Embed Size (px)
Citation preview

Tema 7. Estructura y organización delADN
Genética CC. Mar 2005-06
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 2
Objetivos
•Examinar las propiedades que debetener el material hereditario.
•Comprender la estructura yorganización del material hereditario.
•Entender la replicación del ADN.

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 3
Material hereditario
• El material hereditario debe …– Contener de una forma estable la información
genética.– Transmitir la información fielmente.– Poder cambiar, permitiendo la variación,
adaptación y evolución.
• El material hereditario lo forman:– ¿Proteínas?– ¿Ácidos nucleicos (ADN y ARN)?
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 4
Búsqueda del material hereditario
1890 August Weismann propone que hay una sustancia en elnúcleo que controla las características de los individuos.
1900 Los cromosomas contienen la información hereditaria. Mástarde se comprueba que los cromosomas están hechos deácidos nucleicos (ADN y ARN) y proteínas.
1928 Experimento de transformación de Griffith.1944 Experimento de transformación de Avery, MacLeod y
McCarty.1953 Experimento del bacteriófago de Hershey y Chase.1953 Watson y Crick proponen la estructura en doble hélice del
ADN.1956 Gierer y Schramm demuestran que el ARN es el material
hereditario de algunos virus.1957 Fraenkel-Conrat y Singer confirman el experimento de
Gierer y Schramm.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 5
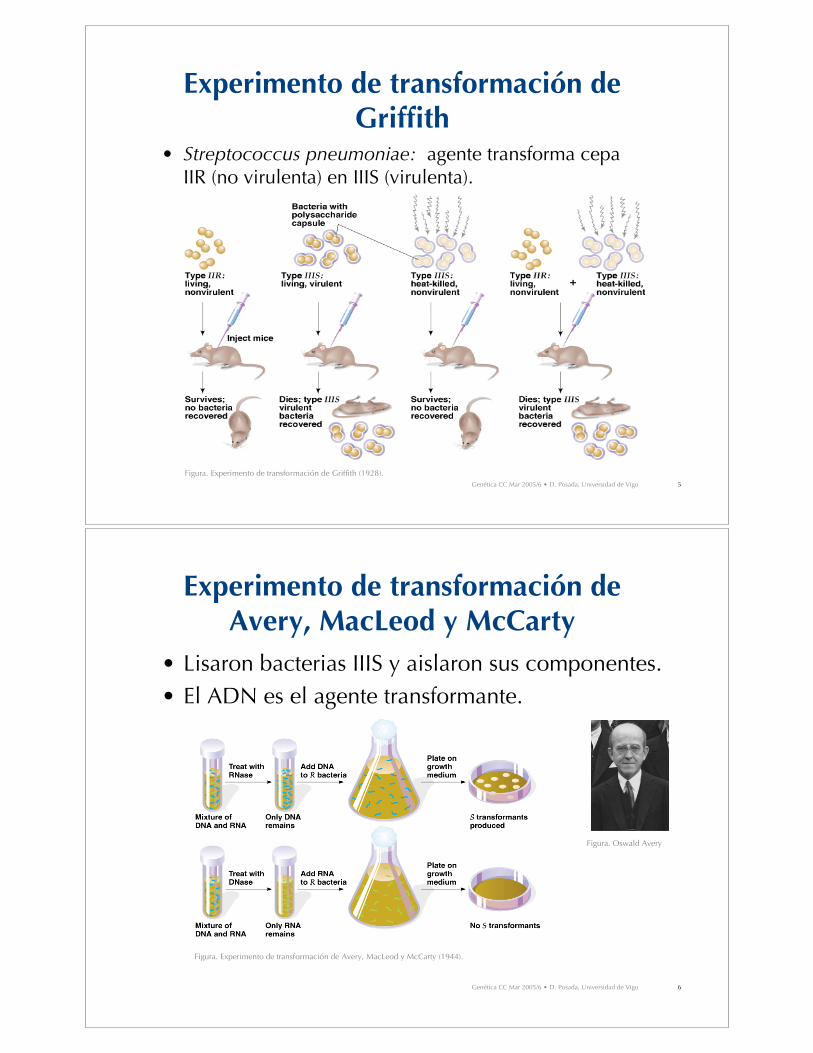
Experimento de transformación deGriffith
• Streptococcus pneumoniae: agente transforma cepaIIR (no virulenta) en IIIS (virulenta).
Figura. Experimento de transformación de Griffith (1928).
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 6
Experimento de transformación deAvery, MacLeod y McCarty
• Lisaron bacterias IIIS y aislaron sus componentes.• El ADN es el agente transformante.
Figura. Oswald Avery
Figura. Experimento de transformación de Avery, MacLeod y McCarty (1944).
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 7
Bacteriófagos
Figura. Fago T2.
Figura. Ciclo lítico del fago T2.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 8
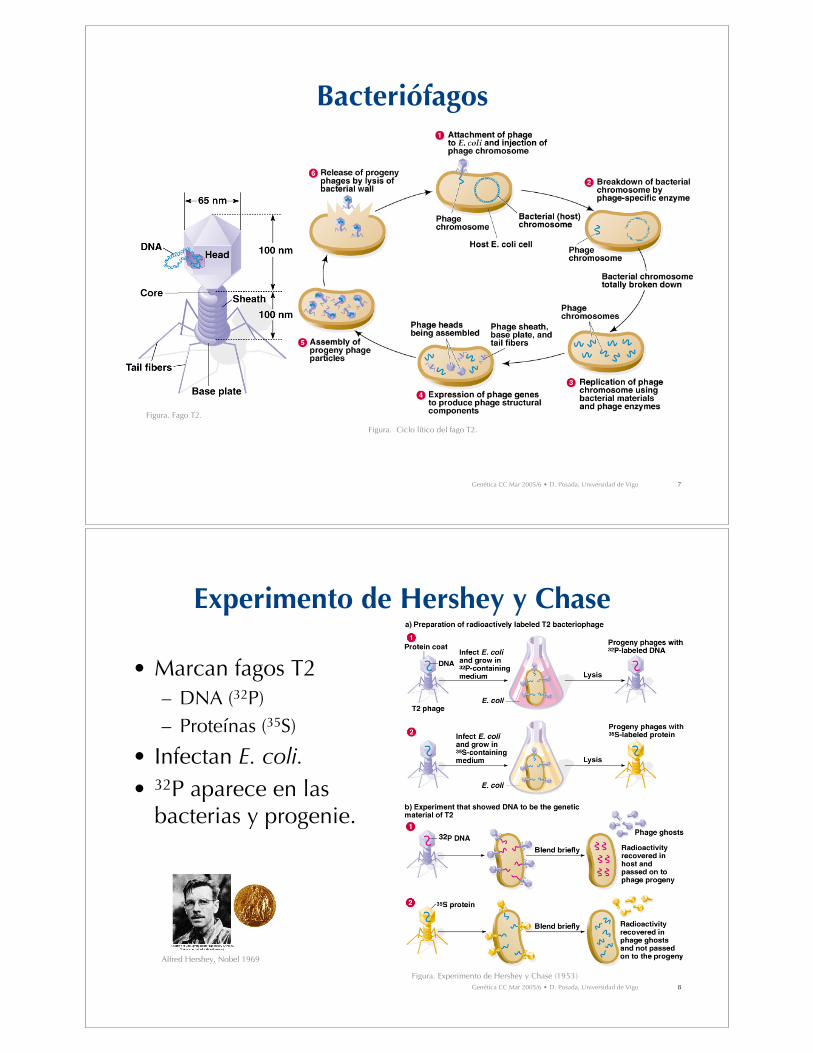
Experimento de Hershey y Chase
• Marcan fagos T2– DNA (32P)– Proteínas (35S)
• Infectan E. coli.
• 32P aparece en lasbacterias y progenie.
Figura. Experimento de Hershey y Chase (1953)
Alfred Hershey, Nobel 1969
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 9
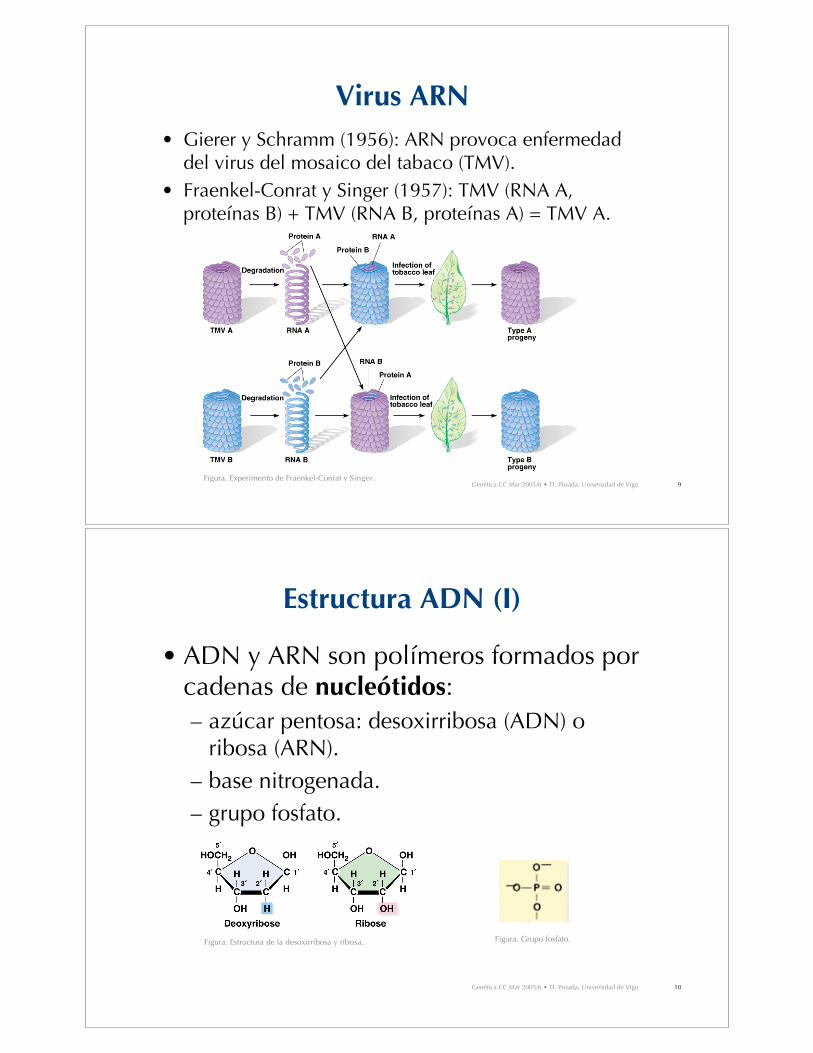
Virus ARN
• Gierer y Schramm (1956): ARN provoca enfermedaddel virus del mosaico del tabaco (TMV).
• Fraenkel-Conrat y Singer (1957): TMV (RNA A,proteínas B) + TMV (RNA B, proteínas A) = TMV A.
Figura. Experimento de Fraenkel-Conrat y Singer.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 10
Estructura ADN (I)
• ADN y ARN son polímeros formados porcadenas de nucleótidos:– azúcar pentosa: desoxirribosa (ADN) o
ribosa (ARN).– base nitrogenada.– grupo fosfato.
Figura. Estructura de la desoxirribosa y ribosa. Figura. Grupo fosfato.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 11
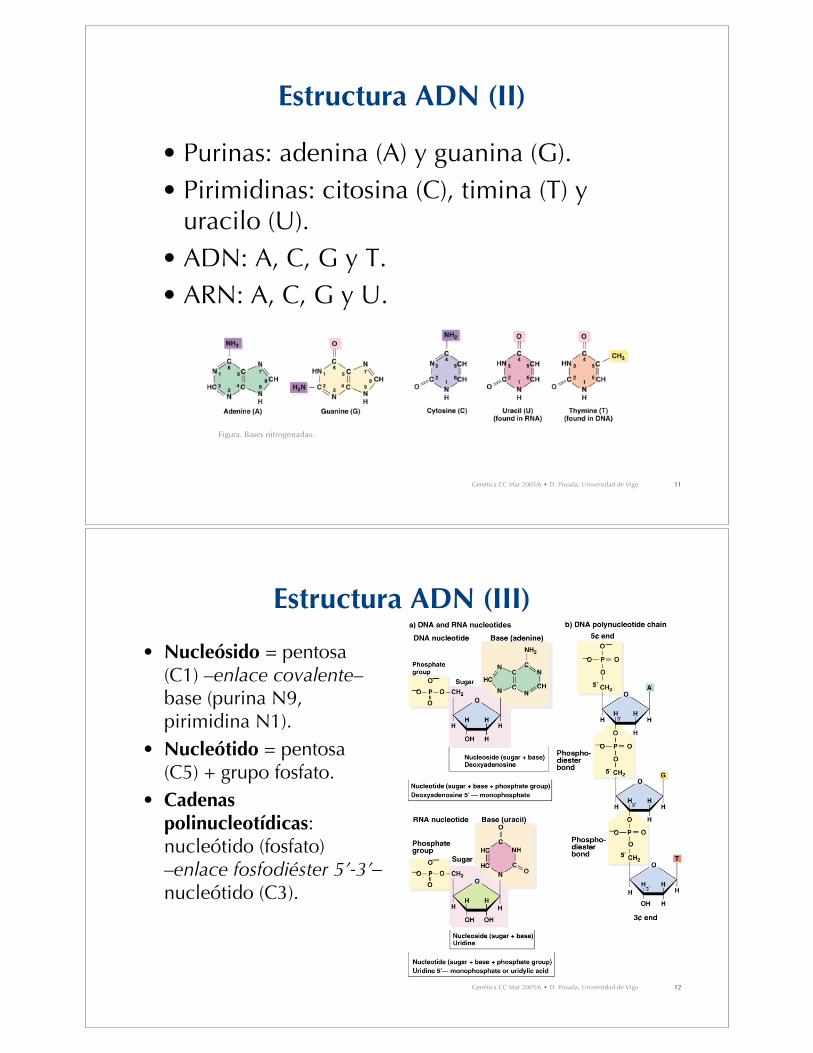
Estructura ADN (II)
• Purinas: adenina (A) y guanina (G).• Pirimidinas: citosina (C), timina (T) y
uracilo (U).• ADN: A, C, G y T.• ARN: A, C, G y U.
Figura. Bases nitrogenadas.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 12
Estructura ADN (III)
• Nucleósido = pentosa(C1) –enlace covalente–base (purina N9,pirimidina N1).
• Nucleótido = pentosa(C5) + grupo fosfato.
• Cadenaspolinucleotídicas:nucleótido (fosfato)–enlace fosfodiéster 5’-3’–nucleótido (C3).

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 13
Estructura del ADN
• Estructura descubierta por James Watsony Francis Crick en 1953.
• Basada en dos fuentes de informaciónprincipales:– Estudios de composición de bases de Erwin
Chargaff.– Estudios de difracción de rayos X de
Rosalind Franklin y Maurice Wilkins.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 14
Reglas de Chargaff
• Chargaff (1949):– (A + G) / (C + T) = 1– A/T = 1– G/C = 1– (A + T) / (G + C) = %GC
• Contenido GC varía a escala evolutiva:– 50-60% procariotas– 45% eucariotas

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 15
Difracción de rayos X
• Los rayos X son difractados de una forma particularsegún la disposición de los átomos en una molécula.
• Franklin y Wilkins (1953), ADN:– hélice no sencilla– diámetro de 2nm– dos regularidades de 0.34 y 3.4 nm a lo largo del
eje de la molécula
Figura. Difracción de rayos X.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 16
Modelo de Watson y Crick (1953)
1. Dos cadenas polinucleotídicas enlazadas en unadoble hélice dextrógira.
2. Las cadenas son antiparalelas, una está orientada endirección 5’-3’ y la otra en dirección 3’-5’.
3. El esqueleto azúcar-fosfato se sitúa en la parteexterna, mientras que las bases nitrogenadas seorientan perpendicularmente hacia el eje interno.
4. Las bases complementarias en cadenas opuestasforman enlaces de hidrógeno. A se empareja con T(puentes 2H), y C con G (puentes 3H).
5. Pares de bases 0.34 nm aparte. Una vuelta completade la hélice requiere 3.4 nm (10 bases por vuelta).
6. Las vértebras de azúcar-fosfato forman surcosmayores y menores.
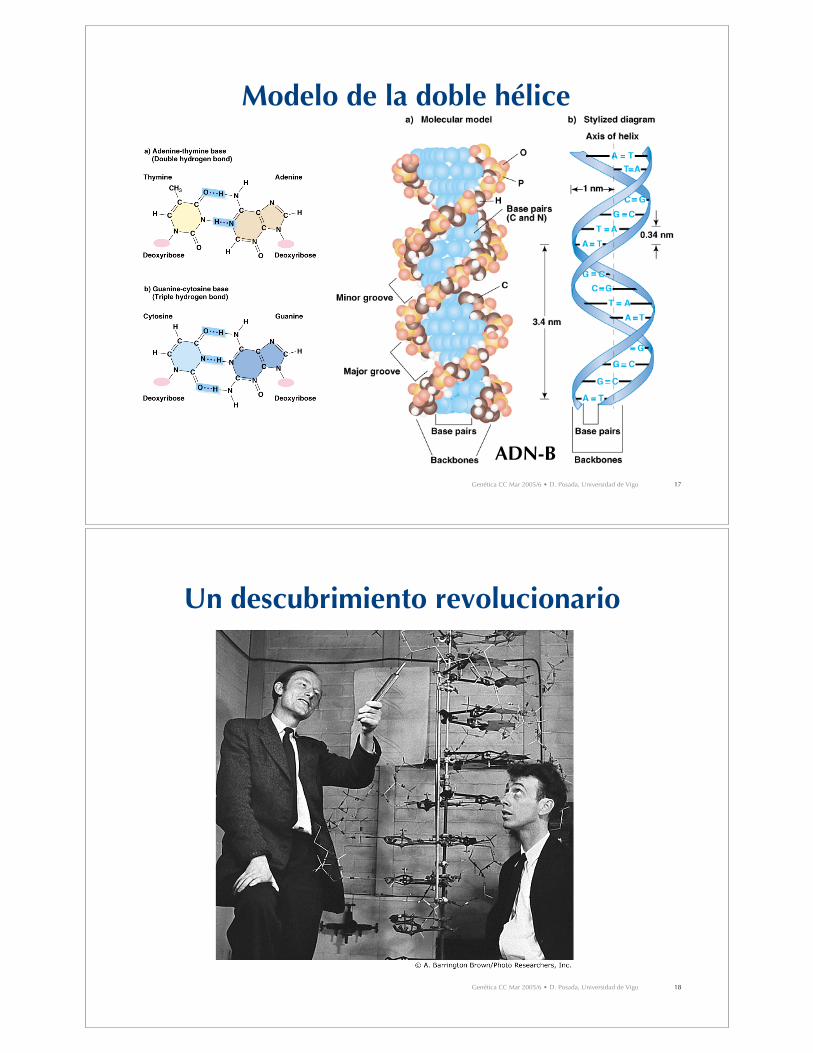
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 17
Modelo de la doble hélice
ADN-B
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 18
Un descubrimiento revolucionario

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 19
Protagonistas
• Crick, Watson y Wilkins recibieron elNobel en 1962.
Rosalind
Franklin
James D.
Watson
Francis H.
Crick
Maurice H. F.
Wilkins
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 20
Tipos diferentes de ADN

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 21
Estructura del ARN
• El ARN es de cadena única,y puede adoptar estructurascomplejas, inclusointramoleculares.
• El ARN utiliza como azúcarla ribosa.
• El ARN utiliza U en vez deT.
• El ARN puede catalizarimportante reaccionesbiológicas.
rRNAtRNA
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 22
Organización del ADN en cromosomas
• El DNA de la célula se organiza cromosomas.• El cromosoma o juego de cromosomas que
contiene todo el DNA de un organismo sedenomina genoma.
• En procariotas el genoma es usualmente uncromosoma circular único.
• En eucariotas el genoma es un juego haploidecompleto de cromosomas contenidos en elnúcleo celular.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 23
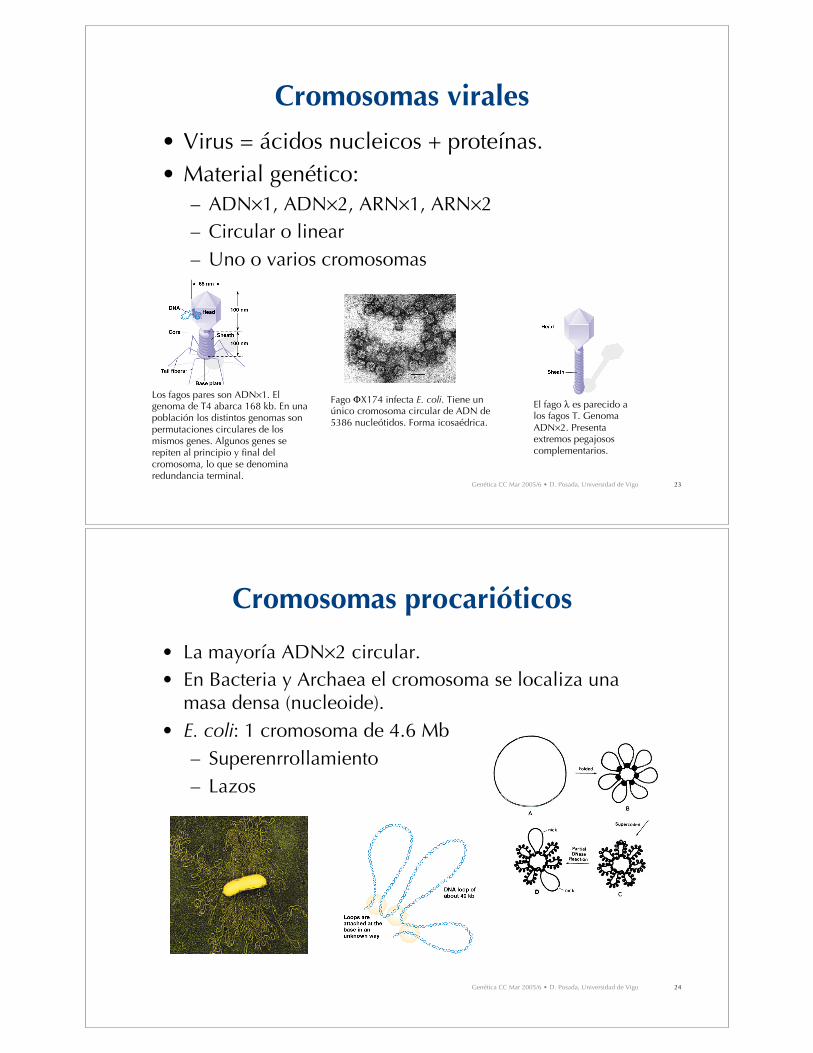
Cromosomas virales
• Virus = ácidos nucleicos + proteínas.• Material genético:
– ADN!1, ADN!2, ARN!1, ARN!2– Circular o linear– Uno o varios cromosomas
Fago "X174 infecta E. coli. Tiene unúnico cromosoma circular de ADN de5386 nucleótidos. Forma icosaédrica.
El fago # es parecido alos fagos T. GenomaADN!2. Presentaextremos pegajososcomplementarios.
Los fagos pares son ADN!1. Elgenoma de T4 abarca 168 kb. En unapoblación los distintos genomas sonpermutaciones circulares de losmismos genes. Algunos genes serepiten al principio y final delcromosoma, lo que se denominaredundancia terminal.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 24
Cromosomas procarióticos
• La mayoría ADN!2 circular.• En Bacteria y Archaea el cromosoma se localiza una
masa densa (nucleoide).• E. coli: 1 cromosoma de 4.6 Mb
– Superenrrollamiento– Lazos

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 25
Valor C
• Valor C = cantidad total de ADN en el genomahaploide de una especie.
• Varía entre las especies sin relación concomplejidad estructural u organizativa (paradojadel valor C).
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 26
Cromatina
• La cromatina es el complejo de ADN y proteínascromosómicas. Parecida en todos los eucariotas.
• En la cromatina encontramos proteínas histonas y nohistonas.
• Las histonas son comunes, pequeñas, básicas (+)– Papel fundamental en el empaquetamiento de la cromatina.– Cinco tipos principales H1, H2A, H2B, H3 y H4,– En peso, histonas/ADN =1 en todos los eucariotas.– Muy conservadas evolutivamente (H4 de vacas y guisantes se
diferencian en 2 aminoácidos).
• Las proteínas no histonas son a menudo acídicas (-)– Difiere mucho entre los tipos celulares y entre los diferentes
organismos.– En peso, no histona / ADN = 50-100%.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 27
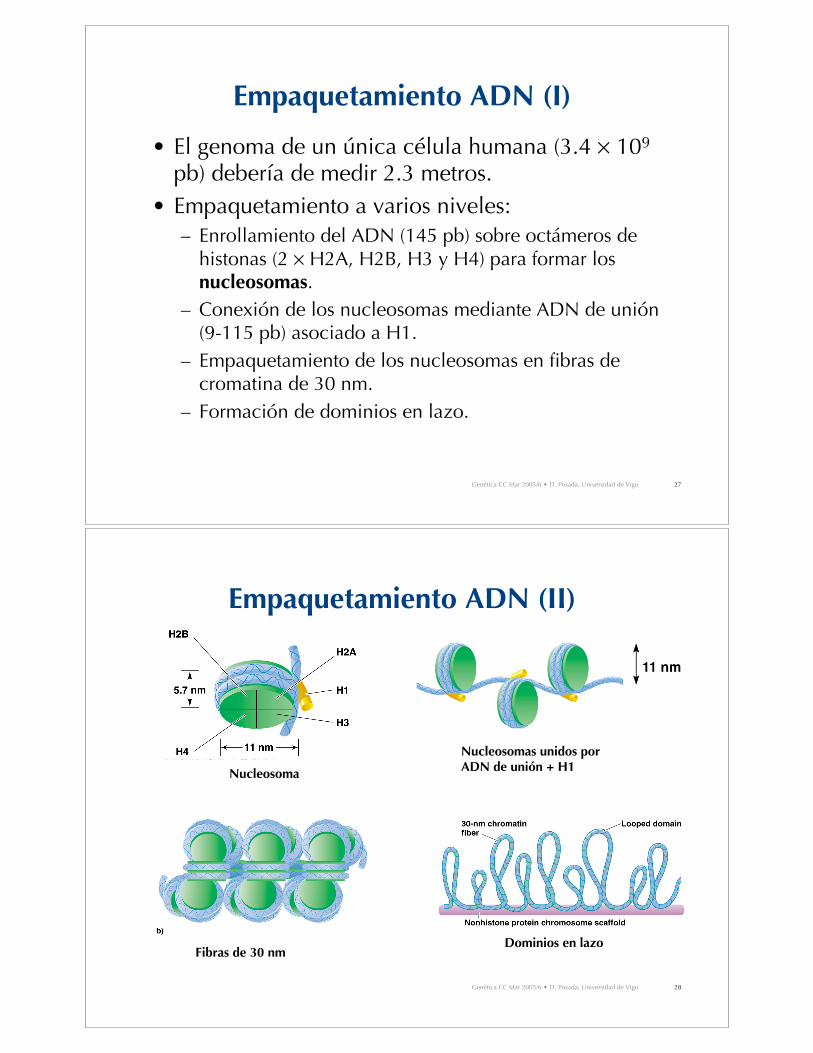
Empaquetamiento ADN (I)
• El genoma de un única célula humana (3.4 ! 109
pb) debería de medir 2.3 metros.• Empaquetamiento a varios niveles:
– Enrollamiento del ADN (145 pb) sobre octámeros dehistonas (2 ! H2A, H2B, H3 y H4) para formar losnucleosomas.
– Conexión de los nucleosomas mediante ADN de unión(9-115 pb) asociado a H1.
– Empaquetamiento de los nucleosomas en fibras decromatina de 30 nm.
– Formación de dominios en lazo.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 28
Empaquetamiento ADN (II)
Nucleosoma
Nucleosomas unidos por ADN de unión + H1
Fibras de 30 nmDominios en lazo

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 29
Empaquetamiento ADN (III)
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 30
Eucromatina y heterocromatina
• Eucromatina: sigue los ciclos habituales decondensación (máxima en metafase)ydescondensación durante el ciclo celular.– La mayoría del genoma de una célula activa es eucromatina.– Transcripción activa.– Secuencias no repetidas.
• Heterocromatina: siempre condensada.– Transcripción inactiva.– La heterocromatina constitutiva está presente en todas las
células en posiciones idénticas cromosomas homólogos.Básicamente ADN repetitivo. Ej. Regiones centroméricas.
– La heterocromatina facultativa varía en estado de célula acélula, durante el desarrollo, y a veces, entre cromosomashomólogos. Ej. corpúsculos de Barr.

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 31
ADN centromérico y telomérico
• En el centrómero se pueden encontrar secuencias deADN especiales (CEN en levaduras) que interactúancon los microtúbulos.
• Las secuencias teloméricas mantienen la estabilidad delos cromosomas y intervienen en su replicación:– Secuencias simples teloméricas: secuencias repetidas en
tándem en los extremos con funciones esencialesestabilizadoras.
– Secuencias asociadas a telómeros, de forma más interna, algomás complejas y con función desconocida.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 32
Propiedades físico-químicas de losácidos nucleicos
• Densidad de los ácidos nucleicos.• Desnaturalización de los ácidos nucleicos:
temperatura de fusión (Tm).• Absorbancia a 260 nm.• Cinética de renaturalización: Curvas Cot.• Hibridación de los ácidos nucleicos.

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 33
Densidad y desnaturalización
• $G+C $densidad del ADN.• Desnaturalización: paso de cadena doble a
sencilla.– $G+C $estabilidad del ADN (+puentes 3H).– Temperatura de fusión (Tm): Tº necesaria para
desnaturalizar la mitad del ADN de una mezcla.– $G+C $Tm
Ab
sorb
anci
a a
26
0 n
m
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 34
Absorbancia UV a 260 nm
• El menor grado de absorción se produce enestado de doble hélice.
• La absorción aumenta cuando se produce ladesnaturalización pasando a estado de hélicesencilla (efecto hipercrómico, aumento de laabsorbancia).
• Si degradamos este ADN de hélice sencilla anivel de nucleótidos libres, de nuevo aumentala absorbancia.

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 35
Cinética de la reasociación (I)
• La renaturalización es el paso de hélice sencilla adoble hélice.
• Curvas Cot o de renaturalización.• Cot ! = tiempo en que se ha reasociado ! del ADN.
– Directamente proporcional a la complejidad del ADN delorganismo.
– $secuencias repetidas $velocidad renaturalización
Figura. Curva Cot.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 36
Cinética de la reasociación (II)
• Las curvas Cot de virus y bacterias tienen un sólopunto de inflexión (secuencias únicas).
• Las curvas Cot de eucariotas poseen varios puntos deinflexión:– Cot ! bajos (10-4 a 10-1): secuencias altamente repetidas– Cot ! 10–102: secuencias moderadamente repetidas.– Cot ! > 103 : secuencias únicas o de bajo número de copias.
Figura. Curvas Cot.

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 37
Hibridación
• Las técnicas de desnaturalización renaturalizaciónpermiten la hibridación ADN+ADN o ADN+ARN.
• Se puede identificar el gen que ha dado lugar a undeterminado ARN mensajero.
• Las técnicas de hibridación "in situ" permiten localizarla posición de un gen de un cromosoma eucariótico.– ADN marcado con fluorescencia (FISH).– Marcaje del genoma (GISH).
Figura. FISH Figura. GISH
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 38
Secuencias únicas y repetidas de ADN
• ADN de secuencia única: una opocas copias.– Genes codificantes– ADN procariota– ~65% genoma humano
• ADN repetitivo:– Moderadamente repetitivo (pocas
o hasta 105 copias) o altamenterepetitivo (105 – 107 copias).
– Disperso (SINEs y LINEs; funcióndesconocida) o en tándem(centrómeros y telómeros; ~65%genoma humano; microsatélites).

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 39
Modelos de replicación del ADN• Semiconservativo: cada molécula de la progenie conserva una de las
hebras parentales.• Conservativo: las hebras parentales sirven como un único molde para la
síntesis de nuevas dobles hélices.• Dispersivo: fragmentos de doble hélice parental sirven como moldes
para la síntesis de nuevos segmentos de doble cadena. Las nuevas hélicesestán formadas por segmentos parentales y de la progenie.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 40
Experimento de Meselson y Stahl (1958)
• Obtuvieron evidencia experimental de que elmodelo semiconservativo era el correcto.
• Marcan ADN de E. coli con 15N.• 15N y 14N (normal) se pueden separar mediante
centrifugación en gradientes de densidad.

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 41
Experimento de Meselson y Stahl (1958)
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 42
ADN polimerasa I
• Arthur Kornberg (1955): la ADN polimerasa Illeva a cabo la síntesis de ADN.
• Los componentes necesarios son:– La ADN polimerasa I– Desoxirribonucleósidos trifosfato (dNTPs = dATP,
dCTP, dGTP, dTTP), precursores de losnucleótidos.
– Un molde de ADN.– Iones de magnesio (Mg2+), que optimizan la
actividad de la ADN polimerasa.
Figura. Arthur Kornberg (Nobel 1959).

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 43
Reacción de la polimerasa I de ADN (I)
• Formación de un enlace fosfodiéster en el 3’-OH de ladesoxirribosa y el fosfato 5’ del dNTP.– La energía proviene de la liberación de 2 de los 3 fosfatos.– La cadena que crece actúa como cebador en la reacción.
• La polimerasa encuentra el dNTP complementario alnucleótido de la cadena molde y lo incorpora– 800 nucleótidos por segundo.– 1 error cada 10-6 nucleótidos.
• La dirección de la síntesis es 5’-3’.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 44
Reacción de la polimerasa I de ADN (II)

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 45
Reacción de la polimerasa I de ADN (III)
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 46
Polimerasas de ADN procarióticas
• E. coli: 3 polimerasas de ADN (I, II, y II).• Actividad exonucleásica 3’-5’: corrección de
errores (tasa error final 10-9).• La polimerasa de ADN I presenta además
actividad exonucleásica 5’-3’.

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 47
Replicación en procariotas• Un único origen de replicación (oriC en E. coli, ~245 pb).• En este sitio el ADN se desnaturaliza en cadenas sencillas
produciendo una horquilla de replicación.• Replicación bidireccional.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 48
Inicio de la replicación en E. coli (I)
1. Una girasa (tipo de topoisomerasa) relaja el superenrollamiento delADN.
2. Proteínas iniciadoras se unen al origen de replicación.3. Una helicasa de ADN se une a las proteínas iniciadoras. La helicasa
avanza hidrolizando ATP desenrollando la doble cadena.4. Las proteínas de unión al ADN de cadena sencilla (SSB), previniendo
que estas cadenas se emparejen de nuevo.5. La primasa de ADN (polimerasa de ADN) se une a la helicasa
formando el primosoma.6. La primasa sintetiza un cebador de ARN (11-12 nt.), al cual puede
añadir nucleótidos la polimerasa de ADN III en dirección 5’-3’.7. El cebador de RNA es más tarde eliminado y reemplazado por ADN
por acción de la polimerasa de ADN I. El hueco es sellado por unaligasa.

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 49
Inicio de la replicación en E. coli (II)
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 50
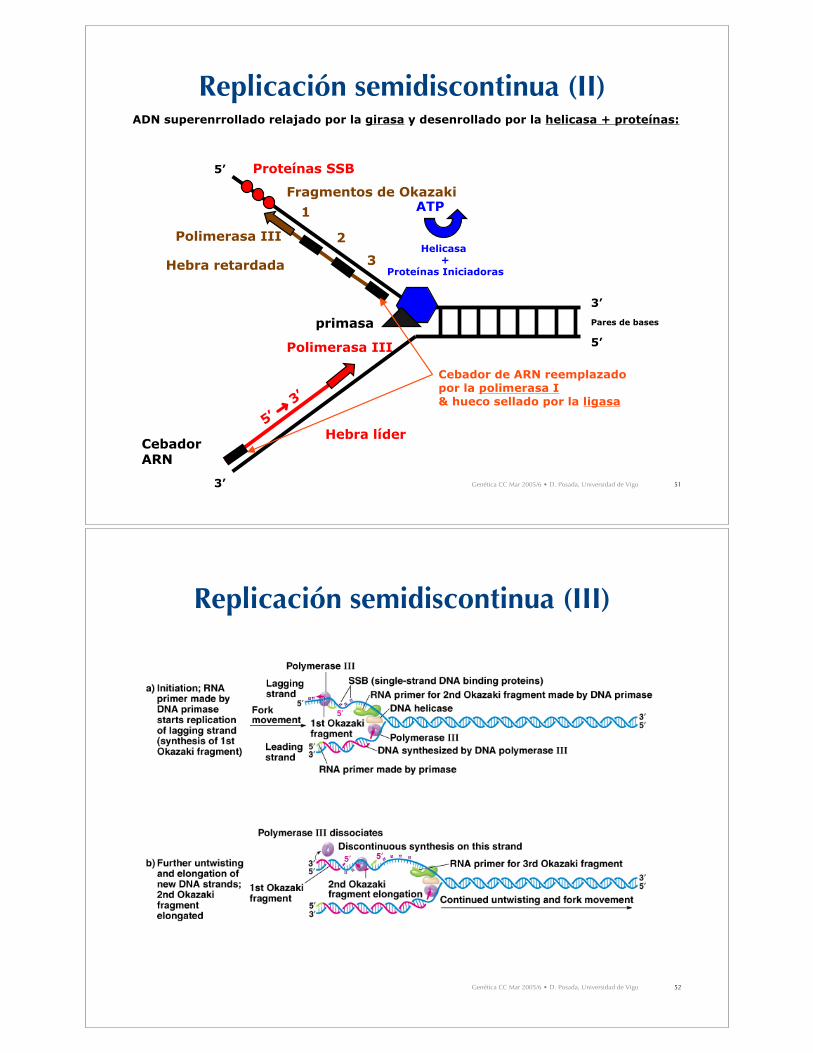
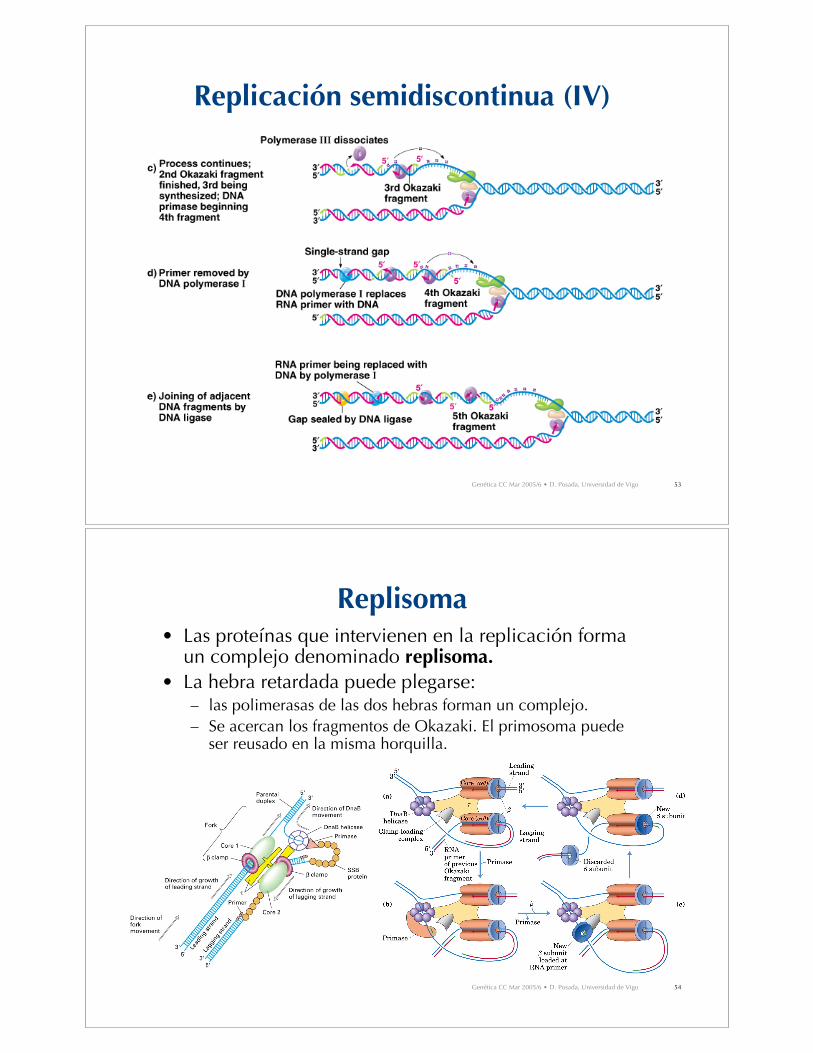
Replicación semidiscontinua (I)
• Para mantener la polaridad 5’-3’, el ADN ha de sintetizarse endirecciones opuestas en ambas hebras molde.
• Hebra líder: síntesis en la misma dirección que el movimiento dela horquilla de replicación.– Síntesis continua.– Sólo necesita 1 cebador
• Hebra retardada: síntesis en dirección opuesta al movimiento dela horquilla de replicación.– Síntesis discontinua (fragmentos de Okazaki).– Necesita varios cebadores de ARN.– Eventualmente los fragmentos de Okazaki son conectados entre sí
por la acción de la polimerasa I de ADN (que reemplaza el cebadorde ARN por ADN) y una ligasa de ADN, que une los fragmentos.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 51
3
Polimerasa III
5’ % 3
’
Hebra líder
Pares de bases
5’
5’
3’
3’
ADN superenrrollado relajado por la girasa y desenrollado por la helicasa + proteínas:
Helicasa +
Proteínas Iniciadoras
ATP
Proteínas SSB
CebadorARN
primasa
2Polimerasa III
Hebra retardada
Fragmentos de Okazaki
1
Cebador de ARN reemplazado por la polimerasa I& hueco sellado por la ligasa
Replicación semidiscontinua (II)
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 52
Replicación semidiscontinua (III)
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 53
Replicación semidiscontinua (IV)
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 54
Replisoma• Las proteínas que intervienen en la replicación forma
un complejo denominado replisoma.
• La hebra retardada puede plegarse:– las polimerasas de las dos hebras forman un complejo.– Se acercan los fragmentos de Okazaki. El primosoma puede
ser reusado en la misma horquilla.

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 55
Replicación ADN circular E. coli
1. Dos horquillas de replicaciónresultan en una estructura enforma de zeta (&).
2. A medida que se separan lashebras, se formasuperenrrollamiento positivoen algún otro lugar de lamolécula.
3. Las topoisomerasas alivian latensión en lossuperenrrollamientos,permitiendo que las doshebras continúen separándose.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 56
Replicación en círculos rodantes
• Varios fagos de ADN,como "X174.
1. Corte en la hebra +.2. El extremo 5’ la hebra +
se desplaza para formarun horquilla.
3. La cadena intacta sirvecomo molde.
4. La síntesis en la la hebra+ desplazada ocurre deforma discontinua haciael final de la cadena.

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 57
Replicación del ADN en eucariotas
• La replicación el ADN es similar en procariotas y eneucariotas (varios cromosomas).
• En cada ciclo celular, cada cromosoma debe de serduplicado fielmente durante la fase S. Tres puntos dechequeo:– Punto START (G1; en levaduras): se comprueba si la célula es
suficientemente grande y el ambiente favorable, para pasar ala fase S.
– Punto de chequeo de la G2: se decidirá si la célula entra enmitosis, para lo que todo el DNA tiene que estar replicado.
– Punto de chequeo durante la mitosis: los cromosomas debende estar propiamente ligados al huso mitótico para que secomplete la mitosis.
– En estos chequeos intervienen ciertas proteínas denominadasciclinas, y quinasas dependientes de ciclinas.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 58
Polimerasas de ADN eucarióticas
• No se sabe mucho sobre estas enzimas, pero almenos 5 han sido identificadas en mamíferos:– Polimerasa ' (alfa): nuclear, replicación del ADN,
sin función correctora.– Polimerasa ( (beta): nuclear, reparación del ADN,
sin función correctora.– Polimerasa ) (gamma): mitocondrial, replicación
del ADN, con función correctora.– Polimerasa * (delta): nuclear, replicación del ADN,
con función correctora.– Polimerasa + (epsilon): nuclear, reparación del ADN
(?),con función correctora.

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 59
Replicón eucariótico
• Cada cromosomaeucariótico es una doblecadena linear de ADN.
• Existen varios orígenes dereplicación (no simultáneos).
• La región de replicación sedenomina replicón o unidadde replicación.– El replicón se mueve de forma
más lenta que en procariotas.– Los fragmentos de Okazaki son
más pequeños (100-150 pb).
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 60
Orígenes de replicación
• En levaduras se han identificado secuenciasespecíficas que parecen conferir la habilidad dereplicación autónoma (secuencias de replicaciónautónoma o ARSs).
• Se cree que por lo menos algunas de estas secuenciascorresponden con orígenes de replicación.
• En eucariotas más complejos los orígenes dereplicación no están bien definidos.

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 61
• Al eliminar los cebadores de los extremos, los huecos no puedenser cebados por otros fragmentos => los cromosomas se acortaríaen cada ronda de replicación.
• Secuencias repetidas en tándem en los telómeros.• La telomerasa (transcriptasa reversa), que contiene proteínas y
ARN complementario a la repetición terminal, añade repeticionesen el extremo del cromosoma, rellenando el hueco dejado por elcebador.
• El resultado de la acción de la telomerasa es el alargamiento delcromosoma.
Replicación de los telómeros (I)
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 62
Replicación de los telómeros (II)

Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 63
Ensamblaje del nuevo ADN ennucleosomas
• La replicación estácoordinada con laformación de nuevosnucleosomas.
• Se requiere la formaciónde nuevas histonas.
• Los nucleosomas seforman casi de formainmediata durante lareplicación.
Figura. Ensamblaje de nucleosomas.
Genética CC Mar 2005/6 • D. Posada, Universidad de Vigo 64
Replicación del ADN mitocondrial
• Modelo del lazo de desplazamiento olazo D.
• Hélice ligera (L) y pesada (H).• Primero se inicia la replicación
unidireccional de la nueva hélice L.• Cuando se ha sintetizado, unos 2/3 de
la hélice L, comienza la síntesis de lanueva hélice H, en otro origen y enuna sola dirección, opuesta a la desíntesis de la hélice L.
Figura. Replicación del ADN mitocondrial





























