Embed Size (px)
Citation preview

Universidad Autónoma de Baja California Sur
Área de Conocimiento en Ciencias del Mar
Departamento Académico de Biología Marina
Tesis
MORFOMETRÍA CRANEAL Y MANDIBULAR EN JUVENILES DE TORTUGA
AMARILLA (Caretta caretta) DEL PACÍFICO NORTE EN BAHÍA MAGDALENA.
BAJA CALIFORNIA SUR. MÉXICO.
QUE COMO REQUISITO PARA OBTENER EL TÍTULO DE
Biólogo Marino
PRESENTA:
Juan Alfredo López Ramírez
Directora:
Dra. Rocío Andrea Franco Moreno
La Paz Baja California Sur, Noviembre de 2015

I
Dedicatoria
A mi madre Juana Ramírez Huidobro, por su incondicional apoyo y eterno cariño, así
como sus consejos, pero sobre todo por ser siempre esa roca que me ha sostenido
cuando he estado a punto de caer. A mi padre Lino López Millán, por mi maravillosa
infancia, su apoyo invaluable para alcanzar mis metas desde siempre, por traerme
aquí y por todo tu amor. No existen palabras para describir todo lo que significan
para mí, mucho menos para agradecerles por todo. Los AMO.
A Israel y Omar, porque nunca han dejado de estar aquí, por sus extraños consejos,
las incontables peleas y juegos. Gracias por las charlas nocturnas, recordarme la
dirección correcta y por ser mis hermanos. A Vianey por escuchar y reír con nosotros
sin importar sus ideas raras.
A mi mamá Adelina, por todo el amor que me diste y que sigo llevando conmigo. A mi
papá Rufo, por la brisca, el conquián, el domino, etc. Gracias por siempre tener una
palabra de aliento para mí, por todo el tiempo que compartimos y por ser mi valedor.
A Emmanuel, por recordarme esos pequeños grandes gustos, por las risas y las
bromas. A Dante, por la oportunidad de mejorar.
A todos ustedes, gracias por su cariño y confianza.
“El trabajo más difícil es nunca darse por vencido”
-Vincent Thomas Lombardi-

II
Agradecimientos
Finalmente se concreta este proyecto, el cual no habría podido realizar sin el apoyo
de muchas personas que me ayudaron a lo largo de las distintas fases de esta tesis.
A mi directora Rocío Andrea Franco Moreno, gracias por el tiempo dedicado a este
proyecto, por sus consejos y por las lecciones que me dio en este tiempo. Gracias
también al Doctor Víctor Cruz Escalona por permitirme participar en los proyectos del
Instituto Politécnico Nacional IPN-SIP 20140234 e IPN-SIP 20150352, “Morfología
Ecológica y Biometría Animal Basadas en Visión por Computadora, con Fines de
Evaluación y Comparación del Desempeño de Especies Acuáticas”.
A mi comité revisor, al doctor Enrique Alejandro Gómez Gallardo Unzueta, por su
gran apoyo durante el trabajo, su paciencia, comprensión, tiempo y consejo así como
las enseñanzas impartidas en quinto semestre de la carrera. Al doctor Gerardo
González Barba, gracias de igual manera por su tiempo y apoyo, también agradezco
sus clases durante la carrera pero sobre todo las prácticas de campo. A la doctora
María Mónica Lara Uc, por sus asesorías, consejos y orientación que ayudaron a
encauzar el proyecto, por sus palabras y su comprensión. También quiero agradecer
a la doctora Liliana Hernández Olalde por su invaluable asesoría y apoyo.
Al grupo Tortuguero de las Californias, por permitirme participar con ellos y por su
apoyo en la colecta de muestras. Al M.C. Juan Manuel Rodríguez Barón, al PhD.
Hoyt Peckham por la oportunidad de participar con en su equipo de trabajo.

III
A la gente de López Mateos, a Esmeralda “Meme”, “picafresa”, Juan “molenco”,
Oliver, Juan “guapo”, Edwin, Doña Graciela, Doña Guille gracias porque por usted
calme durante meses la perra como se dice vulgarmente. Muchas gracias también a
Jessie, Bárbara, Antonio Diego por ser siempre tan amable con migo y siempre hacer
todo de una manera positiva, Athena por los smores, las lecciones de inglés y los
chistes. También a Bladimir de la Tova, quien me trato como un amigo desde que lo
conocí y quien me enseño lo hermoso que es San Lázaro y sus parajes, Don Diego
quien me dejo descansar en su casa en más de una ocasión, al “Guerepo”, Allan, el
“Japo” y sobre todo a Don Víctor de la Tova, por ser un verdadero maestro de campo
y enseñarme más de lo que me imaginaba.
Gracias a la profesora Isabel Palacios, por el gran empujón que me dio para alcanzar
mis metas.
A la palomilla, Ivan, Juan Carlos “JC7”, Peyen, Ember, Tomas “Torreón”, Cesar
“Coshon”, Tavo, Patrick, Alland, “todo el Soccer Mesta”, Leonardo, Héctor, Noé,
Copado y Chynthia. Gracias por apoyarme hasta terminar.
Aquí me permitiré robar las siguientes palabras. A mi familia paceña Carlos, Beni y
Margarita. Por los consejos, regaños, tapiocas, abrazos y lágrimas, que me han
ayudado durante este recorrido, no tengo más para decirles que gracias.
Gracias a la vida y a dios.
“I´d like to be under the sea, in an octopus´s garden…with you”
The Beatles-Octopus´s Garden

IV
ÍNDICE
RESUMEN……………………………………………...…………………………………..1
1. INTRODUCCIÓN………………………………………………………….…………….2
2. ANTECEDENTES……………………………………………………………………….5
3. JUSTIFICACIÓN………………………………………………………………………...9
4. HIPOTESIS……………………………………………………………………………..10
5. OBJETIVOS………………………………………………………………………….....10
5.1 OBJETIVO GENERAL…………………………………………………………...…..10
5.2 OBJETIVOS PARTICULARES………………………………………………..…….10
6. MATERIAL Y METODOS……………………………………………………………...11
6.1 ÁREA DE ESTUDIO………………………………………………………………….11
6.2 CAMPO………………………………………………………………………………..13
6.3 LABORATORIO………………………………………………………………………14
6.4 GABINETE…………………………………………………………………………....20
7. RESULTADOS………………………………………………………………………....26
7.1 ANÁLISIS DE DATOS………………………………………………………………..26
7.2 ANÁLISIS MORFOMÉTRICOS……………………………………………………..28
7.3 ALIMENTACIÓN……………………………………………………………………...38
7.4 CORRELACION ALIMENTACIÓN-TALLA………...……………………………....42
8. DISCUSIÓN………………………………………………………………………….....46
9. CONCLUSIÓNES………………………………………………………………………55
10. RECOMENDACIONES……………………………………………………….……...56
11. BIBLIOGAFÍA………………………………………………………………………….56
12. ANEXO I……………………………………………………………………………….68

V
13.ANEXOII………………………………………………………………………………69
14.ANEXOIII……………………………………………………………………………...80

1
RESUMEN
El cráneo es una estructura ósea compleja, que cumple con dos funciones
principales: uno, el alojamiento y protección del cerebro y órganos sensoriales,
siendo la vista, el olfato y el oído las más relevantes. Y dos su participación en el
proceso de alimentación. El ciclo de vida de las tortugas marinas hace que varíen su
dieta conforme crecen y migran. Dentro de este grupo, Caretta caretta parece ser la
especie más generalista; se encuentra provista de un cráneo ancho con mandíbulas
potentes y se caracteriza por consumir una amplia variedad de presas. Dentro de
estas destacan crustáceos, moluscos y peces. Entre los meses de junio a septiembre
de 2012, se realizó la colecta de cráneos de Caretta caretta provenientes de playa
San Lázaro, en la Isla Magdalena B.C.S. A partir de estos se realizó un estudio
morfométrico lineal, con el fin de identificar las estructuras más relevantes en el
proceso de alimentación. Se realizó un análisis de correlación que arrojó ocho
variables. Así mismo un análisis de componentes principales indicó que la amplitud
total (AmT), longitud del supraoccipital (LSO), la distancia entre los cóndilos
mandibulares (DCM) y la distancia desde el extremo postero-inferior del cuadrado
hasta el extremo anterior del pico (DEPICP) son las variables que permiten
diferenciar entre individuos. Por otra parte la amplitud total (AmT) y la distancia entre
los cóndilos mandibulares (DCM), podrían relacionarse con la longitud de la
mandíbula inferior (LMI) y con la longitud del escamoso (LES). Se puede concluir que
las correlaciones encontradas entre estas estructuras craneales podrían ser
responsables de la robustez del cráneo de Caretta caretta, sin embargo estas no se
pudieron relacionar con el consumo de alguna presa en particular, por lo que se
recomienda el empleo de otras técnicas en trabajos futuros a fin de encontrar dicha
relación, así como el uso de otras fuentes de información respecto a las presas para
dar mayor robustez a investigaciones futuras. Este trabajo servirá como línea base
en posteriores estudios morfométricos en tortugas marinas.
.

2
1. INTRODUCCIÓN
En México se pueden encontrar siete especies de tortugas marinas;
Lepidochelys olivacea (golfina), Dermochelys coriacea (laúd), Chelonia agassizi
(prieta), Lepidochelys kempi (lora), Chelonia mydas (verde o blanca), Eretmochelys
imbricata (carey) y Caretta caretta (cabezona, caguama o amarilla). Dependiendo de
su distribución, estas especies pueden ser localizadas tanto en la vertiente del
Pacífico como en el Golfo de México y el Caribe (Sánchez, 2011).
Caretta caretta, puede ser localizada en el Golfo de México así como en el
litoral del Pacífico mexicano, específicamente en las costas de Nayarit, Sinaloa, en la
península de Baja California y dentro del Golfo de California. Como las demás
especies, realiza grandes migraciones desde el momento en que los huevos
eclosionan (Johnson et al., 1993 y Eckert y Abreu, 2001). Las migraciones inicia en
las zonas de anidación y forman parte fundamental para el conocimiento del ciclo de
vida de estos organismos; una vez las crías salen del cascarón se alejan de la orilla y
se dispersan con ayuda de las corrientes.
Ya en su estado juvenil pueden diferenciarse dos fases, una fase oceánica y
una pelágica. Hay pocos detalles acerca de la fase oceánica, sin embargo parece
claro que estos se dispersan a través de las cuencas oceánicas; especialmente en
aquellas zonas de afloramiento y convergencia de producción biológica. En esta
etapa la dieta incluye organismos gelatinosos, larvas de invertebrados, algas e
insectos (Eckert y Abreu, 2001; Silman et al., 2002).
Después de diez años aproximadamente comienza la fase pelágica. Al llegar
a la costa los ahora juveniles establecen un rango de residencia (home ranges), en el
que permanecen entre 8 y 20 años hasta alcanzar su estado adulto. Durante esta
etapa su alimentación es similar a la de los adultos, por lo que es posible observar
individuos de ambas clases de edad alimentándose en las mismas zonas;
usualmente consumen caballitos de mar, moluscos, pepinos de mar e incluso

3
corales. Para Caretta caretta esta fase comienza cuando el caparazón alcanza una
longitud entre los 25 y 30 cm. (Eckert y Abreu, 2001).
Algunas tortugas de esta especie que nacen en las costas de Estados Unidos
usan las acumulaciones oceánicas de Sargassum como refugios y zonas de
alimentación, hasta que se transportan a la parte oriental del Océano Atlántico.
Cuando las variaciones en la temperatura son muy fuertes, los juveniles llegan a
desplazarse de sus zonas de residencia. Después se desplazan hacia el sur con los
giros de las corrientes del Atlántico Norte, hasta llegar a orillas de las Islas Canarias;
posteriormente regresan como adultos juveniles con la corriente del Norte Ecuatorial
hasta el Atlántico Occidental (Eckert y Abreu, 2001).
En el caso particular de México los estudios han demostrado que esta especie
puede recorrer grandes distancias en periodos relativamente cortos (Eckert y Abreu,
2001). En la costa del Pacífico, arriban tras realizar una extensa migración desde sus
playas de anidación localizadas en Japón; pasan toda su etapa juvenil en costas
mexicanas hasta regresar a sus costas natales para reproducirse (Bowen et al., 1995
y Seminoff et al., 2004). Estas migraciones se encuentran estrechamente
relacionadas con su alimentación; por ejemplo en las costas de Baja California, se ha
documentado la presencia de juveniles que se congregan para alimentarse del
cangrejo rojo Pleuroncodes planipes (Bowen et al., 1995).
Un aspecto importante es que la alimentación de las tortugas marinas varía
respecto a su especie y su edad. Siendo crías presentan un estilo de vida pelágico y
se alimentan de organismos que forman parte del plancton; sin embargo a partir de
su estado juvenil cambian su alimentación. Chelonia mydas se alimenta
exclusivamente de algas, mientras que Dermochelys coriacea lo hace
primordialmente de medusas. Caretta caretta es una especie generalista, siendo
principalmente carnívora; se alimenta de moluscos bentónicos, crustáceos,
gasterópodos, corales blandos, pepinos de mar, camarones e incluso algas
(Sargassum), entre otras presas. Hay registros que indican que en algunas zonas se
alimentan de organismos provenientes del descarte de la pesca; un ejemplo de esto

4
se da a orillas de la Isla Cumerland en Georgia, donde las tortugas que mueren
ahogadas en las redes de arrastre, aparentemente se alimentan de peces y algunos
crustáceos que caen atrapados en estas mismas (Badillo, 2007 y Bjorndal, 1985).
En México una de las zonas de alimentación más importantes para Caretta
caretta se encuentra en las playas del Pacífico, específicamente en el complejo
lagunar Bahía Magdalena en el Golfo de Ulloa. Debido a las condiciones
oceanográficas que inducen una alta productividad y biodiversidad en esta zona, se
genera una alta concentración de la langostilla roja Pleuroncodes planipes, que
constituye una fuente principal de alimento para esta especie en la región (CNAP,
2011).
Dentro de las adaptaciones que poseen las tortugas marinas encontramos
capacidad de bucear a grandes profundidades, la presencia de glándulas que
permiten excretar el exceso de sal y un cuerpo hidrodinámico. También poseen un
tronco largo dentro de una estructura dorsal convexa y rígida denominada
caparazón, complementado por una sección ventral casi plana llamada plastrón. La
cabeza ha perdido su capacidad para retraerse y las extremidades se encuentran
modificadas en forma de remo, lo cual permite un desplazamiento adecuado dentro
del medio acuático (Gerosa y Ureggi., 2005 y Chacón et al., 2007).
La anatomía de un animal nos puede dar información acerca de su historia de
vida, sus interacciones con el ambiente y su alimentación. En el caso de Caretta
caretta sus hábitos generalistas podrían verse reflejados en la anatomía de su
cráneo. A diferencia de las otras especies, su cabeza es relativamente grande y
ancha hacia la parte posterior y el pico se estrecha anterior a las cavidades orbitales.
Las mandíbulas son robustas y tienen forma de V. Así mismo, tienen un paladar
secundario relativamente largo sin crestas alveolares y los dos huesos maxilares
entran en contacto posterior a los huesos premaxilares (Lutz et al., 2003 y Wyneken,
2004). La cabeza de gran tamaño y la mandíbula potente, hacen posible que esta
especie se alimente de presas fuertemente blindadas como bivalvos y crustáceos
(Spotila, 2004).

5
El estudio de las formas de las estructuras nos ayuda a comprender las
causas de las variaciones anatómicas. El desarrollo ontogénico, las adaptaciones al
medio y los procesos evolutivos, generan diferencias estructurales entre individuos y
especies. Estas diferencias, pueden indicarnos la variedad de funciones que pueden
tener las estructuras como respuesta a una misma presión selectiva, o durante los
procesos de crecimiento y morfogénesis (Zelditch et al., 2004).
Conceptualmente, la ecomorfología se basa en la relación existente entre el
organismo y su ambiente. Es decir, permite analizar los mecanismos por los cuales
las variaciones morfológicas resultan en diferencias funcionales y de desempeño y
da una aproximación para entender la manera en que las especies se reparten los
recursos (Wainwright, 1994 y Bohórquez, 2009). Un ejemplo de esto es la relación
funcional entre la morfología de las estructuras asociadas con la alimentación y las
características del recurso utilizado, como el tamaño de las presas (Granado, 1996;
Franco, 2011). En esencia se puede describir como el entendimiento de la forma y la
función de una estructura (Kardong, 2006).
Durante años se ha reconocido la relación general entre la forma de los
organismos y su ecología. Hutchinson (1959) sugirió que las medidas lineales de las
estructuras tróficas, podrían utilizarse como indicador del grado de traslape o
solapamiento ecológico y esto reforzaría la teoría que diferentes morfologías podrían
adaptarse mejor a diferentes tipos de presas (Wainwright, 1996). Teniendo en cuenta
esto, en el presente trabajo se pretenden determinar cuáles son las estructuras
craneales de Caretta caretta que influyen durante el proceso de alimentación.
2. ANTECEDENTES
La morfometría constituye una forma cuantitativa de abordar las
comparaciones de las formas (Zelditch et al., 2004) y en el caso de las tortugas
marinas, ha sido empleada en distintos estudios. Pritchard y Mortimer (1999)
publicaron “Taxonomía, Morfología Externa e Identificación de las Especies”, donde
mencionan estructuras morfológicas claves para la identificación de las especies de

6
tortugas marinas en campo. Esto para las etapas de cría y adultos. Para ello hacen
uso de características como el número de escamas en la cabeza, de escudos en el
caparazón y de escudos del plastrón.
La morfometría por otro lado también ha sido utilizada para diferenciar entre
sexos. Michel (1995) en su tesis de maestría caracteriza la variabilidad fenotípica de
crías de la tortuga golfina Lepidochelys olivacea, utilizando 30 variables
morfométricas a partir de las cuales busca características particulares para cada
sexo. Mediante el empleo de métodos multivariados (análisis discriminante), sugiere
la posibilidad de inferir con un grado de certeza aceptable (95%) el sexo de las crías,
a partir de nueve variables morfométricas.
Medina et al. (2011) con base en sus análisis morfométricos, determinan la
relación existente entre los escudos dorsales y el largo recto del caparazón (LRC),
para 44 ejemplares de Eretmochelys imbricata (tortuga carey) capturados en Cuba.
Para esto midieron el largo y ancho de los cuatro escudos izquierdos y los cinco
vertebrales. Los autores concluyen que las medidas de los escudos pueden ser
usadas para definir clases de tallas, siempre y cuando los escudos no estén
modificados y usando siempre una ecuación de regresión adecuada. Así mismo
demostraron que las correlaciones entre el crecimiento de los escudos y las tallas de
la tortuga carey, descritas anteriormente para la gran barrera Australiana y Puerto
Rico, también pueden ser aplicadas a las tortugas carey de Cuba y el Caribe.
Otros estudios han usado la morfometría para diferenciar entre poblaciones.
García (2009) realiza su trabajo con morfometría de poblaciones alopátricas de
Chelonia, manejando a C. m. agassizii como subespecie de C. agassizii. En este
estudio se analizaron morfométricamente dos poblaciones reproductoras de C.
agassizii y de C. mydas para determinar posibles diferencias en algunas variables
morfométricas (longitud, amplitud y forma del caparazón y altura del cuerpo), que
sugirieran un posible aislamiento reproductivo mecánico entre ambas poblaciones.
Los datos morfométricos fueron analizados mediante análisis de componentes
principales para determinar la diferenciación entre ambas colonias reproductivas. A

7
pesar de ser encontradas dichas diferencias, se recomienda profundizar al respecto
ya que no se concluye de manera determinante si son suficientes para que se de
dicho aislamiento.
El primer antecedente de morfometría craneal en tortugas marinas es el
trabajo de Nishizawa et al. (2010). En este realizan una comparación morfológica del
cráneo de juveniles y adultos de tortuga verde (Chelonia mydas) y sus implicaciones
en la alimentación. Se encontró que el tamaño de la órbita y la longitud del
supraoccipital en relación con el centro del cráneo, fueron mayores en juveniles que
en adultos. Sin embargo estas diferencias relativas no predijeron una funcionalidad
equivalente en las estructuras tróficas.
En el caso particular de Caretta caretta, se han realizado algunos estudios
morfométricos. Báez et al. (2010) realizan un análisis biométrico de la especie en el
Mediterráneo Occidental, con el fin de encontrar si existen diferencias significativas al
relacionar la longitud recta (LR) y longitud mínima curva del caparazón (LmCC), para
dos poblaciones que confluyen en esta zona. Desarrollaron fórmulas de conversión a
partir de medidas obtenidas de 670 individuos, de un componente multipoblacional.
Se demostró la capacidad predictiva de dichas fórmulas contrastándolas con diez
ejemplares medidos in situ, encontrando que no existen diferencias significativas
entre los valores obtenidos y las medidas estimadas por la fórmula de conversión.
Ferreira et al. (2010) evaluaron los patrones morfométricos de embriones de
Caretta caretta. Mediante marcadores anatómicos sobre el caparazón y el plastrón,
pudieron discriminar la velocidad del desarrollo de las crías. Observaron una
diferencia significativa en la forma del caparazón de las crías, siendo más redondo el
de aquellas que se incubaron por menos tiempo (55 días); en comparación con
aquellas que fueron incubadas por un periodo más largo (67 días). Los resultados de
este estudio indicaron que el tiempo de incubación, explica solo una parte de la
variación total en la forma del caparazón.
Uno de los análisis morfométricos comparativos es el de Jones et al. (2012),
quienes publican un estudio sobre las funciones y variaciones morfológicas del cuello

8
y el cráneo de Caretta caretta y Lepidochelys kempi. Para una mejor observación del
acomodo de los músculos y de la bóveda craneana, hicieron uso de la anatomía
comparada clásica y tomografías. Ellos encontraron que en ambas especies la región
temporal está rodeada de hueso y se encuentra asociada a la mandíbula, haciendo
posible su movimiento; así mismo el aparato hioideo no es conspicuo como en las
tortugas de agua dulce y existen varias similitudes entre los músculos de ambas
especies. Se concluye que la habilidad de succionar a la hora de tragar no se
encuentra bien desarrollada como en las tortugas de agua dulce, sin embargo ambas
poseen un esqueleto capaz de dar poderosas mordidas. Esto podría deberse al tipo
de presas de las cuales se alimentan ambas especies.
Durante las migraciones, las tortugas marinas pueden tener acceso a
diferentes recursos alimentarios. Bowen et al. (1995), demuestran la presencia de
juveniles de C. caretta en las zonas de concentración de Pleuroncodes planipes
(langostilla roja), en las costas de Baja California a partir del empleo de marcadores
mitocondriales. Concluyen que el 95% de las tortugas observadas en el norte de las
costas del Pacífico mexicano provienen de Japón y recorren más de 10,000 km
desde sus zonas de anidación para alimentarse.
Los crustáceos son de las presas más comunes para Caretta caretta. Ocaña
et al. (2006), destacan la importancia del cangrejo Polybius henslowii como principal
recurso en la dieta de la especie en la costa del Estrecho de Gibraltar, África. Los
datos para este estudio fueron obtenidos de 150 tortugas varadas y de un número no
especificado de tortugas capturadas en altamar, de las que se obtuvo el contenido
estomacal (por inducción al vómito). Los autores mencionan que la mayor
concentración de cangrejos, coincide con la abundancia de tortugas, lo que se
produce durante la primavera y el verano. En este periodo P. henslowii representa
entre el 99-100% del contenido estomacal de Caretta caretta. A partir de los
resultados se concluye que esta especie es especialista, ya que al ser cosmopolita
podría alimentarse de diferentes recursos disponibles; sin embargo dados los
resultados obtenidos, estas parecen estar seleccionando su alimento.

9
Báez et al. (2012) realizan “Nuevas aportaciones a la ecología trófica de
Caretta caretta”. El objetivo fue ampliar la información sobre la ecología alimentaria
de esta especie, comparando los contenidos estomacales de dos ejemplares
juveniles capturados: uno perteneciente al Atlántico Norte y otro del Mediterráneo
Occidental. Para el individuo del Atlántico Norte se encontraron moluscos
gasterópodos, anfípodos, cefalópodos y medusas, mientras que para el del
Mediterráneo Occidental se encontraron restos de peces (probablemente de
especies que se utilizan como carnada en los palangres).
En ambos individuos se encontraron objetos antropogénicos como señuelos,
materiales plásticos, cuerdas, trozos de redes, etc. Los autores concluyen que si bien
el número de muestras en este trabajo es meramente anecdótico, los resultados
coinciden con lo ya descrito en la bibliografía. Futuros trabajos podrían mostrar con
mayor claridad las diferencias entre ambas poblaciones de C. caretta; así mismo
mencionan que la presencia de elementos de pesca puede deberse al carácter
oportunista de la especie.
3. JUSTIFICACIÓN
El cráneo es la estructura ósea más compleja de los vertebrados. Este se
encuentra formado por varios huesos que en conjunto, funcionan de una manera
particular dentro de cada grupo animal y acorde a las necesidades de cada especie.
Esta estructura sirve cómo protección para los órganos sensoriales y en conjunto con
la mandíbula trabajan en el proceso de alimentación.
Las tortugas marinas constituyen uno de los grupos de reptiles más antiguos,
por lo que poseen características que podrían definirse cómo primitivas. Poseen un
cráneo anápsido y carecen de dientes, sin embargo esto último no les impide tener
una mordida potente, la cual utilizan para poder capturar sus presas y probablemente
como defensa ante algunos de sus depredadores.
Aunque se tiene conocimiento bibliográfico de que las presas de Caretta
caretta varían respecto a su ubicación geográfica y edad, aún se desconoce si existe

10
una relación entre algunas estructuras craneales y mandibulares específicas y el
consumo de dichas presas. Por esta razón, se considera que un análisis
morfométrico lineal (tradicional) podría permitir la identificación dichas estructuras y
la discriminación de su relevancia. Este constituye un primer acercamiento a la
relación entre la forma y la función de las estructuras cráneo-mandibulares de
Caretta caretta.
4. HIPOTESIS
Dentro de las tortugas marinas Caretta caretta presenta el cráneo más ancho
y robusto, lo cual podría ser atribuido en parte a sus hábitos alimentarios. Si la
morfología del cráneo y la mandíbula restringen el espectro de presas que un
organismo puede consumir, entonces se considera que al realizar un análisis
morfométrico básico, se podría vislumbrar una relación entre las estructuras
craneales y mandibulares medidas, con el tamaño de las presas consumidas por la
especie.
5. OBJETIVOS
5.1 Objetivo general
Identificar las principales características morfométricas del cráneo de los
individuos Caretta caretta distribuidos en Baja California Sur y su posible
relación con los hábitos alimentarios de la especie.
5.2 Objetivos particulares
Realizar una búsqueda bibliográfica exhaustiva acerca de los hábitos
alimentarios de Caretta caretta.
Recopilar la información existente acerca de las tallas de las presas de Caretta
caretta.
Identificar y discriminar las principales estructuras anatómicas del cráneo y la
mandíbula de Caretta caretta, empleando morfometría lineal.

11
Determinar si existe una relación entre las principales estructuras anatómicas
del cráneo y la mandíbula de C. caretta, con sus hábitos alimentarios.
6. MATERIAL Y METODOS
6.1 Área de Estudio
Bahía Magdalena se encuentra localizada entre los 24°15’N, 111°30’W y
25°20’N, 112°15’W y conforma el sistema lagunar más extenso de Baja California
Sur, México con una extensión de 649.7 km2 (Funes y González., 2002; Fleischer y
Urruchúa, 1986).
Dicho sistema puede ser dividido en áreas plenamente diferenciadas:
Área noroeste: se comunica con el Océano Pacífico a través de una entrada
de un kilómetro de ancho denominada Boca de Soledad, que da paso a un canal que
se prolonga siete kilómetros y termina al sureste en el Puerto Adolfo López Mateos.
Hacia la parte sur el canal posee una profundidad promedio de 11 m y se separa del
mar sólo por un estrecho cordón litoral, formado por dunas de arena fina. Termina 30
km al sur en lo que se conoce como la Curva del Diablo. Área norte: donde Bahía
Magdalena se conecta con el Pacífico por una boca de aproximadamente 38 m de
profundidad y 5.5 km de ancho, cuyos extremos son Punta de Entrada y Punta
Redonda. La parte final del complejo se denomina Bahía Almejas, y se comunica al
océano por una boca de 2.5 Km de ancho aproximadamente y 30 m de profundidad,
formando los extremos de la Isla Margarita e Isla Creciente (Fleischer y Urruchúa,
1986).
El complejo lagunar posee tres islas:
Isla Magdalena: con una longitud de aproximadamente 90 km y con forma de
escuadra, se sitúa al Oeste del litoral del municipio de Comondú a 24° 32’N y
25°16’N y 112°18’W y 112°03’W. Esta tiene como principal asentamiento el Puerto
Magdalena.

12
Isla Margarita: localizada a 24° 31’N y 24° 18’N y 112° 18’W y 112° 00’W, con
33.8 km de largo y 7.2 km de ancho.
Isla Creciente: también llamada Bahía Almejas, formada por una franja
angosta de tierra de 23 km de largo que se localiza a 24° 22’N y 24° 19’N y 111°
26’W y 111° 40’W. Esta isla es considerada como área de refugio para múltiples aves
acuáticas y mamíferos marinos (Funes et al., 2007).
Playa San Lázaro forma parte de Isla Magdalena y se encuentra localizada en
el Golfo de Ulloa, en la costa occidental de la Península de Baja California. En dicha
zona se han realizado diferentes estudios dirigidos a la conservación de Caretta
caretta, debido a que es un área importante de alimentación para estos quelonios;
así mismo es una región en la que la pesca ribereña es destacable, lo que trae como
consecuencia un constante impacto sobre esta especie por efectos antropogénicos
(González., 2008; Peckham et al., 2008).

13
Figura 1. Mapa de la zona de muestreo, en el que se demarcan los 45 km de playa San
Lázaro, BCS, México.
6.2 Campo
Los cráneos utilizados en este estudio fueron obtenidos de organismos
varados, encontrados durante los recorridos realizados de Junio a Septiembre de
2012, a lo largo de 45 km de costa pertenecientes a playa San Lázaro B.C.S.,
México. Las salidas de campo formaron parte del monitoreo realizado anualmente
por el Proyecto Caguama (ProCaguama) del Grupo Tortuguero de las Californias
(Oficio; SGPA/DGVS/04568/11). A los organismos muestreados se les aplicó el
protocolo de identificación, se les tomó el largo y ancho curvo del caparazón, se
realizó el registro de ectoparásitos, grado de descomposición, anomalías
morfológicas (en caso de existir), presencia de daño antropogénico (en caso de
existir), localización por GPS y registro fotográfico.

14
Las cabezas de aquellos especímenes identificados como Caretta caretta y
cuyo estado fuese óptimo para el estudio, fueron separadas con ayuda de un
cuchillo, realizando un corte desde la parte posterior del supraoccipital hasta el
cuello. Cada cabeza recolectada fue empaquetada, etiquetada con plumón indeleble
y refrigerada para su posterior transporte al Laboratorio de Docencia número 2 del
Centro Interdisciplinario de Ciencias Marinas del Instituto Politécnico Nacional
(CICIMAR-IPN). En donde se realizó la limpieza de los cráneos y se tomaron las
medidas morfométricas correspondientes. El total de muestras recolectadas en
campo fue de 34.
6.3 Laboratorio
Cada muestra se descongeló con un poco de agua. Posteriormente se realizó
la limpieza de manera manual empleando equipos de disección, cuchillos, espátulas,
pinzas y cepillos. Se retiró todo el tejido muscular, epitelial y adiposo con la mayor
precaución posible, pues la osamenta de estos quelonios es muy frágil y se rompe
con facilidad. Una vez removido el tejido en su totalidad, cada cráneo se dejó secar a
temperatura ambiente durante varios días para después ser re etiquetado e iniciar
con las respectivas mediciones.
Se empleó la guía de identificación “La Anatomía de las Tortugas Marinas”
(Wyneken, 2004), para diferenciar los cráneos de Caretta caretta que se encontraban
en resguardo en la colección del Centro Interdisciplinario de Ciencias Marinas
(CICIMAR-IPN); algunos estos cráneos fueron incluidos dentro de los análisis (n=26).
La guía también fue empleada para determinar las estructuras craneales a medir,
con base en su posible importancia durante el proceso de alimentación.
Los cráneos fueron medidos empleando dos instrumentos de medición: cinta
métrica (flexible) y vernier digital (Fowler 0-24”). Las medidas se tomaron en las
vistas dorsal, ventral, lateral y mandibular. Sin embargo debido al mal estado de
algunos cráneos, no todos se pudieron medir con ambas herramientas. Los cráneos
medidos con cinta fueron 46, mientras que los medidos con vernier fueron 60. Cada

15
medida fue registrada en una base de datos realizada en Excel para los posteriores
análisis exploratorios y estadísticos.
A continuación se muestran las medidas tomadas a los cráneos de C. caretta,
(Figuras 2, 3, 4 y 5).
Figura 2. Medidas tomadas a los cráneos en la vista dorsal. a) Medidas de 1 a 6. b) Medidas de 7 a
12. Modificado de Wyneken, 2004.
1) longitud total, 2) longitud desde el borde posterior de las narinas hasta el
extremo posterior, 3) longitud de las narinas externas, 4) longitud del rostro (extremo
más anterior del cráneo hasta el borde posterior del frontal), 5) longitud desde el
borde posterior del frontal (unión frontal/ parietal) hasta el extremo posterior, 6)
longitud de la órbita, 7) ancho del frontal, 8) longitud desde la unión
frontal/postorbital/parietal hasta el extremo posterior, 9) longitud del postorbital 10)
ancho del postorbital, 11) longitud del escamoso y 12) longitud del supraoccipital.
a) b)
16
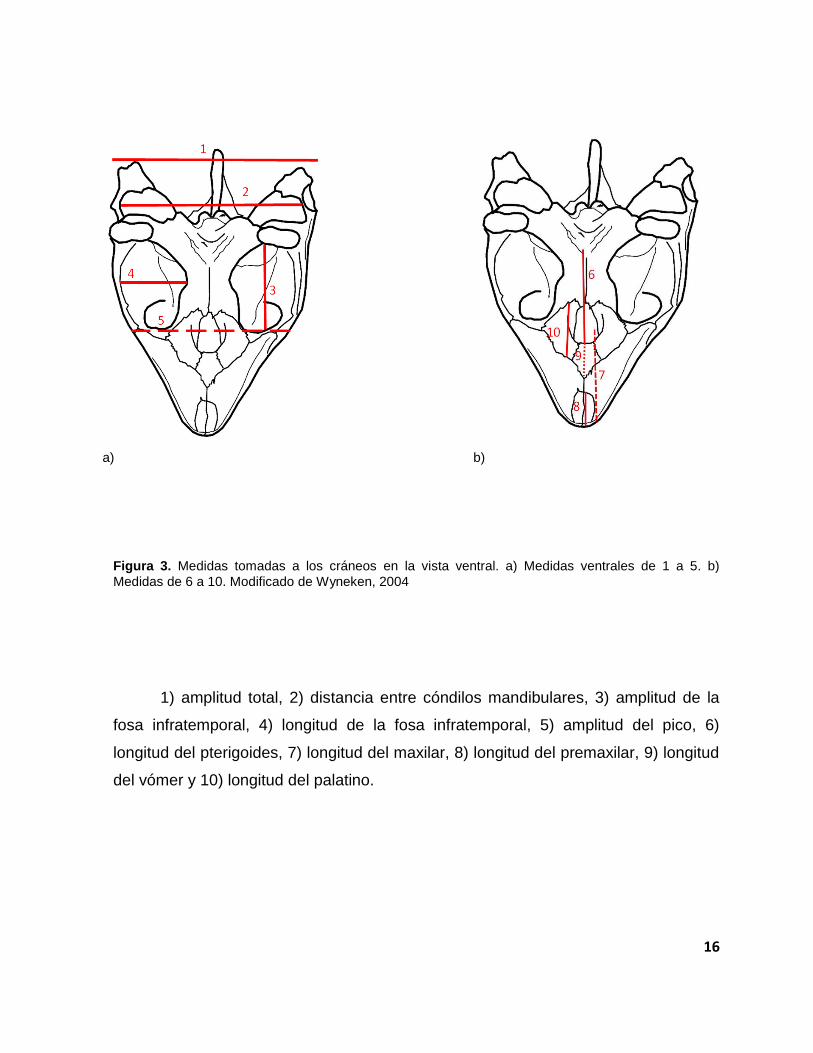
Figura 3. Medidas tomadas a los cráneos en la vista ventral. a) Medidas ventrales de 1 a 5. b)
Medidas de 6 a 10. Modificado de Wyneken, 2004
1) amplitud total, 2) distancia entre cóndilos mandibulares, 3) amplitud de la
fosa infratemporal, 4) longitud de la fosa infratemporal, 5) amplitud del pico, 6)
longitud del pterigoides, 7) longitud del maxilar, 8) longitud del premaxilar, 9) longitud
del vómer y 10) longitud del palatino.
a) b)

17
Figura 4. Medidas tomadas a los cráneos en la vista lateral. a) Medidas laterales de 1 a 4. b) Medidas
laterales de 5 a 7. Modificado de Guido y Aureggi, 2005.
1) distancia desde el extremo póstero-inferior del cuadrato (cóndilo
mandibular) hasta el extremo anterior del pico, 2) longitud del cuadrato-yugal, 3)
altura del cuadrato-yugal, 4) amplitud del cóndilo del cuadrato-yugal (extremo del
proceso del cuadrado debajo del conducto auditivo), 5) longitud de la órbita ocular, 6)
distancia desde el extremo póstero-inferior del cuadrato-yugal (cóndilo mandibular)
hasta la unión póstero parietal/ escamoso y 7) longitud total.
a) b)

18
Figura 5. Medidas tomadas a las mandíbulas. Modificado de Wyneken, 2004.
1) longitud de la mandíbula inferior, 2) longitud del articular y 3) longitud del
dentario.
Para cada variable se asignó una abreviatura. Éstas son presentadas en la
siguiente tabla.
Tabla 1. Abreviaturas aplicadas a las variables.
VARIABLE ABREVIATURA VARIABLE ABREVIATURA
1.-Longitud total. (LT) 17.-Amplitud del pico. (AmP)
2.-Longitud desde el borde
posterior de las narinas
hasta el extremo posterior.
(LNEEP) 18.-Longitud del pterigoides. (LPT)
3.-Longitud de las narinas
externas.
(LNEX) 19.-Longitud del maxilar. (LM)

19
4.-Longitud del rostro. (LR) 20.-Longitud del premaxilar. (LPM)
5.-Longitud desde el borde
posterior del frontal hasta el
extremo posterior.
(LBPFEP) 21.-Longitud de vómer. (LV)
6.-Longitud de la órbita. (LO) 22.-Longitud del palatino. (LP)
7.-Ancho del frontal. (AnF) 23.-Distancia desde el extremo
postero-inferior del cuadrado
hasta el extremo anterior del
pico.
(DEPICP)
8.-Longitud desde la unión
frontal/postorbital parietal
hasta el extremo posterior.
(LUFEP) 24.-Longitud del cuadrato-
yugal.
(LCY)
9.-Longitud del postorbital. (LPO) 25.-Altura del cuadrato-yugal. (AlCY)
10.-Ancho del postorbital. (AnPO) 26.-Amplitud del cóndilo
cuadrato-yugal.
(AmCCY)
11.-Longitud del escamoso. (LES) 27.-Longitud del orbita ocular. (LOO)
12.-Longitud del
supraoccipital.
(LSO) 28.-Distancia desde el extremo
postero-inferior del cuadrato-
yugal, hasta la unión
posteroparietal-escamoso.
(DCMPES)
13.-Amplitud total. (AmT) 29.-Longitud total. (LTL)
14.-Distancia entre cóndilos
mandibulares.
(DCM) 30.-Longitud del la mandíbula
inferior.
(LMI)
15.-Amplitud de la fosa
infratemporal.
(AmFI) 31.-Longitud del articular. (LA)
16.-Longitud de la fosa
infratemporal
(LFI) 32.-Longitud del dentario. (LD)

20
6.4 Gabinete
Análisis de Datos
Se tomaron un total de 32 medidas del cráneo y la mandíbula (12 dorsales, 10
ventrales, 7 laterales y 3 mandibulares), a los individuos que presentaban todas o al
menos la mayoría de estructuras óseas (n=42). Estas fueron tomadas empleando
dos instrumentos de medición y registradas por dos observadores.
Para analizar si se presentaban diferencias significativas entre instrumentos
de medición y observadores, se realizó una prueba t en la paquetería PAST con un
95% de confianza y tomando las medidas como dos grupos de datos diferentes (cinta
métrica vs. vernier y observador 1 vs. observador 2).
Con el fin de conocer el comportamiento de los datos, se obtuvieron algunos
valores descriptivos para cada una de las 32 variables medidas (medias, desviación
estándar, máximos y mínimos).
Estos cálculos se realizaron para cada una de las variables medidas
empleando Excel. Posteriormente para obtener el comportamiento y tipo de
distribución de los datos de manera gráfica, se realizaron histogramas de frecuencia.
Debido a que el procedimiento se realizó para cada variable, los histogramas
variaron en cuanto al tamaño de muestra (n); ya que como se mencionó
anteriormente, no todos los cráneos presentaban la totalidad de las estructuras a
medir. Sucesivamente se les realizo una prueba de normalidad para conocer la
naturaleza de los datos.
Corrección del factor talla
Debido a que las diferencias en el tamaño tienden a ser muy pronunciadas, la
información acerca de las diferencias en la forma podría quedar enmascarada
(Samuels y Valkenburgh, 2009); adicionalmente las medidas craneales están
fuertemente correlacionadas con el tamaño del cráneo (Cabral et al., 2003). Por esta
razón, corregir o eliminar el factor talla nos permite obtener información acerca de la

21
variación en estructuras craneales específicas, que quedarían enmascaradas de no
eliminar dicho efecto.
Para eliminar el efecto de la talla se empleó la fórmula utilizada por Cabral et al.,
(2003).
b
Donde:
Mc= medida estandarizada
Mx= medida de cada variable (longitud de cada variable)
TL= longitud total del cráneo
= media de las longitudes totales (media de todos los cráneos)
b= pendiente
Este modelo es efectivo puesto que puede ser aplicado a variables
dependientes e independientes (Cabral et al., 2003). Al obtener la matriz con las
medidas estandarizadas, se aplicaron los análisis de datos mencionados (media,
desviación, varianza e histogramas de frecuencia), para conocer su comportamiento
y analizar si éste se diferenciaba con respecto al de las medidas no estandarizadas.
Análisis Morfométricos
Correlación
Para identificar aquellas medidas craneales que se encontraban
autocorrelacionadas y que nos darían información redundante, se calcularon los
coeficientes de correlación por parejas de variables. Para ello se evaluaron todas las
correlaciones posibles entre las variables. La correlación puede aplicarse cuando se
tienen variables continuas o discretas de muchos valores y se desea saber si se
encuentran asociadas o no (Pedrosa y Dicovskyi, 2007). Esta es medida en una
escala que varía entre -1 (correlación inversa) y 1 (correlación directa). Un valor de 0

22
significa ausencia de correlación, lo que indica que las variables son independientes
entre sí.
Para efectos de este análisis, se utilizaron únicamente los cráneos que
presentaban la totalidad de estructuras medidas; lo que redujo el número de muestra
a 26 individuos. Se utilizó el coeficiente de correlación de Spearman, debido a que
permite determinar si hay asociación o interdependencia entre variables aleatorias
continuas, cuando no existe una distribución normal de los datos (tal como sucede
con nuestras medidas).
Análisis Multivariados
Después de identificar y descartar aquellas medidas que aportaban
información redundante, se realizó un Análisis de Componentes Principales (ACP).
Este análisis permite condensar la información dada por un conjunto de K variables
en un conjunto de W componentes, siendo cada uno de los W factores una
combinación de las K variables (Álvarez, 1995).
En esta parte del trabajo al igual que en el análisis anterior, se utilizaron
solamente los cráneos que tuvieran la totalidad de estructuras óseas presentes. Se
aplicó este análisis exploratorio debido a que no es necesario suponer una
distribución normal de los datos y nos muestra aquellas agrupaciones que pudieran
existir entre los organismos. El ACP no considera ninguna estructura a priori de los
datos y trabaja bajo el supuesto que las variables no están correlacionadas; realiza
una serie de combinaciones de las variables originales y las reordena con base a su
variabilidad (Álvarez., 1995).
Análisis de varianza multivariado (MANOVA)
Los análisis multivariados son ampliamente utilizados por dos razones
principales: en primer lugar, porque los cuestionamientos ecológicos siempre
involucran interacción entre las respuestas de más de una variable y en segundo,
porque las diferencias existentes entre los grupos pueden deberse no solamente a

23
una variable, sino ser el resultado de todo un conjunto de variables (Scheiner y
Gurevitch, 2001).
Para evaluar si las diferencias encontradas a partir del ACP eran
estadísticamente significativas, se realizó un Análisis de Varianza Multivariado No
Paramétrico de Krustal-Wallis (MANOVA), ya que la prueba de normalidad indicó que
las variables craneales evaluadas no son normales (Anexo III). El MANOVA nos
permite realizar el análisis conjunto de la relación entre dos o más variables
independientes y dos o más variables dependientes; es decir, nos permite determinar
si las variables independientes influyen de manera simultánea en el valor de
diferentes variables dependientes (Hernández et al., 1991 y Trespalacios et al.,
2005).
Alimentación
En este trabajo se pretendió indagar la relación entre las estructuras craneales
de Caretta caretta con sus hábitos alimentarios. Sin embargo, debido a las
dificultades logísticas para conseguir los contenidos estomacales de individuos
capturados en el área de estudio, se realizó una búsqueda bibliográfica exhaustiva
para caracterizar la dieta de C. caretta. Algunos autores consideran esta especie
como oportunista, debido a la variedad de presas que puede consumir, sin embargo
la mayor parte de su dieta parece estar constituida por crustáceos.
Tras la revisión bibliográfica se determinaron 23 presas principales,
pertenecientes a tres fila diferentes. Posteriormente se consultaron algunas
características físicas de las presas que se consideró pudieran influir en el evento
alimentario de la especie y relacionarse con las estructuras craneales medidas, como
talla y dureza, para esta última se usaron las tres categorías propuestas por
Vanhooydonck et al (2007). Dura, Intermedia y Suave.
La mandíbula de los depredadores debe cumplir con dos rasgos
fundamentales: 1) permitir el cierre rápido de la apertura bucal en el momento en que
la presa sea capturada y 2) producir la fuerza suficiente para triturar y desgarrar las

24
presas (Franco, 2011). En este sentido la talla y dureza de las presas son
características a considerar, pues estas pueden influir en la alimentación de Caretta
caretta. En su ingesta existe una gran variedad de taxones, dentro de los que se
encuentran moluscos y crustáceos que son presas fuertemente blindadas, así como
algunos peces de natación lenta (Baez et al., 2012 y Spotila, 2004).
La talla o tamaño de un organismo condiciona muchas de las características
del mismo (Garcia, 1999). Por ejemplo un mayor tamaño podría dar ventaja para
poder escapar de los depredadores (Kardong, 2007). Por otro lado podemos definir
la dureza como la resistencia a la penetración dada por un objeto determinado
(Valera, 2005). En los animales la dureza se encuentra dada por las características
de los tejidos (Molist et al., 2011). Por ejemplo la consistencia del exoesqueleto de
los crustáceos se da por la quitina, mientras que en los vertebrados encontramos
colágeno, fibras musculares y carbonato de calcio, este último también se encuentra
en la concha de los moluscos, los cuales poseen una concha sólida, interna o
externa dependiendo la especie (Martínez et al., 2011 y Brusca y Brusca, 2005).
Es importante considerar estas dos características, ya que gran parte de las
presas si bien no son muy grandes, si están fuertemente protegidas contra la
depredación, por lo que sería factible encontrar la relación que guardan algunas
estructuras del cráneo Caretta caretta con el consumo de dichas presas. Estas
características aunque no se analizan más a fondo si son consideradas como
factores que influyen en la elección de las presas.
Una vez recopilada esta información se realizó una matriz de presencia
ausencia (1 presencia, 0 ausencia) de presas. Esta fue construida a partir de los
intervalos de clase generados con base en la longitud total de los cráneos, a los
cuales se les asignó la presencia (1) o ausencia (0) de las presas. La asignación fue
hecha asumiendo que las tortugas tendrían cierta preferencia por las presas acorde a
su tamaño: presas grandes - depredadores grandes, presas pequeñas –
depredadores pequeños.
Correlación alimentación – talla

25
Para evaluar si existe una correlación entre las variables morfológicas y las
ecológicas (presas), se desarrolló una prueba de Mantel. Para esto fue necesario
realizar una transformación previa a los grupos de datos (matriz de presencia-
ausencia de las presas y matriz de medidas lineales), debido a que tienen diferente
naturaleza. Esta transformación se realizó mediante el índice de similitud de Bray-
Cutis (Bray y Curtis., 1957), el cual trabaja con un rango de 0 a 1, permitiendo un
traspaso fácil de disimilaridades a afinidades (Salazar et al., 2001; Ramírez, 2006).
Una vez transformados los dos grupos de datos, para evaluar la significancia
de la correlación entre la alimentación de Caretta caretta y sus rasgos craneales, se
realizó la prueba de Mantel con 1.000 aleatorizaciones. Este análisis nos permitió
contrastar un grupo de datos diferentes, con el fin de encontrar la posible relación
entre ambos; es decir una relación causa-efecto (Diez, 2008). El primer grupo de
datos estuvo constituido por las medidas lineales de las que se obtuvo mayor
información y que fueron identificadas a partir de la correlación de Spearman y el
ACP. Para el segundo grupo se utilizó la matriz de presencia-ausencia de las presas,
generada a partir de la información bibliográfica recolectada y que se describe en el
apartado de alimentación de esta misma sección.
Regresión Lineal
Se realizó la regresión lineal para determinar la relación existente entre pares
de matrices (medidas craneales vs presencia-ausencia de presas), para obtener una
representación gráfica y un valor representativo de la misma. Para ello se tomaron
los valores transformados mediante la aplicación del índice de Bray-Curtis,
mencionado anteriormente.
Todos los análisis (correlación de Spearman, ACP, prueba de Mantel,
MANOVA y Bray-Curtis), fueron realizados empleando el programa PAST
(Paleontological Statistics 3.0).

26
7. RESULTADOS
7.1 Análisis de datos
Las medidas tomadas con uno y otro instrumento y por uno y otro observador,
se manejaron como dos grupos de datos diferentes (n1= medidas tomadas con cinta
métrica y n2= medidas tomadas con vernier; n1=medidas tomadas por el observador
1 y n2=medidas tomadas por el observador 2). Teniendo en cuenta un 95% de
confianza (probabilidad de 0.05) y partiendo de la siguiente hipótesis:
H0: Xn1 = Xn2
Ha: Xn1 ≠n2
En cuanto a los instrumentos de medición y observadores, se aceptó la
hipótesis nula: no hay diferencias significativas (t=-0.011, p=0.99 y t=0.87, p=0.38),
respectivamente. Esto nos indica que los datos tomados con una u otra herramienta
o por cualquiera de los observadores, pueden ser utilizados en los análisis
posteriores.
Estadísticos descriptivos
Se realizaron estadísticos descriptivos para ambos grupos de medidas
(estandarizadas y sin estandarizar), a fin conocer su comportamiento (tipo de
distribución) y observar si éste variaba con la estandarización. No se encontraron
diferencias, lo que nos indicó que se podía trabajar con cualquier grupo de datos
para los análisis posteriores. Se optó por trabajar con los datos estandarizados y
provenientes del vernier, por lo que de aquí en adelante los resultados que se
mostrarán corresponden a este grupo de datos. Los valores descriptivos de los datos
sin estandarizar pueden ser revisados en el Anexo I.
En la Tabla 2 se muestran los valores descriptivos de nuestros datos
estandarizados.

27
Tabla 2. Valores descriptivos de los datos estandarizados. 2) longitud desde el borde posterior de las narinas hasta el extremo posterior, 3) longitud de las narinas externas, 4) longitud del rostro, 5) longitud desde el borde posterior del frontal hasta el extremo posterior, 6) longitud de la órbita, 7) ancho del frontal, 8) longitud desde la unión frontal/postorbital parietal hasta el extremo posterior, 9) longitud del postorbital, 10) ancho del postorbital, 11) longitud del escamoso, 12) longitud del supraoccipital, 13) amplitud total. 14) distancia entre cóndilos mandibulares, 15) amplitud de la fosa infratemporal, 16) longitud de la fosa infratemporal, 17) amplitud del pico, 18) longitud del pterigoides, 19) longitud del maxilar, 20) longitud del premaxilar, 21) longitud de vómer, 22) longitud del palatino, 23) distancia desde el extremo postero-inferior del cuadrado hasta el extremo anterior del pico, 24) longitud del cuadrato-yugal, 25) altura del cuadrato-yugal, 26) amplitud del cóndilo cuadrato-yugal, 27) longitud del orbita ocular, 28) distancia desde el extremo postero-inferior del cuadrato-yugal, hasta la unión posteroparietal-escamoso, 29) longitud total, 30) longitud de la mandíbula inferior, 31) longitud del articular, 32) longitud del dentario.
VARIABLE MAX MIN PROM σ VARIABLE MAX MIN PROM σ
2(LNEEP) 15.19 13.95 14.77 0.26 18(LPT) 3.88 2.26 2.90 0.38
3(LNEX) 2.94 2.06 2.49 0.21 19(LM) 7.59 5.86 6.62 0.41
4(LR) 6.11 4.84 5.60 0.28 20(LPM) 2.36 1.59 1.90 0.16
5(LBPFEP) 11.73 9.74 10.85 0.50 21(LV) 2.48 1.41 1.93 0.26
6(LO) 5.25 4.08 4.70 0.27 22(LP) 3.67 2.30 2.93 0.29
7(AnF) 6.23 4.53 5.24 0.37 23(DEPICP) 14.58 12.34 13.24 0.44
8(LUFEP) 11.56 9.39 10.52 0.51 24(LCY) 6.21 4.15 5.10 0.50
9(LPO) 7.79 5.39 6.74 0.58 25(AlCY) 5.98 3.87 4.74 0.49
10(AnPO) 2.44 1.26 1.77 0.24 26(AmCCY) 2.92 2.06 2.49 0.22
11(LES) 5.92 3.59 4.87 1.00 27(LOO) 5.43 4.23 4.78 0.31
12(LSO) 3.64 1.56 2.63 0.47 28(DCMPES) 8.76 6.42 7.64 0.57
13(AmT) 11.13 8.35 9.38 0.76 29(LTL) 16.90 16.09 16.45 0.14
14(DCM) 13.28 11.05 12.07 0.59 30(LMI) 8.17 0.45 9.02 7.10
15(AmFI) 4.44 3.03 3.66 0.38 31(LA) 5.12 0.49 6.29 4.07
16(LFI) 6.13 4.64 5.40 3.86 32(LD) 3.77 0.34 4.44 3.18
17(AmP) 9.03 6.75 7.85 0.53

28
Para entender la tendencia de las variables (grado de homogeneidad o
variabilidad) y tener una visión general de su distribución mediante representaciones
gráficas, se realizaron histogramas para cada una de las 32 medidas morfométricas
tomadas a los cráneos de los individuos. Estos pueden ser revisados en el anexo II
(Figuras 6 a 36).
El tamaño de muestra (n) varió, debido a que no todos los cráneos
presentaron todas las estructuras tomadas en cuenta para el análisis morfométrico.
Sin embargo se trabajó con la mayor cantidad de cráneos posible para tener
robustez en los resultados. Aunque la mayoría de las variables presenta varianzas
relativamente pequeñas (que nos indica un grado de dispersión bajo), no fue posible
observar una distribución normal. Esto pudo constatarse con la prueba de
normalidad, realizada a un 95% de confianza (Anexo III).
7.2 Análisis morfométricos
Correlación
En la Tabla 3 se observan los valores de las correlaciones obtenidas con su
respectiva significancia; a partir de éstos determinamos las variables a eliminar,
debido a su autocorrelación. En total se eliminaron 8 variables: longitud de las
narinas externas (LNEX), longitud desde el borde posterior del frontal hasta el
extremo posterior (LBPFEP), ancho del frontal (AnF), longitud del postorbital (LPO),
longitud de la fosa infratemporal (LFI), amplitud del pico (AmP), longitud del maxilar
(LM), distancia desde el extremo postero-inferior del cuadrado hasta el extremo
anterior del pico (DCMPES).
Los valores de las variables correlacionadas estuvieron muy cercanos a 1(la
mayoría por encima de 0.8), lo que significa que la correlación es positiva o directa.

29
Tabla 3. Correlaciones entre las variables con su respectivo valor y significancia.
VARIABLE CORRELACIONES VALOR SIGNIFICANCIA
Longitud de las narinas
externas (LNEX).
Longitud del maxilar (LM). 0.80 5.58E-03
Altura del cuadrato-yugal (AlCY). 0.81 4.96E-03
Longitud desde el borde
posterior del frontal
hasta el extremo
posterior (LBPFEP).
Longitud desde la unión frontal/postorbital
parietal hasta el extremo posterior (LUFEP).
0.88 2.68E-05
Longitud del postorbital (LPO.) 0.82 2.80E-03
Longitud del dentario (LD). 0.82 1.74E-03
Ancho del frontal (AnF). Longitud del postorbital (LPO). 0.80 6.52E-003
Distancia desde el extremo postero-inferior
del cuadrato-yugal hasta la unión
posteroparietal-escamoso (DCMPES).
0.82 2.68E-03
Longitud del postorbital
(LPO)
Amplitud de la fosa infratemporal (AmFI). 0.80 7.04E-04
Amplitud del pico (AmP). 0.84 6.00E-04
Longitud del cuadrato-yugal (LCY). 0.80 5.81E-03
Altura del cuadrato-yugal (AlCY). 0.83 1.05E-03
Distancia desde el extremo postero-inferior
del cuadrato-yugal hasta la unión
posteroparietal-escamoso (DCMPES).
0.85 2.15E-04
Longitud del dentario (LD). 0.85 1.82E-04
Longitud de la fosa
infratemporal (LFI)
Longitud del maxilar (LM). 0.81 3.74E-03
Longitud del cuadrato-yugal (LCY). 0.85 3.45E-04
Altura del cuadrato-yugal (AlCY). 0.82 1.67E-03
Longitud del articular (LA). 0.85 2.03E-05

30
Amplitud del pico (AmP) Distancia desde el extremo postero-inferior
del cuadrato-yugal hasta la unión
posteroparietal-escamoso (DCMPES).
0.94 7.27E-09
Longitud del dentario (LD) 0.83 9.23E-04
Longitud del maxilar
(LM)
Longitud del cuadrato-yugal (LCY). 0..80 8.52E-03
Altura del cuadrato-yugal (AlCY). 0.80 9.18E-03
Longitud de la mandíbula inferior (LMI) 0.80 5.58E-03
Distancia desde el
extremo postero-inferior
del cuadrato-yugal hasta
la unión posteroparietal-
escamoso (DCMPES).
Longitud del articular (LA) 0.80 7.04E-03
Longitud del dentario (LD) 0.88 2.86E-05
Análisis multivariados
Antes de realizar el ACP se asignaron categorías a los individuos con base en
la longitud total de su caparazón; además se obtuvieron los valores máximos,
mínimos, promedio y desviación estándar (Tabla 4). Cada categoría se representó
con un color para facilitar la visualización en el espacio de ordenación: morado (50 a
59 cm), rojo (60 a 69 cm), azul (70 a 79 cm) y amarillo para los que se desconocía la
longitud total del caparazón. Tras realizar el ACP (Figura 37), notamos una clara
agrupación entre los individuos, con base en dichos patrones de coloración; salvo por
un individuo de longitud de 60 a 69 cm (rojo), el cual se encuentra distante respecto a
su grupo.
También es notable la aparente dispersión de los individuos de color amarillo
(que corresponden a aquellos de los que se desconoce su longitud total). Esta
aparente dispersión en el espacio podría constituir en realidad un acercamiento a su
clasificación con base en la longitud total del caparazón. Debido a su distribución,
suponemos que la mayoría de los individuos indeterminados pudieron presentar una

31
longitud total de 60 a 69 cm; sólo se observan tres individuos ubicados a los
extremos del plano de ordenación: dos que podrían agruparse dentro aquellos que
miden entre 50 y 59 cm y uno dentro de los que miden de 70 a 79 cm. Finalmente
tras observar el área de dispersión se evidencia que la mayoría de individuos se
encuentran dentro del rango de talla que va desde los 60 a los 69 cm.
La selección de los componentes se hace con base en la proporción de
varianza que representan. Usualmente se seleccionan hasta cubrir una porción del
80, 90 o 100%; esta regla es arbitraria y debe aplicarse con cierto cuidado (Peña,
2003). El componente principal uno (CPI) representó el 62.98% de la varianza,
mientras que el segundo (CPII) el 11.64%; ambos explicaron el 74.62%. Sin embargo
al tomar en cuenta el tercer componente (CPIII), que nos explica el 7.19% de la
varianza, se alcanza un 81.81% de la varianza total de nuestros datos.
Tabla 4. Valores máximos, mínimos, promedio y desviación estándar de las longitudes totales de los
caparazones.
LONGITUD TOTAL DEL CAPARAZÓN
VALOR MÁXIMO VALOR MÍNIMO PROMEDIO DESVIACIÓN ESTANDAR
77 54 65.11 6.44

32
Figura 37. ACP de los datos estandarizados tomando en cuenta CPI y CPII. Se observa la agrupación de los individuos con base en la longitud total de los caparazones y la varianza explicada por cada componente principal.
Dado que se utilizó el tercer componente principal, era necesario ubicar las
variables que determinaron su gradiente de variación. Al observar gráficamente el
comportamiento de CPI y CPIII (Figura 38), notamos un cambio en la configuración
espacial de los individuos. Sin embargo se mantiene la agrupación basada en el
tamaño total de los caparazones; incluso aquellos organismos que se encuentran
alejados de sus agrupaciones permanecen distribuidos de la misma manera, al igual
que los organismos de los que se desconoce su longitud (amarillo).

33
Figura 38. ACP de los datos estandarizados, tomando en cuenta CPI y CPIII se observa la agrupación de los individuos con base en la longitud total de los carapachos y la varianza explicada por cada componente principal.
Cabe mencionar que al observar la distribución espacial de nuestros
individuos (figuras 37 y 38), notamos la mayoría sobrepasan los 60 cm de longitud de
acuerdo con las medidas de los caparazones, esto significa que la mayoría de
nuestros individuos son sub adultos. Para Caretta caretta los adultos presentan un
rango va desde 70cm (Peckham, 2008). Aunque en este trabajo se pretendía trabajar
sólo con juveniles, la presencia de los sub adulos no afecta el resultado del trabajo
más allá de la distribución antes mencionada. Además se debe considerar que las
muestras fueron obtenidas de organismos varados, por lo que su clase de edad fue
azarosa.
34
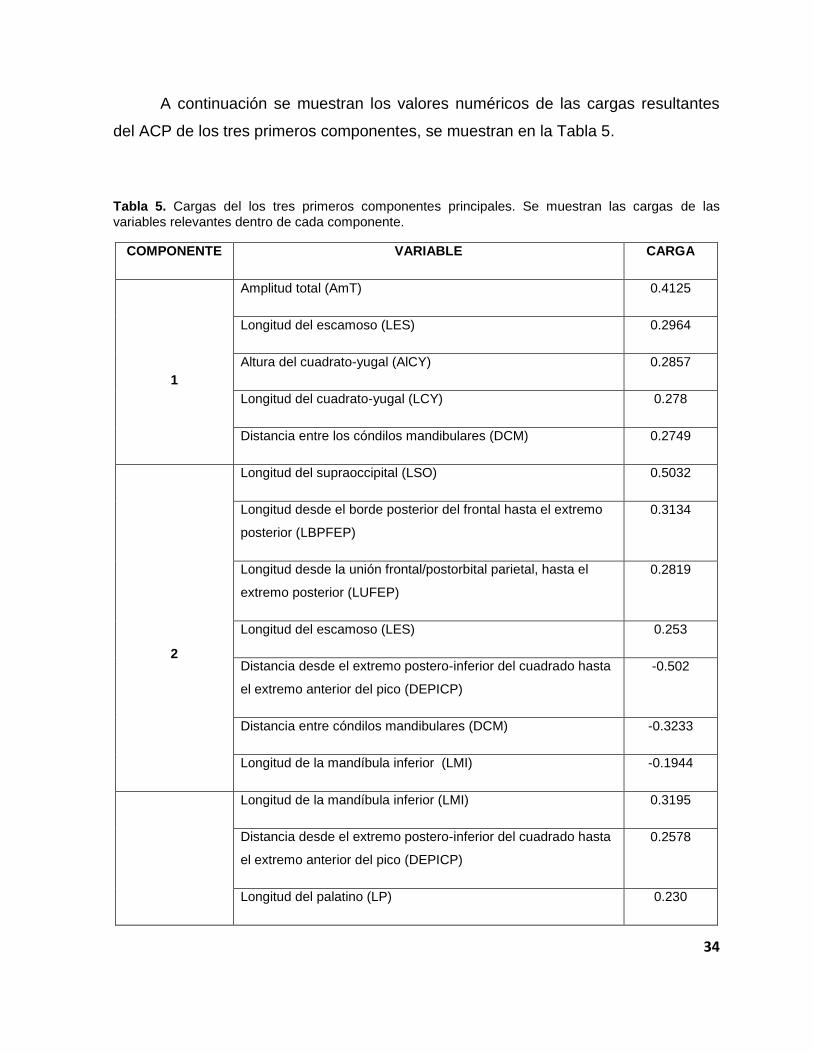
A continuación se muestran los valores numéricos de las cargas resultantes
del ACP de los tres primeros componentes, se muestran en la Tabla 5.
Tabla 5. Cargas del los tres primeros componentes principales. Se muestran las cargas de las
variables relevantes dentro de cada componente.
COMPONENTE VARIABLE CARGA
1
Amplitud total (AmT) 0.4125
Longitud del escamoso (LES) 0.2964
Altura del cuadrato-yugal (AlCY) 0.2857
Longitud del cuadrato-yugal (LCY) 0.278
Distancia entre los cóndilos mandibulares (DCM) 0.2749
2
Longitud del supraoccipital (LSO) 0.5032
Longitud desde el borde posterior del frontal hasta el extremo
posterior (LBPFEP)
0.3134
Longitud desde la unión frontal/postorbital parietal, hasta el
extremo posterior (LUFEP)
0.2819
Longitud del escamoso (LES) 0.253
Distancia desde el extremo postero-inferior del cuadrado hasta
el extremo anterior del pico (DEPICP)
-0.502
Distancia entre cóndilos mandibulares (DCM) -0.3233
Longitud de la mandíbula inferior (LMI) -0.1944
Longitud de la mandíbula inferior (LMI) 0.3195
Distancia desde el extremo postero-inferior del cuadrado hasta
el extremo anterior del pico (DEPICP)
0.2578
Longitud del palatino (LP) 0.230

35
3 Longitud del articular (LA) 0.2118
Longitud de la órbita (LO) 0.2065
Amplitud total (AmT) -0.5476
Distancia entre los cóndilos mandibulares (DCM) -0.4219
Tras revisar las cargas del primer componente resaltan cinco variables
principales: amplitud total (AmT), longitud del escamoso (LES), altura del cuadrato-
yugal (AlCY), longitud del cuadrato-yugal (LCY) y distancia entre los cóndilos
mandibulares (DCM). Gráficamente observamos que las cargas poseen valores
positivos (Figura 39); destacamos aquellas que mostraron mayor proximidad a 1,
debido a que esto significa que dentro del primer componente tienen mayor peso y
su relación es positiva. La variable de la amplitud total fue la que mostró mayor
relevancia.
Figura 39. Cargas del primer componente principal. Las variables AmT, LES, AlCY, LCY y DCM,
fueron las que mostraron mayor proximidad a 1.

36
Las cargas del segundo componente muestran siete variables representativas:
longitud del supraoccipital (LSO), longitud desde el borde posterior del frontal hasta
el extremo posterior (LBPFEP), longitud desde la unión frontal/postorbital parietal,
hasta el extremo posterior (LUFEP), longitud del escamoso (LES), distancia desde el
extremo postero-inferior del cuadrado, hasta el extremo anterior del pico (DEPICP),
distancia entre cóndilos mandibulares (DCM), longitud de la mandíbula inferior (LMI)
(Tabla 5).
Al representar gráficamente las cargas del segundo componente (Figura 40)
se observan las siete variables relevantes. De éstas, tres presentan una relación
negativa, implicando una relación inversa con las variables restantes.
Figura 40. Cargas del segundo componente principal. Se encontraron correlaciones negativas en las
variables DEPICP, DCM y LMI.
En las cargas del tercer componente (Tabla 8) resaltan nuevamente un par de
variables con cargas negativas, lo que indica una relación inversa. Aunque existen
variables con cargas positivas, las dos más representativas se encuentran hacia el

37
componente negativo del plano. En el caso particular de la distancia entre los
cóndilos mandibulares (DCM), ésta aparece con una carga negativa representativa
en el segundo y tercer componente; mientras que en el primero presenta valores
positivos. Otra variable con un comportamiento opuesto es la amplitud total (AmT);
en el CPIII aparece con valores negativos, pero posee la carga más representativa (-
0.5476). En el CPI de igual modo es la más representativa pero toma valores
positivos (0.4125).
Figura 41. Cargas del tercer componente principal. En este componente las cargas más
representativas las presentaron las variables AmT y DCM.
De este modo podemos decir que las variables más representativas para
diferenciar por grupos son: la amplitud total (AmT), longitud del supraoccipital (LSO),
la distancia entre los cóndilos mandibulares (DCM) y la distancia desde el extremo
postero-inferior del cuadrado hasta el extremo anterior del pico (DEPICP).

38
MANOVA
Para tener un sustento estadístico respecto a nuestros resultados, se realizó
un análisis de varianza múltiple o MANOVA, utilizando las variables morfométricas
utilizadas en el ACP. Esto nos indicaría si las diferencias entre las variables usadas
para diferenciar entre individuos son o no estadísticamente significativas. Partiendo
de la siguiente hipótesis.
H0: Las diferencias encontradas entre los individuos son significativas.
Ha: Las diferencias encontradas entre los individuos no son significativas.
El MANOVA arrojó una F=1.44 con una significancia p=0.21 (teniendo en
cuenta una probabilidad de 0.05 (95% de confianza)). Por lo que la H0 es rechazada,
lo que significa que las diferencias entre los individuos no son estadísticamente
significativas.
7.3 Alimentación
La dieta de las tortugas marinas cambia con la edad. En su estadio juvenil se
alimentan principalmente de plancton, pero al crecer varían el tipo de presas que
consumen (Badillo, 2007). Para Caretta caretta podemos definir su alimentación en 3
grandes grupos: crustáceos, moluscos y peces. De acuerdo con la bibliografía
consultada, en ocasiones también se alimentan de equinodermos (pepinos de mar) y
algas (Sargassum spp).
A continuación se muestra la agrupación por fila de las principales presas de
las que se alimenta Caretta caretta, con su respectiva consistencia y talla (Tabla 6).

39
Tabla 6. Presas principales de Caretta .caretta ordenadas por fila; se muestran las tallas y su
consistencia.
FILUM ESPECIE CONSISTENCIA Talla Cm
Max Prom Min
CRUSTACEA Arenaeus cribrarius (Andrade et
al, 2013).
Dura (Quitina) 15.4 14.1 X
Cancer irroratus (Bigford, 1979). Dura (Quitina) 14.1 3.0 X
Callinectes sapidus (Sutton y
Wagner, 2007).
Dura (Quitina) 21 18 X
Hemisquilla ensigera
californiensis (Basch y Engle,
1987).
Dura (Quitina) 30 20 18
Hepatus epheliticus (Hernáez et
al, 2012).
Dura (Quitina) 7.6 6.4 X
Menippe mercenaria (Lindberg y
Marshal, 1984).
Dura (Quitina) 17.0 13.0 7.0
Persephona mediterránea
(Martínez et al, 2009).
Dura (Quitina) 4.4 X 1.0
Platymera gaudichaudii
(Retamal et al, 2013).
Dura (Quitina) 9.0 5.0 X
Pleuroncodes planipes
(Longhurst y Seibert, 1971).
Dura (Quitina) 1.5 1.3 0.9
Polybius henslowii (Munilla,
1997).
Dura (Quitina) 4.2 3.1 2.4
Phronima sedentaria (Báez et
al, 2012).
Dura (Quitina) 4.0 1.6 0.8
Limulus polyphemus (Walls et
al, 2002).
Dura (Quitina) 60 40 25
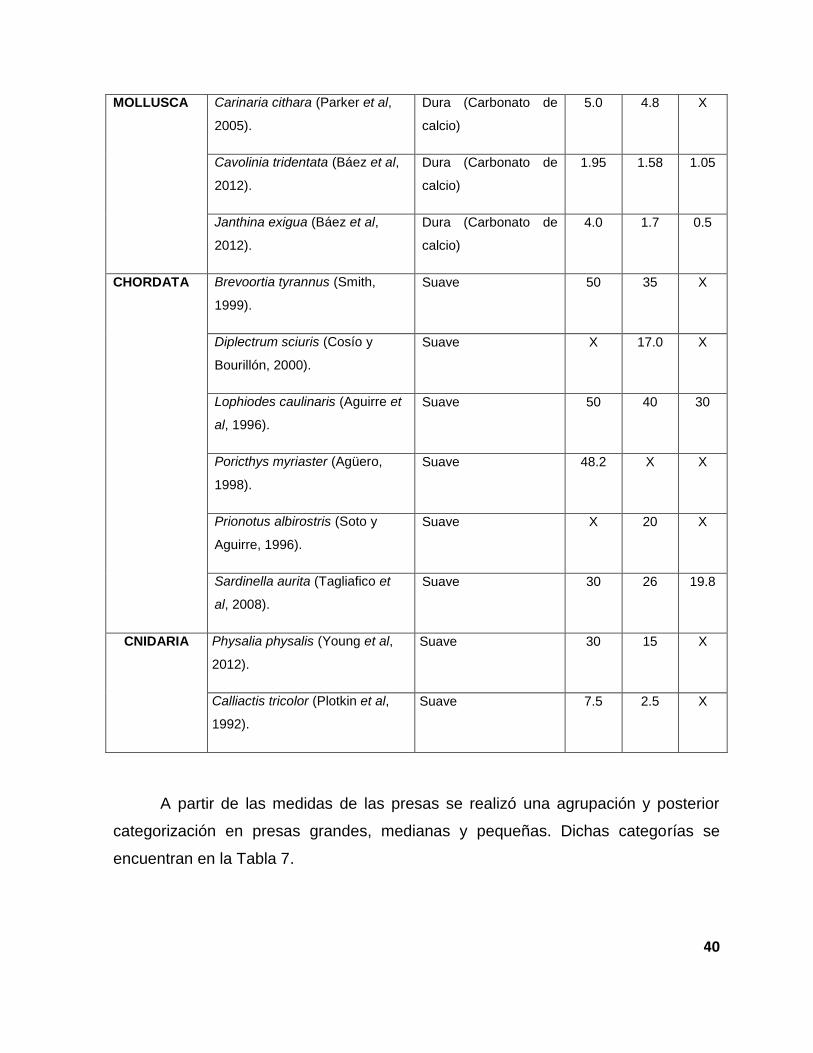
40
MOLLUSCA Carinaria cithara (Parker et al,
2005).
Dura (Carbonato de
calcio)
5.0 4.8 X
Cavolinia tridentata (Báez et al,
2012).
Dura (Carbonato de
calcio)
1.95 1.58 1.05
Janthina exigua (Báez et al,
2012).
Dura (Carbonato de
calcio)
4.0 1.7 0.5
CHORDATA Brevoortia tyrannus (Smith,
1999).
Suave 50 35 X
Diplectrum sciuris (Cosío y
Bourillón, 2000).
Suave X 17.0 X
Lophiodes caulinaris (Aguirre et
al, 1996).
Suave 50 40 30
Poricthys myriaster (Agüero,
1998).
Suave 48.2 X X
Prionotus albirostris (Soto y
Aguirre, 1996).
Suave X 20 X
Sardinella aurita (Tagliafico et
al, 2008).
Suave 30 26 19.8
CNIDARIA Physalia physalis (Young et al,
2012).
Suave 30 15 X
Calliactis tricolor (Plotkin et al,
1992).
Suave 7.5 2.5 X
A partir de las medidas de las presas se realizó una agrupación y posterior
categorización en presas grandes, medianas y pequeñas. Dichas categorías se
encuentran en la Tabla 7.

41
Tabla 7. Categorización de las presas por su talla.
PRESAS PEQUEÑAS (0.5-9cm) PRESAS MEDIANAS (10-24cm) PRESAS GRANDES (25-50 cm)
Carinaria cithara Arenaeus cribrarius Brevoortia tyrannus
Calliactis tricolor Callinectes sapidus Limulus polyphemus
Cavolinia tridentata Cancer irroratus Lophiodes caulinaris
Hepatus epheliticus Diplectrum sciuris Poricthys myriaster
Janthina exigua Hemisquilla ensigera californiensis Sardinella aurita
Persephona mediterránea Menippe mercenaria
Phronima sedentaria Physalia physalis
Platymera gaudichaudii Prionotus albirostris
Pleuroncodes planipes
Polybius henslowii
Del mismo modo, tomando en cuenta la longitud total del caparazón de los
individuos, se clasificaron en tres categorías diferentes: I (50-59 cm), II (60-69 cm) y
III (70-79 cm).
Después de categorizar los individuos y las presas con base en su tamaño, se
generó la siguiente relación (Tabla 8). Esta contrasta el tamaño de las presas con
respecto al tamaño de los individuos y nos da un panorama general en cuanto al
posible consumo de las presas por tamaño.

42
Tabla 8. Contraste entre las tallas de las presas y la talla de las tortugas.
INDIVIDUOS PRESAS PEQUEÑAS
(0.5-9cm)
PRESAS MEDIANAS (10-
24cm)
PRESAS GRANDES (25-
50 cm)
Talla de
Caparazón
I(50-59cm)
X
Talla de
Caparazón II
(60-69cm)
X X
Talla de
Caparazón III
(70-79cm)
X X
7.4 Correlación alimentación-talla
En la Tabla 9 se observan los valores obtenidos al realizar los intervalos de
clase para construir la matriz de presas (presencia-ausencia). La obtención de
valores descriptivos básicos, permitió observar el comportamiento de los datos de
forma general y para cada una de las clases formadas.

43
Tabla 9. Valores máximos, mínimos, promedio y desviación estándar de las longitudes totales de los
cráneos. Nótese la formación de clases con sus respectivos valores promedio y desviación estándar.
LONGITUD TOTAL DEL CRÁNEO
MAXIMO 19.86 CLASE LIM. INF LIM. SUP FREC PROM D. EST
MINIMO 14.14 I 14.14 16.04 11 14.95 0.61
PROMEDIO 16.32 II 16.04 17.94 12 16.87 0.57
D. ESTANDAR 1.51 III 17.94 19.84 2 18.83 0.70
IV 19.84 21.74 1 19.86 X
El arreglo obtenido para representar la matriz de presas se observa a
continuación (Tabla 10):

44
Tabla 10. Matriz de presencia-ausencia incluyendo las 23 presas. Se observan los intervalos de clase formados a partir de las longitudes totales de los cráneos, la frecuencia y el consumo (1) ó no (0) de las presas. El número en las columnas representa cada una de las siguientes presas.(1) Arenaeus cribrarius, (2) Polybius henslowii, (3) Phronima sedentaria,(4) Pleuroncodes planipes, (5) Cavolinia tridentata, (6) Janthina exigua, (7) Callinectes sapidus, (8) Cancer irroratus, (9) Brevoortia tyrannus, (10) Sardinella aurita, (11) Limulus polyphemus, (12) Prionotus albirostris, (13) Hemisquilla ensigera californiensis, (14) Lophiodes caulinaris, (15) Poricthys myriaster, (16) Diplectrum sciuris, (17) Hepatus epheliticus, (18) Menippe mercenaria, (19) Persephona mediterránea, (20) Platymera gaudichaudii, (21) Carinaria cithara, (22) Physalia physalis, (23) Calliactis tricolor.
Organismo Clase 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23
1 I 0 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 1 0 1 1 1 0 1
2 I 0 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 1 0 1 1 1 0 1
3 I 0 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 1 0 1 1 1 0 1
4 I 0 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 1 0 1 1 1 0 1
5 I 0 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 1 0 1 1 1 0 1
6 I 0 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 1 0 1 1 1 0 1
7 I 0 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 1 0 1 1 1 0 1
8 I 0 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 1 0 1 1 1 1 1
9 I 0 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 1 0 1 1 1 1 1
10 I 0 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 1 0 1 0 1 1 1
11 I 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 1 1 1
12 II 1 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 1 1
13 II 1 1 0 1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0
14 II 1 0 0 1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0
15 II 1 0 0 1 0 0 0 0 0 0 0 1 1 0 0 1 0 0 0 0 0 1 0
16 II 1 0 0 1 0 0 0 1 0 0 0 1 1 1 0 1 0 0 0 0 0 1 0
17 II 1 0 0 1 0 0 0 1 0 0 0 1 1 1 0 1 0 0 0 0 0 1 0
18 II 1 0 0 1 0 0 0 1 0 0 0 1 1 1 0 1 0 0 0 0 0 1 0
19 II 1 0 0 1 0 0 0 1 0 0 0 1 1 1 0 1 0 0 0 0 0 1 0
20 II 1 0 0 1 0 0 1 1 0 0 0 1 1 1 1 1 0 0 0 0 0 1 0
21 II 1 0 0 1 0 0 1 1 1 0 0 1 1 1 1 1 0 1 0 0 0 1 0
22 II 1 0 0 1 0 0 1 1 1 0 0 1 1 1 1 1 0 1 0 0 0 1 0
23 II 1 0 0 1 0 0 1 1 1 1 1 1 1 1 1 1 0 1 0 0 0 1 0
24 III 1 0 0 1 0 0 1 1 1 1 1 1 1 1 1 1 0 1 0 0 0 1 0
25 III 1 0 0 1 0 0 1 1 1 1 1 1 1 1 1 1 0 1 0 0 0 1 0
26 IV 1 0 0 1 0 0 1 1 1 1 1 1 1 1 1 1 0 1 0 0 0 1 0

45
Teniendo en cuenta que la ecomorfología propone que la morfología de las
estructuras de las especies determina una porción importante de la ecología
alimentaria de las mismas, la correlación entre las variables morfológicas y los
hábitos alimentarios, definirá el posible espectro de presas que Caretta caretta podría
consumir. Sin embargo el comprobar exitosamente este supuesto, dependerá de la
calidad de los resultados que han sido analizados por separado para el presente
trabajo (alimentación y variables craneales).
El resultado de la prueba de Mantel nos muestra que no existe una correlación
entre nuestras matrices. A partir de 9999 permutaciones, se obtuvo un valor de r=-
0.01 con un p=0.58; que implica una relación sin significancia estadística. Este
análisis indica de manera cuantitativa la influencia de la variación de las
características que determinan la habilidad de C. caretta para alimentarse de tipos de
presas de tamaños específicos; en este caso, no encontramos dicha influencia.
Adicional a la prueba de Mantel, se desarrolló una regresión lineal entre las
variables lineales y tróficas obtenidas a partir del cálculo del análisis de Bray-Curtis.
El resultado indica que no existe una correlación alta entre ambas matrices de datos;
coeficiente de correlación de R2 0.00137 y pendiente de 0.461. La regresión nos
permite observar gráficamente (Figura 42) que no existe una relación entre ambas
matrices de datos.

46
Figura 42. Regresión Lineal que muestra la asociación entre las medidas lineales y la presencia-ausencia de las presas con base en el análisis de Bray-Curtis. El coeficiente de correlación R2 dice que no existe un buen ajuste de los datos.
8. DISCUSIÓN
Los valores descriptivos obtenidos para las variables, arrojaron entre cuatro y
cinco intervalos de clase y también mostró que, salvo por las variables; longitud de la
órbita (LO), longitud de la unión frontal postorbital/parietal/ hasta el extremo posterior
(LUFEP), longitud del supraoccipital (LSO), distancia entre los cóndilos mandibulares
(DCM), amplitud del pico (AmP), longitud del pterigoides (LPT), distancia desde el
extremo postero-inferior del cuadrato-yugal, hasta la unión posteroparietal-escamoso
(DCMPES) y longitud total (LTL) (Figuras 10, 12, 16, 18, 21, 22, 32 y 33
respectivamente), no hay una distribución normal en nuestros datos. Todo esto pudo
deberse a que no todos los cráneos presentaron todas las estructuras tomadas en
cuenta para el análisis morfométrico. La distribución de los datos podría deberse al
tamaño de muestra, el cual podría parecer bajo al apenas superar el tamaño mínimo
de 30, comúnmente usado en estadística.

47
Sin embargo se debe considerar que no siempre se obtendrá una distribución
normal. Habitualmente se considera que para obtener una distribución normal,
cualquier tamaño de muestra por encima de 30 es suficiente en casi todos los casos
(Norman y Streiner, 2005). Aunque este tamaño varió, no podemos afirmar que haya
influido en la distribución observada, ya que sólo dos variables presentaron tamaños
de muestra menores a 30: la amplitud total (AmT) y la distancia entre los cóndilos
mandibulares (DCM) (figuras 17 y 18 respectivamente); esta última ajustándose a la
distribución normal. Esto indica que la ausencia de normalidad se debe a la
naturaleza misma de los datos. Aunque idealmente se espera trabajar con
distribuciones normales, esto no siempre se aplica a todas las características de la
naturaleza (Daniel, 2008).
Las altas correlaciones encontradas para las ocho variables (Tabla 3), pueden
deberse a que en conjunto forman parte del dermatocráneo. El dermatocráneo es la
parte externa del cráneo de los vertebrados, forma la caja ósea que protege al
cerebro, la limitante del techo de la boca y encierra parte del esplacnocráneo
(Kardong, 2007). En el caso de las tortugas, el cráneo se compone por el
neurocráneo, donde se aloja el cerebro y una estructura ósea externa, el
esplacnocráneo. Este último al igual que en el resto de los reptiles, contribuye a la
formación de la mandíbula y el aparato hioideo (Wyneken, J., 2004; González y
Acosta., 2006). Al ser todas, parte de una misma estructura funcional, es coherente
el que estuvieran relacionadas, lo que ocasionaría que arrojaran información
redundante para los análisis posteriores. Recordemos que el cráneo es una
estructura formada distintas partes, que fusionadas forman una unidad armoniosa
(Kardong, 2007). Por lo que al trabajar con una sola estructura de este conjunto en
particular, se obtendrá información del resto.
La longitud de las narinas externas (LNEX) se encontró relacionada con la
longitud del maxilar (LM) y la altura del cuadrato-yugal (AlCY). La relación entre estas
dos últimas podría hablarnos de la vía alimentarias del organismo. A diferencia de
otras especies, los maxilares de Caretta caretta entran en contacto con el paladar,

48
que separa total o parcialmente los canales respiratorios de los alimenticios
(Wyneken, 2004). Por su parte, el cuadrato-yugal forma parte de las estructuras de
fijación de los músculos aductores, responsables de cerrar la mandíbula al capturar a
las presas. Aunque es difícil asegurar una relación biomecánica de estas estructuras,
la relación encontrada tiene cierta coherencia si se piensa que una mayor superficie
del cuadrato-yugal implicaría una mayor área de inserción muscular y una mayor
superficie de contacto con el maxilar, teóricamente ambas características permitirían
la generación de una mordida fuerte.
La longitud desde el borde posterior del frontal hasta el extremo posterior
(LBPFEP), se relacionó con tres variables; longitud desde la unión frontal/postorbital
parietal hasta el extremo posterior (LUFEP), longitud del postorbital (LPO) y longitud
del dentario (LD). Estas estructuras salvo por la longitud del dentario (LD), se
encuentran muy cercanas una de otra y básicamente difieren solo en el punto de
unión con el frontal. Por otro lado el ancho del frontal (AnF) se correlacionó con la
longitud del postorbital (LPO) y la distancia desde el extremo postero-inferior del
cuadrato-yugal hasta la unión posteroparietal-escamoso (DCMPES). La relación
entre estas estructuras es una de las más relevantes, biológicamente hablando, ya
que estas forman parte de la bóveda craneal.
Aunque algunas de las correlaciones analizadas se centran en las estructuras
presentes en la parte dorsal del cráneo, otras nos muestran las relaciones entre
aquellas que se ubican hacia la parte ventral y lateral del mismo. Estas son de
relevancia en este trabajo, debido que podrían tener más participación en el proceso
de alimentación, particularmente en el caso del dentario, el maxilar y el cuadrato-
yugal.
Se encontró que la longitud del postorbital (LPO) se correlacionó con la
amplitud de la fosa infratemporal (AmFI), amplitud del pico (AmP), longitud del
cuadrato-yugal (LCY), altura del cuadrato-yugal (AlCY), distancia desde el extremo
postero-inferior del cuadrato-yugal hasta la unión posteroparietal-escamoso
(DCMPES) y la longitud del dentario (LD). En algunas de estas estructuras se

49
insertan músculos que ayudan al proceso de alimentación. El músculo depresor
mandibular, se origina en los huesos cuadrado, cuadrato-yugal y escamoso
insertándose en la mandíbula inferior (Wyneken, 2004). Nuevamente encontramos
estructuras que coinciden con las que se analizaron en el estudio.
Así mismo la longitud de la fosa infratemporal (LFI) se correlacionó con
longitud del maxilar (LM), longitud del cuadrato-yugal (LCY), altura del cuadrato-yugal
(AlCY) y longitud del articular (LA). En este conjunto de variables, el maxilar es la
estructura que podría tener la inserción muscular. El intermandibular se extiende
desde la mandíbula inferior hasta el tendón del músculo pseudotemporal, que
continua hasta el hueso parietal, constrictor colli que es una prolongación del
intermandibular, debajo de este se encuentran los genihioideo, que corren
oblicuamente entre las mandíbulas, todos estos músculos ayudan a tragar y a
bombear la garganta (Wyneken, 2004). La fosa infratemporal como tal también
podría estar participando en la inserción de muscular, puesto que esto fue observado
al momento de la limpieza de las muestras, es probable que alguno de los músculos
antes mencionados también se inserte en parte dentro de este amplio espacio.
La amplitud del pico (AmP) se encontró correlacionada con la distancia desde
el extremo postero-inferior del cuadrato-yugal hasta la unión posteroparietal-
escamoso (DCMPES) y la longitud del dentario (LD), mientras que la longitud del
maxilar (LM) con la longitud del cuadrato-yugal (LCY), altura del cuadrato-yugal
(AlCY) y la longitud de la mandíbula inferior (LMI). Estas relaciones podrían indicar
que las estructuras mencionadas juegan un papel primordial durante la alimentación
para esta especie en particular. De acuerdo con Wyneken (2004), aunque los huesos
craneales en las tortugas presentan una estructura similar, la forma de los huesos del
paladar presenta diferencias entre especies. Estas diferencias en el paladar además
de ser diagnosticas podrían indicar algún tipo de estrategia alimentaria. Esto podría
hablar de la eficiencia alimentaria debajo del mar, pues esta especie expulsa agua
por los nostrilos mientras se alimenta (Bjorndal, 1985). Su paladar, único entre las

50
tortugas marinas, probablemente evite que la tortuga trague agua mientras se
alimenta, prolongando así su tiempo de buceo.
Otra correlación encontrada es la distancia desde el extremo postero-inferior
del cuadrato-yugal hasta la unión posteroparietal-escamoso (DCMPES), con la
longitud del articular (LA) y la longitud del dentario (LD). Estas relaciones hacen
referencia a las estructuras asociadas a la mandíbula inferior, en estas estructuras
también encontramos referencias de inserciones musculares. El músculo depresor
mandibulae, que se encarga de la apertura mandibular y que se origina en los
huesos cuadratal, cuadrato-yugal y escamoso; insertándose en el articular de la
mandíbula inferior, a excepción de D. coriacea en la que se inserta en el conducto
auditivo (Wyneken, 2004). La relevancia de estas estructuras se encuentra en el
hecho que parecen ser las responsables de la fuerza mandibular.
Sin embargo debemos tomar en cuenta que el tipo de inserción muscular
juega un papel importante en la eficiencia de los músculos, en el caso de las
mandíbulas, la fuerza de una mordida depende de la dirección y punto de aplicación
de las fuerzas ejercidas por los músculos que cierran la mandíbula (Kane y
Sternheim, 1989). De esta forma se observa la asociación funcional (apertura
mandibular) en este conjunto de estructuras correlacionadas.
Tras revisar las correlaciones obtenidas entre nuestras variables en un sentido
anatómico, podemos vislumbrar la influencia que tiene cada estructura sobre otra u
otras (en mayor o menor medida); lo que explica las correlaciones altas y positivas.
Se debe considerar que el cráneo es una estructura por demás compleja, resultado
de la unión de varios huesos (Parker et al., 1991). Las características óseas
seleccionadas tras el análisis de correlación, se agrupan conformando el
dermatocráneo, por lo que cada una se involucra en diferentes funciones, que se
desarrollan en esta estructura. De esta manera se constituyen lo que consideramos
como componentes funcionales, los cuales pueden ser descritos de acuerdo a las
funciones primarias que cumplan: alimentario, respiratorio y alojamiento del cerebro
(bóveda craneana).

51
La longitud de las narinas externas (LNEX), en conjunto con la longitud del
maxilar (LM) y la altura del cuadrato-yugal (AlCY) podrían actuar en la funciones
respiratorias durante la alimentación., Así mismo, la longitud desde el borde posterior
del frontal hasta el extremo posterior (LBPFEP) en conjunto con la longitud desde la
unión frontal/postorbital parietal hasta el extremo posterior (LUFEP), la longitud del
postorbital (LPO), la longitud del dentario (LD) y el ancho del frontal (AnF)
relacionado con la longitud del postorbital (LPO) y la distancia desde el extremo
postero-inferior del cuadrato-yugal hasta la unión posteroparietal-escamoso
(DCMPES) actuarían funcionalmente en conjunto como parte de la bóveda craneal.
Mientras que la longitud de la fosa infratemporal (LFI), en conjunto con la longitud del
maxilar (LM), la longitud del cuadrato-yugal (LCY), la altura del cuadrato-yugal (AlCY)
y la longitud del articular (LA) formarían parte del componente alimentario del
organismo, encargado de la captura de la presa.
El primer componente principal (CPI) explicó un porcentaje de variación
bastante alto (62.98%). Al observar las variables que lo conforman, notamos
nuevamente la relevancia del ancho del cráneo; la variable con el valor más
representativo en cuanto a las cargas fue la amplitud total (AmT). Al respecto, se
menciona que el cráneo de C.caretta es más ancho hacia su parte posterior en
comparación con su parte anterior, lo que le da su forma característica (Wyneken,
2004); esto podría indicar una mordida potente, necesaria para poder cubrir una dieta
como la que se reporta para esta especie. Aunque dentro de su dieta se encuentran
peces, algas y medusas, la mayor parte de su alimentación se centra en el consumo
de crustáceos y moluscos (Peckham, 2008); presas que debido a la constitución de
sus tejidos, requieren una mayor fuerza de mordida para poder ser capturadas.
Pareciera que el CPII destaca las variables que dan indicios sobre el tamaño
general del cráneo. Este componente estuvo conformado principalmente por la
distancia desde el extremo postero-inferior del cuadrado hasta el extremo anterior del
pico (DEPICP) y la distancia entre cóndilos mandibulares (DCM). Variables que al
parecer fueron fundamentales para la conformación de las agrupaciones (Figura 37);

52
es probable que a partir de estos rasgos anatómicos pudiéramos definir el tamaño
aproximado de los individuos de esta especie. Resultados similares se han
encontrado en otros trabajos, en los que definen que el tamaño de la órbita y del
supraoccipital varía y puede diferenciar organismos juveniles y adultos (Nishizawa et
al., 2010).
Por su parte, el CPIII resalta variables que se descartaron a partir de los dos
primeros componentes. Aunque la varianza explicada es baja (7.19%), podría
confirmar las variables de importancia en la diferenciación entre individuos de C.
caretta: amplitud total (AmT) y distancia entre los cóndilos mandibulares (DCM)
(Figura 38). Ambas nos indican mandíbulas anchas, que permitirían la captura de
presas más grandes.
De acuerdo con varios autores, la dieta de Caretta caretta varía conforme a
su edad (Bjorndal.,1985, Lutz et al.,1997 y Badillo., 2007). Sin embargo la selección
de presas también podría estar influenciada por el desarrollo de los cóndilos
mandibulares y en este caso, los organismos no cambiarían su dieta hasta que estas
estructuras les permitieran generar la fuerza suficiente como para triturar crustáceos
y bivalvos o sujetar peces de mayor tamaño.
Esencialmente, el ACP destacó las variables relacionadas con la robustez del
cráneo, la cual parece estar definida por la amplitud total (AmT) y la distancia entre
los cóndilos mandibulares (DCM). Estas a su vez podrían afectar la longitud de la
mandíbula inferior (LMI) y en menor medida la longitud del escamoso (LES), lo cual
puede deducirse por las cargas obtenidas en el ACP en los componentes II y III
(Figuras 41 y 42) ya que presentan valores opuestos entre sí. Bjorndal (1985)
menciona que para C. mydas el tipo de alimentación puede determinar la interacción
entre el hábitat y el organismo; en este sentido podría ser que el desarrollo de las
tortugas marinas se encuentre ligado al uso de sus hábitats y que para Caretta
caretta (al igual que en otras especies), el óptimo desarrollo de estructuras craneales
como los cóndilos mandibulares, el escamoso y el articular tal vez tengan una
repercusión en la eficiencia para aprovechar los recursos disponibles en el medio.

53
Sin embargo esto no pudo ser comprobado en este trabajo puesto que no se
encontró una relación entre las mediadas lineales y las presas.
Igualmente, la consistencia de las presas podría relacionarse con la fuerza
generada por las estructuras craneales. Tras analizar las presas de Caretta caretta,
encontramos que una parte considerable de ellas se encuentran blindadas; como es
el caso de algunos crustáceos como Arenaeus cribrarius, Cancer irroratus,
Callinectes sapidus, y Hemisquilla ensigera californiensis o los bivalvos Carinaria
cithara y Cavolinia tridentata, por mencionar algunos.
Este blindaje, proporcionado ya sea por una concha o una armadura de
quitina, puede hacer difícil su consumo por la dureza de su consistencia. Por otro
lado, aquellas presas que hacen parte de los hábitos alimentarios de esta especie y
que son blandas, se caracterizan porque presentan tamaños considerables:
Sardinella aurita (26 cm), Lophiodes caulinaris (40 cm) y Brevoortia tyrannus (35 cm).
En ambos casos una mordida poderosa proporcionada por estructuras craneales
robustas como las mencionadas en este trabajo sería sumamente efectiva ya sea
para abatir a las presas grandes, o para triturar aquellas que se encuentran
fuertemente blindadas.
El sistema generado en la mandíbula de los depredadores debe cumplir con
dos rasgos fundamentales: 1) permitir el cierre rápido de la boca en el momento en
que la presa entre a la apertura bucal o cuando puede ser atrapada en algún punto
entre los dientes (en este caso en algún punto del pico queratinoso) y 2) producir una
fuerza de mordida suficiente para lograr inmovilizar, triturar o desgarrar piezas de la
presa (Franco, 2011). De acuerdo con Silman et al., (2002) esta especie es
principalmente carnívora y suele alimentarse de almejas y cangrejos entre otros
invertebrados, empleando sus poderosas mandíbulas para capturar a sus presas.
Sin embargo, no se debe descartar la posibilidad de que las presas blandas
sean obtenidas por oportunismo. Aparentemente los peces que en ocasiones se
registran en la dieta de la especie, parecen proceder del aprovechamiento de los
descartes de la pesca (Bjorndal, 1985), ya que se duda que en realidad Caretta

54
caretta pudiera atrapar peces vivos. De igual manera, sugieren que las algas que
ocasionalmente aparecen dentro de su dieta, son consumidas de forma accidental
pues éstas aparentemente no le aportan ningún valor nutritivo (Sagarminaga et al.,
2006).
Después de todo lo expuesto anteriormente, se puede decir aunque de
manera muy general, que existen estructuras óseas encargadas de la robustez del
cráneo y que podrían cobrar importancia para la alimentación de Caretta caretta. Sin
embargo la prueba de Mantel mostró una correlación negativa entre las medidas
craneales y el tamaño de las presas, impidiendo relacionar alguna de las estructuras
con alguna de las presas.
Esto podemos atribuirlo a distintos factores;
En cuanto a las presas podría ser la fuente de los datos; ya que el trabajo se
realizó con valores tomados de la bibliografía para las tallas de las presas. Se podría
obtener un resultado más preciso al trabajar con presas obtenidas directamente en
los contenidos estomacales de los organismos y al determinar índices alimentarios
para conocer el aporte energético (índice gravimétrico) y la disponibilidad de las
presas (índice numérico).
En cuanto a los organismos usados en el presente estudio podría ser el
tamaño de muestra; pese a que nuestros datos no presentaron una distribución
normal, un mayor tamaño de muestra le daría más robustez a los resultados; también
las variables seleccionadas, ya que posiblemente se omitieron variables importantes
en los análisis realizados en el presente trabajo y por último las técnicas utilizadas;
ya que se podría encontrar una correlación más alta entre las variables de forma y la
alimentación de los individuos, haciendo uso de técnicas como la Morfometría
Geométrica.
Aunque el modelo de regresión lineal supone que es posible la predicción
exacta del comportamiento de los datos, en la práctica esto casi nunca es alcanzable
(Ross, 2007). Es posible observar (Figura 42) que los valores se encuentran

55
dispersos alrededor de la línea de tendencia, mostrando una relación muy pobre.
Esto en definitiva puede ser causa del origen de las matrices de datos, ya que una es
meramente bibliográfica, lo que en si dificultó el poder encontrar alguna relación.
Caretta caretta presenta una buena variedad de presas dentro de su dieta, tal
como se comprobó tras la revisión bibliográfica, sin embargo resulta difícil determinar
si las presas forman parte de la dieta de adultos o juveniles. Lo anterior debido a que
no existe información que especifique en qué momento las crías deben ser
consideradas juveniles, además durante esta etapa es cuando las tortugas inician su
integración a los hábitos de adulto, pues hasta la fecha no se encuentra definida la
duración en años del estado juvenil (Márquez, 1996). Así pues se pueden observar
organismos juveniles y adultos compartiendo zonas de alimentación (Eckert y Abreu,
2001). Esto indica que no es posible definir una dieta exclusiva para adultos o para
juveniles.
Es importante destacar que en el presente trabajo no se caracterizó de
manera directa la dieta de la especie; por lo que la robustez de la información acerca
del espectro alimentario, se limita a los datos de presencia- ausencia generados a
partir de la búsqueda bibliográfica realizada y no se cuenta con información acerca
del aporte energético o la disponibilidad de las presas en el medio; los cuales
seguramente generarían una caracterización más robusta. Esto último también es
causal para que no se haya logrado la aproximación deseada entre la relación de las
medidas lineales y las presas.
9. CONCLUSIÓNES
Existen correlaciones generales de las estructuras craneales, las cuales
arrojaron una visión básica sobre el ancho del cráneo de Caretta caretta y la
robustez del mismo.
Aunque se exploraron las estructuras craneales de Caretta caretta, no fue
posible saber con certeza cuál o cuáles de estas están relacionadas con el
consumo de alguna presa en particular.

56
A pesar de no encontrar una relación medidas-presas, la distancia entre
cóndilos mandibulares (DCM) y la amplitud total (AmT) nos dan indicios de la
importancia del ancho de la mandíbula, fundamental para atrapar a sus presas
y triturar aquellas de textura dura.
10. RECOMENDACIONES
1. Aunque pudieron encontrarse algunas estructuras de importancia en el cráneo
de Caretta caretta, es probable que alguna se haya queda fuera de este
estudio, por lo que sería recomendable redefinir con que estructuras trabajar.
2. Probablemente la relación entre las medidas y las presas no haya sido
definida por la diferencia en la fuente de los datos, se debe considerar el
trabajar con datos provenientes de contenidos estomacales para así tener una
mayor certeza en cuanto a las tallas de presas y depredadores.
3. Dado que algunos de los análisis utilizados en este estudio no arrojaron
información concluyente, tal vez se debe utilizar otro tipo de técnicas y análisis
para encontrar las relaciones entre las medidas lineales y las presas.
4. Aunque los datos no tenían distribución normal, por la naturaleza misma de los
datos, sería altamente aconsejable dar más robustez a las matrices en
trabajos similares a este.
11. BIBLIOGRAFÍA
Aguirre, C. J.L., Acosta, G. A. F., Agüero. J.C y R.M. Estrada. 1996.
Ictiofauna marina-costera del Pacífico central mexicano: análisis
preliminar de su riqueza y relaciones biogeográficas. Marco Ambiental:
Biología de peces.

57
Agüero de la Cruz. J y Cota, G. V. M. 1998. Ictiofauna de la laguna San
Ignacio, Baja California Sur, México: Nuevos registros y ampliaciones de
ámbito. Ciencias Marinas. 24(3):353-358.
Álvarez, C. R. 1995. Estadística Multivariante y no Paramétrica con SPSS
Aplicación a las Ciencias de la Salud. Díaz de Santos, S.A. España. 408 pp.
Andrade. S, L., Fransozo, V., Bertini. G., Fransozo, N. L. M y Greco, L. S. L.
2013. Reproductive plasticity in the Speckled Crab Arenaeus cribratus
(Decapodoa, Brachyura, Portunidae) Associated with a population
decline. Journal of Coastal Research. Vol 0.
Anderson, R. E., Hair, J. F., Tatham, R. L y Black, W. C. 1999. Análisis
Multivariante. Prentice Hall. 348 pp.
Badii, M. H., Catillo, J., Cortez, A. W y Villalpando, P. 2007. Análisis de
correlación canónica (ACC) e investigación científica. Innovaciones de
Negocios. 4(2):405-422.
Báez, C. J, D. Macías, M. A. Puerto, J. A. Camiñas y J. M. Ortiz de Urbina.
2010. Análisis Biométrico de la Tortuga Boba Caretta caretta (Linnaeus
1758), en el Mediterráneo Occidental. Collec. Vol. Sci. Pap. ICCAT. 65(6):
2305-2309.
Báez, C. J., S. García-Barcelona., J. L. Rueda y D. Macías. 2012. Nuevas
aportaciones a la ecología trófica de Caretta caretta. Bol. Asoc. Herpetol.
Esp. 23 (1): 33-36.
Baillo, M. A y Grané, C. A. 2008. 100 Problemas resueltos de estadística
multivariante; implementados en MATLAB. Delta. España. 192 pp.
Badillo, A. F. J. 2007. Epizoítos y parásitos de la tortuga boba (Caretta
caretta) en el mediterráneo occidental. Tesis doctoral. Universidad de
Valencia. Facultad de Ciencias Biológicas. Departamento de Zoología. 262 p.

58
Basch, V. L y Engle, M. J. 1987. Biogeography of Hemisquilla ensigera
californiensis (Crustacea: Stomatopoda) with emphasis on Southern
California Populations. Third California Islands Symposium.211-220.
Bigford, E. T. 1979. Synopsis of Biological data on the rock crab, Cancer
irroratus say. NOAA Technical Report NMFS Circular 426. 26 pp.
Bjorndal, K. A. 1985. Nutritional ecology of sea turtles. American Society of
Ichthyologist and Herpetologist. 1985 (3): 736-751.
Bohórquez, H. J. 2009. Ecomorfología alimentaria de algunas especies de
peces asociadas a fondos blandos. Tesis de Maestría. CICIMAR. La Paz
Baja California Sur. 132p.
Bowen, B.W., F. A. Aubreu-Grobois., G. H. Balaz., N. Kamezaki., C.J. Limpus
y R. J. Ferl. 1995. Trans-pacific migrations of the loggerhead turtle
(Caretta caretta) demonstrated with mitochondrial DNA markers.
Population Biology. (92): 3731-3734.
Brusca, C. R y Brusca, J. G. 2005. Invertebrados. Mcgraw Hill
Interamericana. España. 1005pp.
Cabral, N. H., Márquez, F. J., Rego, A. L., Catarino, A. I., Figueiredo. J y
Garcia, J. 2003. Genetic and Morphological variation of Synaptura
lusitánica Capello, 1868, along the Portuguese coast. Journal of Sea
Research. (50): 167-175.
Chacón, D., Sánchez, J., Calvo, J. J. y Ash, J. 2007. Manual para el manejo y
la conservación de las tortugas marinas en Costa Rica; Con énfasis en la
operación de proyectos en playa y viveros. Sistema Nacional de Áreas de
Conservación (SINAC), Ministerio de Ambiente y Energía (MINAE). Gobierno
de Costa Rica. San José. 103p.

59
Comisión Nacional de Áreas Naturales Protegidas. 2011. Programa de
acción para la conservación de la especie: Tortuga Caguama, Caretta
caretta. Dirección general de operación regional. Dirección de especies
prioritarias para la conservación. Programa nacional de conservación de
tortugas marinas. 44pp.
Cosío, J. T y L. Bourillón. 2000. Inventario y monitoreo del canal del
Infiernillo para el comanejo de os recursos marinos en el territorio Seri,
Golfo de California. Conservation International México AC. Informe final
SNIB-CONABIO proyecto No. L179.76pp.
Danemann, D. G y E. Ezcurra. 2008. Bahía de los Ángeles: recursos
naturales y comunidad, Línea base 2007. Instituto Nacional de Ecología.
México. 740pp.
Daniel, W. W. 2008. Bioestadística: Base para el análisis de las ciencias
de la salud. Limusa Wiley. México. 755 pp.
Diez, M. F., Zapico, F. F., Álvarez, Ll. J. F., González, V. I., Getino, R. J y
Expósito, E. M. 2008. Formación superior en prevención de riesgos
laborales 3.a ed. Lex Nova. España. 698 pp.
Eckert, K. L. y F. A. Abreu Grobois. 2001. Proceedings of the regional
meeting “Marine turtle conservation in the wider Caribbean region: A
dialogue for effective regional management” Santo Domingo, 16-18
November 1999. WIDECAST, IUCN-MTSG, WWF y UNEP Caribbean
Environment Programme. 154 pp.
Ferreira-Júnior, R.L Treichel, T. L. Scaramussa y J.T. Scalfoni. 2010.
Morphometric pattern in Caretta caretta (Linnaeus, 1758) (Cheloniidae)
hatchlings from nest with different embryo development rates. Braz. J.
Biol. 71 (1):151-156.

60
Fleischer, A. L. y J. C. Urruchúa. 1986. Censos de ballenas grises
(Eschrichtius robustus) en Bahía Magdalena, B.C.S. México. Ciencia
Pesquera. Instituto Nacional de Pesca. Secretaría de Pesca. México. (5): 51-
62.
Funes, R. R., J.G. Gutiérrez y R. P. García. 2007. Estudios Ecológicos en
Bahía Magdalena. Gobierno del Estado de Baja California Sur. Secretaría de
Turismo de Baja California Sur. FOMAR. CICIMAR. 311 pp.
Funes, R. R y R. González A. 2002. Diversidad taxonómica de las larvas de
peces de Bahía Magdalena y zona nerítica adyacente (área prioritaria
costera No. 4). IPN. CICIMAR. Informe final SNIB CONABIO proyecto No.
S102. México D.F. 17p.
Franco, M. R. A. 2011. Morfología y desempeño del aparato mandibular de
seis especies de peces ictiófagos demersales, asociados a los fondos
blandos de la plataforma continental de Nayarit-Sinaloa. Tesis de
Maestría. CICIMAR. La Paz Baja California Sur. 127 p.
García, B. E. 1999. Implicaciones ecológicas y evolutivas del tamaño en
los artrópodos. Departamento de Biología (Zoología). Universidad Autónoma
de Madrid. (26): 657-678.
García, R. L. I. 2009. Tesis de Licenciatura “Morfometría de dos
poblaciones alopátricas de Chelonia, en el mar Caribe la tortuga verde
(Chelonia mydas) y en el Pacífico mexicano la tortuga negra (Chelonia
agassizii). Universidad Michoacana de San Nicolás de Hidalgo-Facultad de
Biología. 49p.
Gerosa, Guido y Aureggi Mónica. 2005. Guía para pescadores sobre el
manejo de las tortugas marinas: Manual del profesor. Programa de
Investigación y Conservación de las Tortugas Marinas. 49pp.

61
González, T y Acosta, N. 2006. Patrones morfológicos de los cordados.
Universidad Nacional de Colombia. Colombia. 197 pp.
González, R. A. 2005. Ecología aplicada: diseño y análisis estadístico.
Colección de Estudios de Ecología. Universidad de Bogotá, Jorge Tadeo
Lozano.325 pp.
González, R. E. 2008. Variabilidad de la productividad primaria en el Golfo de
Ulloa, costa occidental de Baja California. Tesis de Doctorado CICIMAR. La
Paz Baja California Sur. 143 p.
Granado, L. C. 1996. Ecología de peces: volumen 45 de serie de ciencias.
Universidad de Sevilla. España. 353pp.
Grupo Tortuguero de las Californias. 2012. Propuesta integral para la
conservación y protección de la tortuga caguama del Pacífico (Caretta
caretta) dentro de su hábitat crítico de alimentación y desarrollo en Golfo
de Ulloa, Baja California Sur. CONABIO. México. 26 pp.
Hammer, Ø. Harper, D.A.T. and P.D. Ryan. 2001. PAST: Paleontological
Statistics Software Package for Education and Data Analysis.
Paleontología Electronica 4(1): 9pp.
Hernáez, P., Rombeso, A., Pinheiro, A. A. M y Nuno. Simoes. 2012.
Population structure and sexual maturity of the calico box crab Hepatu
epheliticus Linnaeus (Brachyura, Hepatidae) from Yucatan Peninsula,
México. Latin American Journal of Aquatic Research. 40(2):480-486.
Hernández. S, R., Fernández. C. C Y Baptista. L, P. 1991. Metodología de la
investigación. McGraw-Hill Interamericana. Colombia. 505 pp.
Johnson, D. K. A., Sierra. C. J. L. y Erosa. S. A. I.1993. Un Tesoro de la
Naturaleza: Las Tortugas Marinas. EDAMEX. México. 117pp.

62
Jones. M. E. H., Werneburg. I., Curtis. N., Penrose. R., O´Higgins. P., Fagan J.
M y Evans .E. S. 2012. The head and neck anatomy of sea turtles
(Cryptodira: Chelonidae) and skull shape in testudines. PLoS ONE
7(11):1-28.
Kane, W. J y Sternheim, M. M. 1989. Física. REVERTÉ. España. 824 pp.
Kardong, K. V. 2006. Vertebrados: Anatomía comparada función y
evolución. Mc Graw Hill. España. 782 pp.
Lindbeg, J. W y Marshall, J. M. 1984. Species profiles: Life histories and
environment requirements of coastal fishes and invertebrates (South
Florida), Stone Crab. U.S. Fish and Wildlife Service. 17pp.
Longhurst, R. A y Seibert. L. R. Don. 1971. Breeding in an Oceanic
population of Pleuroncodes planipes (Crustacea, Galatheidae). Pacific
Science. (25):426-428.
Lutz, L. P., J. A. Musick. 1997. The biology of sea turtles. CRC PRESS.
EUA. 411 pp.
Lutz, L. P., J. A. Musick y J. Wyneken.2003. The biology of sea turtles vol. II.
CRC PRESS. EUA. 456 pp.
Machado, A y J. A. Bermejo. 2013. Estado de conservación de la tortuga
boba (Caretta caretta) en islas canarias, 2012. Santa Cruz de Tenerife:
Observatorio Ambiental Granadilla. 154 pp.
Márquez, René. 1996. Las tortugas marinas y nuestro tiempo. Fondo de
Cultura Económica. México. 104pp.
Martín, A. A y del Castillo, L. J.D. 2004. Bioestadística para las ciencias de
la salud. NORMA-CAPITEL. España. 647 pp.

63
Martínez, E. A., Ballesté. C. J. C., Castro. R. L., Navarro. I. F., Reguero. A. M.,
Medel. M. R. A., Ruiz. M. J., Ros. L. M. V y San José. H. N. 2011. Materiales
didácticos de biología. Guía práctica de maturita. Secciones bilingües de
Eslovaquia. Agregaduría de educación. España-Eslovaquia. 329pp.
Martínez, G., F. Scarabino y E. Delgado.2009. New records of the
brachyuran crabs Hepatus pudibundus (Aethridae) and Persephona
mediterranea (Leucosiidae) in their southernmost Western Atlantic
distribution. Pan-American Journal of Aquatic Sciences. 4(3):279-282.
Medina, Y., F. Moncada y G. Nodarse. 2011. Relaciones morfométricas de
la tortuga Carey (Eretmochelys imbricata) con vista a su aplicación en el
manejo. Revista Cubana de Investigaciones Pesqueras. 28(1):64-68.
Michel, M. J. E. 1995. Tesis de Maestría “Variabilidad Fenotípica en Crías
de Tortuga Marina Lepidochelys olivacea. CICIMAR. 99 p.
Molist, P., Pombal. A. M., Megías. M. 2011. Atlas de Histología Vegetal y
Animal. Departamento de Biología Funcional y Ciencias de la Salud. Facultad
de Biología. Universidad de Vigo. España. 30pp.
Moreno, C. E. 2001. Métodos para medir la biodiversidad. M y T-Manuales
y Tesis SEA. Vol 1. Zaragoza. 84 pp.
Munilla, Ignacio. 1997. Henslow´s swimming crab (Polybius henslowii) as
an important food for yellow-legged gulls (Larus cachinnans) in NW
Spain. Journal of Marine Science. 54: 631-634.
Nishizawa, H.,M. Asahara, N. Kamezaki y N. Arai. 2010. Differences in the
skull morphology between juvenile and adult green turtles: Implications
for the ontogenetic diet shift. Current Herpetology. 29(2):97-101.
Norman, G. R. y Streiner, L. D. 2005. Bioestadística. Harcourt International.
España. 261 pp.

64
Ocaña, O., A. G. de los Ríos y Los Huertos y A. Brito. 2006. The Crab
Polybius henslowii (Decapoda: Brachyura) as a main resource in the
Loggerhead turtle (Caretta caretta) diet from North Africa. Revista.
Académica. Canar. Cienc. 17 (4):103-116.
Olson, L. E., Salomon, K. A., Wirsing, J. A y Heithaus, R. M. 2012. Large-
scale movement patterns of male loggerhead sea turtles (Caretta caretta)
in shark bay, Australia. Marine and Freshwater Research. 63:1108-1116.
Parker, M. D., Cooke, J. W y Balazs. H. G. 2005. Diet of oceanic loggerhead
sea turtles (Caretta caretta) in the central North Pacific. Fishery Bulletin.
103 (1):142-152.
Parker, T. J., Haswell, W. A y Nadal, J. (revisor). 1991. Zoología: Cordados.
REVERTÉ, S. A. España. 983 pp.
Peckham, S. H. 2008. Demographic and conservation implications of
alternative foragin strategies in juvenile loggerhead turtle (Caretta
caretta) of the North Pacific Ocean. University of Santa Cruz. 85 p.
Peckham, S. H., Maldonado, D. D., Volker. K., Manici. A., Gaos. A., Tinker. M.
T y Nichols. W. J. 2008. High mortality of loggerhead turtles due to bycatch,
human consumption and stranding at Baja California Sur, México, 2003 to
2007. Endangered Species Research.
Pedrosa, H y Dicovskyi, L. 2007. Sistema de análisis estadístico con SPSS.
Instituto Nicaragüense de Tecnología Agropecuaria. Nicaragua. 153 pp.
Peña, Daniel. 2003. Análisis de Datos Multivariante. Mc Graw Hill
Interamericana. España. 539 pp.
Plotkin, P.T., Wicksten, M. K y Amos, A.F.1992. Feeding ecology of
loggerhead sea turtle Caretta caretta in the Northwestern Gulf of México.
Marine Biology. 115: 1-15.

65
Pritchard, H. C. P y J. A. Mortimer. 1999. Taxonomía, Morfología Externa e
Identificación de las Especies. Técnicas de Investigación y Manejo para la
Conservación de las Tortugas Marinas. K. L. Eckert, K. A. Bjorndal, A. Abreu-
Grobois, M. Donnelly (editores) UICN/CSE Grupo Especialista en Tortugas
Marinas Publicación No.4. 23-42.
Ramírez, G. A. 2005. Ecología aplicada; diseño y análisis estadístico.
Universidad de Bogotá, Jorge Tadeo Lozano. Colombia. 325 pp.
Ramírez, G. A. 2006. Ecología: Métodos de muestreo y análisis de
poblaciones y comunidades. Pontificia Universidad Javeriana. Colombia.
273pp.
Retamal, A. M., Ernst. B., Chamorro. J y Manríquez P. 2013. Descripción y
distribución geográfica de una nueva especie para el archipiélago Juan
Fernández: Platymera gaudichaudii (H. Milne-Edwards, 1837) (Decapoda,
Calappidae). Latin American Journal of Aquatic Research. 41(5):1010-1012.
Ross. M, Sheldon. 2007. Introducción a la Estadística. REVERTE. España.
809 pp.
Sagarminaga. R., Cañadas. A., Urquiola. E y Vázquez. J, A. 2006. Propuesta
de plan de conservación para la tortuga boba (Caretta caretta) en el
mediterráneo español (Región de Andalucía y Murcia). Sociedad Española
de Cetáceos. España. 94 pp.
Salazar, R. M. E., Hernández, A. S y Núñez, B. E. 2001.Coeficientes de
asociación. Plaza y Valdez. México. 168 pp.
Samuels, M. J y Valkenburg, V. B. 2009. Craniodental indicators of prey
size preference in the Felidae. Biological Journal of the Linnean Society.
(96): 784-799.

66
Sábado, T. J. 2009. Fundamentos de bioestadística y análisis de datos
para enfermería. Servei de Publicacions. Universidad Autónoma de
Barcelona. España. 147pp.
Sánchez y A. Gándara. 2011. Conceptos básicos de gestión ambiental y
desarrollo sustentable. SEMARNAT. México. 333pp.
Seminoff, J. A., A. Resendiz., B. Resendiz y J. Nichols. 2004. Ocurrence of
loggerhead sea turtles (Caretta caretta) in the Gulf of California, México:
evidence of life-history variation in the Pacific Ocean. Herpetological
review. 35 (1):24-27.
Shigeto, A. J., Mangel, C.J., Seminoff y P. H. Dutton. 2008. Demography of
loggerhead turtles Caretta caretta in the southeastern Pacific Ocean:
fisheries-based observations and implications for management.
Endangered Species Research. 5:129-135.
Scheiner, M. S y Gurevitch, J. 2001. Design and Analysis of Ecological
Experiments, second division. Oxford University Press. E.U.A. 432pp.
Silman, R., Vargas. I y Troëng. S. 2002. Tortugas Marinas: Guía Educativa.
Corporación Caribeña para la Conservación. 38 pp.
Smith, Josep W. 1999. Distribution of Atlantic menhaden, Brevoortia
tyrannus, purse-seine sets and catches from southern New England to
North Carolina, 1985-96. U.S Dep. Commer. NOAA Technical Report. 22 pp.
Soto, S. J. J y Aguirre, C. J.L. 1996. Trophic comparison amog Triglidae
(Pisces: Scorpaeniformes) off Baja California Sur, México. Revista de
Biología Tropical. 44 (2):803-811.
Spotila, R. J. 2004. Sea Turtles: A complete guide to their biology,
behavior and conservation. JHU Press. 240 pp.

67
Sutton Glen y Wagner Tom. 2007. Stock assessment of blue crab
(Callinectes sapidus) in Texas coastal waters. Management Data Series
No.249. 45pp.
Tagliafico. A., González. L. W y Eslava. N. 2008. Estimación de los
parámetros de crecimiento y reproducción de la sardina (Sardinella
aurita) del sureste de la isla de margarita, Venezuela. Boletín de
Investigaciones Marinas y Costeras. 37 (2):45-54.
Trespalacios. G., J. A., Bello, A. L y Vázquez. C. R. 2005. Investigación de
mercados: métodos de recogida y análisis de la información para la toma
de decisiones en marketing. Paraninfo. España. 353 pp.
Urroz Carlos. 1991. Elementos de Anatomía y Fisiología Animal. EUNED.
268 pp.
Valera, N. J. P. A. 2005. Apuntes de Física General. Universidad Nacional
Autónoma de México. México. 381pp.
Vanhooydonck, B., Herrel. A y R. Van Damme. 2007. Interactions between
habitat use, behavior, and the trophic niche of lacertids lizards. Pp 427-
449. En: Aldape. L. C. T., Lazcano. H. E. y Martínez. C. M. (Edits).
Composición de la dieta de Aspidoscelis sacki de Zaachila, Oaxaca.
Boletín de la Sociedad Herpetológica Mexicana. 17(2):73-887.
Wainwright, P. C. 1996. Ecological explanation through functional
morphology: The feeding biology of sunfishes. Ecology. 77 (5):1336-1343.
Walls, A. E., Berkson, J y Smith, A. S. 2002. The Horseshoe Crab, Limulus
polyphemus: 200 million years of existence, 100 years of study. Fisheries
Science. 10(1):39-73.
Wyneken, J. 2004. La anatomía de las tortugas marinas U.S. deparment of
commerce NOAA technical memorándum.NMFS-SEFSC-470. 172 pp.
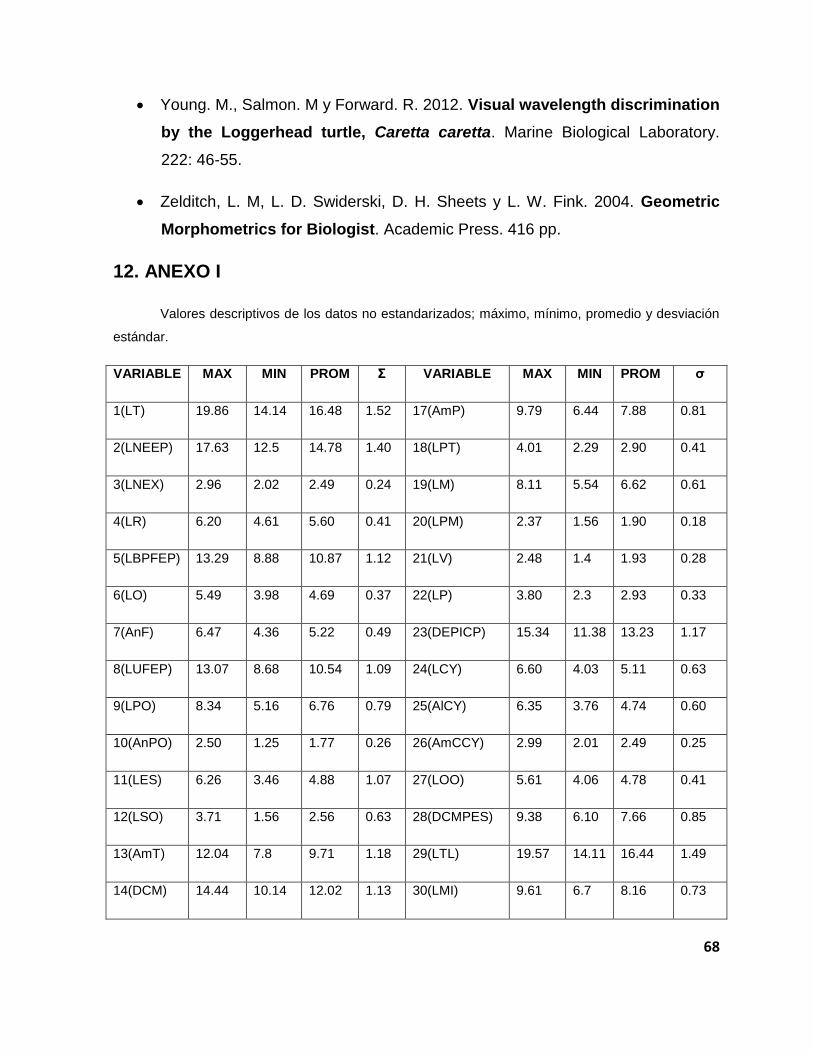
68
Young. M., Salmon. M y Forward. R. 2012. Visual wavelength discrimination
by the Loggerhead turtle, Caretta caretta. Marine Biological Laboratory.
222: 46-55.
Zelditch, L. M, L. D. Swiderski, D. H. Sheets y L. W. Fink. 2004. Geometric
Morphometrics for Biologist. Academic Press. 416 pp.
12. ANEXO I
Valores descriptivos de los datos no estandarizados; máximo, mínimo, promedio y desviación
estándar.
VARIABLE MAX MIN PROM Σ VARIABLE MAX MIN PROM σ
1(LT) 19.86 14.14 16.48 1.52 17(AmP) 9.79 6.44 7.88 0.81
2(LNEEP) 17.63 12.5 14.78 1.40 18(LPT) 4.01 2.29 2.90 0.41
3(LNEX) 2.96 2.02 2.49 0.24 19(LM) 8.11 5.54 6.62 0.61
4(LR) 6.20 4.61 5.60 0.41 20(LPM) 2.37 1.56 1.90 0.18
5(LBPFEP) 13.29 8.88 10.87 1.12 21(LV) 2.48 1.4 1.93 0.28
6(LO) 5.49 3.98 4.69 0.37 22(LP) 3.80 2.3 2.93 0.33
7(AnF) 6.47 4.36 5.22 0.49 23(DEPICP) 15.34 11.38 13.23 1.17
8(LUFEP) 13.07 8.68 10.54 1.09 24(LCY) 6.60 4.03 5.11 0.63
9(LPO) 8.34 5.16 6.76 0.79 25(AlCY) 6.35 3.76 4.74 0.60
10(AnPO) 2.50 1.25 1.77 0.26 26(AmCCY) 2.99 2.01 2.49 0.25
11(LES) 6.26 3.46 4.88 1.07 27(LOO) 5.61 4.06 4.78 0.41
12(LSO) 3.71 1.56 2.56 0.63 28(DCMPES) 9.38 6.10 7.66 0.85
13(AmT) 12.04 7.8 9.71 1.18 29(LTL) 19.57 14.11 16.44 1.49
14(DCM) 14.44 10.14 12.02 1.13 30(LMI) 9.61 6.7 8.16 0.73

69
15(AmFI) 4.60 2.97 3.66 0.45 31(LA) 6.55 3.97 5.12 0.60
16(LFI) 6.40 4.43 5.41 0.55 32(LD) 4.48 3.1 3.77 0.41
13. ANEXO II
Histogramas de frecuencia para las medidas lineales tomadas de los cráneos.

70
Figura.6.Histograma de la variable 2. (LNEEP). Figura.7.Histograma de la variable 3. (LNEX).
Figura.8.Histograma de la variable 4. (LR). Figura.9.Histograma de la variable 5. (LBPFEP).

71
Figura.10.Histograma de la variable 6. (LO). Figura.11.Histograma de la variable 7. (AnF).

72
Figura.12.Histograma de la variable 8. (LUFEP). Figura.13.Histograma de la variable 9. (LPO).

73
Figura.14.Histograma de la variable 10. (AnPO). Figura.15.Histograma de la variable 11. (LES).
Figura.16.Histograma de la variable 12. (LSO). Figura.17.Histograma de la variable 13. (AmT).

74
Figura.18.Histograma de la variable 14. (DCM). Figura.19.Histograma de la variable 15. (AmFI).
Figura.20.Histograma de la variable 16. (LFI). Figura.21.Histograma de la variable 17. (AmP).

75
Figura.22.Histograma de la variable 18. (LPT). Figura.23.Histograma de la variable 19. (LM).

76
Figura.24.Histograma de la variable 20. (LPM). Figura.25.Histograma de la variable 21. (LV).

77
Figura.26.Histograma de la variable 22. (LP). Figura.27.Histograma de la variable 23. (DEPIC).
Figura.28.Histograma de la variable 24. (LCY). Figura.29.Histograma de la variable 25. (AlCY).
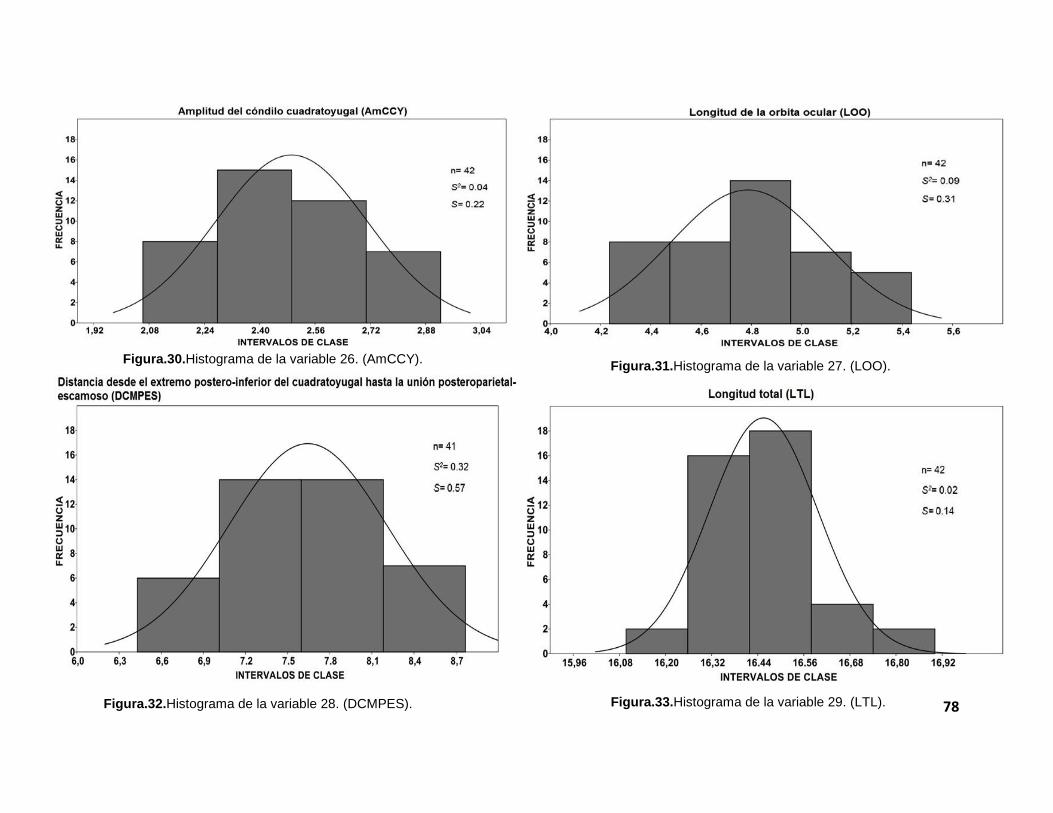
78
Figura.30.Histograma de la variable 26. (AmCCY). Figura.31.Histograma de la variable 27. (LOO).
Figura.32.Histograma de la variable 28. (DCMPES). Figura.33.Histograma de la variable 29. (LTL).
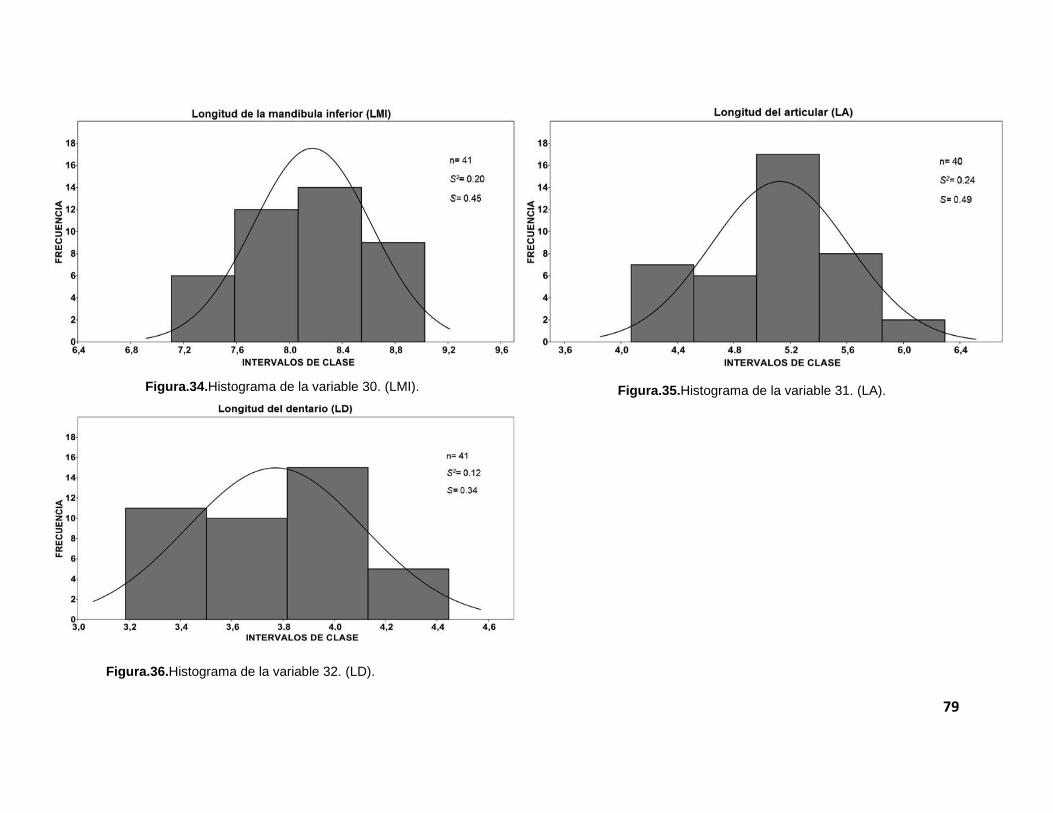
79
Figura.34.Histograma de la variable 30. (LMI). Figura.35.Histograma de la variable 31. (LA).
Figura.36.Histograma de la variable 32. (LD).

80
14. ANEXO III
Valores de normalidad de las 32 medidas craneales.
VARIABLE p-valor VARIABLE p-valor
1.-Longitud total (LT). 0.0015 17.-Amplitud del pico (AmP). 0.0027
2.-Longitud desde el borde
posterior de las narinas hasta el
extremo posterior (LNEEP).
0.0051 18.-Longitud del pterigoides
(LPT).
0.0022
3.-Longitud de las narinas
externas (LNEX).
0.0073 19.-Longitud del maxilar (LM). 0.0030
4.-Longitud del rostro (LR). 0.0028 20.-Longitud del premaxilar
(LPM).
0.0048
5.-Longitud desde el borde
posterior del frontal hasta el
extremo posterior (LBPFEP).
0.0030 21.-Longitud de vómer (LV). 0.0023
6.-Longitud de la órbita (LO). 0.0022 22.-Longitud del palatino (LP). 0.0021
7.-Ancho del frontal (AnF). 0.0037 23.-Distancia desde el extremo
postero-inferior del cuadrado
hasta el extremo anterior del
pico (DEPICP).
0.0019
8.-Longitud desde la unión
frontal/postorbital parietal hasta
el extremo posterior (LUFEP).
0.0028 24.-Longitud del cuadrato-
yugal (LCY).
0.0017
9.-Longitud del postorbital
(LPO).
0.0030 25.-Altura del cuadrato-yugal
(AlCY).
0.0034
10.-Ancho del postorbital
(AnPO).
0.0029 26.-Amplitud del cóndilo
cuadrato-yugal (AmCCY).
0.0030
11.-Longitud del escamoso 0.0029 27.-Longitud del orbita ocular 0.0028

81
(LES). (LOO).
12.-Longitud del supraoccipital
(LSO).
0.0056 28.-Distancia desde el extremo
postero-inferior del cuadrato-
yugal, hasta la unión
posteroparietal-escamoso
(DCMPES).
0.0021
13.-Amplitud total (AmT). 0.0015 29.-Longitud total (LTL). 0.0070
14.-Distancia entre cóndilos
mandibulares (DCM).
0.0019 30.-Longitud del la mandíbula
inferior (LMI).
0.0033
15.-Amplitud de la fosa
infratemporal (AmFI).
0.0011 31.-Longitud del articular (LA). 0.0050
16.-Longitud de la fosa
infratemporal (LFI).
0.0024 32.-Longitud del dentario (LD). 0.0018