Embed Size (px)
Citation preview

SUPPRESSION OF INTRACELLULAR RESISTANCE FACTORS BY ADRIAMYCINAUGMENTS HEAT-INDUCED APOPTOSISVIA INTERLEUKIN-1b-CONVERTINGENZYME ACTIVATION IN PANCREATIC CARCINOMA CELLSDaisuke KOBAYASHI1, Naoki WATANABE1*, Hiroyoshi SASAKI2, Tetsuro OKAMOTO2, Naoki TSUJI2, Tsutomu SATO2,Naofumi YAMAUCHI 2 and Yoshiro NIITSU2
1Department of Laboratory Diagnosis, Sapporo Medical University, School of Medicine, Sapporo, Japan2Fourth Department of Internal Medicine, Sapporo Medical University, School of Medicine, Sapporo, Japan
Combination of heat and various anticancer drugs canexert a synergistic antitumor effect in vitro and in vivo, thoughthe mechanism is not clear. We have previously shown thatendogenous tumor necrosis factor (enTNF) acts as an intra-cellular resistance factor to inhibit the cytotoxic effect of heatby scavenging oxygen-free radicals via the induction of manga-nous superoxide dismutase (MnSOD). Consequently, we ex-amined whether the suppression of these resistance factorsby combining anticancer drugs and heat causes an augmenta-tion of heat-induced cytotoxicity. The human pancreaticcarcinoma cell line, PANC-1, constitutively expresses appre-ciable amounts of enTNF and also demonstrates heat resis-tance. After treatment of these cells for 15 hr with adriamy-cin (ADM), the expression of enTNF was decreased by 43%,and MnSOD activity was suppressed by 55%. The cytotoxiceffects of the treatment of PANC-1 cells with ADM followedby heat were greater than the sum of those observed with theagents administrated individually. Heat-induced apoptosiswas also augmented by pretreatment with ADM. Further-more, the interleukin-1b-converting enzyme inhibitor, Ac-YVAD-CMK, reversed the augmented cytotoxicity. Our re-sults indicate that suppression of intracellular resistancefactors such as enTNF and MnSOD plays an important role inapoptosis seen after heat and ADM combined therapy. Int. J.Cancer 76:552–555, 1998.r 1998 Wiley-Liss, Inc.
Hyperthemia is known to posseses a potent antitumor activity(Maedaet al., 1991). However, its antitumor effect is somewhatlimited (Roizin and Pizzo, 1988; Mivechi and Ross, 1990). Anumber of studies have attempted to augment the antitumor effectsby combining hyperthermia with anticancer drugs such as cisplatinand adriamycin (ADM)in vivo (Kakehiet al.,1990; Kusumotoetal., 1995) andin vitro (Hermanet al., 1982). However, little isknown about the mechanism of the augmentation of heat-inducedcytotoxicity by anticancer drugs. Elucidation of the factors regulat-ing heat-induced cytotoxicity may improve the efficacy of hyper-thermia in anticancer therapy.
We have reported previously that intracellular oxygen freeradical (OFR) production is involved in the mechanism of heat-induced cytotoxicity (Yamauchiet al., 1992). We also observedthat enTNF exerts an intracellular protective effect against not onlyexogenous TNF (Okamotoet al.,1992), but also heat (Tsujiet al.,1992; Watanabeet al.,1997) by scavenging OFR via the inductionof manganous superoxide dismutase (MnSOD) (Himenoet al.,1992).
In the present study, we first examined whether ADM, which hasbeen widely used to treat solid tumors, downregulates intracellularresistance factors such as enTNF and MnSOD in heat-resistantPANC-1 (human pancreatic carcinoma) cells (Watanabeet al.,1997). We then went on to define the optimal sequence of heattreatment in combination with anticancer drugs. It has beenreported that apoptosis plays a major role in hyperthermic cytotox-icity (Yonezawaet al.,1996). We demonstrate here that sequentialtreatment with heat and ADM augments heat-induced apoptosis. Inaddition, the effect of the combination of heat and ADM on thefactors regulating the signal transduction of apoptosis such asinterleukin-1b-converting enzyme (ICE)-like protease, p53, andp21/WAF-1 were also examined.
MATERIAL AND METHODS
Cell culture
PANC-1 and MIAPaCa-2 (human pancreatic carcinoma) cellswere purchased from the ATCC (Rockville, MD) and cultured inDulbecco’s modified Eagle’s minimal essential medium (DMEM;GIBCO, Grand Island, NY) supplemented with 10% fetal bovineserum (FBS; Flow, North Ryde, Australia) at 37°C in a humidified5% CO2 incubator.
In vitro assay for cytotoxic effectsCytotoxic activity was assessed by a dye-uptake method as
described previously (Tsujiet al.,1992).
Assay for enTNF expressionThe expression of enTNF was measured by a indirect method
using a fluorescein isothiocyanate (FITC)-labeled second antibody,according to a modification of the method described earlier(Pitzurraet al.,1990). Briefly, cells (13 104/100 µl) were added tothe wells of a 96-well microculture plate (Costar, Cambridge, MA)and incubated at 37°C in 5% CO2 for 18 hr. Cells were treated byADM (0.5 µM) and washed twice with PBS and fixed withparaformaldehyde solution (4% v/v in PBS) for 20 min at roomtemperature. Next, cells were treated with Triton X-100 (0.2% v/vin PBS) to permeabilize the cell membrane. For blocking, 1% goatserum in PBS was added to each tube for 30 min at 37°C. As a firstantibody, appropriately diluted rabbit anti-recombinant humanTNF polyclonal antibody (Asahi, Tokyo, Japan) was added to eachtube. Following 1 hr incubation at room temperature, the cells werewashed with 1% BSA (Seikagaku, Tokyo, Japan)-PBS 3 times, andthen treated with appropriately diluted FITC-labeled goat anti-rabbit IgG (E.Y. Laboratories, San Mateo, CA) as a secondantibody. Following an additional 30 min incubation at roomtemperature, cells were washed 3 times with 1% BSA-PBS.Intensity of cell fluorescence was measured in a Titertek Fluoros-kan II (Skatron, Lier, Norway; excitation 485 nm, emission 538nm). The relative fluorescence intensity (F. I.) was determinedcompared to the fluorescence intensity of non-treated PANC-1cells.
Determination of MnSOD activityMnSOD activity was assayed by the nitroblue tetrazolium
method (Oberley and Spitz, 1985). Protein concentrations weredetermined by the Bio-Rad (Hercules, CA) DC protein assay.MnSOD activity was calculated as units per mg of protein, andexpressed relative MnSOD activity was determined compared tothe activity of non-treated cells.
Grant sponsor: Ministry of Education, Science, and Culture of Japan.
*Correspondence to: Department of Laboratory Diagnosis, SapporoMedical University, School of Medicine, South-1, West-16, Chuo-ku,Sapporo 060, Japan. Fax: (81)11-622-7502. E-mail: [email protected]
Received 6 October 1997; Revised 19 December 1997
Int. J. Cancer:76,552–555 (1998)
r 1998 Wiley-Liss, Inc.
Publication of the International Union Against CancerPublication de l’Union Internationale Contre le Cancer
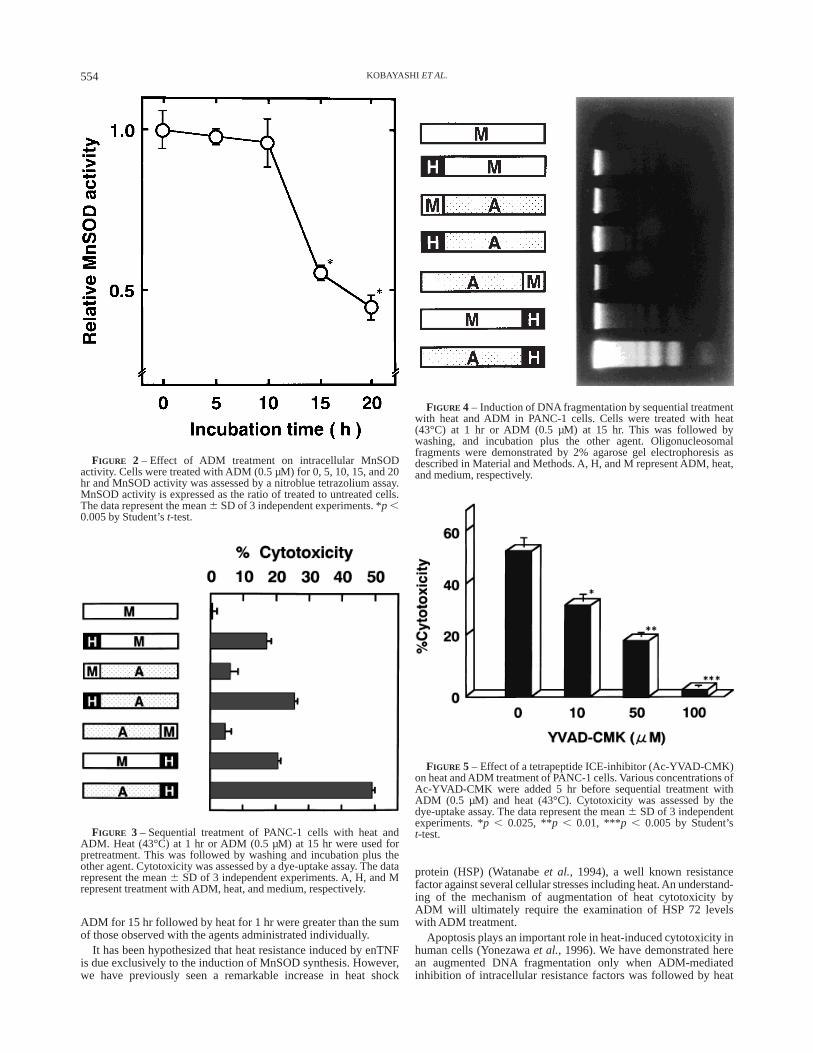
Agarose gel electrophoresis of DNALow m.w. DNA was extracted and agarose gel electrophoresis
was performed. Briefly, 13 106 cells were added to each well of a100 mm culture dish (Costar) and heated or treated with ADM.Following test incubations, cells were washed twice with PBS andlysed with 100 µl of lysis buffer (10mM Tris-HCl (pH7.4), 10 mMEDTA, 0.5% Triton X-100). The lysate was centrifuged at 12,000gfor 20 min, and the supernatant was collected in an Eppendorf tube.Next, the supernatant was incubated with 2 µl of proteinase K(Sigma, St. Louis, MO) at 37°C for 1 hr, and incubation proceededfurther at 37°C for 1 hr after 2 µl of RNase A (20 mg/ml) had beenadded. After overnight precipitation with 0.5 M NaCl and 50%isopropanol at220°C, the pellets were centrifuged at 16,000g for15 min at 4°C and resuspended in 40 µl H2O. Each DNA sample(10 µg) was electrophoresed at 50 V for 1 hr on a 2% agarose gelimpregnated with ethidium bromide, and DNA fragments werevisualized under UV light.
RESULTS
Heat sensitivity of PANC-1 and MIAPaCa-2 cells
The heat sensitivity of 2 lines derived from human pancreaticcarcinoma cells was assessed by incubating these cells at 43°C for1, 2, and 3 hr, and then at 37°C for a total incubation time of 48 hr.PANC-1 cells constitutively expressing appreciable amounts ofenTNF (Watanabeet al., 1996) were more resistant to heattreatment than MIAPaCa-2 cells (data not shown).
Effect of ADM treatment on enTNF expression and MnSODactivity in PANC-1 cells
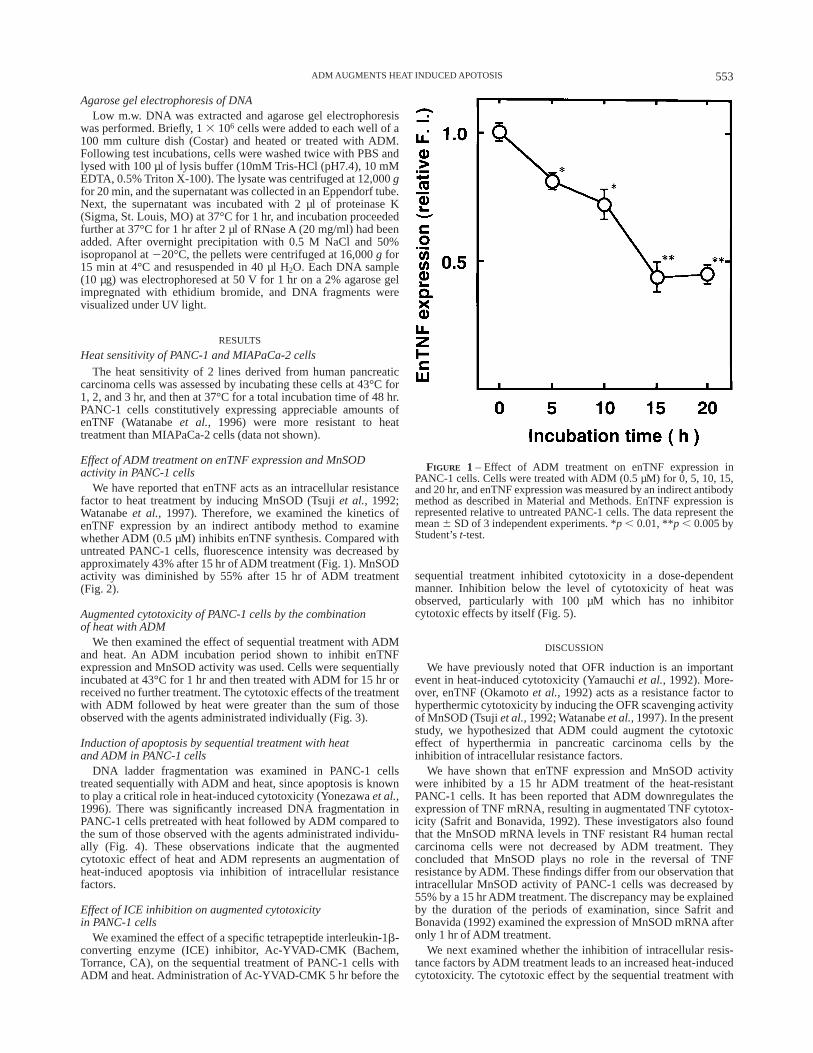
We have reported that enTNF acts as an intracellular resistancefactor to heat treatment by inducing MnSOD (Tsujiet al., 1992;Watanabeet al., 1997). Therefore, we examined the kinetics ofenTNF expression by an indirect antibody method to examinewhether ADM (0.5 µM) inhibits enTNF synthesis. Compared withuntreated PANC-1 cells, fluorescence intensity was decreased byapproximately 43% after 15 hr of ADM treatment (Fig. 1). MnSODactivity was diminished by 55% after 15 hr of ADM treatment(Fig. 2).
Augmented cytotoxicity of PANC-1 cells by the combinationof heat with ADM
We then examined the effect of sequential treatment with ADMand heat. An ADM incubation period shown to inhibit enTNFexpression and MnSOD activity was used. Cells were sequentiallyincubated at 43°C for 1 hr and then treated with ADM for 15 hr orreceived no further treatment. The cytotoxic effects of the treatmentwith ADM followed by heat were greater than the sum of thoseobserved with the agents administrated individually (Fig. 3).
Induction of apoptosis by sequential treatment with heatand ADM in PANC-1 cells
DNA ladder fragmentation was examined in PANC-1 cellstreated sequentially with ADM and heat, since apoptosis is knownto play a critical role in heat-induced cytotoxicity (Yonezawaet al.,1996). There was significantly increased DNA fragmentation inPANC-1 cells pretreated with heat followed by ADM compared tothe sum of those observed with the agents administrated individu-ally (Fig. 4). These observations indicate that the augmentedcytotoxic effect of heat and ADM represents an augmentation ofheat-induced apoptosis via inhibition of intracellular resistancefactors.
Effect of ICE inhibition on augmented cytotoxicityin PANC-1 cells
We examined the effect of a specific tetrapeptide interleukin-1b-converting enzyme (ICE) inhibitor, Ac-YVAD-CMK (Bachem,Torrance, CA), on the sequential treatment of PANC-1 cells withADM and heat. Administration of Ac-YVAD-CMK 5 hr before the
sequential treatment inhibited cytotoxicity in a dose-dependentmanner. Inhibition below the level of cytotoxicity of heat wasobserved, particularly with 100 µM which has no inhibitorcytotoxic effects by itself (Fig. 5).
DISCUSSION
We have previously noted that OFR induction is an importantevent in heat-induced cytotoxicity (Yamauchiet al.,1992). More-over, enTNF (Okamotoet al., 1992) acts as a resistance factor tohyperthermic cytotoxicity by inducing the OFR scavenging activityof MnSOD (Tsujiet al.,1992; Watanabeet al.,1997). In the presentstudy, we hypothesized that ADM could augment the cytotoxiceffect of hyperthermia in pancreatic carcinoma cells by theinhibition of intracellular resistance factors.
We have shown that enTNF expression and MnSOD activitywere inhibited by a 15 hr ADM treatment of the heat-resistantPANC-1 cells. It has been reported that ADM downregulates theexpression of TNF mRNA, resulting in augmentated TNF cytotox-icity (Safrit and Bonavida, 1992). These investigators also foundthat the MnSOD mRNA levels in TNF resistant R4 human rectalcarcinoma cells were not decreased by ADM treatment. Theyconcluded that MnSOD plays no role in the reversal of TNFresistance by ADM. These findings differ from our observation thatintracellular MnSOD activity of PANC-1 cells was decreased by55% by a 15 hr ADM treatment. The discrepancy may be explainedby the duration of the periods of examination, since Safrit andBonavida (1992) examined the expression of MnSOD mRNA afteronly 1 hr of ADM treatment.
We next examined whether the inhibition of intracellular resis-tance factors by ADM treatment leads to an increased heat-inducedcytotoxicity. The cytotoxic effect by the sequential treatment with
FIGURE 1 – Effect of ADM treatment on enTNF expression inPANC-1 cells. Cells were treated with ADM (0.5 µM) for 0, 5, 10, 15,and 20 hr, and enTNF expression was measured by an indirect antibodymethod as described in Material and Methods. EnTNF expression isrepresented relative to untreated PANC-1 cells. The data represent themean6 SD of 3 independent experiments. *p , 0.01, **p , 0.005 byStudent’st-test.
553ADM AUGMENTS HEAT INDUCED APOTOSIS
ADM for 15 hr followed by heat for 1 hr were greater than the sumof those observed with the agents administrated individually.
It has been hypothesized that heat resistance induced by enTNFis due exclusively to the induction of MnSOD synthesis. However,we have previously seen a remarkable increase in heat shock
protein (HSP) (Watanabeet al., 1994), a well known resistancefactor against several cellular stresses including heat.An understand-ing of the mechanism of augmentation of heat cytotoxicity byADM will ultimately require the examination of HSP 72 levelswith ADM treatment.
Apoptosis plays an important role in heat-induced cytotoxicity inhuman cells (Yonezawaet al., 1996). We have demonstrated herean augmented DNA fragmentation only when ADM-mediatedinhibition of intracellular resistance factors was followed by heat
FIGURE 2 – Effect of ADM treatment on intracellular MnSODactivity. Cells were treated with ADM (0.5 µM) for 0, 5, 10, 15, and 20hr and MnSOD activity was assessed by a nitroblue tetrazolium assay.MnSOD activity is expressed as the ratio of treated to untreated cells.The data represent the mean6 SD of 3 independent experiments. *p ,0.005 by Student’st-test.
FIGURE 3 – Sequential treatment of PANC-1 cells with heat andADM. Heat (43°C) at 1 hr or ADM (0.5 µM) at 15 hr were used forpretreatment. This was followed by washing and incubation plus theother agent. Cytotoxicity was assessed by a dye-uptake assay. The datarepresent the mean6 SD of 3 independent experiments. A, H, and Mrepresent treatment with ADM, heat, and medium, respectively.
FIGURE 4 – Induction of DNA fragmentation by sequential treatmentwith heat and ADM in PANC-1 cells. Cells were treated with heat(43°C) at 1 hr or ADM (0.5 µM) at 15 hr. This was followed bywashing, and incubation plus the other agent. Oligonucleosomalfragments were demonstrated by 2% agarose gel electrophoresis asdescribed in Material and Methods. A, H, and M represent ADM, heat,and medium, respectively.
FIGURE 5 – Effect of a tetrapeptide ICE-inhibitor (Ac-YVAD-CMK)on heat and ADM treatment of PANC-1 cells. Various concentrations ofAc-YVAD-CMK were added 5 hr before sequential treatment withADM (0.5 µM) and heat (43°C). Cytotoxicity was assessed by thedye-uptake assay. The data represent the mean6 SD of 3 independentexperiments. *p , 0.025, **p , 0.01, ***p , 0.005 by Student’st-test.
554 KOBAYASHI ET AL.

treatment. The augmentation of heat cytotoxicity by ADM is thusreflected by enhanced apoptotic cell death. There is only one singlestudy that has examined factors regulating apoptosis in heat-treatedcells (Ohnishiet al., 1996). These investigators observed thatp53-dependent p21/WAF-1 accumulation was induced by heat inglioblastoma cell lines carrying the wild typep53gene, but not inthose with a mutantp53gene. The PANC-1 cells used in our studyhave a mutantp53and demonstrated no p21/WAF-1 induction afterheat treatment (data not shown). PANC-1 cells may thus use otherpathways to mediate heat-induced apoptosis.
ICE-like proteases are known to be an important factor in thesignal transduction that underlies apoptosis caused by severalcauses of cellular stress, including anticancer drugs (Mashimaetal., 1995). As described above, OFR production has an importantrole in heat-induced cell death. The activation of ICE-like proteasesis directly involved in OFR production (Shimizuet al.,1996),i.e.,activation of ICE-like proteases, CPP32/Yama-like proteases, OFRproduction, and apoptosis occur during reoxygenation in primarycultured hepatocytes. Furthermore, activation of these proteasesand apoptosis is inhibited by treatment with ICE and CPP32inhibitors, as well as by the antioxidantN-acetylcysteine. Activa-
tion of ICE-like proteases thus appears to play an important role inreoxygenation-induced apoptosis via the induction of OFR.
However, the role of ICE-like proteases in heat-induced celldeath remains unclear. Therefore, we examined whether ADM-augmented heat cytotoxicity involved ICE-related signal transduc-tion. Near complete inhibition of the augmented cytotoxicity wasobserved by the use of the ICE-inhibitor, Ac-YVAD-CMK. Thisresult suggests that the heat-induced death signal of PANC-1 cellsis mainly transducted through ICE-like protease activation, ratherthan by cell cycle-related proteins such as p53 and p21/WAF-1.Taken together, our results indicate that heat-induced apoptosis viaICE-like proteases is augmented by ADM, and that the inhibition ofintracellular resistance factors such as enTNF and MnSOD is animportant underlying mechanism.
ACKNOWLEDGEMENTS
This work was supported by the Grants-in-Aid for CancerResearch from the Ministry of Education, Science, and Culture ofJapan.
REFERENCES
HERMAN, T.S., SWEETS, C.C., WHITE, D.M. and GERNER, E.W., Effect ofheating on due to hyperthermia and selected chemotherapeutic drugs.J. nat.Cancer Inst.,68,487–491 (1982).HIMENO, T., WATANABE, N., YAMAUCHI , N., MAEDA, M., OKAMOTO, T.,TSUJI, N., TSUJI, Y., AKIYAMA , S., SASAKI, H. and NIITSU, Y., Induction ofsynthesis of manganous superoxide dismutase in L-M(pNTnF) cellscarrying an inducible TNF gene.Int. J. Cancer,50,458–462 (1992).KAKEHI, M., UEDA, K., MUKOJIMA, T., HIRAOKA, M., SETO, O., AKANUMA ,A., and NAKATSUGAWA, S., Multi-institutional clinical studies on hyperther-mia combined with radiotherapy or chemotherapy in advanced cancer ofdeep-seated organs.Int. J. Hyperthermia,6, 719–740 (1990).KUSUMOTO, T., HOLDEN, S.A., ARA, G. and TEICHER, B.A., Hyperthermiaand platinum complexes: time between treatments and synergyin vitro andin vivo. Int. J. Hyperthermia,11,575–586 (1995).MAEDA, M., WATANABE, N., YAMAUCHI , N., TSUJI, Y. and NIITSU, Y.,Successful treatment of a case of hepatocellular carcinoma with tumornecrosis factor and local hyperthermia.Gastroenterol. Jpn.,26, 774–778(1991).MASHIMA, T., NAITO, M., KATAOKA , S., KAWAI , H. and TSURUO, T.,Aspartate-based inhibitor of interleukin-1b-converting enzyme preventsantitumor agent-induced apoptosis in human myeloid leukemia U937 cells.Biochem. biophys. Res. Comm.,209,907–915 (1995).MIVECHI, N.F. and ROSSI, J.J., Use of polymerase chain reaction to detectthe expression of theMr 70,000 heat shock genes in control or heat shockleukemia cells as correlated to their heat response.Cancer Res.,50,2877–2884 (1990).OBERLEY, L.W. and SPITZ, D.R., Nitroblue tetrazolium. In:Handbook ofmethods for oxygen radical research,R. Greenwald (ed.), pp. 217–225,CRC, Boca Raton, FL (1985).OHNISHI, T., WANG, X., OHNISHI, K., MATSUMOTO, H. and TAKAHASHI , A.,p53-dependent induction of WAF-1 by heat treatment in human glioblas-toma cells.J. biol. Chem.,271,14510–14513 (1996).OKAMOTO, T., WATANABE, N., YAMAUCHI , N., TSUJI, Y., ITOH, Y., NEDA, H.and NIITSU, Y., Endogenous tumor necrosis factor exerts its protectivefunction intracellularly against the cytotoxicity of exogenous tumor necro-sis factor.Cancer Res.,52,5278–5281 (1992).PITZURRA, L., BLASI, E., BARTOLI, A., MARCONI, P. and BISTONI, F., A rapidobjective immunofluorescence microassay: Application for detection ofsurface and intracellular antigens.J. Immunol. Methods,135,71–75 (1990).
ROIZIN, T.L. and PIRRO, J.P., Thermotolerance in human cells of normal andneoplastic origin.Int. J. Hyperthermia,4, 665–675 (1988).
SAFRIT, J.T. and BONAVIDA , B., Sensitivity of resistant human tumor celllines to tumor necrosis factor and adriamycin used in combination:correlation between down-regulation of tumor necrosis factor-messengerRNA induction and overcoming.Cancer Res.,52,6630–6637 (1992).
SHIMIZU , S., EGUCHI, Y., KAMIIKE , W., AKAO, Y., KOSAKA, H., HASEGAWA, J.,MATSUDA, H. and TSUJIMOTO, Y., Involvement of ICE family proteases inapoptosis induced by reoxygenation of hypoxic hepatocytes.Amer. J.Physiol.,271,G949–958 (1996).
TSUJI, Y., WATANABE, N., OKAMOTO, T., TSUJI, N., SASAKI, H., AKIYAMA , S.,YAMAUCHI , N. and NIITSU, Y., Endogenous tumor necrosis factor functionsas a resistant factor against hyperthermic cytotoxicity.Cancer Res.,52,6258–6262 (1992).
VENETIANER, A., PIRITY, M. and HEVER-SZABO, A., The function ofheat-shock proteins in stress tolerance.Cell Biol. Int.,18,605–615 (1994).
WATANABE, N., AKIYAMA , S., TSUJI, N., SASAKI, H., YAMAUCHI , N.,OKAMOTO, T., KOBAYASHI, D and NIITSU, Y., Induction of synthesis of heatshock protein 72 in tumor necrosis factor gene-transduced cells.Jpn. J.Cancer Res.,85,997–999 (1994).
WATANABE, N., TSUJI, N., AKIYAMA , S., KOBAYASHI, D., YAMAUCHI , N.,SASAKI, H., SATO, T., OKAMOTO, T. and NIITSU, Y., Endogenous tumornecrosis factor functions as a resistant factor against hyperthermic cytotox-icity in pancreatic carcinoma cells via enhancement of the heat shockelement-binding activity of heat shock factor 1.Chemotherapy,43,406–414 (1997).
WATANABE, N., TSUJI, N., TSUJI, Y., SASAKI, H., OKAMOTO, T., AKIYAMA , S.,KOBAYASHI, D., SATO, T., YAMAUCHI , N. and NIITSU, Y., Endogenous tumornecrosis factor inhibits the cytotoxicity of exogenous tumor necrosis factorand adriamycin in pancreatic carcinoma cells.Pancreas,13, 395–400(1996).
YAMAUCHI , N., WATANABE, N., MAEDA, M., OKAMOTO, T., SASAKI, H., TSUJI,N., TSUJI, Y., UMENO, H., AKIYAMA , S. and NIITSU, Y., Mechanism ofsynergistic cytotoxic effect between tumor necrosis factor and hyperther-mia.Jpn. J. Cancer Res.,83,540–545 (1992).
YONEZAWA, M., OTSUKA, T., MATSUI, N., TSUJI, H., KATO, K.H., MORIYAMA ,A. and KATO, T., Hyperthermia induces apoptosis in malignant fibroushistiocytoma cellsin vitro. Int. J. Cancer,66,347–351 (1996).
555ADM AUGMENTS HEAT INDUCED APOTOSIS





























