Embed Size (px)
Citation preview

The sesquiterpene lactone parthenolide in combinationwith docetaxel reduces metastasis and improvessurvival in a xenograft model of breast cancer
Christopher J. Sweeney,1 Sanjana Mehrotra,2
Miral R. Sadaria,3 Suresh Kumar,3
Nicholas H. Shortle,3 Yaritzabel Roman,3
Carol Sheridan,3 Robert A. Campbell,3
Daryl J. Murry,7 Sunil Badve,2
and Harikrishna Nakshatri3,4,5,6
Departments of 1Medicine, 2Pathology, 3Surgery, and4Biochemistry and Molecular Biology, 5Walther Oncology Center,Indiana University School of Medicine; 6Walther Cancer Institute,Indianapolis, Indiana; and 7College of Pharmacy, The Universityof Iowa, Iowa City, Iowa
AbstractParthenolide, a sesquiterpene lactone, shows antitumoractivity in vitro, which correlates with its ability to inhibitthe DNA binding of the antiapoptotic transcription factornuclear factor KB (NF-KB) and activation of the c-Jun NH2-terminal kinase. In this study, we investigated the chemo-sensitizing activity of parthenolide in vitro as well as inMDA-MB-231 cell–derived xenograft metastasis model ofbreast cancer. HBL-100 and MDA-MB-231 cells wereused to measure the antitumor and chemosensitizingactivity of parthenolide in vitro. Parthenolide was effec-tive either alone or in combination with docetaxel inreducing colony formation, inducing apoptosis and reduc-ing the expression of prometastatic genes IL-8 and theantiapoptotic gene GADD45b1 in vitro. In an adjuvantsetting, animals treated with parthenolide and docetaxelcombination showed significantly enhanced survival com-pared with untreated animals or animals treated witheither drug. The enhanced survival in the combination armwas associated with reduced lung metastases. In addition,nuclear NF-KB levels were lower in residual tumors andlung metastasis of animals treated with parthenolide,docetaxel, or both. In the established orthotopic model,there was a trend toward slower growth in the partheno-
lide-treated animals but no statistically significant findingswere seen. These results for the first time reveal thesignificant in vivo chemosensitizing properties of parthe-nolide in the metastatic breast cancer setting and supportthe contention that metastases are very reliant on activa-tion of NF-KB. [Mol Cancer Ther 2005;4(6):1004–12]
IntroductionIntrinsic or acquired resistance to chemotherapy is a majorclinical problem. Several factors including expression levelsof multidrug resistance genes, and proapoptotic and anti-apoptotic genes determine the response to chemotherapy(1, 2). Antiapoptotic genes that determine response tochemotherapy include cIAP-1, cIAP-2, XIAP, TRAF-1,TRAF-2, Bcl-2, Bfl-1/A1, Bcl-xL, c-FLIP, and Mn-SOD (3, 4).The transcription factors nuclear factor nB (NF-nB), activatorprotein 1, and signal transducers and activators of tran-scription 3 are the major transcription factors that control theexpression of antiapoptotic and proapoptotic genes thatdetermine the sensitivity of cancer cells to therapy (2, 4, 5).Aberrant activation of these transcription factors, therefore,may lead to changes in antiapoptotic and proapoptotic genethreshold and resistance to chemotherapy.
NF-nB is an extracellular signal–activated transcriptionfactor, which usually resides in the cytoplasm of resting cellsdue its association with inhibitor of nB proteins (3, 6). Onexposure of cells to cytokines and growth factors such asinterleukin (IL)-1, tumor necrosis factor (TNF), or epidermalgrowth factor, a series of signaling events trigger phosphor-ylation and degradation of inhibitors of nB. NF-nB, liberatedfrom inhibitor of nB, translocates to the nucleus, bindsspecific response elements in the promoter region of targetgenes, and regulates gene expression (6). A distinctphosphorylation network controls subcellular distributionand/or activity of activator protein 1 and signal transducersand activators of transcription 3 transcription factors (7, 8).
Constitutive activation of NF-nB is observed in a numberof cancers including breast cancer (3). It is likely thatconstitutively active NF-nB contributes to chemotherapeu-tic resistance by up-regulating the expression of antiapop-totic genes. Consistent with this possibility, we and othershave observed that inhibition of NF-nB through eitheroverexpression of inhibitors of nB or prior exposure tochemical inhibitors, such as parthenolide, sensitize cancercells to chemotherapeutic drugs such as paclitaxel andCPT11 (9, 10).
Parthenolide is the major focus of this investigation. It isderived from the herb feverfew, which is being used formigraine prophylaxis (11). Apart from inhibiting NF-nB,parthenolide inhibits signal transducers and activators oftranscription 3 activity (12, 13). Depending on the cell type,
Received 1/31/05; revised 3/26/05; accepted 4/11/05.
Grant support: American Institute for Cancer Research grants 00A047 and03A069-REN and grants-in-aid from Aventis Pharmaceuticals(H. Nakshatri) and National Cancer Institute grant CA37403 (S. Badve).
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.
Note: H. Nakshatri is a Marian J. Morrison Investigator in Breast CancerResearch.
Requests for reprints: Harikrishna Nakshatri, R-202, Indiana CancerResearch Institute, 1044 West Walnut Street, Indianapolis, IN 46202.Phone: 317-278-2238; Fax: 317-274-0396. E-mail: [email protected]
Copyright C 2005 American Association for Cancer Research.
1004
Mol Cancer Ther 2005;4(6). June 2005
Research. on November 22, 2018. © 2005 American Association for Cancermct.aacrjournals.org Downloaded from

it can activate or repress c-Jun NH2-terminal kinase (JNK),which modulates the activity of activator protein 1 (14–16).We and others have shown the requirement of JNKactivation in parthenolide-mediated sensitization of cancercells to TNF and TNF-related apoptosis inducing ligand(15, 16). The ability of parthenolide to alter the function ofthree transcription factors makes it an ideal antitumoragent that can sensitize cancer cells to chemotherapy byreducing the level/activity of antiapoptotic proteins.Despite numerous studies showing antitumor activityin vitro , there are few in vivo studies with parthenolide(9, 16–18). In this report, we analyze the chemosensitizingproperties of parthenolide in vitro and in a xenograft modelof breast cancer focusing on its effects on NF-nB andcytokines and antiapoptotic proteins under its control.
Materials and MethodsBreast Cancer Cell LinesMDA-MB-231 and HBL-100 cells were obtained from
American Type Tissue Culture Collection (Rockville, MD)and grown in MEM plus 10% FCS (9).
In vitro DrugTreatment and Apoptosis AssaysSensitivity of cells to parthenolide, docetaxel, or combi-
nation was measured by clonogenic assay (15). Briefly, cells(100 cells per well in six-well plates, assayed in triplicate)were treated with the indicated drugs for 24 hours. Thedrugs were removed, cells were washed, and fresh mediawas added. Number of colonies after 21 days of drugtreatment was counted after staining with Giemsa stain.The amount of colony formation for the treated groups wasexpressed as a percentage of controls (relative colonyformation). The carboxyfluorescein FLICA apoptosis de-tection kit (Immunohistochemistry Technologies, LLC,Bloomington, MN) was used to measure typical apoptosis,atypical apoptosis, and necrosis (19). Briefly, 2 � 105 cellsgrown overnight on a 60-mm plate were treated withindicated drugs for 48 hours. Both attached and floatingcells were collected by trypsinization and incubatedwith carboxyfluorescein-labeled pan-caspase inhibitorcarboxyfluorescein-benzyloxycarbonyl-valine-alanine-aspartic acid-fluoromethyl ketone for 2 hours at 37jC.Labeled cells were rinsed twice in PBS and resuspendedin 300 AL of PBS containing 0.3 Ag of propidium iodide.Apoptotic cells were identified by FACScan analysis aspreviously described (19). All apoptosis assays weredone two to five times and representative data arepresented in the text.
Northern Blot AnalysisCells were treated with indicated drugs and RNA
was isolated using RNAeasy kit from Qiagen Sciences(Valencia, CA). Northern blotting was done as previouslydescribed (20).
Mammary Fat Pad Injection of Tumor Cells andTreatment
MDA-MB-231 cells, which have been passed once throughmammary fat pad (TMD231), were used for mammary fatpad injection because these cells formed tumors reproduc-
ibly and were metastatic. Cells, 106, were injected into themammary fat pad of 6- to 7-week-old female nude mice. Inthe metastasis survival model, tumors (f100 mm3) wereremoved after 6 weeks and treatment was initiated a dayafter removal of the tumors and mice were assigned to eachtreatment group based on average tumor size and weight.Average weight of animals and size of tumors were similaramong different groups. In the orthotopic model, thetumors were left in place and treatment commenced 14days after cell implantation. In both experiments, parthe-nolide (40 mg/kg as a slurry in 10% ethanol) was givendaily by oral gavage. Docetaxel (5 mg/kg in 13% ethanol)was given i.p. once weekly. Experiment was done thricewith 8 to 11 animals per treatment group in eachexperiment. Lungs, primary tumor regrowth, and/or bloodwas collected when the animals died or were sacrificed asper the advice of attending Veterinarian or at the end of theexperiment. Mice were treated for up to 45 days after tumorremoval in the metastasis survival model and the number ofmice euthanized at this time point was scored. In theorthotopic mammary fat pad model, mice were treated for6 weeks, starting 2 weeks after implantation when thetumors were established. The tumors were measuredweekly with vernier calipers. Tumor volume was calculatedusing the following formula: sagittal dimension (mm) �[cross dimension (mm)]2 / 2, and expressed in cubicmillimeter (21).
Analysis ofTumor, Lung, and Serum SamplesLungs were analyzed for metastasis by H&E staining.
Metastasis index was calculated as previously described(22). Briefly, the number and size of metastasis in two to fivefields per sample were calculated and a score of 4+ wasgiven to a sample with the highest metastasis and relativemetastasis in other samples was calculated. LINCOplexmultiplex immunoassay system (Linco Research, Inc.,St. Charles, MO) was used to measure levels of cytokinesin the serum and the samples were analyzed in duplicate.
Immunohistochemistry for p65Sections, 4 Am in thickness, were deparaffinized and
hydrated. Antigen retrieval was done in 140 mmol/Lcitrate buffer (pH 6.0) in a decloaking chamber (BioCare,Walnut Creek, CA) at 115jC for 3 minutes. The slides werethen transferred to boiling deionized water and allowed tocool for 20 minutes at room temperature. The endogenousperoxidase activity was blocked by Peroxo-Block (Zymed,San Francisco, CA) for 2 minutes. The slides were thenincubated with rabbit polyclonal p65 antibody (Lab Vision-NeoMarkers, Fremont, CA; diluted 1:100) for 3 hours atroom temperature, washed in Optimax (Biogenex, SanRamon, CA), followed by incubation for 10 minutes withhorseradish peroxidase polymer conjugate (Zymed). Thestain was visualized using Dako liquid DAB plus substratechromogen solution and hematoxylin QS (Vector Labora-tories, Burlingame, CA) counterstain. Ten random micro-scopic fields were identified and the number of nucleistaining for p65 was documented and expressed as percentof total nuclei in the 10 fields. Nuclear counts were doneusing a hemocytometer counter.
Molecular Cancer Therapeutics 1005
Mol Cancer Ther 2005;4(6). June 2005
Research. on November 22, 2018. © 2005 American Association for Cancermct.aacrjournals.org Downloaded from
Statistical AnalysisFor the metastasis survival model, Prism 3 (version 3.02)
for Windows (GraphPad Software, San Diego, CA) was usedto generate a Kaplan-Meier survival curves. The log-ranktest was then used to compare the survival of theparthenolide alone, docetaxel alone, and docetaxel plusparthenolide groups with the survival of the control group.All analyses used a two-sided P value. For the orthotopicmammary fat pad experiment, analysis was done using SASVersion 8 (Cary, NC). Tumor volume (mm3) was obtainedfrom 80 mice (20 per group) treated in four treatment groupsacross eight time points. A linear growth curve model was fitusing a mixed model with a variance-covariance model thatincorporates correlations of observations within a cage andacross time. To compare the rates of change of the tumorvolume (mm3) between mice treated with parthenolide,docetaxel, parthenolide and docetaxel, and control, the slopeestimates from the model were compared. No adjustmentfor multiple comparisons was made.
To statistically compare the results between groups ofthe lung metastasis index, nuclear NF-nB staining in tumorcells, and cytokine levels in the serum, the Kruskal-Wallistest (nonparametric ANOVA) with Dunn’s post test formultiple comparisons was done using GraphPad InStatversion 3.00 for Windows 95 (GraphPad Software). Graph-Pad program was also used for statistical analysis ofclonogenic and apoptosis assay results.
ResultsParthenolide Increases the Sensitivity of Breast
Cancer Cells to Docetaxel In vitroPrevious studies have shown that JNK increases whereas
NF-nB reduces the sensitivity of cancer cells to docetaxel
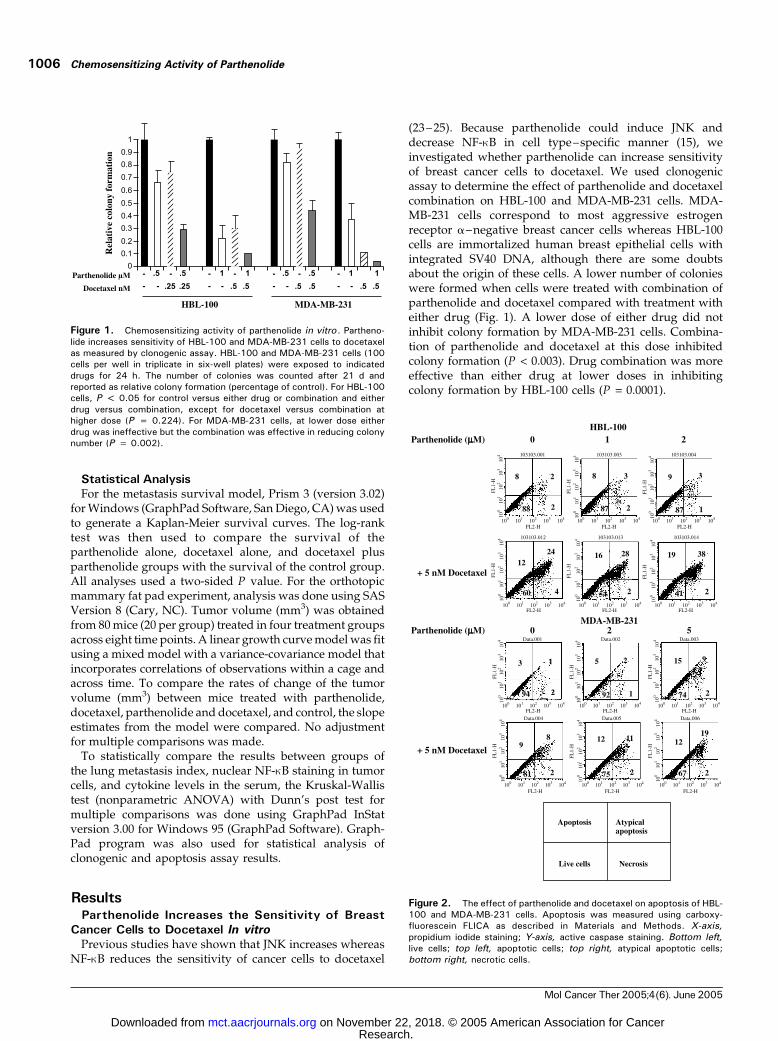
(23 –25). Because parthenolide could induce JNK anddecrease NF-nB in cell type–specific manner (15), weinvestigated whether parthenolide can increase sensitivityof breast cancer cells to docetaxel. We used clonogenicassay to determine the effect of parthenolide and docetaxelcombination on HBL-100 and MDA-MB-231 cells. MDA-MB-231 cells correspond to most aggressive estrogenreceptor a–negative breast cancer cells whereas HBL-100cells are immortalized human breast epithelial cells withintegrated SV40 DNA, although there are some doubtsabout the origin of these cells. A lower number of colonieswere formed when cells were treated with combination ofparthenolide and docetaxel compared with treatment witheither drug (Fig. 1). A lower dose of either drug did notinhibit colony formation by MDA-MB-231 cells. Combina-tion of parthenolide and docetaxel at this dose inhibitedcolony formation (P < 0.003). Drug combination was moreeffective than either drug at lower doses in inhibitingcolony formation by HBL-100 cells (P = 0.0001).
Figure 1. Chemosensitizing activity of parthenolide in vitro . Partheno-lide increases sensitivity of HBL-100 and MDA-MB-231 cells to docetaxelas measured by clonogenic assay. HBL-100 and MDA-MB-231 cells (100cells per well in triplicate in six-well plates) were exposed to indicateddrugs for 24 h. The number of colonies was counted after 21 d andreported as relative colony formation (percentage of control). For HBL-100cells, P < 0.05 for control versus either drug or combination and eitherdrug versus combination, except for docetaxel versus combination athigher dose (P = 0.224). For MDA-MB-231 cells, at lower dose eitherdrug was ineffective but the combination was effective in reducing colonynumber (P = 0.002).
Figure 2. The effect of parthenolide and docetaxel on apoptosis of HBL-100 and MDA-MB-231 cells. Apoptosis was measured using carboxy-fluorescein FLICA as described in Materials and Methods. X-axis,propidium iodide staining; Y-axis, active caspase staining. Bottom left,live cells; top left, apoptotic cells; top right, atypical apoptotic cells;bottom right, necrotic cells.
Chemosensitizing Activity of Parthenolide1006
Mol Cancer Ther 2005;4(6). June 2005
Research. on November 22, 2018. © 2005 American Association for Cancermct.aacrjournals.org Downloaded from

Parthenolide and Docetaxel Combination EnhancesAtypical Apoptosis of Cancer Cells
We used carboxyfluorescein FLICA to measure apoptosisof drug-treated cells (15). This assay simultaneouslymeasures cells undergoing typical apoptosis, atypicalapoptosis (apoptosis with loss of plasma membraneintegrity), and necrosis. However, this assay is not assensitive as clonogenic assay, thus requiring higherconcentration of drugs to detect apoptosis. Docetaxel(5 nmol/L), but not parthenolide (2 Amol/L), inducedatypical apoptosis of HBL-100 cells (Fig. 2A). Atypicalapoptosis was further enhanced in cells treated withboth parthenolide and docetaxel. Total apoptosis/atypicalapoptosis/necrosis is shown in Table 1. Parthenolidetreatment alone caused apoptosis of MDA-MB-231 cells(Fig. 2B). Cells treated with both parthenolide and doce-taxel displayed elevated levels of atypical apoptosis. Thus,increased cell death observed in cells treated with acombination of parthenolide and docetaxel is mostly dueto a shift in cell death from typical apoptosis to atypicalapoptosis.
Parthenolide and Docetaxel Combination Is Effectivein Enhancing Survival and Reducing Metastases in aXenograft Model of Breast Cancer
To determine whether parthenolide and docetaxelcombination is effective in an in vivo metastasis model,we used MDA-MB-231 xenograft model to examine theeffect of drugs in an adjuvant setting. In this series ofexperiments, primary tumors were surgically removedwhen tumors reached f100 mm3 (f6 weeks afterimplantation) and treatment was initiated a day aftersurgery. We have observed that surgical removal ofprimary tumor is necessary to detect metastasis andterminal progression of disease in this model. Experimentswere done thrice with 8 to 12 animals per treatment groupin each experiment. The 30-day survival data from the firsttwo experiments, in which docetaxel was given at 5 mg/kgonce a week and parthenolide at 40 mg/kg daily, are
shown in Fig. 3A. It is notable that in the two experiments,the average number of mice alive at 30 days was f60% incontrol, docetaxel, and parthenolide (Fig. 3A). However,the average number of mice alive for the combination armin both experiments was 90%. The unpaired Student’st test with a two-sided P value showed that this wasstatistically significant (P = 0.0489). For the long-termsurvival analysis with mice treated until 45 days aftertumor removal, a Kaplan-Meier survival curve wasgenerated and is shown in Fig. 3B. Animals treated withdocetaxel and parthenolide showed significantly enhancedsurvival with 90% of mice alive at day 45 compared with73% (8 of 11) in the docetaxel alone arm, 64% (7 of 11) inthe parthenolide arm, and 34% (4 of 11) in the control arm.The improvement in overall survival in the combinationarm was found to be statistically significant (log-rank test,two-sided P = 0.0171). It should also be noted that thesurvival of animals treated with docetaxel alone orparthenolide alone was not statistically significantlydifferent from control animals but was numerically better.Taken together, these results indicate that parthenolide,when given daily at the doses described, can function asa chemosensitizer in vivo .
Table 1. Parthenolide and docetaxel– induced apoptosis/necro-sis of HBL-100 and MDA-MB-231 cells
Cell line/drug % Apoptosis/necrosis F SE
P
HBL-100Vehicle 13.5 F 1.5 —Parthenolide 2 Amol/L 13.5 F 0.5 —Docetaxel 5 nmol/L 40.5 F 1.5 0.006*Parthenolide + docetaxel 55.5 F 3.51 0.008,* 0.008c
MDA-MB-231Vehicle 6.5 F 0.5 —Parthenolide 5 Amol/L 24 F 2.0 0.02*Docetaxel 5 nmol/L 16 F 3.0 —Parthenolide + docetaxel 36 F 3.0 0.01,* 0.04b
*Versus vehicle.cVersus parthenolide.bVersus docetaxel.
Figure 3. The effect of parthenolide and docetaxel treatment on survivalin a metastasis xenograft model. A, 30-day survival rate from twoexperiments (n = 20–24). B, long-term survival analysis (n = 10–11).Tumor cell implantation, surgery, and treatment protocol are described inMaterials and Methods.
Molecular Cancer Therapeutics 1007
Mol Cancer Ther 2005;4(6). June 2005
Research. on November 22, 2018. © 2005 American Association for Cancermct.aacrjournals.org Downloaded from
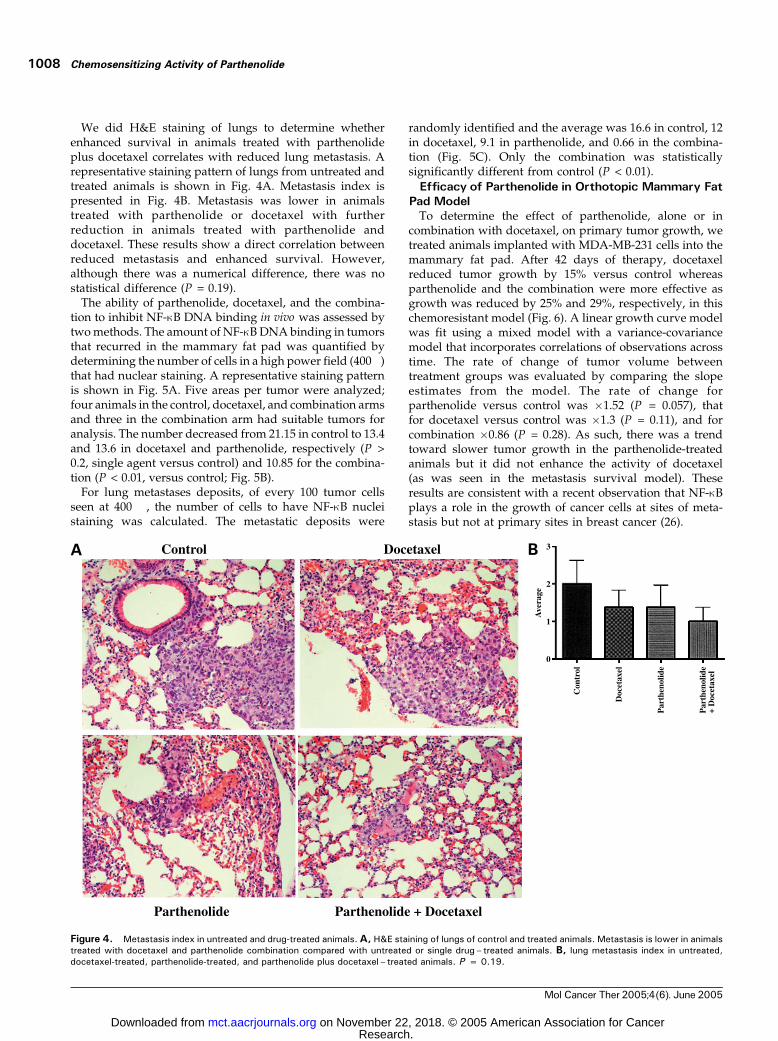
We did H&E staining of lungs to determine whetherenhanced survival in animals treated with parthenolideplus docetaxel correlates with reduced lung metastasis. Arepresentative staining pattern of lungs from untreated andtreated animals is shown in Fig. 4A. Metastasis index ispresented in Fig. 4B. Metastasis was lower in animalstreated with parthenolide or docetaxel with furtherreduction in animals treated with parthenolide anddocetaxel. These results show a direct correlation betweenreduced metastasis and enhanced survival. However,although there was a numerical difference, there was nostatistical difference (P = 0.19).
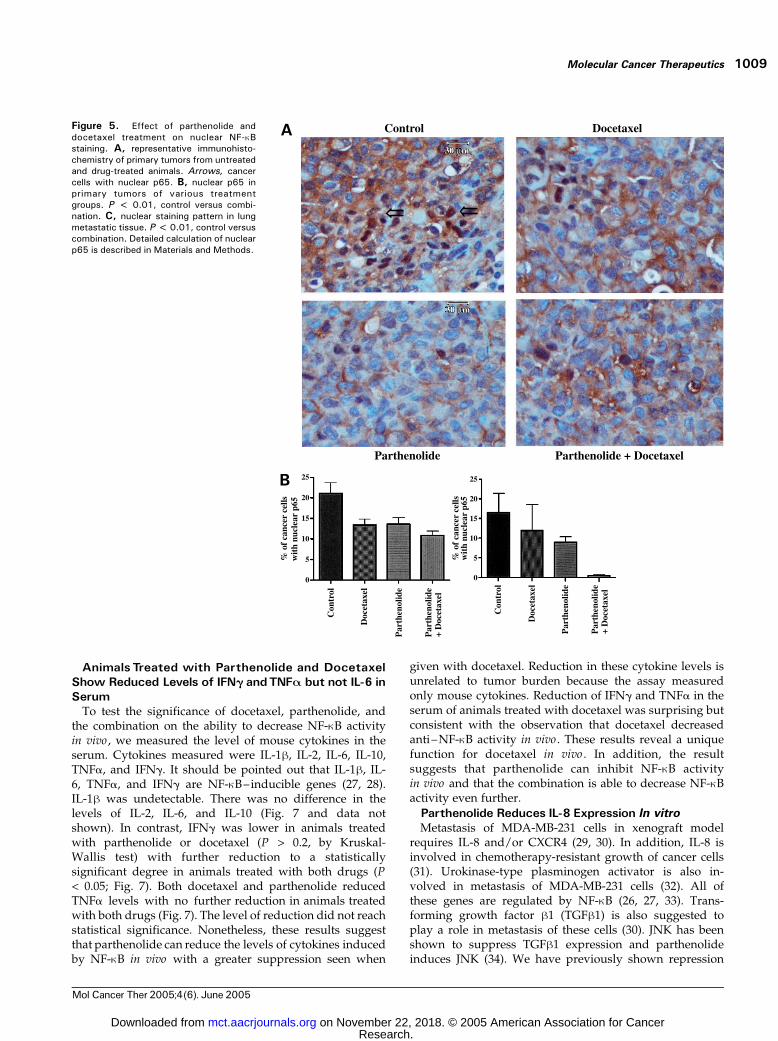
The ability of parthenolide, docetaxel, and the combina-tion to inhibit NF-nB DNA binding in vivo was assessed bytwo methods. The amount of NF-nB DNA binding in tumorsthat recurred in the mammary fat pad was quantified bydetermining the number of cells in a high power field (400�)that had nuclear staining. A representative staining patternis shown in Fig. 5A. Five areas per tumor were analyzed;four animals in the control, docetaxel, and combination armsand three in the combination arm had suitable tumors foranalysis. The number decreased from 21.15 in control to 13.4and 13.6 in docetaxel and parthenolide, respectively (P >0.2, single agent versus control) and 10.85 for the combina-tion (P < 0.01, versus control; Fig. 5B).
For lung metastases deposits, of every 100 tumor cellsseen at 400 �, the number of cells to have NF-nB nucleistaining was calculated. The metastatic deposits were
randomly identified and the average was 16.6 in control, 12in docetaxel, 9.1 in parthenolide, and 0.66 in the combina-tion (Fig. 5C). Only the combination was statisticallysignificantly different from control (P < 0.01).
Efficacy of Parthenolide in Orthotopic Mammary FatPad Model
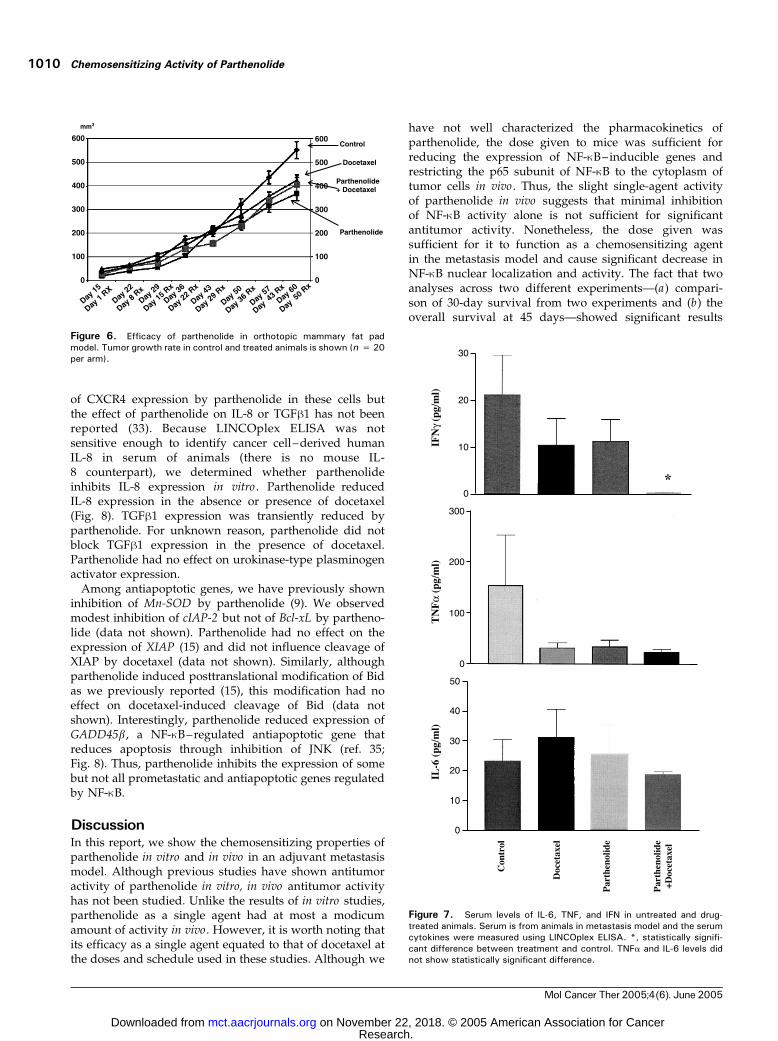
To determine the effect of parthenolide, alone or incombination with docetaxel, on primary tumor growth, wetreated animals implanted with MDA-MB-231 cells into themammary fat pad. After 42 days of therapy, docetaxelreduced tumor growth by 15% versus control whereasparthenolide and the combination were more effective asgrowth was reduced by 25% and 29%, respectively, in thischemoresistant model (Fig. 6). A linear growth curve modelwas fit using a mixed model with a variance-covariancemodel that incorporates correlations of observations acrosstime. The rate of change of tumor volume betweentreatment groups was evaluated by comparing the slopeestimates from the model. The rate of change forparthenolide versus control was �1.52 (P = 0.057), thatfor docetaxel versus control was �1.3 (P = 0.11), and forcombination �0.86 (P = 0.28). As such, there was a trendtoward slower tumor growth in the parthenolide-treatedanimals but it did not enhance the activity of docetaxel(as was seen in the metastasis survival model). Theseresults are consistent with a recent observation that NF-nBplays a role in the growth of cancer cells at sites of meta-stasis but not at primary sites in breast cancer (26).
Figure 4. Metastasis index in untreated and drug-treated animals.A, H&E staining of lungs of control and treated animals. Metastasis is lower in animalstreated with docetaxel and parthenolide combination compared with untreated or single drug– treated animals. B, lung metastasis index in untreated,docetaxel-treated, parthenolide-treated, and parthenolide plus docetaxel – treated animals. P = 0.19.
Chemosensitizing Activity of Parthenolide1008
Mol Cancer Ther 2005;4(6). June 2005
Research. on November 22, 2018. © 2005 American Association for Cancermct.aacrjournals.org Downloaded from
Animals Treated with Parthenolide and DocetaxelShow Reduced Levels of IFN; andTNFA but not IL-6 inSerum
To test the significance of docetaxel, parthenolide, andthe combination on the ability to decrease NF-nB activityin vivo , we measured the level of mouse cytokines in theserum. Cytokines measured were IL-1h, IL-2, IL-6, IL-10,TNFa, and IFNg. It should be pointed out that IL-1h, IL-6, TNFa, and IFNg are NF-nB–inducible genes (27, 28).IL-1h was undetectable. There was no difference in thelevels of IL-2, IL-6, and IL-10 (Fig. 7 and data notshown). In contrast, IFNg was lower in animals treatedwith parthenolide or docetaxel (P > 0.2, by Kruskal-Wallis test) with further reduction to a statisticallysignificant degree in animals treated with both drugs (P< 0.05; Fig. 7). Both docetaxel and parthenolide reducedTNFa levels with no further reduction in animals treatedwith both drugs (Fig. 7). The level of reduction did not reachstatistical significance. Nonetheless, these results suggestthat parthenolide can reduce the levels of cytokines inducedby NF-nB in vivo with a greater suppression seen when
given with docetaxel. Reduction in these cytokine levels isunrelated to tumor burden because the assay measuredonly mouse cytokines. Reduction of IFNg and TNFa in theserum of animals treated with docetaxel was surprising butconsistent with the observation that docetaxel decreasedanti–NF-nB activity in vivo . These results reveal a uniquefunction for docetaxel in vivo . In addition, the resultsuggests that parthenolide can inhibit NF-nB activityin vivo and that the combination is able to decrease NF-nBactivity even further.
Parthenolide Reduces IL-8 Expression In vitroMetastasis of MDA-MB-231 cells in xenograft model
requires IL-8 and/or CXCR4 (29, 30). In addition, IL-8 isinvolved in chemotherapy-resistant growth of cancer cells(31). Urokinase-type plasminogen activator is also in-volved in metastasis of MDA-MB-231 cells (32). All ofthese genes are regulated by NF-nB (26, 27, 33). Trans-forming growth factor h1 (TGFh1) is also suggested toplay a role in metastasis of these cells (30). JNK has beenshown to suppress TGFh1 expression and parthenolideinduces JNK (34). We have previously shown repression
Figure 5. Effect of parthenolide anddocetaxel treatment on nuclear NF-nBstaining. A, representative immunohisto-chemistry of primary tumors from untreatedand drug-treated animals. Arrows, cancercells with nuclear p65. B, nuclear p65 inprimary tumors of various treatmentgroups. P < 0.01, control versus combi-nation. C, nuclear staining pattern in lungmetastatic tissue. P < 0.01, control versuscombination. Detailed calculation of nuclearp65 is described in Materials and Methods.
Molecular Cancer Therapeutics 1009
Mol Cancer Ther 2005;4(6). June 2005
Research. on November 22, 2018. © 2005 American Association for Cancermct.aacrjournals.org Downloaded from
of CXCR4 expression by parthenolide in these cells butthe effect of parthenolide on IL-8 or TGFh1 has not beenreported (33). Because LINCOplex ELISA was notsensitive enough to identify cancer cell–derived humanIL-8 in serum of animals (there is no mouse IL-8 counterpart), we determined whether parthenolideinhibits IL-8 expression in vitro . Parthenolide reducedIL-8 expression in the absence or presence of docetaxel(Fig. 8). TGFh1 expression was transiently reduced byparthenolide. For unknown reason, parthenolide did notblock TGFh1 expression in the presence of docetaxel.Parthenolide had no effect on urokinase-type plasminogenactivator expression.
Among antiapoptotic genes, we have previously showninhibition of Mn-SOD by parthenolide (9). We observedmodest inhibition of cIAP-2 but not of Bcl-xL by partheno-lide (data not shown). Parthenolide had no effect on theexpression of XIAP (15) and did not influence cleavage ofXIAP by docetaxel (data not shown). Similarly, althoughparthenolide induced posttranslational modification of Bidas we previously reported (15), this modification had noeffect on docetaxel-induced cleavage of Bid (data notshown). Interestingly, parthenolide reduced expression ofGADD45b, a NF-nB–regulated antiapoptotic gene thatreduces apoptosis through inhibition of JNK (ref. 35;Fig. 8). Thus, parthenolide inhibits the expression of somebut not all prometastatic and antiapoptotic genes regulatedby NF-nB.
DiscussionIn this report, we show the chemosensitizing properties ofparthenolide in vitro and in vivo in an adjuvant metastasismodel. Although previous studies have shown antitumoractivity of parthenolide in vitro, in vivo antitumor activityhas not been studied. Unlike the results of in vitro studies,parthenolide as a single agent had at most a modicumamount of activity in vivo. However, it is worth noting thatits efficacy as a single agent equated to that of docetaxel atthe doses and schedule used in these studies. Although we
have not well characterized the pharmacokinetics ofparthenolide, the dose given to mice was sufficient forreducing the expression of NF-nB–inducible genes andrestricting the p65 subunit of NF-nB to the cytoplasm oftumor cells in vivo . Thus, the slight single-agent activityof parthenolide in vivo suggests that minimal inhibitionof NF-nB activity alone is not sufficient for significantantitumor activity. Nonetheless, the dose given wassufficient for it to function as a chemosensitizing agentin the metastasis model and cause significant decrease inNF-nB nuclear localization and activity. The fact that twoanalyses across two different experiments—(a) compari-son of 30-day survival from two experiments and (b) theoverall survival at 45 days—showed significant results
Figure 6. Efficacy of parthenolide in orthotopic mammary fat padmodel. Tumor growth rate in control and treated animals is shown (n = 20per arm).
Figure 7. Serum levels of IL-6, TNF, and IFN in untreated and drug-treated animals. Serum is from animals in metastasis model and the serumcytokines were measured using LINCOplex ELISA. *, statistically signifi-cant difference between treatment and control. TNFa and IL-6 levels didnot show statistically significant difference.
Chemosensitizing Activity of Parthenolide1010
Mol Cancer Ther 2005;4(6). June 2005
Research. on November 22, 2018. © 2005 American Association for Cancermct.aacrjournals.org Downloaded from

strongly supports this conclusion. This very interestingand unique finding is supported by the recent datashowing exquisite dependence of breast cancer metasta-ses, but not of primary tumor growth, on NF-nB (26).
Although constitutive NF-nB activation is reported in avariety of cancers (36, 37), its role in specific stages ofcancer progression is just beginning to be defined. Forexample, recent studies suggest the requirement of NF-nBin tumor promotion of the colitis-associated cancer and themalignant phenotype of squamous cell carcinoma (38, 39).In contrast, NF-nB is not required in the early phases ofhepatocyte transformation but is required for progressionto hepatocellular carcinoma (40). Our study and a study byHuber et al. (26) suggest the requirement of NF-nB inmetastasis but not in proliferation of breast cancer cells atprimary sites. Thus, anti-NF-nB–based therapies may havecancer type–specific effects.
We observed two distinct effects of parthenolide in vivo:reducing metastasis and increasing the sensitivity todocetaxel. Reduced metastasis in parthenolide-treatedanimals may be a result of reduced IFNg production. IFNg
has been shown to reduce the growth of primary breastcancers but increases metastasis (41). Similarly, lower levelsof TNFa in parthenolide-treated animals may have resultedin reduced growth of cancer cells at sites of metastasis.TNFa level is elevated in the serum of patients with lymphnode–positive breast cancer and it is believed to play a rolein angiogenesis (42, 43).
Other possible parthenolide targets that play a rolein reducing metastasis include IL-8, TGFh1, and CXCR4(Fig. 8).
Exactly how parthenolide sensitizes to docetaxel in vivo isnot known but may involve direct modulation of anti-apoptotic gene expression or is a consequence of reducedprometastatic gene expression under NF-nB control. MDA-MB-231 cells used in this study express very high levels
of NF-nB–regulated antiapoptotic genes, including Bcl-xL,TRAF-1, cIAP-2, and Mn-SOD , compared with breast cancercells that lack constitutively active NF-nB (9). With theexception of Mn-SOD and cIAP-2 , to a lesser extent,parthenolide had minimum effect on the expression of thesegenes (data not shown). In this study, we have identifiedGADD45b, an antiapoptotic gene, as a target of partheno-lide. Others have shown regulation of proapoptotic proteinsBax and Bak by parthenolide (44). Thus, the chemo-sensitizing activity of parthenolide could be secondary toreduced expression levels of select antiapoptotic genes.Alternatively, the chemosensitizing action of parthenolidemay be an indirect consequence of repression of CXCR4,IL-8 , and TGFb1 expressions. These genes have beenshown to modulate antiapoptotic gene expression, apartfrom playing a role in metastasis (31, 45–47).
Docetaxel is a very important therapy for breast cancer assingle agent or in combination with other agents (48, 49).However, docetaxel resistance is observed clinically. Ourstudies showing the beneficial effects of the docetaxel andparthenolide combination in an adjuvant setting in ananimal model raise the hope for a similar combinationtreatment of docetaxel-resistant breast cancers. We have notobserved any apparent toxicity in animals treated withparthenolide alone or in combination with docetaxel evenafter 10 weeks of continuous treatment. Additional pre-clinical studies in a syngeneic model may be required toconfirm the effectiveness as well as the lack of toxicity ofthe combination treatment before our studies can betranslated to clinic. The other major issue to resolve isimprovement in the solubility and bioavailability ofparthenolide. Poor solubility prevented us from dosinganimals at more than 40 mg/kg of parthenolide. Prelimi-nary pharmacokinetic studies have shown low nanomolarconcentrations of parthenolide (f200 nmol/L; data notshown) in serum of treated animals, well below the targetplasma concentrations of 5 Amol/L that we were aiming forbased on our in vitro data. Efforts to make analogues withbetter bioavailability but maintaining the same degree ofactivity are under way. It is also of note that viable cancercells were seen despite low amounts of NF-nB aftertreatment with docetaxel and parthenolide. This probablyindicates that other survival pathways are important andthat combining parthenolide derivatives with improvedpharmacologic properties with other agents that blocksurvival pathways will be needed.
Acknowledgments
We thank Joan Dunn for technical assistance.
References
1. Johnstone RW, Ruefli AA, Lowe SW. Apoptosis: a link between cancergenetics and chemotherapy. Cell 2002;108:153–64.
2. Pommier Y, Sordet O, Antony S, Hayward RL, Kohn KW. Apoptosisdefects and chemotherapy resistance: molecular interaction maps andnetworks. Oncogene 2004;23:2934–49.
3. Baldwin AS. Control of oncogenesis and cancer therapy resistance bythe transcription factor NF-nB. J Clin Invest 2001;107:241–6.
Figure 8. Parthenolide inhibits IL-8 and TGFh1 expression in MDA-MB-231 cells in vitro . MDA-MB-231 cells were treated with parthenolide (5Amol/L) either alone or in combination with docetaxel (5 nmol/L) for theindicated time. IL-8, TGFh1, urokinase-type plasminogen activator (uPA ),and GADD45h expressions were measured by Northern blot. The sameblot was reprobed with 36B4, a ribosomal protein gene, to ensure equalloading.
Molecular Cancer Therapeutics 1011
Mol Cancer Ther 2005;4(6). June 2005
Research. on November 22, 2018. © 2005 American Association for Cancermct.aacrjournals.org Downloaded from

4. Karin M, Lin A. NF-nB at the crossroads of life and death. Nat Immunol2002;3:221–7.
5. Darnell JE Jr. Transcription factors as targets for cancer therapy. NatRev Cancer 2002;2:740–9.
6. Ghosh S, May MJ, Kopp EB. NF-nB and Rel proteins: evolutionarilyconserved mediators of immune responses. Annu Rev Immunol 1998;16:225–60.
7. Shaulian E, Karin M. AP-1 as a regulator of cell life and death. Nat CellBiol 2002;4:E131–6.
8. Levy DE, Darnell JE Jr. Stats: transcriptional control and biologicalimpact. Nat Rev Mol Cell Biol 2002;3:651–62.
9. Patel NM, Nozaki S, Shortle NH, et al. Paclitaxel sensitivity of breastcancer cells with constitutively active NF-nB is enhanced by InBa super-repressor and parthenolide. Oncogene 2000;19:4159–69.
10. Wang CY, Cusack JC Jr., Liu R, Baldwin AS Jr. Control of induciblechemoresistance: enhanced anti-tumor therapy through increased apo-ptosis by inhibition of NF-nB. Nat Med 1999;5:412–7.
11. Murphy JJ, Heptinstall S, Mitchell JR. Randomised double-blindplacebo-controlled trial of feverfew in migraine prevention. Lancet 1988;2:189–92.
12. Hehner SP, Hofmann TG, Droge W, Schmitz ML. The antiinflamma-tory sesquiterpene lactone parthenolide inhibits NF-nB by targeting the InBkinase complex. J Immunol 1999;163:5617–23.
13. Sobota R, Szwed M, Kasza A, Bugno M, Kordula T. Parthenolideinhibits activation of signal transducers and activators of transcription(STATs) induced by cytokines of the IL-6 family. Biochem Biophys ResCommun 2000;267:329–33.
14. Won YK, Ong CN, Shi X, Shen HM. Chemopreventive activity ofparthenolide against UVB-induced skin cancer and its mechanisms.Carcinogenesis 2004;25:1449–58.
15. Nakshatri H, Rice SE, Bhat-Nakshatri P. Antitumor agent parthenolidereverses resistance of breast cancer cells to tumor necrosis factor-relatedapoptosis-inducing ligand through sustained activation of c-Jun N-terminalkinase. Oncogene 2004;23:7330–44.
16. Zhang S, Lin ZN, Yang CF, Shi X, Ong CN, Shen HM. Suppressed NF-nB and sustained JNK activation contribute to the sensitization effect ofparthenolide to TNF-a-induced apoptosis in human cancer cells. Carcino-genesis 2004;25:2191–9.
17. deGraffenried LA, Chandrasekar B, Friedrichs WE, et al. NF-nBinhibition markedly enhances sensitivity of resistant breast cancer tumorcells to tamoxifen. Ann Oncol 2004;15:885–90.
18. Wen J, You KR, Lee SY, Song CH, Kim DG. Oxidative stress-mediatedapoptosis. The anticancer effect of the sesquiterpene lactone partheno-lide. J Biol Chem 2002;277:38954–64.
19. Pozarowski P, Halicka DH, Darzynkiewicz Z. NF-nB inhibitor sesqui-terpene parthenolide induces concurrently atypical apoptosis and cellnecrosis: difficulties in identification of dead cells in such cultures.Cytometry 2003;54A:118–24.
20. Bhat-Nakshatri P, Campbell RA, Patel NM, et al. Tumour necrosisfactor and PI3kinase control oestrogen receptor a protein level and itstransrepression function. Br J Cancer 2004;90:853–9.
21. Kumar S, Kishimoto H, Chua HL, et al. Interleukin-1 a promotes tumorgrowth and cachexia in MCF-7 xenograft model of breast cancer. Am JPathol 2003;163:2531–41.
22. Muller A, Homey B, Soto H, et al. Involvement of chemokine receptorsin breast cancer metastasis. Nature 2001;410:50–6.
23. Kim R, Ohi Y, Inoue H, Toge T. Enhancement of chemotherapeuticagents induced-apoptosis associated with activation of c-Jun N-terminalkinase 1 and caspase 3 (CPP32) in bax-transfected gastric cancer cells.Anticancer Res 2000;20:439–44.
24. Wang Q, Wieder R. All-trans retinoic acid potentiates Taxotere-induced cell death mediated by Jun N-terminal kinase in breast cancercells. Oncogene 2004;23:426–33.
25. Zhang H, Morisaki T, Nakahara C, et al. PSK-mediated NF-nBinhibition augments docetaxel-induced apoptosis in human pancreaticcancer cells NOR-P1. Oncogene 2003;22:2088–96.
26. Huber MA, Azoitei N, Baumann B, et al. NF-nB is essential forepithelial-mesenchymal transition and metastasis in a model of breastcancer progression. J Clin Invest 2004;114:569–81.
27. Baeuerle PA, Henkel T. Function and activation of NF-nB in theimmune system. Annu Rev Immunol 1994;12:141–79.
28. Sica A, Dorman L, Viggiano V, et al. Interaction of NF-nB and NFATwith the interferon-g promoter. J Biol Chem 1997;272:30412–20.
29. Bendre MS, Gaddy-Kurten D, Mon-Foote T, et al. Expression ofInterleukin 8 and not Parathyroid Hormone-related Protein by HumanBreast Cancer Cells Correlates with Bone Metastasis In vivo. Cancer Res2002;62:5571–9.
30. Kang Y, Siegel PM, Shu W, et al. A multigenic program mediatingbreast cancer metastasis to bone. Cancer Cell 2003;3:537–49.
31. De Larco JE, Wuertz BR, Manivel JC, Furcht LT. Progression andenhancement of metastatic potential after exposure of tumor cells tochemotherapeutic agents. Cancer Res 2001;61:2857–61.
32. Kim J, Yu W, Kovalski K, Ossowski L. Requirement for specificproteases in cancer cell intravasation as revealed by a novel semiquanti-tative PCR-based assay. Cell 1998;94:353–62.
33. Helbig G, Christopherson KW II, Bhat-Nakshatri P, et al. NF-nBpromotes breast cancer cell migration and metastasis by inducing theexpression of the chemokine receptor CXCR4. J Biol Chem 2003;278:21631–8.
34. Ventura JJ, Kennedy NJ, Flavell RA, Davis RJ. JNK regulatesautocrine expression of TGF-h1. Mol Cell 2004;15:269–78.
35. De Smaele E, Zazzeroni F, Papa S, et al. Induction of gadd45h by NF-nBdownregulates pro-apoptotic JNK signalling. Nature 2001;414:308–13.
36. Karin M, Cao Y, Greten FR, Li ZW. NF-nB in cancer: from innocentbystander to major culprit. Nat Rev Cancer 2002;2:301–10.
37. Nakshatri H, Goulet RJ Jr. NF-nB and breast cancer. Curr Probl Cancer2002;26:282–309.
38. Greten FR, Eckmann L, Greten TF, et al. IKKh links inflammation andtumorigenesis in a mouse model of colitis-associated cancer. Cell2004;118:285–96.
39. Loercher A, Lee TL, Ricker JL, et al. Nuclear factor-nB is an importantmodulator of the altered gene expression profile and malignant phenotypein squamous cell carcinoma. Cancer Res 2004;64:6511–23.
40. Pikarsky E, Porat RM, Stein I, et al. NF-nB functions as a tumourpromoter in inflammation-associated cancer. Nature 2004;431:461–6.
41. Lollini PL, Bosco MC, Cavallo F, et al. Inhibition of tumor growth andenhancement of metastasis after transfection of the g-interferon gene. IntJ Cancer 1993;55:320–9.
42. Sheen-Chen SM, Chen WJ, Eng HL, Chou FF. Serum concentration oftumor necrosis factor in patients with breast cancer. Breast Cancer ResTreat 1997;43:211–5.
43. Leek RD, Landers R, Fox SB, Ng F, Harris AL, Lewis CE. Association oftumour necrosis factor a and its receptors with thymidine phosphorylaseexpression in invasive breast carcinoma. Br J Cancer 1998;77:2246–51.
44. Zhang S, Ong CN, Shen HM. Involvement of proapoptotic Bcl-2 familymembers in parthenolide-induced mitochondrial dysfunction and apopto-sis. Cancer Lett 2004;211:175–88.
45. Balkwill F. Cancer and the chemokine network. Nat Rev Cancer 2004;4:540–50.
46. Abdollahi T, Robertson NM, Abdollahi A, Litwack G. Identification ofinterleukin 8 as an inhibitor of tumor necrosis factor-related apoptosis-inducing ligand-induced apoptosis in the ovarian carcinoma cell lineOVCAR3. Cancer Res 2003;63:4521–6.
47. Teicher BA. Malignant cells, directors of the malignant process: role oftransforming growth factor-h. Cancer Metastasis Rev 2001;20:133–43.
48. Nowak AK, Wilcken NR, Stockler MR, Hamilton A, Ghersi D.Systematic review of taxane-containing versus non-taxane-containingregimens for adjuvant and neoadjuvant treatment of early breast cancer.Lancet Oncol 2004;5:372–80.
49. Yardley DA. Gemcitabine and docetaxel in metastatic and neo-adjuvant treatment of breast cancer. Semin Oncol 2004;31:37–44.
Chemosensitizing Activity of Parthenolide1012
Mol Cancer Ther 2005;4(6). June 2005
Research. on November 22, 2018. © 2005 American Association for Cancermct.aacrjournals.org Downloaded from

2005;4:1004-1012. Mol Cancer Ther Christopher J. Sweeney, Sanjana Mehrotra, Miral R. Sadaria, et al. xenograft model of breast cancerdocetaxel reduces metastasis and improves survival in a The sesquiterpene lactone parthenolide in combination with
Updated version
http://mct.aacrjournals.org/content/4/6/1004
Access the most recent version of this article at:
Cited articles
http://mct.aacrjournals.org/content/4/6/1004.full#ref-list-1
This article cites 47 articles, 8 of which you can access for free at:
Citing articles
http://mct.aacrjournals.org/content/4/6/1004.full#related-urls
This article has been cited by 12 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://mct.aacrjournals.org/content/4/6/1004To request permission to re-use all or part of this article, use this link
Research. on November 22, 2018. © 2005 American Association for Cancermct.aacrjournals.org Downloaded from





























