Embed Size (px)
Citation preview

The Transforming Growth Factor-b SuperfamilyCytokine Macrophage Inhibitory Cytokine-1 Is Presentin High Concentrations in the Serum ofPregnant Women*
A. G. MOORE†, D. A. BROWN†, W. D. FAIRLIE, A. R. BAUSKIN, P. K. BROWN,M. L. C. MUNIER, P. K. RUSSELL, L. A. SALAMONSEN, E. M. WALLACE, AND
S. N. BREIT
Centre for Immunology (A.G.M., D.A.B., W.D.F., A.R.B., P.K.B., M.L.C.M., P.K.R., S.N.B.), St.Vincent’s Hospital and University of New South Wales, Sydney, New South Wales, Australia;Department of Obstetrics and Gynecology (E.M.W.), Monash University, Clayton, Victoria, Australia;and Prince Henry’s Institute of Medical Research (L.A.S.), Clayton, Victoria, Australia
ABSTRACTMacrophage inhibitory cytokine-1 (MIC-1) is a recently described
divergent member of the transforming growth factor-b superfamily.MIC-1 transcription up-regulation is associated with macrophageactivation, and this observation led to its cloning. Northern blotsindicate that MIC-1 is also present in human placenta. A sensitivesandwich enzyme-linked immunosorbent assay for the quantificationof MIC-1 was developed and used to examine the role of this cytokinein pregnancy. High levels of MIC-1 are present in the sera of pregnantwomen. The level rises substantially with progress of gestation.
MIC-1 can also be detected, in large amounts, in amniotic fluid andplacental extracts. In addition, the BeWo placental trophoblastic cellline was found to constitutively express the MIC-1 transcript andsecrete large amounts of MIC-1. These findings suggest that theplacental trophoblast is a major source of the MIC-1 present in ma-ternal serum and amniotic fluid. We suggest that MIC-1 may promotefetal survival by suppressing the production of maternally derivedproinflammatory cytokines within the uterus. (J Clin EndocrinolMetab 85: 4781–4788, 2000)
THE TRANSFORMING GROWTH factor-b (TGF-b) su-perfamily consists of an increasing number of mole-
cules that regulate a variety of cellular processes such asgrowth, differentiation, and oncogenesis. Members of theTGF-b superfamily have been classified into major familygroupings, which include TGF-b, bone morphogenetic pro-tein, growth and differentiation factor, inhibin/activin, mul-lerian inhibitory substance, glial-derived neurotrophic factor(GDNF), and (more recently) macrophage inhibitory cyto-kine-1 (MIC-1) (1–3). Involvement of the TGF-b superfamilyin human pregnancy is suggested by the presence of manyof the TGF-b superfamily members (TGF-b1, TGF-b2, TGF-b3, activin, and inhibin) in amniotic fluid. Additionally, TGF-b1, activin, and inhibin have been localized to the placentalvilli (4–12).
Cytokines in the TGF-b superfamily are thought to havemultiple functions during pregnancy. The ability of the
TGF-b isoforms to modulate cell-cell adhesion, cell migra-tion, and tissue remodeling has led some authors to suggestthat this molecule may control trophoblast invasion and im-plantation early in pregnancy. Other possible roles includeregulation of fetal growth and suppression of the maternalimmune responses.
Placental cells are a major source of TGF-b superfamilycytokines and are regulated by them. For example, activinsuppresses the production of inhibin and enhances secretionof progesterone, human CG (hCG), and GnRH by placentalcells (13). Inhibin suppresses placental hCG, GnRH, and ac-tivin-induced progesterone release (13), whereas TGF-b1suppresses human placental lactogen (PL) production. Ac-tivin and TGF-b3 have also been shown to have opposingeffects in regulating extravillous trophoblast invasion inearly pregnancy (11, 14). These findings suggest that TGF-b1,TGF-b3, activin, and inhibin regulate the growth and differ-entiation of the placenta in an autocrine manner. TGF-b1,activin, and inhibin are also present in the embryo proper,where they have been demonstrated to regulate growth anddifferentiation. In particular, TGF-b superfamily membersare well known for their ability to promote mesoderminduction.
TGF-b superfamily proteins may promote fetal survival.Experimental evidence suggests that the amniotic fluid con-centration of the proinflammatory cytokines interleukin(IL)-1, IL-6, and tumor necrosis factor rise during labor. Fur-thermore, proinflammatory cytokine production accompa-
Received December 9, 1999. Revision received May 12, 2000. Rerevi-sion received August 11, 2000. Accepted August 11, 2000.
Address all correspondence and requests for reprints to: Dr. SamuelN. Breit, Centre for Immunology, St. Vincent’s Hospital, Victoria Street,Sydney, New South Wales 2010, Australia. E-mail: [email protected].
* This work has been funded in part by grants from St. Vincent’sHospital and by Meriton Apartments Pty Ltd. through an R&D syndi-cate arranged by Macquarie Bank Limited. In addition, this project waspartially funded by a New South Wales Health Research and Devel-opment infrastructure grant.
† Contributed equally to this work.
0021-972X/00/$03.00/0 Vol. 85, No. 12The Journal of Clinical Endocrinology & Metabolism Printed in U.S.A.Copyright © 2000 by The Endocrine Society
4781
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 10 August 2014. at 21:31 For personal use only. No other uses without permission. . All rights reserved.

nying intrauterine infection has been associated with fetalrejection or preterm labor (15–17). TGF-b1 and inhibin havebeen shown to suppress production of proinflammatory cy-tokines by macrophages and lymphocytes, respectively (18,19), whereas activin has proinflammatory effects on macro-phages and the amnion (20, 21). This has led to the suggestionthat TGF-b1 and inhibin promote fetal survival by suppress-ing the production of potentially harmful proinflammatorycytokines by the maternal immune system.
We have recently cloned and characterized a divergentmember of the TGF-b superfamily, MIC-1 (1). Its expressionis associated with macrophage activation. Like several othermembers of this superfamily, MIC-1 messenger RNA(mRNA) was identified as being highly expressed in placenta(22–24) and may also be involved in pregnancy. To betterunderstand the role of MIC-1 in pregnancy, we have devel-oped a sensitive sandwich enzyme-linked immunosorbentassay (ELISA) for its quantitation. Using this assay, we in-vestigated the temporal relationship between maternal se-rum MIC-1 concentrations and gestational age. We also mea-sured its concentration in amniotic fluid and placentalextracts. To further delineate the origins of this cytokine, wehave also assessed the capacity of a placental trophoblasticcell line (BeWo) to synthesize MIC-1.
Materials and MethodsSerum and amniotic fluid samples
Serum samples were obtained from 22 healthy pregnant women withnormal singleton pregnancies and on no medications. Approval fromThe Human Research and Ethics Committee of Monash Medical Centerwas obtained for all human sample collection. In each case, gestationalage was determined by an early pregnancy ultrasound scan. All womensubsequently had a normal vaginal delivery, at term (37–41 weeks), ofa healthy normally grown infant. Serum samples were collected from 6women between 10–14 weeks of pregnancy, and 8 women between26–30 weeks and 37–40 weeks of pregnancy. The time periods indicatedcorrespond to the end of each trimester. Samples corresponding to eachtrimester were pooled before measurement of MIC-1 levels. Serial ma-ternal serum samples were also taken, on a weekly basis, from another4 women, from 30 weeks of gestation to delivery. Again, all 4 womenwere healthy, with a normal singleton pregnancy, and had a normalvaginal delivery (at term) of a normal healthy infant. In addition, am-niotic fluid was obtained from 10 women undergoing amniocentesis, at15–17 weeks of gestation, for fetal karyotyping. In all cases, the indi-cation for karyotyping was advanced maternal age (.37 yr). Amnioticfluid samples were also pooled before measurement of MIC-1 levels.Normal nonpregnant serum samples were obtained from 6 healthywomen with a regular menstrual cycle and not on hormonal contra-ception. These women were in the luteal phase of their cycle whensampled.
Placental extracts
Between 100–150 mg placental tissue, obtained from normal termdeliveries, (rinsed 4–5 times in saline solution and frozen in liquidnitrogen and stored at 280 C) was homogenized in 1 mL PBS. Homog-enates were centrifuged at 10,000 rpm for 30 sec, and the supernatantwas transferred to tubes. Total protein was measured by the BCA totalprotein assay (Pierce Chemical Co.), following the manufacturer’s in-structions. BSA solutions, ranging between 0–1000 mg/mL, were usedas standard solutions.
BeWo cell culture
The human choriocarcinoma trophoblastic cell line (BeWo) was pur-chased from ATCC (Rockville, MD). Cells were seeded into 96-well
tissue-culture plates, at 5000 cells per well, in 250 mL DMEM (containing4.5 g/L d-glucose, 110 mg/L sodium pyruvate, 0.584 g/L l-glutamine,4 mg/L pyridoxine hydrochloride) (Life Technologies, Inc.) and 13Nutridoma-SR (Roche Molecular Biochemicals, Germany) and culturedat 37 C in the presence of 5% carbon dioxide for 1–5 days. The cultureplates were then spun at 1000 rpm for 10 min, and the supernatant wasremoved and stored at 220 C until quantification of MIC-1.
RT-PCR analysis of MIC-1 mRNA synthesis
Total RNA was isolated from BeWo cell monolayers, in 96-well plates,using Tri-Pure Reagent (Roche Molecular Biochemicals), method asprovided by the manufacturer. RT was carried out in a total reaction volof 20 mL using 1 mg RNA, a poly(T)15 primer, and 50 U of Expand ReverseTranscriptase (Roche Molecular Biochemicals) using the manufacturer’srecommended conditions. A 5-mL aliquot of the RT reaction was am-plified in a PCR reaction using Pfu polymerase (Promega Corp.) andprimers MSB-1 (59-AGGACCTGCTAACCAGGCTGCGGGCCAACC-AGAGC-39) and MSB-5 (59-GGCTAACAAGTCATCATAGGTCTG-GAGCGACAC-39), which flank the single intron of MIC-1. PCR con-ditions were as follows: an initial denaturation step at 95 C for 1 min,followed by 35 cycles at 95 C for 30 sec, 60 C for 30 sec, and 72 C for 2min. An RT reaction in which the RNA was omitted was used as anegative control, and a plasmid carrying the MIC-1 pre-pro-MIC/FLAGcoding sequence (1) was included as a positive control. PCR productswere separated on 0.8% (wt/vol) agarose gels.
Recombinant human (rh)MIC-1
Recombinant MIC-1 was expressed in the yeast Pichia pastoris andpurified to homogeneity, using a multistep procedure, culminating inreverse-phase high-performance liquid chromatography (25).
Generation of MIC-1 antibodies
The sheep anti-MIC-1 polyclonal antibody (PAb) 233B3 was gener-ated by immunization with rhMIC-1 in Complete Freund’s Adjuvant.Additional boosts were given over a period of 6 months, and the sheepwere bled 10 days after the final injection. An enriched IgG fraction ofnormal sheep serum and 233B3 were prepared by caprylic acid precip-itation, followed by ammonium sulfate precipitation. The IgG-enriched233B3 fraction was designated 233B3-P.
The mouse anti-MIC-1 monoclonal antibody (MAb) 13C4H3-secret-ing hybridoma was generated from mice immunized with rhMIC-1.Hybridomas were cultured in DMEM (Life Technologies, Inc.) contain-ing 4.5 g/L d-glucose, 110 mg/L sodium pyruvate, 0.584 g/L l-glutamine, and 4 mg/L pyridoxine hydrochloride supplemented with20% FCS (CSL Melbourne). For MAb collection, the hybridomas weretransferred into fresh DMEM-hi glucose, supplemented with Nutri-doma-SR (Roche Molecular Biochemicals), for 7 days. The culture su-pernatants were spun at 2000 rpm for 10 min, to remove cell debris, andfrozen until used. The sensitivity of the PAb and MAb preparations wereexamined by direct ELISA.
Direct ELISA
Ninety-six-well Maxisorp ELISA plates (Nunc) were coated (100 mL/well) with either 18 ng/mL rhMIC-1 or 20 ng/mL rhTGF-b1 (R&DSystems) in coating buffer (0.1 mol/L carbonate in distilled H20, pH9.4–9.8) at 4 C for 24 h. Plates were washed three times with 300 mL/wellwash buffer [PBS containing 0.05% (vol/vol) Tween-20 (Sigma)]. Non-specific binding was blocked with addition of 250 mL/well of 1% (wt/vol) BSA (Roche Molecular Biochemicals), in PBS for 2 h at 37 C.Hybridoma-conditioned, serum-free media, containing the MAb13C4H3; sheep PAb 233B3-P diluted 1:500,000 in antibody diluent (Abdil) [PBS containing 1% (wt/vol) BSA and 0.05% (vol/vol) Tween-20];culture media conditioned by the mouse myeloma cell line SP2/0;DMEM1Nutridoma; IgG-enriched normal sheep serum diluted1:500,000 in Ab dil; 200 ng/mL mouse IgG1 (R&D Systems) inDMEM1Nutridoma, or Ab dil alone were then added to the plates (100mL/well) and incubated for 1 h at 37 C. The plates were washed threetimes, followed by the addition of 100 mL/well biotinylated donkeyantisheep IgG (Jackson ImmunoResearch Laboratories, Inc.) or biotin-
4782 MOORE ET AL. JCE & M • 2000Vol. 85 • No. 12
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 10 August 2014. at 21:31 For personal use only. No other uses without permission. . All rights reserved.
ylated goat antimouse IgG (Jackson ImmunoResearch Laboratories, Inc.)diluted 1:10,000 in Ab dil and incubated for 1 h at 37 C. The plates werewashed three times, and 100 mL/well of horseradish peroxidase-conjugated strepavidin (Genzyme), diluted 1:2000 in Ab dil, was addedto the plates and incubated for 30 min at 37 C. Plates were washed fourtimes, followed by the addition of 100 mL/well of peroxidase substrate(1 mg/mL o-phenylenediamine dihydrochloride (Sigma) in 0.05 mol/Lphosphate-citrate buffer containing 0.014% H2O2, pH5.0 (Sigma). Colordevelopment was allowed to proceed for 5–15 min and was terminatedby the addition of 100 mL/well of 4N H2SO4. The absorbance wasmeasured at 490 nm in a microplate reader (Pasteur Diagnostics).
MIC-1 sandwich ELISA
A MIC-1 sandwich ELISA was established using the mouse MAb13C4H3 for antigen capture; and the sheep PAb 233B3-P, for detection.The optimum concentration of both antibodies was determined, thenused for all subsequent studies. Ninety-six-well Maxisorp ELISA plateswere coated with MAb 13C4H3 supernatant diluted 1:5 (final Ig con-centration was approximately 20 ng/mL) in coating buffer at 4 C for 24 h.ELISA plates were then washed three times with 300 mL/well of washbuffer. Nonspecific binding was blocked with 250 mL/well of 1% (wt/vol) BSA in PBS for 2 h at 37 C. rhMIC-1 standards, tissue culturesupernatant, maternal serum, placental extracts, or amniotic fluid in Abdil were then added to the plates (100 mL/well) and incubated for 1 hat 37 C. The plates were washed three times, followed by the additionof 100 mL/well of the sheep PAb 233B3-P diluted 1:5000 in Ab dil andincubated for 1 h at 37 C. ELISA plates were then washed three times,and 100 mL/well of biotinylated donkey antisheep IgG diluted to 1:5000
in Ab dil was added and incubated for 1 h at 37 C. The plates were thendeveloped, as for the direct ELISA. The concentration of hMIC-1 in thesamples was determined by comparison with the rhMIC-1 standardcurve. The standard curve was constructed using standard curve-fittingsoftware supplied with the microplate reader (Pasteur Diagnostics). Thelevel of rhMIC-1 in the standard curve was determined on the basis ofa comparison of this standard to a master standard of highly purifiedrecombinant MIC-1. The master standard protein concentration wasdetermined by an average of eight estimations of total amino acid com-position. All samples were assayed in triplicate on at least two occasions.Results are presented as the mean 6 sd.
Immunohistochemistry
Term placenta was fixed overnight in 10% formaldehyde (vol/vol) inPBS and paraffin embedded. Four-micrometer sections were placed onsialine-coated slides. Sections were brought to water and microwavedfor 20 min in 0.1 mol/L citrate (Univar) buffer, pH 6.0. Nonspecificbinding was blocked using 4% BSA (wt/vol) in PBS for 2 h at 21 C. Slideswere incubated with the sheep PAb 233B3 or IgG-enriched normal sheepserum, diluted 1:5000 in 4% BSA (wt/vol) in PBS, overnight at 4 C. Toblock nonspecific peroxidase activity, sections were incubated in 0.03%peroxide (Analar) (vol/vol) in methanol (Analar) for 30 min at 21 C.Biotinylated donkey antisheep IgG (Jackson ImmunoResearch Labora-tories, Inc.) diluted to 1:200 in 4% BSA (wt/vol) in PBS and was incu-bated with slides for 45 min at 21 C. Streptavidin-HRP (DAKO Corp.,large volume kit) was incubated with samples for 10 min at 21 C, andthe slides were developed using DAB (Pierce Chemical Co.), 1:10 (vol/vol), in stable peroxide buffer (Pierce Chemical Co.). A light counterstain
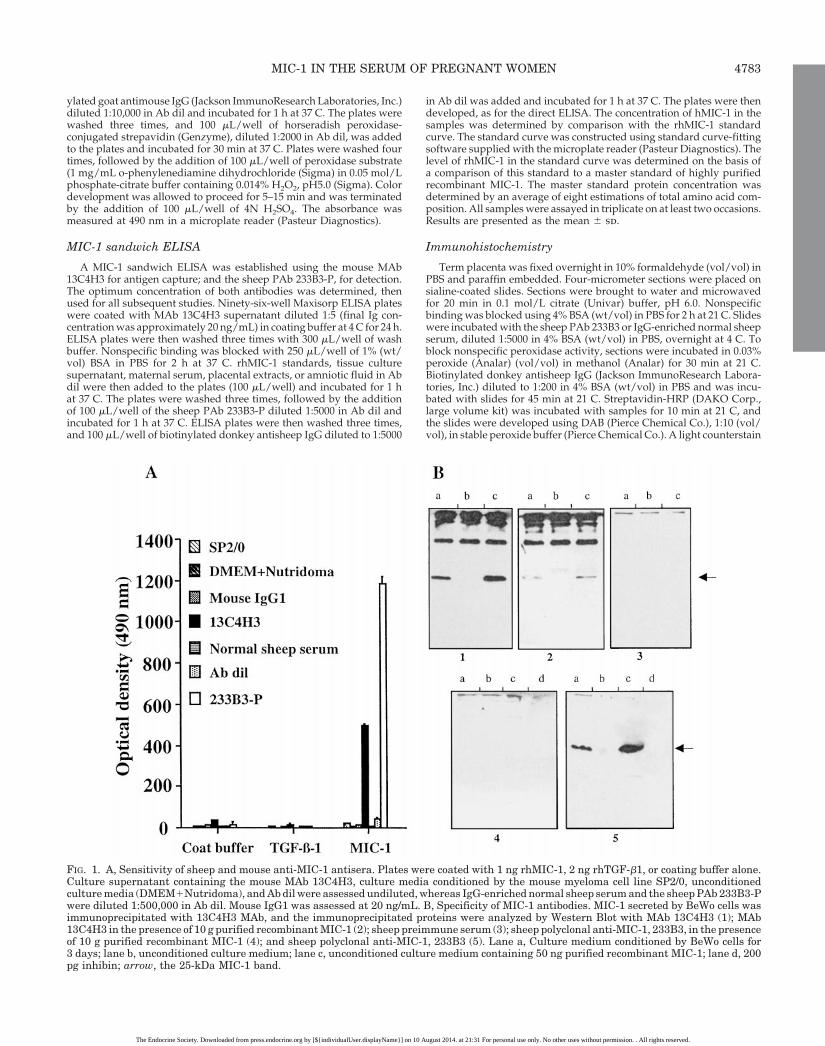
FIG. 1. A, Sensitivity of sheep and mouse anti-MIC-1 antisera. Plates were coated with 1 ng rhMIC-1, 2 ng rhTGF-b1, or coating buffer alone.Culture supernatant containing the mouse MAb 13C4H3, culture media conditioned by the mouse myeloma cell line SP2/0, unconditionedculture media (DMEM1Nutridoma), and Ab dil were assessed undiluted, whereas IgG-enriched normal sheep serum and the sheep PAb 233B3-Pwere diluted 1:500,000 in Ab dil. Mouse IgG1 was assessed at 20 ng/mL. B, Specificity of MIC-1 antibodies. MIC-1 secreted by BeWo cells wasimmunoprecipitated with 13C4H3 MAb, and the immunoprecipitated proteins were analyzed by Western Blot with MAb 13C4H3 (1); MAb13C4H3 in the presence of 10 g purified recombinant MIC-1 (2); sheep preimmune serum (3); sheep polyclonal anti-MIC-1, 233B3, in the presenceof 10 g purified recombinant MIC-1 (4); and sheep polyclonal anti-MIC-1, 233B3 (5). Lane a, Culture medium conditioned by BeWo cells for3 days; lane b, unconditioned culture medium; lane c, unconditioned culture medium containing 50 ng purified recombinant MIC-1; lane d, 200pg inhibin; arrow, the 25-kDa MIC-1 band.
MIC-1 IN THE SERUM OF PREGNANT WOMEN 4783
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 10 August 2014. at 21:31 For personal use only. No other uses without permission. . All rights reserved.

was performed by incubating the slides in Mayer’s hematoxylin solution(Sigma) for 2 min, followed by Scott’s bluing solution for 30 sec. Threewashes with wash buffer [Tris-phosphate 0.1 molfl (pH 7.6) buffer 0.1%Tween-20] were performed between all steps, after the primary antibodyincubation, until counter staining, where tap water was used.
Immunoprecipitation
MAb 13C4H3 was adsorbed to protein-A Sepharose beads. Serumand medium samples (1 mL) were immunoprecipitated by incubatingwith beads overnight at 4 C and then washing 5 times with PBS con-taining 1% (vol/vol) Triton X-100. Bound proteins were eluted, usingnonreducing SDS sample buffer, and analyzed by SDS-PAGE (26), fol-lowed by immunoblot analysis (1) with the sheep PAb 233B3 or MAb13C4H3, in the presence or absence of 10 g purified recombinant MIC-1,or sheep preimmune serum. The 233B3 and sheep preimmune serummix was used at a dilution of 1:14,000. Donkey antisheep IgG-biotin, ata dilution of 1:2,000 was used as the secondary antibody. Immunoblotanalysis with Mab 13C4H3 was undertaken using serum free hybridomasupernatant, diluted 1:1 with PBS containing 0.1% Tween 20. In this case,goat antimouse IgG-biotin (at a dilution of 1:2,000) was used as thesecondary antibody.
ResultsSensitivity and specificity of anti-MIC-1 PAb and MAb
The ability of the sheep PAb 233B3-P and the mouse MAb13C4H3 to bind to rhMIC-1 was examined by direct ELISA.It was found that both undiluted tissue culture supernatant(containing the MAb 13C4H3) and the sheep PAb 233B3-P (ata dilution of 1:500,000 in Ab dil) bound strongly to 1.8 ngimmobilized rhMIC-1 (Fig. 1A). No reaction was observedbetween rhMIC-1 and culture media conditioned by themouse myeloma cell line SP2/0, unconditioned culture me-dia, mouse IgG1, Ig-enriched normal sheep serum, or Ab dil.Minimal background binding to uncoated wells was ob-served for all samples examined. No reactivity was detectedwhen either 13C4H3 or 233B3-P were incubated with im-mobilized rhTGF-b1.
Specificity of the antibodies was determined by immuno-precipitation of purified recombinant MIC-1 with MAb13C4H3, followed by immunoblot analysis with variousMIC-1 specific antibodies (Fig. 1B). All the MIC-1 antibodiesspecifically recognized the 25-kD dimeric MIC-1. Addition-ally, blocking of the antibodies was performed by preincu-bating the antibody with purified recombinant MIC-1, beforeWestern Blot analysis. This greatly reduced the interaction ofthe antibody with the MIC-1 specific 25-kD band, confirmingspecificity of the antibodies Mab 13C4H3 (Fig. 1B2) and233B3-P (Fig. 1B4). Furthermore, those antibodies testedfailed to recognize inhibin (Fig. 1B, 4d and 5d), anothermember of the TGF-b superfamily, elevated in pregnancy.
MIC-1 sandwich ELISA
A sandwich ELISA, employing MAb 13C4H3 and the PAb233B3-P, was established. This assay could accurately quan-tify rhMIC-1 in the range of about 20–900 pg/mL (Fig. 2).Above this range, there was saturation demonstrated withthe assay (Fig. 3). To examine the effect of factors present inhuman serum and culture media on estimation of this cy-tokine, 900 pg/mL rhMIC-1 was added to Ab dil containing
FIG. 2. A, rhMIC-1 standard curve generated by sandwich ELISA using the mouse MAb 13C4H3 for capture and the sheep PAb 233B3-P fordetection. Diluent used was sample buffer, DMEM, DMEM 1 10% FCS. Curve fitting using polynomial transformation. B, Parallelism of MIC-1sandwich ELISA, measuring serial dilutions of serum and culture supernatant samples in culture medium and sample diluent.
FIG. 3. Saturation curve generated for MIC-1 by sandwich ELISAusing the mouse MAb 13C4H3 for capture and the sheep PAb 233B3-Pfor detection.
4784 MOORE ET AL. JCE & M • 2000Vol. 85 • No. 12
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 10 August 2014. at 21:31 For personal use only. No other uses without permission. . All rights reserved.

either 10% (vol/vol) normal human serum or 10% (vol/vol)DMEM1FBS, and then quantified. It was found that thesandwich ELISA was accurate to within 5% of the correctvalue. Run-to-run variation was less than 5%. TGF-b1 andinhibin, assayed in the sandwich ELISA at a concentration of500 pg/mL each, yielded optical density readings no differ-ent from negative controls in the sandwich ELISA system,indicating the total absence of cross-reactivity. Parallelism ofthe sandwich ELISA was confirmed with serial dilutions ofculture and serum samples diluted in culture medium andAb dil (Fig. 2B).
MIC-1 levels in staged pregnancy sera increase
Pooled serum samples were diluted between 1:5 and 1:20in Ab dil before MIC-1 quantitation by sandwich ELISA. Itwas determined that pooled normal human sera containedapproximately 0.36 (6 0.04) ng/mL MIC-1 (Fig. 4A). Inpooled maternal serum, the MIC-1 concentration was foundto increase dramatically during pregnancy. Maternal serumsamples, corresponding to the first trimester, containedapproximately 6.3 (6 0.02) ng/mL MIC-1, which rose to 12.2(6 0.5) ng/mL during the second trimester and peaked at15.3 (6 1.3) ng/mL during the third trimester. The rise inserum MIC-1 during pregnancy was found to be statisticallysignificant (P , 0.001), using the Student’s t test.
Immunoprecipitation was used to confirm the presence ofMIC-1 in pooled maternal serum samples during pregnancy.MIC-1 was visualized by immunoprecipitation with MAb13C4H3, followed by immunoblot analysis with PAb233B3-P. A band corresponding to the disulphide-linked ma-ture MIC-1 peptide (approximately 25 kDa) can be observedin the second and third trimester serum samples (Fig. 4B,lanes 3–4). The highest level of MIC-1 was found in the thirdtrimester sample. No similar band was observed in normalserum or the sample corresponding to the first trimester,presumably because of the lower sensitivity of immunoblotanalysis (Fig. 4B, lanes 1–2).
Maternal serum MIC-1 concentrations were also examinedin serial samples from four pregnant women. At 30 weeks ofgestation, serum from all four women examined containedapproximately 3.6 ng/mL MIC-1 (Fig. 5). Maternal serumMIC-1 levels were found to increase from 30 weeks of ges-tation until birth. Subjects designated MH and JB exhibiteda slight decrease in MIC-1 maternal serum levels over the lastweek of pregnancy.
MIC-1 can be detected in amniotic fluid
In additional to maternal serum, amniotic fluid collectedfrom 10 women during the second trimester for karyotypingpurposes was pooled before quantification of MIC-1 levelsby sandwich ELISA. It was determined that the pooled am-niotic fluid sample contained approximately 13.68 (6 0.16)ng/mL MIC-1 (Fig. 4A). Immunoprecipitation and Westernblot analysis of pooled amniotic fluid revealed a band ofapproximately 25 kDa, which corresponds to the disulphide-linked mature MIC-1 peptide (Fig. 4B, lane 5).
Placental MIC-1 is detected by immunohistochemistry
Term placental immunohistochemistry reveals that MIC-1is produced predominantly by the trophoblastic cells. Al-though there is production by stromal cells, the result shown(Fig. 6.) indicates that the trophoblast cells are the majorproducers of MIC-1 in the placenta.
FIG. 4. MIC-1 is present in maternalserum and amniotic fluid during preg-nancy in women. A, Estimation ofMIC-1 concentrations in pooled normalhuman serum, pooled staged maternalserum, and pooled amniotic fluid (AF),as determined by sandwich ELISA; B,immunoprecipitation and Western blotanalysis of MIC-1 in pooled normal hu-man serum (lane 1), pooled staged ma-ternal serum (lane 2–4), and pooled am-niotic fluid (lane 5).
FIG. 5. Maternal serum MIC-1 concentrations determined serially infour pregnant women (IT, PT, JB, MH) from 30 weeks of gestationuntil birth.
MIC-1 IN THE SERUM OF PREGNANT WOMEN 4785
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 10 August 2014. at 21:31 For personal use only. No other uses without permission. . All rights reserved.

MIC-1 can be detected in human placental extracts
Whole-tissue Northern blot analysis indicates that MIC-1transcripts are expressed abundantly in the placenta (22–24),suggesting that the placenta is a major source of circulatingMIC-1 in the serum of pregnant women. To test further thepresence of MIC-1 in placental material, we have examinedfive human placental extracts, for the presence of MIC-1, bysandwich ELISA. All five samples were found to be positivefor MIC-1 (Fig. 7), ranging in concentration from 5–54ng/mL.
Cultured BeWo cells constitutively express MIC-1 RNA andsecrete mature MIC-1
Because high levels of MIC-1 were detected in placentalextracts and in placental immunohistochemistry, it seemedlikely that the placental trophoblastic cell line, BeWo, mayalso produces this cytokine. To confirm this, we examinedtissue culture media, conditioned by BeWo cells under rest-ing conditions, for the presence of secreted MIC-1, by sand-wich ELISA. BeWo-conditioned media (24 h) contained ap-proximately 21.6 (62.95) ng/mL MIC-1 (Fig. 8A). Theconcentration of MIC-1 in the culture media, after a 5-dayincubation, increased to approximately 117 (67.2) ng/mL.The ability of unstimulated BeWo cells to secrete MIC-1 wasalso examined by immunoprecipitation and Western blotanalysis. High levels of secreted mature MIC-1, as indicatedby a band at approximately 25 kDa, were observed in mediaconditioned by BeWo cells for 5 days (Fig. 8B). Additionalbands, migrating at 55-kDa and 12.5-kDa bands, were ob-served, which most likely represent incompletely processedMIC-1 hemidimer and monomer, respectively. Culture me-dia, which had not been exposed to BeWo cells, contained nodetectable MIC-1 when examined by sandwich ELISA or byimmunoprecipitation.
To confirm that the 25-kDa band present in Fig. 8B was infact MIC-1, the detection antibodies 233B3 and Mab 13C4H4were preincubated with purified recombinant MIC-1, as pre-viously described, and compared with nonabsorbed detec-tion antibodies. This resulted in markedly diminished stain-ing (Fig. 1B: 1a, 2a, 4a, and 5a), confirming that the band doesrepresent MIC-1.
RT-PCR was used to confirm the presence of the MIC-1transcript in unstimulated BeWo cells. Total RNA was ex-tracted from BeWo cells cultured for 24 h and subjected toRT-PCR as described. A single product of 0.4 kbp was ob-served, indicating that the MIC-1 transcript was present inBeWo cells (Fig. 8C). No product was detected in the absenceof BeWo or plasmid DNA.
Discussion
Here we describe, for the first time, a sensitive sandwichELISA for the quantitation of the recently described TGF-bsuperfamily member, MIC-1. The evidence presented in thisreport indicates that MIC-1 is present in large amounts in theserum of pregnant women, and in increasing concentrationsas the pregnancy progresses. Similar observations have beenmade in the case of activin and inhibin concentrations duringpregnancy in humans (9, 21, 27).
Although elevated levels of MIC-1 in maternal serum donot necessarily indicate fetal exposure, detection in the am-niotic fluid does. These findings indicate that the developingfetus is exposed to substantial concentrations of MIC-1. Thelevel of MIC-1 in amniotic fluid was comparable with thatpresent in second- and third-trimester maternal serum andwas well in excess of the serum level of nonpregnant women.
FIG. 6. Placental immunohistochemis-try using antibody 233B-P as the pri-mary antibody. Arrow, Trophoblast.
FIG. 7. MIC-1 concentration in five different human placental ex-tracts, as assessed by sandwich ELISA.
4786 MOORE ET AL. JCE & M • 2000Vol. 85 • No. 12
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 10 August 2014. at 21:31 For personal use only. No other uses without permission. . All rights reserved.

The fetus is bathed in amniotic fluid, and there is significantrecirculation of this ingested fluid, which is excreted renallyduring the pregnancy. This provides multiple routes of fetalexposure to MIC-1, topically to the gut and epidermis, as wellas systemically through ingested amniotic fluid.
It has previously been demonstrated, both in our labora-tory (22) and by others (23, 24, 28, 29), that the MIC-1 tran-script is expressed most abundantly in placenta. To deter-mine the source of MIC-1 in pregnancy, we measured MIC-1concentrations in human placental extracts and undertookimmunohistochemical localization. Placental extracts con-tain substantial amounts of MIC-1, in excess of pooled serumor amniotic fluid levels. Using immunohistochemistry, weconfirmed previous observations (29) that the major sourceof placental MIC-1 is the trophoblast. We have also shownthat the BeWo human trophoblastic cell line synthesizes andsecretes MIC-1 constitutively. Together, these findings sug-gest that the trophoblastic cells within the placenta are amajor source of the MIC-1 present in maternal serum andamniotic fluid. However, the localization of the MIC-1 tran-script and protein to the developing epidermis, in day-18 ratembryos (29), suggests that the embryo may also contributeto the MIC-1 levels observed.
The precise role of MIC-1 during pregnancy is unknown.Experimental evidence suggests that MIC-1 has immuno-modulatory activity. For example, rhMIC-1 inhibits the re-lease of proinflammatory cytokines from LPS-activated mac-rophages (1) and suppresses the formation of erythrocyteand granulocyte/macrophage cell lineages from normal hu-man nonadherent T-cell-depleted marrow cells (28). Thesefindings suggest that MIC-1 may have wider modulatoryeffects, possibly suppressing both the development of themonocyte/macrophage lineage and/or their ability to pro-duce proinflammatory mediators. Intrauterine inflammationaccompanying proinflammatory cytokine production hasbeen associated with fetal rejection or preterm labor (15–17).In this context, it is possible that MIC-1, present in the pla-centa and amniotic fluid, acts to maintain pregnancy bysuppressing the production of proinflammatory cytokines.
The finding that a placental extract, derived from a prema-ture labor, contained depressed concentrations of MIC-1,compared with normal pregnancies, supports this hypoth-esis. Intensive examination of MIC-1 levels in maternal se-rum and amniotic fluid during multiple and abnormal preg-nancies should help clarify the precise role of MIC-1 duringpregnancy.
References
1. Bootcov MR, Bauskin A, Valenzuela SM, et al. 1997 MIC-1, a novel macro-phage inhibitory cytokine, is a divergent member of the transforming growthfactor-b superfamily cluster. Proc Natl Acad Sci USA. 94:11514–11519.
2. Bottner M, Laaff M, Schechinger B, Rappold G, Unsicker K, Sutter-Craz-zolara C. 1999 Characterization of the rat, mouse, and human genes of growthdifferentiating factor-15/macrophage inhibiting cytokine-1 (GDNF-15/MIC-1). Gene. 237:105–111.
3. Bottner M, Sutter-Crazzolara C, Scober A, Unsicker K. 1999 Expression of anovel member of the TGF-beta superfamily, growth differentiating factor-15/macriophage inhibiting cytokine-1 (GDNF-15/MIC-1) in adult rat tissues. CellTissue Res. 297:103–110.
4. Graham CH, Lysiak JJ, McCrae KR, Lala PK. 1992 Localisation of transform-ing growth factor at the human fetal-maternal interface: role of trophoblastgrowth and differentiation. Biol Reprod. 46:561–572.
5. Petraglia F, Woodruff TK, Botticelli G, et al. 1993 Gonadotropin-releasinghormone, inhibin, and activin in human placenta: evidence for a commoncellular localisation. J Clin Endocrinol Metab. 74:1184–1188.
6. Petraglia F, Anceschi M, Calza L, et al. 1993 Inhibin and activin in human fetalmembranes: evidence for a local effect on prostaglandin release. J Clin Endo-crinol Metab. 77:542–548.
7. Minami S, Yamoto M, Nakano R. 1992 Immunohistochemical localisation ofinhibin/activin subunits in human placenta. Obstet Gynecol. 80:410–414.
8. Lang AK, Searle RF. 1994 The immunodomodulatory activity of human am-niotic fluid can be correlated with transforming growth factor-b1 and trans-forming growth factor-b2 activity. Clin Exp Immunol. 97:158–163.
9. Qu J, Thomas K. 1992 Changes in bioactive and immunoactive inhibin levelsaround human labor. J Clin Endocrinol Metab. 74:1290–1295.
10. Altman DJ, Schneider SL, Thompson DA, Cheng HL, Tomasi TB. 1990 Atransforming growth factor beta 2-like immunosuppressive factor in amnioticfluid and localisation of the TGF-beta 2 mRNA in the pregnant uterus. J ExpMed. 172:1391–1401.
11. Caniggia I, Grisaru-Gravnosky S, Kuliszewsky M, Post M, Lye SJ. 1999Inhibition of TGF-b3 restores the invasive capacity of extravillous trophoblastsin pre-eclamptic pregnancies. J Clin Invest. 103:1641–1650.
12. Wallace EM, Riley SC, Crossley JA, et al. 1997 Dimeric inhibins in amnioticfluid, maternal serum and fetal serum in human pregnancy. J Clin EndocrinolMetab. 82:218–222.
13. Petraglia F, Vaughan J, Vale W. 1989 Inhibin and activin modulate the releaseof GnRH, hCG, and progesterone from cultured human placental cells. ProcNatl Acad Sci USA. 86:5114–5117.
FIG. 8. MIC-1 expression and secretion by the human trophoblastic cell line BeWo. A, MIC-1 secretion by BeWo cells after 1 and 5 days in culture,as determined by sandwich ELISA. B, Immunoprecipitation and Western blot analysis of secreted MIC-1 by BeWo cells. Lane 1; Unconditionedculture media; lane 2, culture media that had been conditioned by BeWo cells for 5 days. Bands marked: 1, Heavy chain; 2, unprocessed monomer;3, mature dimer; 4, monomer. C, RT-PCR analysis of MIC-1 expression by unstimulated BeWo cells. Lane 1, RT-PCR on total RNA from BeWocells cultured for 24 h; lane 2, negative control (no total RNA); lane 3, positive PCR control.
MIC-1 IN THE SERUM OF PREGNANT WOMEN 4787
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 10 August 2014. at 21:31 For personal use only. No other uses without permission. . All rights reserved.

14. Caniggia I, Lye SJ, Cross JC. 1997 Activin is a local regulator of humancytotrophoblast cell differentiation. Endocrinology. 138:3976–3986.
15. Romero R, Mazor M, Sapulveda W, Avila C, Copeland D, Williams J. 1992Tumour necrosis factor in preterm and term labour. Am J Obstet Gynecol.166:1576–1587.
16. Hillier SL, Witkin SS, Krohn MA, Watts DH, Kiviat NB, Eschenbach DA.1993 The relationship of amniotic fluid cytokines and preterm delivery, am-niotic fluid infection, histologic chorioamnionitis, and chorioamnion infection.Obstet Gynecol. 81:941–948.
17. Opsjon S-L, Wathen N, Tingulstad S, et al. 1993 Tumor necrosis factor,interleukin-1, and interleukin-6 in normal human pregnancy. Am J ObstetGynecol. 169:397–404.
18. Bogdan C, Nathan C. 1993 Modulation of macrophage function by transform-ing growth factor beta, IL-4 and IL-10. Ann NY Acad Sci. 685:713–739.
19. Petraglia F, Sacerdote P, Cossarizani A, et al. 1991 Inhibin and activin mod-ulate human monocyte chemotaxis and human lymphocyte interferon-gammaproduction. J Clin Endocrinol Metab. 72:496–502.
20. Nusing RM, Barsig J. 1997 Inflammatory potency of activin A. Effect onprostanoid and nitric oxide formation. Adv Exp Med Biol. 407:243–248.
21. Petraglia F, Garg S, Florio P, et al. 1993 Activin A and activin B measured inmaternal serum, cord blood serum, and in amniotic fluid during humanpregnancy. Endocr J. 1:323–328.
22. Fairlie WD, Moore AG, Bauskin AR, Russell PK, Zhang HP, Breit SN. 1999MIC-1 is a novel transforming growth factor-beta superfamily cytokine asso-ciated with macrophage activation. J Leukoc Biol. 651:2–5.
23. Lawton LN, de Fatima Bonaldo M, Jelenc PC, et al. 1997 Identification of anovel member of the transforming growth factor-b superfamily highly ex-pressed in human placenta. Gene. 203:17–26.
24. Yokoyama-Koyabashi M, Saeki M, Sekine S, Kato S. 1997 Human cDNAencoding a novel transforming growth factor-b superfamily protein highlyexpressed in placenta. J Biochem. 122:622–626.
25. Fairlie WD, Zhang HP, Brown PK, Russell PK, Bauskin AR, Breit SN. 2000Expression of a TGF-b superfamily protein, macrophage inhibitory cytokine-1,in the yeast Pichia pastoris. Gene. Submitted.
26. Laemmli UK. 1970 Cleavage of structural proteins during the assembly of thehead of bacteriophage T4. Nature. 227:680–685.
27. Fowler PA, Evans LW, Groome NP, Templeton A, Knight PG. 1998 A lon-gitudinal study of maternal serum inhibin A, inhibin B, activiv A, activin AB,pro-aC and follistatin during pregnancy. Hum Reprod. 13:3530–3536.
28. Hromas R, Hufford M, Sutton J, Xu D, Li Y, Lu L. 1997 PLAB, a novel placentalbone morphogenetic protein. Biochim Biophys Acta. 1354:40–44.
29. Paralker VM, Vail AL, Grasser WA, et al. 1998 Cloning and characterisationof a novel member of the transforming growth factor-beta/bone morphogenicprotein family. J Biol Chem. 273:13760–13767.
4788 MOORE ET AL. JCE & M • 2000Vol. 85 • No. 12
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 10 August 2014. at 21:31 For personal use only. No other uses without permission. . All rights reserved.





























