Embed Size (px)
Citation preview

UNIVERSIDAD A U T ~ N O M A METROPOLITANA UNIDAD IZTAPALAPA
DIVISIóN DE CIENCIAS BIOLóGICAS Y DE LA SALUD
LICENCIATURA EN BIOLOGÍA EXPERIMENTAL
REPORTE DEL PROYECTO DOCENTE DE INVESTIGACIóN
ALUMNA EN FORMACIóN:
Srita. María del Pilar Jimenez Aguilera Alumna de la licenciatura en Biología Experimental, con número de matrícula 96228389, Division de Ciencias Biol6gicas y de la Salud. UAM - Iztapalapa, Te1 2 1-57- 12-82.
TÍTULO DEL PROYECTO:
EVALUACI6N DE ALGUNOS FACTORES QUE MODIFICAN LA EXPRESI6N DE LA AROMATASA EN CÉLULAS DE LEYDIG DE RATAS MACHO DE
DISTINTAS EDADES.
ASESOR:
M. en B.E. Joaquín Fernando Herrera Muñoz Prof. Titular C, Medio Tiempo Departamento de Ciencias de la Salud, Universidad Autónoma Metropolitana - Iztapalapa.
Investigador Titular A Unidad de Investigaci611 en Medicina Reproductiva Hospital Luis Castelazo Ayala Instituto Mesicano del Seguro Social
LUGAR DE R E A L I Z A C I ~ N DEL PROYECTO:
Laboratorio de Biología Molecular y Regulaci6n Testicular Departamento de Ciencias de la Salud, Universidad Aut6noma Metropolitana - Iztapalapa.
Unidad de Investigaci6n en Medicina Reproductiva Hospital Luis Castelazo Ayala Instituto Mexicano del Seguro Social
Julio de 200 1.

E V A L U A C I ~ N DE ALGUNOS FACTORES QUE MODIFICAN LA EXPRESIONDE LA AROMATASA EN CGLULAS DE LEYDIG DE
RATAS MACHO DE DISTINTAS EDADES.
INTRODUCCIbN
ANATOMIA DEL APARATO REPRODUCTOR MASCULINO
En la mayoría de los mamíferos el sistema reproductor masculino se encuentra constituido por las glándulas sexuales y los 6rganos accesorios. Est& formado por 4 componentes con funciones diferentes (Fig. 1).
1. Los testiculos o g6nadas que cumplen con una doble funci611, endocrina y de producción celular; ya que actúan tanto en la producci6n de células germinales (espermatozoides) como en la producci6n de hormonas sexuales.
2. Una complicada red de túbulos, consistentes en conductos eferentes, epidídimo, conducto deferente y conducto eyaculador.
3. Un grupo de gldndulas exocrinas que contribuyen a la secreción de fluidos seminales, tales como: vesícula seminal, glkndula prostdtica y un par de glhndulas bulbouretrales accesorias.
4. Un 6rgano copulador, el pene (Fawcett, 1995).
Figura 1. Aparato Rcproductor Masculino.
ANATOMIA TESTICULAR 1
Los testículos de desarrollan en la cavidad abdominal, descendiendo hacia el escroto poco antes o despues del nacimiento, en la parte interna de esta proyeccidn de la piel se encuentran revestidos anterior y lateralmente por una cavidad serosa que es derivada del peritoneo parietal, la trirlicn vngilznlis; inmediatamente despub hay una cavidad virtual que separa a la túnica vaginalis del testículo, el cual estd cubierto por una capa fibrosa llamada trirricn nlbugilzen, y finalmente lo

cubre la t?ílricn vascularis que está formada por redes de vasos sanguíneos incluidos en el tejido conectivo fibroso. En la superficie posterior de la túnica albuginea existe una zona de tejido conjuntivo denso que se extiende a corta distancia desde la propia túnica albuginea y que forma el ?nedinstim testicztlnr, a traves del cual se introducen los vasos sanguíneos y salen los condlrctillos e j r o l t e s del 6rgano. En el interior, desde el mediastino hasta la túnica albuginea se extiende una serie de tabiques fibrosos llamados séptupla testis que dividen al parenquima testicular en aproximadamente 250 compartimentos de forma piramidal, denominados lobzllillos fesficulares; los lobulillos se comunican entre sí a traves de perforaciones existentes en los tabiques fibrosos, pero están completamente separados en la zona de convergencia del mediastino (Fawcett, 1995).
Cada lobulillo contiene entre 1 y 4 tlibltlos s e l ~ ~ i n f e r o s , que son tortuosos tienen un digmetro de 150 -250 pm y una longitud de 30 a 70 cm. En el estrecho Apex de cada lobulillo piramidal se produce la transici6n abrupta de sus túbulos seminiferos hacia los hihulos rectos, los cuales convergen en la denominada rete festis. Desde los túbulos rectos sale una serie de conductillos eferentes a traves de los cuales los espermatozoides alcanzan el coldzrcfo e,uididi?lrnrio. Cada túbulo seminíferos está rodeado por una densa red capilar y linfática, y el tejido conjuntivo laxo areolar que lo rodea contiene células mesenquimales perivasculares, fibroblastos y algunos macrófagos, que aparecen sobre una trama de finas fibras reticulares y de colágeno (Fawcett, 1995). (Fig.?)..
Figura 2 Esquema de la arquitectura del testículo y del sistema ductal cscretor tornado de Martin Dym 1975.
En el adulto, el testículo es una glándula con funciones endocrinas y de producción celular, presenta dos componentes funcionales importantes: A) los túbulos seminiferos (compartimiento tubular), y B) el compartimiento intersticial.
A) COMPARTIMIENTO TUBULAR.
Los túbulos seminiferos esthn formados por una gruesa Ihmina basal que a su vez esta rodeada por 3 6 4 capas de las aplanadas céfdns Pcritubulares (CP), y que guardan alguna semejanza con las celulas de músculo liso, por lo que tambien se denominan ttrioides; por dentro de la lamina basal los túbulos se encuentran revestidos por epitelio estratificado que contiene dos estirpes celulares: las céhllns de In linen gemritznl (CG) y las multifuncionales célztlns de Sertoli (S); las cuales generan y mantienen tanto la arquitectura tubular requerida, como el microambiente especial necesario para el desarrollo y maduraci6n de las celulas de la linea germinal, a traves de la

produccidn de esteroides activos, gomo la 5a-dihidro testosterona y el estradiol (Ez); de la síntesis regulada de y uiía grin variedadr de polipéptidos con actividad biol6gica ya sea de mensaje o actividad enzimdtica, amen de otros productos metab6Iicos que se piensa nutren a las células de la línea germinal (Jégou, 1992).
Las celulas de Sertoli secretan proteínas fijadoras de andr6genos (ABP), inhibina, y en etapas embrionarias la sustancia inhibidora de conductos de Muller (SIM). Desde el aspecto endocrino, no sintetizan andr6genos de muo, pero contienen aromatasa, que es la enzima causante de la conversi611 de los andr6genos a estr6genos. La ABP actúa probablemente para mantener un abastecimiento alto y estable de andr6genos en el líquido seminal tubular. La inhibina impide la secreci6n de la FSH en la hip6fisis. La SIM produce regresibn de los conductos mullerianos en machos durante la vida embrionaria (Ganon, 1998).
B) COMPATIMIENTO INTERSTICIAL
El compartimiento intersticial se encuentra formado principalmente de tejido conectivo laxo conteniendo vasos linfdticos y sanguíneos y las llamadas célrrlns de teydig (CL), las cuales son células poligonales, que generalmente se encuentran en cúmulos asociados a vasos sanguíneos y que representan aproximadamente el 5% del total celular. (Ewing y Keeney, 1993). Las CL son los principales componentes endocrinos del testículo (Wilson & Foster, 1992). Las CL no presentan gránulos de secrecibn, ya que su secreci6n es de naturaleza lipídica (Fawcett, 1995).
Las CL sintetizan y liberan la hormona masculina llamada Testosterona (T). Esta estirpe celular involucra al menos dos generaciones de celulas: (a) la generaci6n celular de tipo fetal que es responsable de la masculinizaci6n y por lo tanto de la aparici6n de las características sexuales primarias durante la vida fetal y neonatal, y (b) l a poblaci6n de tipo adulto, la cual es responsable de la masculinizaci6n durante la pubertad, la aparici6n y permanencia de las características sexuales secundaras y del funcionamiento masculino en general (Gondos, 1980).
En las ratas las CL fetales aparecen aproximadamente a los 13 días derla regi6n intersticial, por diferenciaci6n de las células somdticas mesenquimales. Estas celulas contienen grandes mitocondrias, gotas de lípidos y retículo endopldsmico liso (Magres y Jost, 1980). Estas células a los 30 días de desarrollo fetal alcanzan un máximo de 1Oj células por testículo y posteriormente después del nacimiento y durante las dos semanas de vida postnatal, su número disminuye claramente (Zirkin y Ewing, 1987).
La evoluci6n de las CL de tipo fetal y las evidencias de su regresi6n funcional y morfol6gica después de estas semanas son desconocidos; por lo tanto, tampoco se tiene de una explicaci6n de este fen6men0, por lo que se ha sugerido que la regresibn tanto morfol6gica como funcional de las CL fetales es controlada por una regulaci6n autocrina o paracrina (Saez, 1994).
Las CL de tipo adulto aparecen en el dkimo día de nacimiento, e incrementan considerablemente su número despues de día 15 hasta llegar a 2.5 x 106 celulas por testículo al finalizar la pubertad. Las CL son formadas principalmente por diferenciacibn de células mesenquimales, pero tambien por divisi6n de CL preexistentes (Hardy y cok, 1989).
En el humano la diferenciaci6n de las g6nadas masculina aparece a la sexta semana de gestaci6n, y su citodiferenciaci6n comienza con un incremento en el volumen citoplasmdtico, desarrollo del retículo endopldsmico liso, un incremento en el número y tamaiio de las mitocondrias, agrandamiento del núcleo y acumulaci6n de gotas de lípidos. Los cristales de Reinke nunca son formados en las CL fetales y son solamente observados en el adulto. Durante la infancia, el espacio intersticial es poblado por cklulas mesenquimales estrelladas y fusiformes con núcleos

elongados y citoplasma visible que son considerados precursores celulares no difernciadas. Durante la pubertad estas células se diferencian a CL adultas bajo el control de la Hormona Luteinizante (LH) (Prince, 1984).
Este proceso de diferenciaci6n involucra la aparici6n de un marcado incremento del retículo endoplásmico liso, el cual consiste en túbulos con membranas que lo limitan y que contiene a la mayoría de las enzimas que participan en la biosíntesis de Testosterona a partir del colesterol (Martin y cols, 1990). El retículo endopldsmico rugoso también est6 disperso en la célula y se interconecta con el liso. Las mitocondrias son de un tamaño moderado y contienen crestas de forma laminar así como tubulares. El aparato de Golgi se encuentra sumamente desarrollado, y en la CL se presentan gotas de lípidos, hosomas, cristales de Reinke, así como tipicos cambios en la morfología nuclear (Connell y Connell, 1977).
En el hombre las CL de tipo adulto aparecen en el décimo día de nacimiento e incrementan considerablemente su número después de día 15 hasta llegar a 2.5 x 106 celulas por testiculo, al finalizar la pubertad el número de CL se incrementa y hacia los 20años llega a alcanzar un mdximo de 5 x 10s células por testiculo, y aunque su número gradualmente disminuye a los 60 años de edad, se encuentran funcionales hasta mas alld de los 80 años (Neaves y cols, 1984). Las CL son formadas principalmente’ por diferenciaci6n de células mesenquimales, pero también por divisi6n de CL preesistentes (Hardy y cols, 1989).

Como se observa en la Figura3, las CL son capaces de sintetizar andr6genos de lmuo a partir del colesterol (Col), el cual es bansformado en el interior de la mitocondria por la enzima llamada C20,22 desmolasa, dependiente del Citocromo P450 (CytP45o SCC), para dar origen a la Pregnenolona (Preg), la cual se transforma a su vez en T mediante la acci6n de un conjunto de enzimas, agrupadas dentro de los intermediarios de las vías A4 o A5- (Risbridger, 1992). En el testículo de los roedores, los humanos u otros mamíferos, la vía de los A 4 intermediarios es predominante, y comprende la transformaci6n inicial de la Preg (un compuesto A5) en progesterona (Prog), mediada por la acci6n de la 3P-hidroxiesteroide deshidrogenasa / A4 - A5-isomerasa. La Prog es posteriormente hidroxilada en posici6n 17a y convertida en androstendiona (A) por el complejo enzimático microsomal de la 17a-hidroxilaqa/Cl7,20 desmolasa dependiente del Citocromo PJjo (CytP4sol7P) y finalmente la A es convertida en T por la 170-hidroxiesteroide deshidrogenasa (Miller, 1988).
La actividad enzimhtica responsable de la síntesis de estr6genos es llamada complejo de la aromatasa o estr6geno sintetasa (dependiente del CytPJwaro) y se encuentra localizada en el retículo endoplásmico. La aromatasa cataliza la conversi6n de los andr6genos (A y T) a los estrógenos correspondientes (Estrona (El) y Ez), a través de una hidroxilaci6n en el grupo metilo del C19, seguida de una oxidaci6n con perdida de Hidr6genos para formar un aldehído en el C19 y una tercera que involucra al C1, las cuales resultan en la pérdida del primer C y la consecuente aromatizaci6n del anillo A de los andrbgenos (Fishman 6r Goto, 1981).
EJE DE REGULACION HIPOTALAMO-HIPOFISISTESTICULO.
La funci6n testicular está controlada por el eje hipotálamo-hipófisis-g6nada, en donde el hipotálamo actúa como el centro integrador de la informacibn que es recibida de la corteza cerebral Y que modula la actividad de las neuronas peptidérgicas hipotalámicas, las cuales se encargan de l a síntesis y secreci6n de la Hormona Liberadora de Gonadotropinas (GnRH), la cual es transportada por el sistema porta hipofisiario hasta la hip6fisis, donde se une a receptores de membrana con alta afinidad en la superficie celular de los gonadotropos, células encargadas de sintetizar y secretar las gonadotropinas las cuales viajan por la circulación general hasta encontrar s u órgano blanco, el testículo (\Vilson y Foster, 1992) (Fig. 4).
Las gonadotropinas liberadas a la circulaci6n periférica modulan la actividad de sus células blanco a través de su interacci6n con los receptores membranales específicos que en forma general están ligados al sistema de la Adenilato Ciclasa.; haciendo que se manifieste la actividad enzimatica, por lo que se incrementa el contenido intracelular del nucle6tido cíclico AMPc, el cual actúa como segundo mensajero; esto desencadena cambios en los patrones de fosforilaci6n proteica mediados por las proteínas cinasas A, activando o desactivando rutas metab6licas y por lo tanto generando modificaciones en las funciones particulares de cada tipo celular.

Tirbulo Sernlnlfero I
Fig4. Eje de regulación Hipot¿ilarno-Hipófisis-G6nada hlodificado de Pesconitz 1994.
La L H se une a los receptores específicos en la membrana plasmática de las CL, presentando elevadas constantes de afinidad y un numero limitado de sitios de unión. Estas proteínas receptoras transducen el mensaje hacia el interior celular interactuando un sistema de proteínas que unen nucle6tidos de guanina (proteínas G estimuladoras (Gs)) que son capaces de activar la adenilato ciclasa e incrementar los niveles intracelulares de AMPc. Este nucleótido se une a una subunidad catalítica libre de la proteína cinasa A (PKA), la cual cataliza la fosforilación de algunas proteínas intracelulares, activando la secuencia de eventos involucrados en la biosíntesis de esteroides.
La liberación de L H por parte de la adenohip6fisis es regulada por la concentración serica de T mediante un mecanismo de retroalimentaci6n negativo: Esto ocurre a través de la acci6n inhibidora directa sobre la adenohip6fisis o bien mediante una inhibici6n de la síntesis hipotalamica de GnRH (Cheng y cols.,1976).
Por su parte la FSH se une a los receptores específicos en la membrana plasmhtica de las C S , y a traves de las protefnas G, ya sea estimuladoras (Gs) 6 inhibidoras (Gi), estas activan a la adenilato ciclasa provocando que se activen protefnas cinasas dependientes de AMPc desencadenado una cascada de eventos de fosforilaci6n proteica. Se dice que al regularse la adenilato ciclasa por medio de las proteínas G, se estimulan tanto la actividad de la RNA Polimerasa, así como la síntesis de la proteína unidora de andr6genos (ABP) y la transferrina.;

también puede activar canales de Calcio directa 0 indirectamente de la fosforilaci6n de sus proteínas vía la subunidad catalítica de la p m dependiente de AMPc.
ANTECEDENTES
Los experimentos realizados por diversos investigadores utilizando CS aisladas a partir de ratas de 20 días de edad, y posteriormente mantenidas i n vitro permitieron plantear la hip6tesis de que éstas eran la fuente principal de la actividad de aromatasa, y que la producci6n de E2 a partir de T ex6gena añadida a estos cultivos primarios de CS podía ser estimulada por la presencia en el medio de hormona folículo estimulante (FSH) (Dorrington ¿k Armstrong, 1975; Dorrintong et al, 1978; Mercante et al 1979). Asimismo, varios autores han sugerido que la capacidad de síntesis de E. en las CS se pierde con la edad, un fenómeno relacionado probablemente con la pérdida de la capacidad mitótica de esta estirpe celular (Rommerts et al, 1982; Payne et al, 1987; Jégou, 1992), si bien existen datos que señalan que las CS mantienen una producci6n considerable de E. aún en animales maduros (Promerantz, 1980; Tsai-Morris et al, 1985; Mendieta et al 1995).
Otros estudios demostraron que la producci6n de E. podía ser detectada en el tejido intersticial de ratas prepúberes (Tsai-Morris et al, 1985) y adultas (Valladares ¿k Payne, 1979). A diferencia de lo que sucede en los túbulos seminíferos, la actividad de la aromatasa en las CL parecería incrementarse con la edad (Valladares & Payne, 1981; Rommerts et al, 1982), así como por la acci6n de diversos factores, como por ejemplo LH o gonadotropina cori6nica humana (hCG), 3’,5’-adenosinmonofosfato cíclico (AMPc), insulina (Ins) o factor de crecimiento epidérmico (EGF) (Payne et al, 1987; Rigaudiere et al, 1989; Syed et al, 1991).
Como se sabe la actividad secretora de las CL se encuentra regulada por la LH, mientras que la FSH influye principalmente sobre la actividad de las CS por Io tanto, sobre el mantenimiento del microambiente que permite la diferenciación de las células de l a línea germinal (Skinner, 1991; l‘erhoeven, 1992).
Existe evidencia cada vez más concluyentes de que la regulaci6n final de la función testicular se lleva a cabo en forma local, a través de interacciones de tipo paracrino y autocrino que involucran la existencia de contactos intracelulares, así como las interacciones mediadas a través de factores de diversa naturaleza, presumiblemente mediante la participaci6n de algunos de los mecanismos de traducci6n descritos en otros modelos celulares (Risbridger, 1992).
En los últimos arios en el grupo de investigaci6n al que me estoy incorporando se han interesado en tratar de resolver algunos aspectos de fisiología testicular. Con relaci6n a la funci6n de las CL se ha demostrado la presencia de al menos un factor proveniente de las CS capaz de disminuir la secreci6n de T en las CL (Herrera y Cols 1996), tambien se ha demostrado que ese factor es de naturaleza proteica (Bustos Rosas, 1991) y que tiene un peso molecular aparente de 21 KD (Ruiz-Paniagua, 1994).
Paralelo a este estudio, el inter& se ha dirigido hacia la determinacih de los mecanismos finos de regulaci6n de la producci6n de la actividad moduladora de esteroidoghesis (AME), y en consecuencia al estudio del control de las diferentes actividades funcionales tanto de las CS, como de las CL. Así como parte de los factores que regulan a la producci6n de la AME por las CS (Perdomo Esquive1 et al, 1996).
Mientras que en las CL, tambien se han realizado estudios acerca del mecanismo a traves del cual la AME modula la actividad de la vía esteroidogenica (Bustos et al, 1996), así como tanlbien en lo que corresponde a la regulaci6n de producci6n de T por factores tales como la melatonina

(Mendieta et al, 1991) el E2 (Herrera et al, 1991), o algunos neurotransmisores (Herrera et al, 1995). Txnbién s6 ha demostrado el efecto de algunos factores que influyen sobre la actividad de la aromatasa presente en las C& como es el dibutiril-AMPc, el cual aumenta su actividad despues de 24 hrs. de manera dosis dependiente de CS humanas en el cultivo. Como ya se sabe el AMPc es el segundo mensajero del mecanismo de regulaci6n de las células de Leydig dependiente de la LH. Lo cual no pasa en CS de ratas inmaduras ya que no presentan respuesta a hCG (Gonadotropina cori6nica humana) o a dibutiril-AMPc (Rommerts, 1987).
Otro agente como la insulina, la cual es una hormona de uso terapéutico muy importante y se tienen reportes de que en las células de la granulosa de rata y humano, puede estimular la actividad de la aromatasa, haciendo que disminuya la producci6n de testosterona, esto puede deberse a que la insulina afecta la concentración intracelular de AMPc haciendo que disminuya.
También se ha observado que al adicionar al factor de crecimiento semejante a la Insulina-I (IGF-I) en cultivos de CL humanas procedentes de pacientes maduros orquidectomizados y estimuladas con hCG, se observ6 un efecto potenciado por IGF-I sobre la producci6n de testosterona sugiriendo un papel regulador en la esteroidogénesis, en CL de rata adulta se ha evidenciado la existencia de receptores específicos para este factor ademis de ser importante en la maduración testicular (Gelhr., 1992; Lin, 1987).
El factor de crecimiento epidérmico (EGF) es un potente agente mitogénico, siendo sus principales células blanco las células del epitelio, mesenquimatosas y de la glia. Sin embargo, en otros tipos celulares puede no estimular la proliferaci6n, pero sí regular funciones específicas; tal es el caso de los cultivos primarios de células testiculares y algunas líneas clonadas de CL tumorales, en donde provocan la disminuci6n de la produccidn de T estimulada por gonadotrofinas en células de rata o en la producci6n progesterona en CL tumorales de rata. En cultivos primarios de células testiculares disminuye la actividad de 17a-hidroxilasa y C17, 20 liasa; en CL tumorales reduce él numero de receptores para hCG (Helding, 1989; Welsh, 1982)
Recientemente se ha realizado un estudio en CL de ratas maduras con el fin de saber como influyen algunos compuestos en la producción de Estradiol, así como también en la expresi6n del gen del citocromo P4joarom; se ObSeN6 que la producci6n de E? aumenta de manera tal que se relacióna con la dosis. En este estudio se utili26 testosterona, DI", 17P-estradiol, medios condicionados con híbulos seminíferos(STM), LHo (LH de ovino) Ó BU)^ AMPc.
Se observ6 que al agregar 200 ng/mL de T, había una producci6n mhxima de Ez; esto es que se incrementa un 28%; siendo semejante al adicionar 1 ng/mL de LHo ; esto también sucedi6 al agregar (Bu)zAMPc a una concentraci6n de ImM. Esto tambien se correlacion6 con los niveles de RNAm de la Pwarom, observindose un decremento en la presencia de una baja concentraci6n (50 y 100 ng/mL) de T; mientras que se registr6 un incremento del 20% en la cantidad de la transcripci61-1, a concentraciones altas (200 - 500 ng/mL) de T. Se obtuvo el mismo resultado con la presencia de 5a- dihidrotestosterona (un andr6geno resistente a la actividad de aromatasa).
Pero al adicionar LH de ovino (LHo 0.1 -5.0 ng/mL), los niveles de RNAm cambian totalmente; esto es que decrecen escasamente en presencia de bajas concentraciones de LHo (0.1 -1 ng /mL). Los niveles de RNAm son incrementados por LHo (5 - 10 ng/mL) en un 35 y 75 X respectivamente en la ausencia y presencia de testosterona siendo el control CL incubadas sin tratamiento. Agregando concentraciones de 50 ng/mL de LHo , se observ6 un incremento al doble en el nivel del RNAm de P4soarom , ya sea con o sin testosterona. El dirbutiril AMPc (1ntM) minimiza el efecto de LHo.

Dentro de este mismo estudio se tom6 en cuenta la vida media del RNAm para saber como influía en la expresi6n del gene. La vida medía del RNAm de la P450arom es incrementada al doble en presencia de testosterona y LH; esto se compar6 con la vida media en la ausencia del tratamiento (5.8 f 0.6 hs). Tomando todos los datos, se demostr6 que en las CL recien aisladas de testículos de ratas maduras; la regulacibn de la expresidn de la aromatasa, así como su actividad enzim&tica, estdn bajo la influencia de LH (por medio del AMPc) y el control esteroideo, ademds de los factores secretados por los túbulos seminíferos que tambien esthn involucrados en este proceso. Por Io tanto la aromatasa de las CL de ratas es controlada en ambos pasos transcripcional y postranscripcionalmente por la producci6n de moduladores endocrinos y/o locales (Genissel y cols., 2001).

JUSTIFICACION
Tomando en consideraci6n a lo planteado, el grupo de trabajo al cual me he integrado, tiene como- objetivo, el estudio en forma sistemhtica de los mecanismos de regulaci6n de la actividad de la aromatasa en la CL; así como demostrar que la expresibn de la aromatasa se modifica en las CL con forme a la edad
OBJETIVOS.
OBJETIVO GENERAL.
Demostrar la existencia de la actividad de la aromatasa en cklulas de Leydig de ratas de 30, 40, 50, 60, 70, 80 y 90 días de nacidas; así como tambien determinar los factores que activan a la enzima aromatasa y a la expresibn de su mensajero.
OBJETIVOS PARTICULARES.
1.
2.
3.
4.
5.
Obtener las fracciones enriquecidas de las cklulas de Leydig; en ratas macho de dispntas edades.
Demostrar y evaluar la actividad de aromatasa en fracciones enriquecidas de las células de Leydig; en ratas macho de distintas edades.
Evaluar los niveles de Estradiol en celulas de Leydig en ratas macho de distintas 6 p.
Obtener el RNAm de la aromatasa en las celulas de Leydig, en ratas macho de distintas edades.
Determinar los factores que pueden modificar la actividad de la aromatasa en celulas de Leydig de distintas edades.

Se utilizaron ratas macho de la cepa Wistar de 30, 40, 50, 60, 70, 80 y 90 días de edad, mantenidas en el Bioterio de la Divisi6n de Ciencias Biol6gicas y de la Salud de la Universidad Aut6noma Metropolitana, Unidad Iztapalapa, con periodos de luz-obscuridad de 12 hrs. c/u, y alimentaci6n y agua ad libitunr.
~~ ~~
O B T E N C I 6 N DE LAS MUESTRAS
Se diseñ6 un sistema simple para la obtenci6n de las muestras requeridas para cada objetivo: Se hizo la selecci6n de los animales por cada edad, se sacrificaron y se obtuvieron las fracciones enriquecidas & CL, con las fracciones en KRBG se hicieron las incubaciones para el tiempo cero y el tiempo de una hora, separhdolas en grupos por quintuplicado, (a) para evaluar la biotransformaci6n de precursores radiomarcados, (b) para la evaluaci6n de los contenidos end6genos de Estradiol y (c) para la evaluaci6n del RNAm de la aromatasa a las que se les adicion6 0.5 mL de Trisol. Al tkrmino de cada tiempo se conge16 la muestra en un baño crioscópico y se mantuvieron en congelaci6n hasta ser utilizadas.
Los animales fueron sacrificados por dislocaci6n cervical, se extirparon los testículos por vía abdominal. Los testículos fueron descapsulados y la sangre se elimin6 mediante varios lavados con una soluci6n Krebs-Ringer (NaCl 125 mM, KC1 5 mM, MgS04 1 mM, Tris 35 mM, NaH2P04 lmM, pH 7.4), adicionada con glucosa al 0.2% y gentamicina al 0.05%.
OBTENCI6N DE LAS FRACCIONES CELULARES.
Separación celular (Bermúdez et al, 1988).
Cada par de testículos se colocaron en 7 mL de colagenasa (lmg/mL en KRBG) durante 18 minutos a 37°C con agitaci6n peribdica, al tkrmino de este período se suspendi6 la actividad enzimática por diluci6n con 15 mL de soluci6n salina isot6nica (NaCl al 0.9%), mezclando y dejando reposar durante 10 minutos a temperatura ambiente.
La suspensi6n celular se filtr6 a travks de una malla estkril de Nylon; el filtrado se utili26 para obtener la fracci6n enriquecida de cklulas de Leydig.
Purificación de la fracci6n intersticial cruda.
La fracci6n intertisial cruda fue centrifugada a 1,500 x g durante 10 minutos. Se desech6 el sobrenadante y la pastilla celular se resuspendi6 en 1 mL de KRBG para ser colocado sobre 8 mL de una soluci6n de KRBG - Ficoll-Albúmina Serica Bovina (Ficoll400 13% - albúmina 0.2%, pH 6.5) y posteriormente se sediment6 a 1,000 x g durante 15 minutos. El segundo sobrenadante se desech6 y el b o t h celular se resuspendi6 en KRBG, ajustando la concentraci6n a 105 CL por mL.
Biotransformaci6n de esteroides radiomarcados.
Las CL se incubaron en presencia de T marcada [I, 2, 6, 7, 16, 17 -3H, con A.E. de 109 Ci/mol] durante O 6 1 hora a 34"C, pasado este tiempo los medios y las celulas se recuperaron para determinar el efecto de la incubacibn sobre la conversi6n de testosterona (T) a estradiol (Ez), y la

posible conversi611 de androstendiona (A) a estrona (El). Las muestras fueron mantenidas en congelacibn a -20°C hasta ser utilizadas.
Para analizar la biotransformaci6n se reaIiz6 una purificacibn cromatogrAfica de cada esteroide la cual consistid en: descongelar los tubos que tenían las muestras; estas se transfirieron a tubos de centrífuga con tap6n (de 12 mL aproximadamente); con el fin de realizar un enjuague, a los tubos que contenían las muestras se les adicion6 0.5 mL de agua destilada tratando de colectar los residuos de la muestra.
A los tubos de centrífuga se les adicion6 5 mL de eter etilico, se agitaron vigorosamente un minuto y se dejaron reposar algunos minutos con el fin de que se diera la separaci6n de las fases org6nica y acuosa, posteriormente la última se conge16 en un baño criosc6pico (hielo seco y acetona), esto se realiz6 para obtener la fracci6n etérea donde se encontraban los esteroides, la cual fue decantada a otro tubo de centrífuga sin tap6n. La operaci6n se realiz6 dos veces.
Se dej6 evaporar el éter a sequedad en una campana de extracci6n; posteriormente sé procedi6 a concentrar a los esteroides, para lo cual se agregaron por las paredes del tubo 10 gotas de éter etilico, repitiendo la operaci6n por tres ocasiones, dejdndose evaporar el @ter en cada vez
El paso siguiente fue colocar estas muestras en las zonas definidas para ello en las cromatoplacas de silicagel 60 F w para cromatografía en placa fina, esto se realiz6 agregando nuevamente éter etilico a cada tubo, pero en este caso fueron solo 5 gotas pero repitiendo la operaci6n tres veces. Así mismo en las zonas correspondientes se colocaron eskindares auténticos de los esteroides a evaluar, T, A, Ez y El, una vez adicionados los esteroides, las cromatoplacas fueron sometidas a tres tratamientos de desarrollo: 1) Benceno; 2) Benceno-acetato de etilo (8:2) y 3) Benceno - Metano1 (95-5); posteriormente y ya secas completamente, fueron irradiadas con UV (254 nm) con el fin de que se evidenciaran los esteroides de la serie A43cetona en la fluorescencia verde . .
de la cromatoplaca. Para que se evidenciaran los estr6genos se adicion6 reactivo de Oertel, marcando las zonas coloridas que aparecieron después de calentar la zona con aire caliente, y corroborando su presencia por la fluorescencia a 266 nm; posteriormente las placas se dejaron secar y se marcaron las zona a colectar tomando como criterio las partes mas alta y mas baja de las manchas del estdndar correspondiente, se realiz6 el raspado en las placas para obtener los esteroides que se deseaba cuantificar.
El raspado fue colectado por aspiraci6n con pipetas Pasteur, las cu6les contenían una capa de fibra de vidrio la cual sirvi6 como filtro al polvo de la silica gel, ya que a las pipetas se les adicion6 1 mL de éter etílico para disolver al esteroide; el filtrado que se obtuvo se colect6 en viales. Una vez evaporado el eter, a los viales se les adicionaron 5 mL de líquido de centelleo (Fluor) para que posteriormente fueran evaluados en el contador de centelleo Packard Tri Carb 2900 TR
EVALUACION DE LOS CONTENIDOS END6GENOS DE ESTRADIOL
R A D I O I N M U N O A N A L I S I S .
Preparaci6n de la curva patr6n.
Se prepararon soluciones etan6licas de concentraciones conocidas en diluciones progresivas de un orden de magnitud. La curva patr6n se prepar6 tomado volúmenes fijos de Ias soluciones mencionadas como se describe en la tabla 1.
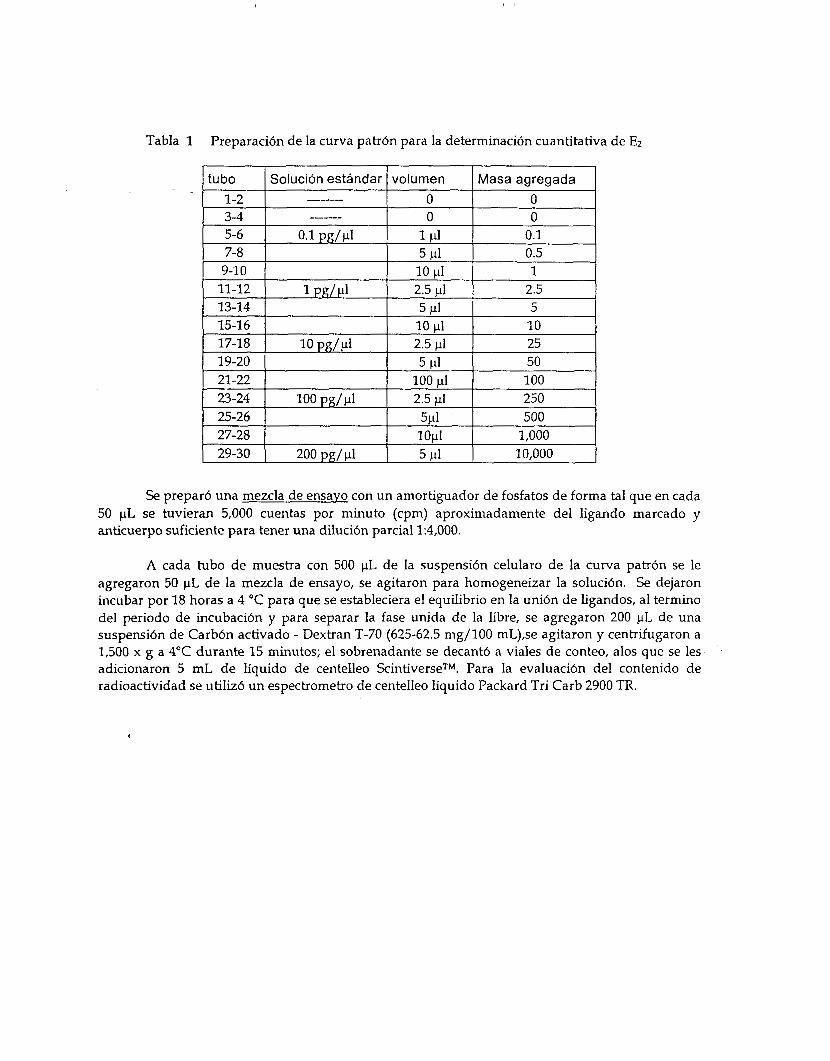
Tabla 1 Preparaci6n de la curva patr6n para la determinaci6n cuantitativa de E2
tubo Masa agregada volumen Solución estándar 1-2
O O """ 3 4 O O
I 19-20 I 50 I I 21-22 I I 100 u1 I 100 I c ~ r -
i
23-24
1,000 1 op1 27-28 500 5P1 25-26 250 2.5 pl 100 pg/p1
1 29-30 1 200 pg/pl 10,000 5 Pl
Se prepar6 una mezcla de ensayo con un amortiguador de fosfatos de forma tal que en cada 50 pL se tuvieran 5,000 cuentas por minuto (cpm) aproximadamente del ligando marcado y anticuerpo suficiente para tener una diluci6n parcial 1:4,000.
A cada tubo de muestra con 500 pL de la suspensi6n celularo de la curva patrón se le agregaron 50 pL de la mezcla de ensayo, se agitaron para homogeneizar la soluci6n. Se dejaron incubar por 18 horas a 4 "C para que se estableciera el equilibrio en la uni6n de ligandos, al termino del periodo de incubaci6n y para separar la fase unida de la libre, se agregaron 200 pL de una suspensi6n de Carb6n activado - Dextran T-70 (625-62.5 mg/100 mL),se agitaron y centrifugaron a 1,500 x g a 4°C durante 15 minutos; el sobrenadante se decant6 a viales de conteo, alos que se les adicionaron 5 mL de líquido de centelleo ScintiverseTM. Para la evaluaci6n del contenido de radioactividad se utiliz6 un espectrometro de centelleo líquido Packard Tri Carb 2900 TR.

RESULTADOS
Obtención de la fraccibn enriquecida de Células de Leydig.
Despues del tratamiento mecdnico-enzimdtico; la fracci6n intersticial proveniente de ratas macho de 30, 40, 50, 60, 70, 80 y 90 días de edad, mostraron porcentajes de pureza y viabilidad semejantes a los antes obtenidos por el grupo de trabajo (Bermúdez y col, 1988).
La pureza fie superior al 60 % y los contaminantes de la fracci6n obtenida fueron eritrocitos con un 35 % y el resto, celulas de origen tubular, posiblemente espermatogonias. Es importante hacer notar que el contaminante indeseable son las celdas de Sertoli, las cuales no se hicieron presentes a pesar de las edades tempranas.
La viabilidad medida utilizando el metodo de excIusi6n de azul Tripan, present6 porcentajes mayores al 85 % tras una hora de incubaci6n.
Efecto de la edad en la transformación de T* a Ez'..
Cromatografía en capa fina.
En cuanto a los pardmetros de calidad de la cromatografía y datos relativos al experimento se obtuvieron los siguientes:
Los frentes relativos promedio de los esteroides heron, en orden de polaridad, 0.82 para El, 0.74 para A, 0.55 para E1 y 0.45 para T, la variaci6n de ellos fue menor al 2 %.
Del total de esteroide marcado agregado en la incubaci6n 13,047 cpm se recuperaron de las cromatoplacas al final de todo el experimento, 11,916 cpm lo que representa el 91.33 %, considerAndolo adecuado.
En los tubos incubados durante O horas, pero sometidos al proceso cromatográfico, , se encontr6 un contenido constante de F con un promedio de 11,952 k 35 cpm con una n de 80, se consider6 que no hubo transformacibn de esa T*.
En los tubos incubados con KRBG, sin celulas, se obtuvo una supuesta transformaci6n de T* a E2* del 0.002 % que definitivamente f ie no significativa con un valor de p = 0.00001022.
La biotransformacibn hacia androstendiona y de esta hacia estrona, determinada por los contenidos de radiactividad fue menor al 0.01 %, por lo que se consider6 no sigruficativa (p = 0.00003781) y por lo mismo innecesaria su expresi6n grdfica.
En cuanto al experimento de biotransformaci6n de la T*, en la grdfica 1 se puede apreciar en primer lugar que la T* si es transformada por las CL en el periodo de una hora, puesto que los contenidos del precursor radiomarcado van disminuyendo en las preparaciones celulares de las diversas edades, así mismo la cantidad de E2 se incrementa tras incubar las cClulas por 1 hora, por lo que la transformaci6n se debe a la actividad de la aromatasa presente en las preparaciones celulares.

También se puede observar que la actividad de la aromatasa se hace presente desde los 30 días de edad; también se puede observar que en la preparaci6n de los 60 días de edad, la cantidad de E2 presente (2,653.6 cpm) cs nlucho menor comparada con la cantidad de T* (8571.6 cpm); a los 70 días se observa que la canlidad de Ez ( 5 , 8 5 1 ~ ~ ~ ~ ) presente no es significativamente mayor que la de T* (5,243 cpm); pero en las CL. de ratas de 80 y 90 días, la cantidad de EZ formado (6,933 y 9,187 C P ~ I respectivamente) es significativamente mayor con respecto a la cantidad de T* presente (4,047 y 2,088 cpm respectivamente).
12000 -
10000 -
8000 --
6000 -
4000 -
2000 -
0 -
TRANSFORMACIóN DE T* ~
T
T
30 40 50 60 70 80 90
EDAD (dias) . . ~ "" ~. ~~ ~ ~ ~ - ~ .
Gráfica 1 Muestra los resultados crudos de la biotransformaci6n de T* a E:*. Los resultados expresan el promedio y la Desviaci6n Estándar de los quintuplicados.
En la misma gráfica 1 se observa que los cambios mAs importantes en la actividad de la aromatasa ocurren entre los 50 y los 70 días de edad de los animales.
Se hace notar que la dispersi6n en los resultados de la biotransformaci6n es adecuada a pesar de que en esta se implican posibilidades de variaci6n relacionadas con los procesos de incubacibn, viabilidad de las preparaciones, el proceso de obtenci6n de las CL, así como tambien la variabilidad biol6gica de los animales, cabe la pena hacer notar que por las condiciones en el manejo del Bioterio no siempre se puede definir con exactitud la fecha de nacimiento de los animales, esta se define con f un dfa, por lo que para cada edad fueron seleccionados los machos de una camada en particular.

Evaluación de los contenidos end6genos de Estradiol.
Con la seguridad de que había actividad de aromatasa en las preparaciones celulares, se procedi6 a determinar el contenido de Estradiol en las mismas y al mismo tiempo, con la incubaci6n tratar de definir si los contenidos del estr6geno eran ahí formados o bien dependían de su síntesis en otro compartimiento y su migraci6n hasta las CL.
Las determinaciones del E2 se hicieron por un Radioinmunoanálisis 'altamente' específico con un antisuero obtenido de acuerdo al concepto de Bermúdez y cols. 1975. La diluci6n final del mismo de 1 : 40,000 asegura alta sensibilidad, raz6n por la que la curva patr6n se inici6 con 0.1 pg de masa desplazante de un trazador con seis sitios de sustituci6n de H por 3H, es decir de alta Actividad Específica, (109 Ci/mmol). El resultado se expres6 en la gráfica 2,
Se observa en la gráfica que el desplazamiento de las dosis de 0.1 y 0.5 pg son significativas respecto del cero, lo cual tiene mucha importancia en relaci6n con la sensibilidad del ensayo. Así mismo se observa que hubo un desplazamiento proporcional que, en terminos de la gráfica se manifiesta con un Coeficiente de Regresi6n de 0.973. Por lo que las determinaciones de las muestras son válidas mientras se encuentren en los límites de detecci6n de la curva patr6n de 0.1 a 1,000 pg.
RIA estradiol
-1 -0.5 O 0.5 1 1.5 2 2.5 3
Log dosis - - ~- ~.~~ __ ____ - "-1
Gráfica 2. Curva de dosis respuesta para el RIA de Estradiol, trazador agregado 5,000 cpm de Ez - JH , diluci6n final del anticuerpo 1:40,000, límites de detecci6n 0.1 a 1,000 pg.
Contenido y producci6n de EZ
En este caso el tiempo cero s610 define el contenido inicial de E2 presente en las CL, las cifras encontradas muestran contenidos que van de 2.82 a 9.47 pg por tubo, cifras claramente significativas con relaci6n a la sensibilidad del RIA, aunque la diferencia entre los contenidos iniciales del estr6geno entre leas edades de 30 y 90 días es estadfsticamente significativa, no va de

acuerdo con la capacidad de transformaci6n mostrada en los resultados de la incubaci6n de una hora de las mismas celdas.
6 1 HORA
""""""
EDAD (dias) . ~ " ~ ~ ~. ~~ ." "" . ~. ~ "" ~ .
Gráfica 3. Muestra la cantidad de Ez presente en el tiempo O y 1 hr como funci6n de la edad de los animales. Los resultados Promedio y Desviación Estándar se expresan en pg/ tubo.
En la gráfica 3 se puede observar que después de 1 hora de incubaci6n la cantidad de E? formado como resultado de la actividad de la aromatasa inicia desde la preparaci6n celular de ratas de 30 días, pero donde se puede apreciar un incremento considerable y significativo (p< 0.00001) es en las ratas de 60 días, donde la cantidad de E? sobrepasa los 100 pg, después de esta edad la cantidad de Ez formado por la incubaci6n continua incrementándose pero no en una forma tan significativa como en las ratas de 60 días, también es claro que la producción del estrógeno presenta gran variabilidad como lo demuestran las amplias barras de dispersi6n.
Es por demás notorio en la gráfica 3 que los contenidos del estrógeno para las preparaciones sin incubaci6n y las correspondientes a las edades más tempranas no pueden ser debidamente apreciadas por razones de la escala, por lo que se definen mejor al expresar, en la gráfica 4 las mismas cifras en escala logaritmica.
Efecto de la edad en la cantidad inicial y produccih de E2.
En este caso se puede observar que en las CL de ratas de 30 días de edad la producci6n de E2 en una hora no es significativa al compararse con el tiempo cero; mientras que para las provenientes de ratas de 40 días, la cantidad de E2 presente en las celulas que se incubaron 1 hora comienza incrementase significativamente con respecto a las de tiempo cero; en las Clobtenidas de ratas de 50 dias la diferencia en la cantidad de E2 presente en las celulas incubadas 1 hora es un poco m6s significativa con respecto a las de tiempo cero. Pero donde realmente se observa una diferencia altamente significativa entre el tiempo cero y 1 hora es a la edad de 60 dfas ya que en esta edad el incremento que se observa en el tiempo 1 hora es de unas 14 veces. En las CL de ratas de edades mayores la diferencia en la cantidad de E2 presente, sigue siendo bastante significativa entre el tiempo cero y 1 hora; pero si s610 se comparan las cantidades obtenidas tras de 1 hora de

incubacih, se podría decir que el incremento entre una edad y la otra no es muy grande ( esto es en las edades de 70,80 y 90 días); confirmado al comparar las edades de 80 y 90 días donde se puede ver que el incremento es mucho menor, l o que podría llevarnos ha considerar que la producci6n de E? se comienza hacer estable a partir de esta edad, o bien que la actividad de aromatasa en las CL comienza a decrecer, o que simplemente la capacidad de transformaci61-1 ha llegado a su máximo: (GrAfica 4).
." .~
1000 -
- 100 - n CD
Y
-I o s I-
v) w 10 -
1 '
CONTENIDO Y PRODUCC16N
~"
T
.""
T
""
" _
"
30 40 50 60 70 80
EDAD (dias)
" . .
Gráfica 4 Muestra en escala logaritmica el contenido de EZ inicial y la cantidad que se produce después de 1 hr. de incubaci6n en cada grupo experimental.
Cambio en la capacidad de síntesis de E2 entre 10s periodos de edad.
Como se ha venido diciendo, al parecer la capacidad de síntesis de Estradiol está presente en las preparaciones celulares de Leydig desde los 30 días de edad, pero también se ha venido observando en los resultados que dicha capacidad no es constante, por lo que se calcularon las diferencias entre cada dos edades, tanto de los contenidos iniciales como de las cantidades de EZ formado por la incubaci6n y se expresaron en la grdfica 5.

l o 0 0 -
149 49 I
o m z 1 97
30-40 40-9 53-60 60 - 70 70 - 80 80 - 90 I PERIODOS DE CAMBIO I
. 1
Gráfica 5. hluestra la cantidad total de E2 producida en cada grupo experimental entre los periodos de edad.
Se observa que los cambios entre los diferentes periodos en los contenidos endógenos de EZ no son significativos a excepción del periodo de 70 - 80 días, pero los resultados de la incubación son muy diferentes; en este caso se puede observar que el periodo en el cual se encuentra un cambio mayor en la capacidad de formación de EZ es en el período de 50 -60 días (149.49 pg), en donde la cantidad de E2 formado es mucho mayor; posteriormente se observ6 que este valor o esta diferencia en la actividad de aromatasa comienza a declinar conforme a la edad, ya que la diferencia en E2 formado es m6s pequeña; no obstante no se puede decir que alcance a los mismos niveles que en etapas tempranas de edad.

DISCUSI6N
En cuanto al aspecto general del planteamiento experimental, se defini6 claramente en la evaluaci6n de la presencia y actividad de la aromatasa; definir primero si dicha actividad estaba presente por medio de la biotransformación de un precursor radiomarcado, confirmando tal presencia por medio de la evaluaci6n de los contenidos end6genos del esteroide producto, el Estradiol y finalmente tener la ‘garantia’ para proseguir con las tecnicas de Biología Molecular en aquellos grupos experimentales donde claramente se hubiera demostrado la ausencia o la poca presencia y actividad de la enzima y en aquellos donde se hubiera demostrado de forma clara la actividad de la misma. Por lo pronto se reportan las dos primeras etapas, ya que los procedimientos y resultados de la última e s t h en proceso.
Hay que hacer notar que la obtenci6n de los animales experimentales es un serio problema ya que como se comentó en el capítulo de resultados, el manejo del Bioterio tiene sus pequeños problemas, la definición de la edad real de los animales, es uno de ellos y crucial para este tipo de experimento; otro problema es la alimentación, ya que una vez destetados, son alimentados con el preparado para roedores comercial, pero en el transcurso del presente experimento les fue cambiado por dos ocasiones de marca, lo que trajo cambios en el peso de los animales en los correspondientes periodos de adaptacih, y si reconocemos que el estado nutricional de los organismos es parte importante de la omeostasis requerida para que se presenten los cambios implicados en la madurez sexual, resulta importante reconocer que los resultados fueron satisfactorios a pesar de dichos cambios, puesto que se tom6 la decisi6n de no utilizar aquellos animales que presentaran gran variación en el peso. LOS datos correspondientes no se agregaron al reporte, pero si fueron tomados en consideración.
Con respecto a los observado en las graficas de biotransformaci6n se podría decir que desde los 30 días de edad se presenta alguna actividad de aromatasa, lo cual se comprueba con la cantidad de E2 end6geno presente y se puede decir que esta actividad se va incrementando conforme a la edad. En este caso el incremento en la cantidad de E2 presente puede deberse a que al ir creciendo las ratas, se presentan cambios tanto morfológicos como fisiológicos, lo cual nos indicaría que esta entrando en la pubertad en donde la producción de T se incrementa haciendo que la actividad de la aromatasa sea mAs acelerada; siendo por esto que en las ratas de 60 días es más notorio el incremento en la producci6n de E?.
En este experimento no se tomaron en cuenta los otros dos esteroides analizados (Androstendiona y Estrona) por que las cantidades en que aparecían eran muy pequeñas, demostrando que la esteroidogenesis se encuentra funcional, y que todo el procesamiento para la obtencibn de las c6lulas y su incubacibn durante una hora en KRBG, no alter6 sustancialmente ese estado, ya que de haber aparecido androstendiona marcada se podría pensar que se hubiera alterado el patr6n biosintetico por desequilibrio en la oxido-reducci6n del sistema enzimAtico de la 17 p hidoxiesteroide deshidrogenasa.
Los resultados obtenidos en este trabajo mostraron que la actividad de la aromatasa estA presente desde los 30 días, concordando con otros estudios que demostraron que la producci6n de E: podía ser detectada en el tejido intersticial de ratas prepúberes (Tsai-Morris et al, 1985) y que se incrementa conforme la edad (Valladares & Payne, 1981; Rommerts et al, 1982) y es clara en las adultas (VaIladares & Payne, 1979).
S610 que nuestros resultados experimentales no implican ningún tipo de estimulaci6n, son s610 resultados de una incubacidn en un medio ‘simple‘ de mantenimiento, sin la adici6n de diversos factores utilizados por diversos autores, como por ejemplo hormona Luteinizante (LH) o

gonadotropina cori6nica humana (hCG), 3’,5’-adenosinmonofosfato cíclico (AMPc), insulina (Ins) o factor de crecimiento epidermic0 (EGF) (Payne et al, 1987; Rigaudiere et al, 1989; Syed et al, 1991).
Como se describi6 en resultados, es notorio que la cantidad de EZ endbgeno presente (tiempo cero) cambia muy poco entre las ratas de 30 y las de 90 días, en tanto que la capacidad de transformaci6n en una hora de ‘simple’ incubaci6n demuestra la posibilidad de formar una mayor cantidad; la explicacibn de este hecho podría estar en la posibilidad de que el estr6geno formado por las CL se hubiera salido del compartimiento celular distribuyhdose por el espacio intercelular y perdiendose en el procesamiento de la separaci6n celular, mientras que durante la incubaci611, el estr6geno s610 se acumula, dentro o fuera de las celulas pero en el mismo tubo de incubación, no obstante las cantidades formadas fueron al menos 10 veces superiores a las antes encontradas por otro participante del grupo de investigaci6n (Experimentos del Doctorado del M. en B.E. Joaquín Herrera) y es necesaria al menos una repetici6n del diseño experimental para corroborar los resultados.
En cuanto al gran cambio que ocurre en la capacidad de síntesis de EZ a la edad de 50 y 60 dias, es importante señalar que posiblemente los animales estdn entrando a la madurez sexual, es decir son animales en pubertad y que de acuerdo a la literatura, se incrementa la síntesis de Testosterona y con ello, ademds de haber m8s substrato, se confirma el hecho de la posible replaci6n de la aromatasa por el andr6geno propuesto recientemente (Genissel y cols., 2001).

BIBLIOGRAFÍA
BERMÚDEZ J.A., CORONADO V., MIJARES A., LEON C., VELÁZQUEZ A. & MATEOS J.L., 1975. Steroidochemical approach to increase the specificity of steroid antibodies. J Steroid Biochem; 6:283-2901
BERMÚDEZ J.A., MENDIETA E & HERRERA J. 1988. Evaluaci6n de los métodos de aislamiento y Purificaci6n de celulas de Leydig y Sertoli. Archivo de Investigación Médica 2:291.
BLOOM J. Y FAWCETT D: 1995. Tratado de Histología. McGraw- Hill, México D.F.
BUSTOS ROSAS J. 1991. Efecto de los tratamientos térmicos y enzimdticos sobre la actividad moduladora de la esteroidogénesis presente en el medio condicionado de las células de Sertoli. Tesis para obtener éI titulo de Biólogo, ENEP- Zaragoza, UNAM, 33 pp.
BUSTOS J., HERRERA J & MENDIETA E., 1996. Efecto de la actividad moduladora de la esteroidogénesis (AME) sobre los sistemas de transducci6n membranal de células de Leydig (CL). Encuen-tro para el fortalecimiento de la Docencia e Investigación Agropecuaria en Biología de la Reproducción, UAM- Iztapalapa, México, D.F.
CONNELL C. Y CONNELL G. 1977 The interstitial tissue of the test in: Johnson A.D. y Gomes W.R. (Ends) the testis, Volt IV Academic Press N. Y. Pp 33-369.
DORRINGTON J.H. & ARMSTRONG D.T. 1975. Follicle-stimulating hormone stimulate estradiol 17-p synthesis in cultured Sertoli cell. Proc Nat Acad. Sci USA 2: 6551.
DORRINGTON J.H., FRITZ I.B. & ARhlSTRONG D.T. 1978. Control of testicular estrogen synthesis. Biol. Reproduction 18: 55.
EWING R. AND KEENEY P. 1993. Evidence that the FSH receptor itselt is not a calcium chanell: Endocrinology 131 2:979-981.
FAUSET.1995 Fisiología medica
FISHMAN J & GOT0 J. 1981. Mechanism of estrogen biosynthesis: Participation of multiple enzyme sites in placental aromatase hydroxylation. J. Biol. Chem. 256: 4466.
GANON 1998. Fisiologia medica.
GELBER S.J., HARDY M. P., MENDISHANDAGAMA S.M.L. 1987. Direct stimulatory effects of insulin-like growth factor-I on androgen production by highly purified puberty and adult rat Sertoli cell. J Andrology 184: 478.
GENISSEL C., LAVALLET J. & CARREAU S. 2001. Regulation of citochrome p450 aromatae gene expression in adult rat Leydig cell: comparison with estradiol production. J. Endocrinology 168:95- 105.
GONDOS B. 1980. Development and differentiation of the testis and male reproductive tract. in: steinberger a: testicular development, structure, and function. Raven Press, N.Y:, pp 3-20.

HARDY M., ZIRKIN B Y EWING 1989. kinetic studies on the development of the adult population of Leydig cell in testis of the pubertal. Endocrinology 124: 762-770.
HELDING C.H. & WESTERMARK B. 1989. Growth factors as transforming proteins. Eur J Biochem - 184: 478.
HERRERA J, RODRfGUEZ E, GONZÁLEZ E, BERMÚDEZ J.A. & MENDIETA E., 1991. Efecto del estradiol sobre la síntesis de testosterona en células de Leydig en cultivos. XXXI Reunión Anual de la Sociedad Mexicana de Nutrición y Endocrinología, AC, Oaxaca.
~ ~~
HERRERA J, NAVARRO M, MENDIETA E & BERMÚDEZ J.A., 1995. Los neurotransmisores como factores de modulaci6n de la esteroidoghesis en las células de Leydig. XXXV Reunión Anual de la Sociedad Mexicana de Nutrición y Endocrinología, AC, Guanajuato, Gto.
HERRERA J., GARCÍA-CASTRO J, RODRÍGUEZ-MALDONADO E, MENDIETA E & BERMÚDEZ J.A., 1996. Sertoli cell conditioned media modulate the androgen biosynthetic pathways i n rat Leydig cell primary cultures. Arch Androls:127.
JEGOU B., 1992. The Sertoli cell. Bailliere Clin Endocrine Metab 6273.
LIN T., HASKELL J., WINSON N. & TERRACIO L. 1987. Direct stimulatory effects of on Leydig cell steriodogenesis in primary culture. Biochem Biophys Res. Commune 1 3 7 : 950.
hlAGRES AND JOST A. 1980. The initial phases of testicular organogenesis in the rat. Arch Anat Microsc. Morpho1 Exp., 69:297-318.
MERCANTE M.L., FLORID1 A., NATAL1 P.G., CITRO G., CIOLI V. & DE MARINO C. 1979. Sertoli cell of adult rats ilz vitro. L1. Effect of different steroid precursors on oestradiol 17-p synthesis. Experiential 3 5 : 563.
hlARYIN W; MAYERS P., RODWELL V. AND GRANNER K. 1990 Bioquimica de Harper, 10a Ed., Editorial El Manual Moderno, México D. F., pp 543-547.
hENDIETA E, SALAME A, HERRERA J & ANTON-TAY F., 1995. Melatonina mhibition of androgen biosynthetic pathway in Leydig cell (LC)-enriched cell fraction from normal adult rats. hlolecular Andrology 2:5
hdILLER WL., 1988. Molecular biology of steroid hormone synthesis. Endocrine Rev2:295.
NEAVES M. JOHONSON L. PORTER J., PARKER C. Y PETTY C. 19%. Leydig cell number, daily sper production and serum gonadotropin in ageing men. J. Clin. Endocrinol, Metab 59:756-762.
PAPADOPOULOS V., KATAHOVING P: DROSDOWSKI M. Y HOCHEREAU DE REVERS M. 1987. Adult rat Sertoli cells secrete a factor or factors which modulate Leydig cell function. J. Endocrinol. 114: 459-468.
PAYNE AH, PERKINS LM, GEORGIOU M & QUINN PG., 1987. Intratesticular site of aromatase activity and possible function of testicular estradiol. Steroids 50:435.
PERDOMO ESQUIVEL M, ACEVEDO VARGAS J, HERRERA MUÑOZ J & BERMÚDEZ JA., 1996. Evaluaci6n de la vfa adenilatociclasa dependiente de las celulas de Leydig y Sertoli. X X I Reunibn Anual de la Academia de Investigacibn en Biología de la Reproduccibn, AC; Mazatlán, Sin.

PROMERANTZ DK., 1980. Developmental changes in the ability of follicle stimulating hormone to simulate estrogen synthesis in vivo by the testis of the rat. Biology Reproduction 23.948.
PRINCES F 1984. Ultrastucture of in mature Leydig cell in the human prepubertal testis. Anat rec 209: 165-1 76.
RIGAUDIERE N, GRIZARD G & BOUCHER D., 1989. Aromatase activity in purified Leydig cell from adult rat. Comparative effects of insulin, IGF-I and hCG. Acta Endocrinology 1 2 1 : 677.
RISBRIDGER G.P. , 1992. Local regulation of Leydig cell function by inhibitors of steroidogenic activity. Cell Biol. Int. Rep s:399.
ROMMERTS F. F.G., TEERDS K, THEMMEN A.P.N., & VAN NOORT M., 1992. Multiplied regulation of testicular steriodogenesis. J. Steroid Biochem g:398.
RUIZ PANIAGUA LSC. 1994. Purificaci6n parcial de un factor inhibidor de la síntesis de testosterona, a partir de medios condicionados por células de Sertoli. Tesis para obtener éI grado de maestro en Biología Experimental, Universidad Autónoma Metropolitana, Unidad Iztapalapa, 13 pp.
SAEZ J. 1993. Leydig cell: endocrine, paracrine, and autocrine regulation. Endocrine Reviews. Vol. 15: 574 -626.
SKINNER ME;. 1991. Cell-cell interactions in the testis. Endocrine Rev 12:45.
SYED V, KHAN SA & NIESCHLAG E. 1991. Epidermal growth factor stimulate testosterone production of human Leydig cell in vitro. J. Endocrinol Invest 14:93.
TSAI-MORRIS CH, AQUILANO DR & DUFAU ML. 1979. Cellular localization of rat testicular aromatase activity during development. Endocrinology E : 4 3 1 .
LE & PAYNE AH. 1979. Induction of testicular aromatization by luteinizing hormone in mature rat. Endocrinology =:431.
VALLADARES LE & PAYNE AH. 1981. Effects of hCG and cyclic AMP on aromatization in purified Leydig cell of immature and mature rats. Biol. Repro. 2:725.
VERHOEVEN G. 1992 Local control systems within the testis. Bailliere Clin Endocrinol Metab 6:313.
WELSH T.H. Jr. & HSUEH A.J.W. 1982. Mechanism of the inhibitory action of epidermal growth factor on testicular androgen biosynthesis i n vivo. Endocrinology 4 7 : 99.
WILSON, J:D. AND FOSTER, D.W. 1992. Williams textbook of endocrinology 8* Edition, W.B. Saunders.
ZIRKIN B. Y EWING L. 2987. Leydig cell differentiation during maturation of the rat testis: a stereological study of cell number and ultrastructure. Anat. Sc Amer. 253: 125 -135.
ZWAIN I.H., MORRIS T& CHENG H. 1991. Identification of an inhibitory factor from a Sertoli clonal cell line (tm4) that modulate adult rat Leydig cell. Biochem and Biophys Acta; 1334:338-348.

DIAGRAMA DE LA METODOLOGfA
OBTENCI6N DE LAS FRACCIONES CELULARES
Ratas macho cepa Wistar de diferentes edades (30,40,50,
60 y 90 días)
Se sacrificaron a las ratas por dislocaci6n cervical
testiculos
de la vena central
Se lavaron con soluci6n
amortiguadora de KREBS.
+ Se colocaron en 7 mL de KRBG- colagenasa 18
min. a 37OC L
c 15 mL de soluci6n salina por 10 min.

Se fragmentaron (con bisturí a fragmentos de 1 -
5 mm).
Se realiz6 una digesti6n enzimdtica (20 mL de KRBG - pancreatina, 20
min.) , 20°C, con agitaci6n.
Se realiz6 una sedimentaci6n (lg, 10
min. 4°C).
Se decant6 el sobrenadante
Se sediment6 a través de un gradiente de sacarosa del 2 a 6 % (lg, 30 min. ,
4"C\
Se recuperaron las fracciones de 4 y 5 %
Se filtr6 (malla de 50 - 100 pm).
Se sediment6 (1500g, lOmin., 5°C).
I I
Se sedimentaron (1500 g, 10 min. a 5' C
Sé resuspendieron (ImL de KRBG) .
Se sedimentaron a través de Ficoll - BSA (1000 g, 15 min.
, 5°C).
Célula de Leydig.
de CClulas de Sertoli
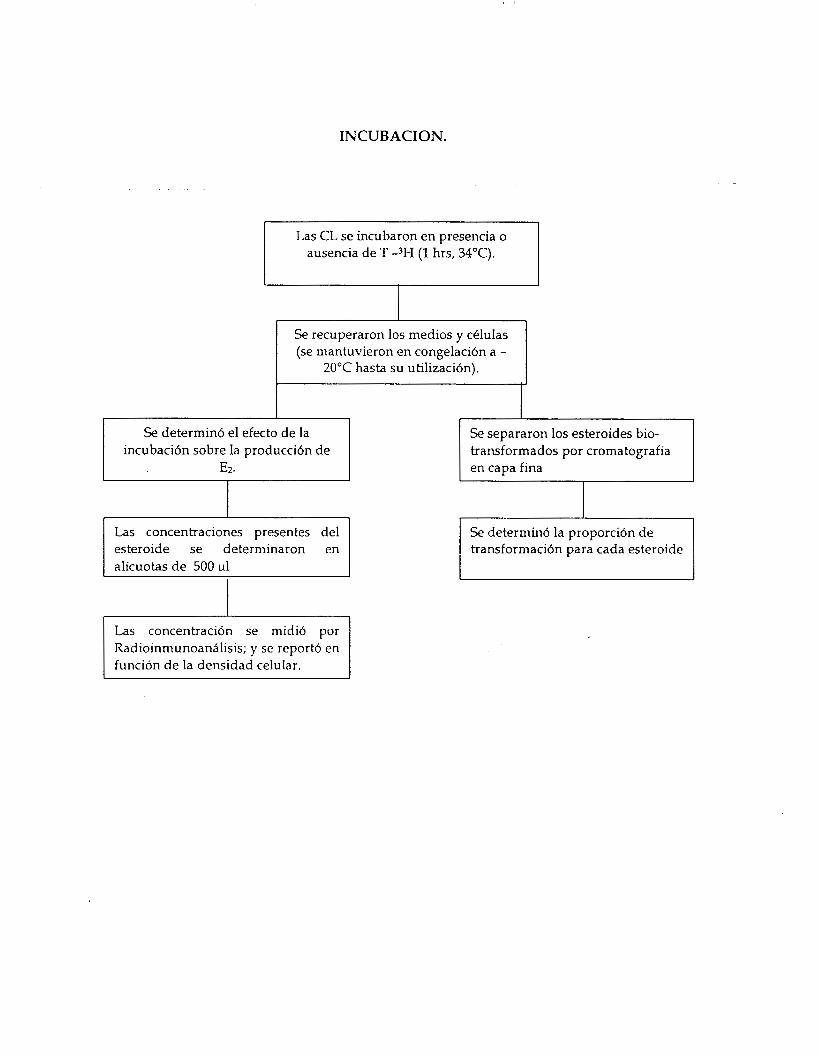
INCUBACION.
Las CL se incubaron en presencia o ausencia de T -3H (1 hrs, 34°C).
Se recuperaron los medios y c6lulas (se mantuvieron en congelaci6n a -
20°C hasta su utilizaci6n).
I Se determin6 el efecto de la
incubacibn sobre la producci6n de E2.
Las concentraciones presentes del
alícuotas de 500 111
Las concentraci6n se midi6 por Radioinmunoantilisis; y se report6 en función de la densidad celular.
I
Se separaron los esteroides bio- transformados por cromatografía en capa fina
Se determin6 la proporción de transformaci6n para cada esteroide





























