主細胞に吸着する段階を選択的に阻害するこ
とが明らかになった.38,39)
3-3-3. Nosto‰an と Oseltamivir との併用効果
FU について検討したように, IFV に対する
nosto‰an と oseltamivir との併用効果を評価したと
ころ,in vitro 及び in vivo のいずれの場合にも両者
の間に相乗作用が確認された.一方,BALB/c マウ
スを用いて,IFV を経鼻感染させ,nosto‰an (1 回
投与量 0.2 mg 又は 0.5 mg)と oseltamivir (1 日投
与量 0.02 mg, 0.05 mg, 0.2 mg)を種々の用量で組
み合わせて投与した.感染 3 日後の肺のウイルス量
を調べたところ,Fig. 13 から分かるように,osel-
tamivir を 0.02 mg/day/mouse 単独投与した場合に
は対照群との有意差は認められなかったが,同用量
の oseltamivir と nosto‰an (0.2 mg)を併用した場
合,肺のウイルス量は,対照群と比較して有意に減
少した.Nosto‰an の投与量を 0.5 mg にした場合
は,さらに顕著なウイルスの増殖抑制効果が認めら
れた.以上の結果から,単独投与で無効量の osel-
tamivir であっても,nosto‰an と併用すれば IFV の
増殖を抑制できることが分かった.このことは,両
者を併用すれば,oseltamivir の低用量での使用が
可能になり,結果的に耐性ウイルスの出現や副作用
などのリスクを回避できる可能性があることを示唆
している.
3-4. 食用藻類由来新規糖鎖分子の有用性と今後
の課題 本稿で紹介した 3 種の食用藻類は通常の
陸上植物と異なる環境下に生育しているが,これら
の素材から抽出・分離された新規抗ウイルス活性分
子は,いずれもユニークな化学構造を持つ酸性多糖
であった.従来,報告された多くの硫酸化多糖体の
作用標的はウイルスの宿主細胞への吸着・侵入段階
であり,糖鎖に結合している硫酸基の陰イオンが作
用発現に重要な役割を果たしているとの説明がなさ
れていた.24,25)
しかしながら,今回評価した多糖分子の抗ウイル
ス活性の強弱は,硫酸基の含量のみでは説明でき
ず,分子の長さや硫酸基の対イオン(金属イオン)
の種類によって著しく影響を受けたことから,ウイ
ルスが宿主細胞表面の受容体に吸着する際の多糖分
子の三次元構造を含めた形状及び大きさが重要であ
ることが示唆された.また,IFV 感染動物を用いた
FU と nosto‰an の有効性評価の結果から,糖鎖分
子は既存の抗ウイルス薬と併用することにより,既
に問題となっている合成薬の副作用の発生や耐性ウ
イルスの出現を阻止できる可能性が期待される.
一方,最近,生体の細胞表面に存在している糖鎖
が種々の細胞の増殖,炎症,がん転移,細菌・ウイ
ルスの感染,免疫,受精などにおけるシグナル伝達
に係わっていることが判明してきた.他方,天然物
由来の多糖など外来性の糖鎖分子は,その構造の違
いに応じて細胞機能を調節し,多様な生物活性を発
現することが報告されている.そこで,スピルリナ
由来硫酸化多糖(Na-SP や Ca-SP)について,種
々の生物活性を調べたところ,抗ウイルス活性のほ
かに,がん転移抑制作用,40)血液凝固抑制作用,41)
動脈硬化抑制作用42)などの生物活性が認められた.
これらの生物活性の作用機序については,多糖分子
の立体構造など詳細な糖鎖構造が未解明であるの
で,なお不明な点が多く残されているが,近年進歩

hon p.18 [100%]
7878 Vol. 128 (2008)
が著しいプロテオミクス,メタボロミクスやニュー
トリゲノミクスなどを駆使することにより,外来性
糖鎖分子の機能の解明と応用が可能になると思われ
る.
謝辞 本研究は,本学大学院医学薬学研究部の
林 京子博士及び李 貞範博士のほか,当研究室に
所属した大学院生(川崎 勝博士,兼清健志博士,
Preeprame Srisomporn 博 士 , Xiao-Long Hou 修
士)や多くの学生諸君の協力を得て行われたもので
あり,心より感謝申し上げます.また,パラグァイ
薬草の調査研究の機会を与えて頂きました森田直賢
富山医科薬科大学名誉教授を始め,共同研究の推進
に直接ご協力いただきました浅野真司先生(現在,
立命館大学教授),鍛冶利幸先生(北陸大学教授),
菊地 徹先生(富山医科薬科大学名誉教授),小島
一郎博士,前田昌徹先生(埼玉大学名誉教授),仲
野隆久博士(理研ビタミン),竹中裕行博士(マイ
クロアルジェコーポレーション),竹口紀晃先生
(富山医科薬科大学名誉教授),早川由美子博士,山
本千夏先生(北陸大学准教授),手塚康弘先生(富
山大学和漢医薬学総合研究所准教授),宮原龍郎博
士,中井智恵子博士,米谷芳枝先生(星薬科大学教
授)及び豊岡尚樹先生(富山大学准教授)に厚くお
礼申し上げます.
REFERENCES
1) Pneumocystis pneumonia-Los Angeles, NMWRMorb. Mortal. Wkly. Rep., 30, 250252 (1981).
2) Barre-Sineoussi F., Chermann J. C., Rey F.,Nugeyre M. T., Chamaret S., Gruest J., Dau-guet C., Axler-Blin C., Vezinet-Brun F., Rou-zioux C., Rozenbaum W., Montagnier L.,Science, 220, 868871 (1983).
3) Silberner J., Sci. News, 127, 148 (1985).4) Ezzell C., Nature, 329, 751 (1987).5) Lalezari J. P., Henry K., O'Hearn M., Mon-
taner J. S., Piliero P. J., Trottier B.,Walmsley S., Cohen C., Kuritzkes D. R., EronJr. J. J., Chung J., DeMasi R., Donatacci L.,Drobnes C., Delehanty J., Salgo M., N. Eng.J. Med., 348, 21752185 (2003).
6) Hammer S. M., Squires K. E., Hughes M. D.,Grimes J. M., Demeter L. M., Currier J. S.,Eron J. J., Feinberg J. E., Balfour H. H.,
Deyton L. R., Chodakewitz J. A., Fischl M.A., N. Engl. J. Med., 337, 725733 (1997).
7) Hirsch M., Steigbigel R., Staszwski S., Mel-lors J., Scerpella E., Hirschel B., Lange J.,Squires K., Rawlins S., Meibohm A., LeavittR., J. Infect. Dis., 180, 659665 (1999).
8) Hayashi T., Kishi M., Kawasaki M., ArisawaM., Shimizu M., Suzuki S., Yoshizaki M.,Morita N., Tezuka Y., Kikuchi T., BerganzaL. H., Ferro E., Basualdo I., TetrahedronLett., 28, 36933696 (1987).
9) Hayashi T., Kawasaki M., Miwa Y., Taga T.,Morita N., Chem. Pharm. Bull., 38, 945947(1990).
10) Hayashi T., ``Studies in Natural ProductsChemistry, 21, Bioactive Natural Products(Part B),'' ed. by Atta-ur-Raman, Elsevier,Amsterdam, 2000, pp. 689727.
11) Hayashi K., Niwayama S., Hayashi T., NagoR., Ochiai H., Morita N., Antiviral Res., 9,345354 (1988).
12) Elion G. B., Furman P. A., Fyfe J. A., Miran-da P. D., Beauchamp L., SchaŠer H. J., Proc.Natl. Acad. Sci. U.S.A., 74, 57165720(1997).
13) Hayashi K., Hayashi T., Antiviral Chem.Chemother., 7, 7985 (1996).
14) Chen S.-H., Shine H. D., Goodman J. C.,Grossman R. G., Woo S. L. C., Proc. Natl.Acad. Sci. U.S.A., 91, 30543057 (1994).
15) Hayashi K., Lee J.-B., Maitani Y., ToyookaN., Nemoto H., Hayashi T., J. Gene Med., 8,10561067(2006).
16) Hayashi K., Hayashi T., Recnt Res. Devel.Chem. Pharm. Sci., 2, 223236 (2002).
17) Freeman S. M., Abboud C. N., Whartenby K.A., Packman C. H., Koeplin D. S., MooltenF. L., Abraham G. N., Cancer Res., 53, 52745283 (1993).
18) Asano S., Mizutani M., Hayashi T., MoritaN., Takeguchi N., J. Biol. Chem., 265, 2216722273 (1990).
19) Hayashi T., Asano S., Mizutani M.,Takeguchi N., Kojima T., Okamura K., Mori-ta N., J. Nat. Prod., 54, 802809 (1991).
20) Miyahara T., Hayashi T., Matsuda S., Yama-da R., Ikeda K., Tonoyama H., KomiyamaH., Matsumoto M., Nemoto N., Sankawa U.,Bioorg. Med. Chem. Lett., 6, 10371042

hon p.19 [100%]
7979No. 1
(1996).21) Overman K. E., Ricca D. J., Tran V. D., J.
Am. Chem. Soc., 115, 20422044 (1993).22) Overman L. E., Ricca D. J., Tran V. D., J.
Am. Chem. Soc., 119, 1203112040 (1997).23) Ziegler F. E., Wallace O. B., J. Org. Chem.,
60, 36263636 (1995).24) Witvrouw M., DeClerk E., Gen. Pharmacol.,
29, 497511 (1997).25) Damonte E. B., Matulewicz M. C., Cerezo A.
S., Curr. Med. Chem., 11, 23992419 (2004).26) Nakaya N., Honma Y., Goto Y., Nutr. Rep.
Int., 37, 13291337 (1988).27) Schwartz J., Shklar G., Reid S., Trickler D.,
Nutr. Cancer, 11, 127134 (1988).28) Hayashi O., Katoh T., Okuwaki Y., J. Nutr.
Sci., Vitaminol., 40, 431441 (1994).29) Lee J.-B., Hayashi T., Hayashi K., Sankawa
U., Maeda M., Nemoto T., Nakanishi H., J.Nat. Prod., 61, 11011104 (1998).
30) Lee J.-B., Hayashi T., Hayashi K., SankawaU., J. Nat. Prod., 63, 136138 (2000).
31) Lee J.-B., Srisomporn P., Hayashi K., TanakaT., Sankawa U., Hayashi T., Chem. Pharm.Bull., 49, 108110 (2001).
32) Hayashi T., Hayashi K., Maeda M., KojimaI., J. Nat. Prod., 59, 8387 (1996).
33) Hayashi K., Hayashi T., Kojima I., AIDSRes. Hum. Retroviruses, 12, 14631471(1996).
34) Lee J.-B., Hou X., Hayashi K., Hayashi T.,Carbohydr. Poly., 69, 651658 (2007).
35) Lee J.-B., Hayashi K., Hashimoto M., Naka-no T., Hayashi T., Chem. Pharm. Bull., 52,10911094 (2004).
36) Kiso M., Mitamura K., Sakai-Tagawa Y.,Shiraishi K., Kawakami C., Kimura K., Hay-den F. G., Sugaya N., Kawaoka Y., Lancet,364, 759765 (2004).
37) Hayashi T., Hayashi K., Kanekiyo K., OhtaY., Lee J.-B., Hashimoto M., Nakano T.,``Combating the Threat of Pandemic In‰uen-za: Drug Discovery Approaches,'' ed. by Tor-rence P. F., John Wiley & Sons, Inc., NewYork, 2007, pp. 166182.
38) Kanekiyo K., Lee J.-B., Hayashi K., Takena-ka H., Hayashi T., J. Nat. Prod., 68, 10371041 (2005).
39) Kanekiyo K., Hayashi K., Takenaka H., LeeJ.-B., Hayashi T., Biol. Pharm. Bull., 30,15731575 (2007).
40) Mishima T., Murata J., Fujii H., Hayashi T.,Kato T., Saiki I., Clin. Exp. Metastasis, 61,541550 (1998).
41) Hayakawa Y., Hayashi T., Lee J.-B., OzawaT., Sakuragawa N., J. Biol. Chem., 275,1137911382 (2000).
42) Kaji T., Okabe M., Shimada S., YamamotoC., Fujiwara Y., Lee J.-B., Hayashi T., LifeSci., 26, 24312439 (2004).