Embed Size (px)
Citation preview

XENOBIOTICA, 1983, VOL. 13, NO. 9, 513-520
The metabolism of 4-substituted-N-ethyl-N-methyl- anilines
11. Some factors influencing a-G and N-oxidation
J. W. GORROD and L. H. PATTERSON? Department of Pharmacy, Chelsea College (University of London), Manresa Road, London SW3 6LX, UK
Received 29 October 1982; accepted 1 September 1983
1 . A g.1.c. method for the quantification of N-demethylated and/or N-de-ethylated (a-C-oxidation) and N-oxidation products of several 4-substituted-N-ethyI-N- methylanilines is described.
2. Factors affecting the metabolism of these tertiary anilines in vitro have been studied and conditions which allow maximal metabolism established.
3. The a-C- and N-oxidase activity was detected principally in the liver, lung, kidney and bladder microsomal fraction.
4. A species difference in the extent of metabolism was evident, the order of activity not being the same for a-C- as that for N-oxidation.
5 . The pK, but not log P values determined for the tertiary anilines appear to influence the apparent K,,, for both a-C- and N-oxidation.
Introduction Numerous studies on the metabolism of tertiary aromatic amines have shown
that a-C-( N-dealkylation) and N-oxidation are major pathways of metabolism (McMahon 1966, Bickel 1969). Studies on u-C- and N-oxidation have often been carried out using N, N-dimethylaniline as substrate with assay methods of low specificity. The development of a specific g.1.c. assay which allowed the simul- taneous determination of N-demethylated and N-de-ethylated products as well as the N-oxide from N-ethyl-N-methylaniline allowed the metabolism and enzy- mology of the processes to be studied in more detail (Gorrod, Temple and Beckett 1975 a, b).
The N-oxidation of amines can be mediated by either cytochrome P-450 or a flavoprotein amine oxidase and it has been suggested (Gorrod 1973, 1974) that tertiary aromatic amines may be substrates for both enzymes, the quantitative importance of either enzyme being influenced by the pK, of the nitrogen under metabolic attack. It was the intention of this study, in part, to establish the quantitative relationship between the pK, and/or log P values and kinetic constants K, and V,,, for the N- and u-C-oxidation of some 4-substituted-N-ethyl-N- methylanilines. Such compounds have differing physicochemical properties without affecting sterically the nitrogen centre. The occurrence of u-C and N-oxidation products in oitro of these 4-substituted-N-ethyl-N-methylanilines has recently been reported (Cowan et al. 1982).
t Present address: School of Pharmacy, Leicester Polytechnic, Leicester LE1 9BH, UK.
Xen
obio
tica
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
orth
Car
olin
a on
05/
14/1
3Fo
r pe
rson
al u
se o
nly.

514 J.’W. Gorrod and L . H . Patterson
Materials and methods Materials
N-Ethyl-N-methylaniline and the 4-substituted derivatives used as substrates and metabolites were synthesized and checked for purity as previously described (Cowan et al. 1982). Glucose 6-phosphate, glucose 6-phosphate dehydrogenase and NADP were purchased from Boehringer Corporation (London) Ltd. All other chemicals were obtained from B D H Ltd (Poole, Dorset, UK).
T h e animals used throughout this investigation were as follows: male albino Wistar rats (30WOOg) fed on a normal laboratory diet (Diet 86; E. Dixon and Sons Ltd, Ware, UK); male albino Dunkin-Hartley guinea-pigs (400-6OOg) fed on diet R G P (E. Dixon); male New Zealand White rabbits (2.5-3.0kg) fed on diet S G I (E. Dixon); male Syrian hamsters (80-lOOg) and male L.A.C.A. stock mice ( 3 W O g ) fed on diet E F G M (E. Dixon). T h e animals were starved overnight before tissue preparation but were given drinking water ad libitum. Organ homogenates and various tissue fractions were prepared as described previously (Gorrod et al. 1975 a).
Incubation of substrates Substrates (5 pmol), in water (0.5 ml), in 25 ml stoppered Erlenmeyer flaskes with 0.1 M phosphate
buffer p H 7.4 (2 ml) containing N A D P (2pmol), glucose 6-phosphate (lOpmol), glucose 6-phosphate dehydrogenase (1 unit) and MgCI, (20pmol) were mixed and preincubated at 37°C for five minutes before the addition of hepatic microsomal preparations equivalent to 0.5 g of original liver (in 1 ml tris-KCI buffer, p H 7.4). Enzymic activity was terminated by the addition of 0.5 m l 5 M HC1. For most experiments a standard incubation time of 1Omin was used and the complete incubation system was as described above. In certain experiments the incubation time or the concentration of substrate or various co-factors or amount and source of enzyme preparation was varied.
Extraction of metabolites from incubation mixtures T h e acidified incubation mixture was diluted with 20ml water and a 4.0ml aliquot transferred to a
screw-capped tube containing an appropriate internal standard (usually a higher homologue of the 4- substituted-N-ethyl-N-methylaniline substrate) in water. T h e mixture was made alkaline with 0 2 m l 5 M NaOH and immediately extracted with freshly double-distilled diethyl ether (3 x 5 ml). T h e extracts were pooled, evaporated to about loop1 and stored on ice. T h e aqueous phase remaining was acidified with 0 1 ml 5 M HCI and 0.2 ml titanous trichloride added. T h e resulting mixture was shaken for 30min, the excess TiCI, decomposed by the addition of 0.3 ml 5 M NaOH, further internal standard added, and the alkaline solutions re-extracted with freshly double distilled ether (3 x 5 ml). These extracts were pooled, concentrated to about loop1 and stored on ice before g.1.c. analysis.
Gas-liquid chromatography A Perkin-Elmer F33 gas chromatograph equipped with a flame ionization detector and a 0-10 mV
Hitachi-Perkin-Elmer 56 chart recorder was used. For analysis of N-ethyl-N-methyl-4-aminobiphenyl and its metabolic products the following was used: a precoiled glass column (1 m, 0.5 cm int. diam.) packed with 3% OV-1 on 80-100 mesh AW-DMDCS-treated Chromosorb G silanized with 20pl hexamethyl disilazane. For analysis of all other tertiary amines the following was used: a precoiled glass column (1 m, 0.5cm int. diam.) packed with 8% Apiezon L plus 10% K O H on 80-100 mesh AW-DMDCS-treated Chromosorb G. All columns were conditioned at 225°C for 48 h before use. Gas pressures were N,, 140 kN/m2; H,, 119 kN/m2; and air, 175 kN/m2; detector and injection port temp. were set at 50°C above oven operating temp.
Linear calibration curves for the various authentic metabolites were obtained by adding to typical microsomal incubation mixtures known amounts of respective compounds and the appropriate internal standard. A range of concentrations of between 0.25 and 5 pmo1/3.5 ml for the 4-substituted tertiary anilinrs, 0.075 and 1.5 /1mol/3.5 ml for the 4-substituted N-ethylanilines and 0.025 and 0.5 pmo1/3.5 ml for the 4-substituted N-methylanilines and tertiary aniline-N-oxides was thus prepared. Internal standards (1Opg) were added in 0.5 ml solution.
Determination of log P and p K, values T h e determination of n-octanol/O.l M NaOH partition coefficients was carried out using glass tubes of
100ml capacity sealed at both ends and having a central stoppered sampling port. T h e 4-substituted-N- ethyl-N-methylanilines were dissolved in the n-octanol phase (1-3 ml) which had previously been saturated with the alkaline phase to provide concentrations of 2 4 m g l m l . This together with the alkaline phase (50 ml) was introduced into the tubes and allowed to rock through 90” (10 times per min) for 16 h a t 22 1°C. after which time the contents were centrifuged to break any emulsion formed. An aliquot of the aqueous phase was then suitably diluted and assayed by U . V . spectroscopy for ‘aniline’ content. Linear calibration had previously been established for each compound in 0.1 M NaOH presaturated with n-octanol at the wavelength of maximum absorbance. T h e partition coefficient (P) was calculated from the final concentrations in the octanol and aqueous phase. T h e pK, values of the anilines were determined by the spectrophotometric method of Albert and Serjeant (1971) where advantage is made of the difference in
Xen
obio
tica
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
orth
Car
olin
a on
05/
14/1
3Fo
r pe
rson
al u
se o
nly.

I-Substituted N-ethyl-N-methylaniline metabolism 515
U.V. spectra between the ionized and unionized forms. Standard solutions of the anilines were made in buffers of various pH values in the range f 0.6 of the expected pK, value. (Coarse checks of this value were made beforehand.) Likewise, solutions of the same strength were made up in 01 M HCl and 0 1 M NaOH. The absorbance at the wavelength of max. absorption of the unionized form (see table 5) was measured using a Hilger-Watts ‘Uvispek’ H-700 manual spectrophotometer. The pK, was calculated from the following equation:
where d is the measured absorbance at the pH, d,,, is the absorbance of the molecular species and d, is the absorbance of the ionized species.
Results and discussion Organ and cellular distribution of a-C- and N-oxidases
The animals used in these experiments were either male adult rabbits or guinea- pigs. The substrate used was N-ethyl-N-methyl-4-chloroaniline. Metabolic a-C- and N-oxidation of N-ethyl-N-methyl-4-chloroaniline was shown in a variety of rabbit-tissue microsomes, notably liver, lung, kidney and bladder (see table 1). In all tissues, N-demethylation occurred to a greater extent than N-deethylation and the liver was the most active organ. N-Oxidation by the lung was found to be three-fold greater than that found in the liver. Using various cell fractions from guinea-pig- liver homogenate, the a-C- and N-oxidation of N-ethyl-N-methyl-4-chloroaniline in this species was found to reside predominantly in the microsomal fraction (see table 2). Therefore, washed microsomal preparations fortified with a NADPH- regenerating system was used for all further work on the assumption that this would also be the most active cell fraction in the other species examined.
Effect of varying protein concentration The a-C- and N-oxidation of N-ethyl-N-methylaniline and 4-methyl- and 4-
chloro-derivatives using guinea-pig hepatic microsomes was linear with respect to protein concentration up to 2.4mg per ml incubate. This corresponded to a total protein content of 8.3 mg per incubation mixture which was equivalent to 0 5 g of
Table 1. The a-C- and N-oxidation of N-ethyl-N-methyl-4-chloroaniline by rabbit-tissue microsomal preparations.
Organ
a-C-Oxidation
N-Demethylation N-De-ethylation N-Oxidation
Liver Lung Bladder Kidney Brain Spleen Heart Intestines Testes
428 (1 00) 289 (68) 51 (12) 44 (10) 37 (9) 21 (5) 11 (3) 5 (1) 5 (1)
91 (100) t - 7 (8) 6 (7)
n.d. ~
3 (3) n.d.- 4 (4) 7 (8)
35 (100) 113 (323)
10 (29) 24 (69) n.d. - 3 (9)
n.d. - n.d. - n.d. -
t Interference peak in assay. n.d., Not detected. Enzyme activity is expressed in nmol product/0.5 g tissue wet weight per 10min incubation
Results are the means of two separate assays that could be reproduced within & 10%. (percentage of activity of liver in parentheses).
Xen
obio
tica
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
orth
Car
olin
a on
05/
14/1
3Fo
r pe
rson
al u
se o
nly.
516 J . W. Gorrod and L . H . Patterson
Table 2. The a-C- and N-oxidation of N-ethyl-N-methyl-4-chloroaniline by various cell fractions from guinea-pig liver homogenate.
a-C-Oxidation
Cell fraction N-Demethylation N-De-ethylation N-Oxidation ~~~~
Whole liver homogenate 810 172 155 9000g supernatant 996 227 385 140000g supernatant (soluble fraction) 63 10 19 Microsomal fraction 673 159 314 'Washed' microsomal fraction 629 144 321 Mitochondria1 fraction 32 n.d. n.d.
n.d., Not detected. Results are expressed in nmol product/O.Sg original liver wet wt. per 10min incubation and are the
NADPH-generating system as described in text was added to all subcellular fractions. means of two separate assays that could be reproduced within f 10%.
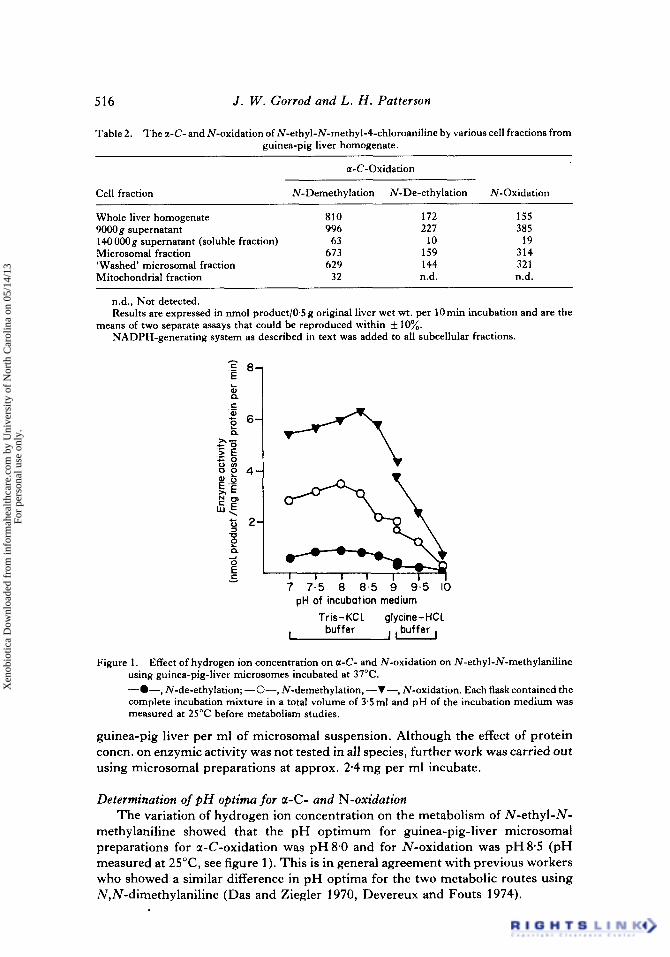
7 7.5 8 8.5 9 9-.5 I 0 pH of incubation medium
Tris- K C l glycine- HCl buffer Lbuffer, I
Figure 1. Effect of hydrogen ion concentration on a-C- and N-oxidation on N-ethyl-N-methylaniline using guinea-pig-liver microsomes incubated at 37°C. -0-, N-de-ethylation; -0-, N-demethylation, -V-, N-oxidation. Each flask contained the complete incubation mixture in a total volume of 3.5 ml and pH of the incubation medium was measured at 25°C before metabolism studies.
guinea-pig liver per ml of microsomal suspension. Although the effect of protein concn. on enzymic activity was not tested in all species, further work was carried out using microsomal preparations at approx. 2.4 mg per ml incubate.
Determination of p H optima for a-C- and N-oxidation The variation of hydrogen ion concentration on the metabolism of N-ethyl-N-
methylaniline showed that the pH optimum for guinea-pig-liver microsomal preparations for a-C-oxidation was pH8.0 and for N-oxidation was pH8.5 (pH measured at 25"C, see figure 1). This is in general agreement with previous workers who showed a similar difference in pH optima for the two metabolic routes using N,N-dimethylaniline (Das and Ziegler 1970, Devereux and Fouts 1974).
Xen
obio
tica
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
orth
Car
olin
a on
05/
14/1
3Fo
r pe
rson
al u
se o
nly.

4-Substituted N-ethyl-N-methylaniline metabolism 517
Effect of varying incubation time and substrate concentration The metabolism of the 4-substituted-N-ethyl-N-methylanilines was linear with
respect to time for at least 10 min using liver microsomes from all the species and is typified by N-ethyl-N-methyl-4-chloroaniline as shown in figure 2. In all species, N-demethylation was quantitatively a more significant pathway than N- deethylation, whilst the extent of N-oxidation varied considerably with species. For example, whilst N-oxidation and N-deethylation of N-ethyl-N-methyl-4- chloroaniline occurred to about the same extent in the rabbit, the extent of N- oxidation was similar to N-demethylation in the guinea-pig. The effect of substrate concentration on the metabolism of the 4-substituted-N-ethyl-N-methylanilines in vitro is shown in figure 3. In general, although the rate of a-C- and N-oxidation falls after 2pmollflask the amount of metabolites formed continues to rise over the concentrations used.
Apparent K, and V,,, values for a-C- and N-oxidation and species differences Both a-C- and N-oxidation of the 4-substituted-N-ethyl- N-methylanilines used
was first order up to at least 2 pmol substrate concentration after which they tended
l o0 l 90 i
30
20
10
I I I I I 1 0 10 20 30 40 50 60
20 - 4 10 -
I I 1 0 10 20 30 40 50 60
Time of incubation (min) Time of incubation ( m i d
(4 (6 ) Figure 2. Effect of incubation time on (a) a-C-oxidation, and (b ) N-oxidation, of N-ethyl-N-methyl-4-
chloroaniline by liver microsomes from various species. (a) Black symbols = N-demethylation; open symbols = N-de-ethylation (see N-oxidation legend for species). (b) t, rabbit; -0-, guinea-pig; -V-, rat; -m-, hamster; A , mouse. Each flask contained the complete incubation mixture in a total volume of 3.5 ml.
Xen
obio
tica
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
orth
Car
olin
a on
05/
14/1
3Fo
r pe
rson
al u
se o
nly.

518 J . W. Gorrod and L . H . Patterson
1 %
0 2 4 6 8
1
I I 0 2 4 6 0 2 4
Substrate concn.(pmol/ flask)
(4 ( b ) (4 Figure 3 . Effect of substrate concentration on a-C- and N-oxidation of (a ) N-ethyl-N-methyl-4-
toluidine, (b ) N-ethyl-N-methylaniline, (c) N-ethyl-N-methyl-4-chloroaniline by guinea-pig- liver microsomes.
-0-, N-demethylation; -0-, N-de-ethylation; -V-, N-oxidation. Each flask contained the complete incubation mixture in a total volume of 3.5ml.
to zero-order kinetics at 8pmol. When S/Vo (S=substrate concn. (mol) and Vo = initial rate of reaction (nmol product/mg protein per min)) was plotted against S, a linear relationship was obtained which, when extrapolated to the abscissa, allowed the determination of K , and V,,, (Hanes Plot). The K , and V,,, values for N-ethyl-N-methylaniline and the 4-chloro and 4-methyl derivatives for a variety of species are summarized in table 3. From V,,, evaluation there is a considerable species variation in the specific activity for both N-oxidation and N-dealkylation. In general, the order of decreasing N-dealkylase activity was rabbit > hamster > guinea- pig> mouse = rat. The species difference for N-oxidation was not so well defined and tended to be dependent on the substrate used. For example, the N-oxidation of N-ethyl-N-methylaniline was equal in the rabbit and rat, but considerably less than in the hamster, mouse and guinea-pig.
Influence of log P and pK, on metabolism Partition coefficients for the tertiary amines used in this study range from log
P = 2.74 for N-ethyl-N-methylaniline to log P = 4.54 for N-ethyl-N-methyl-4- aminobiphenyl reflecting the effect that the 4-substituent has on their lipophilicity (see table 4).
The pK, values range from 5.1 5 for N-ethyl-N-methyl-4-chloroaniline, the least basic compound, to 6.59 for N-ethyl-N-methyl-4-anisidine, the most basic com- pound (see table 4).
A comparison of the kinetic constants calculated for the metabolism of N-ethyl- N-methylaniline and the .l-methoxy, 4-phenyl, 4-chloro and 4-methyl derivatives by guinea-pig hepatic microsomal preparations with measured log P and pK, values
Xen
obio
tica
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
orth
Car
olin
a on
05/
14/1
3Fo
r pe
rson
al u
se o
nly.

&Substituted N-ethyl-N-methylaniline metabolism 519
Table 3. The apparent K, and V,,, values for the a-C- and N-oxidation of 4-substituted-N-ethyl-N- methylanilines by liver microsomes from various species.
Route of metabolism
N-Demethylation N-De-ethylation N-Oxidation
Substrate Species K m Vmax K m Vmax K m Vmax
N-Ethyl-N-rnethyl- aniline Rabbit 7.0 8.8 4 4 0.9 1 .o 1.4
Hamster 7.0 8.8 4.4 2.4 2 6 10.4 Guinea-pig 8.4 5.4 4.4 1.4 2.6 6.5 Rat 6.0 5.7 4 4 1.1 1 .o 1.4 Mouse 6.0 5.7 4.4 2.2 2.6 10.4
4-toluidine Rabbit 9.2 14.6 6.7 2.7 3.0 6.0 Hamster 9.2 108 8.8 3.7 4.6 5.8 Guinea-pig 9.2 11.5 8.8 3.0 1.4 2.3 Rat 9.2 7.1 8.8 1.4 4 6 1.6 Mouse 9.2 6.1 8.8 1.3 2 0 1.7
chloroaniline Rabbit 3.2 107 2.8 1.6 4 0 5.7 Hamster 5.5 13.8 6.4 2.5 3.6 4.0 Guinea-pig 5.5 8.5 5.3 3.8 4.0 6.7 Rat 5.5 7.7 2.8 0.9 2 2 1.6 Mouse 108 9.4 90 1.7 6.0 1.5
N-Ethyl-N-methyl-
N-Ethyl-N-methyl-4-
All animals were male. Kinetic constants were calculated from the mean results of two determinations. K, values are all x 1 0 - 4 ~ , Vmax values are expressed as nmol productlmg microsomal protein
per min.
Table 4. Partition coefficients, pK, values, apparent K, and V,,, values for some 4-substituted-N- ethyl-N-methylanilines for N- and a-C-oxidation by guinea-pig-liver microsomes.
N-Oxidation N-Demethylation NaOH/
A,,, (nm) octanol Km K m Compound major peak pK, iogp x10-4M vmaX x ~ o - ~ M vmaX N-Ethyl-N-methyl-4-
N-Ethyl-N-methyl-4-
N-Ethyl-N-methyl-
N-Ethyl-N-methyl-4-
N-Ethyl-N-methyl-4-
chloroaniline 255 5.1 5 3.60 4.0 6,7 5.5 8.5
aminobiphenyl 290 5.41 454 2.4 1.3 6 9 19.7
aniline 245 5.95 2.74 2.6 6.5 8.4 5.4
toluidine 245 6.39 2.85 1.4 2 3 9.2 11.5
anisidine 241 6.59 2.30 1 .o 3.3 15.6 17.3
The pK, values are the means nf seven determinations which did not differ by more than kO.05. Partition coefficient values are the means of five determinations. K, and VmaX are calculated from the mean results of two determinations. V,,,,x values quoted as nmol product formed/mg microsomal protein per min.
(see table 4) shows that an inverse relationship between the K,,, for N-oxidation and the pK, of the respective substrate is apparent. However, the 4-phenyl derivative does not fit into this pattern: in this case the extensive further metabolism of N - ethyl-N-methyl-4-aminobiphenyl-N-oxide suggests this compound to be atypical of the series (Patterson and Gorrod 1978). From the results it would appear that as the basicity of the substrate increases (increasing pK,), so does the affinity of the
Xen
obio
tica
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
orth
Car
olin
a on
05/
14/1
3Fo
r pe
rson
al u
se o
nly.

5 20 &Substituted N-ethyl-N-methylaniline metabolism
enzymesubstrate complex (decreasing K,). However, this is not always accom- panied by increasing specific activity of the reaction (increasing V,,,,,). In contrast to the above relationship, the N-demethylation of the tertiary amines showed that the increasing pK, resulted in increasing K,.
These results indicate that the pK, of the nitrogen group is some measure of the reactivity of that centre with regard to enzymic a-C- and N-oxidation. However, it seems likely that this parameter together with other molecular properties including lipid solubility (partition behaviour) act in concert to determine the extent of metabolism observed as measured by K, and V,,, values. In this respect some success has been achieved in structureactivity relationships for the N- demethylation of tertiary amines using equations based on both basicity and lipophilicity of the parent compound (Galliani, Rindone and Salmona 1978).
Acknowledgements This work was supported by the Science Research Council during a studentship
awarded to L. H. Patterson. The authors thank M r L. G. Disley for preparing the tissue fractions.
References ALBERT, A., and SERJEANT, E. P., 1971, Determination of Ionisation Constants (London: Chapman and
BICKEL, M. H., 1969, Pharmac. Rev. , 21, 325. COWAN, D., PATTERSON, L. H., DAMANI, L. A,, and GORROD, J. W., 1982, Bio-Med. Mass Spectrom.,
DAS, M. L., and ZIEGLER, D. M., 1970, Archs. Biochem. Biophys., 140, 300. DEVEREUX, T. R., and FOLJTS, J . R., 1974, Chemico-Biological Interactions, 8, 91. GORROD, J. W., 1973, Chemico-Biological Interactions, 7 , 289. GORROD, J. W., 1974, Dtsch. Apoth. Ztg. , 1887. GORROD, J. W., TEMPLE, D. J., and BECKETT, A. H., 1975 a, Xenobiotica, 5, 453. GORROD, J . W., TEMPLE, D. J., and BECKETT, A. H., 1975 b, Xenobiotica, 5 , 465. GALLIANI, G., RINDONE, B., and SALMONA, M., 1978, Bio-organic Chem., 1, 103. MCMAHON, R. E., 1966, J. Pharm. Sci., 55,457. PATTERSON, L. H., and GORROD, J . W., 1978. In Biological Oxidation of Nitrogen, edited by J . W. Gorrod
Hall).
9, 233.
(Amsterdam: Elst.vier/North-Holland), p. 471.
Xen
obio
tica
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
orth
Car
olin
a on
05/
14/1
3Fo
r pe
rson
al u
se o
nly.





























