Embed Size (px)
DESCRIPTION
celula
Citation preview

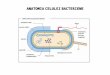
Suprafaţa celulei

Organizarea membranei celulare Definiţie: membrana este o peliculă foarte subţire
(câţiva nm) şi foarte flexibilă ce acoperă şi delimiteazăcompartimentul citoplasmatic, controlează schimburile cu mediul înconjurător şi se comportă ca un sistem de recepţie-transducţie şi constituie substratul adezivităţii celulare.
Membrana prezintă şi diferenţieri speciale cum sunt dispozitivele joncţionale, cilii, microvilii, flagelii etc. Astfel Robertson 1959 emitea teoria membranei „unit” pe baza cercetărilor la microscopul electronic, prin care toate membraneleplasmatice au o organizare ultrastructurală unitară după modelul trilaminat ce implică o foiţă centrală clară flancată de două foiţe dense la fluxul de electroni în grosime totală de 7,5 nm.
Foiţa centrală clară corespunde axului lipidic central hidrofob, iar foiţele dense corespund feţelor hidrofile lipidice pe care se ataşează proteinele.

Dar modelul membranei „unit” s-a dovedit incomplet deoarece ignora dinamismul moleculelor componente şi a proteinelor transmembranare. În 1972 Singer şi Nicolson elaborează teoria mozaicului fluid, astfel moleculele componente – lipide şi proteine – difuzează liber în planul continuumului lipidic şi aceasta implică o organizare întâmplătoare a distribuţiei moleculare lipoproteice.
Modelul actual de membrană celulară este o preluare şi dezvoltare a ideilor principale din modelele precedente, cât şi o concepţie cu mult mai cuprinzătoare faţă de cele precedente. În esenţă continuumul bilipidic, asimetric şi fluid reprezintă axul întregului edificiu molecular membranar, pe cele două feţe hidrofile proteinele sunt distribuite asimetric şi în aranjamente foarte caracteristice, spre deosebire de celelalte teorii ce se referă doar la plasmalemă, conceptul actual arată că membrana celulară este alcătuită din trei părţi componente principale:
- plasmalema – complexul lipoproteic în grosime de 7,5 nm, - glicolema – complexul glicoproteic superficial în grosime de 50
nm, - citoscheletul membranei – reţeaua de proteine (în special
proteine fosforilate) aşezate pe faţa internă a plasmalemei în grosime de câţiva nm.

Bazele moleculare ale organizării membranei celulare
La alcătuirea membranei celulare participă apa cu aprox. 30%, substanţe organice aproximativ 70% şi săruri minerale în cantităţi foarte mici. Peste 90% din masa uscată este reprezentată de lipoproteine, iar glucidele numai în câteva procente.
Totalitatea lipidelor din membrană sunt localizate în plasmalemă unde constituie foiţa continuă hidrofobă pe care se organizează celelalte populaţii de molecule din membrană.
Glucidele sunt prezente exclusiv în glicolemă numai sub formă de fragmente de molecule relativ scurte, fie conjugate cu proteine, fie ataşate la capetele hidrofile a unor lipide din plasmalemă.
Proteinele sunt reprezentate în toate cele trei straturi ale suprafeţei celulare: în plasmalemă în special de o parte şi de alta a continuumului lipidic, ăn glicolemă alcătuind ţesătura pe caru sunt ataşate glucidele, în citoscheletul membranei unde realizează o reţea foarte fină şi suplă.
Apa înbibă glicocalixul, citoscheletul membranei şi foiţele externă şi internă ale plasmalemei, vom găsi şi ioni anorganici care deşi participă cu un procent foarte mic la masa membranei, joacă un rol important în biologia acesteia.

Plasmalema
Fosfolipidele formează marea majoritate a lipidelor din endomembrane şi aprox. 75% din lipidele citoplasmatice (ex.: fosfatidilcolina, sfingomielina, fosfatidiletanolamina etc.).
Fiecare moleculă are aspectul unui „cui” în care capul este hidrofil şi este denumit grup polar pentru că el poartă sarcini electrice, iar reziduurile de acizi graşi sunt hidrofobe şi sunt de obicei desemnate ca segment apolar.
Este partea centrală a membranei celulare şi este prima componentă care a fost identificată la microscopul electronic.
Principalele tipuri de lipide ce intră în alcătuirea membranelor sunt: fosfolipide, colesterol, glicolipide.

Deoarece fosfo-lipidele concentrează la un capăt radicalii hidrofili şi la celălalt radicalii hidrofobi sunt denumite molecule amfipate.
Cullis şi Kruijff (1980) au descoperit că unele lipide cum sunt fosfatidiletanolamina, digli-ceridele şi în special cardiolipina sunt capabile ca în prezenţa Ca²+ să se detaşeze din bistratul lipidic şi să se organizeze în aşa numita fază hexagonală ce realizează canale lungi apoase în interiorul hidrofob.
Proteinele plasmalemei – o mare parte din proteinele membranei celulare sunt localizate la nivelul plasmalemei şi ele se află în relaţii speciale cu bistratul lipidic. Proteinele reprezintă substratul specificităţii suprafeţei celulare; există două grupe de proteine: extrinseci şi intrinseci.
Proteinele extrinseci – sunt distribuite pe o faţă sau pe alta a continuumului lipidic şi contactează pe suprafaţa lor legături hidrofile cu capetele hidrofile ale lipidelor. Ele sunt mai numeroase pe faţa internă unde edifică citoscheletul membranei celulare.
Proteinele intrinseci – se extrag foarte greu din continuumul lipidic deoarece dezvoltă şi legături hidrofobe. Ele servesc ca dispozitiv de recepţie-transducţie (ex: aprox. 15-20% din masa moleculelor de rodopsină este cufundată în regiunea hidrofobă a continuumului lipidic din membrana bastonaşelor retiniene).

Citoscheletul membranei celulare
Este o reţea de proteine extrinseci ce formează partea internă a membranei celulare, cu o grosime de 5-9 nm, care conferă flexibilitate şi rezistenţă suprafeţei celulare.
Un model de aranjare a proteinelor citoscheletale este de reţea, în care ochiurile reţelei sunt alcătuite de dimerii de spectrină, iar nodurile reţelei sunt alcătuite din două tipuri de complexe moleculare.

Glicolema
Este partea externă a membranei celulare cu o grosime medie de 50 nm, este laxă şi greu de identificat în microscopie electronică de rutină, conferă o încărcătură predominant electric negativă suprafeţei celulare şi joacă un rol major în controlul schimburilor ionice transmembranare.
Termeni sinonimi: glicolema, glicocalix, înveliş de suprafaţă „fuzzy coat” – pufoasă, scămoasă.
Din punct de vedere chimic, glicolema este alcătuită indeosebi din glicoproteine, este vorba de o ţesătură delicată şi laxă de lanţuri proteice pe care sunt ancorate reziduurile glucidice (cel mai fecvent este galactoza, iar în cantităţi mici manoza, fructoza, glucoza).

Matricea extracelulară
Matricea extracelulară sau intercelulară constituie mediul interstiţial în care îşi desfăşoară activitatea diferite tipuri de celule; ea este alcătuită din elemente greu identificabile morfologic şi eterogene chimic. Matricea extracelulară controlează adezivitatea, creşterea şi diferenţierea şi joacă rolul principal în organizarea celulelor sub formă de ţesuturi şi în distribuirea acestora în mod specific în diferite organe ale omului.
Matricea extracelulară se află în relaţii de continuitate şi contiguitate moleculară cu glicolema.
La microscopul luminiscent în matricea extracelulară pot fi observate trei tipuri de elemente: membranele bazale, fibre intercelulare şi substanţa fundamentală a matricei bogat coloidală.
Moleculele organice care stau la baza edificării diferitelor structuri microscopice sau ultramicroscopice sunt de natură glicoproteică. Dintre acestea fac parte colagenul şi elastina, glicoproteine sulfatate necolagene şi proteoglicanii.

Colagenul – e o familie de proteine cu foarte mare forţă de extensibilitate ce reprezintă 25% din proteinele omului adult. Au fost identificate în matricea extracelulară 5 izotipuri de molecule de colagen.
Elastina – prezintă deosebiri faţă de molecula de colagen ce îi conferă proprietăţi fizico-mecanice deosebite.
Proteoglicanii – sunt cea de-a II-a componentă importantă a matricei extracelulare, aprox. 95% din molecula de proteoglicani este constituită de glicozaminoglicani şi 5% din proteine. Proteoglicanii datorită componentei polizaharidice formează polianioni foarte mari ce leagă apa şi cationii alcătuind aşa-zisa substanţă fundamentală.
A III-a grupă de molecule organice a matricei extracelulare e reprezentată de glicoproteinele sulfatate necolagene – au greutate moleculară mare şi sunt responsabile de ataşarea celulelor de matricea extracelulară. Au fost identificate fibronectina, laminina, dronectina, entactina, etc. Glicoproteinele necolagene intră în alcătuirea diferitelor tipuri de membrane bazale alături de colagen de tip IV şi glicozaminoglicani.

Receptorii de membrană
Sunt dispozitive moleculare specializate, aşezate pe faţa externă a membranei celulare, cu ajutorul cărora se interceptează semnale sau mesaje venite pe cale nervoasă sau umorală.
Termenul generic de ligand defineşte toate substanţele care, vehiculate pe cale umorală sau nervoasă, se leagă de receptori şi modulează funcţia celulară. Din categoria liganzilor fac parte mesagerii extracelulari sau mesageri de ordinul I (ex: neurotransmiţătorii şi hormonii).

Tipuri de receptori de membrană:- Receptori pentru substanţe endogene – produse de celulele
proprii organismului care pot induce modificări mediate de receptori şi transmise pe cale nervoasă sau umorală.
☺ pentru cale nervoasă – mai cunoscuţi sunt receptorii pentru acetilcolină, noradrenalină, serotonină, histamină, dopamină, epinefrină, acid gamaaminobutiric (GABA).
☺ pentru calea umorală – receptorii hormonii – cei mai bine caracterizaţi sunt cei pentru insulină (izolate în celulele adipoase şi hepatocite).
Receptorii pentru antigenele endogene – se găsesc la suprafaţa celulei implicate în răspunsul imun, fie la suprafaţa celulei neimunogene.
- Receptorii pentru substanţele exogene: - receptori pentru viruşi, - receptori pentru antigene nonself, - receptori pentru toxine microbiene, - receptori pentru droguri, - receptori pentru lecitine etc.

Joncţiunile intercelulare
Sunt dispozitive de legătură intercelulare discrete cu rol de adezivitate intercelulară, de control al scurgerii moleculelor prin spaţiul dintre celule, dar şi cu rol în împărţirea suprafeţei celulare în sectoare separate.
Joncţiunile celulare pot fi sistematizate în două grupe: 1) joncţiuni simple, sunt de trei tipuri: - spaţiile intercelulare, - legăturile intercelulare denticulare, - legăturile intercelulare digitiforme. Membranele a două celule ce alcătuiesc joncţiuni
simple se află la distanţă de cel puţin 30 nm, iar prin spaţiile delimitate de astfel de joncţiuni se pot scurge liber toate tipurile de molecule din mediul interstiţial.

Joncţiunile intercelulare
2) joncţiunile speciale, includ 5 tipuri:
- joncţiuni strânse,
- desmozomii,
- joncţiuni GAP,
- complexele joncţionale,
- legăturile celulei cu membrana bazală. Grupul de complexe joncţionale speciale cum
sunt sinapsele şi discurile intercalare fac obiectul studiului ţesuturilor în cadrul disciplinei de histologie.

Joncţiunile strânse Sunt dispozitive de adezivitate cu o dispoziţie „în
panglică”, impermeabile pentru markerii intercelulari, separând net compartimentele tisulare cu compoziţii chimice diferite.
Denumiri sinonime: zonula occludens, joncţiuni pentalaminate sau joncţiuni heptalaminate.
În organizarea joncţiunilor strânse, membranele adiacente se apropie complet una de alta pentru a edifica structuri pentalaminate, obliterând stfel spaţiul intercelular şi împiedicând scurgerea moleculelor printre membrane. La edificarea acestor dispozitive participă şi proteine integrate care traversează membranele adiacente, se dispun în şiruri gemene pentru a organiza dispozitive de sutură „în fermoar”. Proteinele integrate sunt ancorate prin intermediul unor microfilamente de citoscheletul matricei (sunt întâlnite la nivelul tubului proximal renal, vezică urinară, epiteliu intestinal).

Principalele proprietăţi ale joncţiunilor strânse:
- dispozitive de adezivitate intercelulară, flexibile şi sigure,
- bariere fizice şi chimice intercelulare,
- conferă polaritate celulelor angajate în astfel de joncţiuni,
- apar timpuriu între celulele embrionare, de obicei sub formă de macule pentru ca apoi să se transforme în zonule.

Desmozomii Sunt dispozitive de adezivitate intercelulară foarte puternice. Ele
nu sunt implicate în schimburile intercelulare şi nu împiedică deplasarea moleculelor prin spaţiile pe care le delimitează.
Denumiri sinonime: zonula adherens, macula adherens, lamina desmozomica.
Se întâlnesc 3 tipuri: în formă de pată, în formă de bandă şi hemidesmozomi.
Desmozomii în pată sunt alcătuiţi din următoarele elemente componente:
- membranele plasmatice adiacente aşezate paralel la o distanţă de 25-30 nm,
- un material intercelular dens la fluxul de electroni, proteic, de aspect filamentos, bisectat de o densificare centrală bogată în glucide şi calciu,
- densificări în formă de disc pe frontul citoplasmatic,- dispozitive de legătură sau „linkeri” reprezentate de microfilamente
ce se ancorează de scheletul matriceal,- elemente citoscheletale reprezentate fie de tonofilamente, fie de
microfilamente de actină.

Ei sunt prezenţi între diferite tipuri de celule a epiteliului de acoperire.
Desmozomii în bandă sunt prezenţi la nivelul segmentelor transversale ale discurilor intercalare. Ultrastructura asemănătoare cu a desmozomilor în spot, cu următoarele deosebiri:
- spaţiul intercelular este de 15-25 nm şi mai sărac în material dens,
- densificările de pe faţa internă a plasmalemei nu au formă de disc, ci se extind în mod nedefinit.
Hemidesmozomii sunt variante ale desmozomilor în spot ce realizează joncţiuni cu membrana bazală.

Joncţiunile de tip GAP Sunt diferenţieri ale membranelor celulare în formă de
macule cu diametrul de 0,1-10 nm, care realizează transferul direct de mesageri chimici sau fizici de la o celulă la alta.
Termeni sinonimi: nexus sau macule communicans. La organizarea lor participă membrane adiacente care
se apropie la o distanţă de 2-3 nm, spaţiul dintre membrane nu este obliterat ci poate fi străbătut de molecule mici.
Cel de-al II-lea component sunt conexonii – dispozitive proteice complexe ce traversează membranele adiacente în formă de prisme haxagonale cu diametru de 7-8 nm şi dispuse în registru pe suprafaţa maculei joncţionale. Conexonii sunt alcătuiţi din 6 subunităţi proteice în formă de bastonaşe cu un diametru de 2,5 nm şi lungime de 7,5 nm. Subunităţile sunt dispuse în inel pentru a delimita un canal hidrofil cu diametru reglabil între 0,4-2 nm.

Joncţiunile GAP sunt foarte răspândite la diferite tipuri de celule şi reprezintă principala cale de comunicare intercelulară. Caracteristica lor principală este că permit schimburi rapide de molecule între celulele angajate în joncţiune.

Complexele joncţionale
În mod obişnuit celulele epiteliale secretorii sau celulele din epiteliul intestinal şi renal sunt solidare între ele la polul apical printr-o serie de elemente joncţionale numite complexe joncţionale. Ele cuprind joncţiuni strânse la polul apical şi elemente desmozomale spre extremitatea bazală. Adeseori în poziţii intermediare sunt plasate joncţiuni GAP.

Schimburile prin membrane Funcţii:- preia din mediul extracelular o gamă variată de
combustibili metabolici necesari menţinerii vieţii celulei şi activităţilor ei metabolice,
- reglează volumul celular şi menţinerea pH-ului,- generează gradiente ionice care permit
desfăşurarea unor activităţi complexe ca excitabilitatea nervului sau muşchiului.
După mecanismele care intervin în mişcarea moleculelor prin membrane avem: transport pasiv şi transport activ.
Transportul pasiv – are loc fie prin difuziune simplă, fie prin difuziune facilitată.

Transportul pasiv prin difuziune simplă Pentru cele mai multe tipuri de molecule
trecerea prin difuziune simplă se face lent. Pentru moleculele mici neionizate difuziunea simplă prin membranele biologice se face in cazul celor liposolubile prin continuumul lipidic, iar pentru cele hidrosolubile prin mici orificii sau pori hidrofili situaţi în structurile bimoleculare lipidice.
După legile difuziunii de pe o faţă pe alta a membranei se face din regiunea cu concentraţie mai mare spre cea cu concentraţie mai mică.

Transportul pasiv prin difuziune facilitată Este mecanismul prin care unele substanţe greu solubile
în lipide şi cu o masă moleculară relativ mare sunt transportate prin traversul structurilor membranei mult mai rapid decât prin difuziunea simplă (100.000x). Frecvent întâlnită la hepatocite difuziunea facilitată este modalitatea de transport pentru glucoză, aminoacizi, purine, glicerol etc.
Pentru transportul facilitat se presupune că în grosimea membranei există molecule proteice cu rol de cărăuşi care preia substanţele pe o faţă a membranei şi le eliberează pe cealaltă faţă.
În mod obişnuit, ca şi în cazul difuziunii simple, transportul prin difuziune facilitată se efectuează în sensul gradientului de concentraţie şi duce la echilibrarea concentraţiilor pe ambele feţe ale membranei.

Transportul activ
Este modalitatea de transport transmembranar caracteristică celulelor vii. El asigură schimburi foarte rapide prin membrană a unor molecule şi ioni de importanţă vitală pentru celule împotriva gradientelor de concentraţie sau a celor electrochimice, realizând o creştere a concentraţiei de câteva ori pe o faţă a membranei. Întrucât transportul de molecule şi ioni împotriva gradientelor se face cu consum de energie liberă rezultată din metabolism – este metabolic-dependent.

Transportul activ prin pompe ionice. Pentru menţinerea gradientului ionic intracelular trecerea ionilor prin
membrana celulară se face de cele mai multe ori în direcţia termodinamică nefavorabilă, împotriva gradientului de concentraţie sau a celui electrochimic. Transportul ionilor se face cu ajutorul unor protein-enzime din plasmalemă contrar gradientului de concentraţie şi poartă denumirea de pompe ionice.
Pompele ionice pot transporta un singur ion (ex: pompa de Ca²+, pompa de Mg²+) sau concomitent doi ioni (pompa de Na+ şi K+). De asemenea mai există şi sisteme de transport cuplat ce transferă ioni, apă sau alte molecule necesare metabolismului celular.
Cea mai importantă şi mai cunoscută pompă de ioni este pompa de Na-K; astfel este demonstrat că în mediul intracelular există o mare cantitate de K şi mult mai puţin Na contrar mediului extracelular fapt datorat prezenţei la nivelul membranei celulare a unei enzime ce hidrolizează ATP-ul numai în prezenţa Na şi K şi a unei concentraţii favorabile de Mg. Enzima numită Na-K ATP-aza face parte integrantă din pompa de Na-K.
Căile furnizoare de energie necesară pentru transportul activ sunt depemdente de tipul de celule. Astfel, celulele înalt diferenţiate (fibra musculară, neuroni etc.) folosesc în principal căile aerobe producătoare de ATP. În alte tipuri de celule (hematiile) sunt folosite pentru transport molecule de ATP furnizate prin glicoliză – metabolism anaerob.

Transportul în masă
Este procesul prin care celulele preiau sau elimină în mediul extracelular particule de natură diferită, fie ca atare, fie împreună cu cantităţi variabile de lichid. Transportul se face cu ajutorul unor vezicule formate pe seama membranei celulare. În funcţie de direcţia în care se deplasează veziculele deosebim două tipuri de transport în masă: exocitoza şi endocitoza, iar endocitoza se poate face fără lichid interstiţial – fagocitoza sau cu – pinocitoza.

Endocitoza particulelor fără fluid sau fagocitoza Prin fagocitoză (phagein = a mânca) celulele transportă
din mediul extracelular în citoplasmă particule de natură foarte diferită, utilizând regiuni specializate din membrana celulară purtătoare de receptori. Celulele ce realizează un astfel de transport poartă denumirea de fagocite. După mărimea partuculelor pe care fagocitele sunt capabile să le înglobeze distingem microfage, ce înglobează numai particule mici (leucocite granulare) şi macrofage care înglobează în special particule mari (fagocitele mononucleare).
Fagocitele prezintă la suprafaţa celulei receptori cu ajutorul cărora recunosc elementele „self”, proprii organismului, şi „non-self” sau macromolecule străine de organism (antigeni). Receptorii recunosc şi molecule „self-alterat” (celule degenerate, celule maligne, celule îmbătrânite, resturi tisulare etc.).

Etapele procesului de fagocitoză. Primul element este recunoaşterea antigenelor de către receptori cu fixarea primelor la membrana celulară, determinând activarea receptorilor şi agragarea proteinelor contractile de la nivelul citoscheletului cu emiterea de pseudopode. Ele înconjoară strâns particula, astfel apar interacţiuni receptor particulă pe toată suprafaţa ei, „în fermoar”, fixând particula la membrana pseudopodelor.
Pseudopodele care inconjoară particule se unesc şi la locul de unire fuzionează rezultând o veziculă ce cuprinde particula şi se numeşte fagozom.
În citoplasmă, fagozomii se unesc cu lizozomii ce-şi varsă conţinutul de enzime şi digeră particulele introduse în fagozom.
Fagocitoza, întrucât comportă emiterea de pseudopode, se face cu cheltuială de energie; este deci un proces energodependent.

Endocitoza particulelor în fază fluidă sau pinocitoza Este procesul de transport în masă a unei cantităţi variabile de fluid
tisular împreună cu particulele pe care le conţine (pinos = a bea) prin intermediul unor vezicule pinozomale.
Pinocitoza fără receptori este forma de înglobare a substanţelor din mediul extracelular în vezicule formate din învelişul celular fără fixarea prealabilă a lor de suprafaţa celulară prin receptori. Ea reprezintă forma de endocitoză cel mai frecvent întâlnită la celulele organismului.
Procesul are loc în mai multe etape, începând cu contactul particulelor din fluidul tisular cu învelişul celular, sun activate anumite situsuri anionice de suprafaţă, proces ce induce agragarea proteinelor contractile citoscheletale, cu invaginarea membranei şi formarea de cripte adânci. Buzele superioare fuzionează formând vezicule sau vacuole pinozomale denumite pinozomi. După mărime aceştia pot fi micropinozomi sau macropinozomi. Ulterior pinozomii se unesc cu lizozomii şi-şi descarcă conţinutul în aceştia, apoi membranele pinozomale se reintegrează la nivelul membranei citoplasmatice.
Pinocitoza cu receptori are un mecanism asemănător cu fagocitoza. Viteza de transport prin pinocitoza cu receptori este mare şi se apreciază că în 2-5 minute sunt înglobate în citoplasmă molecule diferite din mediul extracelular prin vezicule pinocitotice.



![LISTA OBIECTIVELOR - satu-mare.ro file2 Nr. crt. Denumire Strada Numar postal Amplasament Numar imobil Suprafata imobil [mp] Suprafata SV [mp] Suprafata AF [mp] Suprafata PD [mp] Suprafata](https://img.pdfslide.tips/doc/110x75/5e112fdf6ac32e2f534236df/lista-obiectivelor-satu-marero-nr-crt-denumire-strada-numar-postal-amplasament.jpg)