Embed Size (px)
Citation preview
Tab. 1.3 Funzioni degli osteociti
Funzioni meccaniche ! Percepiscono l’osso obsoleto e le microfratture e dirigono il flusso di osteoclasti verso il sito che necessita di rimodellamento
! Mediano l’adattamento omeostatico dell’osso alle necessità meccaniche
Funzioni regolatorie locali ! Producono fattori che influenzano la generazione di osteoblasti e osteoclasti
! Controllano la mineralizzazione
Funzioni endocrine ! Regolano l’omeostasi minerale
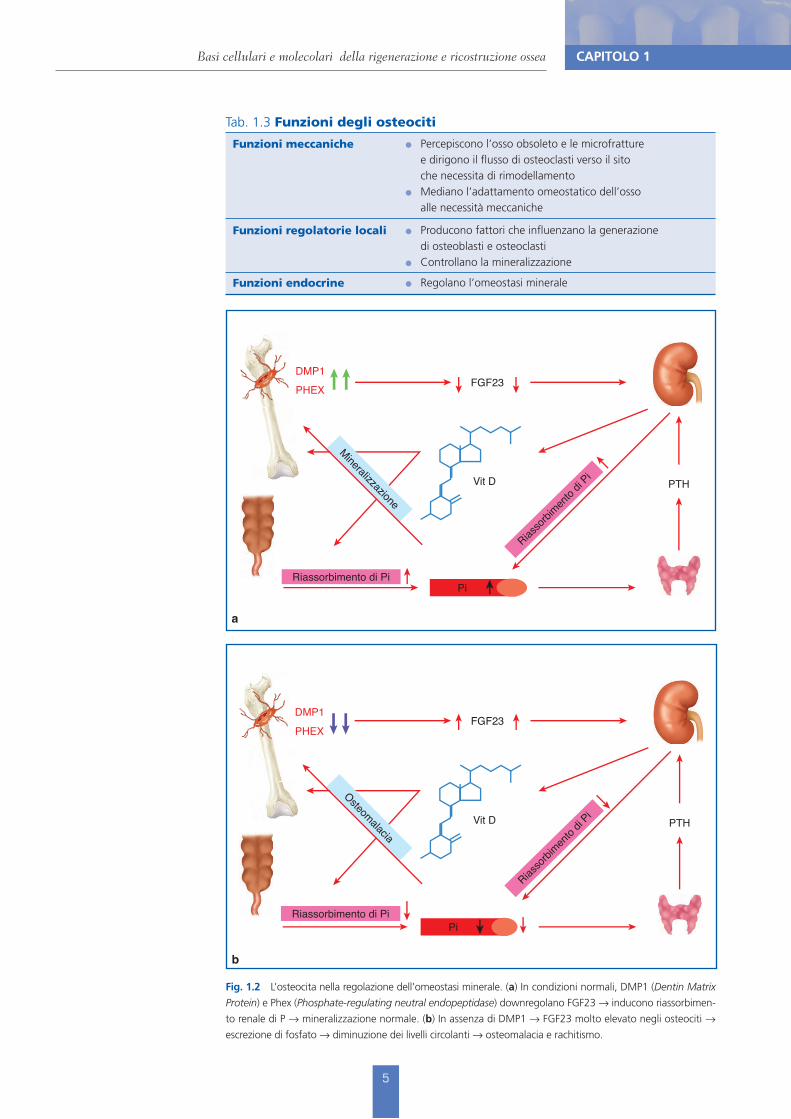
Fig. 1.2 L’osteocita nella regolazione dell’omeostasi minerale. (a) In condizioni normali, DMP1 (Dentin Matrix Protein) e Phex (Phosphate-regulating neutral endopeptidase) downregolano FGF23 ! inducono riassorbimen-to renale di P ! mineralizzazione normale. (b) In assenza di DMP1 ! FGF23 molto elevato negli osteociti ! escrezione di fosfato ! diminuzione dei livelli circolanti ! osteomalacia e rachitismo.
DMP1PHEX FGF23
PTH
Mineralizzazione
Pi
Vit D
Riassor
biment
o di Pi
Riassorbimento di Pi
a
DMP1PHEX
FGF23
PTH
Osteomalacia
Pi
Vit D
Riassor
biment
o di Pi
Riassorbimento di Pi
b
5
Basi cellulari e molecolari della rigenerazione e ricostruzione ossea CAPITOLO 1
VEGF A-B-C-DaFGF, bFGF
Ihh
PTHrp
Capillari Cartilagineavascolare
Condrocitiipertrofici
Vaso sanguigno
Osteoblasti
ManicottoperiostaleSpongiosa
primaria
Centro diossificazionesecondaria
Ossotrabecolare
Midolloosseo
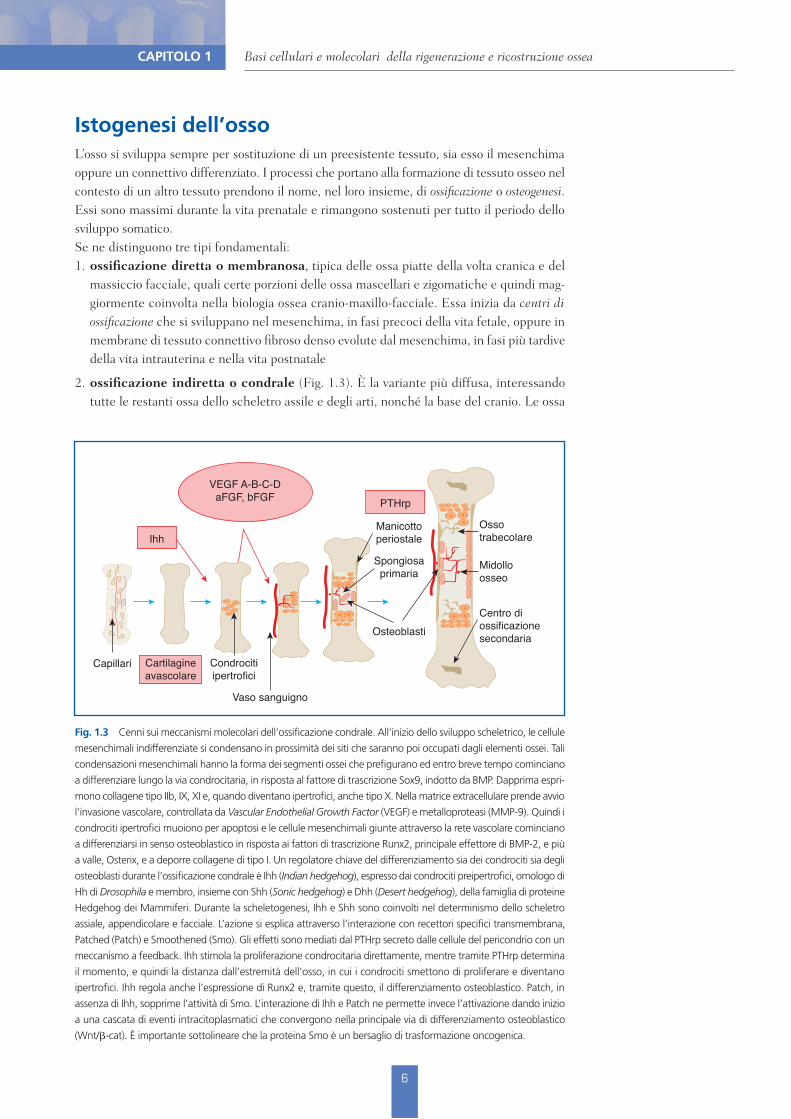
Fig. 1.3 Cenni sui meccanismi molecolari dell’ossificazione condrale. All’inizio dello sviluppo scheletrico, le cellule mesenchimali indifferenziate si condensano in prossimità dei siti che saranno poi occupati dagli elementi ossei. Tali condensazioni mesenchimali hanno la forma dei segmenti ossei che prefigurano ed entro breve tempo cominciano a differenziare lungo la via condrocitaria, in risposta al fattore di trascrizione Sox9, indotto da BMP. Dapprima espri-mono collagene tipo IIb, IX, XI e, quando diventano ipertrofici, anche tipo X. Nella matrice extracellulare prende avvio l’invasione vascolare, controllata da Vascular Endothelial Growth Factor (VEGF) e metalloproteasi (MMP-9). Quindi i condrociti ipertrofici muoiono per apoptosi e le cellule mesenchimali giunte attraverso la rete vascolare cominciano a differenziarsi in senso osteoblastico in risposta ai fattori di trascrizione Runx2, principale effettore di BMP-2, e più a valle, Osterix, e a deporre collagene di tipo I. Un regolatore chiave del differenziamento sia dei condrociti sia degli osteoblasti durante l’ossificazione condrale è Ihh (Indian hedgehog), espresso dai condrociti preipertrofici, omologo di Hh di Drosophila e membro, insieme con Shh (Sonic hedgehog) e Dhh (Desert hedgehog), della famiglia di proteine Hedgehog dei Mammiferi. Durante la scheletogenesi, Ihh e Shh sono coinvolti nel determinismo dello scheletro assiale, appendicolare e facciale. L’azione si esplica attraverso l’interazione con recettori specifici transmembrana, Patched (Patch) e Smoothened (Smo). Gli effetti sono mediati dal PTHrp secreto dalle cellule del pericondrio con un meccanismo a feedback. Ihh stimola la proliferazione condrocitaria direttamente, mentre tramite PTHrp determina il momento, e quindi la distanza dall’estremità dell’osso, in cui i condrociti smettono di proliferare e diventano ipertrofici. Ihh regola anche l’espressione di Runx2 e, tramite questo, il differenziamento osteoblastico. Patch, in assenza di Ihh, sopprime l’attività di Smo. L’interazione di Ihh e Patch ne permette invece l’attivazione dando inizio a una cascata di eventi intracitoplasmatici che convergono nella principale via di differenziamento osteoblastico (Wnt/!-cat). È importante sottolineare che la proteina Smo è un bersaglio di trasformazione oncogenica.
6
Basi cellulari e molecolari della rigenerazione e ricostruzione osseaCAPITOLO 1
Istogenesi dell’ossoL’osso si sviluppa sempre per sostituzione di un preesistente tessuto, sia esso il mesenchima oppure un connettivo differenziato. I processi che portano alla formazione di tessuto osseo nel contesto di un altro tessuto prendono il nome, nel loro insieme, di ossificazione o osteogenesi. Essi sono massimi durante la vita prenatale e rimangono sostenuti per tutto il periodo dello sviluppo somatico.Se ne distinguono tre tipi fondamentali:1. ossificazione diretta o membranosa, tipica delle ossa piatte della volta cranica e del
massiccio facciale, quali certe porzioni delle ossa mascellari e zigomatiche e quindi mag-giormente coinvolta nella biologia ossea cranio-maxillo-facciale. Essa inizia da centri di ossificazione che si sviluppano nel mesenchima, in fasi precoci della vita fetale, oppure in membrane di tessuto connettivo fibroso denso evolute dal mesenchima, in fasi più tardive della vita intrauterina e nella vita postnatale
2. ossificazione indiretta o condrale (Fig. 1.3). È la variante più diffusa, interessando tutte le restanti ossa dello scheletro assile e degli arti, nonché la base del cranio. Le ossa

A B C D
Fig. 1.4 Fasi del rimodellamento osseo. (a) Attivazione degli osteoclasti maturi, reclutamento di nuovi osteo-clasti e riassorbimento della matrice organica e minerale. (b) Passaggio tra periodo di rimozione del tessuto osseo e fase di formazione e attivazione degli osteoblasti. (c) Neoformazione e deposizione della matrice ossea che successivamente andrà incontro a mineralizzazione. (d) Ritorno al precedente stato di riposo con osteociti indovati nella matrice mineralizzata.
7
Basi cellulari e molecolari della rigenerazione e ricostruzione ossea CAPITOLO 1
che si formano con tale modalità sono anche dette ossa di sostituzione. Nelle fasi iniziali si realizza un abbozzo cartilagineo, che richiama la forma del futuro segmento osseo, e che viene gradualmente riassorbito e rimpiazzato dal tessuto definitivo. L’osso si forma sia alla superficie dell’abbozzo cartilagineo, apponendosi all’esterno tra cartilagine e pericondrio (ossificazione pericondrale), sia all’interno di questo (ossificazione endocondrale)
3. ossificazione mantellare. Avviene a livello del corpo della mandibola. Può essere consi-derata una variante di ossificazione diretta in quanto si realizza, seguendo le stesse tappe di questa, nel contesto di un mesenchima e poi di un tessuto connettivo.La peculiarità di questa modalità di ossificazione è che l’osso in formazione si modella intorno a un abbozzo cartilagineo conformato a ferro di cavallo, detto cartilagine di Meckel, che deriva dal mesenchima del primo arco branchiale. A differenza di quanto avviene per gli abbozzi scheletrici cartilaginei nell’ossificazione indiretta, essa non ossifica, ma viene circondata completamente dal tessuto osseo e infine involve, e il vuoto che rimane viene colmato da tessuto osseo. Solo nella regione del mento un piccolo tratto della cartilagine di Meckel viene inglobato nella mandibola e ossifica con meccanismo condrale.
Rimodellamento osseoIl rimodellamento osseo è il processo fisiologico attraverso il quale la massa ossea è man-tenuta costante nei vertebrati tra la fine della pubertà e il termine della funzione gonadica, consentendo il rinnovamento della matrice e il continuo adattamento funzionale a livello di organo e tessuto. Esso dipende da una serie di eventi che si susseguono in maniera sequen-ziale all’interno di unità funzionali autonome denominate unità multicellulari di base (Basic Multicellular Unit, BMU).La singola unità va incontro ciclicamente a un processo caratterizzato da varie fasi (Fig. 1.4):! nascita e formazione di uno stimolo in grado di modificare lo stato di riposo della BMU! attivazione delle cellule osteoclastiche mature accompagnata dalla formazione e dal re-
clutamento di nuovi osteoclasti! riassorbimento della matrice organica e minerale! fase di passaggio (reversal phase) che definisce il periodo di rimozione del tessuto osseo
ed è seguita dalla fase di formazione e attivazione degli osteoblasti! neoformazione e deposizione della matrice ossea che successivamente andrà incontro a
mineralizzazione! ritorno al precedente stato di riposo.

8
Basi cellulari e molecolari della rigenerazione e ricostruzione osseaCAPITOLO 1
L’intero ciclo di eventi sopra esposti avviene simultaneamente in siti scheletrici multipli, con il verificarsi di riassorbimento e formazione in stretta contiguità nelle BMU e si realizza a livello delle superfici interne (endocorticale, intracorticale, trabecolare) e, in misura minore, sulla su-perficie periostale. In ogni dato istante, il rimodellamento delle varie localizzazioni è asincrono. I due eventi hanno durata molto diversa, in quanto il riassorbimento avviene in 3-4 settimane mentre la formazione si compie in 3-4 mesi. Un ciclo di rimodellamento dura 6-9 mesi e in ogni momento il 5-25% dello scheletro è interessato dal fenomeno, con un intervallo tra eventi successivi nello stesso punto stimato in 2-5 anni. È stato calcolato che approssimativamante il 10% del volume dell’osso viene rimpiazzato ogni anno, come media di rinnovamento di osso corticale, che costituisce circa il 75% dello scheletro e ha un tasso di rimodellamento del 4% all’anno, e del trabecolare, che rappresenta il rimanente 25% e ha un tasso del 28%.L’entità di rimodellamento è determinata da una serie di fattori quali l’età, il genere, l’etnia, la stagione e il momento del giorno. Può poi essere alterata dall’esercizio fisico e dal regime dietetico. In particolare, sono associati a minore massa ossea l’età anziana, il sesso femminile e la razza asiatica. Inoltre, è noto che il riassorbimento osseo è aumentato in condizioni di riposo.
Rimodellamento osseo a livello orale. L’osso mandibolare e i mascellari sono sottoposti a continue sollecitazioni meccaniche determinate dal movimento dei denti, principalmente nel corso della masticazione. Il rimodellamento in queste sedi è particolarmente intenso, si parla infatti di ossa a elevato turnover, attività che consente la riparazione delle microfratture e la conservazione della competenza meccanica. Come descritto, gli osteociti rispondono in modo selettivo allo stimolo meccanico, liberando molteplici fattori che stimolano il reclutamento e l’attività osteoclastica da cui prende avvio il processo rimodellativo, ma anche le cellule del lega-mento parodontale, e i fibroblasti gengivali sono in grado di esprimere i fattori osteoclastogenici.È poi da considerare che il rimodellamento scheletrico aumenta in tutti i distretti con l’età e ciò è particolarmente evidente a livello mascellare e mandibolare per la presenza concomitan-te dello stato infiammatorio locale associato a disordini parodontali, anch’essi maggiormente prevalenti nell’anziano. Questo comporta una maggiore perdita di osso alveolare e un minore legame del dente. Si stima che circa il 50% dei soggetti di età superiore ai 50 anni abbia una perdita di osso alveolare superiore a 4-6 mm e il 3-11% delle donne in postmenopausa perda almeno un dente all’anno per tale motivo.
Angiogenesi e ossificazioneLo sviluppo scheletrico, così come la rigenerazione e la riparazione ossea, richiede l’ordinato susseguirsi di molti eventi che comprendono la migrazione, il differenziamento e l’attivazione di tipi e tessuti cellulari multipli. Lo sviluppo di una microvascolarizzazione e di un microcircolo sono essenziali per l’omeostasi e la rigenerazione di tessuto vitale: è noto che ogni alterazione nella vascolarizzazione del tessuto osseo può comportare stati patologici, come si verifica nel caso dell’osteonecrosi. Grazie alla rete vascolare possono essere infatti trasportati tutti gli elementi necessari alla formazione/rigenerazione ossea, come ossigeno, nutrienti, fattori solubili, cellule.I fattori coinvolti nella neoangiogenesi giocano anche un ruolo nella proliferazione delle cellule dell’osso e dei loro progenitori: Vascular Endothelial Growth Factor (VEGF), basic Fibroblast Growth Factor (bFGF), Transforming Growth Factor-! (TGF-!), Platelet Derived Growth Factor (PDGF), Hepatocyte Growth Factor (HGF), Insulin-like Growth Factors (IGFs), Nerve Growth Factor (NGF). Per alcuni di questi, l’effetto a livello scheletrico è un concetto acquisito da tem-po, mentre per altri, quali le neurotrofine, si tratta di osservazioni più recenti. Di fondamentale importanza sono poi il contatto e l’interazione tra le cellule staminali ematopoietiche (HSC) e gli osteoblasti, i quali sono in grado di produrre una varietà di fattori ematopoietici. In effetti, nell’os-so adulto, la bassa tensione di ossigeno che si realizza quando la matrice si accumula e la distanza

9
Basi cellulari e molecolari della rigenerazione e ricostruzione ossea CAPITOLO 1
degli osteociti dalla superficie aumenta, induce negli osteoblasti l’espressione di molecole che iniziano il programma di neovascolarizzazione. Di fatto, l’associazione tra osso e sangue è molto stretta. Gli osteoclasti appartengono alla linea dei monociti-macrofagi ed è stato dimostrato che il sito dell’ematopoiesi cambia nel corso dello sviluppo fino a raggiungere il midollo osseo e un compartimento ben definito al suo interno, la regione endostale all’interfaccia osso/midollo osseo. Come accennato, in questo contesto l’osteopontina è fondamentale nell’ematopoiesi, venendo a rappresentare un “ponte” tra osso e sangue: oltre a fornire, in quanto proteina strutturale, il supporto fisico e le molecole critiche per la realizzazione della rete vascolare, ha anche un’azione diretta nel mantenimento della quiescenza delle HSC che risiedono nella cavità endostale in una nicchia di cui gli osteoblasti rappresentano una componente essenziale. I condrociti iper-trofici esprimono e sintetizzano diverse classi di VEGF (A, B, C, D), i cui recettori si trovano principalmente a livello endoteliale, ma anche su monociti, neuroni, osteoblasti e sui condrociti stessi (VEGF-R2 e 3), realizzando un meccanismo autocrino oltre che paracrino (vedi Fig. 1.3).I condrociti ipertrofici esprimono poi BMP, il cui effettore Runx2 regola l’angiogenesi alla giunzione tra il piatto cartilagineo e le trabecole attraverso la sovraespressione di VEGF e l’inibizione di fattori antiangiogenici. Quando la condensazione si è verificata, il segnale BMP non è più indispensabile per l’osteogenesi ma lo è per la formazione della cavità midollare e per avere la presenza di osso trabecolare e corticale. Durante l’ontogenesi, le cellule endoteliali si sviluppano in una rete immatura che entra nell’osso attraverso i canali cartilaginei già formati. Nell’adulto, invece, la formazione dei canali procede all’unisono con la formazione dei vasi.L’osso possiede la caratteristica unica di rigenerarsi senza dare luogo a una cicatrice fibrosa. Ciò è ottenuto attraverso un processo riparativo che ripercorre in tutte le sue fasi lo sviluppo em-brionale dello scheletro, nel cui ambito la vascolarizzazione svolge una funzione determinante.In conseguenza a un trauma, il deterioramento del sistema vascolare porta all’attivazione di fattori trombotici nella cascata coagulativa, conducendo alla formazione di un ematoma. La risposta infiammatoria e la rottura del tessuto attivano citochine e fattori di crescita che reclu-tano gli osteoprogenitori e le cellule staminali dirigendoli verso il sito della frattura. Nella zona periferica del trauma, il tessuto di granulazione è sostituito da fibrocartilagine e in concomi-tanza il periostio va incontro a osteogenesi intramembranosa che porta alla formazione di un callo esterno, mentre internamente il tessuto viene mineralizzato a formare osso fibroso. Dopo la stabilizzazione del tessuto osseo e della vascolarizzazione nel sito di frattura, il processo di rimodellamento viene attivato e la rimozione di osso necrotico da parte degli osteoclasti è ac-compagnata dall’angiogenesi, che è seguita dalla sostituzione del callo osseo con osso lamellare. Nella sequenza di eventi che portano alla riparazione della frattura gioca un ruolo preminente una pletora di mediatori associati allo sviluppo osseo fetale e postnatale. Tra questi: BMP-2 e 4, VEGF, PDGF, FGF2 ecc. Inoltre, gli osteoclasti rilasciano eparinasi che induce il rilascio di VEGF dall’eparina, attivando quindi non solo l’angiogenesi ma anche il reclutamento osteo-clastico, poiché è stato dimostrato che VEGF promuove le fasi precoci dell’osteoclastogenesi.A volte la riparazione di una frattura può subire ritardo o non realizzarsi a causa dell’interfe-renza di fattori quali farmaci antinfiammatori, steroidi, o carenza di vitamina D, vitamina C e calcio, ma anche l’assenza di una rete vascolare funzionale è un elemento fondamentale della mancata guarigione.
Regolazione del tessuto osseoÈ difficile stabilire una gerarchia tra i meccanismi di controllo del rimodellamento schele-trico. La regolazione avviene a molti livelli, ma è in ultima analisi il risultato dell’azione di fattori principalmente locali perché le diverse fasi attraverso cui passa la BMU nel processo di rimodellamento, nonché i processi di proliferazione e differenziamento cellulare, sono controllati da mediatori prodotti proprio nel microambiente delle BMU.

10
Basi cellulari e molecolari della rigenerazione e ricostruzione osseaCAPITOLO 1
Regolazione localeÈ importante notare che molte delle molecole che agiscono localmente non sono specifiche per il tessuto osseo, ma sono anzi pressoché ubiquitarie. Tale pleiotropismo è giustificato dal fatto che svolgono azioni diverse in contesti diversi: è pertanto spesso la presenza dei recettori ormonali solo in alcuni tessuti che determina una specificità nell’azione locale, modulando sia la produzione sia la risposta cellulare. Inoltre, durante il riassorbimento osseo, citochine e fattori di crescita immagazzinati nella matrice vengono rilasciati in modo graduale dal processo osteolitico, contribuendo in tal modo alla realizzazione della ordinata sequenza delle fasi del rimodellamento.Infine, è ancora importante sottolineare che alcune di queste sostanze, proprio per il ruolo di coordinamento che svolgono, sono in grado di modulare l’attività sia degli osteoblasti sia degli osteoclasti, a seconda del momento in cui intervengono e della necessità di stimolare il processo riassorbitivo o, viceversa, quello neoappositivo (Tab. 1.4)
Specie reattive dell’ossigeno (ROS)L’ossigeno molecolare interviene nella formazione dell’osso non solo in quanto indispensabile per la fosforilazione ossidativa, ma anche come fattore di stimolo sulle cellule ossee. In ogni tipo di ossificazione, il differenziamento delle cellule mesenchimali in osteoprogenitori e poi in osteoblasti, si realizza in stretta concomitanza con la genesi di nuovi vasi sanguigni, che possono assicurare una elevata pressione parziale di ossigeno nelle sedi in cui avviene formazione di osso. Durante il normale metabolismo aerobico si formano molecole altamente reattive contenenti ossigeno, comunemente indicate con il termine ROS (Reactive Oxygen Species), che hanno un ruolo importante nella funzione cellulare (mediano, per esempio, la carbossilazione). In condizioni particolari, tuttavia, come in presenza di stimoli ambientali o durante l’invecchiamento, i ROS possono aumentare in modo drammatico causando danni irreversibili a cellule e tessuti.L’ossido nitrico (NO) è un radicale gassoso prodotto da molte cellule, incluse quelle endote-liali. È stato inizialmente identificato come agente vasodilatatore endogeno e mediatore della citotossicità macrofagica. È prodotto dall’enzima NO sintetasi (NOS), di cui si conoscono una forma costitutiva (cNOS), tipica dell’endotelio e delle cellule nervose, e una inducibile (iNOS), più ampiamente distribuita e presente anche a livello osseo, dove svolge una azione specifica, rappresentando una delle molecole prodotte dagli osteoblasti in grado di modulare direttamente l’attività osteoclastica. È stato anche dimostrato che numerosi fattori proinfiam-matori stimolano la produzione di NO a livello osseo tramite l’induzione dell’espressione di iNOS, confermando il ruolo di NO come effettore in varie patologie scheletriche.
Tab. 1.4 Fattori locali coinvolti nel rimodellamento osseo
Fattori attivi sul riassorbimento osseo
Fattori che stimolano le colonie ematopoieticheColony stimulating factors (CSFs): M-CSF, GM-CSF, IL-3
Fattori che stimolano il differenziamento e l’attività osteoclasticaRANKL, IL-1, TNF, Citochine gp130 (IL-6, IL-11*, LIF*, OSM*, CT-1*, CTNF), PGE2...
Fattori che inibiscono il riassorbimento osseoOPG, IFN, IL-4…
Fattori attivi sulla formazione ossea
Wnt, BMP, TGF!*, IGF-I e II, bFGF, PDGF...
I fattori in grado di agire sia sulla formazione che sul riassorbimento sono inseriti in tabella sulla base del loro ruolo predomi-nante e sono indicati con un asterisco. La tabella fornisce solo un elenco parziale dei fattori coinvolti nel rimodellamento osseo, giacché il progresso della ricerca evidenzia sempre nuovi elementi e vie di segnale.

Inibizionedella funzione
OSTEOCLASTA
EfrinaB2
c-Fos NF-AT RhoA
EphB4
PRE-OSTEOBLASTA
Aumentodel differenziamento
Fig. 1.5 Comunicazione diretta tra cellule ossee. La comunicazio-ne bidirezionale tra efrina B2 sulla superficie osteoclastica e recettore EphB4 sul preosteoblasta inibisce il differenziamento osteoclastico con un meccanismo mediato dal blocco di c-fos e al tempo stesso stimola l’osteoblastogenesi attraverso una diminuzione di RhoA.
11
Basi cellulari e molecolari della rigenerazione e ricostruzione ossea CAPITOLO 1
Comunicazione diretta: le efrineLa segnalazione cellulare bidirezionale è mediata dalle efrine. Queste sono suddivise in due classi: efrine A (1-6), proteine ancorate al glicosil-fosfatidil-inositolo ed efrine B (1-3), che sono proteine transmembrana. I ligandi, indicati con EphA (1-10) ed EphB (1-6), sono recettori tirosin chinasici. Le efrine e i loro recettori sono fattori coinvolti in molti processi di diffe-renziamento, inclusa la guida dell’assone neuronale, lo sviluppo intestinale e l’angiogenesi, nonché in alcuni tipi di cancro. A livello del tessuto osseo un contatto diretto tra osteoblasta e osteoclasta non è ancora stato dimostrato in maniera univoca, sebbene molte ricerche siano state compiute in questo senso. Si realizza invece, nell’ambito delle BMU, tra osteoclasta e preosteoblasta nel corso del processo di rimodellamento.L’interazione dell’efrina A2 con il recettore EphA2 facilita le fasi iniziali del rimodella-mento e l’osteoclastogenesi. Viceversa, la comunicazione tra efrina B2 e recettore EphB4 inibisce il differenziamento osteoclastico, con un meccanismo mediato dal blocco di c-fos, e stimola l’osteoblastogenesi attraverso una diminuzione di RhoA, come schematizzato in Figura 1.5.
Regolazione centraleIl cervello è un regolatore chiave dell’omeostasi di molti organi e tessuti e nell’ultima decade sono stati compiuti grandi passi verso l’identificazione di meccanismi di regolazione centrale anche nell’ambito del rimodellamento scheletrico. Allo stato attuale delle conoscenze sono note due vie metaboliche principali: la prima comprende segnali neuroendocrini generati nell’ipotalamo e successivamente processati tramite l’ipofisi, mentre la seconda consiste di neuroni efferenti ipotalamici che scaricano attraverso le vie nervose.Evidenze scientifiche hanno dimostrato una regolazione coordinata di tipo endocrino tra il rimodellamento osseo e il metabolismo energetico, il cui elemento chiave è stato identificato nella leptina, che svolge un ruolo critico nella regolazione della massa ossea quanto nell’ap-petito e nella spesa calorica: fondamentale ai fini di tale associazione è l’osservazione che dal punto di vista evolutivo essa compare contemporaneamente allo scheletro, ma dopo altri fattori coinvolti nel metabolismo energetico.Studi più recenti hanno chiarito che gli effetti scheletrici periferici della leptina sono mediati dalla serotonina, di cui inibisce la sintesi. Inoltre, è stato dimostrato che la diminuzione dei livelli di serotonina media anche il decremento di massa ossea associato all’anoressia nervosa.

12
Basi cellulari e molecolari della rigenerazione e ricostruzione osseaCAPITOLO 1
L’osso a sua volta interviene nel controllo del metabolismo energetico regolando la secrezione di insulina nelle cellule ! pancreatiche, la sua sensibilità e la spesa energetica attraverso una forma non carbossilata di osteocalcina, che raggiunge più facilmente il torrente circolatorio.
Regolazione sistemica
Il metabolismo del calcio è modulato da sostanze ormonali che ne regolano la biodisponibilità orchestrando i processi di formazione e di riassorbimento osseo. Tra queste vanno elencate il paratormone (PTH), il peptide paratormono-simile (PTHrp), la calcitonina (CT), l’ormone della crescita (GH), e gli ormoni tiroidei e sessuali, per il cui meccanismo d’azione e signifi-cato fisiologico nel tessuto scheletrico si può fare riferimento ai testi universitari preclinici.Un approfondimento sarà qui riservato al gruppo di molecole genericamente indicate come vitamina D.I soggetti che assumono queste sostanze possono essere infatti di frequente riscontro negli ambulatori odontoiatrici, dato l’ampio impiego clinico della vitamina D nella terapia e nella prevenzione di disordini non solo del metabolismo minerale, ma anche di altre problematiche genericamente associate all’invecchiamento e al deterioramento di diversi organi e sistemi.Con il termine sistema endocrino della vitamina D si indica un gruppo di sostanze dotate di attività ormonale, il cui precursore comune è il 7-deidrocolesterolo. La vitamina D può derivare da sintesi endogena (a livello epatico e successiva conversione a livello cutaneo, mediata dal calore e dai raggi UV) o dalla dieta, da sostanze di origine sia animale (colecal-ciferolo, vitamina D3) sia vegetale (ergocalciferolo, vitamina D2).Le tappe successive comprendono una idrossilazione a livello epatico – con formazione di 25-OH vit. D, calcifediolo – e l’idrossilazione, in posizione 1", a livello renale, con formazione di 1,25(OH)2 vit. D (1,25(OH)2D3 o calcitriolo) che è il metabolita biologicamente più attivo. L’attività della 1"-idrossilasi è regolata da molti fattori, quali PTH, estrogeni, corticosteroidi e livelli di fosforo sierico; la diminuzione della calcemia stimola la sintesi renale con un meccanismo PTH-mediato.La sintesi di 25-OH vit. D è regolata tramite un meccanismo di feedback negativo dalla quantità di vitamina D presente nell’organismo e dalle concentrazioni di 1,25(OH)2D3. Poiché rappresenta la maggiore forma circolante dell’ormone, è il metabolita che normalmente viene dosato nel siero, dove riflette lo stato vitaminico D dell’organismo oltre a rappresentare la molecola di riserva per i successivi passaggi metabolici.L’importanza del calcitriolo nel metabolismo scheletrico è da ricercare nella sua azione che si realizza a livello intestinale, osseo e renale, ed è esplicata tramite il legame con un recettore nucleare detto VDR che attiva la trascrizione genica dei vari effettori a valle. La presenza di recettori per il calcitriolo è stata recentemente dimostrata in una varietà di altre cellule e tessuti, incluse cellule muscolari, endoteliali, immunitarie ecc.Di conseguenza, sono numerosissimi gli effetti extrascheletrici, la cui lista va accrescendosi di pari passo con la ricerca. È stato ampiamente dimostrato che la vitamina D interviene, con esiti benefici, nella modulazione di patologie cardiovascolari, autoimmunitarie, degenerative e neoplastiche, e il conseguimento di un adeguato stato vitaminico riveste sempre maggiore importanza.La definizione dei livelli sierici di 25-OH vit. D, che descrivano appunto lo stato vitaminico, sono in continuo aggiornamento (Tab. 1.5). La concentrazione di 75 nmol/L, corrispondente a 32 ng/mL, viene attualmente considerata come il limite inferiore dell’intervallo ottimale che garantisce gli effetti a livello scheletrico ed extrascheletrico.
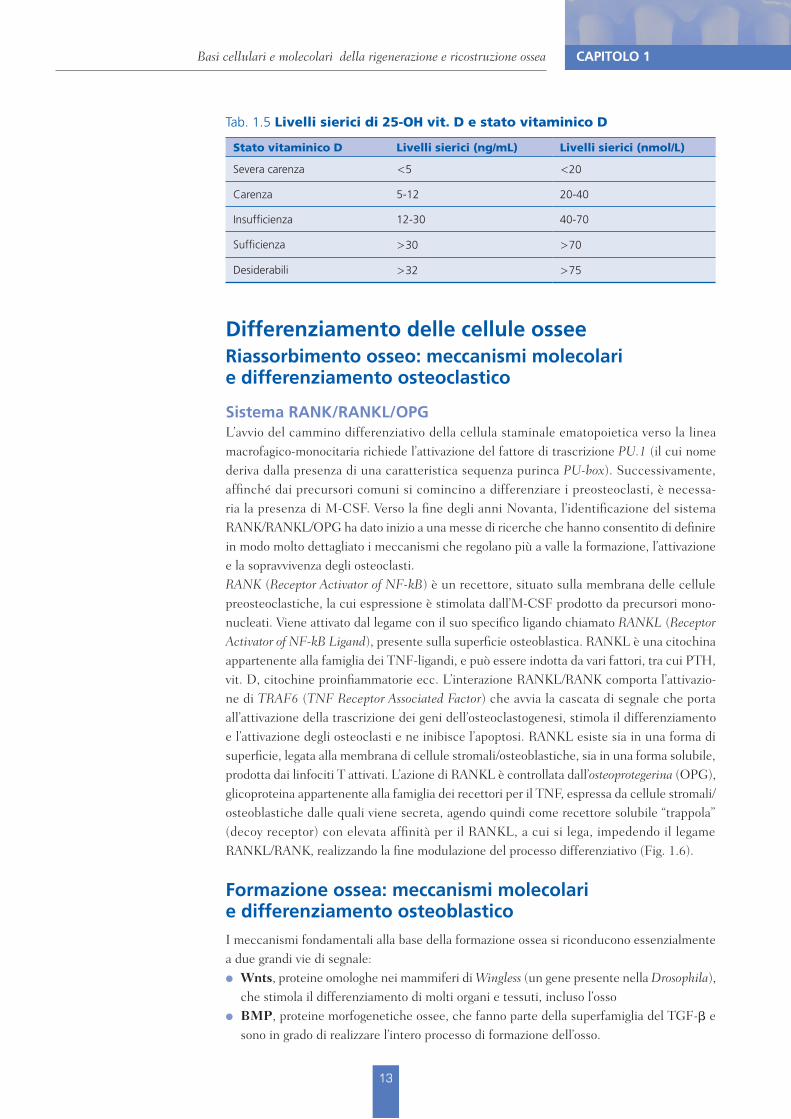
Tab. 1.5 Livelli sierici di 25-OH vit. D e stato vitaminico D
Stato vitaminico D Livelli sierici (ng/mL) Livelli sierici (nmol/L)
Severa carenza <5 <20
Carenza 5-12 20-40
Insufficienza 12-30 40-70
Sufficienza >30 >70
Desiderabili >32 >75
13
Basi cellulari e molecolari della rigenerazione e ricostruzione ossea CAPITOLO 1
Differenziamento delle cellule osseeRiassorbimento osseo: meccanismi molecolari e differenziamento osteoclastico
Sistema RANK/RANKL/OPGL’avvio del cammino differenziativo della cellula staminale ematopoietica verso la linea macrofagico-monocitaria richiede l’attivazione del fattore di trascrizione PU.1 (il cui nome deriva dalla presenza di una caratteristica sequenza purinca PU-box). Successivamente, affinché dai precursori comuni si comincino a differenziare i preosteoclasti, è necessa-ria la presenza di M-CSF. Verso la fine degli anni Novanta, l’identificazione del sistema RANK/RANKL/OPG ha dato inizio a una messe di ricerche che hanno consentito di definire in modo molto dettagliato i meccanismi che regolano più a valle la formazione, l’attivazione e la sopravvivenza degli osteoclasti.RANK (Receptor Activator of NF-kB) è un recettore, situato sulla membrana delle cellule preosteoclastiche, la cui espressione è stimolata dall’M-CSF prodotto da precursori mono-nucleati. Viene attivato dal legame con il suo specifico ligando chiamato RANKL (Receptor Activator of NF-kB Ligand), presente sulla superficie osteoblastica. RANKL è una citochina appartenente alla famiglia dei TNF-ligandi, e può essere indotta da vari fattori, tra cui PTH, vit. D, citochine proinfiammatorie ecc. L’interazione RANKL/RANK comporta l’attivazio-ne di TRAF6 (TNF Receptor Associated Factor) che avvia la cascata di segnale che porta all’attivazione della trascrizione dei geni dell’osteoclastogenesi, stimola il differenziamento e l’attivazione degli osteoclasti e ne inibisce l’apoptosi. RANKL esiste sia in una forma di superficie, legata alla membrana di cellule stromali/osteoblastiche, sia in una forma solubile, prodotta dai linfociti T attivati. L’azione di RANKL è controllata dall’osteoprotegerina (OPG), glicoproteina appartenente alla famiglia dei recettori per il TNF, espressa da cellule stromali/osteoblastiche dalle quali viene secreta, agendo quindi come recettore solubile “trappola” (decoy receptor) con elevata affinità per il RANKL, a cui si lega, impedendo il legame RANKL/RANK, realizzando la fine modulazione del processo differenziativo (Fig. 1.6).
Formazione ossea: meccanismi molecolari e differenziamento osteoblasticoI meccanismi fondamentali alla base della formazione ossea si riconducono essenzialmente a due grandi vie di segnale:
! Wnts, proteine omologhe nei mammiferi di Wingless (un gene presente nella Drosophila), che stimola il differenziamento di molti organi e tessuti, incluso l’osso
! BMP, proteine morfogenetiche ossee, che fanno parte della superfamiglia del TGF-! e sono in grado di realizzare l’intero processo di formazione dell’osso.

14
Basi cellulari e molecolari della rigenerazione e ricostruzione osseaCAPITOLO 1
Wnt e osteoblastogenesi
Segnale Wnt/a-catenina e regolazione della massa osseaLe proteine della famiglia Wnt sono un gruppo di 19 ligandi solubili che si legano a una fa-miglia di 10 recettori cellulari di superficie, chiamati Fzd (Frizzled Receptors) e al corecettore LRP (Lipoprotein Receptor related Protein) 5 o 6, dando avvio alla trasduzione del segnale intracellulare mediata dall’attivazione della !-catenina, molecola implicata nella stabilizza-zione del citoscheletro che realizza anche le giunzioni cellulari attraverso la formazione di un complesso proteico con la caderina. Una volta traslocata nel nucleo, la !-catenina interagisce con i repressori della trascrizione LEF/TCF (Lymphoid Enhancer binding Factor/T Cell specific transcription Factor). L’interazione permette il reclutamento dell’istone CBP/p300 (CREB Binding Protein) e l’attivazione della trascrizione genica e determina il differenziamento e la crescita delle cellule osteoblastiche. Oltre alla via canonica che utilizza la !-catenina, il se-gnale Wnt può essere trasmesso attraverso vie non canoniche (Ca-dipendente e PCP-planar cell polarity), che non sono note con altrettanta chiarezza e comportano una regolazione del citoscheletro attraverso altre molecole di segnale (RhoA, Rac2, Cdc42, e le piccole GTPasi).Il sistema Wnt/!-catenina incrementa la formazione ossea a livello cellulare in modo inte-grato, attraverso meccanismi che includono il rinnovamento delle cellule staminali, la sti-molazione della replicazione preosteoblastica, l’induzione dell’osteoblastogenesi e l’inibizione dell’apoptosi degli osteoblasti e degli osteociti.Il segnale Wnt è finemente regolato e può essere inibito a vari livelli sia extra- sia intracellular-mente. Il principale inibitore è la sclerostina prodotta dagli osteociti, che si lega al corecettore LRP 5/6, rendendolo indisponibile per l’interazione con Wnt. Vari altri fattori possono poi bloccare Wnt direttamente, come nel caso di sFRP (recettore frizzled solubile) o tramite il complesso recetto-riale come Dkk1 (Dikkopf), Kremen1/2 (corecettore di Dkk). All’interno della cellula, la GSK3, catalizzando la fosforilazione della !-catenina, ne determina la degradazione. Dati più recenti indicano che anche le proteine della famiglia Rspo intervengono nell’interazione Wnt/!-catenina.I dettagli dell’attivazione della via canonica Wnt e la sua regolazione sono descritti in Figura 1.7.
OSTEOBLASTA
PRE-OSTEOCLASTA
OPG
NF-kB
RANKL
RANK
TRAF6
Osteoclastogenesi
c-Fos NF-ATc1
Fig. 1.6 Il sistema RANK/RANKL/OPG. Regolazione della formazio-ne, attivazione e sopravvivenza de-gli osteoclasti. L’interazione del re-cettore RANK con il ligando RANKL provoca l’attivazione di TRAF6 e quindi della trascrizione. OPG lega RANKL realizzando il blocco del differenziamento e dell’attivazione osteoclastica.
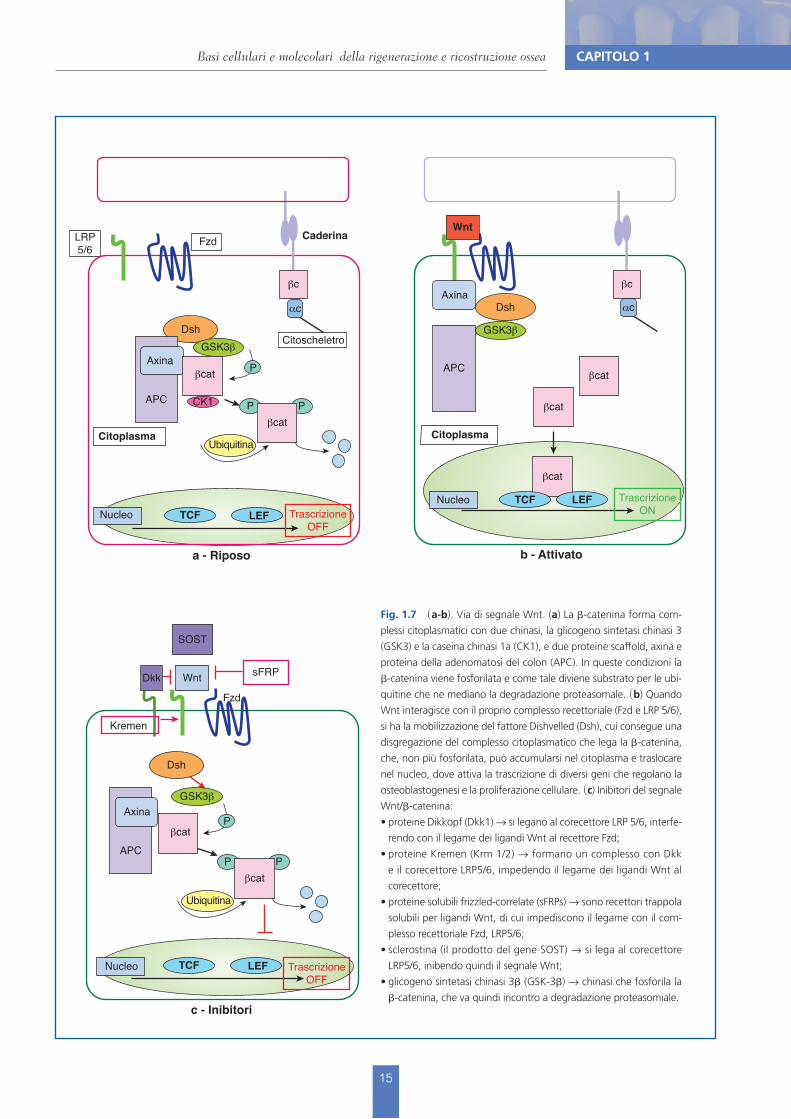
Fig. 1.7 (a-b). Via di segnale Wnt. (a) La !-catenina forma com-plessi citoplasmatici con due chinasi, la glicogeno sintetasi chinasi 3 (GSK3) e la caseina chinasi 1a (CK1), e due proteine scaffold, axina e proteina della adenomatosi del colon (APC). In queste condizioni la !-catenina viene fosforilata e come tale diviene substrato per le ubi-quitine che ne mediano la degradazione proteasomale. (b) Quando Wnt interagisce con il proprio complesso recettoriale (Fzd e LRP 5/6), si ha la mobilizzazione del fattore Dishvelled (Dsh), cui consegue una disgregazione del complesso citoplasmatico che lega la !-catenina, che, non più fosforilata, può accumularsi nel citoplasma e traslocare nel nucleo, dove attiva la trascrizione di diversi geni che regolano la osteoblastogenesi e la proliferazione cellulare. (c) Inibitori del segnale Wnt/!-catenina:
proteine Dikkopf (Dkk1) ! si legano al corecettore LRP 5/6, interfe-rendo con il legame dei ligandi Wnt al recettore Fzd; proteine Kremen (Krm 1/2) ! formano un complesso con Dkk e il corecettore LRP5/6, impedendo il legame dei ligandi Wnt al corecettore; proteine solubili frizzled-correlate (sFRPs) ! sono recettori trappola solubili per ligandi Wnt, di cui impediscono il legame con il com-plesso recettoriale Fzd, LRP5/6; sclerostina (il prodotto del gene SOST) ! si lega al corecettore LRP5/6, inibendo quindi il segnale Wnt; glicogeno sintetasi chinasi 3! (GSK-3!) ! chinasi che fosforila la !-catenina, che va quindi incontro a degradazione proteasomiale.
Fzd
SOST
APCP P
Dsh
Axina
Nucleo TCF LEF
GSK3!
!cat
!cat
TrascrizioneOFF
sFRPWntDkk
Kremen
Ubiquitina
P
LRP5/6
P P
Dsh
Fzd Caderina
Axina
Citoplasma Ubiquitina
Nucleo
Citoscheletro
CK1APC
TCF LEF
GSK3!
!cat
!cat
!c
"c
TrascrizioneOFF
P
a - Riposo
DshAxina
Citoplasma
Nucleo
APC
!cat
TCF LEF
GSK3!
!cat
!cat
!c
"c
TrascrizioneON
Wnt
b - Attivato
c - Inibitori
15
Basi cellulari e molecolari della rigenerazione e ricostruzione ossea CAPITOLO 1

16
Basi cellulari e molecolari della rigenerazione e ricostruzione osseaCAPITOLO 1
Bone Morphogenetic Proteins e osteogenesiLe notevoli capacità rigenerative dell’osso sono note fin dall’antichità: Ippocrate, colpito dall’efficienza dei processi di riparazione dell’osso, aveva postulato la presenza all’interno dell’organismo di sostanze che, possedendo tali proprietà guaritrici, potessero essere poten-zialmente utilizzate in campo clinico. Nel 1945 Pierre Lacroix ipotizzava che l’osso potesse contenere una sostanza, l’osteogenina, in grado di stimolare l’osteogenesi. L’impianto di diafisi ossee demineralizzate in siti eterotopici non scheletrici induce infatti la formazione di osso.La “attività” fu chiamata Bone Morphogenetic Protein (BMP) da Urist nel 1965, ma le pro-teine responsabili dell’induzione di osso rimasero sconosciute fino al 1988, quando furono clonate le BMP 2-4 umane.Le Bone Morphogenetic Proteins, con la sola eccezione di BMP-1 che è una proteasi (procol-lagene C-endopeptidasi), fanno parte della superfamiglia del TGF-!. Sono sintetizzate come grandi (400-525 aa) molecole precursori consistenti in un peptide di segnale, un prodominio e un dominio carbossiterminale con 7 residui di cisteina altamente conservati, che contribuiscono alla formazione del nucleo nel core proteico a livello del quale si stabiliscono le interazioni con altre molecole per realizzare la dimerizzazione, che avviene tramite ponti disolfuro. Dopo l’eliminazione del peptide di segnale, la proteina precursore va incontro a glicosilazione. La proteina matura è quindi secreta e infine dimerizza, a livello intra- o extracellulare.La BMP-2 è stata isolata alla fine del 1987 dall’osso bovino e inizialmente suddivisa in 2a e 2b (quest’ultima successivamente denominata BMP-4), sulla base della diversa affinità dimostrata dal gene codificante durante l’ibridazione con mRNA umano. Viene secreta come precursore di 18 kDa e la sua forma attiva è un omodimero di circa 30 kDa, costituito da due frammenti C-terminali di 14 kDa glicosilati in diversi siti. Nella famiglia delle BMP sono comprese anche le proteine GDF (Growth and Differentiation Factors) o CDMP (Cartilage-Derived Morphogentic Proteins). Di fatto, la nomenclatura di queste classi di composti si sovrappone in molti casi e le designa-zioni sono ridondanti. Tutte contengono un motivo di 7 cisteine separate da una caratteristica spaziatura (cystine-knot motif), tipico della superfamiglia del TGF-!. Nell’insieme formano un gruppo di circa 20 citochine coinvolte in vari stadi e processi dello sviluppo embrionale e del differenziamento di organi e tessuti (Tab. 1.6).
Via di segnale delle BMPIl complesso recettoriale delle BMP (Fig. 1.8) è un sistema tetramerico costituito da due recettori tipo I (BMPrI) e due recettori tipo II (BMPrII), tutti dotati di attività serina-treonina chinasica. I BMPrI sono i membri 1, 2, 3 e 6 della famiglia Activin receptor-Like Kinase (ALK) 1-7, mentre i BMPrII fanno parte della famiglia Activin type IIA e IIB Receptors (ActRIIA e ActRIIB). In seguito al legame con BMP, i BMPrI vengono fosforilati dai BMPrII, costituti-vamente attivi, avviando la trasduzione del segnale che si realizza attraverso la via canonica (Smad-Runx2) o non canonica (MAPK).1. Via canonica. Dopo l’attivazione del complesso recettoriale in seguito all’interazione con
la BMP, la trasduzione del segnale avviene attraverso alcune proteine intracellulari della famiglia Smad, il cui nome deriva dalla combinazione di quelli delle proteine omologhe Mothers Against Decapentaplegic (MAD) di Drosophila e SMA di Caenorhabditis elegans. Le R-Smad 1,3,5 (Receptor-activated Smad, proteine attivate dal recettore) vengono fo-sforilate e in questa forma interagiscono con un altro membro della famiglia Smad, la Smad4 (common partner Smad, co-Smad), a formare un eterotrimero che può traslocare nel nucleo. Questa interazione può essere prevenuta dai membri della famiglia Smad con funzione inibitoria (I-Smad 6 e 7).
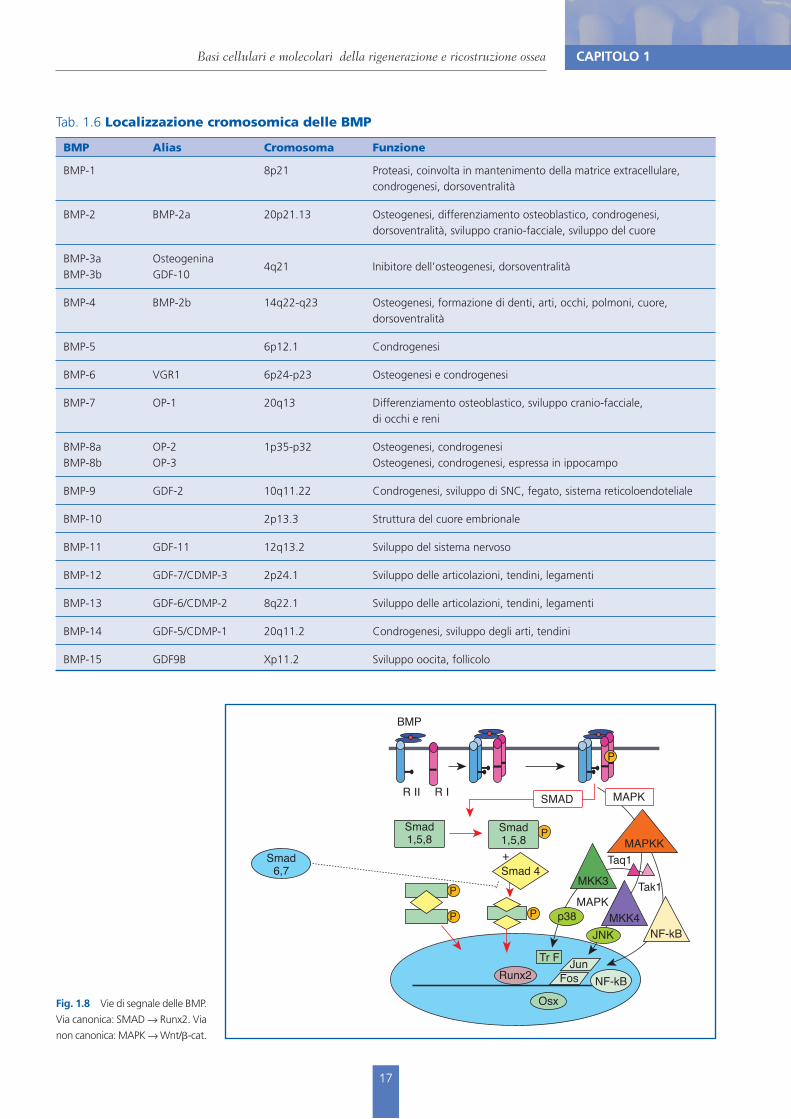
Tab. 1.6 Localizzazione cromosomica delle BMP
BMP Alias Cromosoma Funzione
BMP-1 8p21 Proteasi, coinvolta in mantenimento della matrice extracellulare, condrogenesi, dorsoventralità
BMP-2 BMP-2a 20p21.13 Osteogenesi, differenziamento osteoblastico, condrogenesi, dorsoventralità, sviluppo cranio-facciale, sviluppo del cuore
BMP-3aBMP-3b
OsteogeninaGDF-10
4q21 Inibitore dell’osteogenesi, dorsoventralità
BMP-4 BMP-2b 14q22-q23 Osteogenesi, formazione di denti, arti, occhi, polmoni, cuore, dorsoventralità
BMP-5 6p12.1 Condrogenesi
BMP-6 VGR1 6p24-p23 Osteogenesi e condrogenesi
BMP-7 OP-1 20q13 Differenziamento osteoblastico, sviluppo cranio-facciale, di occhi e reni
BMP-8aBMP-8b
OP-2OP-3
1p35-p32 Osteogenesi, condrogenesi Osteogenesi, condrogenesi, espressa in ippocampo
BMP-9 GDF-2 10q11.22 Condrogenesi, sviluppo di SNC, fegato, sistema reticoloendoteliale
BMP-10 2p13.3 Struttura del cuore embrionale
BMP-11 GDF-11 12q13.2 Sviluppo del sistema nervoso
BMP-12 GDF-7/CDMP-3 2p24.1 Sviluppo delle articolazioni, tendini, legamenti
BMP-13 GDF-6/CDMP-2 8q22.1 Sviluppo delle articolazioni, tendini, legamenti
BMP-14 GDF-5/CDMP-1 20q11.2 Condrogenesi, sviluppo degli arti, tendini
BMP-15 GDF9B Xp11.2 Sviluppo oocita, follicolo
BMP
P
P
P
P P
SMAD
Smad6,7
Runx2
Osx
JunFos
Tr F
Smad1,5,8
Smad1,5,8
Smad 4
MAPK
MAPKK
NF-kB
+
p38JNK
MAPK
Taq1
Tak1MKK3
MKK4NF-kB
R II R I
Fig. 1.8 Vie di segnale delle BMP. Via canonica: SMAD ! Runx2. Via non canonica: MAPK ! Wnt/!-cat.
17
Basi cellulari e molecolari della rigenerazione e ricostruzione ossea CAPITOLO 1

PU.1M-CSF
MSC Condrocita
Preosteoclasta Preosteoblasta
BMPSox9
BMPRunx2
Wnt
DKK-1OPG
HSC
RANKL
Osteoclasta Osteoblasta
Fig. 1.9 Meccanismi molecolari del differenziamento delle cellule del tessuto osseo. Sulla sinistra: osteoclastogenesi. Dalla cellula staminale ematopoietica (HSC) originano i preosteoclasti, in pre-senza di PU.1 e M-CSF, e quindi gli osteoclasti, con l’intervento di RANKL. Sulla destra: osteoblasto-genesi. Dalla cellula mesenchima-le staminale (MSC) BMP induce il differenziamento dei condrociti, tramite Sox9, e degli osteoblasti, tramite Runx2, con la cooperazio-ne di Wnt. Gli osteoblasti regolano l’osteoclastogenesi tramite RANKL e OPG.
18
Basi cellulari e molecolari della rigenerazione e ricostruzione osseaCAPITOLO 1
Una volta nel nucleo, i complessi trimerici legano degli elementi bersaglio presenti nei promotori di diversi geni e reclutano una varietà di coregolatori e corepressori trascrizionali. In questo modo il segnale delle BMP può interessare molti geni contemporaneamente. Tra gli elementi chiave attivati nel nucleo svolgono un ruolo principale: Runx2, Sox9, Msx2 e, più a valle, Osterix (Osx).
Del ruolo di Sox9 nel differenziamento condrocitario si è già accennato nella descrizione dei processi molecolari dell’ossificazione condrale. Runx2 (Runt related transcription factor 2 detto anche Cbfa1, Core binding factor A1) è un fattore di trascrizione della famiglia Runt necessario ma non sufficiente per il completo differenziamento osteogenico, è un marker precoce della linea osteoblastica e modula l’espressione di numerosi fattori (VEGF, RANKL, SOST, DMP1 ecc.) e successivamente quella di geni associati con il differen-ziamento terminale, come l’osteocalcina e l’osteopontina, di cui è il principale regolatore.
2. Via non canonica. In alternativa, il complesso recettoriale attivato dal legame con BMP può fosforilare la chinasi MAPKK (Mitogen Activated Protein Kinase Kinase) avviando la cascata di segnale che, attraverso la fosforilazione di altre proteine come p38 e JNK, e infine di jun, fos e NF-"B, promuove a livello nucleare la trascrizione di geni critici per l’induzione ossea.
È interessante notare che le BMP, attraverso MAPK, bloccano la GSK3 del complesso che fosforila la !-catenina, favorendone l’accumulo e la traslocazione nucleare, cooperando quindi con la via Wnt. Come sopra descritto, le BMP, utilizzando la via canonica che attiva Runx2, stimolano la progressione dell’osteogenesi. Questa è regolata anche da Osx, che agisce più a valle. La BMP-2 è però anche un potente attivatore di Dlx5 e 3 (omologo di Distal less di Drosophila), che a sua volta induce Osterix ma è indipendente da Runx2. Questo meccanismo potrebbe spiegare l’attivazione di Osterix via MAPK, indipendente da Smad.A loro volta i promotori delle BMP hanno sequenze di legame per Runx, spiegando l’azione bifasica di BMP: nella fase iniziale BMP induce Runx2 e successivamente Runx2 aumenta BMP per il completamento del differenziamento attraverso l’attivazione di Dlx (Fig. 1.9).Esistono molteplici antagonisti delle BMP, tra cui Noggin, chordin, follistatina, sclerostina, che agiscono prevenendo il legame con il complesso recettoriale.

19
Basi cellulari e molecolari della rigenerazione e ricostruzione ossea CAPITOLO 1
Ruolo delle BMP nella filogenesi e nello sviluppo embrionale e considerazioni sul loro impiego in ingegneria tessutaleLa formazione di osso ectopico mediata dalle BMP ricapitola lo sviluppo fetale degli arti, nel processo di ossificazione sia condrale sia intramembranoso. Ciò si realizza attraverso l’azione sulle cellule staminali, siano esse embrionali, fetali o adulte, che possiedono molti recettori per BMP. Le Bone Morphogenetic Proteins hanno però un ruolo più ampio nel rego-lare la biologia delle cellule staminali: oltre a promuovere il differenziamento osteoblastico e condrocitario e a regolare il destino delle cellule della cresta neurale, mantengono anche la capacità di autorinnovamento delle cellule staminali embrionali.La sottofamiglia a cui appartengono le BMP include geni critici per lo sviluppo embrionale, come il Dpp (decapentaplegico) e il 60-A della Drosophila, il comune moscerino della frutta, che sono implicati nella formazione del pattern dorso-ventrale. Le BMP sono anche collegate al gene Vg-1 degli Anfibi, coinvolto nell’induzione mesodermica. Giacché dal mesoderma prendono origine la cartilagine, l’osso, il muscolo e il midollo osseo, è importante il fatto che le BMP isolate da osso adulto siano strutturalmente simili a geni coinvolti nel controllo dello sviluppo embrionale.Come accennato, ci sono voluti dieci anni per purificare e clonare la prima BMP, la BMP-3, e attualmente se ne contano circa una ventina. Il significato biologico di tale ridondanza è stato indagato ed è stato dimostrato che, nell’embrione di topo, la BMP-2 è presente anche in tessuti non scheletrici, quali cuore e follicoli piliferi. Le BMP possono quindi avere un ruolo ad ampio spettro come morfogeni durante lo sviluppo e l’organogenesi. Una caratteristica comune delle proteine regolatorie è che la cascata di eventi necessari alla trasduzione del segnale è altamente conservata. Come accennato, nella Drosophila, il gene Dpp è implicato nella determinazione della dorsoventralità e sia la BMP-2 sia la BMP-4 sono omologhe per oltre il 70% al prodotto di Dpp. Nella Drosophila sono assenti osso e cartilagine, ma i prodotti genici di Dpp e di 60-A (omologo di BMP-7) possono indurre la formazione di osso nel ratto. Analogamente, Drosophila mancanti del gene Dpp si sviluppano normalmente in presenza di BMP-2. La sequenza appare dunque essere conservata dagli insetti all’uomo, coprendo un periodo di circa 600 milioni di anni. Per di più, anche il principale effettore della BMP, il sistema Smad, consta di proteine molto conservate dal punto di vista evolutivo.Da questa premessa risulta evidente che la funzione delle BMP nell’organismo va ben oltre l’induzione di osso.Inoltre, i sistemi che regolano il differenziamento e la proliferazione cellulare sono pres-soché ubiquitari e le stesse molecole di segnale sono utilizzate dall’organismo in tessuti e contesti diversi sia in condizioni fisiologiche sia nella trasformazione oncogenica. In questo caso si viene a sbilanciare proprio il critico equilibrio tra differenziamento/quie-scenza da una parte e proliferazione/trasformazione dall’altra. Una vastissima produzione scientifica dimostra l’attivazione del sistema Wnt/!-catenina nelle fasi iniziali di molti tipi di cancro, e anche le vie di segnale della BMP interessano fattori coinvolti altresì nelle attivazioni oncogeniche (MAPK, i proto-oncogeni p38, c-fos e jun, per nominarne solo alcuni). Durante lo sviluppo embrionale, le BMP mediano la transizione epitelio-mesenchimale (EMT), processo grazie al quale le cellule possono migrare verso diversi microambienti per potersi differenziare in diversi tessuti. D’altra parte, esiste anche una EMT oncogenica che media l’invasività e il potenziale metastatico di molti tumori. Le molecole altamente coinvolte nei processi ontogenetici e differenziativi, come le BMP, agiscono spesso sulle cellule bersaglio in modo diverso, a seconda del loro stato proliferativo.

20
Basi cellulari e molecolari della rigenerazione e ricostruzione osseaCAPITOLO 1
Cellule staminali e rigenerazione tessutaleUn discorso particolare dovrebbe essere fatto per l’utilizzo di cellule staminali. Una trattazione dettagliata di questo aspetto esula dagli scopi di questo libro, ma non sarebbe deontologico concludere senza accennare alle problematiche che si possono accompagnare al loro impiego. Non dobbiamo dimenticare che:1. le cellule staminali sono, per le loro caratteristiche intrinseche, resistenti alle comuni
terapie farmacologiche antineoplastiche, oltre a non essere riconosciute dal sistema im-munitario
2. esistono anche cellule staminali cancerose che possono essere somministrate inavverti-tamente nel corso di un innesto di cellule staminali “sane” e rispondono in modo violento agli stimoli mitogenetici
3. l’utilizzo di uno scaffold tridimensionale può indurre la trasformazione perché rappresenta un ambiente permissivo la cui struttura rammenta quella dello stroma tumorale; gli stessi geni che vengono attivati nel processo di meccanotrasduzione che media il differenzia-mento di una MSC in senso osteoblastico quando posta su un substrato rigido, sono anche coinvolti nella trasformazione neoplastica.
La medicina rigenerativa e le potenzialità che l’impiego di fattori di crescita e di cellule staminali lasciano giustamente prevedere rappresentano senza dubbio il futuro verso cui la ricerca e l’applicazione clinica si rivolgeranno. Allo stato attuale delle conoscenze sappiamo però troppo poco sui reali rischi che si associano all’utilizzo di questi strumenti potentissimi ed è saggio impiegarli con cautela, fintantoché trial clinici controllati condotti su un numero di pazienti molto elevato e con follow-up a lungo e lunghissimo termine non ne avranno dimostrato con rigore scientifico la assoluta sicurezza di impiego.
Farmaci attivi sul tessuto osseoRiteniamo opportuno introdurre alcuni cenni sulla regolazione farmacologica del tessuto scheletrico, data la frequenza con cui si incontrano pazienti in trattamento per patologie ossee. Inoltre, poiché i farmaci in questione interagiscono con i processi di rimodellamento, i loro effetti assumono particolare importanza nella pratica odontoiatrica.
Osteoporosi. L’osteoporosi è la più frequente malattia metabolica dello scheletro, oltre a essere una delle patologie di più comune riscontro nella popolazione anziana. È caratterizzata da ridotta massa ossea e deterioramento della microarchitettura, cui consegue un aumento del rischio di frattura. Secondo stime recenti, in Italia, nella fascia di età compresa tra i 50 e i 59 anni, questa patologia interessa il 13% della popolazione femminile, e tale prevalenza aumenta con l’età fino a superare il 40% nelle ultrasettantenni.Attualmente il numero di soggetti di entrambi i sessi con ridotta massa ossea nel nostro Paese supera i 5 milioni, ma è destinato ad aumentare in quanto alla elevata prevalenza attuale andrà ad aggiungersi il progressivo invecchiamento della popolazione. Di conseguenza, la terapia antiosteoporotica, già molto frequente e diffusa, verrà utilizzata da un numero sempre maggiore di soggetti.Oltre all’osteoporosi postmenopausale e senile esistono poi forme di osteoporosi secondaria a malattie specifiche (iperparatiroidismo, morbo di Cushing, celiachia, insufficienza gonadica e renale ecc.) o iatrogene (uso di corticosteroidi, immunosoppressori ecc.).
Malattie reumatiche. In alcuni casi le patologie reumatiche sono direttamente associate e responsabili dell’erosione ossea (artrite reumatoide, artrosi erosiva ecc.), mentre in altri il quadro è secondario alla condizione sistemica o alla terapia impiegata.
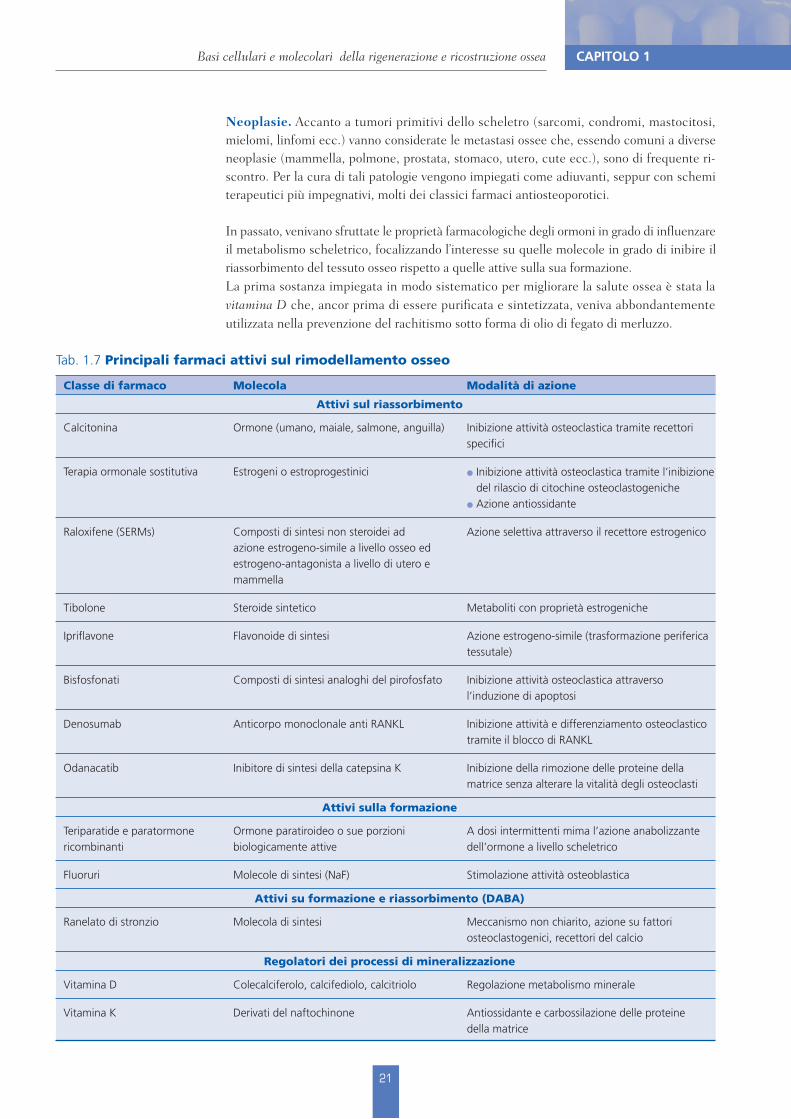
Tab. 1.7 Principali farmaci attivi sul rimodellamento osseo
Classe di farmaco Molecola Modalità di azione
Attivi sul riassorbimento
Calcitonina Ormone (umano, maiale, salmone, anguilla) Inibizione attività osteoclastica tramite recettori specifici
Terapia ormonale sostitutiva Estrogeni o estroprogestinici ! Inibizione attività osteoclastica tramite l’inibizione del rilascio di citochine osteoclastogeniche
! Azione antiossidante
Raloxifene (SERMs) Composti di sintesi non steroidei ad azione estrogeno-simile a livello osseo ed estrogeno-antagonista a livello di utero e mammella
Azione selettiva attraverso il recettore estrogenico
Tibolone Steroide sintetico Metaboliti con proprietà estrogeniche
Ipriflavone Flavonoide di sintesi Azione estrogeno-simile (trasformazione periferica tessutale)
Bisfosfonati Composti di sintesi analoghi del pirofosfato Inibizione attività osteoclastica attraverso l’induzione di apoptosi
Denosumab Anticorpo monoclonale anti RANKL Inibizione attività e differenziamento osteoclastico tramite il blocco di RANKL
Odanacatib Inibitore di sintesi della catepsina K Inibizione della rimozione delle proteine della matrice senza alterare la vitalità degli osteoclasti
Attivi sulla formazione
Teriparatide e paratormone ricombinanti
Ormone paratiroideo o sue porzioni biologicamente attive
A dosi intermittenti mima l’azione anabolizzante dell’ormone a livello scheletrico
Fluoruri Molecole di sintesi (NaF) Stimolazione attività osteoblastica
Attivi su formazione e riassorbimento (DABA)
Ranelato di stronzio Molecola di sintesi Meccanismo non chiarito, azione su fattori osteoclastogenici, recettori del calcio
Regolatori dei processi di mineralizzazione
Vitamina D Colecalciferolo, calcifediolo, calcitriolo Regolazione metabolismo minerale
Vitamina K Derivati del naftochinone Antiossidante e carbossilazione delle proteine della matrice
21
Basi cellulari e molecolari della rigenerazione e ricostruzione ossea CAPITOLO 1
Neoplasie. Accanto a tumori primitivi dello scheletro (sarcomi, condromi, mastocitosi, mielomi, linfomi ecc.) vanno considerate le metastasi ossee che, essendo comuni a diverse neoplasie (mammella, polmone, prostata, stomaco, utero, cute ecc.), sono di frequente ri-scontro. Per la cura di tali patologie vengono impiegati come adiuvanti, seppur con schemi terapeutici più impegnativi, molti dei classici farmaci antiosteoporotici.
In passato, venivano sfruttate le proprietà farmacologiche degli ormoni in grado di influenzare il metabolismo scheletrico, focalizzando l’interesse su quelle molecole in grado di inibire il riassorbimento del tessuto osseo rispetto a quelle attive sulla sua formazione.La prima sostanza impiegata in modo sistematico per migliorare la salute ossea è stata la vitamina D che, ancor prima di essere purificata e sintetizzata, veniva abbondantemente utilizzata nella prevenzione del rachitismo sotto forma di olio di fegato di merluzzo.

22
Basi cellulari e molecolari della rigenerazione e ricostruzione osseaCAPITOLO 1
Il primo farmaco specifico per l’osteoporosi è stata la calcitonina, per la capacità di bloccare selettivamente l’attività osteoclastica. Contestualmente, sono stati valorizzati i benefici in termini di massa ossea e riduzione delle fratture associati alla terapia ormonale sostitutiva. At-tività simil-estrogenica a livello scheletrico è stata poi conseguita con l’impiego del raloxifene, la prima molecola della nuova classe dei modulatori selettivi del recettore estrogenico (SERMs) in grado di regolare le risposte recettoriali a livello tessutale. Nel caso specifico del raloxifene, l’effetto è estrogeno-agonista a livello osseo ed estrogeno-antagonista a livello mammario.Dopo gli agenti antiosteolitici inibitori dell’attività osteoclastica, sono giunti sul mercato farmaci ad azione anabolizzante elettiva quali il teriparatide, corrispondente dalla sequenza N-terminale 1-43 del paratormone, e il PTH ricombinante rappresentato dall’intera molecola ormonale (aa 1-84). Questi vengono iniettati nel sottocute ove, in breve tempo, la molecola proteica viene inattiva-ta consentendole così di svolgere solo la parte iniziale, anabolica appunto, dell’azione ormona-le, eliminando invece l’effetto catabolico che si manifesta a distanza dalla somministrazione.Un’altra classe di farmaci attivi sull’osso è rappresentata dai DABA (Dual Action Bone Agents, agenti ossei a doppia azione), di cui è al momento commercializzato solo lo stronzio ranelato che, di fatto, disaccoppia il processo di rimodellamento scheletrico inibendo l’attività rias-sorbitiva e stimolando contestualmente quella formativa (Tab. 1.7).Nel paragrafo che segue saranno invece discussi più in dettaglio agenti selettivamente an-tiosteolitici quali i bisfosfonati (BP), in assoluto i farmaci per l’osteoporosi più utilizzati, e il Denosumab, che rappresenta il primo anticorpo monoclonale impiegato in questa patologia. Oltre che per l’ampia diffusione dei bisfosfonati, il focalizzare l’attenzione su queste due classi di farmaci trova ragione nelle problematiche e speculazioni circa l’effetto a livello orale del loro impiego ad alte dosi.
DenosumabIl sistema RANK/RANKL/OPG è il principale regolatore della formazione, dell’attivazione e della sopravvivenza degli osteoclasti. Come descritto, RANKL lega RANK sulla superficie di preosteoclasti e osteoclasti, stimolandone e promuovendone il differenziamento e l’atti-vazione. Di conseguenza, RANKL rappresenta un ottimo bersaglio terapeutico per malattie associate ad aumentato riassorbimento osseo. Il Denosumab® è un anticorpo monoclonale umano che lega RANKL con alta affinità e specificità, con un meccanismo simile a quello dell’osteoprotegerina. Questa molecola appartiene alla categoria dei farmaci biologici; è stata recentemente ap-provata dall’EMEA e immessa nel mercato europeo e italiano. In studi condotti su animali ha dimostrato di poter regolare il metabolismo dell’osso, inibendo selettivamente l’attività osteoclastica. Alla fase sperimentale è seguito uno studio clinico controllato che ha coinvolto un numero elevatissimo di pazienti, tra cui donne in postmenopausa, maschi con carcinoma prostatico trattati con inibitori degli androgeni, e pazienti con metastasi scheletriche. I risultati relativi ai primi 3 anni di studio ne hanno confermato l’efficacia clinica in termini di aumento di massa ossea e riduzione dell’incidenza di fratture.Per quanto attiene la sicurezza di impiego non sono stati rilevati effetti collaterali importanti, non c’è accumulo della molecola e la sua farmacocinetica non si modifica nel tempo. Il rischio di insorgenza di osteonecrosi della mandibola, data l’enfasi posta negli ultimi anni su questa problematica, è stato affrontato con particolare attenzione nella fase di estensione dello studio, al momento attuale in corso.
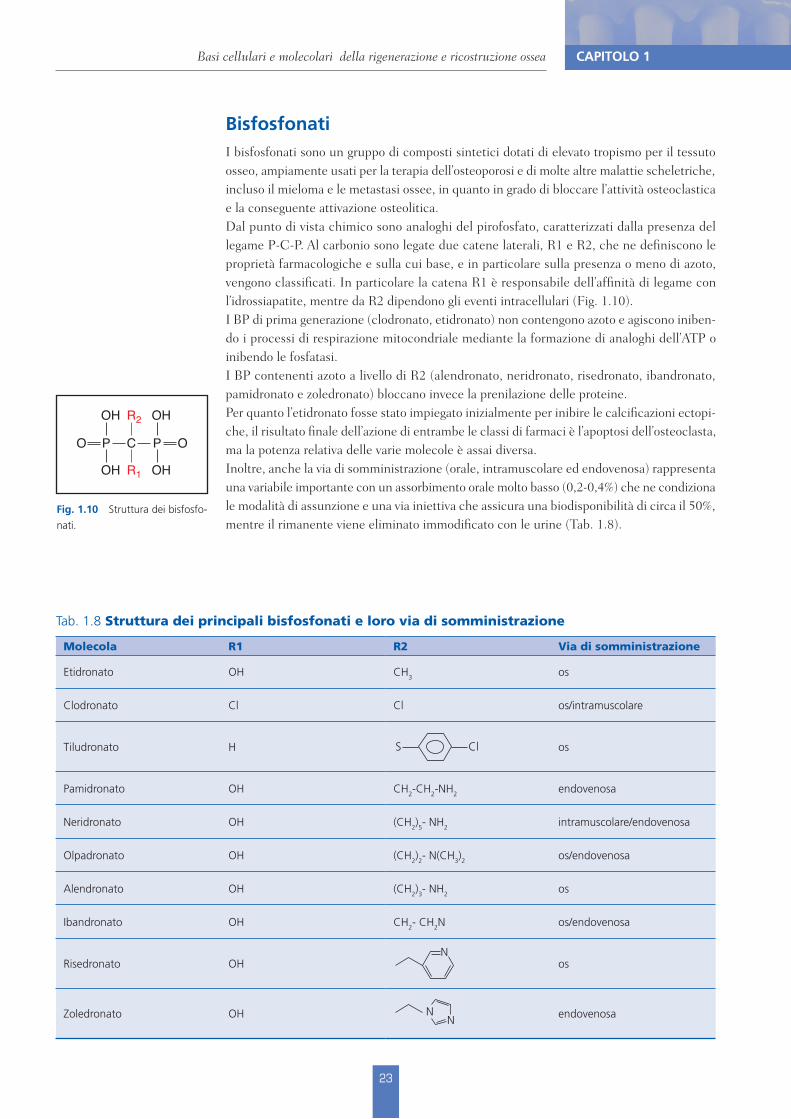
Tab. 1.8 Struttura dei principali bisfosfonati e loro via di somministrazione
Molecola R1 R2 Via di somministrazione
Etidronato OH CH3 os
Clodronato Cl Cl os/intramuscolare
Tiludronato H os
Pamidronato OH CH2-CH2-NH2 endovenosa
Neridronato OH (CH2)5- NH2 intramuscolare/endovenosa
Olpadronato OH (CH2)2- N(CH3)2 os/endovenosa
Alendronato OH (CH2)3- NH2 os
Ibandronato OH CH2- CH2N os/endovenosa
Risedronato OH os
Zoledronato OH endovenosa
S Cl
N
NN
23
Basi cellulari e molecolari della rigenerazione e ricostruzione ossea CAPITOLO 1
OH
PO
R2 OH
OH R1 OH
C OP
Fig. 1.10 Struttura dei bisfosfo-nati.
BisfosfonatiI bisfosfonati sono un gruppo di composti sintetici dotati di elevato tropismo per il tessuto osseo, ampiamente usati per la terapia dell’osteoporosi e di molte altre malattie scheletriche, incluso il mieloma e le metastasi ossee, in quanto in grado di bloccare l’attività osteoclastica e la conseguente attivazione osteolitica.Dal punto di vista chimico sono analoghi del pirofosfato, caratterizzati dalla presenza del legame P-C-P. Al carbonio sono legate due catene laterali, R1 e R2, che ne definiscono le proprietà farmacologiche e sulla cui base, e in particolare sulla presenza o meno di azoto, vengono classificati. In particolare la catena R1 è responsabile dell’affinità di legame con l’idrossiapatite, mentre da R2 dipendono gli eventi intracellulari (Fig. 1.10).I BP di prima generazione (clodronato, etidronato) non contengono azoto e agiscono iniben-do i processi di respirazione mitocondriale mediante la formazione di analoghi dell’ATP o inibendo le fosfatasi.I BP contenenti azoto a livello di R2 (alendronato, neridronato, risedronato, ibandronato, pamidronato e zoledronato) bloccano invece la prenilazione delle proteine.Per quanto l’etidronato fosse stato impiegato inizialmente per inibire le calcificazioni ectopi-che, il risultato finale dell’azione di entrambe le classi di farmaci è l’apoptosi dell’osteoclasta, ma la potenza relativa delle varie molecole è assai diversa.Inoltre, anche la via di somministrazione (orale, intramuscolare ed endovenosa) rappresenta una variabile importante con un assorbimento orale molto basso (0,2-0,4%) che ne condiziona le modalità di assunzione e una via iniettiva che assicura una biodisponibilità di circa il 50%, mentre il rimanente viene eliminato immodificato con le urine (Tab. 1.8).
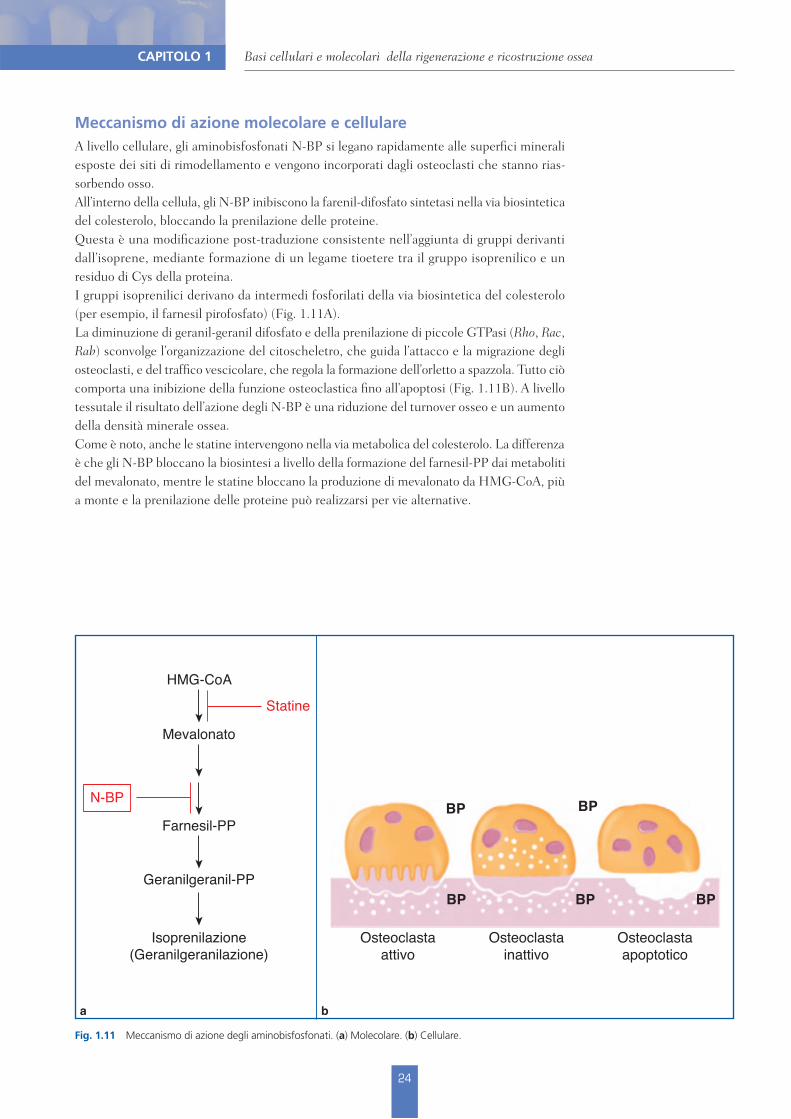
Fig. 1.11 Meccanismo di azione degli aminobisfosfonati. (a) Molecolare. (b) Cellulare.
HMG-CoA
MevalonatoStatine
N-BPFarnesil-PP
Geranilgeranil-PP
Isoprenilazione(Geranilgeranilazione)
BP BP
BP
Osteoclastaattivo
Osteoclastainattivo
Osteoclastaapoptotico
BP BP
a b
24
Basi cellulari e molecolari della rigenerazione e ricostruzione osseaCAPITOLO 1
Meccanismo di azione molecolare e cellulareA livello cellulare, gli aminobisfosfonati N-BP si legano rapidamente alle superfici minerali esposte dei siti di rimodellamento e vengono incorporati dagli osteoclasti che stanno rias-sorbendo osso. All’interno della cellula, gli N-BP inibiscono la farenil-difosfato sintetasi nella via biosintetica del colesterolo, bloccando la prenilazione delle proteine. Questa è una modificazione post-traduzione consistente nell’aggiunta di gruppi derivanti dall’isoprene, mediante formazione di un legame tioetere tra il gruppo isoprenilico e un residuo di Cys della proteina. I gruppi isoprenilici derivano da intermedi fosforilati della via biosintetica del colesterolo (per esempio, il farnesil pirofosfato) (Fig. 1.11A). La diminuzione di geranil-geranil difosfato e della prenilazione di piccole GTPasi (Rho, Rac, Rab) sconvolge l’organizzazione del citoscheletro, che guida l’attacco e la migrazione degli osteoclasti, e del traffico vescicolare, che regola la formazione dell’orletto a spazzola. Tutto ciò comporta una inibizione della funzione osteoclastica fino all’apoptosi (Fig. 1.11B). A livello tessutale il risultato dell’azione degli N-BP è una riduzione del turnover osseo e un aumento della densità minerale ossea.Come è noto, anche le statine intervengono nella via metabolica del colesterolo. La differenza è che gli N-BP bloccano la biosintesi a livello della formazione del farnesil-PP dai metaboliti del mevalonato, mentre le statine bloccano la produzione di mevalonato da HMG-CoA, più a monte e la prenilazione delle proteine può realizzarsi per vie alternative.

MODULO I
25
Osteonecrosi da bisfosfonati
L’osteonecrosi della mandibola (ONJ) è una patologia progressiva con scarsa propensione alla guarigione, associata ad alcune condizioni tra cui la radio e la chemioterapia e, dal 2003, anno della prima segnalazione, anche al trattamento con aminobisfosfonati. In tutto il mondo sono stati segnalati circa 1000 casi di ONJ, ma non sem-pre è facile stabilire una diretta relazione con l’uso di N-BP, per la concomitanza di altre condizioni patologiche o predisponenti e per il tempo che intercorre tra l’assunzione del farmaco e l’insorgenza della malattia. Dei casi descritti, oltre il 90% riguarda pazienti onco-logici in terapia con bisfosfonati per via iniettiva (prevalentemente pamidronato e zoledronato) a dosaggi assai superiori a quelli in uso per altre patologie.In Italia sono attualmente circa 1.200.000 i soggetti trattati con bisfosfonati per patologie ossee non neoplastiche; di questi, oltre un terzo assume non-aminobisfosfonati la cui associazione con l’ONJ è ancora dibattuta. L’incidenza annua di ONJ per i trattamenti per via orale è di circa 1/100.000: è evidente la sproporzione tra il reale impatto di questa patologia (10-15 casi attesi/anno) e il clamore che essa suscita. L’osteonecrosi da bisfosfonati colpisce le ossa della cavità orale per alcune peculiarità: si tratta di segmenti ossei a ele-vato tasso di rimodellamento posti inoltre in una condizione di facile contaminazione con la flora batterica normalmente presente. L’alto turnover, causato da movimento dentario, malattia parodontale e traumi iatrogeni e non, determina il richiamo-fissazione dei bisfosfo-nati che vengono quindi sequestrati in questo distretto. Successivi interventi odontoiatrici (estrazioni, posizionamento di impianti, altra chirurgia) o altri insulti traumatici (protesi incongrue) come anche una malattia parodontale attiva determinano una ripresa del turno-ver; come conseguenza, gli osteoclasti liberano i farmaci inglobati nella matrice ossea che a loro volta possono esplicare il loro effetto terapeutico volto all’inibizione dell’osteoclasta stesso, alterando il normale processo di riparazione della lesione.La presenza costante di osteomielite nell’ONJ è una caratteristica che la differenzia completamente dalle altre forme di osteonecrosi: il germe più frequentemente isolato è l’Actinomyces, responsabile dell’alitosi che rappresenta uno dei sintomi predominanti del quadro clinico.L’esatto meccanismo patogenetico responsabile dell’instaurarsi della lesione non è attualmente noto. Le ipotesi avanzate riguardano un’iniziale azione tossica dei bisfosfonati a livello della mucosa orale, un’alterazione dell’angiogenesi in grado di modificare il turnover osseo, eventuali terapie concomitanti e stato fisico del soggetto. Successivamente compare la caratteristica esposizione di osso in cavità orale che, per poter porre diagnosi, deve persistere per oltre 8 settimane. La mancata guarigione della lesione, indipendentemente da una terapia antibiotico-mirata, evolve poi verso una forma di osteomielite cronica nella quale intervengono poi una serie di altri fattori non necessariamente legati al tipo di farmaco.La complessità della patologia rende fondamentale nella pratica odon-toiatrica l’esecuzione di un corretto screening anamnestico in grado di individuare i pazienti a rischio. Ad esempio, i pazienti oncologici in terapia con zoledronato o pamidronato ad alte dosi non possono e non devono essere considerati e trattati alla stregua dei pazienti che assumono aminobisfosfonati per via orale, a dosaggi annuali di
almeno un ordine di grandezza inferiore. Non dimentichiamo inoltre che i BP rappresentano la classe di farmaci più utilizzata per prevenire le fratture da fragilità. Rivestono in ogni caso un ruolo fondamentale le manovre di prevenzione sia primaria, prima dell’inizio della terapia, che secondaria, accompagnata da controlli odontoiatrici frequenti. Nei pazienti in cui la terapia sia già in corso da tempo, ogni modalità chirurgica dovrebbe essere attentamente ponderata e possibilmente prevenuta. Quando non evitabile, per esempio un’estrazione dentale, occorre attenersi scrupolosamente ai protocolli consigliati. Da ultimo, va ricordato come proprio i bisfosfonati e in particolare l’alendronato e lo zoledronato, somministrati per os o in singolo bolo per via endovenosa, fossero stati inizialmente proposti nella chirur-gia ortopedica protesica dell’anca, proprio per la loro efficacia nella riduzione del rischio di allentamento protesico associato al processo infiammatorio nella zona peri-implantare. Anche nella pratica odon-toiatrica, l’utilizzo di impianti trattati con aminobisfosfonati, sebbene si tratti di casi isolati, ha dato risultati incoraggianti, evidenziando una maggiore superficie di contatto con osso neoformato rispetto ai controlli, ma il clamore suscitato dalle prime rilevazioni di ONJ ha condizionato negativamente il prosieguo degli studi clinici.
Prevenzione e terapiaNon esiste attualmente un trattamento definitivo e risolutivo per la cu-ra dell’osteonecrosi da bisfosfonati. Tuttavia sono stati formulati diversi protocolli terapeutici, che hanno una sempre maggior percentuale di successo e una sempre minore percentuale di recidive.È considerato oggi un successo clinico la chiusura della mucosa sull’e-sposizione ossea, in assenza di segni di infiammazione o infezione: questo è l’obiettivo di tutti gli attuali protocolli terapeutici.Il piano di trattamento è sempre impostato sulla base della diagnosi e della stadiazione. La diagnosi clinica è l’osservazione dell’esposizione ossea, con infiammazione e infezione dei tessuti circostanti associata a una sintomatologia dolorosa importante. L’insorgenza della malattia è subdola, con un osso scarsamente sanguinante, di colore bianca-stro. Con il progredire della lesione, l’osso assume una colorazione grigio-giallastra con una consistenza duro-gessosa, con infezione che si manifesta con pus o fistole. Di fondamentale importanza è la conferma radiografica della lesione con l’utilizzo della TC, la sola in grado di identificare anche le lesioni iniziali e di stabilire con precisione i confini della necrosi. L’uso della scintigrafia ossea, della tomografia a emissione di positroni, il monitoraggio serico del PTH e del CTX, supposto marker del turnover osseo, non sono ancora supportate da chiara evidenza scientifica. Tra le principali stadiazioni proposte, quella di McMahon è basata sia sul quadro clinico-sintomatologico sia sulle immagini radiografiche. In presenza di piccole lesioni con segni scarsi di infiammazione e infezione, è possibile applicare diverse terapie anche combinate tra loro. Si possono usare lavaggi quotidiani con soluzioni di metronidazolo o di clorexidina allo 0,12%. In tutti i casi è associata una terapia antibiotica mirata sui Gram negativi e anaerobi, utilizzando amoxicillina+acidoclavulanico e metronidazolo. In alcuni casi può essere utile un trattamento con strumenti ultrasonici.In presenza di lesioni importanti con dolore e infezione ma senza complicazioni quali fistole, lisi della corticale inferiore o fratture pato-
MODULO I Osteonecrosi da bisfosfonati
2626
logiche, oltre alla terapia antibiotica locale e per via generale, trova indicazione associare una terapia antidolorifica e ricorrere a una terapia chirurgica conservativa con rimozione di osso francamente necrotico.Per lesioni necrotiche che presentano invece delle complicanze si deve ricorrere alla terapia resettiva. Tuttavia va eseguita tenendo conto delle condizioni generali del paziente che spesso lo controindicano. È importante valutare il rapporto rischio-beneficio nell’utilizzo di que-sta terapia chirurgica e, in ogni caso, in letteratura si ritiene che si debba mantenere un approccio il più conservativo possibile.È stato recentemente introdotto l’uso di un fascio laser a bassa po-tenza (Low Level Laser Therapy, LLLT) sulle lesioni osteonecrotiche. Trova indicazione in lesioni di piccola dimensione o in quei casi in cui non si può intervenire chirurgicamente. Ha un effetto antalgico, antiedemigeno e antibatterico, consentendo la totale o parziale riepi-
telizzazione della lesione, ed è ripetibile e senza dolore per il paziente. La prevenzione è in grado di ottenere una drastica riduzione dell’inci-denza dell’osteonecrosi. Prima di iniziare una terapia con bisfosfonati a elevato dosaggio, una visita odontostomatologica è in grado di individuare lesioni infette o elementi che potrebbero dare problemi in futuro e di intervenire prima dell’inizio della cura. A terapia instaurata è indispensabile mantenere un’ottima igiene orale, riducendo il consu-mo di alcool e abolendo il fumo. È necessario controllare che i pazienti abbiano protesi congrue e non traumatizzanti la mucosa. Come già accennato bisogna distinguere pazienti in terapia con bisfosfonati per motivi oncologici da quelli in terapia per l’osteoporosi. Questi ultimi hanno un rischio molto più ridotto di manifestare la malattia: non emergono infatti controindicazioni assolute al trattamento chirurgico di pazienti che assumono bisfosfonati per via orale.
Tab. I.2 Protocolli terapeutici in pazienti con osteonecrosi dei mascellari in terapia con bisfosfonati (Arduino)
Tipo di lesione Aspetti clinici Terapia
Iniziale Dimensioni <2 cmAssenza o scarsa presenza di infiammazione e infezione
Lavaggi topici con metronidazolo, clorexidina o minociclina cloridratoTerapia antibiotica sistemicaStrumentazione con ultrasuoniLLLT
Avanzata Dimensioni >2 cmPresenza di doloreInfiammazione e pus
Lavaggi topici con metronidazolo, clorexidina e minociclina cloridratoTerapia antibiotica sistemicaLLLTTerapia antalgicaChirurgia conservativa
Complicata Dimensione <2 cmPresenza di doloreInfiammazione e pusPresenza di fistole oro-cutaneeComunicazioni oro-nasaliParestesiaLisi corticale inferioreFrattura mandibolare
Lavaggi topiciTerapia antibiotica sistemicaLLLTTerapia antalgicaChirurgia conservativaChirurgia recettiva
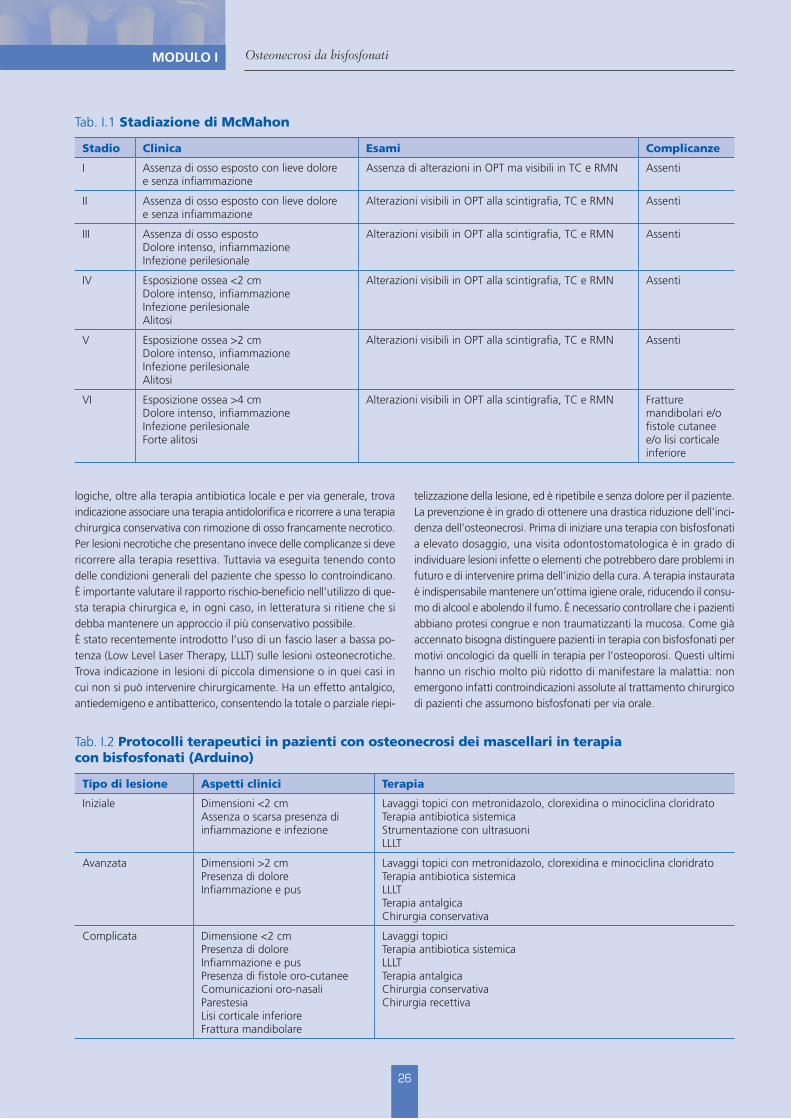
Tab. I.1 Stadiazione di McMahon
Stadio Clinica Esami Complicanze
I Assenza di osso esposto con lieve dolore e senza infiammazione
Assenza di alterazioni in OPT ma visibili in TC e RMN Assenti
II Assenza di osso esposto con lieve dolore e senza infiammazione
Alterazioni visibili in OPT alla scintigrafia, TC e RMN Assenti
III Assenza di osso espostoDolore intenso, infiammazioneInfezione perilesionale
Alterazioni visibili in OPT alla scintigrafia, TC e RMN Assenti
IV Esposizione ossea <2 cmDolore intenso, infiammazioneInfezione perilesionaleAlitosi
Alterazioni visibili in OPT alla scintigrafia, TC e RMN Assenti
V Esposizione ossea >2 cm Dolore intenso, infiammazioneInfezione perilesionaleAlitosi
Alterazioni visibili in OPT alla scintigrafia, TC e RMN Assenti
VI Esposizione ossea >4 cmDolore intenso, infiammazioneInfezione perilesionaleForte alitosi
Alterazioni visibili in OPT alla scintigrafia, TC e RMN Fratture mandibolari e/o fistole cutanee e/o lisi corticale inferiore

Tab. 1.9 Osteoporosi: geni candidati
Ormoni e recettori Citochine e fattori di crescita
Proteine regolatorie e strutturali
Recettore della vitamina D (VDR) IL-1/ IL-1ra Matrice ossea
Recettore degli estrogeni (ESR) IL-6 COLIA1
Recettore degli androgeni TGF-! Collagenasi
PTHr1/ PTH TNFRSF11 Osteocalcina
Recettore della calcitonina (CTR) SOST alfa-2-HS-glicoproteina
Calcium Sensing Receptor (CSR) BMP —
LRP5 PPAR# Altre
Aromatasi — Apolipoproteina E
27
Basi cellulari e molecolari della rigenerazione e ricostruzione ossea CAPITOLO 1
2727
Studi sulla genetica dell’ossoIl metabolismo osseo è estremamente complesso e molti sono i fattori che ne regolano l’andamento fisiologico e la patologia. Di conseguenza, il bilancio scheletrico è il risultato dell’azione combinata di molti geni e della loro interazione con l’ambiente.Il deterioramento della massa ossea che si verifica con l’avanzare dell’età ha una eziologia multigenica e polifattoriale ed è difficile valutare in un anziano il contributo individuale di ciascuno di questi componenti.D’altra parte, numerosi studi effettuati su gemelli nonché indagini epidemiologiche condotte su un amplissimo numero di soggetti hanno messo in evidenza e confermato l’importanza della familiarità (ereditarietà) nel determinismo del picco di massa ossea, del suo deterioramento e della suscettibilità alle fratture.Nel corso degli anni Novanta, in seguito alla disponibilità di nuove, veloci e semplici tec-nologie per lo studio del DNA, è stata dedicata molta attenzione alla ricerca di un gene che potesse essere ritenuto il principale responsabile della perdita di massa ossea. Sono stati indagati molti geni candidati, scelti fra quelli codificanti per gli agenti coinvolti nella regola-zione metabolica: ormoni e loro recettori, citochine e fattori di crescita, proteine strutturali o funzionali (Tab. 1.9).La grande produzione scientifica che ne è derivata non ha portato a un reale avanzamento delle conoscenze. Sono state infatti individuate delle varianti di sequenza che sembravano associate con la variazione della massa ossea, anche se in modo spesso debole e controverso.Si è trattato nella maggioranza dei casi di polimorfismi, cioè di varianti che hanno nella po-polazione una frequenza elevata e che pertanto ben difficilmente possono rappresentare una condizione di reale svantaggio o vantaggio per l’individuo portatore della mutazione. Inoltre, spesso tali polimorfismi si riscontrano a livello intronico e quindi anche il loro significato biologico è di difficile interpretazione.Di un certo rilievo si è dimostrata una variante nel sito di legame Sp1 del promotore del gene codificante per la catena "1 del collagene di tipo I (COLIA1) che è risultata associata a un maggiore rischio di frattura, come conseguenza di una struttura del collagene meno funzionale, in quanto contenente un eccesso di catene "1 ("1-omotrimeri anziché etero-trimeri "1/"1/"2). È importante notare che mutazioni in siti diversi di questo stesso gene sono associate alla patologia osteogenesi imperfecta, caratterizzata da deficienze scheletriche che possono portare a morte già in epoca prenatale.
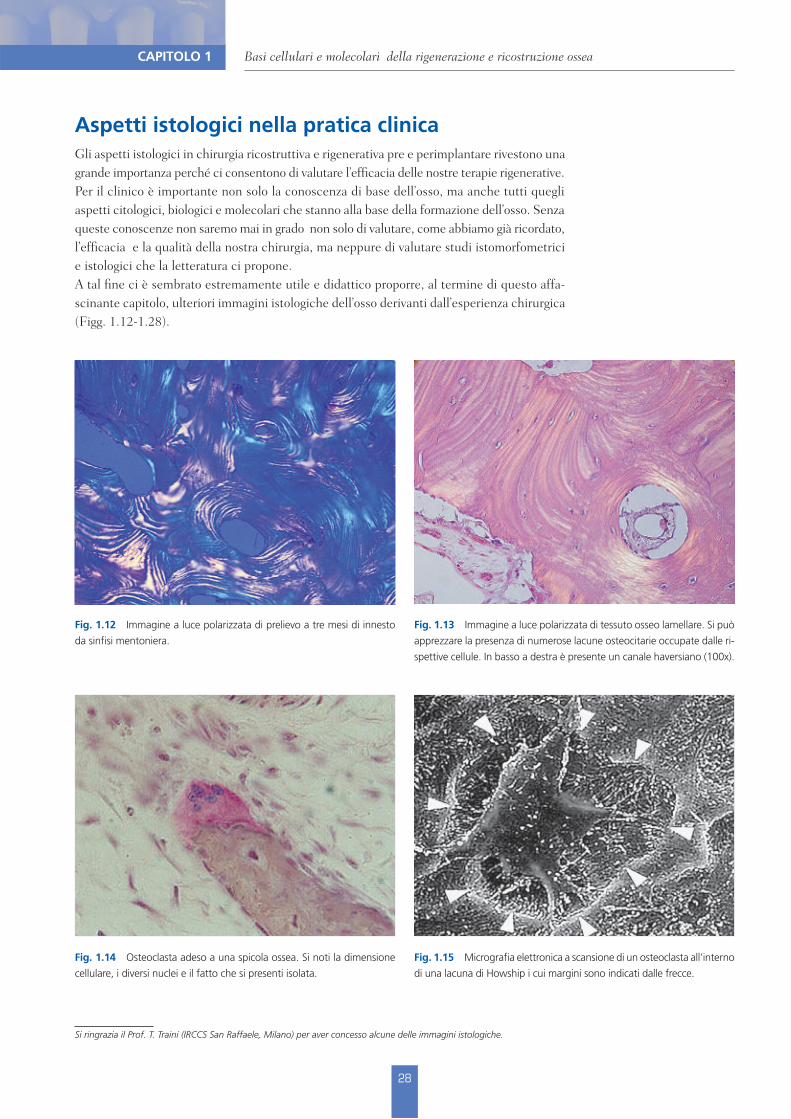
Fig. 1.13 Immagine a luce polarizzata di tessuto osseo lamellare. Si può apprezzare la presenza di numerose lacune osteocitarie occupate dalle ri-spettive cellule. In basso a destra è presente un canale haversiano (100x).
Fig. 1.14 Osteoclasta adeso a una spicola ossea. Si noti la dimensione cellulare, i diversi nuclei e il fatto che si presenti isolata.
Fig. 1.15 Micrografia elettronica a scansione di un osteoclasta all’interno di una lacuna di Howship i cui margini sono indicati dalle frecce.
Fig. 1.12 Immagine a luce polarizzata di prelievo a tre mesi di innesto da sinfisi mentoniera.
Si ringrazia il Prof. T. Traini (IRCCS San Raffaele, Milano) per aver concesso alcune delle immagini istologiche.
28
Basi cellulari e molecolari della rigenerazione e ricostruzione osseaCAPITOLO 1
Aspetti istologici nella pratica clinicaGli aspetti istologici in chirurgia ricostruttiva e rigenerativa pre e perimplantare rivestono una grande importanza perché ci consentono di valutare l’efficacia delle nostre terapie rigenerative.Per il clinico è importante non solo la conoscenza di base dell’osso, ma anche tutti quegli aspetti citologici, biologici e molecolari che stanno alla base della formazione dell’osso. Senza queste conoscenze non saremo mai in grado non solo di valutare, come abbiamo già ricordato, l’efficacia e la qualità della nostra chirurgia, ma neppure di valutare studi istomorfometrici e istologici che la letteratura ci propone.A tal fine ci è sembrato estremamente utile e didattico proporre, al termine di questo affa-scinante capitolo, ulteriori immagini istologiche dell’osso derivanti dall’esperienza chirurgica (Figg. 1.12-1.28).
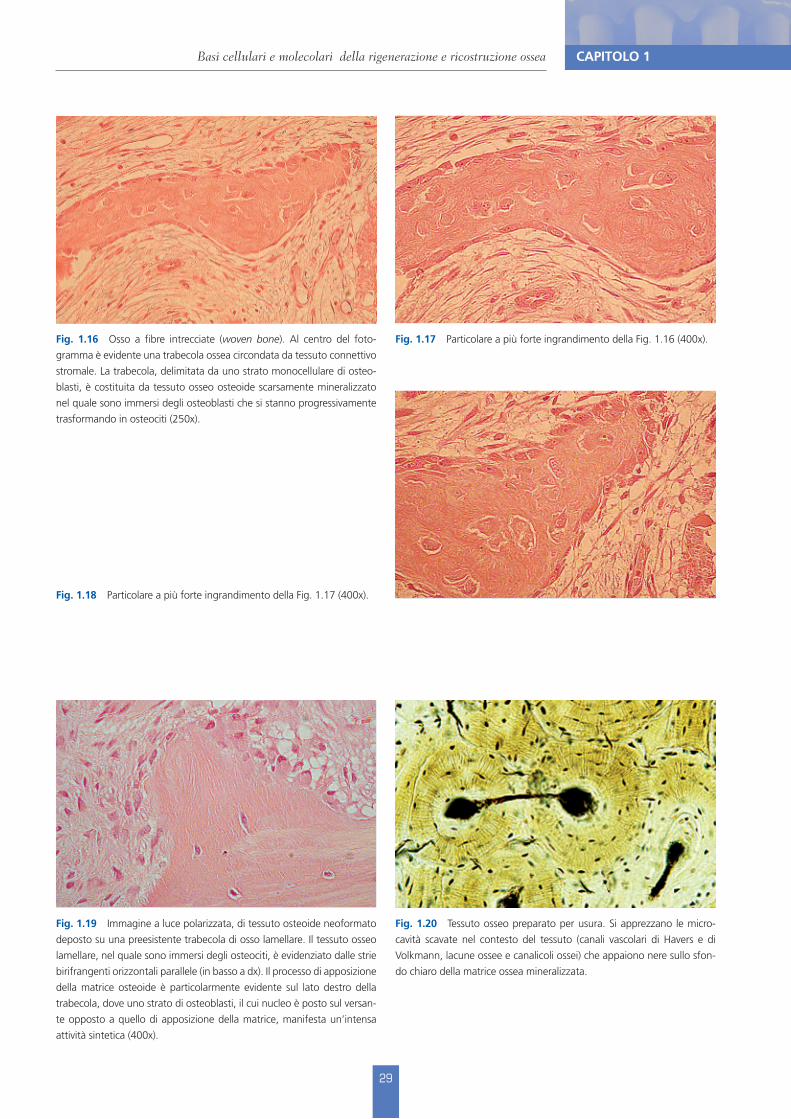
Fig. 1.20 Tessuto osseo preparato per usura. Si apprezzano le micro-cavità scavate nel contesto del tessuto (canali vascolari di Havers e di Volkmann, lacune ossee e canalicoli ossei) che appaiono nere sullo sfon-do chiaro della matrice ossea mineralizzata.
Fig. 1.19 Immagine a luce polarizzata, di tessuto osteoide neoformato deposto su una preesistente trabecola di osso lamellare. Il tessuto osseo lamellare, nel quale sono immersi degli osteociti, è evidenziato dalle strie birifrangenti orizzontali parallele (in basso a dx). Il processo di apposizione della matrice osteoide è particolarmente evidente sul lato destro della trabecola, dove uno strato di osteoblasti, il cui nucleo è posto sul versan-te opposto a quello di apposizione della matrice, manifesta un’intensa attività sintetica (400x).
Fig. 1.18 Particolare a più forte ingrandimento della Fig. 1.17 (400x).
Fig. 1.17 Particolare a più forte ingrandimento della Fig. 1.16 (400x).Fig. 1.16 Osso a fibre intrecciate (woven bone). Al centro del foto-gramma è evidente una trabecola ossea circondata da tessuto connettivo stromale. La trabecola, delimitata da uno strato monocellulare di osteo-blasti, è costituita da tessuto osseo osteoide scarsamente mineralizzato nel quale sono immersi degli osteoblasti che si stanno progressivamente trasformando in osteociti (250x).
29
Basi cellulari e molecolari della rigenerazione e ricostruzione ossea CAPITOLO 1
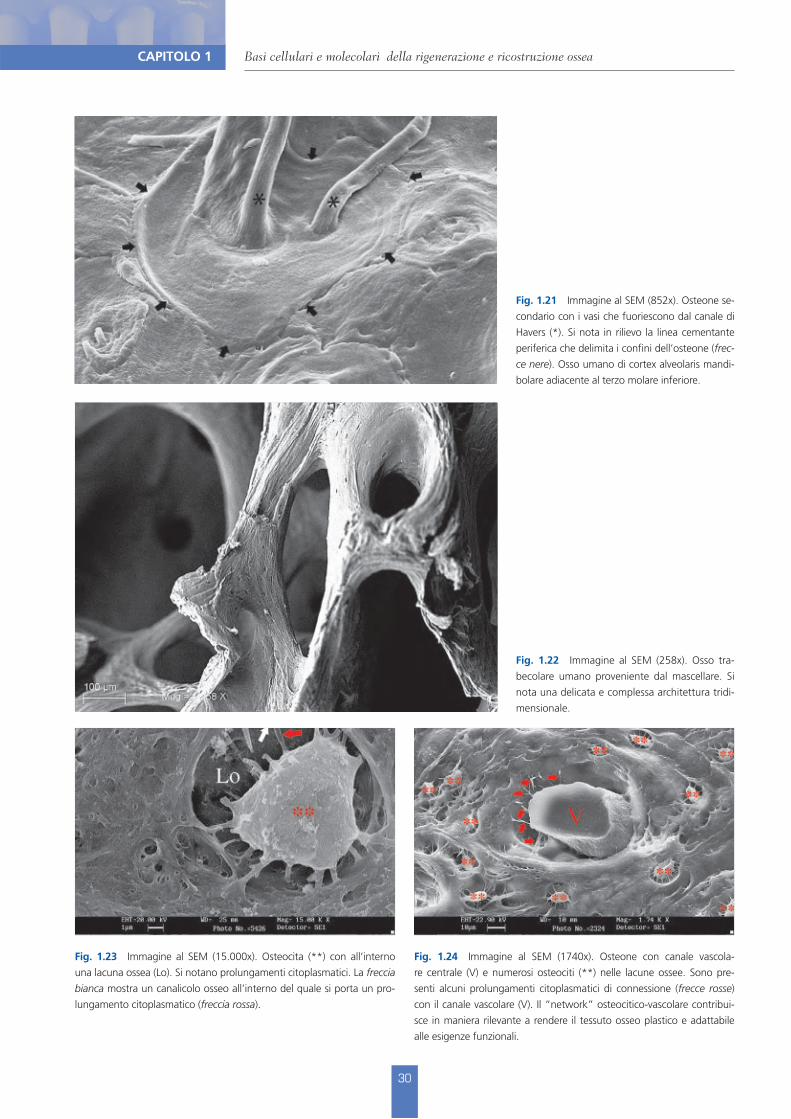
Fig. 1.23 Immagine al SEM (15.000x). Osteocita (**) con all’interno una lacuna ossea (Lo). Si notano prolungamenti citoplasmatici. La freccia bianca mostra un canalicolo osseo all’interno del quale si porta un pro-lungamento citoplasmatico (freccia rossa).
Fig. 1.21 Immagine al SEM (852x). Osteone se-condario con i vasi che fuoriescono dal canale di Havers (*). Si nota in rilievo la linea cementante periferica che delimita i confini dell’osteone (frec-ce nere). Osso umano di cortex alveolaris mandi-bolare adiacente al terzo molare inferiore.
Fig. 1.22 Immagine al SEM (258x). Osso tra-becolare umano proveniente dal mascellare. Si nota una delicata e complessa architettura tridi-mensionale.
Fig. 1.24 Immagine al SEM (1740x). Osteone con canale vascola-re centrale (V) e numerosi osteociti (**) nelle lacune ossee. Sono pre-senti alcuni prolungamenti citoplasmatici di connessione (frecce rosse) con il canale vascolare (V). Il “network” osteocitico-vascolare contribui- sce in maniera rilevante a rendere il tessuto osseo plastico e adattabile alle esigenze funzionali.
30
Basi cellulari e molecolari della rigenerazione e ricostruzione osseaCAPITOLO 1
Fig. 1.25 Immagine al microscopio ottico (200x). Tessuto osseo umano in un sito perimplantare. Le frecce mostrano alcuni osteoclasti in attività (blu di toluidina, fucsina acida).
Fig. 1.26 Immagine al microscopio con focale a scansione laser (CSLM) Area con intensa attività osteogenica. Il tessuto osseo (O) è rivestito da un rim di cellule osteoblastiche (**), con nuclei ben visibili, in attività osteosintetica. Alle spalle degli osteoblasti si nota un vaso di cospicue dimensioni (V) infarcito di globuli rossi. Il processo di deposizione di ma-trice osteoide richiede un apporto costante di ossigeno e sostanze per la funzione e il trofismo cellulare.
Fig. 1.27 (a) Micrografie elettroniche a trasmissione. (b) Micrografie elettroniche a scansione di tessuto osseo lamellare. È evidente l’orienta-mento variabile delle fibre collagene sottili (punte di frecce), più ricche di fibre, che nelle lamelle spesse, più ricche di componente minerale. a b
Fig. 1.28 Microradiografie di osso compatto umano preparato per usura provenienti da soggetti di varia età. Le cavità di riassorbi-mento appaiono nere mentre gli osteoni tanto più chiari quanto maggiore è il loro grado di mine-ralizzazione. (a) Soggetto giovane. (b) Soggetto adulto. (c) Soggetto anziano. a b c
31
Basi cellulari e molecolari della rigenerazione e ricostruzione ossea CAPITOLO 1

“This page intentionally left blank"

33
© 2
012
Els
evie
r Srl.
Tut
ti i d
iritt
i ris
erva
ti.
La corretta diagnosi dell’atrofia è alla base di un adeguato piano di trattamento, specifico per ogni singolo paziente.Le caratteristiche del difetto osseo sono determinate dall’entità e dalle modalità con le quali si è verificato (Figg. 2.1-2.3).
Cause del riassorbimento osseoLa principale causa di riassorbimento osseo è la perdita precoce degli elementi dentari. L’e-dentulia colpisce soprattutto le persone di età avanzata. In questi pazienti si ha un riassorbi-mento dapprima dell’osso alveolare e successivamente dell’osso basale. Tale riassorbimento è influenzato:! dalla mancanza di stimolazione meccanica conseguente alla progressiva perdita degli
elementi dentari! dalla progressiva erosione indotta dall’azione traumatica sui tessuti di protesi mobili tra-
dizionali! dall’alterazione della dinamica masticatoria che porta a un diverso trofismo delle masse
muscolari del complesso maxillo-facciale.
Il riassorbimento osseo crestale in senso verticale si stabilizza in media a circa 0,1 mm all’an-no nell’adulto, con ampie variazioni da individuo a individuo, con una compromissione del microcircolo vascolare che ne riduce la mineralizzazione.La mancanza di stimolazione funzionale del tessuto osseo residuo, che si ha in seguito alla perdita degli elementi dentari, modifica profondamente l’osso alveolare dal punto di vista strutturale. I cambiamenti più importanti sono la diminuzione della trabecolatura dell’osso residuo e una riduzione della densità dell’osso residuo stesso.Non ci sono studi che documentino come le alterazioni sistemiche del metabolismo osseo, che possono manifestarsi negli individui in età avanzata, incidano in maniera significativa sull’involuzione atrofica dei mascellari.
Modalità del riassorbimento osseoEdentulie singoleNelle edentulie singole e intercalari il riassorbimento osseo si manifesta sul piano trasversale soprattutto a carico del versante vestibolare, dove la corticale è più sottile, meno resistente all’azione osteoclastica di rimaneggiamento ed è spesso compromessa da fenomeni infettivo-infiammatori.Le corticali linguale e palatale, con il loro spessore, oppongono maggior resistenza al progres-sivo riassorbimento osseo. Ne consegue una riduzione del volume osseo dapprima in senso trasversale e successivamente in altezza.
Diagnosi e piano di trattamentoD.A. Di Stefano, A. Cazzaniga, P. Azzolini, P. Arosio
CAPITOLO 2
















![L. Milani - MEDIBIOgosa del fiume Begawan [Trinil – Ngawi (Solo – attualmente Surakarta)] pochi re - sti fossili indiscutibilmente “umanoidi”: una calotta cranica completa,](https://img.pdfslide.tips/doc/110x75/60931be823a2bc1b6d229912/l-milani-medibio-gosa-del-fiume-begawan-trinil-a-ngawi-solo-a-attualmente.jpg)










