Embed Size (px)
Citation preview

RESEARCH ARTICLE
The tumor suppressor LKB1 antagonizes WNT signalingpathway through modulating GSK3β activity in cell growthof esophageal carcinoma
Kai Liu & Yue Luo & Hui Tian & Kai-Zhong Yu &
Jin-Xian He & Wei-Yu Shen
Received: 4 July 2013 /Accepted: 23 August 2013 /Published online: 11 September 2013# International Society of Oncology and BioMarkers (ISOBM) 2013
Abstract The tumor suppressor LKB1 gene encodes a serine–threonine kinase that regulates cell proliferation and polarity.Inactivation of LKB1 by mutations in LKB1 or loss of itsexpression is highly correlated with lung, ovarian, and pancre-atic cancers, and WNT/β-catenin pathway is also known to beinvolved in many human malignancies. However, the relation-ship between LKB1 and WNT signaling pathway in esopha-geal carcinoma remains unknown. The expression of LKB1 in62 cases of esophageal cancer patients was determined byquantitative real-time PCR. It was found that LKB1 mRNAlevel was significantly lower than the adjacent normal epithe-lium and that the LKB1 downregulation was correlating withTNM stages. Moreover, the expression of WNT target genessuch as Cyclin D1, C-MYC, MMP2, and FZD2 was signifi-cantly upregulated in esophageal cancer tissues. LKB1overexpression in TE10 cells inhibited TOPFlash luciferasereporter activity and WNT target gene expression even in thepresence of WNT3A. Conversely, LKB1 knockdown en-hanced WNT signaling activity in esophageal cancer cells. Itwas also found that LKB1 antagonized WNT signaling path-way through interaction with GSK3β to downregulateβ-catenin expression level. Functional investigation revealedthat LKB1 suppressed the promotion effects ofWNT3A on thecell growth of TE10 cells. The LKB1 functions in regulatingcell growth and WNT target genes expression were impairedby GSK3β inhibition, suggesting that LKB1 antagonizedWNT-induced cell proliferation through enhancement ofGSK3β activity. Together, the interaction between LKB1 and
GSK3β upregulates GSK3β activity to suppress WNT-induced cell proliferation in esophageal carcinoma cells. Lossof LKB1 expression may result in the deregulation of WNT/β-catenin pathway to promote malignant progression of esopha-geal cancer.
Keywords LKB1 .WNTsignaling pathway . GSK3β .
Esophageal carcinoma
Introduction
Esophageal cancer is one of the most common malignancies,accounting for the leading causes of cancer deaths all over theworld [1, 2]. Esophageal cancer belongs to invasive andmetastatic cancers, which leads to the high rate of cancerdeath. Although the advances in the diagnosis and treatmentof esophageal carcinoma improve patient survival [3, 4],effective approaches to cure esophageal cancer remain elu-sive. Previous reports suggest that esophageal cancer is asso-ciated with the aberrant expression of p53, Cyclin D1, and p16[5–8]. However, the exact molecular mechanisms underlyingthe initiation and development of esophageal cancer remainunclear. Therefore, it will be of great significance to elucidatethe molecular mechanisms for esophageal cancer diagnosisand treatment.
The serine–threonine kinase plays essential roles in thedevelopment and cell polarity [9, 10]. LKB1 has also beenimplicated in cancer development, the mutations of whichcause the inherited human cancer disorder Peutz–Jeghers syn-drome [11, 12]. Subsequent studies demonstrated that LKB1is a target for mutational inactivation or aberrant expression inhuman malignancies including lung, colorectal, testis, mela-noma, cervical, and pancreatic cancer [13–18]. Currently, themechanisms of LKB1-mediated tumor suppression remainlimited. LKB1 inhibits tumor cell cycle progression through
K. Liu :H. Tian :K.<Z. Yu : J.<X. He :W.<Y. Shen (*)Department of Thoracic Surgery, Ningbo Medical Center, LihuiliHospital, No. 57 XingNing Road, Ningbo, Zhejiang 315041, Chinae-mail: [email protected]
Y. LuoDepartment of Paediatrics, Wenzhou Medical College affiliatedYuYing Children’s Hospital, Wenzhou, China
Tumor Biol. (2014) 35:995–1002DOI 10.1007/s13277-013-1133-0

inducing p21 and p53 gene expression dependent on its kinaseactivity [19, 20], and LKB1 deficiency leads to the inductionof MMP2, MMP9, and VEGF [9, 21]. In esophageal cancercells, altered LKB1/CRTC1 signaling promotes cell migrationand invasion [22]. Meanwhile, the functional mechanismunderlying LKB1 functions in esophageal carcinoma cellgrowth and invasion was not fully understood.
The WNT/β-catenin signaling pathway plays essentialroles in cell proliferation and differentiation, and deregulatedWNT/β-catenin pathway leads to many types of human can-cers [23–26]. In esophageal cancer cells, WNT1 inducesβ-catenin/TCF-dependent transcription [27], and alteredα-catenin and β-catenin expression was reported in esopha-geal cancer tissues [28, 29]. In Xenopus development, innateLKB1 has been shown to function by regulating GSK3βactivity [30], while Peutz–Jeghers LKB1 mutants fail to acti-vate GSK3β [31]. As a kinase, LKB1 also triggers phosphor-ylation of PAR1A to regulate WNT/β-catenin signaling path-way [32]. However, it remains unknown whether WNT/β-catenin signaling pathway is involved in progression of esoph-ageal cancer. It is also unclear whether and how LKB1 mod-ulates WNT/β-catenin pathway to regulate esophageal cancercell proliferation or invasion.
In this study, we collected esophageal carcinoma samplesand adjacent non-cancer epithelium from 62 esophageal cancerpatients for quantitative real-time PCR (qRT-PCR) analysis. Itwas found that downregulated LKB1 expression and elevatedWNT target genes were observed in esophageal carcinomatissues accompanying with stage progression. Studies in esoph-ageal carcinoma cell line TE10 cells revealed that LKB1 in-hibits WNTsignaling activity through interaction with GSK3βto result in GSK3β phosphorylation, which leads to subsequentphosphorylation and degradation of β-catenin. Meanwhile,LKB1 knockdown enhances the transcriptional activity ofWNTsignaling. Additionally, LKB1 acts as a tumor suppressorto inhibitWNT-induced cell proliferation through the activationof GSK3β activity in esophageal cancer cells. These findingssuggest that loss of LKB1 in normal cells may result in theinitiation of original esophageal tumors.
Materials and methods
Samples and cell lines
Sixty two esophageal carcinoma specimens and their adjacentnormal epithelium were collected from Lihuili Hospital fromFebruary 2008 to October 2012. The conditions of esophagealcancer patients were summarized in Table 1. Tumors werestaged histologically according to the Guide Lines for theClinical and Pathologic Studies on Carcinoma of theEsophagus. Samples were freshly frozen. When enough speci-mens were collected, each sample was subjected for RNA
extraction and qRT-PCR analysis. Informed consent wasobtained from each patient prior surgery. This study was ap-proved by Lihuili Hospital Clinical Research Ethics Committee.Two-year survival rate was calculated from the date of operationto the date of the latest follow-up visit or death due to recurrentesophageal cancer. TE10 cells were originally obtained from theAmerican Type Culture Collection (ATCC) and culturedaccording to ATCC guidelines.
Quantitative real-time PCR
Total RNA was isolated from cells or tissues with TRIzolReagent (Life Technologies, Carlsbad, CA, USA) accordingto the manufacturer’s recommendations. Five microgramsRNA was reversely transcribed using Superscript III ReverseTranscriptase (Invitrogen). qRT-PCR was performed accordingto the manufacturer’s protocols which is amplified with SYBRGreen real-time PCR master mix (Sigma). Relative expressionlevel of target genes was normalized according to GAPDH. Allprimer sequences are available on request, which were mostlydesigned from Primer bank (http://pga.mgh.harvard.edu/primerbank/).
Western blot analysis
Cells were harvested in lysis buffer (150 mM NaCl, 50 mMTris–HCl (pH 8.0), 0.1 % SDS, 0.5 % sodium deoxycholate,5 mM EDTA, 1 % Nonidet P-40, 0.25 mM phenylmethane-sulfonyl fluoride, and protease inhibitors). Protein extracts werefractionated by SDS-PAGE on polyacrylamide gels and thenelectrophoretically transferred to nitrocellulose membranes,followed by blocking in with 5 % skim milk and immunoblot-ting with anti-GSK3β (1:500, Cell Signaling Technology,Danvers, MA, USA), anti-p-GSK3β (Tyr216) (1:1,000, BDTransduction Laboratories, Lexington, KY, USA), anti-β-catenin (1:1,000, Transduction Laboratories, San Jose, CA,USA), anti-LKB1 (1:400, Santa Cruz Biotechnology, SantaCruz, CA, USA), anti-GAPDH (1:5,000, Abcam, Cambridge,MA, USA), anti-Flag (1:1,000, Sigma), and anti-Myc(Covance). The secondary horseradish peroxidase linked an-tibody to rabbit or mouse IgG was then incubated at roomtemperature for 1–2 h. A commercially available detectionsystem (Pierce) was used for ECL exposure.
Table 1 Survival rate analysis in esophageal carcinoma patients
Phase Male/female Number (%) Two-year survival rate
I 1/4 5 (8 %) 100 % (n =5)
II 14/16 30 (48.2 %) 80 % (n =24)
III 7/11 18 (29 %) 44.4 % (n=8)
IV 4/5 9 (14.5 %) 0 (n =0)
996 Tumor Biol. (2014) 35:995–1002
Transient transfection and viral transduction
TE10 cells were transfected using Lipofectamine 2000 re-agents (Life Technologies, Carlsbad) following the manufac-turer’s instructions. Lentiviral production and transductionwere previously described [33]. pLKO.1-GFP lentiviralLKB1 short hairpin RNA (shRNA) construct was orig-inally from Open Biosystems (Huntsville, AL, USA).The same vectors with random short hairpin RNA wereused as control.
Immunoprecipitation
Immunoprecipitation (IP) was performed as previouslydescribed [34]. Briefly, 293T cells were transfected withplasmids pcDNA3–LKB1–Flag and pcDNA3–pcDNA3–GSK3β–myc using Lipofectamine 2000. TE10 cells ortransfected 293T cells were lysed with lysis buffer(50 mM Tris–HCl pH 8.0, 150 mM NaCl, 1 % TritonX-100 containing a protease inhibitor cocktail (Roche)),and the supernatant was precleared with protein G aga-rose beads (Roche). Subsequently, antibodies were incu-bated with the cell extracts, and 1 h later, the protein Gagarose beads (Roche) were added for incubation. Thewashed agarose beads were resuspended in SDS gel-loading buffer (62 mM Tris–HCl pH 7.4, 2 % SDS,10 % glycerol, 5 % b-mercaptoethanol) and boiled for3 min. The samples were subjected to western blottinganalysis with indicated antibodies.
Cell proliferation assay
Cell proliferation was detected using microculture tetrazolium(MTT) assays as previously described [35]. Cells were seededin 24-well plates at a density of 3×104 cells per well andtransfected with indicated plasmids. After transfection for24–48 h, cells were incubated with 0.5 mg/mLMTT in culturemedium for an additional 4 h. The cell proliferation wasmeasured spectrophotometrically at 570 nm (OD570). Themean values of three wells were calculated as the final ODvalue.
Luciferase assay
TE10 cells were plated in six-well plates at 50 % confluencyand co-transfected with a WNT luciferase reporter vectorTOPFlash, pRL-TK (Renilla, Promega), and pcDNA3-LKB1 or pSuper-LKB1 shRNA (pcDNA3 or pSuper-conserved as control) using transfection reagents. Transfectionswere repeated at least in three wells for each experiment.Firefly and Renilla luciferase activities were determined 24–48 h post-transfection using the Dual Luciferase ReporterAssay System (Promega).
Statistical analysis
All results were obtained from two to three independentexperiments. Student’s t test was used for data analysis.A p value of <0.05(*) was considered statistically significant.
Results
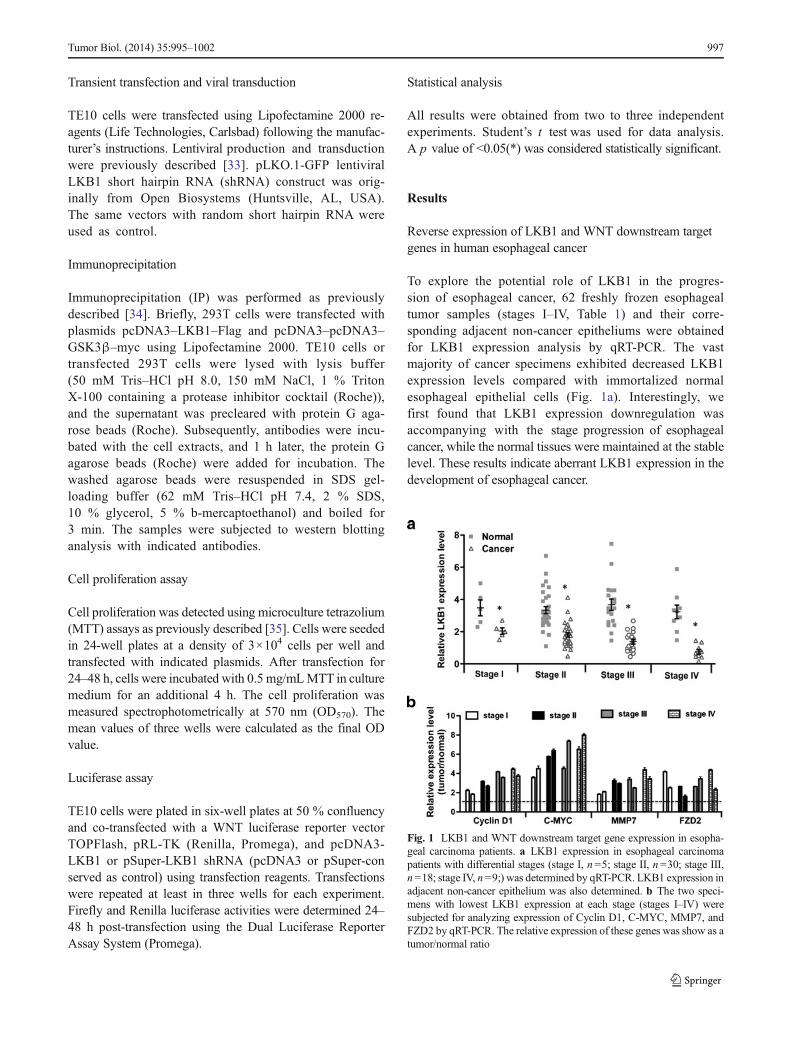
Reverse expression of LKB1 and WNT downstream targetgenes in human esophageal cancer
To explore the potential role of LKB1 in the progres-sion of esophageal cancer, 62 freshly frozen esophagealtumor samples (stages I–IV, Table 1) and their corre-sponding adjacent non-cancer epitheliums were obtainedfor LKB1 expression analysis by qRT-PCR. The vastmajority of cancer specimens exhibited decreased LKB1expression levels compared with immortalized normalesophageal epithelial cells (Fig. 1a). Interestingly, wefirst found that LKB1 expression downregulation wasaccompanying with the stage progression of esophagealcancer, while the normal tissues were maintained at the stablelevel. These results indicate aberrant LKB1 expression in thedevelopment of esophageal cancer.
Fig. 1 LKB1 and WNT downstream target gene expression in esopha-geal carcinoma patients. a LKB1 expression in esophageal carcinomapatients with differential stages (stage I, n=5; stage II, n=30; stage III,n =18; stage IV, n =9;) was determined by qRT-PCR. LKB1 expression inadjacent non-cancer epithelium was also determined. b The two speci-mens with lowest LKB1 expression at each stage (stages I–IV) weresubjected for analyzing expression of Cyclin D1, C-MYC, MMP7, andFZD2 by qRT-PCR. The relative expression of these genes was show as atumor/normal ratio
Tumor Biol. (2014) 35:995–1002 997

WNT/β-catenin pathway is frequently activated in variouscancers [24, 25], which has not been reported in esophagealcancer. WNT signaling is tightly connected with LKB1[30–32], while the specific regulatory network is not elucidat-ed in esophageal cancer. To determine the correlation betweenWNT signaling and LKB1 in vivo, the expression of WNTtarget genes including Cyclin D1, C-MYC,MMP7, and FZD2was determined in the two patient samples with lowest LKB1expression from each stage. We found that the expression ofthe WNT target genes were significantly higher (more thantwofold) in tumors comparing with corresponding normalepithelium (Fig. 1b), suggesting that WNT signaling pathwayis also aberrantly activated in esophageal cancer. Taken to-gether, our data demonstrate that LKB1 expression is gradu-ally lost in esophageal tumorigenesis, and the reverse expres-sion pattern of LKB1 and WNT target genes indicates tightassociation between LKB1 downregulation and abnormalWNT signaling activity.
LKB1 antagonizes WNT signaling pathway in esophagealcarcinoma cells
To investigate the molecular relationship between LKB1 andWNT/β-catenin pathway, esophageal cell line TE10 cells[22], which possess low but detectable level of LKB1 expres-sion, were chosen to perform biological analysis in vitro.When LKB1was overexpressed in TE10 cells,WNT luciferasereport (TOPFlash) activity was obviously suppressed (Fig. 2a).Moreover, WNT3A stimulated TOPFlash activity was alsoimpaired by LKB1 overexpression (Fig. 2a), suggesting thatLKB1 antagonizes WNT signaling pathway in esophagealcancer cells. To further confirm this, the expression of WNTtarget genes Cyclin D1, C-MYC, MMP7, and FZD2 wasdetermined. It was found that forced expression of LKB1inhibited the expression of WNT target genes in the presenceor absence of WNT3A (Fig. 2b).
Subsequently, we used lentivirus-mediated LKB1 shRNAto transfect TE10 cells with high efficiency (Fig. 2c). Westernblotting analysis showed that LKB1 was significantlyknocked down by LKB1 shRNA (Fig. 2d). Consistent withour predication, LKB1 knockdown enhanced TOPFlash lucif-erase activity (Fig. 2e). Additionally, the expression of WNTtarget genes was also upregulated by LKB1 knockdown(Fig. 2f). Collectively, LKB1 antagonizes WNT signalingpathway and inhibits the expression of WNT downstreamtarget genes in esophageal carcinoma cells.
LKB1 interacts with GSK3β to downregulate β-cateninprotein level
The classical WNT signaling cascade inhibits the activity ofthe enzyme GSK3β, augments β-catenin stabilization and
translocation to the nucleus, and promotes the transcriptionof target genes [36, 37]. To determine how LKB1 affectsWNT signaling activity, the effects of LKB1 overexpressionon WNT signaling cascade were determined by western blot-ting analysis. β-catenin protein level was downregulated, andWNT-induced β-catenin upregulation was also impaired byLKB1 overexpression (Fig. 3a,b). Furthermore, phosphorylat-ed GSK3β on Tyr-216 (p-GSK3β (Tyr216)) (enzymaticallyactive form) was increased, and WNT3A-elicited p-GSK3βreduction was partially recovered when LKB1 wasoverexpressed in TE10 cells (Fig. 3a,b). These data suggestthat LKB1 may activate GSK3β activity by promoting itsphosphorylation at Tyr-216. We speculated that LKB1 mightinteract with GSK3β to directly modulating GSK3β activity.
To test this hypothesis, we first performed co-immunoprecipitation in 293T cells. When LKB1-Flag andGSK3β-myc were co-transfected, LKB1-Flag could beimmunoprecipitated by GSK3β-myc (Fig. 3c), demonstratingthe physical association between LKB1 and GSK3β. Next, weused GSK3β antibody to immunoprecipitate endogenic LKB1in TE10 cells, and the endogenic interaction between the twoproteins was observed (Fig. 3d). Moreover, more LKB1 pro-teins were immunoprecipitated by GSK3β in LKB1overexpressed TE10 cells (Fig. 3d), further confirming theinteraction between GSK3β and LKB1 in esophageal cells.
Based on our findings that LKB1 affects β-catenin andp-GSK3β protein levels and that LKB1 physically interactswith GSK3β, we postulate that LKB1might induceβ-catenindownregulation through activating GSK3β. To test this, weused a GSK3β inhibitor LiCl to block GSK3β activity inLKB1 overexpressing TE10 cells. As expected, LKB1-induced β-catenin decrease was fully blocked by GSK3β inhi-bition (Fig. 3e). These results indicate that LKB1 interacts withGSK3β to activate GSK3β activity and further downregulateβ-catenin protein level.
LKB1 antagonizes WNT-induced cell growth of esophagealcarcinoma cells through activating GSK3β
Given that LKB1 antagonizes WNT signaling pathway throughactivation of GSK3β in esophageal cells, we next try to explorethe functional relationship between LKB1 and WNT signalingin the molecular events of esophageal cancer pathogenesis. Cellinvasion, survival, and proliferation are important characteristicsof cancer cells [38], and cell proliferation capacity was deter-mined in TE10 cells using MTT assay. We found that LKB1overexpression significantly suppressed the cell growth of TE10cells (Fig. 4a). Conversely, cell proliferationwas promotedwhenLKB1was knocked down by LKB1 shRNA in esophageal cells(Fig. 4b).
WNT target genes were aberrantly expressed in esophagealcells (Fig. 1a). It was reasonable to speculate that WNT
998 Tumor Biol. (2014) 35:995–1002

signaling plays essential roles in the pathogenesis of esopha-geal cancer. Consistently, WNT3A promoted the cell growthof TE10 cells in a dose-dependent manner (Fig. 4c).Importantly, LKB1 overexpression impaired WNT-inducedpromotion of cell growth (Fig. 4d), suggesting that LKB1functionally antagonizes WNT effects in esophageal cancercell proliferation, which is consistent with our findings ofLKB1 effects on WNT luciferase activity and WNT targetgene expression (Fig. 2).
To determine whether LKB1 inhibits WNT-induced cellgrowth through activation of GSK3β, we used LiCl to blockGSK3β activity in LKB1 overexpressing cells. It was foundthat GSK3β inhibition leaded to the increase of cell growthand that LiCl also fully blocked the inhibitory effects of LKB1in cell proliferation of TE10 cells (Fig. 4e). When LiCl wassubjected to cells, the expression ofWNT target genes was notaffected by LKB1 overexpression (Fig. 4f). Together, LKB1antagonizes WNT-induced cell growth of esophageal cancercells through activation of GSK3β.
Discussion
Esophageal carcinoma is an aggressive and life-threateningtumor with a poor prognosis [39, 40]. It is consistent with ourstatistical analysis of two-year survival rate (Table 1), andstage I patients are very hard to be diagnosed in clinical cases.It is vital to investigate the molecular mechanism underlyingthe pathogenesis of esophageal cancer.
Mutational inactivation or loss of LKB1 has been reportedin several human malignancies [13, 14, 18]; the questionwhether and how LKB1 acts in esophageal cancer was notfully understood. We identified a reduction of LKB1 expres-sion in esophageal cancer samples, and the extent of LKB1decrease was augmented with cancer progression (Fig. 1),suggesting that LKB1 downregulation might be correlatedwith esophageal cancer pathogenesis. It is consistent withprevious report [22], while we first reported the exact inci-dence of LKB1 downregulation at stage I. Based on thisobservation, LKB1 downregulation might be applied as a
Fig. 2 LKB1 inhibits WNT signaling pathway in esophageal carcinomacells. a The activity of WNT luciferase report TOPFlash was determinedwhen TE10 cells were co-transfected with pcDNA3-LKB1 (OV) or emptyvector (Con), supplemented with or without WNT3A (50 ng/mL). b Theexpression of Cyclin D1, C-MYC, MMP7, and FZD2 was examined inTE10 cells under the situation in (a). c Lentivirus with scramble shRNA(Con shRNA) or LKB1 shRNA was used to transfect TE10 cells. The
transfection efficiency is over 85%. d The knockdown efficiency of LKB1shRNA was determined by western blotting analysis. e The TOPFlashluciferase activity was examined in control shRNA (Con) or LKB1 shRNA(KD) expressing cells. WNT3A (50 ng/mL) was added into the mediumfor incubation 24 h after transfection for 24 h. f Determination of theexpression of Cyclin D1, C-MYC, MMP7, and FZD2 in TE10 cells withsame conditions in (e)
Tumor Biol. (2014) 35:995–1002 999

molecular diagnostic marker at early stage of esophageal can-cer. Functional analysis revealed that LKB1 inhibits the cellgrowth of esophageal cells (Fig. 4), confirming the previousfindings of LKB1 functions in cell migration and invasion[22].
Our data provided evidence that LKB1 knockdown has apro-invasive effect on TE10 cell-based assays (Fig. 4), indi-cating that LKB1 downregulation may contribute to the inva-sive property of esophageal cancer cells. However, the exactrole of LKB1 in the initiation and progression of esophagealcancer is unclear. WNT/β-catenin pathway is aberrantly acti-vated in various cancers [23–26], while it was unknown aboutWNT signaling activity in esophageal cancer in vivo. LKB1has been suggested be related to WNT signaling pathway[30–32], we decide to explore whether LKB1 could modulateWNTsignaling in esophageal cancer cells. Detection of WNTtarget genes including Cyclin D1, C-MYC,MMP7, and FZD2revealed that the expression of WNT target genes was appar-ently upregulated in esophageal cancer tissues with very low
LKB1 level (Fig. 1). LKB1 overexpression suppressed WNTluciferase reporter activity and inhibited the expression ofWNT target genes, while LKB1 knockdown resulted in anopposite outcome (Fig. 2). It demonstrates that LKB1 reduc-tion is the main cause of upregulation of WNT signaling.Moreover, we first reported that WNT3A stimulation leadedto the promotion of cell growth (Fig. 4), suggesting that WNTsignaling is functionally involved in esophageal cancer path-ogenesis through upregulating WNT target gene expression.
LKB1 regulates multiple kinases and has the potential tomodulate different signaling activities. LKB1 interacts withseveral partners such as PTEN, CRTC, and AMPK [22, 34,41] to regulate multiple processes. LKB1 has been reported tobe associated with GSK3β to regulate WNT signaling invertebrate development [30]. Here, we found that LKB1 wasable to promote the phosphorylation of GSK3β through in-teraction with GSK3β. LKB1 also reduced the protein level ofβ-catenin even in the presence of WNT stimulation, which isdependent on the activation of GSK3β (Fig. 3). It is possible
Fig. 3 LKB1 interacts withGSK3b to downregulateβ-catenin protein level.a Western blotting analysis of the protein levels of β-catenin, p-GSK3β(Tyr216), GSK3β, LKB1, and GAPDH. TE10 cells were transfected withpcDNA3-LKB1 (OV) or empty vector (con), supplemented with or withoutWNT3A (50 ng/mL). b Quantification of β-catenin relative to GAPDH andquantification of p-GSK3β (Tyr216) relative to GSK3β in (a). c LKB1-Flagand GSK3β were transfected in 293T cells as indicated. Cell extract wassubjected for immunoprecipitation (IP) with anti-myc antibody, and immu-noblotting (IB) was performed using anti-Flag antibody. The input of LKB1-
Flag, GSK3β-myc, and GAPDH expression was also determined with anti-Flag, anti-myc, and anti-GAPDH antibodies, respectively. d TE10 cellstransfected with pcDNA3-LKB1 (OV ) or empty vector (con ) wasimmunoprecipitated (IP) using anti-GSK3β antibody and IgG was used asnegative control. The immunoprecipitates were analyzed by immunoblottingwith anti-LKB1 antibody. The protein level of LKB1, GSK3β, and GAPDHwas also examined in input lysates. e Western blotting analysis of β-catenin,p-GSK3β (Tyr216), andGAPDHexpressionwhen vector (con) or pcDNA3-LKB1 (OV) transfected TE10 cells were treated with 5 mM LiCl for 24 h
1000 Tumor Biol. (2014) 35:995–1002

that GSK3β is the direct substrate of LKB1 kinase andthat LKB1 leads to the degradation of β-catenin throughactivation of GSK3β. Importantly, LKB1 antagonizedWNT-induced cell proliferation of esophageal cancer cells,which is also dependent on the activation of GSK3β (Fig. 4).
In summary, LKB1 downregulation in esophagealcancer was observed, which was reversely related withWNT target gene upregulation. LKB1 antagonizesWNT/β-catenin pathway to negatively regulate esopha-geal cancer cell growth, and the mechanism is brieflysummarized in our hypothetic model (Fig. 4g). LKB1
interacts with GSK3β to promote phosphorylation ofGSK3β. Then, WNT protein-induced β-catenin stabilityis inhibited by activated GSK3β, and β-catenin proteinlevel is decreased. Subsequently, the transcription of WNTtarget genes is partially turned off, and WNT stimulated cellgrowth is inhibited by LKB1. The loss of LKB1 downregulatesthe activity of GSK3β and derepresses WNT signaling path-way to promote cell proliferation of esophageal cancer. TheLKB1-dependent GSK3β signaling pathway might become aneffective drug target for diagnosis and treatment of esophagealcancer.
Fig. 4 LKB1 antagonizes WNT-induced cell growth of esophagealcarcinoma cells throughactivating GSK3b. a MTT assayswere performed in TE10 cellswith vector (con) or pcDNA3-LKB1 (OV) transfected TE10cells at indicated days. OD570
values represent the cellproliferation capacity of TE10esophageal cancer cells. b LKB1knockdown promotes the cellproliferation of TE10 cells. MTTassays were performed in LKB1shRNA expressing cells. c Thecell proliferation was examinedusing MTT assay at day 3 whenTE10 cells were treated withincreasing concentrations ofWNT3A at 10, 20, 50,100 ng/mL, respectively. d LKB1overexpression (OV) antagonizedWNT3A-induced cell growth.Vector (con) or pcDNA3-LKB1(OV) transfected TE10 cellssupplemented with WNT3A(50 ng/mL) were subjected toMTT assays. e The cell growth incells transfected with vector (con)or pcDNA3-LKB1 (OV) wasdetermined using MTT assaywhen cells were treated with5 mM LiCl. f The expression ofCyclin D1, C-MYC, MMP7, andFZD2 was examined in TE10cells with the same conditions in(e). g Hypothetic model of LKB1functions in WNT signalingregulation of esophageal cancercell growth
Tumor Biol. (2014) 35:995–1002 1001

Conflicts of interest None
References
1. Vallbohmer D, Brabender J, Metzger R, Holscher AH. Genetics inthe pathogenesis of esophageal cancer: possible predictive and prog-nostic factors. J Gastrointest Surg. 2010;14 Suppl 1:S75–80.
2. Klein CA, Stoecklein NH. Lessons from an aggressive cancer: evo-lutionary dynamics in esophageal carcinoma. Cancer Res. 2009;69:5285–8.
3. Knisely JP, Burtness BA, Salem RR. Surgical treatment of esophagealcancer. N Engl J Med. 2003;348:1177–9. author reply 1177–1179.
4. Kocher HM, Tekkis PP. Surgical treatment of esophageal cancer. NEngl J Med. 2003;348:1177–9. author reply 1177–1179.
5. JiangW,ZhangYJ,Kahn SM,HollsteinMC, Santella RM, Lu SH, et al.Altered expression of the cyclin d1 and retinoblastoma genes in humanesophageal cancer. Proc Natl Acad Sci U S A. 1993;90:9026–30.
6. Montesano R, Hollstein M, Hainaut P. Genetic alterations in esoph-ageal cancer and their relevance to etiology and pathogenesis: areview. Int J Cancer. 1996;69:225–35.
7. Kuwano H, Kato H, Miyazaki T, Fukuchi M, Masuda N, Nakajima M,et al. Genetic alterations in esophageal cancer. Surg Today. 2005;35:7–18.
8. Igaki H, Sasaki H, Kishi T, Sakamoto H, Tachimori Y, Kato H, et al.Highly frequent homozygous deletion of the p16 gene in esophagealcancer cell lines. BiochemBiophys Res Commun. 1994;203:1090–5.
9. Ylikorkala A, Rossi DJ, Korsisaari N, Luukko K, Alitalo K,Henkemeyer M, et al. Vascular abnormalities and deregulation ofVEGF in LKB1-deficient mice. Science. 2001;293:1323–6.
10. Watts JL,MortonDG, Bestman J, KemphuesKJ. TheC. elegans par-4gene encodes a putative serine-threonine kinase required forestablishing embryonic asymmetry. Development. 2000;127:1467–75.
11. Hemminki A,Markie D, Tomlinson I, Avizienyte E, Roth S, LoukolaA, et al. A serine/threonine kinase gene defective in Peutz-Jegherssyndrome. Nature. 1998;391:184–7.
12. van Lier MG, Wagner A, Mathus-Vliegen EM, Kuipers EJ,Steyerberg EW, van Leerdam ME. High cancer risk in Peutz-Jeghers syndrome: a systematic review and surveillance recommen-dations. Am J Gastroenterol. 2010;105:1258–64. author reply 1265.
13. Avizienyte E, Loukola A, Roth S, Hemminki A, Tarkkanen M,Salovaara R, et al. LKB1 somatic mutations in sporadic tumors.Am J Pathol. 1999;154:677–81.
14. Avizienyte E, Roth S, Loukola A, Hemminki A, Lothe RA, StenwigAE, et al. Somatic mutations in LKB1 are rare in sporadic colorectaland testicular tumors. Cancer Res. 1998;58:2087–90.
15. Sato N, Rosty C, Jansen M, Fukushima N, Ueki T, Yeo CJ, et al.STK11/LKB1 Peutz-Jeghers gene inactivation in intraductalpapillary-mucinous neoplasms of the pancreas. Am J Pathol.2001;159:2017–22.
16. Wang ZJ, Churchman M, Campbell IG, Xu WH, Yan ZY,McCluggage WG, et al. Allele loss and mutation screen at thePeutz-Jeghers (LKB1) locus (19p13.3) in sporadic ovarian tumours.Br J Cancer. 1999;80:70–2.
17. Sanchez-Cespedes M. A role for LKB1 gene in human cancer be-yond the Peutz-Jeghers syndrome. Oncogene. 2007;26:7825–32.
18. Guldberg P, thor Straten P, Ahrenkiel V, Seremet T, KirkinAF, ZeuthenJ. Somatic mutation of the Peutz-Jeghers syndrome gene, LKB1/STK11, in malignant melanoma. Oncogene. 1999;18:1777–80.
19. Tiainen M, Vaahtomeri K, Ylikorkala A, Makela TP. Growth arrestby the LKB1 tumor suppressor: induction of p21(waf1/cip1). HumMol Genet. 2002;11:1497–504.
20. Tiainen M, Ylikorkala A, Makela TP. Growth suppression by LKB1is mediated by a g(1) cell cycle arrest. Proc Natl Acad Sci U S A.1999;96:9248–51.
21. Kim DW, Chung HK, Park KC, Hwang JH, Jo YS, Chung J, et al.Tumor suppressor LKB1 inhibits activation of signal transducer andactivator of transcription 3 (STAT3) by thyroid oncogenic tyrosinekinase rearranged in transformation (RET)/papillary thyroid carcino-ma (PTC). Mol Endocrinol. 2007;21:3039–49.
22. Chen Y, Wu X, Bu S, He C, Wang W, Liu J, et al. Promisingoutcomes of definitive chemoradiation and cetuximab for patientswith esophageal squamous cell carcinoma. Cancer Sci. 2012;103:1979–84.
23. ClementsWM,Wang J, Sarnaik A, KimOJ, MacDonald J, Fenoglio-Preiser C, et al. Beta-catenin mutation is a frequent cause of WNTpathway activation in gastric cancer. Cancer Res. 2002;62:3503–6.
24. Barker N, Clevers H. Catenins, WNT signaling and cancer.Bioessays. 2000;22:961–5.
25. Polakis P. WNTsignaling and cancer. Genes Dev. 2000;14:1837–51.26. Holland JD, Klaus A, Garratt AN, Birchmeier W. WNT signaling in
stem and cancer stem cells. Curr Opin Cell Biol. 2013;25:254–64.
27. Mizushima T, Nakagawa H, Kamberov YG, Wilder EL, Klein PS,Rustgi AK. WNT-1 but not epidermal growth factor induces beta-catenin/t-cell factor-dependent transcription in esophageal cancercells. Cancer Res. 2002;62:277–82.
28. Nakanishi Y, Ochiai A, Akimoto S, Kato H, Watanabe H, TachimoriY, et al. Expression of e-cadherin, alpha-catenin, beta-catenin andplakoglobin in esophageal carcinomas and its prognostic signifi-cance: immunohistochemical analysis of 96 lesions. Oncology.1997;54:158–65.
29. Kadowaki T, Shiozaki H, Inoue M, Tamura S, Oka H, Doki Y, et al.E-cadherin and alpha-catenin expression in human esophageal can-cer. Cancer Res. 1994;54:291–6.
30. Ossipova O, Bardeesy N, DePinho RA, Green JB. LKB1 (XEEK1)regulates WNT signalling in vertebrate development. Nat Cell Biol.2003;5:889–94.
31. Lin-Marq N, Borel C, Antonarakis SE. Peutz-Jeghers lkb1 mutantsfail to activate gsk-3beta, preventing it from inhibitingWnt signaling.Mol Genet Genomics. 2005;273:184–96.
32. Spicer J, Rayter S, Young N, Elliott R, Ashworth A, Smith D.Regulation of the WNT signalling component par1a by thePeutz-Jeghers syndrome kinase LKB1. Oncogene. 2003;22:4752–6.
33. Chen J, Kesari S, Rooney C, Strack PR, Chen J, Shen H, et al.Inhibition of notch signaling blocks growth of glioblastoma cell linesand tumor neurospheres. Genes Cancer. 2010;1:822–35.
34. Mehenni H, Lin-Marq N, Buchet-Poyau K, ReymondA, CollartMA,Picard D, et al. Lkb1 interacts with and phosphorylates PTEN: afunctional link between two proteins involved in cancer predisposingsyndromes. Hum Mol Genet. 2005;14:2209–19.
35. Schmidt AI, Reismann M, Kubler JF, Vieten G, Bangen C,Shimotakahara A, et al. Exposure to carbon dioxide and heliumreduces in vitro proliferation of pediatric tumor cells. Pediatr SurgInt. 2006;22:72–7.
36. Espada J, Calvo MB, Diaz-Prado S, Medina V. WNT signalling andcancer stem cells. Clin Transl Oncol. 2009;11:411–27.
37. Reya T, Clevers H. WNT signalling in stem cells and cancer. Nature.2005;434:843–50.
38. Evan GI, Vousden KH. Proliferation, cell cycle and apoptosis incancer. Nature. 2001;411:342–8.
39. Lu XJ, Luo JD, Ling Y, Kong YZ, Feng LL, Zhou J, et al.Management of small cell carcinoma of esophagus in China. JGastrointest Surg. 2013;17(7):1181–7.
40. Takubo K, Makuuchi H, Arima M, Aida J, Arai T. Vieth M: [lymphnode metastasis in superficial squamous carcinoma of the esopha-gus]. Pathologe. 2013;34:148–54.
41. Shackelford DB, Shaw RJ. The LKB1-AMPK pathway: metabolismand growth control in tumour suppression. Nat Rev Cancer. 2009;9:563–75.
1002 Tumor Biol. (2014) 35:995–1002











![NEAT1 regulates microtubule stabilization via FZD3/GSK3β/P ...€¦ · to control gene expression and epigenetic events [10, 11]. The NEAT1 gene has two isoforms, NEAT1v1 (3.7 kb](https://img.pdfslide.tips/doc/110x75/60e1a9861d33103c6f3754f5/neat1-regulates-microtubule-stabilization-via-fzd3gsk3p-to-control-gene.jpg)

















