Embed Size (px)
Citation preview

FACULTAD DE FARMACIA
UNIVERSIDAD COMPLUTENSE
TRABAJO FIN DE GRADO
INFLUENCIA DE LA CRONOBIOLOGÍA
EN EL METABOLISMO: VÍA mTOR Y RELOJ
CIRCADIANO
Autor: Adrián González Villeta
Tutor: Ángel Agis Torres
Convocatoria: Julio 2017

Resumen El organismo presenta una serie de variaciones rítmicas en su metabolismo y expresión
genética que le llevan a funcionar de una forma más eficaz en consonancia con el
medio. Dichas oscilaciones ocurren en ciclos de 24 horas y determinan gran parte de los
procesos fisiológicos. Su desincronización o alteración producen enfermedades desde el
cáncer a la epilepsia. El reloj central, en el NSQ (núcleo supraquiasmático), es el
marcapasos que coordina los demás relojes periféricos y para ello se vale de hormonas
como la melatonina. Los estímulos externos, como el lumínico, resetean el reloj para
ajustarse a las variaciones del ambiente y en él participan los llamados genes reloj.
Éstos oscilan en ciclos retroalimentativos de forma que ellos mismos modulan su ritmo
y en conjunción con el estímulo luz/oscuridad generan la ritmicidad circadiana. La
inducción lumínica también afecta a la vía de señalización mTOR, con importantes
funciones en cuanto a crecimiento y proliferación celular, así como metabólicas. Dicha
vía, además, modula la expresión de los genes reloj y se ve influida a su vez por ellos,
influyendo la oscilación circadiana a todo tipo de funciones fisiológicas.
Introducción y antecedentes Las variaciones diarias en los procesos comportamentales y fisiológicos están
controlados por una red de relojes circadianos, reseteados por estímulos y enviando
señales circadianas al cerebro y órganos periféricos. (Pevet 2014) (Pevet and Challet
2011) Dicha red se compone por el reloj maestro del NSQ (núcleo supraquiasmático),
ubicado en el hipotálamo, y una multitud de relojes secundarios localizados en el
cerebro y órganos periféricos (Challet 2015).
La razón de un ritmo circadiano no es otra que la de anticiparse a los cambios regulares
diarios para responder más eficazmente y optimizar los recursos. Para que esto tenga
lugar, el ritmo debe ajustarse a factores indicadores externos tales como la luz diaria,
puesto que no es perfectamente preciso. Los ritmos circadianos siguen funcionando
incluso cuando la fuente de la regulación ha desaparecido, siendo su período de libre
funcionamiento ligeramente superior o inferior a 24 horas (Pevet, Agez et al. 2006). Un
ciclo circadiano puede ser reseteado por los factores externos para adaptarse a los
cambios en las oscilaciones diarias. (Golombek and Rosenstein 2010). De esta forma,
los ciclos de luz/oscuridad diarios modulan el ritmo circadiano natural de las neuronas
del NSQ del hipotálamo para ajustarlo al medio ambiente y se complete en 24 horas
(Touitou 2016).
- 2 -

La detección del descenso de intensidad lumínica que modula el ritmo circadiano se
lleva a cabo mediante los conos y bastones, situados en la capa externa de la retina, que
se encargan de la fototransducción, transformando la misma mediante la intervención de
fotopigmentos en un potencial nervioso que se transmite y se modula a medida que se
transmite por las diversas capas celulares de la retina hasta llegar a las células
ganglionares que conectan directamente con el NSQ del hipotálamo a través del tracto
retinohipotalámico (Foster 1998, Berson 2003) No obstante, se ha observado que una
pequeña proporción de éstas células ganglionares tienen capacidad fotorreceptiva
intrínseca, respondiendo directamente a la luz incluso tras haber sido aisladas del resto
de la retina (Berson, Dunn et al. 2002). Las llamadas células ganglionares de la retina
intrínsecamente fotosensibles (ipRGCs) captan la luz gracias a un fotopigmento basado
en la opsina llamado melanopsina (Opn4) (Qiu, Kumbalasiri et al. 2005) (Panda, Nayak
et al. 2005) (Peirson and Foster 2006) (Dominguez-Solis and Perez-Leon 2015) Son los
verdaderos receptores lumínicos del ritmo circadiano.
Desde el núcleo supraquiasmático, las neuronas proyectan sus axones al cuerno lateral
de la médula espinal, donde sinaptan las neuronas postganglionares que modularán
dicho impulso en el ganglio superior cervical y lo transmitirán finalmente a la glándula
Pineal (Que significa forma de piña). Es aquí donde se sintetiza la melatonina a partir de
Triptófano (Trp) (Dominguez-Solis and Perez-Leon 2015) (Schibler, Gotic et al. 2015).
La regulación de la oscilación transcurre a nivel celular, individualmente, aunque se
trate de un organismo pluricelular. Nuestros ciclos diurnos de vigilia y sueño,
temperatura corporal y liberación hormonal están controlados por un reloj que opera en
cada miembro de un grupo especializado de células (neuronas del NSQ) en el
hipotálamo. Estas células constituyen el reloj circadiano en el NSQ (Kalsbeek, Perreau-
Lenz et al. 2006) y continúan oscilando individualmente incluso después de haber sido
separadas del resto del tejido, mostrando un patrón de expresión génica cíclico de
aproximadamente 24 horas (Welsh, Takahashi et al. 2010) Constituyen, así, el reloj
maestro que sincroniza los relojes circadianos secundarios: el marcapasos central.
(Pevet 2014) (Pevet and Challet 2011) (Challet 2015) Si bien recientemente se ha
sugerido que en realidad forma parte de una red mayor en la que su interacción con el
núcleo arcuato (ARC) es esencial para el correcto funcionamiento del sistema
circadiano (Buijs, Guzman-Ruiz et al. 2017). En humanos el NSQ proyecta sus axones a
la glándula pineal mediante las conexiones hipotalámicas al tronco del encéfalo y
médula espinal. De allí la inervación, principalmente simpática, sinapta con el ganglio
- 3 -

superior cervical y pineal. A parte de la melatonina, la glándula pineal contiene también
noradrenalina y serotonina, péptidos como GnRH y TRH, y neurotransmisor GABA.
La proteína mTOR (Mammalian Target Of Rapamycine) se ha revelado como un
componente más del ciclo circadiano, además de sus ya conocidas funciones en el
metabolismo y en la regulación del crecimiento y proliferación celular (Cao, Lee et al.
2008). Su regulación por el estímulo lumínico y su participación en los mecanismos del
reloj, le convierten en un interesante punto para comprender mejor nuestro organismo y
mejorar los tratamientos de las enfermedades derivadas de su desregulación.
Objetivos El presente trabajo tiene por objetivos:
• Revisar los mecanismos que regulan los ritmos circadianos, desde el estímulo
lumínico a la expresión de los genes reloj.
• El papel que desempeña la vía mTOR en dichos mecanismos y cómo se ve
influida por el estímulo luminoso.
Metodología Se ha llevado a cabo una revisión bibliográfica, utilizando de forma preeminente la base
de datos PubMed como principal fuente de artículos y consulta de libros, así como otras
fuentes de forma complementaria como Google Scholar y Web of Science. También se
ha acudido a la revista Chronobiology International y a Endotext, una web de
endocrinología clínica.
Resultados y discusión Melatonina: coordinación con relojes periféricos La síntesis de melatonina es mayor en ausencia de luz. La exposición a luz natural o a
luces artificiales de alta luminosidad inhiben parcialmente la enzima N-acetiltransferasa
(Pevet and Challet 2011). De esta forma, La producción y liberación de melatonina se
relaciona estrechamente con la fase lumínica imperante y el control del reloj NSQ,
siendo mayor durante la noche e inhibida por la exposición lumínica (Pevet 2014)
(Pevet and Challet 2011). Sus funciones son variadas y dependen del tejido donde se
- 4 -

expresen dichos receptores, consistiendo principalmente en distribuir las señales
temporales generadas por el NSQ a dichos tejidos, regular las fases de los ritmos
circadianos al ejercer un efecto sincronizador actuando a nivel de los osciladores
periféricos, la promoción del sueño y la termorregulación (Choi, Kwon et al. 2014)
(Pevet and Challet 2011) (Pevet 2014). Sus acciones a nivel central y periférico parecen
ser importantes en el antagonismo de la inflamación, al reducir significativamente la
activación de p65 y la consiguiente liberación de factores inflamatorios, aunque su
papel en la inflamación aún es controvertido ya que también puede actuar como
activador de la respuesta inmune (Liu, Hong et al. 2014, Manka and Majewska 2016) y
la excitotoxicidad, teniendo incluso actividad oncostática. (Huo, Wang et al. 2017)
(Hardeland, Cardinali et al. 2011) Ejerce sus efectos interactuando con los receptores
MT1 y MT2 además de proteínas intracelulares, los llamados receptores nucleares
huérfanos, tales como la quinona reductasa 2, calmodulina, calreticulina y tubulina. Los
receptores MT1 y 2 son receptores unidos a proteínas G expresados en varias partes del
sistema nervioso central y tejidos periféricos. Sus efectos varían en función del medio
celular donde actúe. (Pandi-Perumal, Trakht et al. 2008) Dichos receptores están
presentes también en el NSQ, de tal forma que la melatonina endógena es capaz de
ejercer un efecto feedback sobre el reloj maestro (Pevet and Challet 2011) Además, la
melatonina también tiene efectos independientes de receptor, poseyendo efecto
detoxificador de radicales libres y otras especies de oxígeno. (Reiter, Tan et al. 2010)
Relojes circadianos: genes reloj Hay una serie de genes que regulan la expresión génica dentro de la célula. Son los
llamados “genes reloj” y generan la ritmicidad circadiana a nivel celular. Están
ampliamente distribuidos en casi todas las células del cuerpo (Gerber, Saini et al. 2015)
y sus productos se relacionan en un ciclo transcripcional y traduccional para dar lugar a
las oscilaciones (Mazzoccoli, Pazienza et al. 2012) Son Per (gen Period) (Per1, Per2,
Per3), Tim (gen Timeless), Cry (gen Cryptochrome) (Cry1, Cry2), Rev-Erb (reverse
erythoblastosis virus)(Rev-Erbα), Ror (Retinoic acid-related Orphan Receptor), CLOCK
(Circadian Locomotor Output Cycles Kaput) (y su análogo neuronal Npas2), y Bmal
(Brain and Muscle ARNT-Like protein) (Bmal1, Bmal2). Tanto en las neuronas del
NSQ como en las células de los tejidos periféricos, el ciclo molecular del reloj
circadiano se compone en general de dos ciclos acoplados y retroalimentativos:
- 5 -

transcripcional (constituido a su vez por tres ciclos de retroalimentación) y post-
transcripcional.
1. Transcripcional: CLOCK/Bmal1 forman un heterodímero de regulación positiva
y Per/Cry otro negativo. Clock/Bmal heterodimerizan y se unen a la región
promotora específica “E-box” (5’-CACGTG-3’) activando al máximo la
transcripción de los genes Per y Cry (además de otros genes reloj-dependientes
en tejidos periféricos), que codifican para las proteínas homónimas. Estas
proteínas, inestables, se van acumulando y estabilizando durante el día y al
alcanzar una concentración crítica (pico máximo al comienzo de la noche)
heterodimerizan en el represor Per/Cry y se traslocan al núcleo donde inhiben la
actividad de CLOCK/Bmal. Hay un segundo ciclo regulador retroalimentativo o
“ciclo estabilizador” constituído por los receptores nucleares huérfanos Rev-
Erbα y RORα: Rev-Erbα compite con RORα por unirse al promotor de Bmal1.
RORα activa su transcripción, mientras que Rev-Erbα la reprime, dando una
expresión cíclica de Bmal1
2. Post-transcripcional: Modificaciones en las proteínas Per y Cry (que inhiben la
transcripción por CLOCK/Bmal) mediante fosforilación y defosforilación, entre
otros. Estas modificaciones pueden acelerar o endentecer su transferencia al
proteosoma para su degradación.
Estos ciclos aseguran el funcionamiento rítmico circadiano (Lincoln, Messager et al.
2002) (Mazzoccoli, Pazienza et al. 2012) (Schibler, Gotic et al. 2015) (Charrier, Olliac
et al. 2017) (Ver Figura 1)
Figura 1. ciclos retroalimentativos de los genes reloj. (Cho et al. 2012)
- 6 -

Regulación de los relojes circadianos
Ajuste nocturno Como ya se ha indicado, el reloj circadiano del NSQ es inducido (sincronizado o ajuste)
por señales ambientales para sincronizarse con las variaciones diarias del medio. La luz
es la principal de esas señales, de forma que el reloj NSQ esta estrechamente regulado
por los ciclos luz/oscuridad diarios. Durante la noche, la exposición a estímulos
luminosos provoca un ajuste rápido del reloj (Cao, Gkogkas et al. 2015). El reloj tiene
una característica llamada “gating” que le permite restringir su respuesta a las señales
ambientales en función del momento del día. Por ejemplo, los mamíferos diurnos son
generalmente insensibles a los pulsos luminosos durante el día, pero durante la noche un
pulso de luz puede retrasar o adelantar el reloj para adaptarse al medio (Goldsmith and
Bell-Pedersen 2013).
El estímulo luminoso llega desde el tracto retinohipotalámico. Los axones de las
ipRGCs sinaptan con el NSQ y liberan glutamato y PACAP (pituitary adenylate
cyclase-activating Polypeptide). PACAP es modulador del glutamato. Éste se une al
receptor NMDA y AMPA, despolarizando la membrana y abriendo los canales de
calcio, lo que activa una compleja red de kinasas, como las MAPK, que termina
fosforilando CREB (Welsh, Takahashi et al. 2010).
Sincronización entre el marcapasos central y los relojes periféricos Si bien la sincronización entre la red multi-osciladora se ejerce a través de la
melatonina, no es ésta la única hormona que ejerce dicha acción. De esta forma, los
glucocorticoides (corticosterona) (Gerber, Saini et al. 2015), la hormona del crecimiento
y la leptina derivada de los adipocitos tienen también su papel. La insulina participa en
la sincronización de la comida de los relojes periféricos, mientras que la grelina
proporciona señales temporales modulando la anticipación comportamental de la hora
de comer (Challet 2015) Sin embargo, a pesar de que la melatonina no es la única señal
endocrina del reloj circadiano, sí que es la única con un papel específico debido a que su
liberación no se ve influenciada por otros factores como el estrés, sino únicamente por
la presencia/ausencia de luz y es por ello la señal eferente endocrina más estable del
reloj circadiano (Pevet, Agez et al. 2006). Es el sincronizador endógeno del organismo y
actúa a diferentes niveles de los sistemas de temporizado, estabilizando los ritmos
circadianos y reforzándolos en sus relaciones mutuas de fase. Tiene, además,
- 7 -

propiedades cronobióticas al actuar sobre el reloj circadiano, ejerciendo una acción
indirecta a todos los niveles de la red circadiana en los mamíferos (Pevet, Agez et al.
2006) Aunque la deficiencia de melatonina no tiene efecto sobre la generación de los
ritmos o el mantenimiento de su oscilación, sí que es esencial para la señalización de los
ritmos en pars tuberalis de la pituitaria (tiene una gran densidad de receptores MT)
(Stehle, von Gall et al. 2003). En los estudios realizados en animales se ha visto que está
involucrada en la regulación de la expresión circadiana de varios genes reloj, como
Per1, Per2, Bmal1, Rev-Erbα, Clock y Cry en los tejidos diana de la melatonina a nivel
central y periférico, resaltando su papel en la regulación de la expresión de estos genes y
permitiendo la sincronización de los osciladores centrales y periféricos (Charrier, Olliac
et al. 2017). Se cree que los receptores nucleares huérfanos, como Rorβ y Rev-Erbα, son
las dianas iniciales de la melatonina, pudiendo ser regulados por el ritmo circadiano en
pars tuberalis y constituir el enlace entre la acción fisiológica de la melatonina y el
núcleo del reloj circadiano molecular en este tejido. (Agez, Laurent et al. 2009) Actúa
junto con la adenosina suscitando ritmicidad en la expresión de los genes reloj.
Sintetizando adenilciclasa, se abre un canal restringido temporalmente y baja el umbral
para que la adenosina induzca a los genes AMPc-depenedientes. Esto determina una
regulación temporal precisa de la expresión génica que puede reflejar un mecanismo
general por el que el reloj maestro en el cerebro sincroniza a las células reloj en los
tejidos periféricos que requieren señales específicas eferentes de fase. (Stehle, von Gall
et al. 2003)
MicroARN La traducción de ARNm es un evento crítico en la inducción lumínica del reloj. Los
micro ARNs (miARNs) son pequeños fragmentos (de aproximadamente 22 nucleótidos)
de ARN no codificante que juegan un papel significativo en la regulación genética y
expresión proteica. Se unen a regiones 3’ no traducidas (3’UTRs) e inducen
degradación, represión o traducción de sus ARNm diana (Krishnan, Nair et al. 2017)
(Pegoraro and Tauber 2008). Se han identificado cientos de tipos de miARN y se
observó que dos de ellos; miR-219 y miR-132 (dos miARN específicos del cerebro)
influían en el proceso sincronizador, revelando una nueva clase de genes regulados por
reloj (Cheng, Papp et al. 2007). Sucesivamente se fueron observando oscilaciones en
otros miARN, afianzándose la evidencia de una nueva regulación circadiana (Pegoraro
- 8 -

and Tauber 2008). Sucesivos estudios evidencian la acción de miARN como
modificadores postraduccionales del ritmo circadiano (Liu and Wang 2012) siendo
potentes reguladores negativos de la traducción del ARNm ya no sólo en el reloj central,
sino en los relojes periféricos (Shende, Kim et al. 2014), elucidándose mecanismos cada
vez más específicos de regulación así como la implicación de más tipos de miARN
(Mendoza-Viveros, Chiang et al. 2017) (Chen and Rosbash 2016) (Chen and Rosbash
2017)
Vía de MAPK Dentro del NSQ, el estímulo luminoso del reloj está mediado por la familia de proteínas
Ser-Thr kinasas MAPK (Proteína Kinasa Activada por Mitógeno) (Cao, Butcher et al.
2013). La cascada MAPK es una ruta compleja, específica del contexto, reguladora de
un vasto abanico de procesos fisiológicos. Dicha cascada está compuesta de forma
canónica por una jerarquía de tres niveles, comenzando con una MAPK kinasa kinasa
(MAMPKKK) que al ser activada por señalización celular fosforila a una MAPK kinasa
(MAPKK) asociada. Esta fosfo-MAPKK se asocia a su cognado MAPK y la fosforila
(Goldsmith and Bell-Pedersen 2013).
La activación de los receptores postsinápticos por la señal que llega desde el tracto
retinohipotalámico provoca la fosforilación y activación de la kinasa efectora ERK
(Kinasa regulada por señal extracelular) (MAPK p42/44). Aquí, ERK puede actuar
sobre tres sustratos para seguir la vía: MNK (MAPK-interacting serine/
threonine-protein kinase), MSK1 (Proteína Kinasa activada por Estrés y Mitógeno) y el
complejo mTORC1, el cual se comentará más adelante. La fosforilación de MSK1 por
ERK conlleva su activación y ésta se trasloca al núcleo donde activa la vía de
transcripción CREB/CRE mediante la fosforilación en Ser-133 de CREB (cAMP
response element-binding protein). Su activación por fosforilación, un complicado
proceso de varios pasos vía AMPc activado por quinasa y calcio, promueve la
transcripción de genes de respuesta temprana dependiente de CRE (Elementos de
respuesta a AMPc) (Lee, Li et al. 2010) (Sakamoto, Norona et al. 2013) como c-FOS y
Per1, al unirse a sus promotores, y también estimula la expresión del miARN miR-132
(Cao, Li et al. 2010). MSK1 también puede fosforilar la transcripción de genes vía
epigenética fosforilando la histona H3 (Ver figura 2) (Cao, Butcher et al. 2013)
- 9 -

El ritmo de producción y degradación de las proteínas PER determina la duración de un
ritmo circadiano (Takahashi, Hong et al. 2008). Un incremento de los niveles de PER
interrumpe el equilibrio dinámico de los ciclos retroalimentativos, lo que lleva a una
fase de reinicio del reloj del NSQ (Golombek and Rosenstein 2010).
La abundancia de Per1 y Per2 también está controlada a nivel de la traducción de
ARNm y a éste nivel es donde interviene otra vía de MAPK; la vía MAPK/MNK. La
expresión de miARN media la señalización MAPK (Alvarez-Saavedra, Antoun et al.
2011) ERK también puede fosforilar a MNK, activándola y ésta fosforila a eIF4E
(Elongation Initiation Factor 4E) en Ser209 (Cao, Gkogkas et al. 2015). Dicho factor
reconoce y se une a la estructura mGpppN (N puede ser cualquier nucleótido) presente
en el extremo 5’ de todos los ARNm (ARNm 5’ cap) transcritos en el núcleo, junto con
eIF4A y eIF4G (Sonenberg and Hinnebusch 2009) iniciando la traducción.
La vía mTOR La proteína mTOR (Mammalian Target Of Rapamycine) codificada en el gen del
mismo nombre, pertenece a una gran familia de proteínas que comparten entre un 40 –
60% de similitud (Janus, Robak et al. 2005) (Sandsmark, Pelletier et al. 2007) la familia
de las kinasas relacionadas con fosfatidilinositol kinasa (PIKK) poseyendo un dominio
Serina/treonina kinasa. Esta proteína actúa como diana del represor del ciclo celular y
los efectos inmunosupresores del complejo FKBP12-Rapamicina. (PubMed NCBI) Las
proteínas TOR fueron descubiertas por primera vez en la Levadura (Sacharomyces
cerevisiae) en 1991, identificadas por screening genético como diana del potente
macrólido inmunosupresor Rapamicina. (Heitman, Movva et al. 1991) Las proteínas
TOR y sus derivados son relativamente ubicuos y altamente conservados en diversas
especies. Toda célula Eucariota posee un gen que codifica para una proteína TOR, de
forma que en las especies Eucariotas superiores sólo hay un único gen, mientras que
algunas especies de levaduras poseen dos (Wullschleger, Loewith et al. 2006). Se han
implicado en una gran variedad de procesos celulares tales como el crecimiento celular,
proliferación, supervivencia/muerte y metabolismo en respuesta a diversos estímulos
externos (Sandsmark, Pelletier et al. 2007, Dowling, Pollak et al. 2009) así como en la
diferenciación y autorregulación de las células madre embrionarias en humanos (Lee,
Yook et al. 2010) Su acción es esencial en la regulación del crecimiento y proliferación
celular y precisamente es este papel el que la relaciona directamente con multitud de
- 10 -

procesos tumorales cuando esta vía se halla sobreexpresada (Dowling, Pollak et al.
2009) (Deng, Jiang et al. 2017).
TOR actúa como un sensor central para nutrientes y energía, modulándose también por
mecanismos dependientes de PI3K (Fosfatidilinositol 3-Kinasa)/Akt (conocida también
como proteína kinasa B) formando la vía de señalización de mTOR: el eje
PI3K/Akt/mTOR (Ver figura 2). Cuando la insulina o el factor insulínico de crecimiento
(IGF) u otras señales nutricionales se unen al receptor tirosina kinasa, éste recluta y
fosforila al sustrato del receptor insulínico (IRS) y recluta también a IP3K. La unión de
IP3K con IRS fosforila a PIP2 (Fosfatidil inositol bifosfato) a PIP3 en la membrana
celular. PIP3 recluta a PSK1 y Akt a la membrana, resultando en la fosforilación y
activación de Akt (Kijak and Pyza 2017), quien fosforila directamente a mTOR y lo
activa, o inhibe el complejo proteico TSC (Tuberous Sclerosis Complex) TSC1/2. Esto
provoca la hiperactivación de la GTPasa Rheb, la cual es un activador directo de
mTOR. A partir de aquí, puede regular la traducción proteica actuando por dos vías
paralelas: fosforilando y activando la S6K1 (Kinasa ribosomal S6-1), la cual termina
fosforilando a la proteína ribosomal S6, y suprimiendo por fosforilación la elongación
de 4E-BP1 (4E Binding Protein-1), que inhibía por fosforilación la actividad de la
proteína de unión a cap eIF4E. Éstos son factores inductores de la traducción,
importantes para la traducción ARNm cap-dependiente que incrementa los niveles de
muchas proteínas necesarias para la progresión del ciclo celular, proliferación,
angiogénesis y rutas de supervivencia. (Sandsmark, Pelletier et al. 2007) (Gibbons,
Abraham et al. 2009) (Dobashi, Watanabe et al. 2011)
- 11 -
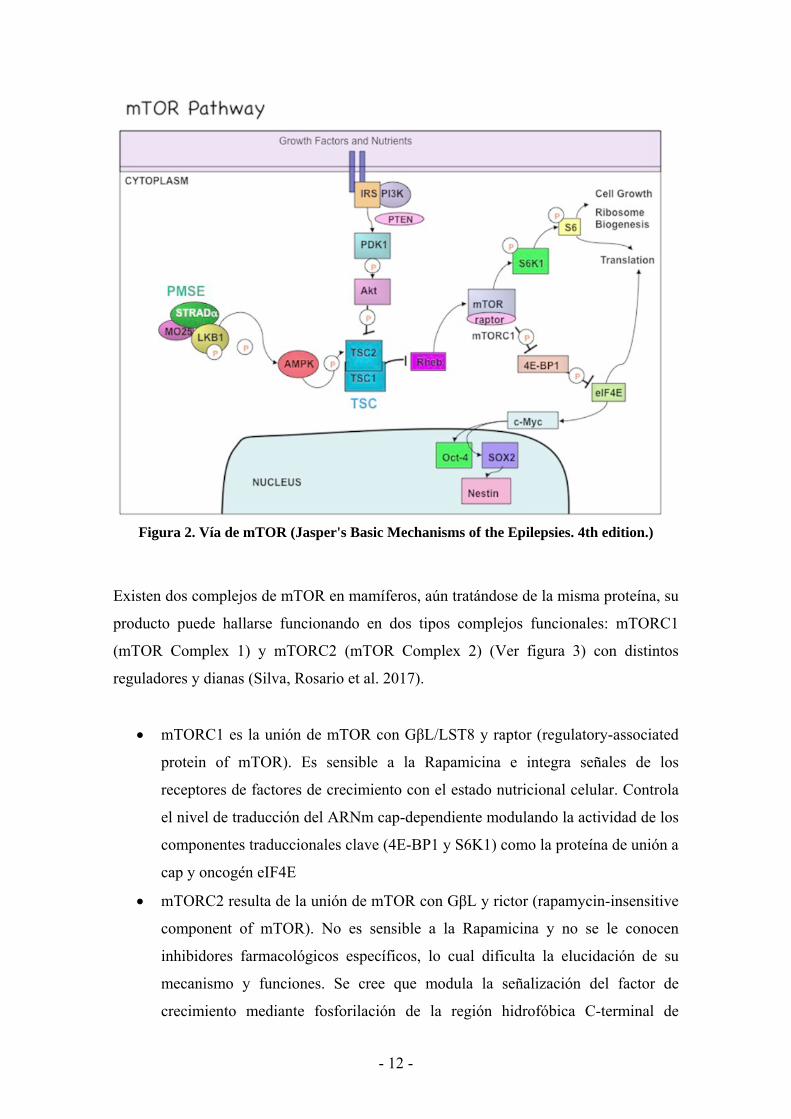
Figura 2. Vía de mTOR (Jasper's Basic Mechanisms of the Epilepsies. 4th edition.)
Existen dos complejos de mTOR en mamíferos, aún tratándose de la misma proteína, su
producto puede hallarse funcionando en dos tipos complejos funcionales: mTORC1
(mTOR Complex 1) y mTORC2 (mTOR Complex 2) (Ver figura 3) con distintos
reguladores y dianas (Silva, Rosario et al. 2017).
• mTORC1 es la unión de mTOR con GβL/LST8 y raptor (regulatory-associated
protein of mTOR). Es sensible a la Rapamicina e integra señales de los
receptores de factores de crecimiento con el estado nutricional celular. Controla
el nivel de traducción del ARNm cap-dependiente modulando la actividad de los
componentes traduccionales clave (4E-BP1 y S6K1) como la proteína de unión a
cap y oncogén eIF4E
• mTORC2 resulta de la unión de mTOR con GβL y rictor (rapamycin-insensitive
component of mTOR). No es sensible a la Rapamicina y no se le conocen
inhibidores farmacológicos específicos, lo cual dificulta la elucidación de su
mecanismo y funciones. Se cree que modula la señalización del factor de
crecimiento mediante fosforilación de la región hidrofóbica C-terminal de
- 12 -
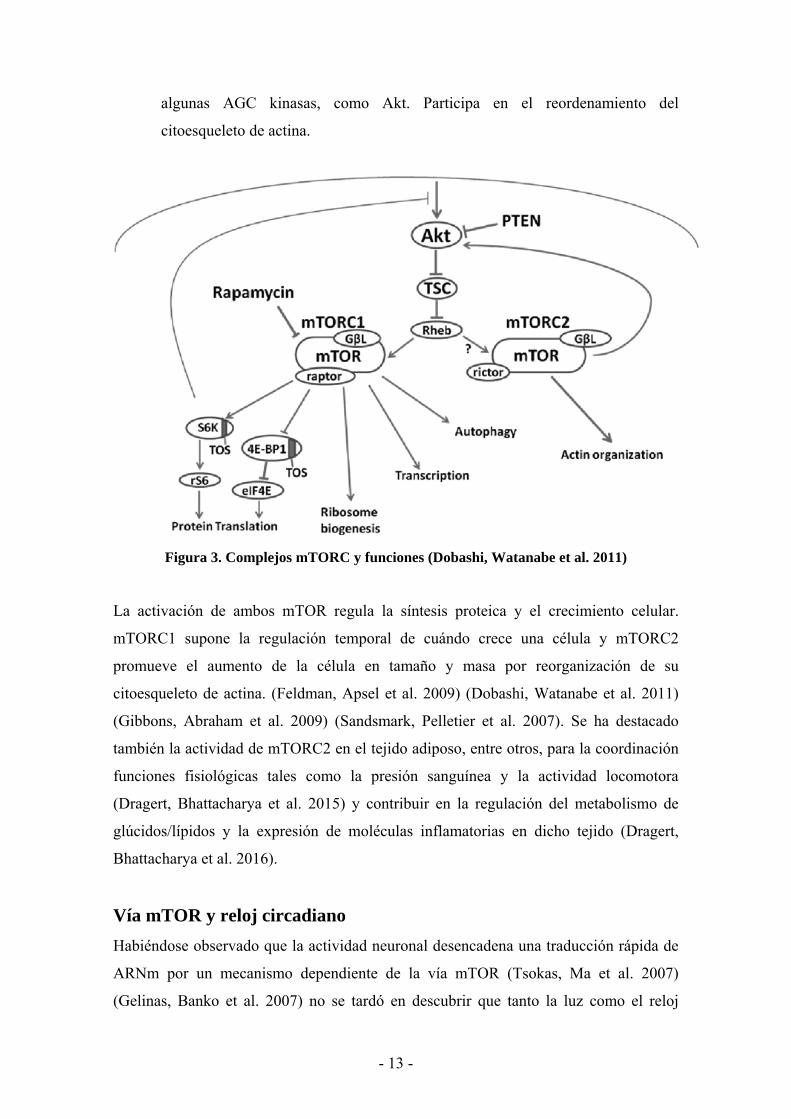
algunas AGC kinasas, como Akt. Participa en el reordenamiento del
citoesqueleto de actina.
Figura 3. Complejos mTORC y funciones (Dobashi, Watanabe et al. 2011)
La activación de ambos mTOR regula la síntesis proteica y el crecimiento celular.
mTORC1 supone la regulación temporal de cuándo crece una célula y mTORC2
promueve el aumento de la célula en tamaño y masa por reorganización de su
citoesqueleto de actina. (Feldman, Apsel et al. 2009) (Dobashi, Watanabe et al. 2011)
(Gibbons, Abraham et al. 2009) (Sandsmark, Pelletier et al. 2007). Se ha destacado
también la actividad de mTORC2 en el tejido adiposo, entre otros, para la coordinación
funciones fisiológicas tales como la presión sanguínea y la actividad locomotora
(Dragert, Bhattacharya et al. 2015) y contribuir en la regulación del metabolismo de
glúcidos/lípidos y la expresión de moléculas inflamatorias en dicho tejido (Dragert,
Bhattacharya et al. 2016).
Vía mTOR y reloj circadiano Habiéndose observado que la actividad neuronal desencadena una traducción rápida de
ARNm por un mecanismo dependiente de la vía mTOR (Tsokas, Ma et al. 2007)
(Gelinas, Banko et al. 2007) no se tardó en descubrir que tanto la luz como el reloj
- 13 -

circadiano modulaban la expresión de miARN en el NSQ (Cheng, Papp et al. 2007). Se
estudió la regulación de la señalización dependiente de mTOR en el reloj del NSQ y
pudo observarse que la estimulación lumínica provocaba un incremento dependiente de
fase en el estado activado de p70, S6K y E4-BP1 (que son efectores directos de mTOR)
y una activación coordinada de CREB y mTOR en las neuronas del NSQ, además de
que mTOR es diana de la vía de señalización de la MAPK p42/44 (Cao, Lee et al.
2008). Resultó que en el NSQ había una fuerte expresión de mTOR, mayor que en los
tejidos circundantes (Cao and Obrietan 2010). Todo ello evidenció que en el NSQ
existía una cascada mTOR sensible a la luz. Otros estudios señalaron el vínculo que
existía en el tejido adiposo, fuertemente regulado por el ritmo diurno, entre ambas vías
evidenciado por la asociación significativa con los transcritos reprimidos por los
inhibidores de mTOR (Loboda, Kraft et al. 2009). Además, mediante el uso de la
proteína ribosomal S6 fosforilada como marcador de la actividad de mTOR, se
demostró que su actividad mostraba una marcada oscilación circadiana en el NSQ,
siendo su actividad máxima durante el día subjetivo y mínima durante la noche
subjetiva (Cao, Anderson et al. 2011).
MAPK e inducción de mTOR por luzHabiéndose descartado la posibilidad de que fuera la ruta PI3K/Akt la que acoplara el
estímulo lumínico a mTOR, se vio que era a través de la ruta MAPK que esto ocurría
(Cao and Obrietan 2010). ERK es la encargada de fosforilar al complejo mTORC1 que,
como ya se ha visto anteriormente, provoca la activación por fosforilación de p70
S6K/S6 que estimulan la traducción de ARNm TOP, así como la inhibición de 4E-BP1
y, consecuentemente eIF4E, lo que disminuye la traducción de ARN dependiente de
CAP (Ver figura 4) (Cao, Li et al. 2010) (Cao, Anderson et al. 2011) La vía de
mTORC1 y MAPK coinciden en la regulación de eIF4E1. El control de la traducción
mediada por mTOR/4E-BP1 es clave en la regulación de la inducción y sincronía del
reloj maestro (Cao, Robinson et al. 2013) (Ver figura 4)
Aunque mTORC1 es el principal implicado en el reloj circadiano, se ha observado que
mTORC2 también desempeña un papel en la regulación del reloj a nivel periférico y no
en el NSQ: Se comprobó en ratones que la ausencia de Rictor reducía fuertemente la
fosforilación de Akt, resultando en una señalización dañada de mTORC2. Se observó
que Rictor/mTORC2 contribuía en la regulación diurna de la presión sanguínea y
- 14 -

actividad locomotora en cerebro y tejido adiposo (Dragert, Bhattacharya et al. 2015).
Además poco después se comprobó una regulación diurna de los componentes de
mTORC2, Rictor y mSIN1 en el tejido adiposo (Dragert, Bhattacharya et al. 2016).
Sin embargo, el luminoso no es el único estímulo circadiano de mTOR. Se halló que
existía un oscilador interno diferente del estimulo lumínico, aunque ligado a él que
regulaba la actividad de la vía mTOR (Khapre, Patel et al. 2014). No tardó en
demostrarse que la deficiencia de BMAL1 in vivo suponía un incremento en la
actividad de mTORC1, demostrándose que era un regulador negativo de dicha vía
(Khapre, Kondratova et al. 2014). Así se pudo proponer que el reloj circadiano era un
regulador maestro del metabolismo.
BMAL1 es, así mismo, sustrato para fosforilación mediada por S6K1, lo que es esencial
para que interactúe con la maquinaria traduccional. BMAL1 es un factor traduccional
que estimula la síntesis proteica, enlazándose así el ritmo circadiano con la misma y con
la vía mTOR (Lipton, Yuan et al. 2015). Pudo comprobarse en ratones que la mayor
activación de mTORC1 y 2 resultaba en una acumulación de BMAL1, restaurándose los
niveles de ésta con la inhibición de mTORC1 y 2 (Matsumoto, Almeida et al. 2016).
Además de BMAL1, recientemente se estudió el papel de Per2 en la vía mTOR en las
células de adenocarcinoma: el brusco descenso de Per2 resultó en un incremento de la
activación de PI3K/Akt/mTOR (Chen, Tan et al. 2017).
La Glucógeno sintasa kinasa 3 β (GSK3), un punto crítico para la regulación de la
síntesis de Glucógeno, está regulada por Akt (Loboda, Kraft et al. 2009)
Figura 4. Inducción de mTOR por la luz y visión integrada con la expresión de genes
circadianos (Cao, Li et al. 2010).
- 15 -

Implicaciones Las implicaciones que el reloj circadiano tiene en el metabolismo son múltiples y la
participación en el mismo de mTOR permite conocer el alcance de las mismas. Por
ejemplo, la proteína Per2 tiene un efecto supresor de tumores y su relación con la vía
PI3K/Akt/mTOR sugiere que está implicada en la aparición de resistencia a fármacos en
tumores (Chen, Tan et al. 2017). La desregulación del reloj circadiano, así como un
aumento en la señalización de la vía mTOR, puede derivar en patologías cancerígenas,
ya que ciertos oncogenes circadianos como MYC, al verse sobreexpresados favorecen la
aparición y desarrollo de las células cancerosas (Dang 2016) En anteriores estudios se
ha podido relacionar la desregulación de mTOR con la aparición de muchos tipos de
cáncer (Bajwa, Nielsen et al. 2017) (Johnson, O'Connell et al. 2017).
Pero los usos de mTOR y su participación circadiana pueden tener variados usos, por
ejemplo mTOR fosforilado (p-mTOR) es un pronóstico positivo en el cáncer de
estómago (Cao, Xu et al. 2016). La oscilación circadiana de la actividad de mTOR,
regulada por el reloj, influye en el efecto antitumoral de los inhibidores de mTOR
(Okazaki, Matsunaga et al. 2014).
A nivel metabólico, se ha observado que la disrupción de los genes reloj resulta en
dislipemia, insulinoresistencia y obesidad, modificando los comportamientos
alimenticios (Engin 2017)
La función circadiana con participación mTOR se ha mostrado de gran impacto en el
desarrollo y tratamiento de enfermedades del sistema nervioso. Por ejemplo, los niveles
disminuidos de BDNF (Brain-derived neurotrophic factor) se asocian con desarrollo de
obesidad y neurodegeneración y su expresión está asociado a la señalización circadiana
y la alimentación (Genzer, Dadon et al. 2016). El factor de supervivencia Akt y BMAL1
han servido para observar que el daño neuronal por infarto cerebral es menor durante la
noche, habiendo más supervivencia neuronal, que durante el día (Beker, Caglayan et al.
2017).
Sus implicaciones pasan, por supuesto por el crecimiento celular y su diferenciación,
recibiendo una influencia circadiana; la señalización PDK1-mTOR-B4BP4-CD122 se
ha probado importante en el desarrollo de las células NK (Samsa, Zhou et al. 2017).
- 16 -

Conclusión Con todo lo revisado se puede concluir que la vía mTOR y el reloj circadiano están muy
íntimamente ligados. La vía de señalización mTOR recibe inducción directa por el
estímulo lumínico y modula el propio reajuste del reloj a través de sus efectores S6k/S6,
implicada además en la fosforilación de proteínas reloj como BMAL1, y sIF4E. Éste, a
su vez influye en la vía mTOR, modulando su actividad mediante la expresión de las
proteínas reloj BMAL1 y Per principalmente. Un punto clave en común del ritmo
circadiano y la vía mTOR son la familia de las kinasas MAPK que forman parte tanto
de la vía mTOR como de la activación de la transcripción circadiana por CREB/CRE.
Las implicaciones de la influencia circadiana de mTOR son múltiples y ayudan a
vislumbrar mecanismos hasta ahora desconocidos en enfermedades cancerígenas,
metabólicas, alimenticias, comportamentales y neurológicas, allanando el camino a
nuevas terapias y prevenciones.
Por todo lo anterior, es interesante continuar investigando y seguir esclareciendo
mecanismos aún ignotos en la conjunción de ambas vías así como de sus implicaciones.
Bibliografía Buijs, F. N., et al. (2017). "Suprachiasmatic Nucleus Interaction with the Arcuate Nucleus; Essential for Organizing Physiological Rhythms." eNeuro 4(2).
Agez, L., et al. (2009). "Endogenous melatonin provides an effective circadian message to both the suprachiasmatic nuclei and the pars tuberalis of the rat." J Pineal Res 46(1): 95-105.
Cao, G. D., et al. (2016). "Phosphorylated Mammalian Target of Rapamycin p-mTOR Is a Favorable Prognostic Factor than mTOR in Gastric Cancer." PLoS One 11(12): e0168085.
Alvarez-Saavedra, M., et al. (2011). "miRNA-132 orchestrates chromatin remodeling and translational control of the circadian clock." Hum Mol Genet 20(4): 731-751.
Cao, R., et al. (2011). "Circadian regulation of mammalian target of rapamycin signaling in the mouse suprachiasmatic nucleus." Neuroscience 181: 79-88.
Bajwa, P., et al. (2017). "Overactive mTOR signaling leads to endometrial hyperplasia in aged women and mice." Oncotarget 8(5): 7265-7275.
Cao, R., et al. (2013). "Mitogen- and stress-activated protein kinase 1 modulates photic entrainment of the suprachiasmatic circadian clock." Eur J Neurosci 37(1): 130-140.
Beker, M. C., et al. (2017). "Time-of-Day Dependent Neuronal Injury After Ischemic Stroke: Implication of Circadian Clock Transcriptional Factor Bmal1 and Survival Kinase AKT." Mol Neurobiol.
Cao, R., et al. (2015). "Light-regulated translational control of circadian behavior by eIF4E phosphorylation." Nat Neurosci 18(6): 855-862.
Berson, D. M. (2003). "Strange vision: ganglion cells as circadian photoreceptors." Trends Neurosci 26(6): 314-320.
Cao, R., et al. (2008). "Photic regulation of the mTOR signaling pathway in the suprachiasmatic circadian clock." Mol Cell Neurosci 38(3): 312-324.
Berson, D. M., et al. (2002). "Phototransduction by retinal ganglion cells that set the circadian clock." Science 295(5557): 1070-1073.
- 17 -

Cao, R., et al. (2010). "Mammalian target of rapamycin signaling modulates photic entrainment of the suprachiasmatic circadian clock." J Neurosci 30(18): 6302-6314. Cao, R. and K. Obrietan (2010). "mTOR Signaling and Entrainment of the Mammalian Circadian Clock." Mol Cell Pharmacol 2(4): 125-130. Cao, R., et al. (2013). "Translational control of entrainment and synchrony of the suprachiasmatic circadian clock by mTOR/4E-BP1 signaling." Neuron 79(4): 712-724. Cassone, V. M. (1992). "The pineal gland influences rat circadian activity rhythms in constant light." J Biol Rhythms 7(1): 27-40. Challet, E. (2015). "Keeping circadian time with hormones." Diabetes Obes Metab 17 Suppl 1: 76-83. Charrier, A., et al. (2017). "Clock Genes and Altered Sleep-Wake Rhythms: Their Role in the Development of Psychiatric Disorders." Int J Mol Sci 18(5). Chen, B., et al. (2017). "Per2 participates in AKT-mediated drug resistance in A549/DDP lung adenocarcinoma cells." Oncol Lett 13(1): 423-428. Chen, X. and M. Rosbash (2016). "mir-276a strengthens Drosophila circadian rhythms by regulating timeless expression." Proc Natl Acad Sci U S A 113(21): E2965-2972. Chen, X. and M. Rosbash (2017). "MicroRNA-92a is a circadian modulator of neuronal excitability in Drosophila." Nat Commun 8: 14707. Cheng, H. Y., et al. (2007). "microRNA modulation of circadian-clock period and entrainment." Neuron 54(5): 813-829. Choi, T. Y., et al. (2014). "Melatonin inhibits voltage-sensitive Ca(2+) channel-mediated neurotransmitter release." Brain Res 1557: 34-42. Dang, C. V. (2016). "A Time for MYC: Metabolism and Therapy." Cold Spring Harb Symp Quant Biol 81: 79-83. Deng, L., et al. (2017). "The PI3K/mTOR dual inhibitor BEZ235 suppresses proliferation and migration and reverses multidrug resistance in
acute myeloid leukemia." Acta Pharmacol Sin 38(3): 382-391. Dobashi, Y., et al. (2011). "Mammalian target of rapamycin: a central node of complex signaling cascades." Int J Clin Exp Pathol 4(5): 476-495. Dominguez-Solis, C. A. and J. A. Perez-Leon (2015). "[Phototransduction mediated by melanopsin in intrinsically photosensitive retinal ganglion cells]." Gac Med Mex 151(6): 764-776. Dowling, R. J., et al. (2009). "Current status and challenges associated with targeting mTOR for cancer therapy." BioDrugs 23(2): 77-91. Dragert, K., et al. (2016). "Basal mTORC2 activity and expression of its components display diurnal variation in mouse perivascular adipose tissue." Biochem Biophys Res Commun 473(1): 317-322. Dragert, K., et al. (2015). "Deletion of Rictor in brain and fat alters peripheral clock gene expression and increases blood pressure." Hypertension 66(2): 332-339. Engin, A. (2017). "Circadian Rhythms in Diet-Induced Obesity." Adv Exp Med Biol 960: 19-52. Feldman, M. E., et al. (2009). "Active-site inhibitors of mTOR target rapamycin-resistant outputs of mTORC1 and mTORC2." PLoS Biol 7(2): e38. Foster, R. G. (1998). "Shedding light on the biological clock." Neuron 20(5): 829-832. Gelinas, J. N., et al. (2007). "ERK and mTOR signaling couple beta-adrenergic receptors to translation initiation machinery to gate induction of protein synthesis-dependent long-term potentiation." J Biol Chem 282(37): 27527-27535. Genzer, Y., et al. (2016). "Effect of dietary fat and the circadian clock on the expression of brain-derived neurotrophic factor (BDNF)." Mol Cell Endocrinol 430: 49-55. Gerber, A., et al. (2015). "The systemic control of circadian gene expression." Diabetes Obes Metab 17 Suppl 1: 23-32. Gibbons, J. J., et al. (2009). "Mammalian target of rapamycin: discovery of rapamycin reveals a signaling pathway important for normal and
18

cancer cell growth." Semin Oncol 36 Suppl 3: S3-S17.
Lee, B., et al. (2010). "CREB influences timing and entrainment of the SCN circadian clock." J Biol Rhythms 25(6): 410-420. Goldsmith, C. S. and D. Bell-Pedersen (2013).
"Diverse roles for MAPK signaling in circadian clocks." Adv Genet 84: 1-39.
Lee, K. W., et al. (2010). "Rapamycin promotes the osteoblastic differentiation of human embryonic stem cells by blocking the mTOR pathway and stimulating the BMP/Smad pathway." Stem Cells Dev 19(4): 557-568.
Golombek, D. A. and R. E. Rosenstein (2010). "Physiology of circadian entrainment." Physiol Rev 90(3): 1063-1102.
Lincoln, G., et al. (2002). "Temporal expression of seven clock genes in the suprachiasmatic nucleus and the pars tuberalis of the sheep: evidence for an internal coincidence timer." Proc Natl Acad Sci U S A 99(21): 13890-13895.
Hardeland, R., et al. (2011). "Melatonin--a pleiotropic, orchestrating regulator molecule." Prog Neurobiol 93(3): 350-384. Heitman, J., et al. (1991). "Targets for cell cycle arrest by the immunosuppressant rapamycin in yeast." Science 253(5022): 905-909.
Lipton, J. O., et al. (2015). "The Circadian Protein BMAL1 Regulates Translation in Response to S6K1-Mediated Phosphorylation." Cell 161(5): 1138-1151.
Huo, X., et al. (2017). "Human transporters, PEPT1/2, facilitate melatonin transportation into mitochondria of cancer cells: An implication of the therapeutic potential." J Pineal Res 62(4).
Liu, C., et al. (2014). "Melatonin synergized with cyclosporine A improves cardiac allograft survival by suppressing inflammation and apoptosis." Mol Med Rep 10(3): 1323-1328.
Janus, A., et al. (2005). "The mammalian target of the rapamycin (mTOR) kinase pathway: its role in tumourigenesis and targeted antitumour therapy." Cell Mol Biol Lett 10(3): 479-498.
Liu, K. and R. Wang (2012). "MicroRNA-mediated regulation in the mammalian circadian rhythm." J Theor Biol 304: 103-110.
Johnson, M. D., et al. (2017). "mTOR activation is increased in pilocytic astrocytomas from older adults compared with children." Surg Neurol Int 8: 85.
Loboda, A., et al. (2009). "Diurnal variation of the human adipose transcriptome and the link to metabolic disease." BMC Med Genomics 2: 7.
Kalsbeek, A., et al. (2006). "A network of (autonomic) clock outputs." Chronobiol Int 23(1-2): 201-215.
Manka, S. and E. Majewska (2016). "Immunoregulatory action of melatonin. The mechanism of action and the effect on inflammatory cells." Postepy Hig Med Dosw (Online) 70(0): 1059-1067.
Khapre, R. V., et al. (2014). "BMAL1-dependent regulation of the mTOR signaling pathway delays aging." Aging (Albany NY) 6(1): 48-57.
Matsumoto, C. S., et al. (2016). "PI3K-PTEN dysregulation leads to mTOR-driven upregulation of the core clock gene BMAL1 in normal and malignant epithelial cells." Oncotarget 7(27): 42393-42407.
Khapre, R. V., et al. (2014). "Metabolic clock generates nutrient anticipation rhythms in mTOR signaling." Aging (Albany NY) 6(8): 675-689. Mazzoccoli, G., et al. (2012). "Clock genes and
clock-controlled genes in the regulation of metabolic rhythms." Chronobiol Int 29(3): 227-251.
Kijak, E. and E. Pyza (2017). "TOR signaling pathway and autophagy are involved in the regulation of circadian rhythms in behavior and plasticity of L2 interneurons in the brain of Drosophila melanogaster." PLoS One 12(2): e0171848.
Mendoza-Viveros, L., et al. (2017). "miR-132/212 Modulates Seasonal Adaptation and Dendritic Morphology of the Central Circadian Clock." Cell Rep 19(3): 505-520.
Krishnan, R., et al. (2017). "Computational study of 'HUB' microRNA in human cardiac diseases." Bioinformation 13(1): 17-20.
Okazaki, H., et al. (2014). "Circadian regulation of mTOR by the ubiquitin pathway in renal cell carcinoma." Cancer Res 74(2): 543-551.
19

Panda, S., et al. (2005). "Illumination of the melanopsin signaling pathway." Science 307(5709): 600-604. Pandi-Perumal, S. R., et al. (2008). "Physiological effects of melatonin: role of melatonin receptors and signal transduction pathways." Prog Neurobiol 85(3): 335-353. Pegoraro, M. and E. Tauber (2008). "The role of microRNAs (miRNA) in circadian rhythmicity." J Genet 87(5): 505-511. Peirson, S. and R. G. Foster (2006). "Melanopsin: another way of signaling light." Neuron 49(3): 331-339. Pevet, P. (2014). "The internal time-giver role of melatonin. A key for our health." Rev Neurol (Paris) 170(11): 646-652. Pevet, P., et al. (2006). "Melatonin in the multi-oscillatory mammalian circadian world." Chronobiol Int 23(1-2): 39-51. Pevet, P. and E. Challet (2011). "Melatonin: both master clock output and internal time-giver in the circadian clocks network." J Physiol Paris 105(4-6): 170-182. Qiu, X., et al. (2005). "Induction of photosensitivity by heterologous expression of melanopsin." Nature 433(7027): 745-749. Reiter, R. J., et al. (2010). "Melatonin: a multitasking molecule." Prog Brain Res 181: 127-151. Sakamoto, K., et al. (2013). "Clock and light regulation of the CREB coactivator CRTC1 in the suprachiasmatic circadian clock." J Neurosci 33(21): 9021-9027. Samsa, W. E., et al. (2017). "Signaling pathways regulating cartilage growth plate formation and activity." Semin Cell Dev Biol 62: 3-15. Sandsmark, D. K., et al. (2007). "Mammalian target of rapamycin: master regulator of cell
growth in the nervous system." Histol Histopathol 22(8): 895-903. Schibler, U., et al. (2015). "Clock-Talk: Interactions between Central and Peripheral Circadian Oscillators in Mammals." Cold Spring Harb Symp Quant Biol 80: 223-232. Shende, V. R., et al. (2014). "MicroRNAs function as cis- and trans-acting modulators of peripheral circadian clocks." FEBS Lett 588(17): 3015-3022. Silva, E., et al. (2017). "mTOR Is a Novel Molecular Mechanism Linking Folate Availability and Cell Function." J Nutr. Sonenberg, N. and A. G. Hinnebusch (2009). "Regulation of translation initiation in eukaryotes: mechanisms and biological targets." Cell 136(4): 731-745. Stehle, J. H., et al. (2003). "Melatonin: a clock-output, a clock-input." J Neuroendocrinol 15(4): 383-389. Takahashi, J. S., et al. (2008). "The genetics of mammalian circadian order and disorder: implications for physiology and disease." Nat Rev Genet 9(10): 764-775. Touitou, Y. (2016). "[The circadian system in man: From the internal clock to melatonin secretion]." Ann Pharm Fr 74(5): 331-334. Tsokas, P., et al. (2007). "Mitogen-activated protein kinase upregulates the dendritic translation machinery in long-term potentiation by controlling the mammalian target of rapamycin pathway." J Neurosci 27(22): 5885-5894. Welsh, D. K., et al. (2010). "Suprachiasmatic nucleus: cell autonomy and network properties." Annu Rev Physiol 72: 551-577. Wullschleger, S., et al. (2006). "TOR signaling in growth and metabolism." Cell 124(3): 471-484.
20