Embed Size (px)
Citation preview

1
The kinesin-like protein TOP promotes Aurora
localisation and induces mitochondrial, chloroplast and
nuclear division
Authors
Yamato Yoshida1, *, Takayuki Fujiwara2, Yuuta Imoto1, 3, Masaki Yoshida4, Mio Ohnuma1,
Shunsuke Hirooka5, 6, Osami Misumi7, Haruko Kuroiwa1, Shoichi Kato8, Sachihiro
Matsunaga8 and Tsuneyoshi Kuroiwa1, ‡
Affiliations
1Graduate School of Science, Rikkyo University, 3-34-1 Nishiikebukuro, Toshima-ku,
Tokyo 171-8501, Japan
2Chromosome Dynamics Laboratory, RIKEN Advanced Science Institute, 2-1 Hirosawa,
Wako, Saitama 351-0198, Japan
3Department of Integrated Biosciences, Graduate school of Frontier Sciences, The
University of Tokyo, 5-1-5 Kashiwanoha, Kashiwa, Chiba 277-8562, Japan
4Integrative Environmental Sciences, Graduate School of Life and Environmental Sciences,
University of Tsukuba, 1-1-1 Tennoudai, Tsukuba, Ibaraki 305-8572, Japan
5Center for Frontier Research, National Institute of Genetics, 1111 Yata,
Mishima, Shizuoka 411-8540, Japan
6Core Research for Evolutional Science and Technology (CREST),
Japan Science and Technology Agency, Gobancho, Chiyoda-ku, Tokyo
102-0076, Japan
7Graduate School of Medicine, Faculty of Science, Department of Biological
Science and Chemistry, Yamaguchi University, 1677-1 Yoshida, Yamaguchi, Yamaguchi
753-8512, Japan
© 2012. Published by The Company of Biologists Ltd.Jo
urna
l of C
ell S
cien
ceA
ccep
ted
man
uscr
ipt
JCS Advance Online Article. Posted on 2 April 2013

2
8Department of Applied Biological Science, Faculty of Science and Technology, Tokyo
University of Science, 2641 Yamazaki, Noda, Chiba 278-8510, Japan
*Present address: Department of Plant biology, Michigan State University, East Lansing,
MI 48824, USA
‡Correspondence and requests for materials should be addressed to T. K. (e-mail:
Running title TOP induces organelle divisions
Key words cell-division cycle / Cyanidioschyzon merolae / kinesin / mitochondrial
division / plastid division
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

3
The cell cycle usually refers to the mitotic cycle, but the cell-division cycle in the plant
kingdom consists of not only nuclear but also mitochondrial and chloroplast division
cycle. However an integrated control system that initiates division of the three
organelles has not been found. We first report that a novel C-terminal kinesin-like
protein, three-organelle divisions inducing protein (TOP), controls nuclear,
mitochondrial and chloroplast divisions in red alga Cyanidioschyzon merolae. A
proteomics revealed that TOP was contained in the complex of mitochondrial-dividing
(MD) and plastid-dividing (PD) machineries (MD/PD machinery complex) just prior
to constriction. After TOP localized on the MD/PD machinery complex, mitochondrial
and chloroplast divisions were performed and the components of the MD/PD
machinery complexes were phosphorylated. Furthermore, TOP down-regulation
impaired both mitochondrial and chloroplast divisions. MD/PD machinery complexes
were formed normally at each division site but they were neither phosphorylated nor
constricted in these cells. Immunofluorescence signals of Aurora kinase (AUR) were
localized around the MD machinery before constriction whereas AUR was dispersed
in cytosol by TOP down-regulation, suggesting that AUR is presumably required for
the constriction. Taken together, TOP is likely to induce protein phosphorylation of
MD/PD machinery components to accomplish mitochondrial and chloroplast divisions
prior to nuclear division by transferring of AUR. Concurrently, the involvement of
TOP in mitochondrial and chloroplast division, given the presence of TOP homologs
throughout eukaryotes, may illuminate the original function of C-terminal kinesin-
like proteins.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

4
Introduction
Almost all eukaryotic cells in the plant kingdom possess three kinds of organelles that
contain DNA and have double-membranes: one nucleus, many mitochondria and
chloroplasts (plastids). Since mitochondria and chloroplasts were derived from free-living
α-proteobacterial and cyanobacterial ancestors, respectively, they are never synthesized de
novo and their continuities are maintained by division, same as the nucleus. The cell cycle,
therefore, consists of not only the mitotic cycle but also mitochondrial and chloroplast
division cycles (Suzuki et al., 1994; Imoto et al., 2010). In plant cells, mitochondrial and
plastid DNA replications take place before nuclear DNA replication (Kobayashi et al.,
2009; Imoto et al., 2010). Recently, Kobayashi et al. (2009) has shown that the proceeding
of mitochondrial and plastid DNA replications is signalled by the intracellular
accumulation of a tetrapyrrole intermediate, probably Mg-ProtoIX, resulting in the
activation of cyclin-dependent kinase A (CDKA, also known as Cdc2 in fission yeast) and
the consequent initiation of nuclear DNA replication in Cyanidioschyzon merolae and
tobacco BY-2 cells (Kobayashi et al., 2009). Therefore, it seems that an integrated control
system which induces division of the three types of double-membrane organelles may be
hidden in the initiation step of mitochondrial and plastid divisions.
In the last two decades, it has been shown that the mitochondrial and chloroplast
divisions are performed by following three steps: formation of mitochondrial-dividing
(MD) machinery and plastid-dividing (PD) machineries at each division site (Kuroiwa et al.,
1998), constriction of the division site, and pinching-off the bridge of daughter organelles
(Fig. 1A). During the formation step, MD and PD machineries are connected with each
other and form a complex structure (MD/PD machinery complex), but the complex
structure of MD and PD machineries separates in the constriction step (Fig. 1A) (Yoshida
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

5
et al., 2009). Both MD and PD division machineries comprised a chimera of inner rings of
bacterial-derived proteins, such as FtsZ (Osteryoung and Nunnari, 2003; Kuroiwa et al.,
2008) and eukaryote specific proteins, such as the MD ring, PD ring and dynamin proteins
(Bleazard et al., 1999; Miyagishima et al., 2003; Gao et al., 2003; Nishida et al., 2003;
Osteryoung and Nunnari, 2003; Kuroiwa et al., 2008). Recent studies showed that dynamin
are localized between the PD ring filaments and are essential to generate the motive force
for contraction (Yoshida et al., 2006). In addition, the PD ring is constructed by a bundle of
glycosyltransferase protein PDR1 mediated-polyglucan filaments. Thus, the contraction of
the PD machineries is caused by the sliding movement between dynamin and polyglucan
filaments (Yoshida et al., 2010). Similarly, it has been thought that the contraction of the
MD machineries is probably driven in the same way as the PD machinery (Nishida et al.,
2003; Yoshida et al., 2009; Kuroiwa et al., 2008).
Besides, reports thus far showed that a pre-spindle structure designated as
mitochondrial spindle was formed from each spindle pole to the division site of
mitochondria in C. merolae which precedes nuclear division (Fig. S1, white arrowheads)
(Nishida et al., 2005; Imoto et al., 2010). Similar to spindle for cell division (Hirokawa,
1998; Walczak and Heald, 2008), the mitochondrial spindle may be also organized by
tubulin mediated microtubule polymers, several types of kinesin-superfamily proteins, and
many relating factors such as mitotic-kinases.
Our recent study showed that a mitotic-serine/threonine kinase Aurora kinase (AUR),
which was encoded a single-copy gene in the genome C. merolae, was localized at not only
spindle poles and spindle for cell division but also the mitochondrial spindle (Kato et al.,
2011). Specifically, AUR was accumulated from the mitochondrial spindle to the
mitochondrial division site in contraction phase of mitochondrial division, implying that
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

6
involves in the activation of the MD machinery. Thus, these facts suggest the existence of
an uncharacterized pathway or factor which coordinates timing of mitochondrial and
chloroplast division and links mitochondrial- and chloroplast-division cycles with the cell-
division cycle.
C. merolae offers advantages for studying the regulation of integrated initiation of
nuclear, mitochondrial and chloroplast divisions. The cell contains just one chloroplast,
one mitochondrion and one nucleus (Matsuzaki et al., 2004), the division of which occurs
in that order in highly synchronized cells by light/dark cycles (Fig. S1) (Suzuki et al., 1994).
In addition, availability of the complete genome sequence of C. merolae facilitates highly
sensitive transcriptomic and proteomic analyses (Matsuzaki et al., 2004; Nozaki et al.,
2007; Yagisawa et al., 2009; Fujiwara et al., 2009; Yoshida et al., 2009, 2010).
In this study, we revealed that a novel kinesin-like protein TOP induces three
organelle divisions of nuclear, mitochondrion, and chloroplast. TOP localized on the
MD/PD machinery complex to transfer AUR just before the contraction of the MD and PD
machineries. By this process, mitochondrion and chloroplast were divided by the MD and
PD machineries, respectively. And then, nuclear division was also accomplished by TOP-
mediated spindle. Thus, TOP regulates three organelle divisions of nuclear, mitochondrion,
and chloroplast through transferring of AUR proteins.
Results
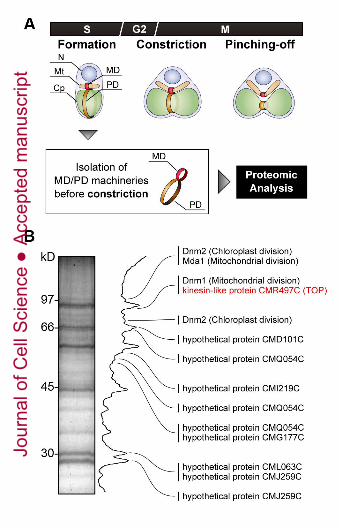
Identification of a novel kinesin-like protein TOP in the MD/PD machinery complexes
To identify proteins that regulate the integrated initiation of nuclear, mitochondrial
and chloroplast division, we performed a proteomic analysis of isolated MD/PD machinery
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

7
complex from cells in early S phase (Fig. 1B, Fig. S2 and Table S1, see methods) because
division of the organelles and organization of the mitochondrial spindle begins immediately
after this phase. For isolation of non-constricting MD/PD machinery complexes,
synchronised cells were harvested at the start of the second dark period (S phase) and
treated with Nonidet P-40 and n-octyl-β-D-glucopyranoside (see methods). After isolation
of the non-constricting MD/PD machinery complexes, proteomic analysis was performed
by using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry
(MALDI-TOF-MS). In this fraction, known components of the MD and PD machinery,
Dynamin 1 (Dnm1), Mda1 and Dnm2 were certainly identified. Comparing previous results
of proteomic analyses of the MD/PD machineries isolated from cells in M phase (Yoshida
et al., 2009, 2010), we identified a novel S phase specific protein TOP (three-organelle
divisions inducing protein) (Fig. 1B). By the completely sequenced genomic information of
the C. merolae (http://merolae.biol.s.u-tokyo.ac.jp/), we distinguished that TOP was one of
the COOH-terminal motor domain-type kinesin-superfamily proteins (C-terminal KIFs,
reviewed in Hirokawa et al., 2009) (Fig. 2A), which had never been detected in isolated
constricting MD and PD machineries from cells in M phase (Yoshida et al., 2009, 2010).
By phylogenetic analysis, we showed that a kinesin motor domain of TOP was classified in
classical C-terminal KIFs and TOP homologs are widely conserved throughout eukaryotic
cells, especially plants (Fig. S3). Thus far, some groups of C-terminal KIFs have been
reported to transport organelles (Saito et al., 1997; Xu et al., 2002; Bananis et al., 2004;
Hirokawa et al., 2009). Also, one of the known functions of C-terminal KIFs is the
assembly of spindle poles by the cross-linking of parallel microtubules in each half-spindle,
where they focus minus ends into spindle poles (Walczak et al., 1997; Hirokawa, 1998),
however, the involvement of TOP in mitochondrial and/or chloroplast division has never
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

8
previously been reported. We examined the function of TOP to reveal the regulation
mechanism of nuclear, mitochondrial and chloroplast division.
The genes of known MD and PD machinery-associated proteins (Mda1 for MD
machinery and Dnm2 for PD machinery) are selectively transcribed before organelle
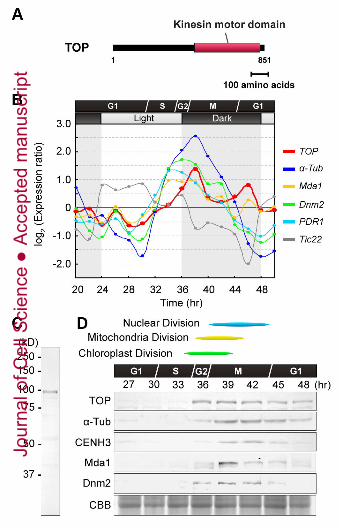
division; therefore, we examined the level of TOP transcription during the cell cycle using
C. merolae microarray data (Fujiwara et al., 2009). The level of transcription of TOP
increased during S to M phase and this expression profile coincided with that of other
known MD and PD machinery-associated proteins (Fig. 2B). We then generated anti-TOP
antibodies via bacterially expressed TOP protein. Anti-TOP antibodies detected the
predicted molecular mass of TOP (~96 kD) (Fig. 2C). Also, immunoblot analyses of total
protein from synchronized cultures showed that TOP was detected during chloroplast,
mitochondrial and nuclear divisions (Fig. 2D). Next, we examined the intracellular
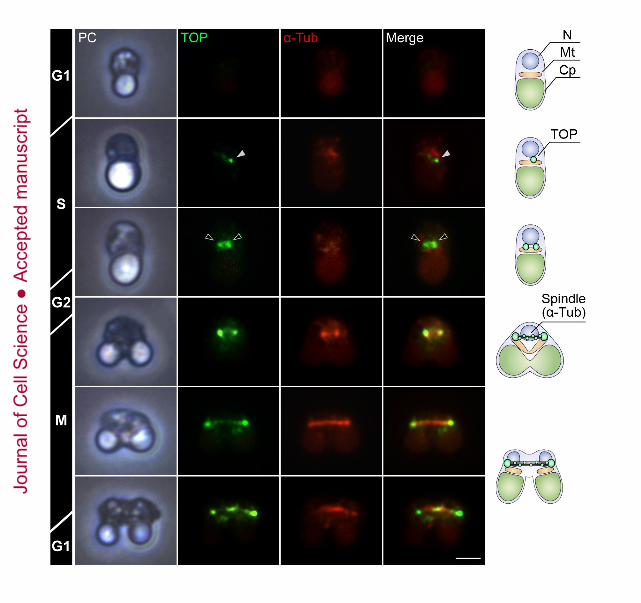
localization of TOP during the cell cycle by immunofluorescence microscopy using the
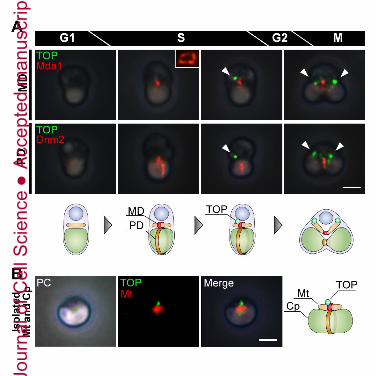
anti-TOP antibody (Fig. 3). Although TOP was not detected during the G1 phase, it was
observed in S phase, indicating that TOP was located near the mitochondrial division site
(Fig. 3, white arrowhead). Then, in the S-G2 phase, fluorescence signal of TOP-assembled
small spindle pole increased in size and split into two spindle poles (Fig. 3, black
arrowheads). During M phase, TOP was localized on microtubules in addition to at the two
spindle poles. Therefore, we conclude that TOP is involved in the spindle poles and the
spindle microtubules for nuclear division in M phase.
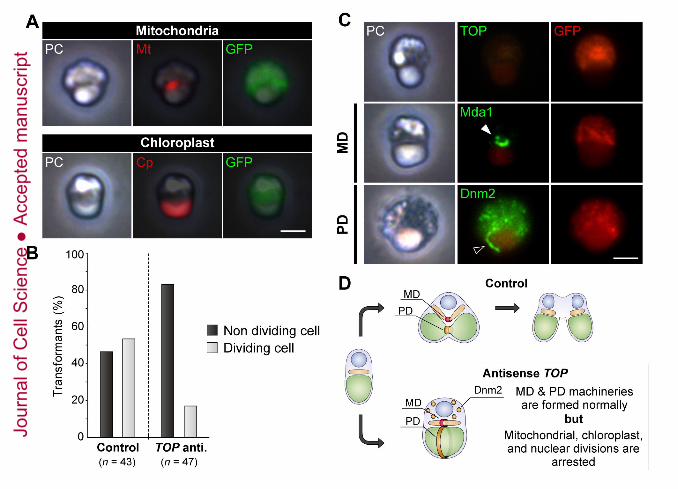
Since TOP was identified in the fraction of the isolated MD/PD machinery complex
by proteomic analysis (Fig. 1B) and appeared near the mitochondrial division site (Fig. 3,
white arrowhead), we next inspected whether TOP was involved in mitochondrial and/or
chloroplast division in addition to nuclear division. For the purpose, we examined the
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

9
effects of TOP down-regulation using antisense suppression (Fig. 4). 24 hours after
transformation, antisense-TOP induced the defects in mitochondrial and chloroplast
divisions (p < 0.001; Fisher’s exact test) (Fig. 4A, B). In these cells, both MD and PD
machineries were found at the mitochondrial and chloroplast division sites, respectively,
although constriction had not commenced in either (Fig. 4C, D). On the other hand,
inhibition of microtubule organization by oryzalin treatment arrested nuclear division of the
cells, but both mitochondrial and chloroplast divisions are performed normally (Fig. S4)
(Nishida et al., 2005). In addition, immunofluorescence microscopy showing that a
centromere marker protein, CENH3, appeared as multiple discrete speckles in nuclear, TOP
down-regulated cells was arrested in S/G2 phase (Fig. S5). Therefore, the obstruction of
mitochondrial and chloroplast divisions by TOP down-regulation are not caused by spindle
checkpoint. However, we cannot completely exclude the possibility that the S/G2 arrest
was caused by activation of other checkpoints.
TOP binds directly to the MD machinery complex
As TOP was required for the initiation of three-organelle divisions and seemed to be
located near on the mitochondrial division site, we examined whether the timing and
manner of localization of TOP related to the MD/PD machinery complex by using
immunofluorescence microscopy. Both MD and PD machineries were formed prior to the
expression of TOP (Fig. 5A). Immunofluorescence signal of TOP seems to be localized
only on the MD machinery in vivo (Fig. 5A) and could be recognized on mitochondrial
division site of the isolated dividing mitochondria and chloroplast (Fig. 5B). To reveal
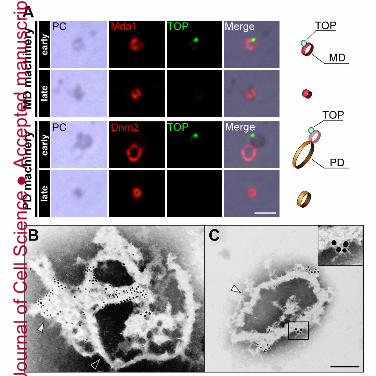
whether TOP binds directly to the MD machinery complex, we isolated the MD/PD
machinery complexes from cells in S or M phase and examined the localization of TOP by
immunofluorescence microscopy (Fig. 6A). We also measured the circumference of
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

10
isolated PD machineries to detect the stage of division because the circumference of the PD
machinery reduces as chloroplast division progresses (Miyagishima et al., 2001). The
average circumference of the PD machineries containing TOP was 2.6 μm (n=25) (Fig. S6),
therefore, we concluded that TOP bound to the MD/PD machinery complexes which were
derived from cells in the early phase of mitochondrial and chloroplast divisions. On the
other hand, TOP did not associate with small MD/PD machinery complexes derived from
cells in late phase of mitochondrial and chloroplast division (Fig. 6A). These results were
also confirmed by immuno-electron microscopy (immuno-EM) (Fig. 6B, C). Immunogold
particles indicating TOP were localized as a spot-like structure on the MD machinery (Fig
6C, inset). These results indicate that the direct interaction between TOP and larger MD
machineries plays an important role on the initiation of constriction of MD and PD
machineries.
TOP is required for the phosphorylation of proteins in both MD and PD machineries
Reports thus far imply that protein phosphorylation is involved in at least the
mitochondrial division. WD40 protein Mda1 with the MD ring assembles the outer ring of
MD machinery, and then Mda1 is phosphorylated. In the contraction phase of the
mitochondrial division, phosphorylated Mda1-oligomers on the MD machinery are
disassembled by GTP hydrolysis of Dnm1 (Nishida et al., 2007). And finally, the
mitochondrial dynamin ring mediated by Dnm1 fission the bridge of dividing
mitochondrion (Nishida et al., 2003). Since phosphorylation of Mda1 is induced with the
progress of the mitochondrial division phases, protein phosphorylation is likely to mediate
changes in the organization of the MD machinery for constriction. Thus, it is thought that
phosphorylation is one of the most important modifications for the MD and PD machineries.
Based on these results and hypothesis, we examined whether proteins in the isolated
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

11
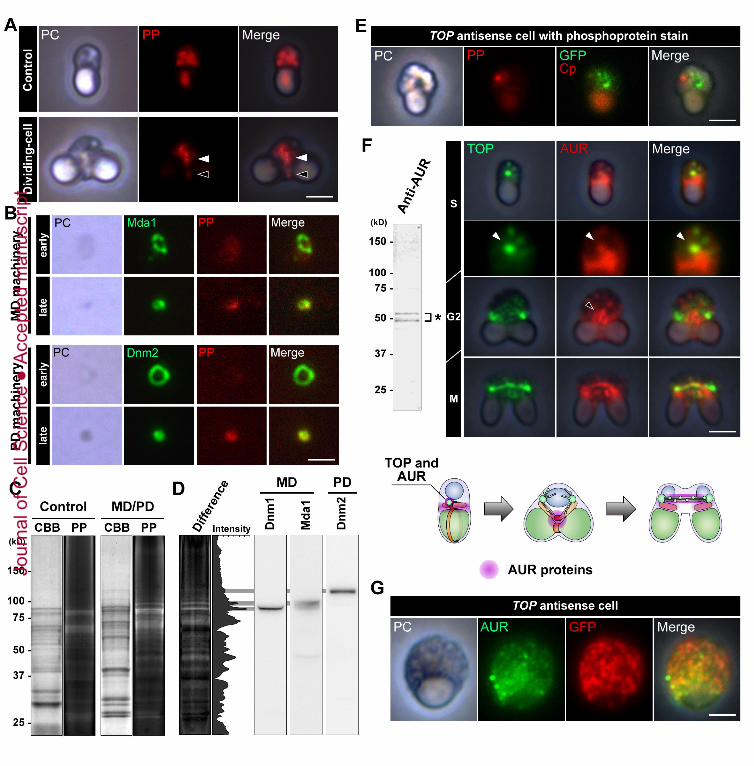
MD/PD machinery complexes were phosphorylated in the early and late phases of
mitochondrial and chloroplast division using cytochemical and MALDI-TOF-MS methods.
Large MD/PD machinery complexes which were derived from the cells in early phase of
division had few fluorescence signals derived from phosphorylation, but small MD and PD
machineries which were derived from the cells in late phase of division had strong
fluorescence signals (Fig, 7A, B). Moreover, differences in fluorescence signals between
the control fraction and the isolated MD/PD machinery complex fraction after staining for
phosphorylated proteins showed that many proteins, including Dnm1, Mda1 (for
mitochondrial division) and Dnm2 (for chloroplast division) in the isolated MD/PD
machinery complex fraction, were phosphorylated (Fig, 7C, D). By the MALDI-TOF-MS
analyses, phosphorylated peptides were estimated in matched peptide fragments for Dnm1,
Mda1, and Dnm2, respectively (Tables S2-S4). Therefore, both MD and PD machineries
were highly phosphorylated according to the progress of mitochondrial and chloroplast
division. In addition, we showed that antisense-TOP cells contained the non-constricted
MD and PD machineries at each division sites (Fig 4C), and besides, we revealed that they
were not phosphorylated (Fig 7E). Lastly, we examined immunofluorescence microscopy
to confirm the responsible kinases of the MD/PD machinery complex. Similar to C-
terminal KIFs, it is known that Aurora kinase is also involved in spindle pole maturation
because Aurora kinases were originally identified as being required for accurate spindle
pole structure (Glover, et al., 1995; Andrews et al., 2003). In addition, very recent study
showed that Aurora kinase in C. merolae (named AUR) related with not only mitotic
spindle formation but also mitochondrial division in C. merolae (Kato et al., 2011). The
level of transcription of AUR increased in S and M phase and this expression profile
coincided with that of TOP (Fig. S7). Therefore, we investigated whether AUR is related
with TOP during mitochondrial and chloroplast divisions. Immunofluorescence microscopy
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

12
found out that AUR was co-localized with TOP and localized around the mitochondrion
before the constriction phase of mitochondrial and chloroplast divisions (Fig. 7F, white
arrowhead). And then, AUR were accumulated into the mitochondrial division site (Fig. 7F,
black arrowhead). These results implied that AUR mediate phosphorylation of the MD and
PD machineries likewise spindle pole maturation. Next, we investigated that localization of
AUR in the TOP down-regulated cells to examine whether AUR is involved in the
maturation of the MD and PD machineries. As a result, AUR proteins were not localized
around mitochondrial division site but scattered in cytosolic region in the TOP down-
regulated cells (Fig. 7G). In addition, the proteomic analysis of isolated MD/PD machinery
complexes suggested that some of the phosphorylated peptides of Dnm1, Dnm2 and Mda1
were processed by Aurora kinase (Table S2-S4). In conclusion, TOP is a regulator of
MD/PD machinery complexes for mitochondrial and chloroplast divisions probably by
transferring of Aurora kinase to the machinery.
Discussion
Thus far, it has been revealed that mitochondrial and chloroplast division is performed by
the MD and PD machineries which are large protein complexes. Although the molecular
mechanisms how the MD and PD machineries divide mitochondria and chloroplast still
remain partially unknown, recent studies showed that FtsZ and dynamin proteins can
generate a force for contraction of membrane by GTP hydrolysis (Osawa et al., 2008;
Mears et al., 2011). Especially, a series of analyses of isolated single-PD and single-MD
machineries showed that Dnm2 and Dnm1 are required to generate constriction force by
sliding-movement of the PD or MD ring filaments, respectively (Yoshida et al., 2006,
2009). In addition, it was also revealed that phosphorylation of Mda1 may be required to
change the conformation of Dnm1 in the MD machinery to constrict mitochondrial division
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

13
site (Nishida et al., 2007). Thus, protein modification of the components of the MD and PD
machineries were strictly important to accomplish mitochondrial and chloroplast division.
However, such regulation factor for protein modification of the MD and PD machineries
has not been identified so far. In this study, we identified a kinesin-like protein TOP by the
proteomic analyses of isolated MD/PD machinery complexes before constriction (Fig. 1B,
Fig. S2). Since immunofluorescence microscopy showed that TOP was localized on the
MD/PD machinery complex at just before constriction of mitochondrial and chloroplast
division (Fig. 5, 6), it was hypothesized that TOP is required for activation of the MD and
PD machineries to perform constriction of each division sites. Finally, a series of analyses
of the antisense suppression of TOP (Fig. 4) and staining phosphoproteins in the MD and
PD machineries (Fig. 7) showed that TOP was required to induce contraction of the MD
and PD machineries by phosphorylation. TOP is directly bound to the MD machinery in
late S phase, then, TOP may induce activation of the MD/PD machinery complex through
protein phosphorylation by transferring of mitotic-kinase AUR from cytosol to the MD/PD
machinery complex (Fig. S8). Combined with the results of down-regulation of TOP (Fig 4)
and localization of TOP on the MD/PD machinery complex (Fig. 5), it is thought that
mitotic-kinases need to interact with a small part of the MD/PD machinery complex which
are slightly exposed in cytosol to activate the MD/PD machinery complex. Actually,
immunofluorescence signals of AUR was accumulated in this part of the MD/PD
machinery complex according to the progresses of mitochondrial and chloroplast division
(Fig. 7F). To interact mitotic-kinases with the MD/PD machinery complex, TOP may be
required as a mediator (Fig. S8). Mitotic-kinases on TOP can easily move to the periphery
of the MD machinery and can also move to the periphery of the PD machinery along the
MD machinery (Fig. S8). Although we showed that Aurora kinase may be responsible for
the MD and PD machineries, other kinases would be needed for the regulation of
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

14
machineries. Mammalian CDK1, an orthologue of plant CDKA, regulates phosphorylation
of mitochondrial dynamin, Drp1, to enhance mitochondrial division during mitosis
(Taguchi et al., 2007). CDKA and CDKB are cell cycle regulators in C. merolae
(Kobayashi et al., 2011) and mRNA accumulation of CDKB is specifically detected in
mitochondrial and chloroplast division phase of C. merolae (Fig. S9). Thus, CDKA and
CDKB in C. merolae are candidates to phosphorylate MD/PD machinery complexes in
addition to Aurora kinase.
Thus far, it was revealed that kinesin-superfamily members were important molecular
motors that directionally transport various cargos, including single/double-bounded
membranous organelles and large protein complexes (Hirokawa et al., 2009). Also, it was
well known that kinesin proteins which are expressed in M phase are involved in the
organization and function of the mitotic spindle (Walczak and Heald, 2008). These mitotic
kinesins typically act in chromosome alignment and segregation. However, the involvement
of a kinesin-like protein in mitochondrial and/or chloroplast division has never been
reported until now. Thus, this report is the first evidence that a member of the kinesin-
superfamily proteins is involved in mitochondrial and chloroplast division. TOP induced
protein phosphorylation of MD and PD machineries to accomplish mitochondrial and
chloroplast divisions prior to nuclear division. And finally, nuclear division is performed by
the TOP-mediated spindle structure (Fig. 3 and Fig. S8). Thereby, mitochondria- and
chloroplast-division cycles are combined with the cell-division cycle.
Recently, it is revealed that the nuclear division which is performed by TOP-mediated
spindle poles (Fig. 3) is accompanied by the inheritance of the endoplasmic reticulum (ER)
and Golgi apparatus in association with the mitotic spindle (summarized in Fig. S10)
(Imoto et al., 2011; Yagisawa et al., 2012a, 2012b). Although the interaction of plant C-
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

15
terminal KIFs with organelles is still unclear (Cai and Cresti, 2012), it was found that
animal C-terminal KIFs related with intracellular transportations of these single-bounded
membranous organelles. KIFC3 transports Golgi apparatus and KIFC2 transports early
endosomes as cargo, respectively (Saito et al., 1997; Hirokawa, 1998; Xu et al., 2002;
Bananis et al., 2004; Hirokawa et al., 2009). Moreover, we showed that the initiation of
both mitochondrial and chloroplast divisions are regulated by TOP, and recent studies
revealed that the divided mitochondria are required as the carrier of microbodies and
lysosomes which are connected with divided mitochondria (Fujiwara et al., 2010; Imoto et
al., 2010, 2011). Thus, TOP is related with all of single- and double-membrane bounded
organelle proliferations (Fig. S10). Concurrently, the involvement of TOP in mitochondrial
and chloroplast division, given the presence of TOP homologs in many members of
eukaryotes (Fig. S3), may indicate the original function of C-terminal kinesin-like proteins
which control all of double- and single-membrane bounded organelle proliferations.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

16
Materials and methods
Cell culture and isolation of MD/PD machinery complexes
The 10D strain of Cyanidioschyzon merolae was used (Matsuzaki et al., 2004), and
was cultured in flasks, with shaking, under continuous light (40 W/m2) at 42°C. For
synchronisation, the cell cultures were subcultured to <1 × 107 cells/mL in a flat-bottomed
flask and subjected to a 12-h light/12-h dark cycle at 42°C using an automatic light/dark
cycle CM incubator (Fujimoto Rika, Tokyo, Japan) (K. Suzuki et al., 1994). Synchronised
cells were harvested at the start of the second dark period (S phase) or after 2 hours of the
second dark period (M phase). Dividing chloroplasts and mitochondria and intact PD and
MD machineries were isolated as described previously (Yoshida et al., 2009).
MALDI-TOF-MS analysis and MASCOT search
Samples were analysed by a peptide mass fingerprinting (PMF) search using a
MALDI TOF AXIMA TOF2 mass spectrometer (Shimadzu, Kyoto, Japan) in reflectron
mode. Database searches were performed using the software program MASCOT v2.2.01
(Matrix Science, MA, USA) running on the local server against the C. merolae genome
database (including 5014 sequences) based on the FASTA file distributed by the
Cyanidioschyzon merolae Genome Project (http://merolae.biol.s.u-tokyo.ac.jp/). The
permissible value of missed cleavages was set at one. MS tolerance values were set at 0.2–
0.4 Da. Identified proteins (a MASCOT score of more than 50) are listed in Supplementary
Fig. S2 and in Supplementary Table S1.
Phylogenetic analysis
Additional amino acid sequences of C-terminal kinesin-superfamily members were
obtained from GenBank (http://www.ncbi.nlm.nih.gov/Genbank/) and were automatically
aligned using CLUSTAL X version 2.0.9 (http://www.clustal.org/download/current/)
(Larkin et al., 2007). For phylogenetic analyses, ambiguously aligned regions were
manually arranged or deleted using BioEdit Sequence Alignment Editor version 4.8.10
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

17
(http://www.mbio.ncsu.edu/BioEdit/bioedit.html), resulting in 363 amino acids (including
inserted gaps) being used. Phylogenetic tree construction and bootstrap analyses were
performed using PHYLIP version 3.66
(http://evolution.genetics.washington.edu/phylip.html), PROTDIST and NEIGHBOR for
neighbor-joining (NJ), PROTPARS for maximum parsimony (MP), and PROML for
maximum likelihood (ML). The JTT + Γ model (among-site rate variation model with four
rate categories) was selected as the probability model for NL and ML analyses. Multiple
datasets for bootstrap analyses (1000 replicates for NJ and MP, 100 replicates for ML) were
calculated using CONSENSE. No outgroup was used to root the tree.
Antibodies
To generate an anti-TOP antiserum in guinea pig, amino acids 1 to 431 of the
predicted 431-aa sequence of the CMR497C protein was amplified by PCR using the
following primers: 5’-cgggatccatgattcgagacagggttccag-3’ and 5’-
cccaagctttgcgcgaagttccatgatc-3’. The resultant DNA fragment was cloned into pQE80L
(QIAGEN, Hilden, Germany) following restriction digestion at BamHI and HindIII sites.
Protein expression and purification was performed as previously described (Nishida et al.,
2005). To produce a protein expression vector for AUR, the full length of a coding
sequence of AUR was amplified by PCR with following primer: 5’-
atggtaccatgcaggcgacaccaggcct-3’, 5’-atagaagcttctattgttccgcagcgtgca-3’. The fragment was
cloned into the KpnI/HindIII site of pCold-GST (Hayashi and Kojima, 2008). The vector
was transformed into OverExpressTM C43 (DE3) (Lucigen, WI, USA). The recombinant
protein of GST-AUR was purified with GSTrap HP (GE Healthcare, Buckinghamshire,
UK). The protein was immunized to a rabbit and the antiserum was subjected to affinity
purification (Protein Purify Ltd, Gunma, Japan)
Immunofluorescence microscopy, immunoblotting analysis and staining of
phosphorylated protein
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

18
The anti-TOP guinea pig antibody was used at a dilution of 1:1000 for
immunoblotting or at 1:100 for immunofluorescence. Antibodies against Dynamin 1
(Dnm1), Dynamin 2 (Dnm2), Mda1, α-tubulin, CENH3, and mitochondrial porin were used
as previously described (Nishida et al., 2003; Miyagishima et al., 2003; Nishida et al.,
2005; Maruyama et al., 2007; Nishida et al., 2007; Takayuki Fujiwara et al., 2009).
Secondary antibodies used for immunofluorescence were, Alexa Fluor 488 or Alexa Fluor
555 goat anti-guinea pig, anti-mouse, or anti-rabbit IgG, highly cross-adsorbed (Molecular
Probes, Eugene, OR). Images were captured using a BX51 fluorescence microscope
(Olympus, Tokyo, Japan), additionally equipped with an XF37 narrow bandpass filter
(Omega, Tokyo, Japan) and a C7780-10 three charge-coupled device (CCD) camera system
(Hamamatsu Photonics, Shizuoka, Japan). Primary antibody reactions were performed for 1
h at 4°C. Secondary antibody reactions were performed for 1 h at 4°C. For the staining of
phosphorylated protein, cells or isolated MD/PD machineries were stained with Pro-Q
Diamond phosphoprotein gel stain (Molecular Probes, Eugene, OR) for 15 min at 4°C.
Negative staining and immunoelectron microscopy
For immunoelectron microscopy (immuno-EM), primary reactions were performed
for 1 h at 4°C with guinea pig anti-TOP or rabbit anti-Mda1 diluted 1:100 in Can Get
Signal Immunostain Solution B (TOYOBO, Osaka, Japan), and labeled with 15-nm for
TOP or 10-nm gold particle-conjugated secondary antibody (British BioCell International,
Cardiff, UK) for Mda1 at a dilution of 1/20. After MD and PD machineries with outer
membranes were incubated in organelle membrane dissolution buffer (OMD buffer; PBS
containing 100 mM n-octyl-β-D-glucopyranoside [OG], 6 mM sodium lauryl sulphate, 20
mM urea) for 2 min on ice, the lysate was negatively stained with 0.5% phosphotungstic
acid (pH 7.0). The samples were then examined with an electron microscope (model JEM-
1230; JEOL, Tokyo, Japan).
Plasmid Construction of pCPG and pCPG-TOP-AS
PCR reactions were performed with KOD FX (TOYOBO, Osaka, Japan) using the
oligonucleotide primers, 5’-cttaaccgtactgatcgtact-3’ and 5’-tagtctaaactgagaacagcc-3’ and
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

19
pBSHAb-T3’ as a template (Ohnuma et al., 2008), and 5’-
tctcagtttagactactgcactcaaagtgagtgtccg-3’ and 5’-atcagtacggttaagtcatgtttgacagcttatcatc-3’
and pI050P-GFP as a template (Ohnuma et al., 2009). The resultant DNA fragments were
combined and cloned to make pCPG (catalase promoter with sGFP) using the In-FusionTM
advantage PCR cloning kit (Clontech, CA, USA). The 5’-flanking region (1,500 bp) and
the open reading frame (ORF) of the TOP gene (2,553 bp) was amplified using the
oligonucleotide primers, 5’-ggcggccgctctagagaatggaaatcgcgcgcttctcc-3’ and 5’-
tgggtaattaattaatggctcctggaaagagactcgttg-3’. The pCPG vector fragment was amplified using
the oligonucleotide primers, 5’-ttaattaattacccatacgatgttcctgactatgcggg-3’ and 5’-
tctagagcggccgccaccg-3’ and pCPG as a template. The resultant DNA fragments were
combined and cloned to make pCPG-TOP-S using the In-FusionTM advantage PCR cloning
kit (Clontech, CA, USA). The antisense strand of the TOP ORF (2,553 bp) was amplified
using the oligonucleotide primers, 5’-aagtgcgcctgcgcatggctcctggaaagagactcgttg-3’ and 5’-
tgggtaattaattaaatgattcgagacagg-3’. Using the oligonucleotide primers, 5’-
ttaattaattacccatacgatgttcctgac-3’ and 5’-tgcgcaggcgcacttg-3’ and pCPG-TOP-S as a
template, DNA fragments were PCR amplified, then combined and cloned to make pCPG-
TOP-AS using the In-FusionTM advantage PCR cloning kit (Clontech, CA, USA). pCPG-
TOP-AS has the antisense strand of the TOP ORF instead of the sense strand of the TOP
ORF. The resultant plasmids, pCPG as a control and pCPG-TOP-AS for antisense
suppression of TOP, have a catalase promoter-sGFP fused sequence; therefore,
transformed cells could be detected by green fluorescence under microscopic observation.
Transformation of C. merolae cells was performed as described previously (Ohnuma et al.,
2009).
Acknowledgements
We thank T. Shimada for technical help with MALDI-TOF analysis. This work was
supported by a Grant-in-Aid for Scientific Research on Priority Areas (A) (22247007 to
T.K.); a Grant-in-Aid for Challenging Exploratory Research (22657061 to T. K.). This
research was supported by a Grant-in-Aid for X-ray Free Electron Laser Priority Strategy
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

20
Program (MEXT) to S.M. and MEXT/JSPS KAKENHI to S.M. (23370029, 23120518).
Y.Y. is supported by a Human Frontier Science Program Long Term Fellowship.
Author Contributions
Y.Y., T. F., Y. I., M. O., S. H., O. M., H. K., S. K., S. M. and T. K. designed the research,
and Y.Y., T. F., Y. I., M. O., S. K. and S. M. carried out the research. Y.Y., M.Y. and Y. I.
performed the proteomics analyses. Y.Y. and T.F analysed the C. merolae microarray data.
Y.Y., T. F., Y. I. and M. O. performed TOP down-regulation analysis. Y.Y. and T. K.
wrote the paper.
Conflict of interest
The authors declare that they have no conflict of interest.
References
Andrews, P. D., Knatko, E., Moore, W. J., Swedlow, J. R. (2003). Mitotic mechanics: the auroras come into view. Cur. Opin. Cell Biol. 15, 672–683.
Bananis, E., Nath, S., Gordon, K., Satir, P., Stockert, R. J., Murray, J. W., Wolkoff, A. W. (2004). Microtubule-Dependent Movement of Late Endocytic Vesicles In Vitro�: Requirements for Dynein and Kinesin. Mol. Biol. Cell, 15, 3688–3697.
Bleazard, W., McCaffery, J. M., King, E. J., Bale, S., Mozdy, A., Tieu, Q., Nunnari, J., Shaw, J. M. (1999) The dynamin-related GTPase Dnm1 regulates mitochondrial fission in yeast. Nat. Cell Biol. 1, 298-304.
Cai, G., Cresti, M. (2012). Are kinesins required for organelle trafficking in plant cells? Front. Plant Sci. 3, 1–10.
Felsenstein, J. (2002) PHYLIP: Phylogeny Inference Package, Ver 3.6. University of Washington, Seattle, WA.
Fujiwara, T., Misumi, O., Tashiro, K., Yoshida, Y., Nishida, K., Yagisawa, F., Imamura, S., Yoshida, M., Mori, T., Tanaka, K., Kuroiwa, T. (2009) Periodic gene expression patterns during the highly synchronized cell nucleus and organelle division cycles in the unicellular red alga Cyanidioschyzon merolae. DNA Res. 16, 59-72.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

21
Fujiwara, T., Kuroiwa, H., Yagisawa, F., Ohnuma, M., Yoshida, Y., Yoshida, M., Nishida, K., Misumi, O., Watanabe, S., Tanaka, K., Kuroiwa, T. (2010) The coiled-coil protein VIG1 is essential for tethering vacuoles to mitochondria during vacuole inheritance of Cyanidioschyzon merolae. Plant Cell 22, 772-781.
Gao, H., Kadirjan-Kalbach, D., Froehlich, J. E., Osteryoung, K. W. (2003) ARC5, a cytosolic dynamin-like protein from plants, is part of the chloroplast division machinery. Proc. Natl. Acad. Sci. USA 100, 4328-4333.
Glover, D. M., Leibowitz, M. H., Mclean, D. A., & Parry, H. (1995). Mutations in aurora prevent centrosome separation leading to the formation of monopolar spindles, Cell 81, 95–105.
Hayashi, K., Kojima, C. (2008) pCold-GST vector: a novel cold-shock vector containing GST tag for soluble protein production. Protein Expr. Purif. 62, 120-127.
Hirokawa, N. (1998). Kinesin and Dynein Superfamily Proteins and the Mechanism of Organelle Transport. Science 279, 519–526.
Hirokawa, N., Noda, Y., Tanaka, Y., Niwa, S. (2009). Kinesin superfamily motor proteins and intracellular transport. Nature Rev. Mol. Cell Biol. 10, 682–696.
Imoto, Y., Fujiwara, T., Yoshida, Y., Kuroiwa, H., Maruyama, S., Kuroiwa, T. (2010) Division of cell nuclei, mitochondria, plastids, and microbodies mediated by mitotic spindle poles in the primitive red alga Cyanidioschyzon merolae. Protoplasma 241, 63-74.
Imoto, Y., Yoshida, Y., Yagisawa, F., Kuroiwa, H., Kuroiwa, T. (2011) The cell cycle, including the mitotic cycle and organelle division cycles, as revealed by cytological observations. J. Electron microsc. 60 Suppl 1, S117-S136.
Kato, S., Imoto, Y., Ohnuma, M., Matsunaga, T. M., Kuroiwa, H., Kawano, S., Tsuneyoshi, K., Matsunaga, S. (2011). Aurora Kinase of the red alga Cyanidioschyzon merolae is related to both mitochondrial division and mitotic spindle formation. Cytologia 76, 465–472.
Kobayashi, Y., Kanesaki, Y., Tanaka, A., Kuroiwa, H., Kuroiwa, T., Tanaka, K. (2009) Tetrapyrrole signal as a cell-cycle coordinator from organelle to nuclear DNA replication in plant cells. Proc. Natl. Acad. Sci. USA 106, 803-807.
Kobayashi, Y., Imamura, S., Hanaoka, M., Tanaka, K. (2011) A tetrapyrrole-regulated ubiquitin ligase controls algal nuclear DNA replication. Nat. cell biol. 13, 483-487.
Kuroiwa, T. (1998). The primitive red algae Cyanidium caldarium and Cyanidioschyzon merolae as model system for investigating the dividing apparatus of mitochondria and plastids. BioEssays 20, 344–354.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

22
Kuroiwa, T., Misumi, O., Nishida, K., Yagisawa, F., Yoshida, Y., Fujiwara, T., Kuroiwa, H. (2008) Vesicle, mitochondrial, and plastid division machineries with emphasis on dynamin and electron-dense rings. Int. Rev. Cell Mol. Biol. 271, 97-152.
Larkin, M. A., Blackshields, G., Brown, N. P., Chenna, R., McGettigan, P. A., McWilliam, H., Valentin, F., Wallace, I. M., Wilm, A., Lopez, R., Thompson, J. D., Gibson, T. J., Higgins, D. G. (2007) Clustal W and Clustal X version 2.0. Bioinformatics 23, 2947-2948.
Maruyama, S., Kuroiwa, H., Miyagishima, S., Tanaka, K., Kuroiwa, T. (2007) Centromere dynamics in the primitive red alga Cyanidioschyzon merolae. Plant J. 49, 1122-1129.
Matsuzaki, M., Misumi, O., Shin-I, T., Maruyama, S., Takahara, M., Miyagishima, S., Mori, T., Nishida, K., Yagisawa, F., Nishida, K., Yoshida, Y., Nishimura, Y., Nakao, S., Kobayashi, T., Momoyama, Y., Higashiyama, T., Minoda, A., Sano, M., Nomoto, H., Oishi, K., Hayashi, H., Ohta, F., Nishizaka, S., Haga, S., Miura, S., Morishita, T., Kabeya, Y., Terasawa, K., Suzuki, Y., Ishii, Y., Asakawa, S., Takano, H., Ohta, N., Kuroiwa, H., Tanaka, K., Shimizu, N., Sugano, S., Sato, N., Nozaki, H., Ogasawara, N., Kohara, Y., Kuroiwa, T. (2004) Genome sequence of the ultrasmall unicellular red alga Cyanidioschyzon merolae 10D. Nature 428, 653-637.
Mears, J. a, Lackner, L. L., Fang, S., Ingerman, E., Nunnari, J., Hinshaw, J. E. (2011). Conformational changes in Dnm1 support a contractile mechanism for mitochondrial fission. Nature Struct. Mol. Biol. 18, 20–26.
Miyagishima, S., Takahara, M., Mori, T., Kuroiwa, H., Higashiyama, T., Kuroiwa, T. (2001) Plastid division is driven by a complex mechanism that involves differential transition of the bacterial and eukaryotic division rings. Plant Cell 13, 2257-68.
Miyagishima, S., Nishida, K., Mori, T., Matsuzaki, M., Higashiyama, T., Kuroiwa, H., Kuroiwa, T. (2003) A plant-specific dynamin-related protein forms a ring at the chloroplast division site. Plant Cell 15, 655–665.
Nishida, K., Takahara, M., Miyagishima, S., Kuroiwa, H., Matsuzaki, M., Kuroiwa, T. (2003) Dynamic recruitment of dynamin for final mitochondrial severance in a primitive red alga. Proc. Natl. Acad. Sci. USA 100, 2146-2151.
Nishida, K., Yagisawa, F., Kuroiwa, H., Nagata, T., Kuroiwa, T. (2005) Cell cycle-regulated, microtubule-independent organelle division in Cyanidioschyzon merolae. Mol. Biol. Cell 16, 2493-2502.
Nishida, K., Yagisawa, F., Kuroiwa, H., Yoshida, Y., Kuroiwa, T. (2007) WD40 protein Mda1 is purified with Dnm1 and forms a dividing ring for mitochondria before Dnm1 in Cyanidioschyzon merolae. Proc. Natl. Acad. Sci. USA 104, 4736-4741.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

23
Nozaki, H., Takano, H., Misumi, O., Terasawa, K., Matsuzaki, M., Maruyama, S., Nishida, K., Yagisawa, F., Yoshida, Y., Fujiwara, T., Takio, S., Tamura, K., Chung, S. J., Nakamura, S., Kuroiwa, H., Tanaka, K., Sato, N., Kuroiwa, T. (2007) A 100%-complete sequence reveals unusually simple genomic features in the hot-spring red alga Cyanidioschyzon merolae. BMC Biol. 5, 28.
Ohnuma, M., Yokoyama, T., Inouye, T., Sekine, Y., Tanaka, K. (2008) Polyethylene glycol (PEG)-mediated transient gene expression in a red alga, Cyanidioschyzon merolae 10D. Plant Cell Physiol. 49, 117-120.
Ohnuma, M., Misumi, O., Fujiwara, T., Watanabe, S., Tanaka, K., Kuroiwa, T. (2009) Transient gene suppression in a red alga, Cyanidioschyzon merolae 10D. Protoplasma 236, 107-112.
Osawa, M., Anderson, D. E., Erickson, H. P. (2008). Reconstitution of contractile FtsZ rings in liposomes. Science 320, 792–794.
Osteryoung, K. W., Nunnari, J. (2003) The division of endosymbiotic organelles. Science 302, 1698-1704.
Saito, N., Okada, Y., Noda, Y., Kinoshita, Y., Kondo, S., Hirokawa, N. (1997). KIFC2 is a novel neuron-specific C-terminal type kinesin superfamily motor for dendritic transport of multivesicular body-like organelles. Neuron 18, 425–38.
Suzuki, K., Ehara, T., Osafune, T., Kuroiwa, H., Kawano, S., Kuroiwa, T. (1994) Behavior of mitochondria, chloroplasts and their nuclei during the mitotic cycle in the ultramicroalga Cyanidioschyzon merolae. Eur. J. Cell Biol. 63, 280-288.
Taguchi, N., Ishihara, N., Jofuku, A., Oka, T., Mihara, K. (2007) Mitotic phosphorylation of dynamin-related GTPase Drp1 participates in mitochondrial fission. J. Biol. Chem. 282, 11521-11529.
Walczak, C. E., Heald, R. (2008). Mechanisms of mitotic spindle assembly and function. Int. Rev. Cytol. 265, 111–158.
Walczak, C. E., Verma, S., Mitchison, T. J. (1997). XCTK2: A kinesin-related protein that promotes mitotic spindle assembly in Xenopus laevis egg extracts. J. Cell Biol. 136, 859–870.
Xu, Y., Takeda, S., Nakata, T., Noda, Y., Tanaka, Y., Hirokawa, N. (2002). Role of KIFC3 motor protein in Golgi positioning and integration. J. Cell Biol. 158, 293–303.
Yagisawa, F., Nishida, K., Yoshida, M., Ohnuma, M., Shimada, T., Fujiwara, T., Yoshida, Y., Misumi, O., Kuroiwa, H., Kuroiwa, T. (2009) Identification of novel proteins in isolated polyphosphate vacuoles in the primitive red alga Cyanidioschyzon merolae. Plant J. 60, 882-893.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

24
Yagisawa, F., Fujiwara, T., Kuroiwa, H., Nishida, K., Imoto, Y., Kuroiwa, T. (2012a) Mitotic inheritance of endoplasmic reticulum in the primitive red alga Cyanidioschyzon merolae. Protoplasma 249, 1129-1135.
Yagisawa, F., Fujiwara, T., Ohnuma, M., Nishida, K., Imoto, Y., Yoshida, Y., Kuroiwa, H., Kuroiwa, T. (2012b) Golgi inheritance in the primitive red alga, Cyanidioschyzon merolae. Protoplasma in press.
Yoshida, Y., Kuroiwa, H., Misumi, O., Nishida, K., Yagisawa, F., Fujiwara, T., Nanamiya, H., Kawamura, F., Kuroiwa, T. (2006) Isolated chloroplast division machinery can actively constrict after stretching. Science 313, 1435-1438.
Yoshida, Y., Kuroiwa, H., Hirooka, S., Fujiwara, T., Ohnuma, M., Yoshida, M., Misumi, O., Kawano, S., Kuroiwa, T. (2009) The bacterial ZapA-like protein ZED is required for mitochondrial division. Curr. Biol. 19, 1491-1497.
Yoshida, Y., Kuroiwa, H., Misumi, O., Yoshida, M., Ohnuma, M., Fujiwara, T., Yagisawa, F., Hirooka, S., Imoto, Y., Matsushita, K., Kawano, S., Kuroiwa, T. (2010) Chloroplasts divide by contraction of a bundle of nanofilaments consisting of polyglucan. Science 329, 949-953.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

25
Figure legends
Fig. 1. A proteomic analysis of MD/PD machinery complexes before constriction. (A)
Schematic depiction of mitochondrial and chloroplast division processes in C. merolae cells.
Division of mitochondria and chloroplasts is performed by the MD and PD machineries,
respectively, following the three steps; formation, constriction, and pinching-off. (B)
Proteomic analysis of isolated MD/PD machinery complexes before constriction. Identified
proteins were shown in Supplemental Fig. S2 and Supplemental Table S1.
Fig. 2. mRNA and protein expression profiles of TOP. (A) Molecular structures of kinesin-
like protein TOP. Red bars indicate the kinesin motor domain. (B) mRNA levels of TOP
(red), α-Tubulin (α-Tub, blue), Mda1 (yellow), Dnm2 (green) and PDR1 (light blue) at
different points of the cell cycle, determined using microarray data. The light-dependent
gene Tic22 (grey) is also shown as a control. (C) Total protein from synchronized M phase
cells was blotted with anti-TOP antibody. (D) Protein levels of TOP, α-Tub, CENH3, Mda1
and Dnm2 at different points of the cell cycle.
Fig. 3. Immunofluorescence images and models of TOP (green) and α-Tubulin (α-Tub, red)
in cells. White arrowhead indicates TOP protein near the mitochondrial division site and
black arrowheads indicate TOP-assembled two spindle poles. Scale bar, 1 μm.
Fig. 4. Down-regulation of TOP. (A) Phase contrast (PC) and fluorescence images of
antisense-TOP cells. Transiently transformed cells express sGFP (green) in the cytosolic
space. Mitochondria (Mt, red) were immunolabeled using an anti-mitochondrial porin
antibody and chloroplasts (Cp, red) emitted red autofluorescence. (B) In cells which TOP
expression was down-regulated by antisense suppression, the frequency of dividing cells
was reduced (p< 0.001; Fisher’s exact test). Data are the total number of transformants
examined (n) in more than five replicates. (C) PC and immunofluorescence images of TOP,
mitochondria (Mt), MD machinery (Mda1, white arrowhead) and PD machinery (Dnm2,
black arrowhead) of cells treated with antisense-TOP. (D) Schematic model of the
antisense-TOP cells. Scale bars, 1 μm.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

26
Fig. 5. TOP localizes on near the mitochondrial division site. (A) Sequential
immunofluorescence images and explanations of the formation of MD machinery (Mda1,
red), PD machinery (Dnm2, red), and TOP (green, arrowhead). (B) Immunofluorescence
images and a model of TOP on an isolated dividing mitochondrion (red) and chloroplast.
Scale bars, 1 μm.
Fig. 6. Direct interaction of TOP on MD/PD machinery complex. (A) Immunofluorescence
images and explanations of direct interaction of TOP on isolated MD and PD machineries
from early phases of division. (B, C) Immuno-EM images of large isolated MD/PD
machinery complex (B) and large isolated MD machinery (C). Small immunogold particles
indicating Mda1 and large immunogold particles indicating TOP. Arrowheads indicate MD
machinery (white arrowhead) or PD machinery (black arrowhead), respectively. Scale bars,
1 μm in (A) and 200 nm in (B, C).
Fig. 7. Phosphorylation of MD and PD machineries during mitochondrial and chloroplast
division. (A) Phase contrast (PC) and fluorescence images of cells stained for
phosphorylated protein (PP, red). The cells were stained by Pro-Q Diamond dye.
Arrowheads indicate MD machinery (white arrowhead) or PD machinery (black
arrowhead). (B) Immunofluorescence images of isolated MD machineries (Mda1, green)
and PD machineries (Dnm2, green) stained for phosphorylated protein. MD and PD
machineries were isolated from cells in early M phase (upper set) or late M phase (bottom
set). (C) Coomassie brilliant blue (CBB)-stained and phosphoprotein-stained gels in the
isolated MD/PD machinery complex (right) and control fraction (left). Each gel was stained
by Pro-Q Diamond dye before CBB-staining. (D) A gel image indicates differences in
fluorescence signals between the gel image of the isolated MD/PD machinery complex
fraction and the gel image of the control fraction in (C). Three phosphorylated proteins in
the gel are identified as Dnm1, Mda1 and Dnm2 by immunoblotting and MALDI-TOF-MS
analyses. Matched peptide sequence and estimated phosphopeptides were shown in
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

27
Supplemental Tables S2-S4. (E) Immunofluorescence images of antisense-TOP cells with
staining for phosphorylated proteins. After staining phosphorylated proteins (PP, red), cells
were immunolabeled using anti-GFP antibody (GFP, green). (F) Immunoblotting and
immunofluorescence images of AUR and schematic model of localization of AUR proteins
(pink) during cell division. Anti-AUR antibodies detected two major polypeptides (asterisk).
A higher form of AUR is probably phosphorylated-AUR proteins. (G) Phase contrast and
immunofluorescence images of AUR (green) in TOP down-regulated cell. Transiently
transformed cells express AUR and sGFP (red) in the cytosolic space. Scale bars, 1 μm.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t





























