Embed Size (px)
Citation preview

UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR
ÁREA DE CONOCIMIENTOS DE CIENCIAS DEL MAR
DEPARTAMENTO DE BIOLOGÍA MARINA
TESIS
INFLUENCIA DEL BRIOZOARIO Membranipora tuberculata SOBRE EL
METABOLISMO DEL ALGA ROJA Gelidium robustum
QUE COMO REQUISITO PARA OBTENER EL TÍTULO DE
BIÓLOGO MARINO
PRESENTA
BERNARDO VEYRAND QUIRÓS
DIRECTOR:
DR. JESÚS IVÁN MURILLO ÁLVAREZ
LA PAZ BAJA CALIFORNIA SUR, NOVIEMBRE DE 2013

II

III
El hecho que un misterio esté más allá del alcance de la comprensión humana no
significa que no debamos intentar comprenderlo, hasta donde ello sea posible,
dentro del dominio del intelecto.
Taimni

IV
Dedicatoria
En memoria de Rodolfo M. Quirós Moreno, quien siempre me dio su apoyo y
atención en mi desarrollo académico. Aunque te nos adelantaste en este camino
llamado vida, y no pudiste ver culminado mi estudio, sé que me apoyas e impulsas
ahora desde algún otro lado. Estés donde estés, siempre serás parte de la energía
que impulsa a este motor para salir adelante. Espero estés orgulloso de mi como
siempre lo estuviste, gracias por todo Abuelo.

V
Agradecimientos
Para llegar hasta este punto se recorrieron varias etapas a lo largo de mi
vida, siendo este trabajo un conjunto de situaciones y hechos que se enlazaron.
Este proyecto reúne el trabajo y esfuerzo de muchos, conjuntando sueños y
anhelos de varias personas que se relacionaron a lo largo de estos años por lo
que el crédito no puede ser sólo mío.
Primeramente me gustaría agradecer a mi familia, mis padres, esas
personas quienes sin ellos, esto no hubiera sido posible. A mi madre, por tener su
apoyo incondicional desde siempre, alentando mis sueños y pasiones,
enseñándome a no rendirme y cumplir con las cosas que me proponga, ser fuerte
y llegar alto, sin embargo también a cómo mantener los pies en tierra y ser
objetivo. A mi padre, esa persona que siempre fue mi ejemplo a seguir, que con
tropezones y todo siempre nos sacó adelante. Ese hombre que me enseñó a tener
principios y ser una persona recta, cumplir mis metas y objetivos y siempre
terminar lo que uno comenzó; este sueño de convertirme en Biólogo marino
también se lo atribuyo a él. En conjunto les debo ser la persona de bien que soy
en la actualidad y en la que podré convertirme en un futuro. Gracias por todos
estos años de apoyo y enseñanzas, valores y sentido de la responsabilidad.
A mi pequeña hermana Dominique, que aunque no intervino directamente
con la tesis, si lo hace en mi vida cotidiana, gracias por su apoyo y todos esos
buenos momentos que hemos tenido y espero tener para siempre; sin ella la vida
hubiera sido muy aburrida y no hubiera aprendido el valor de compartir, te quiero.
A mis abuelos, quienes son una parte fundamental en mi vida, mi abuela
Esperanza quién siempre me recibe con una gran sonrisa y mucha comida. Mi
abuelo Rodolfo que aunque se adelantó con su partida y no pudo ver esta tesis
concluida siempre se interesó por mi carrera universitaria y me apoyó en lo que
pudo. A mi abuela Clara Elena quien además de apoyarme y alentarme me ayudó
en la revisión y corrección de estilo de este manuscrito, muchas gracias.
A mis tíos biologuescos, en especial a Alberto y Lucía Flores, quienes me
han enseñado que la familia no sólo se crea por lazos de sangre, si no por esas
personas que están siempre para ti y te dan su apoyo en todo momento, gracias
por guiarme por una vida en relación a la biología marina y sus buenas comidas.

VI
A mis amigos y compañeros de generación, aquellos que se volvieron mi
familia durante los años de carrera, en especial a Roberto, Pablo y Edmundo (las
perras) con quienes siempre pude contar. A Priscila por siempre ser una buena
amiga y escuchar, y Luna por soportarnos a todos con nuestras continuas bromas
y molestias, así como a Fernando que siempre estuvo ahí para apoyar.
Quisiera agradecer por supuesto a mi comité de tesis; mi Sensei, el Dr. Iván
Murillo, por haberme recibido y adoptado ya desde hace algún tiempo en el
laboratorio e instruirme en el arte de los productos naturales, coordinar el
desarrollo y logística de la investigación, así como aguantarme y ayudarme con la
realización de este escrito y sus constantes revisiones, gracias. Al Dr. Mauricio
Muñoz, mi director adoptivo, quien siempre con buena fe me apoyaba e instruía en
todo lo que podía, sin mencionar que siempre tenía algo interesante que contar o
al menos una carcajada que formar. Al Dr. Rafael Riomena, Dr. Juan Manuel
López y la M. en C. Erika Torres, por las molestias y las revisiones de este escrito
gracias.
Un agradecimiento importante a los miembros del laboratorio de Química de
Algas en CICIMAR por su apoyo diario en mantener la cordura, ya que si no se
podía ayudar con algo al menos siempre tenían una sonrisa que sacar. Un
agradecimiento especial a Araceli, quién me salvó y ayudó en múltiples ocasiones.
A la M.C. Elizabeth Rodríguez y M.C. Dora Arvizu por consejo en laboratorio. A
Ana, Ilse, Miguel, Valerie, Valeria, Dania y Stephanie gracias por la compañía y
espero verlos por ahí el próximo semestre. A Mario Vergara gracias por la ayuda y
corrección del planteamiento en la parte del laboratorio.
Agradezco a la Universidad Autónoma de Baja California Sur, por el
aprendizaje dado en estos años de carrera y a CICIMAR y la Secretaría de
Investigación y Posgrado del Instituto Politécnico Nacional por su apoyo y
financiamiento en la realización de este proyecto.
A todos los que han creído en mí, así como los que no, los que no he
mencionado a causa de mi Alzheimer, así como todas las personas que han sido
transeúntes en esta historia denominada vida, una disculpa y muchas gracias.

VII
Índice
Lista de figuras……………………………………………………….......................................VIII
Lista de Tablas…………………………………………………………………………..............VIII
Resumen…………………………………………………………………………………………...IX
1. Introducción……………………………………………………………………………………1
2. Antecedentes…………………………………………………………………………………..3
2.1 Estudios de la relación de algas y briozoarios……………….…………………………….5
2.2 Metabolitos secundarios aislados de algas rojas con actividad antiepífita…………….7
2.3 Características biológicas de Gelidium robustum………………………………………....9
3. Justificación…………………………………………………………………………….…….11
4. Objetivos………………………………………………………………………….…………..13
4.1 Objetivo general…………………………………………………………………….………..13
4.2 Objetivos específicos……………………………………………………………..…..……..13
5. Material y métodos…………………………………………………………………………..14
5.1 Recolección del alga……………………………………………………………….………..14
5.2 Obtención de extractos etanólicos…………………..…………………………………….14
5.2.1 Pruebas cromatográficas de extractos…………………………………………………15
5.2.2 Fraccionamiento de extractos etanólicos……………………………………………...16
5.3 Evaluación de actividad antibacteriana…………………………………………….……..16
5.4 Identificación estructural por análisis espectroscópico……………………………..…...18
5.5 Cálculos y análisis estadísticos……………………………………………….……………19
6. Resultados……………………………………………….…………………………………...20
7. Discusión……………………………………………….…………………………………….28
8. Conclusiones…………………………………………………….…………………………...34
9. Literatura citada……………………………………………….……………………………..36
10. Anexos……………………………………………….……………………………………….43

VIII
LISTA DE FIGURAS
Figura 1. Alga Gelidium robustum…………………………………………………….9
Figura 2. Ciclo de vida de Gelidium robustum (Hernández-Guerrero,1997)…..…11
Figura 3. Cromatografía de extractos crudos de G. robustum……...……………...21
Figura 4. Fracciones 12-001234-40F1, 12-001234-40F2 y 12-001234-40F3…….21
Figura 5. Espectro infrarrojo de los extractos crudos de G. robustum con
diferentes porcentajes de cobertura de M. tuberculata y sus respectivos números
de banda…………………………..……………………………………………………...24
Figura 6. Espectro infrarrojo de la fracción 12-001234-40 F3……………..………25
Figura 7. Análisis de varianza de las diferentes bandas con disminución de
área………………………………………………………………………………………..27
LISTA DE TABLAS
Tabla I. Actividad antimicrobiana de los extractos y fracciones de G. robustum,
empleando el método de difusión en agar. El número es el diámetro promedio
expresado en mm; NA significa que no presentan actividad………………………..22

IX
Resumen
En las últimas tres décadas, la ecología química tomó relevancia como ciencia,
dando lugar a un nuevo campo de investigación que involucra tanto la química de
productos naturales, como el estudio de relaciones biológicas y ecológicas. La
epibiosis es un tipo de simbiosis donde un organismo vive sobre la capa superficial
de otro en una fase de su vida sin considerarse parasitismo, ya que generalmente
no causan un efecto negativo al hospedero; los briozoarios son un grupo de
invertebrados que cubren las frondas de las algas interfiriendo en sus procesos
fotosintéticos. El objetivo de este trabajo fue observar el efecto metabólico del
briozoario Membranipora tuberculata sobre el alga roja Gelidium robustum para
entender la relación química y ecológica de estos organismos. Para ello se
obtuvieron cinco extractos etanólicos de G. robustum con diferente porcentaje de
cobertura de M. tuberculata (0-30%, 30-60%, >60%, cobertura de 100% y 100%
de cobertura posteriormente limpiada de epífitos) y se compararon sus perfiles de
metabolitos por medio de cromatografía de capa fina, observándose la presencia
de un metabolito polar, cuya concentración varió en relación inversa al porcentaje
de cobertura. Como resultado del fraccionamiento del extracto etanólico se
observó que una fracción (12-001234-40F3) inhibió el crecimiento de
Staphylococcus aureus y Vibrio parahaemolyticus, ambas bacterias relacionadas
al microepifitismo. Mediante espectroscopia de infrarrojo se analizaron los
extractos etanólicos contrastando la composición relativa de metabolitos mediante
la cuantificación de bandas diagnósticas. Se observó una relación entre la
disminución del área de bandas de absorción y el aumento de porcentaje de
briozoario. M. tuberculata, sin embargo por medio de análisis de ANDEVA las
diferencias resultaron no significativas.
Palabras clave: Epibiontes, Microepifitismo, Gelidium robustum, Membranipora
tuberculata, Espectrometría Infrarrojo.

1. Introducción
La ecología química es rama multidisciplinaria reciente con gran impacto en
las áreas de ciencias químicas y biológicas. Esta ciencia conduce al estudio de
sustancias químicas involucradas en las interacciones ecológicas intra e
interespecíficas de organismos vivos (Bergström, 2007). Durante las últimas tres
décadas, los químicos de productos naturales han empleado la ecología química
para determinar funciones e interacciones ecológicas entre los organismos,
estudiando los mecanismos químicos subyacentes y usando condiciones
ecológicas reales (Williams et al., 1989; Hay, 1996).
Actualmente el campo de la ecología química marina presenta la
integración de diferentes ciencias creando un modelo interdisciplinario que estudia
tópicos como interacciones de depredador-presa, competencia, comunicación
química y evolución de metabolitos secundarios (McClintock y Baker, 2001). Las
sustancias químicas involucradas en las diferentes interacciones ecológicas se
denominan semioquímicos, del griego semion, que significa marca o señal (Law y
Regnier, 1971). Sin embargo recientemente se propuso el término infoquímicos,
refiriéndose a compuestos que transmiten información entre individuos y que
producen en el receptor una respuesta en su fisiología o comportamiento,
mostrando que las interacciones entre seres vivos por sustancias químicas
pueden presentarse de manera perjudicial o beneficiosa, interviniendo en
procesos evolutivos (Dick y Sabelis, 1988; Cortez, 2013).

2
La epibiosis es un tipo de simbiosis donde un organismo vive sobre la capa
superficial de uno de mayor tamaño utilizándolo como sustrato por al menos una
fase de su ciclo vital. Estos organismos epibiontes no se consideran parásitos, ya
que generalmente no le causan algún efecto negativo al organismo hospedero
(Wahl, 1989). Las macroalgas que se encuentran en la zona intermareal son
frecuentemente recubiertas por diferentes conjuntos de organismos epibiontes, a
los cuales las algas les proporcionan tanto sustrato como alimentos para
desarrollarse (Hurd et al., 1994). Dentro de estos organismos podemos encontrar
briozoarios, un grupo de invertebrados que coloniza cubriendo extensas áreas de
las frondas de las algas e interfieren en la captación de luz, afectando así su
fisiología, lo que produce diferentes efectos tales como la modificación del costo
energético del alga mediante la reducción de la tasa de fotosíntesis, disminuyendo
la densidad del flujo fotónico que recibe el alga, así como la alteración de la
calidad de la luz (Cancino et al., 1987; Molina et al., 1991). Como consecuencia de
ello, los talos del alga con epibiontes presentan tasas fotosintéticas menores que
los talos sin epibiontes. A su vez limita el tamaño de las algas y altera los tejidos
más susceptibles ocasionando el desplazamiento o ruptura de las frondas (Dixon
et al., 1981; Molina et al., 1991).
Sin embargo, algunos estudios sugieren que la colonización por briozoarios
no siempre interfiere con los costes metabólicos en las algas asociadas, sino que
reducen la cantidad de luz que llega a la superficie del alga bajo irradiancias
similares a las que ocurren en el entorno natural del alga, siendo la luz que

3
penetra en el organismo suficiente para permitir la fotosíntesis máxima. Estos
estudios muestran que algas rojas del género Gelidium J.V. Lamouroux, 1813
aumentan las concentraciones de pigmentos fotosintéticos en los tejidos como
respuesta a la baja irradiancia causada por la colonización, aumentando así su
eficiencia fotosintética (Cancino et al., 1987; Hurd et al., 1994).
El presente trabajo tiene como propósito establecer la diferencia relativa de
la composición de metabolitos de la especie Gelidium robustum (N.L. Gardner)
Hollenberg & I.A. Abbott 1965 en diferentes porcentajes de cobertura por el
briozoario epífito Membranipora tuberculata (Bosc, 1802) para observar si existe
una influencia negativa real sobre el hospedero por parte del huésped y, de ser
así, comprender si emplean un medio de defensa anti-incrustante o una
compensación metabólica fotosintética.
2. Antecedentes
Mundialmente existe un gran interés por las especies de algas Rodophytas
del género Gelidium, debido a su alta calidad de agar y los derivados que se
obtienen de ellas. Estos productos tienen aplicaciones en la industria alimenticia y
farmacéutica debido a sus propiedades estabilizadoras, emulsificadoras y como
agentes gelificantes, así como en bacteriología para la elaboración de medios de
cultivo (Casas-Valdez y Fajardo-León, 1990; Hernández-Guerrero, 1997).

4
En México se tienen registros de 13 especies de Gelidium en la región del
Pacífico, de las cuales hay 10 representantes en el Pacífico de Baja California, 7
especies en el Golfo de California y 5 especies en el Pacífico Tropical Mexicano.
Sin embargo sólo la especie G. robustum es empleada como materia prima para la
industria del agar en México, colocándolo dentro de los principales países
productores de agarofitas (Rodríguez-Vargas et al., 1993). La explotación de G.
robustum en México se inició en 1945 manteniéndose estable hasta llegar
actualmente al segundo lugar en producción de algas marinas en el mundo
(Molina, 1986). Sin embargo no fue objeto de estudio hasta finales de los sesenta,
siendo los pioneros Guzmán del Próo y Granados (1968) quienes realizaron un
reconocimiento de la costa de la península de Baja California y mencionaron que
se distribuye desde Isla Coronado hasta Bahía Magdalena, siendo los mantos de
mayor densidad únicamente hasta Punta Abreojos. Posteriormente se
comenzaron a hacer estudios de la velocidad de crecimiento bajo diferentes
sistemas de cosecha (Guzmán del Próo y De La Campa, 1969).
Aguilar y Aguilar (1990) establecieron la problemática de la relación de G.
robustum con el briozoario Membranipora tuberculata denominado comúnmente
como “conchilla”, el cual provoca la presencia de una cubierta calcárea en los ejes
y frondas basales de las plantas (Casas y Hernández, 1996). Esta cobertura
calcárea determina el valor comercial de G. robustum debido a que disminuye la
calidad de su agar y por lo tanto su precio en el mercado; el Gelidium de primera
calidad no presenta ninguna placa calcárea, teniendo un costo aproximado de

5
1,600 dólares por tonelada seca, mientras que algas con gran cantidad de
cobertura calcárea se denomina de tercera o cuarta calidad y disminuye su valor a
900 dólares por tonelada seca; por lo que se determina que entre menos cobertura
de conchilla, mejor es la calidad de G. robustum como materia prima (Aguilar-
Rosas et al., 1998).
2.1 Estudios de la relación de algas y briozoarios
La mayoría de las publicaciones relacionadas con epibiontes sugieren que
tienen un efecto negativo en el crecimiento o sobrevivencia de las algas
colonizadas, cuando en realidad no se han presentado evidencias que lo afirmen;
incluso podría presentarse una posible relación de beneficio mutuo (De Burg y
Frankboner, 1978).
Dixon et al. (1981) desarrollaron un experimento en el cual comparaban el
tiempo de caída entre las frondas de Macrocystis pyrifera (Linnaeus) C. Agardh
1820 cubiertas por el briozoario Membranipora membranacea (Linnaeus, 1767) y
otras sin cobertura, obteniendo que no se muestra una diferencia significativa en
los tiempos de caída. A su vez Cancino et al. (1987) reportaron los cambios que
ocurren en la intensidad y calidad del paso de luz a través de colonias de
briozoarios Membranipora tuberculata en el alga roja Gelidium rex Santelices &
Abbott 1965, observando que la densidad del flujo de fotones disminuye hasta en
un 56 %; sin embargo aunque el briozoario reduce la tasa fotosintética del alga, no
presenta un efecto neto sobre el crecimiento de las algas.

6
Molina et al. (1991) analizaron la concentración de pigmentos en talos de
Gelidium rex conforme a cambios de colonización del epibionte M. tuberculata
tanto en condiciones naturales como en laboratorio. Se obtuvo una diferencia
significativa en ficobilinas, clorofila α y carotenos totales en algas, siendo mayor en
talos con briozoarios, indicando que las algas pueden compensar pigmentos como
respuesta a la disminución de la densidad de flujo fotónico causado por el
epibionte.
Muñoz et al. (1991) midieron en condiciones de laboratorio los efectos de
M. tuberculata en la eficiencia fotosintética de G. rex., demostrando que los
briozoarios incrustantes pueden reducir la luz incidental hasta un 44%, sin
embargo los pigmentos accesorios son significativamente mayores en talos
incrustados, por lo que la eficiencia es casi idéntica en ambos talos. Así que
aunque los libres de epífitos mostraron una mayor fotosíntesis, los talos con
incrustaciones exhibieron un valor semejante, por lo que el efecto perjudicial se
puede compensar por CO2 liberado por el briozoario como fuente inorgánica
fotosintética.
Hurd et al. (1994) estudiaron los efectos de la colonización de briozoario
Membranipora membranacea en las algas Agarum fimbriatum Harvey, 1862 y
Macrocystis integrifolia Bory de Saint-Vincent 1826 (Vancouver, British Columbia,
Canadá) con relación a estimaciones de rangos de nitratos y amonio. Obtuvieron
que el briozoario se puede presentar como un factor positivo, presentándose como

7
una fuente importante de nitrógeno inorgánico para M. integrifolia mas no para A.
fimbriatum.
Actualmente los estudios que muestran la relación entre el género del alga
Gelidium y el briozoario M. tuberculata sólo hacen alusión a la especie Gelidium
rex, pero no a Gelidium robustum, siendo esta especie estudiada únicamente por
la producción de agar, así como sus cultivos correspondientes, y no desde una
perspectiva ecológica. A su vez los estudios sobre epifitismo en las costas de Baja
California se desarrollan en su mayoría a un nivel bacteriológico, por lo que no se
estudia el aspecto de la sucesión ecológica de especies.
2.2 Metabolitos secundarios aislados de algas rojas con actividad antiepífita.
Para que se produzca el asentamiento de diferentes especies se debe dar
primeramente un proceso de colonización bacteriana, la cual fomentará una
sucesión ecológica (Gómez-Alcalá, 2012). Con el paso de los años se han
presentado diversos estudios que demuestran que extractos obtenidos de
especies pertenecientes al grupo de las Rodophytas presentan una gran
efectividad contra bacterias; dentro de los primeros trabajos se encuentra el de
Mautner et al. (1953) donde reportan la actividad antibacteriana del alga
Rhodomela larix (Turner) C. Agardh 1822 (actualmente sinónimo de
Neorhodomela larix (Turner) Masuda 1982) (Zaleta-Pinet, 2009).
De Nys et al. (1995) investigaron la actividad anti-incrustante del alga roja
Delisea pulchra (Greville) Montagne 1844, probando metabolitos aislados en

8
ensayos contra larvas de balano Balanus amphitrite Darwin 1854, alga Ulva
lactuca Linnaeus 1753 y una bacteria marina (Cepa SW8). Como resultado se
obtuvo la inhibición de larvas de balano e incluso de gametos del alga D. pulchra,
así como de la cepa bacteriana. El efecto inhibitorio presentó un mejor resultado
en conjunto con un antibiótico, gentamicina común, presentando a la vez el mismo
efecto que pinturas anti incrustantes.
En los trabajos de antibacterianos regionales destacan los de Durán-Riverol
(2009) y Zaleta-Pinet (2009), los cuales probaron la actividad antibacteriana de
diferentes fracciones de extractos orgánicos de algas del género Gracilaria
Greville 1830 contra diferentes cepas de bacterias de los géneros Vibrio y
Staphylococcus, presentando actividad contra Vibrio parahaemolyticus y
Staphylococcus aureus, siendo estas especies bacterianas las que intervienen en
el proceso de epifitismo.
Ríos et al. (2009) evaluaron las propiedades bioactivas antibacterianas y
antimicóticas de diferentes extractos de especies de algas marinas en San Juan
de Los Cayos, Venezuela, obteniendo que tanto Gracilaria sp. como Gelidium sp.
resultaron bioactivas contra diferentes cepas.
Águila-Ramírez et al. (2012) evaluaron el efecto de extractos orgánicos de
diferentes macroalgas en la inhibición del crecimiento de epibiontes, empleando
siete especies recolectadas en el arrecife de Punta Arenas, BCS, México, y
obtuvieron que el alga roja Laurencia johnstonii Setchell & Gardner 1924 resultó

9
con la mayor actividad anti-incrustante frente a cepas de bacterias marinas y
terrestres, así como con microalgas marinas y cepas de hongos.
2.3 Características biológicas de Gelidium robustum
Gelidium robustum (Gardner) Hollenberg y Abbott, denominada
comúnmente como gelidio y sargazo rojo, perteneciente al orden de las
Gelidiales, es un alga recta de color rojo vino con ejes inferiores no ramificados
(Fig. 1). La parte superior presenta ramificación lateral, pinada, plana y de
contorno piramidal. Su altura promedio radica entre los 31 y 37 cm, aunque
pueden encontrarse organismos de hasta un metro de longitud (Guzmán del Próo
et al., 1986).
Figura 1. Alga Gelidium robustum

10
Esta especie de alga se caracteriza por presentarse en fondos rocosos o
constituidos por sustrato duro, en zonas con oleaje intenso y movimiento continuo;
forma mantos densos en hábitats sombreados distribuyéndose desde el límite
inferior de la marea baja hasta profundidades entre los 15 y 17 metros. Suele
encontrarse como vegetación asociada a los mantos de Macrocystis pyrifera
(Guzmán del Próo et al., 1986).
Gelidium robustum tiene un ciclo de vida con alternancia de generaciones
isomórfico y trifásico (Fig. 2). Presenta gametofitos haploides masculinos y
femeninos de vida libre y dos fases diploides, una de vida libre (tetraesporofito) y
otra (carposporofito) que parasita al gametofito femenino. Las estructuras
reproductoras se presentan durante todo el año, siendo considerablemente más
abundantes las plantas tetraspóricas respecto a las carpospóricas a lo largo del
año; estas últimas tienen una mayor fecundidad y un período de actividad
reproductora más prolongado (Guzmán del Próo et al., 1972; Cole y Sheath,
1990).
La distribución de esta especie en la costa occidental de Norteamérica
comprende desde el sur de la Columbia Británica (Canadá) hasta Isla Margarita en
Baja California Sur (Guzmán del Próo et al., 1986).

11
Figura 2. Ciclo de vida de Gelidium robustum (Hernández-Guerrero, 1997).
3. Justificación
En el mundo existe un gran interés en las especies de algas rojas del
género Gelidium debido a la alta calidad del agar que se obtiene de éstas, así
como la demanda en el mercado de sus derivados para aplicación en la industria
alimenticia y farmacéutica como estabilizador, emulsificador y gelificante. El alga
roja Gelidium robustum es la principal fuente de agar en México, siendo la
segunda alga más importante para su explotación.
Aunque la importancia económica de este recurso es conocida, no se han
desarrollado estudios ecológicos en torno a su fauna de acompañamiento y las
interacciones que presenta con relación a ella para entender los efectos negativos
o positivos que podrían tener directamente con el alga. Uno de los mayores
problemas que presenta G. robustum es la relación con el briozoario epífito
Tricogino
Carpogonio
Tetrasporofito

12
Membranipora tuberculata denominado “conchilla”, el cual se presenta como una
cubierta calcárea en los ejes y frondas basales del alga. Esto genera problemas
en la industria de Gelidium debido a la baja calidad de un porcentaje y rendimiento
del producto. El valor comercial de G. robustum está determinado por la presencia
del briozoario, disminuyendo su calidad así como su precio en el mercado; el
Gelidium de primera calidad no presenta ninguna placa calcárea, mientras que
algas con gran cantidad de placas calcáreas se denominan de tercera o cuarta
calidad.
Algunos estudios de relaciones entre algas y epibiontes muestran que
dentro de los efectos de la epifauna en cultivos comerciales de algas marinas, se
encuentra la pérdida parcial o total de las frondas debido al peso que adquieren
las algas con epibiontes o una disminución en el crecimiento del hospedero, así
como disminución en sus procesos de captación fotosintéticos. La realización de
este estudio interviene en la comprensión de la relación entre los epibiontes y su
hospedero, sirviendo para plantear en lo futuro diferentes metodologías para la
prevención contra el epibionte en cultivos comerciales.
La comprensión de las funciones ecológicas por medio de los metabolitos
secundarios ayuda a definir el rol ecológico de cada organismo, formulando los
nichos ecológicos de los mismos, así como la composición de una comunidad y
facilitando la comprensión de las interacciones ecológicas. Establecer si el alga
presenta sus metabolitos secundarios como medio de defensa o como

13
compensación de pigmentos fotosintéticos puede plantear una representación
evolutiva bajo la selección natural por parte de ambos organismos, pudiendo
elucidar una relación de coevolución. Asimismo, el estudio de organismos
epibiontes es de importancia debido a la obtención de compuestos con actividad
anti-incrustante, sentando bases en lo futuro para el desarrollo de productos
comerciales como pinturas o insecticidas no contaminantes que produzcan
alteraciones bioquímicas al ambiente.
4. Objetivos
4.1 Objetivo general
Establecer si existen diferencias en composición de los metabolitos de
Gelidium robustum asociada a la colonización del briozoario epífito Membranipora
tuberculata.
4.2 Objetivos específicos
Obtener diferentes extractos etanólicos a partir del alga G. robustum.
Fraccionar los extractos orgánicos por medio de métodos cromatográficos.
Probar los diferentes extractos/fracciones para observar si presentan
actividad antibacteriana.
Establecer la diferencia en la composición relativa de metabolitos entre
extractos empleando técnicas espectroscópicas.

14
5. Material y métodos
5.1 Recolección del alga
Se recolectó material algal de la especie Gelidium robustum con diferentes
coberturas de briozoario Membranipora tuberculata en aguas superficiales de 7-10
m por medio de buceo SCUBA, en Puerto Escondido, al sur de Bahía Tortugas,
Baja California Sur, México encontrado en las coordenadas 27º41'30" N,
114º53'45" O. La identificación taxonómica fue realizada por el Dr. Rafael
Riosmena Rodríguez en el laboratorio de Botánica Marina de la Universidad
Autónoma de Baja California Sur.
El material recolectado se lavó y se le retiró el exceso de sal, dejando el
epibionte presente en el alga. Posteriormente se trasladó a las instalaciones del
Centro Interdisciplinario de Ciencias Marinas, donde se separó con base en su
porcentaje de cobertura en 0% de epífito, 0-30%, 30-60%, >60%, 100% y algas
con una cobertura de 100% limpiadas posteriormente de epífitos; las muestras se
etiquetaron y guardaron en bolsas plásticas a temperatura ambiente hasta el
momento de su extracción.
5.2 Obtención de extractos etanólicos
Para la obtención de extractos crudos etanólicos se pesaron 800 g de alga
seca y molida para cada caso de porcentaje de cobertura, siendo 0-30% (12-001-

15
40), 30-60% (12-002-41), >60% (12-003-40), 100% (12-04-40) y 100% con epífitos
retirados posteriormente (12-004-40A), así como uno adicional empleando sólo el
briozoario retirado (12-004-40B). Se realizaron extracciones empleando 5 ml de
etanol destilado (EtOH) por cada gramo de muestra, filtrando la mezcla para
extraer el residuo en dos ocasiones más empleando las mismas condiciones. Los
extractos resultantes se concentraron a sequedad en rotavapor a 40°C (±2 °C) y
se transfirieron a viales previamente tarados y etiquetados como 12-001-40, 12-
002-40, 12-003-40, 12-004-40, 12-004-40A y 12-004-40B.
5.2.1 Pruebas cromatográficas de extractos
Para determinar la influencia del porcentaje de cobertura del briozoario M.
tuberculata sobre la producción de metabolitos secundarios en el alga G. robustum
y diferenciar la producción de metabolitos entre cada uno, se analizó la
composición química relativa a través de sus respectivos perfiles cromatográficos
empleando una disolución de 10 mg de muestra en 1 ml de EtOH por cada
tratamiento (12-001-40, 12-002-41, 12-003-40, 12-04-40, 12-004-40A y 12-004-
40B) sobre placas de sílica gel. Para los análisis por cromatografía de capa fina
(CCF) se probaron diferentes sistemas de elución en cámaras de saturación
empleando disolventes puros y posteriormente mezclas para obtener el mejor
gradiente de concentración. Para el revelado de las placas se utilizó ácido
sulfúrico (H2SO4) al 10 % y vainillina empleando calor con una plancha eléctrica.

16
5.2.2 Fraccionamiento de extractos etanólicos
Una vez seleccionados los metabolitos secundarios de interés de los
extractos crudos 12-001-40, 12-002-40, 12-003-40 y 12-004-40 se unieron con el
fin de obtener una mayor cantidad de compuesto para aislar (14 g, separando 100
mg de muestra por tratamiento para la posterior realización de pruebas anti-
incrustantes y análisis espectroscópico), dando lugar a un nuevo extracto
denominado 12-001234-40, el cual se sometió a un fraccionamiento por medio de
una extracción en fase sólida sobre sílica gel fase normal (muestra:sílica 1:10), la
elución se realizó con diclorometano, mezclas de diclorometano y etanol (9:1, 7:3
y 5:5) y finalmente etanol. A partir de ese procedimiento se obtuvieron 20 eluatos,
los cuales se emplearon para realizar un perfil de elución por medio de
cromatografía de capa fina. De acuerdo a las diferencias de concentración del
metabolito secundario se unieron los eluatos semejantes formando tres fracciones,
12-001234-40 F1 (1-10), 12-001234-40 F2 (11-14) y 12-001234 F3 (15-20), las
cuales fueron llevadas a sequedad en rotavapor a 40°C (±2°C), para
posteriormente realizar pruebas de cromatografía de capa fina empleando una
disolución de 10 mg mL-1 de EtOH.
5.3 Evaluación de actividad antibacteriana
Con el fin de comprobar si G. robustum presenta actividad biológica anti-
incrustante contra epibiontes no briozoarios, se determinó el efecto antibacteriano

17
de los extractos crudos 12-001-40, 12-002-40, 12-003-40, 12-004-40 y las tres
fracciones 12-001234-40 F1, 12-001234-40 F2 y 12-001234 F3 contra diferentes
cepas bacterianas tanto terrestres (Staphylococcus aureus, Pseudomonas
aeruginosa y Escherichia coli) como marinas (Vibrio parahaemolyticus y Vibrio
harveyi), por el método de difusión en agar con discos.
Para reactivar las cepas, fueron sembradas en agar marino [1 g de extracto
de levadura (DIFCO), 5 g de peptona de carne (DIFCO), 15 g de Bacto agar
(DIFCO) en 1 L de agua de mar artificial (Coral Reef)] e incubadas durante 14 h.
Una vez transcurrido el tiempo de incubación, se sembraron en tubos de agar
inclinado (dos tubos de trabajo, uno de reserva) y se incubaron nuevamente
durante 24 horas y posteriormente fueron almacenadas a 11 °C.
Para la preparación de los sensidiscos se empleó el método de difusión en
agar descrito por Brock y Madigan (1993) para la realización de pruebas de
sensibilidad antibacteriana, utilizando discos de papel filtro Whatman No. 4., de 7
mm, esterilizados previamente en autoclave. Estos discos se impregnaron con una
disolución de 2 mg mL-1 de los extractos y fracciones a probar diluidos en
disolventes apropiados. La preparación se llevó a cabo en una campana de flujo
laminar desinfectada con solución de cloruro de benzalconio, sobre papel aluminio
lavado con etanol para asegurar la esterilidad. En esas condiciones los discos se
dejaron secar y se almacenaron en frascos de vidrio esterilizados de 7 mL, a -
18°C por un período no mayor a 48 h. De igual manera se prepararon controles

18
negativos con los disolventes puros utilizados durante la extracción y
fraccionamiento. Se utilizaron sensidiscos impregnados con oxitetraciclina como
control positivo de inhibición.
Para la preparación de inóculos se sembraron cultivos masivos en agar,
utilizando agar marino para las bacterias marinas y agar tripticaseina-soya (TSA)
(DIFCO) para las bacterias no marinas, incubándolas durante 24 h. Una vez
obtenida biomasa suficiente de cada especie bacteriana, se tomó una parte de ella
con un hisopo estéril y se suspendió en solución salina estéril (2.5 % NaCl para
bacterias marinas y 0.85 % para bacterias no marinas), hasta obtener una
densidad óptica de aproximadamente 1.00 UA, a una longitud de onda 585 nm.
Con cada especie bacteriana se inocularon masivamente placas de agar
Muller-Hilton (Merck ®) (2.5 % NaCl), sobre las cuales se colocaron los
sensidiscos con los extractos y fracciones de prueba y los controles positivos y
negativos. Estas placas con sensidiscos se dejaron a temperatura ambiente (24°C
aproximadamente) por 1.5 h para permitir la difusión del compuesto en el agar y
posteriormente se incubaron a 30°C por 24 h. El efecto sobre las bacterias fue
registrado como negativo cuando no se observó inhibición del crecimiento y en los
casos positivos, las zonas de inhibición alrededor de cada disco fueron medidas
en milímetros. Las pruebas fueron realizadas por duplicado.

19
5.4 Identificación estructural por análisis espectroscópico
Con el fin de observar la diferencia metabólica entre los porcentajes de
cobertura, se realizó un análisis de las diferencias en la composición relativa de
las bandas de los extractos (16 picos) 12-001-40, 12-002-40, 12-003-40 y 12-004-
40 de G. robustum esperando observar una diferenciación entre las áreas de los
picos de cada extracto para obtener alguna banda diagnóstica. A su vez se analizó
la estructura de la fracción 12-001234-40 F3, la cual presentaba la mayor
concentración del metabolito polar encontrado. Para ello se empleó un análisis de
espectroscopia de infrarrojo utilizando un espectrómetro FTIR- Modelo Spectrum
TwoTM Perkin-Elmer y se leyó en un espectro entre 4,000 y 250 cm-1 realizando
una corrección por medio del Software Spectrum de Perkin-Elmer. Los resultados
obtenidos se analizaron y compararon con bases de datos de espectros infrarrojos
así como con literatura especializada.
5.5 Cálculos y análisis estadísticos
Para la realización de una comparación cuantitativa entre los extractos
crudos se calculó el área de cada banda por medio del Software Spectrum de
Perkin-Elmer; estos datos fueron sometidos a un análisis de proporciones
comparando cada banda del espectro IR con las demás, formando una matriz por
cada extracto. Posteriormente los datos entre extractos se compararon para
determinar el aumento o disminución de compuestos conforme aumentaba el

20
porcentaje de cobertura por M. tuberculata en el alga. Estas mediciones fueron
realizadas con el programa Microsoft Excel 2010.
Finalmente las bandas que mostraban diferencia entre sus valores de áreas
de bandas fueron analizadas por medio de Análisis de Varianza (ANDEVA) de una
vía con ayuda del Software STATISTICA 7, para ver si existían diferencias
verdaderamente significativas.
6. Resultados
Para los análisis cromatográficos de los diferentes extractos etanólicos de
G. robustum a diferentes porcentajes de cobertura por el briozoario M. tuberculata,
se obtuvo que el mejor disolvente fue una mezcla de diclorometano:etanol (7:3), el
cual reveló en la placa un metabolito altamente polar (mancha gris) en todos los
extractos algales; sin embargo se mostró evidencia de que dicho metabolito
disminuía conforme aumentaba el porcentaje de cobertura por briozoario en el
alga (Fig. 3).
A su vez se pudo apreciar un posible aumento de otros metabolitos
secundarios conforme aumentó el porcentaje de cobertura (Fig. 4), siendo
posiblemente metabolitos como medio de defensa. En el extracto conformado sólo
por el briozoario M. tuberculata no se mostró efecto metabólico alguno, por lo que
ninguna de las manchas de compuestos encontradas se le atribuye a este

21
organismo. Al realizar el fraccionamiento del extracto 12-001234-40 se mostró que
la fracción 12-001234-40 F3 conformada por los eluatos 15-20, eluída en etanol al
100%, resultó altamente enriquecida con el metabolito polar (RF de 0.3), a
diferencia de las otras dos fracciones.
Figura 3. Cromatografía de extractos crudos de G. robustum
Figura 4. Fracciones 12-001234-40F1, 12-001234-40F2 y 12-001234-40F3
Donde: 1 Extracto 12-001234-40 (0-30%) 2 Extracto 12-001234-40 (30-60%) 3 Extracto 12-001234-40 (>60%) 4 Extracto 12-001234-40 (100%) Fase estacionaria: Placas de sílica gel fase normal. Fase móvil: CH2Cl2: EtOH (7:3) Revelador: H2SO4 10% y vainillina
F1 F2 F3
22
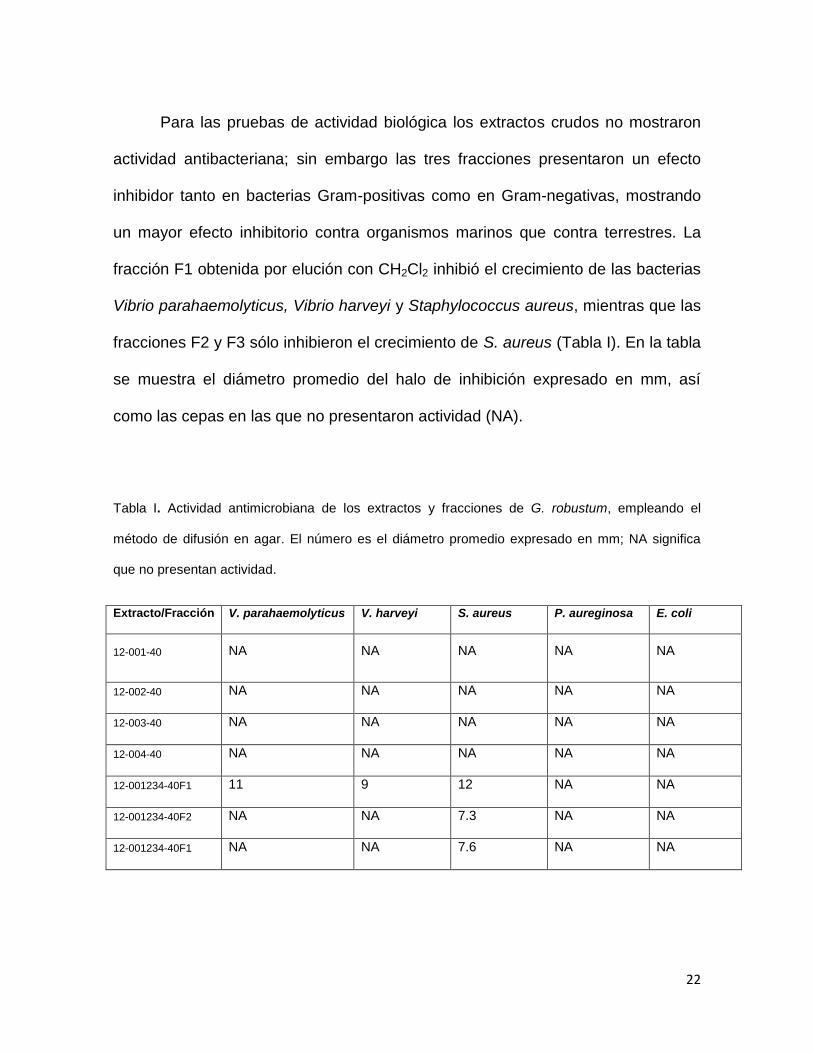
Para las pruebas de actividad biológica los extractos crudos no mostraron
actividad antibacteriana; sin embargo las tres fracciones presentaron un efecto
inhibidor tanto en bacterias Gram-positivas como en Gram-negativas, mostrando
un mayor efecto inhibitorio contra organismos marinos que contra terrestres. La
fracción F1 obtenida por elución con CH2Cl2 inhibió el crecimiento de las bacterias
Vibrio parahaemolyticus, Vibrio harveyi y Staphylococcus aureus, mientras que las
fracciones F2 y F3 sólo inhibieron el crecimiento de S. aureus (Tabla I). En la tabla
se muestra el diámetro promedio del halo de inhibición expresado en mm, así
como las cepas en las que no presentaron actividad (NA).
Tabla I. Actividad antimicrobiana de los extractos y fracciones de G. robustum, empleando el
método de difusión en agar. El número es el diámetro promedio expresado en mm; NA significa
que no presentan actividad.
Extracto/Fracción V. parahaemolyticus V. harveyi S. aureus P. aureginosa E. coli
12-001-40 NA NA NA NA NA
12-002-40 NA NA NA NA NA
12-003-40 NA NA NA NA NA
12-004-40 NA NA NA NA NA
12-001234-40F1 11 9 12 NA NA
12-001234-40F2 NA NA 7.3 NA NA
12-001234-40F1 NA NA 7.6 NA NA

23
De las señales observadas en los espectros infrarrojos de los extractos 12-
001-40, 12-002-40, 12-003-40 y 12-004-40 se distinguen 4 bandas de absorción
principales: la banda en el rango de 3400 cm-1 que corresponde un grupo hidroxilo,
mientras que las bandas 1634 y 1407 a enlaces de grupo carboxilo. En la zona
denominada huella dactilar que se ubica entre los 600 y 1400 cm-1 se puede
distinguir una banda de 1029.32 cm-1 indicando la presencia de un anillo de
azúcar; sin embargo otras señales no identificadas pueden indicar la presencia de
alcanos, alcoholes, esteres, éteres o ácidos carboxílicos (Fig. 5). Los espectros
infrarrojos mostraron semejanza entre los cuatro extractos, siendo encontrados los
mismos compuestos entre ellos, con 16 bandas de absorción, así como
presentando las señales propias de un compuesto orgánico indicadas por las
bandas de absorción principales. Asimismo el análisis de proporciones indicó una
posible banda diagnóstica, presentando un pico a los 820.21 cm-1, la cual
disminuye con base en la proporción de áreas conforme aumenta el porcentaje de
cobertura, por lo que se muestra una posible diferencia metabólica entre los
diferentes extractos. Sin embargo estos análisis de proporción no mostraron ser
significativos en cuanto a correlaciones entre bandas, por lo que no se pudo
determinar una relación lineal entre la disminución de estas proporciones y el
aumento de cobertura por conchilla.

24
% T
ransm
itan
cia
Longitud de onda (cm-1)
2 3 4 5
1 8
7
6
12 13 9 10 11
15 16 14
Figura 5. Espectro infrarrojo de los extractos crudos de G. robustum con diferentes porcentajes de
cobertura de M. tuberculata y sus respectivos números de banda.

25
Figura 6. Espectro infrarrojo de la fracción 12-001234-40 F3.
La fracción 12-001234-40 F3, que es la que presentó la mayor
concentración del metabolito polar, muestra un gran parecido con los extractos
crudos, presentando a diferencia de las otras dos fracciones un grupo hidroxilo en
la banda 3307 cm-1 y un prolongado pico en 1028.07 cm-1 e indicando la presencia
de anillo de azúcar, lo que posiblemente podría atribuirse al metabolito polar
aislado.
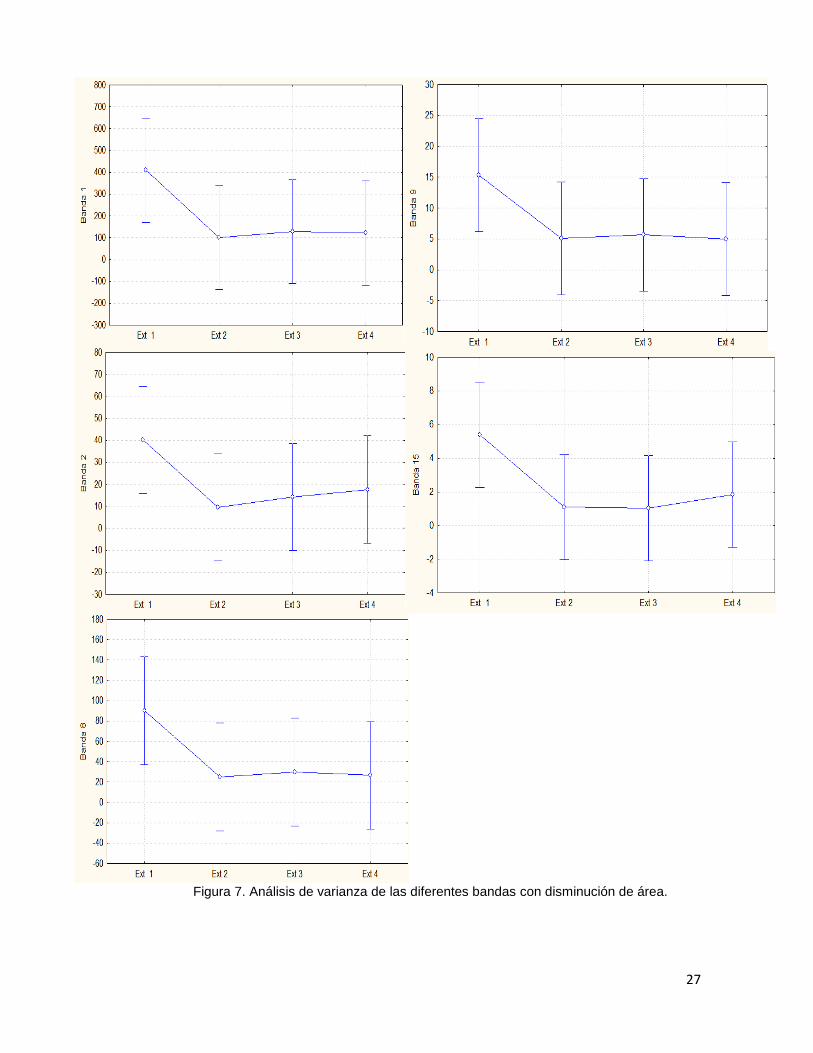
La determinación del área de las diferentes bandas mostró una variación
entre extractos, en donde se aprecia una disminución a partir del extracto 2 (mayor
a 30% de cobertura). Los picos que presentaron una disminución notable entre
tratamientos fueron las bandas 1 (3339 cm-1), 2 (1634 cm-1), 8 (1029 cm-1), 9 (976
cm-1) y 15 (601 cm-1). Sin embargo los resultados obtenidos para las pruebas de

26
análisis de varianza (Fig. 7) indicaron que no existe una diferencia significativa por
áreas de bandas entre extractos; obteniendo para banda 1 (ANOVA, F=1.5074, P=
.22178), banda 2 (F=1.2564, P=29747), banda 8 (F= 1.4255, P=24421), banda 9
(F=1.2200, P= .31029) y banda 15 (F=1.7573, P=.16500).
27
Figura 7. Análisis de varianza de las diferentes bandas con disminución de área.

28
7. Discusión
Las pruebas de cromatografía de capa fina revelaron la presencia de un
metabolito altamente polar, el cual se presenta en todos los extractos crudos
conformados por el alga G. robustum (Fig. 3). Sin embargo se observó que dicho
metabolito disminuye conforme aumenta el porcentaje de cobertura del epibionte
Membranipora tuberculata; esto indica que el briozoario presenta un efecto
negativo sobre el alga alterando las concentraciones en los metabolitos presentes
en ésta; y que el metabolito secundario puede ser un producto derivado de la
fotosíntesis. Hay estudios que muestran que el briozoario M. tuberculata presenta
efectos negativos de forma directa en la fotosíntesis de algas del género Gelidium,
donde al cubrir la superficie de este organismo disminuye el flujo de fotones hasta
en un 56% alterando la intensidad y calidad del paso de luz, así como la reducción
de luz incidental hasta en un 44% (Cancino et al., 1987; Muñoz et al., 1991).
En la placa revelada (Fig. 3) se puede observar también la aparición de
otros posibles compuestos que aumentan conforme se incrementa el porcentaje
de cobertura, lo que podría ser interpretado como respuesta del mecanismo de
defensa química del alga contra el epibionte. En algas los metabolitos secundarios
parecen funcionar en general como mecanismos de defensa contra depredadores
o como cierta capacidad regenerativa al ser dañados o consumidos. Dentro de
estos metabolitos podemos encontrar compuestos de origen biogenético, como
aminoácidos, terpenodies, florotaninos, derivados del ácido acrílico, esfingolípidos,
esteroides, entre otras (Newman et al., 2003; Amsler, 2008). Esta respuesta de

29
defensa de mecanismo puede explicarse por dos teorías: la teoría de la defensa
óptima, propuesta por Rhoades (1979), donde se dice que las plantas asignan
recursos de defensa al valor adaptativo de sus tejidos; o la teoría más empleada
en la actualidad, estableciendo que el efecto de la respuesta metabólica del alga
va en función de las variables ambientales (McClintock y Baker, 1997).
Otra opción podría ser que los compuestos que van en aumento (Fig. 3) no
sean mecanismos de defensa, sino una compensación fotosintética como
respuesta a la colonización de M. tuberculata, tratándose de un aumento de
pigmentos auxiliares. Molina et al. (1991) establecieron que talos de Gelidium rex
con un mayor porcentaje de colonización por briozoario presentaban una mayor
cantidad de pigmentos fotosintéticos como ficobilinas, clorofila α y carotenos,
indicando que las algas pueden presentar una compensación fotosintética como
respuesta a la disminución de la densidad de flujo fotónico causado por el
epibionte. Para establecer e identificar de cuáles compuestos se trata realmente,
se debe continuar el trabajo con aislamiento y purificación de metabolitos
secundarios presentes en el alga. A su vez los análisis de varianza (Fig. 7) no
mostraron que algún compuesto aumentara de manera significativa, por el
contrario, los que presentaron diferencia fue en relación con una disminución, por
lo que no se podría decir que algún compuesto esté en aumento de concentración.
Aunque ya se había reportado actividad anti-incrustante en algas rojas
(Águila-Ramírez et al., 2012), no se muestra otro trabajo anteriormente procedente

30
de Gelidium robustum con aplicación antibacteriana. En su trabajo Águila-Ramírez
et al., emplearon algas Rhodophytas de la misma subclase que G. robustum,
contra la bacteria S. aureus, donde Gymnogongrus martinensis no presentó
actividad y Laurencia johnstonii presentó con 18.7 mm un halo de inhibición mayor
que G. robustum con sólo de 12 mm. Con base en esto se puede determinar que
G. robustum es un alga capaz de sintetizar compuestos activos contra organismos
terrestres y marinos, siendo tanto Gram-positivos como Gram-negativos.
Al presentarse un aumento en el briozoario M. tuberculata, el cual coloniza
las frondas del alga, y no presentarse un efecto de defensa, se puede decir que G.
robustum no presenta un efecto anti-incrustante contra briozoarios; sin embargo
no se puede percibir ningún otro epífito encontrado en el alga, por lo que podría
tener actividad contra otro tipo de organismos invasores presentando algún tipo de
infoquímico como medio de defensa. En el presente trabajo las fracciones F1, F2 y
F3 resultaron activas contra la bacteria terrestre (Gram-positiva) Staphylococcus
aureus y la bacteria marina (Gram-negativa) Vibrio parahaemolyticus, las cuales
están involucradas en el proceso del micro-epifitismo, produciendo un mucílago o
sustancia exopolimérica que les permite a los nuevos epibiontes establecerse
sobre un sustrato más adecuado, dando lugar a una sucesión ecológica (Bakus et
al., 1986; Henschell y Cook, 1990).
En el trabajo se mostró que el alga Gelidium puede presentar actividad anti-
incrustante contra organismos epífitos; sin embargo no se presentó efecto contra

31
el briozoario M. tuberculata. Esto se puede deber a que G. robustum no presenta
un compuesto específico para el briozoario o que este organismo presenta
resistencia hacia los metabolitos del alga. Existen estudios en los cuales se
demuestra que los hospederos son excepcionalmente resistentes a incrustaciones
de organismos debido a sus compuestos; sin embargo éstos suelen presentarse
en su superficie y no transmitidos en el agua, por lo que en ocasiones no afecta al
epibionte que la coloniza (Hay, 1996).
La mayoría de las publicaciones sugieren que los organismos epibiontes
presentan una marcada y negativa influencia en el alga hospedera al interferir en
la captación de luz o de nutrientes (Dixon et al., 1981). Sin embargo hay autores
que indican que no existen evidencias de efectos negativos en el crecimiento,
sobrevivencia y fecundidad del alga colonizada; afirmando, por el contrario, que
existe una relación de beneficio donde el epibionte brinda amonio y dióxido de
carbono cuando las concentraciones de estas moléculas se ven limitadas en el
ambiente (De Burg y Frankboner, 1978; Taylor y Ress, 1988).
Los análisis de espectro IR (Fig. 5) mostraron similitudes entre los
diferentes porcentajes de cobertura, presentando distintos grupos en común como
el enlace hidroxilo encontrado en la banda de 3399 cm-1, así como el anillo de
azúcar en la banda 1029 cm-1 ,los cuáles podrían establecer que se presenta un
compuesto de origen glúsido como carbohidrato. Los grupos carboxilos
encontrados en las bandas 1634.01 y 1407. 92 cm-1, podrían tratarse de derivados

32
de polisacáridos, como ácido glucorónico o ácido pirúvico (Vilar et al., 2011).
Aunque el trabajo de Nelson et al. (1994) establece que en la zona de la huella
digital podríamos encontrar galactanos sulfatados, pertenecientes al grupo éster, y
agarocoloides cercanos a 930 cm-1, esto no es posible debido al tipo de extracción
empleando etanol en lugar de agua. Sin embargo podría tratarse de algún tipo de
carbohidrato de bajo peso molecular como reserva, como almidón floridiano
(Ascêncio, et al., 2006).
Los análisis de varianza de bandas de absorción mostraron que no existe
una diferencia significativa (Fig. 7); sin embargo en la placa de cromatografía de
capa fina se puede observar cómo disminuye la concentración de un compuesto
(Fig. 3) mientras más porcentaje de conchilla presente. A su vez en las áreas de
bandas indican una posible disminución muy representativa en el grupo hidroxilo,
así como en ciertos grupos carboxilo y en el pico del anillo de azúcar. La
disminución en las bandas de grupos de azúcares podría indicar que al interferir el
briozoario en el proceso fotosintético, el alga presenta problemas en la producción
de azúcares que constituyen sus diferentes estructuras celulares. Aguilar-Rosas et
al. (2003) presentaron en su trabajo mantos de Gelidium robustum en las costas
de Baja California Sur, con una cobertura de sus talos por conchilla en un 70 %,
obteniendo aun así una talla record de 180 cm. Esto podría indicar que aunque se
presenten efectos negativos en los metabolitos secundarios del alga, no afecta su
crecimiento al obtener tallas muy grandes. Esto podría corroborar los estudios que
sugieren que la colonización no interfiere en los costes metabólicos al aumentar

33
las algas la concentración de sus pigmentos fotosintéticos, aumentando su
eficiencia fotosintética (Cancino et al., 1987; Hurd et al., 1994).
Pelegrín et al. (1999) indicaron que aunque la calidad del agar se veía
disminuida entre mayor concentración de conchilla, sus características de gel no;
mientras que Hurtado et al. (2011) indicaron que no encontraron relación entre la
calidad del agar y la cobertura de conchilla debido a la baja concentración que
encontraron en sus muestras. El efecto negativo podría encontrarse en el
rendimiento del alga al presentar gran colonización en lugar de encontrarse en la
calidad del agar del mismo, siendo que en la producción un gran porcentaje se
pierde al ser sólo placa calcárea de la conchilla.
La actividad biológica que presentan los extractos etanólicos y las
fracciones de G. robustum difieren; esto se debe a que las sustancias activas se
van separando y concentrando en ciertas fracciones, siendo por eso que las
fracciones tienden a inhibir el crecimiento de microorganismos mientras que los
extractos crudos no. Este mismo problema se ve reflejado en el trabajo anti-epífito
de Gómez-Alcalá (2012), donde puede presentar un gran problema al no
emplearse una metodología muy sensible o si se utilizan bajas concentraciones
del principio activo, en donde puede pasar inadvertida la actividad del extracto.
También puede ocurrir que una fracción no sea activa mientras que el extracto
crudo sí por cuestiones de concentración, porque se pierde la actividad debido que
al reaccionar los grupos funcionales originan subproductos inactivos, o porque la

34
actividad se debe a la sinergia entre los componentes de la mezcla, perdiendo la
efectividad al separarse (Bhakuni y Rawat, 2005).
El uso de técnicas espectroscópicas de infrarrojo no se había empleado con
anterioridad para establecer diferencias en la composición relativa de compuestos
de algas utilizando las bandas de absorción como patrones diagnósticos. La
técnica resultó efectiva para cuantificar el área de dichas bandas, sin embargo no
se presentó como una técnica que presentara resultados concluyentes de manera
robusta para la diferenciación significativa entre los diferentes porcentajes de
cobertura, dando un resultado similar al de los métodos de cuantificación de
pigmentos fotosintéticos al no presentar resultados de manera significativa (Dixon
et al, 1981; Cancino et al., 1987).
8. Conclusiones
A partir de los extractos etanólicos de G. robustum se logró el aislamiento
de un metabolito polar. El análisis cualitativo de los extractos crudos por medio de
técnicas cromatográficas mostró diferencias en la concentración del metabolito
aislado, encontrando que disminuye conforme aumenta el porcentaje de cobertura
de briozoario M. tuberculata.
Se observó una relación de aumento de unos compuestos en capa fina
conforme aumenta el porcentaje de cobertura. Sin embargo se desconoce si se

35
trata de una respuesta de defensa metabólica o compensación fotosintética, por lo
que se sugiere continuar con fraccionamiento y purificación de extractos.
La fracción 12-001234-40F1 mostró actividad antibacteriana contra cepas
de V. parahaemolyticus, V. harveyi y S. aureus, las cuales están involucradas en
el proceso de epifitismo. G. robustum presentó compuestos antibacterianos que se
aplican tanto a bacterias marinas como a terrestres, así como para Gram-positivas
y para Gram-negativas.
El análisis espectroscópico de IR mostró similitudes entre los diferentes
extractos etanólicos; sin embargo aunque tras una cuantificación del área de
bandas de absorción se observó que ciertas bandas en relación con los azúcares
disminuyen conforme aumenta el porcentaje de cobertura del briozoario, no se
presentaron diferencias significativas por medio de un análisis de varianza.
El briozoario M. tuberculata podría presentar un efecto negativo sobre el
alga G. robustum interfiriendo en el proceso fotosintético, provocando alteraciones
en su metabolismo y disminuyendo sus compuestos derivados por estos procesos;
sin embargo parece no alterar el crecimiento del alga.
El método de cuantificación de bandas diagnósticas para establecer la
diferencia en la composición relativa de metabolitos, no se presentó como técnica
robusta capaz de predecir la diferencia metabólica entre extractos.

36
9. Literatura citada
Aguilar Rosas, L.E.; R. Aguilar Rosas y D. Lora Sánchez. 2003. Registro de talla
record para Gelidium robustum (Gardner) Hollenberg et Abbot
(GELIDIALES; RHODOPHYTA) en la costa Pacífico de México. Oceánides.
18 (2): 83-85.
Aguilar Rosas, R. y L. E. Aguilar Rosas. 1990. La Conchilla contra el sargazo rojo.
Conciencia. 5: 5-6.
Águila-Ramírez, R. N.; A. Arenas-González y C. J. Hernández-Guerrero. 2012.
Actividades antimicrobiana y anti-incrustante obtenidas de los extractos de
algas marinas del Golfo de California, México. Hidrobiología. 22 (1): 8-15.
Aguilar-Rosas, L.E.; R. Aguilar-Rosas y D, Lora-Sánchez. 1998. El Sargazo rojo:
un alga marina de importancia económica en Baja California. Divulgare. 23:
4-17.
Amsler, C. D. 2008. Algal Chemical Ecology. Springer. Alemania. 313 pp.
Ascêncio, S. D.; A. Orsato, R. A. Franca, M. E. Duarte y M. D. Noseda. 2006.
Complete 1H and 13C NMR assignment of digeneaside, a low-molecular-
mass carbohydrate produced by red seaweeds. Carbohydrate research. 341
(5): 677-682.
Bakus, G. J.; N. M. Targett y B. Shulte. 1986. Chemical ecology of marine
organisms: an overview. Journal of Chemical Ecology. 12 (5): 951-987.

37
Bergström, G. 2007. Chemical ecology = chemistry + ecology. Pure Appl. Chem.
79 (12): 2305-2323.
Bhakuni, D. S. y D. S. Rawat. 2005. Bioactive Marine Natural Products. Springer.
New Delhi, India. 400 pp.
Brock, T. D. y M. T. Madigan. 1993. Microbiología. Pretince Hall
Hispanoamericana, S. A. México. 956 p.
Cancino, J. M.; J. Muñoz, M. Muñoz y M. C. Orellana. 1987. Effects of the
bryozoan Membranipora tuberculata (Bosc.) on the photosynthesis and
growth of Gelidium rex Santelices et Abbott. J. Exp. Mar. Biol. 113: 105-112.
Casas V. M. y C. J. Hernández G. 1996. Pesquería de Gelidium robustum. En:
Casas V. M., y G. Ponce (Eds.). Estudio del Potencial Pesquero y Acuícola
de Baja California Sur Vol. II. SEMARNAP, Gobierno del Estado de Baja
California Sur, FAO, UABCS, CIBNOR, CICIMAR, Inst. Nal. de la Pesca y
CETMAR 419-429.
Casas-Valdez, M. M. y C. Fajardo-León. 1990. Análisis preliminar de la explotación
de Gelidium robustum (Gardner) Hollenberg y Abbot en Baja California Sur,
México. Investigaciones Marinas CICIMAR. 5(1): 83-86.
Cole, M. K. y R. G. Sheath. 1990. Biology of the red algae. Cambridge University
Press. E.U.A. 517 p.
Cortez, Vieyle. 2013. Ecología química y perspectiva de su aplicación en la
conservación de la biodiversidad. Cuadernos de Biodiversidad. 41: 16-21.

38
De Burg, M. E. y P.V. Frankboner. 1978. A nutritional association between the bull
kelp Nereocystis luetkeana and its epizoic bryozoan Membranipora
membranacea. En: Hepburn, C. D. y C. L. Hurd. 2005. Conditional
mutualism between the giant kelp Macrocystis pyrifera and colonial
epifauna. Marine Ecology Progress Series. 302: 37-48.
De Nys, R.; P. D. Steinberg, P. Willemsen, S. A. Dworjanyn, C. L. Gabelish y R. J.
King. 1995. Broad spectrum effects of secondary metabolites from the red
alga Delisea pulchra in antigouling assays. Biofouling: The Journal of
Bioadhesion and Biofilm Research. 8 (4): 259-271.
Dicke, M. y M. W. Sabelis. 1988. Infochemical terminology: base on cost-benefit
analysis rather than origin of compounds? Func. Ecol. 2: 131-139.
Dixon, J.; S. C. Schroeter y J. Kastendiek. 1981. Effects of the encrusting bryozoa,
Membranipora membranacea, on the loss of blades and fronds by the giant
Kelp, Macrocystis pyrifera (Laminariales). Journal of Phycology. 17 (4): 341-
345.
Durán-Riverol, L. M. 2009. Estudio de la actividad frente a bacterias patógenas de
cinco especies algales: pros y contras del protocolo de aislamiento guiado
por bioensayo. Tesis de Maestría. CICIMAR-IPN. La Paz, Baja California
Sur. 126 pp.
Freile-Peregrín, Y.; D. Robledo y E. Serviere-Zaragoza. 1999. Gelidium robustum
agar: quality characteristics from exploited beds and seasonality an
unexplited bed at Southern Baja California, México.

39
Gómez-Alcalá, 2012. Efecto in vitro del extracto etanólico de Macrocystis pyrifera
sobre el desarrollo de microepífitos. Tesis de Licenciatura. Universidad
Autónoma de Baja California Sur. La Paz, B.C.S. 49 pp.
Guzmán del Próo, S. A. y L. Granados. 1968. Programa nacional de algas
marinas. Serie Trab. De Divul. Direc. Gral. Pesca. 13 (130): 3-26.
Guzmán del Próo, S. A. y S. De la Campa. 1969. Investigaciones sobre Gelidium
cartilagineum en la costa occidental de Baja California, México. Proc. Sixth
Int. Seaweed Symp. 6: 179-186.
Guzmán del Próo, S. A.; M. Casas, A. Díaz, J. Pineda y M. E. Sánchez. 1986.
Diagnóstico sobre las investigaciones y explotación de las algas marinas en
México. Inv. Mar. CICIMAR. 3 (2): 1-63.
Guzmán del Próo, S. A.; S. De la Campa y J. Pineda. 1972. Shedding rhythm and
germination of spores in Gelidium robustum. Proc. Int. Seaweed Symp. 7:
221-228.
Hay, M. E. 1996. Marine chemical ecology: what’s known and what´s next?.
Journal of Experimental Marine Biology and Ecology. 200: 103-134.
Henschell, J.R. y P.A. Cook. 1990. The development of a marine fouling
community in relation to the primary film of microorganisms. Biofouling. 2
(1): 1-11.
Hernández-Guerrero, C. J. 1997. Análisis de la variación estacional e interanual de
la cosecha de Gelidium robustum en Baja California Sur, México. CICIMAR-
IPN. Tesis de Maestría. México. 60 pp.

40
Hurd, C. L.; K. M. Durarte, F.S. Chia y P. J. Harrison. 1994. Effect of bryozoan
colonization on inorganic nitrogen acquisition by the kelps Agarum
fimbriatum and Macrocystis integrifolia. Marine Biology. 121: 167-173.
Hurtado, M. A.; M. Manzano-Sarabia, E. Hernández-Garibay, I. Pacheco-Ruiz y J.
A. Zertuche-González. 2011. Latitudinal variations of the yield and quality of
agar from Gelidium robustum (Gelidiales, Rhodophyta) from the main
commercial harvest bedsalong the western coast of the Baja California
Peninsula, Mexico. J. Apply Phycology. 23: 727-734.
Law, J.H. y F. E. Regnier. 1971. Pheroones. Annual. Rev. Biochem. 40: 533-548.
McClintock, J. B. y B. J. Baker. 1997. Marine Chemical Ecology. 1997. CRC Press.
EUA. 610 pp.
Molina, M. 1986. Notas sobre tres especies de algas marinas: Macrocystis
pyrifera, Gelidium robustum y Gigartina canaliculata, de interés comercial en
la costa occidental de Baja California, México. Documento Técnico
Informativo. Sepesca. 3: 16-39.
Molina, X.; J. M. Cancino y V. Montecino. 1991. Cambios en los pigmentos
fotosintetizadores de Gelidium rex (Rhodophyta) inducidos por el epibionte
Membranipora tuberculata (Bryozoa). Revista Chilena de Historia Natural.
64: 289-297.
Muñoz, J., J. M. Cancino y M. X. Molina. 1991. Effect of Encrusting Bryozoans on
the Physiology of Their Algal Substratum. Journal of the Marina Biological
Association of the United Kingdom. 71 (4): 877-882.

41
Nelson, W. A.; G. A. Knight, R. Falshaw, R.H. Furneaux, A. Falshaw y S. M. Lynds.
1994. Characterisation of the enigmatic, endemic red alga Gelidium allani
(Gelidiales) from northern New Zeland – morphology, distribution, agar
chemistry. Journal of Applied Phycology. 6 (5): 497-507.
Newman, D. J.; G. M. y K. M. Snader. 2003. Natural products as sources of new
drugs over the period 1981-2002. Journal of Natural Product. 66 (7): 1022-
1037.
Rhoades, D. F. 1979. Evolution of plant chemical defense against herbivores. En:
Herbivores: their interaction with secondary plant metabolites. Academic
Press. EUA. 353 pp.
Ríos, N.; G. Medina, J. Jiménez, C. Yánez, M. Y. García, M. L. Di Bernardo y M.
Gualtieri. 2009. Actividad antibacteriana y antifúngica de extractos de algas
marinas venezolanas. Rev. peru.biol. 16 (1): 97-100.
Rodríguez-Vargas, D. J.; J. González-González y E. Serviere-Zaragoza. 1993.
Gelidáceas (Rhodophyta) en el Pacífico Tropical. En: Salazar-Vallejo y N. E.
González (eds) Biodiversidad marina y costera de México. Com. Nac.
Biodiversidad y CIQRO, México. 445-455 p.
Taylor, R. B. y T. A. Rees. 1988. Excretory products of mobile epifauna as a
nitrogen source for seaweeds. Limnology & Oceanography. 43: 600-606.
Vilar, V. J.; C. M. Botelho y R. A. Boaventura. 2011. Environmental fruendly
technologies for wastewaters treatment: Biosorption of heavy metals using

42
low cost materials and solar photocatalysis. Security of Industrial Supply and
Management. 159-173.
Wahl, M. 1989. Marine epibiosis I. Fouling and antifouling: some basic aspects.
Marine Ecology Progress Series. 58: 175-189.
Williams, D. H.; M. J. Stone, P. R. Hauck y S. K. Rahman. 1989. Why are
secondary metabolites (Natural Products) biosynthesized?. J. Nat. Prod. 52
(6): 1189-1208.
Zaleta-Pinet D.A. 2009. Actividad antimicrobiana contra 6 especies del género
Vibrio de extractos orgánicos y fracciones de alga roja Gracilaria
vermiculophylla (Ohmi) Papenfuss. Tesis de Licenciatura. Universidad
Autónoma de Baja California. La Paz, B. C. S.

43
10. Anexos
Anexo 1. Áreas de las bandas analizadas por espectro infrarrojo para cada extracto.
Espectro Área Altura Eje X Eje Y
Banda 1
12-001-40 -12690.56 -31.84 3340.43 68.16
12-002-40 -16338.73 -38.81 3336.33 61.19
12-003-40 -16150.29 -38.68 3348.06 61.32
12-004-40 -17021.41 -39.17 3339.03 60.83
Banda 2
12-001-40 -1249.25 -14.25 1634.04 85.75
12-002-40 -1557.91 -15.94 1642.73 84.06
12-003-40 -1781.2 -17.41 1642.69 82.59
12-004-40 -2457.83 -22.31 1634.01 77.69
Banda 3
12-001-40 -180.25 -4.28 1436.57 95.72
12-002-40 -461.83 -9.53 1464.84 90.47
12-003-40 -398.56 -8.67 1464.47 91.33
12-004-40 -568.8 -11.15 1465.56 88.85
Banda 4
12-001-40 -350.4 -6.02 1417.06 93.98
12-002-40 -731.53 -12.64 1411.33 87.36
12-003-40 -670.72 -11.47 1410.29 88.53
12-004-40 -890.44 -15.26 1407.92 84.74
Banda 5
12-001-40 -195.5 -4.93 1336.8 95.07
12-002-40 -369.19 -9.24 1338.81 90.76
12-003-40 -342.14 -8.54 1339.13 91.46
12-004-40 -404.82 -10.07 1339.67 89.93
Banda 6
12-001-40 -681.4 -9.79 1206.53 90.21
12-002-40 -1856.14 -25.14 1209.07 74.86
12-003-40 -1669.29 -22.59 1208.83 77.41
12-004-40 -2053.32 -27.51 1190.86 72.49
Banda 7
12-001-40 -498.67 -14.66 1147.52 85.34
12-002-40 -993.04 -29.34 1146.28 70.66
12-003-40 -905.33 -26.72 1146.61 73.28
12-004-40 -1002.62 -28.47 1147.09 71.53
Banda 8
12-001-40 -2800.85 -35.42 1028.04 64.58
12-002-40 -4029.17 -51.76 1025.9 48.24
12-003-40 -3772.76 -47.95 1026.48 52.05
12-004-40 -3733.12 -48.61 1029.32 51.39

44
Banda 9
12-001-40 -474.56 -14.8 983.03 85.2
12-002-40 -819.97 -22.17 976.42 77.83
12-003-40 -712.33 -19.63 976.85 80.37
12-004-40 -691.47 -18.34 976.7 81.66
Banda 10
12-001-40 -10.04 -0.86 907.55 99.14
12-002-40 -50.02 -3.02 906.96 96.98
12-003-40 -39.2 -2.45 907.28 97.55
12-004-40 -50.08 -2.85 906.17 97.15
Banda 11
12-001-40 -14.79 -0.97 862.85 99.03
12-002-40 -109.19 -4.49 861.32 95.51
12-003-40 -80.9 -3.49 861.83 96.51
12-004-40 -71.33 -2.9 860.61 97.1
Banda 12
12-001-40 -3.64 -0.38 819.26 99.62
12-002-40 -27.57 -1.84 819.29 98.16
12-003-40 -21.42 -1.44 819.71 98.56
12-004-40 -19.08 -1.22 820.21 98.78
Banda 13
12-001-40 -72.94 -2.54 783.1 97.46
12-002-40 -195.44 -6.2 780.97 93.8
12-003-40 -157.98 -5.21 781.74 94.79
12-004-40 -197.35 -5.96 771.4 94.04
Banda 14
12-001-40 -40.5 -2.15 744.56 97.85
12-002-40 -122.44 -6.55 744.01 93.45
12-003-40 -90.81 -4.95 744 95.05
12-004-40 -155.93 -7.42 743.35 92.58
Banda 15
12-001-40 -167.59 -5.14 603.14 94.86
12-002-40 -178.69 -6.6 601.49 93.4
12-003-40 -130.68 -4.93 599.86 95.07
12-004-40 -257.22 -8.78 601.81 91.22
Banda 16
12-001-40 -104.96 -3.63 531.11 96.37
12-002-40 -186.93 -7.19 524.23 92.81
12-003-40 -146.81 -5.64 523.6 94.36
12-004-40 -227.14 -8.38 524.17 91.62

45
Anexo 2. Análisis de proporción de área de bandas de extracto 12-001-40.
12-001-40 Banda 1 Banda 2 Banda 3 Banda 4 Banda 5 Banda 6 Banda 7 Banda 8 Banda 9 Banda 10 Banda 11 Banda 12 Banda 13 Banda 14 Banda 15 Banda 16
Banda 1 1.000 0.098 0.014 0.028 0.015 0.054 0.039 0.221 0.037 0.001 0.001 0.000 0.006 0.003 0.013 0.008
Banda 2 10.159 1.000 0.144 0.280 0.156 0.545 0.399 2.242 0.380 0.008 0.012 0.003 0.058 0.032 0.134 0.084
Banda 3 70.405 6.931 1.000 1.944 1.085 3.780 2.767 15.539 2.633 0.056 0.082 0.020 0.405 0.225 0.930 0.582
Banda 4 36.217 3.565 0.514 1.000 0.558 1.945 1.423 7.993 1.354 0.029 0.042 0.010 0.208 0.116 0.478 0.300
Banda 5 64.913 6.390 0.922 1.792 1.000 3.485 2.551 14.327 2.427 0.051 0.076 0.019 0.373 0.207 0.857 0.537
Banda 6 18.624 1.833 0.265 0.514 0.287 1.000 0.732 4.110 0.696 0.015 0.022 0.005 0.107 0.059 0.246 0.154
Banda 7 25.449 2.505 0.361 0.703 0.392 1.366 1.000 5.617 0.952 0.020 0.030 0.007 0.146 0.081 0.336 0.210
Banda 8 4.531 0.446 0.064 0.125 0.070 0.243 0.178 1.000 0.169 0.004 0.005 0.001 0.026 0.014 0.060 0.037
Banda 9 26.742 2.632 0.380 0.738 0.412 1.436 1.051 5.902 1.000 0.021 0.031 0.008 0.154 0.085 0.353 0.221
Banda 10 1264.000 124.427 17.953 34.900 19.472 67.869 49.668 278.969 47.267 1.000 1.473 0.363 7.265 4.034 16.692 10.454
Banda 11 858.050 84.466 12.187 23.692 13.218 46.072 33.717 189.375 32.087 0.679 1.000 0.246 4.932 2.738 11.331 7.097
Banda 12 3486.418 343.201 49.519 96.264 53.709 187.198 136.997 769.464 130.374 2.758 4.063 1.000 20.038 11.126 46.041 28.835
Banda 13 173.986 17.127 2.471 4.804 2.680 9.342 6.837 38.399 6.506 0.138 0.203 0.050 1.000 0.555 2.298 1.439
Banda 14 313.347 30.846 4.451 8.652 4.827 16.825 12.313 69.157 11.718 0.248 0.365 0.090 1.801 1.000 4.138 2.592
Banda 15 75.724 7.454 1.076 2.091 1.167 4.066 2.976 16.713 2.832 0.060 0.088 0.022 0.435 0.242 1.000 0.626
Banda 16 120.909 11.902 1.717 3.338 1.863 6.492 4.751 26.685 4.521 0.096 0.141 0.035 0.695 0.386 1.597 1.000

46
Anexo 3. Análisis de proporción de área de bandas de extracto 12-002-40.
12-002-40 Banda 1 Banda 2 Banda 3 Banda 4 Banda 5 Banda 6 Banda 7 Banda 8 Banda 9 Banda
10
Banda
11
Banda
12
Banda
13
Banda
14
Banda
15
Banda
16
Banda 1 1.000 0.095 0.028 0.045 0.023 0.114 0.061 0.247 0.050 0.003 0.007 0.002 0.012 0.007 0.011 0.014
Banda 2 10.488 1.000 0.296 0.470 0.237 1.191 0.637 2.586 0.526 0.032 0.070 0.018 0.125 0.079 0.115 0.146
Banda 3 35.378 3.373 1.000 1.584 0.799 4.019 2.150 8.724 1.775 0.108 0.236 0.060 0.423 0.265 0.387 0.492
Banda 4 22.335 2.130 0.631 1.000 0.505 2.537 1.357 5.508 1.121 0.068 0.149 0.038 0.267 0.167 0.244 0.310
Banda 5 44.256 4.220 1.251 1.981 1.000 5.028 2.690 10.914 2.221 0.135 0.296 0.075 0.529 0.332 0.484 0.615
Banda 6 8.803 0.839 0.249 0.394 0.199 1.000 0.535 2.171 0.442 0.027 0.059 0.015 0.105 0.066 0.096 0.122
Banda 7 16.453 1.569 0.465 0.737 0.372 1.869 1.000 4.057 0.826 0.050 0.110 0.028 0.197 0.123 0.180 0.229
Banda 8 4.055 0.387 0.115 0.182 0.092 0.461 0.246 1.000 0.204 0.012 0.027 0.007 0.049 0.030 0.044 0.056
Banda 9 19.926 1.900 0.563 0.892 0.450 2.264 1.211 4.914 1.000 0.061 0.133 0.034 0.238 0.149 0.218 0.277
Banda 10 326.644 31.146 9.233 14.625 7.381 37.108 19.853 80.551 16.393 1.000 2.183 0.551 3.907 2.448 3.572 4.541
Banda 11 149.636 14.268 4.230 6.700 3.381 16.999 9.095 36.901 7.510 0.458 1.000 0.252 1.790 1.121 1.637 2.080
Banda 12 592.627 56.507 16.751 26.534 13.391 67.325 36.019 146.143 29.741 1.814 3.960 1.000 7.089 4.441 6.481 8.239
Banda 13 83.600 7.971 2.363 3.743 1.889 9.497 5.081 20.616 4.196 0.256 0.559 0.141 1.000 0.626 0.914 1.162
Banda 14 133.443 12.724 3.772 5.975 3.015 15.160 8.110 32.907 6.697 0.409 0.892 0.225 1.596 1.000 1.459 1.855
Banda 15 91.436 8.719 2.585 4.094 2.066 10.387 5.557 22.548 4.589 0.280 0.611 0.154 1.094 0.685 1.000 1.271
Banda 16 71.932 6.859 2.033 3.221 1.625 8.172 4.372 17.739 3.610 0.220 0.481 0.121 0.860 0.539 0.787 1.000
47
Anexo 4. Análisis de proporción de área de bandas de extracto 12-003-40.
12-003-40 Banda 1 Banda 2 Banda 3 Banda 4 Banda 5 Banda 6 Banda 7 Banda 8 Banda 9 Banda
10
Banda
11
Banda
12
Banda
13
Banda
14
Banda
15
Banda
16
Banda 1 1.000 0.110 0.025 0.042 0.021 0.103 0.056 0.234 0.044 0.002 0.005 0.001 0.010 0.006 0.008 0.009
Banda 2 9.067 1.000 0.224 0.377 0.192 0.937 0.508 2.118 0.400 0.022 0.045 0.012 0.089 0.051 0.073 0.082
Banda 3 40.522 4.469 1.000 1.683 0.858 4.188 2.272 9.466 1.787 0.098 0.203 0.054 0.396 0.228 0.328 0.368
Banda 4 24.079 2.656 0.594 1.000 0.510 2.489 1.350 5.625 1.062 0.058 0.121 0.032 0.236 0.135 0.195 0.219
Banda 5 47.204 5.206 1.165 1.960 1.000 4.879 2.646 11.027 2.082 0.115 0.236 0.063 0.462 0.265 0.382 0.429
Banda 6 9.675 1.067 0.239 0.402 0.205 1.000 0.542 2.260 0.427 0.023 0.048 0.013 0.095 0.054 0.078 0.088
Banda 7 17.839 1.967 0.440 0.741 0.378 1.844 1.000 4.167 0.787 0.043 0.089 0.024 0.174 0.100 0.144 0.162
Banda 8 4.281 0.472 0.106 0.178 0.091 0.442 0.240 1.000 0.189 0.010 0.021 0.006 0.042 0.024 0.035 0.039
Banda 9 22.672 2.501 0.560 0.942 0.480 2.343 1.271 5.296 1.000 0.055 0.114 0.030 0.222 0.127 0.183 0.206
Banda 10 411.997 45.439 10.167 17.110 8.728 42.584 23.095 96.244 18.172 1.000 2.064 0.546 4.030 2.317 3.334 3.745
Banda 11 199.633 22.017 4.927 8.291 4.229 20.634 11.191 46.635 8.805 0.485 1.000 0.265 1.953 1.122 1.615 1.815
Banda 12 753.982 83.156 18.607 31.313 15.973 77.931 42.266 176.133 33.255 1.830 3.777 1.000 7.375 4.239 6.101 6.854
Banda 13 102.230 11.275 2.523 4.246 2.166 10.566 5.731 23.881 4.509 0.248 0.512 0.136 1.000 0.575 0.827 0.929
Banda 14 177.847 19.615 4.389 7.386 3.768 18.382 9.969 41.546 7.844 0.432 0.891 0.236 1.740 1.000 1.439 1.617
Banda 15 123.587 13.630 3.050 5.133 2.618 12.774 6.928 28.870 5.451 0.300 0.619 0.164 1.209 0.695 1.000 1.123
Banda 16 110.008 12.133 2.715 4.569 2.330 11.370 6.167 25.698 4.852 0.267 0.551 0.146 1.076 0.619 0.890 1.000

48
Anexo 5. Análisis de proporción de área de bandas de extracto 12-004-40.
12-004-40 Banda 1 Banda 2 Banda 3 Banda 4 Banda 5 Banda 6 Banda 7 Banda 8 Banda 9 Banda 10 Banda 11 Banda 12 Banda 13 Banda 14 Banda 15 Banda 16
Banda 1 1.000 0.144 0.033 0.052 0.024 0.121 0.059 0.219 0.041 0.003 0.004 0.001 0.012 0.009 0.015 0.013
Banda 2 6.925 1.000 0.231 0.362 0.165 0.835 0.408 1.519 0.281 0.020 0.029 0.008 0.080 0.063 0.105 0.092
Banda 3 29.925 4.321 1.000 1.565 0.712 3.610 1.763 6.563 1.216 0.088 0.125 0.034 0.347 0.274 0.452 0.399
Banda 4 19.116 2.760 0.639 1.000 0.455 2.306 1.126 4.192 0.777 0.056 0.080 0.021 0.222 0.175 0.289 0.255
Banda 5 42.047 6.071 1.405 2.200 1.000 5.072 2.477 9.222 1.708 0.124 0.176 0.047 0.488 0.385 0.635 0.561
Banda 6 8.290 1.197 0.277 0.434 0.197 1.000 0.488 1.818 0.337 0.024 0.035 0.009 0.096 0.076 0.125 0.111
Banda 7 16.977 2.451 0.567 0.888 0.404 2.048 1.000 3.723 0.690 0.050 0.071 0.019 0.197 0.156 0.257 0.227
Banda 8 4.560 0.658 0.152 0.239 0.108 0.550 0.269 1.000 0.185 0.013 0.019 0.005 0.053 0.042 0.069 0.061
Banda 9 24.616 3.554 0.823 1.288 0.585 2.969 1.450 5.399 1.000 0.072 0.103 0.028 0.285 0.226 0.372 0.328
Banda 10 339.884 49.078 11.358 17.780 8.083 41.001 20.020 74.543 13.807 1.000 1.424 0.381 3.941 3.114 5.136 4.536
Banda 11 238.629 34.457 7.974 12.483 5.675 28.786 14.056 52.336 9.694 0.702 1.000 0.267 2.767 2.186 3.606 3.184
Banda 12 892.107 128.817 29.811 46.669 21.217 107.616 52.548 195.656 36.241 2.625 3.738 1.000 10.343 8.172 13.481 11.905
Banda 13 86.250 12.454 2.882 4.512 2.051 10.404 5.080 18.916 3.504 0.254 0.361 0.097 1.000 0.790 1.303 1.151
Banda 14 109.161 15.762 3.648 5.711 2.596 13.168 6.430 23.941 4.434 0.321 0.457 0.122 1.266 1.000 1.650 1.457
Banda 15 66.175 9.555 2.211 3.462 1.574 7.983 3.898 14.513 2.688 0.195 0.277 0.074 0.767 0.606 1.000 0.883
Banda 16 74.938 10.821 2.504 3.920 1.782 9.040 4.414 16.435 3.044 0.220 0.314 0.084 0.869 0.686 1.132 1.000





























