Embed Size (px)
Citation preview

Di r ecci ó n:Di r ecci ó n: Biblioteca Central Dr. Luis F. Leloir, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Intendente Güiraldes 2160 - C1428EGA - Tel. (++54 +11) 4789-9293
Co nta cto :Co nta cto : [email protected]
Tesis Doctoral
Trazas fósiles de coleópterosTrazas fósiles de coleópteroscoprófagos del cenozoico de lacoprófagos del cenozoico de laPatagonia Central. SignificadoPatagonia Central. Significado
evolutivo y paleoambientalevolutivo y paleoambiental
Sánchez, María Victoria
2009
Este documento forma parte de la colección de tesis doctorales y de maestría de la BibliotecaCentral Dr. Luis Federico Leloir, disponible en digital.bl.fcen.uba.ar. Su utilización debe seracompañada por la cita bibliográfica con reconocimiento de la fuente.
This document is part of the doctoral theses collection of the Central Library Dr. Luis FedericoLeloir, available in digital.bl.fcen.uba.ar. It should be used accompanied by the correspondingcitation acknowledging the source.
Cita tipo APA:
Sánchez, María Victoria. (2009). Trazas fósiles de coleópteros coprófagos del cenozoico de laPatagonia Central. Significado evolutivo y paleoambiental. Facultad de Ciencias Exactas yNaturales. Universidad de Buenos Aires.
Cita tipo Chicago:
Sánchez, María Victoria. "Trazas fósiles de coleópteros coprófagos del cenozoico de laPatagonia Central. Significado evolutivo y paleoambiental". Facultad de Ciencias Exactas yNaturales. Universidad de Buenos Aires. 2009.

�
�
����������������������
Facultad�de�Ciencias�Exactas�y�Naturales�
�
�
�
�
������������������ ����������������
������������� ������� �������������
����� ������������� �
�
������������������������� �����������������������������������
�����������������������������������
�
������ ����������
�
�
�
�
Directores�de�Tesis:������������� !"�#����$%&�$�����''�"!
Consejero�de�Estudios:�����$�&�$�����(����
�
�
�
Lugar�de�trabajo:�División�Icnología���Museo�Argentino�de�Ciencias�
Naturales�“Bernardino�Rivadavia”�
�
Buenos�Aires,�2009�

� ��
������������������ ����������������
������������� ������� �������������
����� ������������� �
�
�
�������
Las� bolas� de� cría� de� Scarabaeinae� se� conservan� en� el� registro� geológico�
como� trazas� pertenecientes� al� icnogénero� �����������.� Este� incluía� cinco�
icnoespecies:���� ���������,�����������,����������,�����������y����������� ��.�
Se� realizó� una� revisión� icnotaxonómica� que� resultó� en� la� descripción� de� una�
icnoespecie�nueva�(���isp.�B),�se�estableció�que���� ����������Sauer�1955�es�
un� sinónimo� junior� de� ��� �������� Roselli� 1939,� mientras� que� ��� ��������
������� pasó� a� tener� un� nuevo� nombre� provisorio,� ��� isp.� A.� Para� adjudicar�
estas�icnoespecies�a�posibles�productores�actuales,�se�estudiaron�13�especies�de�
Scarabaeinae� pertenecientes� a� 4� de� las� 5� tribus� presentes� en� la� Argentina.� La�
micro��y�macromorfología�de�sus�bolas�de�cría�permitió�adjudicar�a�����������y�
����������a�Phanaeini�y����isp.�B�a�Canthonini.�Estos�registros�estarían�predatando�
en� 30�40�millones� de� años� a� los� fósiles� corpóreos�más� antiguos� conocidos� de�
estas� tribus,� contribuyendo� significativamente� al� conocimiento� de� su� historia�
evolutiva�en�Sudamérica.��
Con� el� objetivo� de� realizar� un� estudio� de� icnoestratigrafía� de� alta�
resolución,� se� colectaron� 1200� ejemplares� en� 14� niveles� estratigráficos�
(paleosuelos)� de� la� Formación� Sarmiento� (Eoceno� Medio�� Mioceno� inferior)� en�
Gran�Barranca�(Chubut,�Argentina).�Se�midió�la�densidad�de�ejemplares�por�m2,�
abundancia�relativa�de�icnoespecies,�diámetro�ecuatorial�y�espesor�de�la�pared.�
El�análisis�de�estos�datos�permitió�estimar�el�número�de�especies�productoras�así�

� ��
como�también�distinguir�6�episodios,�en�la�evolución�de�la�fauna�de�escarabajos�
coprófagos� y� de� los� pastizales� asociados,� para� Patagonia.� Se� registraron� y�
describieron� dos� trazas� fósiles� (�������� ����� ���� y� ��� ����� ����������)�
excavadas�en�las������������,�que�revelaron�la�presencia�de�cleptoparásitos�y�
detritívoros� en� la� comunidad� asociada� al� excremento.� Se� analizó� el� contenido�
fitolítico� de������������� y� las� diferencias� encontradas� en� el� número� total� y�
frecuencias� de� morfotipos� de� fitolitos,� entre� pared,� relleno� y� paleosuelo,�
mostraron�que� la�primera�estaba�adicionada�con�bosta.�Esta�evidencia�permitió�
inferir� aspectos� del� comportamiento� de� los� coprófagos� y� la� dieta� de� los�
herbívoros.��
�
�&'&)�&" *'&+�, Scarabaeinae,� bolas� de� cría,� icnogénero� �����������,�
registro�fósil,�Cenozoico,�Patagonia�Central,�fitolitos,�trazas�asociadas.���
�
�
�
�
�
�
�
�
�
�
�
�
�

� ��
������������������������ ������� ��
������������� ��� �����
�������������� �������� �����
�����������
�
�
�� �� �
Scarabaeinae� brood� balls� preserved� in� the� geological� record� as� traces�
belong� to� the� ������������ ichnogenus,� which� included� five� icnospecies:� ���
���������,� ��� �������,� ��� ������,� ��� ������� y� ��� ������� ��.� The�
ichnotaxonomic�revision�resulted�in:�a)�the�description�of�a�new�ichnospecies�(���
isp.� B),� b)� ��� ���������� Sauer� 1955� is� a� junior� synomym� of� ��� ��������
Roselli�1939,�and�c)�������������������has�a�new�and�provisory�name,����isp.�
A.� Thirteen� Scarabaeinae� species,� belonging� to� 4� of� the� 5� tribes� inhabiting�
Argentina,� were� studied� in� order� to� attribute� these� ichnospecies� to� possible�
extant� producers.� Micro�� and� macromorphology� of� the� brood� balls� allowed� to�
attribute�����������and�����������to�Phanaeini,�and����isp.�B�to�Canthonini.�These�
records,� which� predate� 30�40�millon� of� years� the� oldest� body� fossils� of� these�
tribes,�contribute�significatively�to�its�evolutionary�history�in�South�America.�
A�high�resolution� ichnostratigraphic� study�was�performed�based�on�1200�
specimens� collected� from� 14� stratigraphic� levels� (paleosols)� in� the� Sarmiento�
Formation� (Middle� Eocene�Lower� Miocene)� at� Gran� Barranca� locality� (Chubut,�
Argentina).� Density� per� square� meter,� relative� abundance� of� ichnospecies,�
equatorial� diameter,� and�wall� thickness�were�measured.� These�data� allowed� to�
estimate�the�number�of�producers�and�to�dintinguish�6�episodes�in�the�evolution�
of� dung� beetles� and� of� the� related� grasslands� of� Patagonia.� Two� trace� fossils,�

� ��
excavated� in������������� (�������� ��������� and���� ����� ����������),�
were�described.�They� indicated� the�presence�of� cleptoparasites�and�detritivores�
from�the�dung�community.�The�analysis�of�the�differences�in�the�total�number�of�
phytoliths� and�morphotype� frequencies� among������������� wall,� filling,� and�
paleosol,� showed� that� the� original� soil� wall� was� added� with� dung� fibers.� This�
evidence�revealed�aspects�of�dung�beetle�behavior�and�mammal�diets.��
����
-�./��$",Scarabaeinae,�brood�balls,� icnogenus������������,�fossil�record,�
Cenozoic,�Central�Patagonia,�phytoliths,�associated�traces.���

� ��
���������� ��
Quiero�agradecer�en�primer�lugar�a�Jorge�F.�Genise,�mi�director�de�Tesis,�por�
darme�la�oportunidad,�por�acompañarme�y�ayudarme�en�el�campo,�por�cada�uno�de�
los�trabajos�que�emprendimos�y�concluimos�juntos,�por�el�sostén�de�todos�los�días,�y�
por�enseñarme�a�disfrutar�y�amar�esta�profesión…�
A�Eduardo�S.�Bellosi,�mi�codirector�de�Tesis,�por�todo�el�apoyo�que�me�brindó�
siempre,� por� transmitirme� sus� conocimientos� geológícos� en� el� campo� y� en� el�
laboratorio,�y�por�su�ayuda�cotidiana…���
A�ambos,�gracias�por�confiar�en�mí�y�por�todo�lo�que�aprendí�y�crecí�junto�a�
Uds.� durante� este� tiempo,� por� su�guía� permanente� y�por� estar� presentes�hasta� el�
último�momento.�Gracias�por� las� increíbles� salidas�de� campo,� las� viví� y�aproveché�
tanto�que�las�conservaré�siempre�en�mis�recuerdos�(y�en�mi�“Querido�Diario”)…��
A� “Pepe”� (José� Laza),� por� enseñarme� la� preparación� de� las������������,�
por�trasmitirme�sus�conocimientos�y�su�pasión�por�los�escarabajos,�por�pasarme�toda�
su�bibliografía�y�su�costumbre�de�guardar�cajitas,�por�la�música�clásica�y�los�mates�
compartidos�cada�mañana…�
A�Mirta�González,�“Mirchus”,�por�su�permanente�ayuda�en�la�micromorfología�
de� las�trazas,�por� trabajar�a� la�par�mío�en�el�análisis� fitolítico,�por�su�paciencia�de�
docente� y� por� su� enseñanza� de� la� geología� general,� y� fundamentalmente,� por� su�
compañía�cotidiana�y�por�las�charlas�compartidas�cada�día�de�escritorio�a�escritorio...�
A� Lauri� (Laura� Sarzetti),� por� estar� siempre� predispuesta� a� salir� al� campo�
conmigo,�por�esas�tardes�de�tierra�debajo�de�las�uñas�y�bosta�en�las�alpargatas,�por�
los�almuerzos�con�Victorinox,�por�los�nervios�pre�charlas�de�congresos,�por�las�horas�
de�canto�en�bondis�y�camionetas,�y�por�convertirse�en�una�gran�amiga...�
�A�Marce� (Marcelo� Krause),� por� las� tardes� de���� ����� y� tablas� excell,� por�
todo�lo�compartido�en�Gran�Barranca,�mi�primera�salida�al�campo,�por�las�caminatas�
eternas�en�las�tardes�calurosas�de�Anillaco�y�las�noches�de�filmación,�y�por�todos�los�
buenos�momentos�compartidos�y�vividos�juntos…�
A�Juan�Farina�y�Daniel�Speranza�por�estar�presentes�y�continuamente�atentos�
a�mi�trabajo,�por�colaborar�conmigo�en�el�campo�y�en�el� laboratorio,�por�hacer� los�
coros�en�los�maravillosos�viajes�de�campo,�por�los�“parches�en�los�ojos”�y�los�“tecitos�
danzantes”…�
A�Ricardo�Melchor,�Mariano�Verde�y�Emilio�Bedatou,�por�Baqueró,��Formosa,�
Ushuaia,�San�Luis,�y�demás�viajes�compartidos…todos�fantásticos…�

� ��
A�Guillermo�Morra�y�Silvia�Erdmann�por�permitirme� trabajar,�durante�varias�
temporadas�de�verano,�en�su�campo�en�Navarro�(Buenos�Aires),�y�por�ser�siempre�
tan�atentos�conmigo…�
�� A�Patricio�Fidalgo,�a�su�Sra.�Chongui�y�a�su�nieto�Nicolás,�por�su�permanente�
asistencia�en�el�trabajo�de�campo�en�Anillaco�(La�Rioja),�por�las�cenas�caseras�y�los�
“icecream”,� por� las� charlas� y� los�momentos� compartidos,� y� por� el� afecto� que�me�
brindaron�siempre…�
A�Enrique�Rodríguez�por�estar,�por�guiarme�en�los�análisis�estadísticos�y�por�
mantener�las�puertas�de�su�laboratorio�abiertas�para�mis�consultas…�
A� Federico� Ocampo� por� compartir� conmigo� sus� conocimientos� en�
Scarabaeidae,� por� guiarme� en� las� identificaciones,� por� estar� siempre� presente� y�
predispuesto�a�colaborar�con�mi�trabajo…��
A�Wilfredo�Fernández�y� Jorge�Frogoni�por�permitirme� trabajar� tan�cómoda�y�
acogedoramente�en�el�Museo�Lucas�Roselli�(Nueva�Palmira,�Uruguay)…�
A�David�Edmonds�y�Frank�Krell,�por�resolver�mis�dudas,�por�su�apoyo�y�por�la�
ayuda�que�me�ofrecieron�desde�el�primer�momento…��
�� A�Alejandra�Molina�por�compartir�conmigo�sus�observaciones�de�campo�sobre�
el�comportamiento�de�los�escarabajos�y�por�regalarme�una�de�las�experiencias�más�
lindas�de�mi�vida,�el�abrazo�de�un�oso�hormiguero�bebé!...�
A� Sergio� Gómez� y� a� Jimena� González� Naya� por� ser� mis� fuentes� de� agua�
destilada�(de�la�mejor�calidad!)�y�por�su�amistad…�
A� Carmen� Huerta� por� responder� generosamente� a� mis� consultas� sobre� los�
Aphodinae…�
A�Luciano�Patitucci�y�Pablo�Mulieri�por�la�determinación�de�larvas�y�pupas�de�
Dipteros�halladas�en�La�Rioja...�
A� Rosario� Romero� por� la� pintura� presentada� aquí� como� reconstrucción� del�
ecosistema�en�Gran�Barranca…��
A� Ei� (Eider� Ruiz� Manzanos)� y� Mai� (Margarita� Rocca)� por� los� increibles�
recuerdos�que�tengo�junto�a�ellas…�
A�Ariel,�Diego,�Agustín,�Fede�y�Martín,�vecinos�de�laboratorio�y�amigos,�por�las�
charlas�y�el�apoyo�mutuo�de�todos�los�días…�
Y� finalmente,� quiero� agradecer� a� toda� esa� gente� extraordinaria� que� forma�
parte�de�mi�vida:��
A�Sher,�mi�hermana�del�alma,�gracias�por�darme�confianza,�por�cuidarme,�por�
empujarme�a�hacer�aquéllas�cosas�a�las�que�le�temí�siempre,�por�darme�la�mano�y�
por�recorrer�juntas�todo�este�camino…te�quiero�y�te�extraño…�

� ��
A�Van,�amiga,�sos�increíble,�te�admiro�y�te�adoro,�y�me�alegra�enormemente�
haber�hecho�embriología…jajaja…ahí�te�encontré!...�
A� Gabi� (Titi),� cuantas� cosas� vivimos� en� la� Facu!!!� Y� cuantas� otras� por� la�
vida!!...siempre�estás�conmigo…�
A� Cande,� Pacipaz,� Pichu� y� Mariela� que� aunque� no� nos� veamos�
permanentemente,�están�siempre…�
A�Pol,�mi�gran�amigo,�gracias�por�disfrutar�mi�trabajo�conmigo�y�por�estar!�
Anita,�Seba,� Luli,�Amil,� Toto,�Noe,�Tavo�y� Juli,� amigos�gracias�por�aguantar�
mis�historias�de�campaña,�mis�fotos�y�videos�eternos,�mis�“platelmintos”,�y�mis�tiqui�
tiquis…sin�terminar�de�entender�todavía�para�qué�revuelvo�bosta!!!!�los�quiero!...��
A�Patricia�y�Carlos�por�todo�el�cariño�que�me�dieron�siempre…��
A�Juan,�Lili,�Jorgelina,�Anselmo�y�a�la�Abuela�Ñata…�
A�Pepe,�Santi�y�Gabi,�mis�hermanos,�y�a�Pao,�Gri,�Lidia�y�Celi,�mis�cuñadas…�
A�mis�sobris:�Fede,�Valen,�Gero,�Nacho,�Tomás,�Lauti,�Tomy,�Luli,�Cami,�Oti,�
Lucia�y�Camila…�
A�mi�tía�Estela,�a�Pauli,�Marce,�Gini�y�flia.�…�
A�mis�tíos,�Marta�y�Chin,�y�a�Mari…�
A�Sasha�y�Lostris…mis�soles�
A� Jolie�y�Mingo,�mis� “papuchos”,�por�amarme,�por�estar�siempre�a�mi� lado,�
por� apoyar� mi� carrera� y� por� creer� que� soy� una� “grosa”,� gracias� por� sentirse�
orgullosos�de�mi!...�
A� Pabli,�mi� amor,� por� todos� estos� años� recorriendo�caminos,� que� aunque� a�
veces�no�son�los�mismos,�siempre�lo�hacemos�juntos,�uno�al� lado�del�otro.�Gracias�
por� todas� las� horas� en� las� que� dormiste� solo,� por� esperarme,� por� cuidarme,� por�
apoyarme,�por�sentirte�orgulloso�de�mí�y�por�tu�amor�incondicional…�
�
�
�����000
�!1.

� �
������
2���� ��������� 23�
�
2�2� �� Icnoestratigrafía� de� alta� resolución� de������������� en� Gran� Barranca�
(Chubut,�Argentina),�Formación�Sarmiento�(Eoceno�medio�Mioceno�inferior)…..27�
�
2�4� ��Trazas�de�organismos�cleptoparásitos�y�detritívoros�en�bolas�de�cría�fósiles�
de�escarabajos�coprófagos�(�����������)……………………………………………………….30�
�
2�5� ��Análisis�del�contenido�fitolítico……………………………………………………………………32�
�
4����� ���� 56
�
4�2��Objetivos�generales………………………………………………………………………………………35�
�
4�4��Objetivos�particulares.......................................................................35
5��������������57
�
8��� ��������9 ����87�
�
8�2 �� Comportamiento� de� nidificación� de� distintas� especies� de� escarabajos�
coprófagos�de�la�Argentina……………………………………………………………………………………49�
�
8�2�2��Selección�de�las�especies�de�estudio………………………………………………….49�
�
8�2�4��Trabajo�de�campo……………………………………………………………………………….50�
8�2�4�2���Reconocimiento�de�bostas�con�actividad……………………………………53�
8�2�4�4��Excavación�de�nidos�y�extracción�de�masas/bolas�de�cría……….54�
8�2�4�5���Registro�de�la�arquitectura�de�los�nidos�y�del�comportamiento�de�
las�especies�estudiadas……………………………………………………………………………….55�
8�2�4�8��Colecta�de�escarabajos……………………………………………………………….56�

� ��
8�2�5��Trabajo�de�laboratorio…………………………………………………………………………56�
�
8�2�5�2��Material�colectado……………………………………………………………………….56�
8�2�5�4 �� Mediciones� y� observaciones� de� masas/bolas� de� cría� y�
descripciones�de�los�nidos…………………………………………………………………………..57�
8�2�5�5��Observación�del�desarrollo�larval�en�el�laboratorio……………………58�
8�2�5�8��Ensayos�de�nidificación�en�el�laboratorio,�o�en�el�campo�pero�en�
condiciones�experimentales�controladas……………………………………………………59�
�
8�4�� Icnotaxonomía�e� icnoestratigrafía�de�alta�resolución�de�������������en�
Gran� Barranca� (Chubut,� Argentina),� Formación� Sarmiento� (Eoceno� medio�
Mioceno�inferior)…………………………………………………………………………………………………….64�
�
8�5� �� Micromorfología� de� masas/bolas� de� cría,� cámaras� pupales,� fósiles� y�
actuales,�y�de�trazas�halladas�en�ejemplares�de�Coprinisphaera………………………..74�
�
8�8���Análisis�del�contenido�fitolítico:�selección�de�muestras,�extracción,�conteo�y�
clasificación…………………………………………………………………………………………………………….75�
�
8�8�2��Bolas�de�cría�actuales�y�suelo�circundante…………………………………………75�
�
8�8�4��Bolas�de�cría�fósiles�y�paleosuelo�circundante……………………………………76�
�
6������ ���:4�
�
6�2��Icnología�Sistemática……………………………………………………………………………………82�
�
6�2�2��Bolas�de�cría�fósiles�de�escarabajos�coprófagos…………………………………82�
6�2�4��Trazas�fósiles�de�organismos�asociados�a������������………………107��
6�4 �� Estudios� neoicnológicos:� comportamiento� de� nidificación� de� especies� de�
escarabajos�coprófagos�de�la�Argentina…………………………………………………………….116�
6�4�2�������������sp.�………………………………………………………………………………117�

� �
�
6�4�2�2��Estructura�de�la�masa�de�cría………………………………………………….117�
6�4�2�4���Micromorfología�de�la�masa�de�cría………………………………………..118�
6�4�2�5���Estructura�del�nido…………………………………………………………………..119�
�
6�4�4����������� �������Brullé………………………………………………………………120�
�
6�4�4�2���Estructura�de�la�masa�de�cría………………………………………………….120�
6�4�4�4��Micromorfología�de�la�masa�de�cría…………………………………………123�
6�4�4�5���Estructura�del�nido……………………………………………………………………124�
�
6�4�5#���� ������������(Laporte)……………………………………………………126�
�
6�4�5�2��Estructura�de�la�bola�de�cría…………………………………………………….126��
6�4�5�4���Micromorfología�de�la�bola�de�cría……………………………………………130�
6�4�5�5���Estructura�del�nido……………………………………………………………………133�
6�4�8������ ���������������(Chevrolat)………………………………………………134�
�
6�4�8�2���Estructura�de�la�bola�de�cría…………………………………………………….134�
6�4�8�4���Micromorfología�de�la�bola�de�cría……………………………………………139�
6�4�8�5��Estructura�del�nido……………………………………………………………………144�
�
6�4�6����� ������������(Harold)…………………………………………………………146�
�
6�4�6�2���Estructura�de�la�bola�de�cría…………………………………………………….146�
6�4�6�4��Micromorfología�de�la�bola�de�cría…………………………………………..148�
6�4�6�5��Estructura�del�nido……………………………………………………………………150�
6�4�3�������������� ����(Fabricius)……………………………………………………………151
�
6�4�3�2��Estructura�de�la�bola�de�cría…………………………………………………….151�
6�4�3�4��Micromorfología�de�la�bola�de�cría……………………………………………154�
6�4�3�5���Estructura�del�nido……………………………………………………………………157��

� ��
6�4�;– ��������! ���������"���� �� ������(Blanchard)……………….158
�
6�4�;�2��Estructura�de�la�bola�de�cría…………………………………………………….158�
6�4�;�4���Micromorfología�de�la�bola�de�cría……………………………………………161�
6�4�;�5���Estructura�del�nido……………………………………………………………………163�
6�4�:#� ��������! �������"��������(Gillet)…………………………………164��
�
6�4�:�2���Estructura�de�la�bola�de�cría…………………………………………………….164�
6�4�:�4��Micromorfología�de�la�bola�de�cría……………………………………………167�
6�4�:�5���Estructura�del�nido………….……………………………………………………….169�
6�4�7���������sp.1…….……………………………………………………………………………….171
�
6�4�7�2���Estructura�de�la�bola�de�cría…………….…………………………………....171�
6�4�7�4���Micromorfología�de�la�bola�de�cría……………………………………………173�
6�4�7�5��Estructura�del�nido……………………………………………………………………174�
6�4�2<����������sp.�2………………………………………………………………………………….174
�
6�4�2<�2���Estructura�de�la�bola�de�cría………………………………………………….174�
6�4�2<�4���Micromorfología�de�la�bola�de�cría……….……………………………….176�
6�4�2<�5���Estructura�del�nido…………………………………………………………………178�
6�4�22–�$������������������(Burmeister)……..%%%�…….……………………….178�
�
6�4�22�2���Estructura�de�la�cámara�pupal……………..……………………………….178�
6�4�22�4���Micromorfología�de�la�cámara�pupal………………..…………………..180�
6�4�22�5���Estructura�del�nido………………………………………………….……………..181�
6�4�24–�$���������������� �&��(Blanchard)……%�…….………………………..182�
�
6�4�24�2���Estructura�del�nido………………………………………………….………….….182�
6�4�25–�'� ������ & ������Burmeister………%%%%�…….…………..………….185�

� ��
�
6�4�25�2���Estructura�del�nido…………………………………………………………..…….185�
6�5 �� Icnoestratigrafía� de� alta� resolución� de� ������������ en� Gran� Barranca�
(Chubut,� Argentina),� Formación� Sarmiento� (Eoceno� medio�Mioceno�
inferior)…………………………………………………………………………………………………………….….185
�
6�5�2� �� Aparición,� densidad,� abundancia� relativa� y� desaparición� de�
������������en�la�secuencia…………………………………………………………………….186�
�
6�5�4� �� Análisis� del� tamaño� de� los� ejemplares� de� ������������ (DE:�
Diámetro�Ecuatorial)………….………………………………….………………………………………191�
6�5�5 �� Análisis� de� las� icnoespecies� agrupadas� por� clases� de�
tamaño...………………………………………………………………………….…………………………….197�
�
6�5�8 �� Análisis� del� Índice� de� Espesor� de� la� Pared� (IEP)� de�
�����������……………………………………………………………………………………………….199
�
6�8��Análisis�del�contenido�fitolítico…………………………………………………………………202
6�8�2��Bolas�de�cría�actuales�y�suelo�circundante…..………..……………………….202�
6�8�4��Bolas�de�cría�fósiles�y�paleosuelo�circundante…..…………………………….202�
�
6�8�4�2 �� Comparación� del� número� total� de� fitolitos� entre� el� relleno� de�
������������y�el�paleosuelo�circundante…………………..……………………...202�
6�8�4�4 �� Comparación� del� número� total� de� fitolitos� entre� la� pared� de�
������������y�el�paleosuelo�circundante�(o�el�relleno)…………...........206�
6�8�4�5��Comparación�del�número�total�de�fitolitos�entre�las�dos�partes�de�
la�pared�(interna�y�externa)�de�Coprinisphaera………………………………………207�
6�8�4�8���Comparación�de�las�frecuencias�de�morfotipos�de�fitolitos�entre�
la� pared� y� el� relleno� de� �����������,� y� el� paleosuelo�
circundante…………………………………………………………………………………………….….208�
�

� ��
3����������425�
3�2 �� Icnotaxonomía�de������������� y� cambios� en� la� posición�de� la� cámara�
ovígera………………………………………………………………………………………………………………….213�
3�4� �� Estudios� neoicnológicos:� comportamiento� de� nidificación� de� especies� de�
escarabajos�coprófagos�de�Argentina……………………..………………………………………….217�
3�4�2�������������sp.�…….………………………......………………….…………………….218�
�
3�4�4����������� ������……………......................................………………219�
�
3�4�5����� �������spp.�….............................................................223�
�
3�4�8������������� ���...................................................................227
3�4�6�� ��������spp.�……………………………………………………………………………..231�
�
3�4�3����������spp.�………………………………………………………..…………………….……234�
3�4�;��$������������spp.�y�'� ������sp.�……………………….……..…………...236�
3�5#�Posible�atribución�de�las�icnoespecies�de�������������a�taxones�actuales�
y�la�comparación�con�el�registro�fósil�de�escarabajos……………………………….………238
3�8 �� Icnoestratigrafía� de� alta� resolucion� de������������� en� Gran� Barranca�
(Chubut,� Argentina),� Formación� Sarmiento� (Eoceno� medio�Mioceno�
inferior)………………………………………………………………………………………………………………..241
3�8�2 �� Aparición,� densidad,� abundancia� relativa� y� desaparición� de�
������������en�Gran�Barranca�(Formación�Sarmiento)……………….…….……241�
3�8�4 �� Cambios� registrados� en� las� icnoespecies:� diversidad� y� abundancia�
relativa……………………….……………………………………………………………………………………248�
�

� ��
3�8�5 #El� incremento�de� tamaño�de� los�ejemplares�de������������� (DE:�
Diámetro� Ecuatorial)� y� su� relación� con� el� tamaño� de� los�
mamíferos………………………….………………….……………………………………………………….249�
�
3�8�8 �� El� tamaño� de� las� icnoespecies� de� �����������:� análisis� de� la�
diversidad�de�escarabajos�coprófagos�productores……………….…………………….252�
���
3�8�6 �� Cambios� del� espesor� de� la� pared� de� ������������ (IEP):� un�
indicador�paleoambiental?...................................................................254�
3�6��Trazas�de�organismos�cleptoparásitos�y�detritívoros�en�bolas�de�cría�fósiles�
de�escarabajos�coprófagos�(�����������)……………………………………………………..256�
�
3�3�Análisis�del�contenido�fitolítico…..….………………………………………………………….268
�
3�3�2��El�relleno�de�������������y�el�paleosuelo�circundante�muestran�un�
número�similar�de�fitolitos………………………………………………………………………….….268
3�3�4��La�pared�de�������������concentra�mayor�número�de�fitolitos�que�
el�relleno�y�el�paleosuelo�circundante…………………………………………….….………..271�
3�3�5 �� El� número� de� fitolitos� de� la� parte� interna� de� la� pared� de�
������������es�similar�a�la�de�la�parte�externa……………………………………….274�
�
3�3�8��Inferencia�de�la�dieta�de�los�herbívoros�productores�de�bosta�a�partir�
de�las�diferencias�halladas�en� las�frecuencias�de�morfotipos�de�fitolitos�entre�
la� pared� y� el� paleosuelo� circundante� (o� el�
relleno)……………………………………………………………………………………………………………274�
�
;�������������4;;�
�
:�����������4:8�
�
�
�

� ��
2��� ���������
�
Las� bolas� de� cría� fósiles� de� escarabajos� coprófagos,� incluidas� en� el�
icnogénero� ������������ Sauer,� han� sido� registradas� en� paleosuelos� y�
conglomerados� de� diferentes� edades� y� en� distintos� continentes� (Genise,� 2004;�
Laza,� 2006� y� referencias� alli).� En� el� Hemisferio� Norte,� Asia� y� Europa,� se� han�
hallado�pocos�registros,�tales�como�los�del�Mioceno�de�Pakistan�(Retallack,�1991)�
y�del�Eoceno�de�Italia�(Dieni�y�Genise,�2004).�Basilewsky�(1951)�mencionó�estas�
trazas� del� Pleistoceno� de� Burundi,� en� África,� mientras� que� el� resto� de� los�
hallazgos� están� relacionados� con� afloramientos� del� Plioceno� en� Sudáfrica�
(Kitching,�1980),�Tanzania�(Sands,�1987)�y�Tchad�(Duringer����.,�2000a,�b).�Es�
probable� que� el� escaso� registro� de������������� en� estas� regiones,� se� deba�
fundamentalmente�a�las�pocas�investigaciones�realizadas,�en�algunos�casos,�o�a�
la� ausencia� real� de� los� escarabajos� productores� de�estas� trazas,� en� otros.� Los�
depósitos� de� América� del� Sur,� en� cambio,� son� los�más� ricos� en� icnofaunas� de�
escarabajos� coprófagos� y� se� han� registrado� ������������ en� diferentes�
formaciones� de� Uruguay,� Ecuador� y� Argentina,� desde� el� Eoceno� hasta� el�
Cuaternario.�En�Uruguay,�Roselli�las�describió�y�nominó�por�primera�vez�en�1939,�
como� (��� ���,� un� nombre� preocupado� y� que� de� todos� modos� pasó�
desapercibido� en� la� literatura,� y� posteriormente� describió� otros� tipos� en� dos�
trabajos� más� (1976,� 1987).� Siguiendo� la� revisión� histórica� hecha� por� Laza�
(2006),�en�Ecuador,�Estrada,�Bruet�y�Sauer�publicaron�diferentes�contribuciones�
sobre� estas� trazas.� Estrada� (1941)� las� registró� por� primera� vez� para� el�
Pleistoceno� de� Ecuador,� y� las� atribuyó� a� escarabajos� coprófagos.� Luego,� Bruet�
(1950)�confirmó�esta�atribución�y�Sauer�(1950,�1955,�1956,�1959,�1965)�aportó�
el�marco�geológico�y�postuló�su�origen,�creando�el�icnogénero�������������en�

� ��
1955.�En�Argentina,�Frenguelli�(1938a,�b;�1939a,�b;�1940;�1941),�Fossa�Mancini�
(1941)� y� Andreis� (1972),� registraron� estas� trazas� en� formaciones� de� distintas�
edades�y�diferentes�regiones.�En�Argentina,�existen�además�algunas�menciones�
previas�pero�vagamente�documentadas,�particularmente�aquéllas�de�Patagonia,�
lugar�donde�estas�trazas�son�muy�comunes.�Como�menciona�Laza�en�su�trabajo�
de�������������del�2006,�entre�estos�registros,�se�destacan�las�descripciones�
hechas� por� C.� Ameghino� en� 1897� en� una� carta� a� su� hermano� Florentino;� las�
menciones�hechas�por�McCartney� (1933),�geólogo�adjunto�de� las�campañas�de�
G.�G.�Simpson,�donde�las�interpretó�como�frutos�en�la�descripción�de�los�perfiles,�
mientras�que�el�paleontólogo�las�describió�como�clastos�de�origen�desconocido�en�
sus� anotaciones� de� campo� (Laza,� 2006).� Luego� de� un� paréntesis� desde� las�
menciones� y� publicaciones� originales,� en� los� últimos� años� se� retomó� la�
investigación� y� se� publicaron� desde� breves� resúmenes� en� congresos,� hasta�
publicaciones�en�revistas�periódicas�de�gran�importancia,�asi�como�la�creación�de�
la� icnofacies� de� ������������ (Genise� �� ��,� 2000)� y� de� la� icnofamilia�
��������������(Genise,�2004).��
A� pesar� de� todos� estos� descubrimientos,�menciones� y� registros� previos,�
recién� en� el� 2006,� Laza� publicó� la� primera� revisión� icnotaxonómica� del�
icnogénero�utilizando�criterios�claros�y�abarcativos,�como�resultado�de�más�de�30�
años� de� trabajo� de� investigación,� tanto� en� el� campo,� en� colecciones� y� con� los�
ejemplares,�como�en�el�conocimiento�de�la�evolución,�biología�y�comportamiento�
de�los�escarabajos�coprófagos�actuales.�No�obstante�lo�reciente�de�esta�revisión,�
la� investigación� realizada� para� esta� Tesis� doctoral,� basada� en� la� morfología�
(macro� y�micro)� de� las� trazas� fósiles� y� en� nuevos� conocimientos� adquiridos� a�
partir�del�trabajo�de�campo�y�de�laboratorio�de�las�bolas�de�cría�actuales�arrojó�
como� resultado� algunas� novedades� icnotaxonómicas� y� nomenclatoriales� de�

� ��
importancia,� asi� como� un� panorama� nuevo� sobre� la� atribución� de� estas� trazas�
fósiles�a�coprófagos�actuales.�
La� familia� Scarabaeidae� se� encuentra� conformada� por� escarabajos�
pertenecientes� a� las� siguientes� subfamilias:� Scarabaeinae,� Geotrupinae� y�
Aphodiinae.� A� pesar� que� todos� ellos� se� alimentan� de� detritos� orgánicos,� tanto�
adultos�como�larvas,�se�los�conoce�comúnmente�como�“escarabajos�coprófagos�o�
estercoleros”� (dung� beetles).� Sin� embargo,� sólo� en� los� Scarabaeinae,� los�
escarabajos� de� la� mayoría� de� las� especies� tienen� como� hábito� alimentario� la�
coprofagia,�usando� fundamentalmente�como�fuente�de�alimento,�el�excremento�
producido�por�los�mamíferos.�En�los�Geotrupinae,�sólo�los�escarabajos�de�la�tribu�
Geotrupini� son� coprófagos,� mientras� que� la� mayoría� de� las� especies� de�
Aphodiinae�son�saprófagos�(Halffter�y�Edmonds,�1982).�A�lo�largo�de�esta�Tesis,�
los�términos�“escarabajos�coprófagos”�y�“escarabajos�estercoleros”�serán�usados�
para�hacer�referencia,�especialmente,�a�los�Scarabaeinae.��
Las�adaptaciones�más�destacadas�de�esta�subfamilia�son:�comportamiento�
de� reubicación� del� alimento;� mayor� desarrollo� de� caracteres� músculo�
esqueletales� que� intensifican� su� capacidad� fosorial;� fortalecimiento� y� uso� de�
distintas� estructuras� del� cuerpo� como� herramientas� para� la� movilización� de�
material�del�suelo�y�la�excavación;�comportamiento�de�nidificación�subterráneo;�
reducción�del�comportamiento�de�cortejo;�frecuente�cooperación�macho�hembra�
durante� la� nidificación;� extrema� reducción� de� la� fecundidad� (reducción� de� las�
gónadas� de� la� hembra� a� una� sola� ovariola);� adaptación� de� larvas� y� pupas� al�
desarrollo� confinado� a� un� espacio� cerrado;� piezas� bucales�modificadas� para� la�
manipulación� de� un� alimento�más� suave,� altamente� dúctil� y� semilíquido;� y� un�
tracto� digestivo� en� los� adultos� modificado� para� procesar� rápidamente� grandes�
cantidades�de�alimento�(Halffter�y�Edmonds,�1982).��������

� �
Los� escarabajos� coprófagos� son� mundialmente� conocidos� debido� a� su�
complejo�comportamiento�de�nidificación,�y�han�sido�usados�como�herramienta�
de� estudio� durante� décadas� debido� a� su� importante� rol� en� el� reciclado� del�
excremento�de�vertebrados�y�de�su�incorporación�al�suelo�(Fincher�����,�1970;�
Waterhouse,� 1974;� Hughes,� 1975;� Halffter� y� Edmonds,� 1982).� Se� cree� que� el�
comportamiento�de�nidificación�ha�surgido�en� la�evolución�de�estos�organismos�
como�consecuencia�de�otro�comportamiento,�el�de�reubicación�del�alimento�por�
medio� del� adulto,� y� que� ambos� han� sido� además� influidos� por� las� mismas�
limitaciones� ecológicas� (Halffter� y� Edmonds,� 1982).� El� proceso� de� nidificación�
puede� resumirse� de� la� siguiente�manera:� búsqueda� de� la� fuente� de� alimento;�
construcción�de�una�bola�de�excremento�para�ser�transportada�o�simplemente�el�
enterramiento� de� una� porción� de� bosta;� y� la� construcción� subterránea� de�
túneles,� cámaras� y� de� las� masas/bolas� de� cría� dentro� de� las� cuales� son�
depositados�los�huevos.�El�paso�inicial�de�la�nidificación,�es�decir�la�búsqueda�de�
alimento,�suele�ser�através�del�vuelo,�durante�el�cual�pueden�“oler”�la�fuente�de�
origen.�Luego�del�vuelo�de� reconocimiento,�y�una�vez�hecho�el� contacto�con�el�
alimento,�el�adulto�rápidamente�reubica�una�porción�de�excremento�y�la�entierra,�
asegurándose�con�ello�el�menor�tiempo�posible�de�exposición�en�la�superficie,�y�
en� consecuencia,� disminuyendo� la� competencia� por� el� alimento,� y� a� su� vez,� la�
predación.�Asimismo,�el�excremento�producido�por�los�herbívoros�sufre�un�rápido�
proceso�de�deshidratación�en� la�superficie�del�suelo,� lo�que� lo�convierte�en�una�
fuente�menos�atractiva�para�estos�escarabajos.�Por�lo�tanto,�al�ser�enterrada�en�
condiciones� microclimáticas� que� favorecen� su� vida� útil,� las� propiedades� de� la�
bosta�se�conservan�por�más�tiempo,�en�particular�su�grado�de�hidratación,�y�por�
otro�lado,�se�disminuye�así�su�disponibilidad�para�otros�coprófagos�competidores�
(Halffter� y� Edmonds,� 1982).� Esta� es� otra� ventaja� adaptativa� que� resulta� del�

� ���
comportamiento� de� reubicación� del� alimento,� en� comparación� con� una�
alimentación� ��� ����.� En� relación� con� este� comportamiento� y� basándose� en� la�
ubicación�de�la�fuente�de�alimento�(bosta)�y� la�distancia�a�la�cual� los�nidos�son�
excavados,� Bornemissza� (1969,� 1976)� creó� una� clasificación� simple� del�
comportamiento�de�nidificación�de� los�Scarabaeinae,� reconociendo�tres�posibles�
tipos�de�nidos:�1)�endocópridos:�aquéllos�nidos�creados�dentro�de� la� fuente�de�
alimento� (excremento),�o�en�una�cavidad�poco�profunda� justo�por�debajo�y�en�
contacto�con�ella;�2)�paracópridos:�nidos�excavados�en�el�suelo�por�debajo�del�
excremento�que�es�usado�como�fuente�de�alimento�y�al�que�se�comunican�por�un�
sistema� de� galerías;� y� finalmente,� 3)� telecópridos:� nidos� excavados� a� una�
determinada� distancia� de� la� fuente� de� alimento� de� la� cual� se� ha� extraído� una�
porción� para� ser� transportada� al� nido� por� medio� del� rolado� de� una� bola� de�
excremento,�o�empujando�un�fragmento.�Halffter�y�Matthews�(1966)�fueron� los�
primeros�en�intentar�clasificar� los�diferentes�comportamientos�de�nidificación�de�
los� Scarabaeinae,� basándose� fundamentalmente� sobre� la� secuencia� de� pasos�
comportamentales�que�deben�ser�alcanzados�por�los�escarabajos�para�completar�
un� nido.� Distinguieron� así� cuatro� patrones� o� grupos� de� comportamiento� de�
nidificación,�de�los�cuales�los�tres�primeros�incluyen�excavadores�(paracópridos),�
y� sólo� el� Grupo� IV� incluye� a� los� roladores� (telecópridos).� En� 1977,� Halffter�
propone� dos� grupos� adicionales� (Grupo� V� y� VI)� para� reacomodar� una�
modificación� hecha� en� el� Grupo� IV,� y� para� incluir� un� comportamiento� nuevo.�
Posteriormente,� Halffter� y� Edmonds� (1982)� redefinen� y� extienden� las�
clasificaciones�de�Halffter�y�Matthews�(1966),�y�de�Halffter�(1977),�usando�para�
ello� más� caracteres,� tales� como� la� estructura� de� la� cámara� de� cría� y� la�
arquitectura� de� los� nidos,� y� conservando� el� criterio� de� Bornemissza� sobre� la�
ubicación� de� los� nidos,� como� un� carácter� de� identificación� de� los� tipos� de�

� ��
nidificación.� En� esta� clasificación,� actualmente� en� uso,� se� reconocen� siete�
patrones�de�comportamiento�de�nidificación�y�se�propuso�un�esquema�general�de�
su�evolución.�Los�caracteres�usados�como�diagnósticos�para�la�determinación�de�
dichos�patrones�y� los�cuales�fueron�considerados�en�los�estudios�neoicnólogicos�
presentados� aquí,� se� detallan� a� continuación� de� manera� resumida:� tipo� de�
provisión�de�la� larva�(bola�o�masa�de�cría);�posición�del�nido�en�relación�con�la�
superficie� (superficial� o� subterráneo)� y� en� relación� con� la� fuente� de� alimento�
(criterio� de� Bornemissza:� endocóprido,� paracóprido,� telecóprido);� grado� de�
complejidad� del� nido,� superficial� o� subterráneo� (cantidad� de� bolas� de� cría� por�
cámara,� cantidad�de�masas/bolas�de� cría�por� túnel,�cantidad�de� túneles,� etc.);�
posición�de�las�masas/bolas�de�cría�en�nidos�subterráneos�y�compuestos�(lineal,�
en�racimo,�agrupada);�manipulación�de�la�provisión�de�la�larva�(trabajada�o�no,�
es�decir,�separando�o�no�los�grandes�fragmentos�de�hojas�o�pastos�incluidos�en�
la�bosta�de�los�herbívoros�antes�de�ser�usada�para�la�fabricación�de�la�masa/bola�
de�cría);� tipo�de�aprovisionamiento�de�nidos�subterráneos�(directo,� la�provisión�
es�usada�rápidamente�para�la�construcción�de�masas/bolas�de�cría;�o�retardado,�
cuando� la�bosta�es�aprovisionada�en�una� cámara�o� túnel� antes�de� ser�usada);�
superficie� externa� de� la� bola� de� cría� (con� o� sin� capa� de� material� del� suelo�
agregada� sobre� la� provisión� por� los� progenitores);� ubicación� de� la� cámara�
ovígera�en�relación�con�la�provisión�(en�contacto�con�la�provisión,�o�separada�por�
medio�de�un�recubrimiento�con�material�del�suelo,�o�contenida�en�una�esfera�de�
provisión� altamente� compactada)� y� en� relación� con� el� túnel� de� acceso� a� la�
cámara� nido� (distal� cuando� la� cámara� ovígera� es� creada� durante� los� primeros�
pasos�de�la�construcción�de�la�bola�de�cría,�o�proximal�cuando�es�creada�durante�
los� últimos� estadios);� cooperación�macho�hembra� (“ausente”� cuando� el�macho�
no� participa,� “escaso”� cuando� el� macho� colabora� alcanzándole� provisión� a� la�

� ���
hembra�mientras�excava,�o�durante�el�rolado,�y�“considerable”�cuando�el�macho�
participa� en� el� aprovisionamiento� o� rolado,� y� en� las� otras� etapas� de� la�
nidificación);�y�cuidado�parental�(“ausente”�cuando�la�hembra�abandona�el�nido�
después�de�crear�la�última�masa/bola�de�cría,�o�“presente”�cuando�la�hembra�se�
queda�en�el�nido�una�vez�creadas�todas�las�bolas�de�cría�y�se�mantiene�allí�hasta�
su�muerte�o�hasta�la�emergencia�de�la�progenie).��
Teniendo�en�cuenta�los�caracteres�previamente�mencionados,�los�patrones�
de�nidificación�extraídos�de�Halffter�y�Edmonds�(1982),�pueden�resumirse�de�la�
siguiente�manera:�
�&=�> �,� desarrollo� larval� en� una� masa� de� cría;� nidos� subterráneos� y�
paracópridos,�simples�o�compuestos�(masas�de�cría�con�distribución� lineal�o�en�
racimos);�aprovisionamiento�directo;�cámara�ovígera�proximal�y�en�contacto�con�
la� provisión;� cooperación� macho�hembra� escasa� o� ausente;� cuidado� parental�
ausente.�Algunos�géneros�incluidos�son:�����������,��������,�(� ��������.�
�&=�> ��,�desarrollo�larval�en�una�bola�de�cría�de�material�de�la�provisión�
trabajado� y� recubierta� por� una� capa� compacta� de� material� del� suelo;� nidos�
subterráneos� y� paracópridos� (raramente� telecópridos),� simples� o� compuestos�
(bolas� de� cría� en� cámaras� separadas,� con� distribución� lineal� o� en� racimos);�
aprovisionamiento�retardado,�el�alimento�de�la� larva�es�previamente�acumulado�
en� un� lugar� del� túnel;� cámara� ovígera� proximal� aislada� por� recubrimiento� con�
material� del� suelo,� o� en� una� esfera� de� provisión� especialmente� fabricada;�
cooperación� macho�hembra� considerable;� cuidado� parental� ausente.� Algunos�
géneros� incluidos� son:� )����,� ��� ������,� *������� (*�� ��������),�
(� ��������,���������(����+� ���).��
�&=�> ���,� desarrollo� larval� en� una� bola� de� cría� de� material� de� la�
provisión� trabajado� y� recubierta� generalmente� por� una� capa� delgada,� poco�

� ���
compactada,� de� material� del� suelo� depositada� por� la� hembra� luego� de� la�
oviposición;�nidos�subterráneos�y�paracópridos,�compuestos;�aprovisionamiento�
retardado,�el�alimento�de�la�larva�es�previamente�acumulado�en�la�cámara�nido�
como� una�masa� compacta� denominada� “torta”;� cámara� ovígera� proximal� y� en�
contacto� con� la� provisión;� cooperación� macho�hembra� considerable;� cuidado�
parental� presente.� Algunos� géneros� incluidos� son:� ������,� ,��� �����,�
�����������.�����
�&=�> ��, desarrollo� larval� en� una� bola� de� cría� de� material� de� la�
provisión,� recubierta� o� no� por� material� del� suelo;� nidos� subterráneos� y�
telecópridos,� simples� (las� bolas� de� cría� son�moldeadas� en� la� bosta,� roladas� y�
enterradas�a�determinada�distancia�de�la�fuente�de�alimento);�aprovisionamiento�
directo;�cámara�ovígera�proximal�(superior),�en�contacto�o�no�con� la�provisión;�
cooperación� macho�hembra� considerable;� cuidado� parental� ausente.� A� este�
patrón�pertenecen�la�mayoría�de�las�especies�roladoras.��
�&=�> �,�desarrollo�larval�en�una�bola�de�cría�de�material�de�la�provisión,�
recubierta�por�material�del�suelo;�nidos�superficiales�y�telecópridos,�compuestos�
(las�bolas�de�cría�son�modeladas�en�la�bosta�por�la�hembra�y�el�macho,�roladas�y�
sucesivamente�agrupadas�en�una�cavidad�poco�profunda�a�determinada�distancia�
de�la�fuente�de�alimento);�aprovisionamiento�directo;�cámara�ovígera�superior�y�
aislada;�cooperación�macho�hembra�considerable;�cuidado�parental�presente.�El�
ejemplo�más�conocido�es�el�de�������� &������ &�����.����
�&=�> ��,� desarrollo� larval� en� una� bola� de� cría� de� material� de� la�
provisión,� � recubierta� por� material� del� suelo� antes� de� ser� fabricada;� nidos�
superficiales� y� endocópridos,� compuestos� (bolas� de� cría� agrupadas� en� una�
cavidad);�aprovisionamiento�directo;�cámara�ovígera�superior�y�en�contacto�con�
la� provisión;� cooperación� macho�hembra� escasa� o� ausente;� cuidado� parental�

� ���
presente.�Es�el�patrón�de�nidificación�más�raro,�conocido�solamente�para�varias�
especies�de�'��&������.���
�&=�> ���,� desarrollo� larval� en� una� bola� de� cría� de� material� de� la�
provisión,� � recubierta� o� no� por� material� del� suelo;� nidos� superficiales� y�
endocópridos,� compuestos� (bolas� de� cría� agrupadas� en� una� concavidad� a� un�
costado�de� la� bosta);� aprovisionamiento�directo;� cámara�ovígera� superior� y� en�
contacto�con�la�provisión;�cooperación�macho�hembra�ausente;�cuidado�parental�
presente� o� ausente.� Este� patrón� de� nidificación� es� conocido� solamente� para�
algunas�especies�de������ �����y������ ��.�
En�relación�a�las�especies�de�Scarabaeinae�estudiadas�para�esta�Tesis,�los�
antecedentes� son� escasos.� Judulien� (1899),� Fabre� (1899),� Richter� (1918),�
Halffter� y� Matthews� (1966),� Halffter� y� Edmonds� (1982),� Cabrera�Walsh� y�
Gandolfo� (1996),� Morelli� �� ��� (1996)� y� Barbero� �� ��� (1999)� registraron�
diferentes� aspectos� de� su� comportamiento.� Sin� embargo,� estas� contribuciones�
carecen�de�descripciones�completas�y�detalladas�de�las�masas/bolas�de�cría�y/o�
de� la� arquitectura� de� los� nidos,� e� incluso� algunas� de� ellas� se� contradicen.� Por�
ello,�se�resúmen�a�continuación�los�aspectos�signifcativos�de�estos�trabajos,�los�
cuales� serán� discutidos� posteriormente� en� combinación� con� los� resultados�
obtenidos�de�las�observaciones�de�campo�y�de�laboratorio.�
Respecto� al� género� �������,� Judulien� (en� Fabre,� 1899)� registró� por�
primera�vez�los�nidos�de���������� �������Brullé,� los�cuales�describió�como�
ramificados� y� conformados�por� una�masa�de� cría� cilíndrica� en� cada�una�de� las�
ramas.� Halffter� y� Matthews� (1966),� en� cambio,� los� describieron� como� nidos�
simples�y� los� incluyeron�en�el�patrón�I�de�nidificación.�La�contribución�posterior�
de� Cabrera�Walsh� y� Gandolfo� (1996)� para� esta� especie,� incluyó� descripciones�

� ���
detalladas� de� la� estructura� de� las� masas� de� cría,� las� cuales� fueron� además�
obtenidas�en�condiciones�de�laboratorio�controladas.�
Halffter�y�Edmonds�(1982)�describieron�por�primera�vez�las�bolas�de�cría�y�
el� comportamiento� de� nidificación� de� una� especie� de��������,���� �+� ����
Harold.�En�dicho�trabajo,�incluyeron�a�esta�especie�en�el�patrón�II�de�nidificaciòn�
junto�con� la�mayoría�de� los�)����,�aclarando�que� las�diferencias�entre�ellos�
se�debían�fundamentalmente�a�la�forma�de�la�bola�de�cría�y�a�su�posición�dentro�
de� la� cámara� nido.� En� relación� con� este� género,� Cabrera�Walsh� y� Gandolfo�
(1996)�describieron�las�bolas�de�cría�de�una�especie�sudamericana�de��������,�
��� ��� ���� (Fabricius),� y�mencionaron�allí� que� los� escarabajos� de�esta� especie�
construyen� sus� bolas� de� cría� de� la�misma�manera� que� lo� hacen� los� Phanaeina�
incluidos� en� el� patrón� II� de� nidificación.� En� dicho� tabajo,� señalaron� además�
diferencias�significativas�entre�las�bolas�de�cria�construidas�por������� ����y����
�+� ���,�sin�embargo,�no�hicieron�ninguna�mención�sobre� la�arquitectura�de�
los�nidos.�
Dentro�del�género�����������,�se�han�registrado�hasta�el�momento� los�
nidos� de� un� gran� número� de� especies,� todos� ellos� incluidos� en� el� patrón� I� de�
nidificación,� y� los� cuales� pueden� ser� diferenciados� en� simples� o� compuestos�
(Halffter� y� Edmonds,� 1982).� En� el� primer� tipo,� los� nidos� se� encuentran�
conformados�por�un�túnel�con�una�única�masa�de�cría�cilíndrica�con�su�cámara�
ovígera�construida�en�el�polo�superior.�Se�incluyen�en�este�tipo,�los�nidos�de����
��� ����� (Halffter� y� Edmonds,� 1982,� Cabrera�Walsh� y� Gandolfo,� 1996).� En� el�
segundo� tipo,� en� cambio,� los� nidos� son� ramificados�y� poseen� varias�masas� de�
cría�por�cada�rama,�las�cuales�además�son�bien�ovaladas�y�más�elaboradas�que�
las� anteriores.� Los� nidos� de���� ����� y���� ����� son� ejemplos� de� este� tipo�
(Halffter�y�Edmonds,�1982).�

� ���
Judulien� (1899)� fue� también� el� primero� en� estudiar� los� nidos� de�
��� ������������(Laporte),�y�describió�las�bolas�de�cría�como�esferas�con�
una� calota� semiesférica� en� uno� de� sus� polos,� donde� la� hembra� construye� una�
cámara� ovígera� cónica.� Cabrera�Walsh� y� Gandolfo� (1996),� estudiaron� y�
describieron� las� bolas� de� cria� de� esta� especie,� más� las� de� ��� �������
��������(Chevrolat)�y���� ������������(Harold),�obtenidas�en�condiciones�
de�laboratorio�controladas.�Según�sus�observaciones,�las�tres�especies�poseen�un�
comportamiento� de� nidificación� similar� (patrón� II)� y� construyen� bolas� de� cría�
esféricas� que� recubren� externamente� con� un� “cemento”,� luego� de� excavar� la�
cámara�ovígera�en�uno�de�sus�polos.�Luego�de�poner�el�huevo�allí,�la�hembra�usa�
este�mismo�material�para�terminar�la�cámara,�formando�un�techo�perforado�que�
recubre� con� una� mezcla� de� este� cemento� y� fibras� vegetales� secas.� En� 1996,�
Morelli������describen�de�manera�detallada,�y�contribuyendo�con�nuevos�datos,�
el�desarrollo,�el�comportamineto�de�nidificación�y�las�bolas�de�cría�de��������-�
donde�se�destaca�el�resultado�del�comportamiento�de�la�larva�en�la�construccion�
de�la�cámara�pupal.��
En� relación� con� �������� sp.,� Barbero� �� ��� (1999),� estudiaron� en�
condiciones� de� laboratorio,� el� comportamiento� de� nidificación� de� ��������
������ �Balthasar,�1939.�De�sus�observaciones�se�destaca�la�presencia�de�una�
única� bola� de� cría,� piriforme� y� recubierta� por� una� capa� de�material� del� suelo,�
alojada�en�una�cámara�nido�donde�es�cuidada�por�la�hembra�hasta�que�la�larva�
alcanza�su�tercer�estadio.��� �
Respecto�al�género�������,�se�conoce�desde�tiempos�de�Judulien�(1899),�
y� hasta� la� actualidad,� el� comportamiento� de� nidificación� de� varias� de� sus�
especies,� tanto� necrófagas� como� coprófagas.� Sin� embargo,� el� caso� mejor�
estudiado�es�el�de���� &������ &������LeConte�(Halffter,�1977;�Halffter�����,�

� ���
1983;� Favila� y� Díaz,� 1996;� Favila,� 2001;� etc.),� del� cual� puede� destacarse� la�
forma� de� sus� bolas� de� cría,� típicamente� biesférica,� donde� la� cámara� ovígera�
ubicada� en� la� esfera� más� pequeña,� se� mantiene� aislada� de� la� provisión,� y� el�
comportamiento�de�la�larva�que�se�desarrolla�en�su�interior,�que�suele�expulsar�
hacia� la� pared,� pequeños� ������ fecales� esféricos� que� decoran� la� superficie�
externa�de�la�bola�de�cría.���
� El� comportamiento� de� relocación� del� alimento,� tan� característico� de� las�
especies� de� Eucraniini,� ha� sido� ampliamente� estudiado� por� diferentes� autores�
(Zunino� �� ��,� 1989;� Monteresino� y� Zunino,� 2003;� Ocampo,� 2004;� Ocampo� y�
Philips,� 2005).� Sin� embargo,� no� se� conocen� hasta� el� momento� demasiados�
detalles�de�la�arquitectura�de�sus�nidos,�y�mucho�menos�de�las�cámaras�pupales�
que�sus�larvas�construyen�antes�de�empupar.���
Esta�Tesis�doctoral�arroja�nuevos�datos�sobre�la�estructura�de�los�nidos�y�
de� las� masas/bolas� de� cría,� a� través� de� las� observaciones� de� campo� de� las�
distintas� estructuras,� como� así� también� del� trabajo� de� laboratorio� macro�� y�
micromorfológico.�Esto�permitió�hacer�comparaciones�con�las�bolas�de�cría�fósiles�
de�escarabajos�coprófagos,������������,�para�entender�el�comportamiento�de�
sus�productores.�
�
2�2 # ������ � ����� �� � ���������� �� ������������ �� ���
����� ?����� @ ���� ��A@ �������� ������ � ?������ �����#
���������������A
�
� El� origen� y� la� rápida� evolución� de� los� hábitats� dominados� por� pastos�
(“GDH:�Grass�Dominated�Habitats”)�que�incluyen�estepas,�pastizales�templados,�
y� sabanas� tropicales� y� subtropicales� (Retallack,� 1990,� 2001;� Shaw,� 2000;�

� ���
Strömberg,�2002,�2004),�han�sido�de�gran�interés�en�los�últimos�años.�Diferentes�
evidencias�paleontológicas,�tales�como�el�análisis�de�fitolitos,�la�hipsodoncia�y�la�
cursorialidad�en�mamíferos,�y�los�paleosuelos,�entre�otros,�han�sido�usados�para�
demostrar�su�presencia�debido�a�la�escacez�de�registros�paleobotánicos�directos�
(Retallack,� 2001;� Wooler� y� Beijing,� 2002;� Strömberg,� 2004).� En� América� del�
Norte,� los� ungulados� fósiles� del� Mioceno� serían� los� primeros� indicadores� del�
origen�de�pastizales�(Janis�����,�2000),�al�igual�que�lo�postulado�para�el�Mioceno�
de� Australia,� África,� y� Eurasia� (Retallack,� 2001)� pero� usando� para� ello� datos�
paleobotánicos�(Jacobs�����,�1999).�Los�hábitats�más�antiguos�dominados�por�
pastos� fueron� registrados� en� el� Oligoceno� tardío� de� Oregon� sobre� la� base� del�
estudio� de� los� paleosuelos� (Retallack,� 2004).� En� América� del� Sur,� diferentes�
evidencias,�tales�como�la�temprana�hipsodoncia�de�los�mamíferos,�la�abundancia�
de� fitolitos� de� Poaceae,� y� los� paleosuelos� ándicos� con� peds� granulares,�
estudiados� en� el� Eoceno�Medio�Mioceno� Inferior� de� la� Formación�Sarmiento� en�
Gran� Barranca� (Patagonia� central),� apoyan� la� idea� del� origen� de� los� hábitats�
dominados� por� pastos,� en� el� Eoceno� Medio� (Pascual� y� O.� Jaureguízar,� 1990;�
Zucol,�2005;�Madden�����,�2005;�Bellosi�y�González,�2005,�en�prensa).��
Gran�Barranca�es�una�localidad�clásica�para�la�paleontología�de�mamíferos�
de� la� Formación�Sarmiento,� y� está� ubicada� en� el�margen� sur� del� lago�Colhue�
Huapi,�a�50�km�al�este�de�Sarmiento�en�la�provincia�de�Chubut,�Argentina.�Esta�
localidad� fue� ampliamente� explorada� por� Carlos� Ameghino� durante� la� última�
década� del� siglo� XIX,� quien� descubrió� además� varias� capas� con� abundantes� y�
sucesivas� faunas� de� mamíferos� que� fueron� estudiadas� por� su� hermano,�
Florentino� (1906).� Estudios� posteriores� llevados� a� cabo� en� Gran� Barranca� por�
paleontólogos� y� geólogos� de� diferentes� países� (Windhausen,� 1924;� Frenguelli,�
1929;� Simpson,� 1940;� Feruglio,� 1949;� Pascual��� ��,� 1985,� 1996;�Marshall���

� ��
��,� 1983,� 1986;� Kay� �� ��,� 1999;� Madden� �� ��,� 2003)� han� demostrado� la�
notable� importancia� paleontológica� de� esta� localidad.� De� esta� manera,� la�
localidad�de�Gran�Barranca,�y�áreas�vecinas,�son�considerados�de�los�sitios�más�
significativos�en�mamíferos�Cenozoicos�del�mundo�(Cifelli,�1985),�ya�que�se�han�
podido� reconocer� ocho� Edades�Mamífero� de�América� del� Sur� (“South�American�
Land�Mammal�Ages”)�(Flynn�y�Swisher,�1995;�Madden�����,�2005).��
Respecto�a�la�icnología,�Gran�Barranca�es�uno�de�los�mejores�ejemplos�de�
la� gran� abundancia� y� diversidad� que� las� construcciones� hechas� por� insectos�
alcanzaron�en�paleosuelos�sudamericanos�en�el�Paleógeno�(Genise,�2004;�Bellosi�
����,�en�prensa),�cuando�todos� los�grupos�de� insectos�modernos�ya�se�habían�
diversificado�(Grimaldi�y�Engel,�2005).�Allí� también�se�da�uno�de�los�casos�más�
típicos� de� la� Icnofacies� de� ������������ (Genise� �� ��,� 2000),� debido� a� la�
presencia�de�numerosos�paleosuelos�con�diferentes�asociaciones�de�trazas�fósiles�
producidas� por� himenópteros� y� coleópteros.� Estos� paleosuelos� estudiados� por�
Bellosi�y�González�(2005,�en�prensa),�fueron�utilizados�también�para�mostrar�las�
modificaciones� de� las� capas� originales� de� los� depósitos� debido� a� icno�� y�
pedofábricas� (Genise�����,� 2004).�Además,�Gran�Barranca�posee�un�detallado�
armazón�lito�y�cronoestratigráfico�y�paleoambiental�basado�sobre�varios�perfiles�
sedimentológicos� y� magnetoestratigráficos,� y� sobre� numerosas� edades�
radimétricas� (Bellosi� y�Madden,�2005;�Madden�����,�2005),�que� junto�a�otras�
evidencias� paleontológicas� y� paleopedológicas� proveen� un� excelente� caso� de�
estudio�para�analizar�cambios�en�la�composición�de�las�trazas�fósiles�a�través�de�
los� sucesivos� paleosuelos� que� las� contienen.� Este� conjunto� de� información�
multidisciplinaria�es� integrado�con� los�resultados�del�análisis� icnológico�a� fin�de�
lograr�interpretaciones�paleoambientales�con�mayor�soporte.��

� ���
En�relación�con�el�contenido�icnológico,�las�trazas�fósiles�comprendidas�en�
el� icnogénero� �����������,� son� unas� de� las� mejores� representadas� en� el�
registro�icnofósil�(Bellosi�y�Genise,�2004;�Bellosi�����,�en�prensa).�Las�bolas�de�
cría�de�escarabajos�coprófagos�actuales,�semejantes�a�las�fósiles�incluidas�en�las�
icnoespecies� de� �����������,� son� el� resultado� del� comportamiento� de�
reubicación� del� alimento� mencionado� previamente.� En� hábitats� abiertos,� el�
excremento�depositado�sobre�la�superficie�por�los�mamíferos,�es�decir,�la�fuente�
de�alimento�mayormente�empleada�por�estos�escarabajos,�está�expuesto�a�una�
rápida�desecación� (y�a�una� fuerte� competencia).� Por�ello,� para� ser�protegido�y�
para�evitar�la�pérdida�de�agua,�el�excremento�es�transportado�y�enterrado�en�el�
sitio� donde� posteriormente� serán� construidas� las� bolas� de� cría� (Halffter� y�
Edmonds,� 1982).� Los�mamíferos� involucrados� son�más�abundantes� en�hábitats�
abiertos� dominados� por� pastos� (“open�GDH”),� y� de� acuerdo� con� esto,� los�
escarabajos� coprófagos� actuales� son� considerados� típicos� representantes� de�
estos�ecosistemas�(Halffter�y�Mathews,�1966;�Halffter�y�Edmonds,�1982;�Hanski�
y�Cambefort,�1991).��
En� esta� Tesis� se� analizarán� entonces,� los� cambios� en� la� fauna� de�
escarabajos�coprófagos,�desde�el�Eoceno�medio�al�Mioceno�temprano,�reflejados�
en�sus�trazas�fósiles�y�en�relación�con�ello,�se�inferirá�la�evolución�de�los�hábitats�
dominados�por�pastos�en�la�localidad�de�Gran�Barranca.��
�
2�4# ������������������ ������ ����� �� �����������������
��������������������������?�����������A
�
Existen�muy�pocos�antecedentes�de�trazas�de�un�organismo�producidas�en�
trazas�de�otros�organismos�en�el� registro� icnofósil� continental,�y� la�mayoría�de�

� ��
ellos�tratan�sobre�túneles�en�coprolitos,�o�sobre�depresiones,�agujeros�o�galerías�
hechos� en� nidos� fósiles� de� insectos� (Chin� y� Gill,� 1996;� Chin,� 2007;� Mikuláš� y�
Genise,� 2003;� y� referencias� en� ellos).� En� el� registro� geológico,� pueden�
reconocerse� dos� tipos� de� trazas� en� trazas:� en� un� caso� se� trata� de� trazas� que�
fueron� construidas� accidentalmente� adentro� o� en� contacto� con� otras� trazas;�
mientras� que� en� el� segundo� tipo� la� co�ocurrencia� de� las� trazas� se� debe� a� la�
interacción� entre� dos� organismos� que� se� encuentran� relacionados�
biológicamente.�Se�observan�ejemplos�del�primer�tipo,�en�trazas�de�escarabajos�
coprófagos�(�����������)�usadas�como�sustrato�por�las�de�abejas,�cigarras�y�
otros� escarabajos� (Laza,� 2006;� y� en� esta� Tesis).� Sin� embargo,� es� sólo� en� el�
segundo� tipo� de� trazas� en� trazas� donde� pueden� verse� reflejados� aspectos�
paleobiológicos� significativos� de� los� productores.� En� la� mayoría� de� ellos� se�
reconocen� diferentes� tipos� de� parasitismos,� donde� los� insectos,� son� los�
principales�actores.�Los� trabajos�publicados�hasta�el�momento,�muestran�casos�
de�cleptoparasitismo�entre�escarabajos,�donde�los�túneles�de�provisión�de�un�tipo�
de� escarabajo� son� surcados� por� los� de� los� cleptoparásitos;� o� casos� donde� los�
nidos�de�diferentes�tipos�de�insectos�han�sido�atacados�por�parasitoides�(Chin�y�
Gill,� 1996;� Martin,� 2001;� Mikul� y� Genise,� 2003;� Genise� y� Cladera,� 2004).�
Mikuláš�y�Genise�(2003),�estudiaron�varios�tipos�de�cámaras�fósiles�de�insectos,�
en� las� que� identificaron� dos� tipos� de� trazas� excavadas� en� sus� paredes�
construidas.� Como� resultado� de� estas� observaciones,� crearon� por� primera� vez,�
dos�icnogéneros�de�trazas�en�trazas,��������� �����y���� ����.�Los�agujeros�
y�depresiones�excavados�en�la�pared�construida�de�los�nidos�fósiles�de�insectos,�
fueron�incluídos�en��������� ����-�mientras�que�los�agujeros�excavados�en�la�
pared�que�se�encuentran�conectados�con�galerías�en�el�relleno,�fueron�incluidos�
en���� ����.�Por�otro�lado,�sugirieron�también�que�estos�tipos�de�trazas�fósiles�

� ���
podrían�ser�el�resultado�de�la�actividad�de�organismos�parasitoides�(cuyas�larvas�
se� desarrollan� junto� con� la� larva� del� constructor� del� nido,� parasitándola� hasta�
matarla),� cleptoparásitos� (organismos�que� roban� las�provisiones� colectadas�por�
otros�para�el�desarrollo�de�sus�propias�larvas),�o�detritívoros�(organismos�cuyas�
larvas�se�alimentan�de�los�restos�orgánicos�de�los�nidos).�
� Se� presentan� en� esta� Tesis� los� distintos� tipos� de� trazas� halladas� en� las�
bolas� de� cría� fósiles� de� escarabajos� coprófagos� (�����������),� las� cuales�
podrían�ser,�a�priori,�atribuidas�a�organismos�cleptoparásitos�o�detritívoros.��
�
2�5#������������� ������� ��� ���
�
Los�fitolitos�se�conocen�desde�mediados�del�siglo�XIX�y�han�sido�utilizados�
por�científicos�de�diferentes�disciplinas�(Piperno,�2006).�En�Argentina,�el�análisis�
de� la� producción� de� fitolitos,� su� taxonomía� y� preservación� en� suelos� y�
paleosuelos,� tiene� más� de� treinta� años� de� historia� (Bertoldi� de� Pomar,� 1971,�
1975;�Spalletti�y�Mazzoni,�1977,�1979;�Mazzoni,�1979;�Zucol�����,�1999;�Zucol�
y�Osterrieth,�2002).�Sin�embargo,�las�investigaciones�llevadas�a�cabo�con�fitoltos�
extraídos�de�otros�sustratos,�diferentes�de�plantas�y�rocas�sedimentarias,�como�
las�trazas�fósiles,�son�más�recientes�en�la�literatura�y�la�mayoría�de�ellas�están�
más� relacionadas� con� la� inferencia� de� la� dieta� de� los� herbívoros� a� partir� del�
análisis�del�contenido�fitolítico�en�coprolitos�(Horrocks�����,�2002,�2003,�2008;�
Prasad�����,�2005;�Piperno�y�Sues,�2005).���
Los�hábitos�alimentarios�y�la�dieta�de�herbívoros�fósiles�pueden�deducirse�
a� partir� de� diferentes� fuentes,� tales� como� la� evolución� de� la� hipsodoncia,�
microscopía� de�desgaste�en�esmalte�de�dientes� (“tooth�microwear”),�desarrollo�
de� facetas� de� las� superficies� oclusales� de� los� dientes� (“mesowear� análisis”),�

� ���
análisis� isotópicos� y� coprolitos,� entre� otros� (e.g.� Chin� y� Gill,� 1996;� Kingston� y�
Harrison,� 2007;� Rivals� y� Athanassiou,� 2008;� Valli� y� Palombo,� 2008).� Entre� los�
estudios� que� involucran� fitolitos,� el� contenido� microfósil� vegetal� (incluyendo�
fitolitos)�en�coprolitos�es�usado�y�analizado�ampliamente�para�inferir�el�uso�dado�
por�hombres�prehistóricos�en�la�dieta�de�determinadas�plantas�(Horrocks�����,�
2002,�2003).�En�otras� contribuciones,� se�ha�analizado�el� contenido� fitolítico�en�
coprolitos� tanto� de� vertebrados� extintos� (Prasad��� ��,� 2005;� Piperno� y� Sues,�
2005),� como� de� vertebrados� con� representates� modernos� (Horrocks� �� ��,�
2008).�En�todos�estos�trabajos,�se�muestran�preservados�fragmentos�de�tejidos�
vegetales�silicificados,�a�partir�de�los�cuales�se�infiere�la�dieta�de�los�vertebrados�
basada� sobre� la� diversidad� de� morfotipos� de� fitolitos� hallados.� Prassad� y�
colaboradores� (2005),� extrajeron� fitolitos� correspondientes� a� pastos,�
dicotiledóneas,� coníferas� y� palmeras,� preservados� en� coprolitos� de� saurópodos�
titanosauridos�del�Cretácico�tardío�de�India,�concluyendo�que�el�posible�productor�
de� la� bosta� analizada,� se� alimentaba� de� manera� indiscriminada� de� diferentes�
tipos� de�plantas.�Horrocks� y� colaboradores� (2008),�estudiaron� fitolitos,� y� otros�
restos� vegetales,� extraídos� de� coprolitos� de� una� especie� de� loro� endémico� de�
Nueva� Zelanda,� ��������� ����������,� concluyendo� que� debido� a� la� diversidad�
hallada,� la� especie� de� ave� analizada� mostraba� un� comportamiento� alimenticio�
muy�versátil,�nutriéndose�de�un�amplio�espectro�de�plantas.��
Existe�hasta�el�momento�un�único�registro�relacionado�con�el�análisis�que�
se�presenta�en�esta�sección.�Strömberg�y�Stidham�(2001)�publicaron�un�resúmen�
mostrando�el� contenido� fitolítico� hallado� en������������� de�Patagonia,� en� el�
que� concluyen� que� de� acuerdo� a� los� fitolitos� extraídos,� correspondientes� a�
dicotiledóneas� arbóreas� y� herbáceas,� palmeras� y� otras� monocotiledóneas,� los�
herbívoros� productores� de� la� bosta� habrían� tenido� una� dieta�muy� diversa.� Sin�

� ���
embargo,� en� este� resúmen,� la� pared� y� el� relleno� de� los� ejemplares� de�
������������ fueron� procesados� de� manera� conjunta,� en� lugar� de� ser�
considerados� como� muestras� independientes,� y� además� no� se� hicieron�
comparaciones�con�el�contenido�fitolítico�presente�en�el�paleosuelo�circundante.�
Esta� comparación� imprescindible,� permite� evaluar� si� el� contenido� fitolítico�
obtenido�de�las�bolas�de�cría�fósiles,�corresponde�a�la�bosta�usada�originalmente�
por�los�escarabajos,�o�si�en�cambio,�refleja�únicamente�el�contenido�fitolítico�del�
material�del�suelo�usado�para�la�construcción�de�las�bolas.�Por�ello,�es�necesario�
analizar� como� muestras� independientes� tanto� la� pared� como� el� relleno� de� los�
ejemplares� de� �����������,� para� poder� determinar� si� el� contenido� fitolítico�
corresponde�a� la�bosta�aprovisionada�originalmente�o�al�material�del� suelo�que�
rellenó�las�bolas�como�consecuencia�de�la�emergencia�del�nuevo�adulto.�Por�otro�
lado,� también� es� crítico� determinar� si� los� especímenes� de� ������������
utilizados�corresponden�a�ejemplares�cerrados,�es�decir,�donde�no�tuvo�lugar� la�
emergencia� del� adulto,� o� a� ejemplares� abiertos.� En� el� primer� tipo� de�
������������(��� isp.�A,����������,����������),�se�podría�encontrar�preservada�
en�su�interior� la�provisión�original,�en�cambio�en�el�segundo�tipo�(����������),�
es� decir� en� ejemplares� abiertos,� una�gran� cantidad�de�material� del� suelo� pudo�
entrar�a�la�cámara�vacía�luego�de�la�emergencia�del�nuevo�adulto.��
El� objetivo� general� de� este� análisis� es� el� de� comprobar� si� el� contenido�
fitolítico� de� los� ejemplares� de������������-� los� cuales� representan� bolas� de�
cría�fósiles�de�escarabajos�coprófagos�aprovisionadas�con�bosta,�puede�ser�usado�
para�inferir�el�comportamiento�de�estos�escarabajos,�la�dieta�de�los�herbívoros,�y�
otros�aspectos�del�paleoambiente�involucrado.��
�
�

� ���
4����� ����
�
4�2#���� ������������
�
Los� objetivos� generales� de� este� trabajo� son� ?2A� la� reconstrucción� de� la�
historia� evolutiva� de� los� Scarabaeinae� y� organismos� asociados� durante� el�
Paleogeno�tardío�e�inicios�del�Neógeno,�y�?4A�la�caracterización�ambiental�de�los�
paleoecosistemas�del�Eoceno�medio�Mioceno�temprano�de�la�Patagonia�Central�y�
particularmente� sobre�el� origen�y�evolución�de� los�hábitats�abiertos�dominados�
por�pastos.�
�
4�4#���� ������ �������
�
2A� Relevar� y� revisar� icnotaxonómicamente� las� trazas� fósiles� del� icnogénero�
������������de� la� Formación�Samiento� (Eoceno�medio�Mioceno� inferior)�
en�la�localidad�de�Gran�Barranca,�Chubut,�Argentina.�
4A� Estudiar� nidos� y� cámaras� de� cría� de� diferentes� taxones� de� escarabajos�
coprófagos�actuales�(Coleoptera:�Scarabaeidae:�Scarabaeinae).�La�hipótesis�
de�este�objetivo�es�que�las�comparaciones�macro�y�micromorfológicas�entre�
las� bolas� de� cría� fósiles� y� actuales� de� la� misma� región� sudamericana�
permitirán�reconocer�a�los�productores�de�las�icnospecies�de������������.��
5A� Estudiar� el� comportamiento� de� nidificación� de� diferentes� taxones� de�
escarabajos�coprófagos�actuales�(Coleoptera:�Scarabaeidae:�Scarabaeinae).�
La� hipótesis� de� este� objetivo� es� que� el� estudio� del� comportamiento� de�
coprófagos�actuales�y�su�reflejo�morfológico�en�las�trazas�fósiles,�permitirá�
inferir�el�comportamiento�del�escarabajo�productor�de�las�trazas�fósiles.��

� ���
8A� Completar�el�registro�fósil�de�los�Scarabaeinae�con�la�información�obtenida�a�
partir� de� las� bolas� de� cría� fósiles� atribuídas� a� escarabajos� coprófagos�
(�����������).�La�hipótesis�de�este�objetivo�es�que�a�partir�de�las�trazas�
fósiles�se�podrán�sumar�nuevos�datos�que�no�se�obtienen�con� los�cuerpos�
fósiles,� algunos� de� importancia� filogenética� como� edades� máximas� de�
algunos�de�sus�comportamientos�y�taxones�productores.�
6A� Interpretar�los�cambios�ambientales,�y�particularmente�los�relacionados�con�
el� origen� y� evolución� de� los� hábitats� abiertos� dominados� por� pastos� en� la�
Patagonia� Central,� sobre� la� base� de� la� aparición,� abundancia� relativa� y�
diversidad� de� �����������.� Esta� meta� es� llevada� a� cabo� mediante� un�
trabajo� de� icnoestratigrafía� de� alta� resolución� por� primera� vez� en� la�
icnología� continental,� a� lo� largo� de� un� significativo� intervalo� temporal� de�
más�de�23�millones�de�años�representado�en�la�secuencia�de� la�Formación�
Sarmiento�aflorante�en�Gran�Barranca�(Chubut).�Por�lo�tanto,�las�hipótesis�a�
testear� son:� (1)� que� la� aparición,� densidad,� abundancia� relativa,� y�
desaparición�de�los�ejemplares�de������������;�(2)�que�el�incremento�de�
tamaño� de� ������������ en� relación� con� el� tamaño� corporal� de� los�
mamíferos� asociados;� y� (3)� que� el� tamaño� y� espesor� de� la� pared� de� las�
icnoespecies�de�������������permitirán� inferir� la�diversidad�potencial�de�
especies� de� escarabajos� coprófagos,� y� reconocer� los� cambios�
paleoambientales,�particularmente,�los�hábitats�dominados�por�pastos.��
3A� Reconocer� trazas� de� cleptoparásitos� y� otros� organismos� asociados� a�
������������y�su�significado�paleobiológico.�Las�hipótesis�de�esta�sección�
son:�(1)�una�nueva�icnoespecie�de��������� �����refleja�un�novedoso�caso�
potencial� de� cleptoparasitismo;� (2)� el� posible� productor� de� ��� �����
����������,� sería� un� organismo� con� comportamiento� cleptoparásito� o�

� ���
detritívoro;� y� (3)� las� trazas� en� trazas� permitirán� mostrar� aspectos�
paleoecológicos�de� la� comunidad�de� invertebrados�asociada�a� las�bolas�de�
cría�fósiles�de�escarabajos�coprófagos.�
;A� Evaluar� el� contenido� fitolítico� de� ������������ como� herramienta� para�
contribuir� al� conocimiento� del� comportamiento� de� los� escarabajos�
coprófagos,� la� dieta� de� los� mamíferos� con� cuyos� excrementos� se�
aprovisionaban�las�bolas�y�el�paleoambiente.�Para�ello,�se�testearon�en�esta�
sección� las� siguientes� hipótesis:� 1)� el� relleno� de� los� ejemplares� de�
������������ posee� un� número� de� fitolitos� similar� al� del� paleosuelo,�
debido�a�que� los�ejemplares�abiertos� fueron� rellenados�activamente�por�el�
nuevo�adulto�al�emerger,�o�posteriormente�por�otros�procesos�pasivos,�con�
material�del�suelo�circundante;�2)�el�número�de�fitolitos�de�la�pared�de�los�
ejemplares�de�������������difiere�del�número�hallado�en�el�relleno�y�en�
el� paleosuelo� circundante,� debido� a� que� queda� preservada� una� capa�
remanente� de� la� bosta� original.� Este� remanente� de� bosta� adherido� a� la�
pared�construida�con�material�del� suelo,�podría� incrementar�o�disminuir� la�
concentración�de�fitolitos�en�la�pared�en�función�del�paleoambiente,�debido�
principalmente� a� la� presencia� y� abundancia� de� plantas� con� fitolitos� en� el�
suelo� antes� de� su� soterramiento.� Un� suelo� rico� en� plantas� con� fitolitos,�
tendría� una� mayor� concentración� de� fitolitos� que� una� simple� muestra� de�
bosta,� que� al� preservarse� en� la� pared�de� las� bolas�de� cría,� disminuiría� su�
concentración�en�esa�muestra,�y�viceversa;�3)�el�número�de� fitolitos�en�el�
espesor� de� la� pared� de� los� ejemplares� de� ������������ muestra� un�
gradiente�de�concentración.�Si�las�hipótesis�1�y�2�son�verdaderas,�entonces:�
4)�la�diversidad�de�fitolitos�de�la�pared,�o�las�diferencias�en�sus�frecuencias�
relativas,�reflejarían�la�diversidad�de�plantas�consumidas�por�los�herbívoros,�

� ���
cuyos� fitolitos� se� habrían� conservado� en� la� bosta;� y� 5)� si� las� frecuencias�
relativas� de� los� diferentes� morfotipos� de� fitolitos� es� similar� entre� las�
muestras,� es� decir,� entre� pared,� paleosuelo� y� relleno,� entonces� los�
herbívoros� involucrados� no� mostrarían� preferencias� entre� las� plantas� con�
fitolitos� disponibles;� pero� si� las� frecuencias� entre� las� muestras� difieren,�
podría� revelarse� la� preferencia� de� los� herbívoros� por� algunas� plantas� con�
fitolitos�más�que�por�otras.��

� ��
5��������������
La�Formación�Sarmiento�es�una�sucesión�piroclástica�de�origen�continental�
del�Eoceno�Medio�Mioceno�Inferior,�expuesta�en�el�centro�y�norte�de�Patagonia�y�
conocida,�principalmente,�por�la�abundancia�y�diversidad�de�vertebrados�fósiles.�
Sus� asociaciones� de� mamíferos� son� consideradas� los� estándares�
bioestratigráficos� para� las� distintas� Edades� Mamífero� Sudamericanas� (“South�
American� Land� Mammal� Ages”� o� SALMAs)� (Ameghino,� 1906;� Simpson,� 1940;�
Pascual,� 1984;� Cifelli,� 1985).� Esta� unidad� es� relativamente� delgada� con� un�
espesor�promedio�regional�de�60�m,�y�esta�compuesta�por�diferentes�litologías:�
chonitas� (cenizas� volcánicas� finas� y� muy� finas),� bentonitas,� areniscas�
piroclásticas,�y�conglomerados�intraformacionales,�las�cuales�suelen�encontrarse�
modificadas�por�pedogénesis�(Spalletti�y�Mazzoni,�1979;�Mazzoni,�1979;�Bellosi,�
en�prensa).�Los�paleosuelos,�generalmente�conteniendo� trazas� fósiles,� son�otra�
característica� significativa� y� bien� conocida� de� esta� formación� (Andreis,� 1972;�
Spalletti� y� Mazzoni,� 1979;� Bellosi� �� ��,� 2001;� Genise� �� ��,� 2004).� Los�
afloramientos� a� lo� largo� de� Gran� Barranca,� situada� en� la� provincia� de� Chubut�
(Fig.� 1),� comprenden� la� sección� tipo� de� la� Formación�Sarmiento,� cuyo� espesor�
integrado�alcanza�355�m.��
Los�piroclastos�finos�y�félsicos�de�la�Formación�Sarmiento�fueron�aportados�
desde�dos�arcos�andinos�ubicados�en�el�noroeste�de�la�Patagonia�(a�500�km�de�
Gran� Barranca),� donde� tuvo� lugar� un� prolongado� vulcanismo� explosivo� tipo�
pliniano,�desde�el�Paleoceno�tardío�hasta�el�Mioceno�temprano�(Mazzoni,�1985;�
Rapela�����,�1988;�Bellosi,�en�prensa).�En�consecuencia,�el�material�parental�de�
los� paleosuelos� es� ceniza� volcánica� fina� compuesta� por� trizas� vítreas� riolíticas�
dacíticas� y� plagioclasa� subordinada� (andesita).� La� mayoría� de� los� paleosuelos�
� ���
están� bioturbados� (Genise� �� ��,� 2004),� muestran� un� horizonte� argílico,�
contienen� nódulos� de� óxidos� de� Fe� y�Mn,� o� carbonato� de� calcio,� una� b�fábrica�
moderada�a�bien�desarrollada,�y�argilanes�y�ferromanganes�(Bellosi�y�González,�
2005,� en� prensa).� De� acuerdo� con� sus� características,� estos� paleosuelos� se�
corresponden� con� Andisoles,� Alfisoles,� Alfisoles�Ultisoles,� Aridisoles,� Entisoles� y�
Vertisoles.��
�
�
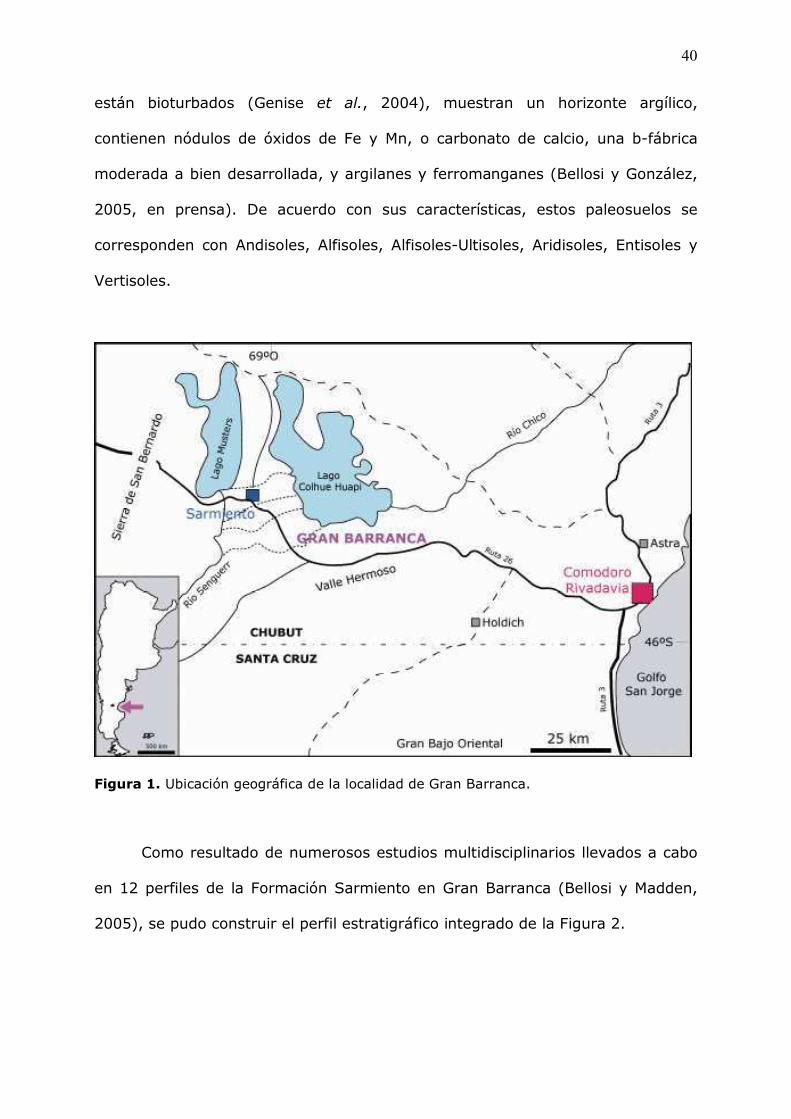
�!�%�&2��Ubicación�geográfica�de�la�localidad�de�Gran�Barranca.�
�
Como�resultado�de�numerosos�estudios�multidisciplinarios�llevados�a�cabo�
en�12�perfiles�de� la�Formación�Sarmiento�en�Gran�Barranca�(Bellosi�y�Madden,�
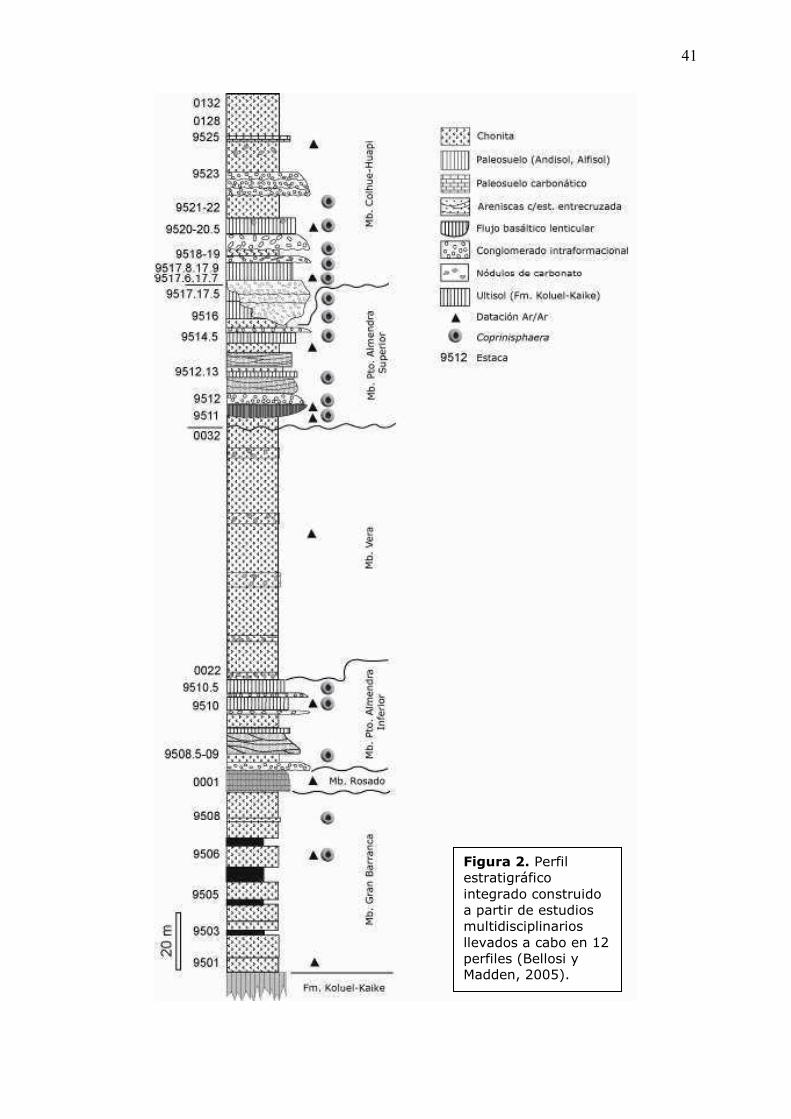
2005),�se�pudo�construir�el�perfil�estratigráfico�integrado�de�la�Figura�2.�
�
�
� ��
�!�%�&4��Perfil�estratigráfico�integrado�construido�a�partir�de�estudios�multidisciplinarios�llevados�a�cabo�en�12�perfiles�(Bellosi�y�Madden,�2005).�

� ���
En� este� perfil� simplificado,� las� muestras� y� los� niveles� se� encuentran�
identificados�por�estacas�numeradas.�La�mayoría�de�estas�estacas�están�referidas�
al� perfil� MMZ� ubicado� en� el� lado� occidental� de� Gran� Barranca� que,� aunque� es�
incompleto,�corresponde�al�perfil�originalmente�estudiado�por�Spalletti�y�Mazzoni�
(1979).� Los� niveles� estratigráficos� que� no� poseen� estacas� en� la� secuencia�
corresponden� a� otros� perfiles� (J,� K� o� M)� situados� hacia� el� este,� que�
coincidentemente� no� son� significativos� para� el� análisis� de� trazas� fósiles,� a�
excepción�de�la�muestra�de�la�estaca�9512�13�colectada�en�el�perfil�M.��
La� Formación� Sarmiento� en� Gran� Barranca� se� divide� en� seis� miembros�
delimitados�por�superficies�de�discontinuidad�(Bellosi�y�Madden,�2005).�Spalletti�
y�Mazzoni� (1979)� definieron� formalmente� tres� de� ellos� en� el� perfil� MMZ:�Gran�
Barranca�(Fig.�3�A�y�B),�Puesto�Almendra�(Fig.�3�D,�F�y�G),�y�Colhue�Huapi�(Fig.�
3�H).�Recientemente,�Bellosi�y�Madden�(2005,�en�prensa)�identificaron�un�cuarto�
miembro,�el�Rosado�(Fig.�3�C),�y�subdividieron�al�Miembro�Puesto�Almendra�en�
Inferior� (Fig.� 3� D)� y� Superior� (Fig.� 3� F� y� G)� debido� a� la� presencia� de� una�
discordancia�significativa�y�de�otro�nuevo�Miembro�(Vera,�Fig.�3�E)� intercalado.�
Las�edades�de�los�distintos�niveles�de�la�Formación�Sarmiento�fueron�obtenidas�
siguiendo� un� detallado� esquema� litoestratigráfico� que� incluye� superficies� de�
discontinuidad�(Bellosi,�en�prensa),�y�basado�sobre�numerosas�dataciones�Ar/Ar�
en� vídrio� volcánico� y� plagioclasas,� y� varios� perfiles�magnetoestratigráficos� a� lo�
largo�de�Gran�Barranca�(Madden�����,�2005;�Re�����,�en�prensa).�
El�Miembro�Gran�Barranca� (Fig.�3�A�y�B)�está� compuesto�por�chonitas�y�
bentonitas� macizas,� que� estan� pobremente� estratificadas� o� en� capas� muy�
gruesas.� Son� comunes� las� concreciones� diagenéticas� de� sílice� y� yeso� en� la�
sección� inferior�donde� los�paleosuelos�no�son�abundantes�y�además�estan�poco�
desarrollados� (estacas� 9501� a� 9505).� En� la� sección� superior� (estacas� 9506� a�

� ���
9508),�los�paleosuelos�están�mejor�desarrollados�y�muestran�también�horizontes�
de� nódulos� y� costras� de� manganeso,� potentes� y� continuos,� y� trazas� fósiles,�
incluyendo� ������������ (Bellosi� �� ��,� en� prensa).� Durante� el� tiempo� que�
abarca� el� Miembro� Gran� Barranca,� prevaleció� una� sedimentación� loessica�
subaérea� en� planicies� con� ondulaciones� y� cursos� fluviales� reducidos,� y� bajos�
ocupados� por� pequeños� lagos� efímeros.� Las� loessitas� téfricas,� con� sus�
paleosuelos� asociados,� se� formaron� en� un� clima� estacional� subhúmedo� a�
semiárido�(Bellosi,�en�prensa).�La�abundancia�de�fósiles�de�mamíferos�y�reptiles�
hallada� en� este� miembro,� corresponde� al� (SALMA)� Barranquense�
(Casamayorense� tardio)� del� Eoceno� Medio� (Kay� �� ��,� 1999;� Madden� �� ��,�
2005;� Re��� ��,� en� prensa).� Por� consiguiente,� las�muestras� icnológicas� de� las�
estacas�9506�y�9508�del�Miembro�Gran�Barranca�son�asignadas�al�Eoceno�Medio�
(alrededor�de�39�ma).��
El�Miembro�Rosado�(Fig.�3�C)�es�un�paleosuelo�con�carbonato�(calcrete)�de�
7� m� de� espesor,� que� posee� una� diversa� asociación� de� microgasterópodos�
terrestres� (Bellosi� �� ��,� 2002a;� Miquel� y� Bellosi,� 2007)� y� mamíferos� fósiles�
pertenecientes� al� (SALMA)� Mustersense� (Eoceno� Medio)� (Kay� �� ��,� 1999;�
Madden� �� ��,� 2005).� Las� trazas� fósiles� son� muy� escasas� y� no� se� registran�
ejemplares� de� ������������ (Bellosi� �� ��,� en� prensa).� La� sedimentación�
loéssica� ocurre� también� en� este� tiempo,� y� los� paleosuelos� indican� condiciones�
semiáridas� a� áridas� durante� las� etapas� tardías� de� pedogénesis� (Bellosi� �� ��,�
2002a).�
El�Miembro� Puesto� Almendra� Inferior� (30�m�de� espesor)� (Fig.� 3�D)� está�
bien� estratificado� y� posee� capas� lenticulares� y� tabulares� de� conglomerados�
intraformacionales,� areniscas� con� estratificación� entrecruzada,� chonitas� y�
bentonitas,� algunas� de� ellas� con� paleosuelos� (Andisoles,� Alfisoles).� Esta�

� ���
asociación� de� facies� se� acumuló� en� un� ambiente� fluvial� entrelazado,�
probablemente�por� ríos� no�permanentes.� Las� planicies� subaéreas� adyacentes� y�
los� lagos�pequeños�y�poco�profundos,�habrían�agradado�por�acumulación�eólica�
de� sedimentos� volcaniclásticos� finos.� Los� alfisoles� ándicos,� moderada� a�
pobremente� desarrollados,� sugieren� condiciones� sub�húmedas� para� este�
miembro� (Bellosi,� en�prensa).� Los� fósiles�de�mamíferos�y�de�otros�vertebrados�
preservados�en�facies�fluviales�son�también�asignados�al�(SALMA)�Mustersense,�y�
el�análisis�cronológico�sugiere�que�la�edad�de�este�miembro�es�Eoceno�medio�a�
tardío.� Las� trazas� fósiles� de� insectos� son� abundantes,� particularmente�
�����������,� las� cuales� fueron� colectadas� en� tres� niveles� estratigráficos�
(estacas�9508.5�9509;�9510�y�9510.5).�
El�Miembro�Vera�(Fig.�3�E),�desarrollado�en�el�lado�Este�de�Gran�Barranca�
(perfiles�J�a�M),�es�una�sucesión�homogénea�de� loessitas� téfricas�de�105�m�de�
espesor,� con� unas� pocas� capas� delgadas� de� bentonitas� y� paleosuelos� cálcicos�
muy� débilmente� desarrollados.� Las� trazas� fósiles� de� invertebrados� estan�
prácticamente�ausentes�en�este�miembro�(Bellosi�����,�en�prensa),�y�los�restos�
de� mamíferos� colectados� de� la� sección� media,� pertenecen� al� (SALMA)�
Tinguirririquense� del� Oligoceno� temprano� (Kay� �� ��,� 1999).� La� asociación� de�
depósitos� loéssicos� y� paleosuelos� con� nódulos� calcáreos� débilmente�
desarrollados,� registran� una� importante� modificación� del� clima� y� del� ambiente�
durante� la� transición� Eoceno�Oligoceno,� que� involucró� un� decrecimiento� en� las�
precipitaciones�y�en�la�cobertura�vegetal�(clima�semiárido).�Edades�Isotópicas�y�
perfiles�magnetostratigráficos�sugieren�que� la�edad�de�este�miembro�es�Eoceno�
tardío�Oligoceno�temprano�(Madden�����,�2005).�
El�Miembro� Puesto� Almendra�Superior� (46�m�de� espesor)� (Fig.� 3� F� y�G)�
está� compuesto� por� conglomerados� intraformacionales,� chonitas� y� areniscas,�

� ���
bentonitas,�paleosuelos�apilados,�pobre�a�fuertemente�desarrollados�(Andisoles,�
Alfisoles,� Alfisoles�Ultisoles),� y� flujos� de� lava� lenticulares.� La� sedimentación�
fluvial� estuvo� restringida� a� pequeños� canales� sinuosos� que� retransportaron�
materiales�piroclásticos.�El�desarrollo�de�los�paleosuelos�en�este�miembro�sugiere�
condiciones� estacionales� subhúmedas� a� húmedas,� y� paisajes� más� estables�
(Bellosi,�en�prensa).�Los�fósiles�de�mamíferos�no�son�abundantes�y�corresponden�
al� (SALMA)� Deseadense� del� Oligoceno� (Flynn� y� Swisher,� 1995).� Los� estudios�
geocronológicos� basados� sobre� edades� isotópicas� de� basaltos� y� rocas�
piroclásticas,� junto� con� los� perfiles� paleomagnéticos,� sugieren� una� edad�
oligocena�tardía,�para�este�miembro�(Madden�����,�2005;�Re�����,�en�prensa).�
De� acuerdo� con� las� significantes� variaciones� en� la� diversidad,� densidad� y�
morfología�de�������������registrados�en�este�miembro,�se�identificaron�tres�
intervalos� correspondientes� con� las� siguientes�estacas�y�edades:�9512�y�9512�
9513�(Oligoceno�tardio�bajo);�9514.5�y�9516�(Oligoceno�tardío�medio);�y�9517�
9517.5�(Oligoceno�tardio�alto).�
El� Miembro� Colhue�Huapi� (64� m� de� espesor)� (Fig.� 3� H)� se� encuentra�
separado� del� Miembro� Puesto� Almendra� Superior� por� una� discordancia� erosiva�
profunda�(Bellosi�����,�2002b),�la�cual�involucra�un�hiato�de�3.1�a�3.5�millones�
de�años� (Re�����,� en�prensa).�Se� reconocen�en�el�Miembro�Colhue�Huapi�dos�
secciones.� La� inferior� se� encuentra� compuesta� por� conglomerados�
intraformacionales,� chonitas� macizas� y� paleosuelos� apilados,� pobre� a�
moderadamente� desarrollados,� con� abundantes� trazas� fósiles.� Esta� sucesión�
rellena� un� paleovalle� inciso� en� la� antigua� llanura� costera� de� Patagonia� central,�
que�fue�posteriormente� inundado�por�el�mar�Superpatagoniano�más�al�Este.�La�
incisión�del�paleovalle�estuvo�relacionada�con�una�caída�significativa�del�nivel�del�
mar�en�21�ma.�Durante�una�primera�etapa,�la�sedimentación�ocurrió�en�canales�

� ���
fluviales� (probablemente� efímeros)� y� en� planicies� edafizadas.� La� asociación� de�
facies� y� paleosuelos� (Alfisoles)� sugieren� un� clima� subhúmedo� estacional� y� un�
incremento�en�la�tasa�de�sedimentación�(Bellosi,�en�prensa).�Abundante�cantidad�
de� fósiles� de� mamíferos,� asignados� al� (SALMA)� Colhuehuapense� del� Mioceno�
temprano� (Marshall��� ��,� 1986;� Flynn� y� Swisher,� 1995;� Kay��� ��,� 1999)� se�
encuentran� preservados� en� cuerpos� de� canales� fluviales� y� paleosuelos� de� la�
sección� inferior.� La� sección� superior� está� constituida� por� loessitas� téfricas�
gruesas� y� escasos� paleosuelos� cálcicos,� débilmente� desarrollados� y�
prácticamente� sin� trazas� fósiles,� que� representan� un� ambiente� eólico� y�
semiárido.� En� la� parte� superior� de� esta� sección,� los� restos� de� mamíferos�
corresponden� al� (SALMA)� Pinturense� del� Mioceno� temprano� a� medio� (Bown� y�
Fleagle,� 1995).� Los� estudios� geocronológicos� indican� una� edad� miocena�
temprana� para� el� Miembro� Colhue�Huapi� (Madden��� ��,� 2005).� En� la� sección�
inferior� de� este� miembro� se� colectaron� ������������ en� cinco� niveles�
estratigráficos:� estacas� 9517.6�9517.7,� 9517.8�9517.9,� 9518�9519,� 9520�
9520.5� y� 9521�9522.� En� la� sección� superior� no� se� observaron� trazas� fósiles�
(estacas�9523�a�9525.5)�(Bellosi�����,�en�prensa).�
En� la� Formación�Sarmiento,�en�Gran�Barranca,� los� fitolitos�de�gramíneas�
(braquiolitas,� prismatolitas,� aculeolitas,� flabelolitas,� halteriolitas,� doliolitas,� y�
estrobilolitas)� y� de� palmeras� (globulolitas)� son� comunes� y� hasta� muy�
abundantes.� Spalletti� y� Mazzoni� (1979)� y� Mazzoni� (1979),� en� los� primeros�
análisis� fitolíticos� llevados�acabo�en�esta� localidad,�mencionan�que�braquiolitas,�
prismatolitas,� aculeolitas� y� flabelolitas,� son� los� morfotipos� más� comunes�
encontrados�a�lo� largo�de�la�secuencia,�siendo�los�primeros�los�más�frecuentes.�
Tiempo� después,� Zucol� y� colaboradores� (1999,� 2001,� 2005)� analizaron� la�
secuencia,� desde� la� base� hasta� el� tope,� y� encontraron� asociaciones� dominadas�

� ���
por� fitolitos� de� palmeras� (globulolitas� esféricas� a� subesféricas)� junto� con�
prismatolitas,�aculeolitas�y�fitolitos�indeterminados�durante�el�Eoceno�Medio,�las�
cuales,� hacia� arriba� en� la� secuencia,� son� reemplazadas� por� asociaciones� con�
mayor� abundancia� de� fitolitos� graminoides� (halteriolitas,� aculeolitas� y�
flabelolitas).� De� estas� asociaciones,� aquéllas� con� componentes� panicoides�
dominaron�en�el�Eoceno�tardío�y�aquéllos�con�componentes�chloridoides�pooides,�
dominaron� en� el� Oligoceno� –� Mioceno� temprano.� La� sección� superior� de� la�
secuencia� (Mioceno� temprano)� muestra� una� distribución� heterogénea� de�
morfotipos� de� fitolitos,� con� niveles� estratigráficos� dominados� por� globulolitas�
esféricas�a�subesféricas,�y�fitolitos�panicoides,�pooides�y�chloridoides.��
��
���
�
�
�
�
�
�
�
�
�
�

� ���
�
�!�%�& 5� �� Miembro� Gran� Barranca� que� se� extiende� en� la� imagen� hasta� la� linea� de�color;���� Colecta� de� especimenes� en� el� Miembro� Gran� Barranca;���� Miembro� Rosado,�expuesto�a�partir�de� la� linea�de�color;����La� flecha� indica�el�Miembro�Puesto�Almendra�Inferior;����Miembro�Vera;���La�flecha�indica�el�Miembro�Puesto�Almendra�Superior;����Colecta�de�especimenes�en�el�Miembro�Puesto�Almendra�Superior;���Miembro�Colhue�Huapi.��
�

� ��
8�� ��������9 ����
8�2#������ ���� ������������������� �� �������������������
���������������� ��
� �
� Los�estudios�neoicnológicos�llevados�a�cabo�para�esta�investigación,�tienen�
como� fin� la� obtención� de� datos� relacionados� con� el� comportamiento� de�
escarabajos� coprófagos� actuales,� a� través� del� estudio� de� sus� nidos� y�
principalmente�de�bolas�de�cría,� las�que�fueron�posteriormente�comparadas�con�
bolas�de�cría�fósiles.��
�
8�2�2#��'�**!> $�'&"�"B�*!�"$��"=%$!�
�
� Como� criterios� de� selección� de� las� especies� a� estudiar,� se� tuvieron� en�
cuenta� características� tales� como� el� tipo� de� comportamiento� de� nidificación�
(Patrón� I,� II,� etc.),� el� tipo� de� nido� (para� o� telecóprido)� y� cámaras� de� cría�
construidas� por� cada� especie� (masa� o� bola� de� cría),� etc.,� en� función� de� su�
aplicación�en�la�comparación�con�bolas�de�cría�fósiles.�No�obstante,�se�estudiaron�
también�especies�no�productoras�de�bolas�con�el�objeto�de�obtener�un�panorama�
general� propio� de� los� distintos� patrones� de� nidificación� y� poder� realizar� las�
comparaciones�micromorfológicas� ausentes� en� la� bibliografía.� De� esta�manera,�
las�especies�seleccionadas�y�estudiadas�fueron�las�siguientes:�
�
Onthophagini:������������sp.��
Phanaeini:���������� �������Brull�
��� ������������(Laporte)�

� ���
��� ���������������(Chevrolat)�
��� ������������(Harold)��
Dichotomiini:������������ ����(Fabricius)��
Canthonini:� ��������! ���������"���� �� ������(Blanchard)�
� ��������! �������"��������(Gillet)��
��������sp.1��
��������sp.2��
Eucraniini:�$������������������(Burmeister)�
$���������������� �&��(Blanchard)�
'� ������ & ������Burmeister�
�
8�2�4# �&)&C�$�*&(B�
� Los� nidos� de� �������� � ������,� �������� ��� ���� y� ��� �������
����� fueron�estudiados�durante�enero,� febrero,�marzo,�abril�y�diciembre�de�
2006,� y� marzo� de� 2007,� meses� en� los� que� se� registra� la� mayor� actividad� de�
adultos.� Para� ello,� se� realizaron� reiterados� viajes� y� estadias,� en�un� campo� con�
importante� cantidad� de� vacas� y� caballos� de� la� localidad� de� Navarro,� Buenos�
Aires,�y�con�abundante�cobertura�vegetal�dominada�por�pastos.�Se�examinaron�y�
midieron� ��� ����,� 6�nidos�de���� � ������� conteniendo�un� total�de�3�masas�de�
cría,�27�nidos�de������� ����de�los�que�se�obtuvieron�9�bolas�de�cría,�y�17�nidos�
de���������de�los�cuales�sólo�se�hallaron�3�bolas�de�cría.�Las�masas�y�bolas�
de�cría�colectadas�fueron�transportadas�al�laboratorio�para�su�posterior�estudio.��
� Los� nidos� de� ��� ������� �������,� ��� ����,� ��������
! ���������"� ��� �� �����,� �� ! �������"� ������,� ������� sp.1,�
'� ������ & �����,�$���������������� �&��y�$�������,�fueron�estudiados�

� ��
durante�viajes�de�campaña�en�los�meses�de�enero�de�2007�y�febrero�de�2008�a�
distintas�localidades�de�La�Rioja:�Anillaco,�Vera�Cruz,�Chuquis,�Antinaco,�Pituil�y�
Pampa� de� la� Viuda,� dependiendo� de� cada� caso.� Se� revisaron� lugares� con�
diferente� disponibilidad� de� bosta� y� con� distinta� cobertura� vegetal,� incluso� en�
terrenos�al�costado�de�la�ruta,�caminos�de�tierra�entre�los�pueblos,�y�en�caminos�
en�altura.�Se�examinaron�y�midieron��������,�7�nidos�de������������en�Chuquis,�
de� los� que� se� extrajeron� dos� bolas� de� cría,� una� de� ellas� en� muy� malas�
condiciones�de�preservación,�5�en�Anillaco�(camino�a� la�Quebrada)�y�3�en�Vera�
Cruz�de� los�cuales�no�se�obtuvieron�bolas�de�cría.��������� fue�encontrado�en�
Pampa�de� la�Viuda,� donde� los�nidos�no� fueron�estudiados�en�detalle�porque�el�
trabajo�de�campo�se�restringió�a�la�colecta�de�bolas�de�cría�(3�bolas�de�cría),�y�
en�Vera�Cruz,�donde�se�examinaron�5�nidos�en�los�que�no�se�hallaron�bolas�de�
cría.� Se� estudiaron� 2� nidos� de� ��������� ! ���������"� ��� �� ������ en�
Vera�Cruz,�y�1�en�Chuquis,�de�los�cuales�se�extrajeron�bolas�de�alimentación.�A�
su�vez,� los�nidos�de�esta�especie,�fueron�también�estudiados�en�condiciones�de�
laboratorio,�detalladas�mas�adelante,�usando�para�ello�escarabajos�colectados�en�
Anillaco.�Los�nidos�de� ��������! �������"��������fueron�directamente�
estudiados�en�condiciones�de� laboratorio,� como�se�describe�mas�abajo,�usando�
para� ello� escarabajos� colectados� por� la� noche� en� las� calles� de� Anillaco.� Sin�
embargo,�las�bolas�de�cría�usadas�para�las�descripciones�fueron�colectadas���������
en� Chuquis� al� costado� de� la� ruta� provincial� Nº1� (km� 83),� en� un� ambiente�
semiárido�caracterizado�por�una�cobertura�vegetal�irregular,�y�con�poco�ganado,�
en�el�cual�se�encontraron�en�la�superficie�del�suelo�bolas�de�cría�removidas�por�
roedores�cavícolas.�De�la�misma�manera�fueron�estudiados�los�nidos�de��������
sp.1� colectados� en� Antinaco.� Por� último,� los� nidos� de� Eucraniini� fueron�
examinados�y�estudiados��������,�en�diferentes�localidades�de�La�Rioja.�Los�nidos�

� ���
de�'� ������ & ������fueron�estudiados�en�Chuquis�(1)�y�en�la�Quebrada�en�
Anillaco� (1);� los�de�$������������������en�Pituil� (2),�y� los�de�$������������
���� �&�� fueron� estudiados� también� en� Pituil� (1)� asociados� a� nidos� de�
roedores,�y�finalmente�en�Vinchina�(2).�
� La�bola�de�cría�de��������sp.�2�fue�colectada�en�la�Reserva�Experimental�
de�Horco�Molle�(REHM),�en�el�departamento�de�Yerba�Buena,�al�pie�del�parque�
Sierra�de�San�Javier�y�a�15�km�al�Oeste�de�San�Miguel�de�Tucumán�durante�el�
mes� de� noviembre� de� 2007,� y� donada� a� la� colección� de� icnología� del� Museo�
Argentino�de�Ciencias�Naturales�por�Paula�Cristini�y�Juan�Farina.�La�bola�de�cría�
fue� encontrada� debajo� de� una� roca� junto� a� un� adulto,� sobre� la� superficie� del�
suelo�y�en�posición�lateral,�es�decir,�con�su�eje�mayor�paralelo�al�suelo.�El�sitio�
donde�se�halló�dicho�material�es�un�sector�de�unos�500�metros�cuadrados,�con�
abundante�pasto�y�árboles,�que�se�encuentra�en�el�acceso�al�parque,�cerca�de�la�
ruta.� Dada� la� cercanía� con� el� lugar� donde� se� encuentran� los� animales� en� la�
reserva,�y� la�diversidad�de� los�mismos,�estos�escarabajos�podrían�estar�usando�
bosta� tanto� de� herbívoros� como� de� carnívoros.� Se� los� ha� visto� rolar� trozos� de�
bosta� de� yaguarundí� ()��� &���������,� Felidae),� e� incluso� se� los� ha� visto�
acarrear� también�pichones�de�bicho� feo� ()������� ����������,� Tyrannidae)� (A.�
Molina,�comunicación�personal)�(Fig.�28�D).�����
� Los�nidos�de������������sp.�fueron�estudiados�durante�una�campaña�en�
el� mes� de� mayo� de� 2007� al� Parque� Nacional� Río� Pilcomayo,� Formosa.� La�
excavación� de� los� nidos� se� realizó� en� un� terreno,� fuera� del� Parque,� inundable�
debido� a� estar� parcialmente� circundado� por� una� laguna� (Laguna� Blanca).� Esta�
parcela�tiene�vacas�y�caballos,�y� la�cobertura�vegetal�es�abundante�y�dominada�
por� pastos.� Se� examinaron� y� midieron� ��� ����,� 3� nidos� de� de� los� cuales� se�

� ���
extrajeron� 4� masas� de� cría� que� fueron� transportadas� al� laboratorio� para� su�
estudio.��
� En� todos� los� casos� presentados� aquí,� las� descripciones� detalladas� de� la�
arquitectura� de� los� nidos� fueron� basadas� en� los� nidos� más� completos;� sin�
embargo,� los� datos� obtenidos� de� las� mediciones� y� morfología� de� los� nidos�
fragmentados� fueron� usados� también� como� complemento� de� estructuras�
faltantes�o�incompletas.�
�� El� trabajo� de� campo� empleado� fue� similar� en� todos� los� casos,� con�
pequeñas� variaciones� dependiendo,� principalmente,� del� tipo� de� suelo� y� de� la�
abundancia� de� herbívoros� productores� de� la� fuente� de� alimento� de� los�
escarabajos� en� estudio.� La� metodología� empleada� puede� resumirse� de� la�
siguiente�manera:�
�
8�2�4�2�#��*� �*!(!� =�$�)�"=&"*� &*=!+!$&$
�
� Las� bostas� de� vacas� y� caballos� recientemente� atacadas� por� escarabajos�
paracópridos,�se�identifican�debido�a�la�presencia�de�material�del�suelo�removido�
(en�forma�de�“rulitos”)�sobre�las�bostas�o�en�su�periferia.�Por�otro�lado,�en�todos�
los� casos,� la� bosta� escogida� por� estos� escarabajos� mostró� siempre� una�
consistencia� interior� más� bien� fresca� y� húmeda,� en� combinación� con� una�
cobertura� superficial� externa� de� pocos� milímetros,� y� de� apariencia� seca,�
formando�una�cáscara.���
� En�el�caso�de�las�especies�con�nidos�telecópridos,�el�trabajo�consistió�en�la�
identificación�del�tipo�de�bosta�escogida�por�estos�escarabajos�(por�ejemplo,�si�es�
fresca� o� vieja;� que� herbívoro� la� produciría;� etc.),� mediante� la� observación� de�
ejemplares� en� actividad� y� la� posterior� marcación� del� sitio� elegido� para� la�

� ���
nidificación.� En� algunos� casos,� se� capturaron� a� los� escarabajos� e�
independientemente� se� tomaron� muestras� de� bostas,� para� recrear�
posteriormente�su�actividad�en�condiciones�experimentales�particulares.�
�
8�2�4�4#�D*&+&*!> $� !$�".�D=�&**!> $�(&"&"E)�'&"$�*�F&
#�&=��!&' �(B'�&$�,� palas� de� distinto� tamaño,� cuchillos� grandes� (tipo�
de� cocina),� cortaplumas� (tipo� Victorinox),� cánulas� de� goma� de� diferentes�
diámetros,� pinceles� con� cerdas� de� distinta� dureza,� papel� higiénico,� algodón,�
papel� aluminio,� cinta� métrica,� cajas� y/o� envases,� y� material� entomológico� de�
campo.��
#��=�$�'��F&,Los� nidos� fueron� excavados� de� dos�maneras� diferentes,�
alternando�sus�usos�de�acuerdo�al�caso,�y�dependiendo,�particularmente,�de�las�
características�del�suelo.�Uno�de�los�métodos�consistió�en�realizar�una�excavación�
alrededor�y�por�debajo�de�la�bosta�marcada,�de�manera�tal�de�poder�extraer�el�
nido� contenido� en� una� porción� aislada� de� suelo� (“tocón”).� Antes� de� realizar� la�
excavación,�fue�necesario�el�reconocimiento�de�alguna�estructura�del�nido,�como�
por�ejemplo,�las�perforaciones�de�sección�circular�en�el�suelo�justo�por�debajo�de�
la�bosta�o�en�su�periferia,�que�indican�la�presencia�de�algún�túnel.�El�recorrido�y�
la�longitud�de�estos�túneles�fueron�seguidos�por�el�uso�cuidadoso�de�cánulas�que�
permitieron,�además,�estimar� la�profundidad�de� los�nidos.�Este� instrumento�fue�
de�uso�fundamental�en�todo�el�proceso�de�excavación,�ya�que�no�sólo�sirvió�de�
guía�a� lo� largo�del�nido,�sino�que�también�permitió� la� limpieza�de� los�túneles�y�
cámaras,� al� ser� usado� como� soplador.� Por� lo� general,� las� excavaciones� de� los�
“tocones”�fueron�cilíndricas,�y�alcanzaron�valores�de�25�a�30�cm�de�diámetro�en�
una� circunferencia� que� contenía� la� bosta� en� el� centro,� y� de� 40� a� 60� cm� de�

� ���
profundidad,�aproximadamente.�Una�vez�removido,�cada�“tocón”�fue�cortado�en�
capas� delgadas� usando� un� cuchillo� bien� afilado,� para� poder� reconocer� la�
arquitectura� del� nido� (túneles,� cámaras,� etc.)� y� poder� colectar� los� escarabajos�
adultos� y� las�masas/bolas� de� cría� construidas� por� ellos.� Este� proceso� permitió�
tomar�medidas�y�fotografías�de�las�distintas�partes�del�nido,�y�simultáneamente,�
permitió� esquematizarlas,� facilitando� así� su� posterior� reconstrucción� en� el�
laboratorio.�Esta�metodología�fue�aplicada,�fundamentalmente,�en�la�excavación�
de�los�nidos�de������� ���,����� �������y��������.�
Una�vez�reconocida�la�bosta�elegida�por�los�escarabajos,�la�otra�manera�de�
trabajar�fue�por�medio�de�la�excavación�de�calicatas.�En�este�caso,�al�igual�que�la�
metodología�ya�descripta,�fue�necesario�el�reconocimiento�previo�de�estructuras�
que� permitieran� confirmar� la� presencia� de� un� nido.� Una� vez� reconocida� una�
perforación,�se�colocó�delicadamente�la�cánula�para�poder�seguir�el�recorrido�y�la�
profundidad� del� túnel.� Con� la� cánula� en� dicho� lugar,� se� procedió� a� excavar�
paralelamente� al� túnel� hasta� alcanzar� el� primer� tope� de� la� cánula.� Luego� se�
realizó� una� excavación� horizontal� al� piso,� como� un� escalón,� del� cual� se� fue�
retirando� con� cuidado� el� material� del� suelo� con� una� palita� chica,� hasta�
reencontrar�el�recorrido�del�túnel�por�visualización�de�la�cánula.�Este�proceso�se�
repitió� hasta� localizar� las� masas/bolas� de� cría,� y/o� hasta� completar� la�
arquitectura� del� nido.� Esta� metodología� fue� aplicada� para� la� mayoría� de� las�
especies�estudiadas,�fundamentalmente,�en�La�Rioja�y�Formosa.�
�
8�2�4�5 # ���!"=�� $� '& &�G%!=�*=%�& $� '�" !$�" . $�'
*�(B��=&(!� =�$�'&"�"B�*!�"�"=%$!&$&"
�

� ���
� En�cada�excavación,�el�proceso�completo�fue�registrado�por�medio�del�uso�
de� cámaras� fotográficas� digitales,� filmadoras,� y� dibujos� acompañados� por�
medidas,� cuya� información� fue� usada� posteriormente� en� el� laboratorio� para� la�
reconstrucción�de�los�nidos,�como�así�también�para�el�análisis�y�descripción�del�
comportamiento�de�nidificación�de�los�escarabajos�coprófagos.�Las�anotaciones�y�
dibujos�de�campo�se�completaron�con�el�análisis�de�los�videos�digitalizados.��
�
8�2�4�8#��'�*=&$��"*&�&)&C�"
�
� Este� ítem� tuvo� como� fin� la� identificación� de� los� escarabajos� coprófagos�
como�registro�del�organismo�productor�de�determinado�tipo�de�nido�y�masa/bola�
de� cría.� Para�ello,� los� insectos� capturados�en� los�nidos�excavados�en�el� campo�
fueron� transportados� al� laboratorio� en� etanol� 75� %,� donde� luego� fueron�
preparados,�rotulados�y�montados�en�cajas�entomológicas�para�su�determinación�
y�posterior�conservación.�
�
8�2�5# �&)&C�$�'&)��&=��!�
8�2�5�2#�&=��!&'*�'�*=&$�
� El�material� de� estudio� extraído� del� campo,� tanto�masas� y� bolas� de� cría,�
como�así�también�diferentes�estructuras�de�los�nidos�(túneles,�cámaras,�bolas�de�
alimentación,� etc.)� fueron� depositados� en� la� colección� de� icnología� del� Museo�
Argentino�de�Ciencias�Naturales�(Buenos�Aires).�De�la�misma�colección�se�estudio�
material�que�Cabrera�Walsh�y�Gandolfo�(1996)�usaron�para�su�publicación�sobre�
la�nidificación�de�escarabajos�coprófagos�de�Argentina.���

� ���
�
8�2�5�4 # ��$!*!� �" . �)"��+&*!� �" $� (&"&"E)�'&" $� *�F& .
$�"*�!B*!� �"$�'�" !$�"
�
� Las� masas� y� bolas� de� cría� colectadas� fueron� cortadas� en� sección�
longitudinal� y/o� transversal� con� el� fin� de� observar� y� medir� los� caracteres�
internos.�Los�parámetros�generales�medidos�fueron:�ancho�y�largo�de�las�masas�
de�cría,�diámetro�ecuatorial�de�las�bolas�de�cría,�diámetro�de�la�cámara�ovígera,�
espesor� de� la� pared� de� la� bola� de� cría� y� espesor� de� la� pared� de� la� cámara�
ovígera.� En� algunos� casos� pudieron� registrarse� otros� caracteres,� tales� como� el�
canal�de�aireación�de�la�cámara�ovígera.�Los�ejemplares�que�no�fueron�cortados,�
fueron� incorporados� enteros� a� la� colección� como� ejemplos� de� su� morfología�
externa�y�su�forma�general.������
� Para� todas� las� descripciones,� se�usó� la� nomenclatura�de�masas/bolas� de�
cría� y� nidos� de� escarabajos� coprófagos� propuestas� por� Halffter� y� Matthews�
(1966)� y� Halffter� y� Edmonds� (1982),� y� se� definieron� dos� nuevos� términos,�
“cámara�de�cría”�y�“túnel�de�almacenamiento”.�El�primero�los�usamos�para�incluir�
tanto� a� las�masas� como� a� las� bolas� de� cría,� y� no� debe� ser� confundido� con� el�
término�“cámara�nido”,�que�hace�referencia�a�la�cámara�excavada�alrededor�de�
(o�en�donde�se�aloja)� la�o� las�bolas�de�cría.�El�segundo�término�es�usado�para�
hacer� referencia� a� los� túneles� aprovisionados� con� bosta� en� los� que� no� se�
encontraron�adultos�alimentándose�(túnel�de�alimentación),�lo�que�sugiere�que�la�
bosta� almacenada�podría� estar� destinada� tanto� a� ser� usada� como�provisión�de�
una�cámara�de�cría,�como�a�la�alimentación�de�los�adultos.��
�
�

� ���
8�2�5�5H�)"��+&*!> $�'$�"&���''�'&�+&'� �''&)��&=��!�
�
� Algunas� bolas� de� cría� colectadas� en� el� campo,� fueron�mantenidas� en� el�
laboratorio�en�condiciones�de�humedad�y�temperatura�controladas�para�facilitar�
el� desarrollo� de� las� crías� hasta� la� emergencia� del� nuevo� adulto.� El� objetivo�
principal�fue�el�de�detectar�posibles�caracteres�en�la�morfología�de� las�bolas�de�
cría� producidos� por� las� larvas� en� desarrollo,� como� fue� ya� observado� en� otras�
especies� de� escarabajos� coprófagos� (Claude� Joseph� 1929,� Klemperer� 1978,�
Barattini�y�Sáenz�1953),�o�generadas�por�el�adulto�al�emerger,�que�pudieran�ser�
usadas�para�su�comparación�con�las�estructuras�presentes�en�las�trazas�fósiles.�
� Las�bolas�de�cría�fueron�cortadas�en�sección�longitudinal,�pasando�el�corte�
por� el� agujero� de� emergencia� o� el� canal� de� aireación,� para� el� estudio� de� su�
morfología.� Se� encontró� a� la� larva� en� el� interior� de� diez� bolas� de� cría� de����
��� ���,�en�una�de���������y�en�una�de��������,�y�se�halló�un�huevo�en�la�
cámara� ovígera� de� una� bola� de� cría� de� ��� �������.� Todas� ellas� fueron�
seleccionadas� para� estudiar� el� desarrollo� de� las� larvas� en� condiciones� de�
laboratorio� controladas.� Luego� de� ser� fotografiadas,� las� dos� mitades� fueron�
ensambladas� y� sujetadas� por� una� envoltura� de� papel� higiénico� humedecido,� y�
colocadas�en�recipientes�plásticos�con�tapa�(o�de�tergopol,�para�bolas�de�cría�de�
mayor�tamaño)�conteniendo�tierra�levemente�humedecida.�El�telgopor,�a�su�vez,�
tiene�como�objetivo�aislar�a�las�bolas�de�cría�térmicamente.�El�estado�general�del�
interior�de� las�cámaras,�y�el�desarrollo�de� las� larvas,� fueron� revisados�una�vez�
por�semana,�abriendo�cada�una�de�las�bolas�de�cría�por�el�mismo�sitio�donde�ya�
habían�sido�cortadas.�Se�fotografiaron�y�anotaron�las�modificaciones�observadas�
en�cada�caso,�y�aquellas�bolas�con� larvas�o�pupas� todavía�en�desarrollo� fueron�
envueltas� con� nuevo� papel� humedecido� y� colocadas� en� el� recipiente�

� ��
correspondiente.�Esto�proceso�fue�repetido�en�cada�caso�hasta�la�emergencia�del�
nuevo�adulto,�o�en�su�defecto,�hasta�la�muerte�de�la�larva.�
� El� resto�de� las�bolas�colectadas�y� transportadas�al� laboratorio�no�usadas�
en� este� tratamiento,� fueron� incorporadas� a� la� colección� una� vez� que� fueron�
expuestas� al� calor� de� lamparas�para�que� perdieran� la� humedad�original� con� el�
objeto� de� evitar� el� desarrollo� de� hongos� y� de� esta� manera� favorecer� la�
conservacion� de� su� forma,� apariencia� y� morfología� externa� originales.� A� este�
material�se�le�extrajo,�previamente,�los�huevos�y�las�larvas�para�ser�conservados�
en�etanol�75�%.�
�
8�2�5�8#� "&.�"$� !$!I!*&*!> � �''&)��&=��!�@�� �'*&(B�B���
� *� $!*!� �"�DB��!(� =&'�"*� =��'&$&"
�
�� ������������������
� Cuatro�especímenes�de��������,�dos�hembras�y�dos�machos,�colectados�
en�Vera�Cruz� (La�Rioja)� fueron� colocados� sobre�bosta� de� caballo� de�un�día� de�
deposición,�en�un�ambiente�natural�(parques�internos�del�CRILAR),�pero�limitado�
por�una�caja�de�marcos�de�madera,�paredes�de�tul,�sin�base�y�de�60�cm�de�lado�
(base�x�altura)�(Fig.�4�A).�Este�tipo�de�estructura�permitió� la�entrada�del�sol,�y�
no�modificó�la�temperatura�ni�la�humedad�del�aire�en�su�interior�ni�en�el�suelo.�
� A�lo�largo�de�siete�días,�la�actividad�de�los�escarabajos�fue�controlada�dos�
veces� al� día,� una� vez� a� la� mañana� y� otra� a� la� noche.� La� bosta� colocada�
originalmente�no�fue�removida�ni�modificada�por�el�observador.�Se�registraron�y�
anotaron�caracteres�externos�típicos,�tales�como�la�presencia�de�agujeros�en� la�
bosta�y/o�en�la�tierra,�material�del�suelo�removido,�como�así�también�todo�otro�
registro� de� actividad.� El� séptimo� día� se� procedió� a� la� excavación� de� los� nidos�

� ���
mediante�calicatas�y�siguiendo�la�metodología�descripta�para�el�estudio���������de�
los�nidos.�
�
�� ������ �������������������������� ��
� Dos� ejemplares� de� �������� ! ���������"� ��� �� �����,� una�
hembra�y�un�macho,�que� fueron�encontrados�rolando�una�bola�de�provisión�en�
las� calles� de� Anillaco� (La� Rioja),� fueron� colocados� en� un� terrario� de� plástico�
cilíndrico,�de�15�cm�de�base�x�20�cm�de�alto,�colmado�hasta�los�15�cm�con�tierra�
del�lugar,�levemente�humedecida�(Fig.�4�B).�La�pareja�fue�colocada�en�el�terrario�
junto� con� la� bola� de� provisión� previamente� construida.� Durante� la�
experimentación�no�se� incorporó�ninguna� fuente�de�alimentación�y� la�tierra� fue�
mantenida�siempre�en�condiciones�moderadas�de�humedad.�La�actividad�de� los�
escarabajos,�como�así� también� los�cambios�observables�en� la�superficie,� fueron�
registrados�a�los�largo�de�5�días,�luego�de�los�cuales�la�tierra�fue�cuidadosamente�
removida�para�poder�observar�las�características�del�nido�y�poder�recuperar�a�los�
escarabajos�y�a�la�bola�de�cría�fabricada.�
��
�� ������ ����� ������ ������������
� En�dos�viajes�de�campo�diferentes�a�Anillaco�(el�primero�durante�el�mes�de�
enero�del�2007�y�el�segundo�durante�febrero�del�2008),�se�colocaron�cinco�y�dos�
ejemplares�de� ��������! �������"�������,� respectivamente,�en�bosta�
de� caballo� de� un� día� de� deposición,� en� condiciones� ambientales� naturales�
(parques�internos�del�CRILAR)�y�en�el�horario�de�la�máxima�actividad�registrada�
para�estos�escarabajos�en�observaciones�anteriores�(de�10�de�la�noche�a�6�de�la�
mañana)�(Fig.�4�C).�Por�medio�de�fotos�y�filmaciones,�se�registró�todo�el�proceso�
de�selección�de�la�bosta,�producción�y�transporte�de�la�bola�de�provisión,�hasta�

� ��
su�enterramiento.�Se�marcó�el�sitio�donde�fue�enterrada�y�luego�de�un�día,�en�la�
primera�oportunidad,�y�de�seis�días�en� la�segunda,�se�procedió�a� la�excavación�
mediante� calicatas.� En� la� primera� oportunidad,� el� escarabajo� y� la� bola� fueron�
removidos� del� sitio� y� colocados� en� un� terrario.� Las� caracteristicas� del� terrario�
empleado�y� la�metodología�aplicada,�coinciden�con�las�descriptas�anteriormente�
para� ����������� �� �����.��
� Por� otro� lado,� tres� ejemplares� de� �������� ! �������"� �������
colectados� en� Anillaco� durante� el� mes� de� febrero� del� 2008,� fueron� colocados�
sobre�bosta�de�caballo�de�un�día�de�deposición,�separados�(una�hembra�sola,�y�
una�pareja�en�el�otro)�en�dos�terrarios�de�acrílico�transparente�de�30�cm�de�lado�
(base�x�altura)�con�una�abertura�en�el�techo�cerrada�con�tul,�conteniendo�tierra�
extraída�del� lugar�y� ligeramente�humedecida� (15�cm�de�altura)� (Fig.�4�D�y�E).�
Del� terrario�con�dos� individuos,�el�macho� fue�retirado� rápidamente,�para�evitar�
que� su� actividad� entorpeciera� el� proceso� de� nidificación,� ya� que� no� mostró�
ninguna� colaboración� con� la� hembra.� Una� vez� por� día� a� lo� largo� de� 6� días� de�
experimentación,�se�realizaron�observaciones�y�anotaciones�del�comportamiento�
de� los�escarabajos�y�de� las�modificaciones�externas�de� la�bosta�y�de� la� tierra.�
Durante� dicho� periodo,� la� tierra� fue� mantenida� siempre� en� condiciones�
moderadas� de� humedad� y� la� bosta� fue� retirada� rápidamente,� luego� del�
enterramiento� de� los� escarabajos.� Cumplido� dicho� periodo� de� tiempo,� la� tierra�
fue�removida�con�una�pala�pequeña�en�capas�paralelas�a�las�paredes�del�terrario�
y�usando�como�guía�el�agujero�dejado�por�los�escarabajos�al�excavar,�con�el�fin�
de�reconocer�las�cámaras�nido�y�poder�extraer�de�ellas�las�posibles�bolas�de�cría�
producidas.��
� �
�

� ���
�������"B�2
� Once� especímenes� de� ������� sp.� colectados� en� Antinaco� (La� Rioja)�
fueron�colocados�sobre�bosta�de�vaca�de�un�día�de�deposición,�en�terrarios�con�
las� mismas� características� de� los� ya� mencionados� para� �������� sp.,�
conteniendo� tierra� extraída� del� lugar� (Fig.� 4� F).� A� lo� largo� de� 4� días� de�
experimentación,� el� terrario� fue� expuesto� a� luz� natural� a� partir� del�mediodía� y�
durante� aproximadamente� tres� horas,� horario� al� cual� se� había� registrado� la�
mayor�actividad�de�los�escarabajos�en�condiciones�naturales�(Fig.�4�F).�Durante�
dicho�tiempo�de�exposición,�la�bosta�se�cambió�sola�una�vez,�se�controló�siempre�
la�humedad�de�la�tierra,�y�se�registró�digitalmente�y�anotó�toda�actividad�de�los�
escarabajos.�El�último�día� se� removió� la� tierra� y� se� tamizó,�debido�al�pequeño�
tamaño�de�los�escarabajos,�para�recuperar�los�insectos�y�para�extraer�todas�las�
posibles�bolas�de�cría�fabricadas.�����
�

� ���
�
�!�%�&8��� Terrario� colocado�al� aire� libre�para� ser�usado� con��������;���� Terrarios�cilíndricos� usados� con� �� ��� �� �����;� ��� Trabajo� al� aire� libre� con� �� ������,��colaboradores:�Lic.�Marcelo�Krause�y�Nicolás�Fidalgo;��#���Terrario�de�acrílico�usado�con� �� ������,� colaborador:� Lic.� Marcelo� Krause;� ��� Terrarios� de� acrílico� expuestos� a�condiciones�naturales�de�luz�usados�con��������sp.��
�
�

� ���
8�4 # ���� J����� � ������ � ����� �� � ���������� ��
����������� �� ��� ����� ?����� @ ���� ��A@ ��������
������ �?�����������#���������������A
�
Los� especímenes� de� ������������ colectados� en� el� campo� fueron�
preparados� en� el� laboratorio� bajo� microscopio� estereoscópico,� usando�
instrumentos�manuales�y�neumáticos,�y�removidos�de�la�roca�de�caja,�o�de�sus�
restos.�Algunos�ejemplares�muy�fragmentados�fueron�pegados�con�cianocrilato�y�
reforzados� con� laca� a� la� piroxilina.� Posteriormente,� cada� ejemplar� de�
������������fue�seccionado�para�ser�examinado�y�medido,�como�asi�también,�
para� poder� identificar� en� ellos� evidencias� de� la� actividad� de� otros� organismos.�
Las� icnoespecies� correspondientes� fueron�determinadas� siguiendo�el� trabajo� de�
Laza�(2006)�y�la�actualización�icnotaxonómica�propuesta�aquí.�
Para� el� análisis� icnoestratigráfico,� todas� las� trazas� fósiles� colectadas� y�
todas� las� observaciones� tomadas� en� el� campo,� fueron� correlacionadas� con�
estacas�que�indican�niveles�estratigráficos�específicos�en�varios�perfiles�de�Gran�
Barranca�(Formación�Sarmiento)�(ver�Materiales�y�Métodos).�La�Tabla�1�muestra�
la� relación� que� existe� entre� las� estacas,� las� edades,� las� SALMAS� y� las�
subunidades�litoestratigráficas�de�la�Formación�Samiento�en�Gran�Barranca.��
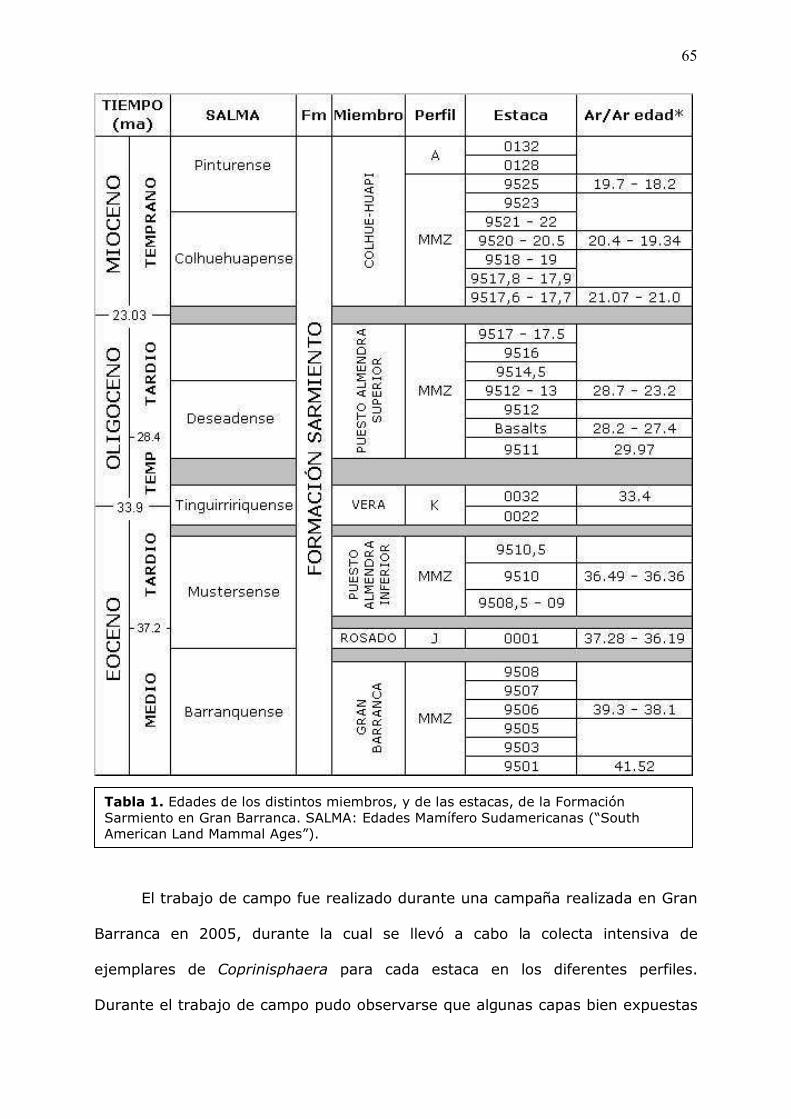
� ���
El�trabajo�de�campo�fue�realizado�durante�una�campaña�realizada�en�Gran�
Barranca� en� 2005,� durante� la� cual� se� llevó� a� cabo� la� colecta� intensiva� de�
ejemplares� de� ������������ para� cada� estaca� en� los� diferentes� perfiles.�
Durante�el�trabajo�de�campo�pudo�observarse�que�algunas�capas�bien�expuestas�
&)'&2�Edades�de�los�distintos�miembros,�y�de�las�estacas,�de�la�Formación�Sarmiento�en�Gran�Barranca.�SALMA:�Edades�Mamífero�Sudamericanas�(“South�American�Land�Mammal�Ages”).�

� ���
poseen�menos�ejemplares�que�otras�que�afloran�pobremente,�sugiriendo�que�el�
número� de� especímenes� colectados� (i.e.� abundancia� de� �����������)� es�
independiente� de� la� extensión� o� exposición� de� la� capa.� Sin� embargo,� esto�
probablemente�no�sea�válido�para�la�estaca�9516,�en�la�cual�la�poca�cantidad�de�
ejemplares�colectada�se�deba�principalmente�a�la�pobre�exposición�y�a�su�difícil�
acceso.� Otro� caso� particular� es� la� estaca� 9511,� donde� se� colectó� un� elevado�
número�de�especímenes�de������������,� pero� todos� ellos� con� evidencias� de�
retrabajo,�como�por�ejemplo,� las�superficies�pulidas.�Este�material� transportado�
no�fue�considerado�para�el�análisis�icnoestratigráfico.�Por�otro�lado,�en�el�perfil�M�
del� Miembro� Vera� (estacas� 0022� y� 0032)� se� registraron� sólo� dos� o� tres�
ejemplares�en�el�tope�que�no�pudieron�ser�colectados�debido�a�su�difícil�acceso,�
mientras� que� en� el� perfil� K� del� mismo� miembro,� donde� se� encuentra�
extensivamente� expuesto,� no� se� hallaron� ejemplares.� Por� ello,� y� para� los�
objetivos�de�este�análisis,�se�consideró�que�no�hay�registros�de�������������
en�el�Miembro�Vera.�Cabe�destacar�que�para�las�colectas�intensivas:�(1)�se�usó�
por� nivel,� el� trabajo� de� varios� colectores� durante� uno� o� más� días,� (2)� en� los�
casos� que� en� el� gráfico� no� aparecen� ejemplares� colectados� es� porque� no� se�
encontraron�ejemplares�expuestos�en�esos�niveles,�(3)�en�aquellos�niveles�donde�
el�número�está�por�debajo�de�100,�los�especímenes�colectados�fueron�todos�los�
expuestos,� y� (4)� en� aquellas� estacas� con� valor� de� 100� o� más,� la� colecta� fue�
detenida� arbitrariamente� después� de� alcanzar� o� pasar� dicho� número.� De� esta�
manera,�el�número�de�especímenes�colectado�es�usado�como�una�estimación�de�
la� abundancia� general� de� icnoespecies� de������������� presente� por� estaca.�
Además,�la�densidad�máxima�de�especímenes�para�muchas�estacas�fue�estimada�
en� el� campo� contando� los� especímenes� por� metro� cuadrado� junto� con� la�
distribución� uniforme� o� irregular� de� especímenes� a� lo� largo� del� nivel�
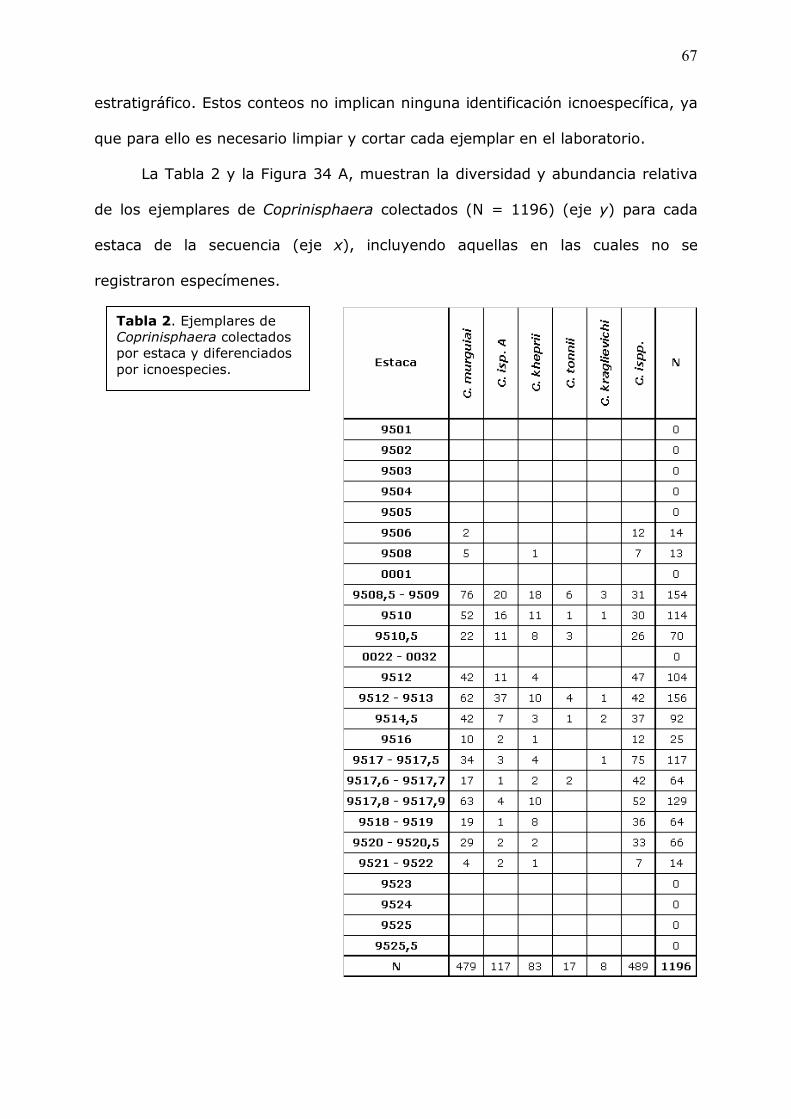
� ���
estratigráfico.�Estos�conteos�no�implican�ninguna�identificación�icnoespecífica,�ya�
que�para�ello�es�necesario�limpiar�y�cortar�cada�ejemplar�en�el�laboratorio.�
La�Tabla�2�y�la�Figura�34�A,�muestran�la�diversidad�y�abundancia�relativa�
de� los� ejemplares� de������������� colectados� (N�=� 1196)� (eje�&)� para� cada�
estaca� de� la� secuencia� (eje� +),� incluyendo� aquellas� en� las� cuales� no� se�
registraron�especímenes.��
�
&)'&4.�Ejemplares�de�������������colectados�por�estaca�y�diferenciados�por�icnoespecies.���

� ���
La�Tabla�3�(al�final�de�esta�sección)�muestra�el�diámetro�ecuatorial�(DE),�el�
espesor�de�la�pared�(EP),�y�el�índice�que�relaciona�a�estos�dos,�EP/DE�(IEP),�de�
todos� los� ejemplares� medidos� (DE,� N� =� 1065;� EP,� N� =� 982).� Las� posibles�
relaciones� existentes� entre� estos� parámetros,� y� que� fueron� analizadas,� están�
representadas� en� cuatro� gráficos� (Figs.� 35� a� 37).� La� Figura� 34� A� es� un�
histograma�de�barras�donde� las�barras� representan�el�número�de�especímenes�
colectado� por� estaca,� y� los� distintos� patrones� o� colores� dentro� de� cada� barra,�
representan�las�diferentes�icnoespecies.�En�este�gráfico�se�incluyeron�también�los�
especímenes�indeterminados�(��������).�El�histograma�de�la�Figura�34�B�muestra�
el�número�máximo�de�especímenes�colectado�por�m2�(densidad,�eje�&)�por�cada�
nivel�estratigráfico�(eje�+).��
La�Figura�35�A�muestra�el�DE�promedio� (eje�&),� como� representante�del�
tamaño�de�todos�los�especímenes�de�������������para�cada�estaca�(eje�+).�El�
DE� fue� medido� en� el� ecuador� de� las� bolas� de� cría� fósiles,� usando� como� polo�
superior�el�que�posee�el�agujero�de�emergencia�o� la�cámara�ovígera.�La�Figura�
35�B�muestra�el�mismo�parámetro,�pero�diferenciado�por�icnoespecie.�La�Figura�
36� muestra� para� cada� estaca,� un� histograma� de� barras� donde� cada� barra�
representa�el�número�total�de�especímenes�(diferenciado�por�icnoespecies)�de�un�
determindo� rango� de� DE� (de� I� a� V� clases� de� tamaño).� Estas� categorías� de�
tamaño,� fueron� definidas� considerando� que� cada� especie� de� escarabajo�
coprófago�construye�bolas�de�cría�con�un�rango�de�tamaño�del�30�%�(Lengerken,�
1954;�Klemperer�y�Boulton,�1976;�Klemperer,�1983;�Sato�y�Imamori,�1987).�De�
esta�manera,�el�menor�diámetro�ecuatorial�medido�(21�mm)�es�tomado�como�el�
menor�valor�de� la�primera�clase,�y�en�consecuencia,�el� límite�superior�de�dicha�
categoría� corresponde� a� 27,3� mm� (30� %� más).� Para� crear� el� resto� de� las�
categorías� se� usa� como� límite� inferior� el� valor� mayor� correspondiente� a� la�

� ��
categoría�anterior,�al�cual�se�le�suma�el�30�%�para�obtener�el�límite�superior�de�
la�nueva�clase.�El�mayor�valor�de�DE�medido�(66,4�mm),�determina�el�número�
total� de� categorías� (cinco),� quedando� incluído� dentro� de� la� última� clase� de�
tamaño� (60,5� –� 78,6� mm).� Siguiendo� esta� metodología,� se� obtuvieron� las�
siguientes� cinco� clases� de� tamaño:� (I)� 21�27,3� mm,� (II)� 27,4�35,6� mm,� (III)�
35,7�46,4�mm,�(IV)�46,5�60,4�mm,�y�(V)�60,5�78,6�mm.�En�resúmen,�teniendo�
en� cuenta� los� valores� máximo� y� mínimo� medidos� de� ED,� y� el� porcentaje� de�
dispersión�del�tamaño�de�las�bolas�de�cría�actuales�que�existe�por�cada�especie�
de�escarabajo�coprófago,�pueden�reconocerse�un�total�de�5�potenciales�especies�
productoras.�Para�cada�estaca,� la�presencia�y�abundancia�de�estos�productores�
se�encuentran�representadas�en�histogramas�individuales�de�la�Figura�36�(A�O).�
Sin� embargo,� la� potencial� diversidad�de�productores�no� sólo� está� representada�
por� el� tamaño� de� los� especímenes� de� �����������,� sino� también� por� la�
presencia�de�las�diferentes�icnoespecies.�Por�esta�razón,�en�los�histogramas�de�la�
Figura�36,�cada�icnoespecie�se�encuentra�diferenciada�por�un�determinado�patrón�
o�color�dentro�de�cada�barra.�Las�siguientes�icnoespecies�de������������,����
������� ��,����������,����isp.�A�y����������,�poseen�determinadas�características�
morfológicas� que� las� diferencian� entre� sí� y� que� son� interpretadas� aquí� como�
resultado�de�la�actividad�de�diferentes�productores;�mientras�que������������y�
��� ������ requieren� un� análisis� aparte.� Dado� que� ��� �������� se� encuentra�
representada� por� bolas� de� cría� fósiles� con� un� agujero� de� emergencia� sin� otra�
característica� distintiva,� especímenes� que� originalmente� hayan� tenido�
características� morfológicas� comparables� con� las� de� ��� isp.� A� o� ��� ������,�
podrían� estar� siendo� incluidos� dentro� de� esta� icnoespecie.� De� esta�manera,����
�������� no�puede�ser� considerada� como�una� icnoespecie�distintiva�de�un� sólo�
tipo� de� productor,� pero� sí� como� un� icnotaxón� para� bolas� de� cría� fósiles�

� ���
emergidas.�Lo�mismo�ocurre�con�los�ejemplares�incluidos�en���������,� los�cuales�
representan�especímenes� indeterminados�que�podrían�haber� sido�originalmente�
atribuídos� a� cualquiera� de� las� icnoespecies.� Entonces,� para� evaluar� el� número�
total�potencial�de�especies�de�escarabajos�coprófagos�productoras,�las�diferentes�
icnoespecies�fueron�diferenciadas�por�clases�de�tamaño,�con�la�excepción�de�����
�������� y� ��� �����,� que� fueron� sólo� consideradas� cuando� ninguna� otra�
icnoespecie�se�encontraba�representada�en�la�categoría.�La�diversidad�potencial�
de�productores�estimada�siguiendo�esta�metodología�debe�ser�considerada�como�
un�valor�mínimo,�ya�que�una�mayor�diversidad�podría�estar�escondida�en�bolas�
petenecientes�a�la�misma�icnoespecie�y�tamaño.��
La�Figura�37�A�muestra� los�cambios�del� índice� (IEP�=�EP/DE�x�200)�que�
relaciona�el�espesor�de�la�pared�(EP)�con�el�diámetro�ecuatorial�(DE)�(eje�&),�a�lo�
largo� de� la� seccuecia� estudiada� (eje� +).� La� Figura� 37� B� muestra� el� mismo�
parámetro,�pero�diferenciado�por�icnoespecies.�Este�índice�surge�como�resultado�
del�siguiente�razonamiento.�Una�pared�de�5�mm�de�espesor�podría� representar�
una�pared�muy�delgada�en�una�bola�de�cría�de�50�mm�de�diámetro,�pero�podría�
representar�también�una�pared�gruesa�en�una�bola�de�20�mm�de�diámetro.�Esto�
sugiere�que�el�espesor�de�la�pared�(EP)�no�es�un�dato�significacativo�����.�En�
cambio,� la� relación� entre� el� espesor� de� la� pared� y� el� diámetro� ecuatorial� es�
considerada�más�representativa.�La�relación�EP/DE�es�multiplicada�por�100�para�
evitar�las�fracciones,�y�por�2�debido�a�que�en�el�diámetro�ecuatorial�de�una�bola,�
el� espesor� de� la� pared� está� representado� dos� veces.�De� esta�manera,� el� valor�
resultante�del�índice�refleja�el�porcentaje�de�la�sección�de�una�bola�ocupado�por�
la�pared.�Es�decir,�un�valor�grande�de� IEP� representa�una�pared�gruesa.�Dado�
que�el�espesor�de�la�pared�es�irregular,�y�difiere�alrededor�de�la�bola,�el�espesor�
usado� como� valor� final� es� el� promedio� de� tres� valores,� de� los� cuales� dos� son�

� ��
medidos�en�el�ecuador�y�el�tercero�en�el�polo�opuesto�al�que�posee�el�agujero�de�
emergencia�o�la�cámara�ovígera.��
Los� valores� estimados� del� IEP� fueron� analizados� para� cada� potencial�
productor�reconocido�en�la�Figura�36.�El�objetivo�de�este�análisis�es�el�de�evaluar�
si� la�variación�en�el�grosor�de� la�pared� también�pudiera� resultar�de�diferencias�
comportamentales� que� reflejarían� la� presencia� de� más� productores� de� los� ya�
reconocidos�en�la�Figura�36.�La� idea�es�que�si�un�posible�productor� identificado�
en� la� Figura� 36�muestra� diferencias� significativas� en� este� índice� para� un� nivel�
estratigrafico� en� particular,� el� cual� indicaría� condiciones� paleoambientales�
similares,� entonces� las� diferencias� podrían� ser� atribuidas� al� comportamiento,� y�
por� consiguiente� a� un� productor� diferente.� En� algunos� pocos� casos,� pudo�
reconocerse� un� “gap”� entre� los� valores� de� IEP� dentro� de� una�misma� clase� de�
tamaño�de������������y����������en�las�estacas�9506,�9508,�9516�y�9517�17.5,�
sugiriendo� que� más� de� un� productor� estaría� involucrado.� Sin� embargo,� este�
análisis� no� puede� ser� sustentado� por� cálculos� estadísticos� debido� a� la�
fragmentación�de�la�muestra,�y�por�ello�no�está�incluido�detalladamente�aquí.�En�
resúmen,� no� se�pueden�agregar,� en�ningún� caso,� nuevos�productores� a� los� ya�
estimados�en�la�Figura�36.�
Las�medias�de�DE�y�el�IEP�fueron�analizadas�por�un�ANOVA�de�una�vía�y�
las�comparaciones�entre�estacas,�fueron�evaluadas�por�el�Test�de�Tukey�(Sokal�y�
Rohlf,� 1991).� Todos� los� análisis� estadísticos� fueron� llevados� a� cabo� con� el�
software�Statistica�7.0�(©�StatSoft,�Inc.),�usando�siempre�un�nivel�de�confianza�
del�95�%.
������

� ���
�

� ���
�
�
�

� ���
����
8�5#�������������������E���������@�����������@��������
� ����@��� ��������������������������������
Se� realizaron� cortes� delgados� de� al� menos� una� masa,� bola� de� cría� o�
cámara�pupal,�de�cada�una�de�las�especies�de�escarabajos�estudiadas,�como�así�
también� de� algunos� ejemplares� de� �����������,� �������� ����� ���� y�
��� ����� ���������,� � con� el� fin� de� estudiar� su� micromorfología.� Los� cortes�
delgados�obtenidos�a�partir�de�dichas�muestras�impregnadas�en�resina�poliéster�
(Murphy,�1986),�fueron�observados�con�microscopio�petrográfico�(Nikon�HFX�DX�
Optiphot�pol).�Los�distintos�caracteres�micromorfológicos�fueron�observados�con�
luz� transmitida,� mientras� que� el� iso� y� anisotropismo� y� la� birrefringencia� del�
material� fino,� fueron� observados� con� luz� polarizada.� Para� las� descripciones�
micromorfológicas,�se�usó�la�terminología�extraida�de�la�nomenclatura�propuesta�
por�Bullock������(1985)�para�micromorfología�de�suelos.��
�
&)'&5��Medidas� tomadas�a�cada�ejemplar�de������������,� agrupadas� por� miembro.� DE:�diámetro� ecuatorial;� EP:� espesor� de� la� pared;�IEP:�índice�espesor�de�la�pared.��

� ���
8�8#������������� ������� ��� ���,��������������� ��@�J ������@
��� ��������������
8�8�2#��'&"$�*�F&&*=%&'�"."%�'�*!�*% $& =�
Se� extrajeron� fitolitos� de� dos� porciones� de� suelo� muestreadas� en� dos�
ambientes�diferentes,�Navarro�(Buenos�Aires)�y�Chuquis�(La�Rioja),�durante� las�
distintas�campañas�ya�mencionadas�en�la�sección�8�2�de�esta�Tesis.�Del�material�
colectado�en�dichas� localidades,�se�seleccionaron�en�el� laboratorio�dos�bolas�de�
cría� actuales,� una� de� ellas� perteneciente� a� �������� ��� ���� y� la� otra� a�
��� ������� �������-�y�se�extrajeron�muestras�de� las�diferentes�partes�de�
estas�bolas�de�cría�(pared,�restos�de�bosta�de� la�provisión�original�conservados�
en�el�interior�de�las�bolas,�y�tapíz�de�la�cámara�pupal,�cuando�estaba�presente).�
Para�ello,�se�limpio�externamente�cada�una�de�las�bolas�de�cría�usando�un�pincel�
de�cerdas�blandas,�para�retirar�restos�de�material�del�suelo�no�pertenecientes�a�
la� pared� y� para� poder� tomar� una� muestra� de� la� misma� lo� más� pura� posible.�
Luego�fueron�seccionadas�con�sierra�de�mano,�por�el�eje�del�canal�de�aireación,�y�
se�tomaron�las�muestras�correspondientes�a�la�bosta�aprovisionada�en�el�interior�
y�al�tapíz�o�pared�de�la�cámara�pupal,�dependiendo�el�caso.�El�mismo�volúmen�
de�material�de� todas� las�muestras,� fue� tomado�con�una�cuchara�de� laboratorio�
colmada� de� 0,02�ml� de� capacidad,� y�montado� directamente� en� un� portaobjeto�
con� Bálsamo� de� Canadá� sintético.� Cada� preparado� fue� observado� luego� en� un�
microscopio�petrográfico�Nikon�Optiphot�pol.���
Se�extrajeron�también�fitolitos�del�interior�del�tracto�digestivo�de�una�larva�
de������� ����en�desarrollo�conservada�en�alcohol,�removida�de�una�bola�de�cría�
proveniente�de�Navarro.�Para�ello�se�disectó�la�larva�y�se�extrajo�el�contenido�del�

� ���
interior�del�tramo�final�del�mesenterón,�para�comprobar�la�presencia�y�en�dicho�
caso,�el�estado�de�preservación�de�los�fitolitos,�luego�del�proceso�de�digestión.�El�
material�extraído�fue�tomado�con�una�cuchara�de�laboratorio�colmada�de�0,02�ml�
de�capacidad,�montado�directamente�en�un�portaobjeto�con�Bálsamo�de�Canadá�
sintético�y�observado�en�microscopio�petrográfico�Nikon�Optiphot�pol,�como�con�
el�resto�de�los�preparados.�
Los� resultados� obtenidos� a� partir� de� estas�muestras,� serán�usados� en� la�
discusión� sobre� la� comparación� del� contenido� fitolítico� del� paleosuelo� y� de� la�
pared�de�los�ejemplares�de������������.��
�
8�8�4H��'&"$�*�F&I>"!'�".B&'��"%�'�*!�*% $& =�
�
Se� colectaron� uno� o� dos� especímenes� de� �����������,� incluídos� en�
muestras� respectivas� del� paleosuelo� circundante,� por� cada� uno� de� los� 14�
diferentes� niveles� estratigráficos� de� la� sección� más� completa� de� la� Formación�
Sarmiento,�en�Gran�Barranca.�Una�vez�transportados�al�laboratorio,�se�seleccionó�
para� este� análisis� un� ejemplar� por� cada� nivel� estratigráfico,� y� los� restantes�
fueron� guardados� en� la� colección� para� futuros� estudios.� Los� ejemplares� de�
������������ seleccionados� (MPEF�IC� 720�733),� fueron� cortados� en� dos�
mitadaes�y�preparados�para� la� identificación�de� las� icnoespecies.�En� la�mayoría�
de� los� casos,� cada� ejemplar� fue� dividido� en� tres� submuestras:� pared� externa�
(PE),� pared� interna� (PI),� y� relleno� (R).� La� pared� fue� dividida� en� dos�mitades,�
denominadas� interna�y�externa,�considerado�como� límite�entre�ellas�a� la�mitad�
del�espesor�total�de�la�pared.�En�algunos�casos,�donde�los�ejemplares�mostraron�
paredes�muy�delgadas,�fue�imposible�realizar�una�separación�en�dos�partes�como�
se� explicó� más� arriba,� por� ello� toda� la� pared� fue� utilizada� como� una� única�

� ���
muestra�y�se�la�denominó�simplemente�pared.�La�pared�externa�(PE)�es�la�parte�
de� la� pared� que,� en� la� bola� de� cría� original,� se� encontraba� en� contacto� con� el�
suelo�circundante,�mientras�que�la�pared�interna�(PI)�es�la�parte�de�la�pared�que�
contactaba� con� la�bosta�aprovisionada�en�el� interior�de� la�bola.�El�material� del�
relleno� usado� para� el� análisis� fue� removido� del� interior� de� la� cámara� principal�
dividida� en� las� dos�mitades� de� cada� ejemplar.� En� todos� los� casos,� se� usó� del�
paleosuelo� (P)� de� cada�muestra,� una�porción�que� estuviera� en� contacto� con� la�
bola� fósil,� debido�a�que�el�productor�habría�usado�esta�parte�del� suelo�original�
para� construir� la� pared� de� dicha� traza.� Se� descarta� de� esta� manera,� posibles�
resultados�confusos�introducidos�por�una�distribución�heterogénea�de�los�fitolitos�
en� el� suelo.� Finalmente,� para� la� extracción� de� fitolitos,� se� usaron� 10� g� de�
material�de�cada�una�de�estas�cuatro�muestras�(P,�PE,�PI,�y�R),�a�excepción�de�
unos�pocos�casos�donde�se�utilizó�menor�cantidad�debido�al�pequeño�tamaño�de�
los�ejemplares�de������������.��
Los� ejemplares� de������������� usados� para� la� extracción� de� fitolitos,�
pertenecen�a� tres�de� las�seis� icnoespecies.�De�un� total�de�14�especímenes,�10�
corresponden�a�����������-�uno�a����isp.�A,�otro�a�����������(Fig.�6�A,�G,�B,�H,�
C,� I,� respectivamente),�y� los�dos� restantes�no�pudieron�ser� identificados.�Cabe�
recordar,� que������������ es� interpretada� como� una� bola� de� cría� fósil� que� se�
encuentra� “abierta”� en� el� polo� superior� por� un� agujero� que� representa� la�
emergencia� del� nuevo� adulto� (Fig.� 6� A� y� G).� En� cambio,� las� otras� dos�
icnoespecies�han�sido� interpretadas�como�bolas�de�cria� fósiles�en� las�cuales�se�
preservan� restos� de� la� cámara� ovígera� pero� que� caracen� de� agujero� de�
emergencia,� lo� que� sugiere� que� el� adulto� nunca� emergío,� y� por� ello� son�
consideradas�como�bolas�“cerradas”.�Las�tres�icnoespecies�mencionadas,�poseen�
relleno�en�el�interior�de�la�cámara�principal,�el�cual�pudo�haber�sido�introducido�

� ���
por� la� actividad� del� adulto� al� emerger� en� el� caso� de� ����������,� o� debido� a�
procesos�pasivos,�en�las�otras�dos��
Los�fitolitos�fueron�extraídos�de�cada�una�de�las�cuatro�muestras�de�cada�
ejemplar� de� ������������ (pared� interna� y� externa,� relleno� y� paleosuelo)�
siguiendo� la� metodología� estandarizada� por� Zucol� y� Osterrieth� (2002).� Los�
procedimientos� llevados� a� cabo� en� el� laboratorio� pueden� resumirse� de� la�
siguiente�manera:�10�gr�de�material�seco�y�finamente�molido�fue�dispersado�en�
una� solución� de� hexametafosfato� de� sodio� (Na4P2O7)� (Fig.� 5� A).� Las� etapas�
siguientes,� indicadas� en� la� metodología� estandarizada,� de� disolución� de�
carbonatos�con�ácido�clorhídrico� (HCl),�y�de�oxidación�de�materia�orgánica�con�
agua� oxigenada� (H2O2),� no� fueron� realizados� debido� a� que� los� paleosuelos�
muestreados� no�muestran� cantidades� significativas� de� estos� dos� componentes,�
como�fue�previamente�reportado�para�esta� localidad�por�Bellosi�y�González�(en�
prensa).�Luego�de�la�dispersión,�se�pasó�el�material�por�dos�tamices�(Fig.�5�B),�el�
primero� de� 250� ^� y� el� siguiente� de� 63� ^,� obteníendose� así� tres� fracciones�
diferentes:�gruesa�FG�(mayor�a�250�^),�media�FM�(250�a�63�^),�y�fina�FF�(63�a�
3,9�^).�La�FG�se�caracteriza�por�poseer�muy�escasa�cantidad�de�fitolitos,�por�ello�
no�fue�usada�en�este�análisis�y�fue�reservada�para�futuros�trabajos,�mientras�que�
las� otras� dos� fracciones,� media� (FM)� y� fina� (FF),� ambas� ricas� en� contenido�
fitolítico,�fueron�usadas�para�la�extracción,�identificación�y�conteo.�La�FF�obtenida�
del�proceso�de�tamizado,�fue�expuesta�al�método�de�sedimentación�gravimétrica�
(Fig.� 5� C� y� D),� bajo� la� Ley� de� Stokes,� para� descartar� de� las� muestras� las�
partículas�menores�a�3,9�^,�antes�de�la�identificación�y�conteo�de�fitolitos.�Luego�
de�la�sedimentación,�el�material�retenido�en�las�probetas�es�filtrado�(Fig.�5�E)�y�
dejado� secar� antes� de� ser� usado� para� la� construcción� de� los� preparados.� Para�
estandarizar�el�conteo�de�fitolitos�en�los�preparados,�se�tomó�el�mismo�volumen�

� ��
de�material� de� las� fracciones�media� (250�63�^)� y� fina� (63�3,9�^)� de� todas� las�
muestras,�usando�para�ello�una�cuchara�de�laboratorio�de�0,02�ml�de�capacidad,�
colmada�de�material.�Una�vez�medida�la�cantidad�de�material�a�utilizar,�se�colocó�
en�el�portaobjeto�distribuyéndolo�los�más�homogeneamente�posible,�y�se�montó�
con�Bálsamo�de�Canadá�sintético�(Fig.�5F).�Finalmente,�se�colocó�el�cubreobjeto,�
se� selló,� se� dejó� secar,� y� se� observó� bajo� microscopio� petrográfico� (Nikon�
Optiphot�pol).��
�

� ���
�!�%�& 6�� Pasos� empleados� en� la� metodología� de� extracción� de� fitolitos� de������������.���Dispersión�del�material�en�una�solución�de�hexametafosfato�de�sodio�(Na4P2O7)� durante� 24hs.;� ��� Tamizado;� � . ��� Sedimentación� gravimétrica,�colaboradora:� Lic.� Mirta� González;� ��� Filtrado� y� secado� del� material� retenido� en� las�probetas;� ��� Elaboración� de� preparados� para� su� observación� bajo� microscopio�petrográfico.�����
En� todos� los� casos,� la� frecuencia� absoluta� de� fitolitos� fue� estimada�
contando� el� número� de� fitolitos� de� cada�morfotipo� presente� en� 10� puntos� del�
preparado� de� cada� muestra,� tomados� al� azar.� A� pesar� que� el� análisis� de� las�
diferencias� estadísticas� entre� estos� puntos� no� formó� parte� de� los� objetivos�
principales�de�este�estudio,�pudo�observarse�durante�los�conteos�que�los�valores�
fueron� similares� entre� los� distintos� puntos� de� un�mismo� preparado,� sugiriendo�
que�el�material�fue�homogéneamente�distribuído�durante�la�elaborarión�de�cada�
preparado.�Para�cada�punto�se�contaron�todos�los�fitolitos�incluidos�en�el�campo�
visual� del� microscopio� a� 400x� para� la� FF� y� a� 200x� para� la� FM.� Los� fitolitos�
pertenecientes�a�dos�clases�de�tamaño�de�partículas,�es�decir,�FF�y�FM,� fueron�
contados� de� manera� independiente,� siguiendo� el� procedimiento� usual.� Este�
conteo�independiente,�fue�aprovechado�como�réplica�de�los�resultados.�
Los� diferentes� morfotipos� de� fitolitos� fueron� identificados� siguiendo� las�
clasificaciones� propuestas� por� Bertoldi� de� Pomar� (1971),� Twiss� (1992)� y� Zucol�
(2001).� En� algunos� casos,� tipos� particulares� de� fitolitos� fueron� también�
identificados� por� comparación� con� morfotipos� fósiles� y� actuales� descriptos� e�
ilustrados�por�diferentes�autores�(Barboni�����,�1999;�Strömberg,�2002;�Gallego�
y� Distel,� 2004;� Zucol� y� Brea,� 2005;� Fernández� Honaine� �� ��,� 2006;� Piperno,�
2006).�����
Para� testear� las� hipótesis� que� se� postulan� en� esta� sección,� la� frecuencia�
absoluta� de� fitolitos� observada� por� muestra,� fue� utilizada� para� estimar� la�
frecuencia� esperada� en� cada� caso,� y� sus� distribuciones� fueron� comparadas�

� ��
estadísticamente� con� un� test� de� Bondad� de� ajuste� de� Chi�cuadrado� (Sokal� y�
Rohlf,�1991),�usando�para�ello�el�software�Statistica�7.0�(©�StatSoft�Inc.).�
�En�éste�análisis,�se�usó�como�variable�el�tipo�de�muestra�(P,�PE,�PI�y�R),�y�
se� postuló� como� hipótesis� nula� que� los� valores� obtenidos� (frecuencias�
observadas)�estan�uniformemente�distribuídos�entre�las�muestras.�Para�el�sexto�
objetivo� postulado� en� esta� Tesis,� la� frecuencia� absoluta� de� cada� categoría� de�
morfotipo� observada� por� muestra,� fue� utilizada� para� estimar� la� frecuencia�
esperada�en�cada�caso,�y�para�testear�si�estas�dos�variables�(tipo�de�muestra:�P,�
PE,�PI�y�R,�y�categoría�de�fitolito:�Gl:�globulolita,�Br:�braquiolita,�Pr:�prismatolita,�
O:�otros)�son�independientes�entre�sí�(hipótesis�nula).�Para�ello,�se�corrió�un�Test�
de�Independencia�de�Chi�cuadrado�con�el�software�Instat.�El�objetivo�de�correr�
este� test� fue� el� de� conocer� si� los� distintos� morfotipos� de� fitolitos� están�
representados�en�igual�proporción�en�las�muestras�estudiadas.�
Para�las�dos�últimas�hipótesis�postuladas�en�el�séptimo�objetivo,�y�usando�
los� resultados� obtenidos� del� análisis� anterior,� aquéllos� especímenes� que�
mostraron�diferencias� significativas� en� la� frecuencia� absoluta� de� los�morfotipos�
de� fitolitos� entre� la� pared� y� el� paleosuelo� (o� relleno),� fueron� usados� para�
comparar� estas� frecuencias� y� para� inferir� preferencias� en� la� dieta� de� los�
herbívoros� productores� de� la� bosta.� Estas� comparaciones� permitieron� conocer�
también� si� los� herbívoros� involucrados� preferían� los� tipos� de� plantas� (con�
fitolitos)�más�abundantes�en�el�ambiente�para�ese�momento,�o�si�escogían�otros�
tipos.��
�

� ���
6������ ���
6�2#����������� ��� ��
��
6�2�2���'&"$�*�!&I>"!'�"$��"*&�&)&C�"*�B�>I&��"
�
Se�examinó�un�total�de�1231�ejemplares�de������������,�de�los�cuales�
385� (������������ isp.)� no� pudieron� ser� identificados� a� nivel� icnoespecífico,�
fundamentalmente�por�estar�fragmentados,�y/o�por�no�presentar�clara�evidencia�
de� los� caracteres� diagnósticos.� De� este� material,� 1196� ejemplares� de�
������������ (Tabla� 2)� fueron� identificados� y� medidos� para� el� análisis�
icnoestratigráfico� que� se� muestra� en� la� sección�6�5� de� “Icnoestratigrafía”;� 14�
ejemplares�(MPEF�IC�720�733)�fueron�utilizados�para�la�extracción�de�fitolitos,�y�
mencionados� en� la� sección� 6�8� de� “Análisis� del� contenido� fitolítico”;� y,�
finalmente,�26�de�estos�ejemplares�(MPEF�IC�600,�634,�636,�638,�649,�665,�667,�
669,� 671,� 673,� 675,� 677,� 679,� 681,� 683,� 685,� 687,� 689,� y� 691,� y� MACN�Icn�
1373,� 1382,� 1392,� 1605,� 1620,� 2074� y� 2300)� presentaron� trazas� de� otros�
organismos�asociadas,�y�por�ello�son�mencionados�en�la�sección�6�2�4�de�“Trazas�
de�organismos�asociados�a������������”.��
El�material�examinado�está�depositado�en�las�colecciones�de�Icnología�del�
Museo�Paleontológico�Egidio�Feruglio�(Chubut,�Argentina)�(MPEF�IC),�del�Museo�
Argentino� de� Ciencias� Naturales� “Bernardino� Rivadavia”� �� Division� Icnología�
(Buenos� Aires,� Argentina)� (MACN�Icn),� del� Museo� Municipal� “Lucas� Roselli”�
(Nueva� Palmira,� Departamento� de� Colonia,� Uruguay)� (MLR)� y� del� Museo�
Municipal� de� Mar� del� Plata� “Lorenzo� Scaglia”� (Mar� del� Plata,� Buenos� Aires,�
Argentina)� (MMP).� Algunos� de� los� ejemplares� colectados� en� la� Formación�

� ���
Sarmiento,� localidad� Gran� Barranca� (Chubut,� Argentina),� se� encuentran�
depositados� temporalmente� a� préstamo� en� el� Museo� Argentino� de� Ciencias�
Naturales,� hasta� ser� ingresados� en� la� colección� de� Icnología� del� Museo�
Paleontológico� Egidio� Feruglio.� Cada� ejemplar� lleva� como� rótulo� un� número� de�
campo�(MPEF�Nº�de�campo).�
�Los� ejemplares� provenientes� de� la� Formación� Sarmiento� (Chubut,�
Argentina)�que�no�pudieron�ser�identificados�a�nivel�icnoespecifico�y�por�lo�tanto�
no�aparecen�mencionados�en�ninguna�de�las�secciones�correspondientes,�son�los�
siguientes:�Localidad�Gran�Barranca:�MACN�Icn�1305,�1366,�1372,�1373,�1382,�
1408,� 1410�1413,� 1416,� 1418,� 1441,� 1443,� 1445,� 1488,� 1491,� 1493,� 1589,�
1605,�1702,�2128,�2261,�MPEF�IC�665,�667,�669,�671,�673,�675,�677,�679,�681,�
683,�685,�687,�689,�691,�693�699,�723,�733,�MPEF�Nº�de�campo�1060,�1063,�
1065,� 1066,� 1069,� 1070,� 1072,� 1104�1109,� 1111,� 1113,� 1114,� 1116�1118�
(Miembro�Gran�Barranca),�MPEF�Nº�de�campo�17,�21,�29,�32,�41,�45,�46,�49,�51�
53,�56,�61�67,�69,�75�80,�82,�88,�92,�93,�100,�102,�104,�111,�116,�126,�129,�
146,� 147,� 149,� 153,� 156,� 158,� 162,� 167,� 169,� 170,� 171,� 174,� 181,� 184,� 185,�
187,�189�191,�302�304,�306,�308�311,�313,�314,�316�318,�446,�452,�470,�472,�
485,�496,�505,�508,�515,�516,�520,�522,�523,�526,�528,�531,�538,�547�(Miembro�
Puesto�Almendra�Inferior),�MPEF�Nº�de�campo�3,�4,�6,�10,�12,�13,�15,�193,�205,�
214,�216�218,�255�258,�319�321,�326,�402,�403,�592,�600,�602,�629,�659,�664,�
667,�671,�676,�688,�695,�704,�705,�714,�719,�856,�858,�861,�864�870,�874�876,�
879,�880,�887,�896,�954,�962,�964�967,�970,�971,�973�975,�977�980,�982,�984,�
985,� 989,� 990,� 992,� 993,� 995�1000,� 1093,� 1094,� 1125,� 1126,� 1128,� 1130,�
1134�1137,� 1139,� 1140,� 1143,� 1147,� 1149,� 1150,� 1152,� 1156,� 1162,� 1167,�
1170,� 1173,� 1179,� 1184,� 1187,� 1188,� 1193,� 1200�1202,� 1204�1206� (Miembro�
Puesto� Almendra� Superior),�MPEF�Nº� de� campo� 229,� 232,� 233,� 237�240,� 268,�

� ���
274,�279�281,�286,�287,�290,�292,�728�739,�741�746,�749,�751,�752,�759,�760,�
762,�767,�769,�773,�774,�777,�779,�780,�783,�787�800,�802�809,�811,�820,�822,�
825,�833�836,�838,�839,�904,�906,�909,�911,�915,�917,�920,�923,�925,�928�933,�
935,� 937�939,� 941,� 943,� 945,� 947,� 949,� 953,� 1003,� 1010,� 1016,� 1017,� 1019,�
1021,�1024,�1028,�1037,�1042,�1044,�1047,�1048,�1056,�1057,�1059�(Miembro�
Colhue�Huapi).�Localidad�Sierra�de�Talquino:�MACN�Icn�2074�y�MPEF�IC�700.��
�* �I&(!'!&��B�! !"BK&��!$&��� !"�4<<8
����������������&%��2766
�
������������ Sauer� 1955,� p.� 123� (icnoespecie� tipo� por� monotipia:� ������������
���������� Sauer);� 1962:� Häntzschel,� W189;� 1975:� Häntzschel,� W52;� 1982:�
Martinez,�p.�48;�1993:�Genise,�p.�50;�1994:�Donovan,�p.�209;�1994:�Hasiotis�����,�
fig.�149;�1995:�Genise�y�Cladera,�p.�78;�1998:�Buatois�����,�p.�226;�1998:�Genise�y�
Laza,�p.�220;�1998:�González�����,�p.�38;�1999:�Genise,�p.�110;�2000:�Buatois���
��;�2000a:�Genise,�p.�50;�2000c:�Genise,�p.�115;�2000:�Genise����.,�p.�49;�2000a:�
Krell,�p.�890;�2000:�Verde,�p.�112;�2001:�Bellosi�����,�p.�31;�2001:�Genise�����,�p.�
45;�2001:�Retallack,�p.�291;�2002:�De,�p.�65;�2002:�Hasiotis,�p.�79;�2003:�Genise,�p.�
19;�2003:�Mikulás�y�Genise,�p.�341;�2004:�Aramayo�����-�p.�13;�2004:�Bellosi,�p.�
19;� 2004:� Bellosi� y� Genise:� p.� 15;� 2004:� Bellosi� �� ��,� p.� 33;� 2004:� Buatois� y�
Mángano,�p.�312;�2004a:�Dieni�y�Genise,�p.�29;�2004b:�Dieni�y�Genise,�p.�31;�2004:�
Genise,�p.�421;�2004:�Genise�y�Cladera,�p.�632;�2004a:�Genise�����,�p.�366;�2004b:�
Genise� �� ��,� p.� 39;� 2004:�Genise� y� Bellosi,� p.� 41;� 2004:� Hasiotis,� p.� 184;� 2004:�
Hembree� y� Hasiotis,� p.� 63;� 2004:� Pieńkowski,� p.� 8;� 2005:� Bellosi� �� ��,� p.� 306;�
2005:�De,�p.�233;�2005:�Sánchez�����,�p.�120;�2006:�Chure�����,�p.�243;�2006:�
Genise�����,�p.�243;�2006:�Hasiotis,�p.�401;�2006:�Hasiotis�y�Bourke,�p.�217f;�2006:�
Laza,� p.� 220;� 2006:� Sánchez� �� ��,� p.� 20;� 2007:� Bromley� �� �.-� p.� 144;� 2007:�
Buatois� y�Mángano,� p.� 286;� 2007:�Duringer��� ��,� p.� 333;� 2007:� Ekdale��� ��,� p.�
570;� 2007:� Genise,� p.� 12;� 2007:� Hasiotis,� p.� 265;� 2007:� Hasiotis� �� ��,� p.� 174;�
2007:�Hunt�y�Lucas,�p.�59;�2007:�Krause�����,�p.�38;�2007a:�MacEachern�����,�p.�
54;�2007b:�MacEachern��� ��,� p.� 114;�2007:�Melchor��� �.,� p.� 16;� 2007:�Verde� y�
Genise,� p.� 40;� 2007:� Sánchez� �� ��,� p.� 41;� 2007:� Verde� �� ��,� p.� 342;� 2008:�
Comisión� Internacional� de�Nomenclatura�Zoológica,� p.� 318;� 2008:�Genise��� ��,� p.�

� ���
41;� 2008:�Krell,� p.� 39;� 2008:� Sánchez��� ��,� p.� 162;� 2009:� Sánchez� y�Genise,� p.�
837.�
“bolas�de�Cangagua”�Estrada�1941,�p.�433,�figs.�1,�2–7;�1950:�Sauer,�p.�344.�
�������Lengerken�1955,�p.�937�(especie�tipo�por�monotipia:�������� ����������
Sauer).�
�����������Sauer�1956,�p.�550;�1966:�Halffter�y�Matthews,�p.�154;�1991:�Retallack,�
p.�182;�2000b:�Duringer�����,�p.�264;�2006:�Laza,�p.�220�(lapsus�calami).�
“Nido�fósil�de�escarabajo”�Camacho�1966,�p.�488.�
“Paleonidos�de�escarabéidos”�Spalletti�y�Mazzoni�1977,�p.�265.�
“nidos�de�escarabajos�peloteros”�Fontaine����.�1995,�p.�12.��
“nódulos� [...]� como� producto� de� la� actividad� de� escarabajos� estercoleros”� Iriondo� y�
Kröhling�1996,�p.�43.�
(��� ���� Roselli� 1939,� p.� 81� (especie� tipo� por� monotipia:� (��� ���� ��������
Roselli;�preocupado�por�(��� ����Kraglievich�1932�(1994:�Genise�y�Bown,�p.�109).�
�������Roselli�1939,�p.�79�(especie� tipo�por�monotipia:��������������� ���Roselli);�
1998:�Genise�y�Laza,�p.�213;�1998:�González�����,�p.�39;�2000c:�Genise,�p.�115;�
2000:�Genise�����,�p.�49;�2000a:�Krell,�p.�891;�2000:�Verde,�p.�112;�2004:�Genise,�
p.�421;�2006:�Genise�����,�p.�243;�2006:�Laza,�p.�220;�2007:�Buatois�y�Mángano,�p.�
288;�2007a:�MacEachern�����,�p.�58.�
(�� ��� ����� Roselli� 1976,� p.� 167� (especie� tipo� por� monotipia:� (�� ��� �����
�������� Roselli);� 1982:� Martinez,� p.� 48;� 1998:� Buatois� �� ��,� p.� 226;� 1998:�
González�����,�p.�39;�2000a:�Genise,�p.�53.�
������ ����� Roselli� 1976,� p.� 167� (especie� tipo� por� monotipia:� ������ �����
������� ���Roselli);�1990:�Retallack,�p.�219;�1994:�Donovan,�p.�209;�1998:�Buatois�
����,�p.�226;�2006:�Genise�����,�p.�243.�
(�� ����Roselli�1987,�p.�21�(lapsus�calami).�
������ ����� Roselli� 1987,� p.� 22� (especie� tipo� por�monotipia:� ������ ����� ��� ����
Roselli);�1994:�Hasiotis�����;�1995:�Genise�y�Cladera,�p.�78;�1998:�Genise�y�Laza,�p.�
213;� 1998:� Buatois��� ��,� p.� 226;� 1998:� González��� ��,� p.� 39;� 1999:�Genise,� p.�
110;�2000a:�Genise,�p.�53;�2000:�Genise�����,�p.�54;�2000a:�Krell,�p.�891;�2000:�
Verde,�p.�112.�
���� ����� Roselli� 1987,� p.� 23� (especie� tipo� por�monotipia:� ���� ����� �������
Roselli);�1994:�Hasiotis�����;�1998:�Genise�y�Laza,�p.�213;�1998:�Buatois�����,�p.�
226;� 1998:� González� �� ��,� p.� 39;� 1999:� Genise,� p.� 110;� 2000a:� Genise,� p.� 53;�
2000:�Genise�����,�p.�54;�2000a:�Krell,�p.�891;�2000:�Verde,�p.�112.�
� ��� �� �����Roselli�1987,�p.�49�(especie�tipo�por�monotipia:� � ��� �� ��������� ���
Roselli);�1994:�Hasiotis�����;�1998:�Buatois�����,�p.�226;�1998:�Genise�y�Laza,�p.�

� ���
213;� 1998:� González� �� ��,� p.� 39;� 1999:� Genise,� p.� 110;� 2000a:� Genise,� p.� 53;�
2000:�Genise-�p.�54;�2000a:�Krell,�p.�891;�2000:�Verde,�p.�112.�
(������� ����� Genise� 1993,� p.� 50;� 1994:� Hasiotis� �� ��,� fig.� 148;� 1998:� Genise� y�
Laza,�p.�213.�
�
�* ��"B�*!� =!B�,� ������������ �������� (Roselli� 1939)� (non� Laza�
2006)�(=� ����������Sauer�1955)�
�!&� �"!":�Cámaras�esféricas,� subesféricas,�piriformes�y�biesféricas�que�
tienen�una�pared� construída,� la� cual�puede�estar� completamente�perforada�por�
un� agujero� de� tamaño�medio,� o� pueden�mostrar� una� cámara� secundaria,�más�
pequeña,�conectada�con�la�cámara�principal�por�medio�de�un�pasaje�estrecho.�La�
cámara� secundaria� puede� estar� también� conectada� con� el� exterior� a� través� de�
otro� agujero� de� tamaño� medio.� Las� cavidades� internas� contienen,� en� algunos�
casos,� rellenos� fundamentalmente� pasivos,� y� en� otros� casos� se� encuentran�
vacías.� Los� ejemplares� son� encontrados� aislados� y� en� contacto� con� la� roca� de�
caja,�o�dentro�de�una�cavidad�(Laza�2006).�
��(� =&�!�",�Recientemente,�Laza�(2006)�realizó�una�detallada�revisión�
de�este�icnogénero,�en�la�que�señala�que�si�bien�������������Sauer�1955�es�
un�sinónimo�junior�de��������Roselli�1939,�debido�al�extenso�uso�que�se�le�ha�
dado�al�primer�nombre�desde�su�creación,�lo�considera�allí�un�nombre�válido�para�
este�icnogénero,�como�adelanto�de�la�propuesta�hecha�por�Genise������(2006a)�
a� la� Comisión� Internacional� de� Nomenclatura� Zoológica.� En� ella� se� propuso,�
mantener�el�uso�del� nombre�������������a�pesar�de� ser�un� sinónimo� junior�
subjetivo�de�������,�para�el�cual�se�planteó�además�su�supresión�e�inclusión�en�
el� Índice� Oficial� de� Nombres� Genéricos� Rechazados� e� Inválidos� en� Zoología.�
Dicha�propuesta�se�apoyó�en�el�hecho�que�������������fue�usado�como�válido�
por� más� de� 25� autores� durante� los� últimos� 50� años,� y� que� es� además� el�

� ���
icnogénero� tipo� de� la� icnofamilia� �������������� Genise� 2004� y� de� la�
icnofacies� de� ������������ (Genise� �� ��,� 2000).� Krell� (2008)� apoyó� esta�
propuesta,�aportando�más�citas�del�icnogénero,�pero�sugiriendo�que��������no�
fuera� suprimido� sino� que� se�diera� precedencia� al� nombre������������� sobre�
este� último,� cuando� ambos� fueran� considerados� sinónimos.� Dicha� postura� fue�
avalada� por� Genise� �� ��� (2008).� Finalmente,� la� Comisión� Internacional� de�
Nomenclatura� Zoológica� (2008)� aceptó� la� precedencia� de� ����������� Sauer�
1955�sobre��������Roselli�1939.��
�
������������������������?��"�''!2757A
(Fig.�6�A�y�G)�
(Fig.�7�A�E,�G�M)�
�
“Bolas�de�escarabeidos”�Frenguelli�1938a,�p.�348.�
“Pallotole�di�Scarabeidi”�Frenguelli�1938b,�p.�77.�
“Nidos�fósiles�de�Escarabeidos”�Frenguelli�1939a,�p.�270.�
“Nidos�fósiles�de�Escarabeidos”�Frenguelli�1939b,�p.�379,�figs.�4�9,�pls.�I�II.�
“Nidos�fósiles�de�Escarabeidos”�Frenguelli�1940,�p.�70.��
(��� ������������Roselli�1939,�p.�81,�figs.�26�–�28,�31(5�6)�(non�Kraglievich�1932);�
1993:�Genise,�p.�50.�
“Nidos�de�escarabeidos”�Frenguelli�1941,�p.�87;�Laza�y�Reguero�1990:�245.��
“Bolas�de�Cangagua”�Bruet�1950,�p.��280,�pl.�I,�figs.�2,�3.�
������������ ����������Sauer�1955,�p.�123;�1956:�Sauer,�p.�555;�1959:�Sauer,�p.�
119;�1962:�Häntzschel,�W189;�1965:�Sauer,�p.�271;�1975:�Häntzschel,�W52;�1982:�
Martinez,�p.�48;�1994:�Genise�y�Bown,�p.�109;�1998:�Buatois�����,�p.�227;�1998:�
Genise�y�Laza,�p.�220;�1999:�Genise,�p.�110;�2000b:�Genise,�p.�28;�2000:�Genise���
��,�p.�54;�2000a:�Krell,�p.�890;�2001:�Retallack,�p.�142;�2002:�Buatois�����,�p.�189;�
2004:�Genise,�p.�429;�2005:�Bellosi�����,�p.�306;�2006:�Genise�����,�p.�243;�2006:�
Laza,�p.�217;�2007:�Krell,�p.�40;�2007:�Sánchez�����,�p.�41;�2008:�Sánchez�����,�
p.�162;�2009:�Sánchez�y�Genise,�p.�838.�
������� ����������Lengerken�1955,�p.�937.�
����������� ����������Sauer�1956,�p.�550.�

� ���
“Nidos�de�Scarabaeidae”�Halffter�1959,�p.�174.�
����������� ���������Halffter�y�Matthews�1966,�p.�154.�
“Nidos�de�escarabajos”�Andreis�1972,�p.�91;�1981:�Andreis,�p.�34.�
(�� ��� �������������Roselli�1976,�p.�167;�1982:�Martinez,�p.�48.�
(��� ��� �������������Roselli�1976,�p.�166;�1982:�Martinez,�p.�48;�1993:�Genise,�p.�
50;�1994:�Genise�y�Bown,�p.�109.��
������������ ����������Martinez�1982,�p.�48.�
“Icnofósiles�de�Scarabaeinae”�Laza�1986a,�p.�13.�
“Nidos�de�Scarabaeinae”�Laza�1986b,�p.�19.�
(�� ��� �������������Roselli�1987,�p.�21,�pl.�I,�f�.�
������ ����� ��� ���� Roselli� 1987,�p.� 22,� pl.� I,� figs.� 2,� 2a.� (syn�nov)(�����);�1995:�
Genise�y�Cladera,�p.�78;�2000a:�Krell,�p.�891;�2006:�Laza,�p.�227.�
���� ������������Roselli�1987,�p.�23;�2000a:�Krell,�p.�891;�2006:�Laza�,�p.�227.�
� ��� �� ��������� ���Roselli�1987,�p.�49,�fig.�8.�(syn�nov)�
(��� ��� ������������Retallack�1990,�p.�219.�
“Dung�beetle�trace�fossils”�Retallack�1990,�p.�436.
������������ ���������Genise�1993,�p.�50.�
����������������������Genise�y�Bown�1994,�p.�110,�figs.�3,�4.�1994:�Hasiotis�����,�
fig.�149;�1998:�Buatois�����,�p.�226;�2000a:�Genise,�p.�51;�2000a:�Krell,�p.�891;�
2006:�Laza,�p.�221.�
(������� ������������Donovan�1994,�p.�209.�
������������isp.�Genise�y�Cladera�1995,�p.�78,�figs.�1E,�F,�izquierda�y�derecha.�
�������������������� Roselli:� 1994:� Genise� y� Bown,� p.� 109;� 2000a:� Krell,� p.� 891;�
2000:� Krell,� p.� 890;� 2004:� Genise,� p.� 429;� 2005:� Sánchez� �� ��,� p.� 120;� 2006:�
Bellosi�����,�p.�306;�2006:�Laza,�p.�227;�2007:�Sánchez�����,�p.�41;�2008:�Sánchez�
����,�p.�162;�2009:�Sánchez�y�Genise,�p.�838.�
�
��'�=!B�,� Un� ejemplar� rotulado� como�(�� ���� �������� Roselli� 1939�
(figura� 26� A),� e� ilustrado� también� en� la� Lámina� I,� figura� 1,� de� su� trabajo� de�
1987,�depositado�en�el�Museo�“Lucas�Roselli”� (Roselli� col.,�MLR�479,�Fig.�7�A),�
Nueva�Palmira,�Dpto.�de�Colonia,�Uruguay,�proveniente�de�una�cantera�próxima�
al� Arroyo� de� Las� Flores� (Formación� Asencio)� en� la�misma� localidad.� Los� datos�
consignados� por� Roselli� en� sus� dos� trabajos� (1939,� 1987),� la� identificación�

� ��
posterior� de� Laza� (2006)� y� el� material� depositado� en� dicho� Museo,� no� dejan�
dudas�acerca�de�la�identidad�de�este�ejemplar�como�holotipo�de�la�icnoespecie.�
�&�&=!B�",Roselli�(1939)�menciona�tres�ejemplares�además�del�holotipo�e�
ilustra� y� describe� a� dos� de� ellos� (figuras� 27� y� 28).� En� su� trabajo� de� 1987,�
menciona� “paratipos”� de� 5� canteras� diferentes,� agregando� entonces�
erróneamente�2�ejemplares�más�con�esta�denominación.�Laza�(2006)�consideró�
erróneamente�a�estos�5�ejemplares,�2�de�ellos�no�contemplados�en�la�descripción�
original�como�paratipos,�y�le�adjudicó�letras�a�cada�uno�de�ellos.�Sin�embargo,�no�
especificó�en�su�contribución�cual�de�los�ejemplares�de�la�serie�típica�era�el�a,�ni�
los� rotuló� cuando� examinó� la� colección� de� Roselli,� por� lo� tanto� las� letras�
agregadas� por� Laza� (2006)� carecen� de� sustento.� En� la� colección� del� Museo�
“Lucas� Roselli”� se� conservan� actualmente� solo� 5� ejemplares� (Roselli� col.,� MLR�
479,� Fig.� 7� A�F),� contando� el� holotipo,� con� la� denominación� MLR� 479,� y� sin�
ningún� otro� dato� complementario� que� permita� discernir� cuales� eran� los�
ejemplares�originalmente�examinados�por�Roselli,�a�excepción�del�holotipo.���
De�los�tres�ejemplares�válidos�para�ser�considerados�como�paratipos,�solo�
dos� son� ilustrados� y� descriptos� en� el� trabajo� original� (Roselli,� 1939).� Uno� de�
estos�ejemplares�(Roselli,�1939,�fig.�28,�ejemplar�C)�pudo�ser�individualizado�en�
la� coleccción�del�Museo� “Lucas�Roselli”� (Fig.� 7� F),� y� es� considerado�aquí� como�
������������ ������� ��.� El� otro� ejemplar� ilustrado� (Roselli,� 1939,� fig.� 27,�
ejemplar�B)�es�más�dificil�de�reconocer�ya�que�ninguno�en�la�colección�se�ajusta�
estrictamente�al�dibujo,�que�pudo�haber�sido�algo� idealizado.�No�obstante,�uno�
de� los� ejemplares� (Fig.� 7� B� y� C)� que� presenta� actualmente� varias� fracturas�
pegadas,�coincide�con�la�breve�descripción�de�Roselli�y�una�de�sus�mitades�con�
su�dibujo,�por� lo�que�es� considerado�aquí� como�paratipo.�El� tercer�paratipo�no�
fue� descripto� ni� ilustrado� originalmente� (Roselli,� 1939),� y� en� el� Museo� se�

� ��
conservan� dos� ejemplares� asignables� a� ��� �������� rotulados� también� con� el�
número�479�(Fig.�7�D�y�E).�Al�no�poder�identificar�por�el�momento�a�ninguno�de�
los� dos� con� certeza� como� el� ejemplar� mencionado� por� Roselli� (1939)�
originalmente,� y� aun� cuando� ambos� son� efectivamente� ��� �������,� no� se�
considera�a�ninguno�de�ellos�como�paratipos.�
�&=��!&' �D&(! &$�,Holotipo� de� ���� ����� ������� (Roselli� col.,�
MLR�641,� Fig.� 7�G)�proveniente� de� la� cantera�próxima�al� Arroyo�de� Las� Flores�
(Formación� Asencio),� Nueva� Palmira,� Dpto.� de� Colonia,� Uruguay.� Holotipo� y�
paratipo�de� � ��� �� ��������� ���(Roselli�col.,�MLR�717�y�717a,�Fig.�7�H�e�I)�
provenientes� de� las� canteras� Maldonado� y� Chileno� (Formación� Asencio),�
respectivamente,� Nueva� Palmira,� Dpto.� de� Colonia,� Uruguay.� Holotipo� y� 2�
paratipos� de� ������ ����� ��� ���� (Roselli� col.,� MLR� 607,� Fig.� 7� J,� L� y� M)�
provenientes�de�la�cantera�Las�Flores�(Formación�Asencio),�Nueva�Palmira,�Dpto.�
de�Colonia,�Uruguay.�Un�tercer�ejemplar�(MLR�714)�ilustrado�aquí�en�la�Figura�7�
K,� no� comparte� el� número� de� catálogo� con� el� resto,� y� está� indicado�
erróneamente�como�paratipo�en�el�libro�del�Museo.�Algunos�de�estos�ejemplares�
fueron�identificados�con�una�letra�por�Laza�(2006),�pero�al�igual�que�con�el�resto�
del� material,� no� pudo� reconocerse� a� que� harían� mención� estas� letras� que� no�
aparecen� en� los� trabajos� de� Roselli� (1939,� 1987),� ni� en� el� catálogo,� ni� en� las�
etiquetas�del�material�depositado.�Un�ejemplar�(MACN�Icn�2300)�proveniente�de�
la� Formación� Sarmiento,� localidad� Sierra� de� Talquino� (Chubut,� Argentina).� Se�
examinó�un�total�de�490�ejemplares�provenientes�de�la�misma�formación,�pero�
colectados� en� la� localidad� Gran� Barranca� (Chubut,� Argentina).� Dicho�material,�
discriminado� por� miembros,� es� el� siguiente:� 9� ejemplares� (MACN�Icn� 1061,�
1062,� 1068,� 1112,� 1115,� MPEF�IC� 720,� 721,� MPEF�Nºcampo� 416,� 417)�
provenientes� del� Miembro� Gran� Barranca� (Eoceno� medio),� 151� ejemplares�

� �
(MACN�Icn�1346�1348,�1352,�1353,�1356,�1358�1365,�1368�1370,�1378,�1381,�
1383,�1386�1390,�MPEF�IC�722,�MPEF�Nº�de�campo�19,�20,�22,�24�26,�31,�33,�
34,�36�40,�42,�44,�47,�50,�54,�55,�59,�60,�73,�74,�84,�86,�91,�94�99,�113,�114,�
117�119,�125,�130,�131,�133,�134,�136,�137,�139�141,�186,�299�301,�307,�315,�
437�440,�443,�444,�447�450,�453,�455,�456,�458�460,�462�465,�468,�469,�471,�
473�76,�478,�487,�489,�492,�498,�499,�500,�501,�503,�504,�507,�511�514,�517,�
527,�529,�530,�533,�536,�537,�539,�540,�542,�543,�549�551,�553�565,�568,�569)�
provenientes� del� Miembro� Puesto� Almendra� Inferior� (Eoceno� medio�Eoceno�
tardío),�194�ejemplares�(MACN�Icn�1397,�1398,�MPEF�IC�724�727,�MPEF�Nº�de�
campo�1,�5,�7�9,�193,�194,�197,�198,�200�202,�204,�206�208,�213,�215,�221,�
252,�254,�294,�322,�323,�325,�574�576,�578,�580�582,�584,�585,�590,�591,�593�
597,� 599,� 633,� 634,� 637�640,� 645,� 648,� 649,� 651,� 654,� 656,� 657,� 660,� 662,�
666,� 669,� 670,� 673,� 679,� 681,� 683,� 685�687,� 689,� 696,� 699,� 701,� 702,� 707,�
709,� 711,� 713,� 716,� 854,� 857,� 859,� 860,� 862,� 863,� 871�873,� 877,� 878,� 881,�
882,�884�886,�888�892,�894,�895,�897,�898,�900,�955�961,�963,�968,�969,�972,�
981,� 983,� 986,� 987,� 991,� 1074,� 1075,� 1077�1082,� 1084�1092,� 1095,� 1096,�
1098�1103,� 1119�1121,� 1124,� 1129,� 1133,� 1138,� 1141,� 1142,� 1144,� 1145,�
1148,� 1151,� 1153�1155,� 1157�1160,� 1164�1166,� 1168,� 1169,� 1171,� 1172,�
1174�1177,� 1180�1182,� 1185,� 1186,� 1191,� 1192,� 1194,� 1195,� 1197�1199,�
1203)� provenientes� del� Miembro� Puesto� Almendra� Superior� (Oligoceno� medio�
temprano�Oligoceno�tardío),�136�ejemplares�(MACN�Icn�1342�1345,�1407,�1409,�
1413�1415,� 1440,� 1442,� 1444,� 1489,� 1490,� 1492,� MPEF�IC� 729,� 731,� 732,�
MPEF�IC�634,�MPEF�Nº�de�campo�223,�225,�226,�228,�230,�234�236,�267,�269,�
272,�275,�276,�278,�282,�284,�285,�288,�293,�723�727,�747,�753,�755�758,�763�
766,� 768,� 770�772,� 775,� 776,� 778,� 785,� 786,� 801,� 810,� 814,� 818,� 819,� 824,�
826�828,�831,�837,�840,�901�903,�905,�907,�908,�910,�912,�914,�916,�918,�919,�

� ��
921,� 922,� 927,� 936,� 942,� 944,� 946,� 948,� 950,� 951,� 952,� 1005�1009,� 1011,�
1014,�1015,�1018,�1020,�1022,�1023,�1025�1027,�1029�1036,�1038�1041,�1043,�
1045,�1046,�1049�1055,�1058)�provenientes�del�Miembro�Colhue�Huapi�(Mioceno�
temprano�Mioceno�medio).��
�!&� �"!" �(� $&$&,� Cámaras� aisladas,� esféricas� a� subesféricas,� con�
una�pared�construída,�relativamente�gruesa.�En�uno�de�los�polos,�la�pared�de�la�
cámara� está� completamente� perforada� por� un� agujero� de� tamaño� medio�
(alrededor�de�¼�del�diámetro�ecuatorial),�el�cual�posee,�en�sección�longitudinal,�
un�contorno�cilíndrico.�No�hay�restos�de�una�cámara�secundaria�o�de�estructuras�
adicionales� rodeando� el� agujero.� El� interior� de� la� cámara� posee,� como� norma,�
relleno�pasivo�(modificado�de�Laza�2006).�
��(� =&�!�",�durante�el�mes�de�enero�de�2009,�se�realizó�una�visita�al�
Museo� Lucas� Roselli� en� Nueva� Palmira� (Uruguay)� para� revisar� la� colección� de�
trazas�fósiles,�durante� la�cual�se�examinó�el�material�preservado�en�ese�Museo�
como�(��� ������������ (MLR� 479,� Fig.� 7� A�F).� Durante� dicha� revisión,� se�
observó�que�el�holotipo,�el�cual�fue�considerado�por�Laza�(2006)�como���������
Roselli,�comparte�sus�caracteres�con�la�icnoespecie������������� ����������
posteriormente� creada� por� Sauer� (1955).� Debido� a� ello,� ������������
���������� Sauer� 1955� pasa� a� ser� un� sinónimo� junior� subjetivo� nuevo� de�
��������������������(Roselli�1939),�la�cual�además�se�mantiene�sin�cambios�
como�icnoespecie�tipo�del�icnogénero.�
Es� posible� que� Laza� (2006)� le� haya� dado� importancia� diagnóstica� a� una�
segunda�perforación�más�pequeña�en�uno�de�los�ejemplares�de�esta�serie�(MLR�
479,� Fig.� 7� B� y� C),� la� cual� es� considerada� aquí� como� un� ejemplar� de�
�������� ����� �����,� es� decir,� la� entrada� o� salida� de� algún� otro� organismo�
invasor� (Mikuláš�y�Genise,�2003).�El�agujero�de�emergencia�de� tamaño�medio,�

� ��
diagnóstico�de�la�icnoespecie,�se�encuentra�fragmentado�en�este�ejemplar,�por�lo�
que�tal�vez�Laza�(2006)�no�lo�haya�reconocido,�separando�asi���������Roselli�de�
���������� Sauer.� Laza� (2006)� además� incluyó� en������������ (Laza� 2006,�
non�Roselli�1939)�una�serie�de�ejemplares�de�Patagonia�con�rasgos�distintivos�y�
fácilmente�separables�de������������(Roselli,�1939),�a�los�cuales�se�les�da�aquí�
el�nombre�de�isp.�A.�
De�la�misma�revisión�del�material�surge�que� ������ �������� ����Roselli�
1987,� considerado�por� Laza� (2006)� como������������ (Laza�non�Roselli� 1939),�
incluye�en�su�serie�típica�bajo�el�numero�de�catálogo�MLR�607�cuatro�ejemplares�
(Fig.� 7� J,� L�N),� de� los� cuales� tres� de� ellos� en� realidad� poseen� agujero� de�
emergencia�de� tamaño�medio,�por� lo�que� son� incluidos�aquí� como������������
(Roselli,� 1939)� (Fig.� 7� J,� L� y�M).� El� restante� ejemplar� es� considerado� como����
������� ��� (Roselli,� 1939)� (Fig.� 7�N).�A� diferencia� del� caso�de������������ no�
existe�en�el�trabajo�de�Roselli�(1987)�ninguna�indicación�que�permita�identificar�
un�holotipo�de�esta�serie,�por�lo�que�se�realiza�la�sinonimia�sobre�la�base�de�los�
tres�ejemplares�de�la�serie�típica,�cuyos�ejemplares�comparten�la�totalidad�de�los�
caracteres.� Las� medidas� obtenidas� de� estos� cuatro� ejemplares� (35� mm,� 35,5�
mm,� 35,6� mm,� 40� mm)� tampoco� coinciden� con� la� proporcionada� por� Roselli�
(1987)�(26�mm)�para�el�holotipo.� � ��� �� ��������� ���Roselli�1987,�también�
considerado� por� Laza� (2006)� como���� �������� (Laza� non� Roselli� 1939),� está�
basado� sobre� un� holotipo� (MLR� 717,� Fig.� 7� H)� que� presenta� un� agujero� de�
emergencia,� por� lo� que� esta� icnoespecie� es� considerada� aquí� como� sinónimo�
junior�de������������(Roselli,�1939).�El�paratipo�mencionado�por�Roselli�(1987)�
(MLR�717a)�se�encontró�en�la�colección�mal�rotulado,�como�734�(Fig.�7�I),�el�cual�
pudo� ser� identificado� por� comparación� con� el� ejemplar� representado� junto� al�
holotipo�en�la�Lámina�I,�figura�8,�de�su�trabajo�de�1987.��

� ��
��"*�!B*!> ,el�holotipo�(MLR�479,�Fig.�7�A)�de������������mide�46�mm�
de� diámetro� ecuatorial,� 42� mm� de� alto,� tomado� en� el� eje� del� agujero� de�
emergencia,�el�cual�posee�13�mm�de�diámetro,�y�el�espesor�de�la�pared�es�de�5�a�
7� mm.� El� paratipo� (MLR� 479,� Fig.� B� y� C),� y� los� dos� ejemplares� también�
catalogados�como�479�pero�que�no�tendrían�status�de�paratipos�(Fig.�7�D�y�E),�
miden�37,7�mm,�40�mm�y�42,2�mm�de�diámetro�ecuatorial,�24�mm,�38,1�mm�y�
37,8�mm�de�alto�en� el� eje�del� agujero�de�emergencia,� el� cual�mide�10,3�mm,�
12,2� mm� y� 13�mm� de� diámetro,� respectivamente,� y� la� pared,� que� solo� pudo�
medirse�en�dos�ejemplares,�es�de�5�mm�y�6,3�mm�de�espesor.�El�paratipo�(Fig.�7�
B�y�C)�y�otro�de�los�ejemplares�(Fig.�7�D)�se�encuentran�fragmentados,�e�incluso�
uno�de�ellos�posee�en�la�pared�un�ejemplar�de��������� �����������(Fig.�7�C).�
El� holotipo� de� ���� ����� ������� (MLR� 641,� Fig.� 7�G)�mide� 36,6�mm�de�
diámetro�ecuatorial,�30�mm�de�alto�en�el�eje�del�agujero�de�emergencia,�el�cual�
mide�13�mm�de�diámetro.�El�Holotipo�y�el�paratipo�de� � ��� �� ����� ���� ���
(MLR� 717� y� 717a,� Fig.� 7� H� e� I)� miden� 25,4� mm� y� 22,3� mm� de� diámetro�
ecuatorial,�25,5�mm�y�20�mm�de�alto,�y�poseen�un�agujero�de�emergencia�de�5,3�
mm� y� 6,6� mm� de� diámetro,� respectivamente.� Los� tres� ejemplares� de�
������ �������� ���,�rotulados�como�MLR�607�e�incluidos�aquí�en������������
(Roselli,�1939)�(Fig.�7�J,�L�y�M),�miden�35�mm,�35,5�mm�y�40�mm�de�diámetro�
ecuatorial,� 32,3� mm,� 34,5� mm� y� 39,5� mm� de� alto� en� el� eje� del� agujero� de�
emergencia,� el� cual� mide� 7,8� mm,� 7,7� mm� y� 6,6� mm� de� diámetro,�
respectivamente.�En�ninguno�de�ellos�se�pudo�medir�el�espesor�de�la�pared�por�
ser�ejemplares�completos,�no�cortados�ni� fragmentados.�El�ejemplar�(MLR�714)�
ilustrado� aquí� en� la� Figura� 7� K,� considerado� erróneamente� como� paratipo� de�
������ ����� ��� ���,�mide�30�mm�de�diámetro�ecuatorial,�37,8�mm�de�alto,�
con�posee�un�agujero�de�emergencia�de�7�mm�de�diámetro�y�un�espesor�de� la�

� ��
pared�de�7,2�mm.�Las�dimensiones�y�caracteres�morfológicos�pertenecientes�al�
resto�del�material�estudiado�se�encuentran�descriptos�en�diferentes�secciones�de�
los�resultados�de�esta�Tesis.�
���������������!"B� �+
(Fig.�6�B�y�H)�
�
�������������������� Laza� (non� Roselli� 1939)� 2006,� p.� 217,� figs.� 3B,� 5E,� Table� 2;�
2007:�Krell,�p.�41;�2007:�Sánchez�����,�p.�41;�2008:�Sánchez�����,�p.�162;�2009:�
Sánchez�y�Genise,�p.�838.��
����������������������
��'�=!B�,� Un� ejemplar� (MACN�Icn� 2058)� examinado� e� incluido� por� Laza�
(2006)� en� ��� �������� (Roselli,� 1939),� depositado� en� la� Divisón� Icnología� del�
Museo� Argentino� de� Ciencias� Naturales� (Buenos� Aires,� Argentina),� proveniente�
del� Miembro� Puesto� Almendra� de� la� Formación� Sarmiento� (Eoceno� medio�
Oligoceno�tardío)�de�Sierra�de�Talquino,�Chubut,�Argentina.��
�&�&=!B�",Dos�ejemplares�(MPEF�Nº�de�campo�577�y�641)�provenientes�
del� Miembro� Puesto� Almendra� Inferior� (Eoceno� medio�Eoceno� tardío)� de� la�
Formación�Sarmiento�en�Gran�Barranca,�Chubut,�Argentina.�
�&=��!&' �D&(! &$�,� Un� ejemplar� (MACN�Icn� 2392)� proveniente� de� la�
Formación� Sarmiento,� localidad� Sierra� de� Talquino� (Chubut,� Argentina).� Se�
examinó�un�total�de�124�ejemplares�provenientes�de�la�misma�formación,�pero�
colectados� en� la� localidad� Gran� Barranca� (Chubut,� Argentina).� Dicho�material,�
discriminado� por� miembros,� es� el� siguiente:� 48� ejemplares� (MACN�Icn� 1349,�
1357,�1367,�1384,�MPEF�IC�636,�MPEF�Nº�de�campo�23,�28,�30,�35,�48,�57,�58,�
83,�85,�87,�89,�90,�103,�105,�109,�120�122,�124,�127,�128,�132,�142,�305,�441,�
445,� 451,� 454,� 461,� 466,� 480,� 483,� 486,� 488,� 493,� 494,� 524,� 534,� 544,� 548,�

� ��
566,� 567)� provenientes� del�Miembro� Puesto� Almendra� Inferior� (Eoceno�medio�
Eoceno� tardío),� 62� ejemplares� (MACN�Icn� 1620,�MPEF�IC� 600,� 638,� 649,� 728,�
MPEF�Nº�de� campo�2,�196,�199,�203,� 571�573,�579,�583,� 587�589,� 628,�630�
632,� 636,� 643,� 644,� 647,� 650,� 652,� 655,� 661,� 663,� 665,� 668,� 674,� 675,� 677,�
680,� 682,� 684,� 693,� 698,� 706,� 708,� 712,� 715,� 717,� 718,� 855,� 883,� 893,� 899.�
988,�1083,�1097,�1122,�1123,�1127,�1178,�1183,�1189,�1196)�provenientes�del�
Miembro� Puesto� Almendra� Superior� (Oligoceno� medio� temprano�Oligoceno�
tardío),�10�ejemplares�(MPEF�Nº�de�campo�224,�227,�231,�291,�754,�830,�832,�
924,� 934,� 1013)� provenientes� del� Miembro� Colhue�Huapi� (Mioceno� temprano�
Mioceno�medio).��
�!&� �"!" �(� $&$&, Cámaras� aisladas,� esféricas� a� subesféricas,� con�
una�pared�construída,�relativamente�gruesa.�En�uno�de�los�polos,�la�pared�de�la�
cámara� incluye� una� cavidad� o� cámara� secundaria,� hemiesférica� y� pequeña,�
abierta�al�exterior.�Un�pasaje�muy�estrecho�en�el�piso�de�esta�cámara,�la�conecta�
con� la� cámara�principal.� El� interior� de� esta�última�posee,� como�norma,� relleno�
pasivo�(modificado�de�Laza�2006).�
��(� =&�!�",�como�fue�mencionado�previamente�durante la�revisión�del�
material� de� Roselli� (enero� 2009),� se� concluyó� que� � ��� �� ����� ���� ���
Roselli�1987�y� ������ �������� ����Roselli�1987,�considerados�por�Laza�(2006)�
como� ��� �������� (Laza� non� Roselli� 1939),� son� sinónimos� junior� subjetivos�
nuevos� de� ��� �������� (Roselli� 1939),� por� ello� ninguno� de� sus� nombres�
icnoespecíficos�esta�disponible�para�nominar�a�la�nueva�icnoespecie.���
��"*�!B*!> ,� El� holotipo� (MACN�Icn� 2058)� de������������� isp.� A,� de�
4,2� cm� de� diámetro� ecuatorial,� posee� una� cámara� hemiesférica� incluída� en� la�
pared,�de�1,1�cm�de�diámetro,�la�cual�presenta�en�la�base�una�perforación�de�4,5�
mm�de�diámetro.�La�pared�mide�de�6,3�mm�a�8,6�mm�de�espesor.�Los�paratipos�

� ��
(MPEF�Nº� de� campo� 577� y� 641)� miden� 31,6� mm� y� 36,1� mm� de� diámetro�
ecuatorial,�y�la�cámara�hemiesférica�mide�11,6�mm�y�11,2�mm�de�diámetro,�con�
una�perforación�en�la�base�de�6,4�mm�y�2�mm�de�diámetro,�respectivamente.�El�
segundo�ejemplar�no�fue�cortado,�para�conservar�un�ejemplar�entero,�por�lo�que�
el�espesor�de�la�pared�solo�fue�medido�en�el�primero,�el�cual�es�de�5,5�mm�a�6,5�
mm.� Las�medidas� y� otros� caracteres�morfológicos� del� resto� de� los� ejemplares�
estudiados�se�encuentran�detallados�en�diferentes�secciones�de�los�resultados.�
������������������������&L&4<<3
(Fig.�6�C�e�I)�
�
�������������������Laza�2006,�p.�229;�2007:�Krell,�p.�40;�2007:�Sánchez�����,�p.�
41;�2008:�Sánchez�����,�p.�162.�
��'�=!B�, Un� ejemplar� (MACN�Icn� 1670)� depositado� en� la� División�
Icnología� del�Museo�Argentino� de� Ciencias� Naturales,� proveniente� del�Miembro�
Colhue�Huapi� de� la� Formación� Sarmiento� (Mioceno� inferior)� de� Gran� Barranca,�
Chubut,�Argentina.�
�&�&=!B�",Dos� ejemplares� (MACN�Icn� 1720� y� 1724)� depositados� en� la�
División� Icnología� del� Museo� Argentino� de� Ciencias� Naturales,� proveniente� del�
Miembro� Colhue�Huapi� de� la� Formación� Sarmiento� (Mioceno� inferior)� de� Gran�
Barranca,�Chubut,�Argentina.
�&=��!&' �D&(! &$�, Se� examinó� un� total� de� 84� ejemplares,�
provenientes� de� la� Formación� Sarmiento,� localidad� Gran� Barranca� (Chubut,�
Argentina),� los� cuales,� discriminados� por� miembros,� son� los� siguientes:� 1�
ejemplar� (MPEF�Nº� de� campo� 1064)� proveniente� del� Miembro� Gran� Barranca�
(Eoceno� medio),� 37� ejemplares� (MACN�Icn� 1354,� 1355,� 1377,� 1380,� 1385,�

� ��
MPEF�Nº�de�campo�18,�27,�58,�71,�72,�81,�106�108,�110,�112,�135,�138,�144,�
312,�467,�477,�482,�490,�502,�506,�510,�518bis,�519,�521,�532,�535,�541,�545,�
552)�provenientes�del�Miembro�Puesto�Almendra�Inferior�(Eoceno�medio�Eoceno�
tardío),� 22� ejemplares� (MPEF�Nº� de� campo� 11,� 14,� 195,� 253,� 295,� 296,� 324,�
586,�598,�601,�672,�690,�691,�694,�700,�703,�710,�720,�976,�1161,�1076,�1190)�
provenientes� del� Miembro� Puesto� Almendra� Superior� (Oligoceno� medio�
temprano�Oligoceno� tardío),� 24� ejemplares� (MACN�Icn� 1340,� 1417,� MPEF�IC�
730,�MPEF�Nº�de�campo�241,�266,�270,�271,�276bis,�277,�289,�740,�748,�750,�
761,� 784,� 815�817,� 821,� 823,� 913,� 1004,� 1035)� provenientes� del� Miembro�
Colhue�Huapi�(Mioceno�temprano�Mioceno�medio).�
�!&� �"!" �(� $&$&, Cámaras� aisladas,� subesféricas� a� piriformes,�
mostrando� una� protuberancia� o� mamelón,� con� una� pared� construída,�
relativamente� gruesa.� Un� agujero� de� tamaño�medio� perfora� completamente� la�
protuberancia,�comunicando�el�interior�de�la�cámara�principal�con�el�exterior.�La�
pared� de� la� cámara,� en� el� piso� de� la� protuberancia,� incluye� una� cavidad�
hemiesférica�y�pequeña,�que�se�abre�hacia�el�interior�de�la�cámara�principal.�La�
cámara�principal�posee,�como�norma,�relleno�pasivo�(modificado�de�Laza�2006).
��"*�!B*!> , El� holotipo� y� los� paratipos� de� ��� ������� ya� han� sido�
descriptos�en�detalle�por�Laza�(2006),�motivo�por�el�cual�no�serán�repetidos�aquí.�
Las� medidas� y� otros� caracteres� morfológicos� del� resto� de� los� ejemplares�
estudiados�se�encuentran�detallados�en�diferentes�secciones�de�los�resultados.�
������������������� �� ����?��"�''!@2757A
(Fig.�6�D�y�J)�
(Fig.�7�F,�N�Q)��
�

� �
�������������� ���Roselli�1939,�p.�79;�1993:�Genise,�p.�53;�1994:�Genise�y�Bown,�p.�
112;�2000a:�Genise,�p.�51;�2000a:�Krell,�p.�891;�2004:�Genise,�p.�427;�2006:�Genise�
����,�p.�243.�
������ ������������ ���Roselli�1976,�p.�167;�1982:�Martínez�,�p.�58;�1987:�Roselli,�p.�
34;�1990:�Retallack,�p.�219;�1994:�Donovan,�p.�209.�
������ �������� ����Roselli�1987,�p.�22,�pl.�I,�figs.�2,�2a.�(syn�nov)�(�����).�
�������������� ���Roselli:�1993:�Genise,�p.�53;�1994:�Genise�y�Bown,�p.�112;�2000:�
Genise�����,�p.�54;�2000:�Krell,�p.�891;�2004:�Genise,�p.�426.�
������������������� ���Laza�2006,�p.�228;�2007:�Krell,�p.�40;�2007:�Sánchez�����,�
p.�41;�2008:�Sánchez�����,�p.�162.�
�
��'�=!B�,Un� ejemplar� depositado� en� el�Museo� Lucas�Roselli� (MLR� 219)�
(Fig.�7�O),�Nueva�Palmira,�Dpto.�de�Colonia,�Uruguay,�proveniente�de�la�cantera�
Arroyo�Las�Flores�(Formación�Asencio)�en�Las�Flores.�Este�ejemplar�fue�ilustrado�
por�Roselli�en�su� trabajo�de�1939�(figuras�23�y�24)�cuando�creó� la� icnoespecie�
bajo�el�nombre�de��������������� ��.�
�&=��!&'�D&(! &$�,�Un�ejemplar�mencionado�por�Roselli�erróneamente�
como�holotipo�(MLR�319)�(Fig.�7�P�y�Q)�en�el�trabajo�de�1987,�proveniente�de�la�
cantera� Maldonado� cuya� designación� como� tal� no� tiene� valor� nomenclatorial�
debido�a�que�en�el�trabajo�de�1939,�la�descripción�de��������������� ���está�
basada� sobre� otro� ejemplar,� único,� el� cual� puede� identificarse� por� las� figuras�
originales�(Roselli,�1939,� figs.�23�y�24),�y�se�encuentra�alojado�en� la�colección�
bajo� el� número�MLR� 219� (Fig.� 7�O).� Un� ejemplar� catalogado� como�(�� ����
�������� bajo� el� número� MLR� 479� comentado� previamente� (Fig.� 7� F).� Un�
ejemplar�de� la� serie� tipica�de� ������ ����� ��� ���� (MLR�607)� (Fig.�7�N).�Se�
examinó� además� un� total� de� 8� ejemplares,� provenientes� de� la� Formación�
Sarmiento,� localidad� Gran� Barranca� (Chubut,� Argentina),� los� cuales,�
discriminados� por� miembros,� son� los� siguientes:� 4� ejemplares� (MPEF�Nº� de�
campo�43,� 479,� 481,� 546)� provenientes� del�Miembro� Puesto�Almendra� Inferior�

� ���
(Eoceno� medio�Eoceno� tardío),� 4� ejemplares� (MPEF�Nº� de� campo� 653,� 1132,�
1146,� 1163)� provenientes� del� Miembro� Puesto� Almendra� Superior� (Oligoceno�
medio�temprano�Oligoceno�tardío).��
�!&� �"!" �(� $&$&, Cámaras� aisladas,� esféricas� a� subesféricas,� con�
una�pared�construída,�gruesa,�relativamente.�En�uno�de�los�polos,�la�pared�de�la�
cámarapuede�mostrar�un�agujero,�cilíndrico�en�sección�longitudinal,�que�se�abre�
al�exterior,�o�una�pequeña�perforación�en�la�base�de�un�embudo,�el�cual�también�
se�abre�al�exterior,�en�medio�de�una�superficie�externa�más�o�menos�aplanada,�
la�cual�posee�un�borde�o�anillo�que�apenas�sobrepasa�la�superficie�de�la�pared�de�
la� cámara.� El� interior� de� esta� última� posee,� como� norma,� relleno� pasivo�
(modificado�de�Laza�2006).�
��(� =&�!�", en� el� trabajo� de� 1939,� Roselli� crea� la� nueva� icnoespecie�
������� ������� ��,� basando� su� descripción� y� diagnosis� sobre� un� único�
ejemplar�proveniente�de�la�canterra�Arroyo�Las�Flores�de�la�Formación�Asencio,�
Uruguay.� En� dicho� trabajo� no� menciona� el� número� de� catálogo� del� ejemplar�
desripto,�pero�muestra�unas�figuras�del�mismo�(figuras�23�y�24).�Posteriormente,�
en� su� trabajo� de�1987,� designa�al� ejemplar� 319� como�holotipo� y� al� 219� como�
paratipo.�Ambas�designaciones�son�inválidas,�por�ser�posteriores�a�la�creación�de�
la�icnoespecie,�y�además�generan�confusión,�ya�que�el�ejemplar�que�ilustra�como�
219�(paratipo�en�este�trabajo�de�1987,�Lámina�II,� figura�6�y�Lámina�III,� figura�
5),� es� el�mismo� que� utiliza� originalmente� en� la� creación� de� ��� ������� ��,� es�
decir� el� holotipo,� motivo� por� el� cual� aquí� se� reconoce� como� holotipo� de� ���
������� ��� (Fig.� 7� O).� En� la� revisión� icnotaxonómica� de� ������������
presentada�por�Laza�(2006),�se�conserva�el�error�de�esta�designación�posterior�
hecha� por� Roselli,� mencionando� al� ejemplar� MLR� 319� como� holotipo� y� al� 219�
como�paratipo�de����������� ��.�

� ��
��"*�!B*!> , El� holotipo� de� ��� ������� ��� (MLR� 219,� descripto� en� el�
trabajo� de� Roselli� de� 1939� como� ������� ������� ��),� posee� tres� pequeñas�
cámaras� alargadas� excavadas� en� la� superficie� interna� de� la� pared,� que� él�
interpretó�como�“cámaras�de�incubación�de�himenópteros”.�La�presencia�de�estas�
cámaras� permitió� reconocer� a� dicho� ejemplar,� durante� la� revisión� del� material�
depositado�en�el�Museo�Lucas�Roselli�(enero�de�2009)�y�por�comparación�con�las�
figuras�ilustradas�en�sus�trabajos,�como�el�holotipo.�Estas�estructuras,�excavadas�
en�la�parte�interna�de�la�pared,�podrían�ser�trazas�de�organismos�cleptoparásitos�
o�detritívoros�(Sánchez�y�Genise,�2009).�El�ejemplar�catalogado�como�(�� ����
�������� bajo� el� número� de� la� serie� MLR� 479� (Fig.� 7� F),� incluído� aquí� en� ���
������� ��,� se�encuentra� fragmentado,�y�mide�37�mm�de�diámetro�ecuatorial,�
con�un�agujero�de�13,3�mm�de�diámetro,�aproximadamente,�y�una�pared�de�4,5�
mm�de� espesor.� El� ejemplar� de� la� serie� típica� de� ������ ����� ��� ���� (MLR�
607)� (Fig.� 7� N),� también� incluído� aquí� en� ��� ������� ��,� se� encuentra�
parcialmente�fragmentado,�con�una�diámetro�ecuatorial�de�35,6�mm,�un�agujero�
de�7,2�mm�y�una�pared�de�5,5�mm�de�espesor.�Las�medidas�y�otros�caracteres�
morfologicos�del�resto�de�los�ejemplares�estudiados�de�Patagonia�se�encuentran�
detallados�en�diferentes�secciones�de�los�resultados.�
�����������������������&L&4<<3�
(Fig.�6�E�y�K)�
�
�������������������Laza�2006,�p.�230�(�����);�2007:�Krell,�p.�41;�2007:�Sánchez���
��,�p.�41;�2008:�Sánchez�����,�p.�162.�
��'�=!B�, Un� ejemplar� (MACN�Icn� 1769)� depositado� en� la� División�
Icnología� del�Museo�Argentino� de� Ciencias� Naturales,� proveniente� del�Miembro�

� ���
Colhue�Huapi� de� la� Formación� Sarmiento� (Mioceno� inferior)� de� Gran� Barranca,�
Chubut,�Argentina.��
�&�&=!B�, Un� ejemplar� (MACN�Icn� 1682)� depositado� en� la� División�
Icnología� del�Museo�Argentino� de� Ciencias� Naturales,� proveniente� del�Miembro�
Puesto�Almendra�de�la�Formación�Sarmiento�(Eoceno�medio�Oligoceno�tardío)�de�
Gran�Barranca,�Chubut,�Argentina.�
�&=��!&' �D&(! &$�, Se� examinó� un� total� de� 17� ejemplares,�
provenientes� de� la� Formación� Sarmiento,� localidad� Gran� Barranca� (Chubut,�
Argentina),� los� cuales,� discriminados� por� miembros,� son� los� siguientes:� 10�
ejemplares� (MPEF�Nº� de� campo� 70,� 101,� 115,� 123,� 442,� 457,� 484,� 495,� 509,�
518)�provenientes�del�Miembro�Puesto�Almendra�Inferior�(Eoceno�medio�Eoceno�
tardío),�5�ejemplares�(MACN�Icn�2258,�MPEF�Nº�de�campo�635,�697,�994,�1131)�
provenientes� del� Miembro� Puesto� Almendra� Superior� (Oligoceno� medio�
temprano�Oligoceno� tardío),� 2� ejemplares� (MPEF�Nº� de� campo� 1001,� 1002)�
provenientes�del�Miembro�Colhue�Huapi�(Mioceno�temprano�Mioceno�medio).��
�!&� �"!"�(� $&$&,Cámaras�aisladas,� subesféricas�a�piriformes,� con�
una�pared�construída,�relativamente�gruesa.�Internamente�compuestas�por�una�
cavidad� o� cámara� principal� esférica� conectada� por� un� pasaje� estrecho� a� otra,�
secundaria,� más� pequeña� y� tambien� esférica.� Hacia� uno� de� los� polos� de� la�
cámara,� se� observa� externamente� un�mamelón,� o� protuberancia,� dentro� de� la�
cual� internamente� se� ubica� la� segunda� cavidad� menor,� completa.� La� cámara�
principal,� como�norma,� posee� en�el� interior� relleno�pasivo� (modificado�de� Laza�
2006).
��"*�!B*!> ,El�holotipo�y�el�paratipo�de�����������ya�han�sido�descriptos�
en� detalle� por� Laza� (2006),� motivo� por� el� cual� no� serán� repetidos� aquí.� Las�

� ���
medidas�y�otros�caracteres�morfológicos�del�resto�de� los�ejemplares�estudiados�
se�encuentran�detallados�en�diferentes�secciones�de�los�resultados.�
�
���������������!"B�� �+�
(Fig.�6�F�y�L)�
�
������������isp.�Mikul�y�Genise�2003,�p.�341.�
�������������������Laza�2006,�p.�230�(�����).��
�
��'�=!B�,Un�ejemplar�(MMP�4046)�depositado�en�el�Museo�Municipal�de�
Mar�del�Plata�“Lorenzo�Scaglia”,�proveniente�de�la�Formación�Vorohué�(Plioceno),�
Necochea,�Buenos�Aires,�Argentina.��
�&�&=!B�,Un� ejemplar� (MMP� 114)� depositado� en� el�Museo�Municipal� de�
Mar�del�Plata�“Lorenzo�Scaglia”,�proveniente�de�la�Formación�Vorohué�(Plioceno),�
Necochea,�Buenos�Aires,�Argentina.�����
�&=��!&' �D&(! &$�,Se� examinó� un� total� de� 3� ejemplares� (MMP� 108,�
111,�115),�provenientes�de�la�Formación�Vorohué�(Plioceno),�Necochea,�Buenos�
Aires,�Argentina.��
�!&� �"!", Cámaras� aisladas,� biesféricas,� con� una� pared� construida,�
relativamente� delgada.� Internamente� compuestas� por� una� cavidad� o� cámara�
principal� esférica� conectada� por� un� pasaje� a� otra,� secundaria,�más� pequeña� y�
también�esférica.�Externamente,�la�pared�de�esta�estructura�biesférica�posee�un�
estrangulamiento� muy� pronunciado,� que� delimita� ambas� cámaras.� Las� dos�
cámaras,�como�norma,�poseen�en�el�interior�relleno�pasivo.
��(� =&�!�",luego�del�trabajo�publicado�por�Laza�(2006),�se�realizó�una�
nueva�revisión�del�material�proveniente�de� las� formaciones�Sarmiento�(Chubut,�
Argentina)� y� Vorohué� (Buenos� Aires,� Argentina).� Como� resultado� de� esta�

� ���
revisión,�se�consideró�necesaria� la�separación�del�material� incluído�por�él�en����
������� en� dos� icnoespecies� diferentes.� El� carácter� fundamental� que� permitió� la�
creación�de�esta�nueva�icnoespecie�(���isp.�B�����)�que�comparte�con�����������la�
completa�separación�de�las�dos�cámaras�que�las�conforman,�es�la�forma�general�
claramente� biesférica� delimitada� por� un� estrangulamiento� pronunciado� entre�
ambas�cámaras�de�las�estruturas�incluídas�aquí,�en�comparación�con�una�forma�
subesférica�a�piriforme�de�los�ejemplares�de�����������Laza�2006.�
��"*�!B*!> ,El� holotipo� de������������� isp.� B� (MMP�114),� posee� un�
eje�mayor�de�5,7�cm�de�largo,�el�cual�atraviesa�verticalmente�las�dos�cámaras,�y�
uno�menor�de�4,4�cm�de�largo,�el�cual�se�corresponde�con�el�diámetro�ecuatorial�
de� la� cámara� principal.� La� cámara� secundaria,� de� 1,6� cm� de� diámetro,� se�
encuentra�comunicada�con�la�principal�por�medio�de�una�perforación�de�9�mm�de�
diámetro�excavada�en�su�base.�La�pared�de�esta�estructura�biesférica,�varía�de�2�
mm�a� 3,7�mm�de� espesor,� y� en� la� cámara� principal� posee� tres� ejemplares� de�
�������� ����� ������ (Mikul� y� Genise,� 2003).� La� superficie� externa� de� la�
pared,� presenta� muchas� irregularidades� y� pequeñas� depresiones� alargadas,�
algunas� de� las� cuales� parecen� ser� improntas� de� la� actividad� del� escarabajo�
generadas� durante� la� construcción� de� la� traza.� El� paratipo� (MMP� 114),�
conservado�aún�en�la�roca�de�caja,�posee�un�eje�mayor�de�5�cm�de�largo,�y�uno�
menor�de�3,7�cm�de� largo� (los�ejes�son� tomados�de� la�misma�manera�que� los�
descriptos�para�el�holotipo).�La�cámara�secundaria�mide�1,2�cm�de�diámetro�y�se�
comunica�con�la�mayor�por�medio�de�una�perforación,�excavada�en�la�base,�de�9�
mm�de�diámetro.�La�pared,�de�2�mm�a�3�mm�de�espesor,�sólo�puede�ser�medida�
en�la�cámara�principal,�ya�que�en�la�cámara�secundaria�se�confunde�con�la�roca�
de� caja.� Todo� el� material� examinado,� incluídos� los� ejemplares� tipo,� son�
estructuras�muy�frágiles,�donde�la�cámara�secundaria�se�encuentra�preservada,�

� ���
en�general,�solo�en�parte;�incluso�en�uno�de�ellos�(MMP�115)�se�preserva�la�base�
de� la�misma,� y� por� ese�motivo� la�mayoría� no�pudo� ser� extraído�de� la� roca�de�
caja.��
�!�%�&3�Icnoespecies�de������������.�������������;�������isp.�A;�������������;������� ������� ��;������� ������;� ������ isp.� B.;��#��� Vista� interna� de� las� seis� icnoespecies�cortadas� por� el� eje� del� agujero� de� emergencia� o� canal� de� aireación,� respectivamente.�Escalas:�1�cm.��

� ���

� ���
�!�%�&;���Holotipo�de������������(MLR�479);��#���Serie�conservada�como�MLR�479,��� y� �� corresponden� a� dos� vistas� distintas� de� un� mismo� ejemplar� mostrando,� en� la�segunda� (�),� un� ejemplar� de� �������� ����� �����,� �� representa� un� ejemplar�considerado�aquí�como����������� ��;����Holotipo�de� ���� ������������(MLR�641)�incluido�en�����������;��#���Holotipo�y�paratipo�de� � ��� �� ��������� ���(MLR�717�y�717a)�incluidos�en�����������;����Holotipo�de� ������ �������� ����(MLR�607)�incluido�en� ��� �������; -� Ejemplar� MLR� 714,� catalogado� erróneamente� como� paratipo� de� ������ ����� ��� ���;��#��� Serie� típica� de� ������ ����� ��� ���� (MLR� 607),��� y���representan�ejemplares�incluidos�en������������y���en����������� ��;����Holotipo�de����������� ���(MLR�219);��#M��Dos�vistas�distintas�del�ejemplar�MLR�319.�Escalas:�1�cm.���
6�2�4# �&L&"I>"!'�"$����& !"(�"&"�*!&$�"&��������������
!����"��������!1%'NO.�� !"�4<<5
�
�������� ����� Mikuláš� &� Genise� 2003,� p.� 339� (icnoespecie� tipo� por� designación�
original:� �������� ����� ������ Mikulás� &� Genise);� 2006:� Laza,� p.� 227;� 2009:�
Sánchez�y�Genise,�p.�837.�
�
�!&� �"!" � (� $&$&�� Perforaciones� pasantes� (perforaciones�
completas)�de�sección�circular�a�sub�circular�en�corte�transversal,�o�depresiones�
internas� o� externas� (perforaciones� incompletas),� hechos� en� las� paredes� de� las�
cámaras�construidas�con�material�del�suelo�aglutinado.�La�presencia�de�una�sóla�
perforación,� sin� importar� su� tamaño,� no� es� diagnóstica� para� el� icnogénero,� el�
cual�sólo�puede�identificarse�cuando�más�de�una�perforación�está�presente�en�la�
pared.�Cuando�las�cámaras�muestran�una�sola�perforación�mayor�y�una�o�más�de�
menor�calibre,�la�primera�debe�ser�considerada�como�parte�del�sustrato,�es�decir,�
como� parte� de� la� cámara� construida.� Sin� embargo,� la� presencia� de� una� sola�
depresión�en� la�superficie�externa�de� la�pared�si�debe�ser�usada�como�carácter�
diagnóstico�del�icnogénero�(enmendado�de�Mikuláš�y�Genise�2003).�
��(� =&�!�"��La�diagnosis�fue�enmendada�para�poder�incluir�depresiones�
en� la� superficie� interna� de� paredes� construidas� de� cámaras.� Estas� trazas� en�

� ���
trazas,� compatibles� con� la�morfología� representada�por��������� ����,� no� se�
conocían�en�el�momento�que�Mikuláš� y�Genise� (2003)�definieron�el� icnogénero�
por�primera�vez.���
�
!����"���������������N *K�L.�� !"�4<<7
(Fig.�8�A�E)�
(Fig.�9�A)�
�
�������� ���������Sánchez�y�Genise�2009,�p.�837.�
�
�=!(�'��F&��Dedicado�a�José�“Pepe”�Laza,�un�pionero�en�la�paleoicnología�
de� insectos,� quien� además� descubrió� el� material� tipo� de� esta� icnoespecie,� así�
como� otras� tantas� joyas� de� la� icnología� durante� 30� años� de� colección,�
preparación�y�observación�de�especímenes�de������������.����
��'�=!B��� Un� espécimen� (MACN�Icn� 2346)� en� un� ejemplar� de�
������������ isp.� (MACN�Icn�2074)�de�Sierra�de�Talquino,�Chubut,�Argentina�
(Formación�Sarmiento).�
�&�&=!B�"�� Tres� especímenes� (MACN�Icn� 2342,� 2345� y� 2355)� en� un�
ejemplar�de�������������isp.�(MACN�Icn�2074)�de�Sierra�de�Talquino,�Chubut,�
Argentina�(Formación�Sarmiento).�Un�espécimen�(MPEF�IC�601)�en�un�ejemplar�
de������������� isp.� A� (MPEF�IC� 600)� de� Gran� Barranca,� Chubut,� Argentina�
(Eoceno�Medio,�Miembro�Gran�Barranca,�Formación�Sarmiento).�
�&=��!&' �D&(! &$��� Diecinueve� especímenes� (MACN�Icn� 2342�2360),�
ocho� especímenes� (MACN�Icn,� 2379�2386)� y� cinco� especímenes� (MACN�Icn�
2393�2397)� en� un� ejemplar� de������������ isp.� (Laza� col.� MACN�Icn� 2074),�
�������������������(Laza�col.�MACN�Icn�2300)�y������������ isp.�A�(Laza�

� ��
col.�MACN�Icn�2392)�respectivamente,�de�Sierra�de�Talquino,�Chubut,�Argentina�
(Formación� Sarmiento).� Setenta� y� cinco� especímenes� (MPEF�IC� 601�632,� 639�
648,�650�664�y�MACN�Icn�2361�2378)�en�cuatro�especímenes�de�������������
isp.� A� (MPEF�IC� 600,� 638,� 649� y� MACN�Icn� 1620)� respectivamente,� de� Gran�
Barranca,�Chubut,�Argentina�(Eoceno�Medio�Oligoceno,�Miembros�Gran�Barranca�
y�Puesto�Almendra,�Formación�Sarmiento).�Cinco�especímenes�(MACN�Icn�2387�
2391)� en� un� ejemplar� de� ������������ isp.� (MACN�Icn� 1382)� de� Gran�
Barranca,� Chubut,� Argentina� (Eoceno� Superior,� Miembro� Puesto� Almendra,�
Formación� Sarmiento).� Un� espécimen� (MPEF�IC� 637)� en� un� ejemplar� de�
������������ isp.� A� (MPEF�IC� 636)� de� Gran� Barranca,� Chubut,� Argentina�
(Eoceno�Medio,�Miembro�Gran� Barranca,� Formación� Sarmiento).� Un� espécimen�
(MPEF�IC� 635)� en� un� ejemplar� de� ������������ �������� (MPEF�IC� 634)� de�
Gran� Barranca,� Chubut,� Argentina� (Eoceno� Medio,� Miembro� Gran� Barranca,�
Formación�Sarmiento).�
�!&� �"!"���������� �����representado�por�una�depresión�elongada�en�
la�superficie�interna�de�la�pared�de�especímenes�de������������,�que�en�corte�
muestra� una� sección� circular� a� elíptica.� Esta� depresión� puede� alojarse� en� el�
centro�o�hacia�un�costado�de�un�montículo�ovoide,�que�no�siempre�está�presente.�
��(� =&�!�"�� �������� ����� ���� se� diferencia� de� ��� ������ ���
debido�a�que�se�aloja�en� la�superficie� interna�de� la�pared�de� los�ejemplares�de�
�����������.� Sumado� a� ésto,� muchos� especímenes� muestran� un� montículo�
que�está�ausente�en���������� ��.��
��"*�!B*!> �� El� holotipo� (MACN�Icn� 2346)� y� algunos� otros� especímenes�
(MACN�Icn�2342�2360,�2387�2391)�se�observan�en�el�interior�de�ejemplares�de�
������������que�no�pueden�ser� identificados�ya�que�no�muestran�evidencias�
de�agujero�de�emergencia�o�de� cámara�ovígera� (Fig.�8�A� y�B).�El� resto�de� los�

� ��
especímenes� se� alojan� en� la� superficie� interna� de� la� pared� de� ejemplares� de�
��������������������o����isp.�A�(Fig.�8�D�y�E).�En�la�mayoría�de�los�casos�los�
especímenes� son� alargados� con� el� eje� mayor� perpendicular,� oblicuo� o� casi�
tangencial�a� la�superficie�de�la�pared.�Los�pocos�especímenes�semiesféricos�son�
interpretados� como� ejemplares� parcialmente� preservados.� Las� depresiones�
miden� de� 1�mm� a� 6,8�mm�de� largo� (N�=� 45),� y� aquéllos� que� poseen� sección�
circular,�presentan�2,2�mm�a�4,6�mm�de�diámetro�(N�=�19).�En�los�especímenes�
de�sección�elíptica,�el�eje�mayor�mide�de�2,6�mm�a�5,3�mm�y�el�eje�menor�de�2�
mm�a�4,1�mm�de�largo�(N�=�26).�Todas�las�depresiones�muestran�una�superficie�
interna�lisa.�En�los�ejemplares�más�completos,�la�depresión�es�cerrada�y�se�abre�
en� el� centro� de� un�montículo� elongado� constituido� por� el� mismo�material� que�
conforma�la�pared�de�la�cámara�de�cría�(Fig.�8�C).�El�montículo�mide�de�1�mm�a�
4,4�mm�de�alto,�4,2�mm�a�6,7�mm�de�largo�y�4,1�mm�a�4,9�mm�de�ancho�(N�=�
14).�
� Micromorfológicamente,�el� corte� transversal�de�una�depresión�muestra� la�
superficie�interna�recubierta�por�una�capa�muy�delgada�de�50�325�^�de�espesor�
(Fig.� 9� A)� que� difiere� en� composición� de� la� pared� de� �����������.� La�
microestructura� de� esta� capa� es�microgranular,� con� un� 10�%� de� porosidad,� y�
está� compuesta� por� gránulos� de� arcilla� que� no� muestran� birrefringencia.� La�
fracción� gruesa� es� tamaño� arena� fina�limo,� y� esta� compuesta� por� trizas�
volcánicas,� cuarzo� y� fragmentos� líticos.� La� fracción� fina� esta� compuesta� por�
abundante� cantidad� de� arcilla,� amarilla� y� birrefringente,� que� rodea� a� los�
microgránulos� y� rellena� los� poros.� La� pared� del� ejemplar� de� ������������
donde� se� aloja� esta� traza,� muestra� una� microestructura� maciza� con� una�
porosidad�menor�al�1�%.�La�fracción�gruesa�es�mayor�(40�%)�que�en�la�capa�que�
recubre�a�������,�y�además,�entre�los�minerales�que�la�componen�se�observa�

� �
también�plagioclasa.�La�fracción�fina�esta�compuesta�por�arcilla�color�pardo�y�con�
poca�birrefringencia,�oscurecida�por�óxido�(Fig.�9�A).�
�
#�$���������!1%'NO.�� !"�4<<5
#�$��������%���� �����!1%'NO.�� !"�4<<5
(Fig.�8�F�y�G)�
(Fig.�9�B�y�C)�
��� �����Mikuláš�&�Genise,�2003:�339;�2006:�Laza,�p.�224,�fig.�5F;�2007:�Sánchez���
��,�p.�41;�2009:�Sánchez�y�Genise,�p.�837.��
�
�&=��!&' �D&(! &$��� Dieciséis� especímenes� (MPEF�IC� 666,� 668,� 670,�
672,�674,�676,�678,�680,�682,�684,�686,�688,�690,�y�692,�y�MACN�Icn�2398�y�
2399)�en�dieciséis�especímenes�de�������������isp.�(MPEF�IC�665,�667,�669,�
671,�673,�675,�677,�679,�681,�683,�685,�687,�689,�y�691,�y�MACN�Icn�1373�y�
1605)�de�Gran�Barranca,�Chubut,�Argentina�(Eoceno�Medio�Oligoceno,�Miembros�
Gran�Barranca�y�Puesto�Almendra,�Formación�Sarmiento).��
�!&� �"!"�Agujeros�de�sección�circular�a�sub�circular�que�atraviesan� las�
paredes�de�cámaras�hechas�de�material�del�suelo�aglutinado�y�que�se�conectan�
con�galerías�internas�en�el�relleno�de�dichas�cámaras.�Un�solo�agujero�conectado�
con�una�cavidad�sin�importar�el�tamaño�que�tenga,�no�es�un�carácter�diagnóstico�
de�este�icnogénero�(Mikuláš�y�Genise�2003).�
��"*�!B*!> �� Los� agujeros� y� galerías� ocurren� en� las� paredes� y� rellenos,�
respectivamente,� de� especímenes� de� ������������ que� han� sufrido� una�
meteorización� intensa,� y� que� por� tal� motivo� han� perdido� los� caracteres� que�
permiten� identificar� las� icnoespecies� (Fig.� 8� F).� No� se� observa� un� patrón� o� un�

� ��
arreglo�particular�de�los�agujeros�en�la�pared�ni�de�las�galerías�en�el�relleno,�sin�
embargo,� si� se� observa�una� conexión� con�una� red�de� túneles�en� el� paleosuelo�
(Fig.� 8� G).� Los� agujeros� y� los� túneles� de� las� galerías� muestran� en� corte�
transversal�una�sección�circular�a�sub�circular.�En�los�primeros�el�diámetro�mide�
de�1,3�mm�a�3,5�mm�(N�=�46),�y�en�los�de�sección�sub�circular�se�observan�dos�
ejes,�uno�mayor�de�2,9�mm�a�3,5�mm�de�largo,�y�uno�menor�de�2,2�mm�a�2,5�
mm� de� largo� (N� =� 4).� Los� túneles� pueden� estar� rellenos� o� vacíos,� siempre�
poseen� un� tapiz� de� textura� lisa� que� cubre� su� interior,� y� en� algunos� casos,� un�
ensanchamiento�de�5�mm�a�6,4�mm�de�diámetro�hacia�el�extremo�mas�distal�(N�
=� 4).� Algunos� de� los� ejemplares� de� ������������ examinados,� muestran�
túneles� con�y� sin�meniscos�excavados� solo�en�el� relleno� (Fig.� 8�H�J),�mientras�
que�otros�ejemplares�muestran�el�caso�contrario�ya�que�solo�poseen�agujeros�en�
la� pared� y� ninguna� estructura� en� el� relleno� (Fig.� 8� K).� Estas� trazas,� tal� vez�
producidas�por�el�mismo�organismo,�no� son� consideradas���� ����� �����������
ya�que�icnotaxonómicamente�carecen�de�agujeros�en�la�pared�en�un�caso,�o�de�
galerías�en�el�relleno�en�el�otro.������
Micromorfológicamente,� el� relleno� de�������������muestra� una� banda�
de�material�oscuro�muy�delgada,�de�25�50�^�de�espesor�(Fig.�9�B),�depositada�
alrededor� de� los� túneles� que� conforman� las� galerías.�Sin� embargo,� fragmentos�
cortos� de� estas� bandas� oscuras,� también� se� observan�dispersos� por� el� relleno.�
Cada�túnel�posee�un�tapiz�de�arcilla�muy�delgado�de�25�75�^�de�espesor�(Fig.�9�
C).�Observado�al�microscopio�petrográfico�y�con� luz�polarizada,� la�arcilla�que� lo�
conforma� es� amarilla,� birrefringente� y� ópticamente� orientada� (Fig.� 9� C).� El�
relleno�de� los�túneles�es�más�poroso�que�el�relleno�de� la�������������y�está�
compuesto� por� trizas� vítreas,� plagioclasa,� cuarzo� y� arcilla� irregularmente�
distribuida(Fig.�9�C).�Los�especímenes�de���������������están�conectados�con�un�

� ��
“��+����”� en� el� paleosuelo,� el� cual� muestra� además,� túneles� y� cámaras� con�
rellenos�meniscados�(Fig.�9�D�y�E).�Micromorfológicamente,�el� túnel�meniscado�
mas�completo�mide�5,75�mm�de� largo�y�de�1,12�mm�a�1,25�mm�de�diámetro.�
Los� meniscos� miden� de� 125� ^� a� 175� ^� de� espesor,� están� compuestos� por�
material� similar� al� del� paleosuelo,� y� los� separa� una� capa� de� arcilla,� que� al� ser�
observada�con�luz�polarizada,�es�amarilla�y�birrefringente�(Fig.�9�D).�La�cámara�
más�completa�es�ovoide�con�un�eje�mayor�de�3,62�mm�de�largo�y�uno�menor�de�
2,75�mm�de� largo�(Fig.�9�E).�La�superficie� interna�de�una�cámara�se�encuentra�
recubierta�por�un�tapiz�de�arcilla�de�75�^�a�500�^�de�espesor,�discontinuo,�que�
forma�en�algunos�sectores�estructuras�meniscadas�de�75�^�de�espesor�orientadas�
con�la�concavidad�hacia�el�lúmen�de�la�cámara.�La�arcilla�que�forma�este�tapiz�y�
las�estructuras�meniscadas,�es�amarilla,�birrefringente,�y�ópticamente�orientada�
cuando�se�lo�observa�con�luz�polarizada.�La�cámara�está�rellena�parcialmente�por�
material� del� suelo� y� también� por� meniscos� de� arcilla� que� muestran� el� mismo�
espesor�y�birrefringencia�que�los�del�tapiz.�Algunos�de�estos�meniscos�muestran�
una� cierta� orientación� y� un� leve� arreglo� en� forma� de� anillo,� sin� embargo� la�
mayoría�de�ellos�se�encuentran� irregularmente�orientados�dentro�de� la�cavidad�
de�la�cámara�(Fig.�9�E).
�
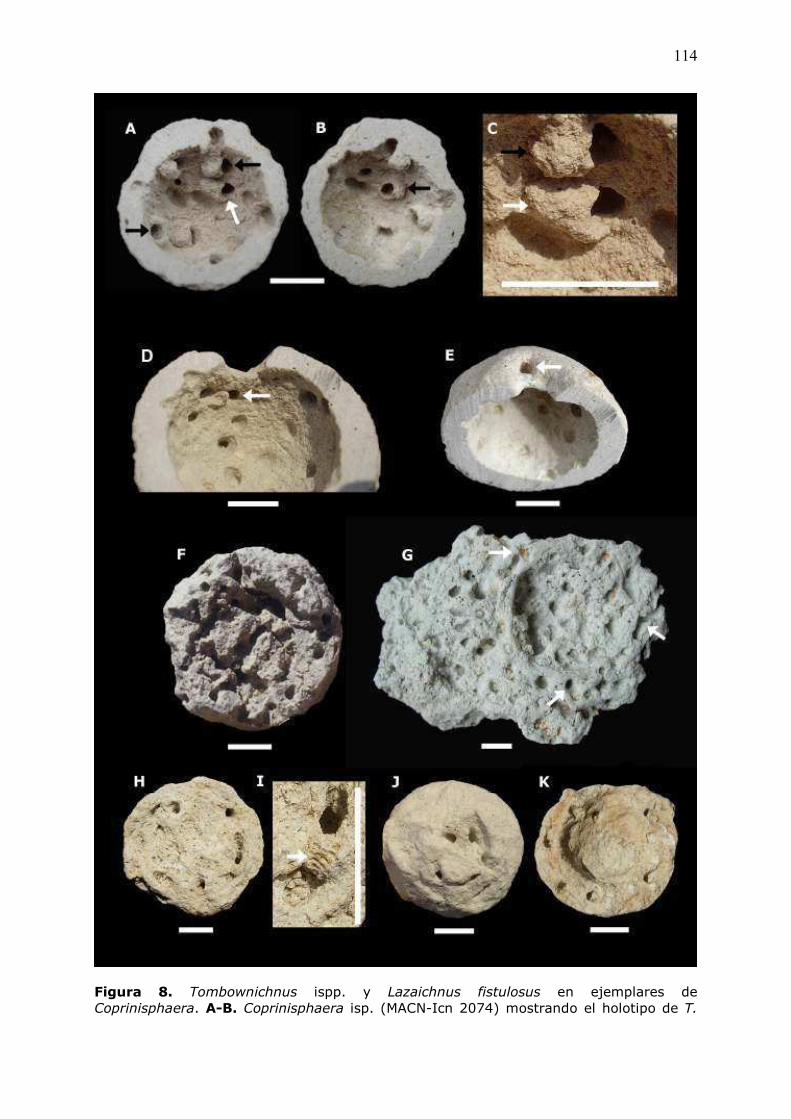
� ��
�!�%�& :� �������� ����� ispp.� y� ��� ����� ����������� en� ejemplares� de������������.#�������������� isp.� (MACN�Icn�2074)�mostrando�el�holotipo�de����
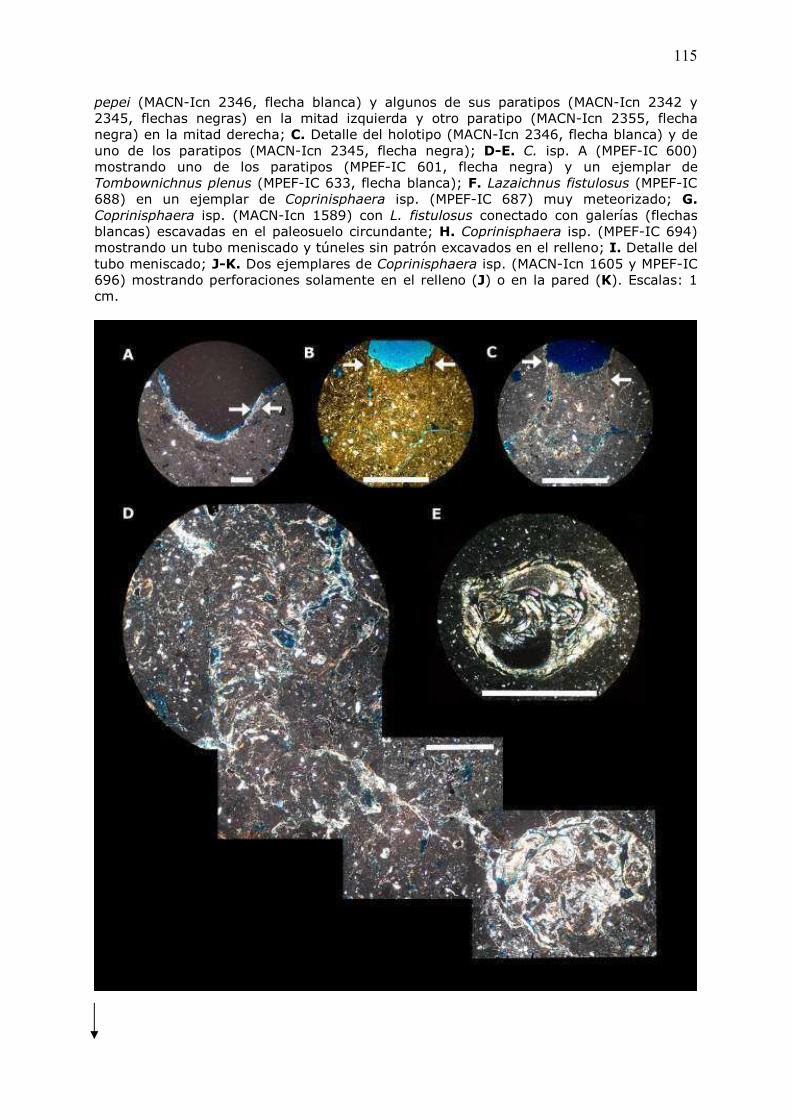
� ��
���� (MACN�Icn� 2346,� flecha� blanca)� y� algunos� de� sus� paratipos� (MACN�Icn� 2342� y�2345,� flechas� negras)� en� la� mitad� izquierda� y� otro� paratipo� (MACN�Icn� 2355,� flecha�negra)�en�la�mitad�derecha;����Detalle�del�holotipo�(MACN�Icn�2346,�flecha�blanca)�y�de�uno� de� los� paratipos� (MACN�Icn� 2345,� flecha� negra);� �#�� ��� isp.� A� (MPEF�IC� 600)�mostrando� uno� de� los� paratipos� (MPEF�IC� 601,� flecha� negra)� y� un� ejemplar� de��������� �����������(MPEF�IC�633,�flecha�blanca);������� ����������������(MPEF�IC�688)� en� un� ejemplar� de� ������������ isp.� (MPEF�IC� 687)� muy� meteorizado;� ��������������� isp.� (MACN�Icn� 1589)� con� ��� ����������� conectado� con� galerías� (flechas�blancas)�escavadas�en�el�paleosuelo�circundante;��������������� isp.�(MPEF�IC�694)�mostrando�un�tubo�meniscado�y�túneles�sin�patrón�excavados�en�el�relleno;����Detalle�del�tubo�meniscado;��#-��Dos�ejemplares�de�������������isp.�(MACN�Icn�1605�y�MPEF�IC�696)�mostrando�perforaciones�solamente�en�el�relleno�(�)�o�en�la�pared�(-).�Escalas:�1�cm.����

� ��
�!�%�&7�Micromorfología�de��������� �������� ()� y���� ����� ����������� (�#�).���Corte�transversal�de�������,�visto�con�luz�polarizada,�mostrando�una�capa�delgada�de�arcilla�birrefringente�cubriendo�la�superficie�interna�de�la�depresión�(flechas�blancas).�Escala:� 700� ^;� �#��� Túnel� excavado� en� el� relleno� de� un� ejemplar� de� ������������delimitado�por�una�capa�muy�delgada�de�material�oscuro�(�,�flechas�blancas),�y�una�capa�interna�muy�delgada�de�arcilla�birrefringente�visto�con�luz�polarizada�(�,�flechas�blancas).�Escala:� 2�mm;���Túnel� excavado� en� el� paleosuelo� circundante,�mostrando� un� relleno�meniscado�con�arcilla�amarilla�y�birrefringente�depositada�entre� los�sucesivos�meniscos�(visto� con� luz� polarizada),� conectado� con� una� cámara� (����� ����� �� ������).� Escala:�1,5�mm;�������� ����� �� �������mostrando� una� capa� interna� conformada� por�������meniscados�de�arcilla�birrefringente,�y�relleno�de�material�del�paleosuelo�disperso�y�una�hilera�de�meniscos�de�arcilla�birrefringente.�6�4#�� �������������������,������ ���� ������������������������
�������������������������� ��
�
Para�describir�la�estructura�de�las�masas/bolas�de�cría,�como�se�presenta�a�
continuación,� se� seleccionaron:� (1)� tres� masas� de� cría� de� ��� � ������� en�
estadios�poco�avanzados�del�desarrollo,�dos�de�ellas�con�un�huevo�cada�una,�y�
una�tercera�con�una�larva;�(2)�masas�de�cría�con�cámara�ovígera,�donadas�a�la�
colección� por� Cabrera�Walsh� y� Gandolfo;� (3)� cinco� masas� de� cría� de�
�����������sp.,�cada�una�con�un�huevo�en�su�interior;�(4)�nueve�bolas�de�cría�
��� ��� ���� en� distintos� estadios� de� desarrollo� (desde� huevo� hasta� pupa);� (5)�
siete�bolas�de�cría�de��������,�tres�de�ellas�con�un�huevo�cada�una�y�el�resto,�
depositadas�previamente�en� la�colección,�emergidas;�(6)�cinco�bolas�de�cría�de�
��� �������,� tres� de� ellas� con� una� larva� cada� una� en� distintos� grados� de�
desarrollo,� y� cada� una� de� las� restantes� con� una� adulto� no� emergido;� (7)� � dos�
bolas�de�cría�de��������,�cada�una�con�una�larva�en�desarrollo;�(8)�una�bola�de�
cría�de� ��������! ���������"���� �� ������con�una�larva�en�su�interior,�y�
seis� bolas� de� provisión� rodadas;� (9)� dos� bolas� de� cría� de� ��������
! �������"� ������,� una� con� una� larva� y� la� otra� con� un� adulto,� muerto�
durante�su�emergencia,�y�conservado�en�dicha�condición,�y�una�bola�de�provisión�

� ��
rodada;� (10)� una�bola�de� cría� de� ������� sp.1� con�un�huevo,� y� tres� bolas� de�
provisión� rodadas;� (11)� una� bola� de� cría� de� ������� sp.2� con� una� larva;� y�
finalmente� (12)�una� cámara�pupal�de� la� cual� emergió�$�����������������.�A�
continuación�se�describen� los�nidos,�masas�y�bolas�de�cría,�y�cámaras�pupales,�
observados�para�cada�una�de�las�especies�estudiadas.�
�
6�4�2#������������"B�
�
5.2.1.1���Estructura�de�la�masa�de�cría��
Todas� las� masas� de� cría� estudiadas� poseen� cámara� ovígera,� y� pueden�
definirse�como�estructuras�sub�cilíndricas�con�ambos�extremos�redondeados�que�
miden�de�2,1�a�3,7�cm�de�largo�y�de�1�a�1,4�cm�de�diámetro�(N�=�5)�(Fig.�10�A�y�
B).�Se� encuentran� al� final� de� túneles� cuya� longitud�no�pudo� ser�medida.�Cada�
masa� de� cría� se� encuentra� conformada� por� bosta� depositada� en� forma� de�
meniscos,� que� a� ojo� desnudo� y� en� el� material� fresco,� son� difícilmente�
individualizados.�En�el�extremo�superior,�el�más�próximo�a�la�superficie�del�suelo,�
cada�masa�de�cría�posee�una�cámara�ovígera�excavada�en�la�bosta�aprovisionada�
como� relleno� (Fig.� 10� A� y� B).� La� cámara� ovígera� es� ovalada,� completamente�
cerrada,�con�un�eje�mayor�de�1,1�cm�a�0,6�cm�de�largo,�y�un�eje�menor�de�0,5�
cm�a�0,8�cm�de�largo.�Dicha�cámara�posee�una�superficie�interna�lisa�y�brillante�
en� el�material� fresco,� y� de� color�más� oscuro� que� el� del� resto� de� la� bosta,� sin�
embargo,�no�se�distingue�con�claridad�una�pared�o�un�tapiz�(Fig.�10�C).�En�todas�
las�masas�de�cría�estudiadas,�se�encontró�en�el�interior�de�la�cámara�ovígera,�un�
huevo�sujeto�a�la�parte�lateral�de�la�pared�(Fig.�10�A�y�B).�El�huevo�es�de�color�
amarillo�pálido,�de�2�cm�a�2,2�mm�de�largo�y�de�0,8�mm�a�1�mm�de�diámetro�(N�
=�3)�(Fig.�10�A�C).�

� ��
�
�!�%�&2<��Nido�y�masas�de�cría�de������������sp.��Tres�masas�de�cría�conformando�un�nido�compuesto.�Se�observa�el�interior�de�la�cámara�ovígera�(flechas�blancas)�de�las�tres�masas,�en�una�de�ellas�se�conserva�el�huevo�(flecha�negra);����Una�masa�de�cría�conformando� un� nido� simple.� La� cámara� ovígera� cortada� longitudinalmente� (flecha�blanca),�muestra�un�huevo�en�su�interior�(flecha�negra);����Vista�interna�de�una�cámara�ovígera� donde� no� se� distingue� ni� un� tapíz� ni� una� pared.� Se� observa� el� huevo� en� su�interior�(flecha�negra).�Escalas:�1�cm.������
�
5.2.1.2���Micromorfología�de�la�masa�de�cría�
En�el�contacto�entre�el�suelo�y�el� relleno�de� la�masa�de�cría�con�cámara�
ovígera,�se�observa�un�oscurecimiento�del�material�del�suelo�de�100�^�de�espesor�
aproximadamente,�debido�a�la�concentración�de�materia�orgánica�en�dicha�zona�
(Fig.� 11� A).� El� material� del� suelo,� desde� el� contacto� con� el� relleno� y� con� un�
espesor�de�500�^,�aproximadamente,�presenta�un�arreglo,�pobremente�definido,�
en�paquetes�ahusados�que�se�yuxtaponen�y�cuyos�límites�están�marcados�por�la�
concentración�de�materia�orgánica�color�castaño�oscuro�(Fig.�11�B).�Este�material�
posee� las� mismas� características� texturales� y� composicionales� del� suelo.� El�
relleno� esta� compuesto,� fundamentalmente,� por� fibras� vegetales� de� la� bosta,�
elongadas,� de� 300� ^� a� 1,4� mm� de� largo,� las� cuales� se� acomodan� formando�
meniscos�de�600�^�a�1�mm�de�espesor�máximo,�aproximadamente�(Fig.�11�C).�Si�
bien�en�el�límite�entre�cada�uno�de�estos,�el�material�se�encuentra�orientado,�las�
fibras�de�bosta�en�el�interior�de�cada�menisco�presenta�una�distribución�azarosa�
y�con�alta�porosidad.��

� �
La� cámara�ovígera� es� esférica� y� la� pared,� de�250�^�a�300�^�de� espesor�
(Fig.� 11� A),� se� compone� del� mismo� material� que� conforma� los� meniscos� del�
relleno,� pero� a� diferencia� de� los�mismos,� el� material� de� la� pared� presenta� un�
mayor�oscurecimiento�y�una�porosidad�casi�nula.��
�
�
�!�%�&22�Micromorfología�de�masas�de�cría�de������������sp.���Zona�de�contacto�entre� el� material� del� suelo,� oscurecido� por� materia� orgánica� (flechas� amarillas),� y� las�fibras� vegetales� del� relleno� de� la� masa� de� cría� que� se� encuentran� orientadas�paralelamente�a�esta�(flecha�blanca).�Se�distingue�también�el�contacto�entre�el�relleno�y�la� pared� de� la� cámara� ovígera� (flecha� negra),� la� cual� se� encuentra� conformada,�fundamentalmente,� por� fibras� vegetales� de� la� bosta.� Escala:� 350� ^;���� Detalle� de� los�paquetes� de� material� del� suelo,� ahusados� y� yuxtapuestos� unos� con� otros� (flechas�amarillas),�delimitados�por�materia�orgánica�de�color�castaño�oscuro.�Escala:�800�^;����Detalle�de�los�meniscos�del�relleno�(flechas�blancas)�conformados�por�fibras�vegetales�de�la�bosta�y�materia�orgánica�de�color�castaño�oscuro.�Escala:�2�mm.��
�
5.2.1.3���Estructura�del�nido�
Se�estudio�un�total�de�tres�nidos�hallados�debajo�de�tres�distintas�bostas�
de�vaca,�de�los�cuales�dos�corresponden�a�nidos�simples�(con�una�masa�de�cría�
cada�uno,�Fig.�10�B)�y�uno�corresponde�a�un�nido�compuesto�(conformado�por�
tres� masas� de� cría,� Fig.� 10� A).� En� ninguno� de� los� casos� se� encontraron� los�
túneles� de� almacenamiento,� y� los� de� nidificación� pudieron� ser� inferidos�
solamente�por�la�presencia�de�la/las�masa/as�de�cría�en�su�extremo�mas�distal.�
Las�masas�de� cría� fueron�halladas� a�13� cm�de�profundidad�promedio� (N�=�2),�
siendo�15�cm�la�máxima�profundidad�medida�en�uno�de�los�nidos�simples,�donde�

� ���
el�fondo�de�la�masa�de�cría�coincide�con�el�límite�entre�los�horizontes�A�y�B�del�
suelo.�Los�dos�nidos�simples�mostraron�una�masa�de�cría�cada�uno�alojada�en�el�
extremo�más� distal� del� túnel� principal,� del� cual� no� se� pudo� distinguir� hacia� la�
superficie� el� extremo� opuesto,� en� ninguno� de� los� dos� casos,� posiblemente� por�
estar� rellenos� con� material� del� suelo.� Sin� embargo,� en� uno� de� los� nidos� se�
encontró� un� remanente� de� bosta� depositado� en� forma�de�meniscos,� 1� cm� por�
encima�de�la�masa�de�cría�y�separado�de�ésta�por�material�del�suelo�(Fig.�10�B),�
que�podría�estar�indicando�tanto�la�ubicación�del�túnel�de�almacenamiento�como�
la� presencia� de� una� segunda�masa� de� cría� alineada� con� la� primera� dentro� del�
mismo�túnel.�En�el�único�nido�compuesto�estudiado�no�se�encontraron�túneles�de�
ningún�tipo,�y�se�hallaron�tres�masas�de�cría�construidas�una�al�lado�de�la�otra�en�
forma�de�racimo�y�separadas�entre�si�por�material�del�suelo.�Formando�parte�del�
racimo,� la�masa�de�cría�central�se�ubica�perpendicular�a� la�superficie�del�suelo,�
mientras�que�las�masas�de�cría�laterales�poseen�una�inclinación�mínima�respecto�
de�la�central�de�aproximadamente�10���15º.��
�
6�4�4#&�������� ����'��������%''P
5.2.2.1���Estructura�de�la�masa�de�cría�
Las�masas� de� cría� estudiadas� con� cámara� ovígera,� son� estructuras� sub�
cilíndricas�construidas�en�los�túneles�de�nidificación�en�el�extremo�opuesto�al�de�
la�superficie�del�suelo�(Fig.�12�A�C),�y�miden�de�cm�3�cm�a�3,7�cm�de�largo�y�de�
1,5�cm�a�2�cm�de�diámetro�(N�=�2).�Estas�estructuras�poseen�una�pared�discreta�
que� no� puede� ser� reconocida� a� ojo� desnudo� en� el� campo,� pero� que� tiempo�
después,� se� distingue� claramente� en� el� mismo�material,� cuando� ha� perdido� la�
humedad.� Al� secarse� toda� la� muestra,� se� observa� una� fisura� que� separa�

� ��
parcialmente� la�masa�de�cría�del� suelo�que� la�circunda�(Fig.�12�G).�Esta�pared�
puede� reconocerse� también,� y� con�mas� detalle,� en� los� cortes� delgados� de� las�
masas�de�cría�que�revelan�además�su�micromorfología�(Fig.�13�B).�Las�masas�de�
cría�muestran�un�pequeño�poro�en�su�extremo�superior,�que�conecta�el� interior�
de� la� cámara� ovígera� con� el� del� túnel� de� nidificación,� y� que� además� posee� un�
tapón�conformado�por�fragmentos�sueltos�de�bosta�y�de�material�del�suelo.�Las�
fibras�vegetales�del�tapón�penetran�el�poro�hacia�el�interior�de�la�cámara�ovígera.�
La� provisión� esta� compuesta� por� bosta� depositada,�mayormente,� en� forma� de�
meniscos�con�su�concavidad�dirigida�hacia�arriba,�es�decir�hacía�la�superficie�del�
suelo.�La�cámara�ovígera,�construida�en�el�extremo�superior�de�la�masa�de�cría,�
es�esférica�de�1�cm�a�1,1�cm�de�diámetro�(N�=�3)�y�se�encuentra�aislada�de�la�
provisión� por� una� pared� delgada.� Esta� pared� posee� en� su� superficie� interna,�
estrías�delgadas,�curvas,�concéntricas�y�poco�profundas,�pero�claramente�visibles�
(Fig.�12�F).�En�dos�de�las�masas�de�cría�estudiadas,�se�encontró�en�el�interior�de�
la� cámara� ovígera� y� depositado� sobre� una� pequeña� base,� un� huevo� de� color�
amarillo�pálido,�de�5�mm�a�5,5�mm�de�largo�y�3�mm�de�diámetro�(Fig.�12�C�y�D).�
�

� ���
�

� ���
�!�%�&24��Nido�y�masas�de�cría�de���������� ������.���Túnel�de�almacenamiento�en�corte�longitudinal;����Segundo�estadio�en�la�construcción�del�nido�donde�se�observan�unos�pocos�centímetros�de�bosta�aprovisionados�en�el�fondo�de�un�túnel�de�nidificación�vertical,�en�el�que�se�encuentra�el�adulto�(flecha�negra).�Se�observa�también�la�conexión�con� el� túnel� de� almacenamiento� (flecha� blanca);� ��� Masa� de� cría� completa� (en� corte�longitudinal)�construída�en�el�fondo�del�túnel�de�nidificación.�Se�observa�también�el�túnel�de�almacenamiento�todavía�abierto�y�relleno�parcialmente�con�bosta�(flecha�blanca);����Escarabajo�adulto�en�el�túnel�de�nidificación;����Escarabajo�adulto�eliminando�sus�heces;���Vista�interna�de�la�pared�de�la�cámara�ovígera�con�surcos�finos,�curvados,�concéntricos�y�poco�profundos�excavados�en�su�superficie�interna�(flecha�blanca);���Pared�de�la�masa�de� cría� separada� parcialmente� del� suelo� circundante� por� una� fisura� (flechas� blancas).�Escalas:�1�cm.��
�
5.2.2.2���Micromorfología�de�la�masa�de�cría�
La� pared� tiene� un� espesor� de� 4,25� –� 4,75� mm� (N� =� 1),� y� una�
microestructura�maciza� con� una� porosidad� de� 5� �� 7�%.� La� fracción� gruesa� es�
mayoritariamente� tamaño� limo,� pero�presenta� también� algunos�granos� tamaño�
arena.�Representa�el�50�%�del�volumen�total�y�esta�compuesta�por�cuarzo,�K���
feldespato,� plagioclasa,� mica,� fragmentos� líticos,� trizas� vítreas,� y� minerales�
pesados� (epidoto,� piroxeno,� anfíbol).� La� fracción� fina� esta� compuesta� por�
material�opaco,�de�granos� tamaño�arcilla,� color�castaño�claro�a�castaño�oscuro�
hacia� el� relleno,� debido� al� alto� contenido� de�materia� orgánica� (Fig.� 13� A).� La�
pared�posee,�externamente,�una�lámina�delgada�de�10�mm�a�20�mm�de�espesor,�
de�material�tamaño�arcilla�con�alto�contenido�de�materia�orgánica,�la�cual�define�
el�límite�entre�la�pared�y�el�suelo�que�contiene�a�la�masa�de�cría�(Fig.�13�B).�El�
relleno� esta� compuesto� únicamente� por� fibras� vegetales� de� la� bosta,�
birrefringentes�y�elongadas,�de�aproximadamente�2�mm�de�largo.�Estas�fibras�se�
encuentran�orientadas�de�forma�paralela�a�la�pared�de�la�masa�de�cría,�formando�
una� capa� de� 0,75� mm� a� 1� mm� de� espesor,� mientras� que� pierden� dicha�
orientación� hacia� el� centro� (Fig.� 13� A),� o� en� algunos� casos,� se� encuentran�
formando�meniscos.������

� ���
La�pared�de� la�cámara�ovígera,�de�3,2�mm�a�6,6�mm�de�espesor,�posee�
una�microestructura�maciza�con�una�porosidad�de�5�%�(Fig.�13�C).�La� fracción�
gruesa� representa� el� 45� %� de� la� pared,� es� tamaño� arena� fina� �� limo� grueso�
(selección� moderada),� y� se� encuentra� compuesta� por� cuarzo,� K� �� feldespato,�
plagioclasa,� trizas� volcánicas,� mica� (en� orden� de� importancia),� y� muy� escaso�
piroxeno.� La� fracción� fina� está� compuesta� por� material� tamaño� arcilla� opaco,�
color�castaño�oscuro,�que�refleja�un�alto�contenido�de�materia�orgánica.�
�
�
�!�%�& 25�� Micromorfología� de� masas� de� cría� de� �������� � ������.� �� Fibras�vegetales�del�relleno�orientadas�paralelamente�(fp)�a�la�pared�(p)�de�la�masa�de�cría,�que�pierden� dicha� orientación� hacia� el� interior� distribuyéndose� desordenadamente� (fd);����Capa�delgada�de�material�fino�tamaño�arcilla�con�un�alto�contenido�de�materia�orgánica�(flechas�negras),�que�delinea�el�límite�entre�la�pared�de�la�masa�de�cría�y�el�material�del�suelo;����Pared�de�la�cámara�ovígera.�Escalas:�500�^.��
5.2.2.3���Estructura�del�nido�
Se� estudió� un� total� de� seis� nidos� los� cuales� fueron� hallados� debajo� de�
cuatro� bostas� de� vaca.� Tres� de� ellos� fueron� encontrados� debajo� de� una� sola�
bosta,�mientras�que�de�los�restantes,�se�halló�solo�uno�por�bosta.�Del�análisis�de�
la� estructura� de� estos� nidos,� se� encontraron� distintos� estadios� de� su�

� ���
construcción.� El� primer� estadio� distinguible,� corresponde� a� nidos� compuestos�
únicamente�por�un�túnel�de�almacenamiento�inclinado,�conectado�a�la�superficie�
del�suelo�por�un�extremo,�y�completamente�aprovisionado�por�bosta�depositada�
en�meniscos�desde�el�extremo�opuesto�(Fig.�12�A).�En�algunos�casos�se�encontró�
un� ejemplar� adulto� reposando� sobre� la� bosta� aprovisionada� en� el� túnel.� En� un�
estadio� más� avanzado,� los� nidos� se� encuentran� conformados� por� un� túnel�
vertical,�con�bosta�depositada�desde�la�base�hasta�pocos�centímetros�por�debajo�
de�la�superficie�del�suelo�(Fig.�12�B).�En�esta�etapa�de�construcción�del�nido,�el�
túnel�se�encuentra�conectado�con�el�de�almacenamiento.�En�el�último�estadio,�se�
observa� una� masa� de� cría� completa� construida� en� el� extremo� más� distal� del�
túnel.� Hacia� el� extremo� opuesto,� se�mantiene� la� comunicación� con� el� túnel� de�
almacenamiento,� el� cual� se� conserva� parcialmente� relleno� y� en� el� que� se�
encuentra�el�adulto� (Fig.�12�C�y�D).�Los� tres�nidos�encontrados�debajo�de�una�
misma�bosta�mostraron�dos�de� los� tres�estadios�de� construcción�descriptos:�el�
estadio�intermedio�en�dos�nidos,�y�el�tercer�estadio�en�uno.�
En�los�nidos�completos,�el�túnel�donde�el�adulto�construye�la�masa�de�cría,�
se�caracteriza�por�ser�recto�y�de�posición�vertical,�de�5�–�8�cm�de�largo�y�1,7�–�2�
cm� de� diámetro� (N� =� 3).� Este� túnel� pudo� ser� trazado� hasta� la� superficie� del�
suelo,� a� pesar� de� encontrarse� rellenado� con� bosta� y�material� del� suelo� suelo,�
probablemente�por�el�adulto�una�vez�terminada�la�masa�de�cría�(Fig.�12�C�y�D).�
El� túnel� de� almacenamiento,� de� 10� cm� de� largo� y� 1,3� –� 1,5� cm� de� diámetro,�
posee�una�inclinación�de�45º�respecto�del�túnel�principal,�y�la�comunicación�entre�
ellos� se� ubica� a� 5� –� 6� cm� de� la� superficie� del� suelo� (N�=� 2).� No� se� observan�
características� particulares� en� las� paredes� de� los� túneles.� En� el� túnel� de�
almacenamiento,�la�bosta�se�encuentra�depositada�en�forma�de�meniscos�con�la�

� ���
parte�superior�cóncava�orientada�hacia�el�extremo�más�externo�del�túnel�(Fig.�12�
A).�
�
6�4�5#�� ��������������� ��?�&B��=�A
�
5.2.3.1���Estructura�de�la�bola�de�cría��
Entre�las�bolas�de�cría�examinadas,�se�distinguen�dos�tipos,�unas�cerradas�
y�otras�abiertas.� Las�bolas�de� cría� cerradas�no�poseen�agujero�de�emergencia,�
son� esféricas,� pero� se� estrechan� en� el� polo� superior� adquiriendo� una� forma�
general�de�gota�(Fig.�14�A).�El�eje�mayor,�es�decir�el�alto�de�la�bola�de�cría,�mide�
de�2,5�a�3,5�cm�de�largo,�y�el�eje�menor�o�diámetro�ecuatorial,�de�2,5�a�3,2�cm�
de�largo�(N�=�3).�La�pared�es�continua�en�toda�su�superficie,�esta�compuesta�por�
material� del� suelo�y� fibras�vegetales�que�pueden�verse�desde�el� exterior,� y�en�
aquellos�casos�que�pudo�ser�medida,�posee�un�espesor�que�varia�de�2�a�4�mm�(N�
=�2).�La�superficie�externa�posee�una�textura�particular,�distinguiéndose�en�ella�
depresiones� poco� profundas� que� se� encuentran� irregularmente� distribuidas,�
probablemente�hechas�por�el�adulto�durante�la�construcción,�y�el�modelado,�de�la�
bola� de� cría� (Fig.� 14� A).� El� polo� superior� de� las� bolas� de� cría,� en� el� que�
internamente�se�aloja�la�cámara�ovígera,�posee�un�conjunto�de�fibras�vegetales�
que�atraviesan�toda�la�pared�desde�el�exterior�de�la�bola�de�cría�hacia�el�interior�
de�la�cámara�ovígera,�formando�parte�del�canal�de�aireación�(Fig.�14�A�y�B).�La�
cámara� ovígera� es� esférica,� de� 3� mm� a� 6� mm� de� diámetro� (N� =� 2),� y� se�
encuentra�excavada�en�el�relleno�de�la�bola�de�cría�(Fig.�14�C).�A�ojo�desnudo,�
no� se� distingue� con� claridad� una� pared,� pero� puede� apreciarse� un� tapiz�
constituido�por�un�material�brillante�y�levemente�más�oscuro�que�el�de�la�bosta�
aprovisionada�en� la�bola.�La�cámara�ovígera�no�está�completamente�cerrada,�y�

� ���
en� el� techo� se� observan� las� mismas� fibras� vegetales� que� se� ven� en� el� polo�
superior� externo� de� la� bola� de� cría� (canal� de� aireación).� En� el� interior� de� la�
cámara� ovígera,� ocupando� todo� el� espacio� disponible,� se� encuentra� depositado�
verticalmente�un�huevo�de�color�amarillo�pálido�(Fig.�14�C),�de�6�mm�de�largo�y�
2� mm� a� 3� mm� de� diámetro� (N� =� 1).� El� relleno� se� encuentra� constituido�
principalmente� por� bosta,� aunque� a� simple� vista� puede� observarse� también,�
material�del�suelo�disperso�(Fig.�14�C).��
En� un� ejemplar� colectado� (2,3� cm� de� diámetro� ecuatorial),� también�
cerrado� (Fig.� 14� D),� pero� en� el� cual� la� larva� había� alcanzado� un� estadio� más�
avanzado� del� desarrollo,� se� conservaba� en� su� interior� la� cámara� pupal.� Esta�
cámara� se� compone� de� una� cavidad� interna� esférica,� de� 1� cm� de� diámetro�
(donde�se�conservaba� la�pupa),� y�una�pared�doble�de�3,6�mm�de�espesor� con�
características� particulares.� La� parte�más� externa� de� dicha� pared� (3,3�mm� de�
espesor)�(Fig.�14�E),�se�encuentra�en�contacto�con�la�pared�de�la�bola�de�cría�y�
está�constituida�por�������de�un�material�color�verde�muy�oscuro�y�de�material�
del�suelo,�que�se�acomodan�de�manera�imbricada�formando�un�helicoide�de�6,1�
mm�a�8,4�mm�de�ancho�por�cada�vuelta�del�mismo�(Fig.�14�F).�Este�helicoide�se�
origina�en�la�base�de�la�bola�de�cría,�y�gira�formando�bandas�(Fig.�14�F)�que�se�
superponen� una� sobre� otra� hacia� el� polo� superior� de� la� bola.� En� este� polo,� el�
helicoide�queda�abierto,�dejando�expuesto�el�“techo”�de�la�cámara�pupal�(Fig.�14�
F).� La� superficie� interna� de� la� cavidad� interna� de� esta� cámara,� se� encuentra�
tapizada�por�otro�material� de� color�marrón�grisáceo�de�no�más�de�0,3�mm�de�
espesor,�que�conforma�la�segunda�capa�(Fig.�14�E),�y�la�más�interna,�de�la�pared�
de� la� cámara� pupal.� Esta� pared� se� continua� hacia� el� polo� superior� con� el� ya�
mencionado�“techo”�de�la�cámara,�el�cual�se�compone�de�un�material�color�gris�
mas�claro�y�posee�tanto�interna�como�externamente,�una�textura�particular�(Fig.�

� ���
14� D� y� E).� Hacia� el� lado� interno,� el� material� que� lo� compone� se� encuentra�
depositado�en�forma�de�“ojuelas”�o�“lentejas”�que�se�continúan�hacia�el�exterior�
formando�en�la�superficie�crestas�poligonales�(Fig.�14�D�y�E).��
Las�bolas�de�cría�abiertas�son�esféricas,�de�2,2�cm�a�2,9�cm�de�diámetro�
ecuatorial,�y�poseen�un�agujero�de�emergencia,�de�al�menos�2�cm�de�diámetro�
(N�=�3).�En�este�tipo�de�bolas,�generalmente,�no�se�conserva�la�cámara�ovígera�
y�sólo�en�algunos�casos�aislados�pueden�encontrarse�restos.�Todas�ellas�poseen�
cámara�pupal,�de�1,1�cm�a�1,3�cm�de�diámetro�(N�=�3),�y�conservan�las�mismas�
características�que�las�ya�descriptas�para�las�bolas�de�cría�cerradas�y�con�cámara�
pupal.�La�única�excepción�es�que�en�las�abiertas�no�se�conserva�el�techo�de�dicha�
cámara�ya�que�este�se�rompe�con�la�emergencia�del�nuevo�adulto.�La�pared�de�
este� tipo� de� bolas,� las� cuales� han� sido� abandonadas,� suele� desgastarse� con� el�
tiempo�y�la�desecación,�dejando�expuesta�la�superficie�externa�de�la�espira�que�
conforma�la�parte�más�externa�de�la�pared�de�la�cámara�pupal�(Fig.�14�F).��

� ��
�
�!�%�&28��Nido�y�bolas�de�cría�de���� �����������.���Vista�externa�de�una�bola�de�cría.�Pueden�distinguirse�externamente�depresiones�poco�profundas�e�irregularmente�distribuidas�en�su�superficie�(flecha�blanca)�y�un�conjunto�de�fibras�vegetales�en�el�polo�superior� que� atraviesan� internamente� el� canal� de� aireación� (flecha� negra);� ��� Vista�

� ���
superior� y� externa� del� conjunto� de� fibras� vegetales� en� el� canal� de� aireación� (flecha�negra);���� Corte� de� una� bola� de� cría,� por� el� eje� del� canal� de� aireación,�mostrando� un�huevo� depositado� en� el� interior� de� la� camara� ovígera� (flecha� blanca)� y� el� relleno�constituido� por� bosta� y� material� del� suelo� disperso� (flecha� negra);��#��� Bola� de� cría�cerrada,�en�un�estadio�del�desarrollo�larval�avanzado,�donde�se�distingue�externamente�la�cámara�pupal�conformada�por�una�pared�doble�con�una�capa�interna�(�,�flecha�negra)�y� otra� capa� más� externa� (�,� flecha� blanca),� esta� última� conformada� por� bandas� (�,�flechas�blancas)�que�constituyen�un�helicoide,�y�un� techo�(F,� flecha�negra)� con�crestas�poligonales�(��y��,�flechas�amarillas).�Escalas:�1�cm;��#���Nido�profundo�constituído�por�una�cámara�nido�con�una�bola�de�cría�en�su� interior.�Escalas:�1�cm;����Nido�superficial�constituido� por� una� cámara� nido� (flecha� blanca)� abierta� hacia� un� túnel� curvo� (flecha�negra)�paralelo�a�la�superficie�del�suelo.�Escala:�5�cm;��#���Distintos�tipos�de�túneles.���Dos� tramos,�primero� recto� y� luego� tramo�paralelo� a� la� superficie� relleno� con�bosta,�-�corto� y� oblícuo� con� el� escarabajo� adulto� (flecha� blanca)� retenido� en� un� hueco� creado�entre� la� bosta� aprovisionada,���� Túnel� bifurcado� con� ambas� ramas� rellenas� con� bosta.�Escalas:�5�cm.��
�
5.2.3.2���Micromorfología�de�la�bola�de�cría�
En�la�bola�de�cría�con�cámara�ovígera,�la�pared�posee�un�espesor�que�varía�
de�1�mm�a�4�mm�y�se�compone�tanto�de�material�del�suelo�como�de�bosta�(Fig.�
15�A).�La�abundancia� relativa�de�estos�dos�materiales�es�diferente�en�distintas�
partes� de� la� pared,� encontrándose� fundamentalmente�material� del� suelo� en� la�
parte�más�externa,�y�más�fibras�vegetales�de�la�bosta�hacia�la�parte�más�interna�
(Fig.�15�A).�La�microestructura�de�la�pared�es�maciza�con�una�porosidad�de�5�%,�
que�llega�a�un�25�%�en�la�parte�más�interna,�debido�a�la�incorporación�de�fibras�
vegetales� de� la� bosta.� La� fracción� gruesa� es� tamaño� limo� �� arena� fina� (buena�
selección),� representa� el� 60�%�de� la� pared,� y� está� compuesta� por� cuarzo,�K� ��
feldespato,�plagioclasa,� fragmentos� líticos�y�escasas� trizas�vítreas.�Se�observan�
también�en�esta�fracción,�fibras�vegetales�de�la�bosta,�birrefringentes�y�de�800�^�
de� longitud� promedio,� que� se� hacen� gradualmente� más� abundantes� hacia� el�
relleno.� En� algunas� de� estas� fibras� pueden� distinguirse� fitolitos� en� su� interior�
(Fig.� 15�B).�El� relleno�esta� conformado�en�un�85�%�por� fibras�vegetales�de� la�
bosta�y�materia�orgánica,�y�en�el�15�%�restante�por�fragmentos�de�material�de�
suelo�y�por�granos�minerales�dispersos.�El�material�del� relleno�está�depositado�

� ��
en� capas� concéntricas� de� 1,5� �� 2� mm� de� espesor,� las� cuales� pueden� ser�
identificadas�por�la�orientación�paralela�de�las�fibras�elongadas�de�la�bosta�(Fig.�
15� C).� Hacia� la� zona�mas� interna� del� relleno,� tanto� las� capas� como� las� fibras�
vegetales� pierden� la� orientación,� y� su� distribución� se� hace� más� caótica.� El�
ejemplar� original� conservaba�parte� de� la� cámara�ovígera,� la� cual� fue�destruída�
durante�el�proceso�de�preparación�del�material�para�corte�micromorfológico.��
En�la�bola�de�cría�con�cámara�pupal,�desde�el�exterior�hacia�el�interior,�se�
observa�una�zona�de�1�mm�de�espesor�máximo�con�una�microestructura�maciza�
y�una�porosidad�de�5�%�aproximadamente,�conformada�fundamentalmente�por�
material�del�suelo�(Fig.�15�D).�La�fracción�gruesa�representa�el�25���30�%�de�esta�
zona,�es�tamaño�limo�y�está�compuesta�por�plagioclasa,�K���feldespato,�cuarzo,�
mica� (muscovita),� fragmento� líticos� y� trizas� volcánicas� (en� orden� de�
importancia).�La� fracción� fina�esta�compuesta�por�material� tamaño�arcilla,�muy�
embebido�en�materiales�opacos�(materia�orgánica�y�óxidos).�A�continuación,�se�
distingue�una�zona�intermedia�de�3�mm�de�espesor�máximo,�dentro�de�la�cual�se�
observa� un� aumento� de� la� cantidad� de� fibras� vegetales� de� la� bosta,� desde� el�
exterior�(en�contacto�con�la�zona�más�externa)�hacia�el�interior,�acompañado�de�
una� disminución� relativa� de� material� del� suelo� (Fig.� 15� D).� En� esta� zona�
intermedia� las� fibras� vegetales� presentan� una� orientación� caótica� con� una�
porosidad�de�25�%,�y�el�material�del�suelo�posee� las�mismas�características�ya�
mencionadas� para� la� zona� externa.� Hacia� la� parte� más� interna� de� la� bola,� se�
observa�una�zona�de�2,25�mm�de�espesor�máximo,�con�una�porosidad�del�20�%,�
que� se� reconoce� por� el� ordenamiento� del� material� en� paquetes� de� tamaño�
variable� (8,2� mm� a� 1� cm� de� largo,� y� 200� ^� a� 1,2� mm� de� ancho),� que� se�
yuxtaponen�unos�con�otros�(Fig.�15�D�y�E).�Cada�paquete�está�conformado�por�
abundante�cantidad�de�fibras�vegetales�de� la�bosta,�materia�orgánica,�y�granos�

� ���
minerales� dispersos,� cuya� composición� y� textura� son� similares� a� los� ya�
descriptos.� En� el� interior� de� los� paquetes,� estos� materiales� se� encuentran�
formando� ������ alargados,� con� extremos� ahusados,� yuxtapuestos� y� muy�
ordenados�(Fig.�15�E).�Cada������puede�ser�identificado�por�la�orientación�de�las�
fibras� vegetales,� que� se� acomodan�paralelamente� al�borde.� El� tamaño�máximo�
promedio,�aproximado,�de�estos�������es�de�1,5�mm�de�largo�y�250�^�de�ancho.�
�
�
�!�%�&26�Micromorfología�de�bolas�de�cría�de���� �����������.���Pared�de�la�bola�de�cría�constituida�mayormente�por�material�del�suelo�(flecha�negra)�y,�en�menor�medida,� por� fiibras� vegetales�de� la�bosta� (flechas�blancas).� Escala�50�^;���Detalle� de�una�fibra�vegetal.�Pueden�distinguirse�fitolitos�en�su�estructura�interna�(flechas�blancas).�Escala:�700�^;����Relleno�de�la�bola�de�cría�constituido�mayormente�por�fibras�vegetales�orientadas�en�capas�concéntricas�(flechas�blancas).�Escala:�100�^;����Pared�de�una�bola�de� cría� con� cámara� pupal� donde� se� distinguen� tres� sectores� que� se� diferencian� por� la�composición�y�diistribución�del�material�que�las�componen�(flechas�blancas).�Escala:�0,5�cm;����Detalle�de�los�paquetes�(2)�de������� (flechas�blancas)�que�conforman� la�pared�de�la�cámara�pupal.�Escala:�1,5�mm.��

� ���
5.2.3.3���Estructura�del�nido�
Se�estudió�un�total�de�dieciocho�nidos�los�cuales�fueron�hallados�debajo�de�
nueve�bostas,�cinco�de�vaca�y�cuatro�de�caballo.�Se�encontraron�como�máximo�
cuatro�nidos�por�bosta�(N�=�1),�pero�en�la�mayoría�de�las�bostas�se�hallaron�uno�
(N�=�4),�dos�(N�=�2)�o�tres�(N�=�2).�Del�total�de�nidos�estudiados,�sólo�en�dos�
casos� se� hallaron� cámaras� nido� (Fig.� 14� G� y� H),�mientras� que� en� el� resto� se�
hallaron�distintos�tipos�de�túneles.�En�cuanto�a�las�cámaras�nido,�se�encontraron�
solo� dos,� excavadas� a� distintas� profundidades.� La� más� superficial� (Fig.� 14� I),�
excavada� a� 2� �� 3� cm�de� profundidad,� es� esférica� con� un� eje�mayor� (diámetro�
ecuatorial)�de�3,5�cm�y�uno�menor�de�2,8�cm,�y�contenía�en�su�interior�la�bola�de�
cría�con�el�canal�de�aireación�paralelo�a�la�superficie�del�suelo.�Esta�cámara�esta�
abierta�y�se�comunica�con�un�túnel�lateral�de�1,3�cm�de�diámetro,�vacío,�corto,�
curvo� y� del� cual� no� puede� distinguirse� el� recorrido� completo� (Fig.� 14� I).� La�
segunda�cámara�(Fig.�14�G),�se�encuentra�excavada�a�29�cm�de�profundidad,�es�
esférica,�de�4�cm�de�diámetro,�y�se�encuentra�completamente�cerrada.�La�bola�
de� cría� se� encuentra� alojada� en� su� interior� con� el� canal� de� aireación�
perpendicular� a� la� superficie� del� suelo� y� levemente� inclinado,� respecto� de� la�
vertical�(Fig.�14�H).�
En� la�diversidad�de� túneles�observada,� se�distinguen� tres� tipos:�1)� túnel�
curvo�compuesto�por�dos�tramos�(N�=�7)�(Fig.�14�J),�donde�el�primero�comienza�
desde�la�superficie�del�suelo�con�un�agujero�que�se�comunica�hacia�el�interior�con�
un�tramo�recto�y�perpendicular�de�7,5�cm�de�largo�promedio�(N�=�3),�el�cual�se�
continúa�con�el�segundo�tramo�de�5�cm�de�largo�promedio�(N�=�3)�por�medio�de�
una� curvatura� que� forma� un� ángulo� de� aproximadamente� 45º� con� la� vertical.�
Este� tipo� de� túnel� alcanza� una� profundidad� promedio� de� 15� cm,� y� suele�
encontrarse�en�ellos�a� los�escarabajos�adultos�(N�=�5)�dentro�o�sobre� la�bosta�

� ���
aprovisionada�en�los�últimos�4�cm�del�segundo�tramo�(N�=�3);�2)�túnel�recto�e�
inclinado�(N�=�4)�(Fig.�14�K),�conformado�por�un�solo�tramo�de�15�cm�de�largo�
promedio� (N� =� 3)� que� comienza� en� la� superficie� del� suelo� con� un� diámetro�
promedio�de�1,5�cm�(N�=�2)�y�que�se�ensancha�en�1�cm�aproximadamente�hacia�
su� extremo� más� distal.� La� característica� más� llamativa� de� este� tipo� de� túnel�
radica� en� el� relleno,� ya� que� en� la� mayoría� (N� =� 3)� el� relleno� se� encuentra�
depositado� en� los� últimos� 3� �� 4� cm� del� extremo� mas� distal� y� también� en� el�
extremo� superior,� dejando� un� hueco� entre� ellos� de� 3� �� 4� cm� de� largo,� donde�
suele�encontrarse�una�hembra�activa� (N�=�3)� (Fig.�14�K);�y�3)� túnel�bifurcado�
(Fig.�14�L),�en�el�cual�las�dos�ramas�del�túnel�se�parecen�entre�sí,�y�también�se�
asemejan�en�tamaño�y� forma�a� los� túneles�descriptos�en�el� item�anterior�(N�=�
3).� Las� dos� ramas� poseen� un� tercio� de� su� capacidad� ocupado� por� bosta�
aprovisionada� en� forma� de�meniscos.� En� dos� de� los� tres� casos� observados,� se�
encontró�un�escarabajo�adulto�alojado�sobre�la�bosta�aprovisionada�en�una�de�las�
dos�ramas.��
6�4�8#�� ��������������������?�K�+��'&=A
�
5.2.4.1���Estructura�de�la�bola�de�cría�
Todas� las� bolas� de� cría� estudiadas� (N� =� 5)� poseen� una� protuberancia�
externa�en�el�polo�superior�(donde�internamente�se�aloja�la�cámara�ovígera),�que�
se� distingue� claramente� de� la� masa� central� esférica� por� la� presencia� de� un�
angostamiento� o� cuello� entre� ambas� estructuras� (Fig.� 16� A� y� B).� El� mayor�
diámetro�ecuatorial�medido�alcanza�4,9�cm,�y�el�menor�2,5�cm�(N�=�5),�mientras�
que�el�eje�vertical,�que�pasa�por�la�cámara�ovígera,�alcanza�4,5�cm�como�valor�
máximo,�y�2,5�cm�como�mínimo�(N�=�5).�La�protuberancia�mide�1,2�cm�a�1,6�cm�

� ���
de� diámetro� en� la� base,� 4,4� cm� a� 6� mm� de� alto� (N� =� 4),� y� se� angosta�
aproximadamente�0,5�cm�en�su�extremo�más�distal,�donde�en�el�centro�se�abre�
una� perforación� de� 3� mm� a� 5� mm� de� diámetro� (N� =� 2).� Esta� perforación� no�
posee� fibras� vegetales,� ni� ninguna� otra� estructura� que� la� obture,� y� en� los�
ejemplares�con�cámara�ovígera,�puede�verse�el�huevo�en�su�interior�a�través�de�
ella�(Fig�16�B).�Las�bolas�de�cría�poseen�una�pared�externa�de�material�del�suelo,�
que� en� una� bola� con� cámara� ovígera� mide� 2,3� mm� de� espesor� (Fig.� 16� C),�
mientras�que�en�las�bolas�con�cámara�pupal�el�espesor�cambia,�haciéndose�más�
gruesa�hacia�el�polo�superior�(de�1,5�a�4,3�mm�promedio,�N�=�2).�Tanto�en�las�
bolas� de� cría� con� larva� en� desarrollo,� como� en� aquéllas� en� las� que� la� cámara�
pupal�ha�sido�construida�completamente,�se�observa�pegado�a�la�pared�material�
del�relleno�en�su�estado�original�que�no�ha�sido�utilizado�por� larva.�En�el�único�
ejemplar�donde�se�conservó�intacta�la�cámara�ovígera,�esta�es�esférica,�de�1�cm�
de�diámetro,�y�se�encuentra�abierta�en�el�polo�superior�por�una�perforación�que�
comunica�el�interior�de�dicha�cámara�con�el�exterior�de�la�bola�(Fig.�16�C�y�D).�La�
cámara�posee�una�pared�de�material�oscuro,�de�1,5�mm�de�espesor,�que�la�aísla�
de� la� provisión� y� que� en� el� polo� superior� se� pone� en� contacto� con� la� pared�
externa�de�la�bola,�sugiriendo�un�origen�diferente�para�ambas�(Fig.�16�D).�En�el�
interior� de� la� cámara� ovígera,� se� encuentra� depositado� horizontalmente,� un�
huevo�de�color�amarillo�pálido,�de�9�mm�de�largo�y�3���4�mm�de�diámetro�(N�=�
1)� (Fig.� 16� C� y� D).� El� relleno� se� ve,� a� ojo� desnudo,� conformado�
fundamentalmente� por� bosta� sin� un� arreglo� particular� (Fig.� 16� C).� En� los� dos�
ejemplares�con�larvas�en�desarrollo,�solo�se�observa�parte�de�la�cámara�ovígera,�
y� la�pared�de�la�futura�cámara�pupal,�en�diferentes�estadios�de�su�construcción�
(Fig.�16�E).�Esta�pared�se�distingue�claramente�por�una�fisura�que�se�forma�entre�
ella�y�el�material�remanente�del�relleno�(que�se�conserva�pegado�a�la�pared�de�la�

� ���
bola),� en� los� ejemplares� que�han�perdido� la� humedad�original� (Fig.� 16�E� y� F).�
Puede�observarse�en�el�corte�de�estos�ejemplares,�que�esta�pared�se�hace�cada�
vez�más�gruesa,� de� bordes�más� redondeados,� y� de� forma�específica� a�medida�
que� la� larva� va� completando� su� elaboración� (Fig.� 16� E).� El� material� que� la�
conforma�no�se�reconoce�a�simple�vista,�pero�sí�se�lo�distingue�más�oscuro�que�el�
resto� del� relleno.� En� los� dos� ejemplares� restantes,� sólo� parte� del� techo� de� la�
cámara� ovígera� se� encuentra� preservado,� pero� se� conserva� intacta� la� cámara�
pupal,� ya�que� se� trata�de�bolas�de� cría�de� las� cuales�el�nuevo�adulto�no� logró�
emerger�(Fig.�16�G).�Estos�datos�se�lograron�cortando�las�bolas�por�el�eje�de�la�
protuberancia.�Una�vez�abiertas�se�observaron�las�cámaras�pupales,�que�poseen�
una� cavidad� interna� de�1� cm�a�1,4� cm�de�diámetro� y�una�pared�que� tiene�un�
espesor�de�3�mm�a�5�mm�promedio�(N�=�2)�(Fig.�16�G).�La�pared�de�las�cámaras�
pupales� posee� dos� capas,� una�más� interna,� delgada� y� de� espesor� uniforme,� y�
otra�más� externa,� gruesa� y� de� espesor� irregular� (Fig.� 16� G).� La� capa� externa�
posee� un� arreglo� particular� del� material,� con� ������ imbricados� que� forman�
bandas�de�0,7�cm�a�1�cm�de�ancho�(N�=�3),�de�un�helicoide�que�puede�verse�con�
claridad�desde�el�exterior,�al�remover�la�pared�de�la�bola�de�cría�(Fig.�16�F).�En�el�
corte�de�la�bolas�de�cría�se�observa�que�hacia�el�exterior�del�helicoide,�la�pared�
de�la�bola�y� los�restos�del�relleno�que�no�han�sido�usados�por�la�larva�y�que�se�
mantienen�unidos�a�esta,�se�separan�de�dicha�espira�por�medio�de�una�fisura�que�
se�forma�entre�ellos�(Fig.�16�F).�Por�ello,�la�pared�de�las�bolas�de�cría�puede�ser�
removida�fácilmente�sin�que�se�rompa�la�capa�más�externa�de�la�cámara�pupal,�
es�decir,�el�helicoide.�En�solo�uno�de�estos�dos�ejemplares,�se�conserva�el�techo�
de�la�cámara�pupal,�mientras�que�en�el�otro�está�ausente�debido,�posiblemente,�
a�que�el�adulto�alcanzó�a�romperlo�antes�de�morir.�En�el�primer�caso,�este�techo�
se� localiza� justo�por�debajo�de� los� restos�de� la� cámara�ovígera,� separados�por�

� ���
una�cavidad�hueca,�y�se�encuentra�conformado�por�un�material�que�se�diferencia�
en�color�y�textura�del�resto�del�material�que�conforma�la�capa�interna�de�la�pared�
de�la�cámara�pupal�(Fig.�16�G�y�H).�La�superficie� interna�del�techo�es�rugosa�y�
hacia� el� lado� externo� posee� pequeñas� crestas� formadas� por� el�mismo�material�
(Fig.�16�H).����
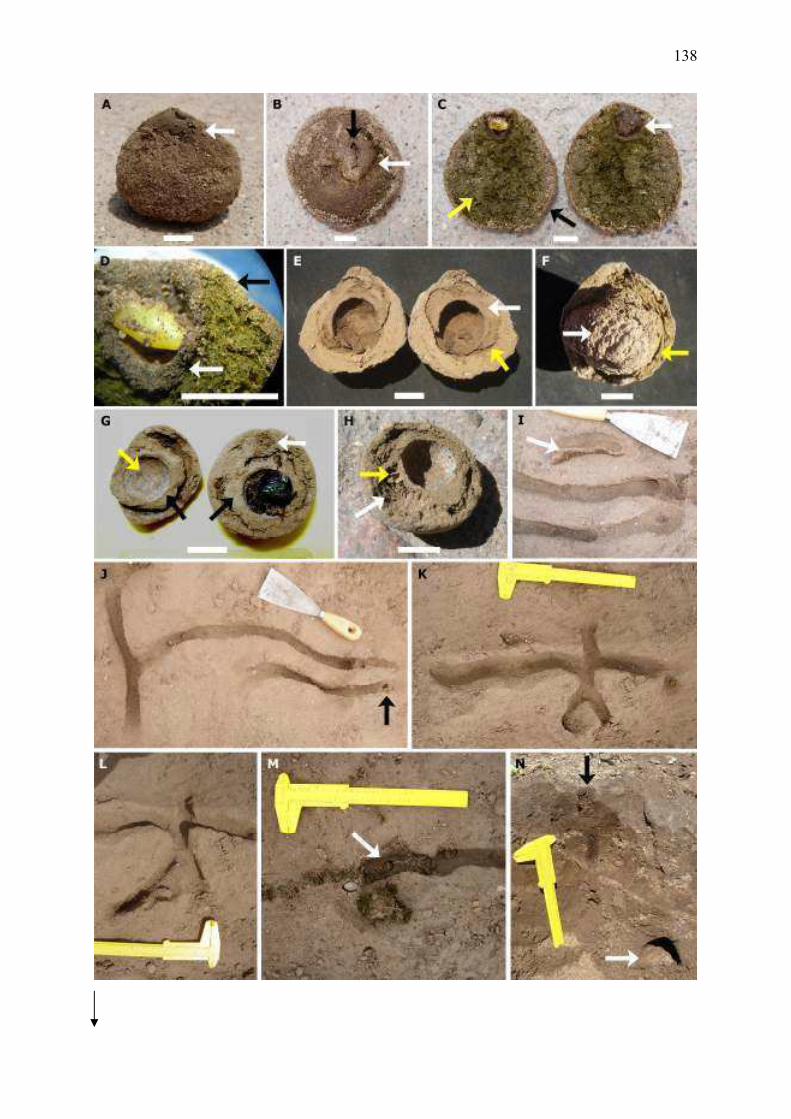
� ���
�

� ��
�!�%�&23�Nido�y�bolas�de�cría�de���� ��������������.#���Vista�externa�lateral�()� y� superior� (�)� de� una� bola� de� cría.� Se� distingue� la� protuberancia�marcada� por� un�angostamiento�de� la�pared�(flechas�blancas)�y�el�huevo�a�través�del�canal�de�aireación�(�,� flecha�negra);���� Vista� interna�de�una�bola�de� cría� cortada�por� el� eje�del� canal� de�aireación.�Se�distinguen�la�pared�(flecha�negra),�la�cámara�ovígera�(Flecha�blanca)�con�el�huevo�y�el�relleno�(flecha�amarilla);����Detalle�de�la�cámara�ovígera�donde�se�destaca�la�pared�(flecha�blanca)�constituida�por�un�material�más�oscuro�y�diferente�al�de�la�pared�de�la�bola�de�cría�(flecha�negra);����Bola�de�cría�en�un�estadio�avanzado�del�desarrollo�larval,�cortada�por�el�eje�del�canal�de�aireación.�Se�distingue�la�pared�de�la�cámara�pupal�en�construcción�(flecha�blanca)�separada�de�la�pared�de�la�bola�y�restos�del�relleno�por�medio� de� una� fisura� (flecha� amarilla); ��� Vista� externa� de� la� pared� helicoidal� de� la�cámara� pupal� separada� por� una� fisura� (flecha� amarilla)� del� resto� del� material.� Se�distingue� en� primer� plano,� una� banda� del� helicoide� conformada� por� ������ (flecha�blanca);���Bola�de�cría�cortada�por�el�eje�del�canal�de�aireación,�con� la�cámara�pupal�intacta�y�un�adulto�no�emergido�conservado�en�su�interior.�Se�dintinguen�la�parte�externa�de�la�pared�de�la�cámara�pupal�(flechas�negras)�y�la�superficie�interna�(flecha�amarilla),�como� asi� también� la� porción� superior� de� la� cámara� ovígera� (flecha� blanca);���� Vista�superior� y� externa� del� techo� de� la� cámara� pupal� con� crestas� (flecha� amarilla)� que� se�extienden� hacia� el� hueco� que� la� separa� de� los� restos� de� la� cámara� ovígera� (flecha�blanca);� Escalas:� 1� cm;��#��� Nido� conformado� por� dos� túeneles� conectados� en� “T”,� Se�distingue� la� bosta� aprovisionada� en� el� extremo� más� distal� de� uno� de� ellos� (�,� flecha�blanca),� y� un� tercer� túnel� paralelo� con� una� hembra� (�,� flecha� negra);� -#��� Nido�conformado�por�dos�túneles�principales�que�se�cruzan�formando�una�“X”,�uno�se�bifurca�en� uno� de� sus� extremos� (-� y� �)� y� el� otro� posee� una� hembra� junto� con� la� bosta�aprovisionada� (�,� flecha�blanca);����Nido� conformado�por�una� cámara� cerrada� (flecha�blanca)�y�un�túnel�relleno�con�material�del�suelo�(flecha�negra)�que�se�comunica�con�la�superficie.��������
5.2.4.2���Micromorfología�de�la�bola�de�cría�
En�la�bola�de�cría�con�cámara�ovígera,� la�pared�posee�un�espesor�de�3,4�
mm�a�7�mm,�y�pueden�distinguirse�dos�sectores,�uno�más�externo�y�delgado�de�
800�^�de�espesor�y�un�sector�más�interno�que�ocupa�el�resto�de�la�pared�(Fig.�17�
A� y� B).� La� pared� se� compone� tanto� de�material� del� suelo� como� de� bosta� y� la�
abundancia�relativa�de�estos�dos�componentes�difiere�en�las�distintas�partes�de�
la�pared�(Fig.�17�B).�La�microestructura�de�la�parte�mas�externa�de�la�pared�es�
maciza�con�una�porosidad�de�5�%�(Fig.�17�B).�La�fracción�gruesa�es�tamaño�limo�
�� arena� muy� fina,� representa� un� 80%� de� esta� parte� de� la� pared� y� esta�
compuesta,� en� orden� de� importancia,� por� mica� (biotita� y� muscovita),� cuarzo,�
plagioclasa�y�minerales�pesados�(piroxeno,�etc.).�En�esta�fracción�de�la�pared�se�
observan� muy� dispersas� fibras� vegetales� de� la� bosta.� La� fracción� fina� esta�
compuesta� por� material� arcilloso,� opaco,� y� por� materia� orgánica.� La�

� ���
microestructura�de�la�parte�mas�interna�de�la�pared�es�simple,�con�granos�sin�un�
arreglo�específico�y�con�una�porosidad�de�25�%�(Fig.�17�B).�La�fracción�gruesa�es�
tamaño�arena�fina���arena�media,�representa�el�90�%�de�esta�parte�de�la�pared�y�
esta� compuesta� por� fragmentos� líticos,� plagioclasa,� K� �� feldespato,� cuarzo,� y�
mica�(biotita�y�muscovita).�Esta�fracción�también�posee�materia�orgánica�y�fibras�
vegetales�de�la�bosta,�pero�en�mayor�proporción�que�en�la�porción�externa�de�la�
pared.�Se�trata�principalmente�de�fibras�elongadas,�birrefringentes,�de�120�^�de�
longitud� y� dentro� de� las� cuales� pueden� verse� fitolitos.� Se� encuentran�
parcialmente� orientadas� de� forma� paralela� a� la� parte� externa� de� la� pared.� La�
fracción�fina�es�arcillosa�y�se�encuentra�depositada�mayormente�alrededor�de�los�
granos.�El� relleno�esta� formado�por������� lenticulares�y�yuxtapuestos�(Fig.�17�
C),�que�muestran�una�porosidad�del�15�%.�Están�compuestos�principalmente�por�
materia� orgánica� y� fibras� de� bosta,� y� una� fracción� mineral� muy� pobremente�
seleccionada�tamaño�limo�–�arena�gruesa.�Esta�fracción�representa�el�60�%�del�
relleno�y�esta� compuesta�principalmente�por� fragmentos� líticos,�mica� (biotita�y�
muscovita),�K���feldespato,�plagioclasa,�y�cuarzo.�Hacia�el� interior�de�la�bola�de�
cría,�se�observa�una�cavidad�dentro�del�relleno,�en�la�cual�se�encontraba�la�larva�
en� desarrollo,� que� esta� parcialmente� tapizada� por� materia� orgánica,�
probablemente�heces�de�la�larva�(Fig.�17�D).��
La�pared�de� la�cámara�ovígera�posee�un�espesor�de�1,75�mm�y�muestra�
las� mismas� características� micromorfológicas� que� las� descriptas� previamente�
para� la�parte�externa�de� la�pared�(Fig.�17�E).�La�fracción�gruesa�de�esta�pared�
posee� también� materia� orgánica� y� fibras� vegetales� de� la� bosta,� en� muy� baja�
proporción.�El�material�que�la�conforma�se�dispone�en�forma�de�grandes�������
lenticulares�que�pueden�ser�identificados�por�la�orientación�de�los�granos�de�mica�
al�usar�luz�polarizada�(Fig.�17�F) ��

� ��
�En� la�bola�de� cría� con� cámara�pupal� (Fig.�17�G),� la�pared�posee�similar�
microestructura,�composición�y�textura�que�las�ya�descriptas�para�la�bola�de�cría�
con�cámara�ovígera.�Sin�embargo,�entre�la�pared�de�la�bola�de�cría�y�la�capa�más�
interna�de�la�pared�de�la�cámara�pupal,�se�distinguen�más�claramente�los�������
lenticulares�y�yuxtapuestos�(Fig.�17�H)�de�la�capa�más�externa�de�dicha�pared,�
los�cuales�presentan�una�porosidad�del�15�%.�Están�compuestos�principalmente�
por�materia�orgánica�y�fibras�vegetales�de�la�bosta,�y�una�fracción�mineral�muy�
pobremente�seleccionada�tamaño�limo�–�arena�media.�Esta�fracción�representa�el�
40�%�y�esta�compuesta�principalmente�por�mica� (biotita�y�muscovita),� cuarzo,�
plagioclasa,�K���feldespato�y�fragmentos�líticos.�La�parte�más�interna�de�la�pared�
de�la�cámara�pupal�posee�un�espesor�que�mide�de�150�^�a�625�^�y�recubre�2/3�
de� la� superficie� interna� de� dicha� cámara.� Está� compuesta� también� por� ������
lenticulares� y� yuxtapuestos� (Fig.� 17� I),� pero�más� pequeños� que� los� descriptos�
para�el� relleno.�Los�������de� la�pared�poseen�una�porosidad�del�15� ��20�%�y�
están� compuestos� por� materia� orgánica� y� fibras� vegetales� de� la� bosta,� y� una�
fracción� mineral� pobremente� seleccionada� tamaño� limo� –� arena� fina.� Esta�
fracción� representa�el�25� ��30�%�de� la�pared�y�esta�compuesta�principalmente�
por� mica� (biotita),� cuarzo,� K� �� feldespato,� plagioclasa� y� fragmentos� líticos.� El�
techo�de�la�cámara�pupal�posee�un�espesor�de�1�mm�a�2,08�mm,�recubre�1/3�de�
la� cámara,� y� presenta� un� límite� neto� tanto� con� el� material� de� la� pared� de� la�
cámara� pupal� como� con� el� del� relleno� (Fig.� 17� J� y� K).� Presenta� una�
microestructura�maciza,�con�una�porosidad�de�10�%.�El�material�se�dispone�en�
paquetes� triangulares� no� muy� bien� definidos,� los� cuales� se� extienden� en�
prolongaciones�aciculares�hacia�el�espacio�hueco�existente�entre�éste�y�el�techo�
de� la� cámara� ovígera� (Fig.� 17� K).� La� fracción� gruesa� es� tamaño� limo� –� arena�
gruesa� pobremente� seleccionada,� representa� un� 35� %� y� esta� compuesta� por�

� ���
fragmentos� líticos,� mica� (biotita� y� muscovita),� cuarzo,� K� �� feldespato� y�
plagioclasa.�La� fracción� fina�está�compuesta�por�material�arcilloso,�opaco�y�por�
materia�orgánica.�En�este�ejemplar�se�preservan�restos�de�la�pared�de�la�cámara�
ovígera� (Fig.� 17� L)� los� cuales� muestran� la� mismas� características�
micromorfológicas� que� las� descriptas� para� el� techo� de� la� cámara� pupal,� solo�
difieren�en�la�porosidad,�siendo�menor�en�la�última.��
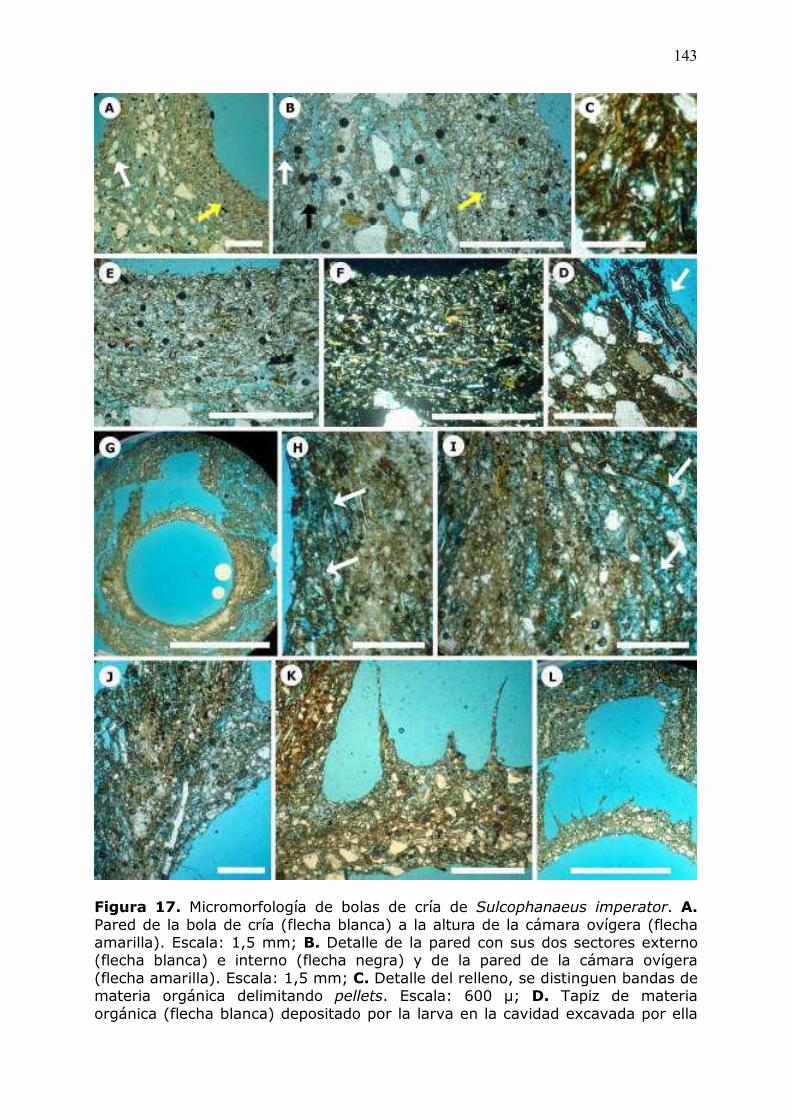
� ���
�
�!�%�& 2;�Micromorfología� de� bolas� de� cría� de� ��� ������� �������.��Pared�de�la�bola�de�cría�(flecha�blanca)�a�la�altura�de�la�cámara�ovígera�(flecha�amarilla).�Escala:�1,5�mm;��Detalle�de� la�pared�con�sus�dos�sectores�externo�(flecha� blanca)� e� interno� (flecha� negra)� y� de� la� pared� de� la� cámara� ovígera�(flecha�amarilla).�Escala:�1,5�mm;��Detalle�del�relleno,�se�distinguen�bandas�de�materia� orgánica� delimitando� �����.� Escala:� 600� ^;� ��� Tapiz� de� materia�orgánica�(flecha�blanca)�depositado�por�la�larva�en�la�cavidad�excavada�por�ella�

� ���
en�el� relleno�de� la� bola� de� cría.� Escala:� 1�mm; �#��Detalle� de� la� pared�de� la�cámara� ovígera� donde� se� distinguen� los� granos� de� mica� paralelamente�orientados,�al�observarlos�sin�?�Ay�con?�A luz�polarizada.�Escala:�1,5�mm;��Bola�de�cría�con�cámara�pupal�(cavidad�esférica�y�central)�que�preserva�parte�de�la�cámara�ovígera�(cavidad�hemiesférica�y�superior).�Escala:�1�cm;��Detalle�de�la� capa�más� interna� de� la� pared� de� la� cámara� pupal.� Se� ditinguen� los� ������delimitados�por�bandas�de�materia�orgánica�(flechas�blancas).�Escala:�350�^;��Detalle�de� la�capa�más�externa�de� la�pared�conformada�por�pellets� lenticulares�delimitados�por�bandas�de�materia�orgánica�(flechas�blancas).�Escala:�350�^;��Contacto�entre�las�dos�capas�de�la�pared�de�la�cámara�pupal�y�el�techo.�Escala:�1�mm;-�Detalle�del�techo�de�la�cámara�pupal�conformado�por�������triangulares�que� se� extienden� en� proyecciones� aciculares.� Escala:� 1� mm; �� Techo� de� la�cámara� pupal� (inferior)� separado� por� una� cavidad� hueca� de� los� restos�preservados�de�la�cámara�ovígera�(superior).�Escala:�5�mm.����
5.2.4.3���Estructura�del�nido�
Se�estudió�un�total�de�seis�nidos,�todos�ellos�hallados�debajo�de�bostas�de�
vaca,�de�los�cuales�cuatro�se�encuentran�constituidos�solo�por�túneles,�con�y�sin�
bosta,� y� en� los� dos� restantes� se� encontraron�además,� las� cámaras�nido.� En� el�
primer� tipo� de� nido,� solo� dos� se� encuentran� constituidos� por� una� galería� de�
túneles,� mientras� que� los� restantes� son� túneles� inconexos� y� dispersos� en� el�
terreno.� De� las� dos� galerías,� una� de� ellas� se� encuentra� constituida� por� tres�
túneles�excavados�en�su�origen�a�muy�poca�profundidad�(3���5�cm),�de�los�cuales�
dos�se�conectan�entre�sí�formando�una�“T”�paralela�a�la�superficie�(Fig.�16�J).�La�
rama�menor� de� esta� estructura� es� recta,� de� 2,5� cm� de� diámetro� y� 26� cm� de�
largo,�pero�hacia�la�mitad�de�su�recorrido,�se�hace�más�profunda�acercándose�a�
la�superficie�nuevamente�en�el�extremo�opuesto�donde�se�encontraron�restos�de�
bosta� removidos.� Aproximadamente,� a� la� mitad� de� su� largo,� en� la� zona� más�
profunda,�esta�rama�se�comunica�con�la�mayor,�de�56�cm�de�largo�y�2,7�cm�de�
diámetro,�la�cual�posee�un�recorrido�sinuoso�y�alcanza�una�profundidad�máxima�
de� 20� cm.� Este� extremo� del� túnel,� posiblemente� de� almacenamiento,� se�
encuentra�ocupado�por�10�cm�de�bosta�(Fig.�16�I).�El�tercer�túnel�del�nido,�de�28�
cm�de�largo�y�3�cm�de�diámetro,�se�encuentra�excavado�paralelamente�y�a�5�cm�

� ���
de�la�rama�mayor,�y�con�la�misma�forma�y�recorrido�que�esta�(Fig.�16�J).�Hacia�
el� extremo� más� distal� de� este� túnel,� se� observan� restos� de� bosta� y� una�
bifurcación� corta� dirigida� hacia� la� zona�más� profunda� donde� se� encuentra� una�
hembra� junto�a� la�bosta�prolijamente�aprovisionada�(túnel�de�almacenamiento,�
Fig.�16�J).�La�segunda�galería�se�encuentra�constituida�por�cuatro�túneles,�de�los�
cuales�tres�se�conectan�entre�sí�formando�una�“cruz”�(poco�profunda�y�paralela�a�
la�superficie,�Fig.�16�K),�y�uno�de�ellos�se�bifurca� lateralmente�dando�origen�al�
cuarto�túnel.�El�túnel�principal�(de�almacenamiento)�es�relativamente�recto,�con�
poca�sinuosidad,�de�36�cm�de�largo�y�2,5���3�cm�de�diámetro,�y�posee�en�uno�de�
sus�extremos�una�hembra�junto�a�20�cm�de�bosta�(Fig.�16�M)�aprovisionada�en�
grandes�paquetes�meniscados�de�aproximadamente�1�cm�de�espesor.�El�extremo�
opuesto,� levemente� curvado,� posee� 3� cm�de� bosta� aprovisionada� de� la�misma�
manera.�A�dos�tercios�de�su�largo�total,�el�túnel�principal�se�comunica,�hacia�un�
lado�y�con�un�ángulo�de�45º,�con�un�túnel�recto�y�corto,�de�aproximadamente�7�
cm� de� largo,� aprovisionado� con� bosta� desde� su� origen� hasta� 6� cm� de�
profundidad,� donde� se� encuentra� alojado� un�macho.� Hacia� el� lado� opuesto,� el�
túnel�principal�se�comunica�con�otro�túnel,�bifurcado,�una�de�las�ramas�(8�cm�de�
largo)�con�su�entrada�bloqueada�por�restos�de�bosta�desmenuzada,�y�la�otra�(12�
cm�de� largo)�más�profunda�y�vacía� (Fig.� 16�L).� Las�tres� ramificaciones�poseen�
cierta�inclinación�respecto�de�la�superficie,�y�alcanzan�mayor�profundidad�que�el�
túnel�principal,�que�se�mantiene�paralelo�a�la�superficie�en�todo�su�recorrido.�
De�los�dos�nidos�con�cámara,�en�el�más�incompleto�de�ellos,�pudo�trazarse�
un�túnel�recto�de�2,2�cm�de�diámetro�y�12�cm�de�profundidad,�relleno�desde�su�
origen�con�5�cm�de�material�del�suelo�removida�y�hacia�el�extremo�más�distal,�
con�7�cm�de�material�del�suelo�compactado�(Fig.�16�M).�A�menos�de�10�cm�de�
distancia�de�este�túnel,�y�a�una�profundidad�de�32�cm,�se�ubica�la�cámara�nido�

� ���
(Fig.� 16� M)� justo� en� el� límite� entre� dos� horizontes� del� suelo.� No� se� observa�
conexión�entre�estas�dos�estructuras,�y� la�cámara�no�se�comunica�con�ninguna�
otra.�La�cámara�es�esférica,�de�6,2���6,5�cm�de�diámetro,�y�en�su�interior�posee�
una� bola� de� cría� muy� mal� preservada.� El� nido� más� completo� se� encuentra�
constituido�por�un�túnel�vertical�que�se�abre�justo�por�debajo�de�la�bosta�y�que�
alcanza�una�profundidad�de�12�cm,�donde�se�continúa�con�un�tramo�paralelo�a�la�
superficie,�de�24�cm�de�largo,�que�desemboca�en�la�parte�superior�y�lateral�de�la�
cámara�nido.�La�cámara�es�esférica,�de�9�cm�de�diámetro�aproximadamente,�y�
dentro�se�aloja�una�bola�de�cría�intacta�con�el�canal�de�aireación�orientado�hacia�
la�superficie�del�suelo.�Cerca�de� la�base�de� la�cámara�nido,�se�abre�otro� túnel,�
inclinado,�de�4�cm�de�profundidad,�en�el�cual�se�encuentra�un�escarabajo�adulto�
junto� con� bosta� aprovisionada� en� su� extremo� más� distal.� Debido� a� las�
condiciones� muy� secas� del� suelo,� las� distintas� partes� del� nido� se� fueron�
desmoronando�con�la�excavación,�y�no�pudieron�ser�correctamente�fotografiadas.������
�
6�4�6#�� �����������������?�&��'$A
�
5.2.5.1���Estructura�de�la�bola�de�cría�
Las�bolas�de�cría�estudiadas�poseen�una�forma�general�de�“gota”�(Fig.�18�
A),�con�un�eje�mayor�de�3,7�cm�de�largo�(N�=�2),�es�decir�el�alto�de�la�bola,�y�un�
eje�menor�de�3,3�cm�(N�=�2),�medido�en�el�ecuador�de�la�bola,�perpendicular�al�
canal�de�aireación.�En�la�parte�superior�de�estas�bolas,�en�el�polo�más�estrechado�
de�la�“gota”,�pueden�distinguirse�externamente�fibras�vegetales�agrupadas�de�la�
bosta� (Fig.� 18� � A� y� B),� formando� un� canal� de� aireación� de� casi� 8� mm� de�
diámetro.�Estas�fibras�atraviesan�la�pared�de�la�bola�de�cría�y�se�comunican�con�
el� interior�de� la�cámara�ovígera� (Fig.�18�B).�Dado�que�cuando� fueron�halladas,�

� ���
cada� bola� de� cría� contenía� en� su� interior� una� larva� en� desarrollo� (Fig.� 18� B),�
muchas�de�las�características�internas�no�pudieron�ser�registradas�como�en�otros�
casos.�La�pared�de�una�de� las�bolas�de�cría,�constituida�por�material�del�suelo,�
mide� 1� mm� de� espesor� aproximadamente,� y� vista� a� ojo� desnudo,� la� bosta�
aprovisionada�en�su�interior�no�muestra�un�arreglo�particular.�En�esta�bola,�sólo�
se�preserva�el�techo�de�la�cámara�ovígera�(Fig.�18�B),�y�en�el�interior�del�relleno�
se�observa�una�cavidad�esférica�de�1,6�cm�de�diámetro,�tapizada�por�un�material�
oscuro�de�250�^�de�espesor�aproximadamente,�donde�se�hallaba�la�larva�(Fig.�18�
B).��
�

� ���
�!�%�&2:��Nido�y�bolas�de�cría�de���� �����������.��Bola�de�cría�en� forma�de�“gota”.�Se�distinguen�las�fibras�vegetales�del�canal�de�aireación�saliento�del�polo�superior�de� la� bola� (flecha� amarilla);���Bola� de� cría� cortada� por� el� eje� del� canal� de� aireación�(flecha� amarilla),� donde� se� distinguen� restos� de� la� cámara� ovígera� (flecha� negra)� y� la�cámara� pupal� (flecha� blanca)� con� la� larva� en� su� interior.� Escala:� 1� cm;���� Relleno� de�túnel� corto� con� un� adulto� en� su� interior.� Escala:� 3� cm; �#��� Túneles� largos�aprovisionados� parcialmente� con� bosta� y� con� un� adulto� en� el� interior� (flecha� blanca).�Escala:�5�cm.���
5.2.5.2���Micromorfología�de�la�bola�de�cría�
En� la� bola� de� cría� con� larva� en� desarrollo,� de� afuera� hacia� adentro,� se�
observan� unas� pocas� porciones� de� material� del� suelo� cubriendo� su� superficie.�
Más� hacia� el� interior,� se� observa� una� zona� de�material� conformado� por� fibras�
vegetales�de�la�bosta�y�escasa�cantidad�de�minerales�(Fig.�19�A).�La�porosidad�es�
de� 15� �� 20�%� y� la�microestructura� esta� determinada�por� la� disposición� de� las�
fibras� vegetales.� La� fracción� gruesa� representa� el� 50� %� de� este� material,� es�
tamaño�limo,�alcanzando�algunos�pocos�granos,�tamaño�arena.�Está�compuesta�
por�fibras�vegetales�de�la�bosta,�alargadas,�de�500�^�a�2,5�mm�de�largo,�y�por�K�
�� feldespato,� fragmentos� líticos,� cuarzo� y� plagioclasa.� La� fracción� fina� está�
compuesta� por� material� tamaño� arcilla,� opacado� por� materia� orgánica.� En�
algunas�zonas,�ésta�se�concentra�formando�bandas�más�o�menos�paralelas�entre�
si,� y� a� la� superficie� externa� de� la� bola� (Fig.� 19� A),� donde� además� las� fibras�
vegetales�de�la�bosta�se�orientan�conforme�a�dichas�bandas.�Más�hacia�el�interior�
de�la�bola�de�cría,�se�observan�dos�paquetes�en�forma�de�medialuna,�los�cuales�
se� encuentran� imbricados� y� conformados� por� ������ (Fig.� 19� A).� En� estos�
paquetes� la� porosidad� es� del� 5� %� y� la� microestructura� está� eminentemente�
determinada�por� la�distribución�de� las� fibras�vegetales�de� la�bosta.� La� fracción�
gruesa� representa� el� 35� %� y� está� compuesta,� fundamentalmente,� por� fibras�
vegetales� de� la� bosta� y� por� escasos� minerales� dispersos� (K� �� feldespato� y�
cuarzo),�tamaño�limo.�Las�fibras�vegetales�son�elongadas,�de�200�^�a�2,5�mm�de�
largo,�y�se�distribuyen�paralelamente�a�los�bordes�de�los�paquetes�de������.�La�

� ��
fracción� fina� esta� compuesta� por� arcilla� teñida� profusamente� por� materia�
orgánica�castaño�rojiza�a�negra�(Fig.�19�B).��
Hacia�uno�de� los�polos�de� la�bola�de�cría,� se�observa�material�del� suelo,�
acompañado�de�fibras�vegetales�de�la�bosta,�concentrado�en�una�hemiesfera�de�
7,8�mm� de� diámetro� (Fig.� 19� C),� la� cual� se� corresponde� con� los� restos� de� la�
cámara� ovígera.� En� el� interior� de� esta� semiesfera,� el� material� se� encuentra�
depositado� en� capas� concéntricas� (Fig.� 19� C).� La� porosidad� de� dicho�material,�
aumenta� de� 10� %� a� 20� %,� hacia� el� interior� de� la� bola� de� cría,� debido� a� la�
disminución�del�material�del�suelo.�La�fracción�gruesa�representa�el�30�%�y�esta�
compuesta,�fundamentalmente,�por�minerales�tamaño�limo���arena�gruesa,�tales�
como� K� �� feldespato,� plagioclasa,� fragmentos� líticos� y� cuarzo,� y� por� fibras�
vegetales� de� la� bosta.� La� fracción� fina� se� encuentra� conformada� por� arcilla,�
opacada�por�materia�orgánica.���
�
�
�!�%�&27�Micromorfología�de�bolas�de�cría�de���� �����������.���Se�distinguen�las� distintas� capas� de� fibras� vegetales� de� la� bosta� delimitadas� por� materia� orgánica�castaño� rojizo� (flechas� amarillas),� desde� el� exterior� (inferior)� hacia� la� cavidad� hueca�interna� donde� se� desarrollaba� la� larva.� Se� distinguen� también,� los� dos� paquetes� de������� imbrincados�(flechas�blancas).�Escala:�0,5�cm;����Detalle�del�contacto�entre� los�paquetes�(flecha�blanca)�y� la�zona�del�relleno�en�bandas�concéntricas�(flecha�amarilla).�Escala:�3�mm;����Restos�de�la�cámara�ovígera�representados�por�una�capa�de�material�del�suelo�en�forma�de�arco.�Escala:�0.5�cm.���
�

� ���
5.2.5.3���Estructura�del�nido�
Se� estudió� un� total� de� nueve� nidos,� de� los� cuales� siete� fueron� hallados�
debajo�de�bosta�de�vaca�y�uno�debajo�de�excremento�humano.�El�noveno�nido�
fue� estudiado� en� condiciones� naturales� controladas,� como� fue� explicado� en� la�
sección�de�materiales�y�métodos,�usando�bosta�de�vaca�como�fuente�de�alimento�
y�nidificación.�De�todos�estos�nidos,�sólo�en�uno�de�ellos�se�halló�la�cámara�nido�
con�su�bola�de�cría,�mientras�que�en� los� restantes�solo�se�encontraron� túneles�
vacíos� o� de� alimentación.� En� los� nidos� constituidos� solamente� por� túneles,� se�
distinguen� básicamente� dos� tipos.� En� el� primer� tipo,� los� túneles� son� cortos� y�
curvos,�completamente�rellenos�con�bosta,�se�encuentran�excavados�a�10�cm�de�
profundidad�(N�=�2)�y�no�se�comunican�con� la�superficie�del�suelo�(Fig.�18�C).�
Esto�último�no�significa�que�la�conexión�no�exista,�sino�que�en�ninguno�de�los�dos�
casos,�pudo�ser�observada.�El�primer�tramo,�horizontal,�de�6�cm�de�largo�y�2�cm�
de�diámetro�promedio�(N�=�2),�forma�un�ángulo�de�90º�con�el�segundo�tramo,�
vertical,�de�3�cm�de� largo�y�2,6�cm�de�diámetro�promedio�(N�=�2).�En� los�dos�
túneles� excavados� se� encontró� un� escarabajo� macho� adulto� alojado� entre� la�
bosta�aprovisionada� (Fig.�18�C).�En�el� segundo� tipo,� los� túneles� se�encuentran�
excavados� a� una� profundidad� similar� a� la� anterior� (alrededor� de� 10� cm),� son�
largos,�levemente�inclinados,�y�con�su�extremo�más�distal�generalmente�curvado�
(Fig.�18�D).�Se�comunican�con�la�superficie�por�medio�de�un�agujero�que�puede�
estar�parcialmente�obturado�por�bosta�desmenuzada.�La�primera�parte�del�túnel�
es�recta,�mide�2�cm�de�diámetro�y�20�cm�de�largo,�promedio�(N�=�2).�La�porción�
final� es� curva,� con� un� recorrido� paralelo� a� la� superficie,� y� de� 7� cm� de� largo�
promedio� (N� =� 2).� El� último� tercio� del� túnel� se� encuentra� aprovisionado� con�
bosta,�y�en� los�dos�casos�estudiados,�se�encontró�una�hembra�alojada�entre� la�
provisión�(Fig.�18�D�y�E) �Finalmente,�en�los�nidos�en�los�que�se�halló�la�bola�de�

� ��
cría,� las� cámaras� son� esféricas,� de� 5� cm� de� diámetro� aproximadamente,� y� se�
encuentran� excavadas� a� poca� profundidad� (10� cm�aproximadamente),� pero� no�
pudo�reconocerse�la�posición�de�la�bola�de�cría�dentro�de�la�cámara�debido�a�la�
excavación.�No�se�encontraron�túneles�conectados�con�dichas�cámaras.���
�
6�4�3#����������� ������?�&)�!*!%"A
�
5.2.6.1���Estructura�de�la�bola�de�cría��
Las�bolas�de�cría�estudiadas�son�esféricas�(Fig.�20�C),�de�1,5�cm�a�2,7�cm�
de�diámetro�(N�=�14),�conformadas�por�una�pared�delgada�de�1,2�mm�a�2,6�mm�
de� espesor� alrededor� de� un� relleno� interno� de� bosta� (N�=� 13)� (Fig.� 20� F).� La�
superficie� externa� de� la� pared� muestra� depresiones� muy� pequeñas� y� poco�
profundas,� irregularmente� distribuidas,� probablemente� hechas� por� el� adulto�
durante�la�construcción�y�el�modelado�de�la�bola�de�cría�(Fig.�20�C).�En�el�polo�
superior�de�la�bola,�se�aloja�la�cámara�ovígera�aislada�del�relleno�por�una�pared�
(Fig.�20�F).�La�porción�de�bosta�sobre�la�cual�se�encuentra�construida�la�cámara�
ovígera,�posee�fibras�vegetales�paralelas�a� la�base�de�dicha�cámara.�En�el�polo�
superior,�la�cámara�ovígera�posee�una�abertura�que�comunica�su�interior�con�el�
exterior�de�la�bola�de�cría�(Fig.�20�C).�La�pared�de�la�bola�de�cría�se�continua�y�
forma�en�el�polo�superior,�sobre�la�cámara�ovígera,�una�protuberancia�cilíndrica�
de� 6� mm� a� 10� mm� de� diámetro� por� 3� mm� de� alto,� terminada� en� un� borde�
prominente� de� superficie� plana� y� anular� (Fig.� 20� A� y� C).� Dentro� de� dicha�
protuberancia,�que�comunica�el�interior�de�la�cámara�ovígera�con�el�interior�de�la�
cámara� nido,� se� observan� algunas� fibras� vegetales� de� la� bosta� que� se�
entrecruzan,� tanto� vertical� como� horizontalmente,� conformando� un� tapón� de�
aireación�(“aereation�plug”������Halffter�y�Edmonds,�1982)�(Fig.�20�C�y�F,�Fig.�

� ���
21�D).�En�el� interior� de� la� cámara�ovígera,� tapizado�por�material� pardo�oscuro�
que� forma� una� capa� muy� delgada� (Fig.� 20� F� y� Fig.� 21� B),� se� encuentra�
depositado�verticalmente,�un�huevo�de�color�amarillo�pálido,�de�5�mm�a�6�mm�de�
largo� y� 2�mm� de� diámetro� (N�=� 2)� (Fig.� 20� F).� Las� bolas� de� cría� que� fueron�
cortadas�en�el�laboratorio�para�la�determinación�del�estadio�de�desarrollo�larval�y�
su�grado�de�avance,�mostraron�en�su�interior�una�cavidad�excavada�en�el�relleno,�
en� la� que� se� aloja� la� larva,� tapizada� por� una� capa� delgada� de� material� color�
pardo�depositada�sobre�la�provisión�alrededor�de�la�larva�(Fig.�20�G�y�Fig.�21�C).�
Cuando� estas� bolas� fueron� abiertas� por� primera� vez� esta� capa� no� estaba�
presente,� pero� si� se� la� observó� posteriormente,� indicando� que� la� larva� habría�
depositado�este�material�sobre�las�rajaduras�provocadas�por�los�cortes.��

� ���
�
�!�%�& 4<� Nido� y� bolas� de� cría� de� �������� ��� ���.� � Nido� conformado� por� una�cámara� nido� con� una� bola� de� cría� en� su� interior;���� Nido� compuesto� por� un� túnel� de�almacenamiento� simple,� completamente� aprovisionado� con� bosta� depositada� en�

� ���
meniscos;���Vista�externa�de�una�bola�de�cría.�Puede�distinguirse�en�el�polo�superior�la�protuberancia� cilíndrica� (flecha� negra),� por� la� cual� internamente� pasa� el� canal� de�aireación,� y� en� la� superficie� externa� de� la� pared,� depresiones� poco� profundas� e�irregularmente� distribuidas� (flecha� blanca); ��� Túnel� de� nidificación,� completamente�relleno�por�material�del�suelo�(flechas�negras),�conectado�con�la�cámara�nido;���Pared�de� la� cámara� nido� en� la� que� se� distinguen� surcos� inclinados,� delgados,� curvos� y� poco�profundos�excavados�en�la�superficie�interna�(flecha�blanca);�y�una�depresión�de�sección�circular�(flecha�negra)�que�conecta�esta�cámara�con�el� túnel�de�nidificación;���Bola�de�cría,�cortada�por�el�eje�del�canal�de�aireación,�donde�se�distinguen�el�relleno�(derecha),�la�pared�de�la�cámara�ovígera�(izquierda)�y�un�huevo�en�su�interior�(derecha).��Bola�de�cría,� cortada� por� el� eje� del� canal� de� aireación,� donde� se� distingue� el� tapiz� (derecha)�depositado�por�la�larva�en�desarrollo�sobre�la�superficie�interna�de�la�cavidad.�Escalas:�1�cm.�
�
5.2.6.2���Micromorfología�de�la�bola�de�cría�
En�las�bolas�de�cría�con�cámara�ovígera,�la�pared�mide�de�1,2�mm�a�2�mm�
de�espesor�(N�=�4),�y�se�compone�tanto�de�material�del�suelo�como�de�bosta.�La�
abundancia�relativa�de�estos�dos�componentes�es�diferente�en�distintas�partes�de�
la�pared,�encontrándose�mayor�cantidad�de�material�del�suelo�hacia�la�parte�más�
externa,� y� más� fibras� vegetales� de� la� bosta� hacia� la� parte� más� interna.� En�
algunos� casos� estas� fibras� se� encuentran� orientadas� paralelamente� al� relleno�
(Fig.�21�A).�La�microestructura�de�la�pared�es�maciza�con�una�porosidad�de�10���
5�%.�La�fracción�gruesa�es�tamaño�arena�fina�–�limo,�representa�un�70�%�de�la�
pared� y� esta� compuesta�por� cuarzo,�K� �� feldespato,�plagioclasa,�mica,� y� trizas�
vítreas� (en�orden�de� importancia).�El�40�%�de�esta� fracción�esta� representada�
por�fibras�vegetales�de�la�bosta�dispersas�en�la�pared,�birrefringentes�y�de�1�mm�
de� longitud,� aproximadamente.� La� fracción� fina� posee� un� alto� contenido� de�
materia� orgánica,� y� esta� representada� por� granos� castaño� oscuro,� pálidos,�
tamaño�arcilla,�que�se�hacen�más�oscuros�hacia�el�relleno�de�la�bola.�El�relleno�
esta� conformado� en� un� 70� �� 75� %� por� fibras� vegetales� de� la� bosta� sin�
orientación,�y�en�el�30���35�%�restante,�por�fragmentos�de�material�de�suelo.�La�
pared�de�la�cámara�ovígera�posee�un�espesor�de�250�^�a�500�^,�y�se�compone�
principalmente�por�materia� orgánica� color� castaño�oscuro,� y� escasos�minerales�

� ���
dispersos� (Fig.� 21� B).� El� canal� de� aireación� posee,� en� el� interior� de� la�
protuberancia� cilíndrica,� fibras� vegetales� de� la� bosta� orientadas� tanto� vertical�
como�horizontalmente�(Fig.�21�D).�����������
La�pared�de�las�bolas�de�cría�en�la�que�se�encontraba�la�larva�(sin�cámara�
ovígera),�posee�un�espesor�de�1,7�mm�a�2,6�mm�(N�=�8)�y�una�microestructura�
maciza�con�una�porosidad�del�3�%�(Fig.�21�C).�La�fracción�gruesa�es�tamaño�limo�
(selección� moderada),� representa� el� 40� %� de� la� pared� y� esta� compuesta� por�
cuarzo,� K� �� feldespato,� plagioclasa,� trizas� vítreas� (en� orden� de� importancia),�
escasa� mica� (biotita� y� muscovita),� y� algunos� minerales� pesados� (piroxeno� y�
epidoto).�La�fracción�fina�esta�representada�por�material�tamaño�arcilla�de�color�
marrón� oscuro� (alto� contenido� de� materia� orgánica).� El� relleno� posee� una�
porosidad� que� varia� entre� 5� %� y� 10� %,� y� esta� formado� por� paquetes�
(aproximadamente� 1� cm� de� largo� y� 1,8� mm� de� ancho)� que� se� acomodan� de�
forma� imbricada� unos� con� otros� a� la� largo� del� relleno.� Cada� paquete� está�
conformado� por� abundante� cantidad� de� fibras� vegetales� de� la� bosta�
(birrefringentes�y�de�1,5�mm�de� largo,�aproximadamente),�materia�orgánica,�y�
por�granos�minerales�dispersos,�cuya�composición�y�textura�son�similares�a� las�
de� la� pared� ya� descripta.� En� el� interior� de� los� paquetes,� estos� materiales� se�
encuentran� formando� ������ alargados,� algunos� lenticulares,� y� yuxtapuestos�
muy�ordenados.�Cada������puede�ser� identificado�por� las� fibras�vegetales�que�
se� acomodan� paralelamente� al� borde� de� cada� uno� de� ellos,� en� el� cual� se�
concentra� la�materia�orgánica.�El� tamaño�de�estos�������varía,�encontrándose�
������mayores�de�2,5�mm�de�largo�y�500�^�de�ancho,�y�������más�pequeños�
de�1,5�mm�de�largo�y�250�^�de�ancho,�aproximadamente.�Dentro�del�relleno,�la�
larva�se�encuentra�alojada�en�una�cavidad�recubierta�por�una�delgada�capa�de�75�
^� a� 360� ^� de� espesor,� de� color� castaño� oscuro� a� muy� oscuro� (Fig.� 21� C).� El�

� ���
material�de�esta�capa�difiere�del�material�que�se�encuentra�en�la�bola�de�cría�y�
en� el� relleno,� porque� no� posee� minerales� y� esta� compuesto� solamente� por�
materia�orgánica�sin�fibras�vegetales�de�la�bosta.����
�
�
�!�%�&42��Micromorfología�de�bolas�de�cría�de������������ ���.�Pared�de�la�bola�de�cría� compuesta� por�material� del� suelo� y� fibras� vegetales� de� la� bosta� (flechas� negras).�Escala:� 500� ^;���� Pared� de� la� cámara� ovígera� compuesta� principalmente� por� materia�orgánica� (flechas�negras)� y� con�minerales�dispersos� (flecha�blanca).� Escala:�1�mm;���Tapiz�de�materia�orgánica�(flechas�negras),�sin�fibras�vegetales�ni�minerales,�depositado�en� la� cavidad� interna� por� la� larva� en� desarrollo.� Escala:� 1� mm;� �� Interior� de� la�protuberancia�mostrando�el�canal�de�aireación�con�fibras�vegetales�de� la�bosta�(flechas�negras),�la�mayoría�de�las�cuales�fueron�removidas�con�el�corte.����
�
�

� ���
5.2.6.3���Estructura�del�nido��
Los�treinta�nidos�estudiados�fueron�encontrados�debajo�de�dieciséis�bostas�
diferentes:�en�7�casos�se�encontró�un�sólo�túnel�de�almacenamiento�debajo�de�
cada�bosta;�en�3�casos,�una�sola�cámara�nido;�en�5�casos,�más�de�una�cámara�
nido� (hasta� ocho);� y� en� 1� sólo� caso,� se� encontraron� 3� túneles� de�
almacenamiento� pertenecientes� a� las� tres� especies� estudiadas� en� Navarro,����
��� ���,����� �������y��������.���
Debajo�del�total�de�las�bostas�estudiadas�se�encontraron�dos�tipos�de�túneles,�
los� cuales� son� considerados� aquí� como� nidos� (Fig.� 20� A� y� B),� en� sentido�
generalizado.�Sólo�en�un�caso,�los�dos�tipos�de�túneles�fueron�encontrados�juntos�
debajo� de� una� misma� bosta.� El� primer� tipo� de� túnel,� denominado� de�
almacenamiento,� se� encuentra� completamente� aprovisionado� con� bosta�
depositada�en�forma�de�meniscos�(Fig.�20�B),�sobre�la�cual�se�ha�encontrado,�en�
algunos� casos,� un� adulto� en� reposo.� El� segundo� tipo� de� túnel,� se� encuentra�
conectado�con�la�cámara�nido�(Fig.�20�D),�la�cual�aloja�en�su�interior�una�bola�de�
cría�completa.�En�la�mayoría�de�los�nidos�estudiados�de�este�tipo,�los�túneles�de�
nidificación�se�encuentran�rellenos�con�material�del�suelo�que�impide�que�se�los�
pueda� distinguir� del� suelo� circundante.� Sólo� en� un� caso� pudo� distinguirse� el�
relleno�de�un�túnel,�conectado�a�una�cámara�nido,�con�bastante�dificultad�(Fig.�
20� D).� En� este� tipo� nido,� no� se� encontraron� adultos� ni� restos� de� túneles� de�
almacenamiento.���
Los� túneles� de� almacenamiento� pueden� variar� en� su� forma� general,�
pudiendo� ser� completamente� rectos� y� verticales,�mayormente� horizontales� con�
algunos�segmentos�verticales,�u�oblicuos�con�una�o�más�curvas�(Fig.�20�B).�Este�
tipo� de� túnel�mide�de�1,5� cm�a�2,5� cm�de�diámetro,�15� cm�de� largo� y� puede�
alcanzar�de�10�cm�a�20�cm�de�profundidad�(N�=�6).�Como�regla�general,�cuando�

� ���
el� túnel� es� largo,� también� es� oblicuo� y� profundo.� El� relleno� es� de� bosta�
depositada� en� forma� de� meniscos� (Fig.� 20� B),� y� no� poseen� pared� con�
características�distintivas.�En�relación�a�la�cámara�nido,�solo�en�dos�de�los�casos�
observados,� se� encontraron� varias� cámaras� debajo� de� una� misma� bosta,�
separadas� entre� si� por� muy� pocos� centímetros.� En� uno� de� estos� grupos� se�
encontraron�tres�cámaras�nido�excavadas�a� la�misma�profundidad,�y�separadas�
entre� si� por� una� distancia� que� no� supera� 2� �� 3� cm.� En� el� segundo� grupo� se�
encontraron�dos�cámaras�nido�excavadas�a�la�misma�profundidad�y�distanciadas�
entre� sí� por� 1,5� cm.� En� el� resto� de� las� bostas� estudiadas,� los� nidos� se�
encontraron� separados� a� una� distancia� mayor� (de� 5� cm� a� 13� cm).� Todas� las�
cámaras�nido�medidas�se�encuentran�excavadas�de�9�cm�a�19�cm�de�profundidad�
en�relación�con�la�superficie�(N�=�22),�son�esféricas�(Fig.�20�A�y�C),�con�un�eje�
vertical�que�mide�de�2,8�cm�a�3,6�cm�de�largo�y�un�eje�ecuatorial�que�mide�de�
2,3�cm�a�3,7�cm�de� largo�(N�=�17).�En�todos� los�casos�se�observa�un�espacio�
libre�de�3�mm�a�4�mm�(N�=�14)�que� separa� la�bola�de� cría� de� la�pared�de� la�
cámara� nido� (Fig.� 20� A).� La� pared� de� la� cámara� nido� posee� surcos� delgados,�
inclinados,� curvos� y� poco�profundos,� trazados� en� su� superficie� interna� (Fig.� 20�
E).�Cuando�la�cámara�nido�está�terminada,�se�encuentra�completamente�aislada�
y�cerrada�por�material�del�suelo.�En�algunas�cámaras,�se�encontró�una�depresión�
redondeada�de�1�cm�de�diámetro�(Fig.�20�E)�excavada�en�la�pared,�en�posición�
lateral� y� superior,� que� podría� corresponderse� con� la� comunicación� que� existió�
originalmente�entre�la�cámara�nido�y�el�túnel�de�nidificación.��
�
6�4�;H�� ������ �������������������������� ��
�
5.2.7.1���Estructura�de�la�bola�de�cría�

� ��
La�única�bola�de�cría�que�pudo�ser�colectada,�es�piriforme�(Fig.�22�A),�con�
un�eje�mayor,�que�atraviesa�la�cámara�ovígera,�de�2,1�cm�de�alto�y�uno�menor,�
diámetro� ecuatorial,� de� 1,7� cm.� Hacia� el� polo� superior,� la� bola� se� estrecha�
notablemente� alcanzando� un� valor� de� 0,5� cm.� La� pared� de� la� bola� está�
conformada�por�material�del�suelo,�es�muy�delgada�y�no�puede�ser�medida�a�ojo�
desnudo.�En�el�relleno�la�bosta�no�muestra�un�arreglo�particular,�aunque�si�se�lo�
distingue�muy�compactado�(Fig.�22�B).�En�el�polo�superior�de�la�bola�de�cría,�se�
observa� en� el� interior,� la� cámara� ovígera� esférica� (0,5� cm� de� diámetro)�
directamente� excavada� en� el� material� del� relleno� (Fig.� 22� B).� Esta� cámara� se�
encuentra�completamente�cerrada,�ya�que�no�se�distingue�en�ella�ningún�canal�o�
poro�de�aireación.�No�se�observa�pared,�ni�tampoco�se�distingue�un�tapiz,�solo�se�
nota� su� superficie� interna�bien� alisada.� En� el� interior� de� la� cámara�ovígera,� se�
aloja,�horizontalmente,�un�huevo�ovalado,�grande,�de�6�mm�de�largo�y�3�mm�de�
ancho,�color�amarillo�pálido�(Fig.�22�B).�����

� ���
�
�!�%�&44�Nido�y�bolas�de�cría�de� ��������! ���������"���� �� �����.�Bola�de�cría�piriforme;���Bola�de�cría�cortada�por�el�eje�de�la�cámara�ovígera�(flecha�blanca)�en� la� que� se� distingue�el� huevo� depositado� en� su� interior; ���Dos� escarabajos� adultos�encontrados�enterrados�juntos�y�con�una�bola�de�provisión;���Bolas�de�provisión�en�las�que�se�distingue�un�surco�en�uno�de�sus�polos�(flechas�blancas);��Nido�conformado�por�una�cámara�en�la�que�se�encontró�al�adulto�junto�con�una�bola�de�cría�en�construcción;��� Nido� conformado� por� una� cámara� con� una� bola� de� cría piriforme� completa� en� su�interior.�Escalas:�1�cm.����
�

� ��
5.2.7.2���Micromorfología�de�la�bola�de�cría�
En�la�bola�de�cría�con�cámara�ovígera�la�pared�posee�un�espesor�de�500�^�
a�1,5�mm,�y�esta� conformada�por������� grandes�yuxtapuestos� (pueden�verse�
hasta�tres�������superpuestos)�(Fig.�23�A).�La�microestructura�es�maciza,�con�
10�%�de�porosidad,�donde�los�poros�elongados�delimitan� los�bordes�de�algunos�
de� los� �����.� Este� material,� observado� con� menor� aumento,� posee� una�
disposición� aparentemente� concéntrica.� La� fracción� gruesa� es� tamaño� limo� ��
arena� media,� representa� un� 40� �� 45� %� de� la� pared� y� esta� compuesta� por�
plagioclasa,� K� �� feldespato,� mica� (biotita� y� muscovita),� cuarzo� y� fragmentos�
líticos.� En� esta� fracción� se� observan� también� fibras� vegetales� de� la� bosta,�
mayormente�elongadas.�La�fracción�fina�esta�compuesta�por�material�arcilloso�y�
materia� orgánica� de� color� negro� y� castaño� rojizo.� En� el� relleno,� la� zona� en�
contacto� con� la� pared� se� dispone� en� capas� groseramente� concéntricas� que� no�
pueden�distinguirse�hacia� la�parte�más�interna�(Fig.�23�B�y�C).�El�relleno�posee�
una�porosidad�de�20�%,�en�el�que� los�poros�mayores,�elongados,�coinciden�en�
muchos� casos� con� los� límites� de� las� capas� ya� descriptas.� La� fracción� gruesa�
representa�el�35�%�del�relleno,�es�tamaño�limo�–�arena�media�y�esta�compuesta�
por�plagioclasa,�cuarzo,�mica�(biotita�y�muscovita),�K���feldespato�y�fragmentos�
líticos.�Conforman�también�esta�fracción,�fibras�vegetales�de�la�bosta�(Fig.�23�C).�
La�fracción�fina�esta�compuesta�por�material�arcilloso,�materia�orgánica�oscura�y�
pequeñas�ojuelas�de�materia�orgánica�rojiza,�homogéneamente�distribuidas�(Fig.�
23�D).�La�cámara�ovígera�posee�una�pared�de�600�^�de�espesor�(Fig.�23�B�y�E)�
que� la� separa� del� relleno� y� la� cierra� completamente.� Esta� pared,� posee� una�
microestrutura�maciza� con� una� porosidad� del� 15�%,� con� poros� intergranulares�
(Fig.�23�E).�La�fracción�gruesa�es�tamaño�limo�–�arena�media,�representa�un�40�
%� de� la� pared� y� esta� compuesta� por� plagioclasa,� mica� (biotita� y� muscovita),�

� ���
cuarzo,�K���feldespato�y�fragmentos�líticos.�Se�observa�además�en�esta�fracción,�
escasas� fibras� vegetales� de� la� bosta.� La� fracción� fina� se� compone� de�material�
arcilloso� y� materia� orgánica� oscura,� castaña,� y� algunas� ojuelas� de� materia�
orgánica�rojiza.���
�
�
�!�%�& 45�� Micromorfología� de� bolas� de� cría� de� �������� ! ���������"���� �� �����.��� Detalle� de� la� pared� de� la� bola� de� cría� conformada� por� grandes� ������yuxtapuestos�(flechas�blancas).�Escala:�425�^;����Zona�de�contacto�entre�la�pared�de�la�bola�de�cría�y�el� relleno�(flechas�blancas),�y�entre�este�último�y� la�pared�de� la�cámara�ovígera�(de�izquierda�a�derecha).�Escala:�2,5�mm;����Detalle�de�la�zona�de�contacto�de�la�pared�y�el�relleno,�donde�el�material�del�último�se�encuentra�ordenado�en�capas�(flechas�blancas)�relativamente�concéntricas.�Escala:�800�^;����Detalle�de�la�zona�más�central�del�relleno�donde�se�distinguen�ojuelas�de�materia�orgánica�rojiza.�Escala:�350�^;����Detalle�de� la� pared� de� la� cámara� ovígera.� Las� flechas� blancas� indican� el� límite� con� el� relleno.�Escala:�600�^.���

� ���
5.2.7.3���Estructura�del�nido�
Se�estudió�un�total�de�cuatro�nidos,�de�los�cuales�tres�fueron�encontrados�
en�las�cercanías�de�un�corral�de�cabras�(5���10�m),�todos�ellos�hallados�debajo�de�
perforaciones� en� el� suelo� o� montículos� de� tierra� removida.� El� cuarto� nido� fue�
estudiado� en� condiciones� de� laboratorio� controladas,� colocando� en� un� terrario�
una�pareja�de�escarabajos�con�una�bola�de�provisión,�previamente�construida�y�
rolada� por� ellos.� En� este� último� caso,� posiblemente� la� provisión� sea� bosta� de�
caballo,� aunque� se� ha� registrado� también� para� esta� localidad,� escarabajos� de�
este�género�rolando�trozos�de�excremento�de�perros�(Genise,�com.�pers.).�De�los�
cuatro� casos� estudiados,� solo� en� dos� de� ellos� se� hallaron� las� cámaras� nido,�
mientras� que� en� los� dos� restantes,� en� los� que� no� se� distinguen� cámaras� ni�
túneles,�se�encontraron�a�los�escarabajos�enterrados�a�muy�poca�profundidad�(2�
3�cm),�cada�uno�con�una�bola�de�provisión�(Fig.�22�C).�Estas�bolas�son�ovaladas�
(eje�mayor:�19�mm�a�22�mm,�eje�menor:�16�mm�a�18�mm,�N�=�3)�o�piriformes�
(eje�mayor:�23�mm,�eje�menor:�18�mm,�N�=�1)�y�poseen�algunas�características�
particulares,� como� por� ejemplo,� dos� de� las� tres� bolas� halladas,� poseen� una�
muesca�o�surco�en�uno�de�sus�polos�(Fig.�22�D),�que�podría�tener�relación�con�la�
construcción�de�la�cámara�ovígera,�y�en�el�interior�de�una�de�ellas,�se�encontró�
un�pupario�de�mosca�(Sarcophagidae)�(Fig.�45�E).�En� los�nidos�con�cámara,�no�
se� hallaron� indicios� de� ningún� tipo� de� túnel.� En� el� nido� cercano� al� corral,� la�
cámara�se�encuentra�excavada�a�3�cm�de�profundidad,�es�esférica,�de�3�cm�de�
diámetro,�y�posee�un�su�interior�una�bola�de�aprovisionamiento,�esférica�(18�mm�
de�diámetro),� junto� a�un� escarabajo� adulto� (Fig.� 22�E).� La� pared�de� esta� bola�
posee�pelos�de� cabra�que�pueden�verse�desde�el� exterior,� y�al� ser� cortada,� se�
encontró�en�su�interior�una�larva�de�mosca,�también�Sarcophagidae�(Fig.�45�D).��
En�el�nido�contenido�en�un�terrario,�la�cámara�se�encuentra�excavada�a�13�cm�de�

� ���
la� superficie�y�a�3�cm�del� lugar�por�el� cual� fue�esnterrada� la�bola�de�provisión�
(Fig.� 22� F).� La� cámara� nido� es� relativamente� esférica,� amplia,� existiendo� un�
espacio�libre�de�2�cm�a�3�cm�entre�su�pared�y�la�bola�de�cría,�espacio�en�el�cual�
se� halló� un� escarabajo� adulto.� La� bola� de� cría� es� piriforme� y� se� encuentra� en�
orientada� con� el� polo� que� lleva� la� cámara� ovígera,� hacia� arriba� y� levemenete�
inclinado�(Fig.�22�F).��
�
6�4�:H�� ������ ����� ������ ������������
�
5.2.8.1���Estructura�de�la�bola�de�cría�
Las�bolas�de�cría�colectadas�poseen�una�forma�general�ovalada,�tendiendo�
a� “gota”,� con� el� polo� superior� levemente� estrechado� (Fig.� 24� A� y� C).� El� eje�
mayor,�que�pasa�por�el�canal�de�aireación,�mide�3,3�cm�de�largo�promedio�(N�=�
2)�y�el� eje�menor,�diámetro�ecuatorial,�3,1� cm�de� largo�promedio� (N�=�2).� La�
pared�de�la�bola�de�cría�es�muy�delgada,�de�1,6�mm�de�espesor�promedio�(N�=�
2),�y�está�constituida�fundamentalmente�por�material�del�suelo�(Fig.�24�B�y�D).�
Se� ha� observado� en� este�material,� tiempo� después� de� haber� sido� colectado� y�
guardado� en� la� colección,� que� al� perder� la� humedad� original,� la� pared� de� las�
bolas�se�desprende�con�mucha�facilidad,�dejando�expuesta�la�superficie�externa�
del� relleno� (Fig.� 24� B� y� D).� En� los� dos� ejemplares� estudiados,� esta� superficie�
muestra�un�arreglo�particular�del�relleno�(Fig.�24�D)�que�no�ha�sido�utilizado�por�
la� larva,� y� que� se� conserva� en� la� bola,� constituyendo� la� pared� de� la� cámara�
pupal.� Este� sector� del� relleno� se� encuentra� conformado� por� pequeños� �����,�
apenas� distinguibles,� que� se� acomodan� unos� sobre� otros� formando� bandas� de�
material� (Fig.� 24�D)�que� se� superponen� formando�un�helicoide� alrededor� de� la�
cavidad� esférica,� hueca,� del� interior� de� la� bola� de� cría.� Uno� de� los� ejemplares�

� ���
(Fig.� 24� A� y� B),� aún� cerrado,� contenía� conservada� en� su� interior� una� larva� a�
punto�de�empupar,�ya�que�se�observa�en�esta�bola,�la�espira�terminada�(de�1,2�
mm� a� 3,2� mm� de� espesor)� y� la� cavidad� interna� (de� 2,6� cm� de� diámetro)�
íntegramente� tapizada� por� un�material� oscuro,� posiblemente�materia� orgánica.�
De�la�cámara�ovígera,�solo�se�conserva�el�canal�de�aireación�y�el�espacio�donde�
esta� se� encontraba,� ambos� completamente� rellenos� por� bosta.� En� el� segundo�
ejemplar�(Fig.�24�C�y�D),�se�conserva�el�adulto�en�el�proceso�de�emergencia,�es�
decir�con�el�protórax�en�el�exterior�de�la�bola�(la�cabeza�ausente)�y�el�resto�del�
cuerpo� en� el� interior.� Al� ser� cortada,� se� distingue� en� esta� bola,� un� helicoide�
completo�(2,2�mm�a�4�mm�de�espesor)�rodeando�la�cavidad�interna�(2,2�cm�de�
diámetro),�y�el� tapiz�de�200�^�de�espesor,�aproximadamente.�En�este�ejemplar�
no� se� encontraron� restos� de� la� cámara� ovígera,� posiblemente� debido� a� la�
emergencia�del�adulto�por�dicha�zona.�
�
�
�

� ���
�
�!�%�&48�Nido�y�bolas�de�cría�de� ��������! �������"�������.#��Bola�de�cría�con�forma�general�ovalada�()�que�al�ser�cortada�por�el�eje�del�canal�de�aireación�

� ���
(�)�muestra� la� cámara�pupal� completa� con�una� larva�en�el� interior� (flecha�negra)� y� la�pared�(flechas�blancas)�levemente�separada�de�la�de�dicha�cámara.�Escalas:�1�cm;��#���Bola� de� cría� con� forma� general� ovalada� tendiendo� a� “gota”� (�),� la� cual� conserva� un�escarabajo� adulto�muerto� al� emerger.� Al� ser� removida� su� pared� (�),� se� distinguen� las�bandas� de� ������ (flechas� amarillas)� que� conforman� la� pared� helicoidal� de� la� cámara�pupal.� Escalas:� 1� cm;� �#��� Etapas� previas� a� la� nidificación:� selección� de� la� bosta� (�,�escala:�1�cm),�construcción�de�una�bola�de�provisión�en�la�misma�fuente�de�alimento�(�,�escala:�1�cm),�rolado�(flecha�blanca)�de�la�bola�de�provisión�(�,�escala:�5�cm),�selección�del�lugar�de�nidificación�y�enterramiento�de�la�bola�de�provisión�rolada�(�#�,�escalas:�1�cm).�
�
5.2.8.2���Micromorfología�de�la�bola�de�cría�
En� la� bola� de� cría� con� larva� en� desarrollo,� de� afuera� hacia� adentro,� se�
observa�una�capa�constituida�por�material�del�suelo�y�fibras�vegetales�de�la�bosta�
(Fig.�25�A).�Los�primeros�aproximadamente�375�^�de�dicha�capa,�se�encuentran�
constituidos� fundamentalmente� por� material� del� suelo,� el� cual� se� va�
enriqueciendo�hacia� el� interior� con� fibras� vegetales� de� la� bosta,� alcanzando�un�
espesor� de� 1,63�mm,� aproximadamente� (Fig.� 25� B� y� C).� Este� material� posee�
bandas�de�materia�orgánica�que�delimitan�grandes�paquetes�de�fibras�vegetales�
y�minerales,�los�cuales�además�muestran�cierto�grado�de�orientación�(Fig.�25�C).�
La�microestructura�de�la�primera�capa�de�material,�es�maciza�con�una�porosidad�
de�5�%.�La�fracción�gruesa�es�tamaño�arena�fina���limo,�representa�un�40���50�%�
de� esta� capa� y� esta� compuesta,� en� orden� de� importancia,� por�mica� (biotita� y�
muscovita),� cuarzo,� K�� feldespato,� plagioclasa� y� escasos� fragmentos� líticos.� En�
esta� fracción� se� observan� también� escasas� fibras� vegetales� de� la� bosta� y�
pequeñas� ojuelas� de�materia� orgánica� rojiza� (Fig.� 25� C).� La� fracción� fina� está�
compuesta�por�material� arcilloso� y�materia� orgánica.� Luego�de� esta� capa,�más�
hacia� el� interior� de� la� bola� de� cría� y� ocupando� aproximadamente� 750� ^� de�
espesor,�se�observa�un�aumento�en�la�cantidad�de�fibras�vegetales�de�la�bosta�y�
de� materia� orgánica� oscura,� acompañado� del� incremento� de� la� cantidad� de�
material� fino� (arcilla� embebida� en� materia� orgánica)� y� una� disminución� de� la�
fracción�mineral,�la�cual�posee�además�mayor�tamaño�de�grano.�A�partir�de�aquí,�

� ���
se� observa� una� zona� de� 3,75� mm� de� espesor,� aproximadamente,� cuya�
microestructura� está� compuesta� por� paquetes� de� �����,� en� forma� de� media�
luna,�yuxtapuestos�(Fig.�25�A,�B�y�D).�Estos�paquetes�miden�de�1,5�mm�a�2,6�
mm�de�largo,�y�de�1,9�mm�a�2,7�mm�de�ancho.�El�material�que�los�conforma�se�
distribuye� en� su� interior� formando� �����,� donde� la� porosidad,� heterogénea,�
varía� entre� 5� %� y� 15� %.� La� fracción� gruesa� de� estos� ������ (35� %�
aproximadamente)� se� encuentra� constituida,� mayoritariamente,� por� fibras�
vegetales�de� la�bosta,�minerales�dispersos� tamaño� limo� ��arena� (cuarzo,�mica,�
plagioclasa,�K� �� feldespato,� fragmentos� líticos),� y� pequeñas� ojuelas� de�materia�
orgánica�rojiza�(Fig.�25�D).�Las�fibras�vegetales�son�elongadas,�de�40�^�a�2,25�
mm�de� largo,� y� se� encuentran� orientadas� paralelamente� a� los� límites� de� cada�
paquete.�La�fracción� fina�se�encuentra�constituida�por�arcilla,�muy�opacada�por�
materia� orgánica� oscura� (Fig.� 25� D).� Más� hacia� el� interior,� cubriendo�
parcialmente�la�cavidad�interna�(cámara�pupal)�de�la�bola�de�cría,�se�observa�un�
tapiz�de�hasta�150�^�de�espesor�compuesto�por�materia�orgánica�castaño�rojiza�
(Fig.�25�E),�similar�a�la�que�se�observa�en�el�interior�de�los������.��
En�uno�de�los�polos�de�la�bola�da�cría,�las�fibras�vegetales�de�la�bosta�y�el�
material� de� suelo� que� conforman� la� parte� más� externa� de� la� bola,� se� curvan�
hacia�fuera�generando�un�cono�(Fig.�25�F),�el�cual�corresponde�con�los�restos�de�
la�cámara�ovígera.�El� interior�de�este�cono�se�encuentra�parcialmente�ocupado�
por� fibras� vegetales� de� la� bosta� y� escasos� minerales� dispersos,� los� cuales� se�
disponen� desordenadamente� y� sueltos.� La� base� de� este� cono� esta� conformada�
por�un�pequeño�paquete�de������,�con�idénticas�características�a�las�descriptas�
anteriormente�(Fig.�25�F).��
�

� ��
�
�!�%�&46�Micromorfología�de�bolas�de�cría�de� ��������! �������"�������.�Se�distinguen�zonas�de�distinta�composición�en�el�corte�de�la�bola�de�cría,�en�las�que�la�más� externa� (abajo,� flechas� negras)� se� compone� fundamentalmente� de� material� de�suelo,� enriquecida� hacia� el� interior,� con� fibras� vegetales� de� la� bosta.� En� la� zona�más�interna� (arriba,� flechas� blancas),� se� distinguen� paquetes� de� ������ delimitados� por�materia� orgánica� oscura.� Escala:� 3�mm;����Detalle� de� la� transición� entre� la� zona�más�externa,�compuesta�por�paquetes�de�material�del�suelo�delimitados�por�bandas�oscuras�de�materia� orgánica� (flechas� amarillas),� y� la� zona�más� interna� de� paquetes� de�������yuxtapuestos,�también�delimitados�por�materia�orgánica�oscura�(flecha�blanca).�Escala:�2�mm;� ��� Detalle� de� la� zona� más� externa� de� la� bola,� donde� se� distinguen� las� fibras�vegetales�de�la�bosta�(flechas�amarillas)�y�las�ojuelas�de�materia�orgánica�rojiza�(flechas�blancas).�Escala:�0,5�mm;����Detalle�de�los�paquetes�de�������de�la�zona�más�interna�de�la�bola�de�cría.�Escala:�1�mm;����Detalle�del�tapiz�(flechas�amarillas)�que�recubre�la�superficie� interna� de� la� cámara� pupal.� Escala:� 400�^;���� Restos� de� la� cámara� ovígera�representados� por� un� cono� conformado� por� un� paquete� de� ������ en� la� base� (flecha�negra)�y�un�centro�parcialmente�ocupado�por�fibras�vegetales�de�la�bosta�y�por�escasos�minerales�dispersos�(flecha�amarilla),�dispuestos�de�manera�desordenada.�Escala:�1�cm.��
�
5.2.8.3���Estructura�del�nido�
A� pesar� de� las� distintas� metodologías� aplicadas� con� esta� especie� en�
distintas�campañas,�solo�pudieron� reconocerse�algunas�pocas�características�de�
los� nidos.� De� los� casos� estudiados� en� condiciones� ambientales� naturales,� pero�
controladas,� de� cinco� ejemplares� en� el� 2007� y� dos� en� el� 2008,� sólo� dos� en� el�
primer�caso�y�uno�en�el�segundo,�seleccionaron,�rolaron�y�enterraron�una�porción�

� ���
de�bosta�(Fig.�24�E�J).�De�estos�resultados�y�en�relación�con� los�nidos,�solo�se�
observaron� las� excavaciones� hechas� en� el� suelo� en� el� momento� del�
enterramiento�de�la�bola�de�provisión�(Fig.�24�H�J),�que�deja�como�resultado�en�
la�superficie,�un�montículo�cónico�de�material�del�suelo�removido�(Fig.�24�J).�En�
un�solo�caso,�en�el�del�nido�abierto�al�día�siguiente�del�enterramiento,�se�halló,�
justo� por� debajo� de� la� base� de� dicho� montículo� y� alojados� en� una� cámara�
espaciosa� pero� de� forma� no� muy� bien� definida,� al� escarabajo� y� la� bola� de�
provisión�intacta.�En�los�otros�dos,�en�un�caso�se�encontró�al�escarabajo�alojado�
en�un�túnel�pequeño�(5�cm�de�largo)�excavado�a�muy�poca�profundidad�(no�más�
de� 2� cm)� junto� con� restos� de� bosta� removidos,� y� en� el� segundo� caso,� se� lo�
encontró�en�una�pequeña�cámara�esférica�excavada�en�el�piso�de�otra� cámara�
ovalada�mayor� en� la� que� se� hallaron� dispersos� restos� de� bosta.� De� los� casos�
estudiados�en�el� laboratorio,�en�el� terrario�armado�en�el�2007,�el�ejemplar�que�
había�enterrado� la� bola�durante� la�experimentación�hecha�en�ambiente�natural�
controlado,�murió�en�esta�nueva�etapa,�y�el�restante�individuo�fue�el�responsable�
de�este�proceso.�En�este�terrario,�se�halló�una�cámara�(eje�horizontal�mayor:�5�
cm,�eje�vertical�menor:�3�cm)�excavada�a�5�cm�de�profundidad,�dentro�de�la�cual�
se�encontraban�el�escarabajo�y� la�bola�de�provisión� intacta.�No�se�encontraron�
túneles�conectados�con�la�cámara.�En�el�terrario�armado�en�el�2008,�luego�de�6�
días,�el�escarabajo�emergió�a�la�superficie�por�medio�de�una�perforación�de�1,5�
cm� de� diámetro� hecha� en� el� suelo� a� 2� cm� de� distancia� del� montículo� dejado�
durante�el�enterramiento�de�la�bola�de�provisión.�Luego�de�un�delicado�proceso�
de� extracción� manual� del� material� del� suelo� del� terrario,� no� se� encontraron�
indicios� del� nido� ni� de� la� bola� de� cría.� Posteriormente,� pudieron� identificarse� a�
estos� escarabajos� como� machos,� lo� que� explica� el� hecho� que� solo� realizaran�
bolas�de�alimentación�durante�el�periodo�de�experimentación.�

� ��
6�4�7#��������"B�2
�
5.2.9.1���Estructura�de�la�bola�de�cría�
La�única�bola�de�cría�colectada�es�piriforme�(Fig.�26�A),�con�un�eje�mayor�
que�pasa�por� el� canal� de� aireación,� de�1� cm�de� largo,� y� uno�menor,� diámetro�
ecuatorial,� de� 7� mm� de� largo.� El� polo� superior,� donde� se� abre� el� canal� de�
aireación,�se�angosta�hasta�alcanzar�2�mm.�En�el�corte�(Fig.�26�B),�la�pared�de�la�
bola�de�cría�mide�de�800�^�a�1�mm�de�espesor,�y�se�encuentra�constituida�por�
material�del�suelo�y�fibras�vegetales�de�la�bosta.�La�bosta�del�relleno�no�presenta�
un� ordenamiento� particular.� En� el� interior� de� dicho� relleno,� y� hacia� el� polo�
superior�de�la�bola,�se�encuentra�excavada�directamente�en�la�bosta,�la�cámara�
ovígera�(Fig.�26�B�y�C).�Esta�cámara�es�esférica,�de�1,9�mm�de�diámetro,�y�no�
posee�pared,�solo�un�material�oscuro�que�tapiza�su�superficie�interna�(Fig.�26�C).�
La�cámara�no�posee�un�techo,�y�se�abre�a�una�cavidad�hueca�superior�ocupada�
por�fibras�vegetales�que�emergen�de�la�pared�y�se�cruzan�entre�si�formando�un�
canal� de� aireación� (Fig.� 26� C).� El� canal� desemboca� en� una� perforación� muy�
pequeña�en�el�extremo�del�polo�superior�de� la�bola�de�cría.�En�el� interior�de� la�
cámara� ovígera,� se� aloja� horizontalmente,� un� huevo� ovalado,� color� amarillo�
pálido,�de�2�mm�de�largo�y�1,2�mm�de�ancho�(Fig.�26�B).��������
�

� ���
�!�%�&43��Nido�y�bolas�de�cría�de��������sp.1.���Bola�de�cría�piriforme;����Bola�de�cría� cortada� por� el� eje� del� canal� de� aireación,� dende� se� distinguen� la� pared� (flecha�

� ���
negra),�el�relleno�(flecha�amarilla)�y�la�cámara�ovígera�(flecha�blanca)�con�un�huevo�en�el�interior�(mitad�derecha);����Detalle�de�la�cámara�ovígera�(flecha�blanca)�y�del�canal�de�aireación�(flecha�amarilla),�cruzado�internamente�por�fibras�vegetales�de�la�bosta;��#���Bolas�de�provisión�enteras�(�)�y�parcialmente�consumidas�(�);��#��Etapas�previas�a�la�nidificación:�construcción�de�una�bola�de�provisión�en�la�bosta�seleccionada�(�)�y�rolado�de�la�misma�(��y��).�Escalas:�1�cm.����
�
5.2.9.2���Micromorfología�de�la�bola�de�cría�
La�pared�de�la�bola�de�cría�(Fig.�27�A�y�B)�posee�un�espesor�que�varia�de�
425�^�a�500�^.�La�microestructura�es�de�grano�simple,�con�una�porosidad�de�15�
%.�La�fracción�gruesa�representa�el�70�%�de�la�pared,�es�tamaño�limo�–�arena�
fina,� con� algunos� individuos� que� alcanzan� un� tamaño� de� arena� media.� Esta�
fracción� esta� compuesta� por� cuarzo,� K� �� feldespato,� fragmentos� líticos� y�mica�
(muscovita).�Forman�también�parte�de�esta�fracción�abundantes�fibras�vegetales�
de� la�bosta,�birrefringentes�y�de�distintos� tamaños�(Fig.�27�B).�La� fracción� fina�
esta�compuesta�por�materia�orgánica�conformada�por�bosta�degradada�y�ojuelas�
de�materia�orgánica�color�castaño�rojizo.�El�relleno�posee�una�porosidad�menor�a�
la�observada�en�la�pared.�La�fracción�gruesa�representa�el�80�%�del�relleno,�es�
tamaño�arena�fina�–�limo,�y�esta�compuesta�principalmente�por�fibras�vegetales�
de� la�bosta�(Fig.�27�B).�Se�reconocen�también�en�esta�fracción�plagioclasa,�K���
feldespato,� cuarzo,� fragmentos� líticos� y�mica.� La� parte� del� relleno� en� contacto�
con� la� pared,� posee� fibras� vegetales� de� la� bosta� orientadas�de� forma�paralela,�
ordenamiento� que� se� pierde� hacia� el� interior� del� relleno� donde� se� hace� más�
caótico.�
�
�
�

� ���
�
�!�%�&4;��Micromorfología�de�bolas�de�cría�de��������sp.�1.���Corte�longitudinal�de�una�de�las�mitades�de�una�bola�de�cría,�donde�se�distingue�la�pared�(flechas�blancas)�y�el�relleno,� este� último� constituido� fundamentalmente� por� fibras� vegetales� de� la� bosta.�Escala:�700�^;����Detalle�de� la�pared� (indicado�su�espesor�por� la�escala:�450�^)�y�del�relleno,�donde�ambos�muestran�fibras�vegetales�de�la�bosta�(flechas�amarillas)�y�ojuelas�de�materia�orgánca�rojiza�(flecha�blanca),�en�diferente�proporción.��
�
5.2.9.3���Estructura�del�nido�
De� los�once�ejemplares�colocados�en� los�terrarios,�como�ya�fue�descripto�
en�la�metodología,�se�observaron�tres�parejas�y�dos�individuos�solos,�moldelando�
(Fig.� 26� F)� y� rolando� (Fig.� 26� G� y� H)� una� porción� de� bosta.� Se� registraron�
también� otros� comportamientos.� Los� terrarios� fueron� excavados� prolijamente,�
pero� a� pesar� de� ello,� no� se� encontraron� túneles� ni� cámaras,� ni� ninguna� otra�
estructura� relacionada� con� los� nidos.� Del� tamizado� del� material� del� suelo�
removido�de�los�terrarios,�se�halló�un�total�de�nueve�bolas,�de�las�cuales�sólo�una�
es� de� cría,�mientras� que� el� resto� corresponde� a� 6� bolas� de� provisión,� algunas�
intactas,�otras�con�cortes�o�desmenuzadas�(Fig.�26�D�y�E).�
�
6�4�2<#��������"B�4
5.2.10.1���Estructura�de�la�bola�de�cría�

� ���
El�único�ejemplar�estudiado�posee�un�diseño�muy�particular,�donde�pueden�
distinguirse�dos�cámaras�unidas�entre�si,�de�las�cuales�la�inferior�es�más�grande�
que�la�superior�(Fig.�28�A�y�B).�La�cámara�mayor�es�esférica�con�un�eje�vertical�
de�1,3�cm�de�largo�y�un�eje�horizontal,�diámetro�ecuatorial,�de�1,4�cm�de�largo�
(Fig.�28�A�y�B).�La�cámara�más�pequeña,�prácticamente�cilíndrica,�posee�un�eje�
vertical� de� 7,7� mm� de� largo,� una� base� de� 7,7� mm� de� ancho� y� el� extremo�
superior,�donde�se�abre�el�canal�de�aireación,�de�5�mm�de�ancho�(Fig.�28�A�y�B).�
Externamente,�se�observa�material�del�suelo�cubriendo�la�superficie�de�la�bola�de�
cría.�En�la�cámara�menor,�puede�distinguirse�que�este�material�forma�una�línea�
de� relieve�que� la�cruza�diagonalmente,�desde�el�agujero�del� canal�de�aireación�
hasta� la�base�de�dicha�cámara,�junto�en�el�contacto�con�la�cámara�mayor�(Fig.�
28�C).�En�el�corte�de�la�bola�de�cría�(Fig.�28�B),�puede�distinguirse�esta�pared,�
constituida�por�material�del�suelo,�de�2,2�mm�de�espesor,�el�cual�se�conserva�sin�
variación�alrededor�de�toda�la�bola.�La�cámara�ovígera�se�encuentra�ubicada�en�
el�interior�de�la�cámara�menor,�y�posee�dos�comunicaciones,�una�superior�y�otra�
inferior� (Fig.� 28� B).� La� primera� corresponde� con� la� abertura� del� canal� de�
aireación,� se� encuentra� tapada� por� bosta,� y� comunica� el� interior� de� la� cámara�
ovígera�con�el�exterior�de�la�bola�de�cría,�mientras�que�la�segunda�comunica�el�
interior� de� esta� cámara� con� la� cavidad� excavada� por� la� larva� en� la� materia�
orgánica�aprovisionada�en�el�interior�de�la�cámara�mayor�(Fig.�28�B).�
�
�
�
�
�

� ���
�
�!�%�&4:��Bolas�de�cría�de��������sp.2.���Bola�de�cría�biesférica,�conformada�por�una�cámara� inferior� (flecha�amarilla),�mayor� y� esférica,�y�una�superior� (flecha�blanca)�más�pequeña� y� alargada;� ��� Bola� de� cría� cortada� por� el� eje� del� canal� de� aireación.� Se�distingue� la� cámara� ovígera� (flecha� blanca)� conectada� al� exterior� por� una� perforación�pequeña� en� el� techo,� y� con� las� provisiones� por� una� abertura� excavada� por� la� larva�(preservada� en� el� relleno� de� la� cámara� inferior);� ��� Detalle� del� cierre� de� la� cámara�superior,� el� cual� se� distingue� por� su� relieve� curvo� (flecha� blanca)� que� cruza� desde� el�canal� de� aireación� hasta� el� contacto� con� la� cámara� inferior.� Escalas:� 1� cm.� ��� Dos�ejemplares�de�������� sp.� acarreando�un�pichón�de�Bicho�Feo� ()�������� ����������,�Tyrannidae)�en�Horco�Molle�(Tucumán).�
�
5.2.10.2���Micromorfología�de�la�bola�de�cría�
La�pared�de� la�bola�de�cría�posee�un�espesor�que�mide�de�750�^�a�1,25�
mm� (Fig.� 29�A).� La�microestructura� es�maciza,� con�una�porosidad�de�5�%.� La�
fracción�gruesa�es�tamaño�limo�–�arena�media,�encontrándose�algunos�individuos�
tamaño�arena�gruesa�–�arena�muy�gruesa.�Esta�fracción,�que�representa�el�25�%�
de� la� pared,� esta� compuesta� por� fragmentos� líticos,� cuarzo,� plagioclasa,� K� ��

� ���
feldespato,� mica� y� algunos� granos� de� mayor� tamaño� que� se� encuentran� muy�
alterados.�La�fracción�fina�esta�compuesta�por�material�arcilloso�muy�embebido�
en� materia� orgánica.� El� relleno� posee� una� microestructura� maciza� con� una�
porosidad�menor�al�5�%.�La�fracción�gruesa�representa�el�20�%�del�relleno�y�es�
similar�en�composición�y�textura�a�la�ya�descripta�para�la�pared.�El�relleno�esta�
compuesto� por� abundante� materia� orgánica� (no� se� observan� fibras� vegetales)�
(Fig.�29�A�C).�Componen�también�esta�fracción,�pequeñas�estructuras�de�formas�
y� tamaños� (entre� 400� ^� y� 900� ^)� diversos,� los� cuales� son� compatibles� con�
fragmentos� de� hueso� (Fig.� 29� B� y� C).� La� fracción� fina� está� compuesta� por�
material�arcilloso�muy�embebido�en�materia�orgánica�oscura.�La�cámara�ovígera�
presenta,�por�sectores,�fragmentos�que�podrían�ser�porciones�de�la�pared.�Estos�
fragmentos� poseen� las� mismas� características� en� composición,� textura� y�
micromorfología� que� las� descriptas� para� la� pared� de� la� bola,� pero� con� una�
porosidad�apenas�mayor�(6���7�%).��
�
�
�!�%�&47��Micromorfología�de�bolas�de�cría�de��������sp.2.���Relleno�(rico�en�materia�orgánica� oscura)� y� pared� (indicada� con� flechas� amarillas)� de� una� bola� de� cría.� Escala:�800�^;��#���Detalle�del�relleno�sin�(�)�y�con�(�)�nicoles�cruzados,�donde�se�distinguen�pequeñas�estructuras�compatibles�con�fragmenos�de�huesos�(flechas�amarillas)�inmersas�en�la�materia�orgánica�oscura�y�amorfa.�Escalas:�400�^.���
�
�

� ���
5.2.10.3���Estructura�del�nido�
En� el� único� caso,� la� bola� de� cría� fue� hallada� junto� al� escarabajo� adulto�
productor,� debajo� de� una� roca� pequeña.�No� se� encontraron� túneles,� ya� que� el�
nido�es�superficial,�y�la�bola�de�cría�se�encontraba�orientada�horizontalmente,�es�
decir,�con�la�cámara�ovígera�paralela�a�la�superficie�del�suelo.�����
�
6�4�22H���������'����� ���
�
5.2.11.1���Estructura�de�la�cámara�pupal�
Las� cámaras� pupales� usadas� para� esta� descripción� fueron� halladas� de�
manera�aislada,�todas�fueron�colectadas�directamente�en�la�superficie�(Fig.�30�A)�
o� en� el�material� removido�del� suelo� (Fig.� 30�B)� y� ningunda� fue� encontrada� en�
relación�con�un�nido.�Debido�al�hallazgo�de�un�escarabajo�adulto�conservado�sin�
emerger� en� el� interior� de� una� de� estas� cámaras� (Fig.� 30� C� y� D),� pudo�
determinarse� la� especie� productora.� Una� de� las� cámaras� subesféricas� medida,�
posee�un�eje�mayor�de�1,7�cm�de�largo�y�uno�menor�de�1,4�cm�de�largo.�Todas�
ellas� se� encuentran� conformadas� por� pequeños� ������ de� un�material� oscuro,�
que�se�yuxtaponen�constituyendo�una�banda�de�material�que�se�enrolla�alrededor�
de�la�cavidad�central�hueca�formando�un�helicoide�(FIg.�B�y�E).�En�los�ejemplares�
completos,�no�se�observan�perforaciones�ni�canales�de�aireación,�y�el�helicoide�se�
encuentra�cerrado�hacia�los�polos�(Fig.�30�E).�En�los�cortes�de�las�cámaras�puede�
observarse�que� la�cavidad�se�encuentra�tapizada�por�una�fina�capa�de�material�
oscuro�(Fig.�30�B),�que�cubre�y�tapa�el�relieve�de�los�������desde�el�interior.�La�
pared,�formada�por�el�helicoide�y�el�tapiz,�mide�de�1�mm�a�2,8�mm�de�espesor.��������
�
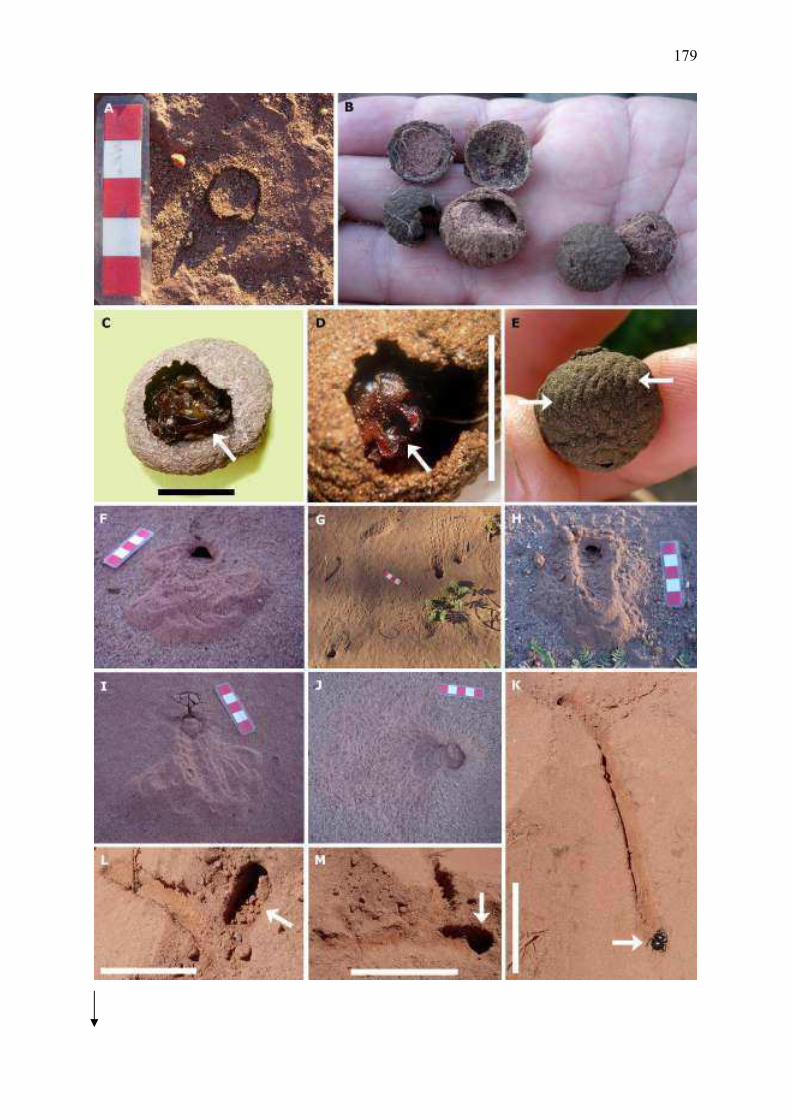
� ��
�

� ���
�!�%�& 5<� Nido� y� cámaras� pupales� de� $������������ �����.� #�� Cámaras� pupales�colectadas� ��� ����� ()� y� de� material� del� suelo� removido� (�);��#�� Cámara� pupal� que�conserva� en� el� interior� un� adulto� (flechas� blancas)� sin� emerger.� Escalas:� 1� cm;� ��Cámara� pupal� completa� y� cerrada.� Se� distinguen� en� la� superficie� externa� los� ������(flechas�blancas)�que�conforman�las�bandas�de�la�pared�helicoidal;��#��Diferentes�tipos�de�entradas�a�los�nidos:���Tipo�1.�Entrada�abierta�y�asociada�a�montículos�de�material�de�suelo�en�forma�de�medialuna,����Tipo�2.�Entrada�abierta�y�asociada�a�rastrilladas,����Tipo�3.�Entrada�que�combina�los�tipos�1�y�2,��#���Tipo�4.�Entrada�cerrada�con�material�del�suelo,�que�además�combina�los�tipos�1�y�2;�-#���Nido�conformado�por�un�túnel�simple,�vacío�y�con�un�adulto�en�su�extremo�más�distal�(-,�flecha�blanca),�que�se�ramifica�cerca�de�la�superficie�en�un�túnel�corto�(�,�flecha�blanca)�conectado�a�ella�por�una�perforación�(�,�flecha�blanca).�����
5.2.11.2���Micromorfología�de�la�cámara�pupal��
La�pared�de�la�cámara�pupal�abierta�posee�un�espesor�más�delgado�(500�
^)�hacia�el�agujero�de�emergencia�(Fig.�31�A),�y�más�grueso�(3,2�mm)�hacia�el�
polo� opuesto� (Fig.� 31� B).� La� microestructura� es� de� grano� simple,� con� una�
porosidad� superior� al� 30� %� (intergranular),� y� los� granos� se� encuentran�
recubiertos� parcialmente� por� materia� orgánica� color� castaño� rojizo,� la� cual� se�
extiende� formando� puentes� entre� estos� (Fig.� 31�A� y�B).� La� fracción� gruesa� es�
tamaño� limo�–�arena�media,�representa�el�60�%�de� la�pared�y�esta�compuesta�
por� fragmentos� líticos,� cuarzo,� plagioclasa,� K� �� feldespato,� mica� (biotita� y�
muscovita)�y�minerales�pesados�(piroxeno).�En�esta�fracción�se�observa�también�
escasa�cantidad�de�fibras�vegetales�de�bosta.�La�fracción�fina�esta�representada�
por� materia� orgánica� color� castaño� rojiza� que� envuelve� gran� parte� de� los�
minerales�y�que�se�continúa�parcialmente�delimitando�groseros������.�Materia�
orgánica� con� estas� mismas� características,� se� encuentra� también� tapizando� la�
superficie� interna� de� la� cámara� pupal� alcanzando� un� espesor�máximo� de� 50� ^�
(Fig.�31�C).�

� ��
�
�!�%�&52��Micromorfología�de�cámaras�pupales�de�$�����������������.�#��Pared�de�la�cámara�pupal,�más�delgada�hacia�el�agujero�de�emergencia�()�y�más�engrosada�hacia�el�lado�opuesto�(�).�Se�distingue�en�ambos�la�materia�orgánica�rojiza�(flechas�amarillas)�que�tapiza�los�granos�y�que�los�conecta�formando�puentes�entre�ellos.�Escalas:�1,5�mm;���� Detalle� del� tapiz� de� materia� orgánica� (flechas� blancas)� que� recubre� la� superficie�interna�de�la�cámara�pupal.�Escala:�120�^.�
�
5.2.11.3���Estructura�del�nido�
En�el� trabajo�de�campo�realizado�en�Pituil,�pudieron�distinguirse�distintos�
tipos� de� entradas� en� los� nidos� de� $�� �����,� según� diferentes� características�
externas:�1)�perforación�en�el�suelo�asociada�a�montículos�pequeños�de�material�
del�suelo�en�forma�de�media�luna,�agrupados�en�la�superficie�en�el�lado�opuesto�
al� cual� el� escarabajo� excava� el� túnel� (Fig.� 30� F);� 2)� perforación� asociada� a�
rastilladas,�de� las� frecuentes�entradas�y�salidas�del� escarabajo�al�nido� (Fig.�30�
G);�3)�1�y�2�juntos�(Fig.�30�H);�4)�1�y�2�juntos,�o�simplemente�2,�en�los�cuales�la�
perforación� ha� sido� cerrada� por� el� escarabajo� desde� el� interior� del� nido,� con�
material�del�suelo�transportado�a�la�superficie�desde�el�fondo�del�túnel�(Fig.�30�I�
y�J).�En�todos�estos�casos,�las�perforaciónes�no�superan�los�2�cm�de�diámetro,�se�
encuentran�excavadas�de�manera�oblicua�a�la�superficie,�y�en�la�mayoría�de�las�
abiertas,�se�observa�material�del�suelo�tapando�parcialmente�la�entrada,�dándole�
a�la�perforación�una�forma�general�de�semicírculo�(Fig.�30�F).��

� ���
El� único� nido� hallado� (Fig.� 30� K)� se� encuentra� conformado� por� un� túnel�
vacío�de�12�cm�de�largo�y�1,3�cm�de�diámetro,�inclinado�aproximadamente�45º�
en�relación�con� la�vertical�y�excavado�a�10�de�profundidad.�En�el�extremo�más�
distal� del� túnel� se� encuentra� un� adulto� activo.� Hacia� la� entrada,� este� túnel� se�
bifurca�en�una�rama�vacía,�corta�y�relativamente�paralela�a�la�superficie�del�suelo�
(Fig.�30�L),�que�se�comunica�con�el�exterior�por�medio�de�una�perforación�(Fig.�
30�M).���
�
6�4�24H���������'���������� (��
�
5.2.12.1���Estructura�del�nido�
Los�dos�nidos�hallados�se�encuentran�conformados�por�túneles�y�por�una�
cámara� de� almacenamiento� pequeña� excavada�muy� cerca� de� la� superficie.� En�
uno� de� los� nidos,� la� entrada� de� 1,8� cm� de� ancho� (Fig.� 32� A),� similar� a� la�
descripta�como�tipo�1�de�$��������(Fig.�30�F),�se�comunica�con�un�túnel�corto�de�
2,3�cm�de�diámetro,�paralelo�a� la�superficie,�poco�profundo,�que�se�hace�curvo�
(Fig.� 32� B)� hacia� su� desembocadura� en� una� cámara� de� forma� no� definida,� de�
aproximadamente�3�cm�de�ancho�(tal�vez�un�ensanchamiento�del�mismo�túnel),�
en�la�que�se�aloja�el�escarabajo�(muy�activo)�junto�con�������fecales�de�roedor�
(Fig.�32�C�y�D).�Esta�cámara�se�comunica�hacia�el� lado�contrario�con�un� túnel�
corto,� recto� y� levemente� inclinado,� de� 8� cm� de� largo� y� excavado� a� mayor�
profundidad,�que�se�encuentra�vacío�(Fig.�32�D�y�E).�En�el�segundo�caso,�se�pudo�
registrar�el�aprovisionamiento�del�nido�por�parte�del�escarabajo�(Fig.�32�F),�que�
al�concluir�este�preceso,�es�cerrado�por�dentro�dejando�una�entrada�(Fig.�32�G)�
similar� a� la� tipo� 4� de� $�� ������ (Fig.� 30� I� y� J).� Esta� entrada� esta� inclinada�
respecto�a�la�superficie�(Fig.�32�H),�mide�1,8�cm�de�ancho�y�se�comunica�con�un�

� ���
túnel� de� 13� cm� de� largo,� con� un� primer� tramo� corto� (3� cm� de� largo�
aproximadamente)� paralelo� a� la� superficie,� y� el� resto� inclinado� 45º,�
aproximadamente�(Fig.�32�I).�Este�túnel�desemboca,�a�una�profundidad�de�5�cm,�
en�una�cámara�muy�pequeña,�de�no�más�de�1,5�cm�de�díametro,�en� la�que�se�
encuentra�un������ fecal�de�oveja�muy�seco,�con�marcas�de�alimentación�(Fig.�
32� I).� Hacia� un� costado� de� esta� cámara,� en� la� pared� del� túnel� original,� se�
encuentra� excavada� una� entrada� circular� de� 1,7� cm� de� diámetro,� la� cual� se�
encuentra� bloqueada� por� pequeños� clastos� rodados� (Fig.� 32� I).� Una� vez�
retirados,� esta� entrada� se� comunica� con� un� nuevo� túnel� curvo,� de� 2,5� cm� de�
diámetro�y�6�cm�de�largo,�en�el�cual�se�aloja�un�escarabajo�adulto�junto�con�otro�
�����fecal�ovino�(Fig.�32�J),�el�cual�también�muestra�marcas�en�su�superficie.�En�
el� extremo� más� distal� de� este� túnel,� se� encontró� una� porción� pequeña� de�
excremento� desmenuzado.� En� ninguno� de� los� casos� mencionados,� se�
encontraron�las�cámaras�pupales�asociadas�ni�cámaras�ovígeras�o�huevos.��
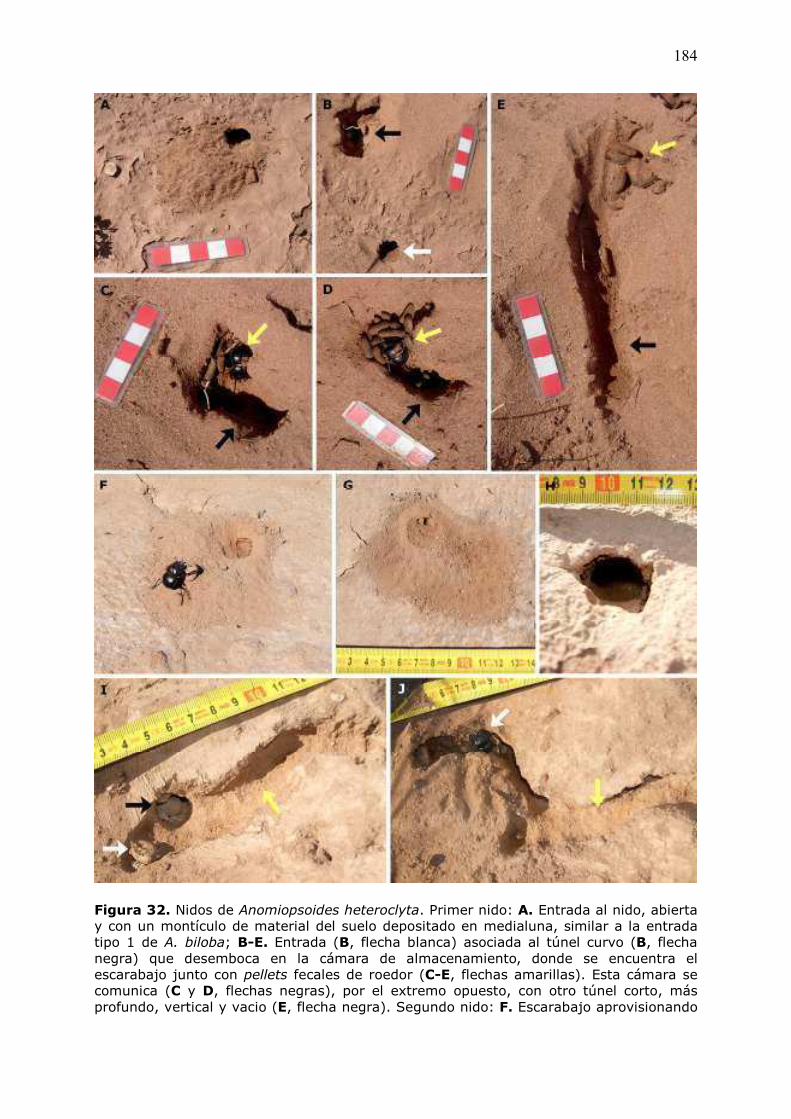
� ���
�
�!�%�&54�Nidos�de�$���������������� �&�.�Primer�nido:��Entrada�al�nido,�abierta�y�con�un�montículo�de�material�del�suelo�depositado�en�medialuna,�similar�a�la�entrada�tipo�1�de�$�������;��#��Entrada� (�,� flecha�blanca)� asociada�al� túnel� curvo� (�,� flecha�negra)� que� desemboca� en� la� cámara� de� almacenamiento,� donde� se� encuentra� el�escarabajo� junto�con������� fecales�de�roedor�(�#�,� flechas�amarillas).�Esta�cámara�se�comunica� (�� y��,� flechas� negras),� por� el� extremo� opuesto,� con� otro� túnel� corto,�más�profundo,�vertical�y�vacio�(�,�flecha�negra).�Segundo�nido:��Escarabajo�aprovisionando�

� ���
su�nido�con�������fecales�ovinos;���Cierre�de�la�entrada�al�nido,�similar�al�tipo�4�de�$�������;��#���La�entrada�al�nido�(�),�al� remover�el�material�del�suelo,�es� inclinada�y�se�comunica�hacia�el� interior�con�un�túnel�corto�y�curvo,�con�estrías�en�su�superficie�(�#�,�flechas�amarillas)�y� con�un������ fecal�depositado�en�el�extremo�más�distal� (�,� flecha�blanca).� Una� segunda� entrada,� de� sección� circular� y� obliterada� por� clastos� (�,� flecha�negra),� se� comunica� con� un� segundo� túnel,� corto� y� curvo,� en� el� que� se� aloja� el�escarabajo�con�otro������fecal�ovino�(�,�flecha�blanca).�������
�
6�4�25H����������(� �����
�
5.2.13.1���Estructura�del�nido�
Los�dos�nidos�estudiados,�se�encuentran�formados�por�un�único�túnel�cada�
uno�(Fig.�33�A�y�B),�los�cuales�son�inclinados�en�relación�con�la�superficie�(35º�a�
45º)� (Fig.� 33� A),� y� cuanto� más� profundos,� típicamente� sinuosos� (Fig.� 33� B).�
Miden�2,5� cm�de�diámetro,� 50� cm�de� largo� y� alcanzan�40� cm�de�profundidad,�
aproximadamente.�En�uno�de�estos�nidos�(túnel�de�52�cm�de�largo,�Fig.�33�A)�se�
encontró� en� la� porción� más� terminal� (4� cm),� junto� con� un� escarabajo� activo,�
bosta�de�vaca�seca�y�desmenuzada.�En�el�otro�nido�(Fig.�33�B),�si�bien�no�pudo�
ser�medida�por�ser�cortada�al�excavar,�se�halló�en�relación�con�el�túnel�principal�
una�cámara�de�provisión�con�������fecales�de�cabra�deshidratados�(Fig.�33�C).�
En�el�extremo�más�distal�de�este�túnel�se�encontró�un�escarabajo�activo�(Fig.�33�
B).��
�

� ���
�!�%�& 55�� Nidos� de� '� ������ & �����. � Nido� conformado� por� un� túnel� largo,�inclinado� y� levemente� sinuoso,� con� bosta� desmenuzada� y� seca� (flecha� blanca)�aprovisionada� en� su� extremo�más� distal;��#��� Nido� conformado� por� un� túnel� principal�sinuoso�(�),�más�vertical,�con�un�escarabajo�activo�en�su�estremo�más�distal�(�,�flecha�amarilla),�asociado�a�una�cámara�(no�se�distingue�por�estar�rota)�con�������fecales�de�cabra�deshidratados�(�,�flechas�negras)�aprovisionados�en�su�interior.�
6�5# ������ � ����� �� � ���������� �� ������������ �� ���
����� ?����� @ ���� ��A@ �������� ������ � ?������ �����#
���������������A
6�5�2 # B&�!*!> @ $� "!$&$@ &)% $& *!& ��'&=!+& . $�"&B&�!*!> $�
��������������� '&"�*%� *!&
�
La� Figura� 34� exhibe� la� abundancia� relativa� de� las� icnoespecies� de�
������������(A),�y�el�número�máximo�de�especímenes�por�m2�(densidad,�B),�a�
lo� largo�de� la� secuencia� estudiada.�Desde� la� estaca�9501�hasta� la� 9505,� en� la�
parte� inferior� de� las� secuencia,� no� se� registra� evidencia� de������������.� El�
primer�registro�(el�más�antiguo)�ocurre�en�la�sección�superior�del�Miembro�Gran�
Barranca� (estaca� 9506,� Eoceno�Medio� superior),� y� está� representado� por� unos�
pocos� y� mal� preservados� ejemplares� de� ��� �������� (14,29� %)� y� varios�
especímenes� indeterminados� (��� �����),� localizados� en� un� parche� con� una�
densidad� máxima� de� 4� especímenes� por� m2.� La� estaca� 9508� muestra� más�
cantidad� de� ejemplares� de� ��� �������� (38,46� %),� menos� especímenes�
indeterminados� (��� �����),� y� el� primer� registro� de���� ������� ��� representado�
por�un�solo�ejemplar.�En�esta�estaca,� los�ejemplares�se�encuentran�distribuidos�
en�parches,�con�una�densidad�máxima�de�3�especímenes�por�m2.�En�el�Miembro�
Rosado�no�se�registraron�ejemplares�de������������.��

� ���
La�parte�inferior�del�Miembro�Puesto�Almendra,�estaca�9508.5�09,�registra�
un�importante�incremento�en�la�abundancia�de�������������(154�especímenes�
colectados)�y�en� la�densidad,�que�alcanza�un�valor�máximo�de�40�especímenes�
por� m2,� y� en� cuanto� a� la� diversidad,� se� registran� por� primera� vez� todas� las�
icnoespecies� de� la� Formación� Sarmiento.� En� esta� estaca,� ��� �������� es� la�
icnoespecie� más� abundante� (49,35� %).� La� abundancia� del� resto� de� las�
icnoespecies,� ��� isp.� A,� ��� ������,� ��� ������,� ��� ������� ��� y� especímenes�
indeterminados�(��������),�es�de�12,99�%,�11,69�%,�3,9�%,�1,95�%�y�20,12��%,�
respectivamente.� La� estaca� 9510�muestra� una� icnodiversidad� similar,� pero� con�
otra�abundancia�relativa:�����������,�45,61�%;���� isp.�A,�14,03�%;����������,�
9,65� %;� ��� ������� y� ��� ������� ��,� 0,88� %� en� cada� caso,� ya� que� estan�
representadas�por�un�solo�ejemplar�cada�una;�y�los�especímenes�indeterminados�
(��������),�26.32�%.�La�máxima�densidad�medida�para�esta�estaca�es�alrededor�
de�100�ejemplares�por�m2,�los�cuales�se�encuentran�uniformemente�distribuidos�
a�lo�largo�del�nivel�estratigráfico.�La�estaca�9510.5�enseña�una�disminución�en�la�
abundancia� de� ������������ (menos� de� 100� especímenes� colectados),� una�
densidad�máxima�de�60� especímenes�por�m2,� uniformemente�distribuídos,� y� la�
presencia� de� todas� las� icnoespecies,� con� la� excepción� de� ��� ������� ��.� La�
abundancia�relativa�de�las�icnoespecies�en�esta�estaca�es:�����������,�31,43�%;�
��� isp.�A,�15,71�%;����������,� 11,43�%;���� ������� 4,29�%;�y� los�especímenes�
indeterminados�(��������),�37,14�%.��
La� estaca� 9512,� en� el� Miembro� Puesto� Almendra� Superior,� exhibe� una�
disminución� de� la� diversidad,� registrándose� solamente���� isp.� A� (10,58�%),����
������� (3,85� %),� ��� �������� (40,35� %)� y� especímenes� indeterminados� (���
�����)� (45.19� %).� Sin� embargo,� la� máxima� densidad� medida� es� de� 60�
especímenes�por�m2�(uniformemente�distribuídos),�similar�al�valor�medido�en�la�

� ���
estaca� anterior.� La� estaca� 9512�13� registra� un� importante� incremento� en� la�
proporción� de� ��� isp.� A� (23,72� %)� en� relación� con� las� icnoespecies� más�
frecuentes� (����������,� 39,24�%;���� ������,� 6,41�%�y���� �����,� 26,92�%),� y�
muestra� además� que� ��� ������� y� ��� ������� ��� se� encuentran� representadas�
nuevamente� por� muy� pocos� ejemplares� (4� y� 1,� respectivamente).� La� máxima�
densidad�medida�en�esta�estaca�es�de�100�especímenes�por�m2,�uniformemente�
distribuidos,�y�la�abundancia�reflejada�en�156�especímenes�colectados,�es�similar�
a� la�observada�en� las�estacas�9508.5�09�y�9510.� La�estaca�9514.5�muestra� la�
misma�icnodiversidad�que�la�estaca�9512�13,�pero�una�disminución�de�la�máxima�
densidad� medida,� la� cual� es� de� 6� �� 8� especímenes� por� m2� (uniformemente�
distribuidos),� y� de� la� abundancia� (menos� de� 100� especímenes� colectados).� En�
esta�estaca�����������y����������� ���se�encuentran�también�en�baja�proporción�
(1� y� 2� especímenes,� respectivamente),� mientras� que� ��� �������� y� ��� ������
poseen�una�proporción�similar�a�la�que�presentan�en�la�estaca�9512�(45,65�%�y�
40,22�%,�respectivamente).�La�estaca�9516�registra�una�importante�disminución�
en� la� abundancia� (25� especímenes� colectados),� máxima� densidad� (6�
especímenes�por�m2,�distribuídos�en�parches),�y�en�la�diversidad,�encontrándose�
presentes������������(40�%),����isp.�A�(8�%),������������(4�%,�1�solo�ejemplar)�
y����������(48�%).��
Desde� la�estaca�9517�17.5�hasta� la�9520�20.5,�se�observan�����������,�
���isp.�A,�����������y���������,�en�proporciones�similares.�En�las�estacas�9517.8�
17.9�y�9518�19,�����������es�más�abundante�que�en� las�otras�estacas�(7,75�y�
12,5�%),�y�en�la�estaca�9517�17.5,����������(64,1�%)�es�más�abundante�que����
�������� (29,06�%).� Las� estacas�9517.6�17.7,� 9518�19�y�9520�20.5�muestran�
similar�abundancia�de������������,�colectándose�un�total�de�64�especímenes�
en� las� dos�primeras,� y� 66� en� la� última.� En� cuanto� a� la� densidad,� los�máximos�

� ��
valores�medidos�entre�estos�niveles�estratigráficos,�en�los�cuales�los�ejemplares�
se� encuentran� uniformemente� distribuidos,� es� alrededor� de� 6� especímenes� por�
m2�para�las�estacas�9517�17.5�y�9518�19,�y�de�10�especímenes�por�m2�para�la�
estaca�9517.8�17.9.�La�estaca�9521�22�registra�una�importante�disminución�de�
la�abundancia�(sólo�14�especímenes�colectados),�de�la��máxima�densidad�medida�
(2� �� 3� especímenes� por� m2,� distribuidos� en� parches),� y� de� la� diversidad� de�
icnoespecies�de������������.�Se�registran�en�esta�estaca,�especímenes�de����
�������,� ��� isp.� A,� ��� ������� e� indeterminados� (��� �����),� en� proporciones�
similares�a�las�observadas�en�la�estaca�9516�(28,57�%,�14,29�%,�7,14�%�y�50�
%,�respectivamente).�Por�encima�de�esta�estaca�no�se�registran�más�ejemplares�
de�������������hacia�la�parte�superior�de�la�secuencia,�desde�la�estaca�9523�a�
la�9525.5.�
El�gráfico�de�la�Figura�34�A,�muestra�además�que�����������,����isp.�A�y�
����������se�registran�en�toda�la�secuencia�desde�su�aparición,�aunque�es�sólo����
�������� la� que� se� encuentra� presente� en� todas� las� estacas.� En� cambio,� ���
������� ���y�����������son�las�icnoespecies�más�raras�en�la�secuencia,�las�cuales�
se� encuentran� dispersas� en� distintas� estacas� y� ausentes� a� partir� de� la� estaca�
9517�17.5�en�el�primer�caso,�y�de�la�9517.6�17.7�en�el�segundo.��
�
�
�
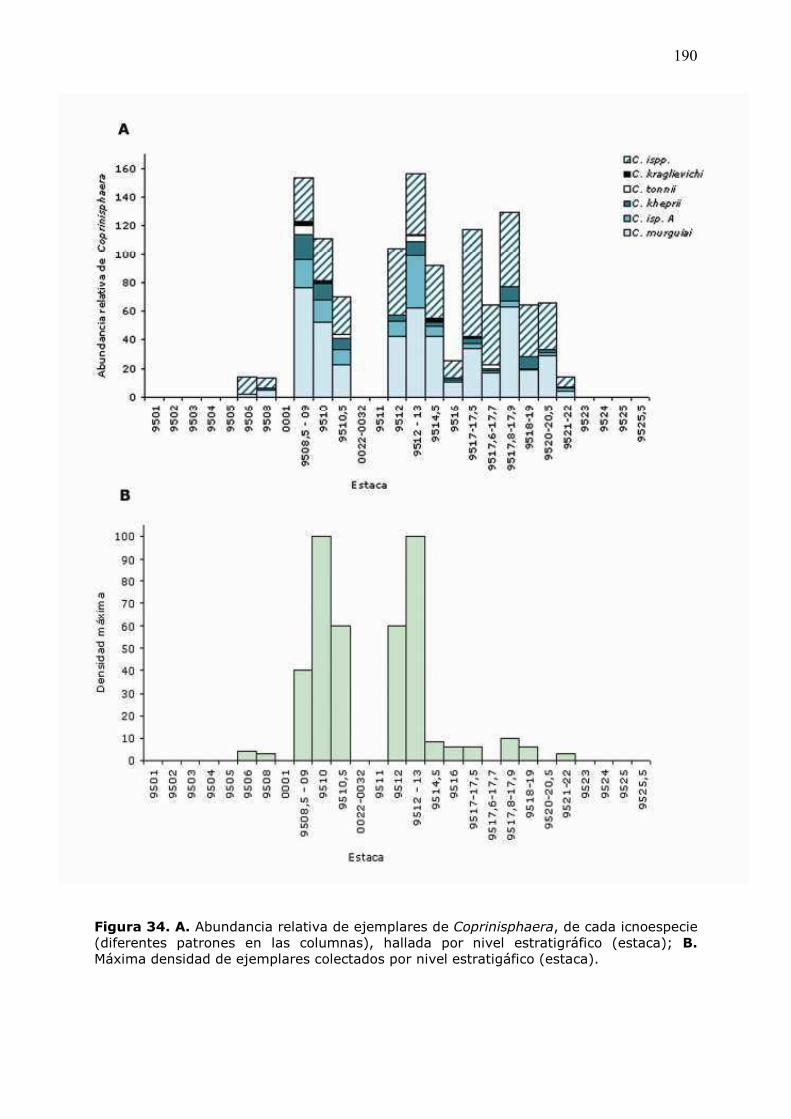
� ��
�
�!�%�&58���Abundancia�relativa�de�ejemplares�de������������,�de�cada�icnoespecie�(diferentes� patrones� en� las� columnas),� hallada� por� nivel� estratigráfico� (estaca);� ���Máxima�densidad�de�ejemplares�colectados�por�nivel�estratigáfico�(estaca).���
�

� �
6�5�4 H N'!"!" $�' =&(&Q� $� '�" �C�(B'&��" $� �������������� ?��,
�!N(�=���*%&=��!&'A
�
La� Figura� 35� A� muestra� que� desde� la� estaca� 9506� a� la� 9512� el� DE�
promedio�es�de�34,1�mm�a�37,61�mm�(N�=�429).�Desde�la�última�estaca�hasta�la�
9516� este� parámetro� se� incrementa� alcanzando� un� valor� de� 48,83� mm� (N� =�
273),� que� disminuye� en� la� estaca� 9517�17.5� hasta� 45,05�mm� (N�=� 61),� y� se�
mantiene� alrededor� de� este� valor� (43,87�mm�–�45,78�mm,�N�=�302)�hasta� la�
estaca�9521�22.�El�análisis�estadístico�refleja�que�sólo�entre�las�estacas�9512�y�
9512�13,�y�entre�esta�última�y�la�9514.5,�el�incremento�en�el�DE�es�significativo�
(�� <� 0,05).� Mientras� que� los� cambios� observados� en� el� DE� entre� las� estacas�
anterior� y� posterior� (9514.5� y� 9517�17.5,� respectivamente)� y� la� 9516,� no� son�
significativos� (��>�0,05).� Las� comparaciones� entre� estacas� en� el�DE�promedio,�
revelaron�que�existen�diferencias� significativas� (��<�0,05)�en�el� tamaño�de� las�
������������ entre� dos� grupos� de� estacas,� de� los� cuales� el� primero� se�
encuentra�conformado�por�los�valores�de�DE�de�las�estacas�9506�a�9512�13,��y�el�
segundo�de�las�estacas�9514.5�a�9521�22.��
La�Figura�35�B�presenta�el�mismo�tipo�de�análisis�que�en�la�parte�A,�pero�
en�este�caso�el�DE�está�discriminado�por�icnoespecies��El�DE�de�����������,����
isp.� A,� y���� ������,� a� lo� largo� de� la� secuencia,� posee� la�misma� tendencia� de�
cambio� que�muestra� este�mismo� parámetro� cuando� es� analizado� considerando�
todas� las� icnoespecies� juntas.�El� análisis� estadístico� refleja�que�para�estas� tres�
icnoespecies,� el� incremento� de� tamaño� registrado� entre� las� estacas� 9512�13� y�
9514.5,�es�significativo�(��<�0,05).�En�cambio,�los�incrementos�de�tamaño�de����
�������registrados�en�las�estacas�9514.5�y�9517.6�17.7,�y�de����������observado�
en�la�estaca�9508,�resultaron�ser�estadísticamente�no�significativos.�����������y����

� ��
������� ��,� a� pesar� de� estar� representadas� por� pocos� ejemplares� que� no�
soportan�un�análisis� estadístico,�mostraron� también� la�misma� tendencia�que� la�
observada� para� el� DE� promedio.� Las� comparaciones� entre� estacas� en� el� DE�
promedio�por� icnoespecie,� revela�que������������y���� ������muestran� también�
diferencias�significativas�(��<�0,05)�entre�dos�grupos�de�estacas.�Para�el�caso�de�
��� �������,� el� primer� grupo� esta� conformado� por� los� valores� de� DE� de� las�
estacas�9508�a�9512�13,�y�el� segundo�por� los�valores� incluidos�en� las�estacas�
9514.5�a�9521�22.�Para���������,�el�primer�grupo�de�valores�de�DE�va�desde�la�
estaca�9506�a�la�9512,�y�el�segundo,�desde�la�9512�13�a�la�9521�22.�Como�fue�
mencionado� más� arriba,� el� bajo� número� de� ejemplares� de� ��� ������� y� ���
������� ��� imposibilita� un� análisis� detallado.� Sin� embargo,� para� ��� ������,� los�
valores� de� DE� obtenidos� en� las� estacas� 9508.5�09� y� 9510.5� difieren�
significativamente�(��<�0,05)�del�valor�obtenido�en�la�estaca�9514.5;�y�para����
������� ��,� los� valores� de� DE� obtenidos� en� las� estacas� 9508.5�09� y� 9510�
difieren�significativamente�(��<�0,05)�del�DE�de�la�estaca�9517�17.5.��
Para� evaluar� la� posibilidad�de�que� el� incremento� o� la� disminución�de� los�
valores� de� DE� promedio� puedan� estar� influidos� por� la� desaparición� de�
especímenes�pequeños�o�la�aparición�de�los�de�mayor�tamaño,�se�requiere�de�un�
análisis�adicional.�Los�histogramas�de� la�Figura�36�evidencian�que�en� la�estaca�
9512�13�(Fig.�36�G)�aparecen�por�primera�vez� los�especímenes�de� la�categoría�
de�mayor�tamaño�(V:�60,5�mm�a�78,5�mm),�y�que�a�partir�de�la�siguiente�estaca�
(9514.5)� (Fig.� 36� H)� hasta� el� tope� de� la� secuencia,� desaparecen� casi� por�
completo� los�especímenes�más�pequeños� incluidos�en� la�clase�I�de� tamaño�(21�
mm�a�27,3�mm),�salvo�por�una�sola�excepción�en�la�estaca�9520�20.5�(Fig.�36�
N)� en� la� que� se� registra� un� solo� ejemplar.� Por� otro� lado,� los� histogramas�
muestran� también� que� hasta� la� estaca� 9512� la� mayoría� de� los� especímenes�

� ��
(moda)�están�incluidos�en�la�clase�II�de�tamaño�(II:�27,4�35,6�mm),�con�un�pico�
secundario�en�la�clase�III�(35,7�46,4�mm).�Sin�embargo,�en�la�estaca�9512�13�la�
moda� se� invierte� y� se� mueve� a� la� clase� III,� quedando� la� clase� II� como� pico�
secundario.�A�partir�de�la�estaca�9514.5,�y�hasta�el�tope�de�la�secuencia,�la�moda�
se� mantiene� en� la� clase� III� con� un� pico� secundario� en� la� clase� IV� (46,5�60,4�
mm),� con� la� excepción� de� la� estaca� 9516� en� la� cual� la� mayoría� de� los�
especímenes�registrados�pertenecen�a�la�clase�IV�de�tamaño.�Este�aumento�del�
tamaño�de� los� ejemplares� de������������,� producido�por� la� desaparición�de�
los�especímenes�más�pequeños,�y� la�aparición�de�los�más�grandes,�y�el�cambio�
en�la�moda�de�la�clase�II�de�tamaño�a�la�III�o�IV,�puede�ser�analizado�para�cada�
icnoespecie�para�saber�si� la� tendencia�es�general,�válida�para� todas�ellas;�o�de�
ser�particular�qué� icnoespecies� se�desvían�o� contribuyen�más�que�otras�a�esta�
tendencia.��
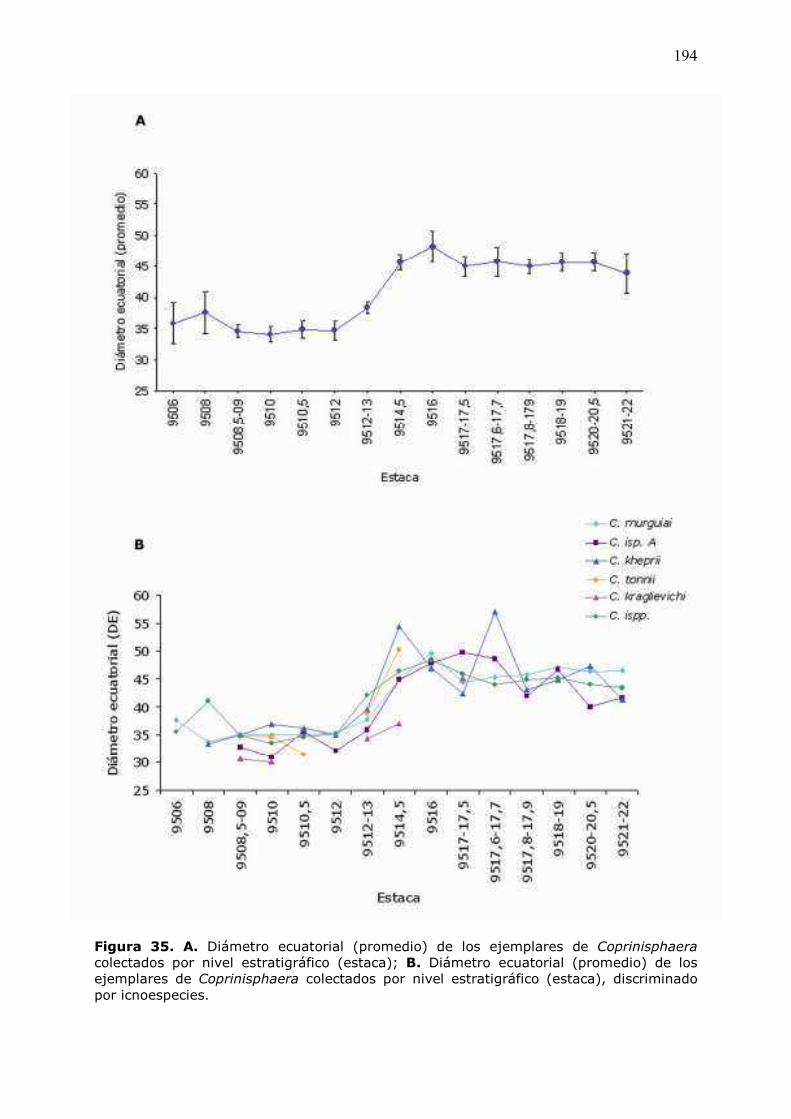
� ��
�
�!�%�& 56� �� Diámetro� ecuatorial� (promedio)� de� los� ejemplares� de� ������������colectados� por� nivel� estratigráfico� (estaca); ��� Diámetro� ecuatorial� (promedio)� de� los�ejemplares� de������������� colectados� por� nivel� estratigráfico� (estaca),� discriminado�por�icnoespecies.���
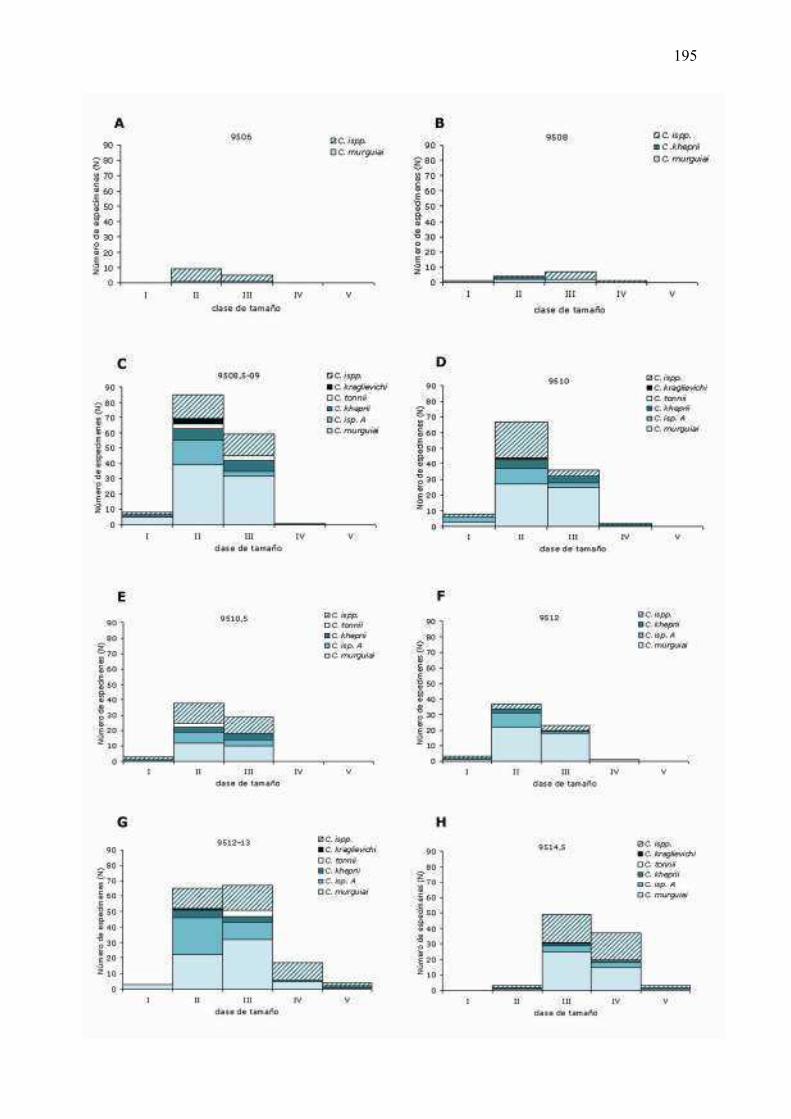
� ��
�
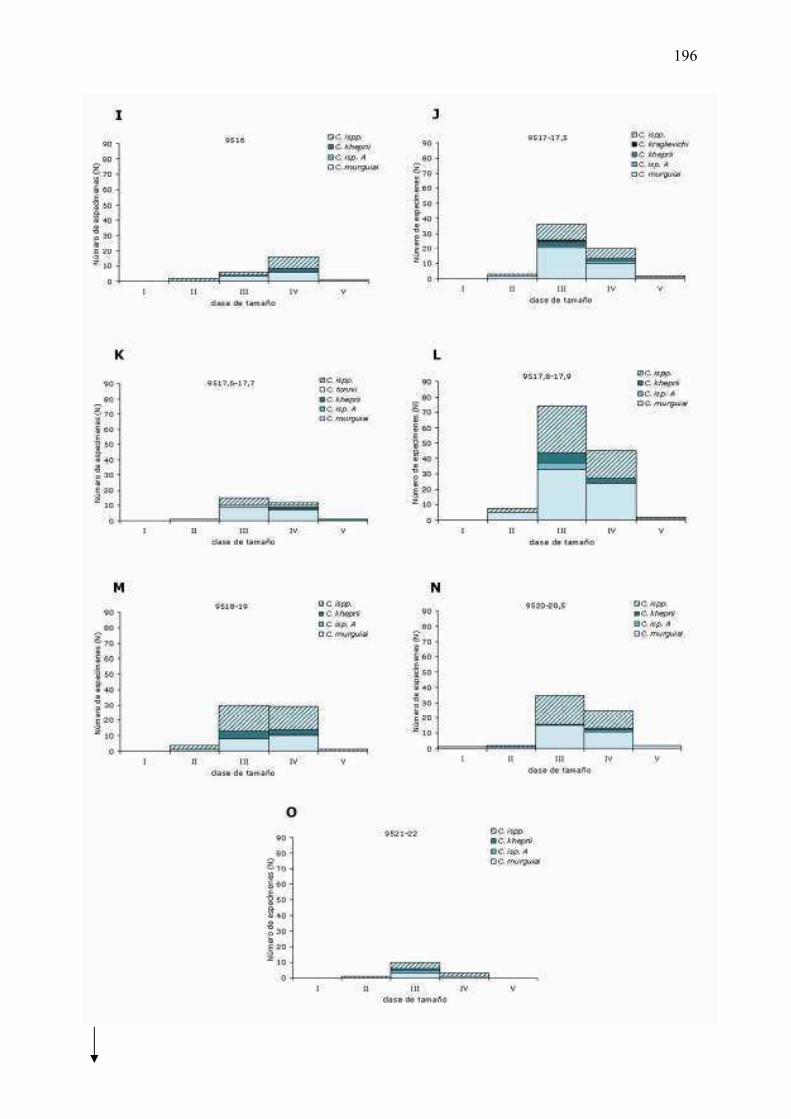
� ��
�

� ��
�!�%�& 53� Especímenes� de� ������������ (diferentes� patrones� en� las� columnas)�agrupados� por� clases� de� tamaño� (I�V),� donde� cada� histograma� representa� un� nivel�estratigráfico�(#�).�(I)�21�27,3�mm,�(II)�27,4�35,6�mm,�(III)�35,7�46,4�mm,�(IV)�46,5�60,4�mm,�y�(V)�60,5�78,6�mm.�
6�5�5H N'!"!"$�'&"!* ��"B�*!�"&��%B&$&"B��*'&"�"$�=&(&Q�
�
La�Figura�36�muestra�también�los�ejemplares�de�diferentes�icnoespecies�de�
������������agrupados�en�distintas�clases�de�DE�(I�V).�En�la�estaca�9506,����
��������y�especímenes� indeterminados�(��� �����)�se�encuentran�agrupados�en�
dos� clases�de� tamaño,� II� y� III,� es�decir� entre�27,4�y�46,4�mm�de�DE.� La�otra�
estaca�del�Mb.�Gran�Barranca,�9508,�posee�dos�nuevos�grupos�de�especímenes�
representados�por������������en�la�clase�de�menor�tamaño�(I:�21�27,3�mm),�y�
��� ������ en� la� clase� mayor� (V:� 60,4�78,6� mm).� La� clase� II� incluye� además�
ejemplares� de� ��� ������.� Las� estacas� 9508.5�09� y� 9510�muestran� las�mismas�
icnoespecies� distribuidas� en� las� cuatro� primeras� clases� de� tamaño� (I� a� IV).� En�
particular,� en� la� estaca� 9508.5�09� se� observan� ejemplares� de� ��� ������� y� ���
�������� en� la� clase� I,� ��� �������-� ��� isp.� A,� ��� ������-� ��� ������� y� ���
������� ���en� la�clase�II,� las�mismas� icnoespecies�menos����������� ��,�en� la�
clase� III,� y� solamente���� ������� en� la� clase� IV.� Mientras� que� la� estaca� 9510�
presenta� además� ejemplares� de� ��� isp.� A� en� la� clase� I� y� ejemplares�
indeterminados�(��������)�en�la�clase�IV,�y�carece�de�����������en�la�clase�III.��
En� la� estaca� 9510.5� la� clase� IV� no� registra� ejemplares,� pero� esta� clase�
vuelve�a�tener�ejemplares�en�la�estaca�9512�representados�por�����������.�En�
estas� dos� estacas���� ������� ��� no� se� encuentra� presente.� La� estaca� 9510.5�
muestra�����������y�especímenes� indeterminados� (��� �����)�en� la�clase� I,�y����
�������,����isp.�A,�����������y����������en�las�clases�II�y�III.�La�clase�II�presenta�
además�ejemplares�de����������.��La�estaca�9512�exhibe�la�misma�distribución�de�

� ��
icnoespecies,�pero�en�la�clase�I������������y����isp.�A�están�presentes�mientras�
que�����������está�ausente,�al� igual�que�sucede�con�����������en�la�clase�II.�La�
estaca� 9512�13� es� la� primera� en�mostrar� especímenes� de� todas� las� clases� de�
tamaño,� incluida� la� última� (clase� V),� la� cual� se� encuentra� representada� por�
especímenes�indeterminados�(��������)�y�por�ejemplares�de����isp.�A�y����������.�
La� clase� I� está� representada� solamente� por� ejemplares� de� ��� �������.� Las�
clases�II�y�III�muestran�ejemplares�de�����������,����isp.�A,����������,�mientras�
que� ��� ������� ��� está� presente� sólo� en� la� segunda� clase� (II),� y� ��� �������
únicamente� en� la� tercera� (clase� III).� La� clase� IV� comprende� únicamente� ���
�������,���� isp.� A� y� especímenes� indeterminados� (��� �����).� Desde� la� estaca�
9514.5�a�la�9518�19�los�especímenes�se�encuentran�distribuidos�entre�todas�las�
clases� de� tamaño,� con� la� excepción� de� la� primera� (clase� I),� y� no� revelan�
diferencias�importantes�entre�ellos�en�la�diversidad�de�icnoespecies.�Entre�estas�
estacas,����������� ���está�presente�solamente�en�las�clases�I�y�II�de�la�estaca�
9514.5,� y� en� la� clase� III� de� la� estaca�9517�17.5.�De�manera� similar,���� �������
está�presente�en� la�clase� IV�de� la�estaca�9514.5�y�en� las�clases� III�y� IV�de� la�
estaca� 9517.6�17.7.� Otra� diferencia� esta� dada� por���� ������� que� se� presenta�
más�abundante�en�las�clases�III�y�IV�de�las�estacas�9517.8�17.9�y�9518�19,�que�
en�las�otras.�En�la�estaca�9520�20.5,�todas�las�clases�de�tamaño�se�encuentran�
representadas,�incluso�la�primera�(clase�I),�con�un�solo�ejemplar�de�����������.��
La�clase� II�muestra�ejemplares�de���� isp.�A�y�especímenes� indeterminados� (���
�����),� al� igual� que� en� la� clase� IV,� la� cual� incluye� además� ejemplares� de� ���
������.� La� clase� III� exhibe� la�misma�diversidad,� con� la� excepción�de���� isp.�A�
que� está� ausente.� La� última� clase� (V)� está� únicamente� representada� por�
ejemplares� de� ��� �������.� Hacia� el� tope� de� la� secuencia,� la� estaca� 9521�22�
muestra� ejemplares� solamente� en� las� tres� clases� centrales� (II� a� IV),�

� �
representadas� por� especímenes� indeterminados� (��� �����)� en� todas� ellas,� ���
��������en�la�clases�III�y�IV,�y����isp.�A�y�����������únicamente�en�la�clase�III.��
�
6�5�8 H N'!"!" $�' � $!*� $� �"B�"�� $� '& �&��$ ?���A $�
��������������
�
La�Figura�37�A�muestra�el�índice�del�Espesor�de�la�Pared�de�������������
(IEP),�como�el�promedio�del�índice�de�todas�icnoespecies,�por�cada�estaca�de�la�
secuencia.� Desde� la� estaca� 9506� a� la� 9512,� el� IEP� estimado� difiere� no�
significativamente� (�� >� 0,05),� de� 44,23� a� 36,34.� En� la� estaca� 9512�13,� este�
índice� disminuye� significativamente� (�� <� 0,05)� alcanzando� el� menor� valor�
estimado� en� toda� la� secuencia� (IEP� =� 32,54;� N� =� 143).� Después� de� esta�
disminución,� el� IEP� muestra� un� incremento� significativo� (�� <� 0,05)� hasta� la�
estaca� 9514.5� (IEP� =� 39,92;� N� =� 89).� En� las� últimas� siete� estacas� de� la�
secuencia,�el�índice�estimado�no�difiere�significativamente�(��>�0,05),�de�37,91�a�
39,85.��
La� Figura� 37� B� revela� el� mismo� análisis,� pero� el� IEP� se� encuentra�
diferenciado�por� icnoespecies.�En� la�mayoría�de� las�estacas� las� icnoespecies�se�
encuentran�representadas�por�muy�pocos�ejemplares.�Por�esta�razón,�el�objetivo�
de� este� análisis� detallado� no� es� el� de� interpretar� los� cambios� del� IEP� en� cada�
icnoespecie,�sino�el�de�mostrar�que� los�cambios�observados�en�el�promedio�del�
índice�se�encuentran�mayormente�soportados�por�cambios�individuales�en�el�IEP�
de�las�icnoespecies.�Al�igual�que�en�el�caso�del�índice�promedio,�la�mayoría�de�las�
icnoespecies�exhiben�una�disminución�del�IEP�entre�las�estacas�9512�y�9514.5.�
Particularmente,�todas�las�icnoespecies�alcanzan�el�mínimo�valor�del�índice�en�la�
estaca�9512�13,�con�la�excepción�de�����������y���������-�las�cuales�lo�alcanzan�

� ����
en� la� estaca� 9520�20.5� y� 9512,� respectivamente.� El� análisis� estadístico� revela�
que� el� IEP� de� ��� �������� disminuye� significativamente� (�� <� 0,05)� entre� las�
estacas� 9512� y� 9512�13,� pero� que� su� incremento� posterior,� desde� la� última�
estaca�hasta� la�9514.5,�es�no�significativo�(��>�0,05).�En�cambio,�el� IEP�de����
isp.� A,� disminuye� no� significativamente� (p� >� 0,05)� desde� la� estaca� 9512� a� la�
9512�13,�pero�muestra�un�incremento�posterior�significativo�(��<�0,05)�desde�la�
última�estaca�hasta�la�9514.5.���
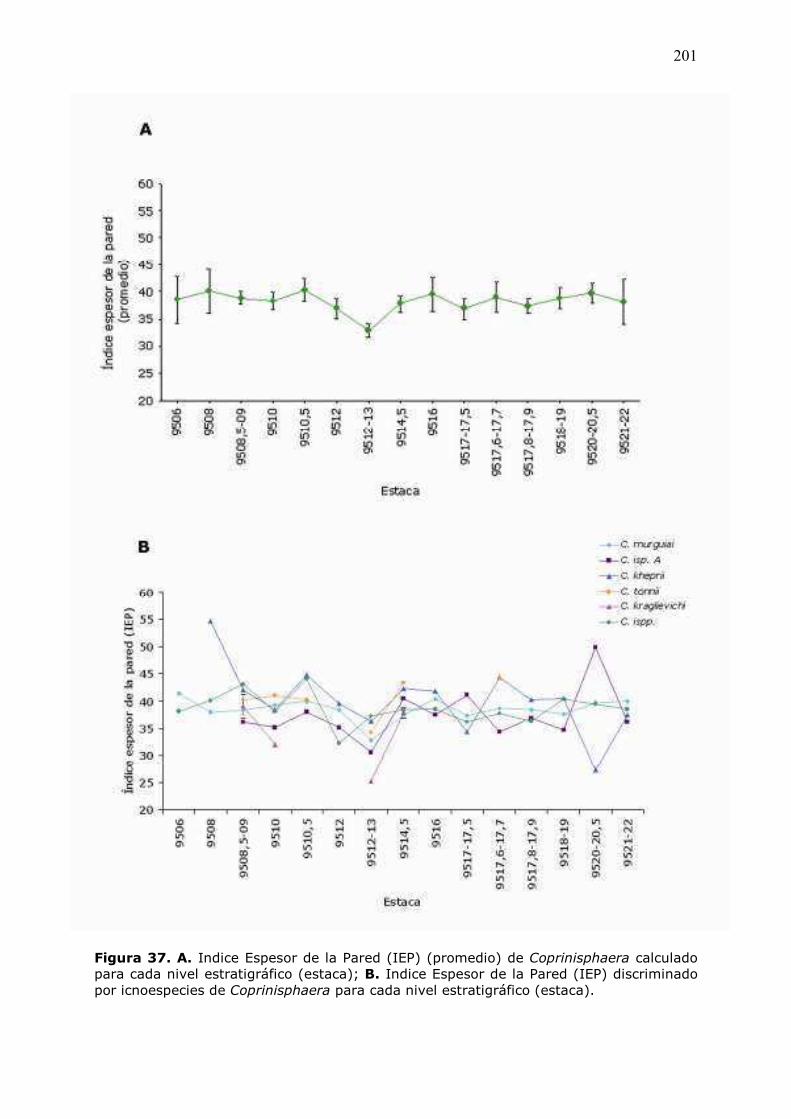
� ���
�
�!�%�&5;���Indice�Espesor�de�la�Pared�(IEP)�(promedio)�de�������������calculado�para�cada�nivel�estratigráfico�(estaca);����Indice�Espesor�de�la�Pared�(IEP)�discriminado�por�icnoespecies�de�������������para�cada�nivel�estratigráfico�(estaca).�

� ����
6�8#������������� ������� ��� ���
6�8�2#��'&"$�*�F&&*=%&'�"."%�'�*!�*% $& =�
�
Los�datos�obtenidos�en�2�bolas�muestreadas�en�Navarro�(Buenos�Aires)�y�
Chuquis� (La� Rioja),�mostraron,� � ������,� importantes� diferencias� entre� ellas� en�
relación�a� la�concentración�de� fitolitos.�En� la�primera� localidad,�en�una�bola�de�
cría� de� ��� ��� ���-� se� encontraron� 123� fitolitos� en� la� pared,� 60� en� la� bosta�
aprovisionada� y� no� consumida� por� la� larva� madura,� y� 235� fitolitos� en� una�
muestra�de�suelo�asociado.�En�cambio,�en�la�segunda�localidad,�en�una�bola�de�
cría� de���� ��������abierta,� se� encontraron� 53� fitolitos� en� la� pared,� 61� en� la�
capa�de�bosta�remanente�y�36�fitolitos�en�una�muestra�del�suelo�circundante.��
�
6�8�4#��'&"$�*�F&I>"!'�".B&'��"%�'�*!�*% $& =�
5.4.2.1� �� Comparación� del� número� total� de� fitolitos� entre� el� relleno� de�
������������y�el�paleosuelo�circundante���
�
� La�Figura�38�A�(y�Tabla�4)�muestra�el�número�total�de�fitolitos�(frecuencia�
absoluta)�resultantes�de�la�suma�de�las�dos�fracciones�(FM�+�FF),�encontrados�en�
el�paleosuelo,� relleno�y�pared�(barras�del�histograma),�de� todos� los�ejemplares�
de� ������������ estudiados.� Las� barras� muestran� también� la� frecuencia�
absoluta� de� cada� morfotipo� de� fitolito� diferenciada� por� distintos� patrones� de�
relleno,�pero�éste�análsis�se�hará�más�adelante.��
� El�análisis�estadístico�entre�el�paleosuelo�y�el�relleno�revela�que�en�once�de�
los�catorce�especímenes�estudiados,�con�la�excepción�de�los�ejemplares�MPEF�IC�

� ����
721,� 724,� y� 731,� la� frecuencia� absoluta� de� fitolitos� del� paleosuelo� no� difiere�
significativamente� (�� >� 0,05)� de� la� hallada� en� el� relleno� de� los� ejemplares� de�
�����������.� Sólo� tres� especímenes�mostraron� un� número� de� fitolitos� en� el�
relleno�menor�al�del�paleosuelo.�Particularmente,�en�el�ejemplar�MPEF�IC�721,�el�
paleosuelo�posee�17,5�%�más�fitolitos�que�el�relleno;�mientras�que�en�los�otros�
dos�ejemplares�(MPEF�IC�724�y�731),� la�diferencia�en� la�frecuencia�absoluta�de�
fitolitos� entre� estas� dos�muestras,� es�menor,�mostrando� 5,1�%� y� 6,2�%�más�
cantidad�de�fitolitos�en�el�paleosuelo�que�en�el�relleno,�respectivamente.�Por�otro�
lado,�el�análisis�de�las�dos�fracciones�(FM�y�FF)�por�separado,�revela�resultados�
similares�en�el�número�total�de�fitolitos�(frecuencia�absoluta)�entre�las�muestras�
de�paleosuelo�y�relleno.�La�Figura�38�B�(FF)�muestra�que�en�trece�de�los�catorce�
ejemplares� estudiados,� con� la� excepción� del� especímen� MPEF�IC� 721,� la�
frecuencia� absoluta� de� fitolitos� hallada� en� el� paleosuelo� no� difiere�
significativamente�(��>�0,05)�de� la�hallada�en�el�relleno.�La�Figura�38�C�revela�
que� analizando� la� fracción� media� (FM),� sólo� en� cuatro� especímenes� (MPEF�IC�
721,� 724,� 726,� y� 731)� la� frecuencia� absoluta� de� fitolitos� encontrada� en� el�
paleosuelo�difiere�significativamente�(��<�0,05)�de�la�encontrada�en�el�relleno,�y�
que� en� los� diez� restantes� las� diferencias� entre� estas� dos� muestras� no� son�
significativas.��
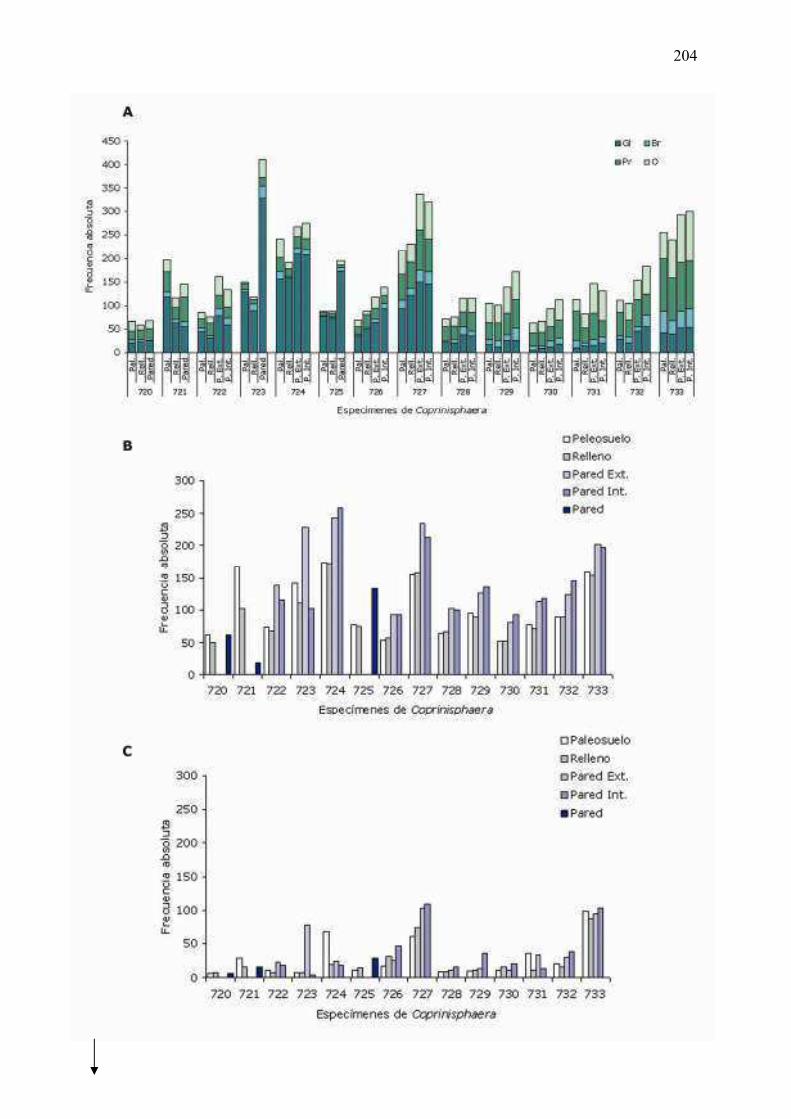
� ����
�
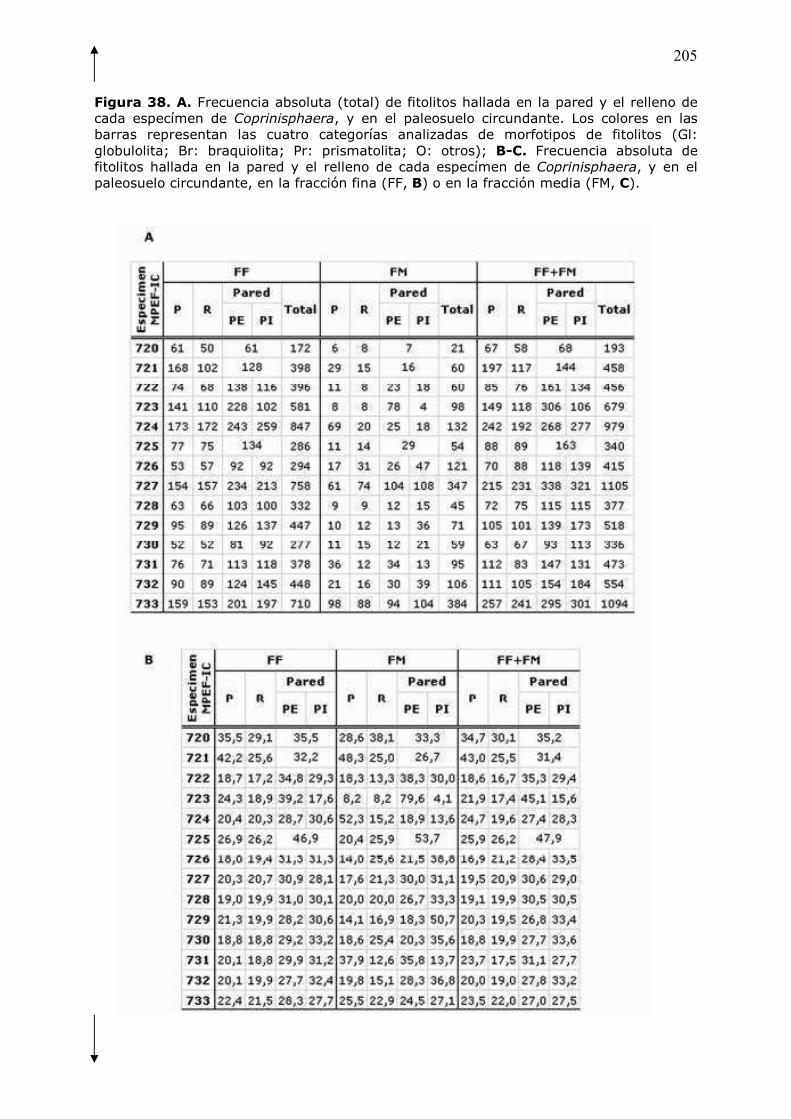
� ����
�!�%�&5:���Frecuencia�absoluta�(total)�de�fitolitos�hallada�en�la�pared�y�el�relleno�de�cada� especímen� de������������,� y� en� el� paleosuelo� circundante.� Los� colores� en� las�barras� representan� las� cuatro� categorías� analizadas� de� morfotipos� de� fitolitos� (Gl:�globulolita;� Br:� braquiolita;� Pr:� prismatolita;� O:� otros);� �#��� Frecuencia� absoluta� de�fitolitos�hallada�en� la�pared�y� el� relleno�de� cada�especímen�de������������,� y� en�el�paleosuelo�circundante,�en�la�fracción�fina�(FF,��)�o�en�la�fracción�media�(FM,��).��
�

� ����
&)'&8���Frecuencia�absoluta�de�fitolitos�hallada�en�el�relleno�(R)�y�en�las�dos�partes�de� la� pared� (PE:� externa,� PI:� interna)� de� cada� ejemplar� de� �����������,� y� en� el�paleosuelo� (P)� � circundante.� FF:� fracción� fina.� FM:� fracción� media. ��� Los� mismos�parámetros�expresados�en�porcentaje.��
5.4.2.2� �� Comparación� del� número� total� de� fitolitos� entre� la� pared� de�
������������y�el�paleosuelo�circundante�(o�el�relleno)�
� �
La�Figura�38�A�(y�Tabla�4)�indica�también�que�en�todos�los�especímenes�de�
������������estudiados,�con� la�excepción�de� los�dos�primeros,�MPEF�IC�720,�
721,�y�de�los�especímenes�MPEF�IC�724�y�731,�la�frecuencia�absoluta�de�fitolitos�
encontrada�en� la�pared�(PE�y�PI)�es�significativamente�mayor�(46�a�66�%�más�
cantidad�de� fitolitos,���<�0,05)�que� la�hallada�en� las�muestras�de�paleosuelo�y�
relleno.� En� el� ejemplar� MPEF�IC� 720,� la� frecuencia� absoluta� de� fitolitos�
encontrada�en� las�muestras�de�paleosuelo� (67),� relleno� (58),� y�pared� (68),�no�
difieren� significativamente�entre� sí� (��>�0,05).�En�el� ejemplar�MPEF�IC�721,� la�
frecuencia�absoluta�de�fitolitos�de� la�pared�(144)�y�del�relleno�(117)�no�poseen�
diferencias�significativas�(��>�0,05)�entre�sí.�En�este�ejemplar,�las�dos�muestras�
mencionadas�difieren�significativamente�de�la�del�paleosuelo�(��<�0,05),�el�cual�
posee� 197� fitolitos.� En� los� otros� dos� casos� excepcionales,� ejemplares� MPEF�IC�
724�y�731,�la�frecuencia�absoluta�de�fitolitos�encontrada�en�las�dos�partes�de�la�
pared� (724:� PE� =� 268� /� PI� =� 277;� 731:� PE� =� 147� /� PI� =� 131)� no� difieren�
significativamente� (�� >� 0,05)� de� las� del� paleosuelo� (724:� P� =� 242;� 731:� P� =�
112),� mientras� que� las� frecuencias� halladas� en� este� último� se� diferencian�
significativamente�(��<�0,05)�de�las�del�relleno�(724:�R�=�192;�731:�R�=�83).��
Los�resultados�obenidos�para�FF�+�FM�que�se�presentan�en�la�Figura�38�A�
se� encuentran� apoyados� parcialmente� por� los� resultados� obtenidos� del� análisis�
separado�de�las�dos�fracciones.�En�la�mayoría�de�los�ejemplares,�la�pared�posee�

� ����
más�cantidad�de�fitolitos�que�las�muestras�de�paleosuelo�y�relleno.�La�Figura�38�
B�(FF)�revela�que�la�frecuencia�absoluta�de�fitolitos�encontrada�en�la�pared�de�la�
mayoría� de� los� ejemplares� estudiados,� con� la� excepción� de� los� dos� primeros�
(MPEF�IC�720,�721),� es�significativamente�mayor� (��<�0,05)�que� la�hallada�en�
las� muestras� de� paleosuelo� y� relleno.� Los� ejemplares� MPEF�IC� 720� y� 721�
muestran� resultados� similares� a� los� hallados� en�el�análisis� conjunto�de� las� dos�
fracciones�(FF�+�FM).�Sin�embargo,�como�se�ve�en�la�Figura�38�C,�la�FM�exhibe�
que�sólo�en�tres�ejemplares�(MPEF�IC�723,�725�y�727)�las�frecuencia�absoluta�de�
fitolitos�de� la�pared�es�significativamente�mayor�(��<�0,05)�que� las�frecuencias�
encontradas�en�las�muestras�de�relleno�y�paleosuelo.���
�
5.4.2.3���Comparación�del�número�total�de�fitolitos�entre�las�dos�partes�de�la�
pared�(interna�y�externa)�de�������������
� �
� La�Figura�39�muestra�el�número�total�de�fitolitos�(frecuencia�absoluta)�de�
FF+FM�hallado�en�las�muestras�de�la�pared�externa�(PE)�e�interna�(PI).�En�esta�
figura� solo� se� representaron� los� valores� correspondientes� a� los� ejemplares� de�
������������cuya�pared�pudo�ser�dividida�en�dos�secciones.�En�la�Tabla�4,�seis�
ejemplares�(MPEF�IC�724,�726,�729,�730,�732�y�733)�muestran�que�la�frecuencia�
absoluta�de�fitolitos�de�la�pared�interna�es�mayor�que�la�de�la�pared�externa.�Sin�
embargo,�el�análisis�estadístico�revela�que�estas�diferencias�no�son�significativas�
(��>�0,05).���
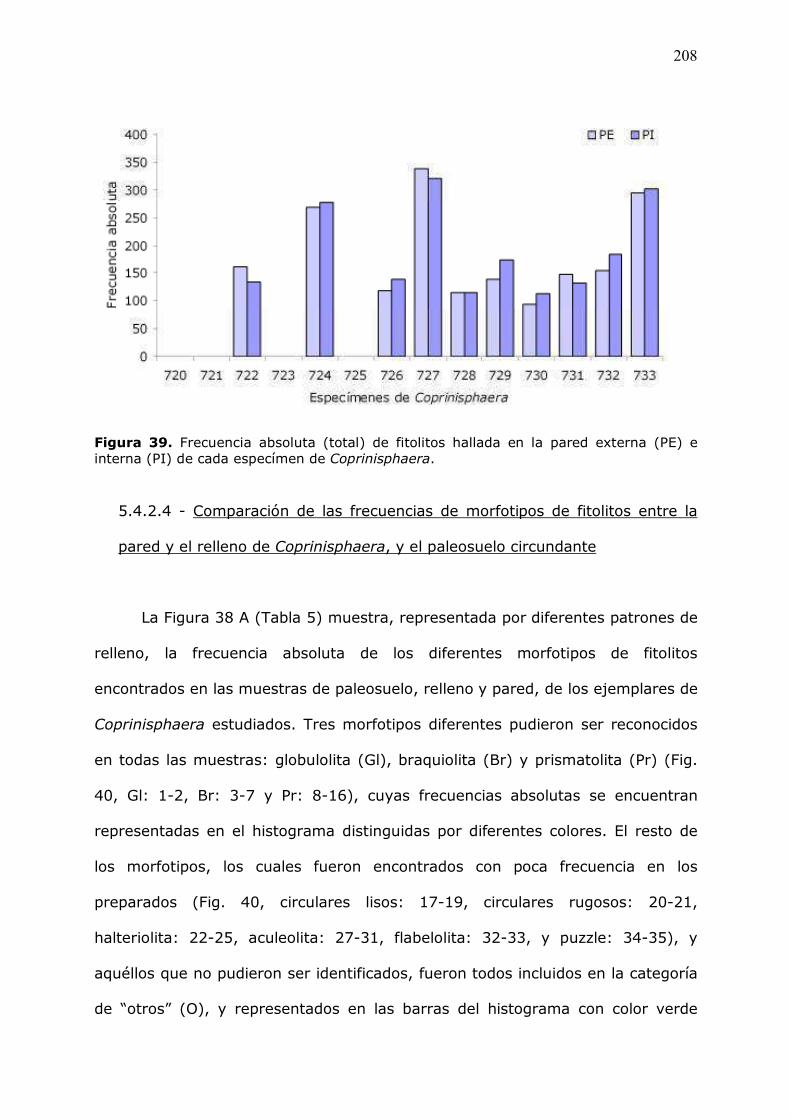
� ����
�
�!�%�& 57�� Frecuencia� absoluta� (total)� de� fitolitos� hallada� en� la� pared� externa� (PE)� e�interna�(PI)�de�cada�especímen�de������������.���
�
5.4.2.4� ��Comparación�de� las� frecuencias� de�morfotipos� de� fitolitos� entre� la�
pared�y�el�relleno�de������������,�y�el�paleosuelo�circundante�
� La�Figura�38�A�(Tabla�5)�muestra,�representada�por�diferentes�patrones�de�
relleno,� la� frecuencia� absoluta� de� los� diferentes� morfotipos� de� fitolitos�
encontrados�en�las�muestras�de�paleosuelo,�relleno�y�pared,�de�los�ejemplares�de�
������������ estudiados.�Tres�morfotipos�diferentes�pudieron� ser� reconocidos�
en�todas�las�muestras:�globulolita�(Gl),�braquiolita�(Br)�y�prismatolita�(Pr)�(Fig.�
40,� Gl:� 1�2,� Br:� 3�7� y� Pr:� 8�16),� cuyas� frecuencias� absolutas� se� encuentran�
representadas�en�el� histograma�distinguidas�por�diferentes� colores.�El� resto�de�
los� morfotipos,� los� cuales� fueron� encontrados� con� poca� frecuencia� en� los�
preparados� (Fig.� 40,� circulares� lisos:� 17�19,� circulares� rugosos:� 20�21,�
halteriolita:� 22�25,� aculeolita:� 27�31,� flabelolita:� 32�33,� y� puzzle:� 34�35),� y�
aquéllos�que�no�pudieron�ser�identificados,�fueron�todos�incluidos�en�la�categoría�
de� “otros”� (O),� y� representados� en� las� barras� del� histograma� con� color� verde�

� ���
claro.� El� análisis� estadístico� revela� que� sólo� uno� de� los� catorce� ejemplares� de�
������������estudiados�(MPEF�IC�723),�muestra�diferencias�significativas�(��<�
0,05)� entre� las� tres� muestras,� en� las� frecuencias� absolutas� de� las� cuatro�
categorías� de� fitolitos.� En� el� ejemplar� MPEF�IC� 721� no� hay� diferencias�
significativas� (�� >� 0,05)� en� la� frecuencia� absoluta� de� las� cuatro� categorías� de�
fitolitos�entre�las�muestras�de�paleosuelo�y�relleno.�Sin�embargo,�las�frecuencias�
de� los�morfotipos�de�estas�dos�muestras�difieren�significativamente� (��<�0,05)�
de�las�de�la�pared.�Los�ejemplares�MPEF�IC�724,�731�y�733�muestran�que�no�hay�
diferencias� significativas� (�� >� 0,05)� en� las� frecuencias� absolutas� de� las� cuatro�
categorías�de�fitolitos,�entre�el�relleno�y�las�dos�secciones�de�la�pared�(PE�y�PI),�
pero� todos� ellos�mostraron� diferencias� con� las� del� paleosuelo.� Contrariamente,�
los�ejemplares�MACN�IC�726�y�732�revelan�que�no�hay�diferencias�significativas�
(��>�0,05)�en�las�frecuencias�absolutas�de�las�cuatro�categorías�de�fitolitos�entre�
el� paleosuelo,� relleno�y�PE,�pero�que� todas�ellas�difieren�de� las�de� la�PI.�En�el�
resto�de�los�ejemplares�(MPEF�IC�720,�722,�725,�727�730),�estas�frecuencias�no�
mostraron� diferencias� (�� >� 0,05)� entre� las� muestras� analizadas� (paleosuelo,�
relleno�y�las�dos�secciones�de�la�pared,�o�la�pared�total),�sugiriendo�que�todos�los�
morfotipos� de� fitolitos� se� encuentran� igualmente� representados� en� todas� las�
muestras�(morfotipos�de�fitolitos�uniformemente�distribuídos).�

� ���
�!�%�& 8<�� Morfotipos� de� fitolitos.� 2#4�� Globulolita� (Gl);� 5#;�� Braquiolita� (Br);� :#23��Prismatolita� (Pr);� 2;#27�� Circular� liso� (Cl); 4<#42�� Circular� rugoso� (Cr);� 44#46��Halteriolita�(Ha);4;#52��Aculeolita�(Ac);�54#55��Flabelolita�(Fl);�58#56��Puzzle�(Pu).��

� ��
�
� &)'&6���Frecuencia�absoluta�(total)�de�los�distintos�morfotipos�de�fitolitos�hallada�en�el�relleno�y�en�las�dos�partes�de�la�pared�de�cada�ejemplar�de������������,�y�en�el�paleosuelo�circundante.���Los�mismos�parámetros�expresados�en�porcentaje.�Gl:�globulolita;�Cl:�circular�liso;�Cr:�circular�rugoso;�Br:�braquiolita;�Pr:�prismatolita;�Ha:�halteriolita;�Ac:�aculeolita:�Fl:�flabelolita;�Pu:�puzzle;�O:�otros.��
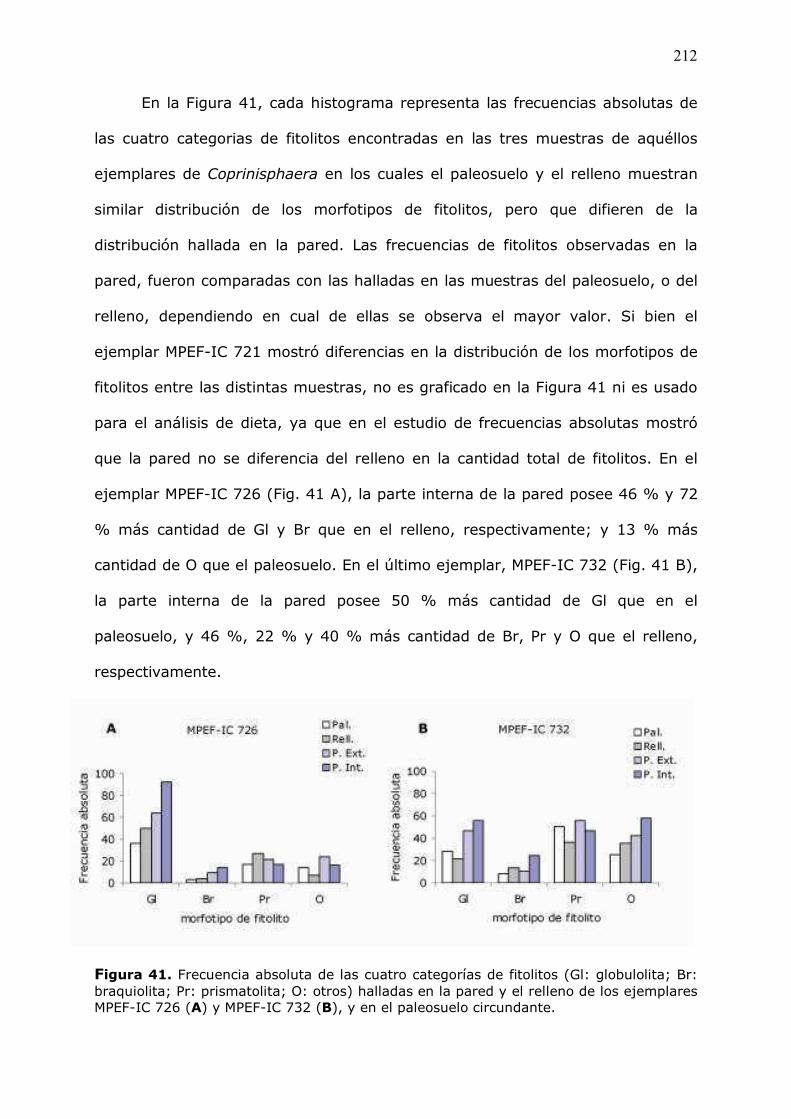
� ���
En� la�Figura�41,�cada�histograma�representa� las�frecuencias�absolutas�de�
las� cuatro� categorias� de� fitolitos� encontradas� en� las� tres�muestras� de� aquéllos�
ejemplares�de�������������en� los�cuales�el�paleosuelo�y�el� relleno�muestran�
similar� distribución� de� los� morfotipos� de� fitolitos,� pero� que� difieren� de� la�
distribución� hallada� en� la� pared.� Las� frecuencias� de� fitolitos� observadas� en� la�
pared,�fueron�comparadas�con�las�halladas�en�las�muestras�del�paleosuelo,�o�del�
relleno,� dependiendo� en� cual� de� ellas� se� observa� el� mayor� valor.� Si� bien� el�
ejemplar�MPEF�IC�721�mostró�diferencias�en�la�distribución�de�los�morfotipos�de�
fitolitos�entre�las�distintas�muestras,�no�es�graficado�en�la�Figura�41�ni�es�usado�
para�el�análisis�de�dieta,�ya�que�en�el�estudio�de�frecuencias�absolutas�mostró�
que� la�pared�no�se�diferencia�del� relleno�en� la�cantidad� total�de� fitolitos.�En�el�
ejemplar�MPEF�IC�726�(Fig.�41�A),�la�parte�interna�de�la�pared�posee�46�%�y�72�
%�más� cantidad� de� Gl� y� Br� que� en� el� relleno,� respectivamente;� y� 13�%�más�
cantidad�de�O�que�el�paleosuelo.�En�el�último�ejemplar,�MPEF�IC�732�(Fig.�41�B),�
la� parte� interna� de� la� pared� posee� 50� %� más� cantidad� de� Gl� que� en� el�
paleosuelo,� y�46�%,�22�%�y�40�%�más�cantidad�de�Br,� Pr�y�O�que�el� relleno,�
respectivamente.�
�
�!�%�&82��Frecuencia�absoluta�de�las�cuatro�categorías�de�fitolitos�(Gl:�globulolita;�Br:�braquiolita;�Pr:�prismatolita;�O:�otros)�halladas�en�la�pared�y�el�relleno�de�los�ejemplares�MPEF�IC�726�()�y�MPEF�IC�732�(�),�y�en�el�paleosuelo�circundante.�

� ���
3����������
3�2 # ���� J����� �� ����������� � ������ �� � �������� �� �
����������
�
� En� el� 2006,� Laza� publicó� por� primera� vez� una� revisión� exhaustiva� del�
icnogénero�������������Sauer�1955,�para�la�que�utilizó�icnotaxobases�nuevas,�
y� bien� definidas,� de� identificación� de� las� distintas� icnoespecies.� Sin� embargo,�
revisiones�posteriores�de�una�mayor�cantidad�de�ejemplares�para�esta�Tesis,�y�
del� material� típico� usado� por� Roselli� (1939,� 1987)� en� la� creación� de� varios�
icnogéneros� e� icnoespecies,� sugirieron� la� necesidad� de� realizar� algunas�
modificaciones� a� dicha� icnotaxonomía.� En� primer� lugar,� la� nueva� revisión� del�
material� de� Roselli,� depositado� en� el� museo� en� Nueva� Palmira� (Uruguay)� que�
lleva� su� nombre,� reveló� que������������� ���������� Sauer� 1955,� era� un�
sinónimo� junior� subjetivo� de� ������������ �������� (Roselli,� 1939).� Roselli�
tenía� por� costumbre� dar� el� mismo� número� de� identificación� a� una� serie� de�
ejemplares,�mientras� que� en�el� texto� se� refería� a� holotipo� en� singular� como� si�
fueran� uno� solo.� Laza� (2006)� reconoció� como� holotipo� de������������ (Roselli�
1939)�al�ejemplar�que�Roselli�(1987)�ilustró�como�tal�en�su�trabajo�posterior,�y�
que� coincide� con� el� que� menciona� como� genotipo� en� su� publiación� de� 1939,�
criterio�que�también�se�sigue�aquí.�Este�ejemplar,�así�como�otros�de�esa�serie,�
poseía� el� agujero� de� emergencia� típico� de� tamaño�medio� que� caracteriza� a����
���������� Sauer,� descripta� posteriormente� (1955).� Posiblemente,� la�
presencia�de�un�segundo�agujero�más�pequeño�en�uno�de�los�ejemplares�típicos�
fue�confundido�por�Laza�(2006)�con�aquel�que�define�a���� isp.�A����,�como�fue�
explicado� en� detalle� en� la� sección�6�2� de� esta� Tesis.� Además� de� este� cambio�

� ���
nomenclatorial,� una� modificación� icnotaxonómica� importante� es� el�
reconocimiento�de�una�nueva�bola�de�cría� fósil�a�partir�de� la�separación�de�dos�
grupos�de�ejemplares�que�habían�sido�originalmente�agrupados�en�����������por�
Laza� (2006).� La� � nueva� icnoespecie,� llamada�aqu��� isp.�B�����,� se� diferencia�
fundamentalmente� de� ��� ������,� en� que� en� la� primera,� la� cámara� ovígera� se�
encuentra�incluida�en�una�protuberancia�de�la�pared�de�la�bola�de�cría,�en�lugar�
de�en�una�cámara�esférica�más�pequeña,�como�se�distingue�en� la�segunda.�La�
ubicación�de�la�cámara�ovígera,� icnotaxobase�fundamental�para�la� identificación�
de� icnoespecies� seguida� por� Laza� (2006)� y� aquí� para� separar� la� nueva�
icnoespecie,� proporciona� a� su� vez� una� hipótesis� a� testear� sobre� la� posible�
evolución�de� la�ubicación�de�esta� cámara� en� relación� con� la�de�provisiones.����
�������� se� caracteriza,� fundamentalmente,� por� la� presencia� de�un�agujero�de�
tamaño� medio� excavado� en� la� pared� y� por� la� ausencia� de� evidencia� de� una�
cámara� ovígera.� Es� por� eso� que� esta� icnoespecie�podría� ser� el� resultado� de� la�
emergencia� de� un� escarabajo� adulto� de� una� bola� de� cría� esférica,� subesférica,�
piriforme�o�con�protuberancia�(���isp.�A,�����������y����������)�las�cuales�habrían�
tenido�su�cámara�ovígera�excavada�en�las�provisiones�(como�pudo�observarse�en�
bolas� de� cría� producidas� por� ��� ������� spp.,� ��� ��� ���� y� ��������
spp.),�o�incluída�en�la�pared,�y�borrada�por�la�emergencia�del�adulto.�El�holotipo�
de����������� ���posee�un�agujero�de�similares�caracteristíscas�excavado�en�la�
pared,� y� carece�de�evidencias� de� la� cámara�ovígera,� pero�dicha�perforación� se�
abre�en�el�centro�de�una�superficie�plana,�y�por� lo�tanto�aún�el�ejemplar�sin�el�
agujero� de� emergencia� no� podría� atribuirse� a� las� icnoespecies� anteriormente�
mencionadas.� Resulta� revelador� que� algunos� ejemplares,� atribuídos� aquí� a� ���
������� ���por�poseer� la�zona�plana�en�un�polo,�provenientes�de� la�Formación�
Sarmiento�en�Gran�Barranca,�muestran�en� la�pared,�en� lugar�de�un�agujero�de�

� ���
tamaño�medio,�una�pequeña�perforación�en�la�base�de�un�embudo,�que�estaría�
indicando�la�no�emergencia�del�adulto�y�la�presencia�de�la�cámara�ovígera�en�la�
provisión.� Roselli� (1939)� en� cambio,� creía� que� tal� zona� plana� era� una� cámara�
ovígera� inconclusa,� como� lo� demuestra� su� comentario� sobre� uno� de� los�
ejemplares�originales�de������������considerados�aqu���������� ��.�Este�tipo�
de� bolas� de� cría� estarían� sugiriendo� tal� vez� la� condición� más� primitiva� en� la�
posición�de�la�cámara�ovígera,�al�ser�excavada�directamente�en�la�provisión.�En�
relación�con�ello,�Halffter�y�Edmonds�(1982)�mencionan�que�en�la�mayoría�de�las�
especies� las� hembras� agregan� secreciones� a� la� cámara� ovígera� durante� su�
construcción,�que� tendrían�propiedades�antibióticas� y�que�actuarían� reduciendo�
las� infecciones� en� el� interior� de� dicha� cámara.� Estas� secreciones� podrían� estar�
relacionadas�con�los�tapices�muy�delgados�que�alisan�la�superficie�interna�de�las�
cámaras�ovígeras�de�muchas�de� las�bolas�de�cría�actuales�estudiadas�aquí.�Por�
otro� lado,� debido� al� alto� riesgo� que� corre� el� huevo� de� ser� atacado� por�
microorganismos,� hongos,� etc.,� es� factible� relacionar� este� hecho� con� la�
construcción� de� una� cámara� ovígera� cada� vez�más� aislada� de� las� provisiones.�
Siguiendo�con�este�razonamiento,� las� icnoespecies�restantes�preservan�restos�o�
toda� la� cámara� ovígera,� en� distintas� posiciones� en� relación� con� la� pared� y� la�
bosta.�����������preservaría�en�la�pared�el�techo�de�la�cámara�ovígera,�sugerido�
por� la� presencia� de� una� cavidad� hemiesférica� y� pequeña� (cóncava� hacia� el�
interior�de�la�bola�de�cría),�excavada�en�el�piso�de�la�protuberancia�y�abierta�en�
su�base�hacia� el� interior,� en� contacto� directo� con� las� provisiones.� Este� sería� el�
primer� indicio� de� una� tendencia� a� aislar� el� huevo� de� las� provisiones,�
construyendo�una�cámara�ovígera�más�retirada�de�la�cámara�principal�de�la�bola�
de�cría,�donde�se�ecuentra�la�provisión.����isp.�A�posee�en�la�parte�externa�de�la�
pared� una� cavidad� o� cámara� secundaria,� hemiesférica� y� pequeña,� con� la�

� ���
concavidad�abierta�al�exterior�y�comunicada�con� la�cámara�principal�por�medio�
de�una�perforación�pequeña,� la� cual� fue� interpretada�por� Laza� (2006)� como�el�
piso�de�la�cámara�ovígera.�Siguiendo�esta�interpretación,�esta�icnoespecie�estaría�
mostrando� un� mayor� aislamiento� de� la� cámara� ovígera� de� las� provisiones,�
reflejado�por�la�cámara�hemiesférica�recién�mencionada.�Sin�embargo,�no�se�han�
encontrado�ejemplos�actuales�hasta�el�momento�que�sustenten�esta�idea�y�sí,�en�
cambio,�sería�posible�relacionar�esta�concavidad�con�el�tapón�de�fibras�vegetales�
presente� en� bolas� de� cría� de�muchas� de� las� especies� actuales� estudiadas� (por�
ejemplo,�las�incluidas�en���� ������),�dejando�de�lado�la�idea�de�una�cámara�
ovígera� incluida�en� la�pared.�En�el�caso�de���� ������,� la�cámara�ovígera�estaría�
completamente� incluida� en� su� pared� y� aislada� de� la� provisión,� sugerido� por� la�
presencia�de�una�cámara�esférica�pequeña�presente�en�la�pared�de�la�bola�en�la�
zona� del�mamelón.� Esta� icnoespecie,� junto� con���� isp.� B,� caracterizada� por� la�
presencia� de� una� segunda� cámara� esférica�más� pequeña� (la� cámara� ovígera),�
comunicada�con�la�cámara�principal�por�medio�de�una�perforación,�serían�los�dos�
casos�más�extremos�de�aislamiento�de�la�cámara�ovígera�de�las�provisiones.�Este�
tipo� de� bolas� de� cría� son� comparables� con� aquéllas� construidas� por� algunas�
especies�de�������,�como�ha�sido�descripto�en�otra�sección�de�esta�Tesis,�en�
las�que�la�presencia�de�una�segunda�cámara,�donde�es�depositado�el�huevo,�es�la�
característica�más�distintiva�de�estas�bolas�de�cría.�Además,�se�ha�registrado�en�
������� &������ &�����,� que� la� emergencia� del� nuevo� adulto� no� borra� las�
evidencias�de�la�cámara�ovígera,�ya�que�estas�bolas�son�depositadas�en�el�nido�
con� el� eje� principal� (el� que� pasa� por� las� dos� cámaras)� paralelo� al� suelo,� y� el�
nuevo� adulto� emerge� generalmente� hacia� arriba� perforando� directamente� la�
pared� de� la� cámara� principal� (Favila,� 1993).� Es� por� ello,� que� las� bolas� de� cría�

� ���
fósiles� como���� isp.� B,� suelen� preservar� la� cámara� ovígera� intacta,�mostrando�
evidencias�del�mayor�aislamiento�del�huevo�en�bolas�de�cría.��
� La� validez� de� esta� hipótesis� sobre� la� evolución� de� este� carácter�
proporcionada�por�las�trazas�fósiles�solo�podrá�ser�testeada�en�la�medida�que�se�
encuentren�trazas�con�caracteres�similares�en�especies�actuales�y�se�conozca�su�
posición�en�la�filogenia�del�grupo.������
�
3�4 # �� ����� ��������������, ������ ���� � �� ����������� ��
��������������������������������� ��
�
Judulien� (1899),� Fabre� (1899),� Richter� (1918),� Halffter� y� Matthews�
(1966),�Halffter�(1976),�Halffter�(1977),�Halffter�y�Edmonds�(1982),�Halffter���
���(1983),�Cabrera�Walsh�y�Gandolfo�(1996),�Favila�y�Díaz�(1996),�Morelli������
(1996),�Barbero������(1999),�y�Favila�(2001),�entre�otros,�han�publicado�sobre�
varios�aspectos�del�comportamiento�de�nidificación�y/o�de�la�arquitectura�de�los�
nidos� y� de� la� estructura� de� las�masas/bolas� de� cría� de� las� diferentes� especies�
estudiadas� para� este� trabajo� de� investigación.� Sin� embargo,� algunos� de� estos�
artículos� se� contradicen� mientras� que� otros� son� fragmentarios� o� bien� se�
complementan,� como� sucede� con� los� nuevos� datos� presentados� aquí.� No�
obstante,� ni� en� estos� trabajos,� ni� en� otros� sobre� Scarabaeinae,� se� utilizaron�
cortes� micromorfológicos� de� las� masas� y� bolas� de� cría,� los� cuales� aportaron�
nuevos� caracteres� y� una� forma� distinta� de� estudiar� la� morfología� de� estas�
estructuras.� Particularmente,� la� micromorfología� permitió� inferir� rasgos�
comportamentales� de� difícil� observación� que� fueron� rastreados� posteriormente�
en�las�bolas�de�cría�fósiles,�arrojando�datos�sobre�el�origen�y�evolución�de�dichos�
comportamientos.���

� ���
3�4�2H�����������"B�
��
� Los�nidos�de������������han�sido� registrados�para�un�gran�número�de�
especies,� todas� ellas� incluidas� en� el� patrón� I� de� nidificación.� De� acuerdo� a� la�
cantidad�de�masas�de�cría�construidas,�estos�nidos�pueden�ser�diferenciados�en�
simples�o�compuestos�(Halffter�y�Edmonds,�1982).�En�el�primer�tipo,�la�hembra�
excava� un� túnel� y� en� el� fondo� del� mismo� deposita� una� masa� cilíndrica� de�
provisión� (bosta)� a� la� que� le� construye,� en� el� extremo� superior,� una� cámara�
ovígera�esférica.�En�el�interior�de�dicha�cámara,�el�huevo�es�depositado�de�forma�
vertical,�adherido�al�piso�e�inmediatamente�después,�la�cámara�es�cerrada.�Son�
un�ejemplo�de�este�tipo,�las�masas�de�cría�de������� �����descriptas�por�Cabrera�
Walsh�y�Gandolfo�(1996).�El�segundo�tipo�de�nido,�el�cual�suele�ser�ramificado,�
difiere�del�anterior,�principalmente,�en�que�cada�túnel�del�nido�posee�una�serie�
de� masas� de� cría,� las� cuales� además� de� ser� claramente� ovaladas,� son� más�
elaboradas�que�las�construidas�en�los�nidos�simples�(Halffter�y�Edmonds,�1982).���
� De�acuerdo�a�lo�observado�en�el�campo,�las�masas�de�cría�de������������
sp.�colectadas�en�Formosa�corresponden�a�nidos�simples,�en�dos�casos�donde�se�
encontró�solo�una�masa�de�cría�por�nido,�y�a�nidos�compuestos,�en�el�caso�donde�
se�encontraron�tres�masas�de�cría�en�un�túnel�ramificado.�A�pesar�de�pertenecer�
a�estos�dos�distintos�tipos�de�nidos,�y�a�diferencia�de�lo�sugerido�por�Halffter�y�
Edmonds� (1982),� las� masas� de� cría� mostraron� ser� muy� similares� entre� sí,�
probablemente� porque� los� nidos� con� una� sola� cámara� fueran� incompletos.� La�
cámara�ovígera�de�todas�ellas�es�ovalada,�se�encuentra�completamente�cerrada,�
y�la�micromorfología�revela�que�posee�una�pared�construida�con�bosta,�como�los�
meniscos�del�relleno,�y�que�a�diferencia�de�éstos,�la�bosta�se�encuentra�mucho�

� ��
más�empaquetada�y�oscurecida.�Si�bien�estas�estructuras�no�poseen�una�pared�
definida,� se� observó� en� la� micromorfología,� un� cierto� grado� de� arreglo� del�
material�del�suelo� limitante,� indicando�que�el�escarabajo�adulto�reubicaría�dicho�
material� durante� la� excavación� del� extremo� más� distal� del� túnel,� para� la�
construcción�de�la�masa�de�cría.�Dadas�estas�características,�y�sumadas�a�las�ya�
descriptas� por� otros� autores,� no� se� duda� la� ubicación� de� los� �����������
estudiados,�en�el�patrón�tipo�I�de�nidificación.�Sin�embargo,�si�bien�este�patrón�
resulta�en�ausencia�de�bolas�de�cría,�esto�se�trataría�de�una�perdida�secundaria,�
ya�que�según� la� filogenia�más�reciente�(Philips�����,�2004b),� los�Onthophagini�
conformarían�uno�de�los�linajes�derivados�más�exitosos,�aún�presentando�uno�de�
los� tipos� más� simples� de� comportamiento� de� nidificación.� No� hay� hasta� el�
momento�registros�de�trazas�fósiles�comparables�con�este�tipo�de�masas�de�cría�
que�puedan�aportar�más�datos�al�orígen�y�evolución�de�su�comportamiento.��
�
3�4�4H&�������� ����'�����
�
El� comportamiento� de� nidificación� y� la� estructura� de� los� nidos� de� ���
� �������han�sido�estudiados�y�registrados�de�manera�incompleta�hasta�el�día�
de� hoy.� Judulien� (en� Fabre,� 1899)� fue� el� primero� en� describir� los� nidos� de����
� ������� destacando�de� ellos� la� presencia� de�una� cámara�ovígera�única:� una�
celda�esférica�recubierta�por�material�del�suelo,�ubicada�en�el�extremo�superior�
de� cada� masa� de� cría.� También� mencionó� que� los� nidos� eran� ramificados� y�
conformados�por�varias�masas�de�cría�cilíndricas.�Tiempo�más� tarde,�Halffter�y�
Matthews� (1966)� los� describieron� como� un� solo� túnel� en� el� cual� se� encuentra�
alojada� una� sola� masa� de� cría,� y� los� consideraron� nidos� con� características�
intermedias�a�las�incluidas�en�los�patrones�I�y�II�de�nidificación.�Por�ejemplo,�son�

� ����
masas�de�cría�como�las�vistas�en�el�patrón�I,�pero�el�huevo�se�encuentra�aislado�
en�una�cámara�de�material�del�suelo,�como�se�ve�en�los�nidos�del�patrón�II.�Sin�
embargo,�en�nuevas�revisiones,�Halffter�y�Edmonds�(1982)�consideraron�que�los�
nidos�de����� �������eran�en�realidad�mucho�más�similares�a�los�incluidos�en�el�
patrón�I,�debido�fundamentalmente�a�su�poca�complejidad.�Finalmente,�Cabrera�
Walsh�y�Gandolfo�(1996),�sin�arriesgar�en�que�patrón�deberían�ser�incluidos,�por�
medio� de� sus� observaciones� consideraron� que� estos� nidos� difieren�
significativamente�de� los�otros� incluidos�en�dicho�patrón�y�que� la�presencia� de�
una�cámara�ovígera�aislada�de�la�provisión�por�una�pared�de�material�del�suelo�
es�una�característica�que�comparten�con����� ��������,�*���������������,�y�
los�Phanaeini.��
� La�información�presentada�aquí,�muestra�por�primera�vez�que�los�nidos�de�
��� � ������� poseen� túneles� de� almacenamiento� donde� la� bosta� es�
aprovisionada�antes�de�ser�usada�para� la�construcción�de� la�masa�de�cría.�Este�
tipo�de�aprovisionamiento� (demorado),�no�es�un� comportamiento� característico�
de� las� especies� incluidas� en� el� patrón� I� de� nidificación,� donde� dicho�
comportamiento�suele�ser�del�tipo�directo�(Halffter�y�Edmonds,�1982).�Sumado�a�
ésto,� se� han� observado� dos� nidos� conformados� por� una� masa� de� cría� ya�
terminada,�en� los�cuales� los� túneles�de�almacenamiento�conservaban�restos�de�
bosta,�e�incluso�a�un�adulto�sobre�ella,�sugiriendo�la�posible�construcción�de�una�
segunda�masa�de� cría� (c.f.,� Fabre,� 1899;� Judulien,�1899;�Halffter� y�Matthews,�
1966;� Halffter� y� Edmonds,� 1982).� Por� otro� lado,� en� ninguno� de� los� estudios�
anteriormente�publicados,�se�ha�mencionado�la�presencia�de�una�pared�discreta�
cubriendo� las�masas�de�cría.�En�su�trabajo�de�1996,�Cabrera�Walsh�y�Gandolfo�
describen�las�masas�de�cría�de����� �������como�una�excavación�al�final�de�un�
túnel,�y�no�hacen�referencia�a�ninguna�característica�de�la�pared.�Sin�embargo,�

� ���
se�observó�que,�al�secarse�en�el� laboratorio,� las�muestras�de� las�masas�de�cría�
tomadas�en�el�campo�en�una�porción�de�suelo�circundante,�se�aislaron�del�suelo�
dejando� una� fisura� entre� ellos,� y� revelaron� la� presencia� de� una� pared� discreta�
cubriendo� a� cada� masa� de� cría� (Fig.� 12� G).� Los� datos� micromorfológicos�
presentados� en� este� trabajo� confirman� ésto,� demostrando� la� presencia� de� una�
pared�discreta�cuyo�límite�puede�ser�distinguido�del�suelo�por�la�presencia�de�una�
capa�muy�delgada�de�alto�contenido�de�materia�orgánica�(Fig.�13�B).�De�acuerdo�
a� lo� revelado�por� la�micromorfología,�esta�pared�es�construida�con�material�del�
suelo�y�con�el�agregado�de�materia�orgánica�fina,�y�sin�el�uso�de�fibras�vegetales�
de�la�bosta�(Fig.�13�B).��
Por� otro� lado,� según� Judulien� (1899),� Fabre� (1899),�Halffter� y�Matthews�
(1966),� Cabrera�Walsh� y� Gandolfo� (1996),� las�masas�de� cría� de���� � �������
poseen�la�cámara�ovígera�aislada�de�la�provisión�por�una�pared.�Algunos�de�estos�
autores�consideraron�que�esta�pared�era�construida�con�“arcilla”,�pero�como�sus�
observaciones�fueron�hechas�a�ojo�desnudo�y�sin�el�uso�de�la�micromorfología,�es�
probable�que�estos�autores�hayan�querido�hacer�referencia�solo�a�la�presencia�de�
material� fino.� Cabrera�Walsh� y� Gandolfo� (1996),� consideraron� que� esta� pared�
estaba�construida�con�un�barro�muy� fino�mezclado�con�una�secreción�cremosa,�
compuesta� por� heces� o� suelo� masticado,� a� la� cual� denominaron� “cemento”.�
Destacaron,�además,�que�este�“cemento”�no�mostraba�la�misma�composición�ni�
textura�que�se�observaba�en�el�suelo�donde�el�nido�era�construido,�debido�a�que�
el� primero� estaba� compuesto� únicamente� por� partículas� finas.� Los� cortes�
delgados�de�las�masas�de�cría�de����� �������colectadas�en�Navarro,�mostraron�
que� la� pared� de� la� cámara� ovígera� está� compuesta� por� material� del� suelo� no�
seleccionado� y� por� materia� orgánica� probablemente� agregada� por� el� adulto�
durante�su�construcción.�Considerando�además�que�la�pared�mostró�un�50�%�de�

� ����
materia�orgánica,�y�que�el�suelo�solo�un�30�%,� la�materia�orgánica�encontrada�
en�la�pared�podría�provenir�de�las�heces�del�escarabajo�adulto.��
�Halffter� y� Matthews� (1966)� incluyeron� a� ��� � ������� en� el� patrón� II�
(variación�1)�de�nidificación�basándose,�principalmente,�en�el�hecho�que�el�huevo�
es� dejado� dentro� de� una� cámara� ahuecada� y� aislado� de� la� provisión,�
característica�que�pudo�ser� confirmada�en� contribuciones�posteriores,� e� incluso�
en� este� estudio.� Halffter� y� Edmonds� (1982),� usando� la� misma� información,�
reubicaron�a�esta�especie�y�la�incluyeron�en�el�patrón�I�de�nidificación,�debido�a�
que�consideraron�como�carácter�más�determinante,�la�similitud�de�sus�nidos�con�
los�de�otras�especies� incluidas�en�dicho�grupo,�en� lugar�de� la�morfología�de� la�
cámara�ovígera.�La�presencia�de�una�pared�en�las�masas�de�cría,�como�pudo�ser�
demostrado� en� este� trabajo,� es� un� carácter� nuevo� que� sugiere� que� el�
comportamiento�de�nidificación�de���� � �������estaría�más�relacionado�con�el�
de� las�especies� incluidas�en�el�patrón�II.�Por�otro� lado,� la�presencia�simultánea�
en� el� nido� de� túneles� de� almacenamiento� y� de� nidificación,� sugiere� un�
aprovisionamiento� demorado,� como� se� observa� en� las� especies� incluidas� en� el�
patrón� II� de� nidificación.� Esta� característica,� podría� estar� relacionada� con� el�
hecho�que�el� adulto� necesitaría� ir� almacenando� la� bosta� en�uno�de� los� túneles�
mientras� construye� la� pared� de� la� masa� de� cría� en� el� túnel� de� nidificación.�
Finalmente,� la� incorporación� de� ��� � ������� en� el� patrón� II� de� nidificación,�
junto� con� los� phanaeinos,� concuerda� con� la� filogenia� recientemente� propuesta�
por�Philips������(2004).��
En� resumen,� �������� � ������� construye� nidos� con� túneles� de�
almacenamiento�que�son�usados�para�la�posterior�construcción�de�las�masas�de�
cría� (almacenamiento� demorado),� las� cuales� a� su� vez,� poseen� una� pared�
discreta.�Estas�dos�características�sugieren�que�el�comportamiento�de�nidificación�

� ����
de�esta�especie�está�más�relacionado�con�el�de�las�especies�incluidas�en�el�patrón�
II� de� nidificación,� que� con� las� del� patrón� I.� Finalmente,� los� estudios�
micromorfológicos�muestran�que����� �������construye�la�pared�de�las�masas�
de�cría,�como�así�también�las�de�las�cámaras�ovígeras,�con�material�del�suelo�no�
seleccionado�mezclado�con�materia�orgánica�fina.�Como�lo�indica�la�filogenia�más�
reciente�de� los�Phanaeini� (Philips�����,� 2004),��������� junto� con����� ����
conforman�el�grupo�más�basal�dentro�de�la�tribu,�pero�especies�de�Dichotomiini�
mas�basales�aún,�construyen�bolas�de�cría,�por�lo�que�las�masas�de���������no�
pueden�ser�consideradas� como�una�condición�primitiva,� sino� como�una�perdida�
secundaria�del� comportamiento�de�construir�bolas.�Esta�condición�un�poco�más�
derivada�del�comportamiento�de���������también�se�apoyaría�en�la�presencia�
de�una�pared�discreta�rodeando� las�provisiones�(Sánchez�y�Genise,�2008),�que�
sugeriría�que�esta�especie�muestra�un�patrón�de�nidificación�tipo�II.��
�
3�4�5H�� �����������"BB�
�
� Judulien� (1899)� fue� también� el� primero� en� estudiar� los� nidos� de�
��� ������������(Laporte),�los�que�describió�como�túneles�verticales,�no�
rectos,�excavados�de�15�cm�a�50�cm�de�profundidad.�Según�sus�observaciones,�
en� determinado� momento,� el� escarabajo� incrementa� el� tamaño� del� fondo� del�
túnel,�y�construye�allí�una�bola�de�cría�esférica�de�2,5���3�cm�de�diámetro,�a�la�
cual�la�hembra�le�superpone�en�uno�de�sus�polos�una�calota�semiesférica�de�0,5�
cm�de�alto.�Esta�calota,�formada�por�filamentos�gruesos�que�la�hacen�permeable�
al� aire,� recubre� una� pequeña� cámara� cónica,� donde� la� hembra� deposita� un�
huevo.�Morelli������(1996),�estudiaron�esta�especie�en�laboratorio�y�observaron�
que�existe�cooperación�hembra�macho�durante�la�construcción�del�nido,�donde�el�

� ����
macho�se�encarga�fundamentalmente�del�aprovisionamiento�y�el�mantenimiento�
de� las� galerías.� Al� iniciar� la� nidificación,� la� pareja� excava� un� túnel� vertical� e�
inclinado� debajo� de� la� fuente� de� alimento� (bosta),� que� es� aprovisionado� con�
bosta.� Luego� la� hembra� construye� la� bola� de� cría� en� el� fondo� ensanchado� de�
dicho� túnel,� o� la� pareja� excava� nuevos� túneles� de� similar� profundidad,� cuyos�
extremos�más�distales�convergen�en�el�primero,�para�modelar�allí�la�bola�de�cría.�
Cada�nido,�construido�en�condiciones�de�laboratorio�controladas,�puede�contener�
de�2�a�3�bolas�de� cría�en� cámaras� independientes,�mientras�que�en�el� campo,�
este�número� se� reduce� a�1� o�2.�Durante� la� construcción�de� la� bola� de� cría,� la�
hembra�moldea�una�esfera�de�bosta�que�cubre� luego�con�una�capa�delgada�de�
material�del�suelo,�y�en�uno�de�los�polos,�forma�una�depresión�donde�deposita�un�
huevo,� que� cubre� luego� en� su� parte� superior� con� una�mezcla� laxa� de� suelo� y�
fibras� vegetales� de� la� bosta.� La� cámara� pupal� es� una� cavidad� esférica,� de�
paredes� lisas,� con�un�casquete�circular�en�el� techo�que�externamente�presenta�
un� diseño� en� espiral,� que� logra� la� larva� como� resultado� de� los� movimientos�
ascendentes� del� clípeo.� Cabrera�Walsh� y� Gandolfo� (1996),� estudiaron� también�
las�bolas�de�cría�de��������,� las�cuales�describieron�en�datalle�y�de�manera�
conjunta� con� las� de� otras� dos� especies� del� mismo� género,� ��� �������
�������� (Chevrolat)� y� ��� ������� ����� (Harold).� Según� sus�
observaciones,� los� escarabajos� de� estas� tres� especies,� excavan� un� túnel� en� el�
que� depositan� en� el� fondo� una� bola� de� provisión� perfecta,� a� la� cual� le� van�
agregando�bosta�al�mismo�tiempo�que�van�incrementado�el�fondo�del�túnel�hasta�
formar� la� cámara� nido.� Una� vez� que� la� bola� alcanza� determinado� tamaño,� la�
hembra� excava� la� cámara� ovígera� en� uno� de� los� polos,� y� agrega� una� delgada�
capa�de�“cemento”�sobre�toda�la�superficie�externa�de�la�bola�de�cría,�incluida�la�
que�será�el�piso�de�la�cámara�ovígera.�Luego�deposita�un�huevo�en�el�interior�de�

� ����
la�cámara,�y�completa�el�techo�con�“cemento”,�dejando�un�orificio�pequeño�en�el�
centro.�Finalmente,� recubre� la�cámara�con� fibras�vegetales�de� la�bosta�secas�y�
con�una�fina�capa�de�“cemento”,�formando�una�especie�de�montículo.�Según�las�
observaciones� dadas� por� los� diferentes� autores,� el� comportamiento� de�
nidificacion�de�las�tres�especies�de���� �������mencionadas,�coincide�con�el�
patrón� tipo� II� de�Halffter� y� Edmonds� (1982),� comportamiento� característico� de�
los�Phanaeina.�
De�acuerdo�con�la�arquitectura�de�los�nidos�observada�en�el�campo�y�con�
la� estructura� de� las� bolas� de� cría� descriptas,� y� a�diferencia� de� lo� sugerido� por�
Cabrera�Walsh�y�Gandolfo� (1996),��������� y��������� poseen�muchas�más�
características� comunes� entre� sí� que� con� ��� �������.� De� hecho,� estas� dos�
especies� fueron� consideradas� durante� mucho� tiempo� como� subespecies� de�
��� �������(Edmonds,�2000).�Las�bolas�de�cría�de�las�dos�primeras�especies,�
poseen�una�forma�general�de�“gota”�con�un�canal�de�aireación�en�el�polo�superior�
que�comunica�el�interior�de�la�cámara�ovígera�con�el�de�la�cámara�nido�y�el�cual�
se�encuentra�constituido�por�fibras�vegetales�de�la�bosta�que�actúan�como�filtro.�
Las� bolas� de� cría� de���� �������,� en� cambio,� poseen�una�protuberancia� en� el�
polo� superior� bien� separada� de� la� cámara� principal� por� medio� de� un�
estrangulamiento� pronunciado� entre� ellas,� y� además� el� canal� de� aireación� no�
posee�estructuras�que�filtren�el�pasaje�del�aire,�quedando�completamente�abierto�
al�exterior.�En�todas�ellas,�la�cámara�ovígera�se�encuentra�excavada�en�la�bosta�
aprovisionada�en�el�interior�de�la�bola�de�cría,�y�sólo�en�el�caso�de�����������,�
la�pared�de� la�misma�se�encuentra�constituida�por�material�del�suelo,�mientras�
que�en�las�bolas�de�cría�de�las�dos�especies�restantes,�la�cámara�ovígera�posee�
sólo�un�tapiz�de�materia�orgánica.�Una�característica�común�a�las�tres�especies,�
está�dada�por� los�cambios�observados�en�la�estructura�de�las�bolas�de�cría�a� lo�

� ����
largo�del�desarrollo�de�la�progenie.�Se�han�podido�distinguir,�que�las�bolas�de�cría�
con�cámara�ovígera�poseen�una�pared�externa�constituida�por�material�del�suelo�
y�fibras�vegetales�de�la�bosta�(cuya�proporción�difiere�de�acuerdo�a�la�especie),�y�
un� relleno� constituido� fundamentalmente� por� bosta� caóticamente� distribuida,� a�
excepción� de� ��� ����,� donde� la� bosta� se� encuentra� ordenada� de� manera�
concéntrica.�En�el�segundo�estadio�de�las�bolas�de�cría,�puede�observarse�a�las�
larvas� en� distintas� etapas� del� desarrollo,� durante� las� cuales� comienza� a�
distinguirse�en�el�interior�de�las�bolas�un�ordenamiento�específico�del�material�del�
relleno�(posiblemente�bosta�no�consumida�mezclada�con�materia�orgánica,�heces�
de� la� larva),� en� pequeños������� yuxtapuestos� formando� bandas� superpuestas�
de� un� helicoide.� Cuanto� más� avanzado� esté� el� desarrollo� de� la� larva,� más� se�
completa�la�estructura�en�hélice,�la�cual�al�ser�terminada,�actuará�como�pared�de�
la�cámara�pupal.�El�tercer�estadio�observado,�corresponde�ya�a�las�bolas�de�cría�
emergidas,�o�a�aquéllas�donde�el�adulto�murió�antes�de�emerger�y�se�encuentra�
conservado�en�el�interior�de�la�cámara�pupal.�En�estas�bolas�de�cría,�la�pared�de�
la�cámara�pupal�se�encuentra�completa�mostrando�un�helicoide�abierto�en�el�polo�
superior,�donde�en�algunos�ejemplares�este�espacio�se�encuentra�ocupado�por�un�
“techo”�que�difiere�en�su�composición�en�dos�de�las�tres�especies�donde�pudo�ser�
observado.�En��������,�este�techo�se�encuentra�constituido�por�ojuelas�de�un�
material�muy�fino�que�presenta�crestas�poligonales�hacia�el�exterior�de�la�cámara�
pupal.� En� cambio,� en� S�� �������,� este� techo� esta� formado� por� ������
triangulares� constituidos� por� material� del� suelo,� los� cuales� también� presentan�
prolongaciones�hacia�el�exterior�de�la�cámara.��
La� presencia� de� una� cámara� pupal� conformada� por� una� pared� helicoidal�
abierta� en� el� polo� superior,� cerrada� por� un� techo� de� características� tan�
particulares� como� las� arriba� mencionadas,� constituyen� dos� datos� nunca� antes�

� ����
mencionadas�para�ninguna�especie�de�escarabajo�coprófago.�Existen�solamente�
dos� menciones� anteriores� relacionadas� con� estas� características.� Morelli� �� ���
(1996),� describieron� la� presencia� de� un� techo� construido� por� la� larva� de� ���
�����antes�de�empupar,�el�cual�según�sus�observaciones,�posee�un�diseño�en�
espiral� que� difiere� significativamente� del� techo� en� “ojuelas”� o� “������
triangulares”�observados�en���������y�����������,�respectivamente.�Y,�por�
otro� lado,� las� cámaras� pupales� de� �������������������� (Halffter� y� Edmonds,�
1982),� las� cuales� son� consideradas� de� las� más� elaboradas,� son� tubulares� y�
redondeadas�en�su�extremo�más�distal,�y�son�construidas�por� la� larva�antes�de�
empupar� a� través� del� agregado� progresivo� de� anillos� conformados� por� sus�
propias�heces.�Sin�embargo,�este�comportamiento�no�es�100�%�comparable�con�
el�descripto�más�arriba�para�las�tres�especies�estudiadas�de���� ������.��
Los� datos� filogenéticos�más�actualizados� sugieren�que���� ������� no�
sería� un� grupo� monofilético� y� que� estaría� conformado� por� uno� monofilético� y�
otros� dos� parafiléticos.� ��� ����� y� ��� �������� se� encuentran� en� clados�
parafiléticos� distintos� (Philips� �� ��,� 2004).���� ����� no� esta� incluido� en� esta�
filogenia�pero�su�similitud�morfológica�con���������(Edmonds,�2000)�sugiere�
que�ocuparía�una�posición�similiar�en�la�filogenia.�Se�muestra�aquí�que���������
y��������,�ademas�de�la�similitud�morfológica�comparten�también�similitudes�
comportamentales.� Estas� especies� conforman� los� grupos� más� derivados� de�
Phanaeini,� y� en� concordancia� con� esto,� muestran� bolas� de� cría� de� cierta�
complejidad�en�las�que�la�cámara�ovígera�no�se�encuentra�totalmente�incluida�en�
las�provisiones.�
�
3�4�8#����������� �����
�

� ����
� Halffter� y� Edmonds� (1982)� fueron� los� primeros� en� describir� el�
comportamiento�de�nidificación�y� las�bolas�de�cría�de�una�especie�de��������,�
��� �+� ���� Harold.� De� acuerdo� con� sus� descripciones,� los� nidos� de� esta�
especie�son�simples�y�presentan�una�sola�bola�de�cría�por�túnel�alojada�en�una�
cámara,�como� los�nidos�de������� ����descriptos�aquí.�Sin�embargo,�no�hacen�
referencia�a�túneles�de�alimentación�o�almacenamiento,�que�sí�han�sido�hallados�
en� los� nidos� de� ��� ��� ���,� particularmente,� los� últimos.� Los� túneles� de�
almacenamiento�de� los� nidos� de���� ��� ���,� no� fueron�hallados� en� simultáneo�
con�los�de�nidificación,�sugiriendo�que�su�función�podría�estar�más�ligada�a�la�de�
alimentación� de� los� adultos,� que� al� aprovisionamiento� de� bosta� para� la�
construcción� de� bolas� de� cría.� Este� comportamiento� sugiere� además� que� las�
bolas� de� cría� son� construidas� por� aprovisionamiento� directo,� como� lo� hacen�
algunos�Phanaeina� incluidos�en�el�patrón�II�de�nidificación�(Halffter�y�Edmonds,�
1982).�Pudo�determinarse�también,�que�los�túneles�de�nidificación�son�rellenados�
con�material� del� suelo�una�vez�que� la�bola�de� cría�ha� sido� terminada.�Dada� la�
ausencia� de� cuidado� parental� en� esta� especie,� el� rellenado� posterior� de� los�
túneles� podría� estar� actuando� como� una� barrera� al� ataque� de� los� nidos� por�
predadores�o�cleptoparásitos.�������
� En� relación� con� la� morfología� de� las� bolas� de� cría,� Halffter� y� Edmonds�
(1982)�describieron�las�bolas�de�cría�de�����+� ����y�definieron�a�la�pared�de�
la� misma� como� una� capa� o� lámina� muy� delgada� compuesta� por� material� del�
suelo.�Asimismo,�Cabrera�Walsh�y�Gandolfo�(1996),�describieron�las�bolas�de�cría�
de� ��� ��� ���� y� definieron� como� “cemento”� al� material� constituyente� de� la�
pared.�Sin�embargo,�los�estudios�micromorfológicos�presentados�en�este�trabajo,�
muestran�que�la�pared�de�la�bolas�de�cría�de������� ���,�esta�compuesta�en�un�
30� %� por� fibras� vegetales� de� la� bosta� y� material� del� suelo� ligeramente�

� ���
enriquecido� con� materia� orgánica.� Por� otro� lado,� las� bolas� de� cría� de� ���
�+� ���� poseen� una� cámara� ovígera� cerrada,� incluida� completamente� en� la�
pared,�y�orientada�en�posición�lateral�hacia�la�entrada�de�la�cámara�nido.�Halffter�
y�Edmonds�(1982),�mencionaron�también�la�presencia�de�un�tapón�de�aireación�
constituido� por� fibras� vegetales� de� la� bosta� (“fibrous� aereation� plug”� �����
Halffter�y�Edmonds,�1982)�incluidas�en�el�interior�de�la�protuberancia,�y�que�no�
entran�en�el�interior�de�la�cámara�ovígera.�En�los�nidos�de������� ���,�la�cámara�
ovígera�de� la�bola�de�cría�se�encuentra�orientada�dentro�de� la�cámara�nido,�en�
posición�lateral�y�superior,�mientras�que�la�entrada�a�la�cámara�es�lateral�como�
en� los� nidos� de� ��� �+� ���.� Otra� diferencia� encontrada� entre� estas� dos�
especies,�es�que�en������� ���� la�cámara�ovígera�no�esta� incluida�en� la�pared�
como�se�ha�descripto�para�����+� ���,�sino�que�esta�construida�dentro�de�la�
provisión�como�en�las�bolas�de�cría�de�las�especies�de����������incluidas�en�el�
patrón� II� de� nidificación.� Este� carácter� podría� estar� relacionado� con� diferentes�
condiciones�de�humedad�de�las�áreas�de�estudio�donde�fueron�hallados�los�nidos�
(Halffter�y�Matthews,�1966),�sin�embargo,�podría�también�deberse�a�diferencias�
en�el�comportamiento.�Cabrera�Walsh�y�Gandolfo�(1996),�quienes�estudiaron�las�
bolas�de�cría�de������� ����por�primera�vez�en�Buenos�Aires�y�bajo�condiciones�
climáticas�similares�a� las�de�Navarro,�describieron�también�a� la�cámara�ovígera�
como�una�excavación�en�la�bosta�aprovisionada�en�la�bola�de�cría.�No�obstante,�
no�hicieron�mención�alguna�sobre�la�presencia�de�una�capa�o�pared�que�aislara�
dicha�cámara�de�la�bosta,�aunque�si�mencionaron�la�presencia�de�una�secreción�
usada� por� el� adulto� para� alisar� su� interior.� En� las� bolas� de� cría� de���� ��� ����
estudiadas� para� este� trabajo,� se� observó� y� documentó� la� presencia� de� una�
lámina�muy�delgada�de�materia�orgánica�fina,�probablemente�heces�del�adulto,�
cubriendo� el� interior� de� la� cámara� ovígera� (Fig.� 21� B).� Este� carácter� pudo� ser�

� ����
observado�en�detalle�en� los�cortes�delgados�de� las�bolas�de�cría,�y�muestra�un�
caracter� que� no� fue� mencionado� hasta� ahora� para� esta� especie,� ni� para� ���
�+� ����(Halffter�y�Edmonds,�1982).�
La� protuberancia� de� las� bolas� de� cría� de���� ��� ���,� también� descripta�
para�����+� ���,� está� sujeta� a� diferentes� interpretaciones.� De� acuerdo� con�
Cabrera�Walsh�y�Gandolfo�(1996),�cuando�la�bola�de�cría�está�casi�terminada,�el�
adulto� excava� la� cámara� ovígera� y� deja� en� ella� un� huevo.� Luego� de� poner� el�
huevo,� completa� la� bola� con� el� agregado� de�más� bosta,� aplastándola� desde� el�
exterior,�y�construye�la�protuberancia�cilíndrica�y�el�tapón.�Ellos�describen�a�esta�
estructura� como�un� “botón� redondeado� y� plano”� (“round� and� flat� button”)� que�
suele� ubicarse� por� encima� de� la� cámara� ovígera,� bloqueando� la� entrada� a� la�
cámara�nido.�Sugieren�además�que�surge�como�resultado�de�la�forma�en�que�la�
cámara� nido� y� la� bola� de� cría� son� construidas,� y� que� este� botón� previene� el�
rolado�de�la�bola�en�el�interior�de�la�cámara.�Los�nidos�de������� ����estudiados�
en� Navarro�mostraron� que� las� bolas� de� cría� nunca� se� encuentran� sujetas� a� la�
cámara�nido�por�ninguna�estructura.�La�entrada�a�la�cámara�es�lateral,�mientas�
que� la� protuberancia� de� las� bolas� de� cría� está� orientada� hacia� arriba� con� un�
ángulo�de�45º,� lo�que�sugiere�que�no�hay�una�relación�directa�entre�la�posición�
de�estas�dos�estructuras.�Sumado�a�ésto,�en�todas�las�bolas�de�cría�colectadas�se�
observó�la�protuberancia�siempre�conectada�con�la�cámara�ovígera.�Los�estudios�
micromorfológicos� muestran� que� la� cámara� ovígera� no� está� completamente�
cerrada�y�que�se�comunica� con� la� cámara�nido,�por�medio�de� la�protuberancia�
cilíndrica.�El�interior�de�esta�protuberancia�es�hueco�y�posee�una�acumulación�de�
fibras� vegetales� de� la� bosta� que� crean� un� poro� de� aireación� (����� Halffter� y�
Edmonds,� 1982),� que� permite� el� intercambio� gaseoso� entre� el� interior� de� la�
cámara�ovígera�y�el�interior�de�la�cámara�nido.��

� ���
En� resumen,� �������� ��� ���� excava� túneles� de� almacenamiento�
probablemente�usados�como�túneles�de�alimentación,�y�construye�bolas�de�cría�
por� aprovisionamiento� directo.� La� cámara� ovígera� es� construida� dentro� de� la�
provisión�de�la�bola�de�cría,�y�está�aislada�de�la�bosta�por�una�capa�delgada�de�
materia�orgánica.�La�bola�de�cría�es�construida�en�la�cámara�nido�con�su�cámara�
ovígera� en�posición� lateral� y� superior,�mientras� que� la� entrada�a� la� cámara� es�
lateral.�Finalmente,� los�estudios�micromorfológicos�muestran�que�los�adultos�de�
������ ����agregan�fibras�vegetales�de� la�bosta�a� la�pared�de� las�bolas�de�cría�
durante� su� construcción,� y� que� solo� utilizan� materia� orgánica� fina�
(probablemente�heces)�para�la�construcción�de�la�pared�de�la�cámara�ovígera.���
Los� Dichotomiini� representan� uno� de� los� linajes� más� antiguos� entre� los�
Scarabaeidae,�y�conforman�además�un�grupo�parafilético�donde�los�(� ���������
se� encuentra�más� emparentados� con� los� Phanaeini� (Philips� �� ��,� 2004a).� Sin�
embargo,� �������� ��� ����muestra� un� comportamiento� complejo,� típico� del�
Patrón�II�de�nidificación�(Halffter�y�Edmonds,�1982).�
���
3�4�6#�� ������ ��"BB�
� En� relación� con� �������� sp.,� Barbero� �� ��� (1999),� estudiaron� una�
especie� en� laboratorio� exclusivamente� coprófaga� y� nocturna,� ��������
������ �Balthasar.�Según�sus�observaciones,�durante�la�nidificación,�la�hembra�
construye�una�cámara�ovalada�a�una�profundidad�de� la�superficie�de�3�cm�a�9�
cm,� que� se� comunica� con� el� exterior� por� medio� de� un� túnel� inclinado,�
parcialmente�obliterado.�Cada�cámara�aloja�una�sola�bola�de�cría,�piriforme,�de�
3,5�cm�de�alto�y�3,8�cm�de�diámetro,�promedio,�la�cual�se�encuentra�recubierta�
por�una�delgada�capa�de�material�del�suelo�de�3�mm�de�espesor�promedio.�De�la�

� ����
cámara�ovígera�sólo�mencionan�su�posición�en�la�bola�de�cría,� la�cual�es�apical.�
Los� autores� destacan� del� comportamiento� de� nidificación� observado� en� esta�
especie�de� �������,�la�construcción�de�una�única�bola�de�cría�por�cámara,�la�
presencia� de� una� capa� de�material� del� suelo� recubriendo� la� bola� de� cría,� y� el�
cuidado�maternal�prolongado�hasta�el�tercer�estadio�larval.�������
� La� especie� de� �������� estudiada� en� Anillaco� (La� Rioja),� ��
( ���������)���� �� �����,�posee�el�mismo�comportamiento�de�nidificación,�e�
incluso� la�misma� arquitectura� del� nido� y� similar� estructura� de� la� bola� de� cría,�
observadas� por� Barbero� �� ��� (1999)� para� �������� ������ .� Las�
observaciones�presentadas�aquí�aportan�nuevos�datos�sobre�algunas�estructuras�
presentes�en�las�bolas�de�cría.�Barbero������(1999)�sólo�mencionan�la�posición�
de� la� cámara� ovígera,� la� cual� al� igual� que� en� �� ��� �� ������ de� Anillaco,� se�
encuentra�excavada�en�el�polo�superior�de�la�bola�de�cría,�dentro�del�relleno.�Sin�
embargo,� no� se�menciona� en� esa� descripción,� que� dicha� cámara� se� encuentra�
completamente�cerrada�y�que�no�posee�ninguna�pared�que�aísle�al�huevo�de� la�
provisión,�como�fue�observado�en� ����� �� �����.�Finalmente,�la�micromorfología�
de� estas� bolas,� revela� la� presencia� de� pequeñas� ojuelas� de� materia� orgánica�
rojiza� en� el� relleno,� ordenada� de�manera� concéntrica,� que� no� fueron� vistas� en�
otrtas� bolas� de� cría,� pero� que� si� bien� se� desconoce� su� origen,� es� posible� que�
tengan�relación�con�el�tipo�de�excremento�usado�por�estos�escarabajos.�
�� Por� otro� lado,� si� bien� los� resultados� de� las� observaciones� del�
comportamiento� de� nidificación� de� �� ! �������"� ������� hechas� en� La�
Rioja� coinciden� en� general,� con� las� descripciones� arriba�mencionadas,� � ciertas�
características,� fundamentalmente�aquéllas� relacionadas�con� la�estructura�de� la�
bola� de� cría,� están� ausentes� en� estas� descripciones.� Por� otro� lado,� no� se� han�
publicado� hasta� el� momento� nuevas� y� detalladas� descripciones� de� dicho�

� ����
comportamiento�en�otras�especies�de� �������,�motivo�por�el�cual�muchas�de�
las�observaciones�presentadas�aquí,�resultan�novedosas.�De�todas�ellas,� la�más�
significativa,� es� la� presencia� de� una� cámara� pupal� helicoidal� construida� por� la�
larva�en�el� interior�de� la�bola�de�cría�antes�de�empupar.�Del�mismo�modo�que�
ocurre� con� las� tres� especies� de� ��� ������� y� �������� estudiadas,� estas�
cámaras� pupales� tan� características� no� habían� sido� mencionadas� hasta� el�
momento� para� ninguna� especie� de� �������,� ni� para� ninguna� especie� de�
ninguna�tribu.�A�pesar�de�no�haber�hallado�bolas�de�cría�en�distintos�estadios�de�
construcción�de�la�pared�helicoidal�de�la�cámara�pupal,�como�sí�fue�observado�en�
��� ������,�la�micromorfología�de�una�bola�de�cría�cerrada�con�cámara�pupal�
terminada,� reveló�además�de� la�estructura�en�hélice,� la�presencia�de� restos�de�
bosta� desordenados� en� contacto� con� ésta� y� con� la� pared� de� la� bola� de� cría,�
correspondientes�a�la�provisión�original,� los�cuales�no�forman�parte�de�la�pared�
de� la� cámara� pupal.� Así� mismo,� las� bolas� de� provisión� roladas� por� estos�
escarabajos� que� pudieron� ser� colectadas,� cortadas� y� observadas� bajo� lupa,�
mostraron� una� zona� externa�muy� delgada� de� fibras� ordenadas� que� pierden� su�
ordenamiento�hacia�el�interior,�y�se�intercalan�generando�zonas�más�porosas�en�
el� relleno.� Estas� evidencias� demuestran� que� el� adulto� construye� bolas� de� cría�
aprovisionando�de�manera�desordenada,�o�al�menos�no�espiralada,� la�bosta�del�
relleno,�y�que�dicha�estructura�es�construida�posteriormente�por�la�larva�durante�
su�desarrollo�como�pared�de� la�cámara�pupal.�Otro�resultado� interesante,�es� la�
similitud� encontrada� en� la� estructura� de� las� bolas� de� cría� de� �� ������,����
����� y��������,� todas�ellas� con�una� forma�general� tendiendo�a�gota,� casi�
piriforme,�y�con�un�canal�de�aireación�constituido�por�fibras�vegetales�que�actuan�
como�filtro�entre�el�interior�de�la�cámara�ovígera�y�el�exterior�de�la�bola�de�cría.���

� ����
� En�cuanto�a�la�filogenia,�los�Canthonini�son�un�linaje�primitivo,�y�a�la�vez�
polifilético,� conformado� por� diferentes� clados� definidos� por� diferentes� “sets”� de�
sinapomorfías� (Philips� �� ��,� 2004).� Es� claro,� dado� su� comportamiento� de�
nidificación�y�por�las�características�de�las�bolas�de�cría�recién�mencionadas,�que�
al�menos� ��������no�debería�ser�incluído�en�el�mismo�clado�que�incluye�a�los�
������� y� que� posiblemente,� en� un� futuro� con� nuevas� investigaciones,�
conformarán�tribus�diferentes.��
�
3�4�3#��������"BB�
�
������� es� un� género� con� muchas� especies� del� cual� se� conocen� tanto�
coprófagas� como� necrófagas,� y� en� relación� con� sus� nidos,� en� 1899� Judulien�
publicó� las� primeras�menciones� de� algunas� carcaterísticas� de� los� nidos� de� tres�
especies�distintas,�������������Germ,�������� ���Har�y������������Har.�De� la�
primera�especie,�remarca�la�estructura�de�la�bola�de�cría,�claramente�biesférica,�
y� además� que� esta� especie� de� ������� nidifica� debajo� de� la� bosta� o� de�
cadáveres,�de�manera�indistinta.�Según�sus�observaciones,�las�bolas�de�cría�de�la�
segunda�especie�son�similares�a� las�de� �����,�y� las�de� la�última,�son�más�
bien�piriformes,�y�según�él,�muy�parecidas�a�las�de��������,�aunque�mucho�
más� pequeñas.� Otra� característica� que� destaca� del� comportamiento� de�
nidificación� de� ������,� es� que� los� adultos� de���� ��������� y� de� ��� �������,�
permanecen� en� el� nido� hasta� que� se� completa� el� desarrollo� de� la� cría.� Otra�
especie� de� ������,� que� ha� sido� ampliamente� estudiada,� es� ��� &�����
&������LeConte.�Halffter�(1977)�describió�por�primera�vez�su�comportamiento�
de� nidificación,� del� cual� remarcó� la� construcción� de� bolas� de� cría� biesféricas,�
decoradas�posteriormente�por�pequeñas�bolas�fecales�que�la�larva�expulsa�desde�

� ����
el� interior�durante�su�desarrollo.�Estas�bolas�de�cría�son�originalmente�una�bola�
esférica�de�provisión,�a� la�que� la�hembra� le�agrega�en�una�segunda�etapa,�una�
bola�más�pequeña�para�construir�allí�la�cámara�ovígera,�con�el�fin�de�mantener�al�
huevo� aislado� de� la� provisión� de� la� cámara� principal.� Esta� especie� construye�
nidos� compuestos,� conformados� por� varias� bolas� de� cría� en� un� mismo� nido�
superficial,�como�resultado�del�esfuerzo�conjunto�de�la�hembra�y�del�macho�que�
conforman� la�pareja.�Este� tipo�de�nido�ha�sido� incluido�por�Halffter�y�Edmonds�
(1982),�en�el�patrón�V�de�nidificación.�
Las�dos�especies�de�������� estudiadas�para�esta�Tesis,� construyen�dos�
tipos� distintos� de� bolas� de� cría.� ������� sp.� 1� es� una� especie� coprófaga� que�
forma�bolas�de�cría�piriformes,�tal�vez�similares�a�las�de������������descriptas�
por� Judulien� (1899),� donde� la� cámara� ovigera� se� encuentra� directamente�
excavada� en� la� bosta� aprovisionada� en� el� relleno,� como� se� vió� en� los� cortes�
delgados,� y� la� cual� se� encuentra� abierta� en� el� polo� superior� y� parcialmente�
protegida�por� fibras�vegetales�que�emergen�de� la� cara� interna�de� la�pared.�En�
cambio,� ������� sp.� 2� es� una� especie� necrófaga� que� construye� bolas� de� cría�
típicamente�biesféricas,�tal�vez�más�parecidas�a�las�de�������� ���descriptas�por�
Judulien� (1899)� y� a� las� de���� &������ &�����,� donde� la� cámara�ovígera� se�
encuentra�construida�en�el�interior�de�una�segunda�bola�pequeña�adherida�a�otra�
de�mayor�tamaño,�donde�se�encuentra�la�provisión.����
Como�fue�mencionado�más�arriba,�es�posible�que��������y� ��������
no�conformen�el�mismo�clado�dentro�del�grupo�polifilético�de�los�Canthonini,�en�el�
que�distintos�tipos�de�comportamiento�han�sido�registrados�para�la�tribu.�Por�otro�
lado,�son�llamativas�las�diferencias�registradas�entre�los�������,�como�el�hecho�
que�algunas�especies�sean�coprófagas�y�otras�necrófagas,�o�que�contruyan�bolas�
de� cría� con� tantas� diferencias� morfológicas� (piriformes� vs.� biesféricas,� etc.)� e�

� ����
incluso� comportamentales� (larvas� que� en� algunas� especies,� como���� &������
&�����,�descartan�sus�heces�en�bolas� fecales�hacia�el�exterior,�vs.�otras�que�
las� depositan� en� el� interior� de� la� bola� como� un� fino� tapiz).� Es� evidente� que�
Canthonini� es� un� grupo� muy� grande,� con� muchas� diferencias� no� sólo� en� la�
morfología�de�sus�individuos,�la�cual�parece�ser�insuficiente�para�definir�grupos,�
sino� también� comportamentales,� las� cuales� posiblemente� analizadas�
conjuntamente�con�las�primeras,�permitirían�una�mayor�definición�de�taxones.��
�
3�4�;#���������'���"BB�.��������"B�
� �
� Si� bien� se� han� publicado� muchos� trabajos� sobre� el� comportamiento� de�
distintas�especies�de�Eucraniini,�la�mayoría�de�ellos�se�basa�fundamentalemnete�
en� la� descripción� de� las� disitntas� variantes� observadas� en� la� relocación� de� la�
provisión� por� parte� de� los� adultos,� para� la� alimentación� y� la� nidificación�
(Monteresino�y�Zunino,�2003;�Ocampo,�2004;�Ocampo�y�Philips,�2005;�Zunino,�
�����1989;�entre�otros),�siendo�muy�escasas�las�menciones�relacionadas�con�la�
arquitectura�de�los�nidos�(Monteresino�y�Zunino,�2003;�Ocampo�y�Philips,�2005).��
� En�relación�con�los�nidos�estudiados�aqui,�se�han�observado�en�el�campo�la�
presencia� de� algunas� estructuras� comunes� a� las� de� otras� especies,� como� por�
ejemplo,� las� presencia� de� antecámaras�de� almacenamiento�del� alimento,� como�
ha� sido�descripto� para�$���������������� �&�,�'� ������� ������� y�'��
���� ���� (Monteresino� y� Zunino,� 2003;� Ocampo� y� Philips,� 2005),� entre� otras.�
Pero,� hasta� el� momento,� no� se� ha� registrado� la� presencia� de� ningún� tipo� de�
cámara� de� cría� (masa/bola� de� cría),� las� cuales� probablemente� no� sean�
construidas,�ni�tampoco�se�habían�registrado�las�cámaras�pupales,�como�las�que�
fueron�halladas�en�los�nidos�de�$������������������estudiados�en�La�Rioja.�En�

� ����
referencia� a� ello,� Halffter� y� Matthews� (1966)� señalaron� en� su� libro� que� de�
acuerdo� a� las� observaciones� inéditas� de� Martínez,� $������������ construiría�
esferas� bien� regulares� donde� es� dejado� el� huevo� y� posteriormente,� se�
desarrollaría� la� larva.�Sin� embargo,� a� partir� de� las� observaciones�hechas�en� el�
campo,� y� de� las� cámaras� pupales� colectadas,� se� deduce� que� los� Eucraniini�
estudiados�no�hacen�ningún�tipo�de�cámara�de�cría,�sino�simplemente�cámaras�
de�aprovisionamiento�de�alimento,�y�que�sus�larvas�construyen�cámaras�pupales.�
Estas� últimas,� se� encuentran� completamente� cerradas,� son� subesféricas� y�
presentan� una� pared� de� forma� general� helicoidal,� similar� a� la� observada� en� el�
interior� de� las� bolas� de� cría� de� ��� ������-� �������� y� �������.� Las�
bandas� que� conforman� este� espiral,� se� encuentran� constituidas� por� pequeños�
������yuxtapuestos�de�materia�orgánica�oscura,�posiblemente�heces�de�la�larva�
con� agregado� de� material� del� suelo.� Dada� la� escasa� información� publicada� al�
respecto,�sería�conveniente�ampliar�el�estudio�de� los�nidos�a�un�mayor�número�
de� especies� para� verificar� si� dicho� comportamiento� se� encuentra� presente� en�
gran�parte�de�ellas,�y�ver�si�existen�variaciones�dentro�de�los�Eucraniini,�como�ya�
ha� sido� registrado� para� el� comportamiento� relacionado� con� la� relocación� del�
alimento.���
� Según� Philips� �� ��� (2004)� las� especies� de� Eucranini� no� voladoras� y�
arenícolas� como� las� estudiadas� aquí� son� miembros� derivados� del� grupo� que�
habrían� perdido� secundariamente� su� comportamiento� de� hacer� bolas,� ya� que�
'�������� ���� �����,� la� especie�más� basal� relacionado� con� los� Phanaeini�
las� construye.� Tendiendo� en� cuenta� que� en� todas� las� especies� de� Phanaeini�
estudiadas�en�esta�Tesis�que�contruyen�bolas�de�cría,�así�como�también���������
��� ����que�pertenece�a�Dichotomiini,�y� ���������������(Canthonini),�las�
larvas�a�su�vez�construyen�en�su� interior�cámaras�pupales�helicoidales,�al� igual�

� ����
que� las�descriptas�aquí�para�Eucraniini,�este�comportamiento�representaría�una�
sinapomorfía� que� habría� surgido� muy� tempranamente� en� la� evolución� de� las�
Scarabaeinae�antes�de�la�separación�de�las�cuatro�tribus.���
�
3�5H������� ������������������������������������� J����
� ������������������������� �����������������
�
Entre�las�especies�actuales�de�escarabajos�coprófagos�estudiadas�en�esta�
Tesis,� algunas� de� ellas� no� presentan� trazas� comparables� a������������.� No�
obstante,� su� estudio� reveló� interesantes� aspectos� aplicables� a� la� evolución� del�
comportamiento�en�Scarabaeinae�como�se�destacó�en�las�secciones�precedentes�
y� también� una� estrecha� relación� con� trazas� fósiles� de� otros� icnogéneros� que�
contribuyen�a�completar�el�registro�fósil�de�esta�subfamilia.��
Por�ejemplo,�si�bien�no�se�encuentran�trazas�similares�a�las�masas�de�cría�
sub�cilindricas� de� �������� � ������� en� Gran� Barranca,� sí� hay� registros,�
incluso� más� antiguos,� de� trazas� fósiles� ( ���� ����� �������)� atribuídas� a�
este�género�(Genise�y�Laza,�1998)�en�la�Formación�Asencio�en�Uruguay�(Eoceno�
inferior)� (Genise� �� ��,� 2002).� De� la� misma� manera,� las� cámaras� pupales�
construidas�con�una�capa�de�bosta�remanente�que�no�es�consumida�por�la�larva�
durante�el�desarrollo,�y�que�ésta�reordena�para�formar�un�helicoide�(conformado�
por� espiras� de� pequeños� ������ documentadas� aquí� en� escarabajos� de� cuatro�
tribus� distintas� de� Scarabaeinae),� son� análogas� a� trazas� fósiles� del� icnogénero�
'���� ����-� conocido� del� Paleoceno�Eoceno� de� Estados� Unidos� (Bown� �� ��,�
1997)� y� la� Argentina� (Genise� �� ��,� 2001).� Ambas� trazas� fósiles� llevarían� el�
registro� más� antiguo� de� los� Scarabaeinae� sudamericanos� hasta� el� Paleoceno�
Eoceno�Inferior.��

� ���
Entre� las� bolas� de� cría� actuales� estudiadas� en� esta� Tesis� y� que� tienen�
correlato�con������������,������������es�comparable�a�ejemplares�que�aun�
cuando�originalmente�pudieron�ser�esféricos,� subesféricos,�piriformes,�e� incluso�
con� protuberancia,� se� transforman� en� bolas� de� cría� abiertas� y� con� agujero� de�
emergencia,�al�salir�el�nuevo�adulto.�Tal�registro�aporta�en�forma�general�al�de�
las�Scarabaeinae�constructoras�de�bolas.��
Las�bolas�de�cría�de���� �������spp.�y� ��������������,�podrían�
mostrar�ciertas�similitudes�con�las�bolas�de�cría�fósiles� incluidas�en�����������y�
���������,�dependiendo�como�se�interprete�la�pared�gruesa�de�estas�icnoespecies.�
Si�consideramos�que�parte�de�esa�pared�fue�en�realidad�resto�de�las�provisiones,�
como�fue�comprobado�por�los�estudios�neoicnológicos,�entonces�la�presencia�de�
parte� de� la� cámara�ovígera,�más� especificamente� el� techo� incluido� en� la� pared�
sería�un�carácter�que�permitiría�atribuir�esas�icnoespecies�a�algunas�especies�de�
Phanaeini�y�Canthonini.��
Respecto� a� ��� ������� ��� y� ��� isp.� A,� no� se� han� encontrado� hasta� el�
momento� representantes� actuales� que� permitan� dilucidar� si� efectivamente� la�
cámara� ovígera� no� se� preserva� debido� a� que� habría� sido� excavada� en� la�
provisión,�o�que,�fundamentalmente�en����isp.�A,�la�cámara�ovígera�se�preserva�
como�una�cámara�hemiesférica�excavada�en� la�cara�externa�de� la�pared,�como�
habría�sugerido�Laza�(2006).�
Las� bolas� fósiles� biesféricas� de�Necochea� incluidas� en���� �����B������ son�
comparables� con� las� bolas� de� cría� construidas� por� la� especie� necrófaga� de�
������,����sp.�2.�No�sólo�la�forma�general�de�estas�bolas�coincide,�sino�también�
el�hecho�de�que�ambas�muestren�paredes�muy�delgadas,�que�en�las�bolas�de�cría�
actuales�se�encuentran�conformadas�únicamente�por�material�del�suelo,�ya�que�
no�se�incorpara�bosta�debido�al�tipo�de�alimentación�necrófaga.�

� ����
El� hecho� que� algunas� de� las� bolas� de� cría� fósiles� no� tiene� análogos�
modernos� entre� las� estudiadas� de� la� Argentina,� podría� sugerir� que� estas�
icnoespecies�de�������������correspondan�a�linajes�de�Scarabaeinae�distintos�
a� los� representados� actualmente� en� esta� región,� y� que� se� habrían� extinguido�
junto�con�los�grandes�mamíferos�cenozoicos�que�producían�la�bosta�utilizada�por�
ellos.��
Finalmente,� es� posible� hasta� el� momento� adjudicar� como� posibles�
productores�de�algunas�de�las�icnoespecies�de�������������de�Gran�Barranca�
a� especies� incluidas� en� dos� tribus� diferentes� de� Scarabaeinae:� Canthonini� y�
Phanaeini.�El�registro�más�antiguo�de�Scarabaeinae�fósiles�proviene�del�Cretácico�
de�Asia� y�del�Eoceno�de�Europa� (Krell,� 2007),� y�en�particular,� los� registros�de�
escarabajos�fósiles�más�antiguos�para�estas�dos�tribus,�provienen�del�Mioceno�de�
Kenia� (Canthonini)� y�del�Pleistoceno�de�USA�(Phanaeini)� (Krell,�2007).�De�esta�
manera,� los� registros� de� ������������ de� la� Formación� Sarmiento� (Eoceno�
medio�Mioceno�inferior)�en�Gran�Barranca�(Chubut),�estarían�predatando�en�30�
40�millones� de� años� al� primer� registro� de� cuerpos� fósiles� de� estas� tribus� (Fig.�
42),� período� para� el� cual� estarían� aportando� además� importante� información�
paleobiológica.�

� ���
�
�!�%�& 84� Esquema� que� muestra� los� primeros� registros� de� �����������,� en�comparación�con�los�de�escarabajos�fósiles�de�las�tribus�Canthonini�y�Phanaeini.��
3�8 H ������ � ����� �� � ���������� �� ����������� �� ���
����� ?����� @ ���� ��A@ �������� ������ � ?������ �����#
���������������A
3�8�2 # B&�!*!> @ $� "!$&$@ &)% $& *!& ��'&=!+& . $�"&B&�!*!> $�
��������������� ��& �&��& *&?���(&*!> �&�(!� =�A
�
En� los� niveles� del� Eoceno� Medio� inferior� (estacas� 9501� a� 9505)� de� la�
Formación� Sarmiento,� en� Gran� Barranca,� no� existe� registro� de� ejemplares� de�

� ����
�����������.� Esta� ausencia� podría� ser� consecuencia� de� los� procesos�
tafonómicos,� los� cuales� suelen� ser� los�primeros� considerados�usualmente� como�
responsables� de� la� ausencia� de� trazas� fósiles� en� un� depósito,� por� lo� que� no�
pueden� ser� descartados� completamente� aqui.� Sin� embargo,� los� paleosuelos� de�
este� intervalo�poseen� litofacies�similares,�y�un�desarrollo� también�similar,�a� los�
observados� en� niveles� más� jóvenes� que� poseen� abundante� cantidad� de�
�����������.� Por� otro� lado,� la� aparición� gradual� de� estas� trazas� en� Gran�
Barranca,� dada� por� el� incremento� del� número� de� icnoespecies� que� se� va�
registrando� a� lo� largo� de� toda� la� secuencia,� sugiere� que� la� ausencia� de�
������������en�los�niveles�inferiores�podría�ser�original�y�no�debido�a�procesos�
tafonómicos,� que� de� ser� responsables� habrían� controlado� de�manera� similar� a�
todas�las�icnoespecies.�Además�de�la�tafonomía,�pueden�analizarse�otras�posibles�
causas,�como�por�ejemplo,�las�condiciones�climáticas.�Junto�con�la�estabilidad�de�
las� condiciones� paleoambientales� mencionadas� anteriormente,� el� Óptimo�
Climático� del� Eoceno� Temprano� (Zachos��� ��,� 2001)� sugiere� que� el� clima� no�
habría� sido� restrictivo� para� el� establecimiento� de� estos� insectos,� los� cuales� se�
encuentran�mejor� representados� en� regiones� templadas� a� tropicales,� donde� la�
Temperatura�Media�Anual�es�alrededor�de�15º�C�y� la�Precipitación�Media�Anual�
excede�los�250�mm�(Halffter,�1991).�
El�registro�de�los�cuerpos�fósiles�de�Scarabaeinae�contribuye�escasamente�
al� entendimiento� del� orígen� y� dispersión� de� estos� escarabajos� coprófagos� del�
Eoceno�de�Patagonia,�productores�de������������.�El�registro�más�antiguo�de�
estos� fósiles� corpóreos� proviene� del� Cretácico� de� Asia� y� del� Eoceno� de� Europa�
(Krell,�2007).�No�existe�registro�de�escarabajos�fósiles�en�América�del�Sur�(Krell,�
2007),� pero� el� registro� más� antiguo� de� bolas� de� cría� fósiles� de� escarabajos�
coprófagos�corresponde�precisamente�a�las�������������del�Eoceno�Medio�de�

� ����
Gran� Barranca� utilizadas� en� este� trabajo� (Sánchez� �� ��,� 2006),� como� fue�
desarrollado�en�la�sección�anterior.�
Para�entender�completamente�la�aparición�de�los�escarabajos�productores�
de� ������������ en� Gran� Barranca,� habría� que� analizar� también� las�
preferencias�ecológicas�de�estos�insectos�(Genise�����,�2000).�Los�escarabajos�
coprófagos� están� adaptados� a� ambientes� abiertos� dominados� por� pastos� y�
habitados�por�grandes�mamíferos,�dos�aspectos�con� los�cuales�su�diversidad�se�
encuentra� totalmente� relacionada� (Halffter� y� Mathews,� 1966;� Halffter� y�
Edmonds,� 1982;� Hanski� y� Cambefort,� 1991).� Tanto� la� morfología� como� el�
comportamiento� de� estos� insectos� se� han� desarrollado� en� respuesta� al�
aprovechamiento�de� los�excrementos�en�hábitats�abiertos� (Halffter�y�Edmonds,�
1982).� Halffter� (1991)� mostró� que� en� Africa� central,� donde� la� megafauna� de�
mamíferos� hoy� día� sigue� siendo� abundante,� la� sabana� es� el� ecosistema� con� la�
mayor�diversidad�de�escarabajos�coprófagos.�Sin�embargo,�en�América�del�Sur,�
la�extinción�reciente�de�los�grandes�mamíferos�provocó�la�diversidad�más�alta�de�
Scarabaeinae�registrada�en�la�selva�tropical�lluviosa,�junto�con�la�adquisición�de�
la�necro�y�saprofagia�(Halffter,�1991;�Hanski�y�Cambefort,�1991).�Por�otro�lado,�
los� escarabajos� coprófagos� juegan� un� rol� ecológico� fundamental� en� la�
descomposición� de� los� excrementos� de� los� mamíferos� en� los� suelos� de� los�
pastizales,� manteniendo� activo� el� ciclo� de� los� nutrientes� y� permitiendo� el�
crecimiento�de�nuevas�gramíneas�(Bornemissza,�1976;�Heinrich�y�Bartholomew,�
1979).�Un�claro�ejemplo�de�la�fuerte�relación�que�existe�entre�estos�escarabajos,�
los� mamíferos� y� los� pastizales,� quedó� demostrado� en� Australia� durante� las�
décadas� del� 60� y� 80.� Fue� necesaria� entonces� la� importación� de� escarabajos�
coprófagos� exóticos� para� descomponer� el� excremento�producido� por� el� ganado�
introducido,� que� al� no� ser� procesado� efectivamente� se� fue� acumulando� en� el�

� ����
suelo�impidiendo�el�crecimiento�de�nuevas�gramíneas�y�favoreciendo�el�desarrollo�
de� los� insectos� plaga� (Bornemissza� 1969,� 1976).� En� climas� más� fríos,� los�
Scarabaeinae� son� aún� más� dependientes� de� hábitats� abiertos� expuestos� a� la�
mayor� insolación� (Halffter� y� Matthews,� 1966;� Halffter� y� Edmonds,� 1982).�
Además,� bolas� de� cría� de� gran� tamaño,� como� las� que� se� describen� en� este�
trabajo,� tuvieron� que� ser� preparadas� de� una� única� bosta� producida� por� un�
mamífero�de�gran�tamaño.��
En�consecuencia,�la�ausencia�de�������������en�las�secciones�inferior�y�
media� del� Miembro� Gran� Barranca� podría� deberse,� en� parte� a� causas�
tafonómicas,� y� a� la� escasez� de� grandes� mamíferos,� que� por� su� parte� estaría�
reflejando� condiciones� desfavorables� para� ellos� en� aquellos� primeros� hábitats�
dominados�por�pastos.�Entre�los�ciclos�faunísticos�reconocidos�por�Pascual������
(1996),� el� Miembro� Gran� Barranca� podría� ser� incluido� en� el� Ciclo� Faunístico�
Prepatagoniano,� el� cual� registra� un� cambio� paleoambiental� importante� de�
ambientes� arbolados,� cálidos� y� húmedos,� con� mamíferos� más� pequeños� a�
ambientes� más� abiertos,� más� fríos� y� secos,� con� mamíferos� más� grandes.� En�
particular,�este�ciclo�muestra�para� los�ungulados,�probablemente� los�herbívoros�
más�grandes�de�aquel�tiempo,�el�aumento�de�tamaño�y�la�hipsodoncia�(Pascual�
����,�1996),�ésta�última�provocada�por�el�incremento�de�pastos�en�la�vegetación�
(Pascual� �� ��,� 1996;� Kay� �� ��,� 1999;� Zucol� y� Brea,� 2005).� Ambas�
características� son� críticas� para� la� producción� de� las� bostas� usadas� por� los�
escarabajos�coprófagos�como� fuente�de�alimento�y�para� la�construcción�de�sus�
nidos.�Pascual������(1996)�mencionan�además�que�el�incremento�de�los�fitolitos�
de�gramíneas�y�de�las�������������durante�este�ciclo�apoya�la�hipótesis�de�los�
cambios�mencionados.�Aún� cuando� los� fitolitos� de�gramíneas� son� conocidos� en�
Patagonia� desde� el� Paleoceno� (Zucol,� 2005),� no� se� conoce� cuándo� los� hábitats�

� ����
dominados�por�pastos�comenzaron�a�expandirse�(Kay�����,�1999),�y�es�aún�más�
difícil� de� establecer� cuándo� estos� ambientes� comenzaron� a� tolerar� el� tipo� y� la�
abundancia�de�comida,�y�el�tamaño�de�los�herbívoros,�apropiados�para�sostener�
la� presencia� de� los� escarabajos� coprófagos.� Por� ejemplo,� el� análisis� de� fitolitos�
muestra� una� disminución� del� componente� arbóreo� durante� el� Eoceno,�
particularmente� de� las� dicotiledóneas� arbustivas� (Zucol� y� Brea,� 2005),� que�
habría� producido� hábitats� cada� vez� más� abiertos� hasta� el� alcance� de� las�
condiciones� óptimas� para� la� aparición� de� los� escarabajos� coprófagos.� Esta�
variación� florística� ha� sido� también� reconocida� por� el� cambio� en� el� tipo� de�
paleosuelos� entre� la� Formación� Koluel�Kaike� (Eoceno� inferior� a� medio),� que�
incluye� Ultisoles� formados� en� ambientes� boscosos� cálido�húmedos� (Krause� y�
Bellosi,� 2008),� y� el�Miembro�Gran�Barranca� (Eoceno�medio� alto)� con�Andisoles�
desarrollados� en� palmares� y� sabanas� subhúmedas� (Bellosi� y� González,� en�
prensa).�
Queda� documentado� en� esta� Tesis� que� el� primer� registro� de�
������������corresponde�al�Eoceno�Medio�alto,�alrededor�de�39.3�ma�(estacas�
9506�y�9508).�Para�ese�momento,�los�ejemplares�de�������������son�escasos�
y�aparacen�en�muy�baja�densidad�(3���4�especímenes�por�m2),�y�también�baja�
diversidad,� sugiriendo� que� los� cambios� ecológicos� del� Ciclo� Faunístico�
Prepatagoniano�habrían�sido�responsables�de�sustentar�un�hábitat�dominado�por�
pastos� que� fuera� conveniente� para� la� aparición� de� los� primeros� escarabajos�
coprófagos.�La�aparición�gradual�de�las�diferentes�icnoespecies�de�������������
a� lo� largo� de� la� secuencia,� parece� reflejar� la� real� aparición� de� los� escarabajos�
coprófagos� en� esta� localidad,� más� que� posibles� cambios� en� las� condiciones�
taxonómicas.��

� ����
Luego�de�este�comienzo�incipiente,�las�condiciones�ambientales�cambiaron�
desfavorablemente� durante� el� tiempo� representado� por� el� Miembro� Rosado�
(Eoceno� Medio� alto,� 37.28� �� 36.19� ma).� Para� entonces,� no� se� registran�
ejemplares� de� �����������,� debido� posiblemente� a� causas� tafonómicas.� En�
relación� con� esto�último,�Bellosi� y� colaboradores� (Bellosi,� en�prensa;�Bellosi���
��,�2002)�propusieron�un�clima�seco�para�aquella�edad�sugerido�por�la�presencia�
de�un�suelo�cálcico�fuertemente�desarrollado�(calcrete).��
A�partir� de� los�36.2�ma,�aproximadamente� (estaca�9508.5�09)� comienza�
una�explosión�de�������������que�se�mantiene�sin�cambios� importantes�a� lo�
largo� del� Eoceno� Superior� bajo� (estaca� 9510.5).� Esta� explosión� se� encuentra�
reflejada� por� (1)� la� gran� abundancia� de� ejemplares,�más� de� 100� especímenes�
colectados� por� estaca� en� la� mayoría� de� los� casos,� (2)� la� elevada� densidad,�
alrededor�de�40�100�especímenes�por�m2,�y�(3)�la�alta�diversidad�(la�mayoría�de�
las� icnoespecies�presentes�en�todas� las�estacas).�En�estos�paleosuelos�(estacas�
9508.5�a�9510.5)�no�se�observan�evidencias�de�una�concentración�secundaria�de�
los�ejemplares�de�������������dada�por�el�retrabajo�mecánico�del�sedimento,�
como� por� ejemplo,� estratificación� cruzada,� superficies� erosivas� internas,� o�
paredes�pulidas�de�las�trazas.�El�depósito�original�es�eólico�y�corresponde�a�una�
loessita�téfrica�maciza�(Bellosi,�en�prensa).�El�pobre�grado�de�madurez�de�estos�
paleosuelos,�también�descarta�la�idea�de�una�alta�concentración�de�trazas�fósiles�
debido�a�un�prolongado�tiempo�de�exposición�subaérea.�De�esta�manera,�durante�
el� Eoceno� Superior� bajo� habrían� ocurrido� cambios� ambientales� favorables� que�
involucraría� la� presencia� de� hábitats� dominados� por� pastos,� herbívoros� y�
escarabajos� coprófagos.� Este� ecosistema� podría� estar� indicado� también� por� el�
aumento� del� contenido� fitolítico� en� los� depósitos� de� pastos� cortos� meso�

� ����
microtérmicos�(tales�como�arundinoides,�estipoides�y�pooides)�durante�el�Eoceno�
Medio�superior�y�tardío�(Zucol�y�Brea,�2005).��
Durante� la� transición� Eoceno�Oligoceno,� solo� dos� o� tres� ejemplares� de�
������������ fueron�observados� en� la� sección� superior� del�Miembro�Vera.� La�
ausencia�de�bolas�de�cría�fósiles�en�este�miembro�podría�deberse�a�la�acción�de�
procesos�tafonómicos,�o�a�condiciones�ambientales�desfavorables,�como�el�clima�
seco,�y�posiblemente�más�frío,�tal�como�sugieren�las�litofacies�y�los�paleosuelos,�
asi� como� también� la� frecuente� caída�de� cenizas� volcánicas� (Bellosi,� en�prensa;�
Bellosi�y�González,�en�prensa).�
Los�parámetros�medidos�para�el�Oligoceno�Superior�bajo�(estacas�9512�y�
9512�13)�muestran� valores� similares� a� los� estimados� para� el� Eoceno� Superior�
bajo� (estacas� 9508.5� a� 9510.5),� sugiriendo� un� retorno� de� las� condiciones�
favorables� para� los� escarabajos� coprófagos� después� de� la� depositación� de� los�
sedimentos� loéssicos�del�Miembro�Vera.�Luego,�en�el�Oligoceno�Superior�medio�
(estaca�9514.5),�la�densidad�de�������������disminuye�significativamente�(6�
8�especímenes�por�m2),�pero�la�diversidad�se�conserva�y�se�mantiene�similar�a�la�
observada� en� estacas� anteriores.� Estos� valores� de� densidad� máxima� y�
relativamente�gran�abundancia,�persisten�hasta�el�Mioceno�Inferior�alto�(hasta�la�
estaca� 9520�20.5),� con� la� excepción� de� la� estaca� 9516� (Oligoceno� Superior�
medio),�que�muestra�una�densidad�máxima�similar�(6�specímenes�por�m2)�pero�
tomada� en� parches� aislados,� y� donde� la� abundancia� general� disminuye�
significativamente� (menos� de� 30� especímenes� colectados).� Sin� embargo,� como�
fue� mencionado� previamente,� en� esta� estaca� se� realizó� un� muestreo� parcial�
debido�a�su�difícil�acceso�y�a�su�escasa�exposición.��
�

� ����
3�8�4 H �&()!�" ���!"=�&$�" � '&" !* ��"B�*!�", $!+��"!$&$ .
&)% $& *!&��'&=!+&
�
Rspecto�a�la�diversidad�de�icnoespecies�y�a�su�abundancia�relativa,�puede�
destacarse�que������������ y���� ������ se�encuentran�presentes� siempre,�y� con�
una�abundancia�relativamente�alta�(entre�66�%�y�100�%�para�todas�las�estacas),�
a� la� largo� de� toda� la� secuencia.� Como� fue� explicado� más� arriba,� ��� ��������
podría� ser� el� resultado� de� la� emergencia� de� los� adultos� de� bolas� de� cría�
correspondientes�a�otras�icnoespecies,�tales�como����isp.�A�y����������,�y�debido�
a�ello�la�posibilidad�de�ser�registrada�es�mayor�que�la�de�las�demás�icnoespecies.�
De�manera�similar,����������es�abundante�en�todas�las�estacas,�debido�a�que�se�
consideran�dentro�de�este�grupo�a� todos� los� fragmentos�de�ejemplares�y�a� los�
especímenes� que� no� pudieron� ser� identificados.� El� número� máximo� de�
icnoespecies�de�������������se�registra�para�el�Eoceno�Superior�bajo�(estacas�
9508.5�9509� y� 9510),� y� para� el� Oligoceno� Inferior� y� Superior� medio� (estacas�
9512�9513�y�9514.5).�Sin�embargo,�en�la�estaca�9510.5�(Eoceno�Superior�bajo),�
y�en� las�estacas�9517�17.5�y�9517.6�17.7�(transición�Oligoceno�Mioceno),� falta�
solo� una� icnoespecie� para� alcanzar� el� máximo� valor.� Las� icnoespecies� que� se�
pierden� son���� ������� ��� o���� ������-�mientras� que���� ������� y� ��� isp.� A� se�
mantienen�presentes�en�todas�las�estacas�desde�su�aparición�en�el�Eoceno�Medio�
alto� y� Superior� bajo� (estacas� 9508� y� 9508.5�09),� respectivamente.� Entre� el�
Eoceno�Superior� alto� y� el�Oligoceno�Superior� bajo� (estacas�9508.5�09�y�9512�
9513)�las�abundancias�relativas�de�estas�dos�icnoespecies,�combinadas,�alcanzan�
el� 25� %� y� el� 30� %,� respectivamente.� Desde� su� aparición� hasta� el� Oligoceno�
Superior�medio�(estaca�9516),����isp.�A�es�en�proporción�mucho�más�abundante�
(13,23�%,)�que�����������(7,25�%).�Mientras�que�desde�el�Oligoceno�más�alto�

� ���
hasta� el� Mioceno� Inferior� alto� (estacas� 9517�17.5� a� 9518�19),� la� abundacia�
relativa� de� ��� isp.� A� es� menor� (2,2� %)� que� la� de� ��� ������� (6,7� %).� En� el�
Mioceno� Inferior� alto� (estaca� 9517.8�17.9),� ��� ������� ��� y� ��� �������
desaparecen� y� no� vuelven� a� registrarse� estas� icnoespecies� a� lo� largo� de� la�
secuencia,� lo� que� brinda� la� primera� evidencia� de� condiciones� ambientales�
desfavorables�para�los�escarabajos�coprófagos�en�Gran�Barranca.��
Como�ya�fue�sugerido�para�la�aparición�de�������������en�la�secuencia,�
los� procesos� tafonómicos� deberían� controlar� de� igual� manera� a� todas� las�
icnoespcies,�y�por�ello�la�disminución�gradual�de�la�icnodiversidad�surgiría�como�
resultado�de�un�deterioro�real�de�las�condiciones�ambientales.�El�último�registro�
de� ������������ (alrededor� de� 20� ma,� estaca� 9521�22)� muestra� una�
abundancia�(14�especímenes)�y�una�máxima�densidad�(2�3�especímenes�por�m2)�
similares� a� las� registradas� al� inicio� de� la� secuencia,� pero� una� diversidad�
levemente�mayor,�ya�que����isp.�A�se�encuentra�también�presente.�Estos�valores�
sugieren� un� incremento� gradual� de� las� condiciones� ambientales� desfavorables�
hacia� la� sección� superior� de� la� secuencia.� A� partir� del� Mioceno� Inferior� alto�
(estacas� 9523� a� 9525.5),� no� se� registran� evidencias� de� especímenes� de�
�����������,� sugiriendo� cambios� ambientales� desfavorables� contínuos� que�
conducen� a� la� desaparición� total� de� los� escarabajos� coprófagos� de� Gran�
Barranca.�Actualmente,�esta�localidad�es�dominada�por�la�vegetación�de�estepa,�
con�un�suelo�escasamente�cubierto�por�pastos,�bajo�un�clima�frío,�con�ovejas�y�
algunas�especies�de�camélidos,�y�sin�registros�de�escarabajos�coprófagos.�
�
3�8�5 H �' ! *��(� =� $� =&(&Q� $� '�" �C�(B'&��" $���������������
?��,�!N(�=���*%&=��!&'A."%��'&*!> *� �'=&(&Q�$�'�"(&(FI���"�
�

� ����
� La�Figura�35�A�muestra�que�desde�el�Eoceno�Medio�(estaca�9506)�hasta�el�
Oligoceno� Superior� bajo� (estaca� 9512)� el� tamaño� promedio� de�������������
(DE)�no�registra�cambios�significativos,�pero�luego,�el�tamaño�aumenta�hasta�el�
Oligoceno� Superior� medio� (estaca� 9514.5),� esto� es� durante� el� Deseadense�
(SALMA).� Después� de� este� aumento� significativo,� el� tamaño� promedio� de�
������������mantiene�valores�similares�a�lo�largo�de�toda�la�secuencia�hasta�
el�último�registro�de�estas�trazas.�Prácticamente�todas�las�icnoespecies�muestran�
estos�cambios�en�el�tamaño�de������������,�con�picos�máximos�(Figura�35�B)�
desde� el� Oligoceno� Superior� medio� (estaca� 9514.5)� hasta� el� Mioceno� más�
temprano� (estaca� 9517.6�17.7),� a� lo� largo� del� Deseadense� (SALMA),�
mayormente.�El�incremento�en�el�tamaño�promedio�se�debe�a�la�desaparición�de�
especímenes�pequeños�durante�el�Oligoceno�Superior�medio�(estaca�9514.5)�y�a�
la�aparición�de�los�ejemplares�más�grandes�a�partir�del�Oligoceno�Superior�bajo�
(estaca�9512�13),�junto�con�el�cambio�de�la�moda�a�clases�de�mayor�tamaño.�El�
máximo�DE�promedio�registrado�durante�el�Oligoceno�Superior�medio�se�debe�a�
la�inclusión�de�la�mayoría�de�los�especímenes�en�la�cuarta�clase�de�tamaño.��
El� tamaño� de� las� bolas� de� cría� de� escarabajos� coprófagos� está�
directamente�relacionado�con�el�tamaño�de�sus�productores�(Halffter�y�Matthews,�
1966;� Lengerken,� 1954;� Klemperer,� 1983;� Klemperer� y�Boulton,� 1976;� Sato� y�
Imamori,� 1987),� como� será� analizado� en� la� siguiente� sección,� pero�
probablemente�también�tenga�relación�con�el�tamaño�de�la�bosta�producida�por�
los�herbívoros.�A�pesar�de�no�haber�estudios�detallados�al� respecto,�Heinrich�y�
Bartholomew� (1979)� reportaron� que� ,��� ������ �������,� una� especie� de� gran�
tamaño�(25�gramos),�utiliza�exclusivamente�la�bosta�producida�por�los�elefantes,�
de�la�cual�otra�especie�grande,�.�������&������(10�gramos),�construye�bolas�
de�cría�que�pueden�alcanzar�el�tamaño�de�una�pelota�de�tenis.�Janssens�(1939)�

� ���
(en� Halffter� y� Matthews,� 1966)� postuló� que� la� distribución� de� las� grandes�
especies�del�género�,��� ������esta�estrechamente�ligada�a�la�de�los�mamíferos,�
particularmente�a�la�de�los�elefantes�(Kingston�y�Coe,�1977;�Hanski�y�Cambefort,�
1991).� La� relación� que� existe� entre� grandes� bolas� de� cría,� grandes� bostas� y�
grandes�mamíferos,�estaría�sustentada�por�el�hecho�que�cada�hembra�o�pareja�
prepara� sus� bolas� de� cría� a� partir� de� una� única� bosta.� De� esta� manera,� la�
construcción�de�bolas�de�cría�grandes�para�permitir�el�desarrollo�de�escarabajos�
coprófagos� grandes,� sólo� es� posible� de� bostas� grandes,� que� por� su� parte� sólo�
pueden�ser�producidas�por�mamíferos�grandes.�
� Esta� información� sugiere� que� el� incremento� en� el� diámetro� de�
������������ estaría� relacionado� con� la� presencia� de� grandes�mamíferos.� En�
concordancia� con� ésto,� Pascual� y� Ortíz� Jauréguizar� (1990)� indicaron� que� las�
faunas�del�Oligoceno�de�Patagonia,�y�otras�regiones�de�Sudamérica�adjudicadas�
al� Deseadense� (28� ma� aproximadamente),� incluyeron� a� los� mamíferos� más�
grandes.� Varios� ungulados� nativos� grandes,� incluyendo�)&��������,� junto� con�
gliptodontes,�mylodontes�y�edentados�y�algunos�otros�depredadores�importantes�
ligados�ecológicamente�con�estos�herbívoros,�son�considerados�como�mamíferos�
gigantescos� (Pascual� �� ��,� 1996;� Ortíz� Jaureguízar� y� Cladera,� 2006).� El� gran�
tamaño�alcanzado�por�algunos�mamíferos�durante�el�Deseadense�fue�claramente�
diferente�del�alcanzado�por� la�fauna�anterior�(i.e.�Tinguirririquense),�y�posterior�
(i.e.�Colhuehuapense).�El�máximo�tamaño�de�������������registrado,�a�partir�
del�Oligoceno�Superior�medio,�sugiere�que�probablemente�a�lo�largo�del�intervalo�
con�los�mamíferos�más�grandes�(Deseadense),�el�tamaño�máximo�alcanzado�por�
los� herbívoros� probablemente� ocurriera� durante� dicha� edad.� Por� último,� y�
tomando� en� cuenta� los� cambios� en� el� tamaño� promedio� de� ������������
observados�en�la�Figura�35�A,�puede�deducirse�que� los�mamíferos�anteriores�al�

� ����
Deseadense� fueron,� en� promedio,� probablemente� más� pequeños� que� los�
posteriores�a�dicha�edad.���
�
3�8�8# �'=&(&Q�$� '&" !* ��"B�*!�"$���������������,& N'!"!"$� '&
$!+��"!$&$$��"*&�&)&C�"*�B�>I&��"B��$%*=���"
�
El�tamaño�de�las�bolas�de�cría�de�escarabajos�coprógafos�es�un�parámetro�
que�está�directamente�relacionado�con�el�tamaño�de�sus�productores,�como�fue�
analizado�por�Halffter�y�Matthews�(1966)�y�por�otros�autores�(Lengerken,�1954;�
Klemperer,�1983;�Klemperer�y�Boulton,�1976;�Sato�y�Imamori,�1987).�Halffter�y�
Matthews�(1966)�estudiaron�la�relación�entre�el�diámetro�de�las�bolas�de�cría�y�la�
longitud�total�de�los�escarabajos�productores�pertenecientes�a�distintas�especies�
de� Scarabaeini,� y� encontraron� que� existe� una� correlación� muy� cercana� entre�
estos� dos� parámetros.� Este� resultado� sugiere� que� grandes� escarabajos�
construyen�bolas�de�cría�más�grandes�que� las�producidas�por�escarabajos�más�
pequeños.� Por� otro� lado,� Lengerken� (1954),� Klemperer� y� Boulton� (1976),�
Klemperer�(1983),�Sato�y�Imamori� (1987),� reportaron�que� las�bolas�de�cría�de�
cada� especie� de� las� que� ellos� estudiaron,� podían� cambiar� en� un� 30%� en� su�
tamaño.� Estos� datos� permiten� estimar� el� número� mínimo� de� especies� de�
escarabajos� coprófagos� en� Gran� Barranca.� De� esta� manera,� la� diversidad�
potencial� de� productores� de� �����������,� fue� estimada� considerando� las�
diferentes�icnoespecies�presentes�en�cada�clase�de�tamaño�(Figura�36).��
En�la�Figura�36,�en�el�Eoceno�Medio,�alrededor�de�39.3�ma�(estaca�9506),�
cuando�������������aparece�por�primera�vez,�el�número�total�de�productores�
que�puede�estimarse�es�2,�porque�en�cada�clase�de�tamaño�sólo�están�presentes�
��� �������� y� ��� �����,� que� en� este� caso� se� consideran� juntos� como� ya� fue�

� ����
explicado�en�la�metodología�(ver�Materiales�y�Métodos).�La�estaca�9508�muestra�
cuatro� posibles� especies� de� escarabajos� coprófagos,� considerando� que� en� la�
segunda�clase�de�tamaño�(II)�el�productor�de������������y����������podría�ser�el�
mismo� que� el� de���� ������,� las� cuales�muestran� además� el� mismo� DE.� En� la�
clase� III,� ��� �������� y� ��� ������ son� consideradas� como� construidas�
potencialmente�por�la�misma�especie�productora,�como�en�la�estaca�anterior.�En�
el� Eoceno� tardio� bajo,� representado� por� la� estaca� 9508.5�09,� se� observa� un�
incremento� significativo� en� el� número� total� de� posibles� productores,� como� se�
observa� también�en� la�estaca�siguiente,�9510.�Para� la�estaca�9508.5�09�puede�
estimarse� la� presencia� de� al�menos�9� especies� de�escarabajos� coprófagos,� y� 8�
para�la�9510.�Las�diferencias�más�importantes�entre�estas�dos�estacas�es�que�en�
la�clase�I�de�la�última,�el�productor�de�����������es�reemplazado�por�el�de����isp.�
A,�y�en�la�clase�III�����������está�ausente.�Seis�posibles�especies�de�escarabajos�
coprófagos�son� registradas�en�el�Eoceno�Superior�bajo� (estaca�9510.5)�y�en�el�
Oligoceno� Superior� bajo,� pero� el� último� registro� incluye� a� un� nuevo� productor�
incluído�en� la�clase�IV,�que�estaba�ausente�hasta�esta�estaca�y�que�además�es�
más�grande�en�tamaño.�En�el�Oligoceno�Superior�bajo,�pero�a�28.7�23.2�ma,�en�
la�estaca�9512�13,�se�observan�10�posibles�especies�de�escarabajos�coprófagos.�
En� el� Oligoceno� Superior� medio,� la� estaca� 9514.5,� muestra� 8� posibles�
productores� de� �����������,� pero� en� este� caso,� y� a� partir� de� aquí,� los�
escarabajos�más�pequeños�incluidos�en�la�clase�I�desaparecen�casi�por�completo,�
con�la�única�excepción�de�la�estaca�9520�20.5.�En�la�estacas�siguientes,�desde�el�
Oligoceno�Superior�medio�hasta�el�Mioceno�temprano�alto,�el�número�de�posibles�
productoress�disminuye�a�5�(estacas�9516,�9517.8�17.9�y�9518�19)�y�6�(estacas�
9517.6�17.7�y�9520�20.5),�con�la�excepción�de�la�estaca�9517�17.5�que�muestra�
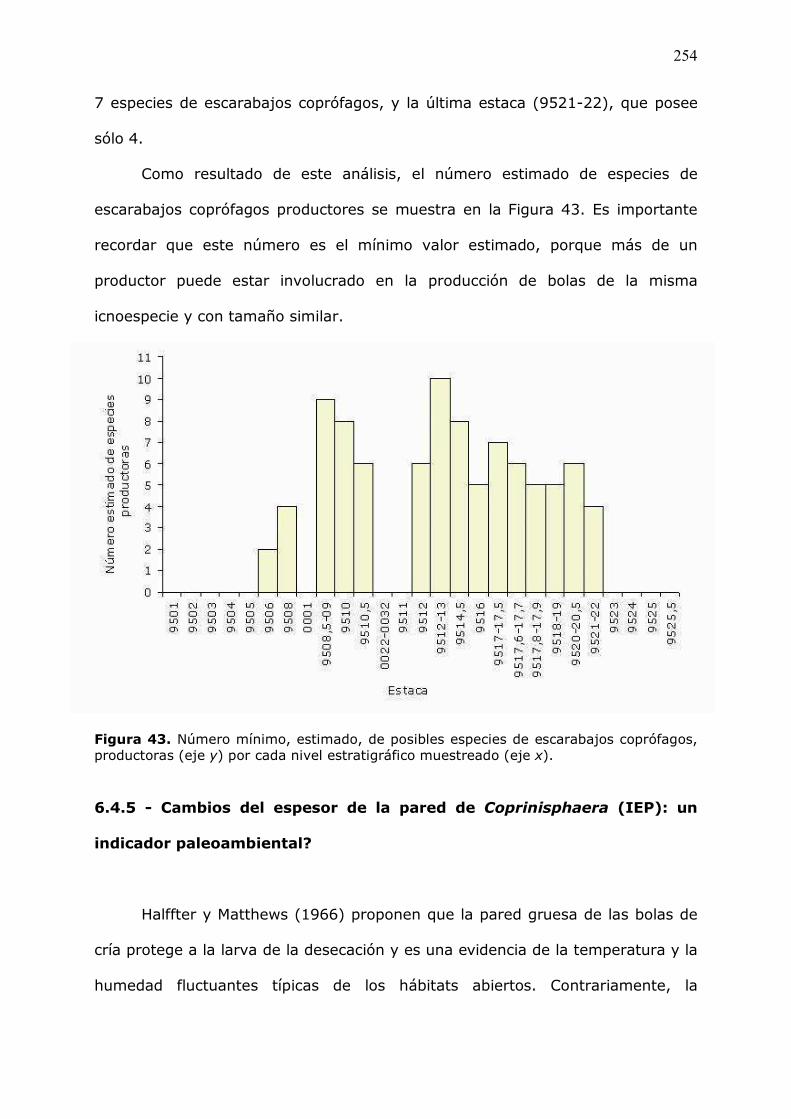
� ����
7�especies�de�escarabajos�coprófagos,�y� la�última�estaca�(9521�22),�que�posee�
sólo�4.��
Como� resultado� de� este� análisis,� el� número� estimado� de� especies� de�
escarabajos� coprófagos�productores� se�muestra�en� la� Figura�43.�Es� importante�
recordar� que� este� número� es� el� mínimo� valor� estimado,� porque� más� de� un�
productor� puede� estar� involucrado� en� la� producción� de� bolas� de� la� misma�
icnoespecie�y�con�tamaño�similar.�
�
�!�%�&85��Número�mínimo,�estimado,�de�posibles�especies�de�escarabajos�coprófagos,�productoras�(eje�&)�por�cada�nivel�estratigráfico�muestreado�(eje�+).�
3�8�6 # �&()!�" $�' �"B�"�� $� '& B&��$ $� �������������� ?���A, %
! $!*&$��B&'��&()!� =&'R
�
Halffter�y�Matthews�(1966)�proponen�que�la�pared�gruesa�de�las�bolas�de�
cría�protege�a�la�larva�de�la�desecación�y�es�una�evidencia�de�la�temperatura�y�la�
humedad� fluctuantes� típicas� de� los� hábitats� abiertos.� Contrariamente,� la�

� ����
estabilidad� en� la� humedad� y� en� la� temperatura� causaría� la� perdida� de� dicha�
protección.�Además,�para�estos�autores,� en�el�patrón� III�de�nidificación� (sensu�
Halffter� y� Mathews,� 1966),� el� cuidado� parental� llevado� a� cabo� por� la� hembra�
sustituye�a�la�pared�gruesa�y�disminuye�la�frecuencia�de�infecciones�microbianas�
(Halffter� y� Edmonds,� 1982).� Según� estos� autores� entonces,� una� pared� más�
gruesa� sugeriría� condiciones� más� secas� del� suelo,� así� como� la� ausencia� del�
cuidado�parental.��
Las� comparaciones� estadísticas� (Figura� 37� A)� muestran� que� no� hay�
diferencias� significativas� en� el� índice� (IEP)� a� lo� largo� de� la� secuencia,� con� la�
excepción�de�una�disminución�significativa�en�el�Oligoceno�Superior�bajo�(estaca�
9512�13)� que� representa� el�mínimo�valor� promedio� de� espesor� de� la� pared�de�
toda� la� secuencia.� Esta� distribución� se� observa� también� en� el� análisis� del� IEP�
diferenciado�por�icnoespecies�(Figura�37�B).�Esta�leve�variación�del�IEP�a�lo�largo�
de� la� secuencia� no� se� corresponde� con� los� cambios� en� las� condiciones� de�
humedad�que�son�registradas�por�litofacies�y�paleosuelos�(Bellosi,�2005;�Bellosi�y�
González,� en� prensa).� Asimismo,� los� depósitos� fluviales� intercalados� con� los�
Alfisoles�Ultisoles� en� el� Perfil� MMZ,� indican� condiciones�más� húmedas� para� las�
estacas� 9516� y� 9517�17.5� (Bellosi,� 2005),� donde� los� ejemplares� de�
������������ muestran� paredes� más� gruesas� que� para� la� estaca� 9512�13,�
donde�se�alcanza�el�mínimo�valor�de�espesor�de� la�pared.�En�consecuencia,�es�
posible� que� este� mínimo� valor� alcanzado� durante� el� Oligoceno� Superior� bajo�
estuviera�más�relacionado�con�elevada�humedad� local�en�el�suelo�debida�a�una�
freática� alta� ligada� a� los� canales� fluviales� próximos,� o� a� suelos� con� menor�
insolación,� que� a� las� condiciones� climáticas� generales.� Por� consiguiente,� la�
variación�del�IEP�no�es�un�indicador�confiable�de�las�condiciones�climáticas,�pero�
en�algunos�casos,�sí�podría�registrar�la�humedad�local�del�suelo.�

� ����
Entre�las�especies�actuales�estudiadas�para�esta�Tesis,�no�se�encontraron�
bolas�de�cría�con�paredes�gruesas�conformadas�solo�por�material�del�suelo,�sino�
por� una� capa� externa� de� este� material� y� bosta,� y� otra� más� interna� de� bosta�
remanente�de�las�provisiones.�La�única�bola�de�cría�construida�por�un�necrófago�
que� pudo� ser� colectada� (������� sp.� 2),� mostró� una� pared� de� suelo� muy�
delgada,�que�sugeriría�que�el�espesor�de�la�pared�podría�estar�relacionado�con�el�
tipo�de�alimentación�de�los�productores�de�las�bolas�de�cría.�Sin�embargo,�Fabre�
(1899)�describió�bolas�de�cría�de�un�escarabajo�necrófago,�)����������,�de�
6,5� cm� de� diámetro� ecuatorial� con� una� gruesa� pared� de� 1,5� cm� promedio� de�
espesor�conformada�por�material�del�suelo.��
�
3�6 # ��� �� ��������� ���� ������ �� � �� �� ������ �� ���� ��
�����������������������������?�����������A
Existen� dos� tipos� de� trazas� en� trazas� en� el� registro� fósil.� El� primer� tipo�
comprende� trazas� que� se� entrecruzan� con� otras� por� casualidad� o� debido� a�
cambios�en� las�condiciones�paleoambientales.�La�presencia�de�agallas�en�hojas�
que� fueron� consumidas� posteriormente� por� otro� insecto� y� que� se� preservan�
juntas�en�hojas�fósiles,�es�un�buen�ejemplo�de�este�tipo�de�trazas�en�trazas�(Wilf�
����,�2005).�Este�tipo�de�relación,�que�involucra�a�un�icnotaxón�que�entrecruza�
a� otro� después� del� establecimiento� de� una� nueva� condición� ambiental,� es� una�
muy� buena� herramienta� para� el� análisis� de� icnofábricas� (Taylor� �� ��,� 2003;�
entre�otros).�El�segundo�tipo�comprende�a�trazas�hechas�sobre�o�dentro�de�otras�
trazas� como� resultado� de� interacciones� biológicas� coetáneas,� tales� como�
parasitismo,� cleptoparasitismo� y� detritivoría,� entre� otras� (Chin� y� Gill,� 1996;�
Martin,�2001;�Mikuláš�y�Genise,�2003;�y�referencias�allí).�Diferentes�especímenes�

� ����
de� ������������ del� Eoceno� Medio�Oligoceno� de� la� Formación� Sarmiento� en�
Gran� Barranca� (y� Sierra� de� Talquino)� mencionados� aquí� en� otras� secciones�
presentan�los�dos�tipos�de�trazas�en�trazas�mencionados.�
En� la� revisión� icnotaxonómica� de� �����������,� Laza� (2006)� ilustra�
algunos� casos� de� trazas� en� trazas� que� pueden� ser� incluidas� en� el� primer� tipo�
mencionado�aquí�(Fig.�44�A�C,�F).�Entre�ellos,�se�observa�la�presencia�de�celdillas�
de� abejas� solitarias,� restos� o� impresiones� de� éstas,� tanto� en� el� relleno� (Laza�
2006,�Fig.�5A)�(Fig.�44�A�B)�como�en�la�superficie�externa�de�la�pared�(Fig.�44�C)�
de� especímenes� de� ������������ de� la� Formación� Sarmiento� (Chubut,�
Argentina).� En� este� ejemplo,� los� escarabajos� habrían� construido� primero� sus�
bolas�de�cría�en�el�suelo�donde,�posteriormente,� las�abejas�habrían�excavado�y�
creado� sus� celdillas� dentro� de� las� bolas,� por� casualidad.� Del� gran� número� de�
ejemplares�de�������������colectados�en�esta�localidad,�un�reducido�número�
de�casos�muestran�celdillas�de�abejas,�sugiriendo�que�esta�interacción�no�estaría�
registrando�cambios�en� las�condiciones�paleoambientales,�sino�más�bien�que� la�
superposición�podría�deberse�a�la�abundancia�de�bolas�de�cría�en�los�suelos.��
En� relación� con� bolas� de� cría� fósiles� de� escarabajos� coprófagos� se�
encontraron�también:���� ����� ���,�recientemente� interpretadas�por�Krause�
���.� (2008)�como�cámaras�de�alimentación�de�ninfas�de�cigarras� (Fig.�44�D);�
���������������,� interpretadas�por�Genise�(2004)�como�cámaras�pupales�de�
escarabajos�(Fig.�44�E)�y�túneles�indeterminados�y�sin�productor�asignado�(Fig.�
44�F).�Otro�caso�particular�ilustrado�por�Laza�(2006,�Fig.�5C),�muestra�pequeñas�
estructuras�ovoides�en�el�interior�de�un�espécimen�de�������������(Fig.�44�G�
H)� de� la� Formación� Sarmiento� (Chubut,� Argentina),� interpretados� por� él� como�
posibles�huevos�de�insectos.�Sin�embargo,�estas�estructuras�se�asemejan�mucho�
a� los������� fecales�producidos�por� los�escarabajos�rizófagos�(Fig.�45�A�B)�que�
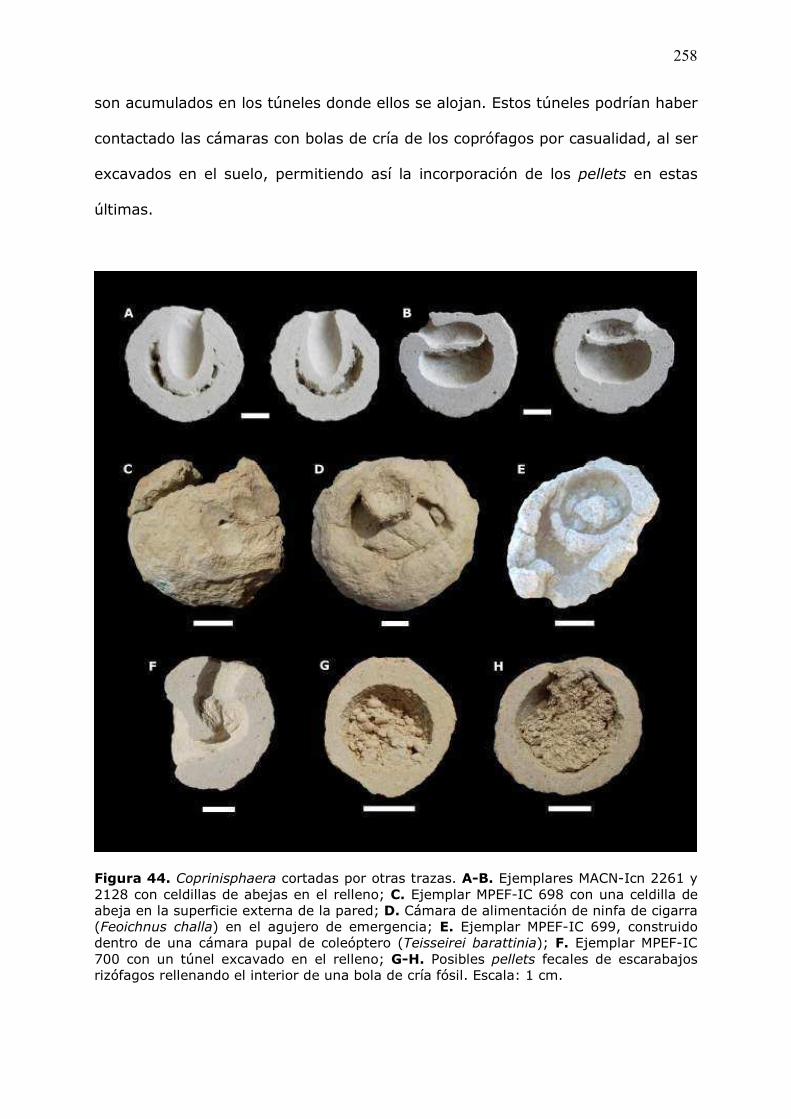
� ����
son�acumulados�en�los�túneles�donde�ellos�se�alojan.�Estos�túneles�podrían�haber�
contactado�las�cámaras�con�bolas�de�cría�de�los�coprófagos�por�casualidad,�al�ser�
excavados�en�el� suelo,�permitiendo�así� la� incorporación�de� los������� en�estas�
últimas.��
�
�
�!�%�&88��������������cortadas�por�otras�trazas.�#���Ejemplares�MACN�Icn�2261�y�2128�con�celdillas�de�abejas�en�el�relleno;����Ejemplar�MPEF�IC�698�con�una�celdilla�de�abeja�en�la�superficie�externa�de�la�pared;���Cámara�de�alimentación�de�ninfa�de�cigarra�(��� ����� ���)� en� el� agujero� de� emergencia;���� Ejemplar�MPEF�IC� 699,� construido�dentro�de�una� cámara�pupal�de�coleóptero� (���������������);����Ejemplar�MPEF�IC�700� con� un� túnel� excavado� en� el� relleno;��#��� Posibles������� fecales� de� escarabajos�rizófagos�rellenando�el�interior�de�una�bola�de�cría�fósil.�Escala:�1�cm.�
�

� ���
El� grupo� de� trazas� en� trazas� del� cual� puede� inferirse� un� significado�
paleobiológico� está� pobremente� representado,� y� analizado,� en� la� literatura�
icnológica.�Muchos�de�ellos� involucran�himenópteros.�Por�ejemplo,� la�presencia�
de� perforaciones� completas� e� incompletas� en� la� pared� de� celdillas� fósiles� de�
abejas,� han� sido� interpretadas� como� agujeros� hechos� por� parasitoides,�
cleptoparásitos�o�detritívoros�al�entrar�a� las�celdillas�desde�el�exterior,�como� lo�
sugieren� las� perforaciones� incompletas� presentes� en� la� superficie� externa� de�
dichas� celdillas� (Houston,� 1987;� Ellis� y� Ellis�Adam,� 1993;� Mikul� y� Genise,�
2003;� Genise� y� Cladera,� 2004;� entre� otros).� Las� avispas� han� sido� propuestas�
también� como� posibles� productores� de� estas� trazas,� debido� a� que� algunos�
Mutillidae�atacan�larvas�en�sus�celdillas,�cavando�a�través�del�suelo�o�perforando�
la�pared�(Evans�y�Eberhard,�1970).�Otro�ejemplo�de�parasitismo�de�avispas�fue�
registrado�por�Bown������(1997),�al�estudiar�capullos�de�avispas�del�Paleoceno�
Eoceno� provenientes� de� las� Formaciones� Claro� y� Colter,� de� Utah� (USA).� Ellos�
encontraron�pequeñas�estructuras�ovoides�dentro�de�otras�de�mayor�tamaño,�las�
cuales� fueron� interpretadas� como� capullos� de� avispas� parasitoides� dentro� de�
capullos� de� otras� avispas.� Martin� (2001)� mencionó� ejemplos� de� capullos� de�
insectos�de� la�Formación�Two�Medicine�(Cretácico�de�USA),�donde�halló�túneles�
de�diámetro�pequeño�conectados�con,�o�saliendo�de,�capullos�de�mayor�tamaño.�
Considerando�la�diferencia�de�tamaño�existente�entre�los�túneles�y�las�cubiertas�
pupales,� y� teniendo� en� cuenta� el� hecho� que� los� parasitoides� actuales� son�
típicamente�más� pequeños� que� sus� hospedadores,�Martin� (2001)� concluyó� que�
los�parasitoides�podrían�haber�sido�los�productores�de�los�túneles�pequeños.�
Otros�ejemplos�involucran�trazas�fósiles�de�escarabajos�coprófagos,�como�
las� que� se� presentan� en� esta� sección.� El� registro�más� antiguo� de� este� tipo� de�
trazas� pertenece� a� Frenguelli� (1938),� quien� mencionó� agujeros� laterales� en�

� ����
masas�de�cría�fósiles�de�escarabajos�coprófagos�del�Oligoceno�de�Patagonia,�las�
que� asignó� a� cleptoparásitos.� Brussaard� (1987)� describió� grandes� túneles�
meniscados�(“back�filling�traces”)�del�Holoceno,�entrecruzados�por�otros�túneles�
similares�pero�de�menor�calibre,�e�interpretó�a�los�primeros�como�construidos�por�
Geotrupinae,� y� a� los� más� pequeños� como� construidos� por� escarabajos�
cleptoparásitos,� como� $�������� ������.� Chin� y� Gill� (1996),� describieron�
túneles� cortados� por� otros� más� pequeños,� similares� al� material� descripto� por�
Brussard� (1987),� pero� en� este� caso� preservados� en� un� coprolito� de� dinosaurio�
herbívoro� del� Cretácico� de� la� Formación� Two� Medicine� (USA).� En� este� trabajo�
sugirieron� que� los� túneles� pequeños� habrían� sido� excavados� por� escarabajos�
paracópridos� en� túneles� de� mayor� tamaño,� posiblemente� producidos� por�
escarabajos� geotrupinos.� Mikul� y� Genise� (2003)� describieron� agujeros� y�
galerías�en�la�pared�de�masas�y�bolas�de�cría�fósiles�de�escarabajos�coprófagos,�
���� �������������y�������������isp.,�respectivamente,�los�cuales�fueron�
interpretados�como�trazas�de�actividad�hechas�por�organismos�cleptoparásitos�o�
detritívoros.��
�Mikul�y�Genise� (2003),�en�ese�mismo�trabajo,�dieron�por�primera�vez�
un� tratamiento� icnotaxonómico� a� las� trazas� hechas� en� cámaras� fósiles� de�
insectos� en� paleosuelos.� Crearon� así� tres� icnotaxones� nuevos:��������� �����
�����,� ��� ������ ��� y� ��� ����� ����������.� La� nueva� icnoespecie,�
�������� ����� ���,� se� separa� claramente� del� resto� y� se� basa� sobre� la�
ubicación� interna� de� las� depresiones,� contrariamente� a� lo� visto� en� las�
icnoespecies�conocidas�hasta�el�momento,�las�cuales�son�externas�o�representan�
agujeros� completos� en� la� pared� de� las� trazas� fósiles� hospedadoras� (Mikuláš� y�
Genise,� 2003).� En� el� material� examinado� para� esta� Tesis� se� han� encontrado�
algunas� diferencias� morfológicas.� El� holotipo� de� ��� ���� (Fig.� 8� A�C)� y� otros�

� ���
ejemplares�creados�en�la�misma�bola�de�cría,�son�depresiones�que�se�abren�en�el�
centro� o� en� el� margen� de� un� montículo� ovoide,� el� cual� podría� ser� material�
removido�de�la�depresión�al�ser�excavada�en�la�pared�de�las�cámaras�de�cría.�Sin�
embargo,� este� montículo� no� siempre� está� presente� en� los� ejemplares� de� ���
���,�como�muestran�algunos�paratipos�y�otros�especímenes�examinados�(Fig.�8�
A� y� B),� los� cuales� son� simples� depresiones� alargadas� y� de� sección� circular� a�
elíptica,� superficialmente� excavadas� en� la� cara� interna� de� la� pared� de� las�
cámaras� de� cría.� De� cualquier� modo,� los� dos� tipos� de� depresiones,� con� o� sin�
montículo,�se�encuentran�presentes�en�un�mismo�espécimen�de�������������
completamente� cerrado,� lo� cual� sugiere� que� ambas� estructuras� habrían� sido�
producidas�por�el�mismo�tipo�de�organismo.�
�������� ��������� puede� ser� interpretado� como�una� traza�hecha�por�
cleptoparásitos.� El� cleptoparasitismo� es� un� comportamiento� común� entre� los�
insectos� pertenecientes� a� la� comunidad� de� la� bosta.� Algunas� especies� de�
escarabajos�coprófagos�no�construyen�sus�nidos�y�utilizan�la�bosta�aprovisionada�
por� otras� especies� coprófagas� (Cambefort� y� Hanski,� 1991).� Estos� escarabajos�
cleptoparásitos� pertenecen� a� dos� subfamilias� de� Scarabaeidae,� Aphodiinae� y�
Scarabaeinae.� Existen� dos� subgrupos� de� cleptoparásitos� reconocidos� por�
Cambefort� (1991):� los� cleptoparásitos� de� escarabajos� coprófagos� rodadores� de�
bolas� (telecópridos),� y� los� cleptoparásitos� de� aquellos� escarabajos� que� las�
fabrican� debajo� de� las� bostas� (paracópridos).� Entre� los� Scarabaeinae,� las�
especies� de� pequeño� tamaño,� tales� como� ����� �����,� son� considerados�
cleptoparásitos� activos� ya� que� se� los� ha� visto� atacar� a� las� bolas� de� bosta�
mientras� están� siendo� rodadas� por� los� aquéllos� escarabajos� que� las� han�
construido� (Cambefort� y� Hanski,� 1991).� Sin� embargo,� las� especies� de� mayor�
tamaño,�tales�como�)����y�����������,�atacan�directamente�los�túneles�de�

� ����
almacenamiento�de�escarabajos�paracópridos�excavados�en�el� suelo� (Halffter� y�
Mathews,� 1966;� Cambefort� y� Hanski,� 1991)� o� sus� bolas� de� cría� (Halffter� y�
Mathews,�1966;�y�referencias�en�ellos).�La�subfamilia�Aphodiinae�incluye�algunas�
especies�de�cleptoparásitos�pasivos,� los�cuales�al�estar�presentes�en� las�bostas�
de� los� herbívoros,� pueden� ser� acarreados� accidentalmente� por� los� escarabajos�
paracópridos� al� extraer� de� allí� una� porción� de� bosta� para� ser� usada� como�
provisión�(Rougon�y�Rougon,�1980;�Cambefort�y�Hanski,�1991).�Otras�especies�
de�Aphodiinae�son�cleptoparásitos�activos,�como�se�ve�en�el�tercer�estadio�larval�
de�$��������������,�quien�excava�túneles�para�alcanzar�y�entrar�a�las�masas�de�
cría� de� �������� �������� (Klemperer,� 1980).� Algunas� especies� de� esta�
subfamilia� son� cleptoparásitos� obligados,� mientras� que� otros� adquieren� este�
comportamiento� de� manera� facultativa� cuando� las� condiciones� ambientales� se�
tornan�desfavorables,�como�por�ejemplo,�durante�la�temporada�seca�(Rougon�y�
Rougon,� 1980;� González�Megías� y� Sánchez�Piñero,� 2003).� Se� ha� visto� que�
especies�de�Aphodinae�y�Scarabaeinae�pueden�atacar�el�mismo�nido.�Cambefort�
(1984)� encontró� cinco� especies� de� cleptoparásitos� de� Scarabaeinae� y� dos� de�
Aphodiinae,�alojadas�en�un�mismo�nido�de�,��� �����.�
Existe�un�grupo�de�moscas�que�se�suma�a�los�organismos�ya�mencionados�
como�integrantes�de� la�comunidad�de� la�bosta,�y� los�cuales�muestran�también,�
distintos�grados�de�cleptoparasitismo�(Halffter�y�Mathews,�1966;�Hanski,�1991).�
Algunas�especies�de�Borboridae�(Diptera),�muestran�un�cleptoparasitismo�activo:�
se�mantienen� adheridas� al� dorso� del� escarabajo�mientras� este� rola� la� bola� de�
bosta,� o� en� otros� casos,� corren� o� vuelan� detrás� de� ellos� durante� el� rolado�
(Halffter� y� Mathews,� 1966).� Observaciones� de� campo� hechas� en� La� Rioja,�
mencionadas� en� otras� secciones,� revelaron� la� presencia� de� varias� especies� de�
moscas� volando� alrededor� de� las� bolas� de� �������� sp.,� mientras� éstas�

� ����
estaban� siendo� roladas� o� enterradas� por� los� escarabajos� adultos� (Fig.� 45� C).�
Sumado� a� estas� observaciones� de� campo,� se� encontró� en� el� laboratorio� que�
algunas� de� las� bolas� rodadas� por� adultos� de� �������� sp.� que� fueron�
colectadas� en� dicha� campaña,� contenían� en� su� interior� larvas� y� pupas�
pertenecientes�a�una�especie�de�Sarcophagidae�(Fig.�45�D�y�E).�En�relación�con�
el� cleptoparasitismo� pasivo,� es� probable� que� este� surja� como� resultado� de� la�
presencia�de� larvas�de�mosca�en�el� interior�de� las�bostas�de�herbívoros,� como�
pudo� observarse� también� durante� los� trabajos� de� campo� (Fig.� 45� F).� Richter�
(1918)�encontró�bolas�de� cría�piriformes�de� ������������ � atacadas�por�
����������������,�una�especie�de�Otitidae�(Diptera)�que�causa�la�muerte�de�
la� larva�del�escarabajo�por� inanición.�Con�estas�observaciones,�propuso�que� los�
huevos�de�la�mosca�habrían�sido�transportados�accidentalmente�con�la�bosta,�por�
el�escarabajo�adulto.��
Los�ejemplares�de��������� ���������fueron�hallados�en�el�interior�de�
especímenes� cerrados�de������������,� es�decir,� bolas�de� cría� fósiles�que�no�
muestran� ninguna� evidencia� de� agujero� de� emergencia.� Esto� sugiere� que� la�
actividad�del�cleptoparásito�productor�de��������habría� interrumpido,�activa�o�
pasivamente,� el� desarrollo� de� la� larva� del� productor� de� la� bola� de� cría.� La�
presencia� de� cleptoparásitos� en� las� bolas� de� cría� de� escarabajos� coprófagos�
podría�ser�el�resultado�de�dos�circunstancias�diferentes.�En�un�caso,�el�productor�
de�estas�trazas�sería�un�organismo�que�habría�estado�alimentándose�en�la�bosta�
del� herbívoro� y� que� habría� sido� acarreado� accidentalmente� por� el� escarabajo�
coprófago�al�tomar�una�porción�de�bosta�para�aprovisionar�en�su�nido.�Este�sería�
un� caso� de� parasitismo� pasivo,� inferencia� que� es� apoyada� por� la� ausencia� de�
agujeros�en�el�espécimen�de�������������que�posee�el�holotipo�de�������.�
En�esta�situación,� los�cleptoparásitos�no�habrían�entrado�activamente�a� la�bola�

� ����
de�cría�desde�el�exterior,�ni� la�habrían�abandonado�luego�de�su�emergencia.�En�
cualquiera� de� los� dos� casos,� puede� inferirse� que,� una� vez� completado� el�
desarrollo� larval,� dichos� cleptoparásitos� dejarían� la� bosta� aprovisionada� en� las�
bolas� de� cría� para� empupar� en� el� suelo,� como� si� se�encontraran� todavía� en� el�
interior� de� las� bostas� de� los� herbívoros.� Sin� embargo,� en� lugar� de� encontrar�
suelo� propiamente� dicho,� en� las� bolas� de� cría� habrían� encontrado� la� pared�
compuesta�por�material�del�suelo�que�las�recubre�externamente,�donde�habrían�
excavado�sus�cámaras�pupales,�como�se�ve�representado�por�los�ejemplares�de�
��� ���� en� el� interior� de� los� de������������.� En� esta� situación,� los� nuevos�
cleptoparásitos�adultos�no�habrían�logrado�salir�de�las�bolas�de�cría,�ya�que�no�se�
encontraron� en� la� pared� de� las� bolas� agujeros� abiertos� al� exterior.� Este� hecho�
sugiere� que� no� tendrían� las� herramientas� apropiadas� para� hacerlo,� lo� que�
apoyaría� la� idea� de� un� cleptoparasitismo� pasivo.� Sin� embargo,� no� pueden�
descartarse� completamente� otras� explicaciones,� tales� como� la� muerte� de� los�
cleptoparásitos� aún� en� estado� larval� por� otras� causas.� Por� otro� lado,� otros�
especímenes�de������������,�dentro�de�los�cuales�se�encontraron�ejemplares�
de��������excavados�en� la�pared,�poseen�agujero�de�emergencia�a�través�del�
cual� los� cleptoparásitos� podrían� entrar� y� salir� de� la� bola.� En� una� de� las�
������������estudiadas�se�encontraron�tanto�ejemplares�de��������como�un�
ejemplar� de� ��� ������ (Fig.� 8� D� y� E).� En� este� caso� particular,� ��� ������
representaría� la� entrada� o� salida� del� cleptoparásito,� mientras� que� los�
�������� ����� ���� representarían� cámaras� pupales� o� intentos� de�
perforaciones�en�la�pared,�según�se�trate�de�una�traza�mas,�o�menos�profunda.�
Algunas�moscas�empupan�dentro�de� la�bosta�(Fig.�45�F),�mientras�que�otras� lo�
hacen�en�el�suelo�justo�por�debajo�de�las�bostas�(Ferrar,�1980),�como�todos�los�
Aphodiinae� (Halffter� y� Edmonds,� 1982).� Los� Scarabaeinae,� por� otro� lado,� lo�

� ����
hacen�dentro� de� cámaras�de� cría� (Halffter� y� Edmonds,� 1982).�De�acuerdo� con�
todo� lo� discutido� hasta� aquí,��������� ����� ����podría� ser� producida� tanto�
por�moscas�que�empupan�en�el�suelo�como�por�Aphodiinae.��
��� ����� ����������� fue� descripto� originalmente� por� Mikul� y� Genise�
(2003)� en� una� traza� denominada� ���� ����� �������,� la� cual� fue�
interpretada� anteriormente� y� por� otros� autores� (Genise� y� Laza,� 1998),� como�
masa� de� cría� de� escarabajos� coprófagos.� Sin� embargo,� en� dicho� trabajo� del�
2003,�Mikul� y�Genise� no� sugieren� posibles� productores� para� estas� trazas.� El�
material�descripto�en�esta�sección,�se�encuentra�en�el�interior�de�ejemplares�de�
������������ isp.� de� la� Formación� Sarmiento,� Gran� Barranca,� extraídos� de�
paleosuelos�que�poseen�una�red�de�galerías�de�características�similares�a�los�que�
se� observan� en� el� interior� de� dichas� trazas.� Bellosi� �� ��� (2001)� realizan� la�
primera� interpretación�tentativa�de�estas� trazas�fósiles�en� los�paleosuelos�de� la�
Formación�Sarmiento,�Gran�Barranca,�y�proponen�a� las� termitas�como�posibles�
productores.�Tiempo�después,�Cosarinsky������(2005),�demuestran�que�dichas�
trazas� habrían� sido� hechas� por� otros� organismos� ya� que,� definitivamente,� no�
poseen� características� que� las� relacione� con� las� termitas.� Algunos� de� los�
caracteres� reportados� en� este� trabajo,� tales� como� “hypocoatings”� e�
intercalaciones� de� bandas� oscuras� en� los� túneles� (bandas� oscuras,� Fig.� 9� B),�
fueron�también�observadas�en�la�micromorfología�del�relleno�de�los�especímenes�
de������������.� La�nueva� información�que�se�presenta�aquí,�demuestra�que�
algunos�de�los�caracteres�vistos�en�la�red�de�galerías�de�los�paleosuelos�y�en�los�
ejemplares�de������������� (Fig.�9),� son�compatibles�con�aquellos�descriptos�
recientemente� por� Verde� �� ��� (2007)� en� trazas� de� lombrices.� Estas� trazas,�
halladas�en�paleosuelos�del�Pleistoceno�tardío�de�la�Formación�Sopas�(Uruguay),�
se� atribuyen� a� cámaras� esféricas� de� estivación� (����� ����� �� ������)�

� ����
conectadas�con������������������.�Las�estructuras�de�los�paleosuelos�de�la�
Formación� Sarmiento,� observadas� tanto� macro� como� micromorfológimante,� y�
que�se�han�descripto�aquí�(Fig.�9�D�E)�son�compatibles�con������ ������� �������
y�con� los� túneles�meniscados�conectados�a�estos,� sugiriendo�que� las� lombrices�
podrían� ser� los� productores� de� estas� redes� de� túneles.� Esta� inferencia� se�
encuentra�apoyada�además�por�la�presencia�de�������lenticulares�que�cubren�la�
superficie�interna�de�la�pared�de�la�cámara,�y�que�se�acomodan�formando�anillos�
en�su�cavidad�interna�(Verde�����,�2007)�(Fig.�9�E).�Estas�trazas,�presentes�en�
los� paleosuelos� de� Gran� Barranca,� se� conectan� con� ��� ����� �����������
presentes� en� los� ejemplares� de� ������������ (Fig.� 8� G),� sugiriendo� que� un�
mismo�productor�habría�sido�responsable�de�la�red�de�túneles�en�el�paleosuelo�y�
de� ��� ����������� en� las� bolas� de� cría� fósiles.� Por� otro� lado,� las� lombrices� son�
consideradas� como� un� integrante� más� de� las� comunidad� de� la� bosta� (Hanski,�
1991)�(Fig.�45�G),�y�su�rol�en�la�descomposición�y�desaparición�de�las�bostas�del�
suelo� es� de� gran� importancia� (Hendriksen,� 1991;� Holter,� 1979).� Finalmente,�
Morelli�(com.�pers.,�2007)�encontró�lombrices�dentro�de�bolas�de�cría�actuales�de�
escarabajos�coprófagos,�y�observó�que�éstas�destruyen�la�bosta�aprovisionada�en�
el�interior�de�las�bolas,�causando�como�consecuencia�la�muerte�de�las�larvas�en�
desarrollo.��
Las� lombrices� productoras� de� ��� ����� ����������� podrían� ser�
consideradas�como�cleptoparásitos�activos,�o�como�organismos�detritívoros.�Los�
agujeros�presentes�en�la�pared�de�������������se�encuentran�conectados�con�
galerías� de� túneles� internas,� lo� que� sugiere� que� estos� organismos� habrían�
entrado�activamente�a� las�bolas�buscando� la�bosta�aprovisionada�en�ellas.�Esta�
teoría� descarta� la� posibilidad� de� que� las� lombrices� se� comportaran� como�
cleptoparásitos�pasivos�transportados�accidentalmente�por�los�escarabajos�de�las�

� ����
bostas� de� los� herbívoros.� Dado� que� las� ������������ con� ��� ����������� se�
encuentran� muy� meteorizadas� y� bioturbadas,� es� difícil� inferir� la� posible�
emergencia�del�escarabajo�adulto,�y�en�consecuencia,�es� imposible�averiguar�si�
las� lombrices� comieron� la�bosta� fresca�aprovisionada� (cleptoparasitismo�activo)�
causando�la�muerte�de�la�larva�por�hambre,�o�si�éstas�entraron�en�bolas�de�cría�
ya� emergidas� con� el� fin� de� conseguir� restos� de� la� provisión� (detritívoros),� o�
ambos.� En� solo� un� caso� se� puede� inferir� que� las� lombrices� actuaron� como�
organismos�detritívoros,�al�usar�el�agujero�de�emergencia�de� la�������������
abierta,�para�entrar�o�salir�de�ésta�(Fig.�8�J).��
Estos� resultados� demuestran� que� el� registro� icnofósil� proporcionado� por�
������������para�los�Scarabaeidae�de�la�Patagonia�Central�durante�el�Eoceno�
Medio�al�Mioceno� Inferior�no� solamente� incluye�a� los�productores�de� las�bolas,�
sino�también�a�parte�de�la�comunidad�de�organismos�asociados�a�éstas,�o�a�las�
bostas�con�las�que�eran�aprovisionadas�como�en�la�actualidad.�

� ����
�
�!�%�&86����Túneles�rellenos�con�������fecales�producidos�por�larvas�de�escarabajos�rizófagos�(Formosa);���Detalle�de�una�larva�y�los�������fecales�producidos�por�esta�(La�Pampa);���� Mosca� posada� en� el� dorso� de� un� ejemplar� de� �������� sp.,� durante� el�enterramiento� de� la� bola� de� provisión� (La� Rioja);� �#��� Larva� (�)� y� pupario� (�)� de�Sarcophagidae�desarrollándose�dentro�de�bolas�de�provisión�construidas�y�enterradas�por� ��������sp.�(La�Rioja);��#���Larva�de�mosca�(�)�y� lombriz�(�)�encontradas�dentro�de�bosta�de�caballo�(Ushuaia).�Escala:�1�cm.�����
3�3#������������� ������� ��� ���
�
3�3�2 # �' ��''� � $� �������������� . �' B&'��"%�'� *!�*% $& =�
(%�"=�& % S(���"!(!'&�$�I!=�'!=�"
�

� ���
Como�ya�fue�mencionado,�las�icnoespecies�de�������������involucradas�
en�este�estudio,�����������,����isp.�A�y�����������(Fig.�6),�poseen�relleno�en�la�
cámara� principal.� El� relleno� de� las� bolas� consideradas� “abiertas”� (����������)�
puede�ser,�en�mayor�medida,�material�proveniente�del�suelo�circundante,�que�es�
empujado� hacia� el� interior� de� la� cámara� vacía� por� el� adulto� al� emerger,� o�
introducido� pasivamente� por� otros� procesos� posteriores.� Por� otro� lado,� los�
fitolitos�de�la�bosta,�la�que�originalmente�ocupaba�la�cámara�principal�de�la�bola�
de�cría,�habrían�sido�ingeridos�por�la�larva�en�desarrollo�y�redepositados�por�ella�
en�la�pared�al�tapizarla�con�sus�propias�heces�antes�de�empupar.�En�dicho�caso,�
se�esperaría�encontrar�un�contenido�fitolítico�en�el�relleno,�similar�al�preservado�
en�el�paleosuelo�circundante.�En�cambio,�en�aquéllas�bolas�donde�el�adulto�no�ha�
emergido,�es�decir,�en�las�bolas�consideradas�“cerradas”�(���isp.�A�y����������),�
es�esperable�encontrar�en�el�relleno�interior�de�la�cámara,�el�contenido�fitolítico�
original�de�la�bosta�completamente�preservado,�o�parte�de�éste�redepositado�en�
la� pared,� dependiendo� del� grado� de� desarrollo� alcanzado� por� la� larva.� Sin�
embargo,�estas�bolas�no�se�encuentran�completamente�cerradas�ya�que�siempre�
presentan� un� canal� de� aireación� por� donde�material� del� suelo� puede� entrar� y�
rellenar� el� interior� de� la� cámara.� En� consecuencia,� en� las� bolas� “cerradas”,� el�
contenido� fitolítico� del� relleno� puede� ser� el� resultado� de� la� combinación� de� los�
fitolitos�presentes�en�la�bosta�original,� los�del�suelo,�y�también�puede�depender�
del�comportamiento�de�la�larva�en�desarrollo�en�su�interior.�
El� análisis� estadístico� (Fig.� 38� A,� Tabla� 4)� revela� que� en� once� de� los�
catorce� ejemplares� de� ������������ estudiados,� de� las� tres� icnoespecies�
mencionadas,�el�número�de�fitolitos�encontrado�en�las�muestras�de�paleosuelo�no�
difiere�significativamente�(��>�0,05)�de� la�cantidad�hallada�en�el�relleno;�y�que�
en� los� tres�ejemplares�restantes�(todos�����������),�el�número�de� fitolitos�del�

� ����
paleosuelo�es�significativamente�mayor�al�del�relleno.�El�hecho�que�el�paleosuelo�
y� el� relleno� muestren� una� cantidad� similar� de� fitolitos� en� la� mayoría� de� los�
ejemplares,� ya� sean� “abiertos”� o� “cerrados”,� sugiere� que� en� los� primeros� el�
contenido� fitolítico� de� ambas� muestras� estaría� representando� al� del� suelo�
original;�y�que�en�los�otros,�el�contenido�fitolítico�del�relleno�similar�al�del�suelo�
sería� consecuencia� de� la� combinación� de� los� tres� factores� previamente�
mencionados.�Los�tres�ejemplares�de������������que�muestran�en�el�paleosuelo�
mayor� número� de� fitolitos� que� en� el� relleno,� sugieren� que� el� relleno� podría�
provenir�de�otro�horizonte�o�nivel�del�suelo�o�de�una�zona�más�distante,�y�con�
otro�contenido�fitolítico,�o�que�algunos�procesos�tafonómicos�habrían�modificado�
el� relleno,� o� en� último� caso,� que� estas� bolas� habrían� sido� removidas� de� su�
ubicación�original�(ver�discusión�mas�abajo).�
Estos�resultados�muestran�que�en� la�mayoría�de� los�casos�el� relleno�y�el�
paleosuelo�poseen�un�contenido�fitolítico�similar,�y�que�en�ningún�caso,�el�relleno�
de�los�ejemplares�de�������������preserva�mayor�cantidad�de�fitolitos�que�el�
paleosuelo� circundante,� incluso� a� pesar� de�haber� sido�un� relleno�originalmente�
conformado�por�bosta.�Esto�podría�suceder�debido�a�que�los�fitolitos�de�la�bosta�
fueron� redepositados� en� la� pared� por� la� larva,� o� debido� a� que� algunos� suelos�
podrían� concentrar� mayor� cantidad� de� fitolitos� que� la� bosta� en� sí� misma.�
Finalmente,� estos� resultados� demuestran� que,�más� alla� del� contenido� fitolítico,�
los�ejemplares�de������������� no�pueden�ser�analizados� como�una�muestra�
única� e� independiente,� sin� tener� en� cuenta� que� parte� de� la� bola� se� está�
analizando,�o�sin�considerar�el�paleosuelo�circundante.�
�
�

� ���
3�3�4#�&B&��$$���������������*� *� =�&(&.�� S(���$�I!=�'!=�"
G%��'��''� �.�'B&'��"%�'�*!�*% $& =�
�
El�análisis�estadístico� (Fig.�38�A,�Tabla�4)� revela�que�diez�de� los�catorce�
ejemplares� de� ������������ estudiados,� poseen� la� pared� con� un� número� de�
fitolitos�significativamente�mayor�que�el�hallado�tanto�en�el�paleosuelo�como�en�
el� relleno.� Estos� resultados� apoyan� la� hipótesis� que� en� las� paredes� de� las�
������������ se� preserva� parte� de� las� provisiones� originales� de� bosta,� como�
surge�de�los�resultados�de�los�estudios�neoicnologicos�presentados�aquí.�Por�otro�
lado,� estos� resultados� demuestran� además� que� la� concentración� de� fitolitos� es�
mayor�en�la�bosta�que�en�el�suelo,�debido�a�que�su�presencia�produjo�en�la�pared�
un� incremento� significativo� de� la� concentración� de� fitolitos.� El� suelo� original�
probablemente� habría� sostenido� una� pobre� a� moderada� cobertura� vegetal� de�
plantas� con� fitolitos,� como�pudo�observarse� en� el� suelo� de�Chuquis� (La�Rioja).�
Por�otra�parte,�con�estos�resultados�es�factible�deducir�la�dieta�de�los�herbívoros,�
o�al�menos�la�ingestión�selectiva�entre�plantas�que�contienen�fitolitos,�ya�que�la�
cantidad� adicional� de� fitolitos� de� la� pared� provendría� directamente� de� la� bosta�
producida�por�los�herbívoros.�
Cuatro�ejemplares�de������������� (MPEF�IC�720,�721,� 724�y�731)�no�
muestran� diferencias� en� el� número� de� fitotlitos� entre� las� muestras� de� pared,�
relleno�y�paleosuelo,� sugiriendo�diferentes� interpretaciones�en�cada�caso.�En�el�
ejemplar�MPEF�IC�720,�el�número�de�fitolitos�encontrado�en�la�pared�es�similar�al�
del� paleosuelo� y� relleno,� sugiriendo� que� podría� tratarse� de� necrofagia;� o� de�
dietas� carentes� en�plantas� con� fitolitos;� o� concentraciones� similares� de� fitolitos�
en� la� bosta� y� el� suelo,� como� consecuencia� de� condiciones� paleoambientales�
particulares.� Los� Scarabaeinae� necrófagos� construyen� bolas� de� cría� similares� a�

� ����
las� de� los� escarabajos� estrictamente� coprófagos,� pero� las� aprovisionan� con�
materia� orgánica� en� descomposición� (Halffter� y� Matthews,� 1966;� Halffter� y�
Edmonds,� 1982).� En� estos� casos,� la� provisión� original� de� la� bola� de� cría� no�
contribuiría�con�el�agregado�de�fitolitos�adicionales�a�la�pared.�Otras�posibilidades�
son� que� las� fibras� vegetales� de� la� bosta� pertenezcan� a� plantas� con� ningún�
contenido� fitolítico,� o� muy� escaso,� lo� cual� no� es� regla� general� para� los�
escarabajos� coprófagos� que� suelen� estar� asociados� a� mamíferos� que� se�
alimentan�de�pastos.�La�última�explicación�probable�consiste�en�que�la�cantidad�
de�fitolitos�concentrada�en�las�bostas�y�en�el�suelo�del�paleoambiente�en�el�cual�
fueron� construidas� estas� bolas� de� cría,� fuera� similar,� sugiriendo� condiciones�
ambientales� intermedias� a� las� registradas� en� las� localidades� de� Navarro� y�
Chuquis.��
En�el�segundo�ejemplar�(MPEF�IC�721),�el�número�de�fitolitos�hallado�en�la�
pared� es� similar� al� del� relleno,� y� ambos� mostraron� significativamente� menor�
cantidad�de�fitolitos�que�el�paleosuelo.�El�conteo�fue�repetido�en�dos�preparados�
diferentes,� que� dieron� los� mismos� resultados,� para� descartar� posibles� errores�
experimentales.�Entonces,�considerando�que�la�pared�es�construída�con�material�
del� suelo� y� que� los� escarabajos� pueden� evitar� el� uso� de� bosta,� pero� que� en�
ningún� caso� podrían� eliminar� o� reducir� selectivamente� la� cantidad� de� fitolitos,�
este� resultado� presenta� dos� posibles� explicaciones.� Una� posibilidad� está� más�
relacionada�con�la�concentración�diferencial�de�fitolitos�entre�el�suelo�y�la�bosta.�
En�un�paleoambiente�similar�al�de�Navarro,�la�bosta�preservada�en�la�pared�de�la�
bola�de�cría�original,�habría�reducido�la�concentración�de�fitolitos�de�esta�parte�de�
la�bola,�en�lugar�de�aumentarla.�Sin�embargo,�el�relleno�de�estas�bolas�“abiertas”�
también�tiene�menos�fitolitos�que�el�paleosuelo,�y�lo�cual�no�puede�ser�explicado�
como�en�el�caso�de�las�bolas�“cerradas"�debido�a�la�presencia�potencial�de�bosta�

� ����
preservada.� En� consecuencia,� otra� posibilidad� es� que� este� ejemplar� de�
������������ corresponda� a� una� bola� de� cría� construida� en� otro� lugar� y�
transportada,�aún�cuando�no�hay�evidencias�sedimentológicas�claras�que�apoyen�
esta� explicación.� La� removilización� de� la� bola� podría� producirse� en� el� mismo�
suelo� sin�mediar� un� transporte� por� agua,� como� fue� documentado� en� Chuquis,�
donde� el� trabajo� de� roedores� cavícolas� produce� una� reubicación� importante� de�
las� bolas� de� cria.� El� relleno� y� la� pared� representarían� entonces� la� composición�
fitolítica� del� suelo� original,� diferente� de� aquella� donde� la� bola� de� cría� fósil� fue�
encontrada.�En� los�dos�últimos�ejemplares� (MPEF�IC�724�y�731),� el� caso�es�el�
inverso:�el�número�de�fitolitos�encontrado�en�las�dos�porciones�de�la�pared�(PE�y�
PI)� no� difiere� significativamente� del� hallado� en� el� paleosuelo,� sin� embargo,� los�
tres� poseen� un� mayor� número� de� fitolitos� que� el� relleno.� La� explicación� sería�
similar�a�la�del�caso�anterior.��
Estos� resultados� muestran� que� en� la� mayoría� de� los� casos,� como� se�
postuló�en�la�hipótesis�correspondiente,� la�pared�posee�un�número�diferente�de�
fitolitos�que�el�relleno�o�el�paleosuelo;�y�que�en�particular�para�los�especímenes�
estudiados,�la�mayor�cantidad�puede�ser�el�resultado�de�fitolitos�preservados�en�
la�capa�de�bosta�remanente,�y�por�aquellos�que�pasan�por�el�tracto�digestivo�de�
la� larva�al� consumir� las�provisiones,�hasta� ser�depositados�en� la�pared� con� las�
heces,�en�un�paleoambiente�con�moderada�a�pobre�cobertura�vegetal�de�pastos�
y/o� palmeras.� Los� cuatro� casos� excepcionales� sugieren� necrofagia,� o� dietas�
carentes� de� plantas� con� fitolitos,� o� bolas� movidas� de� su� posición� original,� o�
condiciones�paleambientales� con�mayor�abundancia�de�pastos�y/o�palmeras.�El�
mayor� número� de� fitolitos� en� la� pared� debido� a� la� preservación� de� fibras�
vegetales�de�la�bosta,�permite�el�uso�de�estas�trazas�fósiles�como�un�instrumento�
para�la�inferencia�de�la�dieta�de�los�herbívoros.�

� ����
��
3�3�5 # �' S(��� $� I!=�'!=�" $� '& B&�=� ! =�� & $� '& B&��$ $�
���������������""!(!'&�&'&$�'&B&�=��D=�� &
�
Los� resultados� obtenidos� en� el� análisis� de� las� dos� capas� de� la� pared� de�
�����������,�muestran�que�si�bien�en�seis�de�los�diez�especímenes�analizados�
la� capa� interna� de� la� pared� posee�mayor� cantidad� de� fitolitos� que� la� externa,�
dicha� diferencia� no� es� significativa� estadísticamente.� La� presencia� de� más�
cantidad� de� fibras� vegetales� de� la� bosta� en� la� capa� interna� de� la� pared,� se�
debería�a�la�preservación�de�los�restos�de�la�provisión�original�o�al�agregado�de�
las�heces�larvales.�Sin�embargo,�y�a�pesar�de�poder�ser�analizadas�y�explicadas,�
las�diferencias�encontradas�entre�las�dos�capas�de�la�pared�no�son�significativas�
estadísticamente� (�� >� 0,05).� Estos� resultados� indicarían� entonces,� que� las�
paredes� externas� construidas� con�material� de� suelo�habrían� sido�delgadas� y� la�
mayor�parte�de�la�pared�de�las�������������estudiadas�estarían�formadas�por�
la�preservación�de�bosta�remanente�de�las�provisiones�originales�como�en�el�caso�
de�las�bolas�actuales�emergidas.�
�
3�3�8H� I��� *!&$� '&$!�=&$� '�"K��)F+���"B��$%*=���"$�)�"=&&
B&�=!� $� '&" $!I��� *!&" K&''&$&" � '&" I��*%� *!&" $� (��I�=!B�" $�
I!=�'!=�"� =��'&B&��$.�'B&'��"%�'�*!�*% $& =�?��'��''� �A
�
� Considerando�entonces,�que� la�pared�de� las������������� se�encuentra�
constituída�por�material�del�suelo�y�por�restos�de�bosta�de�la�provisión�original�no�
consumidos�por� la� larva� (ver� ítem�3�3�5),� las�diferencias�en� las� frecuencias�de�

� ����
morfotipos� de� fitolitos� entre� el� paleosuelo� y� la� pared,� permiten� inferir� la� dieta�
basada�en�plantas�con�fitolitos�de�los�herbívoros�productores�de�bosta.���
� El� resultado� de� este� análisis� revela� que� en� la� mitad� de� los� catorce�
ejemplares�de�������������estudiados�(MPEF�IC�720,�725,�727�730,�732),� la�
distribución� de� frecuencias� de� morfotipos� de� fitolitos� no� difiere� entre� las�
muestras.�Con� la� excepción�del� ejemplar�MPEF�IC�720,� previamente� analizado,�
que�no�muestra�ninguna�prueba�precisa�del�agregado�de�bosta�a�la�pared,�para�
los� otros� ejemplares� los� resultados� sugieren� que� los� herbívoros� no� habrían�
mostrado�preferencias�por�determinadas�plantas,�dentro�de�las�que�poseen�mas�
fitolitos.�En�dichos�casos,�los�herbívoros�habrían�ingerido�plantas�con�fitolitos�con�
la�misma�proporción�en�que�estas�estaban�disponibles�en�el�ecosistema,�como�se�
ve�reflejado�por�la�frecuencia�de�sus�fitolitos�en�el�paleosuelo.��
Los� ejemplares� MPEF�IC� 721,� 726� y� 732� revelan� que� la� distribución� de�
frecuencias�de�morfotipos�de�fitolitos�no�difiere�entre�las�muestras�de�paleosuelo�
y�relleno,�pero�sí�se�diferencian�de�la�distribución�en�la�pared.�El�ejemplar�MPEF�
IC� 721� probablemente� corresponda� a� una� bola� de� cría�movida� de� su� posición�
original,� como� ya� fue� discutido� más� arriba,� y� por� ello� no� se� analizará�
nuevamente.�Los� resultados�de� los�otros�dos�ejemplares� (MPEF�IC�726�y�732),�
sugieren� que� los� herbívoros� involucrados� habrían� sido� más� especialistas,�
seleccionando� determinado� tipo� de� plantas� con� fitolitos.� Las� muestras� de�
paleosuelo� y� relleno,� relacionadas� con� el� primer� ejemplar,� muestran� que� las�
palmeras�(Gl)�(Fig.�40,�fitolitos�1�2)�y�los�pastos�con�prismatolitas�(Pr)�(Fig.�40,�
fitolitos� 8�16)� eran� las� plantas� con� fitolitos� más� abundantes� en� el� ecosistema�
para� ese� momento.� A� pesar� de� ello,� en� este� primer� ejemplar,� los� pastos� con�
braquiolitas� (Br)� fueron� las� plantas� más� seleccionadas� por� los� herbívoros�
productores�de� la�bosta,�a�pesar�de�ser� las�más� raras�en�el�ecosistema�(72�%�

� ����
más� abundantes� en� la� bosta� que� el� suelo),� seguidos� en� segundo� lugar� por�
plantas� más� comunes,� las� palmeras� (46� %� más� abundantes).� En� el� segundo�
ejemplar,� las� muestras� de� paleosuelo� y� relleno,� revelan� que� los� pastos� con�
prismatolitas�(Pr)�(Fig.�40,�fitolitos�8�16),�dicotiledóneas�y�otras�plantas�que�no�
pudieron�ser�identificadas�(Otros:�puzzle�y�otros�morfotipos�indeterminados,�Fig.�
40,� fitolitos� 24�25)� eran� las� plantas� con� fitolitos� más� abundantes� en� el�
ecosistema;� sin� embargo,� las� plantas� mas� escasas,� palmeras� (Gl)� (Fig.� 40,�
fitolitos�1�2),�pastos�con�braquiolitas�(Br)�(Fig.�40,�fitolitos�3�7)�y�otras�plantas�
que� no� pudieron� ser� identificadas� (Otros:� puzzle� y� otros� morfotipos�
indeterminados,� Fig.� 40,� fitolitos� 24�25)� fueron� contrariamente,� las� más�
escogidas�por�los�herbívoros�productores�de�la�bosta�(50�%,�46�%�y�40�%�más�
abundantes�en�la�bosta�que�en�el�suelo,�respectivamente).��
�
�
�
�
�
�
�
�
�
�
�
�
�
�

� ����
;�������������
Esta� Tesis� representa� un� ejemplo� de� las� inferencias� (comportamentales,�
evolutivas,� paleoambientales,� etc.)� que�pueden� ser�extraídas� de�un� icnogénero�
de�trazas�fósiles�de�insectos,������������-�y�como�ellas�pueden�ser�integradas�
en�el�período�de�tiempo�representado�por�la�Formación�Sarmiento�(Eoceno�Medio�
–�Mioceno�Inferior)�en�Gran�Barranca�(Chubut).��
Dichas� inferencias� sólo� fueron� posibles� realizando� primeramente� una�
revisión� icnotaxonómica� actualizada� del� icnogénero,� basada� sobre� 1231�
ejemplares.� Esta� revisión� concluyó� con� la� creación� de� una� icnoespecie� nueva,�
denominada�aqu��� isp.�B,�y�con�cambios�nomenclatoriales� importantes�ya�que�
��� ����������Sauer�1955�pasó�a�ser�un�sinónimo�junior�de������������Roselli�
1939,� mientras� que� ��� �������� Laza� 2006� pasó� a� tener� un� nuevo� nombre,�
denominándosela�aquí�provisoriamente�como����isp.�A.�
Como�segundo�paso�fueron�estudiados�en�el�campo�y/o�en�el�laboratorio,�
el� comportamiento�de�nidificación,� la�estructura�de� los�nidos�y� las�masas/bolas�
de�cría�de�11�especies�actuales�de�escarabajos�coprófagos�pertenecientes�a�5�de�
las�6�tribus�argentinas�(�����������sp.,���������� ������,���� �������
����,�����������,��������,������������ ���,� ����������� �� �����,� ��
������,� ������� spp.,� '� ������ & �����,� $������������ ���� &�,� $��
�����),� con� el� objetivo� de� atribuir� las� 6� icnoespecies� de� ������������ (���
�������,����������,����������� ��,���� ������,���� isp.�A�y���� isp.�B)�a� taxones�
actuales� de� Scarabaeinae.� Se� concluyó� que���� ������� y� ��� ������� podrían� ser�
atribuidas�a�Phanaeini�o�Canthonini,�mientras�que����isp.�B�más�específicamente�
a�alguna�especie�del�género�������.��

� ����
Los� ejemplares� de� ������������ reconocidos� en� estratos� del� Eoceno�
Superior� bajo� de� Gran� Barranca,� estarían� predatando� en� 30� �� 40� millones� de�
años�al�primer�registro�de�Canthonini�y�Phanaeini�fósiles,�que�son�del�Mioceno�de�
Kenia�y�del�Pleistoceno�de�Estados�Unidos�respectivamente.��
La�ausencia�de�������������en� los�niveles�más�bajos�de� la� Formación�
correspondientes�al�Eoceno�Medio�bajo,�y�la�ausencia�de�����������y�����������y�
otras�icnoespecies�en�estratos�que�alcanzan�el�Eoceno�Superior�bajo,�sugiere�que�
la� Patagonia� central� no� habría� sido� demasiado� favorable� para� la� aparición� o�
diversificación�de�los�escarabajos�coprófagos,�debido�probablemente�a�la�escasez�
de�grandes�mamíferos�en�la�Fauna�Barranquense.�Durante�el�Eoceno�Medio�alto�
ocurre� el� primer� registro� de������������,� en� baja� densidad� y� representado�
solamente�por������������y���� �����,� lo�que� sugiere�que� sólo�dos�especies�de�
escarabajos� coprófagos� estarían� presentes� reflejando� las� primeras� condiciones�
ambientales� favorables� para� ellos,� aunque� no� las� óptimas.� Estos� primeros�
escarabajos� coprófagos� probablemente� llegaron� a� Patagonia� desde� localidades�
del�Norte�como�lo�atestigua�la�presencia�de�������������en�el�Eoceno�Inferior�
de� Uruguay.� Esta� etapa� inicial� finaliza� con� la� ausencia� total� del� registro� de� la�
actividad� de� los� escarabajos� coprófagos� durante� un� breve� lapso� de� tiempo�
(alrededor�de�1�ma)�en�el�Eoceno�Medio�alto,�en�el�que�prevalecieron�condiciones�
áridas,�como�lo�indican�las�icnofacies�y�los�paleosuelos.�
Durante�el�Eoceno�Superior�bajo�ocurre�una�“explosión”�en�el�registro�de�
������������reflejada�por�una�gran�abundancia�y�densidad�de�especimenes�de�
todas� las� icnoespecies,� que� documentan� la� máxima� diversidad� de� escarabajos�
coprófagos� alcanzada� en� toda� la� secuencia� (6� a� 9).� Teniendo� en� cuenta� la�
cercana� relación� que� existe� entre� escarabajos� coprófagos,� hábitats� dominados�
por� pastos� y� grandes� mamíferos,� la� mencionada� explosión� en� el� registro� de�

� ���
������������ habría� sido� producida� por� el� establecimiento� de� pastizales� con�
abundancia�de�herbívoros�(Figura�46).�Evidencias�paleontológicas�y�geológicas�de�
diversas� fuentes,� entre� las� que� se� cuentan� el� registro� de� fitolitos� y� el� tipo� de�
paleosuelos,�apoyan�este�resultado�obtenido�del�estudio�de������������.�Este�
cambio� sustancial� del� ecosistema� de� Patagonia� central� tuvo� lugar� junto� con� la�
expansión�de� la� Fauna�Mustersense,�en� la� cual�ha� sido� reconocido�por�primera�
vez�un� significativo� aumento�de� la� hipsodoncia,� principalmente� atribuida� a�una�
dieta�rica�en�gramíneas.��
El� tamaño� promedio� de� las� bolas� de� cría� fósiles� revela� que� la� fauna� de�
escarabajos� coprófagos� estaría� formada� por� muchas� especies� pequeñas.� El�
aumento� en� la� diversificación� no� se� habría� dado� solamente� en� los� coprófagos�
constructores� de� bolas� de� cría� sino� también� en� otros� organismos� relacionados�
con� ellas.� A� partir� del� estudio� de� las� trazas� halladas� en� los� ejemplares� de�
�����������,� es� posible� concluir� que� la� nueva� icnoespecie,� �������� �����
���,� registra� el� comportamiento� cleptoparasítico� de� otros� escarabajos� o�
moscas.�Los�ejemplares�de���� ����������������reconocidos�en�especimenes�de�
�����������,� y� a� su� vez� conectados� con� túneles� meniscados� y� cámaras� de�
estivación� en� el� paleosuelo� circundante,� revelan� que� las� lombrices� se�
comportaban�como�cleoptoparásitos�activos�o�detritívoros�en�las�bolas�de�cría.�La�
asociación� de� �����������,� �������� ����� ���� y� ��� ����� �����������
representa�una�paleocomunidad�de�la�bosta�análoga�a�la�actual,�que�se�registró�
entre�el�Eoceno�Superior�y�el�Oligoceno�en�Patagonia.�
�
�

� ����
�
�!�%�& 83�� Reconstrucción� del� ecosistema� durante� el� Eoceno� Superior� en� Patagonia,�dominado� por� pastos,� con� abundancia� de� herbívoros� y� diversidad� de� escarabajos�coprófagos,� interpretado�a�partir�de� la� “explosión�de������������”.�Creador:�Rosario�Romero.�
�

� ���
Durante� la� Transición� Eoceno�Oligoceno� y� el� Oligoceno� temprano� las�
condiciones� fueron� desfavorables� y� no� se� registraron� �����������.� En� el�
Oligoceno� Superior� bajo,� la� mayoría� de� los� parámetros� estudiados� en�
������������ vuelven� a� mostrar� valores� similares� a� los� registrados� para� el�
Eoceno�Superior�bajo.�La�diversidad�de�potenciales�productores�(6�a�10)�alcanza�
el�máximo�valor�de�toda�la�secuencia,�sugiriendo�una�gran�diversificación�de�los�
escarabajos� coprófagos,� posiblemente� con� las� mismas� especies,� o� con� otras�
nuevas,�ya�que�si�bien�����������,���� isp.�A�y����������� se� registran�durante�
este�tiempo,�����������desaparece.��
La� densidad� y� tamaño� de������������� y� la� diversidad� de� especies� de�
escarabajos�coprófagos�durante�el�Oligoceno�Superior�medio�reflejan�importantes�
cambios�dentro�de�un�hábitat�maduro,�definitivamente�establecido,� tal� como� lo�
sugieren�los�paleosuelos�fuertemente�desarrollados.�El�incremento�de�tamaño�de�
estas� trazas�sugiere� la�aparición�de�grandes�herbívoros,�hecho�corroborado�por�
los� mamíferos� de� la� Fauna� Deseadense.� Esta� fauna� habría� producido� grandes�
cantidades� de� excremento� favoreciendo� la� aparición� de� grandes� escarabajos�
coprófagos� y� la� desaparición� simultánea� de� las� especies�más� pequeñas,� hecho�
sustentado� además� por� la� desaparición� de� ��� ������,� icnoespecie� atribuida� al�
género�������,�conformado�mayormente�por�especies�pequeñas.��
La� diversidad� estimada� de� escarabajos� coprófagos� disminuye� a� 5� en� el�
Oligoceno�Superior�medio.�Más�allá�del�aumento�de�tamaño�de�los�ejemplares�de�
������������y�de�la�aparición�de�mamíferos�más�grandes,�la�disminución�en�la�
densidad�y�diversidad�sugiere�entonces�algún�desmejoraminto�de�las�condiciones�
ambientales.� Hacia� el� Mioceno� Inferior,� se� estabilizaron� las� condiciones�
paleoambientales,�como�se�ve�reflejado�en�la�ausencia�de�cambios�significativos�

� ����
en�la�densidad,� icnodiversidad,�tamaño�y�diversidad�de�especies�de�escarabajos�
coprófagos�(6�a�7).��
Durante� el� Mioceno� Inferior� alto,� se� registra� una� nueva� evidencia� de�
cambios� desfavorables� en� las� condiciones� ambientales� para� los� escarabajos�
coprófagos�en�Gran�Barranca,�la�cual�se�muestra�desde�el�inicio�de�este�periodo�
con�la�desaparición�de�dos�icnoespecies:����������� ���y����������.�El�tamaño�de�
������������no�experimenta�cambios�significativos�desde�aquí�hasta�su�último�
registro,�pero�la�abundancia�se�modifica�y�la�diversidad�de�posibles�productores�
disminuye�de�6�especies�a�4,�posiblemente�en�relación�con�la�desaparición�de�dos�
icnoespecies�vinculada�a�un�ambiente�más�seco,�según�atestiguan�las�litofacies�y�
los�paleosuelos�de�este�tramo�de�la�secuencia.�Estas�características�corresponden�
al�periodo�representado�por�la�Fauna�Colhuehuapense.�
Finalmente,� en� el� Mioceno� Inferior� más� alto,� el� último� registro� de�
ejemplares�de�������������muestra�una�abundancia�y�una�densidad�mínimas,�
similares�a�las�registradas�al�inicio�de�la�secuencia.�Hacia�el�tope�de�la�secuencia�
(fin� del� Mioceno� temprano)� no� se� registran� más� �����������,� sugiriendo�
cambios� ambientales� hacia� condiciones� más� secas� y� pérdida� de� la� cobertura�
vegetal,� tal� como� lo� indican� las� litofacies� y� los� paleosuelos,� que� llevan� a� la�
completa�desaparición�de�los�escarabajos�coprófagos�de�Gran�Barranca.���
El� número� y� diversidad� de� fitolitos� en� ejemplares� de������������� fue�
utilizado� en� esta� Tesis� para� inferir� aspectos� del� comportamiento� de� los�
escarabajos� coprófagos,� la� dieta� de� los� herbívoros� que� producían� los�
excrementos� aprovisionados� y� la� abundancia� de� plantas� con� fitolitos� en� el�
ambiente.�Los�fitolitos�encontrados�en�el�relleno�de�la�mayoría�de�los�ejemplares�
de� ������������ estudiados,� son� similares� a� los� hallados� en� el� paleosuelo�
circundante,� sugiriendo� que� el� relleno� representa� una� muestra� más� del� suelo�

� ����
original�que�fue�introducida�por�el�adulto�al�emerger,�o�más�tarde�por�procesos�
pasivos.� El� número� de� fitolitos� encontrado� en� la� pared� de� la� mayoría� de� los�
ejemplares� de� ������������ es� significativamente� mayor� al� hallado� en� el�
paleosuelo�y�relleno.�Este�resultado�sugiere�que�la�pared�preserva�fitolitos�de�la�
bosta�agregada�a�ella�por�los�constructores�de�las�bolas,�como�se�ha�registrado�
aquí�en�las�especies�actuales.��
El� contenido� fitolítico� de� las� bolas� de� cría� actuales� revela� que� el� mayor�
número� de� fitolitos� en� la� pared� de������������� respecto� de� los� paleosuelos�
asociados,� estaría� evidenciando� paleoambientes� de� pastizales� abiertos� con� una�
cantidad�moderada�de�pastos,�junto�a�palmeras�y�otras�plantas�con�fitolitos.��
Las� comparaciones� de� las� frecuencias� de�morfotipos� de� fitolitos� entre� la�
pared� y� el� paleosuelo� (o� relleno)� en� las������������� donde� se� evidenció� la�
preservación�de�bosta�en�la�pared,�revelan�datos�sobre�la�dieta�de�los�herbívoros.�
Se� concluyó� entonces� que� algunos� herbívoros� habrían� sido� más� generalistas,�
entre� las� plantas� con� fitolitos,� alimentándose�de� la� vegetación�más�abundante,�
palmeras� y� pastos,� mientras� que� otros,� habrían� preferido� vegetación� menos�
frecuente,�alimentándose�de�pastos�con�otro�tipo�de�fitolitos�y�dicotiledóneas.�
�
�
�
�
�
�

� ����
:�����������
�
Ameghino,� F.� 1906.� Les� formations� sédimentaires� du� Crétacé� superieur� et� du�
Tertiaire�de�Patagonie�avec�un�parallèle�entre�leurs�faunes�mammalogiques�
et� celles� de� I'ancien� continent.� Anales� del� Museo� Nacional� de� Historia�
Natural�de�Buenos�Aires,�15:�1�568.��
Andreis,�R.�1972.�Paleosuelos�de�la�Formación�Musters�(Eoceno�Medio),�Laguna�
del�Mate,�Provincia�de�Chubut,�República�Argentina.�Revista�de�la�Sociedad�
Argentina�de�Mineralogía,�Petrografía�y�Sedimentología,�3:�91�97.�
Andreis,� R.� 1981.� Identificación� e� importancia� geológica� de� los� paleosuelos.�
Livro�texto�2.�Univ.�Federal�do�Río�Grande�do�Sul�(ed.),�Porto�Alegre,�67�p.�
Andreis,� R.� R.,� Masón,� M.� M.� y� L.� A.� Spalletti.� 1975.� Estudio� estratigráfico� y�
paleoambiental�de�las�sedimentitas�terciarias�entre�Pico�Salamanca�y�Bahía�
Bustamante,� Provincia� de� Chubut,� República� Argentina.� Revista� de� la�
Asociación�Geológica�Argentina,�30:�85�103.�
Aramayo,�S.�A.,�Schillizzi,�R.�A.�y�B.�Gutiérrez�Téllez.�2004.������������� isp.�
at� the� Irene� Formation� (Early� to� Middle� Pliocene),� Quequén� Salado� river,�
Buenos� Aires� province,� Argentina.� Abstract� Book,� ������ /����������
�����������/ ������&-�$�����0112-�����-�$������:�p.�13.�
Barattini,� L.� P.� y� A.� C.� Sáenz.� 1953.� Nuevos� aportes� para� el� conocimiento� del�
desarrollo� del� )����� ������ (Blanch).� Revista� de� la� Sociedad�
Entomológica�Argentina,�16�(1):�25�30.�
Barbero,�E.,�Palestrini,�C.�y�E.�Monteresino.�1999.�Morfología�preimaginal�y�notas�
de� biología� reproductiva� en� �������� ( ���������)� ������ �
Balthasar,�1939�(Coleoptera:�Scarabaeidae:�Canthonina).�Elytron,�13:�109�
121.�

� ����
Barboni,� D.,� Bonnefille,� R.,� Alexandre,� A.� y� J.� D.�Meunier.� 1999.� Phytoliths� as�
paleoenvironmental� indicators.� West� side� middle� Awash� Valley,� Ethiopia.�
Palaeogeography,�Palaeoclimatology,�Palaeoecology,�152:�87�100.�
Basilewsky,�P.�1951.�Loges�fossiles�de�Coprophages.�Bulletin�et�Annals,�Societe�
Entomologique�d�Belgique,�87�(I�II):�26�27.�
Bellosi,� E.� S.� en� prensa.� Loessic� and� fluvial� pyroclastic� sedimentation� in� the�
Middle� Cenozoic� of� Central� Patagonia� (Sarmiento� Formation).� En� ���
)��������&� ��� ���� *��� 3� '��������� ��� '����������� �����
�������� ��� ����� ������ � ��� )�����,� ed.� Madden,� R.,� Carlini,� A.,�
Vucetich,�M.�G.�y�R,�Kay.�Cambridge�University�Press.�Cambridge,�UK.�
Bellosi,�E.�S.�en�prensa.�Physical�stratigraphy�of�the�Sarmiento�Formation�(Middle�
Eocene� �� Lower� Miocene)� at� Gran� Barranca,� Central� Patagonia.� En� ���
)��������&� ��� ���� *��� 3� '��������� ��� '����������� �����
�������� ��� ����� ������ � ��� )�����,� ed.� Madden,� R.,� Carlini,� A.,�
Vucetich,�M.�G.�y�R,�Kay.�Cambridge�University�Press.�Cambridge,�UK.�
Bellosi,� E.� S.� 2004.� Sedimentologic� control� of� the� ������������ ichnofacies.�
������/���������������������/ ������&-�$�����0112-�����-�$������,�p.�
19.�
Bellosi,� E.� S.� y� J.� F� Genise.� 2004.� Insect� trace� fossils� from� paleosols� of� the�
Sarmiento� Formation� (Middle� Eocene�Lower� Miocene)� at� Gran� Barranca�
(Chubut� Province).� 15–29� en� / ���� 0112�� ������ /������������������ ���
/ ������&���������������������Bellosi,�E.�S.�y�R.�N.�Melchor�(eds).�Museo�
Paleontológico�Egidio�Feruglio,�Trelew,�Argentina,�81�p.��
Bellosi,�E.�S.,�Genise,�J.�F.,�Laza,�J.�H.�y�M.�V.�Sánchez.�2005.�Terrestrial�trace�
fossils� and� ichnostratigraphy� of� the� Sarmiento� Formation:� implications� for�

� ����
the�oldest�grass�dominated�ecosystem.�45/������������6�� ��$�������-�
��)��:�306.�
Bellosi,� E.� S.� y� M.� González,� en� prensa.� Paleosols� of� the� Middle� Cenozoic�
Sarmiento� Formation,� Central� Patagonia.� En� ��� )��������&� ��� ����
*��� 3�'������������'��������������������������� ����������� �
��� )�����,� ed.� Madden,� R.,� Carlini,� A.,� Vucetich,� M.� G.� y� R,� Kay.�
Cambridge�University�Press.�Cambridge,�UK.��
Bellosi,�E.�S.�y�M.�G.�González.�2005.�Paleosuelos�de�la�Formación�Sarmiento�en�
Gran�Barranca� (Chubut,�Argentina).�45/������������6�� ��$�������-���
)��-�4:�433��438.�
Bellosi,�E.�S.,�González,�M.�G.�y�J.�F.�Genise.�2004.�Origen�y�desmantelamiento�
de� lateritas� paleógenas� del� sudoeste� de� Uruguay� (Formación� Asencio).�
Revista�del�Museo�Argentino�de�Ciencias�Naturales,�n.s.�6:�25–40.�
Bellosi,�E.�S.�y�R.�Madden.�2005.�Estratigrafia�física�preliminar�de�las�secuencias�
piroclásticas� terrestres� de� la� Formación�Sarmiento� (Eoceno�Mioceno)�en� la�
Gran� Barranca,� Chubut.� 45/� �������� ���6�� �� $�������-� �� )��-� 4:�
427�432.�
Bellosi,�E.�S.,�Laza,� J.�H.,�Sánchez,�M.�V.�y� J.�F�Genise,�en�prensa.� Ichnofacial�
analysis�of�the�Sarmiento�Formation�(Middle�Eocene�Lower�Miocene)�at�Gran�
Barranca,� Central� Patagonia.� En� ��� )��������&� ��� ���� *��� 3�
'��������� ��� '����������� ����� �������� ��� ����� ������ � ���
)�����,�ed.�Madden,�R.,�Carlini,�A.,�Vucetich,�M.�G.�y�R,�Kay.�Cambridge�
University�Press.�Cambridge,�UK.�
Bellosi,� E.� S.,� Laza,� J.� y� M.� González.� 2001.� Icnofaunas� en� paleosuelos� de� la�
Formación� Sarmiento� (Eoceno�Mioceno),� Patagonia� central.� 5/� 7���6��

� ����
$���������/ �����8�&�//�7���6����/ �����8���� � ����-���������
��0119-��� ��:�-�$������:�p.�31.��
Bellosi,� E.�S.,�Miquel,� S.� E.,�Kay,�R.� F.� y� R.�H.�Madden.� 2002a.�Un�paleosuelo�
mustersense�con�microgastrópodos�terrestres�(Charopidae)�de�la�Formación�
Sarmiento,� Eoceno� de� Patagonia� central:� significado� paleoclimático.�
Ameghiniana.�39:�465�477.�
Bellosi,� E.,� González,� M.,� Kay,� R.� y� R.� Madden.� 2002b.� El� valle� inciso�
colhuehuapense� de� Patagonia� central� (Mioceno� inferior).� /4� 7���6��
$�������������������8-��6����-�49�p.�
Bertoldi� de� Pomar,� H.� 1971.� Ensayo� de� clasificación� morfológica� de� los�
silicofitolitos.�Ameghiniana,�8:�317�328.�
Bertoldi� de� Pomar,� H.� 1975.� Los� silicofitolitos:� sinopsis� de� su� conocimiento.�
Darwiniana,�19:�173�206.�
Bohaty,�S.�M.�y�J.�C.�Zachos.�2003.�Significant�Southern�Ocean�warming�event�in�
the�late�middle�Eocene.�Geology,�31:�1017–1020.�
Bornemissza,�G.�F.�1969.�A�new�type�of�brood�care�observed�in�the�dung�beetle�
����� �������� ����(Scarabaeidae).�Pedobiología,�9:�223�225.�
Bornemissza,� G.� F.� 1976.� The� Australian� dung� beetle� proyect� 1965�1975.�
Australian�Meat�Research�Committee�Review,�30:�1�32.�
Borrelli,� N.,� Osterrieth,� M.� y� M.� Cabello.� 2005.� Degradacion� de� fitolitos� de�
gramíneas.� Experimentacion� in� vitro.� ///� '� ������ �� /������ �����
�����8�� �������������-��� ��:�-�$������-�p.�73.�
Bown,� T.� M.� y� J.� G.� Fleagle.� 1993.� Systematics,� bioestratigraphy� and� dental�
evolution� of� the� Palaeothentidae,� Later� Oligocene� to� Early�Middle� Miocene�
(Deseadan�Santacrucian)� Caenolestoid� Marsupials� of� South� America.�
Paleontological�Society�Memoir,�29:�1�76.�

� ����
Bown,�T.�M.,�Hasiotis,�S.�T.,�Genise,�J.�F.,�Maldonado,�F.�y�E.�M.�Brouwers.�1997.�
Trace� fossils� of� Hymenoptera� and� other� insects� and�paleoinveronments� of�
the� Claron� Formation� (Paleocene� and� Eocene),� Southwestern� Utah.� U.S.�
Geological�Survey�Bulletin,�2153:�42�58.��
Bromley,�R.�G.,�Buatois,�L.�A.,�Genise,�J.�F.,�Labandeira,�C.�G.,�Mángano,�M.�G.,�
Melchor,� R.� N.,� Schlirf,� M.� y� A.� Uchman.� 2007.� Comments� on� the� paper�
"Reconnaissance� of� Upper� Jurassic� Morrison� Formation� ichnofossils,� Rocky�
Mountain�Region,�USA:�Paleoenviromnental,�stratigraphic,�and�paleoclimatic�
significance� of� terrestrian� and� freshwater� ichnocoenoses"� by� Stephen� T.�
Hasiotis.�Sedimentary�Geology�200:�141–150.�
Bruet,� E.� 1950.� Le� loess� de� la� République� del�Equateur� et� ses� nids� fossiles�
d�insects.�Revue��Francaise�d�Entomology,�17�(4):�280�283.�
Brussaard,�L.�1987.�Kleptocopry�of�$�������� ������� (Coleoptera,�Aphodiidae)�
in�nests�of��&����� �&������ (Coleoptera,�Geotrupidae)�and� its�effect�on�
soil�morphology.�Biology�and�Fertility�of�Soils,�3�(1�2):�117�119.�
Buatois,� L.� A.� y� M.� G.� Mángano.� 2004.� Animal�substrate� interactions� in�
freshwater� environments:� applications� of� ichnology� in� facies� and� sequence�
stratigraphic� analysis� of� fluvio�lacustrine� successions.� 311–333.� En� ���
$���� ��������/ ������&����)�������������������������� �$��&���.�
D.�McIlroy�(ed.).�London:�Geological�Society.�
Buatois,� L.� A.� y� M.� G.� Mángano.� 2007.� Invertebrate� ichnology� of� continental�
freshwater�environments.�285–�323.�En��� ������������� ���-�)������-�
)���� ����W.�Millar�(ed.).�Amsterdam:�Elsevier.�
Buatois,� L.� A.,� Mángano,� M.� G.� y� G.� Aceñolaza.� 2002.� Trazas� fósiles.� Museo�
Paleontológico�Egidio�Feruglio.�Trelew.�Edición�Especial�MEF�Nº�2.�

� ���
Buatois,� L.� A.,� Mánagno,� M.� G.� y� J.� F.� Genise.� 2000.� Ichnofacies� models� in�
continental� environments.� ;9��� /���������� ������ �� �������-� 78�� ��
<����-�*����-�$������0111�[CD�ROM].�
Buatois,�L.�A.,�Mángano,�M.�G.,�Genise,�J.�F.�y�T.�N.�Taylor.�1998.�The�ichnologic�
record� of� the� continental� invertebrate� invasión:� evolutionary� trends�
environmental� expansion,� ecospace� utilization,� and� behavioral� complexity.�
Palaios,�13:�217�240.�
Bullock,�P.,�A.�Federoff,�A.�Jongerius,�G.�Stoops,�y�T.�Tursina.�1985.�Handbook�
for� soil� thin� section� description.� Waine� Research� Publications,� Albrighton.�
152�p.�
Cabrera�Walsh,� G.� y� G.� Gandolfo.� 1996.� Nidification� of� thirteen� common�
Argentine� dung� beetles� (Scarabaeidae:� Scarabaeinae).� Annals� of�
Entomological�Society�of�America,�89�(4):�581�588.�
Camacho,�H.�H.�1966.�Invertebratos�Fósiles.�Buenos�Aires:�Eudebra.�XVI+707p.�
Cambefort,� Y.� 1984.� Étude� écologique� des� Coléoptères� Scarabaeidae� de� Cóte�
d´Ivoire.�Travaux�Cherche�Lamto,�3:�1�294.�
Cambefort,�Y.�1991.�Dung�beetles�in�tropical�savannas.�157�177�en�(����*���
' ����&.� Hanski,� I.� y� Y.� Cambefort� (eds.).� Princeton� University� Press,�
Princeton,�N.�J.,�481�p.�
Cambefort,�Y,�y�I.�Hanski.�1991.�Dung�beetle�population�biology.�36�50�en�(����
*���' ����&.�Hanski,�I.�y�Y.�Cambefort�(eds.).�Princeton�University�Press,�
Princeton,�N.�J.,�481�p.�
Chin,�K.�2007.�The�paleobiological�implications�of�herbivorous�dinosaur�coprolites�
from� the� Upper� Cretaceous� Two�Medicine� Formation� of�Montana:�why� eat�
wood?.�Palaios,�22:�554�566.�

� ���
Chin,�K.�y�B.�D.�Gill.�1996.�Dinosaurs,�dung�beetles�and�conifers:�participants�in�
a�Cretaceus�food�web.�Palaios,�11:�280�285.�
Chure,�D.� J.,� Litwin,� R.,�Hasiotis,� S.� T.,� Evanoff,� E.� y�K.�Carpenter.� 2006.� The�
fauna� and� flora� of� the�Morrison� Formation:� 2006.�New�Mexico�Museum�of�
Natural�History�and�Science�Bulletin,�36:�233–249.�
Cifelli,� R.� L.� 1985.� Biostratigraphy� of� the� Casamayoran,� early� Eocene,� of�
Patagonia.�American�Museum�Novitates,�2820:�1�26.�
Comisión�Internacional�de�Nomenclatura�Zoológica.�2008.�OPINION�2211�(Case�
3360).� The� ichnogenus� ������������ Sauer,� 1955� (Ichnotaxa,�
Coprinisphaeridae):� generic� name� given� precedence� over� ������� Roselli,�
1939.�Bulletin�of�Zoological�Nomenclatura,�65�(4):�318�319.�
Cosarinsky,�M.�I.,�Bellosi,�E.�S.�y�J.�F.�Genise.�2005.�Micromorphology�of�modern�
epigean� termite� nests� and� possible� termite� ichnofossils:� a� comparative�
analysis�(Isoptera).�Sociobiology,�45:�1�34.�
De,�C.�2002.�Continental�mayfly�burrows�within�relict�ground�in�inter�tidal�beach�
profile� of� Bay� of� Bengal� coast:� A� new� ichnological� evidence� of� Holocene�
marine�transgression.�Current�Science,�83:�64–67.�
De,� C.� 2005.� Quaternary� ichnofacies� model� for� paleoenvironmental� and�
paleosealevel� interpretations:�a�study�from�the�Banas�River�Basin,�western�
India.�Journal�of�Asian�Earch�Sciences�25:�233�249.�
Dieni,� I.�y�J.�Genise.�2004a.� Il�primo�ritrovamento�[europeo]�di�Coprinisphaera�
Sauer� 1955� (nidi� fossili� di� Scarabei� stercorari)� nell'Eocene� dell'Italia� NE.�
����������)����������0112-�*�����-�09#0;�� ����-�7���������'�� ��
)�� �����3�p.�29.�

� ��
Dieni,� L.� y� J.� F.� Genise.� 2004b.� The� first� European� record� of� ������������
Sauer� 1955.� ������ /���������� �������� ��� / ������&-� 9=>0;?2?0112-�
����-�$������-�p.�31.�
Donovan,� S.� K.� 1994.� Insects� and� other� arthropods� as� trace�makers� in� non�
marine�environments�and�paleoenvironments.�200–220�en����)��������&�
����� ��������.�S.�K.�Donovan�(ed.).�Chichester:�Wiley.�
Duringer,�P.,�Brunet,�M.,�Cambefort,�Y.,�Veauvilain,�A.,�Mackaye,�H.T.,�Vignaud,�
P.� y�M.� Schuster.� 2000a.� Des� boules� de� bousiers� fossiles� et� leurs� terriers�
dans� les� sites� a� Australopitheques� du� Pliocene� tchadien.� Bulletin� de� la�
Societé�Géologique�de�France,�171:�259�269.�
Duringer,�P.,�Brunet,�M.,�Cambefort,�Y.,�Likius,�A.,�Mackaye,�H.T.,�Schuster,�M.�y�
P.� Vignaud.� 2000b.� First� discovery� of� fossil� dung� beetle� brood� balls� and�
nests� in� the� Chadian� Pliocene� Australopithecine� levels.� Lethaia,� 33:� 277�
284.�
Duringer,� P.,� Schuster,� M.,� Genise,� J.� F.,� Mackaye,� H.� T.,� Vignaud,� P.� y� M.�
Brunet.�2007.�New�termite�trace�fossils:�galleries,�nests�and�fungus�combs�
from� the� Chad� basin� of� Africa� (Upper� Miocene–Lower� Pliocene).�
Palaeogeography,�Palaeoclimatology,�Palaeoecology,�251:�323–353.�
Edmonds.� W.� D.� 2000.� Revision� of� the� Neotropical� dung� beetles� genus�
Sulcophanaeus� (Coleoptera:� Scarabaeidae:� Scarabaeinae).� Folia�
Heyrovskyana,�Suppl.�6:�62�p.��
Ekdale,�A.�A.,�Bromley,�R.�G.�y�D.�B.�Loope.�2007.�Ichnofacies�of�an�ancient�erg:�
a� climatically� influenced� trace� fossil� association� in� the� Jurassic� Navajo�
Sandstone,� Southern� Utah,� USA.� 562–576� en� �� � ��������� ��� ���-�
)������-�)���� ��.�W.�Miller�(ed.).�Amsterdam:�Elsevier.�

� ���
Ellis,� W.� E.� y� A.� C.� Ellis�Adam.� 1993.� Fossil� brood� cells� of� solitary� bees� from�
Fuerteventura� and� Lanzarote,� Canary� Islannds� (Hymenoptera:� Apoidea).�
Entomologische�Berichten,�53:�161�173.�
Estrada,�A.�1941.�Contribución�geológica�para�el�conocimiento�de�la�“cangagua”�
de� la� región� interandina� y� del� cuaternario� en� general� del� Ecuador.� Anales�
Universidad�Central,�Quito,�Ecuador,�66�(312):�405�488.�����
Evans,� H.� E.� y� M.� J.� Eberhard.� 1970.� The� wasps.� The� University� of� Michigan�
Press,�Ann�Arbor,�265�p.�
Fabre,�J.�H.�1899.�Souvenirs�Entomologiques.�VI.�Paris:�Librarie�Delagrave.�
Favila,� M.� E.� 1993.� Some� ecological� factors� affecting� the� life�style� of��������
&������ &������(Coleoptera:�Scarabaeidae):�an�experimental�approach.�
Ethology,�Ecology�and�Evolution,�5:�319�328.�
Favila,� M.� E.� 2001.� Historia� de� vida� y� comportamiento� de� un� escarabajo�
necrófago:�������� &������ &������LeConte�(Coleptera:�Scarabaeinae).�
Folia�Entomológica�mexicana,�40�(2):�245�278.�
Favila,�M.�E.�y�A.�Díaz.�1996.�������� &������ &������LeConte�(Coleoptera;�
Scarabaeidae)� makes� a� nest� in� the� field� with� several� brood� balls.� The�
Colepterists�Bulletin,�50�(1):�52�60.��
Fernández� Honaine,� M.,� Zucol,� A.� F.� y� M.� L.� Osterrieth.� 2006.� Phytolith�
assemblages� and� systematic� associations� in� grasslands� species� of� South�
Eastern�Pampean�plains,�Argentina.�Annals�of�Botany,�98:�1155�1165.�
Ferrar,� P.�1980.�Cocoon� formation�by�Muscidae� (Diptera).� Journal�of�Australian�
Entomology�Society,�19:�171�174.�
Feruglio,�E.�1949.�Descripción�geológica�de�la�Patagonia.�Yacimientos�Petrolíferos�
Fiscales,�1:�1�334.��

� ���
Fincher,�G.�T.,�Bonner�Stewart,�T.�y�R.�Davis.�1970.�Attraction�of�coprophagous�
beetles� to� feces� of� various� animals.� Journal� of� Parasitology,� 56� (2):� 378�
383.�
Flynn,�F.�J.�y�C.�C.�Swisher.�1995.�Cenozoic�South�American�Land�mammal�ages:�
correlation� to� global� geochronologies.� En� �� �������&� ���� � ��� ���
���������������� �����������.�Berggren,�W.,�Kent,�D.,�Aubry,�M.�P.�y�J.�
Hardenbol�(eds.).�Society�of�Stratigraphic�Geology,�54:�317�333.�
Fontaine,� J.,� Ballesteros,� J.� M.� y� J.� E.� Powell.� 1995.� Artefactos� del�
comportamiento� de� escarabajos� (Coleoptera,� Scarabaeidae)� como�
evidencias� paleoclimáticas� y� paleoambientales� en� el� Cuaternario�
(Pleistoceno�superior)�del�valle�de�Tafí,�provincia�de�Tucumán,�Argentina.�5�
7���6�� @ ����-� $�� � �6�� $������� �� ��� ��� ��� ������������:�
12.�
Fossa�Mancini,�E.�1941.�Noticias�sobre�hallazgos�de�insectos�fósiles�en�la�América�
del�Sur.�Notas�Museo�de�La�Plata,�Paleontología,�6�(29):�101�140.�
Fraysse,� F.,� Cantais,� F.,� Pokrovsky,� O.� S.,� Schott,� J.� y� L.� D.� � Meunier.� 2006.�
Aqueous� reactivity� of� phytoliths� and� plant� litter:� Physico�chemical�
constraints� on� terrestrial� biogeochemical� cycle� of� silicon.� Journal� of�
Geochemical�Exploration,�88:�202�205.�
Frenguelli,� J.� 1929.� Apuntes� de� Geología� Patagónica.� Descripción� de� algunos�
perfiles� de� la� zona� petrolífera� de� Comodoro� Rivadavia.� Boletín� de�
Informaciones�Petrolíferas,�YPF,�Buenos�Aires,�6:�575�606.��
Frenguelli,� J.� 1938a.�Nidi� fossili� di� Scarabeidi� e�Vespidi.� Bolletino�de� la�Societá�
Geologica�Italiana,�57:�77�96.��
Frenguelli,� J.�1938b.�Bolas�de�escarabeidos�y�nidos�de�véspidos� fósiles.�Physis,�
12�(44):�348�351.�

� ���
Frenguelli,� J.� 1939a.� Sobre�nidos� fósiles� de� insectos� del�Neuquén�y�Río�Negro.�
(Resumen).�Revista�Sociedad�Entomológica�Argentina,�10�(2):�270.�
Frenguelli,�J.�1939b.�Nidos�fósiles�de�insectos�en�el�Terciario�del�Neuquén�y�Río�
Negro.�Notas�Museo�La�Plata,�Paleontología,�4�(18):�379�402.�
Frenguelli,� J.� 1940.� Viaje� a� las� zonas� central� y� andina� de� la� Patagonia�
septentrional.�Revista�Museo�La�Plata,�Sección�Oficial,�1939:�90�106.�
Frenguelli,�J.�1941.�Viaje�a�los�territorios�patagónicos�del�Neuquen�y�del�Chubut.�
Revista�Museo�La�Plata,�Sección�Oficial,�1940:�80�91.��
Gallego,� L.� y� R.� A.� Distel.� 2004.� Phytolith� assemblages� in� grasses� native� to�
Central�Argentina.�Annals�of�Botany,�94:�1�10.�
Genise,�J.�F.�1993.�Trazas�fósiles�de�insectos�en�paleosuelos��R.�N.�Melchor�(ed.).�
Notas�del�curso:�A@�������� ����������������������6����B������
7��-���)��,�p.�49�59.�
Genise,� J.� F.� 1999.� Paleoicnología� de� insectos.� Revista� de� la� Sociedad�
Entomológica�Argentina,�58:�104�116.�
Genise,� J.� F.� 2000a.� Nidos� fósiles� de� insectos� en� paleosuelos.� //� ��������
�������� ��� �� ����������8-� 5///� 7���6�� $������� ��
����������8,� Curso� de� Actualización.� / �����83� ��� ����� �� ��
�����8� ��������� &� �� ��������� ������.� Buatois,� L.� A.,� Genise,� J.,�
Mángano,�M.�G.,�Muñoz,�N.,�Netto,�R.�G.�y�D.�G.�Poiré�(eds.).�Mar�del�Plata,�
Argentina,�p.�46–60.�
Genise,�J.�F.�2000b.�Insect�paleoichnology�in�South�America:�past�and�present.�/�
����6���� *�������� �� )�������������8-� /� ����6���� ������ ��� ��
)�������������8-� /� /���������� ����� ��� )��������������&-�
7����C��)����#��)-�*����-�;#D?=?0111,�p.�28�29.�

� ���
Genise,� J.� F.� 2000c.� Insect� fossil� nests.� /� ����6���� *�������� ��
)�������������8-� /� ��������� ������ ��� �� )�������������8-� /�
/���������� ��������)��������������&-�7����C��)����#��)-�*�����;#
D?=?0111,�p.�114–115.�
Genise,� J.� F.� 2003.� The� palaeontological� significance� of� insect� trace� fossils� in�
palaeosols.�FossilsX3.�)����������$���� ��-�///�/������������������
���)����������&������//�/���������� ��������)��������������&�
���//�E����������������$�����������/� �������-�F>99?0?�011;-�)�����-�
������$��� ,�p.�18–19.�
Genise,� J.� F.� 2004.� Ichnotaxonomy� and� ichnostratigraphy� of� chambered� trace�
fossils� in�palaeosols�attributed�to�coleopterans,�termites�and�ants.�419�453�
en� ��� ���� ����� ��� � ������&� ��� �������������� ��� ���������� �
��&���.�McIlroy,�D.�(ed.).�London,�Geological�Society.�
Genise,� J.� F.� 2007.� Icnoentomología:� el� estudio� integrado�de� trazas� actuales� y�
fósiles� de� insectos.� 5� 7���6�� $������� �� / �����8� &� ///� 7���6�� ��
/ �����8���� � ������G����-��������������-�$������:�p.�12.�
Genise,�J.�F.�y�E.�S.�Bellosi.�2004.�Continental�trace�fossils�of�the�Laguna�Palacios�
Formation� (Upper� Cretaceous)� from� the� San� Bernardo� Range� (Chubut�
Province).�33–43�en�/ ����0112-�������/���������������������/ ������&-�
�����������������.�E.S.�Bellosi�y�R.�N.�Melchor�(eds.).�Trelew,�Argentina:�
Museo�Paleontológico�Egidio�Feruglio.�
Genise,� J.� F.,� Bellosi,� E.� S.� y� M.� G.� González.� 2004a.� An� approach� to� the�
description�and�interpretation�of�ichnofabrics�in�palaeosols.�355�382��en����
$���� ��������/ ������&����)�������������������������� �$��&���.�
D.�McIlroy�(ed.),�Geological�Society�of�London.�

� ���
Genise,� J.� F.� y� T.�M.�Bown.� 1994.�New�Miocene� scarabeid� and�hymenopterous�
nests,� and� Early� Miocene� (Santacrucian)� paleoenvironments,� Patagonian�
Argentina.�Ichnos,�3:�107�117.�
Genise,� J.� F.� y� G.� Cladera.� 1995.� Application� of� computarized� tomography� to�
study�insect�traces.�Ichnos,�4:�77�81.�
Genise,� J.� F.� y� G.� Cladera.� 2004.� ������������� ��������� and� other� wasp�
trace� fossils:� breaking� through� the� taphonomic� barrier.� Journal� of� the�
Kansas�Entomological�Society,�77�(4):�626�638.�
Genise,� J.� F.,� Cladera,� G.� y� S.� Tancroff.� 2001.� La� presencia� de� Eatonichnus�
claronensis�en�el�Paleoceno�del�Chubut�(Argentina).�/5�7���6��$���������
/ �����8� &� ������ 7���6�� �� / �����8� ��� � ����-� ������� ��
0119-��� ���-�$������,�p.�45.�
Genise,�J.�F.�y�J.�H.�Laza.�1998.� ���� �������������Roselli:�a�complex�insect�
trace�fossil�produced�by�two�distinct�trace�makers.�Ichnos,�5:�213�223.�
Genise,� J.� F.,� Laza,� J.� H.,� Fernández,�W.� y� J.� Frogoni.� 2002.� Cámaras� pupales�
fósiles� de� coleópteros:� el� icnogénero� 7������ ����� Roselli.� Revista� del�
Museo�Argentino�de�Ciencias�Naturales,�4:�159�165.�
Genise,� J.� F,� Laza,� J.� H.� y� A.� K.� Rindsberg.� 2006.�Case� 3360.� The� ichnogenus�
������������ Sauer,� 1955� (Ichnotaxa,� Insecta, Coleoptera,�
Coprinisphaeridae):� proposed� conservation.� Bulletin� of� Zoological�
Nomenclature,�63�(4):�243�246.�
Genise,� J.� F,� Laza,� J.� H.� y� A.�K.� Rindsberg.� 2008.�Comments� on� the� proposed�
conservation� of� the� ichnogenus� ������������ Sauer,� 1955� (Ichnotaxa,�
Insecta,�Coleoptera,�Coprinisphaeridae)�(2)�(Case�3360;�see�BZN�63:�243–
246).�Bulletin�of�Zoological�Nomenclature,�65�(1):�54.�

� ���
Genise,�J.�F.,�Mángano,�M.�G.�y�L.�A.�Buatois.�2004b.� Ichnology�moving�out�of�
the�water:� a�model� for� terrestrial� ichnofacies.������� /���������� ��������
���/ ������&-�$�����0112-�����-�$������:�p.�38.�
Genise,�J.�F.,�Mángano,�M.�G.,�Buatois,�L.�A.,�Laza,�J.�H.�y�M.�Verde.�2000.�Insect�
trace� fossil� associations� in� palaeosols:� The� Coprinisphaera� ichnofacies.�
Palaios,�15:�49�64.�
Genise,� J.� F.,� Melchor,� R.� N.,� Bellosi,� E.� S.,� Sanchez,� M.� V.,� Krause,� M.� J.,�
Sarzetti,�L.,�Verde,�M.�y�E.�Bedatou.�2008.�Evolution�of�Mesozoic�Cenozoic�
terrestrial� ichnofaunas� from� southern� South� America.� //� /����������
�����������/ ������&-�)�����-�p.�41.��
González,�M.�1999.�Los�paleosuelos�de� la�Formación�Laguna�Palacios�(Cretácico�
Superior)�de�Patagonia�y�la�Formación�Asencio�(Cretácico�Superior�Terciario�
Inferior)�de�Uruguay.�5�����6��������������: �����*����,�p.�65�70.�
González,�M.,�Tófalo,�O.�R.�y�P.�Pazos.�1998.�Icnología�y�paleosuelos�del�Miembro�
Del�Palacio�de� la� Formación�Asencio� (Cretácico�Superior–Terciario� Inferior)�
del�Uruguay.�//���������G����&����������-�)�������'��-�p.�38–42.�
González�Megías,�A.� y� F.�Sánchez�Piñero.� 2003.� Effects� of� brood�parasitism�on�
host� reproductive� success:� evidence� from� larval� interactions� among� dung�
beetles.�Oecologia,�134�(2):�195�202.�
Grimaldi,�D.�y�M.�S.�Engel.�2005.�Evolution�of�the�Insects.�Cambridge�University�
Press.�
Halffter,�G.�1959.�Etología�y�paleontología�de�Scarabaeinae.�Ciencia,�19:�165�78.�
Halffter,� G.� 1977.� Evolution� of� nidification� in� the� Scarabaeinae� (Coleoptera:�
Scarabaeidae).�Quaestiones�Entomologicae,�13:�231�253.�

� ���
Halffter,�G.�1991.�Historical�and�ecological� factors�determining�the�geographical�
distribution� of� beetles� (Coleoptera:� Scarabaeidae:� Scarabaeinae).� Folia�
Entomológica�Mexicana,�82:�195�238.�
Halffter,� G.� y� W.� D.� Edmonds.� 1982.� ��� ������� �������� ��� ����� �����
!� ����"�� $�� ����� �� ��� �������� ���� ��� Editorial� Galache,�
Instituto�de�Ecología,�México,�D.�F.,�176�p.�
Halffter,�G.�y�E.�G.�Matthews.�1966.�The�Natural�History�of�dung�beetles�of�the�
subfamily� Scarabaeinae� (Coleoptera:� Scarabaeidae).� Folia� Entomológica�
Mexicana,�12�14:�1�312.�
Halffter,�G.,�Halffter,�V.�y�C.�Huerta.�1983.�Comportement�sexuel�et�nidification�
chez�������� &������ ������� LeConte� (Col.�Scarabaeidae).�Bulletin�de�
la�Société�entomologique�de�France,�88:�585�594.�
Häntzschel,�W.�1962.�Trace�fossils�and�problemática�en����������/��������
)��������&�� Part�Miscellanea,�W.�Moore,�R.C.� (ed.),�Geological�Survey�of�
America�and�Kansas�University�Press,�W171�W245.�
Häntzschel,�W.� 1975.� Trace� fossils� and� problematica.� Treatise� on� Invertebrate�
Paleontology�W�Supplement�1:�W1–W269.�
Hanski,� I.� 1991.� The� dung� insect� community.� 5�21� en� (���� *��� ' ����&.�
Hanski,�I.�y�Y.�Cambefort�(eds.),�Princeton�University�Press,�Princeton,�N.�J.,�
481�p.�
Hanski,� I.� y� Y.� Cambefort.� 1991.� (���� *��� ' ����&.� Princeton� University�
Press,�Princeton,�N.�J.,�481�p.��
Häntzschel,�W.�1975.�Trace�fossils�and�problematica�en����������/��������
)��������&.� Part� Miscellanea,� W.� Supplement� 1.� Teichert,� C.� (ed.),�
Geological�Survey�of�America�and�University�of��Kansas�Press,�W269.�

� ��
Hasiotis,� S.� T.� 2002.�Continental� Trace� Fossils.� SEPM� (Society� for�Sedimentary�
Geology)�Short�Course�Notes,�51�(ii),�132�p.�
Hasiotis,� S.� T.� 2004.� Reconnaissance� of� Upper� Jurassic� Morrison� Formation�
ichnofossils,� Rocky� Mountain� Region,� USA:� paleoenvironmental,�
stratigraphic,� and� paleoclimatic� significance� of� terrestrial� and� freshwater�
ichnocoenoses.�Sedimentary�Geology,�167:�177–268.�
Hasiotis,� S.� T.� 2006.� [Book� review� of:]� The� Application� of� Ichnology� to�
Palaeoenvironmental�and�Stratigraphic�Analysis.�D.�McIlroy�(ed.),�2004,�The�
Geological�Society�of�London�Special�Publication�228,�490�p.� [...].�Palaios,�
21:�401–402.�
Hasiotis,�S.�T.�2007.�Continental�ichnology:�fundamental�processes�and�controls�
on�trace� fossil�distribution.�262–278�en��� ������������� ���-�)������-�
)���� ��.�W.�Miller�(ed.).�Amsterdam:�Elsevier.�
Hasiotis,� S.� T.,� Kraus,� M.� J.� y� T.� M.� Demko.� 2007.� Climatic� controls� on�
continental� trace� fossils.� 172–195� en� �� � ��������� ��� ���-� )������-�
)���� ����W.�Miller�(ed.).�Amsterdam:�Elsevier.�
Hasiotis,� S.� T.� y� M.� C.� Bourke.� 2006.� Continental� trace� fossils� and� museum�
exhibits:� displaying� organism� behaviour� frozen� in� time.� The� Geological�
Curator,�8:�211–226.�
Heinrich,�B.�y�G.�A.�Bartholomew�1979.�Roles�of�endothermy�and�size� in� inter��
and� intraspecific�competition�for�elephant�dung� in�an�African�dung�beetles,�
� ���������������.�Physiological�Zoology,�52:�484�496.�
Hasiotis,� S.� T.,� Bown,� T.� M.� y� C.� Abston.� 1994.� Photoglossary� of� marine� and�
continental�ichnofossils.�US.�Geological�Survey�Digital�Data�Series,�23.�[CD�
ROM].��

� ����
Hembree,� D.� L.� y� S.� T.� Hasiotis.� 2004.� Using� paleosols� and� ichnofossils� to�
interpret� the� changing� paleoecology,� paleoenvironments,� and� paleoclimate�
of� the� Eocene–Oligocene� White� River� Formation,� northeastern� Colorado.�
Geological�Society�of�America�Abstracts�with�Programme,�36�(5):�63.�
Hendriksen,�N.�B.�1991.�Consumption�and�utilization�of�dung�by�detritivorous�and�
geophagous�earthworms�in�a�Danish�pasture.�Pedobiologia,�35�(2):�65�70.�
Holter,�P.1979.�Effect�of�dung�beetles�($��������spp.)�and�earthworms�on�the�
disappearance�of�cattle�dung.�Oikos,�32�(3):�393�40.�
Horrocks,�M.� y� G.� J.� Irwin.� 2003.� Pollen,� Phytoliths� and�Diatoms� in� Prehistoric�
Coprolites� from� Kohika,� Bay� of� Plenty,� New� Zealand.� Journal� of�
Archaeological�Science,�30:�13–20.�
Horrocks,�M.,�Jones,�M.�D.,�Beever,�R.�E.�y�D.�G.�Sutton.�2002.�Analysis�of�plant�
microfossils� in� prehistoric� coprolites� from� Harataonga� Bay,� Great� Barrier�
Island,� New� Zealand.� Journal� of� The� Royal� Society� of� New� Zealand,� 32:�
617–628.�
Horrocks,�M.,�Salter,�J.,�Braggin,�J.,�Nichol,�S.,�Moorhouse,�R.�y�G.�Elliott.�2008.�
Plant�microfossil� analysis� of� coprolites� of� the� critically� endangered� kakapo�
(��������� ����������)� parrot� from� New� Zealand.� Review� of� Palaeobotany�
and�Palynology,�149:�229–245.�
Houston,� T.� F.� 1987.� Fossil� brood� cells� of� stenotritid� bee� (Hymenopotera,�
Apoidea)�from�the�Pleoistocene�of�South�Australia.�Transactions�of�the�Royal�
Society�of�South�Australia,�3:�93�97.���
Hughes,�R.�D.�1975.�Assessment�of�the�burial�of�cattle�dung�by�Australian�dung�
beetles.�Journal�of�the�Australian�Entomological�Society,�14:�129�134.�
Hunt,�A.�P.�y�S.�G.�Lucas.�2007.�Tetrapod�ichnofacies:�A�new�paradigm.�Ichnos,�
14:�59–68.�

� ���
Iriondo,� M.� y� D.� M.� Kröhling.� 1996.� Los� sedimentos� eolicos� del� noreste� de� la�
Llanura� Pampeana� (Cuaternario� superior).� 4///� �������� ���6�� ��
$��������&�///�����������'+���� �6����,���� ������-�4:�27–48.�
Janis,�C.�M.,�Damuth,�J.�y�J.�M.�Theodor.�2000.�Miocene�ungulates�and�terrestrial�
primary�productivity:�where�have�all�the�browsers�gone?�Proceedings�of�the�
National� Academy� of� Sciences� of� the� Unites� States� of� America,� 97� (14):�
237–�261.�
Janssens,� A.� 1939.� Coprini� (Coleoptera� Lamellicornia).� Exploration� du� Parc�
National�Albert.�Misión�G.�F.�de�Witte,�29:�1�104.�
Jarzembowski,�E.�A.�y�A.�J.�Ross.�1996.�Insect�origination�and�extinction� in�the�
Phanerozoic�/��M.�B.�Hart,�ed.�Biotic�recovery�from�mass�extinction�events.�
Geological�Society,�102:�65�78.�
Joseph,� H.� C.� 1929.� )������� ���������� Eschsch.� Revista� Chilena� de� Historia�
Natural,�33:�31�46.��
Judulien,� F.� 1899.�Quelques� notes� sur� plusieurs� coprophages� de�Buenos�Aires.�
Revista�del�Museo�de�la�Plata�(Argentina),�9:�371�380.�
Kay,�R.�F.,�Madden,�R.�H.,�Vucetich,�M.�G.,�Carlini,�A.�A.,�Mazzoni,�M.�M.,�Re,�G.�
H.,� Heizler,� M.� y� H.� Sandeman.� 1999.� Revised� geochronology� of� the�
Casamayoran� South� American� Land� Mammal� Age:� Climatic� and� biotic�
implications.� Proceedings� of� the� National� Academy� of� Science,� 96(23):�
13235�13240.�
Kingston,�J.�D.�y�T.�Harrison.�2007.�Isotopic�dietary�reconstructions�of�Pliocene�
herbivores� at� Laetoli:� Implications� for� early� hominin� paleoecology.�
Palaeogeography,�Palaeoclimatology,�Palaeoecology,�243:�272–306.�
Kingston,�T.�J.�y�M.�Coe.�1977.�The�biology�of�a�giant�dung�beetles�(,��� ������
�������)�(Coleoptera:�Scarabaeidae).�Journal�of�Zoology,�181:�243�263.�

� ����
Kitching,� J.� W.� 1980.� On� some� fossil� Arthropoda� from� the� Limeworks,�
Makapansgat,�Potgietersrus.�Palaeontologia�Africana,�23:�63�68.�
Klemperer,� H.� G.� 1978.� The� repair� of� larval� cells� and� other� larval� activities� in�
Geotrupes���������Marchall�and�other�species�(Coleoptera:�Scarabaeidae).�
Ecological�Entomology,�3:�119�131.�
Klemperer,�H.�G.�1980.�Kleptoparasitic�behaviour�of�$���������������(L.)� larvae�
in�nests�of�����������������Marsh.�(Coleoptera,�Scarabaeidae).�Ecological�
Entomology,�5:�143�151.��
Klemperer,� H.� G.� 1983.� Brood� ball� construction� by� the� non�brooding� Coprini�
��� ������� ����+� and� (� ��������� ���������� (Coleoptera,�
Scarabaeidae).�Ecological�Entomology,�8:�61�68.�
Klemperer,�H.�G.�y�R.�Boulton.�1976.�Brood�burrow�construction�and�brood�care�
by� ,��� ������ H����� (Klug)� and� ,��� ������ ����&�� (Fabricius)�
(Coleoptera,�Scarabaeidae).�Ecological�Entomology,�1:�19�29.�
Krause,�M.,�Bellosi,�E.�y�J.�F.�Genise.�2007.�Evolución�de� la� icnofauna�terrestre�
de� la� Patagonia� Central� II.� El� Grupo� Río� Chico:� desde� el� K�T� hasta� la�
explosión�del�Eoceno�Medio.�5�7���6��$���������/ �����8�&�///�7���6��
��/ �����8���� � ����-�G����-�p.�38.�
Krause,� J.�M.,�Bown,�T.�M.,�Bellosi,�E.�S.� y� J.�F.�Genise.�2008.�Trace� fossils�of�
cicadas�from�the�Cenozoic�of�Central�Patagonia�(Argentina).�Palaeontology,�
51�(2):�405�418.�
Krell,� F.� T.� 2000.� The� fossil� record� of� Mesozoic� and� Tertiary� Scarabaeoidea�
(Coleoptera:�Polyphaga).�Invertebrate�Taxonomy,�14:�871�905.�
Krell,� F.�T.�2007.�Catalogue�of� fossil�Scarabaeoidea� (Coleoptera:�Polyphaga)�of�
the�Mesozoic� and�Tertiary.�Denver�Museum�of�Nature�&�Science�Technical�
Report�2007�8,�1�81.�

� ����
Krell,� F.� T.� 2008.� Comments� on� the� proposed� conservation� of� the� ichnogenus�
������������ Sauer,� 1955� (Ichnotaxa,� Insecta,� Coleoptera,�
Coprinisphaeridae)� (1)� (Case� 3360;� see� BZN� 63:� 243–246).� Bulletin� of�
Zoological�Nomenclature,�65�(1):�50�54.�
Labandeira,� C.� C.� y� J.� J.� Sepkoski.� 1993.� Insect� diversity� in� the� fossil� record.�
Science,�261:�310�315.��
Laza,� J.� H.� 1986a.� Icnofósiles� de� paleosuelos� del� Cenozoico� mamalífero� de�
Argentina.� II.� Neógeno.� Boletín� Informativo� Asociación� Paleontológica�
Argentina,�15:�13.�
Laza,� J.� H.� 1986b.� Icnofósiles� de� paleosuelos� del� Cenozoico� mamalífero� de�
Argentina.� I.� Paleógeno.� Boletín� Informativo� Asociación� Paleontológica�
Argentina,�15:�19.�
Laza,� J.� H.� 2006.� Dung�Beetle� Fossil� Brood� Balls:� The� Ichnogenera�
������������ Sauer� and�I������ ����� (Coprinisphaeridae).� Ichnos,� 13:�
217–235.�
Laza,� J.� H.� y� M.� A.� Reguero.� 1990.� Extensión� faunística� de� la� antigua� región�
Neotropical�en� la�península�antártica�durante�el�Eoceno.�Ameghiniana,�26:�
245.�
Lengerken,�H.�V.�1954.�Die�Brutfürsorge��und�Brutpflegeinstinkte�der�Käfer.�2nd�
ed.�Akademische�Verlagsgesellschaft�Geest�&�Portig,�Leipzig.�
Langerken,�H.�V.�1955.�Die�Brutbirnen�von�(��� ������(Coleop.�Lamellicornia).�
Wissenschaftliche� Zeitschrift� der� Martin�Luther�Universitat� Halle�
Wittemberg,�Mathematisch�Naturwissenschaftliche�Klasse,�4�(5):�933�940.�
MacEachern,� J.�A.,�Pemberton,�S.�G.,�Gingras,�M.�K.�y�K.�L.�Bann.�2007a.�The�
ichnofacies� paradigm:� a� fifty�year� retrospective.� 52–77� en� �� � ���������
��� ���-�)������-�)���� ����W.�Miller�(ed.).�Amsterdam:�Elsevier.�

� ����
MacEachern,�J.�A.,�Pemberton,�S.�G.,�Gingras,�M.�K.,�Bann,�K.�L.�y�L.�T.�Dafoe.�
2007b.� Uses� of� trace� fossils� in� genetic� stratigraphy.� 110–134� en� �� �
��������� ��� ���-� )������-� )���� ���� W.� Miller� (ed.).� Amsterdam:�
Elsevier.�
Madden,�R.�H.,�Bellosi,�E.,�Carlini,�A.,�Heizler,�M.,�Vilas,�J.,�Re,�G.,�Kay,�R.�y�M.�
G.� Vucetich.� 2005.� Geochronology� of� the� Sarmiento� Formation� at� Gran�
Barranca�and�elsewhere� in�Patagonia:�calibrating�Middle�Cenozoic�mammal�
evolution�in�South�America.�45/������������6�� ��$�������-���)��,�4:�
411�412.�
Madden,� R.� y� E.� S.� Bellosi,� en� prensa.� Physical� stratigraphy� of� the� Sarmiento�
Formation� (Middle� Eocene�Lower� Miocene)� at� Gran� Barranca,� Central�
Patagonia� en� ��� )��������&� ��� ���� *��� 3� '��������� ���
'���������������� �������� ��� ����������� ����)�����.�Madden,�
R.,�Carlini,�A.�y�M.�G.�Vucetich�(eds.).�Cambridge�University�Press.��
Madden,�R.,�Carlini,�A.,�Vucetich,�M.�G.,�Kay,�R.�F.,�Heizler,�M.,�Vilas,�F.,�Re,�G.,�
Kohn,�M.,�Zucol,�A.� y�E.�Bellosi.�2003.�Gran�Barranca:� the�most� complete�
South� American� Middle� Cenozoic� sequence.� $���� �� J� )������ *���� ���
�&��������������)����-�*�����,�p.�38.�
Marshall,�L.�G.,�Cifelli,�R.,�Drake,�R.�y�G.�Curtis.�1986.�Vertebrate�paleontology,�
geology�and�geochronology�of�the�Tapera�Lopez�and�Scarrit�Pocket,�Chubut�
province,�Argentina.�Journal�of�Geology,�60:�920�951.�
Marshall,� L.�G.,�Hoffstetter,�R.� y�R.� Pascual.� 1983.�Mammals� and� stratigraphy:�
geochronology� of� the� continental� mammal�bearing� Tertiary� of� South�
America.�Palaeovertebrata.�Mémorie�Extraordinaire,�1983:�1�93.��

� ����
Martin,�A.�J.�2001.�Trace�fossil�evidence�for�insect�on�insect�parasitoid�behaviour�
in� the� Upper� Cretaceous� Two� Medicine� Formation� of� Choteau,� Montana.�
$���� ��������������� ���� ��&����$��� � ����-�*�����-�p.�112.�
Martínez,�A.�1959.�Catálogo�de�los�Scarabaeidae�argentinos�(Coleoptera).�Revista�
del� Museo� Argentino� de� Ciencias� Naturales,� Ciencias� Zoológicas,� Tomo� V,�
Vol.�1,�110p.�
Martinez,� S.� 1982.� Catálogo� sistemático� de� los� insectos� fósiles� de� América� del�
Sur.� Revista� Facultad� de� Humanidades� y� Ciencias;� Serie� Ciencias� de� la�
Tierra,�1�(2):�29�83.�
Mazzoni,� M.� M.� 1975.� La� Formación� Sarmiento� y� el� vulcanismo� paleógeno.�
Revista�de�la�Asociación�Geológica�Argentina,�40:�60�68.�
Mazzoni,�M.�M.�1979.�Contribución�al�conocimiento�petrográfico�de�la�Formación�
Sarmiento.� Barranca� sur� del� lago� Colhué� Huapí,� provincia� de� Chubut.�
Revista� de� la� Asociación� Argentina� de� Mineralogía,� Petrología� y�
Sedimentología,�10�(3�4):�33�53.�
Mazzoni,� M.� M.� 1985.� La� Formación� Sarmiento� y� el� vulcanismo� paleógeno.�
Revista�de�la�Asociación�Geológica�Argentina,�40:�60�68.�
McCartney,�G.�C.� 1933.� The�bentonites� and� closely� related� rocks� of� Patagonia.�
American�Museum�Novitates,�630:�1�16.�
Mikuláš,�R.�y�J.�F.�Genise.�2003.�Traces�within�traces.�Holes,�pits�and�galleries�in�
walls�and�fillings�of�insect�trace�fossils�in�paleosols.�Geologica�Acta,�1:�339�
348.�
Miquel,�S.�E.�y�E.�S.�Bellosi.�2007.�Microgasterópodos�terrestres�(Charopidae)�del�
Eoceno� medio� de� Gran� Barranca� (Patagonia� Central,� Argentina).�
Ameghiniana,�44:�121�131.�

� ����
Monteresino,�E.�M.�y�M.�Zunino,�2003.�Sobre�el�comportamiento�de�alimentación�
y� nidificación� de� Eucraniini� (Coleoptera� Scarabaeidae:� Scarabaeinae).�
Monografías�Tercer�Milenio,�3:�75�80.��
Morelli,�E.,�González�Vainer,�P�y�C.�Canziani.�1996.�Nidificación,�ciclo�de�vida�y�
estadios� preimaginales� de� ��� ������� ����� (Laporte,� 1840)�
(Coleoptera:�Scarabaeidae).�Elytron,�10:�11�22.���
Murphy,� C.� P.� 1986.� Thin� sections� preparation� of� soils� and� sediments.� A.� B.�
Academic�Publishers,�London.�149�p.�
Ocampo,� F.� C.� 2004.� Food� relocation� behavior� and� synopsis� of� the� southern�
South� American� genus� ��&�������� Westwood� (Scarabaeidae:�
Scarabaeinae:�Eucraniini).�The�Coleopterists�Bulletin,�58:�295�305.��
Ocampo,�F.�C.�y�T.�K.�Phillips.�2005.�Food�relocation�and�nesting�behavior�of�the�
Argentinian� dung� beetle� genus�'� ������ and� comparison�with� southwest�
African�� �����() �&���)�(Coleoptera:�Scarabaeidae:�Scarabaeinae).�
Revista�de�la�Sociedad�Entomológica�Argentina,�64�(1�2):�53�59.�
Pascual,� R.� 1984.� La� sucesión� de� Edades� mamífero,� de� los� climas� y� del�
diastrofismo�sudamericano�durante�el�Cenozoico:� fenómenos�concurrentes.�
Anales�de�la�Academia�Nacional�de�Ciencias�Exactas,�Físicas�y�Naturales,�36:�
15�37.�
Pascual,�R.�y�E.�Ortíz�Jaureguízar.�1990.�Evolving�climates�and�mammal�faunas�
in�Cenozoic�South�America.�Journal�of�Human�Evolution,�19:�23�60.�
Pascual,�R.,�Ortíz�Jaureguízar,�E.�y�J.�L.�Prado.�1996.�Land�mammals:�Paradigm�
for� Cenozoic� South� American� geobiotic� evolution.� Münchner�
Geowissenschaffen�Abhadlungen,�30�(A):�265�319.�
Pieńkowski,� G.� 2004.� Sołtyków� –�� unikalny� zapis� paleoecologiczny�
wczesnojurajskich�utworów�kontynentalnych.�Tomy�Jurajskie,�2:�1�16.�

� ����
Philips,�T.�K.,�Edmonds,�W.�D.�y�C.�H.�Scholtz.�2004a.�A�phylogenetic�analysis�of�
the� New�World� tribe� Phanaeini� (Coleoptera:� Scarabaeidae:� Scarabaeinae):�
Hypotheses�on�relationships�and�origins.� Insect�Systematics�and�Evolution,�
35:�43�63.��
Philips,� T.� K.,� Pretorius,� E.� y� C.� H.� Scholtz.� 2004b.� A� phylogenetic� analysis� of�
dung� beetles� (Scarabaienae:� Scarabaeidae):� unrolling� and� evolutionary�
history.�Ivertebrate�Systematics,�18:�53�88.��
Piperno,�D.�R.�2006.�Phytoliths:�A�Comprehensive�Guide� for�Archaeologists�and�
Paleoecologists.�AltaMira,�Lanham,�MD.�238�p.�
Piperno,�D.�R.�y�H�D.�Sues.�2005.�Dinosaurs�dined�on�grass.�Science,�310:�1126�
1128.�
Prasad,�V.,�Stromberg,�C.�A.�E.,�Alimohammadian,�H.�y�A.�Sahni.�2005.�Dinosaur�
coprolites� and� the� early� evolution� of� grasses� and� grazers.� Science,� 310:�
1177�1179.�
Ré,�G.,�E.�Bellosi,�M.�Heizler,�J.�Vilas,�R.�Madden,�A.�Carlini,�R.�Kay�y�M.�Vucetich,�
en� prensa.�Geochronology� for� the� Sarmiento� Formation� at�Gran�Barranca.�
En�The�Paleontology�of�Gran�Barranca:�Evolution�and�Environmental�Change�
through� the� Middle� Cenozoic� of� Patagonia,� ed.� Madden,� R.,� Carlini,� A.,�
Vucetich,�M.�&�Kay,�R.�Cambridge�University�Press.�Cambridge,�UK.�
Retallack,�G.�J.�1990.�The�work�of�dung�beetles�and�its�fossil�record.�214�226�en�
'���������&�)��������&����*�������������������.�A.� J.�Boucot� (ed.).�
Amsterdam:�Elsevier.�
Retallack,�G.�J.�1991.�Miocene�paleosols�and�ape�habitats�of�Pakistan�and�Kenya.�
New�York,�Oxford:�Oxford�University�Press,�Claredon�Press,�346�p.�
Retallack,�G.�J.�2001.�Soils�of�the�Past�(2nd.�ed.).�Blackwell,�Oxford,�404p.�

� ����
Retallack,� G.� J.� 2004.� Late� Oligocene� bunch� grassland� and� Early� Miocene� sod�
grassland� paleosols� from� central� Oregon,� USA.� Palaeogeography,�
Palaeoclimatology,�Palaeoecology,�207.�
Richter,� H.� 1918.� Ein� argentinischer� Pillendreher.� Zeitschrift� des� deutschen�
wissenschaftlichen� vereins� zur� Kultur�� und� Landeskunde� Argentiniens,� 5:�
342�347.�
Rivals,�F.�y�A.�Athanassiou.�2008.�Dietary�adaptations�in�an�ungulate�community�
from� the� late� Pliocene� of� Greece.� Palaeogeography,� Palaeoclimatology,�
Palaeoecology,�265:�134–139.�
Roselli,� F.� L.� 1939.� Apuntes� de� Geología� y� Paleontología� uruguayas� y� sobre�
insectos�del�Cretáceo�de�Uruguay�o�descubrimientos�de�admirables�instintos�
constructivos�de�esa�época.�Boletín�de� la�Sociedad�Amigos�de� las�Ciencias�
Naturales�“Kraglievich��Fontana”.�Nueva�Palmira,�Uruguay.�1:�29�102.��
Roselli,� F.� L.� 1976.� Contribución� al� estudio� de� la� geo�paleontología.�
Departamento� de� Colonia� y� Soriano� (República� Oriental� del� Uruguay).�
Edición� especial� de� la� Biblioteca� de� la� Sociedad� Amigos� de� las� Ciencias�
Naturales�“Kraglievich�Fontana”.�Nueva�Palmira,�Uruguay,�173�p.�
Roselli,� F.� L.� 1987.� Paleoicnología.� Nidos� de� insectos� fósiles� de� la� cobertura�
Mesozoica�del�Uruguay.�Publicación�del�Museo�Municipal�Nueva�Palmira,�1:�
1�56.�
Rougon,� D.� y� C.� Rougon.� 1980.� Le� cleptoparasitisme� en� zone� sahélienne:�
phénomène�adaptatif� d'insects�Coléoptères�Coprophages�Scarabaeidae�aux�
climats�arides�et�semi�arides.�Comptes�Rendus�de�l´Academie�des�Sciences,�
Paris,�291:�417�419.��
Sánchez,� M.� V.,� Laza,� J.� H.,� Bellosi,� E.� S.� y� J.� F.� Genise.� 2005.� Comparación�
preliminar�del�registro�fósil�de�Scarabaeinae�(Coleoptera:�Scarabaeidae).5/�

� ���
��������$����������'��������8��� ��:�-�90>9K?=?011K-���� �������
�� ��:�-�$������-�p.�120.��
Sánchez,�M.�V.,�Laza,�J.�H.,�Bellosi,�E.�S.,�y�J.�F.�Genise.�2006.�The�integration�of�
ichnofossil�and�body�record�in�Scarabaeinae�(Coleoptera:�Scarabaeidae).�5//�
7���6�� �������� �� �� � ��������8-� ���� ����� �� �� ����-�
*�����,�p.�20.��
Sánchez,�M.�V.,�Laza,�J.�H.,�Bellosi,�E.�S.�y�J.�F.�Genise.�2007.�Evolución�de� la�
icnofauna� terrestre� de� la� Patagonia� Central� V.� Formación� Sarmiento:� la�
explosión�del�Eoceno�Medio.�5�7���6��$���������/ �����8�&�///�7���6��
��/ �����8���� � ����-�G����-�p.�41.�
Sánchez,�M.�V.,�Laza,�J.�H.,�Bellosi,�E.�S.�y�J.�F.�Genise.�2008.�Icnoestratigrafía�y�
características�de� trazas� fósiles�de�escarabajos� coprófagos� (������������
ispp)�de� la�Formación�Sarmiento� (Cenozoico�medio):�aparición�y�evolución�
de� ecosistemas� de� pastizales� abiertos.� 4//� 7���6�� $������� ��
����������8-�*�����$���-�p.�162.�
Sánchez,�M.�V.�y�J.�F.�Genise.�2008.�Nest�and�brood�chamber�structure�of� two�
South� American� dung� beetles:� �������� � ������� Brullé� and� ��������
��� ���� (Fabricius)� (Coleoptera:� Scarabaeidae:� Scarabaeinae).� The�
Coleopterists�Bulletin,�62:�49–61.��
Sánchez,� M.� V.� y� J.� F.� Genise.� 2009.� Cleptoparasitism� and� detritivory� in� dung�
beetle�fossil�brood�balls�from�Patagonia�(Argentina).�Palaeontology,�52�(4):�
837�848.��
Sands,�W.�A.�1987.�Ichnocoenoses�of�probable� termite�origin� fron�Laetoli.�409�
433�en������-��)��� ������ ���@�����������.� Leakey,�M.�D.�y� J.�M.�
Harris�(eds.),�Clarendon�Press,�Oxford.�

� ���
Sato,�H.�y�M.�Imamori.�1987.�Nesting�behavior�of�a�subsocial�African�ball�roller�
.���� ���&������ (Coleoptera,� Scarabaeidae).� Ecological� Entomology,� 12:�
415�425.�
Sauer,�W.�1950.�Contribuciones�al�conocimiento�del�Cuaternario�en�el�Ecuador.�
Anales�de�la�Universidad�Central,�Quito,�Ecuador,�77�(328):�264�327.�
Sauer,�W.�1955.������������� ���������,�un�fósil�singular�del�Pleistoceno.�
Boletín� del� Instituto� de� Ciencias� Naturales� de� la� Universidad� Central� de�
Quito,�1�(2):�123�129.�
Sauer,�W.�1956.������������ ���������� (Bola�de�cangahua)�y� las�esferas�
elaboradas�actualmente�por�escarabajos�de�la�familia�Scarabaeidae.�Boletín�
de�Informaciones�Científicas�Nacionales�(Quito,�Ecuador),�8:�550�555.�
Sauer,�W.�1959.�Merkwurdige�ugeln�in�Tuffen�Ecuadors�und�ihre�Deutung.�Natur�
und�Volk,�89:�118�124.�
Sauer,�W.�1965.�Geología�del�Ecuador.�Ministerio�de�Educación,�Quito,�383�p.�
Shaw,� R.� B� 2000.� Tropical� Grasslands� and� Savannas� en������3� �&����� ��
���'��������.�Jacobs,�S.�W.�L.�y�J.�Everett�(eds.).�CSIRO:�Melbourne.�
Simpson,�G.�G.�1940.�Review�of�the�mammal�bearing�Tertiary�of�South�America.�
Proceedings�of�the�American�Philosophical�Society,�83:�649�710.�
Sokal,�R.�R.�y�F.�J.�Rohlf.�1979.�Biometry.�Freeman,�New�York.�
Spalletti,�L.�A.�y�M.�M.�Mazzoni.�1977.�Sedimentología�del�Grupo�Sarmiento�en�
un� perfil� ubicado� al� sudeste� del� lago� Colhué�Huapí,� provincia� de� Chubut.�
Obra�del�Centenario�del�Museo�de�La�Plata,�4:�261�283.�
Spalletti,�L.�A.�y�M.�M.�Mazzoni.�1979.�Estratigrafía�de�la�Formación�Sarmiento�en�
la� barranca� sur� del� lago�Colhué�Huapí,� provincia� de�Chubut.�Revista� de� la�
Asociación�Geológica�Argentina,�34:�271�281.�

� ��
Strömberg,�C.�A.�E.�2002.�The�origin�and�spread�of�grass�dominated�ecosystems�
in� the� Late� Tertiary� of� North� America:� Preliminary� results� concerning� the�
evolution� of� hypsodonty.� Palaeogeography,� Palaeoclimatology,�
Palaeoecology,�177�(1–�2):�59–75.�
Strömberg,�C.�A.�E.�2004.�Using�phytolith�assemblages�to�reconstruct�the�origin�
and�spread�of�grass�dominated�habitats�in�the�great�plains�of�North�America�
during� the� late� Eocene� to� early� Miocene.� Palaeogeography,�
Palaeoclimatology,�Palaeoecology,�207:�239–27.�
Strömberg,� C.� A.� E.� y� T.� A.� Stidham.� 2001.� Dung� beetle� brood� balls� and�
notoungulate�diet.�Journal�of�Vertebrate�Paleontology,�21�(3):�105A.�
Taylor,� A.� M.,� Goldring,� R.� y� S.� Gowland.� 2003.� Analysis� and� application� of�
ichnofabrics.�Earth�Science�Reviews,�60:�227�259.�
Twiss,� P.� C.� 1992.� Predicted� world� distribution� of� C3� and� C4� grass� phytoliths.�
113–128�en�)�&������� �&����� �.�Rapp,�Mulholland� (eds.),� Plenum�Press,�
New�York.�
Valli,�A.�M.�F.�y�M.�R.�Palombo.�2008.�Feeding�behaviour�of�middle�size�deer�from�
the�Upper� Pliocene� site� of� Saint�Vallier� (France)� inferred� by�morphological�
and� micro/mesowear� analysis.� Palaeogeography,� Palaeoclimatology,�
Palaeoecology,�257:�106�122.�
Verde,�M.�2000.�Trazas�fósiles�de�artrópodos�en�el�Uruguay.�/�����6����*��������
��)�������������8-�/�����6���������� �����)�������������8-�/�
/���������� ����� ��� )��������������&-� 7����C�� )���� #� �)-� *����-�
;#D?=?0111-�p.�112–113.�
Verde,� M.� y� J.� F.� Genise.� 2007.� Evolución� de� la� icnofauna� terrestre� de� la�
Patagonia� Central� IV.� Formación� Asencio� del� Terciario� temprano� del�
Uruguay:�¿El�antecedente�de�la�explosión�del�Eoceno�Medio�en�la�Patagonia�

� ���
Central?.�5�7���6��$��������� / �����8� &� ///�7���6���� / �����8� ���
� ����-�G����,�p.�40.�
Verde,�M.,�Jimenez,�J.�J.,�Ubilla,�M.�y�J.�F.�Genise.�2007.�A�new�earthworm�trace�
fossil�from�paleosols:�Aestivation�chambers�from�the�late�Pleistocene�Sopas�
Formation�of�Uruguay.�Palaeogeography,�Palaeoclimatology,�Palaeoecology,�
243:�339�347.�
Vucetich,� M.� G.,� Vieytes,� E.� C.,� Kramarz,� A.� y� A.� Carlini.� 2005.� Los� roedores�
caviomorfos� de� Gran� Barranca:� aportes� bioestratigráficos� y�
paleoambientales.�45/������������6�� ��$�������-���)��-�4:�413�418.�
Waterhouse,�D.�F.�1974.�The�biological�control�of�dung.�Scientific�Amercian,�230:�
101�109.�
Watteau,� F.� y� M.� Villemin.� 2001.� Ultrastructural� study� of� the� biogeochemical�
cycle�of�silicon�in�the�soil�and�litter�of�a�temperate�forest.�European�Journal�
of�Soil�Science,�52:�385�396.�
Wilf,� P.,� Labandeira,� C.� C.,� Johnson,� K.� R.� y� N.� R.� Cúneo.� 2005.� Richness� of�
plant�insect�associations�in�Eocene�Patagonia:�A�legacy�for�South�American�
biodiversity.�Proceedings�of�the�National�Academy�of�Sciences,�102:�8944�
8948.�
Windhausen,�A.�1924.�Líneas�generales�de�la�constitución�geológica�de�la�región�
situada�al�oeste�del�Golfo�de�San�Jorge.�Boletín�de�la�Academia�Nacional�de�
Ciencias�de�Córdoba,�23:�167�320.��
Zucol,�A.�F.�2001.�Fitolitos�III.�Una�nueva�metodología�descriptiva.�Asociaciones�
fotolíticas�de�)���� ����������������(Stipeae,�Poaceae).�Boletín�de�la�
Sociedad�Argentina�de�Botánica,�36:�69�85.�

� ���
Zucol,�A.�F.�2005.�Estudios�paleoagrostológicos�en�el�Cenozoico�de�Patagonia,�su�
importancia� en� el� conocimiento� de� la� historia� evolutiva� de� los�
paleopastizales.�45/������������6�� ��$�������-���)��-�4:�403�408.�
Zucol,�A.�F.�y�M.�Brea.�2005.�El�registro�fitolítico�de�los�sedimentos�del�Cenozoico�
de� la� Localidad� de� Gran� Barranca:� su� aporte� a� la� reconstrucción�
paleoecológica.�45/������������6�� ��$�������-���)��-�4:�395�402.�
Zucol,� A.� F.� y� M.� Osterrieth.� 2002.� Técnicas� de� preparación� de� muestras�
sedimentarias�para�la�extracción�de�fitolitos.�Ameghiniana,�39:�379�382.�
Zucol,�A.�F.,�Brea,�M.,�Carlini,�A.�A.y�R.�H.�Madden.�2001.�Análisis�fitolitico�de�la�
secuencia�sedimentariade�Gran�Barranca,�Chubut,�Argentina:�II.�El�Miembro�
Colhué� Huapi� (Formación� Sarmiento).� //� '� ������ �� /������ �����
�������� �������������-�p.�35�36.�
Zucol,� A.� F.,� Mazzoni,� M.� M.� y� R.� H.� Madden.� 1999.� Análisis� fitolíticos� en� la�
secuencia�sedimentaria�de�Gran�Barranca,�Chubut.�Ameghiniana,�36:�43R.�
Zunino,� M.,� Barabero,� E.� y� M.� Luzzatto.� 1989.� Food� relocation� behavior� in�
Eucraniina�beetles�(Scarabaeidae)�and�the�constraints�of�xeric�environment.�
Tropical�Zoology,�2:�235�240.�
�
�
��





























