Embed Size (px)
Citation preview

1
TRABAJO DE INVESTIGACIÓN PARA OPTAR AL
DIPLOMA DE ESTUDIOS AVANZADOS
“Estudios fármaco-genéticos de pacientes con infección por VIH-1
tratados con regimenes antirretrovirales basados en estavudina:
asociación de los polimorfismos de la timidilato sintasa con los niveles
intracelulares de estavudina trifosfato.”
Investigador Principal: Mª del Carmen Cabeza Brasa
Director del trabajo: Dr. Pere Domingo Pedrol
Universitat de Barcelona/ Departament de Medicina Interna
Junio 2010

2
RESUMEN…………………………………………………………………………………. 2
INTRODUCCIÓN………………………………………………………………………..... 4
MATERIAL Y MÉTODOS……………………………………………………………….. 7
RESULTADOS…………………………………………………………………………….. 9
DISCUSIÓN .………………………………………………………………………………. 12
CONCLUSIONES…………………………………………………………………………. 15
BIBLIOGRAFÍA…………………………………………………………………………… 18
TABLAS…………………………………………………………………………… .. 24

3
RESUMEN
Antecedentes: La actividad antirretroviral y la toxicidad de estavudina (d4T) depende
de su metabolito trifosfato, la estavudina trifosfato (d4T-TP). Por tanto, las
modificaciones en los niveles intracelulares de d4T pueden cambiar el perfil de la
toxicidad de la estavudina.
Métodos: Se determinaron los lniveles intracelulares de d4T-TP en células
mononucleares de sangre periférica (PBMC) por la prominencia en el cromatógrafo
líquido conectado a un espectrómetro de masa triple-cuádruple. Los polimorfismos en
los genes de la Timidilato sintasa (TS), Metilenetetrahidrofolato reductasa (MTHFR),
dihidrofolatoreductasa (DHFR), Transportador de folato reducido 1 (RFC1) y ciclina
D1 (CCND1) fueron determinados por secuencia directa usando ABI PRISM 3100
Genetic Analyzer o bien usando Fluidigm’s Biomark system. Para los análisis
estadísticos se usaron el test de la t de Student, el índice de correlación de Pearson,
ANOVA de un factor con correcciones por el método de Bonferroni y una regresión
logística escalonada.
Resultados: Se reclutaron 33 pacientes para este estudio transversal. Los niveles
intracelulares de d4T-TP fueron de 11.50 (RIC: 5.75) fmol/106 cels en pacientes con un
genotipo de alta expresión del TS (*2/*3G, *3C/ *3G and *3G/ 3G) mientras que en
aquellos con un genotipo de baja expresión (*2/ *2, *2/ *3C and *3C/ *3C), los niveles
fueron de 20.65 (12.70) fmol/106 cels (P = 0.0010). Los polimorfismos de los genes de
MTHFR, DHFR, RFC1 y CCDN1 no influyeron en la concentración intracelular de
d4T-TP
Conclusiones: Los niveles intracelulares de d4T_TP son determinados por los
polimorfismos de los genes de timidilato sintasa.
Palabras clave: estavudina, estavudina trifosfato, timidalato sintasa,
metilenetetrahidrofolato reductasa, dihifolato reductasa, ciclina D1, Toxicidad, pools de
nucleótidos, vía metabólica folato, fármaco-genética

4
INTRODUCCIÓN
El síndrome de asociado al tratamiento antirretroviral de gran actividad (TARGA)
lipodistrofia (HALS) es un serio problema de salud en pacientes infectados por el virus
de inmunodeficiencia tipo 1 (VIH-1) con una prevalencia estimada de 24-60% entre los
pacientes tratados [1-3]. Además, en estos pacientes, a menudo se asocia a dislipidemia,
resistencia a la insulina e hipertensión arterial, representando un escenario de riesgo
cardiovascular elevado en estos pacientes [4].
Aunque su patogénesis no es completamente conocida, las teorías de toxicidad por
fármacos han sido consideradas de las más plausibles, y la visión actual sugiere que los
inhibidores de la transcriptasa inversa análogos de los nucleósidos (ITIANs)
contribuyen a este síndrome [5, 6]. Los análogos timidínicos como la zidovudina (AZT)
y estavudina (d4T) has sido los principalmente implicados en el desarrollo de HALS en
ensayos clínicos randomizados [7, 8], y hay un importante número de evidencias en
estudios in vitro y ex vivo que unen el uso de estos fármacos con la aparición de efectos
tóxicos mediados por toxicidad mitocondrial [9-11]. Se sabe que las concentraciones de
ITIAN- trifosfato influyen tanto en el tratamiento antirretroviral como en los efectos
tóxicos, éste último mediante inhibición competitiva de la ADN polimerasa in vivo [12-
14]. Este mecanismo de toxicidad es un problema selectivo y específico, y por lo tanto,
dependerá de manera muy importante, de la dosis del fármaco y de su concentración.
Por esto, elevaciones en las concentraciones de ITIAN trifosfato influirán de manera
significativa en la toxicidad de los ITIANs.
La timidilato sintetasa (TS) es una enzima del metabolismo de las pirimidinas que
cataliza el paso de uridina a timidina para la formación del ADN. Si administramos
nucleótidos exógenos como el d4T que es un análogo de la timidina, favorecemos el uso
de éstos frente a los nucleótidos naturales. La producción del ADN tanto mitocondrial
como celular se llevará a cabo mayoritariamente con los nucleótidos exógenos.
La Timidilato sintasa (TS, N5, N10-metiletilenetetrahidrofolato: dUMP C-
metiltransferasa; EC 2.1.1.45) cataliza de manera no reversible la metilación de
deoxiuridina-5-monofosfato (dUMP) a deoxitimidina-5-monofosfato (dTMP), un
precursor de la síntesis de DNA. Por lo tanto, es un enzima clave en la síntesis de novo
de timidilato y el único enzima responsable de proporcionar los nucleótidos de timina
necesarios para la síntesis de DNA [15]. El gen que codifica este enzima es polimórfico,
teniendo repeticiones dobles (2R) o triples (3R) de una secuencia de 28bp en la región
promotora y se ha demostrado que las células homocigotas 3R/3R sobreexpresan

5
ARNm de la TS comparadas con las células homocigotas 2R/2R [15, 16]. Más
recientemente, se ha descrito un polimorfismo de nucleótido simple G>C en el
nucleótido 20º de la segunda repetición del alelo 3R, que encabeza un locus tri-alélico
(2R, 3RG, y 3RC) [17]. El alelo 3RC muestra una actividad transcripcional similar a la
del alelo 2R.
La TS es un enzima clave en el metabolismo de folato. En esta vía metabólica, otros
enzimas como el transportador de folato reducido 1 (RFC1), la metilenetetra-
hidrofolato reductasa (MTHFR) y la dihidrofolato reductasa (DHFR) también pueden
regular la actividad de la TS de una manera coordinada para un metabolismo eficiente
de folato. Adicionalmente, el incremento del nivel de de transcripción de varios de los
genes anteriormente mencionados (TS, DHFR) se puede asociar con la trascripción de
los factores E2F-1 y DP-1. El CCND1 interviene en la fosforilación de la proteína del
retinoblastoma (RB), liberando estas moléculas, parece necesario en este marco el
estudio del genotipo CCND1. El RFC1, un intercambiador de aniones, es una proteína
transmembrana que transfiere folato hidrofílico a través de la membrana celular. Los
genes que codifican para la proteína RFC1 están localizados en la región 21q. Estudios
recientes han sugerido que el polimorfismo G80A en el RFC1 se asocia con niveles
alterados de folato [18]. La MTHFR cataliza la conversión de 5,10-
metilenetetrahidrofolato a 5-metiltetrahidrofolato. Esta reacción enzimática genera
residuos de metilo esenciales para la función de la TS. El gen de la MTHFR se localiza
en el cromosoma 1p y los polimorfismos mejor caracterizados consisten en i) una
transición 677C>T que resulta en una sustitución de alanina por valina en el
denominado dominio catalítico de MTHFR, y ii) una transición 1298 A>C que resulta
en la sustitución de glutamina por alanina en el supuesto dominio regulador [19]. La
DHFR convierte el dihidrofolato (DHF) generado por la acción de la TS en
tetrahidrofolato (THF). El gen de la DHFR se localiza en el cromosoma 5q. Un
polimorfismo recientemente descrito que consiste en delección/inserción 19 bp que se
encuentra en el intrón 1 ha demostrado influir en las alteraciones de los niveles de folato
[20], y existen 3 polimorfismos en el promotor que fueron analizados (C-1610G/T; C-
680A; A-317G) [21]. El gen de la ciclina D1 (CCND1) se sitúa en 11q13 y codifica la
proteína ciclina D1 que juega un papel importante en la regulación del ciclo celular a
través de sus interacciones con las kinasas dependientes de ciclina. EL CCND1
interviene en la fosforilación de la proteína del retinoblastoma en la transición de la fase
G1 a la S, liberando E2F1 y DP-1 que aumentan la transcripción de varios genes

6
implicados en la replicación del DNA (TS, DHFR). El polimorfismo 870A>G del gen
CCND1 afecta al lugar de la unión con el donante en el límite entre exon4/intrón 4,
modulando la relación entre las isoformas del ARNm del CCND1 [22].
Nuestra hipótesis de trabajo es que las variaciones en los genes que intervienen en las
vías metabólicas anteriormente mencionadas pueden influir en las concentraciones de
niveles intracelulares de d4T trifosfato (d4T-TP) y de esta manera modular los efectos
tóxicos de este fármaco por la alteración de los pools de nucleótidos. Por lo tanto, el
objetivo del presente estudio es encontrar una relación entre los polimorfismos
genéticos de los genes que codifican los enzimas TS, MTHFR, DHFR, RFC1 y CCND1
y los niveles intracelulares de d4T-TP.
MATERIAL Y MÉTODOS
Población estudiada
Todos los pacientes fueron reclutados en la misma clínica en el Hospital de la Santa
Creu i Sant Pau, que atiende a una población de 1457 pacientes con infección por VIH-
1 en seguimiento activo, y eran pacientes con un diagnóstico establecido de infección
por VIH-1 en tratamiento. Los pacientes eran candidatos elegibles si tenían o no
síndrome de lipodistrofia asociado al tratamiento antirretroviral de gran actividad
(HALS) y estaban recibiendo estavudina (d4T) como parte de sus regímenes
antirretrovirales. Los pacientes que estaban hospitalizados o presentaban un deterioro
cognitivo franco como delirium o demencia durante el reclutamiento, no eran elegibles.
Los pacientes con enfermedades oportunistas, neoplasias o fiebre de origen desconocido
también eran excluidos del estudio. También eran excluidos aquellos pacientes que en el
momento del reclutamiento estaban tomando algún fármaco que influyera en el
metabolismo de la glucosa o en la distribución grasa como hormonas anabolizantes o
corticoides sistémicos, hormona del crecimiento recombinante, o estimuladores del
apetito. Se obtuvo consentimiento informado de todos los pacientes cuando entraban a
formar parte del estudio. El diagnóstico de síndrome de inmunodeficiencia humana
(SIDA) se basó en la definición de caso de Centers for Disease Control and Prevention
(CDC) [23] El estudio fue aprobado por el Comité Ético del Hospital de la Santa Creu i
Sant Pau.

7
Medidas de la composición corporal
Los sujetos fueron pesados en básculas calibradas después de quitarse los zapatos, la
ropa de calle y otros artículos pesados. El Índice de masa corporal (BMI, siglas en
inglés), se calculó dividiendo el peso en kilogramos por el cuadrado de la altura en
metros. La cintura se midió en centímetros usando referencias anatómicas definidas en
el “Third National Health and Nutrition Evaluation Survey” [24].
Las DEXAs (dual energy X-ray absorptiometry) de cuerpo entero (Hologic QDR-
4500A Hologic, INc, 590 Lincoln St, Waltham, MA 02154, USA) fueron realizados por
un único operador en todos los pacientes. Se determinó el porcentaje de grasa en los
brazos, piernas y abdomen central (calculado de la masa de grasa versus la masa magra
y la ósea) así como la masa magra corporal total en kilogramos.
Definición de HALS y síndrome metabólico
La presencia o ausencia de lipoatrofia, lipohipertrofia y síndrome mixto fue determinada
según las definiciones del Lipodystrophy Scale Severity Score [referencia del CID, 25].
El síndrome metabólico fue definido de acuerdo a las guías de U.S. National
Cholesterol Education Program (NCEP) Adult Treatment Panel III [26] y
modificaciones recomendadas en el último American Heart Association/National Heart,
Lung, and Blood Institute Scientific Statement [27]. El síndrome metabólico fue
definido como presentar tres o más de los siguientes factores de riesgo metabólico: 1)
obesidad central (circunferencia de la cintura ≥ 80 cm en mujeres y ≥ 90 cm en
hombres); 2) hipertrigliceridemia (triglicéridos en ayuno ≥ 1.69 mmol/l); 3) colesterol
HDL bajo (HDL < 1.29 mmol/l en mujeres y < 1.04 mmol/l en hombres); 4)
hiperglicemia (glucosa en ayunas ≥ 5.6 mmol/l o estar tomando antidiabéticos orales
para el tratamiento de la diabetes tipo 2; e hipertensión (presión arterial sentado, tomada
como media de 2 lecturas después de descansar al menos 10 min, o tomar medicación
antihipertensiva de forma regular).
Medidas del laboratorio de bioquímica
Todos los análisis bioquímicos se realizaron después de un ayuno nocturno de 12 h y al
menos 15 minutos después de la colocación de una vía periférica.
Los pacientes estuvieron sentados durante la extracción sanguínea y evitaron fumar al
menos durante los 15 minutos anteriores a la extracción; el torniquete venoso se evitó
siempre que fue posible y si fue necesario, se mantuvo menos de un minuto. Todas las

8
medidas lipídicas se realizaron con un sistema Hitachi 911 de Roche (Basel, Suiza). El
colesterol total y los triglicéridos se midieron por un método enzimático estandarizado y
el colesterol de las lipoproteínas de alta densidad (HDL) mediante un método directo
usando enzimas de polietilenglicol modificados PEGME) [28]. La fracción de
colesterol de las lipoproteínas de baja densidad se midió después de una
ultracentrifugación de acuerdo con el método recomendado por el Lipid Reserch Clinic,
pero utilizando el método PEGME para el colesterol-HDL en vez de precipitación.
El colesterol de la fracción VLDL fue medido como la fracción de d<1006 Kg/l de la
ultracentrifgación [29]. La resistencia a la insulina se halló por el HOMA-IR
(homeostasis model assessment method) como el producto de las concentraciones de
insulina plasmática (µunidades/ml ) y glucosa plasmática (mmol/l) en ayunas, dividido
entre 22.5 [30]. El valor de corte usado para definir la resistencia a la insulina fue de 3.8
[31]
Medidas de la concentración intracelular de d4T trifosfato
El d4T es de Moravek Biochemicals (CA, USA) y el Cl-ATP (2-cloroadenosin 5'-
trifosfato) usado como estándar interno es de Sigma-Aldrich (St Quentin-Fallavier,
France). En resumen, después de obtener las muestras (sobre 7 mL), las células
mononucleares de sangre periférica (PBMC) serán preparadas mediante centrifugación
con un medio de gradientes (Ficoll Histopaque 1077, Sigma) e inmediatamente serán
almacenados a una temperatura alrededor de -80º, pendientes del análisis. Las PBMC
se lisarán en 1 ml de metanol congelado/ Tris 0.05 M/HCl pH = 5; 70/30 (v/v) que
contendrá IS, y después de la evaporación del metanol, la fracción 40-mL de la solución
remanente se inyectara en el sistema LC-MS/MS system.
El LC-MS/MS consiste en una prominencia cromatógrafo líquido (Shimadzu, Champs
sur Marne, France) conectado a un espectrómetro de masa triple-cuádruple TSQ
Quantum Discovery (Thermo Fisher Scientific, Les Ulis, France) funcionando en el
modo negativo ESI para la detección de ambos d4T – TP e IS [32]. El método
cromatográfico líquido se usará de acuerdo a un método publicado previamente [33]. En
estas condiciones, los tiempos de retención serán de entre 3.95 y 4.05 minutos para d4T-
TP e IS respectivamente. La cantidad de d4T será determinada dentro del rango
calibrado de entre 50 a 3000 femtomoles (fmol) por célula. Finalmente, las CMSP de
cada muestra serán contadas usando un test bioquímico validado tal y como ha sido
descrito previamente [34] para obtener los resultados en mol / 106 PBM

9
Análisis genotípicos
El ADN genómico fue extraído de los leucocitos periféricos mediante el procedimiento
de precipitación de proteína por saturación de la solución con sales neutras [35]. En los
genes de la Timidilato Sintasa (TS) se analizaron un numero variable de repeticiones en
tándem (VNTR) del polimorfismo de 28 bp y el polimorfismo de nucleótido simple
G>C en la primera y segunda repetición. Un fragmento de DNA fue amplificado usando
las condiciones y los primers de la PCR (reacción en cadena de la polimerasa) [15], y
directamente secuenciado usando un analizador ABI PRISM 3100 Genetic Analyzer
(Applied Biosystems, Foster City, CA, USA). Estos cambios, sustituyendo G por C en
un residuo crítico en el elemento de consenso USF E-box, abole la unión de USF-1 y
altera la actividad transcripcional (de la transcriptasa). Los genotipos TS de los
pacientes se clasificaron de acuerdo con Kawakami y Watanabe en 2 grupos: el tipo de
alta expresión (*2/*3G, *3C/ *3G y *3G/ 3G) y el de baja expresión (*2/ *2, *2/ *3C y
*3C/ *3C) [36].
Se analizaron los siguientes polimorfismos en los genes de la dihifolato-reductasa
(DHPR): i) C-1610G/T se determinó por secuencia directa usando un analizador ABI
PRISM 3100 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA); ii) la
inserción/delección en 19bp usando un método de PCR convencional; iii) la sustitución
A-317G y iv) el cambio de C-680ª. Estos dos polimorfismos se analizaron usando un
sistema Fluidigm’s Biomark.
El sistema Fluidigm’s Biomark está diseñado para la discriminación alélica de los
análisis de la 5’ nucleasa. Las muestras y los análisis de la expresión génica TaqMan
Gene (Applied Biosystems, Foster City, CA, USA) se prepararon siguiendo las
instrucciones de los fabricantes. Las matrices dinámicas 48.48 usadas fueron
descargadas usando un IFC Controller (Fluidigm Corporation), y las reacciones a
tiempo real se realizaron y analizaron usando BioMark Real-Time PCR System and
Analysis software (Fluidigm Corporation), respectivamente. Como control de calidad,
se incluyeron muestras heterocigotas y homocigotas normalizadas en cada matriz y para
cada genotipo.
El polimorfismo A80G en el gen RFC1, así como los 2 marcadores (C677T y A1298T)
en el gen de la metil-tetra hidrofolatoreductasa (MTHFR) y el polimorfismo en el gen
CCND1 se analizaron usando la plataforma anteriormente descrita basada en matrices
dinámicas microfluidas.

10
Análisis estadísticos
Los datos se expresaron como medias ± desviación estándar (SD) o medianas con rengo
intercuartílico. Los datos que no siguieron una distribución normal, se determinaron
usando el test Kolmogorov-Smirnov donde fueron transformados logarítmicamente
antes del análisis. El test t de Student se usó para la comparación entre 2 grupos, el
índice de correlación de Pearson o ANOVA con un solo factor y múltiples pruebas se
corrigieron usando el método de Bonferroni. Las frecuencias de los alelos y el genotipo
se compararon entre poblaciones con el test del chi-cuadrado. Todos los valores de P se
definieron de manera bilateral; se consideraron estadísticamente significativos aquellos
análisis con un valor de P menor de 0.05. Todos los análisis se realizaron con SPSS
versión 17.0 (SPSS, Chicago, IL). Los análisis de regresión logística escalonada se
usaron para examinar la asociación entre los niveles de d4T intracelulares y otros
parámetros con los polimorfismos TS. Las variables seleccionadas para esta regresión
escalonada fueron aquellas que se correlacionaban de manera significativa con los
polimorfismos TS (después del método de Bonferroni para múltiples pruebas).
RESULTADOS
Población estudiada
Se reclutaron 33 pacientes caucásicos con infección por el VIH-1 en tratamiento basado
en estavudina. Los datos demográficos y el estatus viro-inmunológico de la población
estudiada se muestran en la Tabla 1. De ellos 27 eran hombres (81.1%) y 6 mujeres
(18.2%) con una media de edad de 45.1 ± 8.4 años (mediana 44.0 [IQR: 7 años]). Las
vías de adquisición de la infección por el VIH-1 fueron: hombres homosexuales (13,
39.4%), uso de drogas por vía parenteral (12, 36.4%), y contacto heterosexual (8,
24.2%). La media de duración de la infección por VIH-1 era de 13.9 ± 5.2 años
(mediana: 13.0 [IQR: 7 años]). Trece pacientes (39.4%) habían presentado previamente
una enfermedad definitoria de SIDA. Once pacientes estaban coinfectados con el virus
de la hepatitis C (33.3%), mientras que únicamente tres pacientes (9.1%) tenían
infección crónica por el virus de la hepatitis B. Sólo dos pacientes (9.1%) estaban
tomando al mismo tiempo sulfamidas y ningún participante tomaba ninguna otra
medicación aparte de los antirretrovirales.
Distribución de los genotipos

11
Los 33 pacientes que cumplían los criterios de inclusión fueron genotipados. En la tabla
2 se muestran las frecuencias de los alelos *2, *3C y *3G de la TS, los alelos C677T y
A1298T de la metilenetetrahidrofolato reductasa (MTHFR), los alelos 19 bp ins/del, A-
317G, C-680A, y C-1610G de la dihidrofolato reductasa (DHFR) y el A80G del
transportador de folato reducido 1 (RFC1)
Con respecto a los genotipos de la TS, 15 pacientes (45.4%) presentaban genotipos de
alta expresión (2/3G, 3C/3G, 3G/3G) y 18 presentaban genotipos de baja expresión (2/2,
2/3C, 3C/3C). La correlación entre el genotipo de la TS y los datos demográficos,
antropométricos, metabólicos, sobre la distribución grasa y las variables viro-
inmunológicas se muestran en la tabla 3.
Fármacos antirretrovirales y situación inmuno-virológica
La mayor parte de los pacientes (28, 84.8%) tenían una carga viral indetectable en el
momento del estudio. La media de la carga viral plasmática de los cinco pacientes que
tenían carga viral detectable, era de 215 copias/ml (rango: 109-473 copies/ml). La
media del recuento de CD4 era de 627 ± 311 cels/mm3 (mediana: 502 [IQR. 408]
cels/mm3). La cifra de CD4 nadir fue < 100 cels/mm3 en 15 pacientes (45.4%), y < 200
cels/mm3 en 21 pacientes (63.4%). La carga viral máxima estaba fue de más de 5 log10
en 15 pacientes (45.4%). La exposición acumulada a fármacos antirretrovirales
relacionada con el genotipo de la TS se muestra en la tabla 4
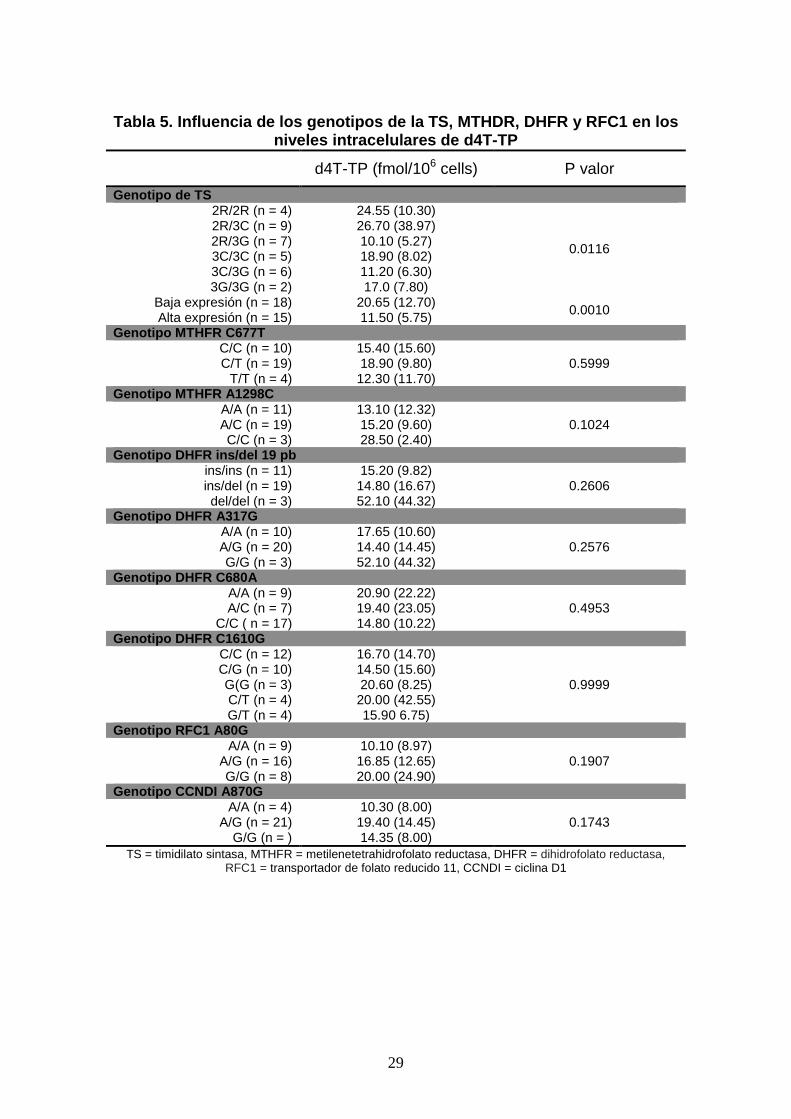
Concentraciones intracelulares de d4T-TP y genotipos
La mediana de las concentraciones de d4T-TP fue de 17 fmol/106 cels (rango
intercuartílico [IQR]: 11.47 fmol/106 cels). La mediana de las concentraciones de d4T-
TP por pacientes con diferente genortipo se muestra en la tabla 5, donde se puede
observar que el genotipo de la TS influencia de manera individual los niveles de d4T-
TP intracelulares de forma estadísticamente significativa. Sin embargo, combinaciones
de genotipos de TS con otros genotipos pueden maximizar o minimizar las diferencias
en los niveles de d4T-TP intracelulares. Así mismo, con respecto a los genotipos de la
MTHFR, la combinación de un genotipo de baja expresión de TS con la MTHFR 677
C/T y MTHFR 1298 A/A o C/C, dieron niveles de d4T-TP de 20.70 (IQR: 24.00)
fmol/106 cels/l, mientras que en la otra parte del espectro, individuos con la
combinación de un genotipo de alta expresión de TS con MTHFR 677 T/T y MTHFR
1298 A/C tenían unos niveles de d4T-TP de 11.50 (IQR: 6.10) fmol/106 cels/l.

12
Para la DHFR, la combinación de genotipos que causan niveles más elevados de d4T
intracelulares eran aquellos de baja expresión para la TS y DHFR del/del, -317 G/G, -
680 C/A y -1610 C/T, con 73.0 fmol/106 cels/l, mientras que en el lado opuesto estaba
la combinación de genotipos de alta expresión de TS y DHFR ins/ins, -317 A/A con
unos niveles de d4T-TP 9.90 (IQR: 2.95) fmol/106 cels/l. ,
Para la RFC1, la combinación de genotipos que causan niveles elevados de d4T-TP eran
aquellos de baja expresión de la TS y RFC1 G/G (20.60 [49.50] fmol/106 cels/l),
mientras que los niveles bajos de d4T-TP intracelular se relacionaban con los genotipos
de alta expresión de la TS y RFC1 A/A (8.65 [2.30] fmol/106 cels/l).
DISCUSIÓN
La fármacogenética es de aplicación reciente en el campo de la terapia antirretroviral.
Hasta ahora se han descrito un número de polimorfismos que modifican el metabolismo
de los fármacos y eventualmente su perfil tóxico. El polimorfismo G5516T en el
CYP2B6 se asoció con unas concentraciones plasmáticas de efavirez aproximadamente
3 veces más altas durante al menos las primeras 24 semanas de tratamiento
antirretroviral y con mayor número de efectos secundarios en el sistema nervioso central
[38,39]. Una frecuencia mayor de este polimorfismo en afro-americanos parece ser la
explicación por la que esta población presenta un menor aclaramiento de efavirenz [40,
41]. Sobre el 5-10% de la población general tiene un defecto en la actividad de la
conjugación de la bilirrubina conocido como síndrome de Gilbert [42,43], causado por
la inserción TA en la secuencia natural (salvaje) A(TA)6TAA del promotor UGT1A1.
La homocigosidad OK de A(TA)7TAA del genotipo del síndrome de Gilbert reduce la
actividad de conjugación de la bilirrubina en un 50% [44]. La hiperbilirrubinemia a
expensas de la bilirrubina no conjugada ocurre a menudo durante el tratamiento con
inhibidores de la proteasa del VIH-1 indinavir y atazanavir [45, 46]. Estos fármacos
compiten con la bilirrubina para unirse a UGT1A1. Aunque habitualmente es
asintomático, algunos pacientes desarrollan una ictericia moderada que es causa de que
abandonen el tratamiento.
El factor de necrosis tumoral (TNF)-α parece que está involucrado en la patogénesis de
la lipodistrofia [47]. Los niveles elevados de TNF-α en el tejido adiposo parecen estar
relacionados con lipodistrofia y el TNF-α puede influir en la diferenciación de los
adipocitos y en la sensibilidad a la insulina [48, 49]. Una variante en la posición
238G/A del alelo del TNF-α se encontró presente en el 15% de los pacientes con

13
lipodistrofia pero no en aquellos sujetos sin lipodistrofia [50]. De manera similar, entre
191 pacientes blancos en Australia, de todos aquellos que presentaban lipoatrofia [51],
el inicio de la pérdida de grasa era más rápido en aquellos que llevaban la variante del
alelo 238 G/A.
La tubulopatía renal proximal ha sido asociada significativamente con una sustitución
simple G>A en la posición 1249 de ABCC2 (también llamado “MRP2”, que codifica la
proteína que media la resistencia a múltiples fármacos [MRP] 2) y con un haplotipo
ABCC2 comparando 4 polimorfismos, incluyendo 1249 G>A [52]. Además, una
sustitución T>A en la posición 3563 y un haplotipo que incluye este polimorfismo, se
relacionaron con la ausencia de toxicidad renal [52].
Nuestro estudio muestra que los polimorfismos funcionales de los genes que codifican
la TS pueden tener implicación cuando el paciente recibe tratamiento antirretroviral que
contiene estavudina. Estos polimorfismos pueden causar variaciones substanciales y
significantes en las concentraciones de d4T-TP, que es la fracción farmacológica activa
(y tóxica) de la estavudina dado que inhibe tanto la transcriptasa inversa del VIH-1[53,
54] y la DNA polimerasa – α de los mamíferos [55]. Hemos encontrado también que
los diferentes polimorfismos que codifican la MTHFR, DHFR, RFC1 y CCND1 no
tienen un impacto significativo en los niveles intracelulares de d4T-TP cuando se
consideran de manera individual, pero se comportan a coadyuvantes con los
polimorfismos de la TS.
Las concentraciones intracelulares de d4T-TP son fidedignas ya que el método
cuantitativo usado presentaba media de precisión (rango) y media de exactitud (SD) de
9.8% (7.1-14.4) y 10.16% (10.0), respectivamente, sobre el rango completo de
calibración (valores de control de calidad de los ensayos). Además, los resultados
obtenidos en este estudio son comparables con los datos previamente publicados [56-
58].
La TS es un enzima clave en la vía de los nucleótidos biosintéticos que metila la
deoxiuridina (dUMP) para producir deoxytimidina (dTMP), y la reacción de la TS es la
única fuente de timidilato en la célula y es esencial para la replicación del ADN [15].
La variabilidad de la expresión de la TS puede estar relacionada con un número
variable de polimorfismos en las repeticiones en tándem (VNTR) de la región
promotora. Recientemente se ha descrito un cambio común G>C (SNP) en la segunda
repetición de los alelos que contiene 3 repeticiones; por lo tanto, la combinación de
SNP y VNRT permite la definición de la diferencia. Los alelos TS que muestran

14
distintos patrones de expresión génica. Nosotros encontramos que los polimorfismos
unidos a enzimas de baja expresión se asocian con un aumento de los niveles de d4T-
TP, lo que puede tener consecuencias en términos de toxicidad, puesto que como un
análogo timidínoco, el d4T es un inhibidor competitivo con el 2’-deoxytimidin-5’-
trifosfato (dTTP) por la polimerasa [59].
Un paso clave en la fisiopatología de la toxicidad y la farmacología de los NRTI es la
regulación del tamaño del pool de desoxirribonucleótidos naturales en la mitocondria
que afecta la replicación del ADN. Por el contrario, la disregulación en la fosforilación
y defosforilación podría alterar la replicación del ADNmt
Se esperaría que los genotipos de la TS asociados con una reducción de la actividad de
la enzima incrementaran la ventaja competitiva del d4T-TP con respecto al pool
(disminuido) del dTTP endógeno [60]. Más aún, los pools disminuidos de dTTP
estimularían la actividad de la timidin kinasa debido al feedback por inhibición del
producto final de esta vía metabólica [61]. Esto puede conllevar un pronunciado
aumento de la ratio intracelular NRTI- trifosfatos/dTP. Hay una evidencia
farmacológica de que los agentes que inhiben la biosíntesis de novo de los nucleótidos
tienen la habilidad de incrementar la fosforilación de d4T [62].
La hidroxiurea, un inhibidor de ribonucleótido- reductasa que también actúa
disminuyendo los pools de dTTP puede causar similares efectos en la fosforilación del
d4T [63]. De manera interesante, la combinación de hidroxiurea con d4T se ha asociado
con un exceso de toxicidad, especialmente pancreatitis y neuropatía periférica, en
diferentes estudios [64-66]. Los diferentes genotipos de los VNRT en la región
promotora de la TS (2R/2R, 2R/3R, y 3R/3R) ocurren a diferentes frecuencias en
diferentes poblaciones. En los caucásicos, es heterocigota aproximadamente el 50% de
la población, y cada uno de los genotipos homocigotos se encuentran en
aproximadamente en el 25% de la población. La distribución es igual en la población
negra y en la población del sudoeste asiático [67, 68]. Sin embargo, los sujetos
homocigotos de la triple repetición eran 2 veces más frecuentes en la población china
(67%) que en los sujetos caucásicos (38%) [67]. Esto puede explicar porqué la
lipodistrofia asociada a la infección por VIH-1, toxicidad que ha sido relacionada
directamente con el uso de análogos timidínicos usados ampliamente por todo el
mundo, ha sido tan escasamente reportada en los países asiáticos del Lejano Oriente
(Norte de China y Japón) [69].

15
La MTHFR cataliza una conversión irreversible de 5-10-metilenetetrahidrofolato a 5-
metiltetrahidrofolato. El paso anterior es fundamental en la síntesis del ADN porque
actúa como cofactor en la conversión de dUMP a dTMP por la TS [70]. Polimorfismos
en los genes de la MTHFR pueden influir en los pools intracelulares de folato, y por
tanto, la actividad de la TS puede modificarse. Hasta el momento se han reconocido 2
polimorfismos C677T, y A1298C, ambos asociados con un descenso de la actividad de
la enzima [71]. Hemos examinado ambos polimorfismos y aunque ninguno de ellos
individualmente determina cambios significativos en los niveles intracelulares de d4T-
TP, ambos modulan los efectos de los genotipos de la TS en mayor o menor medida.
Otros enzimas implicados en el metabolismo del folato, y consecuentemente con un
potencial impacto sobre el funcionalismo de la TS, son la DHFR y la RFC1. La DHFR
está involucrada en la reducción del dihidrofolato, generado durante la síntesis del
timidilato, a tetrahidrofolato para mantener cantidades adecuadas de folato para la
síntesis de DNA y re-metilación de la homocisteína [72]. Hay diversos polimorfismos
de la DHFR descritos. Entre ellos, nosotros hemos analizado 19-bp ins/del, -317 A>G, -
680 C>A, y -1610 C>G [20, 21]. De manera similar a los polimorfismos de la MTHFR,
ninguno de ellos modificaba significativamente los niveles intracelulares de d4T,
aunque sí modulan los efectos de los polimorfismos de la TS. El RFC1, también
llamado SLC19A1, es un transportador esencial de folato y funciona como un
intercambiador de aniones bidireccional, tomando cofactores del folato y exportando
diversos aniones orgánicos. El polimorfismo G80A en el gen del RFC1 ha mostrado
estar asociado con alteraciones del folato y de la homocisteina en sujetos sanos [73].
Encontramos que esta variación genética no inducía modificaciones sustanciales en los
niveles intracelulares de d4T.
También consideramos la implicación de un sistema biológico que regula la
disponibilidad en la célula de 2 factores de transcripción (DP-1 y E2F-1) que aumentan
la transcripción de entre otros, los genes de la TS, de la DHFR y los de la timidin-
kinasa: uno de los miembros de la familia de las ciclinas, CCND1, que junto con las
kinasas dependientes de ciclina, fosforila e inactiva la proteína del retinoblastoma y
promueve la progresión a través de la fase G1-S del ciclo celular.
Durante este proceso, los 2 factores de transcripción anteriormente mencionados, son
liberados en la célula. Hemos encontrado que el polimorfismo G>A en el nucleótido
870 del gen del CCND1 no afectaba a los niveles intracelulares de d4T-TP

16
El presente estudio transversal, que incluye un limitado número de pacientes, ha
mostrado una asociación significativa entre el genotipo de los genes de la TS y los
niveles de d4T intracelulares en pacientes con infección por VIH-1 con regímenes
antirretrovirales basados en d4T.
Hasta donde conocemos, este es el primer estudio farmacogenético que demuestra la
implicación que tiene la vía metabólica del folato en la toxicidad por d4T.
La farmacogenética, y especialmente, la toxicogenética, se están convirtiendo en un
campo de investigación cada vez más importante en la terapia antirretroviral para la
infección por el VIH-1. Los factores genéticos pueden alterar el metabolismo
farmacológico y la actividad y pueden predecir la toxicidad y/o la eficacia de los
fármacos.
La determinación de polimorfismos en enzimas que metabolizan sustancias
xenobióticas antes de la administración de la terapia antirretroviral podría ofrecer
nuevas estrategias para optimizar e individualizar la terapia antirretroviral.
CONCLUSIONES
Durante los últimos años se han descubierto diversos polimorfismos que pueden
modificar el metabolismo de distintos fármacos y por lo tanto, alterar su perfil tóxico.
Este estudio muestra que los polimorfismos funcionales de los genes que codifican la
TS pueden tener un papel importante en pacientes que reciben tratamiento
antirretroviral basado en estavudina debido principalmente a variaciones importantes en
las concentraciones de d4T-TP.
Los diferentes polimorfismos de los genes que codifican los distintos enzimas del
metabolismo del folato, no influyen de manera significativa en los niveles intracelulares
de d4T-TP cuando se consideran de manera individual, pero se comportan como
coadyuvantes a los polimorfismos de la TS.
Los 2 polimorfismos conocidos de los genes de la MTHFR descritos hasta ahora, son el
C677T y el A1298C. Ambos polimorfismos se asocian con un descenso de la actividad
de la enzima a través de la influencia que ejercen sobre los pools intracelulares de
folato.
Este estudio transversal ha mostrado una asociación significativa entre el genotipo de
los genes de la TS y los niveles de d4T intracelulares en pacientes con infección por
VIH-1 con regímenes antirretrovirales basados en d4T. A pesar de que el tamaño

17
muestral es muy pequeño, es el primer estudio farmacogenético que demuestra la
implicación de la vía metabólica del folato en la toxicidad por d4T.
Los factores genéticos pueden alterar el metabolismo farmacológico y la actividad y
pueden predecir la toxicidad y/o la eficacia de los fármacos.
No estamos lejos del día en el que la elección de un determinado tratamiento
antirretroviral se base en los factores genéticos del propio paciente.

18
Ayuda financiera
Este trabajo está financiado en parte por el Fondo de Investigaciones Sanitarias (FIS
02/1280, 05/1591, 07/0976, 08/00256), la Fundación para la Prevención del SIDA en
España (FIPSE 36610, 36572/06) y la Red de Investigación en SIDA (RIS
RD06/006/0022, RD06/0006/1004).

19
BIBLIOGRAFÍA
1. Lichtenstein KA. Redefining lipodystrophy syndrome: risks and impact on clinical decision making. J Acquir Immune Defic Syndr 2005; 39: 395-400.
2. Bernasconi E, Boubaker K, Junghans C, et al. Abnormalities of body fat
distribution in HIV-infected persons treated with antirretroviral drugs: The Swiss HIV Cohort Study. J Acquir Immune Defic Syndr 2002; 31: 50-5.
3. Miller J, Carr A, Emery S, et al. HIV lipodystrophy: prevalence, severity and
correlates of risk in Australia. HIV Med 2003; 4: 293-301. 4. Grinspoon S, Carr A. Cardiovascular risk and body-fat abnormalities in HIV-
infected adults. N Engl J Med 2005; 352: 48-62. 5. Nolan D, Hammond E, James I, McKinnon E, Mallal S. Contribution of
nucleoside-analogue reverse transcriptase inhibitor therapy to lipoatrophy from the population to the cellular level. Antivir Ther 2003; 8: 617-26.
6. Nolan D, Reiss P, Mallal S. Adverse effects of antirretroviral therapy for HIV
infection: a review of selected topics. Expert Opin Drug Saf 2005; 4: 201-18. 7. Gallant JE, Staszewski S, Pozniak AL, et al. Efficacy and safety of tenofovir DF
vs stavudine in combination therapy in antirretroviral-naive patients: a 3-year randomized trial. JAMA 2004; 292: 191-201.
8. Gallant JE, DeJesus E, Arribas JR, et al. Tenofovir DF, emtricitabine, and
efavirenz vs. zidovudine, lamivudine, and efavirenz for HIV. N Engl J Med 2006; 354: 251-60.
9. Lewis W, Day BJ, Copeland WC. Mitochondrial toxicity of NRTI antiviral
drugs: an integrated cellular perspective. Nat Rev Drug Discov 2003; 2: 812-22. 10. Jones SP, Qazi N, Morelese J, et al. Assessment of adipokine expression and
mitochondrial toxicity in HIV patients with lipoatrophy on stavudine- and zidovudine-containing regimens. J Acquir Immune Defic Syndr 2005; 40: 565-72.
11. Caron M, Auclair M, Lagathu C, et al. The HIV-1 nucleoside reverse
transcriptase inhibitors stavudine and zidovudine alter adipocyte functions in vitro. AIDS 2004; 18: 2127-36.
12. Kakuda TN. Pharmacology of nucleoside and nucleotide reverse transcriptase
inhibitor-induced mitochondrial toxicity. Clin Ther 2000; 22: 685-708. 13. Anderson PL, Kakuda TN, Lichtenstein KA. The cellular pharmacology of
nucleoside- and nucleotide-analogue reverse-transcriptase inhibitors and its relationship to clinical toxicities. Clin Infect Dis 2004; 38: 743-53.

20
14. Sales SD, Hoggard PG, Sunderland D, Khoo SH, Hart CA, Back DJ. Zidovudine phosphorylation and mitochondrial toxicity in vitro. Toxicol Appl Pharmacol 2001; 177: 54-8.
15. Horie N, Aiba H, Oguro K, Hojo H, Takeishi K. Functional analysis and DNA
polymorphism of the tandemly repeated sequences in the 5'-terminal regulatory region of the human gene for thymidylate synthase. Cell Struct Funct 1995; 20: 191-7.
16. Kawakami K, Salonga D, Park JM, et al. Different lengths of a polymorphic
repeat sequence in the thymidylate synthase gene affect translational efficiency but not its gene expression. Clin Cancer Res 2001; 7: 4096-101.
17. Mandola MV, Stoehlmacher J, Muller-Weeks S, et al. A novel single nucleotide
polymorphism within the 5' tandem repeat polymorphism of the thymidylate synthase gene abolishes USF-1 binding and alters transcriptional activity. Cancer Res 2003; 63(11):2898-2904.
18. Dervieux T, Furst D, Lein DO, et al. Polyglutamation of methotrexate with
common polymorphisms in reduced folate carrier, aminoimidazole carboxamide ribonucleotide transformylase, and thymidylate synthase are associated with methotrexate effects in rheumatoid arthritis. Arthritis Rheum 2004; 50: 2766-74.
19. Toffoli G, De Mattia E. Pharmacogenetic relevance of MTHFR polymorphisms.
Pharmacogenomics. 2008,9:1195-206. 20. Johnson WG, Stenroos ES, Spychala JR. New 19 bp deletion polymorphism in
intron -1 of dihydrofolate reductase (DHFR): a risk factor for spina bifida acting in mothers during pregnancy? Am J Med Genet 2004; 124A: 339-45.
21. Dulucq S, St-Onge G, Gagne V, et al. DNA variants in dihydrofolate reductase
gene and outcome in childhood ALL. Blood. 2008; 111: 3692-700. 22. Papadimitrakopoulou V, Izzo JG, Liu DD, et al. Cyclin D1 and cancer
development in laryngeal premalignancy patients. Cancer Prev Res 2009; 2: 14-21.
23. Centers for Disease Control. 1993 revised classification system for HIV
infection and expanded surveillance for case definition for AIDS among adolescents and adults. MMWR 1993; 41 (RR-17): 1-13.
24. Lemieux S, Prud'homme D, Bouchard C, Tremblay A, Despres JP. A single threshold of waist girth identifies normal-weight and overweight subjects with excess visceral adipose tissue. Am J Clin Nutr 1996; 64: 685-93.
25. Domingo P, Cabeza MC, Pruvost A, et al. Relationship between HIV/highly active antirretroviral therapy (HAART)-associated lipodystrophy syndrome and stavudine triphosphate intracellular levels in patients with stavudine-based antirretroviral regimes. Clin Infect Dis (In Press)

21
26. Executive Summary of the Third Report of The National Cholesterol Education Program (NCEP) Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults (Adult Treatment Panel III). JAMA 2001; 285: 2486-97.
27. Grundy SM, Cleeman JI, Daniels SR, et al. Diagnosis and management of the metabolic syndrome: an American Heart Association/National Heart, Lung, and Blood Institute Scientific Statement. Circulation 2005; 112: 2735-52.
28. Sugiuchi H, Uji Y, Okabe,K. Direct measurement of high-density lipoprotein cholesterol in serum with polyethylene glycol-modified enzymes and sulphated alpha-cyclodextrin. Clin Chem 1995; 41: 717-23.
29. Hainline A, Karon J, Lippel K. Lipid Research Clinic Program. Manual of Laboratory Operations, Lipids and lipoprotein analysis. 2nd ed.. Washington,D.C.: Govt. Printing Office, 2000.
30. Matthews DR, Hosker JP, Rudenski AS, Naylor BA, Treacher DF, Turner RC.
Homeostasis model assessment. Diabetologia 1985; 28: 412-19. 31. Ascaso JF, Romero P, Real JT, Lorente RI, Martinez-Valls J, Carmena R.
Abdominal obesity, insulin resistance, and metabolic syndrome in a southern European population. Eur J Intern Med 2003; 14:101-6.
32. Pruvost A, Becher F, Bardouille P, et al. Direct determination of phosphorylated
intracellular anabolites of stavudine (d4T) by liquid chromatography/tandem mass spectrometry. Rapid Commun Mass Spectrom. 2001;15: 1401-8.
33. Pruvost A, Théodoro F, Agrofoglio L, Negredo E, Bénech H, Specificity
enhancement with LC-positive ESI-MS/MS for the measurement of nucleotides: application to the quantitative determination of carbovir triphosphate, lamivudine triphosphate and tenofovir diphosphate in human peripheral blood mononuclear cells. J Mass Spectrom. 2008; 43: 224-33.
34. Benech, H., F. Theodoro, A. Herbet, et al. Peripheral blood mononuclear cell
counting using a DNA-detection-based method. Anal. Biochem 2004; 330: 172–4.
35. Miller SA, Dykes DD, Polesky HF. A simple salting out procedure for extracting
DNA from human nucleated cells. Nucleic Acids Res 1988; 16: 1215. 36. Kawakami K, Watanabe G. Identification and functional analysis of single
nucleotide polymorphism in the tandem repeat sequence of thymidylate synthase gene. Cancer Res 2003; 63: 6004-7.
37. Reich DE, Goldstein DB. Detecting association in a case-control study while
correcting for population stratification. Genet Epidemiol 2001; 20: 4-16. 38. Haas DW, Ribaudo HJ, Kim RB, et al. Pharmacogenetics of efavirenz and
central nervous system side effects: an Adult AIDS Clinical Trials Group study. AIDS 2004; 18: 2391-400.

22
39. Marzolini C, Telenti A, Decosterd LA, Greub G, Biollaz J, Buclin T. Efavirenz
plasma levels can predict treatment failure and central nervous system side effects in HIV-1-infected patients. AIDS 2001; 15: 71–5.
40. Barrett JS, Joshi AS, Chai M, Ludden TM, Fiske WD, Pieniaszek HJ Jr.
Population pharmacokinetic meta-analysis with efavirenz. Int J Clin Pharmacol Ther 2002; 40: 507–19.
41. Pfister M, Labbe L, Hammer SM, et al. Population pharmacokinetics and
pharmacodynamics of efavirenz, nelfinavir, and indinavir: Adult AIDS Clinical Trial Group Study 398. Antimicrob Agents Chemother 2003; 47: 130–7.
42. Strassburg CP, Manns MP. Jaundice genes and promoters. J Hepatol 2000; 33:
476–9. 43. Beutler E, Gelbart T, Demina A. Racial variability in the UDP-
glucuronosyltransferase 1 (UGT1A1) promoter: a balanced polymorphism for regulation of bilirubin metabolism?. Proc Natl Acad Sci US 1998; 95: 8170–4.
44. Raijmakers MT, Jansen PL, Steegers EA, Peters WH. Association of human
liver bilirubin UDP-glucuronyltransferase activity with a polymorphism in the promoter region of the UGT1A1 gene. J Hepatol 2000; 33: 348–51.
45. Zucker SD, Qin X, Rouster SD, et al. Mechanism of indinavir-induced
hyperbilirubinemia. Proc Natl Acad Sci USA 2001; 98: 12671–6. 46. Rotger M, Taffe P, Bleiber G, et al. Gilbert syndrome and the development of
antirretroviral therapy-associated hyperbilirubinemia. J Infect Dis. 2005; 192: 1381-6.
47. Domingo P, Vidal F, Domingo JC, et al. Tumor necrosis factor alpha in fat
redistribution syndromes associated with combination antirretroviral therapy: its role in subcutaneous adipocyte apoptosis. Eur J Clin Invest 2005; 35: 771-80.
48. Giralt M, Domingo P, Guallar JP, et al. Human immunodeficiency virus type 1
(HIV-1) infection alters gene expression in adipose tissue, contributing to HIV/highly active antirretroviral therapy-associated lipodystrophy. Antivir Ther 2006; 11: 729-40.
49. Sethi JK, Hotamisligil GS. The role of TNF alpha in adipocyte metabolism.
Semin Cell Dev Biol 1999; 10: 19–9. 50. Maher B, Alfirevic A, Vilar FJ, Wilkins EG, Park BK, Pirmohamed M. TNF-
alpha promoter region gene polymorphisms in HIV-positive patients with lipodystrophy. AIDS 2002; 16: 2013–8.
51. Nolan D, Moore C, Castley A, et al. Tumour necrosis factor-alpha gene -238G/A
promoter polymorphism associated with a more rapid onset of lipodystrophy. AIDS 2003; 17: 121–3.

23
52. Izzedine H, Hulot JS, Villard E, et al. Association between ABCC2 gene
haplotypes and tenofovir-induced proximal tubulopathy. J Infect Dis 2006; 194: 1481–91.
53. Mitsuya H, Weinhold KJ, Furman PA, et al. 3′-Azido-3′-deoxythymidine (BW
A509U): an antiviral agent that inhibits the infectivity and cytopathic effect of human T-lymphotropic virus type III/lymphadenopathy-associated virus in vitro. Proc Natl Acad Sci USA1985; 82: 7096–100.
54. Furman PA, Fyfe JA, St Clair MH, et al. Phosphorylation of 3′-azido-3′-
deoxythymidine and selective interaction of the 5′-triphosphate with human immunodeficiency virus reverse transcriptase. Proc Natl Acad. Sci USA 1986; 83: 8333–7.
55. Lewis, W, Dalakas MC. Mitochondrial toxicity of antiviral drugs. Nature Med
1995; 1: 417–22. 56. Becher F, Pruvost A, Goujard C, et al. Improved method for the simultaneous
determination of d4T, 3TC and ddI intracellular phosphorylated anabolites in human peripheral-blood mononuclear cells using high-performance liquid chromatography/tandem mass spectrometry. Rapid Commun Mass Spectrom 2002; 16: 555-65.
57. Salmon-Céron D, Lassalle R, Pruvost A, et al. Interferon-ribavirin in association
with stavudine has no impact on plasma human immunodeficiency virus (HIV) type 1 level in patients coinfected with HIV and hepatitis C virus: a CORIST-ANRS HC1 trial. Clin Infect Dis 2003; 36: 1295-304.
58. Compain S, Durand-Gasselin L, Grassi J, Benech H. Improved method to
quantify intracellular zidovudine mono- and triphosphate in peripheral blood mononuclear cells by liquid chromatography-tandem mass spectrometry. J Mass Spectrom 2007; 42: 389-404.
59. Lewis W, Simpson JF, Meyer RR. Cardiac mitochondrial DNA polymerase-
gamma is inhibited competitively and noncompetitively by phosphorylated zidovudine. Circ Res 1994; 74: 344–8.
60. Balzarini J. Effect of antimetabolite drugs of nucleotide metabolism on the anti-
human immunodeficiency virus activity of nucleoside reverse transcriptase inhibitors. Pharmacol Ther 2000; 87: 175–87.
61. Bresnick E, Karjala R. End product inhibition of thymidine kinase activity in
normal and leukemic leukocytes. Cancer Res 1964; 24: 841–6. 62. Gao WY, Johns DG, Tanaka M, Mitsuya H. Suppression of replication of
multidrug-resistant HIV type 1 variants by combinations of thymidylate synthase inhibitors with zidovudine or stavudine. Mol Pharmacol 1999; 55: 535-40.

24
63. Gao W-Y, Johns DG, Mitsuya H. Anti-human immunodeficiency virus type 1 activity of hydroxyurea in combination with 29,39-dideoxynucleosides. Mol Pharmacol 1994; 46: 767–72.
64. Rutschmann OT, Vernazza PL, Bucher HC, et al. Long-term hydroxyurea in
combination with didanosine and stavudine for the treatment of HIV-1 infection. Swiss HIV Cohort Study. AIDS 2000; 14: 2145-51.
65. Moore RD, Wong WM, Keruly JC, McArthur JC. Incidence of neuropathy in
HIV-infected patients on monotherapy versus those on combination therapy with didanosine, stavudine and hydroxyurea. AIDS 2000; 14: 273-8.
66. Moore RD, Keruly JC, Chaisson RE. Incidence of pancreatitis in HIV-infected
patients receiving nucleoside reverse transcriptase inhibitor drugs. AIDS 2001; 15: 617-20.
67. Marsh S, Collie-Duguid ESR, Li T, Liu X, McLeod HL. Ethnic variation of the
thymidylate enhancer region polymorphism among Caucasian and Asian populations. Genomics 1999; 58: 310-2.
68. Marsh S, Ameyaw MM, Githang J, Indalo A, Ofori-Adjei D, McLeod HL. Novel
Thymidylate Synthase Enhancer Region Alleles in African Populations. Human Mutation 2000; 16: 528.
69. Chang KH, Kim JM, Song YG, Lee HC, Lim SK. Does Race Protect an Oriental
Population From Developing Lipodystrophy in HIV-infected Individuals on HAART?. J Infect 2002; 44: 33-8.
70. De Mattia E, Toffoli G. C677T and A1298C MTHFR polymorphisms, a
challenge for antifolate and fluoropyrimidine-based therapy personalisation. Eur J Cancer 2009; 45: 1333-51.
71. Sharp L, Little J. Polymorphisms in genes involved in folate metabolism and
colorectal neoplasia: a HuGE review. Am J Epidemiol 2004; 159: 423-43. 72. Gellekink H, Blom HJ, van der Linden IJ, den Heijer M. Molecular genetic
analysis of the human dihydrofolate reductase gene: relation with plasma total homocysteine, serum and red blood cell folate levels. Eur J Hum Genet 2007; 15: 103-9.
73. Chango A, Emery-Fillon N, de Courcy GP, et al. A polymorphism (80G>A) in
the reduced folate carrier gene and its association with folate status and homocysteinemia. Mol Genet Metab 2000; 70: 310–5.
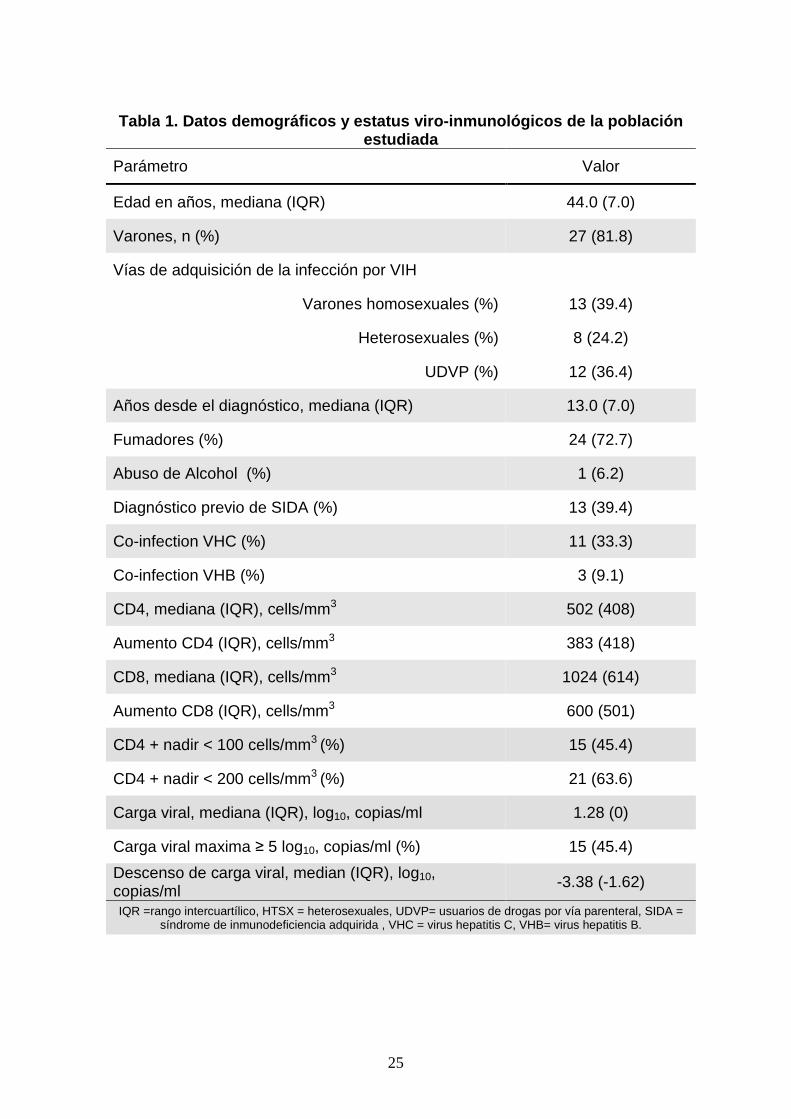
25
Tabla 1. Datos demográficos y estatus viro -inmunológicos de la población
estudiada
Parámetro Valor
Edad en años, mediana (IQR) 44.0 (7.0)
Varones, n (%) 27 (81.8)
Vías de adquisición de la infección por VIH
Varones homosexuales (%) 13 (39.4)
Heterosexuales (%) 8 (24.2)
UDVP (%) 12 (36.4)
Años desde el diagnóstico, mediana (IQR) 13.0 (7.0)
Fumadores (%) 24 (72.7)
Abuso de Alcohol (%) 1 (6.2)
Diagnóstico previo de SIDA (%) 13 (39.4)
Co-infection VHC (%) 11 (33.3)
Co-infection VHB (%) 3 (9.1)
CD4, mediana (IQR), cells/mm3 502 (408)
Aumento CD4 (IQR), cells/mm3 383 (418)
CD8, mediana (IQR), cells/mm3 1024 (614)
Aumento CD8 (IQR), cells/mm3 600 (501)
CD4 + nadir < 100 cells/mm3 (%) 15 (45.4)
CD4 + nadir < 200 cells/mm3 (%) 21 (63.6)
Carga viral, mediana (IQR), log10, copias/ml 1.28 (0)
Carga viral maxima ≥ 5 log10, copias/ml (%) 15 (45.4)
Descenso de carga viral, median (IQR), log10, copias/ml
-3.38 (-1.62)
IQR =rango intercuartílico, HTSX = heterosexuales, UDVP= usuarios de drogas por vía parenteral, SIDA = síndrome de inmunodeficiencia adquirida , VHC = virus hepatitis C, VHB= virus hepatitis B.
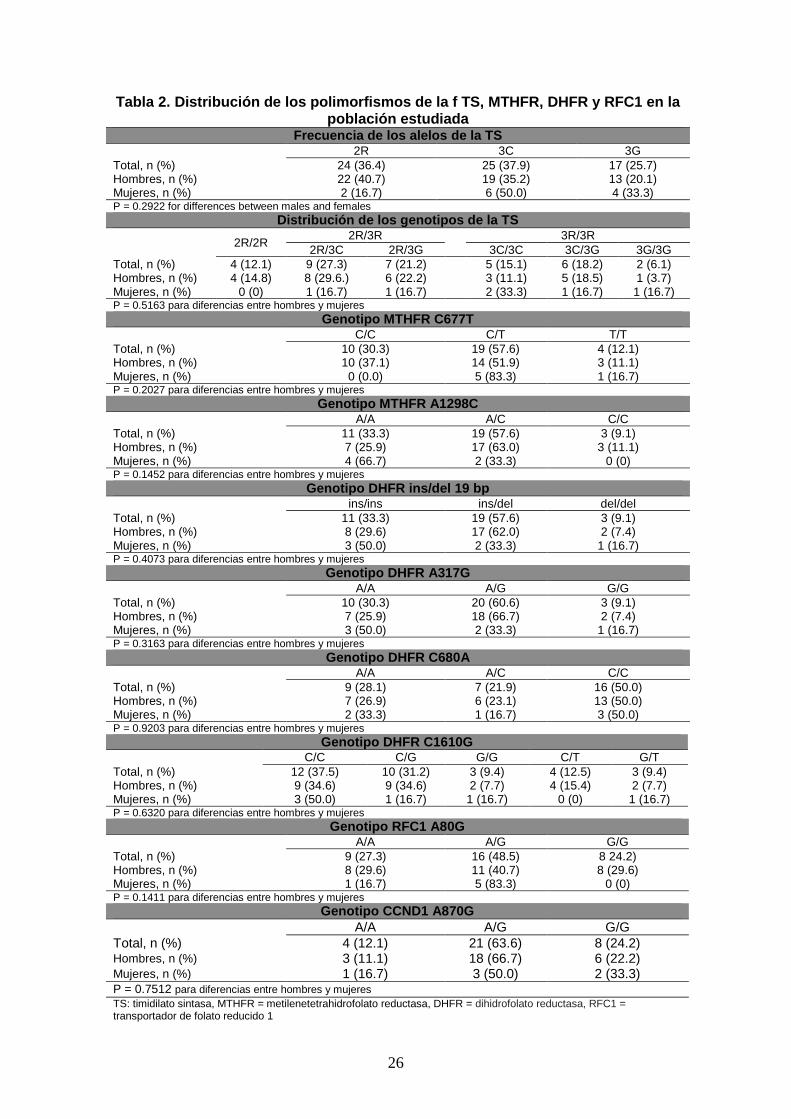
26
Tabla 2. Distribución de los polimorfismos de la f TS, MTHFR, DHFR y RFC1 en la población estudiada
Frecuenc ia de los alelos de la TS 2R 3C 3G Total, n (%) 24 (36.4) 25 (37.9) 17 (25.7) Hombres, n (%) 22 (40.7) 19 (35.2) 13 (20.1) Mujeres, n (%) 2 (16.7) 6 (50.0) 4 (33.3) P = 0.2922 for differences between males and females
Distribución de los genotipos d e la TS
2R/2R 2R/3R 3R/3R
2R/3C 2R/3G 3C/3C 3C/3G 3G/3G Total, n (%) 4 (12.1) 9 (27.3) 7 (21.2) 5 (15.1) 6 (18.2) 2 (6.1) Hombres, n (%) 4 (14.8) 8 (29.6.) 6 (22.2) 3 (11.1) 5 (18.5) 1 (3.7) Mujeres, n (%) 0 (0) 1 (16.7) 1 (16.7) 2 (33.3) 1 (16.7) 1 (16.7) P = 0.5163 para diferencias entre hombres y mujeres
Genotipo MTHFR C677T C/C C/T T/T Total, n (%) 10 (30.3) 19 (57.6) 4 (12.1) Hombres, n (%) 10 (37.1) 14 (51.9) 3 (11.1) Mujeres, n (%) 0 (0.0) 5 (83.3) 1 (16.7) P = 0.2027 para diferencias entre hombres y mujeres
Genotipo MTHFR A1298C A/A A/C C/C Total, n (%) 11 (33.3) 19 (57.6) 3 (9.1) Hombres, n (%) 7 (25.9) 17 (63.0) 3 (11.1) Mujeres, n (%) 4 (66.7) 2 (33.3) 0 (0) P = 0.1452 para diferencias entre hombres y mujeres
Genotip o DHFR ins/del 19 bp ins/ins ins/del del/del Total, n (%) 11 (33.3) 19 (57.6) 3 (9.1) Hombres, n (%) 8 (29.6) 17 (62.0) 2 (7.4) Mujeres, n (%) 3 (50.0) 2 (33.3) 1 (16.7) P = 0.4073 para diferencias entre hombres y mujeres
Genotipo DHFR A317G A/A A/G G/G Total, n (%) 10 (30.3) 20 (60.6) 3 (9.1) Hombres, n (%) 7 (25.9) 18 (66.7) 2 (7.4) Mujeres, n (%) 3 (50.0) 2 (33.3) 1 (16.7) P = 0.3163 para diferencias entre hombres y mujeres
Genotipo DHFR C680A A/A A/C C/C Total, n (%) 9 (28.1) 7 (21.9) 16 (50.0) Hombres, n (%) 7 (26.9) 6 (23.1) 13 (50.0) Mujeres, n (%) 2 (33.3) 1 (16.7) 3 (50.0) P = 0.9203 para diferencias entre hombres y mujeres
Genotipo DHFR C1610G C/C C/G G/G C/T G/T Total, n (%) 12 (37.5) 10 (31.2) 3 (9.4) 4 (12.5) 3 (9.4) Hombres, n (%) 9 (34.6) 9 (34.6) 2 (7.7) 4 (15.4) 2 (7.7) Mujeres, n (%) 3 (50.0) 1 (16.7) 1 (16.7) 0 (0) 1 (16.7) P = 0.6320 para diferencias entre hombres y mujeres
Genotipo RFC1 A80G A/A A/G G/G Total, n (%) 9 (27.3) 16 (48.5) 8 24.2) Hombres, n (%) 8 (29.6) 11 (40.7) 8 (29.6) Mujeres, n (%) 1 (16.7) 5 (83.3) 0 (0) P = 0.1411 para diferencias entre hombres y mujeres
Genotipo CCND1 A870G A/A A/G G/G Total, n (%) 4 (12.1) 21 (63.6) 8 (24.2) Hombres, n (%) 3 (11.1) 18 (66.7) 6 (22.2) Mujeres, n (%) 1 (16.7) 3 (50.0) 2 (33.3) P = 0.7512 para diferencias entre hombres y mujeres TS: timidilato sintasa, MTHFR = metilenetetrahidrofolato reductasa, DHFR = dihidrofolato reductasa, RFC1 = transportador de folato reducido 1
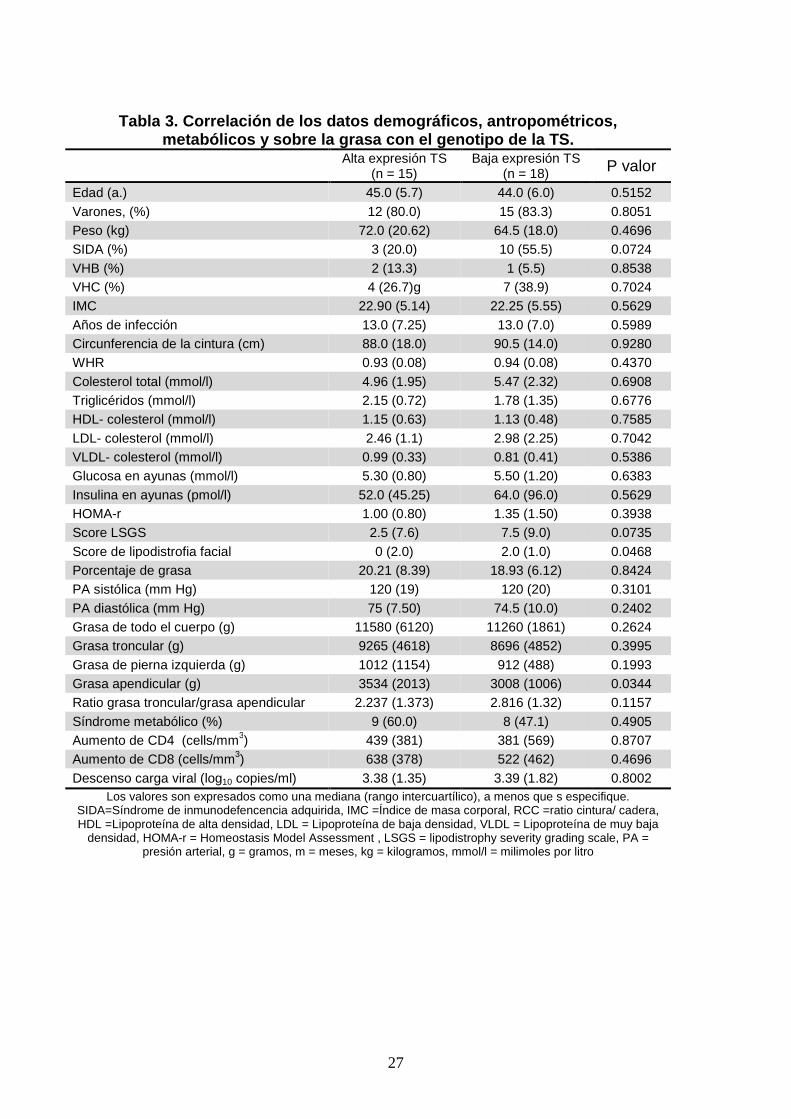
27
Tabla 3. Correlación de los datos demográficos, antropométricos,
metabólicos y sobre la grasa con el genotipo de la TS. Alta expresión TS
(n = 15) Baja expresión TS
(n = 18) P valor
Edad (a.) 45.0 (5.7) 44.0 (6.0) 0.5152 Varones, (%) 12 (80.0) 15 (83.3) 0.8051 Peso (kg) 72.0 (20.62) 64.5 (18.0) 0.4696 SIDA (%) 3 (20.0) 10 (55.5) 0.0724 VHB (%) 2 (13.3) 1 (5.5) 0.8538 VHC (%) 4 (26.7)g 7 (38.9) 0.7024 IMC 22.90 (5.14) 22.25 (5.55) 0.5629 Años de infección 13.0 (7.25) 13.0 (7.0) 0.5989 Circunferencia de la cintura (cm) 88.0 (18.0) 90.5 (14.0) 0.9280 WHR 0.93 (0.08) 0.94 (0.08) 0.4370 Colesterol total (mmol/l) 4.96 (1.95) 5.47 (2.32) 0.6908 Triglicéridos (mmol/l) 2.15 (0.72) 1.78 (1.35) 0.6776 HDL- colesterol (mmol/l) 1.15 (0.63) 1.13 (0.48) 0.7585 LDL- colesterol (mmol/l) 2.46 (1.1) 2.98 (2.25) 0.7042 VLDL- colesterol (mmol/l) 0.99 (0.33) 0.81 (0.41) 0.5386 Glucosa en ayunas (mmol/l) 5.30 (0.80) 5.50 (1.20) 0.6383 Insulina en ayunas (pmol/l) 52.0 (45.25) 64.0 (96.0) 0.5629 HOMA-r 1.00 (0.80) 1.35 (1.50) 0.3938 Score LSGS 2.5 (7.6) 7.5 (9.0) 0.0735 Score de lipodistrofia facial 0 (2.0) 2.0 (1.0) 0.0468 Porcentaje de grasa 20.21 (8.39) 18.93 (6.12) 0.8424 PA sistólica (mm Hg) 120 (19) 120 (20) 0.3101 PA diastólica (mm Hg) 75 (7.50) 74.5 (10.0) 0.2402 Grasa de todo el cuerpo (g) 11580 (6120) 11260 (1861) 0.2624 Grasa troncular (g) 9265 (4618) 8696 (4852) 0.3995 Grasa de pierna izquierda (g) 1012 (1154) 912 (488) 0.1993 Grasa apendicular (g) 3534 (2013) 3008 (1006) 0.0344 Ratio grasa troncular/grasa apendicular 2.237 (1.373) 2.816 (1.32) 0.1157 Síndrome metabólico (%) 9 (60.0) 8 (47.1) 0.4905 Aumento de CD4 (cells/mm3) 439 (381) 381 (569) 0.8707 Aumento de CD8 (cells/mm3) 638 (378) 522 (462) 0.4696 Descenso carga viral (log10 copies/ml) 3.38 (1.35) 3.39 (1.82) 0.8002
Los valores son expresados como una mediana (rango intercuartílico), a menos que s especifique. SIDA=Síndrome de inmunodefencencia adquirida, IMC =Índice de masa corporal, RCC =ratio cintura/ cadera, HDL =Lipoproteína de alta densidad, LDL = Lipoproteína de baja densidad, VLDL = Lipoproteína de muy baja
densidad, HOMA-r = Homeostasis Model Assessment , LSGS = lipodistrophy severity grading scale, PA = presión arterial, g = gramos, m = meses, kg = kilogramos, mmol/l = milimoles por litro
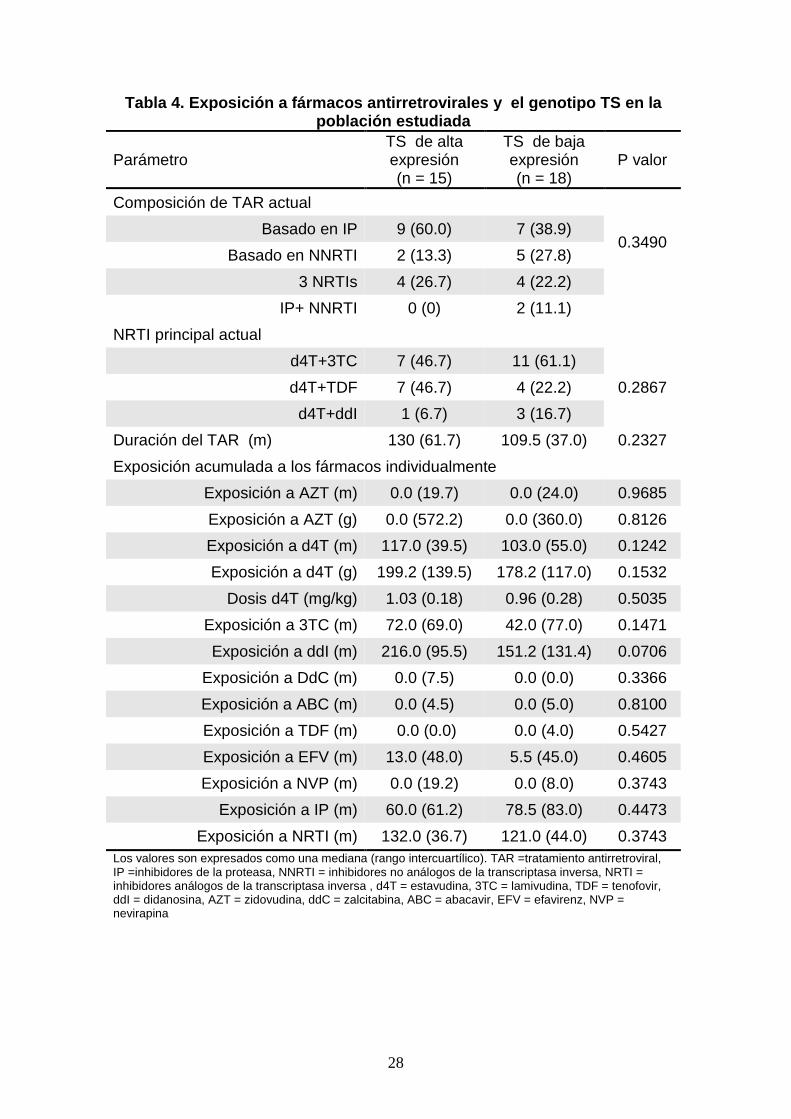
28
Tabla 4. Exposición a fármacos antir retrovirales y el genotipo TS en la población estudiada
Parámetro TS de alta expresión (n = 15)
TS de baja expresión (n = 18)
P valor
Composición de TAR actual
0.3490 Basado en IP 9 (60.0) 7 (38.9)
Basado en NNRTI 2 (13.3) 5 (27.8)
3 NRTIs 4 (26.7) 4 (22.2)
IP+ NNRTI 0 (0) 2 (11.1)
NRTI principal actual
d4T+3TC 7 (46.7) 11 (61.1)
0.2867 d4T+TDF 7 (46.7) 4 (22.2)
d4T+ddI 1 (6.7) 3 (16.7)
Duración del TAR (m) 130 (61.7) 109.5 (37.0) 0.2327
Exposición acumulada a los fármacos individualmente
Exposición a AZT (m) 0.0 (19.7) 0.0 (24.0) 0.9685
Exposición a AZT (g) 0.0 (572.2) 0.0 (360.0) 0.8126
Exposición a d4T (m) 117.0 (39.5) 103.0 (55.0) 0.1242
Exposición a d4T (g) 199.2 (139.5) 178.2 (117.0) 0.1532
Dosis d4T (mg/kg) 1.03 (0.18) 0.96 (0.28) 0.5035
Exposición a 3TC (m) 72.0 (69.0) 42.0 (77.0) 0.1471
Exposición a ddI (m) 216.0 (95.5) 151.2 (131.4) 0.0706
Exposición a DdC (m) 0.0 (7.5) 0.0 (0.0) 0.3366
Exposición a ABC (m) 0.0 (4.5) 0.0 (5.0) 0.8100
Exposición a TDF (m) 0.0 (0.0) 0.0 (4.0) 0.5427
Exposición a EFV (m) 13.0 (48.0) 5.5 (45.0) 0.4605
Exposición a NVP (m) 0.0 (19.2) 0.0 (8.0) 0.3743
Exposición a IP (m) 60.0 (61.2) 78.5 (83.0) 0.4473
Exposición a NRTI (m) 132.0 (36.7) 121.0 (44.0) 0.3743 Los valores son expresados como una mediana (rango intercuartílico). TAR =tratamiento antirretroviral, IP =inhibidores de la proteasa, NNRTI = inhibidores no análogos de la transcriptasa inversa, NRTI = inhibidores análogos de la transcriptasa inversa , d4T = estavudina, 3TC = lamivudina, TDF = tenofovir, ddI = didanosina, AZT = zidovudina, ddC = zalcitabina, ABC = abacavir, EFV = efavirenz, NVP = nevirapina
29
Tabla 5. Influencia de los genotipos de la TS, MTHD R, DHFR y RFC1 en los
niveles intracelulares de d4T-TP d4T-TP (fmol/106 cells) P valor
Genotipo de TS 2R/2R (n = 4) 24.55 (10.30)
0.0116
2R/3C (n = 9) 26.70 (38.97) 2R/3G (n = 7) 10.10 (5.27) 3C/3C (n = 5) 18.90 (8.02) 3C/3G (n = 6) 11.20 (6.30) 3G/3G (n = 2) 17.0 (7.80)
Baja expresión (n = 18) 20.65 (12.70) 0.0010
Alta expresión (n = 15) 11.50 (5.75) Genotipo MTHFR C677T
C/C (n = 10) 15.40 (15.60) 0.5999 C/T (n = 19) 18.90 (9.80)
T/T (n = 4) 12.30 (11.70) Genotipo MTHFR A1298C
A/A (n = 11) 13.10 (12.32) 0.1024 A/C (n = 19) 15.20 (9.60)
C/C (n = 3) 28.50 (2.40) Genotipo DHFR ins/del 19 pb
ins/ins (n = 11) 15.20 (9.82) 0.2606 ins/del (n = 19) 14.80 (16.67)
del/del (n = 3) 52.10 (44.32) Genotipo DHFR A317G
A/A (n = 10) 17.65 (10.60) 0.2576 A/G (n = 20) 14.40 (14.45)
G/G (n = 3) 52.10 (44.32) Genotipo DHFR C680A
A/A (n = 9) 20.90 (22.22) 0.4953 A/C (n = 7) 19.40 (23.05)
C/C ( n = 17) 14.80 (10.22) Genotipo DHFR C1610G
C/C (n = 12) 16.70 (14.70)
0.9999 C/G (n = 10) 14.50 (15.60) G(G (n = 3) 20.60 (8.25) C/T (n = 4) 20.00 (42.55) G/T (n = 4) 15.90 6.75)
Genotipo RFC1 A80G A/A (n = 9) 10.10 (8.97)
0.1907 A/G (n = 16) 16.85 (12.65) G/G (n = 8) 20.00 (24.90)
Genotipo CCNDI A870G A/A (n = 4) 10.30 (8.00)
0.1743 A/G (n = 21) 19.40 (14.45) G/G (n = ) 14.35 (8.00)
TS = timidilato sintasa, MTHFR = metilenetetrahidrofolato reductasa, DHFR = dihidrofolato reductasa, RFC1 = transportador de folato reducido 11, CCNDI = ciclina D1

RESUM
Antecedents:
L’activitat antiretroviral i la toxicitat de l’estavudina (d4T) depèn del seu metabòlit
trifosfat, l’estavudina trifosfat (d4T-TP). Per tant, les modificacions dels nivells
intracel·lulars de d4T-TP poden canviar el perfil de la toxicitat de l’estavudina.
Mètodes:
Es varen determinar els nivells intracel·lulars de d4T-TP a les cèl·lules mononuclears de
sang perifèrica (CBMC) per la prominència al cromatògraf líquid connectat a un
espectròmetre de massa triple - quàdruple. Els polimorfismes dels gens de la Timidilat
sintasa (TS), Metilentetrahidrofolat reductasa (MTHFR), dihidrofolat reductasa ,
transportador de folat reduït 1 (RFC1) i ciclina D1 (CCND1) varen ser determinades per
seqüènciació directa utilitzant ABI PRISM 3100 Genetic Analyzer o bé Fluidigm’s
Biomark system. Per l’anàlisi estadístic es van utilitzar el test de la t de Student, l’índex
de correlació de Pearson, ANOVA d’un factor amb correccions pel mètode de
Bonferroni i una regressió logística esglaonada.
Resultats:
Es van reclutar 33 malalts per aquest estudi transversal. Els nivells intracel·lulars de d4T
van ser de 11.50 (RIC: 5.75) fmol/106 cels als malalts amb un genotip d’alta expressió
del TS (*2/*3G, *3C/ *3G and *3G/ 3G) mentre que per aquells amb un genotip de
baixa expressió (*2/ *2, *2/ *3C and *3C/ *3C), els nivells van ser de 20.65 (12.70)
fmol/106 cels (P = 0.0010). Els polimorfismes dels gens de MTHFR, DHFR, RFC1 i
CCND1 no van influir en la concentració intracel·lular de d4T-TP.
Conclusions:
Els nivells intracel·lulars de d4T són determinats pels polimorfismes de timidilato
sintasa.
Paraules clau: estavudina, estavudina-trifosfat, timidilat sintasa, metilenetetrahidrofolat
reductasa, dihidrofolat reductasa, ciclina D1, toxicitat, pools de nucleòtids, via
metabòlica, farmacogenètica.





























