Embed Size (px)
Citation preview

0
UNIVERSIDADE FEDERAL DO CEARÁ
FACULDADE DE MEDICINA
DEPARTAMENTO DE FISIOLOGIA E FARMACOLOGIA
PROGRAMA DE PÓS-GRADUAÇÃO EM FARMACOLOGIA
RENATA BESSA PONTES
ENVOLVIMENTO DA ENDOTELINA-1, DE RECEPTORES
(TRPV1 E NMDA) E DA SUBSTÂNCIA P NA NEUROPATIA
SENSITIVA PERIFÉRICA INDUZIDA PELO AGENTE
ANTINEOPLÁSICO OXALIPLATINA
FORTALEZA
2015

1
RENATA BESSA PONTES
ENVOLVIMENTO DA ENDOTELINA-1, DE RECEPTORES
(TRPV1 E NMDA) E DA SUBSTÂNCIA P NA NEUROPATIA
SENSITIVA PERIFÉRICA INDUZIDA PELO AGENTE
ANTINEOPLÁSICO OXALIPLATINA
Tese apresentada ao programa de Pós-graduação
em Farmacologia da Faculdade de Medicina do
Departamento de Fisiologia e Farmacologia da
Universidade Federal do Ceará, como parte dos
requisitos para obtenção do título de Doutor em
Farmacologia.
Orientador: Prof. Dr. Ronaldo de Albuquerque Ribeiro
Co-orientadora: Profa. Dra. Mariana Lima Vale
Trabalho desenvolvido no Laboratório de Farmacologia da Inflamação e do Câncer
(LAFICA) da Universidade Federal do Ceará
FORTALEZA
2015

2
RENATA BESSA PONTES
ENVOLVIMENTO DA ENDOTELINA-1, DE RECEPTORES (TRPV1 E NMDA) E DA
SUBSTÂNCIA P NA NEUROPATIA SENSITIVA PERIFÉRICA INDUZIDA PELO AGENTE
ANTINEOPLÁSICO OXALIPLATINA
Essa tese foi submetida como parte dos requisitos necessários à obtenção do Grau de Doutor em Farmacologia, outorgado pela Universidade Federal do Ceará e encontra-se à disposição dos interessados na Biblioteca setorial da referida Universidade. Data da aprovação: ___/___/_____
BANCA EXAMINADORA

3
Dedico essa tese a toda minha família que tanto
me apoia. Em especial aos meus pais que
sempre acreditaram no meu sucesso, aos meus
irmãos que sempre estiveram ao meu lado, ao
meu querido esposo, meu companheiro, pelo
apoio, dedicação e compreensão nos momentos
difíceis e ao meu amado filho que na sua
pequenenice ainda não compreendeu minhas
ausências quando me chamava para brincar.

4
AGRADECIMENTOS
Primeiramente a Deus por todas as oportunidades em minha vida, pelos dons
da ciência e sabedoria tão necessários para o desenvolvimento desse trabalho. E a
Nossa Senhora por sempre guiar meus caminhos.
Muito especialmente ao meu orientador Prof. Dr. Ronaldo de Albuquerque
Ribeiro pela oportunidade e ensinamentos durantes todos esses anos no
laboratório, por ter acreditado em mim e pelo exemplo de competência e
profissionalismo. E hoje, um mês do seu falecimento não tenho como deixar de
mencionar a enorme falta que faz no LAFICA! Saudades eternas!
Agradeço à minha co-orientadora Profa. Dra. Mariana Lima Vale pela sua
paciência e dedicação, transmitindo-me apoio para a realização desse trabalho.
Aos Profs. Thiago Cunha e Fernando Cunha pelo apoio durante o
desenvolvimento da pesquisa na FMRP (USP).
À amiga, grande amiga, Juliana Lino pela disponibilidade e organização dos
experimentos que juntas realizamos, sentirei saudades.
Aos amigos Fábio Bezerra, Ana Paula, Anamaria, Carol, Cibele, Aline,
Deysen pela participação neste trabalho e de um modo geral a todos que fazem o
LAFICA.
Aos Professores Alexandre Havt e Roberto César e aos amigos Deysi
Wong, Karoline Aragão, Marco Clementino, Mário Pontes, Kalina Souza,
Rosinha pela amizade e disposição para ajudar.
À técnica de laboratório Vandinha, que é mais que uma técnica, pelo
reiterado carinho e apoio.

5
Ao corpo docente do programa de Pós-graduação do Departamento de
Fisiologia e Farmacologia pela formação que me legaram e à Dra Margarida, do
Departamento de Patologia, por gentilmente ter cedido espaço em seu laboratório,
assim como ao técnico Adalberto Júnior.
À Unichristus por todo apoio a mim conferido em especial ao Dr. José
Rocha e à Profa. M.Sc. Romina Mourão representando meus amigos na
Instituição.
À Profa. Elzenir Coelho e Prof. Edson Alencar pelo atendimento linguístico,
à Profa. M.Sc. Cymara Kuhener pela bioestatística, à Profa. Dra. Mirizana Alves
pelo apoio metodológico e à Profa. M.Sc. Cintia Rocha pelo apoio psicológico.
À Profa M.Sc. Ediara Rios e Profa. M.Sc. Mylza Carvalho pelo apoio no
início da vida acadêmica como exemplos de profissionais e de docentes.
À Profa. M.Sc. Fabiane Elpídio pelo apoio como coordenação do curso de
Fisioterapia (UFC) e ao colegiado pelo incentivo nesta fase final.
Ao Grupo São Vicente (GSV), in memoriam Ir. Yolanda Brasil, pela formação
pessoal e religiosa a mim conferidos.
Aos meus pais, amigos incondicionais, pelo eterno e imensurável amor, pela
minha existência e o que dela sou.
Aos meus irmãos, cunhadas, sobrinhos, tios, primos, sogros e amigos por
sempre acreditarem na minha dedicação e nos frutos dela colhidos.

6
Ao meu esposo, Júlio César, amigo e confidente, pelo lindo amor que me
dedica, por me tornar uma pessoa mais feliz e por acreditar em meus e, em nossos,
sonhos, fazendo até o impossível para a realização dessa tese.
Ao meu abençoado filho por sempre perguntar, quando a mamãe estava no
computador, se já estava perto de terminar para ir com ele brincar.
À FUNCAP pelo grande apoio financeiro durante os anos de pesquisa.
E a todos que participaram direta ou indiretamente em alguma etapa para a
realização desse trabalho.

7
Buscai primeiro o reino de Deus, e a sua justiça,
e todas as coisas vos serão acrescentadas (Mt
6, 33).

8
RESUMO
ENVOLVIMENTO DA ENDOTELINA-1, DE RECEPTORES (TRPV1 E NMDA) E DA SUBSTÂNCIA P NA NEUROPATIA SENSITIVA PERIFÉRICA INDUZIDA PELO AGENTE ANTINEOPLÁSICO OXALIPLATINA. Renata Bessa Pontes. Doutorado em Farmacologia Departamento de Fisiologia e Farmacologia. Faculdade de Medicina. Universidade Federal do Ceará. Orientador: Prof. Dr. Ronaldo de Albuquerque Ribeiro. Co-Orientadora: Profa. Dra. Mariana de Lima Vale.
Introdução: A neurotoxicidade cumulativa é uma toxicidade que pode advir da terapia à base de
oxaliplatina (OXL), que é a 3ª geração de agentes platinos com amplo espectro de atividade
antitumoral, incluindo câncer colorretal, ovariano e pulmonar. A neurotoxicidade associada à OXL
gera uma toxicidade dose-limitante, crônica, a neuropatia sensitiva periférica (NSP). Objetivo:
Investigar o envolvimento da endotelina-1, de receptores TRPV1 e NMDA e da substância P
envolvidos na patogênese da neuropatia sensitiva periférica induzida pelo agente antineoplásico
oxaliplatina. Materiais e métodos: O estudo foi aprovado pelo Comitê de Ética em Pesquisa Animal
da UFC (protocolo nº 75/12). Camundongos Swiss machos (20g) foram pré-tratados com
antagonistas de receptores de endotelina-1 (Bosentana 100mg/kg, VO; BQ-123 e BQ-788 30µl,
intraplantar) e antagonistas do receptor TRPV1 (capsazepina, 5mg/kg, IP), antagonista do receptor
NK-1 da Substancia P (apreptanto, 1mg/kg, IP) e antagonista de receptores NMDA (MK-801,
0,5mg/kg, IP) 30 minutos antes da administração de OXL (1mg/kg, IV) por 4 semanas e meia.
Paralelamente foram realizados testes nociceptivos para avaliar o desenvolvimento da neuropatia
sensitiva periférica. A hipernocicepção foi avaliada pelo teste de imersão da cauda (TIC) em água fria
(10ºC) ou aquecida (43ºC) e pelo teste Von Frey (HPM). Em seguida, foi realizado
imunofluorescência do segmento medular e gânglio da raiz dorsal e RT-PCR. Resultados: Como
resultados observou-se que com o pré-tratamento ao uso de OXL que houve atenuação da
hiperalgesia da NSP induzida por OXL. Ao realizar a administração de antagonistas seletivos de
endotelina-1 intraplantar na pata direita observou-se redução significativa na hiperalgesia na pata
direita (tratada) em comparação à pata esquerda (controle). Ao analisar a expressão gênica para
cFos, NK-1 e o receptor de endotelina B, observou-se que houve redução significativa da expressão
dos marcadores no grupo pré-tratado com Bosentana ao comparar com o grupo OXL, que
demonstrou a expressão aumentada para esses marcadores. Conclusão: Conclui-se no presente
estudo que há evidências do papel da endotelina-1, de receptores (TRPV1 e NMDA) e da substância
P na patogênese da NSP induzida pelo agente antineoplásico OXL.
Palavras-chave: 1. Compostos de Platina. 2. Endotelina 3. Doenças do Sistema Nervoso Periférico.
4. Camundongos.

9
ABSTRACT
INVOLVEMENT OF ENDOTHELIN-1, RECEPTORS (TRPV1 AND NMDA) AND
NEUROPEPTIDE SP IN PERIPHERAL SENSITIVE NEUROPATHY INDUCED BY
ANTINEOPLASTIC AGENT OXALIPLATIN. Renata Bessa Pontes. PhD in Pharmacology
Department of Physiology and Pharmacology. Faculty of Medicine. Federal University of Ceara.
Advisor: Prof. Dr. Ronaldo Ribeiro de Albuquerque. Co-Advisor: Prof. Dra. Mariana Lima Vale.
Introduction: The cumulative neurotoxicity is a toxicity that can result from oxaliplatin-based therapy
(OXL), which is the 3rd generation platinum agent with broad spectrum of antitumor activity, including
colorectal, ovarian and lung cancer. Neurotoxicity associated with OXL generates a dose-limiting
toxicity, chronic, peripheral sensory neuropathy (NSP). Objective: To investigate the involvement of
endothelin-1, TRPV1 receptors and NMDA and substance P involved in the pathogenesis of
peripheral sensory neuropathy induced by oxaliplatin antineoplastic agent. Methods: Male Swiss mice
(20g) were pre-treated with antagonists of endothelin-1 receptors (Bosentan 100mg / kg orally; BQ-
123 and BQ-788 30μl, intraplantar) and TRPV1 receptor antagonists (capsazepine, 5mg / kg , IP),
antagonist of NK-1 receptor for substance P (apreptanto, 1 mg / kg, IP), and a NMDA receptor
antagonist (MK-801, 0.5mg / kg, IP) 30 minutes before administration of OXL (1mg / kg, IV) for 4.5
weeks. Parallel nociceptive tests performed to assess the development of peripheral sensory
neuropathy. The hyperalgesia assessed by the tail immersion test (ICT) in cold water (10° C) or warm
(43° C) and test Von Frey (HPM). Then it was performed spinal segment, and the dorsal root ganglion
immunofluorescence and RT-PCR the Ethics Committee approved the study for Animal Research
UFC (Protocol 75/12). Results: The results observed when using the antagonists, as a pretreatment
to the use of OXL there was attenuation of the induced hyperalgesia (NSP) OXL. Upon administration
of selective antagonists of endothelin in the right paw was significant reduction in paw hyperalgesia in
the right (treated) compared to the left paw (control). By analyzing the gene expression of cFos, NK-1
and endothelin B receptor, it was observed that there was significant reduction of expression of the
markers in pre-treated bosentan group versus OXL group that showed increased expression for these
markers. Conclusion: It was concluded in this study that there is evidence of the role of endothelin-1
receptors (TRPV1 and NMDA) and substance SP in the pathogenesis of NSP induced antineoplastic
agent OXL.
Keywords: 1. Platinum Compounds. 2. Endothelin 3. Peripheral Nervous System Diseases. 4. Mice.

10
LISTA DE FIGURAS
FIGURA 1 Mecanismo de ação antitumoral da oxaliplatina. 25
FIGURA 2 Modelo do envolvimento do eixo endotelina–
receptores na neuropatia sensitiva periférica induzida
por oxaliplatina.
34
FIGURA 3 Protocolo experimental para o uso dos antagonistas
seletivos para receptores de endotelina-1, TRPV1,
NMDA e da substância P na indução da neuropatia
sensitiva periférica induzida por oxaliplatina.
52
FIGURA 4 Coleta de segmento medular e gânglio da raiz dorsal 55
FIGURA 5 Lista de anticorpos primários e seus respectivos
secundários.
57
FIGURA 6 Sequência de primers utilizados no RT-PCR. 59
FIGURA 7 Avaliação do antagonista não seletivo de endotelina
(Bosentana) na hiperalgesia mecânica plantar
induzida por oxaliplatina.
63
FIGURA 8 Avaliação do antagonista não seletivo de endotelina
(Bosentana) no desenvolvimento de alodínia térmica
ao frio (10 ºC) induzida por oxaliplatina.
64
FIGURA 9 Avaliação do antagonista seletivo de receptor de
endotelina A (BQ-123) na hiperalgesia mecânica
plantar induzida por oxaliplatina.
66
FIGURA 10 Comparação da pata Direita e pata Esquerda da
administração do antagonista seletivo de receptor de
endotelina A (BQ-123) na hiperalgesia mecânica
plantar induzida por oxaliplatina no 28º dia.
67
FIGURA 11 Avaliação do antagonista seletivo de receptor de
endotelina B (BQ-788) na hiperalgesia mecânica
plantar induzida por oxaliplatina.
69
FIGURA 12 Comparação da pata direita e pata esquerda da
administração do antagonista seletivo de receptor de
70

11
endotelina B (BQ-788) na hiperalgesia mecânica
plantar induzida por oxaliplatina.
FIGURA 13 Avaliação do antagonista do TRPV1 (capsazepina) na
hiperalgesia mecânica plantar induzida por
oxaliplatina.
72
FIGURA 14 Avaliação do antagonista do TRPV1 (capsazepina) no
desenvolvimento de alodínia térmica ao calor (43 ºC)
induzida por oxaliplatina.
73
FIGURA 15 Avaliação do antagonista do receptor NK-1
(apreptanto) na hiperalgesia mecânica plantar
induzida por oxaliplatina.
75
FIGURA 16 Avaliação do antagonista do receptor NK-1
(apreptanto) no desenvolvimento de alodínia térmica
ao frio (10 ºC) induzida por oxaliplatina.
76
FIGURA 17 Avaliação do antagonista do receptor de NMDA (MK-
801) na hiperalgesia mecânica plantar induzida por
oxaliplatina.
78
FIGURA 18 Avaliação do antagonista do receptor de NMDA (MK-
801) no desenvolvimento de alodínia térmica ao frio
(10 ºC) induzida por oxaliplatina.
79
FIGURA 19 Fotomicrografias da marcação por
imunofluorescência para cFOS de medula espinhal
de camundongos submetidos ao pré-tratamento com
Bosentana.
82
FIGURA 20 Porcentagem de área imunomarcada para cFOS em
medula espinhal de camundongos submetidos ao
pré-tratamento com Bosentana.
83
FIGURA 21 Fotomicrografias da marcação por
imunofluorescência para cFOS de DRG de
camundongos submetidos ao pré-tratamento com
Bosentana.
84
FIGURA 22 Fotomicrografias da marcação por
imunofluorescência para o receptor NK-1 de medula
86

12
espinhal de camundongos neuropáticos.
FIGURA 23 Fotomicrografias da marcação por
imunofluorescência para o receptor NK-1 de DRG de
camundongos neuropáticos.
87
FIGURA 24 Fotomicrografias da marcação por
imunofluorescência para NK-1 de medula espinhal de
camundongos submetidos ao pré-tratamento com
Bosentana.
89
FIGURA 25 Porcentagem de área imunomarcada para NK-1 de
medula espinhal de camundongos submetidos ao
pré-tratamento com Bosentana
90
FIGURA 26 Fotomicrografias da marcação por
imunofluorescência para NK-1 de DRG de
camundongos submetidos ao pré-tratamento com
Bosentana.
91
FIGURA 27 Fotomicrografias da marcação por
imunofluorescência para o receptor de endotelina B
de medula de camundongos neuropáticos.
93
FIGURA 28 Fotomicrografias da marcação por
imunofluorescência para o receptor de endotelina B
de DRG de camundongos neuropáticos.
94
FIGURA 29 Expressão gênica para Receptor de Endotelina A em
medula espinhal nos grupos controle e oxaliplatina.
97
FIGURA 30 Expressão gênica para Receptor de Endotelina A em
DRG nos grupos controle e oxaliplatina.
98
FIGURA 31 Expressão gênica para Receptor de Endotelina B em
medula espinhal nos grupos controle e oxaliplatina.
99
FIGURA 32 Expressão gênica para Receptor de Endotelina B em
DRG nos grupos controle e oxaliplatina.
100
FIGURA 33 Expressão gênica para TRPV1 em medula espinhal
nos grupos controle e oxaliplatina
102
FIGURA 34 Expressão gênica para TRPV1 em DRG nos grupos
controle e oxaliplatina
103

13
FIGURA 35 Expressão gênica para SP em medula espinhal nos
grupos controle e oxaliplatina
105
FIGURA 36 Expressão gênica para SP em DRG nos grupos
controle e oxaliplatina
106
FIGURA 37 Modelo do envolvimento da endotelina, dos
receptores NMDA e TRPV1 e do neuropeptídeo SP na
neuropatia sensitiva periférica induzida por
oxaliplatina.
117

14
LISTA DE TABELAS
TABELA 1
Agonistas e antagonistas dos receptores de
endotelina usados em pesquisa básica e estudos
clínicos
33
TABELA 2
Descrição dos grupos experimentais 49

15
LISTA DE ABREVIATURAS
ANOVA Análise de Variância
CEPA Comitê de Ética em Pesquisa com Animais
cm Centímetros
CMC Carboximetilcelulose Sal Sódico
CZP Capsazepina
DACH 1-2-Diaminociclohexano
DMSO Dimetil sulfóxido
DNA Ácido desoxirribonucléico
DOPA Dopamina
DRG Gânglio da Raiz Dorsal
ET-1 Endotelina-1
ETA Receptores de endotelina A
ETB Receptores de endotelina B
EPM Erro padrão da média
FU Fluourouracil
HCO3- Bicarbonato de sódio
H2PO4 Dihidrogênio fosfato
IP Via Intraperitoneal
I.pl. Via Intraplantar
IV Via intravenosa

16
lafica Laboratório de Farmacologia da Inflamação e do Câncer
min Minutos
mm Milímetros
MMII Membros inferiores
MMSS Membros superiores
MMR Reparo de má combinação
NaCl Cloreto de sódio
NK-1 Neuroquinina-1
NMDA Ácido N-metil-D-aspartato
NSP Neuropatia Sensitiva Periférica
OXL Oxaliplatina
PFA Paraformaldeído
Pt Platina
RNA Ácido ribonucleico
SNC Sistema Nervoso Central
SNP Sistema Nervoso Periférico
SP Substância P
TRPV1 Receptor de potencial transiente vanilóide 1
VO Via Oral

17
SUMÁRIO
RESUMO VIII
ABSTRACT IX
LISTA DE FIGURAS X
LISTA DE TABELAS XIV
LISTA DE ABREVIATURAS XV
1 INTRODUÇÃO 20
1.1 CÂNCER 21
1.2 FARMACODINÂMICA E FARMACOCINÉTICA DA
OXALIPLATINA E O MECANISMO DE AÇÃO ANTITUMORAL
22
1.3 TOXICIDADE POR OXALIPLATINA 26
1.4 SISTEMA ENDOTELINA-1, RECEPTORES TRPV1 E NMDA E
SUBSTÂNCIA P NA NEUROPATIA SENSITIVA PERIFÉRICA
31
1.5 RELEVÂNCIA E JUSTIFICATIVA 38
2 OBJETIVOS 41
2.1 OBJETIVO GERAL 42
2.2 OBJETIVOS ESPECÍFICOS 42
3 MATERIAIS E MÉTODOS 44
3.1 ANIMAIS 45
3.2 ASPECTOS ÉTICOS 45
3.3 AMBIENTE 46
3.4 HORÁRIO DOS EXPERIMENTOS 46
3.5 OBSERVAÇÕES CLÍNICAS 46
3.6 APARELHOS E INSTRUMENTOS LABORATORIAIS 47
3.7 DROGAS, SOLUÇÕES E OUTROS LÍQUIDOS 47
3.8 PROTOCOLO EXPERIMENTAL E DESENHO DO ESTUDO 49
3.8.1 GRUPOS EXPERIMENTAIS 49
3.8.2 INDUÇÃO DA NEUROPATIA SENSITIVA POR OXALIPLATINA 50
3.8.3 AVALIAÇÃO DO ENVOLVIMENTO DA ENDOTELINA NA NSP
INDUZIDA POR OXALIPLATINA
50

18
3.8.4 AVALIAÇÃO DO DESENVOLVIMENTO DA NEUROPATIA
SENSITIVA PERIFÉRICA EM ANIMAIS TRATADOS COM
ANTAGONISTAS DOS RECEPTORES TRPV1, NK-1 E NMDA
51
3.8.5 TESTES PARA AVALIAÇÃO DE HIPERNOCICEPÇÃO
MECÂNICA E TÉRMICA
53
3.8.5.1 Teste de hiperalgesia mecânica plantar (Von Frey eletrônico) 53
3.8.5.2 Teste de imersão da cauda em água fria (10ºC) ou aquecida
(43ºC)
53
3.8.6 COLETA DO SEGMENTO MEDULAR E GÂNGLIOS DA RAIZ
DORSAL
54
3.8.7 IMUNOFLUORESCÊNCIA EM SEGMENTO MEDULAR E
GÂNGLIO DA RAIZ DORSAL
56
3.8.8 TRANSCRIÇÃO REVERSA - REAÇÃO EM CADEIA DE
POLIMERASE (RT-PCR) PARA SP, NMDA-R, ENDOTELINA A
E ENDOTELINA B DA MEDULA ESPINHAL E DO GÂNGLIO DA
RAIZ DORSAL
58
3.9 ANÁLISE ESTATÍSTICA 59
4 RESULTADOS 60
4.1 ENVOLVIMENTO DO ANTAGONISTA NÃO SELETIVO PARA
OS RECEPTORES A E B DA ET-1 (BOSENTANA)
62
4.2 ENVOLVIMENTO DO ANTAGONISTA SELETIVO PARA O
RECEPTOR A DA ET-1 (BQ-123)
65
4.3 ENVOLVIMENTO DO ANTAGONISTA SELETIVO PARA O
RECEPTOR B DA ET-1 (BQ-788)
68
4.4 ENVOLVIMENTO DO ANTAGONISTA DE RECEPTORES
TRPV1 NA NSP INDUZIDA POR OXL
71
4.5 ENVOLVIMENTO DO ANTAGONISTA DE RECEPTORES NK-1
NA NSP INDUZIDA POR OXL
74
4.6 ENVOLVIMENTO DO RECEPTOR NMDA NA NSP INDUZIDA
POR OXL
77
4.7 IMUNOFLUORESCÊNCIA PARA CFOS EM SEGMENTO
MEDULAR E DRG APÓS PRÉ-TRATAMENTO COM
81

19
BOSENTANA.
4.8 IMUNOFLUORESCÊNCIA PARA O RECEPTOR DE NK-1 EM
SEGMENTO MEDULAR E DRG
85
4.9 IMUNOFLUORESCÊNCIA PARA NK-1 EM SEGMENTO
MEDULAR E DRG APÓS PRÉ-TRATAMENTO COM
BOSENTANA
88
4.10 IMUNOFLUORESCÊNCIA PARA O RECEPTOR DE
ENDOTELINA B EM SEGMENTO MEDULAR E DRG
92
4.11 EXPRESSÃO GÊNICA PARA RECEPTORES DE ENDOTELINA
A E B NA MEDULA ESPINHAL E NO GÂNGLIO DA RAIZ
DORSAL
96
4.12 EXPRESSÃO GÊNICA PARA A TRPV1 NA MEDULA E NO
GÂNGLIO DA RAIZ DORSAL
101
4.13 EXPRESSÃO GÊNICA PARA SP NA MEDULA ESPINHAL E NO
GÂNGLIO DA RAIZ DORSAL
104
5 DISCUSSÃO 107
6 CONCLUSÃO 118
REFERÊNCIAS 120
ANEXO 137

20
INTRODUÇÃO

21
1 INTRODUÇÃO
1.1 CÂNCER
No Brasil, o câncer é um problema de saúde pública e, por isso, seu controle
e prevenção deve ser priorizada no país. O número estimado de canceres para
2014/2015 é de aproximadamente 576 mil casos novos no Brasil. Em homens, os
tipos mais incidentes aproximadamente são os canceres de próstata (69 mil),
pulmão (16 mil), cólon e reto (15 mil), estômago (13 mil) e cavidade oral (11 mil); e
nas mulheres, os de mama (57 mil), cólon do útero (15 mil), pulmão (11 mil), cólon e
reto (17 mil), e glândula tireóide (8 mil) (INCA, 2014).
Essa também ressalta as mudanças no perfil demográfico brasileiro nos últimos anos, denominado de “envelhecimento” da população, que, junto com a transformação nas relações entre as pessoas e seu ambiente, mostrou uma alteração importante no perfil de morbimortalidade, diminuindo a ocorrência das doenças infectocontagiosas e colocando as doenças crônico-degenerativas como novo centro de atenção dos problemas de doença e morte da população brasileira (INCA, 2014, p.63).
As estimativas de câncer são de grande importância, pois a partir delas,
ações para o controle dos tipos de câncer mais esperados podem ser planejadas e
tanto os serviços de saúde quanto os profissionais em saúde podem se preparar
para oferecer as diversas modalidades de atendimento à população.
Nos últimos anos, em nível mundial, tem-se presenciado grandes progressos
no tratamento do câncer. O que traz grande estresse para a pessoa com câncer e
sua família são os efeitos colaterais que são acentuados e são grande desafio para
os profissionais de saúde. Dentre as modalidades de tratamento, encontra-se o uso
de antineoplásicos, cuja toxicidade pode desencadear inúmeros sintomas
estressantes, comprometendo seriamente a qualidade de vida do paciente.
Com a descoberta da solução de Fowler (arsenito de potássio) por Lissauer,
no final do século XIX (1865), e da toxina de Coley (associação de toxinas
bacterianas), em 1890, os agentes antineoplásicos começaram a ser estudados e
utilizados. Porém, foi durante a Segunda Guerra Mundial, após a explosão de um
depósito de gás mostarda em Bari na Itália (1943), que foi observado, pela primeira

22
vez, mielodepressão intensa e morte por hipoplasia de medula óssea entre soldados
acidentalmente expostos (BONASSA; SANTANA, 2005; ROCHA; MARZIALE;
ROBAZZI, 2004).
O gás de mostarda provocou alterações rápidas na medula óssea, para tanto
deveria ter ação similar em células neoplásicas. Após essa observação, iniciaram-se
estudos clínicos para a sua utilização no tratamento do câncer. Em 1940, vários
pacientes foram tratados com esse agente, por via endovenosa, principalmente os
com linfomas avançados. A experiência levou pesquisadores a buscarem outras
substâncias que pudessem ter efeitos semelhantes contra o câncer, pois a melhora
foi espantosa, porém temporária. Como resultado, muitas outras drogas têm sido
desenvolvidas desde então (ACS, 2009).
1.2 FARMACODINÂMICA E FARMACOCINÉTICA DA OXALIPLATINA E O
MECANISMO DE AÇÃO ANTITUMORAL
Dentre as modalidades de tratamento para o câncer encontra-se o uso de
antineoplásicos, cuja toxicidade pode desencadear inúmeros sintomas,
comprometendo seriamente a qualidade de vida do paciente. A oxaliplatina (OXL) é
a terceira geração de compostos platinos amplamente utilizados como um fármaco
de primeira linha em quimioterapia de câncer gastrointestinal, em particular colorretal
metastático que tem atual importância epidemiológica mundial, visto que, a cada
ano, muitos novos casos são diagnosticados em todo o mundo. A principal
toxicidade limitante da dose de OXL é uma neuropatia que pode manifestar-se em
mais de 60% dos pacientes tratados (KAGIAVA et al., 2015).
Este fármaco foi desenvolvido na década de 70 como um dos vários
compostos platinos 1,2-Diamino-Ciclo-Hexano (DACH) na tentativa de se obter
análogos com índices terapêuticos mais favoráveis (STEIN; ARNOLD, 2012). Esses
compostos 1,2-DACH provocaram um entusiasmo imediato por não apresentarem
resistência cruzada com a cisplatina e carboplatina e por terem um perfil pré-clínico
distinto dos análogos mais antigos. Entretanto o interesse nesses agentes diminuiu
após a constatação dos severos efeitos tóxicos neurológicos nos estudos de fase I
(SAIF; READORN, 2005; SHAH; DIZON, 2009; STEIN; ARNOLD, 2012).

23
As diversas combinações quimioterápicas, principalmente as associadas à
OXL para a terapêutica cancerígena, são extremamente úteis visto a gravidade
dessa patologia. As estimativas de câncer têm sua importância, pois ações para o
controle podem ser planejadas e tanto os serviços de saúde quanto os profissionais
em saúde podem se preparar para oferecer as diversas modalidades de
atendimento à população.
Nos últimos anos, em nível mundial, tem-se presenciado progressos no
tratamento do câncer, porém um dos fatores que geram e comprometem o
tratamento para a pessoa com câncer e sua família são os efeitos colaterais que são
acentuados e são um grande desafio para os profissionais de saúde.
A neuropatia periférica é um efeito colateral de diversas classes de
antineoplásicos, como os taxanos (p.e., paclitaxel e docetaxel), os alcaloides da
vinca (p.e., vincristina e vimblastina) e principalmente os derivados de platina (como
a OXL) (SISIGNANO et al. 2014). Esses fármacos causam alterações celulares e
moleculares que interferem na condução e interpretação de impulsos dolorosos
(CAROZZI et al., 2014).
Com a chegada da OXL o valor gasto no mercado para tratamento de câncer
colón retal metastático foi de mais de US$ 1,6 bilhão até 2012. Estudos de Fase III
de uso de OXL também estão em andamento no câncer de pâncreas e câncer
gástrico. O sucesso desses tratamentos tende a aumentar significativamente o valor
gasto com OXL (GRAHAM; MUHSIN; KIRKPATRICK, 2013).
Quanto ao seu mecanismo de ação antitumoral sabe-se que a OXL é
extensamente distribuída por todas as células do corpo, com volume de distribuição
mais elevado que a cisplatina. Ao final da perfusão, 50% da platina (pt) estão fixados
nos eritrócitos e 50% encontram-se no plasma, sendo que 25% na forma livre e 75%
ligados às proteínas plasmáticas. A ligação às proteínas aumenta progressivamente,
estabilizando-se em 95% no quinto dia após a administração (CHU, 2004;
INFARMED, 2010).
A OXL tem rápida captação pelos tecidos e é muito reativa. A eliminação é
bifásica, com meia vida terminal de cerca de 40 horas (h). Sua meia-vida plasmática
é curta (0,2 a 0,3h), média (15 a 16h) e longa (252 a 273h). As concentrações
plasmáticas máximas variam de 1 a 1,5µg de platina/ml para pacientes que recebem
de 80 a 130mg/m2 por via intravenosa (IV) (BRUCE et al., 2007). Um máximo de

24
50% da dose administrada é eliminado na urina em 48h e 55% ao fim de 6 dias. A
excreção fecal é pequena (5% da dose ao final de 11 dias) (INFARMED, 2010).
Quanto à distribuição, é largamente dispersa na maioria dos tecidos, com um
volume mais elevado do que a cisplatina, devido aos metabólitos de OXL, que se
ligam irreversivelmente a proteínas, ácido desoxirribonucleico (DNA) e outras
moléculas celulares. Após duas a cinco horas de infusão de OXL, 40% do platino se
liga de forma irreversível às hemácias, formando produtos não tóxicos. Os 30%
restantes se ligam às proteínas do plasma, e os outros 30% correspondem ao
platino ultrafiltrado. Esse último possui a droga ativa, ou seja, a OXL intacta,
representando o maior componente citotóxico (CHU, 2004; JERREMALM; WALLIN;
EHRSSON, 2009).
Com relação ao metabolismo, a OXL faz uma extensa conversão não
enzimática para ativar espécies citotóxicas, semelhante ao observado com cisplatina
e carboplatina. Essa é uma reação aquosa que ocorre na presença de cloreto e
água, e as espécies formadas Pt (DACH)Cl, Pt (DACH)Cl2 e o complexo de OXL
desidratado, sendo que esses são mais citotóxicos do que a OXL dentro da célula
(JERREMALM; WALLIN; EHRSSON, 2009).
A ligação dos metabólitos da OXL ao DNA resulta em inibição da síntese e da
função, bem como a inibição da transcrição (Figura 1). Apesar da lesão ao DNA pela
OXL aparentar semelhança com a lesão provocada por cisplatina e carboplatina, os
metabólitos 1,2-DACH-Pt produzidos pela OXL são significativamente mais
citotóxicos do que os destes fármacos. Os nucleófilos fracos como o bicarbonato de
sódio (HCO3-) ou dihidrogênio fosfato (H2PO4) do meio intracelular desacoplam o
grupamento oxalato resultando na formação de intermediários não estáveis que são
rapidamente hidrolisados a espécies platinas DACH-pt (PARK et al., 2008).
Consequentemente, os adutos da OXL tornam-se mais eficientes em inibir a síntese
de DNA, sendo geralmente mais citotóxicos ocorrendo apoptose celular
(FOLTINOVÁ et al., 2008).
As interações da platina com o DNA, através da formação de complexos Pt-
DNA, principalmente na posição N7 das bases de guanina no DNA originam ligações
cruzadas inter e intrafilamentares (CHU, 2004; FOLTINOVÁ et al., 2008). Esses
complexos podem interferir na duplicação do DNA através do bloqueio da DNA
polimerase inibindo a transcrição do ácido ribonucleico (RNA), resultando em
bloqueio do ciclo celular e do reparo do DNA (CHO et al., 2008; NADIN et al., 2006).

25
Os complexos Pt-DNA, também, afetam o metabolismo e a organização espacial
celular, a bainha de mielina, os corpos celulares, as células da glia e os axônios
(STILLMAN; CATA, 2006).
FIGURA 1 - Mecanismo de ação antitumoral da Oxaliplatina.
Fonte: AUTOR, 2015
Legenda: Diamino-ciclo-hexano (DACH); Reaparo de má combinação (MMR)
Essas exposições das células tumorais à OXL induzem apoptose celular
programada (LOWE; LIN, 2000; GOURDIER et al., 2002; JOHNSTONE; RUEFLI;
LOWE, 2002). As cascatas de sinalização que conduzem à apoptose podem ser
divididas em dois grandes grupos. A via intrínseca que é caracterizada pelo papel
central das mitocôndrias na iniciação da cascata de caspase (DESAGHER;
MARTINOU, 2000). E a via extrínseca, onde a ativação de caspase é iniciada por
receptores de apoptose na superfície da célula. Os agentes quimioterapêuticos são
conhecidos por induzir apoptose por qualquer um destes dois mecanismos (LOWE;
LIN, 2000; JOHNSTONE; RUEFLI; LOWE, 2002).
Como citado anteriormente, a OXL está associada com graves efeitos tóxicos.
A neurotoxicidade da OXL é muito comum em pacientes tratados com o fármaco,
com 68% passando por algum grau de toxicidade. A toxicidade limitante da dose é
exacerbada pela exposição ao frio ou em doses maiores do que 135mg/m2, com o
desenvolvimento precoce da hipernocicepção a estímulos frios e quentes (ATTAL et
al., 2009).

26
Evidências sugerem que o 1,2-DACH-pt formado a partir da OXL liga-se de
forma mais potente e também é capaz de escapar do reconhecimento do complexo
de enzimas de reparo de má combinação (MMR mismatch repair) que é uma
correção de bases mal pareadas do DNA com maior afinidade por adutos de OXL
guanina-guanina (ZAANAN et al., 2013).
A replicação e posterior separação do DNA são inibidas, da mesma forma
que, secundariamente, é inibida a síntese do RNA e das proteínas celulares. O fato
dos adutos de OXL escaparem do complexo MMR explica a sua excelente atividade
contra tumores que exibem complexos MMR aberrantes, como é o caso dos
carcinomas colorretais e também explica um efeito melhor do que a cisplatina em
tumores sensíveis a essa droga, assim como tumores refratários à cisplatina
possivelmente respondem bem a OXL (CHU, 2004; JERREMALM; WALLIN;
EHRSSON, 2009).
Cada ciclo de quimioterapia dura em geral algumas semanas, mas, no caso
da OXL, durante esses ciclos, pode haver a instalação de um efeito colateral, que
muitas vezes leva a alterar a dose para mais ou menos que é a neuropatia sensitiva
periférica (NSP) (ARETI et al., 2014).
1.3 TOXICIDADE POR OXALIPLATINA
A OXL pode causar dois tipos de sintomas neuropáticos graves: uma
síndrome de dor aguda que desaparece dentro de poucos dias após o tratamento, e
uma neuropatia sensorial distal crônica que ocorre em fases posteriores durante o
tratamento. Os mecanismos celulares da NSP induzida por OXL diferem dos
mecanismos da indução por paclitaxel e vincristina. A OXL tem efeitos neuronais
mais severos, tais como aumento da atividade dos canais iônicos dependentes de
voltagem (PARK et al., 2008).
As toxicidades agudas mais comuns são: laringoespasmo (onde cerca de 1-
2% dos pacientes apresentam dificuldade de respiração e deglutição), náuseas,
vômitos, diarreia e reações de hipersensibilidade. Essa toxicidade aguda é rápida,
transitória, e pode ocorrer durante a infusão ou horas depois. Ocorre em 85-95% dos
pacientes que fazem tratamento com OXL sendo caracterizada pela exacerbação ao

27
frio. Pode ser manifestada toxicidade motora e sensorial que consistem em:
parestesia e/ou disestesia em extremidades, miotomias, espasmos musculares e
fasciculações na região oral (MARSHALL, 2004; SCHIFF; WEN; VAN DEN BENT,
2009).
Os pacientes descrevem as parestesias como sensações de formigamento
nas mãos, pés, garganta e região perioral. É um distúrbio neurosensitivo comumente
provocado pela alteração de sensibilidade na área nervosa atingida, ou seja, em
virtude de lesão do tecido nervoso num dado segmento sensitivo (BENATTI;
GUIMARÃES, 2009). Seu principal sintoma é a ausência de sensibilidade na região
afetada. Em estágios mais evoluídos o paciente poderá relatar alteração de
sensibilidade ao frio, calor e dor, sensação de dormência, choque e prurido (ROSA;
ESCOBAR; BRUTO, 2007). Esses sintomas apresentam início rápido, podendo ser
reversíveis dentro de algumas horas ou alguns dias.
Em geral, os sintomas da NSP aparecem nos membros inferiores (MMII) e em
seguida nos membros superiores (MMSS). Inicialmente do centro dos pés,
tornozelos e, depois, panturrilhas e progridem para os dedos das mãos, palmas e
punhos (SMITH; BECK; COHEN, 2008).
Estudos mostram que a neuropatia em MMSS, em mulheres, gera sensação
de frustração devido à dificuldade de colocar adornos como brincos ou colares. Além
disso, homens e mulheres queixam-se da dependência no momento de se vestir, e
muitos deixam de usar roupas que exigem atividades motoras finas. As sensações
de dor e parestesia nos pés levam a mudanças nos tipos de calçados, suspensão do
uso de tapetes em casa, além de dificuldades para correr, andar de bicicleta ou
permanecer de pé por períodos prolongados. Alguns relatam isolamento pela
dificuldade de andar ou dirigir, fato que impacta significativamente nas atividades
laborais, sociais e na prática de atividades físicas. Impaciência, raiva, cansaço,
frustração e depressão são sentimentos frequentemente relatados pelos pacientes
(BAKITAS, 2007).
Os sintomas crônicos mais comuns, não induzidos pelo frio, são as
disestesias e parestesias de extremidades que persistem entre os ciclos e
aumentam com o acúmulo das doses. Podem ocorrer ataxia, déficit em coordenação
sensório-motora fina e dor neuropática. São caracterizados pela persistência dos
sintomas com degeneração axonal sensorial distal, com perda de mielina que
envolve fibras grandes na ausência de um componente motor (CORIAT, 2014). Os

28
efeitos da neurotoxicidade dependerão de vários fatores diferentes tais como a dose
que uma pessoa foi exposta, as características da neurotoxicidade, da capacidade
para metabolizar e excretar e da capacidade para se recuperar depois de cessada a
aplicação do agente quimioterápico.
A neuropatia como efeito colateral foi descrita há 30 anos com a droga
cisplatina e, a partir de então, não saiu mais do cenário oncológico (VELASCO;
BRUNA, 2010). Essa toxicidade está associada a protocolos de tratamento que
envolvam derivados da platina e taxano atingindo cerca de 60% dos pacientes em
tratamento antineoplásico (CHAUDHRY et al., 1994; VELASCO; BRUNA, 2010).
As taxas de NSP induzidas por quimioterápicos disponíveis na literatura são
muito variáveis. Entre os fatores que podem influenciar essa variabilidade destacam-
se: o tipo de agente citostático; o regime de administração do antineoplásico (dose
total ou por ciclo, assim como o número de ciclos e o tempo de infusão); as
combinações de diferentes quimioterápicos; o tipo de tumor; as características dos
pacientes ou tratamento concomitante com outros fármacos neurotóxicos e a técnica
de diagnóstico ou os critérios de avaliação da NSP (VELASCO; BRUNA, 2010).
A neurotoxicidade pode ser limitada afetando apenas o sistema nervoso
central (SNC) ou o sistema nervoso periférico (SNP), sendo as neuropatias
periféricas as mais comuns. Outros fatores que podem interferir na incidência e
severidade da neurotoxicidade são a idade do paciente, a dose, a via de
administração, as funções renal e hepática, uso concomitante ou prévio de outras
drogas neurotóxicas, administração adequada ou não de agentes neuroprotetores
como cálcio e magnésio e o tratamento radioterápico concorrente do SNC
(BONASSA; SANTANTA, 2005).
A NSP associada à OXL é diferente daquela associada à cisplatina, embora
as duas causem sintomas em “meia e luva” que piora com o aumento da dose a que
o paciente está exposto sendo mais proeminente a toxicidade dose-limitante do
antineoplásico OXL, podendo causar sintomas substanciais que podem durar vários
anos. É a complicação neurológica mais comum do tratamento antineoplásico, e
afeta aproximadamente um terço dos pacientes submetidos a esse tipo de
tratamento.
A OXL não provoca os efeitos tóxicos renais e hematológicos e nem induz
ototoxicidade, comumente observados com cisplatina. Os efeitos eméticos também
são mais bem controlados. A neurotoxicidade, entretanto, apesar de ser na maioria

29
das vezes reversível, parece ser o efeito colateral que mais chama a atenção
(CORIAT t al., 2014).
Assim, a neurotoxicidade causada pela OXL pode ocorrer caracterizada por
neuropatia sensorial em fibras nervosas pequenas e grandes, o que leva a perda ou
alterações sensoriais importantes tais como diminuição da percepção do tato,
temperatura e dor, além da síndrome de Lhermitte’s (sensação similar a choque
elétrico que se irradia pelo membro). Há 28 alterações neurológicas descritas que
podem vir acompanhadas de dor ou deficiências funcionais (BONASSA; SANTANA,
2005; GAMELIN et al., 2002; LEONARD, et al., 2005).
Vários estudos evidenciam algum grau de disfunção neurológica periférica em
quase todos os pacientes que fazem uso da OXL e dependendo da intensidade em
que ocorrem, é recomendada a diminuição da dose em 25% a 50% ou mesmo a
interrupção do tratamento (MCWHINNEY; GOLDBERG; MCLEOD, 2009; VELASCO;
BRUNA, 2010).
Independente da intensidade foi demonstrado que a NSP induzida por
quimioterápicos interfere no estado de saúde geral e psicocognitiva dos pacientes,
tendo ainda um impacto financeiro, devido à incapacidade e a busca por tratamento
(HERSHMAN et al. 2011; MOLS et al. 2015).
Em um âmbito socioeconômico, poucos dados encontram-se disponíveis em
relação aos custos associados ao tratamento da NSP induzida por quimioterápicos.
O tratamento de pacientes com neuropatia custa, para cofres públicos, o triplo do
tratamento de pacientes que não apresentam essa condição (BERGER et al., 2004).
Pacientes com NSP induzida por antineoplásicos geram um gasto anual de US$
21.739,00 a mais do que os pacientes que não desenvolvem este efeito colateral
(PIKE et al., 2012).
Outros aspectos de impacto sobre as condições de vida levam em
consideração a presença de depressão ou ansiedade, transtorno do sono, questões
relacionadas ao trabalho, expectativas do tratamento, necessidades de reabilitação
e disponibilidade de apoio social da família e amigos (DWORKIN et al., 2003) e
também as co-morbidades psiquiátricas (SMITH; BECK; COHEN, 2008).
Apesar de a NSP ser um dos principais fatores limitantes nas dosagens dos
quimioterápicos, ainda não há um protocolo científico e clinicamente aceito para
tratar esta condição (BEIJERS; MOLS; VREUGDENHIL et al., 2014; HERSHMAN et
al. 2014). De fato, estudos clínicos (LOUKAPIS et al., 2014; NAKAYAMA 2015)

30
apresentaram dificuldades na manutenção das doses dos antineoplásicos em
consequência do desenvolvimento de dores de origem neuropática, o que poderia
causar uma diminuição do efeito terapêutico da quimioterapia.
A dor neuropática, por sua complexidade, é um desafio constante para os
profissionais da dor. O tratamento baseado no quadro doloroso não é, entretanto,
eficaz em vista da maior importância representada pelos mecanismos subjacentes à
expressão dolorosa. Contudo, o tratamento da etiologia obviamente deve ser
realizado buscando minimizar os mecanismos e sintomas, os quais serão o centro-
alvo do tratamento (ALVES NETO et al., 2009). Pode ser tratada com abordagens
psicológicas, físicas, cirúrgicas e farmacológicas como: anticonvulsivantes,
antidepressivos, opióides, antiarrítmicos, gabaérgicos, antagonistas de ácido N-
Metil-D-Aspartato (NMDA), bloqueios anestésicos, L-dopamina (DOPA), anti-
inflamatórios não esteroidais (AINEs) e neurolépticos (ALVES NETO et al., 2009).
Medicamentos de primeira linha como gabapentina, lidocaína, opióides,
tramadol e antidepressivos tricíclicos são muito utilizados, assim como os de
segunda linha lamotrigina, carbamazepina e inibidores seletivos da recaptação da
serotonina (DWORKIN et al., 2003). Da mesma forma, dois dos principais
moduladores de canais de Ca2+ (por se ligarem a uma subunidade α2δ) como a
gabapentina e a pregabalina também são utilizados (ALVES NETO et al., 2009;
GILRON, 2015). Atualmente, outros fármacos como Duloxetina (ENGEL, 2014) e
Venlafaxina (RICHARDS et al., 2015) são relatados na literatura para tratamento de
dor neuropática, porém o uso desses fármacos é de forma paliativa para amenizar a
dor da neuropatia. Para tal, surgiu o interesse em saber qual o envolvimento da
endotelina-1, de receptores TRPV1 e NMDA e da substância P na gênese dessa
importante afecção induzida pelo antineoplásico OXL com o intuito de trazer luz para
o desenvolvimento de novas drogas.
1.4 SISTEMA ENDOTELINA-1, RECEPTORES TRPV1 E NMDA E SUBSTÂNICA P
NA NEUROPATIA SENSITIVA PERIFÉRICA

31
A NSP é uma consequência frequente e incapacitante da administração de
OXL sendo o principal determinante de uma má qualidade de vida. Oitenta por cento
dos pacientes com câncer sentem uma forte dor em seus últimos dias de vida
(QUANG; SCHMIDT, 2010). A etiologia da dor do câncer envolve a sinalização por
meio de mediadores (como endotelina) entre as células cancerosas e os neurônios
sensoriais aferentes primários (PETERS et al., 2004; PICKERING et al., 2008;
SCHMIDT et al., 2008).
A ET-1 é produzida por vários canceres, incluindo o da próstata (NELSON et
al., 2000), mama (YAMASHITA et al., 1993), colorretal (INAGAKI et al., 1992;
SHANKAR et al., 1998), hepatocelular (KAR; YOUSEM; CARR, 1995), do pâncreas
(OIKAWA et al, 1994); endometrial (PEKONEN et al., 1992); pulmonar (AHMED et
al., 2000); feocromocitoma (OHNAKA et al., 1993) e carcinoma epidermóide de boca
(PICKERING; JORDAN; SCHMIDT, 2007).
A ET-1 foi descrita pela primeira vez em 1989, pelo grupo de pesquisa do
Prof. Sérgio Ferreira, em seu estudo injetaram ET-1 em ratos que apresentaram
hiperalgesia e ao administrarem por via intradérmica em seres humanos causou
respostas como de mácula e pápula, que foram acompanhadas por prurido. Estes
resultados indicam que a ET-1 pode desempenhar um papel na modulação da dor
(FERREIRA; ROMITELLI, NUCCI, 1989).
Existem dois tipos de receptores de endotelina denominados A (ETA) e B
(ETB). Ambos pertencem à superfamília dos receptores acoplados à proteína G. A
ET-1 ativa principalmente os receptores ETA. O RNA-mensageiro para o receptor
ETA é expresso em muitos tecidos humanos, incluindo o músculo liso vascular, nos
sistemas cardiovascular (BLACK et al., 2007; BOHM; PERNOW, 2007; CERNACEK
et al., 2003; RUSSELL; MOLENAAR, 2000), pulmonar (WATTS, 2009), renal
(GRANGER et al., 2006; KOHAN, 1996; NEUHOFER; PITTROW, 2006), nervoso
central (SCHINELLI, 2006) , gastrointestinal (HUANG, 2005), hipófise (LANGE et al.,
2002), adrenal (DELARUE et al., 2004; HINOJOSA-LABORDE; LANGE, 1999;
ROSSI et al.; 2002; ROSSI et al., 2000), nasal (MULLOL; PICADO, 2000), nervoso
periférico (DAMON, 1999; FUKURODA et al., 1996; MORTENSEN, 1999),
imunológico (MENCARELLI et al., 2009), hepática (MALLAT; LOTERSZTAJN, 1996),
geniturinárias (CAMERON et al., 1995) e endócrino (MEIDEN; LEVY, 2007).
Os receptores ETA são expressos principalmente por neurônios sensoriais de
diâmetro pequeno no gânglio da raiz dorsal (DRG), enquanto que a expressão de

32
receptores de ETB em DRG é restrita a células satélites da glia (POMONIS et al.,
2001). Os ETA são expressos em terminações periféricas do nociceptor, bem como
em axônios e em corpos celulares localizados do DRG (POMONIS et al., 2001)
sendo em uma proporção de 60:40, entre receptores ETA e ETB respectivamente
(PLANT et al., 2007).
O receptor ETB pode estar localizado nas células do musculo liso visceral
sendo um vasoconstrictor e no endotélio é vasodilatador. Os receptores de ETB nas
células endoteliais estimulam a produção de óxido nítrico (NO) e de prostaglandinas
(PGI2) e induz efeitos vasodilatadores e antiproliferativos (GHONEIM et al., 1993;
SAKURAI-YAMASHITA et al., 1997; SHETTY et al., 1993; SCHIFFRIN; TOUYZ,
1998). Também são encontrados nas células satélite da glia e células de Schwann,
mas não são nos nociceptores, podem sim, serem expressos em macrófagos,
queratinócitos na epiderme e derme da pele onde as terminações periféricas dos
nociceptores estão localizados (BERTI-MATTERA et al., 2006; POMONIS et al.,
2001).
No câncer, a ET-1 age nos receptores ETA sobre os nociceptores aferentes
primários que levam à dor (QUANG; BRIAN; SCHMIDT, 2010). O mecanismo de
nocicepção provavelmente envolve regulação por sódio e de tetrodoxina-resistente.
O mecanismo de antinocicepção deve envolver a regulação do receptor opióide de
proteína G por canais de potássio sobre nociceptores aferentes primários (HANSA;
SCHMIDT; STRICHARTZ, 2009).
Sabe-se que a ET-1 e as outras isoformas desencadeiam a dor em humanos
e respostas nociceptivas em animais (FERREIRA; ROMITELLI; DE NUCCI, 1989;
RAFFA et al., 1996). A ET-1 no SNP atua como um algógeno e está relacionada na
patogênese de uma variedade de estados de dor, tais como dor inflamatória, dor
neuropática e dor do câncer. As endotelinas no SNP podem ser expressas e
secretadas por células não neuronais, tais como células endoteliais (YANAGISAWA
et al., 1988), mastócitos (YAMAMOTO et al., 1994), monócitos e macrófagos
(EHRENREICH et al., 1990). Considerando que a literatura sugere que a Pro-ET-1 é
sintetizada por queratinócitos e neutrófilos, não é de se surpreender que o peptídeo
da endotelina tenha sido relacionado com inflamação.
Um inibidor não seletivo dos receptores de ET-1 é a Bosentana que foi o
primeiro antagonista de endotelina desenvolvido para os receptores. Vários ensaios
clínicos em longo prazo já estabeleceram a eficácia da bosentana oral (BRUNTON,

33
2012). Antagonistas seletivos dos receptores ETA e ETB, já foram isolados e incluem
respectivamente BQ-123 e BQ-788 que foram utilizados nesse estudo (Tabela 1)
Fonte: SMITH et al., 2014
Atualmente, a NSP induzida por OXL é tratada de forma empírica com drogas
que são utilizadas para tratamento de outros tipos de neuropatias. É sabido,
entretanto, que a neuropatia induzida por esse agente antineoplásico tem
características clínicas únicas importantes, como citado anteriormente, que suscitam
a real necessidade de um tratamento adequado e baseado em evidências
científicas. Na prática clínica, o que se tem observado é que o tratamento atual tem
tido pouco sucesso e que muitas vezes a neuropatia acaba persistindo dificultando
em demasia as atividades de vida diária, mesmo as mais básicas como caminhar ou
segurar objetos.
Há evidência crescente de que o eixo endotelina–receptores A e B
desempenham um papel no desenvolvimento de alodínia mecânica (tátil), em
condições de dor neuropática, como polineuropatia diabética e neuralgia do trigêmeo
(HANSA; SCHMIDT; STRICHARTZ, 2009), o que sugere que pode ter participação
no desenvolvimento da NSP em condições de lesão periférica ocasionado pelo
tratamento crônico com OXL. Para isso buscou-se identificar a relação entre o
receptor de potencial transiente vanilóide 1 (TRPV1) e receptores de NMDA e o
neuropeptídeo substância P (SP) na neurotoxicidade da OXL (Figura 2).
Receptor Agonistas Antagonistas
Receptor ETA ET-1 BQ-123, Atrasentana,
SB234551, ABT-627,
FR 139317, A-
127722.5, YM598
Receptor ETB ET-1, ET-2, ET-3, IRL-
1620, sarafotoxina S6c
BQ-788, A-192621
Receptores ETA/ETB ET-1, ET-2, ET-3 Bosentana
TABELA 1 – Agonistas e antagonistas dos receptores de endotelinas usados em
pesquisa básica e estudos clínicos.

34
FIGURA 2 – Modelo do envolvimento do Eixo endotelina–receptores na neuropatia sensitiva periférica induzida por oxaliplatina.
Fonte: AUTOR, 2015.
Legenda: endotelina-1 (ET-1); Receptor de potencial transiente vanióide 1 (TRPV-1); Ácido N-metil-D-aspartato (NMDA); Substância P (SP); Neuroquinina 1 (NK-1).
Tem sido relatado que cerca de 50% dos neurônios DRG que expressam o
TRPV1 também exibem imunoreatividade para receptores de ETA (PLANT et al.,
2006). Esses canais iônicos são conhecidos como TRPs que promovem a
sensibilização e ativação de neurônios sensoriais como os nociceptores (ASIA et al.,
2005). Com base na sequência de aminoácidos, os canais TRPs são divididos em
seis subfamílias TRPC (canonical), TRPM (melastatina), TRPV (vanilóide), TRPP
(Polistatina), TRPML (mucolipina), e TRPA (anquirina). São compostos de 700-2000
resíduos de aminoácidos e a abertura desses canais é seguida por fluxo de cátions
monovalentes (Na+, H+) e bivalentes (Ca2+, Mg2+) que causam a despolarização
(RAMSEY et al., 2006; GAUDET, 2009).
O TRPV é subdividido em seis membros que são: TRPV1, TRPV2, TRPV3,
TRPV4, TRPV5 e TRPV6 (ZHENG, 2013). Os canais TRPV estão envolvidos em
uma variedade de funções celulares incluindo sensação de dor e bom gosto, termo
regulação, tensão e manutenção da homeostase óssea, na regulação da água no
rim, motilidade gastrointestinal, digestão, contratilidade uterina e vasculite e
NSP/OXL x ET-1
Lesão Periférica
SP
Nk-1 NMDA
ET-1
TRPV1

35
permeabilidade celular (HOLZER, 2011; JULIUS, 2013; LIEBEN; CARMELIET,
2012).
O TRPV1 é um receptor vanilóide de capsaicina, que é ativado por estímulos
químicos dolorosos, por calor nocivo (por exemplo, a 43 °C) e inflamação
(CATERINA et al., 2000). É expresso principalmente em neurônios peptidérgicos, e
em menor grau nos nociceptores não peptidérgico. A expressão de TRPV1 aumenta
em vários estados de dor crônica humana (SZALLASI, A. et al., 2007). Em modelos
de câncer em ratos é regulado nos neurônios e também tem sido demonstrado que
o TRPV1 pode mediar tanto nocicepção mecânica quanto térmica (BEAUDRY;
DUBOIS; GENDRON, 2011; ASAI et al., 2005; SHINODA et al., 2008) sendo um
importante contribuinte para a dor e seu papel é mais complexo do que se tem sido
relatado na literatura.
Já se sabe que a OXL induz a expressão de TRPV1 e que as mudanças
funcionais contribuem para alterar a excitabilidade neuronal e a neuropatia. A
regulação positiva de TRPV1 reflete aumento na responsividade nos nociceptores
que contribuem para os mecanismos moleculares da hipernocicepção térmica e
mecânica, resultados já observados em camundongos tratados com cisplatina. É,
também, conhecido por desempenhar um papel importante na sensibilidade ao calor
e alguns dados sugerem que o aumento dos níveis de RNAm de TRPV1 que são
encontrados nos corpos de células nervosas sensoriais, poderiam resultar num
aumento dos nociceptores terminais periféricos (TA et al., 2010).
A sensibilidade e expressão de TRPV1 é modulada pelo fator de crescimento
neuronal (AMAYA et al., 2004) e fator neutrófico derivado de linha de células gliais
em roedores (ZHANG; HUANG; MCNAUGHTON, 2005) e nos seres humanos (ASAI
et al., 2005). A dor neuropática é causada por lesão do SNC ou SNP e para seu
tratamento, drogas visando os antagonismos dos receptores TRPV1 estão atraindo
interesse (SINISCALCO et al. 2005; MALMBERG; BLEY 2005; PREVARSKAYA et
al. 2007). Uma vez que tem sido demonstrada a expressão da atividade do TRPV1
nas fibras sensoriais em neuropatias que conduzem a hiperalgesia
(PUNTAMBEKAR et al., 2004; RASHID; INOUE; BAKOSHI, et al., 2003a; RASHID;
INOUE; KONDO, et al. 2003b). Também é expresso em estruturas cerebrais
superiores (CRISTINO et al., 2006), como a substância cinzenta periaquedutal e o
córtex (DE NOVELLIS et al., 2011).

36
Outra ação do TRPV1 é que após a sua a ativação há despolarização
neuronal, que leva à dor e liberação de neuropeptídeos sensoriais, tais como CGRP
e SP a partir das terminações nervosas centrais e periféricas. Os neuropeptídeos
ativam seus receptores de células efetoras e aumentam a sensibilização dos
nociceptores. Esta categoria neuronal expressa SP nas células DRG e em axônios
num grupo de células finamente mielinizadas (A delta), a maioria dos nociceptores
são mecanorreceptores de alto limiar (QUARTU et al., 2014).
A SP é um neuropeptídeo, que tem ação em um receptor que tem em sua
constituição sete domínios transmembrânicos, denominado neuroquinina-1 (NK-1) o
qual está acoplado à proteína G. É responsável por transmitir impulsos de dor para
medula espinhal e para o cérebro, produzir um impulso nervoso gerado que dilata os
vasos sanguíneos e fazer com que os fluidos e proteínas das células migrem para
fora (JANG et al., 2004; LEE; KIM, 2007). É um ligante natural com a afinidade mais
elevada para a NK-1 e a ação biológica da SP é mediada principalmente por este
receptor (MUÑOZ et al., 2015).
A ligação ao receptor NK-1 com a SP é muito importante para o controle de
movimento, para a percepção sensorial, a sobrevivência neuronal e degeneração,
motilidade gástrica, salivação, micção, inflamação, dor, depressão e na regulação do
sistema cardiovascular e dos mecanismos respiratórios (MUÑOZ et al., 2015). NK-1
é expresso em muitas linhagem celulares cancerígenas incluindo carcinomas do
colorretal (MUÑOZ et al., 2012; ROSSO et al., 2008), sendo demonstrado que há
aumento da expressão de RNAm para receptores de NK-1 em tecidos malignos
(SINGH et al., 2000).
O envolvimento do receptor NK-1 em modelo de dor neuropática foi citado em
alguns estudos sugerindo que a SP é responsável pelo desenvolvimento de
hiperalgesia em ratos (JANG et al., 2004; LEE; KIM, 2007) e quando esses
receptores são bloqueados há reversão do quadro (CAHILL; CODERRE, 2002).
Antagonistas do receptor NK-1 estão presentes em atividades antitumorais em
linhas celulares de câncer, estes antagonistas têm sido demonstrados por induzir a
apoptose dessas células (MUÑOZ et al., 2012). O antagonista de receptores de NK-
1, aprepitanto, é um agente não peptídico, bem tolerado, degradado por peptidases
e que pode atravessar a barreira hematoencefálica (BERGER et al., 2004; MUÑOZ
et al., 2012).

37
Outro receptor que pode estar envolvido na NSP induzida por OXL são os
receptores de NMDA, no qual sua sensibilização pode ser induzida com aumento da
plasticidade dendrítica (METZ et al., 2009). É um subtipo de receptores do
glutamato, localizado pré e pós-sinapticamente no DRG da medula espinhal e tem
um papel crucial na transmissão da sinalização de dor. O glutamato é um importante
transmissor excitatório na medula e os receptores de NMDA são conhecidos por
estarem envolvidos em neuropatia dolorosa (MIHARA et al., 2011). A fosforilação
dos receptores de NMDA e um aumento subsequente na sua ativação nos terminais
pós-sinápticos podem contribuir para o desenvolvimento de dor neuropática induzida
por quimioterapia.
Foi demonstrado que os receptores de NMDA desempenham um papel
importante em estados de dor mais prolongados por melhorar, prolongar e alterar a
atividade na via nociceptiva na medula espinhal, pois parece ser responsável pela
hiperalgesia (AHMADI et al., 2014). Os nociceptores enviam seus axônios para o
DRG, onde seus terminais fazem sinapse em neurônios de segunda ordem
(ALMEIDA; ROIZENBLATT; TUFIK, 2004).
Assim, uma vez que o canal dos receptores de NMDA é bloqueado por
magnésio, o receptor NMDA e seu canal não participam da atividade normal dos
circuitos de dor, sob certas condições (WOOLF; SALTER, 2000), porém um
mecanismo possível para a hipersensibilidade em nociceptores é o estímulo térmico,
que por sua vez envia sinais, remove o magnésio dependente de voltagem dos
receptores NMDA permitindo mais rapidamente influxo de Ca2+ nos neurônios de
segunda ordem pós-sinápticos no caminho da dor. Isto por sua vez vai ativar o Ca2+
e a cascata de sinalização intracelular que conduzem à fosforilação do NMDA e
outros canais do receptor iônico, o que pode dar início a aumentos prolongados na
excitabilidade de neurônios da medula espinhal. Assim, os receptores NMDA,
também, têm demonstrado ser uma peça-chave na hipersensibilidade à dor
(WOOLF; SALTER, 2000; INTURRISI, 2005).
Antagonistas do receptor de NMDA parecem inibir a plasticidade neural
subjacente a algumas formas de tolerância à opióides, sensibilização e dependência
física, sugerindo que os receptores de NMDA estão envolvidos no desenvolvimento
de alterações do comportamento induzidas por opióides (TRUJILLO, 2000). No
entanto, a utilização dos antagonistas de NMDA pode ser limitada devido aos seus
efeitos colaterais de, por exemplo, dano neuronal (NIKOLAEV et al., 2012). Os

38
antagonistas são altamente eficazes na redução da dor neuropática, esses agentes
causam efeitos secundários graves em doses terapêuticas, o que limita as suas
aplicações clínicas (CHEN et al., 2009).
Muitos pesquisadores têm explorado diversos outros mecanismos envolvidos
no desenvolvimento da NSP por OXL, a fim de promover uma gestão eficaz dessa
condição (JAGGI; SINGH, 2012). Por isso tentou-se relacionar os receptores
(TRPV1 e NMDA) e o neuropeptídeo SP com a neurotoxicidade sensitiva periférica
da oxaliplatina.
1.5 RELEVÂNCIA E JUSTIFICATIVA
É de conhecimento como as estatísticas de câncer são elevadas e ao
considerar o êxito do tratamento do câncer colorretal com a utilização da oxaliplatina
como antineoplásico de 3ª geração faz-se necessário investigar os mecanismos
envolvidos no principal efeito colateral desse tratamento cuja incidência compromete
o tratamento do câncer. Pois mesmo sendo muito eficaz a toxicidade desse fármaco
é muito limitante ao paciente que muitas vezes interrompe a terapia, para tal busca-
se a prevenção.
Essa pesquisa tem sua relevância em vista que a neurotoxicidade induzida
pela oxaliplatina é extremamente inconveniente podendo causar efeitos prejudiciais
à qualidade de vida do paciente e levando a redução da dose ou até mesmo à sua
descontinuação do uso do quimioterápico, o que em última instância pode trazer
prejuízos ao tratamento do câncer como um todo. Levando-se em consideração o
bom êxito da utilização da oxaliplatina no tratamento do câncer colorretal faz-se
necessário investigar os mecanismos envolvidos na dor neuropática associada à
oxaliplatina para preveni-la ou tratá-la.
Recentemente foi desenvolvido um modelo experimental de NSP por OXL,
objeto de estudo de minha dissertação de mestrado no Laboratório de Farmacologia
da Inflamação e do Câncer (LAFICA) do Departamento de Fisiologia e Farmacologia
da UFC. O modelo foi desenvolvido em camundongos dado ao fato de serem mais
econômicos, tanto no que diz respeito à quantidade de drogas e reagentes gastos

39
no estudo, quanto aos gastos com a manutenção dos animais e dada à possibilidade
de podermos utilizar futuramente animais nocaute para diversos fatores, pois
camundongos são os mais utilizados no desenvolvimento de animais geneticamente
modificados.
No entanto, após o desenvolvimento do modelo questionou-se: quais seriam
os principais mecanismos envolvidos na gênese dessa NSP? De acordo com nossos
estudos preliminares utilizando técnicas de imunohistoquímica observamos que
receptores como NK-1 e NMDA pareciam estar envolvidos no fenômeno. Estes
receptores, principalmente SP, foram expressos na periferia, mesmo após
administração sistêmica de OXL.
Para tanto, havia a necessidade de um estudo aprofundado de receptores e
neuropeptídeos envolvidos na NSP induzida pelo agente antineoplásico OXL em
camundongos, no sentido de esclarecermos o envolvimento e fornecer uma base
para a prevenção e o tratamento adequado desta importante e incapacitante afecção
clínica.
Vários estudos em animais de laboratório e em humanos sugerem o
envolvimento da endotelina na patogênese da dor. Ambos os subtipos de receptores
têm demonstrado ter participação na nocicepção induzida por ET-1, porém não se
sabe os mecanismos através dos quais a ET-1 induz a sensibilização periférica ou
central, o envolvimento com os receptores TRPV1, NMDA e o neuropeptídeo SP e
nem as suas contribuições para o desenvolvimento de nocicepção em condições
cancerosas. No intuito de desvendar os mecanismos envolvidos nessa neuropatia
justifica-se esse estudo, pois é importante identificar o envolvimento da endotelina,
de receptores e da substância P na NSP induzida por OXL, tendo como base esses
resultados preliminares.

40
OBJETIVOS
2 OBJETIVOS

41
2.1 OBJETIVO GERAL
Investigar o papel dos receptores endotelina 1 (ETA e ETB), de receptores
TRPV1 e NMDA e da substância P na neuropatia sensitiva periférica induzida pelo
agente antineoplásico oxaliplatina.
2.2 OBJETIVOS ESPECÍFICOS
Esclarecer a participação dos receptores de endotelina-1 (ETA/ETB) na
patogênese da neuropatia sensitiva periférica induzida pelo agente antineoplásico
oxaliplatina através da avaliação comportamental e imunomarcação para cFOS,
receptor NK-1 e receptor ETB e expressão gênica por PCR;
Estudar o envolvimento do receptor somatossensorial de calor (TRPV1) na
patogênese da neuropatia sensitiva periférica induzida pelo antineoplásico
oxaliplatina através da avaliação comportamental e expressão gênica por PCR;
Estudar o envolvimento do neuropeptídeo Substância P (SP) na patogênese
da neuropatia sensitiva periférica induzida por oxaliplatina através da avaliação
comportamental e imunomarcação para NK-1 e expressão gênica por PCR;

42
Estudar a participação do receptor do ácido N-Metil-D-Aspartato (NMDA) na
patogênese da neuropatia sensitiva periférica induzida pelo antineoplásico
oxaliplatina utilizando testes nociceptivos;
Verificar a expressão gênica por PCR dos receptores de endotelina ETA e
ETB, TRPV1 e da SP nos tecidos (medula espinhal e gânglio da raiz dorsal) em
animais com neuropatia sensitiva periférica induzida por oxaliplatina.

43
MATERIAIS E MÉTODOS
3 MATERIAIS E MÉTODOS

44
3.1 ANIMAIS
Para a realização dos experimentos foram utilizados camundongos Swiss
adultos (Mus muscullus), adultos (n=6, por grupo), machos, pesando 20 gramas(g),
provenientes do Biotério Setorial do Departamento de Fisiologia e Farmacologia da
Universidade Federal do Ceará (UFC). Os animais foram acondicionados em caixas
de polipropileno, à temperatura ambiente, com ciclos de claro/escuro de 12 em 12h,
recebendo ração padrão (Purina Chow) e água “ad libitum”.
Os experimentos foram desenvolvidos no Laboratório de Farmacologia da
Inflamação e do Câncer (LAFICA) da UFC e no Laboratório de Inflamação e Dor da
Faculdade de Medicina de Ribeirão Preto (SP).
3.2 ASPECTOS ÉTICOS
Os protocolos experimentais foram realizados de acordo com os padrões de
uso de animais experimentais e esse projeto foi encaminhado para apreciação pelo
Comitê de Ética em Pesquisa com Animais (CEPA) da UFC, e foi aprovado com o
protocolo número 75/2012. Todos os experimentos foram realizados no sentido de
reduzir o número de animais, a dor, o sofrimento e o estresse dos mesmos
(ANEXO).
3.3 AMBIENTE

45
Os camundongos foram colocados, em número aproximadamente de 20
animais, em gaiolas de plástico medindo 40 centímetros (cm) de comprimento, 31cm
de largura e 17cm de altura. O fundo destas gaiolas foi coberto por raspas de
madeira e o teto consistia em grade de metal com um espaço para serem colocados
comida e água. Os animais foram deixados em ambiente com temperatura de 25ºC,
com exaustão de ar, ciclo claro-escuro de 12h e acesso à agua e comida ad libitum.
Os animais foram alimentados com ração Fri-lab camundongos, tal ração leva
composto de milho moído, farelo de arroz desengordurado, farelo de arroz integral,
farelo de girassol, fosfato monoamônico. Cada 100g de ração contém 23% de
proteína, 22% de água, 4% de extrato etírico, 8% de fibra, 11% de minerais (ferro,
cobalto, cobre, manganês, zinco, iodo), 1% de cálcio, 0,45% de fósforo e vitaminas
(A, B1, B2, B6, ácido nicotínico, ácido pantotênico, C, D3, E, potássio e folato).
3.4 HORÁRIO DOS EXPERIMENTOS
Todos os experimentos, observações clínicas e comportamentais foram
realizados entre 8 e 18h. Os animais foram tratados e testados uma vez por semana
durante dois meses para um dos estímulos no mesmo horário em todos os testes
realizados.
3.5 OBSERVAÇÕES FÍSICAS
Diariamente foram realizados exames clínicos atentando para trofismo
muscular, coloração e aspecto do pelo, cor dos olhos, presença de parasitas, sinais
de infecção local ou generalizada, marcha e sinais de autonomia.
3.6 APARELHOS E INSTRUMENTOS LABORATORIAIS

46
• Alicate;
• Aparelho Von-Frey eletrônico (Insight®);
• Balança analítica (Sartorious modelo BL2105);
• Balança para pesagem de animais modelo ID-1500 (Filizola®);
• Bisturi;
• Capela de fluxo laminar, vertical (modelo TROX do Brasil);
• Cassetes;
• Confocal (LM 710 Zeiss);
• Cronômetro;
• Eppendorf (1ml);
• Equipo (Embramed®);
• Estufa de cultura celular com atmosfera de 5% de CO2;
• Filtro estéril;
• Grade para eppendorf;
• Grade para tubo falcon;
• Lâminas para imunohistoquímica (Fisherbrand);
• Material cirúrgico;
• Micropipetas automáticas (GILSON);
• Micrótomo Olympus;
• Pipetas automáticas de 200 e 1000;
• Ponteiras para as pipetas automáticas estéreis (SIGMA);
• Seringas de 1 e 5ml (BD Plastipak);
• Sonicador (THORNTOW – TT7);
• Termômetro;
3.7 DROGAS, SOLUÇÕES E OUTROS LÍQUIDOS
• Água destilada;
• Água filtrada;
• Albumina bovina (BSA a 1% Sigma-Aldrich®);
• Álcool à 70% (Dinâmica);
• Anticorpos para imunohistoquímica (Santacruz Biotecnology®);
• Aprepitanto (125mg, EMEND; Merck Research Laboratories®);
• Bosentana (125mg, Tracleer® Acetilom);

47
• BQ-123 (200UG, Sigma-Aldrich®);
• BQ-788 (200UG, Sigma-Aldrich®);
• Capsazepina (5mg, Sigma-Aldrich®);
• Carboximetilcelulose Sal Sódico P.A. (CMC, Synth®);
• Dimetil sulfóxido (DMSO, Dinâmica);
• Eosina (Merk Research Laboratories);
• Éter etílico P.A. (Dinâmica – Reagentes Analíticos);
• Formol à 10% (Dinâmica);
• Glicina Sigma-Aldrich®);
• Hematoxilina (Reagen);
• Ketamina (Zoovet®);
• Metanol (Dinâmica);
• MK-801 (Sigma-Aldrich®);
• Oxaliplatina (5mg, Sigma-Aldrich ®);
• Paraformaldeído (PFA 4%, Sigma-Aldrich®);
• Solução glicosada (D-glucose anidra – dextrose synth, Dinâmica);
• Solução salina estéril (NaCl 0,9%, Farmace);
• Solução salina tamponada (PBS 0,1M, Dinâmica);
• Soro fetal bovino (Sigma-Aldrich®);
• Tampão fosfato-salino (PBS, 0,01M (Dinâmica);
• Tampão Hanks com HEPES (Sigma-Aldrich®);
• Tissue Tek® (Sakura, Torrance, CA, EUA)
• Triton X 100 0,1% (Sigma-Aldrich®);
• Tween® 80 (Sigma-Aldrich®);
• Xilazina (Zoovet®).
3.8 PROTOCOLO EXPERIMENTAL E DESENHO DO ESTUDO
48
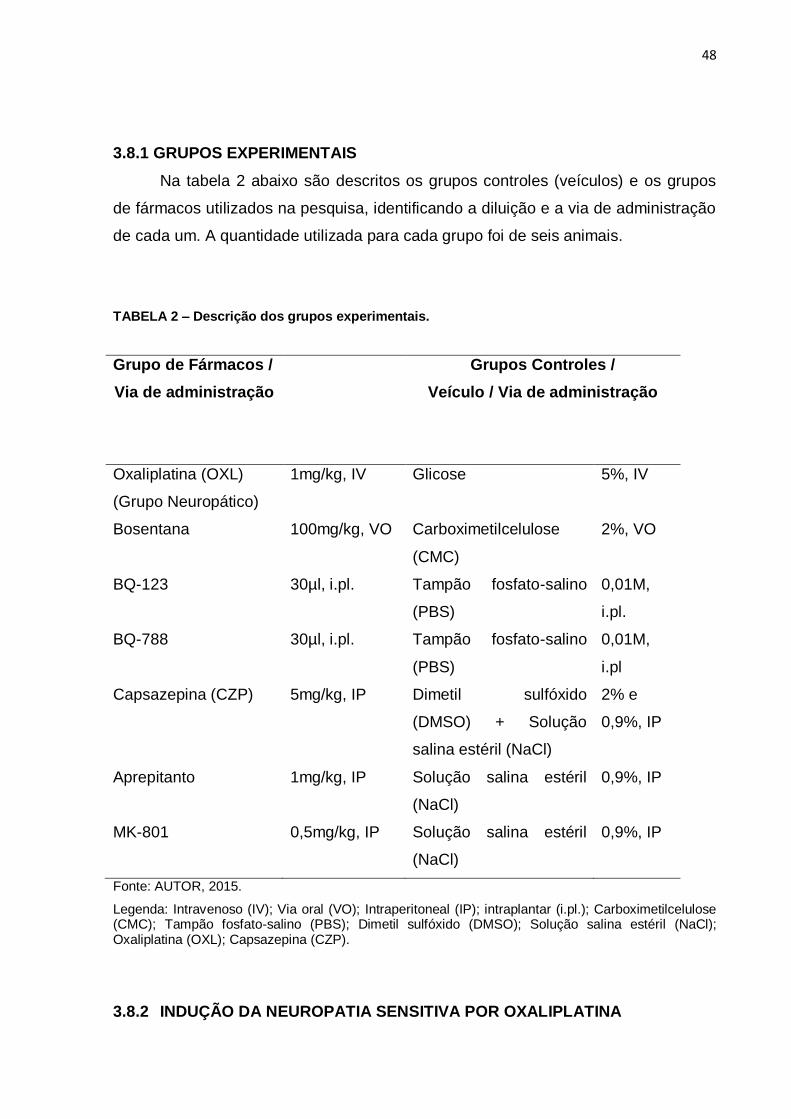
3.8.1 GRUPOS EXPERIMENTAIS
Na tabela 2 abaixo são descritos os grupos controles (veículos) e os grupos
de fármacos utilizados na pesquisa, identificando a diluição e a via de administração
de cada um. A quantidade utilizada para cada grupo foi de seis animais.
TABELA 2 – Descrição dos grupos experimentais.
Grupo de Fármacos /
Via de administração
Grupos Controles /
Veículo / Via de administração
Oxaliplatina (OXL)
(Grupo Neuropático)
1mg/kg, IV Glicose 5%, IV
Bosentana 100mg/kg, VO Carboximetilcelulose
(CMC)
2%, VO
BQ-123 30µl, i.pl. Tampão fosfato-salino
(PBS)
0,01M,
i.pl.
BQ-788 30µl, i.pl. Tampão fosfato-salino
(PBS)
0,01M,
i.pl
Capsazepina (CZP) 5mg/kg, IP Dimetil sulfóxido
(DMSO) + Solução
salina estéril (NaCl)
2% e
0,9%, IP
Aprepitanto 1mg/kg, IP Solução salina estéril
(NaCl)
0,9%, IP
MK-801 0,5mg/kg, IP Solução salina estéril
(NaCl)
0,9%, IP
Fonte: AUTOR, 2015.
Legenda: Intravenoso (IV); Via oral (VO); Intraperitoneal (IP); intraplantar (i.pl.); Carboximetilcelulose (CMC); Tampão fosfato-salino (PBS); Dimetil sulfóxido (DMSO); Solução salina estéril (NaCl); Oxaliplatina (OXL); Capsazepina (CZP).
3.8.2 INDUÇÃO DA NEUROPATIA SENSITIVA POR OXALIPLATINA

49
Os animais foram tratados por via intravenosa (IV) com OXL (1mg/kg)
dissolvida em solução glicosada estéril na veia lateral da cauda do camundongo
utilizando-se agulha de calibre 0,45 x 0,75cm em seringa de 1ml. Os animais
utilizados como controle receberam solução glicosada (5%) estéril na veia lateral da
cauda. Esse modelo utilizado foi o da minha dissertação de mestrado, no qual se
estabeleceu a curva de tempo para se chegar a essa dose.
No início do estudo os animais foram pesados (n=6) utilizando uma balança
de precisão digital Filizola® e aferido os testes nociceptivos em tempo zero. Em
seguida foi realizada a divisão dos grupos, cada grupo recebeu durante quatro
semanas e meia, nove administrações IV de OXL e o grupo controle com solução
glicosada (0,3ml/30g).
Foram realizados testes nociceptivos (descritos a seguir) concomitantemente
às administrações de OXL uma vez por semana durante 30 dias para avaliar o
desenvolvimento dos sintomas de neuropatia. Também foi realizado com esses
grupos imunofluorescência e RT-PCR.
3.8.3 AVALIAÇÃO DO ENVOLVIMENTO DA ENDOTELINA NA NSP INDUZIDA
POR OXALIPLATINA
Para verificação do envolvimento da endotelina na NSP induzida por OXL,
procedeu-se à administração dos antagonistas dos receptores ETA e ETB. Foi
administrado Bosentana (100mg/kg) diluída em CMC (2%) por via oral (VO) 30min
antes da administração de OXL para observação de ambos os receptores visto que
a Bosentana não é seletiva. Os animais utilizados como controle receberam solução
CMC (2%) VO.
Para análise específica administrou-se BQ-123 (antagonista de receptor de
endotelina ETA) e BQ-788 (antagonista de receptor de endotelina ETB) diluidos em
PBS e injetou-se (30µl), intraplantar (i.pl.), na pata direita de cada animal, em grupos

50
de tratamento separados, 30min antes da administração de OXL, obedecendo o
protocolo de duas injeções semanais, totalizando nove injeções, durante 4,5
semanas. Os animais utilizados como controle receberam PBS (0,01M, 30µl) intra-
plantar direita.
Em seguida foram realizados os testes nociceptivos descritos a seguir.
Posteriormente os animais foram eutanasiados para remoção da medula espinhal
lombar e DRG, para posterior análise de imunofluorescência.
3.8.4 AVALIAÇÃO DO DESENVOLVIMENTO DA NEUROPATIA SENSITIVA
PERIFÉRICA EM ANIMAIS TRATADOS COM ANTAGONISTAS DOS
RECEPTORES TRPV1, NK-1 E NMDA
Os camundongos submetidos à NSP induzida por OXL foram tratados por via
sistêmica, 30min antes de cada injeção de OXL, com antagonistas especificos:
capsazepina (antagonista do receptor TRPV1, 5mg/kg, IP), aprepitanto (antagonista
do receptor NK-1 da substância P, 1mg/kg, IP) e MK-801 (antagonista dos
receptores NMDA, 0,5mg/kg, IP). Os animais utilizados como controle receberam
solução salina (0,9%, IP).
Em seguida foram realizados os testes nociceptivos descritos a seguir. A
Figura 3 representa o protocolo experimental utilizado para a indução da NSP pela
OXL e para o uso dos antagonistas seletivos para receptores de endotelina, de
TRPV1, de NMDA e do neuropeptídeo SP.

51
FIGURA 3 - Protocolo experimental para o uso dos antagonistas seletivos para receptores de endotelina-1, TRPV1, NMDA e da substância P na indução da neuropatia sensitiva periférica induzida por oxaliplatina
Fonte: AUTOR, 2015.
Legenda: Medula espinhal (ME); Gânglio da Raiz Dorsal (DRG); Via oral (VO); intra-plantar (i.pl.);
intraperitoneal (IP); oxaliplatina (OXL).
IMUNO PCR
Bosentana (100mg/kg, VO)
ou CMC (2%, VO)
BQ-123 (30µl, i.pl.) ou PBS (30µl, i.pl.)
OXL (1mg/kg, IV, 9x) Grupo neuropático
ME
DRG
BQ-788 (30µl, i.pl.) ou PBS (30µl, i.pl.)
Capsazepina (5mg/kg, IP) ou DMSO (2%) + NaCl
(0,9%, IP)
Aprepitanto (1mg/kg, IP) ou NaCl (0,9%, IP)
MK-801 (0,5mg/kg, IP) ou NaCl (0,9%, IP)
30 min Antes
Testes nociceptivos 1x por semana
(0-56 dias)

52
3.8.5 TESTES PARA AVALIAÇÃO DE HIPERNOCICEPÇÃO MECÂNICA E
TÉRMICA
3.8.5.1 Teste de hiperalgesia mecânica plantar (Von Frey eletrônico)
A intensidade de hiperalgesia foi avaliada pelo limiar de sensibilidade de cada
animal a um estímulo mecânico produzido pela pressão gradual exercida por um
filamento rígido acoplado ao aparelho Von Frey (Insight®) que registra a pressão em
gramas exercida na ponta desse filamento.
A estimulação mecânica foi exercida na região plantar das patas traseiras. O
aparelho eletrônico Von Frey registra a pressão em gramas suficiente para provocar
uma reação descrita como uma flexão da pata seguida por um “flinch” após a
retirada da pata em contato com o aparelho (CUNHA et al., 2004). A hiperalgesia
mecânica (grau de sensibilidade ao estímulo mecânico) foi medida antes (tempo
zero) e após os tratamentos. Foi avaliado o limiar mecânico de cada animal.
Vale ressaltar que para a realização do teste nociceptivo Von Frey foi utilizado
o tipo eletrônico, pois o experimentador não precisa fazer a troca de filamentos de
calibres diferentes, como no Von Frey filamentos, já que o eletrônico registra os
diversos níveis de pressão como uma força em gramas que aparece no visor do
aparelho assim que o animal retira a pata em contato com o sensor. Cabe ao
experimentador interpretar se a retirada da pata foi uma reação de comportamento
nociceptivo ou não, o que depende da forma como ele retira a pata. Em nosso
estudo o mesmo pesquisador realizou a avaliação desse teste em todas as fases
dos experimentos.
3.8.5.2 Teste de imersão da cauda em água fria (10 ºC) ou aquecida (43 ºC)
Baseado no modelo de Authier et al. (2003) a cauda do camundongo foi
imersa em água fria mantida a temperatura de 10 ºC ou em água aquecida (43 ºC)

53
para testar a hipernocicepção, sendo contado o tempo de permanência até o
camundongo levantar a extremidade da cauda em contato com a água.
Outro teste realizado foi o de imersão da cauda que permite tanto o uso de
estimulos quentes como frios. Ficou mais conveniente na hora de comparar qual tipo
de estímulo térmico os animais tratados com OXL respondem com maior
intensidade. Fizemos uma modificação do modelo original no qual os animais são
imobilizados na mão do experimentador. Depois de fazermos várias tentativas de
registro do limiar nociceptivo normal de cada animal com esse tipo de imobilização,
percebemos que a imobilização pelo experimentador causa certo desconforto ao
animal que fica durante um bom tempo tentando fugir, causando reações falso-
positivas. Decidimos lançar mão de outro método de imobilização utilizando um
contensor de acrílico transparente. Com essa metodologia conseguimos realizar
registros de limiar mais fidedignos e que se repetiam.
Observamos que com o contensor de acrílico os animais, depois de alguns
minutos, se acostumavam e desistiam de tentar escapar ficando com a cauda
imóvel, o que permitiu melhores resultados. As temperaturas foram constantemente
mantidas e monitorizadas através do uso de caixas isolantes de isopôr e
termômetros.
Tanto os testes térmicos quentes e frios, assim como o teste mecânico foram
realizados em grupos de animais diferentes, de modo que cada animal foi submetido
somente a um tipo de teste, tentando minimizar o nível de estresse.
Observando todos esses cuidados, bem como o local de experimentação
silencioso e com exaustão e iluminação adequadas podemos garantir que nossos
resultados são fidedignos e com o mínimo de variáveis possíveis para um
experimento de comportamento evitando os vieses de interferência pessoal.
3.8.6 COLETA DO SEGMENTO MEDULAR E GÂNGLIOS DA RAIZ DORSAL
A coleta dos segmentos medulares e gânglios da raiz dorsal foi realizada
após o 56º dia. Os animais (camundongos) foram anestesiados com xilazina (20µl) e
quetamina (20µl/kg) por via intraperitonial (IP) e perfundidos via intracardíaca com
parafolmaldeído 4% em PBS 0,01M. A porção torácica do camundongo foi removida

54
para exposição do coração, então foi introduzida uma agulha descartável de calibre
0,70 x 25cm, sem o bisel, no ventrículo esquerdo cardíaco e foi realizado um
pequeno corte na aurícula direita, para escoamento das soluções. A agulha foi
conectada na seringa de 20ml com equipo simples (Embramed®), onde os animais
foram perfundidos via intracardíaca com solução salina (50ml) por 5min e solução
paraformaldeído (PFA) 4% por mais 5 min.
Após a perfusão, os animais foram fixados sobre uma superfície plana, na
posição em decúbito ventral para retirada do segmento medular e do DRG, como
segue o protocolo abaixo (Figura 4):
Ressecção da pele do dorso do animal com uma tesoura.
Retirada do tecido ao redor da coluna vertebral com pinça, limitada pelos
cortes de bisturi.
Discos vertebrais eram fragmentados com o auxílio de um alicate, até que a
medula fosse totalmente visualizada.
Dissecção da medula com alicate comum.
Observar o DRG na altura da crista ilíaca com a visualização do nervo ciático.
FIGURA 4 – Coleta do segmento medular e gânglio da raiz dorsal.
Fonte: AUTOR, 2015.
O segmento L4-L5 foi identificado pelo alargamento lombar e raízes nervosas.
Após a remoção, esse material foi colocado em gelo seco para congelar
Medula
Espinhal
Gânglio
da raiz
dorsal

55
rapidamente e em seguida colocado em cassetes e fixado na solução de formol PFA
4% por duas horas, seguido de crioproteção em sacarose 30% por um dia.
No dia seguinte, cada tecido foi colocado em um blister com OCT (tissu tek) e
congelado em freezer -80 ºC. Para montagem das lâminas, essas foram cortadas no
criostato (Crycout 1800) com temperatura entre -20 ºC ou –22 ºC (cortes de 10-15
micras) em lâminas silanizadas especiais para imunofluorescência.
Para realização do RT-PCR os animais foram perfundidos com solução salina
dando seguimento após anestesia à retirada da medula espinhal e do DRG. Os
tecidos foram colocados em eppendorf e congelados no freezer -80 ºC.
3.8.7 IMUNOFLUORESCÊNCIA EM SEGMENTO MEDULAR E GÂNGLIO DA
RAIZ DORSAL.
O ensaio de imunofluorescência foi realizado na seguinte sequência:
descongelar as lâminas em temperatura ambiente; fixação em metanol (2min);
hidratação em PBS 0,01M por 3 repetições (5min cada); depois foi realizada a
recuperação antigênica (95 ºC por 18min) no micro-ondas em citrato; resfriar por 20
minutos (min); lavagem em PBS por 3 repetições (5min cada); permeabilização com
Triton X 100 0,1% em PBS-BSA 0,1% (10min) etapa não realizada no anticorpo para
receptor de endotelina B; posteriormente foi realizada a lavagem das lâminas em
PBS por 3 repetições (5min cada); seguido por bloqueio com glicina 0,5M; as
lâminas foram então mergulhadas em uma solução com albumina bovina (BSA a
1%) durante 30min; lavagem em PBS por 5min; em seguida as amostras foram
incubadas em anticorpos primários (cFOS, receptores de endotelina B e NK-1) na
diluição de acordo com descrito na Figura 5 abaixo e ficaram em temperatura de 4
ºC durante uma noite (over night).
No dia seguinte, procedeu-se a seguinte sequência: lavagem das lâminas em
PBS por 3 repetições (5min cada); incubação com anticorpo secundário biotinilado
universal (LSAB – DAKO, anticorpo anti IgG de camundongo da Santa Cruz
Biotechnology) durante 1,5h em temperatura ambiente; lavagem em PBS por 3
repetições (5min cada); incubação do NeuN (1:100) por 1,5h; lavagem em PBS por
5min; colocado o 4´,6-diamidino-2-fenil-indol (DAPI), por 30min; em seguida as

56
lâminas foram montadas, examinadas e fotografadas no microscópio MO Confocal
LM 710 Zeiss.
A quantificação da área marcada nas fotos (pixels) foi realizada pelo
programa Fiji Image com os resultados representados por intensidade de
fluorescência no aumento de 100x, sendo analisados cinco fotos de cada lâmina de
medula espinhal. Foi diferenciado as áreas marcadas pela maior saturação de cor
associada à marcação (vermelha ou verde). Para isso, foi utilizado o procedimento
baseado na saturação da cor associada à marcação positiva para um determinado
marcador. Os limites necessários para definição de pixels marcados e não marcados
foram definidos previamente.
FIGURA 5 – Lista de anticorpos primários e seus respectivos secundários.
Anticorpo primário
(Diluição)
Anticorpo secundário
(Diluição)
NK-1 (1:400)
Rabit anti-NK1
Sigma
Goat anti-rabit (1:400) Alexa-fluor 594
Invitrogen
Endotelina B (1:200)
Rabit anti-EndB
Sigma
Goat anti-rabit (1:400) Alexa-fluor 568
Invitrogen
cFOS (1:200)
Rabit anti-cFOS
Santa Cruz
Goat anti-rabit (1:400) Alexa-fluor 568
Invitrogen
NeuN (1:100)
Anti-NeuN Alexa-fluor 488
Millipore
-
4´,6-diamidino-2-fenil-indol (DAPI),
(1:400)
Invitrogen
-
3.8.8 TRANSCRIÇÃO REVERSA - REAÇÃO EM CADEIA DE POLIMERASE (RT-
PCR) PARA SP, TRPV1, ENDOTELINA A E ENDOTELINA B DA MEDULA
ESPINHAL E NO GÂNGLIO DA RAIZ DORSAL

57
A expressão gênica foi realizada usando um sistema de detecção PCR em
tempo real (síntese de cDNA sobre um molde de RNA catalisada pela transcriptase
reversa, seguida de amplificação por meio de PCR, reação em cadeia de
polimerase). Foi realizada utilizando uma amostra de cDNA proveniente de um pool
de RNA total (produzido a partir de quantidades iguais de RNA extraído do gânglio
da raiz dorsal e segmentos medulares).
Todos os PCRs foram analisados pelo tempo real (CFX96 TouchTM PCR)
com instrumento de sistema de detecção de Bio-Rad (Bio-Rad iQTM SYBR® green
Supermix). Condições de ciclos térmicos incluiu uma passo inicial de desnaturação a
95° C durante 7 min seguido de 45 ciclos (cada ciclo começou com um passo de
desnaturação (20s a 95° C), uma temperatura de recozimento a 60° C durante 20s e
terminou com uma extensão de 45s a 72° C).
Primers DNA foram projetados com base em sequências de RNAm obtidas a
partir do National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov).
que foram utilizados para SP, TRPV-1, receptor de endotelia A e receptor de
endotelina B, como se segue na Figura 6:
FIGURA 6 – Sequência de primers utilizados no RT-PCR.
PRIMERS
SP
AGCCTTTAACAGGGCCACTTG
TRPV1
TCCCCAACGGTGTTATTCAG
Receptor de
Endotelina A
GGCTATTGCCCACAGCAGAC
Receptor de
Endotelina B
ACCTGCGAAATGCTCAGGAA

58
3.9 ANÁLISE ESTATÍSTICA
Os resultados foram expressos como média ± erro padrão da média (EPM).
Para comparação múltipla dos dados paramétricos dos testes comportamentais e
farmacológicos foi utilizado à análise de variância (one way - ANOVA) e o nível de
significância entre os grupos foram determinados pelos testes Newmann-Keuls para
detectar as diferenças entre o grupo controle e os grupos em tratamento.
O número (n) de animais foi de seis em cada grupo experimental. Em todas
as análises estatísticas, considerou-se o nível crítico para rejeição da hipótese de
nulidade menor que 5%. Os asteriscos indicam *p<0,05, **p<0,01 e ***p<0,001.

59
RESULTADOS

60
4 RESULTADOS
Evidências indicam que a endotelina-1 é ativada nos neurônios nociceptivos
após diferentes estímulos nocivos, mas o envolvimento dos mecanismos, como o
papel dos receptores da endotelina-1, de receptores (TRPV1 e NMDA) e da
substância P na patogênese da neurotoxicidade experimental induzida pelo agente
antineoplásico OXL ainda não foi totalmente compreendido, para tanto, buscou-se
fazê-lo de acordo com os resultados descritos abaixo. Para os testes nociceptivos
seguiu-se a metodologia já descrita:
RESULTADOS DOS TESTES NOCICEPTIVOS
Bosentana (100mg/kg, VO) BQ-123 (30µl, intra-plantar em
pata direita) BQ-788 (30µl, intra-plantar em
pata diteita) (2x por semana)
0º dia 30º dia 1º dia
Tempo zero
dos testes
nociceptivos
TIC e Vonfrey (1x por semana
0-56º dia)
56º dia
Capsazepina (5mg/kg, IP) Aprepitanto (1mg/kg, IP) MK-801 (0,5mg/kg, IP)
(2x por semana)
30 min antes
1ª - 9ª injeção de OXL (1mg/kg, IV) /
Controles 2x por semana

61
4.1 ENVOLVIMENTO DO ANTAGONISTA NÃO SELETIVO PARA OS
RECEPTORES A E B DA ET-1 (BOSENTANA)
Para analisar o envolvimento da endotelina na NSP induzida por OXL,
administrou-se o antagonista para os receptores ETA e ETB não seletivo Bosentana
(100mg/kg, VO).
A administração de OXL resultou em diferenças significantes entre os grupos
neuropático e controle (grupo de animais que receberam apenas o veículo). A
administração de bosentana 30 min antes da OXL foi capaz de prevenir a NSP fato
observado no teste de hipernocicepção Von Frey a partir do 28º dia (**p<0,01), no
35º dia a atenuação do efeito da NSP foi de 42,3% (**p<0,01). Nos dias 42º, 49º e
56º dias houve redução no limar (***p<0,001) de 50,9%, 47,9% e 50%
respectivamente no grupo tratado concomitantemente com Bosentana em
comparação ao grupo neuropático, que foi o grupo que recebeu administração IV
somente de oxaliplatina (Figura 7).
Ao realizar o teste TIC a 10ºC, observou-se que a partir do 7º dia houve
aumento significativo (***p<0,001) do tempo da resposta do grupo tratado com
Bosentana em comparação ao grupo neuropático atingindo o máximo no 56º dia, um
tempo de resposta 6 vezes maior (84,5%) que o grupo com NSP por OXL (Figura 8).

62
Figura 7 - Avaliação do antagonista não seletivo de endotelina (Bosentana) na hiperalgesia mecânica plantar induzida por oxaliplatina. O uso de Bosentana (100 mg/kg, VO, n=6) administrada trinta minutos antes de cada uma das nove administrações (setas) de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. Nos grupos controles foi administrado glicose (5%, IV, n=6) e CMC (2%, VO, n=6). O teste nociceptivo (hiperalgesia mecânica plantar – Von Frey eletrônico) foi realizado uma vez por semana durante 56 dias. Os resultados foram expressos como média ± erro padrão da média (EPM) da variação do limiar de retirada da pata em gramas (intensidade de hiperalgesia). Os asteriscos indicam a diferença estatística em relação ao grupo neuropático (** p<0,01; ***p<0,001; Two-way ANOVA seguida do teste de Newman-Keuls). Legenda: neuropatia sensitiva periféria (NSP); Carboximetilcelulose (CMC); Oxaliplatina (OXL).
CMC e OXL
43,5
%%
42,3
%%
50,9
%%
47,9
%%
50%
Teste de hiperalgesia
mecânica
Von Frey
(1x por semana 0-56
dias)
Bosentana
100mg/kg (VO)
30 min antes

63
Figura 8- Avaliação do antagonista não seletivo de endotelina (Bosentana) no desenvolvimento da hipernocicepção térmica ao frio (10 ºC) induzida por oxaliplatina. O uso de Bosentana (100 mg/kg, VO, n=6) administrada trinta minutos antes de cada uma das nove administrações (setas) de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. Nos grupos controles foi administrado glicose (5%, IV, n=6) e CMC (2%, VO, n=6). O teste nociceptivo (teste de imersão da cauda a 10ºC) foi realizado uma vez por semana durante 56 dias. Os resultados foram expressos como média ± erro padrão da média (EPM) da variação do tempo de retirada da cauda (intensidade de alodínia térmica). Os asteriscos indicam a diferença estatística em relação ao grupo neuropático (***p<0,001; Two-way ANOVA seguida do teste de Newman-Keuls). Legenda: Teste de imersão da cauda (TIC); neuropatia sensitiva periféria (NSP); Carboximetilcelulose (CMC); Oxaliplatina (OXL).
CMC e OXL
84,5%
%
Bosentana
100mg/kg (VO)
30 min antes

64
4.2 ENVOLVIMENTO DO ANTAGONISTA SELETIVO PARA O RECEPTOR A
DA ET-1 (BQ-123)
Após a administração intraplantar do antagonista de receptores de endotelina
ETA (BQ-123) observou-se ao realizar o teste de hipernocicepção Von Frey, que a
partir de 35º dia (62,6%) houve redução significativa (***p<0,001) da NSP por
bloqueio do receptor A, sendo de 61,3%, 59,7% e 54,6% nos dias 42º, 49º e 56º
respectivamente ao comparar com o grupo neuropático (OXL) (Figura 9).
Ao analisar a diferença entre as patas direita e esquerda percebeu-se a
notável inibição da NSP na pata direita do grupo BQ-123 e OXL em comparação ao
grupo neuropático (OXL) de 62,6% (p<0,001). Na pata esquerda, que não recebeu a
administração do antagonista BQ-123, houve manutenção da neuropatia (Figura 10).

65
Figura 9 - Avaliação do antagonista seletivo de receptor A de endotelina (BQ-123) na hiperalgesia mecânica plantar induzida por oxaliplatina. O uso de BQ-123 (30µl, i.pl., n=6) administrado trinta minutos antes de cada uma das nove administrações (setas) de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. Nos grupos controles foi administrado glicose (5%, IV, n=6) e PBS (0,01M, i.pl., n=6). O teste nociceptivo (hiperalgesia mecânica plantar – Von Frey eletrônico) foi realizado uma vez por semana durante 56 dias. Os resultados foram expressos como média ± erro padrão da média (EPM) da variação do limiar de retirada da pata em gramas (intensidade de hiperalgesia). Os asteriscos indicam a diferença estatística em relação ao grupo neuropático (***p<0,001; Two-way ANOVA seguida do teste de Newman-Keuls). Legenda: neuropatia sensitiva periféria (NSP); PBS (tampão fosfato-salino); Oxaliplatina (OXL).
62,6%
61,3% 59,7%
54,6%
PBS e OXL
Teste de hiperalgesia
mecânica
Von Frey
(1x por semana 0-56 dias)
BQ-123
30µl (i.pl.)
30 min antes

66
Figura 10 – Comparação entre as patas direita e esquerda com a administração do antagonista seletivo de receptor A de endotelina (BQ-123) na hiperalgesia mecânica plantar induzida por oxaliplatina no 28º dia. O uso de BQ-123 (30µl, i.pl., n=6) administrado trinta minutos antes de cada uma das nove administrações de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. Nos grupos controles foi administrado glicose (5%, IV, n=6) e PBS (0,01M, i.pl., n=6). Os asteriscos indicam a diferença estatística em relação ao grupo neuropático (***p<0,001; One-way ANOVA seguida do teste de Newman-Keuls). Legenda: neuropatia sensitiva periféria (NSP); PBS (tampão fosfato-salino); Oxaliplatina (OXL).
62,6%

67
4.3 ENVOLVIMENTO DO ANTAGONISTA SELETIVO PARA O RECEPTOR B
DA ET-1 (BQ-788)
Após a administração intraplantar do antagonista de receptores de endotelina
ETB (BQ-788) observou-se ao realizar o teste de hipernocicepção Von Frey, que a
partir de 35º dia houve redução (**p<0,01) da NSP por bloqueio do receptor B
(53,5%), sendo de 62,7%, 59% e 57,6% nos dias 42º, 49º e 56º respectivamente ao
comparar com o grupo neuropático (OXL) (Figura 11).
Ao analisar a diferença entre as patas direita e esquerda percebeu-se
redução significativa na pata direita do grupo BQ-788 e OXL em comparação ao
grupo neuropático (OXL) de 18,8% (**p<0,01). Na pata esquerda, que não
administrado o antagonista BQ-788, houve manutenção da neuropatia (Figura 12).

68
Figura 11 - Avaliação do antagonista seletivo de receptor B de endotelina (BQ-788) na hiperalgesia mecânica plantar induzida por oxaliplatina. O uso de BQ-788 (30µl, i.pl., n=6) administrado trinta minutos antes de cada uma das nove administrações (setas) de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. Nos grupos controles foi administrado glicose (5%, IV, n=6) e PBS (0,01M, i.pl., n=6). O teste nociceptivo (hiperalgesia mecânica plantar – Von Frey eletrônico) foi realizado uma vez por semana durante 56 dias. Os resultados foram expressos como média ± erro padrão da média (EPM) da variação do limiar de retirada da pata em gramas (intensidade de hiperalgesia). Os asteriscos indicam a diferença estatística em relação ao grupo neuropático (***p<0,001; Two-way ANOVA seguida do teste de Newman-Keuls). Legenda: neuropatia sensitiva periféria (NSP); PBS (tampão fosfato-salino); Oxaliplatina (OXL).
Teste de hiperalgesia
mecânica
Von Frey
(1x por semana 0-56 dias)
BQ-788
30µl (i.pl.)
30 min antes

69
Figura 12 – Comparação das patas direita e esquerda da administração do antagonista seletivo de receptor B de endotelina (BQ-788) na hiperalgesia mecânica plantar induzida por oxaliplatina. O uso de BQ-788 (30µl, i.pl., n=6) administrado trinta minutos antes de cada uma das nove administrações de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. Nos grupos controles foi administrado glicose (5%, IV, n=6) e PBS (0,01M, i.pl., n=6). Os asteriscos indicam a diferença estatística em relação ao grupo neuropático (**p<0,01; One-way ANOVA seguida do teste de Newman-Keuls). Legenda: neuropatia sensitiva periféria (NSP); PBS (tampão fosfato-salino); Oxaliplatina (OXL).
18,8%

70
4.4 ENVOLVIMENTO DO ANTAGONISTA DE RECEPTORES TRPV1 NA NSP
INDUZIDA POR OXL
Os camundongos submetidos à NSP induzida por OXL foram tratados
previamente, por via sistêmica, concomitantemente às injeções de OXL, com o
antagonista do receptor TRPV1 (Capsazepina, 5mg/kg, IP).
O antagonista do TRPV1, a capsazepina (CPZ), foi capaz de diminuir a
hipernocicepção plantar significativamente (***p<0,001) a partir do 28º dia (30,4%).
Diminuiu também significativamente (***p<0,001) nos dias 35º, 42º, 49º e 56º dias.
As reduções foram de 48,6%; 51,9%; 48,5% e 50,7% respectivamente ao comparar
o grupo pré-tratado com a capsazepina e o grupo neuropático (Figura 13).
No TIC houve aumento significativo (***p<0,001) do limiar de resposta a partir
do 7º dia atingindo o máximo em 56º dia (92,4%) ao comparar o grupo pré-tratado
com a capsazepina e o grupo neuropático (Figura 14).

71
Figura 13 - Avaliação do antagonista do TRPV1 (capsazepina) na hiperalgesia mecânica plantar induzida por oxaliplatina. O uso de Capsazepina (5 mg/kg, IP, n=6) administrada trinta minutos antes de cada uma das nove administrações (setas) de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. Nos grupos controles foi administrado glicose (5%, IV, n=6), DMSO (2%) e salina (NaCl 0,9%, IP, n=6). O teste nociceptivo (hiperalgesia mecânica plantar – Von Frey eletrônico) foi realizado uma vez por semana durante 56 dias. Os resultados foram expressos como média ± erro padrão da média (EPM) da variação do limiar de retirada da pata em gramas (intensidade de hiperalgesia). Os asteriscos indicam a diferença estatística em relação ao grupo neuropático (***p<0,001; Two-way ANOVA seguida do teste de Newman-Keuls). Legenda: neuropatia sensitiva periféria (NSP); Dimetil sulfóxido (DMSO); Solução salina estéril (NaCl); Oxaliplatina (OXL).
30,4%
48,6%
51,9% 48,5%
50,7%
DMSO e NaCl
Teste de hiperalgesia
mecânica
Von Frey
(1x por semana 0-56 dias)
Capsazepina
5mg/kg (IP)
30 min antes

72
Figura 14 - Avaliação do antagonista do TRPV1 (capsazepina) no desenvolvimento de alodínia térmica ao calor (43ºC) induzida por oxaliplatina. O uso de Capsazepina (5 mg/kg, IP, n=6) administrada trinta minutos antes de cada uma das nove administrações (setas) de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. Nos grupos controles foi administrado glicose (5%, IV, n=6), DMSO (2%) e salina (NaCl 0,9%, IP, n=6). O teste nociceptivo (teste de imersão da cauda a 43ºC) foi realizado uma vez por semana durante 56 dias. Os resultados foram expressos como média ± erro padrão da média (EPM) da variação do limiar de retirada da cauda (intensidade de alodínia térmica). Os asteriscos indicam a diferença estatística em relação ao grupo neuropático (***p<0,001; Two-way ANOVA seguida do teste de Newman-Keuls). Legenda: Teste de imersão da cauda (TIC); neuropatia sensitiva periféria (NSP); Dimetil sulfóxido (DMSO); Solução salina estéril (NaCl); Oxaliplatina (OXL).
92,4%
Teste Térmico TIC
(1x por semana 0-56 dias)
Capsazepina
5mg/kg (IP)
30 min antes

73
4.5 ENVOLVIMENTO DO ANTAGONISTA DE RECEPTORES NK-1 NA NSP
INDUZIDA POR OXL
Os camundongos submetidos à NSP induzida por OXL foram tratados
previamente, por via sistêmica, concomitantemente as injeções de OXL, com o
antagonista do receptor de Substância P (Aprepitanto 1mg/kg, IP).
O antagonista do receptor da SP (NK-1), aprepitanto, foi capaz de diminuir a
hipernocicepção plantar significativamente (***p<0,001) no 14º, 21º, 28º, 35º, 42º,
49º e 56º dias, as reduções foram de 42,5%, 44,3%, 47,8%, 55,5%, 56,7%, 50,5%,
52,3% respectivamente ao comparar o grupo pré-tratado com o aprepitanto e o
grupo neuropático (Figura 15).
Ao realizar o teste TIC a 10ºC, observou-se que a partir do 14º dia houve
aumento significativo (**p<0,01) da resposta do grupo tratado com aprepitanto em
comparação ao grupo neuropático, atingindo o máximo no 56º dia, uma resposta de
96,8% comparando com o grupo com NSP por OXL (Figura 16).

74
Figura 15 - Avaliação do antagonista do receptor NK-1 (aprepitanto) na hiperalgesia mecânica plantar induzida por oxaliplatina. O uso de Aprepitanto (1mg/kg, IP, n=6) administrado trinta minutos antes de cada uma das nove administrações (setas) de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. Nos grupos controles foi administrado glicose (5%, IV, n=6) e solução salina (NaCl 0,9%, IP, n=6). O teste nociceptivo (hiperalgesia mecânica plantar – Von Frey eletrônico) foi realizado uma vez por semana durante 56 dias. Os resultados foram expressos como média ± erro padrão da média (EPM) da variação do limiar de retirada da pata em gramas (intensidade de hiperalgesia). Os asteriscos indicam a diferença estatística em relação ao grupo neuropático (***p<0,001; Two-way ANOVA seguida do teste de Newman-Keuls). Legenda: Neuropatia sensitiva periféria (NSP); Solução salina estéril (NaCl); Oxaliplatina (OXL).
Aprepitanto
1mg/kg (IP)
30 min antes
Teste de hiperalgesia
mecânica
Von Frey
(1x por semana 0-56 dias)
55,5%
44,3% 42,5% 47,8%
56,7% 50,5%
52,3%

75
Figura 16 - Avaliação do antagonista do receptor NK-1 (aprepitanto) no desenvolvimento de alodínia térmica ao frio (10 ºC) induzida por oxaliplatina. O uso de Aprepitanto (1mg/kg, IP, n=6) administrado trinta minutos antes de cada uma das nove administrações (setas) de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. Nos grupos controles foi administrado glicose 5%, IV, n=6) e solução salina (NaCl 0,9%, IP, n=6). O teste nociceptivo (teste de imersão da cauda a 10ºC) foi realizado uma vez por semana durante 56 dias. Os resultados foram expressos como média ± erro padrão da média (EPM) da variação do limiar de retirada da cauda (intensidade de alodínia térmica). Os asteriscos indicam a diferença estatística em relação ao grupo neuropático (**p<0,01; Two-way ANOVA seguida do teste de Newman-Keuls). Legenda: Teste de imersão da cauda (TIC); neuropatia sensitiva periféria (NSP); Solução salina estéril (NaCl); Oxaliplatina (OXL).
Teste Térmico TIC
(1x por semana 0-56 dias)
Aprepitanto
1mg/kg (IP)
30 min antes

76
4.6 ENVOLVIMENTO DO RECEPTOR NMDA NA NSP INDUZIDA POR OXL
Os camundongos submetidos à NSP induzida por OXL foram tratados
previamente, por via sistêmica, concomitantemente às injeções de OXL, com o
antagonista do receptor de NMDA (MK-801 0,5mg/kg, IP).
O antagonista dos receptores de NMDA, MK-801, foi capaz de inibir a
hipernocicepção plantar significativamente (**p<0,01) no 42º, 49º e 56º dias, as
reduções foram de 34,2%, 29,4%, 33,2% respectivamente ao comparar o grupo pré-
tratado com o MK-801 e o grupo neuropático (Figura 17).
Ao realizar o teste TIC a 10ºC, observou-se que a partir do 21º dia houve
aumento significativo (**p<0,01) da resposta do grupo tratado com MK-801 em
comparação ao grupo neuropático de 45% no 56º dia (Figura 18).

77
Figura 17 - Avaliação do antagonista do receptor de NMDA (MK-801) na hiperalgesia mecânica plantar induzida por oxaliplatina. O uso de MK-801 (0,5mg/kg, IP, n=6) administrada trinta minutos antes de cada uma das nove administrações (setas) de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. Nos grupos controles foi administrado glicose (5%, IV, n=6) e solução salina (NaCl 0,9%, IP, n=6). O teste nociceptivo (hiperalgesia mecânica plantar – Von Frey eletrônico) foi realizado uma vez por semana durante 56 dias. Os resultados foram expressos como média ± erro padrão da média (EPM) da variação do limiar de retirada da pata em gramas (intensidade de hiperalgesia). Os asteriscos indicam a diferença estatística em relação ao grupo neuropático (** p<0,01, Two-way ANOVA seguida do teste de Newman-Keuls). Legenda: neuropatia sensitiva periféria (NSP); Oxaliplatina (OXL).
34,2% 29,4%
33,2%
Teste de hiperalgesia
mecânica
Von Frey
(1x por semana 0-56 dias)
MK-801
0,5mg/kg (VO)
30 min antes

78
Figura 18 - Avaliação do antagonista do receptor de NMDA (MK-801) no desenvolvimento de alodínia térmica ao frio (10ºC) induzida por oxaliplatina. O uso de MK-801 (0,5mg/kg, IP, n=6) administrada trinta minutos antes de cada uma das nove administrações (setas) de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. Nos grupos controles foi administrado glicose (5%, IV, n=6) e solução salina (NaCl 0,9%, IP, n=6). O teste nociceptivo (teste de imersão da cauda a 10ºC) foi realizado uma vez por semana durante 56 dias. Os resultados foram expressos como média ± erro padrão da média (EPM) da variação do limiar de retirada da cauda (intensidade de alodínia térmica). Os asteriscos indicam a diferença estatística em relação ao grupo neuropático (**p<0,01; Two-way ANOVA seguida do teste de Newman-Keuls). Legenda: Teste de imersão da cauda (TIC); neuropatia sensitiva periféria (NSP); Oxaliplatina (OXL).
MK-801
0,5mg/kg (VO)
30 min antes
Teste Térmico TIC
(1x por semana 0-56 dias)
45%

79
RESULTADOS DAS IMUNOFLUORESCÊNCIAS
30 min antes
30º dia 1º dia
Imunofluorescência
cFOS
Receptor NK-1
Receptor de endotelina B
Bosentana (100mg/kg, VO) (2x por semana)
56º dia
Coleta do segmento medular e gânglios da
raiz dorsal
1ª - 9ª injeção de OXL (1mg/kg, IV) /
Controles 2x por semana
Quantificação da
imunoexpressão no
programa Image J

80
4.7 IMUNOFLUORESCÊNCIA PARA cFOS EM SEGMENTO MEDULAR E DRG
APÓS PRÉ-TRATAMENTO COM BOSENTANA.
Observa-se em verde o marcador NeuN (Anti-NeuN Alexa-fluor 488), que é
uma proteína expressa em neurônios adultos ao qual é conjugado com um fluóforo
(488) marcando a atividade neuronal; em vermelho o Alexa-Fluor 568 que marca o
anticorpo ativado (cFOS) e a co-marcação (Merge). É bem notório observar a
marcação neuronal pelo cFOS bem presente no grupo OXL estando ausente nos
núcleos do grupo controle e bem discreto no grupo pré-tratado com Bosentana no
qual foi induzida a NSP com OXL (Figuras 19 e 20).
Foi realizada análise quantitativa através do programa Image J da
imunoexpressão para cFOS na medula espinhal dos camundongos submetidos à
NSP induzida por OXL, no qual houve redução de 99,9% ao comparar o grupo OXL
com o pré-tratado, demonstrando que o pré-tratamento com Bosentana regride os
sintomas da NSP (Figura 21). Não foi possível realizar a quantificação da
imunoexpressão para cFOS em DRG devido à quantidade insuficiente de amostras
para comparação estatísticas e realização das fotografias.

81
Figura 19 - Fotomicrografias da marcação por imunofluorescência para cFOS de medula espinhal de camundongos submetidos ao pré-tratamento com Bosentana. O uso de Bosentana (100 mg/kg, VO, n=6) administrada trinta minutos antes da administração de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. No grupo controle foi administrado glicose (5%, IV, n=6). Observa-se em verde o marcador NeuN (Anti-NeuN Alexa-fluor 488), em vermelho o Alexa-Fluor 568 (marcação de núcleo neuronal pelo cFOS) e a conjugação de todos (Merge) (Aumento 20x).

82
Figura 20 - Porcentagem de área imunomarcada para cFOS em medula espinhal de camundongos submetidos ao pré-tratamento com Bosentana. O uso de Bosentana (BOS, 100 mg/kg, VO, n=6) administrada trinta minutos antes da administração de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. No grupo controle foi administrado glicose (5%, IV, n=6). Os asteriscos indicam a diferença estatística em relação ao grupo neuropático (***p<0,001; One-way ANOVA seguida do teste de Newman-Keuls).
99,9%
***

83
Figura 21 - Fotomicrografias da marcação por imunofluorescência para cFOS de DRG de camundongos submetidos ao pré-tratamento com Bosentana. O uso de Bosentana (BOS, 100 mg/kg, VO, n=6) administrada trinta minutos antes da administração de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. No grupo controle foi administrado glicose (5%, IV, n=6). Observa-se em verde o marcador NeuN (Anti-NeuN Alexa-fluor 488), em vermelho o Alexa-Fluor (marcação de núcleo neuronal pelo cFOS) e a conjugação de todos (Merge) (Aumento 40x).

84
4.8 IMUNOFLUORESCÊNCIA PARA O RECEPTOR DE NK-1 EM SEGMENTO
MEDULAR E DRG
Observa-se em verde o marcador NeuN (Anti-NeuN Alexa-fluor 488 Observa-
se em verde o marcador NeuN (Anti-NeuN Alexa-fluor 488), que é uma proteína
expressa em neurônios adultos ao qual é conjugado com um fluóforo (488)
marcando a atividade neuronal; em vermelho por Alexa-Fluor 568 que marca o
anticorpo ativado NK-1 e a co-marcação (Merge). É bem notório observar a
marcação de núcleo neuronal para o receptor de NK-1 bem presente no grupo OXL
estando quase ausente nos núcleos do grupo controle ao comparar com o grupo que
foi induzida a NSP com OXL, fato observado tanto no gânglio como na medula
(Figuras 22 e 23).

85
Figura 22 - Fotomicrografias da marcação por imunofluorescência para o receptor NK-1 de medula espinhal de camundongos neuropáticos. Os animais foram tratados com oxaliplatina (OXL, 1mg/kg, IV, n=6) e no grupo controle foi administrado glicose (5%, IV, n=6). Houve a aumento da expressão de NK-1 no grupo neuropático (OXL) comparado com o grupo controle. Observa-se em verde o marcador NeuN (Anti-NeuN Alexa-fluor 488), em vermelho o Alexa-Fluor 568 (expressão do receptor NK-1) e a conjugação de todos (Merge) (Aumento 20x).
Merge
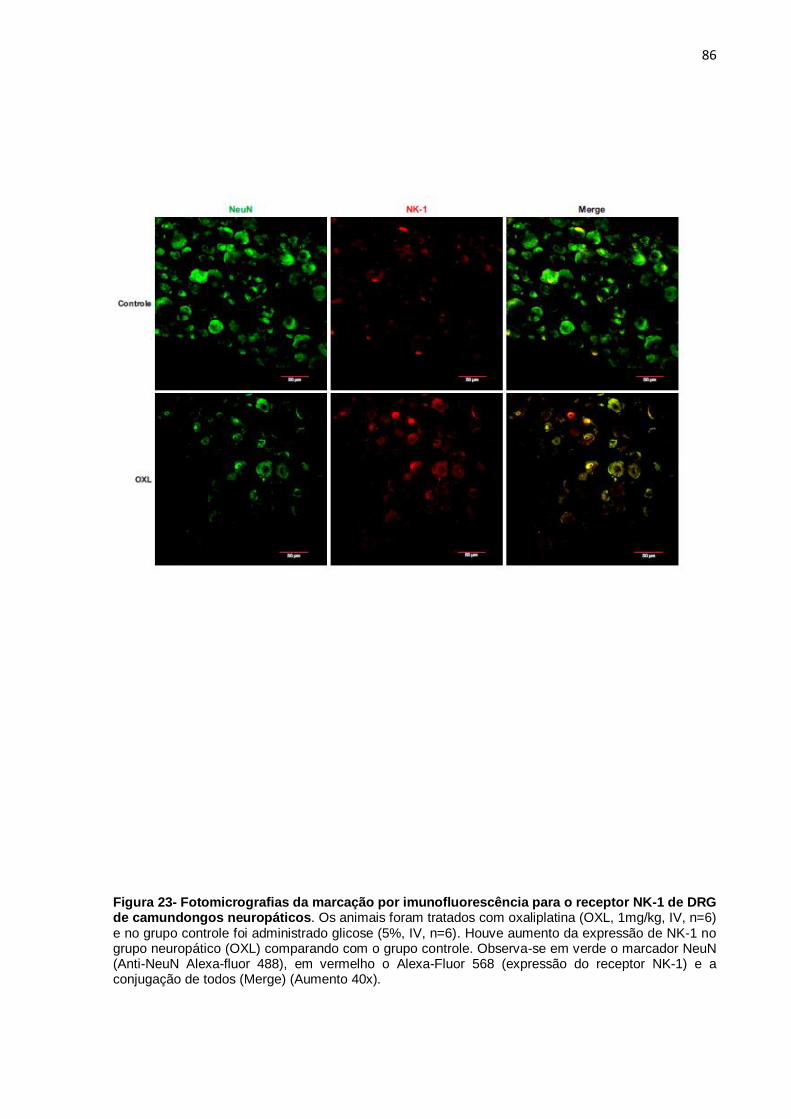
86
Figura 23- Fotomicrografias da marcação por imunofluorescência para o receptor NK-1 de DRG de camundongos neuropáticos. Os animais foram tratados com oxaliplatina (OXL, 1mg/kg, IV, n=6) e no grupo controle foi administrado glicose (5%, IV, n=6). Houve aumento da expressão de NK-1 no grupo neuropático (OXL) comparando com o grupo controle. Observa-se em verde o marcador NeuN (Anti-NeuN Alexa-fluor 488), em vermelho o Alexa-Fluor 568 (expressão do receptor NK-1) e a conjugação de todos (Merge) (Aumento 40x).

87
4.9 IMUNOFLUORESCÊNCIA PARA NK-1 EM SEGMENTO MEDULAR E DRG
APÓS PRÉ-TRATAMENTO COM BOSENTANA.
Observa-se em verde o marcador NeuN (Anti-NeuN Alexa-fluor 488), que é
uma proteína expressa em neurônios adultos ao qual é conjugado com um fluóforo
(488) marcando a atividade neuronal; em vermelho o Alexa-Fluor 594 que marca o
anticorpo ativado e a co-marcação (Merge). É bem notório observar a marcação de
núcleo neuronal pelo NK-1 bem presente no grupo OXL estando ausente nos
núcleos do grupo controle e bem discreto no grupo pré-tratado com Bosentana no
qual induzida a NSP com OXL (Figuras 24 e 25). Houve redução de 94,9% ao
comparar o grupo OXL com o pré-tratado na medula, demonstrando que o pré-
tratamento com Bosentana regride os sintomas da NSP (Figura 26).

88
FIGURA 24 - Fotomicrografias da marcação por imunofluorescência para NK-1 de medula espinhal de camundongos submetidos ao pré-tratamento com Bosentana. O uso de Bosentana (BOS, 100 mg/kg, VO, n=6) administrada trinta minutos antes da administração de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. No grupo controle foi administrado glicose (5%, IV, n=6). Observa-se em verde o marcador NeuN (Anti-NeuN Alexa-fluor 488), em vermelho o Alexa-Fluor 594 (expressão do receptor NK-1) e a conjugação de todos (Merge) (Aumento 20x).

89
Figura 25 - Porcentagem de área imunomarcada para NK-1 em medula espinhal de camundongos submetidos ao pré-tratamento com Bosentana. O uso de Bosentana (BOS, 100 mg/kg, VO, n=6) administrada trinta minutos antes da administração de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. No grupo controle foi administrado glicose (5%, IV, n=6). Os asteriscos indicam a diferença estatística em relação ao grupo neuropático (***p<0,001; One-way ANOVA seguida do teste de Newman-Keuls).
94,9%
***

90
Figura 26 – Fotomicrografias da marcação por imunofluorescência para NK-1 de DRG de camundongos submetidos ao pré-tratamento com Bosentana. O uso de Bosentana (BOS, 100 mg/kg, VO, n=6) administrada trinta minutos antes da administração de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. No grupo controle foi administrado glicose (5%, IV, n=6). Observa-se em verde o marcador NeuN (Anti-NeuN Alexa-fluor 488), em vermelho o Alexa-Fluor 594 (expressão do receptor NK-1) e a conjugação de todos (Merge) (Aumento 40x).

91
4.10 IMUNOFLUORESCÊNCIA PARA O RECEPTOR DE ENDOTELINA B EM
SEGMENTO MEDULAR E DRG.
Observa-se em azul o 4´,6-diamidino-2-fenil-indol (DAPI), que é um marcador
fluorescente que se liga fortemente a núcleos de DNA em células mortas ou vivas.
Em verde o marcador NeuN (Anti-NeuN Alexa-fluor 488). Observa-se em verde o
marcador NeuN (Anti-NeuN Alexa-fluor 488), que é uma proteína expressa em
neurônios adultos ao qual é conjugado com um fluóforo (488) marcando a atividade
neuronal; em vermelho por Alexa-Fluor 568 que marca o anticorpo ativado (receptor
de endotelina B) e a co-marcação (Merge). É bem notório observar a marcação de
núcleo neuronal para receptor de Endotelina B bem presente no grupo OXL estando
ausente nos núcleos do grupo controle e bem discreto no grupo pré-tratado com
Bosentana no qual foi induzida a NSP com OXL (Figuras 27 e 28).

92
Figura 27 - Fotomicrografias da marcação por imunofluorescência para receptor de endotelina B de medula espinhal de camundongos submetidos ao pré-tratamento com Bosentana. O uso de Bosentana (BOS, 100 mg/kg, VO, n=6) administrada trinta minutos antes da administração de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. No grupo controle (veículo) foi administrado glicose (5%, IV, n=6). Observa-se em verde o marcador NeuN (Anti-NeuN Alexa-fluor 488), em azul o DAPI, em vermelho o Alexa-Fluor 568 (expressão do receptor de endotelina ETB) e a conjugação de todos (Merge) (Aumento 20x). Legenda: neuropatia sensitiva periféria (NSP); 4´,6-diamidino-2-fenil-indol (DAPI); endotelina -1 (ET); Oxaliplatina (OXL).
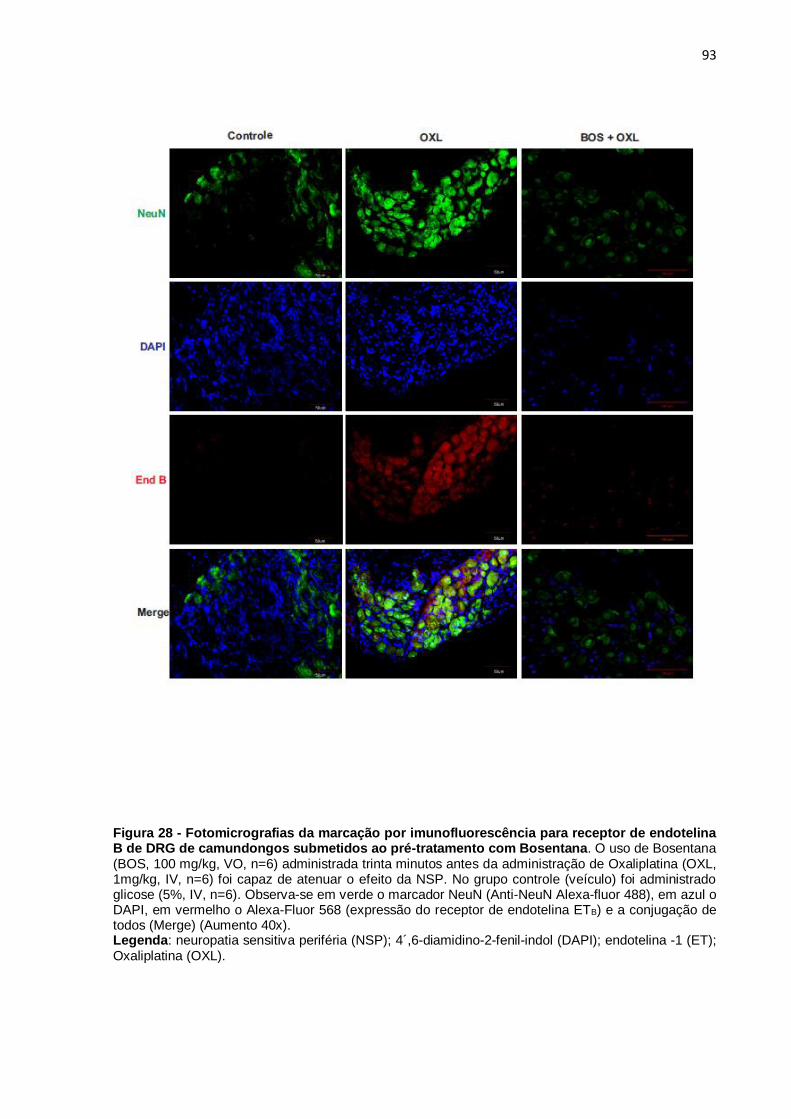
93
Figura 28 - Fotomicrografias da marcação por imunofluorescência para receptor de endotelina B de DRG de camundongos submetidos ao pré-tratamento com Bosentana. O uso de Bosentana (BOS, 100 mg/kg, VO, n=6) administrada trinta minutos antes da administração de Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi capaz de atenuar o efeito da NSP. No grupo controle (veículo) foi administrado glicose (5%, IV, n=6). Observa-se em verde o marcador NeuN (Anti-NeuN Alexa-fluor 488), em azul o DAPI, em vermelho o Alexa-Fluor 568 (expressão do receptor de endotelina ETB) e a conjugação de todos (Merge) (Aumento 40x). Legenda: neuropatia sensitiva periféria (NSP); 4´,6-diamidino-2-fenil-indol (DAPI); endotelina -1 (ET); Oxaliplatina (OXL).

94
RESULTADOS DO RT-PCR
28º dia 49º dia 7º dia
RT-PCR
Receptor de endotelina A
Receptor de endotelina B
TRPV1
SP
Coleta do segmento medular e gânglios da
raiz dorsal
1ª - 9ª injeção de OXL (1mg/kg, IV) / Controle (glicose
5%, IV) 2x por semana
14º dia

95
4.11 EXPRESSÃO GÊNICA PARA RECEPTORES DE ENDOTELINA A E B NA
MEDULA ESPINHAL E NO GÂNGLIO DA RAIZ DORSAL.
Ao analisar a expressão gênica para o receptor de Endotelina A (ETA) na
medula espinhal não se observou significância estatística em comparação ao grupo
controle (Figura 29). No DRG foi significante, também, no 14º e 28º dias (***p<0,001)
em comparação ao grupo controle. A quantificação de RNAm do grupo controle e
grupo OXL no DRG foi 2,5 vezes maior no 14º dia e 2 vezes maior no 28 para a
expressão do receptor ETA, demonstrando um aumento na expressão do receptor no
grupo neuropático (OXL) comparado ao grupo controle tanto na medula espinhal
quanto no DRG (Figura 30).
Ao analisar a expressão gênica para o receptor de endotelina B (ETB) na
medula espinhal não se observou significância estatística do grupo OXL em relação
ao controle (Figura 31). No DRG foi significante no 14º dia (***p<0,001) e no 28º dia
(**p<0,01). A quantificação de RNAm foi 3 vezes maior no 14º dia e 2 vezes maior
no 28º dia ao comparar o grupo neuropático (OXL) com o grupo controle para a
expressão do receptor ETB, demonstrando um aumento na expressão do receptor no
grupo neuropático (OXL) comparado ao grupo controle tanto na medula espinhal
quanto no DRG (Figura 32).

96
Figura 29 - Expressão gênica para Receptor de Endotelina A em medula espinhal nos grupos controle e oxaliplatina. A Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi administrada por 4,5 semanas (9 injeções) e o grupo controle foi tratado com glicose (5%, IV, n=6). Foi realizado o RT-PCR no 7º, 14º, 28º e 49º dias.

97
Figura 30 - Expressão gênica para Receptor de Endotelina A em DRG nos grupos controle e oxaliplatina. A Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi administrada por 4,5 semanas (9 injeções) e o grupo controle foi tratado com glicose (5%, IV, n=6). Foi realizado o RT-PCR no 7º, 14º, 28º e 49º dias. Os asteriscos indicam a diferença estatística em relação ao grupo controle (***p<0,001 no 14º e 28º dias; One-way ANOVA seguida do teste de Newman-Keuls).
***
***

98
Figura 31 - Expressão gênica para Receptor de Endotelina B em medula espinhal nos grupos controle e oxaliplatina. A Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi administrada por 4,5 semanas (9 injeções) e o grupo controle foi tratado com glicose (5%, IV, n=6). Foi realizado o RT-PCR no 7º, 14º, 28º e 49º dias.

99
Figura 32 - Expressão gênica para receptor de endotelina B em DRG nos grupos controle e oxaliplatina. A oxaliplatina (OXL, 1mg/kg, IV, n=6) foi administrada por 4,5 semanas (9 injeções) e o grupo controle foi tratado com glicose (5%, IV, n=6). Foi realizado o RT-PCR no 7º, 14º, 28º e 49º dias. Os asteriscos indicam a diferença estatística em relação ao grupo controle (***p<0,001 no 14º dia e **p<0,01 no 28º dia; One-way ANOVA seguida do teste de Newman-Keuls).
***
**

100
4.12 EXPRESSÃO GÊNICA PARA TRPV1 NA MEDULA ESPINHAL E NO
GÂNGLIO DA RAIZ DORSAL
Ao analisar a expressão gênica para TRPV1 na medula observou-se
significância estatística no 7º dia (***p<0,001) no 14º e 28º dias (**p<0,01) ao
comparar grupo controle com grupo OXL. A quantificação de RNAm do grupo
controle e grupo OXL na medula espinhal foi duas vezes maior no 14º dia em
comparação ao grupo controle (Figura 33).
No DRG foi significante a expressão gênica para TRPV1 no 7º, 14º e 28º dias
(***p<0,001). A quantificação de RNAm do grupo controle e grupo OXL no DRG foi 2
vezes maior, demonstrando um aumento na expressão do receptor no grupo
neuropático (OXL) comparado ao grupo controle tanto na medula espinhal quanto no
DRG (Figura 34).

101
Figura 33 - Expressão gênica para TRPV1 em medula espinhal nos grupos controle e oxaliplatina. A Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi administrada por 4,5 semanas (9 injeções) e o grupo controle foi tratado com glicose (5%, IV, n=6). Foi realizado o RT-PCR no 7º, 14º, 28º e 49º dias. Os asteriscos indicam a diferença estatística em relação ao grupo controle (***p<0,001 no 14º dia; One-way ANOVA seguida do teste de Newman-Keuls).
***

102
Figura 34 - Expressão gênica para TRPV1 em DRG nos grupos controle e oxaliplatina. A Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi administrada por 4,5 semanas (9 injeções) e o grupo controle foi tratado com glicose (5%, IV, n=6). Foi realizado o RT-PCR no 7º, 14º, 28º e 49º dias. Os asteriscos indicam a diferença estatística em relação ao grupo controle (***p<0,001 no 7º, 14º e 28º dias; One-way ANOVA seguida do teste de Newman-Keuls).
*** ***
***

103
4.13 EXPRESSÃO GÊNICA PARA SP NA MEDULA ESPINHAL E NO GÂNGLIO
DA RAIZ DORSAL
A Expressão gênica para SP em medula revelou diferença estatística
significante nos dias 14º e 28º (***p<0,001) entre o grupo controle e o OXL para os
níveis de RNAm. A quantificação de RNAm do grupo controle e grupo OXL na
medula espinhal mostrou um aumento na expressão do receptor no grupo
neuropático (OXL) comparado ao grupo controle duas vezes maior no dia 7 (Figura
35).
A Expressão gênica para SP no DRG revelou diferença estatística significante
no 14º e 28º (***p<0,001) entre o grupo controle e o OXL para os níveis de RNAm. A
quantificação de RNAm ao comparar o grupo neuropático (OXL) com o grupo
controle foi 2 vezes maior no 14º dia e 3 vezes maior no 28º dia (Figura 36).

104
Figura 35 - Expressão gênica para SP em medula espinhal nos grupos controle e oxaliplatina. A Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi administrada por 4,5 semanas (9 injeções) e o grupo controle foi tratado com glicose (5%, IV, n=6). Foi realizado o RT-PCR no 7º, 14º, 28º e 49º dias. Os asteriscos indicam a diferença estatística em relação ao grupo controle (***p<0,001 no 7º dia; One-way ANOVA seguida do teste de Newman-Keuls).
***

105
Figura 36 - Expressão gênica para SP em DRG nos grupos controle e oxaliplatina. A Oxaliplatina (OXL, 1mg/kg, IV, n=6) foi administrada por 4,5 semanas (9 injeções) e o grupo controle foi tratado com glicose (5%, IV, n=6). Foi realizado o RT-PCR no 7º, 14º, 28º e 49º dias. Os asteriscos indicam a diferença estatística em relação ao grupo controle (***p<0,001 no 14º e 28º dias; One-way ANOVA seguida do teste de Newman-Keuls).
*** ***

106
DISCUSSÃO
No estudo foi investigado o envolvimento da endotelina, dos receptores
TRPV1 e NMDA e do neuropeptídeo SP na neuropatia periférica desencadeada pela

107
oxaliplatina, que é um fármaco antineoplásico muito utilizado no manuseio clínico de
diversos tumores como o câncer colorretal. O presente trabalho mostra o
envolvimento desses receptores e do neuropeptídeo SP foi demonstrada, algumas
vezes de forma mais ativa e outras menos, porém com resultados que justificam tal
assertiva, conforme descrito a seguir.
Baseado nos dados da presente investigação foi possível constatar o possível
papel da endotelina-1, provavelmente via ativação dos seus receptores. Tal sugestão
baseia-se no fato de que ao ser realizada a administração não seletiva do antagonista
dos receptores A e B da ET-1 (Bosentana) houve, de fato, significativa atenuação da
hiperalgesia mecânica plantar e alodínia térmica ao frio (10o C), através da
diminuição do limiar de nocicepção do animal, a partir do 28º e 7º dias
respectivamente. E com a administração não seletiva dos antagonistas específicos
dos receptores de endotelina A (BQ-123) e B (BQ-788) houve significativa
atenuação da hiperalgesia mecânica plantar, demonstrada através da diminuição do
limiar de nocicepção do animal, também a partir do 35º dia para ambos os
antagonistas. Esses dois dados conjuntos nos levam a sugerir que a ET-1 deve
exercer algum papel no desencadeamento da resposta hiperalgésica consequente à
administração crônica de oxaliplatina.
Nossos resultados foram coerentes com aqueles mostrados nos estudos de
Coriat (2014) e Blasi et al. (2015) os quais realizaram administração de ET-1 em
ratos e observaram que os animais desenvolveram franca hiperalgesia térmica.
Khodorova et al. (2009) ao realizarem a sensibilização térmica de uma das patas
dos camundongos, tornaram-nas mais sensíveis a ET-1 após a injeção de formalina.
A alodínia tátil também se desenvolveu na pata contralateral, com a sensibilização
após a injeção. Esse fato não foi observado em nossos estudos, pois ao administrar
os antagonistas seletivos de ET-1 (BQ-123 e BQ-788) na pata direita dos animais foi
percebida a atenuação da NSP induzida por OXL nessa pata, mas não na
contralateral (esquerda), que continuou neuropática.
Uma vez que, em boa parte das vezes em que se utiliza a oxaliplatina ela é
feita no tratamento quimioterápico de cânceres metastáticos colorretais, o que
significa que há doença em atividade com presença de tumor, é possível que o
tumor assim como o microambiente tumoral tenha alguma participação na liberação
de ET-1 e esta também possa estar contribuindo no surgimento e/ou amplificação da
resposta hiperalgésica observada na NSP induzida pela oxaliplatina. Embora não se

108
tenha investigado a hipótese supramencionada no presente estudo, a literatura
registra algumas tentativas de associação de mecanismos da hiperalgesia da
endotelina em animais portadores de tumor, porém não encontramos registros que
associem tumor, endotelina e oxaliplatina.
Nesse contexto, nos estudos de Cain et al. (2001) em que foram
simultaneamente realizadas a administração sistêmica do antagonista de receptores
ETA, BQ-123, e a injeção simultânea no tumor de um antagonista de receptores ETB,
BQ-788, antes da injeção da própria ET-1, foi possível se observar que não houve
nenhum efeito sobre o comportamento da resposta nociceptiva. Esses dados
parecem sugerir que ativação local dos receptores de ET-1 parece ser um evento
crucial no surgimento da hiperalgesia consequente à administração de ET-1 e que a
presença de tumor não influencia na magnitude dessa resposta. Nesse ínterim,
deve-se registrar que os dados aqui apresentados frutos de nossa investigação,
sustentam a sugestão da ativação local dos receptores, desde que a atenuação da
resposta hiperalgésica à oxaliplatina, foi observada em animais tratados com
administração local (intraplantar ipsilateralmente) e não sistêmica, dos antagonistas
dos receptores A e B de endotelina-1.
Também são interessantes os dados de Pickering e colaboradores (2008) os
quais ao utilizarem um modelo em ratos, conseguiram demonstrar que a injeção
sistêmica do antagonista de receptores ETA, BQ-123, promoveu uma atenuação da
resposta nociceptiva de ET-1, quando esta foi administrada diretamente no
microambiente tumoral, o que reforça a ideia da ativação periférica dos receptores
pela ET1, neste caso, provavelmente em células residentes presentes nos tecidos
em volta do tumor, possivelmente macrofágos. Esses dados também corroboram
com nossos achados, uma vez que ao administrar os antagonistas seletivos de ET-
1, BQ123 e BQ788, na pata de animais tratados sistemicamente com OXL, foi
percebida uma atenuação da hiperalgesia mecânica, apenas na pata ipsilateral, mas
não na contralateral.
Já nos estudos de Chichorro, Zampronio e Era (2009) a potenciação da
nocicepção induzida por capsaicina foi abolida pela injeção prévia de BQ-123
(antagonista do receptor ETA), mas não foi afetada pelo BQ-788 (antagonista dos
receptores ETB), ao passo que o aumento de hiperalgesia por ET-1 induzida por
capsaicina foi atenuada por ambos os antagonistas. Assim, como em nossos

109
resultados, só com a diferença de que o BQ-788 também atenuou a resposta
hiperalgésica em animais com NSP induzida por OXL.
Em outras pesquisas como as de Schmidt et al. (2008) e Pickering et al.
(2008), os autores concluíram que ET-1 é liberada a partir do próprio tumor e que
este fato está associado a uma amplificação do efeito nociceptivo que se segue
após injeção de ET1. Dessa forma demonstraram que a ET-1 endógena produzida
pelo próprio câncer também desencadeia hiperalgesia mecânica em modelo de
ratos. Assim, é possível que na vigência de NSP consequente ao tratamento
quimioterápico com regimes à base de OXL, os eventos associados à nocicepção
reclamados pelos pacientes possam ter como desencadeantes a presença de ET-1
liberada por células residentes, nos tecidos periféricos pertencentes ou não ao
microambiente tumoral. Novos estudos são necessários, entretanto para comprovar
tal assertiva.
Outro estudo como o de Baamonde (2004) revelou resultados semelhantes
utilizando modelo de constrição crônica do nervo ciático, onde ET-1 e os receptores
de ET-1 foram encontrados regulados no local da constrição em ratos que
apresentavam sinais de dor neuropática. Chichorro, Zampronio e Era (2009) em
seus estudos avaliaram os efeitos de antagonistas dos receptores de ETA e ETB em
um modelo de constrição crônica do nervo infra-orbital em ratos e observaram que a
administração intravenosa de um antagonista seletivo ETB foi capaz de promover
abolição completa da alodínia mecânica ao longo de duas horas, ao passo que a
administração de um antagonista seletivo ETA não o foi. Surpreendentemente, os
mesmos autores relataram em outro estudo que a hiperalgesia ao frio foi aliviada
pelos antagonistas dos receptores ETA e ETB no mesmo modelo de dor neuropática.
Os resultados de Quang, Brian e Schmidt (2010), também indicaram que a
hiperalgesia térmica foi mediada pelos receptores ETA e ETB, enquanto a alodínia
mecânica foi mediada exclusivamente por ETB. Parece que os receptores ETA
promovem sempre a nocicepção, mas o efeito biológico final de ETB é muito mais
complexo. Não apenas este depende da dose exógena de ET-1, mas também de
uma possível condição patológica associada, como no caso da NSP da OXL.
Para Hansa, Schmidt e Strichartz (2009) na inflamação e lesão, os receptores
ETB parecem ser pró-nociceptivos, baseado em anti-hiperalgesia de bloqueio com
BQ-788. Mas a ativação de receptores ETB, por um agonista de ET1, IRL-1620, por

110
exemplo, é também anti-hiperalgésico, o que sugere que depende de onde as
células são expressas, assim mais pesquisas terão de especificar o caminho exato.
Nossos resultados mostraram que os receptores ETA parecem estar mais
envolvidos com NSP induzida por OXL que os receptores ETB. Na confirmação de
observações anteriores, o presente estudo também mostra que ambos os receptores
ETA e ETB medeiam a hipernocicepção mecânica plantar observada quando se
administra o antagonista em uma das patas e se compara. Essa conclusão é
apoiada pela descoberta de que a inibição de ET-1 que pudesse ser alcançada
pelos antagonistas seletivos foi significativamente maior do que a observada com o
não seletivo. Ambos os subtipos de receptores ETA e ETB de ET-1 parecem estar
envolvidos, como pré-tratamento por antagonistas e estão limitados na sua inibição,
mas juntos eles podem atenuar a NSP induzida por OXL.
Com o intuito de avaliar essas hipóteses propusemos ensaios de
imunofluorescência. Observou-se uma notável participação dos receptores de SP
(NK-1), receptores de endotelina B e cFOS. Fato comprovado pela identificação da
ausência dos marcadores ao comparar o grupo pré-tratado com antagonista não
seletivo para os receptores A e B (Bosentana) e o grupo neuropático (OXL).
Há aproximadamente duas décadas, Hunt, Pini e Evan (1987) descreveram
pela primeira vez a expressão do gene cFos e seu produto protéico, a proteína cFos,
em neurônios do corno dorsal da medula espinhal lombar de ratos submetidos à
estimulação nociva. Desde então, a análise da expressão da proteína cFos tem sido
utilizada em estudos de bases neurais de nocicepção.
O cFos, um gene de ativação imediata, vem sendo utilizado como um
marcador anatômico de atividade neuronal, permitindo a avaliação da ativação
nociceptiva de uma grande população de neurônios espinhais, e a determinação do
padrão de ativação neuronal, de acordo com a natureza do estímulo e a localização
da lâmina espinhal onde os neurônios expressam esse gene.
Smits et al., (2009) citam que a proteína cFos é expressa pós-sinapticamente
em núcleos dos neurônios do corno dorsal da medula espinhal. A maioria dos
estímulos utilizados para induzir a expressão de c-Fos na medula é nociva, tais
como estímulos elétricos, tóxicos, químicos e lesão nervosa. Estímulos inócuos
também podem resultar na expressão de cFos na medula, principalmente em casos
de dores crônicas e devido à sensibilização central.

111
Observou-se, no presente estudo, um aumento da expressão de cFos no
DRG e na medula espinhal dos camundongos com neuropatia periférica, sugerindo
haver um aumento da atividade neuronal central nos animais tratados com OXL. O
pré-tratamento com o antagonista não seletivo, Bosentana, promoveu uma redução
importante da expressão de cFos na medula espinhal e no DRG.
Em um modelo de neuropatia periférica por constrição do nervo ciático, em
ratos, Jergova, Kolesar e Cizkova (2008) evidenciaram um aumento da expressão
de c-Fos nos neurônios da região parabraqueal da medula espinhal, relacionado
com a duração da alodínia mecânica. E Khodorova et al. (2009) e Smith et al. (2014)
citam que a ET-1 potencializa a nocicepção induzida por capsaicina através do
receptor ETA e aumenta a liberação de capsaicina estimulada por CGRP em
neurônios sensoriais e expressão de cFos no DRG, fomentando a ideia que
substância P também participa.
Com a realização do pré-tratamento com Bosentana também foi investigado a
expressão dos receptores NK-1, que são os receptores de SP, e a expressão do
receptor de endotelina B. Identificou-se a participação desses receptores ao
comparar o grupo neuropático que foi administrado OXL com o grupo pré-tratado
com Bosentana.
Jamieson et al. (2005) argumentam que mudança na expressão do
neuropeptídeo SP tem sido mostrada em modelos animais. Foi descrito que a SP
tem estado presente em fibras tratadas com cisplatina e OXL. A SP contribui para
sensibilizar os receptores nociceptivos diretamente na periferia ou na membrana
pós-sináptica ou através da interação com outros elementos algiogênicos.
A transmissão de sinais nociceptivos está associada com a liberação de SP
das terminações centrais de nociceptores peptidérgicos que terminam na lamina I e
II do CDME. Carozzi, Marmiroli e Cavaletti (2010) relatam que esses neurônios
também liberam os mesmos peptídeos na periferia para induzir inflamação
neurogênica em vários tecidos-alvo.
Os resultados de Kim, Kim e Yoon (2015) sugerem que a ET-1 está envolvida
no desenvolvimento de alodínia mecânica em síndrome de dor por compressão
regional, com reversão do quadro ao utilizarem o BQ-788, demonstrando que o
receptor de ETB parece estar realmente envolvido. Fato que em nossos resultados
de imunofluorescência pode ser comprovado com a redução da imunomarcação da
desse receptor ao utilizar o antagonista não seletivo Bosentana.

112
Assim, demonstramos a participação de receptores de endotelina B, do
receptor de SP, NK-1, e de cFOS nos quais houve a expressão na medula espinhal
e no DRG no grupo neuropático (OXL) e as suas inibições no grupo com pré-
tratamento com o antagonista não seletivo Bosentana, sugerindo o seu
envolvimento.
Outro receptor analisado foi o TRPV1, um dos membros da superfamília dos
TRPs vanilóide, com função de detecção e regulação da temperatura corporal,
proporcionando sensação de queimação calor e dor (nocicepção). Identificamos o
envolvimento desse receptor na patogênese da neuropatia sensitiva induzida por
oxaliplatina, na qual houve atenuação significativa da hiperalgesia mecânica e
térmica dos animais pré-tratados com o antagonista dos receptores TRPV1,
capsazepina, ao comparar com o grupo oxaliplatina que apresentou a neuropatia
crônica.
O envolvimento do receptor TRPV1, segundo cita Schumacher (2010), já foi
descrito em diversas doenças, sendo que o seu principal foco de estudo é em
patologias dolorosas. Assim, diversas evidências como as de Levine e Alessandrini-
Haser et al. (2007), Patapoutian, Wong e Gavva (2010), levam a crer que o
desenvolvimento de fármacos que possam agir como antagonistas do receptor
TRPV1 em humanos possa ser utilizado em diversos tipos de síndromes dolorosas.
Rashid e colaboradores (2003b), em seus estudos, também citam que a expressão
da atividade TRPV1 nas fibras sensoriais já foi comprovada com hiperalgesia em
neuropatias.
No presente estudo foram mostradas evidências farmacológicas, inicialmente,
acerca da participação do TRPV1 na patogênese de eventos associados à NSP
consequente ao tratamento crônico com oxaliplatina. Nesse sentido, foi possível
mostrar que o pré-tratamento de animais com o antagonista de TRPV1
(capsazepina), promoveu significativa redução na hiperalgesia mecânica plantar e
alodínia térmica observados após tratamento crônico com oxaliplatina a partir do 28º
e 7º dias respectivamente. Estes dados dão sustentação à hipótese de que a
ativação de receptores TRPV1 parece ser evento presente neste tipo de condição
associada à neurotoxicidade da quimioterapia do câncer, particularmente aquela
devida ao uso de oxaliplatina.
Adicionalmente, ao se analisar o papel do neuropeptídeo SP nesta mesma
condição patológica, também foi demonstrado, farmacologicamente, o seu provável

113
envolvimento uma vez que, o pré-tratamento de animais, posteriormente tratados
com oxaliplatina, com o antagonista de receptores NK-1 (apreptanto), promoveu
franca e significativa atenuação da hiperalgesia mecânica plantar e alodínia térmica,
ambas observadas a partir do 21º e 14º dias respectivamente. À semelhança do que
foi dito para o TRPV1, podemos afirmar que tais dados dão sustentação à hipótese
de que a ativação de receptores NK-1 parece ser também um evento presente neste
tipo de condição associada à neurotoxicidade da quimioterapia do câncer,
particularmente aquela devida ao uso de oxaliplatina.
Outra evidência farmacológica sobre a participação de receptores associados
à gênese de respostas nociceptivas, foi a demonstração aqui apresentada do efeito
inibidor parcial, porém significativo do pré-tratamento com o antagonista de NMDA
(MK-801), sobre a hiperalgesia mecânica plantar (a partir do 42º dia) e térmica (no
21º dia) observadas em animais tratados com oxaliplatina. Este resultado sugere a
participação do receptor NMDA nos eventos nociceptivos associados à NSP da
oxaliplatina.
Mihara et al. (2011), em seus estudos com animais de experimentação,
citaram que o antagonista do receptor NMDA reverte completamente a alodínia
mecânica induzida por OXL quando administrada depois do desenvolvimento da
neuropatia. Outros autores demonstraram (QU et al. 2009) que através de um
modelo de neuropatia por ligadura do nervo espinhal em ratos, há a ativação dos
receptores NMDA-2B na medula espinhal e este fato parece, segundo os autores,
ter um papel crucial no desenvolvimento da dor neuropática, especialmente no
estágio inicial da injúria nervosa. Com não muito menos evidência poderíamos
propor o mesmo com base em nossos resultados, para a NSP da OXL.
Nos estudos de Yamamoto et al., (2006) e Khodorova et al. (2006) a ET-1
induz alodínia tátil que é inibida por antagonista de receptores como do TRPV1, a
capsazepina, se administrada em torno de 1-1,5h após a administração de ET-1,
sendo também reduzida ao longo do seu curso por antagonistas dos receptores de
glutamato do tipo NMDA.
Chen, Samoroski e Pan (2009) relataram a participação do receptor NMDA no
desenvolvimento de dor crônica, onde a administração de um antagonista de NMDA
(neramexane) foi capaz de reverter a alodínia e a hiperalgesia mecânicas em um
modelo de neuropatia diabética em ratos semelhante aos nossos achados.

114
A maioria dos estudos vem demonstrando que a ativação excessiva do
receptor NMDA é capaz de produzir uma excitotoxicidade neuronal, e que a
administração de inibidores desses receptores é utilizada para reverter o dano
neuronal de diversas patologias. Em contrapartida, outros estudos vêm mostrando
um papel oposto do receptor NMDA na neuropatia crônica, sugerindo uma ação
antinociceptiva e antiapoptótica na neurotoxicidade induzida por quimioterápicos.
Em seguida verificamos a expressão gênica por análise de RT-PCR nos
tecidos (medula espinhal e gânglio da raiz dorsal) dos animais com neuropatia
sensitiva periférica induzidos por OXL, no qual houve presença marcante de todos
os receptores (ETA, ETB, TRPV1 e SP) nos dias 14 e 28, fato que coincide com o
pico da instalação da NSP induzida por oxaliplatina.
Na análise da quantificação de RNAm nos receptores de endotelina A e B
observou-se maior envolvimento do receptor A na NSP induzida por OXL em
camundongos do que do receptor B no 28º dia, sendo essa expressão maior em
DRG do que na medula espinhal ao comparar o grupo OXL com o controle.
Para Hansa, Schmidt e Strichartz (2009) os receptores ETA são expressos
principalmente por neurônios sensoriais de diâmetro pequeno do DRG, a expressão
de receptores ETB em DRG é restrita a células satélites da glia. Assim, o sistema
endotelina pode estar localizado em vários níveis do neuroeixo da dor e participa de
cascatas de sinalização, fundamentais na fisiologia da dor.
Em uma revisão sobre os mecanismos da NSP induzida por fármacos
antineoplásicos, como OXL, cisplatina, paclitaxel e vincristina os autores Jaggi e
Singh (2012), Descoeur et al. (2011), Ghelardini et al. (2010), Nietro et al. (2008) e
Ling et al. (2007) relataram que esses parecem estar interligados. Postularam que
estes agentes antineoplásicos desencadeiam mudanças na expressão e
características funcionais dos canais de sódio no DRG e em neurônios sensoriais no
corno dorsal. Nos estudos de Lalich et al. (2007), Nelson (2000), Quang e Schmidt
(2010) o RNAm da ET-1 é hipersecretado por muitos tipos de câncer, como próstata,
mama, pâncreas, câncer colorretal, e de carcinoma de células escamosas de boca.
Esses relatos são importantes, pois corroboram com nossos achados visto
que o antagonismo dos receptores de endotelina-1, de TRPV1 e SP atenuaram a
hiperalgesia mecânica e alodínia térmica da NSP nos animais induzidos com OXL,
comprovando a participação desses receptores na análise por PCR e
imunofluorescência. Pode-se comprovar, com os nossos resultados, que o fármaco

115
OXL induz a expressão e mudanças funcionais que contribuem para a sensibilidade
neuronal e alteram a excitabilidade neuropática.
Desse modo é sugerido o seguinte modelo do envolvimento da endotelina-1,
dos receptores TRPV1 e NMDA e do neuropeptídeo SP na gênese da NSP induzida
por OXL (Figura 38), por sua vez essa descoberta pode dar início a uma abordagem
terapêutica inovadora no tratamento da NSP induzida pela OXL e o provável
aumento da resposta dos pacientes com câncer colorretal ao tratamento
quimioterápico.

116
FIGURA 38 - Modelo do envolvimento da endotelina, dos receptores NMDA e TRPV1 e do neuropeptídeo SP na neuropatia sensitiva periférica induzida por oxaliplatina.
Fonte: AUTOR, 2015.
Legenda: Setas (utilização dos respectivos antagonistas); Oxaliplatina (OXL); Endotelina (ET-1); Receptor de endotelina A (ETA); Receptor de endotelina B (ETB); Corno Dorsal da Medula Espinhal (CDME); Receptor do ácido N-metil-D-Aspartato (NMDA); Gânglio da Raiz Dorsal (DRG); Neuropatia Sensitiva Periférica (NSP); Receptor de Potencial Transiente Vanilóide 1 (TRPV1); Substância P (SP); Receptor de substância P neuroquinina -1 (NK-1).
Oxaliplatina

117
CONCLUSÃO

118
Diante dos dados apresentados neste trabalho, foi possível concluir que:
Na patogênese da neuropatia sensitiva periférica induzida por oxaliplatina, a
utilização de antagonistas de endotelina-1, de receptores TRPV1 e NMDA e da
substância P, promoveram uma importante atenuação da hiperalgesia mecânica e
alodínia térmica, além da redução da imunoexpressão de cFos, NK-1 e receptor de
endotelina B no corno dorsal e na medula espinhal dos animais pré-tratados com o
antagonista não seletivo de endotelina-1, fato também comprovado pela
quantificação de RNAm para os receptores de endotelina A e B, TRPV1 e SP.
Portanto, conclui-se com este estudo que o sistema endotelina-1 e os
mecanismos subsequentes da via de dor, com a participação dos receptores NMDA
e TRPV1 e do neuropeptídeo SP estão envolvidos na patogênese da neuropatia
sensitiva periférica por indução da oxaliplatina, e que esses resultados podem ser
utilizados para estudos futuros com a oxaliplatina sugerindo que a neuropatia
sensitiva periférica, ao ser instalada, poderá ser no mínimo atenuada.

119
REFERÊNCIAS

120
AHMADI, S; GOLBAGHI, H.; AZIZBEIGI, R.; ESMAILZADEH, N. N-methyl-D-aspartate receptors involved in morphine-induced hypealgesia in sensitized mice. European Journal of Pharmacology. 737, 85-90, 2014. AHMED, S.I.; THOMPSON, J.; COULSON, J.M.; WOLL, P.J. Studies on the expression of endothelin, its receptor subtypes, and converting enzymes in lung cancer and in human bronchial epithelium. Am. J. Respir. Cell Mol. Biol. 22, 422–
431, 2000. AMAYA, F.; SHIMOSATO, G.; NAGANO, M.; et al. NGF and GDNF differentially regulate TRPV1 expression that contributes to development of inflammatory thermal hiperalgesia. Eur J Neurosci. 20:2303-2310, 2004.
ALMEIDA, T.F.; ROIZENBLATT, S.; TUFIK, S. Afferent pain pathways: a neuroanatomical review. Brain Res., 1000, pp. 40–56, 2004.
ALVES NETO, et al. Dor: princípios e prática, São Paulo: Artmed, 2009. ARAI, H.; HORI, S.; ARAMORI, I.; OHKUBO, H.; NAKANISHI, S. Cloning and expression of a cDNA encoding an endothelin receptor. Nature. 348: 730–732, 1990. ARETI, A.; YERRA, V.G.; NAIDU, V.; KUMAR, A. Oxidative stress and nerve damage: role in chemotherapy induced peripheral neuropathy. Redox Biol. 2:289–295, 2014. ASAI, H.; OZAKI, N.; SHINODA, M.; NAGAMINE, K.; TOHNAI, I.; UEDA, M.; SUGIURA, Y. Heat and mechanical hyperalgesia in mice model of cancer pain. Pain,
117:19–29, 2005. ATTAL, N.; BOUHASSIRA, D.; GAUTRON, M.; VAILLANT, J.N.; MITRY, E.; LEPÈRE, C.;, ROUGIER, P.; GUIRIMAND, F. Thermal hyperalgesia as a marker of oxaliplatin neurotoxicity: a prospective quantified sensory assessment study. Pain, v. 144, n. 3, p. 225-226, 2009. AUTHIER, et al. A new animal modelo f vincristine-induced nociceptive peripheral neuropathy. Neurotoxicology, 24:797-805, 2003. BAAMONDE, A.; LASTRA, A.; VILLAZÓN, M.; BORDALLO, J.; HIDALGO, A.; MENÉNDEZ, L. Involvement of endogenous endothelins in thermal and mechanical

121
inflammatory hyperalgesia in mice. Naunyn Schmiedebergs Arch Pharmacol. 369(2):245–251, 2004. BAKITAS, M.A. Background noise: The experience of Chemotherapy- induced peripheral neuropathy. Nursing Research. v. 56, n 05, p. 323-331, 2007.
BEAUDRY, H.; DUBOIS, D.; GENDRON, L. Activation of spinal mu- and delta-opioid receptors potently inhibits substance P release induced by peripheral noxious stimuli. J Neurosci, 31:13068–13077, 2011. BEIJERS, A.J.; MOLS, F.; VREUGDENHIL, G. A systematic review on chronic oxaliplatin-induced peripheral neuropathy and the relation with oxaliplatin administration. Support Care Cancer. Jul; 22(7):1999-2007, 2014.
BENATTI, G. A.; GUIMARÃES, C. Parestesia do nervo alveolar inferior. 2009.100f. Monografia (Ponte Lima: Portugal, Universidade Fernando Pessoa. Faculdade de ciências da Saúde), 2009. BERGER, A.; DUKES, E.M.; OSTER, G. Clinical characteristics and economic costs of patients with painful neuropathic disorders. J Pain. Apr; 5(3):143-9, 2004. BERTI-MATTERA, L.N.; GARIEPY, C.E.; BURKE, R.M.; HALL, A.K. Reduced expression of endothelin B receptors and mechanical hyperalgesia in experimental chronic diabetes. Exp Neurol. 01(2):399–406, 2006.
BLACK, S.M.; KUMAR, S.; WISEMAN, D.; RAVI, K.; WEDGWOOD, S.; RYZHOV, V.; FINEMAN JR. Pediatric pulmonary hypertension: roles of endothelin-1 and nitric oxide. Clin Hemorheol Microcirc 37: 111–120, 2007. BLASI, F. et al. Late-onset thermal hypersensitivity after focal ischemic thalamic infarcts as a model for central post-stroke pain in rats. J Cer Blood flow & Metab. 35,1100-11003, 2015. BOHM F, PERNOW J. The importance of endothelin-1 for vascular dysfunction in cardiovascular disease. Cardiovasc Res 76: 8–18, 2007.
BONASSA, E.M.A.; SANTANA, T.R. Enfermagem em terapêutica oncológica. 3.
ed. Rio de Janeiro: Atheneu, 2005.538p.

122
BRASIL. Ministério da Saúde. Secretaria de Atenção à Saúde. Instituto Nacional de Câncer. Coordenação de Prevenção e Vigilância de Câncer. Estimativas 2014: Incidência de Câncer no Brasil. Rio de Janeiro: INCA, 2014. Disponível em: http://www1.inca.gov.br/estimativa/2014/index.asp?link=conteudo_view.asp&ID=5 Acesso em: 16/01/2014. BRUCE, A. et al. Agentes antineoplásicos. In: BRUNTON, L. L. Goodman & Gilman: as bases farmacológicas da terapêutica. 11ª ed. Rio de Janeiro: McGraw-Hill, 2007. BRUNTON, L.L. As bases farmacológicas da terapêutica de Goodman & Gilman. 12 ed. Porto Alegre: AMGH, 2012.
CAHILL, C.M.; CODERRE, T.J. Attenuation of hyperalgesia in a rat model of neuropathic pain after intrathecal pre- or pos-treatment with a neurokinin-1 antagonist. Pain. 95(3):277-85, 2002 CAIN, D.M.; WACNIK, P.W.; TURNER, M.; WENDELSCHAFER-CRABB, G.; KENNEDY, W.R.; WILCOX, G.L.; SIMONE, D.A. Functional interactions between tumor and peripheral nerve: changes in excitability and morphology of primary afferent fibers in a murine model of cancer pain. J. Neurosci. 21, 9367–9376, 2001. CAMERON, I.T.; BACON, C.R.; COLLETT, G.P.; DAVENPORT, A.P. Endothelin expression in the uterus. J Steroid Biochem Mol Biol. 53: 209–214, 1995. CAROZZI, V.A.; CANTA, A.; CHIORAZZI, A.; CAVALETTI, G. Chemotherapy-induced peripheral neuropathy: What do we know about mechanisms? Neurosci Lett. Oct 22. pii: S0304-3940(14)00818-0, 2014.
CAROZZI, V.A.; MARMIROLI, P.; CAVALETTI, G. The role of oxidative stress and anti-oxidant treatment in platinum-induced peripherl neurotoxicity. Curr Cancer Drug Targets, 10(7):670-682, 2010. CATERINA, M.J.; LEFFLER, A.; MALMBERG, A.B.; MARTIN, W.J.; TRAFTON, J.; PETERSEN-ZEITZ, K.R.; KOLTZENBURG, M.; BASBAUM, A.I.; JULIUS, D. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science. 5464:306-13, 2000. CERSOSIMO, R.J. Oxaliplatin-associated neuropathy: a review. Ann Pharmacother, 39(1):128-35, 2005.

123
CHAUDHRY, V.; ROWINSKY, E.K.; SARTORIUS, S.E.; DONEHOWER, R.C.; CORNBLATH, D.R. Peripheral neuropathy from taxol and cisplatin combination chemotherapy: Clinical and electrophysiological studies. Annals of Neurology. v. 3, n. 35, p. 304-311, 1994. CHEN, S-R.; SAMORISKI, G.; PAN, H-L. Antinociceptive effects of chronic administration of uncompetitive NMDA receptor antagonists in a rat model of diabetic neuropathic pain. Neuropharmacology, v. 57, p. 121-126, 2009. CHICHORRO, J.G.; ZAMPRONIO, A.R.; ERA, G.A. Endothelin ETB receptor antagonist reduces mechanical allodynia in rats with trigeminal neuropathic pain. Exp Biol Med (Maywood). 231(6):1136–1140, 2009.
CHU, E. et al. Oxaliplatin: the new member of the class. In: MARSHALL, J. The Clinical use of oxaliplatin: case studies and roundtable discussion. New York: Oncology. 2004. CORIAT, R. et al. Treatment of oxaliplatin-induced peripheral neuropathy by intravenous mangafodipir. The journal of clinical investigation. v.124, n.1, january, 2014. CRISTINO, L.; DE PETROCELLIS, L.; PRYCE, G.; BAKER, D.; GUGLIELMOTTI, V.; DI MARZO, V. Immunohistochemical localization of cannabinoid type 1 and vanilloid transient receptor potential vanilloid type 1 receptors in the mouse brain. Neuroscience. 139:1405-1415, 2006.
CUNHA, T.M., et al. An electronic pressure-meter nociception paw test for mice. Braz J Med Biol Res,v. 37(3), p. 401-407, 2004.
DAMON, D.H. Endothelin and post-ganglionic sympathetic neurons. Clin Exp Pharmacol Physiol. 26: 1000–1003, 1999.
DELARUE, C. ; CONLON, J.M. ; REMY-JOUET, I. ; FOURNIER, A. ; VAUDRY, H. Endothelins as local activators of adrenocortical cells. J Mol Endocrinol. 32: 1–7,
2004. DE NOVELLIS, V. ; VITA, D. ; GATTA, L. ; LUONGO, L. ; BELLINI, G. ; DE CHIARO, M. ; MARABESE, I. ; SINISCALCO, D. ; BOCCELLA, S. ; PISCITELLI, F. et al. The blockade of the transient receptor potential vanilloid type 1 and fatty acid amide hydrolase decreases symptoms and central sequelae in the medial prefrontal cortex of neuropathic rats. Mol Pain. 7:7, 2011.

124
DESAGHER, S.; MARTINOU, J.C. Mitochondria as the central control point of apoptosis. Trends Cell Biol. 10: 369 –377, 2000.
DWORKIN, R.H. et al. Advances in neuropathic pain: diagnostic, mechanism, and treatment recommendations. Arch. Neurosc. v.60, n.11, p.1524-1534, 2003.
ENGEL, C.C. Review: In neuropathy, fibromyalgia, or chronic pain, duloxetine reduces pain but increases adverse events. Ann Intern Med. Apr 15;160(8):JC12,
2014. FERREIRA, S.H.; ROMITELLI, M.; DE NUCCI, G. Endothelin-1 participation in overt and inflammatory pain. J. Cardiov. Pharmacol., 13, 220-22, 1989. FOLTINOVÁ, V.; SVIHALKOVA SINDELEROVA, L; HORVATH, V.; SOVA, P.; HOFMANOVA, J.; JANISCH, R.; KOZUBI, K. A. Mechanisms of effects of platinum (II) and (IV) complexes. Comparison of cisplatin and oxaliplatin with saraplatin and LA-12, new Pt(IV)-based drugs. A minireview. Scripta Med. (BRNO), v. 81, n. 2, p. 105-116, 2008. FUKURODA, T.; OZAKI, S.; IHARA, M.; ISHIKAWA, K.; YANO, M.; MIYAUCHI, T.; ISHIKAWA, S.; ONIZUKA, M.; GOTO, K.; NISHIKIBE, M. Necessity of dual blockade of endothelin ET A and ETB receptor subtypes for antagonism of endothelin-1-induced contraction in human bronchi. Br J Pharmacol. 117: 959–999, 1996.
GAMELIN, E.; GAMELIN, L.; BOSSI, L.; QUASTHOFF, S. Clinical aspects and molecular basis of oxaliplatin neurotoxicity: current management and development of preventive measures. Semin Oncol; n. 29; p.21–33, 2002. GAUDET, R. Divide and conquer: high resolution structural information non TRP channel fragments. J.Gen.Physiol.133, 231–237, 2009. GHONEIM, M.A.; YAMAMOTO, T.; HIROSE, S.; NAGASAWA, T.; HAGIWARA, H. Endothelium localization of ETB receptor revealed by immunohistochemistry. J Cardiovasc Pharmacol. 22(Suppl 8): S111–S112, 1993.
GILRON, I.; BARON, R.; JENSEN, T. Neuropathic Pain: Principles of Diagnosis and Treatment. Mayo Clinic Proceedings. V. 90, Issue 4, April, Pages 532–545, 2015. GRAHAM, J.; MUHSIN, M.; KIRKPATRICK, P. Oxaliplatin. Nature Reviews: Drug Discovery. v.3. Jan., 2013.

125
GRANGER, J.P.; ABRAM, S.; STEC, D.; CHANDLER, D.; LAMARCA, B. Endothelin, the kidney and hypertension. Curr Hypertens Rep 8: 298–303, 2006.
GOURDIER, I.; DEL RIO, M.; CRABBE, L.; CANDEIL, L.; COPOIS, V.; YCHOU, M.; AUFFRAY, C.; MARTINEAU, P.; MECHTI, N.; POMMIER, Y.; PAU, B. Drug specific resistance to oxaliplatin is associated with apoptosis defect in a cellular model of colon carcinoma. FEBS Lett 529: 232–236, 2002.
HANSA, G.; SCHMIDT, B.J. STRICHARTZC, G. Nociceptive sensitization by endothelin-1. Brain Research Review. 36–42, 2009.
HERSHMAN, D.L.; WEIMER, L.H.; WANG, A.; KRANWINKEL, G.; BRAFMAN, L.; FUENTES, D.; AWAD, D.; CREW, K.D. Association between patient reported outcomes and quantitative sensory tests for measuring long-term neurotoxicity in breast cancer survivors treated with adjuvant paclitaxel chemotherapy. Breast Cancer Res Treat. Feb;125 (3):767-74, 2011. HERSHMAN, D.L.; LACCHETTI, C.; DWORKIN, R.H.; LAVOI, E.; SMITH, E.M.; BLEEKER, J.; CAVALETTI, G. et al. Prevention and management of chemotherapy-induced peripheral neuropathy in survivors of adult cancers: American Society of Clinical Oncology clinical practice guideline. J Clin Oncol. Jun 20; 32(18):1941-67, 2014. HINOJOSA-LABORDE, C. ; LANGE, D.L. Endothelin regulation of adrenal function. Clin Exp Pharmacol Physiol. 26: 995–999, 1999.
HOLZER, P. Transient receptor potential (TRP) channels as drug targets for diseases of the digestive system. Pharmacol.Ther.131, 142–170, 2011.
HSIEH. Y.L.; FAN, Y.C.; YANG, C.C. Low-level laser therapy alleviates mechanical and cold allodynia induced by oxaliplatin administration in rats. Support Care Cancer. Publish on line, 2014. HUANG, S.C. Endothelin receptor in gastrointestinal smooth muscle. Curr Protein Pept Sci 6: 547–557, 2005. HUNT, S. P.; PINI, A.; EVAN, G. Induction of c-fos-like protein in spinal cord neurons following sensory stimulation. Nature, v. 328, p. 632-634, 1987.

126
INAGAKI, H.; BISHOP, A.E.; EIMOTO, T.; POLAK, J.M. Autoradiographic localization of endothelin-1 binding sites in human colonic cancer tissue. J. Pathol. 168, 263–
267, 1992. INFARMED, Folheto Informativo: informação para o utilizador. Disponível em:
http://www.infarmed.pt/infomed/download_ficheiro.php?med_id=41735&tipo_doc=fi. Acesso em: 28/04/2015, 2010. INTURRISI, C.E. The role of N-methyl-D-aspartate (NMDA) receptors in pain and morphine tolerance Minerva. Anestesiol. 71, pp. 401–403, 2005.
JAGGI, S, SINGH, N. Mechanisms in cancer-chemotherapeutic drugs-induced peripheral neuropathy. Toxicology. v. 291, p. 1-9, 2012.
JAMIESON, S. M.; LIU, J.; CONNOR, B.; MCKEAGE, M. J. Oxaliplatin causes selective atrophy of a subpopulation of dorsal root ganglion neurons without inducing cell loss. Cancer Chemother. Pharmacol., v. 56, n. 4, p. 391-399, 2005. JANG, J.H. et al. Involvement of peripherally released substance P and calcitonin gene-related in mediating mechanical hiperalgesia in a traumatic neuropathy model of the rat. Neurosci Lett. 360(3):129-32, 2004.
JERGOVA, S.; KOLESAR, D.; CIZKOVA, D. Expression of c-Fos in the parabrachial nucleus following peripheral nerve injury in rats. Eur. J. Pain, v. 12, p. 172-179,
2008. JERREMALM, E.; WALLIN, I.; EHRSSON, H. New insights into the biotransformation and pharmacokinetics of oxaliplatin. J. Pharmaceutical Sci., v. 98, n. 11, 2009. JOHNSTONE, R.W.; RUEFLI, A.A.; LOWE, S.W. Apoptosis: a link between cancer genetics and chemotherapy. Cell. 108: 153– 164, 2002. JULIUS, D. TRP channels and pain. Annu.Rev.Cell.Dev.Biol.29, 355–384, 2013.
KAGIAVA, A.A.; THEOPHILIDIS, C.G.; SARGIANNIDOU, A.I.; KYRIACOU, D.K.; KLEOPAS, A. Oxaliplatin-induced neurotoxicity is mediated through gap junction channels and hemichannels and can be prevented by octanol. Neuropharmacology.
97, 289-305, 2015.

127
KAR, S.; YOUSEM, S.A.; CARR, B.I. Endothelin-1 expression by human hepatocellular carcinoma. Biochem. Biophys. Res. Commun. 216, 514–519, 1995.
KHODOROVA, A.; RICHTER, J.; VASKO, M.R.; STRICHARTZ, G. Early and late contributions of glutamate and CGRP to mechanical sensitization by endothelin-1. J Pain.10(7):740–749, 2009. KOHAN, D.E. Endothelins: renal tubule synthesis and actions. Clin Exp Pharmacol Physiol 23: 337–344, 1996. KIM, Y.O.; KIM, I.J.; YOON, M.H. Antiallodynic effect through spinal endothelin-B receptor antagonism in rat models of complex regional pain syndrome. Neuroscience Letters. 584; 45–49, 2015.
LALICH, M.; MCNEEL, D.G.; WILDING, G.; LIU, G. Endothelin receptor antagonists in cancer therapy. Cancer Invest. 25(8):785–794, 2007.
LANGE, M.; PAGOTTO, U.; RENNER, U.; ARZBERGER, T.; OECKLER, R.; STALLA G.K. The role of endothelins in the regulation of pituitary function. Exp Clin Endocrinol Diabetes. 110: 103–112, 2002. LEE, S.E.; KIM, J.H. Involvement of substance P and calcitonin gene-related in development and maintenance of neuropathic pain from spinal nerve injury model of rat. Neuroscience research. 58(3):245-9, 2007.
LEONARD, G. D; WRIGHT, M; QUINN, M; GREM, J. Survey of oxaliplatinassociated neurotoxicity using an interview-based questionnaire in patients with metastatic colorectal cancer. BMC Cancer. v. 5, 2005. Disponível em: <http://www.ncbi.nlm.nih.gov/pubmed/> Acesso em 2 de fevereiro de 2015. LEVINE, JD; ALESSANDRINI-HASER, N. TRP channels: target for the relief of pain. Biochim biophys, v.1772, p. 989-1003, 2007.
LIEBEN, L.; CARMELIET, G. The involvement of TRP channels in bone homeostasis. Front. Endocrinol. 3, 99, 2012.
LING, B.; AUTHIER, N.; BALAYSSAC, D.; ESCHALIER, A.; COUDORE, F. Behavioral and pharmacological description of oxaliplatin-induced painful neuropathy in rat. PAIN. 3:225-34, 2007.

128
LINO, J.A. Efeito protetor da amifostina na neuropatia sensitiva periférica experimental induzida por oxaliplatina. Dissertação (Mestrado). Universidade
Federal do Ceará. Faculdade de Medicina. Departamento de Fisiologia e Farmacologia. Fortaleza, 2011. LOUPAKIS, F.; CREMOLINI, C.; MAIS, G.; LONARDI, S.; ZAGONEL, V.; SALVATORE, L. et al. Initial therapy with FOLFOXIRI and bevacizumab for metastatic colorectal cancer. N Engl J Med. Oct 23; 371(17):1609-18, 2014. LOWE, S.W.; LIN, A.W. Apoptosis in cancer. Carcinogenesis. 21: 485–495, 2000.
MALMBERG, A.B.; BLEY, K.R. Turning up the heat on pain: TRPV1 receptors in pain and inflammation. Basel (Switzerland): Birkhauser. 2005.
MARSHALL, J. The Clinical use of oxaliplatina: case studies and roundtable discussion. New York: Oncology, 2004. MALLAT, A.; LOTERSZTAJN, S. Multiple hepatic functions of endothelin-1: physiopathological relevance. J Hepatol. 25: 405–413, 1996. MEIDEN, R.; LEVY, N. The ovarian endothelin network: an evolving story. Trends Endocrinol Metab. 18: 379–385, 2007.
MENCARELLI, M.; PECORRELI, A.; CARBOTTI, P.; VALACCHI, G.; GRASSO, G.; MUSCETOOLA, M. Endothelin receptor A expression in human inflammatory cells. Regul Pept. 158: 1–5, 2009. MCWHINNEY, S.R.; GOLDBERG, R. M; MCLEOD, H. Platinum neurotoxicity pharmacogenetics. Mol Cancer Ther; n. 1 p.8. 2009. MIHARA, Y.; EGASHIRA,N.; SADA, H.; KAWASHIRI, T.; USHIO, S.; YANO, T.; IKESUE, H.; OISHI, R. Involvement of spinal NR2B-containing NMDA receptors in oxaliplatin-induced mechanical allodynia in rats. Molecular Pain. 7:8, 2011.
MOLS, F.; BEIJERS, A.; VREUGDENHIL, G.; VERHULST, A.; SCHEP, G.; HUSSON, O. Chemotherapy-induced peripheral neuropathy, physical activity and health-related quality of life among colorectal cancer survivors from the PROFILES registry. J Cancer Surviv. Apr 16, 2015. MORTENSEN, L.H. Endothelin and the central and peripheral nervous systems: a decade of endothelin research. Clin Exp Pharmacol Physiol. 26: 980–984, 1999.

129
MULLOL J, PICADO C. Endothelin in nasal mucosa: role in nasal function and inflammation. Clin Exp Allergy. 30: 172–177, 2000.
MUÑOZ, M.; COVEÑAS, R.; ESTEBAN, F.; REDONDO, M. The substance P/NK-1 receptor system: NK-1 receptor antagonists as anti-cancer drugs. J. Biosci. 40, 441–
463, 2015. MUÑOZ, M.; GONZÁLEZ-ORTEGA, A.; COVEÑAS, R. The NK-1 receptor is expressed in human leukemia and is involved in the antitumor action of aprepitant and other NK-1 receptor antagonists on acute lymphoblastic leukemia cell lines. Investig. New Drugs. 30, 529–540, 2012. NADIN, S. B.; VARGAS-ROIG, L. M.; DRAGO, G.; IBARRA, J.;CIOCCA, D. R. DNA damage and repair in peripheral blood lymphocytes from health individuals and cancer patients: A pilot study on the implications in the clinical response to chemotherapy. Cancer Lett., v. 239, n. 1, p. 84-97, 2006. NAKAYAMA, N.; SATO, A.; TANAKA, S.; SHIMADA, K.; KONISHI, K.; SASAKI, E. et al. A phase II study of bevacizumab with modified OPTIMOX1 as first-line therapy for metastatic colorectal cancer: the TCOG-GI 0802 study. Invest New Drugs. May 5,
2015. NELSON, J.B.; HEDICAN, S.P.; GEORGE, D.J.; REDDI, A.H.; PIANTADOSI, S.; EISENBERGER, M.A.; SIMONS, J.W. Identification of endothelin-1 in the pathophysiology of metastatic adenocarcinoma of the prostate. Nat. Med. 1, 944–
949, 2000. NEUHOFER, W.; PITTROW, D. Role of endothelin and endothelin receptor antagonists in renal disease. Eur J Clin Invest 36: 78–88, 2006. OIKAWA, T., KUSHUHARA, M., ISHIKAWA, S., HITOMI, J., KONO, A., IWANAGA, T., YAMAGUCHI, K. Production of endothelin-1 and thrombomodulin by human pancreatic cancer cells. Br. J. Cancer. 69, 1059–1064, 1994.
OHNAKA, K.; OIKE, M.; TAKAYANGI, R.; YANASE, T.; HAJI, M.; NAWATA, H. The role of endothelin in adrenal medulla: identification of endothelin and its receptor in two cases of pheochromocytoma. Fukuoka Igaku Zasshi 84, 97–99, 1993. PARK, S. B. et al. Mechanisms underlying chemotherapy-induced neurotoxicity and the potential for neuroprotective strategies. Curr. Med. Chem. 15, 3081–3094, 2008.

130
PARK, S. B.; KRISHNAN, A. V.; LIN, C. S.; GOLDSTEIN, D.; FRIEDLANDER, M.;
KIERNAN, M. C. Mechanisms underlying chemotherapy-induced neurotoxicity and
the potential for neuroprotective strategies. Curr. Med. Chem., v. 15, n. 29, p. 3081-
3094, 2008.
PATAPOUTIAN, A.; TATE, S.; WOOLF, C.J. Transient receptor potential channel targeting pain at the source . Nat Rev drug discov , v. 8, p. 55-68, 2009
PEKONEN, F.; SAIJONMAA, O.; NYMAN, T.; FYHRQUIST, F. Human endometrial adenocarcinoma cells express endothelin-1. Mol. Cell. Endocrinol. 84, 203–207,
1992. PETERS, C.M.; LINDSAY, T.H.; POMONIS, J.D.; LUGER, N.M.; GHILARDI, J.R.; SEVCIK, M.A.; MANTYH, P.W. Endothelin and the tumorigenic component of bone cancer pain. Neuroscience. 126(4):1043–1052, 2004.
PICKERING, V.; JORDAN, R.C.; SCHMIDT, B.L. Elevated salivary endothelin levels in oral cancer patients—a pilot study. Oral Oncol. 43, 37–41, 2007.
PICKERING, V.; JAY GUPTA, R.; QUANG, P.; JORDAN, R.C.; SCHMIDT, B.L. Effect of peripheral endothelin-1 concentration on carcinoma-induced pain in mice. Eur J Pain.12(3):293–300, 2008. PIKE, C.T.; BIRNBAUM, H.G.; MUEHLENBEIN, C.E.; POHL, G.M.; NATALE, R.B.; Healthcare Costs and Workloss Burden of Patients with Chemotherapy-Associated Peripheral Neuropathy in Breast, Ovarian, Head and Neck, and Nonsmall Cell Lung Cancer. Chemother Res Pract. 2012:913848, 2012.
PLANT, T.D.; ZOLLNER, C.; MOUSA, S.A.; OKSCHE, A. Endothelin-1 potentiates capsaicininduced TRPV1 currents via the endothelin A receptor. Exp. Biol. Med. 231
1161–1164, 2006. PLANT, T.D.; ZÖLLNER, C.; KEPURA, F. et al. Endothelin potentiates TRPV1 via ETA receptor-mediated activation of protein kinase C. Mol Pain. 3(1):35, 2007. POMONIS, J.D.; ROGERS, S.D.; PETERS, C.M.; GHILARDI, J.R.; MANTYH, P.W. Expression and localization of endothelin receptors: implications for the involvement of peripheral glia in nociception. J. Neurosci. 21 999–1006, 2001.

131
PONTES, R.B. Desenvolvimento de modelo experimental de neuropatia sensitiva periférica induzida pelo agente antineoplásico oxaliplatina em camundongos. Dissertação (Mestrado). Universidade Federal do Ceará. Faculdade de Medicina. Departamento de Fisiologia e Farmacologia. Fortaleza, 2009. PREVARSKAYA, N.; ZHANG, L.; BARRITT, G. TRP channels in cancer. Biochim Biophys Acta. 1772:937-946, 2007.
PUNTAMBEKAR, P.; VAN BUREN, J.; RAISINGHANI, M.; PREMKUMAR, L.S.; RAMKUMAR, V. Direct interaction of adenosine with the TRPV1 channel protein. J Neurosci. 24:3663-3671, 2004. QU, X-X.; CAI, J.; LI, M. J.; CHI, Y. N.; LIAO, F. F.; LIU, F. Y.; WAN, Y.; HAN, J. S.; XING, G. G. Role of the spinal cord NR2B-containing NMDA receptors in the development of neuropathic pain. Exp. Neurol., v. 215, n. 2, p. 298-307, 2009.
QUANG, P.N.; SCHMIDT, B.L. Periferal endothelin B receptor agonist-induced antinociception involves endogenous opioids in mice. Pain. 142(2): 254-262, 2010.
QUARTU, M. et al., Bortezomib treatment produces nocifensive behavior and changes in the expression of TRPV1, CGRP and Substance P in the rat DRG, spinal cord, and sciatic nerve. BioMed Reseach International, 2014. RAFFA, R.B.; SCHUPSKY, J.J.; JACOBY, H.I. Endothelin-induced nociception in mice: mediation by ETA and ETB receptors. J. pharmacol. Exp. Ther., 276, 647-651, 1996. RAMSEY, I.S.; DELLING, M.; CLAPHAM, D.E.An introduction to TRP channels. Annu. Rev.Physiol.68,619–647, 2006.
RASHID, M.H.; INOUE, M.; BAKOSHI, S.; UEDA, H. Increased expression of vanilloid receptor 1 on myelinated primary afferent neurons contributes to the antihyperalgesic effect of capsaicin cream in diabetic neuropathic pain in mice. J Pharmacol Exp Ther. 306:709-717, 2003a.
RASHID, M.H.; INOUE, M.; KONDO, S.; KAWASHIMA, T.; BAKOSHI, S.; UEDA, H. Novel expression of vanilloid receptor 1 on capsaicin-insensitive fibers accounts for the analgesic effect of capsaicin creams in neuropathic pain. J Pharmacol Exp Ther
304:940-948, 2003b.

132
RICHARDS, J.S.; BOMBARDIER, C.H.; WILSON, C.S.; CHIODO, A.E.; BROOKS, L.; TATE, D.G.; TEMKIN, N.R.; BARBER, J.K.; HEINEMANN, A.W.; McCULLUMSMITH, C.; FANN, J.R. Efficacy of venlafaxine XR for the treatment of pain in patients with spinal cord injury and major depression: a randomized, controlled trial. Arch Phys Med Rehabil. Apr;96(4):680-9, 2015. ROSA, F. M.; ESCOBAR, C. A. B.; BRUSCO, L. C. Parestesia dos nervos alveolar inferior e lingual pós cirurgia de terceiros molares. RGO, Porto Alegre, v. 55, n.3., p. 291-295, jul./set. 2007. ROSSI, G.P. ; ANDREIS, P.G. ; COLONNA, S. ; ALBERTIN, G. ; ARAGONA, F.; BELLONI, A.S. ; NUSSDORFER, G.G. Endothelin-1[1–31]: a novel autocrine-paracrine regulator of human adrenal cortex secretion and growth. J Clin Endocrinol Metab. 87: 322–328, 2002.
ROSSI, G.P. ; BELLONI, A.S. ; NUSSDORFER, G.G. PESSINA, A.C. Endothelin and the adrenal gland. J Cardiovasc Pharmacol. 35: S17–S20, 2000.
ROSSO, M.; ROBLES-FRÍAS, M.J.; COVEÑAS, R.; SALINAS-MARTÍN, M.V.; MUÑOZ, M. The NK-1 receptor is expressed in human primary gastric and colon adenocarcinomas and is involved in the antitumor action of L-733,060 and the mitogenic action of substance P on human gastrointestinal cancer cell lines. Tumor Biol. 29, 245–254, 2008. RUSSELL, F.D.; MOLENAAR, P. The human heart endothelin system: ET-1 synthesis, storage, release and effect. Trends Pharmacol Sci 21: 353–359, 2000. SAIF, M.W.; REARDON, J. Management of oxaliplatin-induced peripheral neuropathy. Ther Clin Risk Manag. v 1(4); dec, 2005. SAKURAI-YAMASHITA, Y.; YAMASHITA, K.; YOSHIDA, A. et al. Rat peritoneal macrophages express endothelin ET (B) but not endothelin ET (A) receptors. Eur J Pharmacol. 338(2):199–203, 1997.
SCHIFF, D.; WEN, P. Y.; VAN DEN BENT, M. J. Neurological adverse effects caused by cytotoxic and targered therapies. Nat. Rev. Clin. Oncol., v. 6, p. 596-603,
2009. SCHIFFRIN, E.L.; TOUYZ, M.R. Vascular biology of endothelin. J Cardiovasc Pharmacol. 32, S2-13, 1998.

133
SCHMIDT, B.L.; PICKERING, V.; LIU, S.; QUANG, P.; DOLAN, J.; CONNELLY, S.T.; JORDAN, R.C. Peripheral endothelin a receptor antagonism attenuates carcinoma-induced pain. Eur J Pain. 11(4):406–414, 2008.
SHAH, N.; DIZON, D.S. New-generation platinum agents for solid tumors. Future Oncol 5:33-42, 2009.
SHANKAR, A.; LOIZIDOU, M.; ALIEV, G.; FREDERICKS, S.; HOLT, D.; BOULOS, P.B.; BURNSTOCK, G.; TAYLOR, I. Raised endothelin 1 levels in patients with colorectal liver metastases. Br. J. Surg. 85, 502–506, 1998. SHETTY, S.S.; OKADA, T.; WEBB, R.L.; DELGRANDE, D.; LAPPE, R.W. Functionally distinct endothelin B receptors in vascular endothelium and smooth muscle. Biochem Biophys Res Commun. 191(2):459–464, 1993.
SHINODA, M.; OGINO, A.; OZAKI, N.; URANO, H.; HIRONAKA, K.; YASUI, M.; SUGIURA, Y. Involvement of TRPV1 in nociceptive behavior in a rat model of cancer pain. J Pain. 9: 687–699, 2008. SCHUMACHER, M.A.; Transient receptor potential channel in pain and inflammation therapeutic opportunities. Pain pract v. 10: p.185-200, 2010
SINGH, D.; JOSHI, D.D.; HAMEED, M.; QIAN, J.; GASCÓN, P.; MALOOF, P.B; MOSENTHAL, A.; RAMESHWAR, P. Increased expression of preprotachykinin-I and neurokinin receptors in human breast cancer cells: implications for bone marrow metastasis. Proc. Natl. Acad. Sci. USA 97, 388–393, 2000.
SINISCALCO, D.; DE NOVELLIS, V.; ROSSI, F.; MAIONE, S. Neuropathic pain: is the end of suffering starting in the gene therapy? Curr Drug Targets. 6:75-80, 2005.
SISIGNANO, M.; BARON, R.; SCHOLICH, K.; GEISSLINGER, G. Mechanism-based treatment for chemotherapy-induced peripheral neuropathic pain. Nat Rev Neurol.
Dec; 10(12):694-707, 2014. SMITH, T.; BEASLEY, S.; SMITH, S.; MARK, I.; SWEITZER, S.M. Endothelin-1- induced priming to capsaicin in young animals. Neurosci Lett. 567:15–18, 2014. SMITH, E. M. L.; BECK, S. L; COHEN, J. The Total Neuropathy Score: A Tool for Measuring Chemotherapy-Induced Peripheral Neuropathy. Oncology Nursing Forum. v. 35, n. 1,. p . 96-102, 2008.

134
SMITS, H.; KLEEF, M. V.; HONIG, W.; GERVER, J.; GOBRECHT, P.; JOOSTEN, E. A. Spinal cord stimulation induces c-Fos expression in the dorsal horn in rats with neuropathic pain after partial sciatic nerve injury. Neurosci. Lett., v. 450, n. 1, p. 70-73, 2009. STEIN, A.; ARNOLD, D. Oxaliplatin: a review of approved uses. Expert Opin Pharmacother. 13(1):125–37, 2012.
STILLMAN, M.; CATA, J.P. Management of chemotherapy-induced peripheral neuropathy. Curr. Pain Headache Rep., v. 10, p. 279-287, 2006.
SZALLASI, A.; CORTRIGHT, D.N.; BLUM, C.A.; EID, S.R. The vanilloid receptor TRPV1: 10 years from channel cloning to antagonist proof-of-concept. Nat Rev Drug Discov. 5: 357-72, 2007. TA, L.E; BIEBER, A.J.; CARLTON, S.M.; LOPRINZI, C.L.; LOW, P.A.; WINDEBANK, A.J. Transient Receptor Potential Vanilloid 1 is essential for cisplatin-induced heat hiperalgesia in mice. Molecular Pain. 6:15, 2010.
VELASCO; R; BRUNA, J. Chemotherapy-induced peripheral neuropathy: an unresolved issue. Neurología. v. 2, n. 25,.p.116-131. 2010.
WOOLF, C.J.; SALTER, M.W. Neuronal plasticity: increasing the gain in pain Science. 288, pp. 1765–1769, 2000.
WONG, G.Y.; GAVVA, N.R. Therapeutic potential of vanilloid receptor TRPV1 agonists and antagonists as analgesics: Recent advances and setbacks. Brain, v 60,
p 77-267, 2009 YAMAMOTO, H.; KAWAMATA, T.; OMOTE, K.; NAMIKI, A. Endothelin-1 enhances capsaicin-evoked intracellular Ca2+ response via activation of endothelin a receptor in a protein kinase Cepsilon-dependent manner in dorsal root ganglion neuron, Neuroscience. 137. 949–960, 2006.
YAMAMOTO, T.; SHIMOYAMA, N.; ASANO, H.; MIZUGUCHI, T. Analysis of the role of endothelin-A and endothelin-B receptors on nociceptive information transmission in the spinal cord with FR139317, an endothelin-A receptor antagonist, and sarafotoxin S6c, an endothelin-B receptor agonist. J. Pharmacol. Exp. Ther. 271,
156–163, 1994.

135
YAMASHITA, J.; OGAWA, M.; NOMURA, K.; MATSUO, S.; INADA, K.; YAMASHITA, S.; NAKASHIMA, Y.; SAISHOJI, T.; TAKANO, S.; FUJITA, S. Interleukin 6 stimulates the production of immunoreactive endothelin 1 in human breast cancer cells. Cancer Res. 53, 464–467, 1993. YANAGISAWA, M.; KURIHARA, H.; KIMURA, S. et al. A novel potent vasoconstrictor peptide produced by vascular endothelial cells. Nature. 332: 411-415, 1988.
ZAANAN, A. et al. Therapeutic implications of DNA mismatch repair in adjuvant colorectal cancer chemotherapy. Colorectal cancer. 2(1). 51-59-2013.
ZHANG X.; HUANG, J.; MCNAUGHTON, P.A. NGF rapidly increases membrane expression of TRPV1 heat-gated ion channels. EMBO J. 24:4211-4223, 2005.
ZHENG, J. Molecular mechanism of TRP channels. Compr.Physiol.3, 221–242, 2013.

136
ANEXO

137





























