Embed Size (px)
Citation preview

Thermoadaptation-Directed Enzyme Evolution in an Error-ProneThermophile Derived from Geobacillus kaustophilus HTA426
Hirokazu Suzuki,a,b Jyumpei Kobayashi,a,b Keisuke Wada,c Megumi Furukawa,a Katsumi Doid
Functional Genomics of Extremophiles, Faculty of Agriculture, Graduate School, Kyushu University, Fukuoka, Japana; Department of Chemistry and Biotechnology, GraduateSchool of Engineering, Tottori University, Tottori, Japanb; Applied Molecular Microbiology and Biomass Chemistry, Bioscience and Biotechnology, Faculty of Agriculture, KyushuUniversity, Fukuoka, Japanc; Microbial Genetic Division, Institute of Genetic Resources, Faculty of Agriculture, Kyushu University, Fukuoka, Japand
Thermostability is an important property of enzymes utilized for practical applications because it allows long-term storage anduse as catalysts. In this study, we constructed an error-prone strain of the thermophile Geobacillus kaustophilus HTA426 andinvestigated thermoadaptation-directed enzyme evolution using the strain. A mutation frequency assay using the antibioticsrifampin and streptomycin revealed that G. kaustophilus had substantially higher mutability than Escherichia coli and Bacillussubtilis. The predominant mutations in G. kaustophilus were A · T¡G · C and C · G¡T · A transitions, implying that the highmutability of G. kaustophilus was attributable in part to high-temperature-associated DNA damage during growth. Among thegenes that may be involved in DNA repair in G. kaustophilus, deletions of the mutSL, mutY, ung, and mfd genes markedly en-hanced mutability. These genes were subsequently deleted to construct an error-prone thermophile that showed much higher(700- to 9,000-fold) mutability than the parent strain. The error-prone strain was auxotrophic for uracil owing to the fact thatthe strain was deficient in the intrinsic pyrF gene. Although the strain harboring Bacillus subtilis pyrF was also essentially auxo-trophic, cells became prototrophic after 2 days of culture under uracil starvation, generating B. subtilis PyrF variants with anenhanced half-denaturation temperature of >10°C. These data suggest that this error-prone strain is a promising host for ther-moadaptation-directed evolution to generate thermostable variants from thermolabile enzymes.
Enzymes catalyze numerous reactions that are difficult to per-form using chemical catalysts and are commercially utilized in
industry (1). The starch industry is a good example; it has a greatdemand for �-amylase, �-amylase, glucoamylase, pullulanase,and glucose isomerase for producing glucose and its related sug-ars. In addition, biosensors for monitoring of human blood glu-cose levels use glucose oxidase or glucose dehydrogenase. Recentlydeveloped detergents also contain protease, �-amylase, �-amy-lase, lipase, and/or cellulase. Thus, enzymes have potential forcommercial use, but many are not practical despite possessingcatalytic activities suitable for use. One reason is enzyme thermo-lability, which hinders the prolonged use of enzymes as catalystsand their long-term storage at around room temperature. Al-though enzymes identified in extreme thermophiles show excel-lent thermostability, they generally show low activity at moderatetemperatures and accordingly possess low utility in applicationsthat require efficient enzymatic activity at moderate temperatures(e.g., as biosensors and in chemical production and medicalfields). Therefore, thermostability enhancement of thermolabileenzymes while maintaining catalytic activities at high levels atmoderate temperatures is an important approach to expandingthe commercial use of enzymes.
Thermostability enhancement has been achieved by two mainapproaches or their combination (2–4). One approach uses an insilico rational design to predict enzyme variants with enhancedthermostability (3). This approach requires three-dimensionalstructure information for target enzymes but allows the rapid pre-diction of candidates. The other approach uses in vitro randommutations to construct an enzyme variant library from which de-sired variants are screened. This approach is proven but may re-quire in vitro enzymatic characterization of numerous enzymevariants (4). To facilitate the screening process, thermophiles havebeen used as library hosts because they allow in vivo enzymatic
characterization at high temperatures without cell death. Exam-ples are identification of thermoadaptive variants of Escherichiacoli hygromycin B phosphotransferase (5), Streptoalloteichus hin-dustanus bleomycin-binding protein (6), Geobacillus stearother-mophilus �-galactosidase (7), Saccharomyces cerevisiae 3-isopropy-lmalate dehydrogenase (8), and Staphylococcus aureus kanamycinnucleotidyltransferase (9–11) in Thermus thermophilus and/or G.stearothermophilus. Intriguingly, some of these variants have beengenerated, not via in vitro random mutations, but via spontaneousmutations in thermophiles, termed thermoadaptation-directedenzyme evolution (5, 7–9).
Spontaneous mutations primarily occur through formation ofbase-base mismatches caused by replication errors and DNAdamage, such as deamination, 7,8-dihydro-8-oxoguanine (8-oxoG), and depurination lesions (12, 13). Deamination of ade-nine produces hypoxanthine and causes A · T¡G · C transitions(14, 15), whereas deamination of cytosine and 5-methylcytosineproduces U · G and T · G mismatches, respectively, leading to C ·G¡T · A transitions (12, 16–18). Oxidative damage to guanineproduces the aberrant base 8-oxoG, which causes the C · G¡A · T
Received 5 August 2014 Accepted 12 October 2014
Accepted manuscript posted online 17 October 2014
Citation Suzuki H, Kobayashi J, Wada K, Furukawa M, Doi K. 2015.Thermoadaptation-directed enzyme evolution in an error-prone thermophilederived from Geobacillus kaustophilus HTA426. Appl Environ Microbiol 81:149 –158. doi:10.1128/AEM.02577-14.
Editor: R. M. Kelly
Address correspondence to Hirokazu Suzuki, [email protected].
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
doi:10.1128/AEM.02577-14
January 2015 Volume 81 Number 1 aem.asm.org 149Applied and Environmental Microbiology
on August 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

transversion because of the ability to form base pairs with adenineand cytosine (19, 20). The 8-oxoG lesion is also observed in thenucleotide pool as 8-oxo-dGTP, from which 8-oxoG can bemisincorporated in opposing template adenines, causing theA · T¡C · G transversion (20). Replication errors and depurina-tion events can potentially cause any transversion or transition.
Because of the deleterious effects of mutations, base-base mis-matches are corrected by several DNA repair systems. In E. coli,mismatches arising from replication errors are corrected by methyl-dependent mismatch repair that involves MutS, MutL, and MutHproteins (21). Although most organisms lack MutH homologs,MutL from species that lack MutH harbors a latent endonucleaseactivity similar to that of MutH (22), suggesting a MutH-indepen-dent mechanism (23). Mismatches arising from DNA damage arecommonly corrected through base excision repair pathways,which are initiated by DNA glycosylases that excise damagedbases, such as Ung (uracil-DNA glycosylase) for uracil excision(17, 24), alkyladenine DNA glycosylase for hypoxanthine excision(16), MutM for 8-oxoG excision from C · 8-oxoG mismatches(20, 25, 26), and MutY for adenine excision from A · 8-oxoGmismatches (20, 25, 26). In addition, MutT homologs and tran-scription-coupled repair systems have been identified in severalorganisms. MutT degrades 8-oxo-dGTP to prevent 8-oxoG incor-poration during DNA replication (20), and transcription-coupledrepair corrects damage on the transcribed DNA strand, with pref-erence over nontranscribed strands and nontranscribed double-stranded DNA (27). In E. coli, transcription-coupled repair is me-diated by a transcription repair coupling factor encoded by mfd,which facilitates the removal of RNA polymerases stalled at DNAlesions that block transcriptional elongation and the recruitmentof DNA repair proteins to lesions on transcribed strands (27, 28).
Recently, we have studied genetic tools for the aerobic, Gram-positive, Bacillus-related thermophile Geobacillus kaustophilusHTA426 with the aim of using the strain for biotechnological ap-plications and biological studies of the genus Geobacillus (29–32).In these studies, HTA426 cells efficiently produced descendantsconstitutively resistant to the antibiotics rifampin (Rif) and strep-tomycin (Str). Because Rif and Str resistance (Rifr and Strr) oftenarise from spontaneous mutations in certain genes (33–38), thisobservation suggests that the HTA426 strain has considerable mu-tability. Thus, in the present study, we evaluated G. kaustophilusmutability in detail and constructed an error-prone strain fromthe thermophile. We further investigated the generation of en-zyme variants that are more thermostable than the parent enzymeusing the error-prone strain.
MATERIALS AND METHODSBacterial strains and media. Table 1 summarizes G. kaustophilus strains.G. kaustophilus MK242 was constructed from strain MK93 (29) using thesame procedure as for construction of MK244 from MK72 (31). If nototherwise specified, G. kaustophilus strains were grown at 60°C in Luria-Bertani (LB) medium and minimal medium (MM). MM consisted ofK2SO4 (0.3 g liter�1), Na2HPO4 · 12H2O (2.5 g liter�1), NH4Cl (1 gliter�1), MgSO4 (0.4 g liter�1), MnCl2 · 4H2O (3 mg liter�1), CaCl2 ·2H2O (5 mg liter�1), FeCl3 · ·6H2O (7 mg liter�1), 0.1% trace elementsolution (39), 10 mM Tris-HCl (pH 7.0), D-glucose (10 g liter�1), andCasamino Acids (1 g liter�1; Becton, Dickinson and Company, FranklinLakes, NJ). Kanamycin (5 mg liter�1) and uracil (10 mg liter�1) wereadded when necessary. E. coli DH5� (TaKaRa Bio, Otsu, Japan) andpCR4Blunto-TOPO (Life Technologies, Rockville, MD) were used forDNA manipulations. E. coli BL21(DE3) and pET-16b (Merck KGaA,Darmstadt, Germany) were used for recombinant-protein production. E.coli cells were grown at 37°C in LB medium. Ampicillin (50 mg liter�1)
TABLE 1 G. kaustophilus strains
Strain Relevant descriptiona
Doubling time(min)b
MK242 Derivative of the wild-type strain HTA426; �pyrF �pyrR �hsdM1S1R1 �(mcrB1-mcrB2-hsdM2S2R2-mrr)GK0707::Pgk704-bgaB
ND
MK480 Derivative of the strain MK242; �pyrF �pyrR �hsdM1S1R1 �(mcrB1-mcrB2-hsdM2S2R2-mrr) GK0707::Pgk704-bgaB �(mutS-mutL) �mutY �ung �mfd
ND
MK242p70 MK242 derivative; trpE::pGKE70 20.2 � 0.5MK480p70 MK480 derivative; trpE::pGKE70 21.8 � 0.5�mutSLp70 MK242 derivative; �(mutS-mutL) trpE::pGKE70 20.4 � 0.8�mutMp70 MK242 derivative; �mutM trpE::pGKE70 21.6 � 0.8�mutYp70 MK242 derivative; �mutY trpE::pGKE70 20.0 � 1.0�mutTp70 MK242 derivative; �mutT trpE::pGKE70 21.9 � 1.8�ungp70 MK242 derivative; �ung trpE::pGKE70 20.4 � 0.7�mfdp70 MK242 derivative; �mfd trpE::pGKE70 22.7 � 1.6�mutSLcm MK242 derivative; �(mutS-mutL) trpE::pGKE70-mutSL 21.9 � 0.3�mutMcm MK242 derivative; �mutM trpE::pGKE70-mutM 22.6 � 1.5�mutYcm MK242 derivative; �mutY trpE::pGKE70-mutY 22.9 � 1.0�mutTcm MK242 derivative; �mutT trpE::pGKE70-mutT 22.9 � 0.7�ungcm MK242 derivative; �ung trpE::pGKE70-ung 20.5 � 0.9�mfdcm MK242 derivative; �mfd trpE::pGKE70-mfd 20.7 � 0.7MK242BSpyrF MK242 derivative; trpE::pGKE70-BSpyrFwt NDMK480BSpyrF MK480 derivative; trpE::pGKE70-BSpyrFwt NDa �(mutS-mutL) encodes codons mutS1–106 fused with mutL534 – 630. �mutM, �mutY, �mutT, �ung, and �mfd are deleted in codons mutM11–255, mutY14 –347, mutT13–150,ung16 –218, and mfd33–1060, respectively. trpE is an internal gene for anthranilate synthase component I. The strains MK242 and MK480 lack the native pyrF gene encoding orotidine-5=-phosphate decarboxylase, but MK242BSpyrF and MK480BSpyrF harbor BSpyrFwt, which encodes orotidine-5=-phosphate decarboxylase in B. subtilis.b G. kaustophilus strains were cultured at 60°C in LB medium with monitoring of OD600. The binary logarithm of OD600 data during the logarithmic growth phase was plottedagainst incubation time to calculate doubling times. The data are presented as means � standard deviations (SD) (n � 3 to 16). ND, not determined.
Suzuki et al.
150 aem.asm.org January 2015 Volume 81 Number 1Applied and Environmental Microbiology
on August 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

was added when necessary. Bacillus subtilis 168 was obtained from theBacillus Genetic Stock Center (Columbus, OH) and grown in LB mediumat 37°C.
Construction of pGKE25 derivatives. Primer sequences are listed inTable 2. The plasmid constructs p�mutSL, p�mutM, p�mutY, p�mutT,p�ung, and p�mfd were used for in-frame deletions of the mutS-mutL,mutM, mutY, mutT, ung, and mfd genes, respectively, in G. kaustophilusMK242. For p�mutSL construction, the upstream region (1.5 kb) of mutS(GK1306) was amplified using primers 1306upF and 1306upR, and themutL (GK1307) downstream region (1.5 kb) was amplified using primers1307dwF and 1307dwR. The two amplified fragments were combinedusing overlap extension PCR and were cloned into the BamHI site ofpGKE25 (31) to yield p�mutSL. In addition, �mutM (GK2728), �mutY(GK0463), �mutT (GK3067), �ung (GK3421), and �mfd (GK0048) frag-ments were generated using appropriate primers and cloned into pGKE25to yield p�mutM, p�mutY, p�mutT, p�ung, and p�mfd, respectively.
Construction of pGKE70 derivatives. The plasmid pGKE70 (Fig. 1)was constructed to integrate genes into the trpE locus (GK2204) and to
force gene expression under the control of the Pgk704 promoter (29). Toconstruct pGKE70, the Pgk704 fragment was excised from pGAM48 (29)and the trpE internal sequence (positions 157 to 1360) was amplified usingprimers 2204F and 2204R. The Pgk704 fragment was cloned between theHindIII and SphI sites of the plasmid pTK19 (30), and the trpE fragmentwas then cloned in the MunI site to obtain pGKE70. To constructpGKE70-mutSL, the mutS-mutL and Pgk704 fragments were amplified us-ing primers 1306F and 1307R and P704F and 1306R, respectively. Thesetwo fragments were combined to create the Pgk704-mutSL fragment, whichwas trimmed using HindIII and BglII and cloned between the HindIII andBamHI sites of pGKE70 to generate pGKE70-mutSL. In addition, mutM,ung, and mfd were amplified and cloned between SphI and BamHI sites ofpGKE70 to generate pGKE70-mutM, pGKE70-ung, and pGKE70-mfd,respectively. mutY and mutT were amplified and trimmed using Sse8387Iand BamHI and subsequently cloned between the PstI and BamHI sitesof pGKE70 to generate pGKE70-mutY and pGKE70-mutT, respec-tively. The gene encoding orotidine-5=-phosphate decarboxylase of B.subtilis (BSpyrFwt) was amplified from the strain 168 chromosome and sub-
TABLE 2 Primers used in this study
Primer Sequence (5=-3=) Target region Underlined site
0048F GCCGCATGCTTTCGTTGCATCGCTATTTAG mfd full sequence SphI0048R GCCGGATCCTTATGCCGTCACCGACTTCTC mfd full sequence BamHI0048upF GCCGGATCCCTTCGTGAAGGCGGGCGCAC mfd upstream BamHI0048upR GACCATTTCCTCACGCAGCCCGGCGACAAGCTG mfd upstream0048dwF CAGCTTGTCGCCGGGCTGCGTGAGGAAATGGTC mfd downstream0048dwR GGCGGATCCCGACGATGCCATAAATCGGG mfd downstream BamHI0463F GCCCCTGCAGGCACAAGAGAAACAGAGCG mutY full sequence Sse8387I0463R GCCGGATCCTTAATCGGGGCGGCGTAC mutY full sequence BamHI0463upF GCCGGATCCCTTGGATTTGGCGGATGAAC mutY upstream BamHI0463upR CCAAACACGTTGATGAAACTCGCGCGCTGG mutY upstream0463dwF CCAGCGCGCGAGTTTCATCAACGTGTTTGG mutY downstream0463dwR GCCGGATCCACATTCGTCCATGCGTGCTC mutY downstream BamHI1306F GGTGTGTACGATGGCATAATCCCCCTCCTGTC mutSL full sequence1306R GACAGGAGGGGGATTATGCCATCGTACACACC Pgk704 fragment1306upF GCCGGATCCTCCACCAAAACGAAAAAGTG mutS upstream BamHI1306upR CCACGTCGGATGGGAGAGCTGCACAACTTC mutS upstream1307R GCCAGATCTCTACATCACCCGTTTAAATAG mutSL full sequence BglII1307dwF GAAGTTGTGCAGCTCTCCCATCCGACGTGG mutS downstream1307dwR GCCGGATCCATACCGCGTTTTCCAATAGG mutS downstream BamHI2204F GCCGAATTCTGACGAATCGCCGTGGGCGC trpE internal sequence EcoRI2204R GGCGAATTCCTGTCCGGATGGCGATGCAC trpE internal sequence EcoRI2728F GCCGCATGCCGGAATTGCCGGAGGTG mutM full sequence SphI2728R GCCGGATCCCTAGCGCTGGCAGCGCGGGC mutM full sequence BamHI2728upF GCCGGATCCGCCGGAGTGAAAGTGGATAC mutM upstream BamHI2728upR GACCGTTTTTTCAATGATCGTTTCCACCTC mutM upstream2728dwF GAGGTGGAAACGATCATTGAAAAAACGGTC mutM downstream2728dwR GCCGGATCCTAATCGCAGAATGCGGATTC mutM downstream BamHI3067F GCCCCTGCAGGCGTGAACGAATTGCAACGGG mutT full sequence Sse8387I3067R GCCGGATCCTCAGCTCGGATCGAGCCG mutT full sequence BamHI3067upF GCCGGATCCAATACATGTGGCGCCGACAG mutT upstream BamHI3067upR GAGCCGATATGAAAGGTTTGTCACCCGTTG mutT upstream3067dwF CAACGGGTGACAAACCTTTCATATCGGCTC mutT downstream3067dwR GCCGGATCCTTCGCCTTGCATACTTCCTG mutT downstream BamHI3421F GCCGCATGCCGATTCTCAAAAACGAC ung full sequence SphI3421R GCCGGATCCTCATTCAGCGCGGGCGCCGATG ung full sequence BamHI3421upF GCCGGATCCCTTGGATGTACCATGCTTTG ung upstream BamHI3421upR GTTCTCGATTTGCCACTCCTCTTCAAGCAG ung upstream3421dwF CTGCTTGAAGAGGAGTGGCAAATCGAGAAC ung downstream3421dwR GCCGGATCCTTTCATCCGATGCGACGTCC ung downstream BamHIP704F GCCAAGCTTTTTCTTTTTCCTCCTTTGTTATC Pgk704 fragment HindIIIpyrFF GTACATATGCGAAACAACCTGCCCATC BSpyrFwt sequence NdeIpyrFR GCCGGATCCTTAAGATTTGATTCCCTCCC BSpyrFwt sequence BamHI
Evolution in an Error-Prone Thermophile
January 2015 Volume 81 Number 1 aem.asm.org 151Applied and Environmental Microbiology
on August 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

cloned between the SphI and BamHI sites of pGKE70 to generate pGKE70-BSpyrFwt.
Plasmid introduction into G. kaustophilus. Derivatives of pGKE25and pGKE70 were introduced into G. kaustophilus cells using ternary con-jugative transfer from E. coli DH5� (31). Transconjugants integratingpGKE25 derivatives in the chromosome were selected using uracil prot-otrophy. Transconjugants integrating pGKE70 derivatives at the trpE lo-cus were selected using kanamycin resistance and tryptophan auxotrophy.
Construction of G. kaustophilus mutants deficient in DNA repairgenes. DNA repair genes in G. kaustophilus MK242 were deleted by two-step reciprocal crossovers using pyrF-based counterselection (30) withplasmids p�mutSL, p�mutM, p�mutY, p�mutT, p�ung, and p�mfd. Themutants were transformed with pGKE70, yielding the �mutSLp70,�mutMp70, �mutYp70, �mutTp70, �ungp70, and �mfdp70 mutants. StrainMK480 was constructed by simultaneous deletions of mutSL, mutY, ung,and mfd genes in MK242. Strains MK242 and MK480 were also trans-formed with pGKE70 to generate strains MK242p70 and MK480p70, re-spectively. For gene complementation, �mutSL, �mutM, �mutY, �mutT,�ung, and �mfd mutants were transformed with plasmids pGKE70-mutSL, pGKE70-mutM, pGKE70-mutY, pGKE70-mutT, pGKE70-ung,and pGKE70-mfd to generate the �mutSLcm, �mutMcm, �mutYcm,�mutTcm, �ungcm, and �mfdcm mutants, respectively.
Southern blotting. Total DNA was digested using restriction endonu-cleases and separated on a 0.8% (wt/vol) agarose gel by electrophoresis. DNAwas transferred to a nylon membrane and hybridized with digoxigenin(DIG)-labeled DNA probes that were synthesized using a PCR DIG Probesynthesis kit (Roche, Basel, Switzerland). Hybridized DNA was detected bythe chromogenic method using a DIG Nucleic Acid Detection Kit (Roche).
Mutation frequency assay. G. kaustophilus, E. coli, and B. subtilisstrains were cultured overnight in liquid LB medium and mixed withglycerol to a final concentration of 20%. The mixture was divided intoaliquots and stored at �80°C until use. Glycerol stocks were thawed, andan aliquot (106 cells containing �1 Rifr and Strr cell stochastically) wasinoculated into liquid LB medium (20 ml) in an Erlenmeyer flask (100 ml)with a silicon plug. The flask was incubated with shaking at 180 rpm, andthe optical density at 600 nm (OD600) was monitored using an OD-Moni-torA instrument (Taitec, Saitama, Japan). On reaching early stationaryphase (2 h after the OD600 reached 1.0), cells were spread on LB platescontaining efficacious Rif (10 and 50 mg liter�1 for G. kaustophilus and B.subtilis; 50 and 250 mg liter�1 for E. coli) and Str (10 and 50 mg liter�1 for
G. kaustophilus and E. coli; 500 and 3,000 mg liter�1 for B. subtilis). Cellswere also spread on LB plates without Rif or Str to determine viable-cellconcentrations. After subsequent incubation for 24 h, grown colonieswere counted, and the numbers of Rifr or Strr cells per 109 viable cells weredetermined.
Generation of BSpyrFe1 and BSpyrFe2. G. kaustophilus MK242and MK480 were transformed with pGKE70-BSpyrFwt to generateMK242BSpyrF and MK480BSpyrF, respectively. The strains were cultivated inliquid MM (without uracil) at 60°C for 24 h, and then the grown cells werecultivated at 65°C for 24 h. Cells were spread on MM plates and incubatedat 65°C to isolate single clones. From the clones, BSpyrFwt genes wereamplified and sequenced to identify the mutant genes BSpyrFe1 andBSpyrFe2.
Preparation of BSpyrFwt, BSpyrFe1, and BSpyrFe2 proteins. TheBSpyrFwt, BSpyrFe1, and BSpyrFe2 genes were amplified using primerspyrFF and pyrFR and cloned in pCR4Blunt-TOPO. After sequence check-ing, the genes were subcloned between the NdeI and BamHI sites of pET-16b. Using the resulting plasmids, E. coli BL21(DE3) was transformed andcultured at 37°C in liquid LB medium containing 1% lactose. After 6 h ofincubation, the cells were harvested and stored at �80°C until use. All ofthe subsequent operations were performed at 4°C. Cells were suspendedin buffer (20 mM sodium phosphate, 0.5 M NaCl, pH 7.4), followed bysonication. The cell lysates were clarified by centrifugation (18,000 g for10 min at 4°C) and applied to a Talon column (0.2 ml; TaKaRa Bio)equipped with the same buffer. The column was washed with buffer con-taining 20 mM imidazole (2 ml) followed by 50 mM imidazole (0.5 ml).Recombinant proteins were subsequently eluted with buffer containing200 mM imidazole and dialyzed against 10 mM sodium phosphate (pH7.0) with 0.5 M NaCl. The protein purity was evaluated by sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE).
Thermostability assay of BSpyrFwt, BSpyrFe1, and BSpyrFe2 pro-teins. For analysis of thermal denaturation curves, protein solutions (20l; 0.4 g liter�1 protein) were incubated at 30.0 to 55.6°C for 1 h using agradient temperature heater (Thermal Cycler Dice Gradient; TaKaRa Bio)and then centrifuged (18,000 g for 10 min at 4°C) to remove aggregatedproteins. Proteins retained in the supernatant were analyzed by the Brad-ford method using bovine serum albumin as a standard. T1/2, the temper-ature at 50% residual protein, was calculated by linear regression usingthermal denaturation data (BSpyrFwt, 35.6 to 45.6°C; BSpyrFe1, 48.0 to53.2°C; BSpyrFe2, 48.0 to 54.4°C). For time course analysis, protein solu-tions were incubated at 37°C for 7 days. After centrifugation, proteinsretained in the supernatant were analyzed by the Bradford method.
RESULTSG. kaustophilus mutability. G. kaustophilus MK242 was con-structed as a laboratory strain that lacks genes related to pyrimi-dine biosynthesis (pyrF and pyrR) and DNA restriction modifica-tion in the HTA426 strain. It was transformed with pGKE70 forcoordinating analytical conditions with other G. kaustophilus mu-tants (see below). The resulting mutant, MK242p70, was used toassess G. kaustophilus mutability (Fig. 2). The mutability assay wasbased on generation frequencies of Rifr and Strr cells, because Rifr
and Strr often arise from mutations in rpoB (encoding RNA poly-merase � subunit) and rpsL (encoding ribosomal subunit proteinS12) genes, respectively (33–38). The generation frequencies ofRifr cells (per 109 viable cells) from MK242p70 cells were 930 � 350(for 10 mg liter�1 Rif) and 520 � 270 (for 50 mg liter�1 Rif), andthose of Strr cells were 120 � 60 (for 10 mg liter�1 Str) and 70 � 40(for 50 mg liter�1 Str). These frequencies were higher than thoseof E. coli and B. subtilis, confirming substantial mutability of G.kaustophilus.
Mutability of G. kaustophilus mutants. The G. kaustophilusHTA426 genome (40) contains genes that may be involved inDNA repair (mutS, mutL, mutM, mutY, mutT, ung, and mfd). To
FIG 1 pGKE70 structure. pGKE70 was constructed to integrate genes into thetrpE locus and to force gene expression under the control of the Pgk704 pro-moter. AmpR, ampicillin resistance gene; pUC, pUC replicon; oriT, conjuga-tive-transfer origin; TK101, thermostable kanamycin nucleotidyltransferasegene; Pgk704, the Pgk704 promoter functional in G. kaustophilus (29); and trpE,an internal and defective gene for anthranilate synthase component I of strainHTA426. Restriction enzyme sites unique to multiple cloning sites are alsoindicated.
Suzuki et al.
152 aem.asm.org January 2015 Volume 81 Number 1Applied and Environmental Microbiology
on August 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from
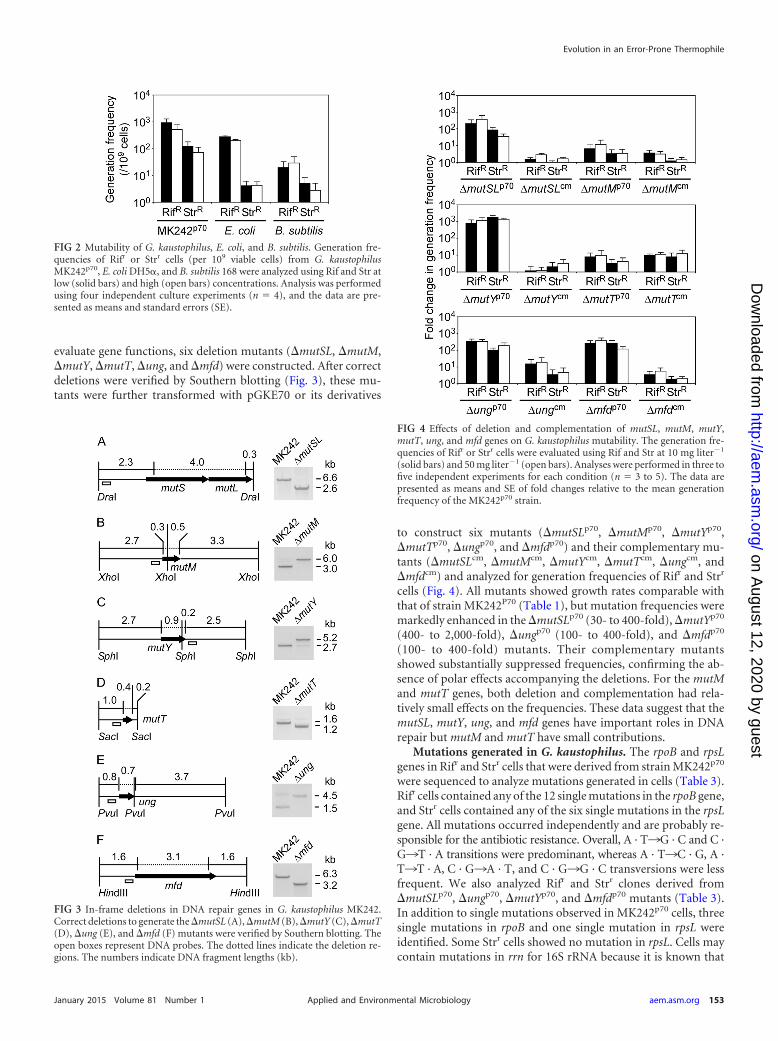
evaluate gene functions, six deletion mutants (�mutSL, �mutM,�mutY, �mutT, �ung, and �mfd) were constructed. After correctdeletions were verified by Southern blotting (Fig. 3), these mu-tants were further transformed with pGKE70 or its derivatives
to construct six mutants (�mutSLp70, �mutMp70, �mutYp70,�mutTp70, �ungp70, and �mfdp70) and their complementary mu-tants (�mutSLcm, �mutMcm, �mutYcm, �mutTcm, �ungcm, and�mfdcm) and analyzed for generation frequencies of Rifr and Strr
cells (Fig. 4). All mutants showed growth rates comparable withthat of strain MK242P70 (Table 1), but mutation frequencies weremarkedly enhanced in the �mutSLp70 (30- to 400-fold), �mutYp70
(400- to 2,000-fold), �ungp70 (100- to 400-fold), and �mfdp70
(100- to 400-fold) mutants. Their complementary mutantsshowed substantially suppressed frequencies, confirming the ab-sence of polar effects accompanying the deletions. For the mutMand mutT genes, both deletion and complementation had rela-tively small effects on the frequencies. These data suggest that themutSL, mutY, ung, and mfd genes have important roles in DNArepair but mutM and mutT have small contributions.
Mutations generated in G. kaustophilus. The rpoB and rpsLgenes in Rifr and Strr cells that were derived from strain MK242p70
were sequenced to analyze mutations generated in cells (Table 3).Rifr cells contained any of the 12 single mutations in the rpoB gene,and Strr cells contained any of the six single mutations in the rpsLgene. All mutations occurred independently and are probably re-sponsible for the antibiotic resistance. Overall, A · T¡G · C and C ·G¡T · A transitions were predominant, whereas A · T¡C · G, A ·T¡T · A, C · G¡A · T, and C · G¡G · C transversions were lessfrequent. We also analyzed Rifr and Strr clones derived from�mutSLp70, �ungp70, �mutYp70, and �mfdp70 mutants (Table 3).In addition to single mutations observed in MK242p70 cells, threesingle mutations in rpoB and one single mutation in rpsL wereidentified. Some Strr cells showed no mutation in rpsL. Cells maycontain mutations in rrn for 16S rRNA because it is known that
FIG 4 Effects of deletion and complementation of mutSL, mutM, mutY,mutT, ung, and mfd genes on G. kaustophilus mutability. The generation fre-quencies of Rifr or Strr cells were evaluated using Rif and Str at 10 mg liter�1
(solid bars) and 50 mg liter�1 (open bars). Analyses were performed in three tofive independent experiments for each condition (n � 3 to 5). The data arepresented as means and SE of fold changes relative to the mean generationfrequency of the MK242p70 strain.
FIG 2 Mutability of G. kaustophilus, E. coli, and B. subtilis. Generation fre-quencies of Rifr or Strr cells (per 109 viable cells) from G. kaustophilusMK242p70, E. coli DH5�, and B. subtilis 168 were analyzed using Rif and Str atlow (solid bars) and high (open bars) concentrations. Analysis was performedusing four independent culture experiments (n � 4), and the data are pre-sented as means and standard errors (SE).
FIG 3 In-frame deletions in DNA repair genes in G. kaustophilus MK242.Correct deletions to generate the �mutSL (A), �mutM (B), �mutY (C), �mutT(D), �ung (E), and �mfd (F) mutants were verified by Southern blotting. Theopen boxes represent DNA probes. The dotted lines indicate the deletion re-gions. The numbers indicate DNA fragment lengths (kb).
Evolution in an Error-Prone Thermophile
January 2015 Volume 81 Number 1 aem.asm.org 153Applied and Environmental Microbiology
on August 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from
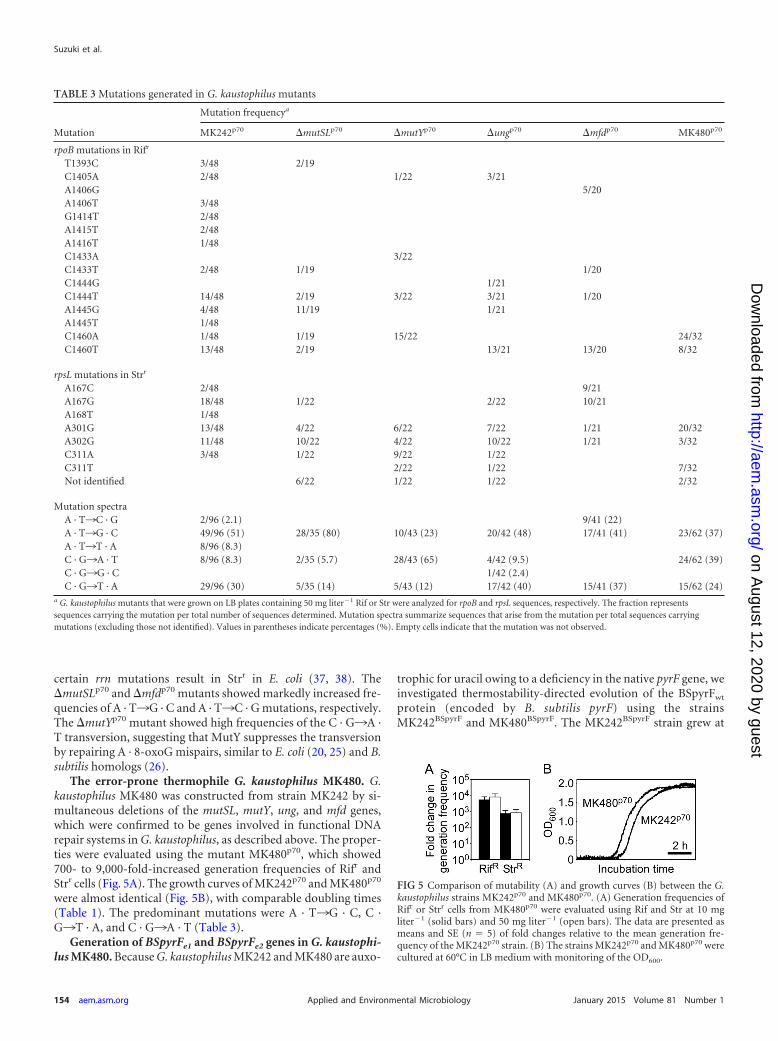
certain rrn mutations result in Strr in E. coli (37, 38). The�mutSLp70 and �mfdp70 mutants showed markedly increased fre-quencies of A · T¡G · C and A · T¡C · G mutations, respectively.The �mutYp70 mutant showed high frequencies of the C · G¡A ·T transversion, suggesting that MutY suppresses the transversionby repairing A · 8-oxoG mispairs, similar to E. coli (20, 25) and B.subtilis homologs (26).
The error-prone thermophile G. kaustophilus MK480. G.kaustophilus MK480 was constructed from strain MK242 by si-multaneous deletions of the mutSL, mutY, ung, and mfd genes,which were confirmed to be genes involved in functional DNArepair systems in G. kaustophilus, as described above. The proper-ties were evaluated using the mutant MK480p70, which showed700- to 9,000-fold-increased generation frequencies of Rifr andStrr cells (Fig. 5A). The growth curves of MK242p70 and MK480p70
were almost identical (Fig. 5B), with comparable doubling times(Table 1). The predominant mutations were A · T¡G · C, C ·G¡T · A, and C · G¡A · T (Table 3).
Generation of BSpyrFe1 and BSpyrFe2 genes in G. kaustophi-lus MK480. Because G. kaustophilus MK242 and MK480 are auxo-
trophic for uracil owing to a deficiency in the native pyrF gene, weinvestigated thermostability-directed evolution of the BSpyrFwt
protein (encoded by B. subtilis pyrF) using the strainsMK242BSpyrF and MK480BSpyrF. The MK242BSpyrF strain grew at
TABLE 3 Mutations generated in G. kaustophilus mutants
Mutation
Mutation frequencya
MK242p70 �mutSLp70 �mutYp70 �ungp70 �mfdp70 MK480p70
rpoB mutations in Rifr
T1393C 3/48 2/19C1405A 2/48 1/22 3/21A1406G 5/20A1406T 3/48G1414T 2/48A1415T 2/48A1416T 1/48C1433A 3/22C1433T 2/48 1/19 1/20C1444G 1/21C1444T 14/48 2/19 3/22 3/21 1/20A1445G 4/48 11/19 1/21A1445T 1/48C1460A 1/48 1/19 15/22 24/32C1460T 13/48 2/19 13/21 13/20 8/32
rpsL mutations in Strr
A167C 2/48 9/21A167G 18/48 1/22 2/22 10/21A168T 1/48A301G 13/48 4/22 6/22 7/22 1/21 20/32A302G 11/48 10/22 4/22 10/22 1/21 3/32C311A 3/48 1/22 9/22 1/22C311T 2/22 1/22 7/32Not identified 6/22 1/22 1/22 2/32
Mutation spectraA · T¡C · G 2/96 (2.1) 9/41 (22)A · T¡G · C 49/96 (51) 28/35 (80) 10/43 (23) 20/42 (48) 17/41 (41) 23/62 (37)A · T¡T · A 8/96 (8.3)C · G¡A · T 8/96 (8.3) 2/35 (5.7) 28/43 (65) 4/42 (9.5) 24/62 (39)C · G¡G · C 1/42 (2.4)C · G¡T · A 29/96 (30) 5/35 (14) 5/43 (12) 17/42 (40) 15/41 (37) 15/62 (24)
a G. kaustophilus mutants that were grown on LB plates containing 50 mg liter�1 Rif or Str were analyzed for rpoB and rpsL sequences, respectively. The fraction representssequences carrying the mutation per total number of sequences determined. Mutation spectra summarize sequences that arise from the mutation per total sequences carryingmutations (excluding those not identified). Values in parentheses indicate percentages (%). Empty cells indicate that the mutation was not observed.
FIG 5 Comparison of mutability (A) and growth curves (B) between the G.kaustophilus strains MK242p70 and MK480p70. (A) Generation frequencies ofRifr or Strr cells from MK480p70 were evaluated using Rif and Str at 10 mgliter�1 (solid bars) and 50 mg liter�1 (open bars). The data are presented asmeans and SE (n � 5) of fold changes relative to the mean generation fre-quency of the MK242p70 strain. (B) The strains MK242p70 and MK480p70 werecultured at 60°C in LB medium with monitoring of the OD600.
Suzuki et al.
154 aem.asm.org January 2015 Volume 81 Number 1Applied and Environmental Microbiology
on August 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from
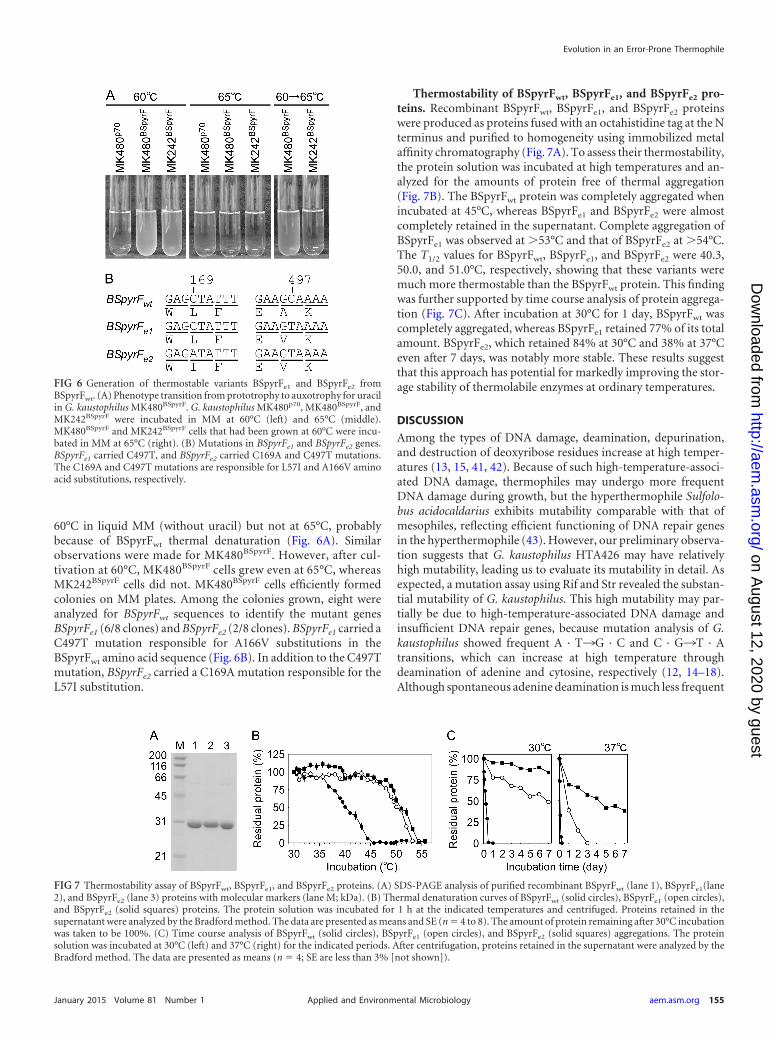
60°C in liquid MM (without uracil) but not at 65°C, probablybecause of BSpyrFwt thermal denaturation (Fig. 6A). Similarobservations were made for MK480BSpyrF. However, after cul-tivation at 60°C, MK480BSpyrF cells grew even at 65°C, whereasMK242BSpyrF cells did not. MK480BSpyrF cells efficiently formedcolonies on MM plates. Among the colonies grown, eight wereanalyzed for BSpyrFwt sequences to identify the mutant genesBSpyrFe1 (6/8 clones) and BSpyrFe2 (2/8 clones). BSpyrFe1 carried aC497T mutation responsible for A166V substitutions in theBSpyrFwt amino acid sequence (Fig. 6B). In addition to the C497Tmutation, BSpyrFe2 carried a C169A mutation responsible for theL57I substitution.
Thermostability of BSpyrFwt, BSpyrFe1, and BSpyrFe2 pro-teins. Recombinant BSpyrFwt, BSpyrFe1, and BSpyrFe2 proteinswere produced as proteins fused with an octahistidine tag at the Nterminus and purified to homogeneity using immobilized metalaffinity chromatography (Fig. 7A). To assess their thermostability,the protein solution was incubated at high temperatures and an-alyzed for the amounts of protein free of thermal aggregation(Fig. 7B). The BSpyrFwt protein was completely aggregated whenincubated at 45°C, whereas BSpyrFe1 and BSpyrFe2 were almostcompletely retained in the supernatant. Complete aggregation ofBSpyrFe1 was observed at �53°C and that of BSpyrFe2 at �54°C.The T1/2 values for BSpyrFwt, BSpyrFe1, and BSpyrFe2 were 40.3,50.0, and 51.0°C, respectively, showing that these variants weremuch more thermostable than the BSpyrFwt protein. This findingwas further supported by time course analysis of protein aggrega-tion (Fig. 7C). After incubation at 30°C for 1 day, BSpyrFwt wascompletely aggregated, whereas BSpyrFe1 retained 77% of its totalamount. BSpyrFe2, which retained 84% at 30°C and 38% at 37°Ceven after 7 days, was notably more stable. These results suggestthat this approach has potential for markedly improving the stor-age stability of thermolabile enzymes at ordinary temperatures.
DISCUSSION
Among the types of DNA damage, deamination, depurination,and destruction of deoxyribose residues increase at high temper-atures (13, 15, 41, 42). Because of such high-temperature-associ-ated DNA damage, thermophiles may undergo more frequentDNA damage during growth, but the hyperthermophile Sulfolo-bus acidocaldarius exhibits mutability comparable with that ofmesophiles, reflecting efficient functioning of DNA repair genesin the hyperthermophile (43). However, our preliminary observa-tion suggests that G. kaustophilus HTA426 may have relativelyhigh mutability, leading us to evaluate its mutability in detail. Asexpected, a mutation assay using Rif and Str revealed the substan-tial mutability of G. kaustophilus. This high mutability may par-tially be due to high-temperature-associated DNA damage andinsufficient DNA repair genes, because mutation analysis of G.kaustophilus showed frequent A · T¡G · C and C · G¡T · Atransitions, which can increase at high temperature throughdeamination of adenine and cytosine, respectively (12, 14–18).Although spontaneous adenine deamination is much less frequent
FIG 6 Generation of thermostable variants BSpyrFe1 and BSpyrFe2 fromBSpyrFwt. (A) Phenotype transition from prototrophy to auxotrophy for uracilin G. kaustophilus MK480BSpyrF. G. kaustophilus MK480p70, MK480BSpyrF, andMK242BSpyrF were incubated in MM at 60°C (left) and 65°C (middle).MK480BSpyrF and MK242BSpyrF cells that had been grown at 60°C were incu-bated in MM at 65°C (right). (B) Mutations in BSpyrFe1 and BSpyrFe2 genes.BSpyrFe1 carried C497T, and BSpyrFe2 carried C169A and C497T mutations.The C169A and C497T mutations are responsible for L57I and A166V aminoacid substitutions, respectively.
FIG 7 Thermostability assay of BSpyrFwt, BSpyrFe1, and BSpyrFe2 proteins. (A) SDS-PAGE analysis of purified recombinant BSpyrFwt (lane 1), BSpyrFe1(lane2), and BSpyrFe2 (lane 3) proteins with molecular markers (lane M; kDa). (B) Thermal denaturation curves of BSpyrFwt (solid circles), BSpyrFe1 (open circles),and BSpyrFe2 (solid squares) proteins. The protein solution was incubated for 1 h at the indicated temperatures and centrifuged. Proteins retained in thesupernatant were analyzed by the Bradford method. The data are presented as means and SE (n � 4 to 8). The amount of protein remaining after 30°C incubationwas taken to be 100%. (C) Time course analysis of BSpyrFwt (solid circles), BSpyrFe1 (open circles), and BSpyrFe2 (solid squares) aggregations. The proteinsolution was incubated at 30°C (left) and 37°C (right) for the indicated periods. After centrifugation, proteins retained in the supernatant were analyzed by theBradford method. The data are presented as means (n � 4; SE are less than 3% [not shown]).
Evolution in an Error-Prone Thermophile
January 2015 Volume 81 Number 1 aem.asm.org 155Applied and Environmental Microbiology
on August 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

than cytosine deamination in general (15, 16), A · T¡G · C tran-sition was more frequent than C · G¡T · A transition in G. kaus-tophilus. In the �mutSLp70 mutant, the frequency of A · T¡G · Cwas as high as 80%. These observations imply a defect in deami-nated-adenine (i.e., hypoxanthine) repair in G. kaustophilus. Inaddition, gene deletion analysis indicated small contributions ofmutM and mutT to DNA repair. In E. coli (20) and B. subtilis (26),MutM suppresses the C · G¡A · T transversion cooperatively withMutY, and MutT suppresses A · T¡C · G transversion. AlthoughMutY and Mfd contribute to suppressing these transversions in G.kaustophilus (Table 3), it is likely that G. kaustophilus is exposed torelatively high risks of the transversions.
We constructed G. kaustophilus MK480 as an error-pronethermophile by deleting functional DNA repair genes that wehad identified and investigated thermoadaptive gene muta-tions in the error-prone strain using BSpyrFwt as a model. G.kaustophilus MK480BSpyrF was originally auxotrophic for uracilat 65°C but became prototrophic after culture in MM at 60°C,followed by 65°C, along with BSpyrFe1 and BSpyrFe2 genera-tion. This phenotype transition can be explained by generationand selection, during incubation at 60°C, of mutant genes thatencode enzyme variants that are more thermostable than theBSpyrFwt protein. The thermostability assay confirmed that thevariants BSpyrFe1 and BSpyrFe2 are actually more thermostablethan BSpyrFwt and that BSpyrFe2 is more thermostable thanBSpyrFe1. These observations indicate that both A166T andL57I substitutions contribute to BSpyrFwt thermostabilization.In the (�/�)8 barrel fold of the BSpyrFwt protein (44), the res-idues L57 (replaced in BSpyrFe2) and A166 (replaced inBSpyrFe1 and BSpyrFe2) constitute �-strand �3 and �-helix �2,respectively (Fig. 8). The side chain of L57 hydrophobicallyinteracts with the side chains of the F39, V47, and L59 residuesto occupy the void between �-strand �3 and �-helix �2 (Fig.8A). Similar hydrophobic interactions were observed for theA166 residue, the side chain of which interacts with Y170 andT179 to occupy the void between �-helix �6 and �-strand �7(Fig. 8B). Therefore, it is possible that thermostability en-hancement by L57I and A166V substitutions involves optimiz-ing hydrophobic interactions between �-helices and �-strandsin the barrel fold, thereby making the structure robust.
Thermoadaptation-directed evolution using strain MK480 re-quires no construction of random mutant libraries because mu-tations are generated in cells during successive culture. Althoughthe mutations may be less frequent than mutations obtained usingin vitro mutagenesis approaches, one amino acid alteration poten-tially confers sufficient enhancement of enzyme thermostability,as shown by BSpyrFe1 and other examples (45). Moreover, strain
MK480 can employ in vivo enzymatic characterization at hightemperatures to select thermoadaptive enzymes, as described be-low: (i) when cell growth is accelerated by catalytic activity oftarget enzymes, candidate genes encoding thermoadaptive en-zymes can be concentrated, because thermolabile enzymes un-dergo thermal denaturation during cell growth at high tempera-tures, as in BSpyrFe1 and BSpyrFe2 generation, and (ii) whencatalytic activities of target enzymes can be detected by chromo-genic or fluorogenic assay in vivo, candidate genes can be selectedby a plate assay without cell death. Even in other cases, candidategenes may be efficiently selected by the activity-independentmethod described by Chautard et al. (46). The availability of high-temperature in vivo assays is a notable advantage of strain MK480over error-prone mesophiles for thermoadaptation-directed evo-lution. In addition, strain MK480 showed 700- to 9,000-fold-in-creased mutability. This enhancement is comparable to thatachieved by abolishing (5,000-fold) or silencing (2,000-fold) DNArepair genes in E. coli (47, 48), while G. kaustophilus had higherintrinsic mutability than E. coli (Fig. 2). Therefore, G. kaustophilusMK480 has higher mutability than E. coli strains engineered forincreased mutability, an additional advantage over error-pronemesophiles.
There are some reports of thermoadaptation-directed en-zyme evolution in thermophiles (5, 7–9), although not using anerror-prone thermophile. A 3-isopropylmalate dehydrogenasevariant having five substitutions with a 12°C-higher T1/2 thanthe parent enzyme was generated by iterative incubation forapproximately 10 days in T. thermophilus (8). T. thermophiluscells were also used to generate an �-galactosidase variant pos-sessing one substitution with an enhanced T1/2 of 3°C by 11days of incubation (7). Compared with these examples, strainMK480 generated thermoadaptive variants with excellent ther-mostability enhancement (�10°C) in a short time (2 days),probably due to high mutability. These data suggest that strainMK480 has an advantage over not only error-prone mesoph-iles, but also thermophiles used for thermoadaptation-directedevolution to date. It is also noteworthy that in strain MK480,every mutation can theoretically occur, given that although theA · T¡C · G, A · T¡T · A, and C · G¡G · C mutations were notobserved in the strain, they were found in the parent strain(MK242p70) or the �ung mutant (�ungp70). Moreover, G. kaus-tophilus allows a wide range of selection temperatures (42 to74°C), which is especially favorable for thermoadaptation-di-rected evolution of very thermolabile enzymes. Thus, strainMK480 is a practical resource for generating thermostable en-zyme variants by thermoadaptation-directed evolution and providesnew opportunities for the facile generation of thermostable enzymes.
ACKNOWLEDGMENTS
This work was supported by the Programme for Promotion of Basicand Applied Researches for Innovations in Bio-Oriented Industry(BRAIN), Japan, and in part by the Institute for Fermentation, Osaka,Japan.
REFERENCES1. Haki GD, Rakshit SK. 2003. Developments in industrially important
thermostable enzymes: a review. Bioresour Technol 89:17–34. http://dx.doi.org/10.1016/S0960-8524(03)00033-6.
2. Bommarius AS, Broering JM, Chaparro-Riggers JF, Polizzi KM. 2006.High-throughput screening for enhanced protein stability. Curr OpinBiotechnol 17:606 – 610. http://dx.doi.org/10.1016/j.copbio.2006.10.001.
FIG 8 Three-dimensional structures surrounding the L57 (A) and A166 (B)residues of the BSpyrFwt protein.
Suzuki et al.
156 aem.asm.org January 2015 Volume 81 Number 1Applied and Environmental Microbiology
on August 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

3. Chen J, Yu H, Liu C, Liu J, Shen Z. 2012. Improving stability of nitrilehydratase by bridging the salt-bridges in specific thermal-sensitive re-gions. J Biotechnol 164:354 –362. http://dx.doi.org/10.1016/j.jbiotec.2013.01.021.
4. Steffler F, Guterl J-K, Sieber V. 2013. Improvement of thermostablealdehyde dehydrogenase by directed evolution for application in syntheticcascade biomanufacturing. Enzyme Microb Technol 53:307–314. http://dx.doi.org/10.1016/j.enzmictec.2013.07.002.
5. Nakamura A, Takakura Y, Kobayashi H, Hoshino T. 2005. In vivodirected evolution for thermostabilization of Escherichia coli hygromycinB phosphotransferase and the use of the gene as a selection marker in thehost-vector system of Thermus thermophilus. J Biosci Bioeng 100:158 –163. http://dx.doi.org/10.1263/jbb.100.158.
6. Brouns SJJ, Wu H, Akerboom J, Turnbull AP, de Vos WM, van der OostJ. 2005. Engineering a selectable marker for hyperthermophiles. J Biol Chem280:11422–11431. http://dx.doi.org/10.1074/jbc.M413623200.
7. Fridjonsson O, Watzlawick H, Mattes R. 2002. Thermoadaptation of�-galactosidase AgaB1 in Thermus thermophilus. J Bacteriol 184:3385–3391. http://dx.doi.org/10.1128/JB.184.12.3385-3391.2002.
8. Tamakoshi M, Nakano Y, Kakizawa S, Yamagishi A, Oshima T.2001. Selection of stabilized 3-isopropylmalate dehydrogenase of Sac-charomyces cerevisiae using the host-vector system of an extreme thermo-phile, Thermus thermophilus. Extremophiles 5:17–22. http://dx.doi.org/10.1007/s007920000168.
9. Liao H, McKenzie T, Hageman R. 1986. Isolation of a thermostableenzyme variant by cloning and selection in a thermophile. Proc Natl AcadSci U S A 83:576 –580. http://dx.doi.org/10.1073/pnas.83.3.576.
10. Hoseki J, Yano T, Koyama Y, Kuramitsu S, Kagamiyama H. 1999.Directed evolution of thermostable kanamycin-resistance gene: a conve-nient selection marker for Thermus thermophilus. J Biochem 126:951–956.http://dx.doi.org/10.1093/oxfordjournals.jbchem.a022539.
11. Matsumura M, Aiba S. 1985. Screening for thermostable mutant of ka-namycin nucleotidyltransferase by the use of a transformation system fora thermophile, Bacillus stearothermophilus. J Biol Chem 260:15298 –15303.
12. Smith KC. 1992. Spontaneous mutagenesis: experimental, genetic andother factors. Mutat Res 277:139 –162. http://dx.doi.org/10.1016/0165-1110(92)90002-Q.
13. Lindahl T. 1993. Instability and decay of the primary structure of DNA.Nature 362:709 –715. http://dx.doi.org/10.1038/362709a0.
14. Parry TE. 2007. On the mutagenic action of adenine. Leuk Res 31:1621–1624. http://dx.doi.org/10.1016/j.leukres.2007.05.002.
15. Karran P, Lindahl T. 1980. Hypoxanthine in deoxyribonucleic acid:generation by heat-induced hydrolysis of adenine residues and release infree form by a deoxyribonucleic acid glycosylase from calf thymus. Bio-chemistry 19:6005– 6011. http://dx.doi.org/10.1021/bi00567a010.
16. Wanner RM, Castor D, Güthlein C, Böttger EC, Springer B, Jiricny J.2009. The uracil DNA glycosylase UdgB of Mycobacterium smegmatis pro-tects the organism from the mutagenic effects of cytosine and adeninedeamination. J Bacteriol 191:6312– 6319. http://dx.doi.org/10.1128/JB.00613-09.
17. López-Olmos K, Hernández MP, Contreras-Garduño JA, Robleto EA,Setlow P, Yasbin RE, Pedraza-Reyes M. 2012. Roles of endonuclease V,uracil-DNA glycosylase, and mismatch repair in Bacillus subtilis DNAbase-deamination-induced mutagenesis. J Bacteriol 194:243–252. http://dx.doi.org/10.1128/JB.06082-11.
18. Krokan HE, Drabløs F, Slupphaug G. 2002. Uracil in DNA—occurrence, consequences and repair. Oncogene 21:8935– 8948. http://dx.doi.org/10.1038/sj.onc.1205996.
19. Shibutani S, Takeshita M, Grollman AP. 1991. Insertion of specific basesduring DNA synthesis past the oxidation-damaged base 8-oxodG. Nature349:431– 434. http://dx.doi.org/10.1038/349431a0.
20. Tajiri T, Maki H, Sekiguchi M. 1995. Functional cooperation of MutT,MutM and MutY proteins in preventing mutations caused by spontane-ous oxidation of guanine nucleotide in Escherichia coli. Mutat Res 336:257–267. http://dx.doi.org/10.1016/0921-8777(94)00062-B.
21. Kunkel TA, Erie DA. 2005. DNA mismatch repair. Annu Rev Biochem74:681–710. http://dx.doi.org/10.1146/annurev.biochem.74.082803.133243.
22. Pillon MC, Lorenowicz JJ, Uckelmann M, Klocko AD, Mitchell RR,Chung YS, Modrich P, Walker GC, Simmons LA, Friedhoff P,Guarné A. 2010. Structure of the endonuclease domain of MutL: un-licensed to cut. Mol Cell 39:145–151. http://dx.doi.org/10.1016/j.molcel.2010.06.027.
23. Simmons LA, Davies BW, Grossman AD, Walker GC. 2008. Beta clampdirects localization of mismatch repair in Bacillus subtilis. Mol Cell 29:291–301. http://dx.doi.org/10.1016/j.molcel.2007.10.036.
24. Lindahl T. 1974. An N-glycosidase from Escherichia coli that releases freeuracil from DNA containing deaminated cytosine residues. Proc NatlAcad Sci U S A 71:3649 –3653. http://dx.doi.org/10.1073/pnas.71.9.3649.
25. Michaels ML, Cruz C, Grollman AP, Miller JH. 1992. Evidence thatMutY and MutM combine to prevent mutations by an oxidatively dam-aged form of guanine in DNA. Proc Natl Acad Sci U S A 89:7022–7025.http://dx.doi.org/10.1073/pnas.89.15.7022.
26. Sasaki M, Kurusu Y. 2004. Analysis of spontaneous base substitutionsgenerated in mutator strains of Bacillus subtilis. FEMS Microbiol Lett 234:37– 42. http://dx.doi.org/10.1111/j.1574-6968.2004.tb09510.x.
27. Selby CP, Witkin EM, Sancar A. 1991. Escherichia coli mfd mutantdeficient in “mutation frequency decline” lacks strand-specific repair: invitro complementation with purified coupling factor. Proc Natl Acad Sci US A 88:11574 –11578. http://dx.doi.org/10.1073/pnas.88.24.11574.
28. Park J-S, Marr MT, Roberts JW. 2002. E. coli transcription repair cou-pling factor (Mfd protein) rescues arrested complexes by promoting for-ward translocation. Cell 109:757–767. http://dx.doi.org/10.1016/S0092-8674(02)00769-9.
29. Suzuki H, Yoshida K, Ohshima T. 2013. Polysaccharide-degrading ther-mophiles generated by heterologous gene expression in Geobacillus kaus-tophilus HTA426. Appl Environ Microbiol 79:5151–5158. http://dx.doi.org/10.1128/AEM.01506-13.
30. Suzuki H, Murakami A, Yoshida K. 2012. Counterselection system forGeobacillus kaustophilus HTA426 through disruption of pyrF and pyrR.Appl Environ Microbiol 78:7376 –7383. http://dx.doi.org/10.1128/AEM.01669-12.
31. Suzuki H, Wada K, Furukawa M, Doi K, Ohshima T. 2013. A ternaryconjugation system for the construction of DNA libraries for Geobacilluskaustophilus HTA426. Biosci Biotechnol Biochem 77:2316 –2318. http://dx.doi.org/10.1271/bbb.130492.
32. Suzuki H, Yoshida K. 2012. Genetic transformation of Geobacillus kaus-tophilus HTA426 by conjugative transfer of host-mimicking plasmids. JMicrobiol Biotechnol 22:1279 –1287. http://dx.doi.org/10.4014/jmb.1203.03023.
33. Jin DJ, Gross CA. 1988. Mapping and sequencing of mutations in theEscherichia coli rpoB gene that lead to rifampicin resistance. J Mol Biol202:45–58. http://dx.doi.org/10.1016/0022-2836(88)90517-7.
34. Nicholson WL, Maughan H. 2002. The spectrum of spontaneous rifam-pin resistance mutations in the rpoB gene of Bacillus subtilis 168 sporesdiffers from that of vegetative cells and resembles that of Mycobacteriumtuberculosis. J Bacteriol 184:4936 – 4940. http://dx.doi.org/10.1128/JB.184.17.4936-4940.2002.
35. Noguchi N, Sasatsu M, Kono M. 1993. Genetic mapping in Bacillussubtilis 168 of the aadK gene which encodes aminoglycoside 6-adenylyl-transferase. FEMS Microbiol Lett 114:47–52. http://dx.doi.org/10.1111/j.1574-6968.1993.tb06549.x.
36. Timms AR, Steingrimsdottir H, Lehmann AR, Bridges BA. 1992. Mu-tant sequences in the rpsL gene of Escherichia coli B/r: mechanistic impli-cations for spontaneous and ultraviolet light mutagenesis. Mol Gen Genet232:89 –96. http://dx.doi.org/10.1007/BF00299141.
37. Montandon PE, Wagner R, Stutz E. 1986. Escherichia coli ribosomes witha C912 to U base change in the 16S rRNA are streptomycin resistant.EMBO J 5:3705–3708.
38. Melançon P, Lemieux C, Brakier-Gingras L. 1988. A mutation in the 530loop of Escherichia coli 16S ribosomal RNA causes resistance to strepto-mycin. Nucleic Acids Res 16:9631–9639. http://dx.doi.org/10.1093/nar/16.20.9631.
39. Amartey SA, Leak DJ, Hartley BS. 1991. Development and optimization ofa defined medium for aerobic growth of Bacillus stearothermophilus LLD-15.Biotechnol Lett 13:621–626. http://dx.doi.org/10.1007/BF01086315.
40. Takami H, Takaki Y, Chee G-J, Nishi S, Shimamura S, Suzuki H,Matsui S, Uchiyama I. 2004. Thermoadaptation trait revealed by thegenome sequence of thermophilic Geobacillus kaustophilus. Nucleic AcidsRes 32:6292– 6303. http://dx.doi.org/10.1093/nar/gkh970.
41. Lindahl T, Nyberg B. 1972. Rate of depurination of native deoxyribo-nucleic acid. Biochemistry 11:3610 –3618. http://dx.doi.org/10.1021/bi00769a018.
42. Lindahl T, Nyberg B. 1974. Heat-induced deamination of cytosine resi-dues in deoxyribonucleic acid. Biochemistry 13:3405–3410. http://dx.doi.org/10.1021/bi00713a035.
Evolution in an Error-Prone Thermophile
January 2015 Volume 81 Number 1 aem.asm.org 157Applied and Environmental Microbiology
on August 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

43. Jacobs KL, Grogan DW. 1997. Rates of spontaneous mutation in anarchaeon from geothermal environments. J Bacteriol 179:3298 –3303.
44. Appleby TC, Kinsland C, Begley TP, Ealick SE. 2000. The crystal structureand mechanism of orotidine 5=-monophosphate decarboxylase. Proc NatlAcad Sci U S A 97:2005–2010. http://dx.doi.org/10.1073/pnas.259441296.
45. Imanaka T. 1990. Enhancement of thermostability of neutral proteases.Ann N Y Acad Sci 613:347–351. http://dx.doi.org/10.1111/j.1749-6632.1990.tb18176.x.
46. Chautard H, Blas-Galindo E, Menguy T, Grand’Moursel L, Cava F,
Berenguer J, Delcourt M. 2007. An activity-independent selection systemof thermostable protein variants. Nat Methods 4:919 –921. http://dx.doi.org/10.1038/nmeth1090.
47. Greener A, Callahan M, Jerpseth B. 1997. An efficient random mutagen-esis technique using an E. coli mutator strain. Mol Biotechnol 7:189 –195.http://dx.doi.org/10.1007/BF02761755.
48. Nakashima N, Tamura T. 2009. Conditional gene silencing of multiplegenes with antisense RNAs and generation of a mutator strain of Esche-richia coli. Nucleic Acids Res 37:e103. http://dx.doi.org/10.1093/nar/gkp498.
Suzuki et al.
158 aem.asm.org January 2015 Volume 81 Number 1Applied and Environmental Microbiology
on August 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from





























