Embed Size (px)
Citation preview

UNIVERSIDAD AUTÓNOMA METROPOLITANA
UNIDAD IZTAPALAPA
DIVISIÓN DE CIENCIAS BIOLOGICAS Y DE LA SALUD
HIDROBIOLOGIA
BIORREMEDIACION DE AGUA CONTAMINADA CON PARATIONY COBRE A TRAVES DE TRES ORGANISMOS ACUATICOS
Roberto de Jesús González Ortiz
Asesores: M en C. Guadalupe Barrera Escorcia
Dra. Laura Martínez Tabche
México, 2003.

DATOS PERSONALES
Nombre: Roberto De Jesús González Ortíz.Matrícula: 93329038.Teléfono: (5)774-24-96.Licenciatura: Hidrobiología.División: Ciencias Biológicas y de la Salud.Unidad: Iztapalapa.
TRIMESTRE: 04-P.
TITULO DEL PROYECTO DE INVESTIGACIÓN: Biorremediación de agua contaminada con paratión y cobre a
través de tres organismos acuáticos.
TITULO DEL TRABAJO DE SERVICIO SOCIAL: Evaluación de la toxicidad por paratión en Hyalella azteca.
NOMBRE DEL ASESOR INTERNO: M. en C. Guadalupe Barrera Escorcia.
NOMBRE DEL ASESOR EXTERNO: Dra. Laura Martínez Tabche.
LUGAR DE REALIZACIÓN: Escuela Nacional de Ciencias Biológicas ( ENCB-IPN ), Departamento de Farmacia, Lab. de Toxicología Acuática.
CLAVE DE REGISTRO: H.18.01
FECHA DE INICIO: 9 noviembre 2001.
FECHA DE TERMINACIÓN: 9 noviembre 2003.

Nombre: Roberto De Jesús González Ortíz.Matrícula: 93329038.Licenciatura: Hidrobiología.
Titulo del proyecto de investigación: EVALUACIÓN DE LA TOXICIDAD POR PARATIÓN EN Hyalella azteca.Clave de registro: : H.018.01.Fecha de registro: 9 noviembre 2001.Fecha de entrega: 9 noviembre 2003.
Nombre y Adscripción de los Asesores:
-Asesor interno: M. en C. Guadalupe Barrera Escorcia, Profr. Titular “C”, T. C., Lab. de Contaminación y Bioensayos, Departamento de Hidrobiología, UAM-Iztapalapa.
-Asesor externo: Dra. Laura Martínez Tabche, Profr. Titular "C", T. C. Jefe del Lab. Toxicología Acuática, Departamento de Farmacia, ENCP-IPN.
El metil-paratión ( MP ) es uno de los plaguicidas organofosforados más ampliamenteutilizados en la agricultura; los cambios en la actividad de los sistemas encargados desu biotransformación pueden modificar la toxicidad inicial producida por esteplaguicida; por ello, para este trabajo se decidió evaluar los efectos agudo y subcrónico( 5 días exposición ) del MP sobre el anfípodo Hyalella azteca, bajo condiciones deinducción e inhibición del metabolismo ( tratamiento con fenobarbital ( PhB ) ycimetidina ( Ci ), respectivamente ). Los biomarcadores utilizados fueron el grado delipoperoxidación ( producción de malondialdehído ó MAD) y la actividad de lasenzimas γ-glutamil transpeptidasa ( GGT ) y etoxiresorufin-O-deetilasa ( EROD ). Losresultados obtenidos fueron los siguientes: CE50 48h MP con 100 ppm Ci ( 40 ppb ) >>CE50 48h MP con 100 ppm PhB ( 1 pbb ) > CE50 48h MP ( 0.17 ppb ). En condiciones deestimulación del metabolismo ( 0.0195 ppb MP con 100 ppm PhB ) se observó unincremento en el efecto lipoperoxidante e inhibición de la GGT y EROD, los cualespodrían ser causados por un aumento en la producción del metil-paraoxón; sinembargo, los valores más altos para estos biomarcadores ( MAD, GGT y EROD ) sepresentaron en el tratamiento inhibidor del metabolismo ( 4 ppb MP con 100 ppm Ci )superando en varios órdenes de magnitud a los controles; en este caso, la toxicidadpodría no ser atribuida a los metabolitos metil-paraoxón ni p-nitrofenol, sinoposiblemente a un intermediario inestable ( fosfooxitiirano ) formado durante labiotransformación de metil-paratión a metil-paraoxón. Los resultados obtenidosmuestran que bajo las condiciones evaluadas ( inducción e inhibición delmetabolismo ) la toxicidad inicial del MP puede ser modificada en forma notable tanto anivel letal como subcrónico.

Alumno: Roberto de Jesús González Ortíz
Asesor interno
Vo. Bo.M. en C. Guagalupe Barrera Escorcia
Asesor externo
Vo. Bo.Dra. Laura Martínez-Tabche


INTRODUCCIÓN.
Generalidades sobre Hyalella azteca.
Hyalella azteca ( Saussure, 1858 ) es un anfípodo epibentónico detritívoro, que seencuentra distribuido de manera permanente en lagos y estanques desdeNorteamérica hasta Sudamérica ( Pennak, 1989 ). Para obtener su alimento, escarba enla superficie del sedimento, ingiriendo preferentemente algas y bacterias ( Hargrave,1970 ). Por su comportamiento y hábitos alimenticios, se considera un excelenteorganismo de prueba para realizar bioensayos con sedimentos ( U. S. EPA, 2000 ); sinembargo, también se han descrito métodos para evaluar la toxicidad de efluentes yxenobióticos sobre los ambientes acuáticos utilizando esta especie ( APHA, 1989; EPA,1991 ).
Los criterios más importantes para considerar a Hyalella azteca como organismo debiomonitoreo son: 1) existe una base de datos toxicológica sobre su relativa sensibilidad( como p. ej., sobrevivencia, crecimiento y reproducción ) a un amplio rango desubstancias de interés, tanto por separado como en mezclas complejas y 2) es fácil decultivar en el laboratorio ( U. S. EPA, 2000 ).
Además, hay antecedentes de estudios en los cuales se ha utilizado a este organismopara evaluar la toxicidad de diversos plaguicidas en la columna de agua, tales comop,p’-DDT, p,p’-DDD, p,p’-DDE, dieldrin y clorpirifos ( Phipps et al., 1995 ).
Por lo anterior, se decidió utilizar a Hyalella azteca como organismo de prueba paraevaluar la toxicidad del metil-paratión en la columna de agua, tanto a nivel letal comosubletal.
Metil-paratión.
El metil-paratión ( O,O-dimetil-O-p-nitrofenil fosforotionato ) es un plaguicidaorganofosforado; se usa más o menos tan ampliamente como el paratión y es muysimilar en sus propiedades físicas y químicas; no obstante, se hidroliza e isomeriza másfácilmente que éste, por lo que es menos estable como insecticida y ligeramente menostóxico para los mamíferos; se usa principalmente en los cultivos de café ycítricos ( Hassall, 1990 ) contra pulgones y catarinitas; se encuentra disponible enfórmulas que contienen desde menos de 1% hasta más del 95% de material puro( Dreisbach y Robertson, 1988 ).
Límites de exposición.
En la legislación mexicana, no se especifica un límite máximo permisible ( LMP ) paraeste plaguicida en cuerpos de agua ( NOM-001-SEMARNAT-1996, Diario Oficial de laFederación del 6 enero 1997) ó agua potable ( NOM-041-SSA1-1993, Diario Oficial de laFederación del 24 marzo 1995 y NOM-127-SSA1-1994, Diario Oficial de la Federacióndel 18 enero 1996 ), aunque se reconoce como residuo peligroso tóxico para el ambiente( NOM-052-SEMARNAT-1993, Diario Oficial de la Federación del 22 octubre 1993 ).
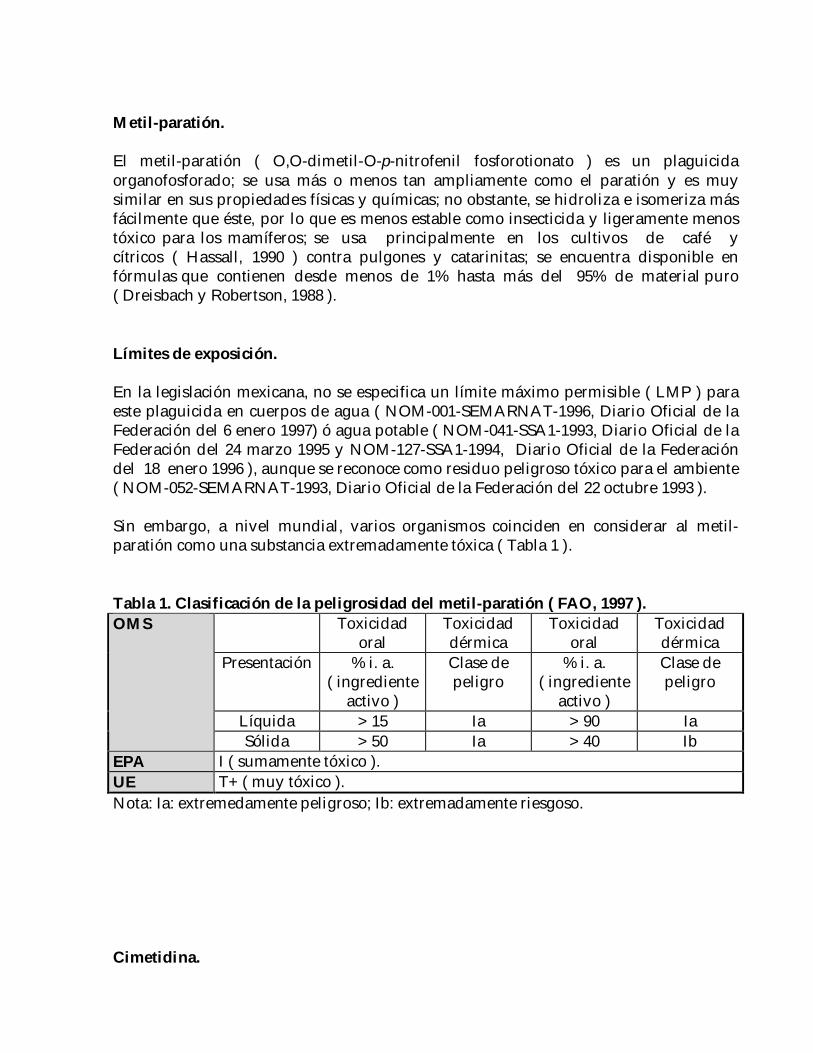
Sin embargo, a nivel mundial, varios organismos coinciden en considerar al metil-paratión como una substancia extremadamente tóxica ( Tabla 1 ).
Tabla 1. Clasificación de la peligrosidad del metil-paratión ( FAO, 1997 ).Toxicidad
oralToxicidaddérmica
Toxicidadoral
Toxicidaddérmica
Presentación % i. a.( ingrediente
activo )
Clase depeligro
% i. a.( ingrediente
activo )
Clase depeligro
Líquida > 15 Ia > 90 Ia
OMS
Sólida > 50 Ia > 40 IbEPA I ( sumamente tóxico ).UE T+ ( muy tóxico ).Nota: Ia: extremedamente peligroso; Ib: extremadamente riesgoso.
Cimetidina.

La cimetidina es un fármaco antagonista de la histamina a nivel de los receptores H2; seusa en el tratamiento de la úlcera péptica y en otros padecimientos en los cuales laacidez gástrica debe ser disminuida; interfiere con la biotransformación de losxenobióticos, al inhibir las oxidaciones mediadas a través de los CYP´shepáticos ( Alvares y Pratt, 1990; Levine et al., 1998 ); se une al Fe del grupo hemo delCYP450 a través de los grupos imidazol y ciano que posee en su estructura ( Fig. 1 )( Rendic et al., 1983 ).
Cimetidina. Fenobarbital sódico.
Fig 1. Estructura química de los fármacos utilizados en este estudio (Merck, 1996 ).
Fenobarbital sódico.
La acción de este fármaco ( Fig. 1 ) induce las enzimas microsomales y aumenta lafunción hepática, modificando de esta manera el metabolismo de diversassubstancias; actúa principalmente sobre la familia de CYP´s IIB ( Tukey y Johnson,1990; Katzung, 1994 ).
Algunos aspectos relevantes de las substancias mencionadas se muestran a continuación( Tablas 2 y 3 ):
Tabla 2. Identificación química de los compuestos empleados en este estudio( Merck, 1996 ).Características Metil-
paratiónMetil-paraoxón Fenobarbital Cimetidina
No.Registro CAS 000298-00-0 000950-35-6 000050-06-6 051481-61-9Nombre O,O-dimetil-
O-p-nitrofenilfosforotionato
O,O-dimetil-O-p-nitrofenilfosfato
Peso molecular 263.21 247.15 232.24 252.34Fórmulamolecular
C8H10NO5PS C8H10NO6P C12H12N2O3 C10H16N6S
Tabla 3. Propiedades físicas y químicas de los compuestos empleados en este estudio( Merck, 1996 ) .
N
N
SNN
NCN
H3CH3C
N O
O
N
NaO
H3C
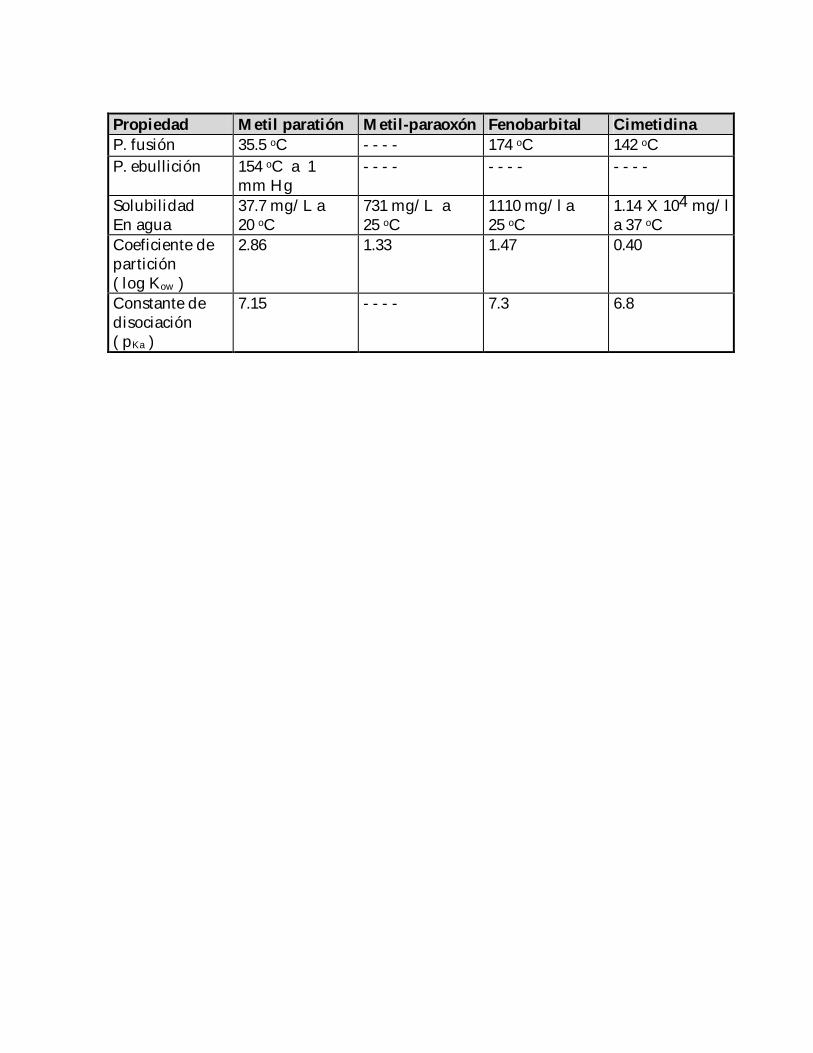
Propiedad Metil paratión Metil-paraoxón Fenobarbital CimetidinaP. fusión 35.5 oC - - - - 174 oC 142 oCP. ebullición 154 oC a 1
mm Hg- - - - - - - - - - - -
SolubilidadEn agua
37.7 mg/L a20 oC
731 mg/L a25 oC
1110 mg/l a25 oC
1.14 X 104 mg/la 37 oC
Coeficiente departición( log Kow )
2.86 1.33 1.47 0.40
Constante dedisociación( pKa )
7.15 - - - - 7.3 6.8

Lipoperoxidación celular y conjugación con glutatión.
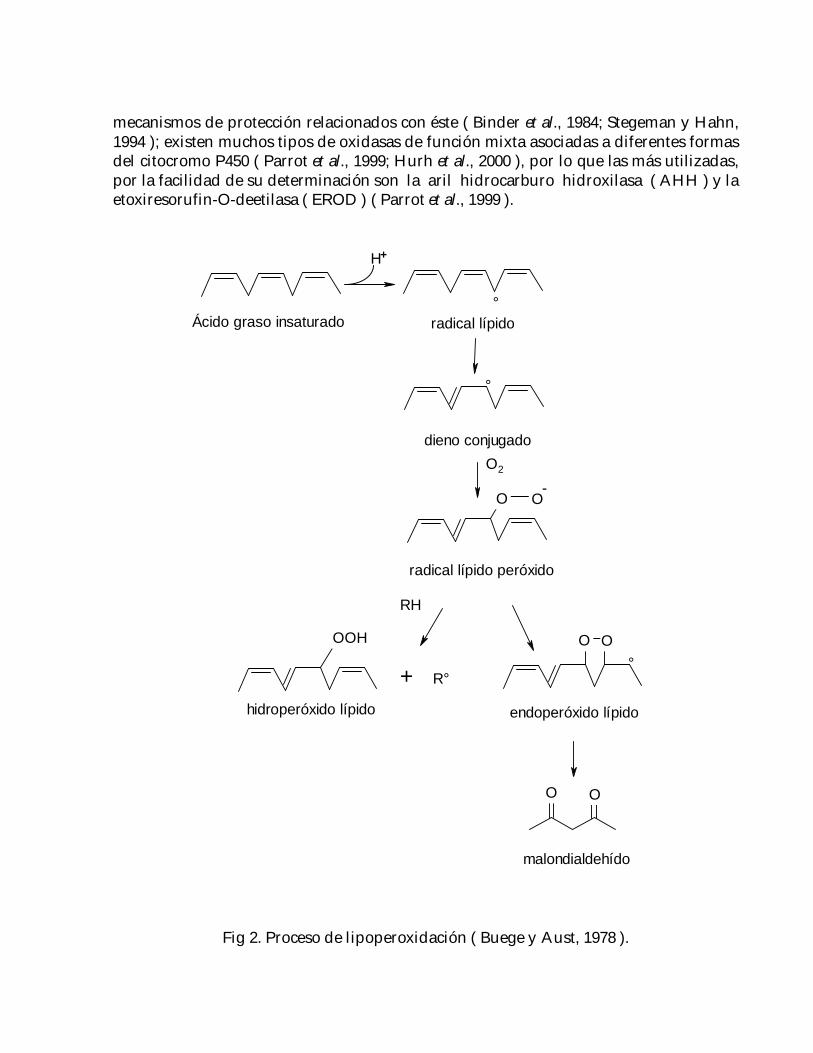
La lipoperoxidación o peroxidación de compuestos lipídicos ( Fig. 2 ) ha sido sugeridacomo uno de los mecanismos moleculares involucrados en la toxicidad inducida porplaguicidas ( Khrer, 1993; Yarsan et al.,1999 ), el cual es un proceso degenerativo queactúa sobre los ácidos grasos poliinsaturados de las membranas celulares, y cuyaconsecuencia más importante es la destrucción paulatina de éstas ( Buege y Aust, 1978;Khrer, 1993 ); dicha ruptura genera una variedad de compuestos ( alcoholes, cetonas,aldehídos y éteres ) entre los que se encuentra el malondialdehído ( MAD ) ( Buegey Aust, 1978; Yarsan et al., 1999 ); existen mecanismos moleculares ( las vitaminas A, C yE, ácido úrico y ceruloplasmina ) y enzimáticos para prevenir este proceso, talescomo la superóxido-dismutasa ( SOD ), catalasa ( CAT ) y las enzimas dependientes delglutatión ( GSH ): γ-glutamil transpeptidasa ( GGT ), glutatión-S-transferasa ( GST ),glutatión-peroxidasa ( GPx ) y glutatión reductasa ( GR ) ( Draper, 1990; Dikshit, 1991;Comporti, 1993; Khrer, 1993; Koner et al., 1998; Yarsan et al., 1999 ).
La γ-glutamil transpeptidasa ( GGT ) es una enzima de membrana, con su sitio activoorientado hacia el exterior de la célula; participa en la degradación de los conjugadosextracelulares del glutatión al separar las moléculas de cisteinilglicina y glutamil ( Fig.3 ), las cuales, de este modo, vuelven a estar disponibles para su transporte a través dela membrana hacia el citoplasma, donde son utilizadas para la resíntesis delglutatión ( Tate y Meister, 1985 ); además, se ha visto que el aumento en los niveles deesta enzima incrementa la resistencia de las células ante substancias tóxicas yestrés oxidativo ( Paolicchi et al., 1997 ).
Ya que tanto el malondialdehído ( MAD ) como la γ- gutamiltranspeptidasa ( GGT ) están relacionados con el proceso de lipoperoxidación, para estetrabajo se escogieron como biomarcadores de este proceso en los organismos expuestos.
Sistema de oxidasas de función mixta ( MFO ).
En los animales, la biotransformación de metil-paratión hacia metil-paraoxón ( Fig. 4 )se realiza a través del sistema enzimático MFO ( oxidasas de función mixta, porsus siglas en inglés ) ( Kamataki y Neal, 1976; Guengerich, 1977; Halpert et al., 1980;Butler y Murray, 1993 ) el cual agrupa a una familia de enzimas inducibles, cuyafunción es ayudar a la excreción de compuestos no polares ( Parrot et al., 1999 ); pararealizar dicha función requieren de oxígeno, un agente reductor ( p. ej. NADP ) y untransporte facilitado de electrones a través del citocromo P450 ( CYP ) ( Martin et al.,1985); la actividad de estas enzimas puede ser alterada por diversos fármacos,estimulándola ( p. ej. en el caso de barbitúricos como el fenobarbital) ó inhibiéndola ( p.ej. por la cimetidina ) ( Mourelle et al., 1986 ), modificando de este modo la toxicidad delcompuesto original, aumentando la producción de metabolitos más tóxicos ( como elmetil-paraoxón, en el caso del metil-paratión ) ó el grado de desintoxicación y los
mecanismos de protección relacionados con éste ( Binder et al., 1984; Stegeman y Hahn,1994 ); existen muchos tipos de oxidasas de función mixta asociadas a diferentes formasdel citocromo P450 ( Parrot et al., 1999; Hurh et al., 2000 ), por lo que las más utilizadas,por la facilidad de su determinación son la aril hidrocarburo hidroxilasa ( AHH ) y laetoxiresorufin-O-deetilasa ( EROD ) ( Parrot et al., 1999 ).
Fig 2. Proceso de lipoperoxidación ( Buege y Aust, 1978 ).
O O
°
°
H+
O2
OO
+ R°
RH
°
O O
-
Ácido graso insaturado radical lípido
dieno conjugado
hidroperóxido lípido endoperóxido lípido
radical lípido peróxido
malondialdehído
OOH
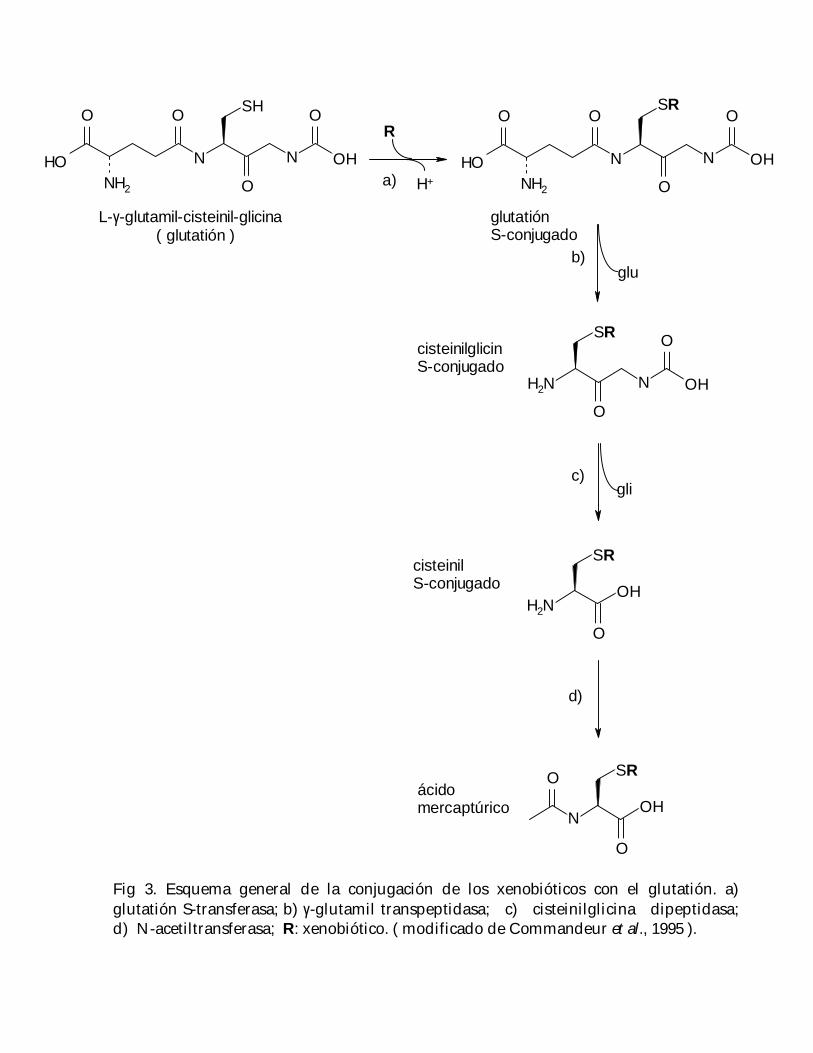
Fig 3. Esquema general de la conjugación de los xenobióticos con el glutatión. a)glutatión S-transferasa; b) γ-glutamil transpeptidasa; c) cisteinilglicina dipeptidasa;d) N-acetiltransferasa; R: xenobiótico. ( modificado de Commandeur et al., 1995 ).
O O
O
O
NNNH2
OHHO
SH O O
O
O
NNNH2
OHHO
SR
a)
O
O
NH2N OH
SR
b)glu
c)gli
O
H2N
SR
OH
cisteinilglicinS-conjugado
glutatiónS-conjugado
L-γ-glutamil-cisteinil-glicina ( glutatión )
R
H+
cisteinilS-conjugado
O
SR
OHN
Oácidomercaptúrico
d)
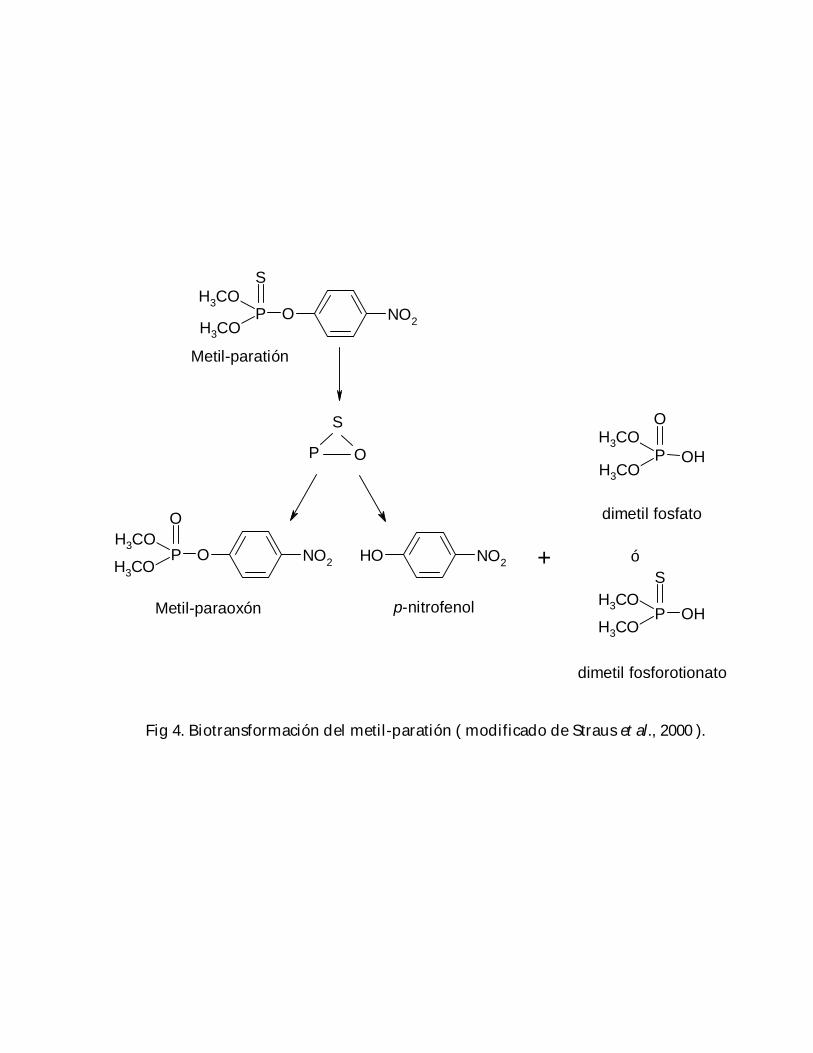
Fig 4. Biotransformación del metil-paratión ( modificado de Straus et al., 2000 ).
P O NO2
S
S
P O
P O NO2
O
OH NO2
O
P OH
S
P OH
Metil-paratión
Metil-paraoxón p-nitrofenol
dimetil fosfato
dimetil fosforotionato
+ ó
H3CO
H3CO
H3CO
H3CO
H3CO
H3CO
H3CO
H3CO

Por esta razón, se determinó emplear a la etoxiresorufin-O-deetilasa ( EROD ) comoindicador de la actividad del CYP450 en presencia de los fármacos fenobarbital ycimetidina.

Uso de programas de cómputo en toxicología acuática.
Para realizar la parte experimental de bioensayos de toxicidad se requiere conocer demanera rutinaria los valores de CE50 48h y CE0 48h de diversas substancias, queincluyen los xenobióticos a probar y los solventes empleados en los bioensayos. Aunqueexisten en el mercado varios programas comerciales para calcular la CE50, su uso esrestringido, debido a que no son fáciles de conseguir o lo son a un precio muy alto ó nose encuentran disponibles en español, de manera que el usuario raras veces tiene formade conocer si el resultado final proporcionado por el programa es el correcto, ya quedesconoce tanto la parte de programación empleada para crear el programa, como elfundamento del método usado para calcular la CE50, lo cual puede afectarnegativamente el desempeño de su trabajo.
Por estos motivos se consideró necesario elaborar un programa de cómputo quecalculara dichos valores a través del método probit, que pudiera ser de aplicacióndirecta en los resultados de los bioensayos y de utilidad para futuras determinaciones .

OBJETIVO GENERAL
Evaluar la toxicidad del metil-paratión en condiciones de estimulación ( tratamiento confenobarbital ) e inhibición del metabolismo ( tratamiento con cimetidina ), utilizandocomo organismo de prueba Hyalella azteca.
OBJETIVOS ESPECÍFICOS
1. Determinar la toxicidad aguda del metil-paratión ( CE50 48h) y de losfármacos fenobarbital y cimetidina.
2. Determinar la toxicidad aguda del metil-paratión ( CE50 48h ) en presencia de lossiguientes tratamientos:
a) un inhibidor del metabolismo ( cimetidina ). b) un estimulador del metabolismo ( fenobarbital ).
3. Determinar la toxicidad subletal del metil-paratión ( CE0 48h ) en los tratamientosarriba mencionados sobre la lipoperoxidación y el sistema de oxidasas de funciónmixta ( MFO ), utilizando como indicadores los niveles de malondialdehído
( MAD) y γ-glutamil transpeptidasa ( GGT ) en el primer caso, y etoxiresorufin-O- deetilasa ( EROD ) en el segundo.
4. Elaborar un programa para calcular la CE50 y su intervalo de confianza, utilizando el lenguaje de programación Turbo Pascal para DOS V. 6.0
METODOLOGÍA
Evaluación de la toxicidad aguda.
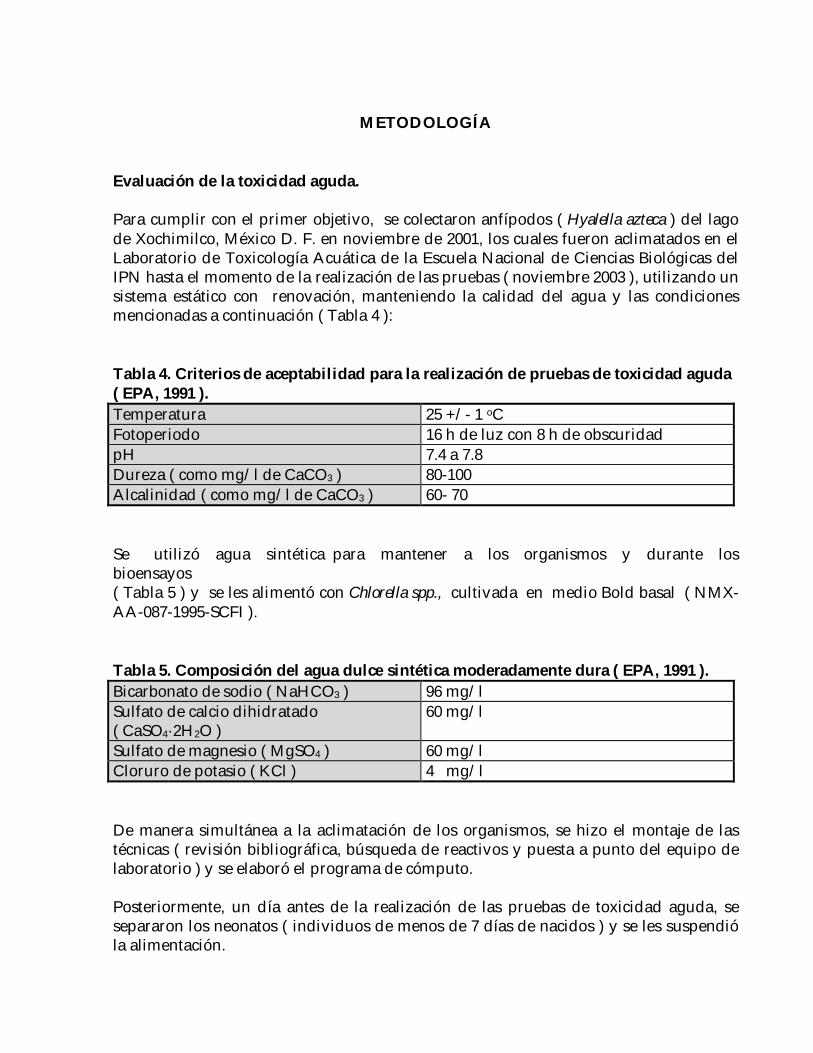
Para cumplir con el primer objetivo, se colectaron anfípodos ( Hyalella azteca ) del lagode Xochimilco, México D. F. en noviembre de 2001, los cuales fueron aclimatados en elLaboratorio de Toxicología Acuática de la Escuela Nacional de Ciencias Biológicas delIPN hasta el momento de la realización de las pruebas ( noviembre 2003 ), utilizando unsistema estático con renovación, manteniendo la calidad del agua y las condicionesmencionadas a continuación ( Tabla 4 ):
Tabla 4. Criterios de aceptabilidad para la realización de pruebas de toxicidad aguda( EPA, 1991 ).Temperatura 25 +/- 1 oCFotoperiodo 16 h de luz con 8 h de obscuridadpH 7.4 a 7.8Dureza ( como mg/l de CaCO3 ) 80-100Alcalinidad ( como mg/l de CaCO3 ) 60- 70
Se utilizó agua sintética para mantener a los organismos y durante losbioensayos( Tabla 5 ) y se les alimentó con Chlorella spp., cultivada en medio Bold basal ( NMX-AA-087-1995-SCFI ).
Tabla 5. Composición del agua dulce sintética moderadamente dura ( EPA, 1991 ).Bicarbonato de sodio ( NaHCO3 ) 96 mg/lSulfato de calcio dihidratado( CaSO4·2H2O )
60 mg/l
Sulfato de magnesio ( MgSO4 ) 60 mg/lCloruro de potasio ( KCl ) 4 mg/l
De manera simultánea a la aclimatación de los organismos, se hizo el montaje de lastécnicas ( revisión bibliográfica, búsqueda de reactivos y puesta a punto del equipo delaboratorio ) y se elaboró el programa de cómputo.
Posteriormente, un día antes de la realización de las pruebas de toxicidad aguda, sesepararon los neonatos ( individuos de menos de 7 días de nacidos ) y se les suspendióla alimentación.

Una vez transcurrido este tiempo, con estos organismos se evaluó la CE50 48h de lassiguientes substancias: metil-paratión ( disuelto en acetona al 0.6% en etanol al 0.4% ),fenobarbital ( disuelto en NaCl al 0.9% ) ( Ohhira et al., 2000 ) y cimetidina ( disueltaen etanol al 0.4% ), utilizando 6 lotes de 20 organismos cada uno ( 5 concentracionesmás un control: 0.01, 0.1, 1, 10, 100 y 0 ppm ) por cada tratamiento y empleandoagua sintética como agua de dilución.
Todas las evaluaciones agudas se realizaron por triplicado y durante el transcurso deéstas, no se suministró alimento a los organismos ni se efectuaron recambios en el aguade tratamiento ( EPA, 1991 ). Para determinar el efecto de estas substancias sobre losorganismos, se usó el criterio de inmovilidad; los valores de CE50 48h fueron calculadosa través del método probit utilizando el programa de cómputo elaboradopreviamente ( CL50 ); si la mortalidad del lote control fue mayor del 10% ó lacorrelación con el método resultó menor a 0.9, se repitió la prueba.
Evaluación de la toxicidad subletal.
Para este fin, se obtuvieron los valores de CE0 48h del metil-paratión con fenobarbital ymetil-paratión con cimetidina por interpolación en la curvas utilizadas para calcular laCE50 48h del metil-paratión con éstos fármacos. A continuación, se expuso a losorganismos a estas concentraciones ( CE0 48h ) durante 5 días ( modificado de Kubitz yGiesy, 1996 ).
Toma de muestras.
Una vez transcurrido el tiempo de exposición, los organismos de cada lote fueronpesados y homogeneizados en búffer NaCl 0.15 M en baño de hielo, y luegocentrifugados, primero a 14 000 rpm durante 15 min ( modificado de Scheuhammer yCherian, 1986 ) y después a 3 500 rpm durante 5 min, para precipitar los núcleos, lasmitocondrias, las proteínas y los fragmentos celulares ( Aïche et al., 1983 ); elsobrenadante obtenido ( fracción S9 ó postmitocondrial ), la cual contiene al citosol y losmicrosomas ( Parrot et al., 1999 ) se utilizó para las determinaciones, por lo que fueseparado, etiquetado, protegido de la luz y mantenido en refrigeración a 4 oC hasta suuso.
Todas las determinaciones fueron referidas respecto a la cantidad de proteína presente,la cual se realizó por el método de Bradford ( 1976 ).

Determinación de lipoperoxidación ( Buege y Aust, 1978 ).
Se cuantificó la concentración de malondialdehído ( MDA ) con el ácido tiobarbitúrico( TBA ), que da como resultado un compuesto de color rosa-anaranjado, el cual esevaluado a 535 nm.
Determinación de la γ-glutamil transpeptidasa ( GGT )( Huseby y Strome, 1974 ).
Esta enzima, cataliza la conversión de γ-glutamil-p-nitroanilina a p-nitroanilina enpresencia de glicil-glicina; la determinación de GGT se realizó evaluando a 407 nm lacantidad de p-nitroanilina que es liberada en la reacción anterior.
Determinación de la etoxiresorufin-O-deetilasa ( EROD ) ( Parrot et al., 1999 ) .
En esta técnica, se hace reaccionar 7-etoxiresorufin en presencia de NADPH; laactividad de la EROD es proporcional a la cantidad de resorufina que es producida enesta reacción, la cual es evaluada en un fluorómetro a 520 nm.
Control de calidad.
Todas las muestras se analizaron por triplicado.
Se evaluó la precisión y exactitud de las técnicas empleadas.
La precisión de las determinaciones, se obtuvo a partir del coeficiente de variación, elcual se expresa como:
C. V. = (desviación estándar X 100)/ media
En las muestras con coeficientes de variación mayores al 10%, se repitió el análisis.
La exactitud se evaluó con base en el porcentaje de recuperación de estándares:
% REC= ( valor obtenido x 100 )/ valor de referencia.

Análisis Estadístico.
Una vez hecho lo anterior, a los datos tratados de esta manera se les aplicaron diversaspruebas estadísticas, con una p<0.01:
• T de Tukey para diferencia de medias.
• t-student para diferencia de medias.
• Análisis de varianza ( ANOVA ) de una sola vía ( Miller y Miller, 1993 ).
Elaboración del programa de cómputo.
Para la realización de este objetivo, se consultó la siguiente bibliografía:
1. Sobre el método probit: Finney ( 1964 ), Finney ( 1971 ), Fulk y Poore ( 1991 ) yNMX-AA-087-1995 ( Diario Oficial de la Federación del 1 de diciembre de 1996 ).
2. Desarrollo del algoritmo: Spiegel ( 1976 ), Miller y Miller ( 1993 ) y Burden yFaires ( 2002 ).
3. Implementación del algoritmo en Turbo Pascal: O’Brien ( 1991 ).
4. Validación del programa: Finney ( 1971 ), Fulk y Poore ( 1991 ), Hotos y Vlahos( 1998 ) y datos para determinar la CE50 48h de diferentes xenobióticos en Daphniapulex, provenientes de otros bioensayos realizados como parte del proyecto.

OBJETIVOS Y METAS ALCANZADOS
Los objetivos planteados inicialmente en el proyecto se pudieron cumplir en sutotalidad, excepto el de la evaluación de la toxicidad subletal del metil-paratión cuandosu biotransformación se halla inhibida, ya que no se encontró que la cimetidina afectarala actividad de la EROD.
Los productos generados con este trabajo fueron los siguientes:
1. Una presentación en el 1er Congreso Mesoamericano de Ecotoxicología y Química Ambiental, A. C. el cual se realizó del 22 al 26 de marzo del 2004 en Jiutepec,Morelos.
2. Una publicación en la revista "Bulletin of Environmental Contamination and Toxicology" en este año.
3. Un programa de cómputo para calcular la CE50.

RESULTADOS
1. Toxicidad aguda.
La CE50 48h de los tratamientos evaluados, en orden ascendente de toxicidad fue lasiguiente: metil-paratión con cimetidina, metil-paratión con fenobarbital y metil-paratión ( 40.4, 1 y 17 ppb, respectivamente ) ( Tabla 6 ).
Tabla 6. CE50 48h de las substancias empleadas en este estudio evaluadas en Hyalellaazteca.metil-paratión. 0.172 ppb ( 0.0843, 0.3512 ).cimetidina. *fenobarbital. *metil-paratión con 100 ppm cimetidina. 40.4 ppb ( 19.8, 82.4 ).metil-paratión con 100 ppm fenobarbital. 1 ppb ( 0.4899, 2.0413 )( * ) No se observó respuesta en concentraciones de hasta 100 ppm.
Ya que durante la duración de las pruebas ( 48 h ) ni el fenobarbital ni lacimetidina mostraron un efecto visible sobre los organismos en ninguna de lasconcentraciones empleadas ( 0.01, 0.1, 1, 10 y 100 ppm ), para determinar la CE50 48h delmetil-paratión en presencia de estos fármacos se decidió usar la concentración más altade ellos empleada en las determinaciones ( 100 ppm ).
2. Toxicidad subletal.
Posteriormente, interpolando en las rectas de regresión ( Fig. 5 ) utilizadas para calcularla CE50 48h de cada uno de los tratamientos ( Tabla 6 ), se obtuvieron los valorescorrespondientes de CE0 48h: ( Tabla 7 ):
Tabla 7. CE0 48h de los tratamientos empleados en este estudio evaluados en Hyalellaazteca.metil-paratión. 0.0003 ppb ( 0.0002, 0.0006 ).metil-paratión con 100 ppm cimetidina. 0.4 ppb ( 0.2, 0.8 ).metil-paratión con 100 ppm fenobarbital. 0.0195 ppb ( 0.0095, 0.0397 ).
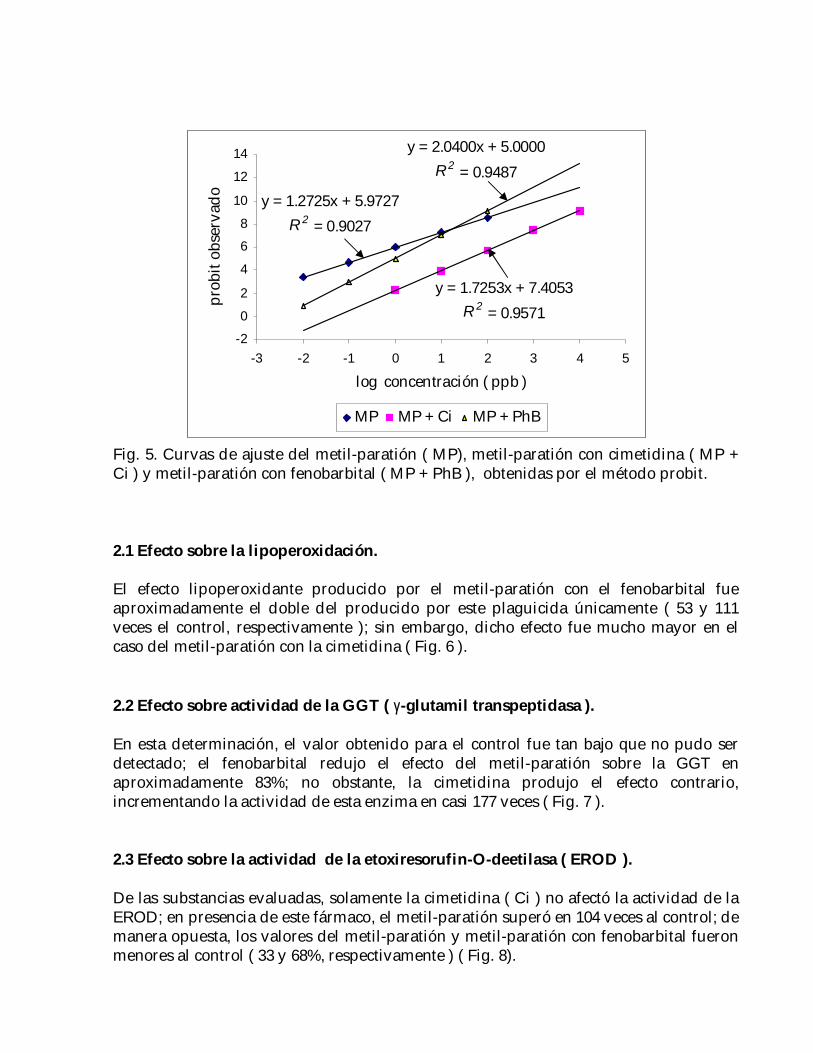
Fig. 5. Curvas de ajuste del metil-paratión ( MP), metil-paratión con cimetidina ( MP +Ci ) y metil-paratión con fenobarbital ( MP + PhB ), obtenidas por el método probit.
2.1 Efecto sobre la lipoperoxidación.
El efecto lipoperoxidante producido por el metil-paratión con el fenobarbital fueaproximadamente el doble del producido por este plaguicida únicamente ( 53 y 111veces el control, respectivamente ); sin embargo, dicho efecto fue mucho mayor en elcaso del metil-paratión con la cimetidina ( Fig. 6 ).
2.2 Efecto sobre actividad de la GGT ( γ-glutamil transpeptidasa ).
En esta determinación, el valor obtenido para el control fue tan bajo que no pudo serdetectado; el fenobarbital redujo el efecto del metil-paratión sobre la GGT enaproximadamente 83%; no obstante, la cimetidina produjo el efecto contrario,incrementando la actividad de esta enzima en casi 177 veces ( Fig. 7 ).
2.3 Efecto sobre la actividad de la etoxiresorufin-O-deetilasa ( EROD ).
De las substancias evaluadas, solamente la cimetidina ( Ci ) no afectó la actividad de laEROD; en presencia de este fármaco, el metil-paratión superó en 104 veces al control; demanera opuesta, los valores del metil-paratión y metil-paratión con fenobarbital fueronmenores al control ( 33 y 68%, respectivamente ) ( Fig. 8).
y = 1.7253x + 7.4053R 2 = 0.9571
y = 1.2725x + 5.9727R 2 = 0.9027
y = 2.0400x + 5.0000R 2 = 0.9487
-2
0
2
4
6
8
10
12
14
-3 -2 -1 0 1 2 3 4 5
log concentración ( ppb )
prob
it ob
serv
ado
MP MP + Ci MP + PhB
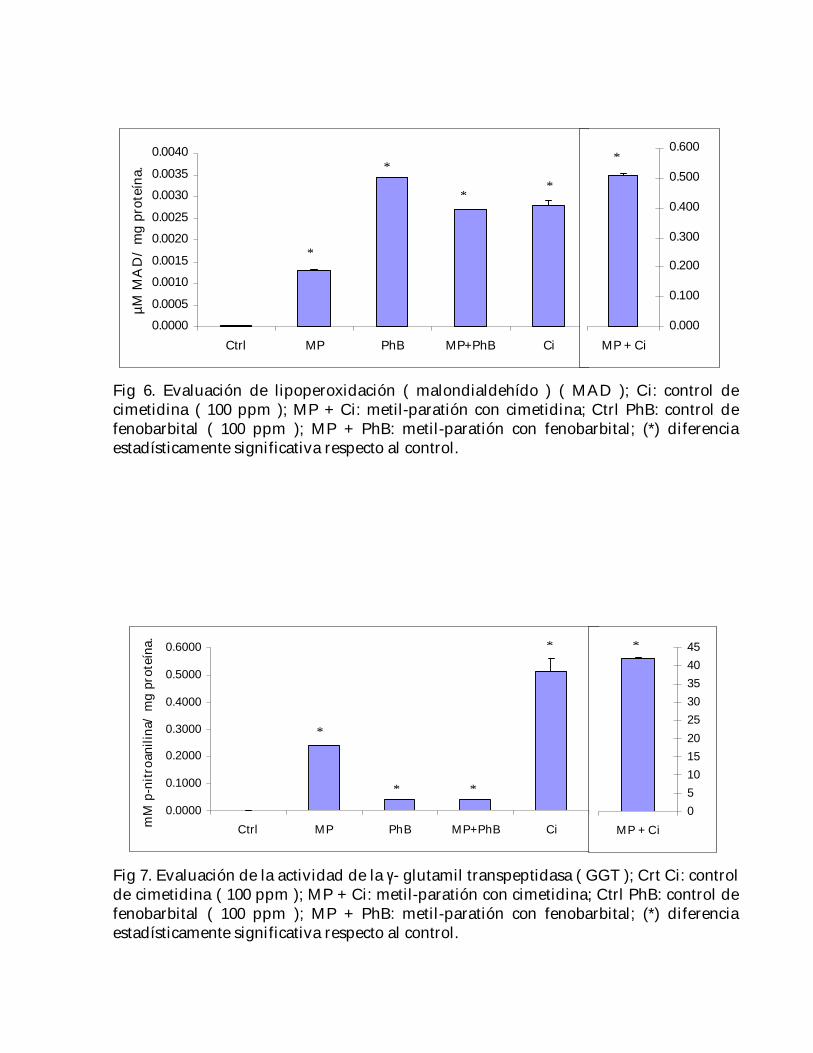
Fig 6. Evaluación de lipoperoxidación ( malondialdehído ) ( MAD ); Ci: control decimetidina ( 100 ppm ); MP + Ci: metil-paratión con cimetidina; Ctrl PhB: control defenobarbital ( 100 ppm ); MP + PhB: metil-paratión con fenobarbital; (*) diferenciaestadísticamente significativa respecto al control.
Fig 7. Evaluación de la actividad de la γ- glutamil transpeptidasa ( GGT ); Crt Ci: controlde cimetidina ( 100 ppm ); MP + Ci: metil-paratión con cimetidina; Ctrl PhB: control defenobarbital ( 100 ppm ); MP + PhB: metil-paratión con fenobarbital; (*) diferenciaestadísticamente significativa respecto al control.
0.0000
0.0005
0.0010
0.0015
0.0020
0.0025
0.0030
0.0035
0.0040
Ctrl MP PhB MP+PhB Ci
µM M
AD
/ m
g pr
oteí
na.
MP + Ci0.000
0.100
0.200
0.300
0.400
0.500
0.600
*
*
*
*
*
0.0000
0.1000
0.2000
0.3000
0.4000
0.5000
0.6000
Ctrl MP PhB MP+PhB CimM
p-n
itroa
nilin
a/ m
g pr
oteí
na.
MP + Ci051015202530354045**
*
*
*
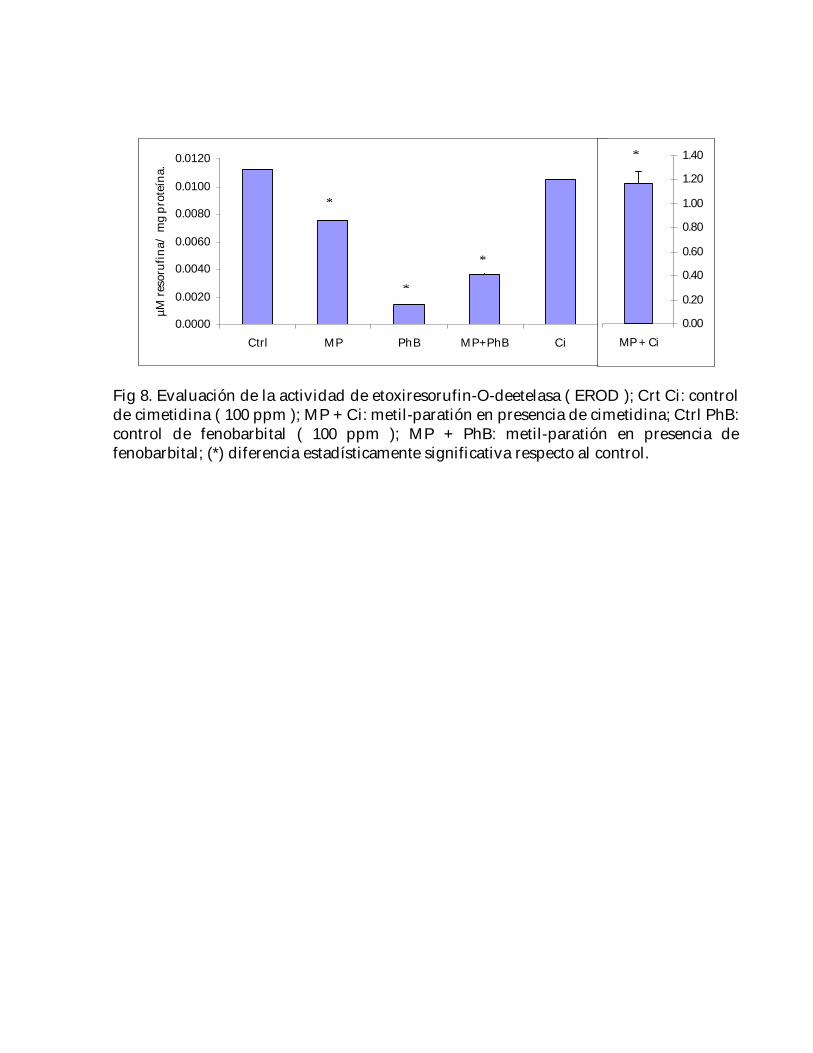
Fig 8. Evaluación de la actividad de etoxiresorufin-O-deetelasa ( EROD ); Crt Ci: controlde cimetidina ( 100 ppm ); MP + Ci: metil-paratión en presencia de cimetidina; Ctrl PhB:control de fenobarbital ( 100 ppm ); MP + PhB: metil-paratión en presencia defenobarbital; (*) diferencia estadísticamente significativa respecto al control.
0.0000
0.0020
0.0040
0.0060
0.0080
0.0100
0.0120
Ctrl MP PhB MP+PhB Ci
µM re
soru
fina/
mg
prot
eína
.
MP + Ci0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40*
*
*
*

3. Salida del programa.
3.1 Metil-paratión.
Mortalidad del grupo control: 0/ 5
Concentracion No. organismos Prop. mortalidad afectados/ ajustada No. organismos totales ------------- -------------- ---------------- 1. 100.000 5/ 5 1.0000 2. 10.000 5/ 5 1.0000 3. 1.000 2/ 5 0.4000 4. 0.100 1/ 5 0.2000 5. 0.010 1/ 5 0.2000
log Concentracion Probit observado Probit estimado----------------- ---------------- --------------- 2.0000 8.4000 8.5176 1.0000 8.4000 7.2452 0.0000 4.7467 5.9727 -1.0000 4.1584 4.7002 -2.0000 4.1584 3.4277
La ec. de la recta es y= 1.2725x +5.9727el coef. de correl. es 0.9027
El valor de la CL50 es 0.1720El intervalo de confianza al 95% es ( 0.0843,0.3512 )
El valor de la CL0 es 0.0003El intervalo de confianza al 95% es ( 0.0002,0.0006 )

3. 2 Metil-paratión con 100 ppm cimetidina.
Mortalidad del grupo control: 0/ 5
Concentracion No. organismos Prop. mortalidad afectados/ ajustada No. organismos totales ------------- -------------- ---------------- 1. 10.000 6/ 6 1.0000 2. 1.000 5/ 5 1.0000 3. 0.100 3/ 5 0.6000 4. 0.010 2/ 5 0.4000 5. 0.001 0/ 5 0.0000
log Concentracion Probit observado Probit estimado----------------- ---------------- --------------- 1.0000 8.4000 9.1307 0.0000 8.4000 7.4053 -1.0000 5.2533 5.6800 -2.0000 4.7467 3.9547 -3.0000 1.6000 2.2293
La ec. de la recta es y= 1.7253x +7.4053el coef. de correl. es 0.9571
El valor de la CL50 es 0.0404El intervalo de confianza al 95% es ( 0.0198,0.0824 )
El valor de la CL0 es 0.0004El intervalo de confianza al 95% es ( 0.0002,0.0008 )

3. 3 Metil-paratión con 100 ppm fenobarbital.
Mortalidad del grupo control: 0/ 6
Concentracion No. organismos Prop. mortalidad afectados/ ajustada
No. organismos totales------------- -------------- ---------------- 1. 100.000 6/ 6 1.0000 2. 10.000 6/ 6 1.0000 3. 1.000 3/ 6 0.5000 4. 0.100 0/ 6 0.0000 5. 0.010 0/ 6 0.0000
log Concentracion Probit observado Probit estimado----------------- ---------------- --------------- 2.0000 8.4000 9.0800 1.0000 8.4000 7.0400 0.0000 5.0000 5.0000 -1.0000 1.6000 2.9600 -2.0000 1.6000 0.9200
La ec. de la recta es y= 2.0400x +5.0000el coef. de correl. es 0.9487
El valor de la CL50 es 1.0000El intervalo de confianza al 95% es ( 0.4899,2.0413 )
El valor de la CL0 es 0.0195El intervalo de confianza al 95% es ( 0.0095,0.0397 )

DISCUSIÓN.
Toxicidad aguda.
En presencia de los fármacos fenobarbital y cimetidina, el valor de la CE50 48h del metil-paratión aumentó en 6 y 238 veces, respectivamente ( Tabla 6 ), reduciendo de estamanera la toxicidad inicial producida por este plaguicida.
Toxicidad subletal.
Los resultados obtenidos sugieren que el fenobarbital estimula la conversión de esteplaguicida hacia metabolitos más tóxicos como el metil-paraoxón, como lo muestra elincremento en la lipoperoxidación producido por el metil-paratión con este fármaco( Fig. 6 ) y la disminución en la actividad de la EROD con este tratamiento ( Fig. 8 ). Esprobable que esto sea debido a que los CYP´s poseen una mayor afinidad por elmetil-paratión que por su oxón ( Kamataki y Neal, 1976 ) o bien porque dichaconversión es autolimitante, a causa del ataque e inactivación de los CYP´s por el oxón( Kamataki y Neal, 1976; Halpert et al., 1980; Butley y Murray, 1993; Keizer et al., 1993;Fabrizi et al., 1999 ) ó a una combinación de ambos factores; sin embargo, también seincrementa la degradación hacia compuestos con menor toxicidad, tales como el p-nitrofenol; el resultado global, es una reducción en la toxicidad inicial producidapor el metil-paratión ( Binder et al., 1984; Stegeman y Hahn, 1994 ) en presencia defenobarbital.
En el caso de la cimetidina, se puede observar que cuando el metil-paratión estáacompañado de este compuesto, hay una gran actividad de la EROD ( Fig. 8 ),implicando la presencia del CYP 1A1 en Hyalella azteca; estos dos resultados sonrelevantes, ya que no habían sido reportados con anterioridad en la literatura; sinembargo, la cimetidina por sí sola no afectó la actividad de la EROD, lo cual concuerdacon los resultados obtenidos por Levine et al. ( 1998 ), en los que utilizando microsomashepáticos in vivo obtenidos de roedores a los que se les administró 150 mg/kg decimetidina, no observaron efecto de este fármaco sobre el CYP 1A1, dependiente de laEROD; por otro lado, se ha visto que en el caso de otros CYP’s ( como p. ej. el 2E ), lacimetidina incluso puede actuar como un inductor ( Tukey y Johnson, 1990 ).
Considerando que la actividad de la EROD del metil-paratión con cimetidina es muchomayor a la que presentan estas substancias por separado ( Fig. 8 ), se puede decir queeste fármaco ejerció un efecto potenciador sobre el metil-paratión; además, ya que elfenobarbital estimula la conversión de este plaguicida hacia su oxón, pero la afinidadde los CYP´s hacia éstos últimos es menor que hacia el compuesto original ( Kamataki yNeal, 1976 ), dicho efecto potenciador no sería atribuible al metil-paratión ni a su oxón,sino posiblemente a un compuesto intermediario entre ambos, el cual puede

corresponder a una ó varias formas de un fosfooxitiirano altamente reactivo ( Neal yHalpert, 1982 ); sin embargo, según Kamataki et al. ( 1976 ) la forma más probable deeste compuesto podría ser la mostrada en la Fig. 9.
La formación de intermediarios inestables similares a éste durante la activación deplaguicidas análogos al paratión en sus correspondientes oxones ( Kamataki y Neal,1976; Fabrizi et al., 1999) podría explicar la inhibición de algunas enzimas que poseen Feó Cu en su estructura debido a la exposición a estos plaguicidas, como p. ej. lasuperóxido dismutasa de Cu-Zn ( SOD Cu-Zn ) en eritrocitos de ratón por elmalatión ( Yarsan et al., 1999 ), la citocromo c oxidasa en hepatopáncreas de camarónMetapenaeus monoceros por el metil-paratión ( Reddy y Rao, 1991 ) ó el CYP 450( Buege y Aust, 1978; Halliwell y Gutteridge, 1999 ); el ataque de estas enzimas por elintermediario reactivo, podría liberar elementos capaces de iniciar ó promover elproceso de lipoperoxidación, tales como los metales Fe ( Buege y Aust, 1978 ) y Cu( Halliwell y Gutteridge, 1984 ) ( principalmente si se hallan en estado reducido ) ó laNADPH- citocromo P450 reductasa, el NADPH y el oxígeno molecular ( O2 )necesarios para el funcionamiento normal de las oxidasas de funciónmixta ( Buege y Aust, 1978; Alvares y Pratt, 1990; Katzung, 1994 ), los cuales promuevenla generación de radicales extremadamente tóxicos, como O2- y OH- ( Halliwell yGutteridge, 1984; Alvares y Pratt, 1990; Commandeur et al., 1995 ).
Con la ruptura de la membrana celular a consecuencia de este proceso lipoperoxidante,se pueden liberar nuevos elementos prooxidantes, como p. ej. iones Cu+1 y Fe+3
intracelulares, pero también defensas antioxidantes, tales como el glutatión y la GGT( la cual es una enzima de membrana ) ( Halliwell y Gutteridge, 1999 ); sin embargo, yaque normalmente los tejidos poseen mecanismos de defensa contra estas especiesreactivas de oxígeno, el efecto lipoperoxidante neto sería el resultado de un balanceentre los efectos agonistas y antagonistas de las substancias prooxidantes yantioxidantes; sin embargo, cuando estas defensas son superadas, p. ej., debido a laexposición subletal a metil-paratión, la respuesta tóxica ( lipoperoxidación celular )puede ser incrementada ( Alvares y Pratt, 1990 ); lo anterior, podría explicar tanto elaumento en la actividad de la GGT ( efecto antioxidante ) como el grado delipoperoxidación ( resultado de un efecto prooxidante ) observado en el tratamiento demetil-paratión con cimetidina ( Figs. 7 y 8 ).
Los estudios ecotoxicológicos con poblaciones silvestres ( como en el caso del presentetrabajo ) son importantes, ya que aunque Hyalella azteca posee una amplia distribuciónen los ambientes lénticos de agua dulce en el continente americano ( Pennak, 1989 ), larespuesta de los individuos de diferentes poblaciones ( ya sean silvestres ó provenientesde cultivos de laboratorio ) ante la exposición a metil-paratión puede no ser la misma( U. S. EPA, 2000 ), debido a la variabilidad genética, las condiciones del medio, lapresencia de otros contaminantes, etc.

Otra razón más para el estudio de los mecanismos de biotransformación de losxenobióticos en los organismos acuáticos, es que a nivel filogenético, algunasinvestigaciones sugieren que a partir de ellos se originaron los sistemas dedesintoxicación presentes en los mamíferos ( incluido el ser humano ), como respuesta alas necesidades particulares de la vida terrestre; además, los plaguicidasorganofosforados tienen menos de 100 años utilizándose, lo cual es un tiempo muybreve para que “surjan” otros sistemas de desintoxicación en los organismos acuáticos,ó los ya existentes se vuelvan más eficientes, por lo que este tipo de estudios refleja lacapacidad de adaptación de dichos sistemas para degradar “nuevos” compuestos.
Además, ya que no existen estudios previos en este tipo de organismos sobre el efectolipoperoxidante del metil-paratión cuando su biotransformación se halla estimulada óinhibida, la información disponible al respecto aún es muy escasa; una forma deincrementar nuestro conocimiento sobre este tema, sería p. ej. determinar laparticipación de los procesos de desulfuración y dearilación en la biotransformación deeste plaguicida, evaluando la producción de metil-paraoxón ( por el grado deinhibición de la acetilcolinesterasa ) y p-nitrofenol.
También se desconoce la posible participación de otras vías ó mecanismos, tales comolas esterasas ( Straus et al., 2000 ) o la glutatión S-transferasa ( Snyder, 2000 ). Esta últimaenzima podría ser un mejor biomarcador que la GGT, al ser la primera en interactuarcon los xenobióticos en la vía del ácido mercaptúrico ( Fukami, 1976;Commandeur et al., 1995 ), poder conjugarse con el metil-paraoxón ( Hollingworth et al.,1973 ) y ser inducida por fenobarbital en el cladócero Daphnia magna ( Baldwin yLeblanc, 1994 ).
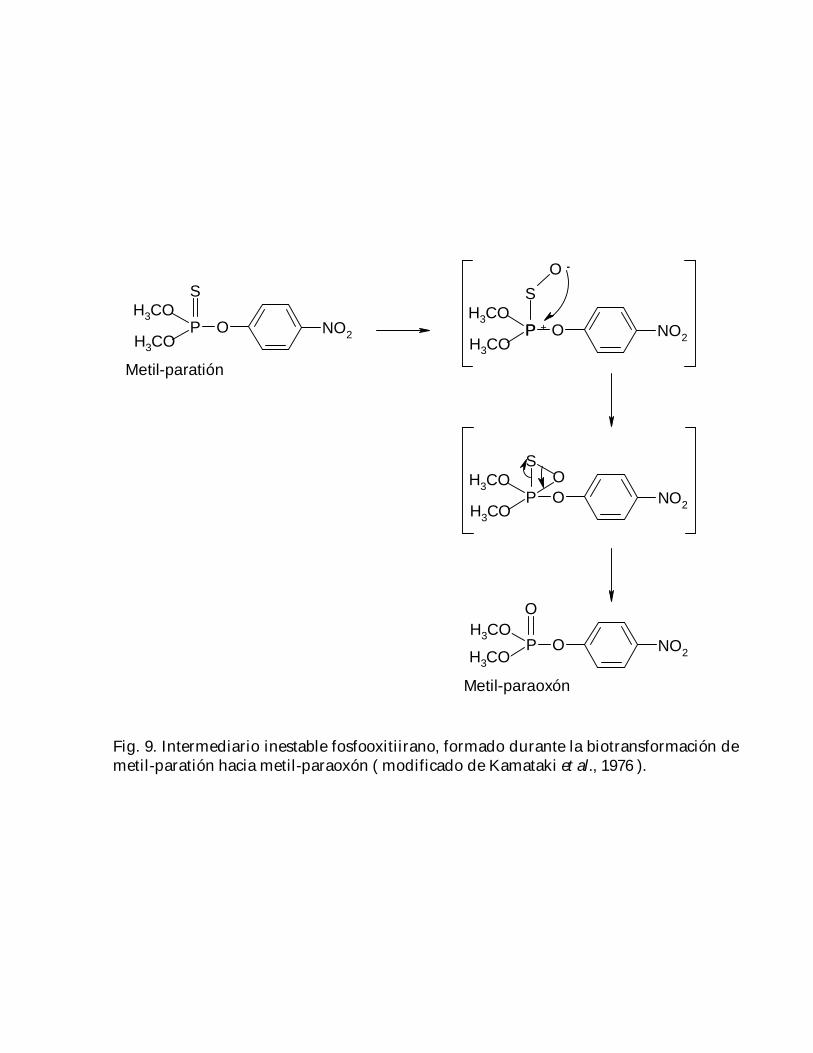
Fig. 9. Intermediario inestable fosfooxitiirano, formado durante la biotransformación demetil-paratión hacia metil-paraoxón ( modificado de Kamataki et al., 1976 ).
P O NO2
S
P O NO2
S
P O NO2
S
P O NO2
O
Metil-paratión
H3CO
H3CO
H3CO
H3CO
O
Metil-paraoxón
H3CO
H3CO
O -
P +
H3CO
H3CO

CONCLUSIONES.
1) El valor de CE50 48h del metil-paratión con Hyalella azteca en la columna de agua( 0.17 ppb ), es similar al reportado en la literatura para Daphnia magna ( 0.14 ppb )( U. S. EPA, 1998 ).
2) En los tres biomarcadores empleados ( grado de lipoperoxidación, actividad de laGGT y EROD ), la cimetidina ejerció un efecto potenciador sobre el metil-paratión;es posible que en esta interacción esté implicado un intermediario inestablealtamente reactivo ( fosfooxitiirano ), formado durante la biotransformación delmetil-paratión en su oxón.
3) A nivel subletal, no se pudo lograr la inhibición del CYP 1A1 ( dependiente de laEROD ) con el uso de la cimetidina.
4) Sin embargo, de manera indirecta, se pudo inferir la presencia del CYP 1A1 en esteanfípodo.
5) Por último, para una misma serie de datos ( que se ajusten al método probit ), losvalores de CE50 proporcionados por el programa “CL50” y los obtenidos por otrosmedios ( como el uso de tablas probit u otros programas de cómputo ) no difierieronestadísticamente entre sí, por lo que dicho programa cumple con la función para lacual fue diseñado.

LITERATURA CITADA
Aïche, J. M., Devissaguet, J. y Guyot-Hermann, A. M., ( 1983 ). Biofarmacia, El ManualModerno, México.
Alvares, A. P. y Pratt, W. B., ( 1990 ). Pathways of Drug Metabolism. In: Pratt, W. B. yTaylor, P., ( Eds. ). Principles of Drug Action. The Basis of Pharmacology, 3rd ed.,Churchill Livingstone, UK, 365- 422.
APHA, AWWA, WPCF, ( 1989 ). Standard Methods for the examination of Waste and WasteWater, 17th ed., American Public Health Association, American Water WorksAssociation and Water Pollution Control Federation, USA.
Baldwin, W. S. y LeBlanc, G. A., ( 1996 ). Expression and induction of aninmunochemically related class of glutathione S-transferases in Daphia magna. Comp.Biochem. Physiol. Part B Biochem. Mol. Biol., 113 ( 2 ): 261-267.
Binder, R. L., Melancon, M. J. y Lech, J. J., ( 1984 ). Factors influencing the persistenceand metabolism of chemicals in fish. Drug Metab. Rev., 15 ( 4 ): 697-724.
Bradford, M. M., ( 1976 ). A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dye binding. Anal.Biochem., 72: 248-254.
Buege, J. A. y Aust, S. D., ( 1978 ). Microsomal lipid peroxidation. Methods. Enzymol., 52:302-310.
Burden, R. L. y Faires, J. D., ( 2002 ). Análisis Numérico, 6ª ed., International ThomsonEditores, México.
Butler, A. M. y Murray, M., ( 1993 ). Inhibition and inactivation of constitutivecytochromes P450 in rat liver by parathion. Mol. Pharmacol., 43: 902-908.
Commandeur, J. N. M., Stijntjes, G. J. y Vermeulen, N. P. E., ( 1995 ). Enzymes andTransport Systems Involved in the Formation and Disposition of Glutathione S-Conjugates. Pharmacol. Rev., 47 ( 2 ): 271-330.
Comporti, M., ( 1993 ). Lipid peroxidation: Biopathological significance. Molec. AspectsMed., 14: 199-207.
Diario Oficial de la Federación. NMX-AA-087-1995. Análisis de agua. Evaluación detoxicidad aguda con Daphnia magna Straus ( Crustacea: Cladocera ). Método de prueba( 1 de diciembre de 1996 ).

Diario Oficial de la Federación. NOM-041-SSA1-1993. Bienes y servicios. Aguapurificada envasada. Especificaciones sanitarias ( 24 marzo 1995 ).
Diario Oficial de la Federación. NOM-127-SSA1-1994 ). Salud ambiental, agua para usoy consumo humano. Límites permisibles de calidad y tratamientos a que debesometerse el agua para su potabilización ( 18 enero 1996 ).
Diario Oficial de la Federación. NOM-001-SEMARNAT-1996 ( antes NOM-001-ECOL-1996 ), que establece los límites máximos permisibles de contaminantes en las descargasresiduales en aguas y bienes nacionales ( 6 enero 1997 ).
Diario Oficial de la Federación. NOM-052-SEMARNAT-1993 ( antes NOM-052-ECOL-1993 ), que establece las características de los residuos peligrosos, el listado de losmismos y los límites que hacen a un residuo peligroso por su toxicidad al ambiente( 22 octubre 1993 ).
Dikshit, T. S. S., ( 1991 ). Toxicology of pesticides in animals. CRC Press, Boca Raton,Boston.
Draper, H. H., ( 1990 ). Advances in Nutritional Research. Vol. 8, Plenum Press, New York.
Dreisbach, R. H. y Robertson, W. O., ( 1988 ). Manual de Toxicología Clínica. Prevención,Diagnóstico y Tratamiento, 6a ed., Manual Moderno, México, 95-104.
EPA, ( 1991 ). Methods for measuring the acute toxicity of effluents and receiving waters tofreshwaters and marine organisms, 4th ed., Environmental Protection Agency, EPA-600/4-90-027.
Fabrizi, L., Gemma, S., Testai y E. Vittozzi, L., ( 1999 ). Identification of the cytochromep450 isoenzymes involved in the metabolism of diazinon in the rat liver. J. Biochem.Molecular Toxicology, 13 ( 1 ): 53-61.
FAO, ( 1997 ). Plaguicidas que suscitan problemas bajo condiciones de empleo existentes enpaíses en desarrollo: Paratión-metilo. FAO, Secretaría Provisional del Convenio deRotterdam sobre el proceso del Consentimiento Previo aplicable a ciertos Plaguicidas yProductos Químicos Peligrosos Objeto del Comercio Internacional. FAO & PNUMA,Roma.
Finney, D. J. ( 1964 ). Statistical method in biological assay, 2ª ed., Compton PrintingLimited, London.
Finney, D. J., ( 1971 ). Probit Analysis, 3rd ed., Cambridge University Press, Great Britain.

Fulk, F. y Poore, C., ( 1991 ). Acute toxicity data analysis. In: Weber, C. I. ( Ed. ).Methods for measuring the acute toxicity of effluents and receiving waters to freshwaterand marine organisms, 4th ed., Environmental Protection Agency, EPA-600/4-90-027.
Fukami, J. I., ( 1976 ). Metabolism of several insecticides by glutathione S-transferase.Pharmac. Ther., 10: 473-514.
Guenguerich, F. P., ( 1977 ). Separation and purification of multiple forms ofmicrosomal cytochrome P450: activities of diferent forms of cytochrome P450 towardsseveral compounds of environmental interest. J. Biol. Chem., 252: 3970-3979.
Halpert, J. A., Hammond, D. y Neal, R. A., ( 1980 ). Inactivation of purified rat livercytochrome P450 during the metabolism of parathion ( diethyl-p-nitrophenyl-phosphorothionate ). J. Biol. Chem., 255: 1080-1089.
Halliwell, B. y Gutteridge, J. M. C., ( 1984 ). Oxygen toxicity, oxygen radicals, transitionmetals and disease. Biochem. J., 219: 1-14.
Halliwell, B. y Gutteridge, J. M. C., ( 1999 ). Oxidative stress, adaptation, damage, repairand death. In: Halliwell, B. y Gutteridge, J. M. C. ( Eds. ). Free radicals in biology andmedicine. 3rd ed., Oxford University Press, NY, 246-350.
Hargrave, ( 1970 ). The utilization of benthic microflora by Hyalella azteca. J. AnimalEcology, 39: 427-437.
Hassall, K. A. ( 1990 ). The biochemistry and uses of pesticides, 2nd ed., Houndmills, McMillan.
Hollingworth, R. M., Alstott, R. L. y Litzenberg, R. D., ( 1973 ). Glutathione S-aryltransferase in the metabolism of parathion and its analogs. Life Sci., 13: 191-199.
Hotos, G. N. y Vlahos, N., ( 1998 ). Salinity tolerance of Mugil cephalus and Chelonlabrosus ( Pisces: Mugilidae ) fry in experimental conditions. Aquaculture, 167: 329-338.
Hurh, E., Lee, E. J., Lee, A. K., Kim, Y. G., Kim, S. H., Kim, S. G. y Lee, M. G., ( 2000 ).Effects of enzyme inducers or inhibitors on the pharmacokinetics of intravenousparathion in rats. Biopharm. Drug Dispos., 21: 193-204.Huseby, N. E. y Stromme, J. H., ( 1974 ). Practical points regarding routinedetermination of gamma-glutamyl transferase ( gamma-GT ) in serum with a kineticmethod at 37 degrees C. Scand. J. Clin. Lab. Invest., 34 ( 4 ): 357-363.
Kamataki, T. y Neal, R. A., ( 1976 ). Metabolism of diethyl-p-nitrophenyl-phosphorothionate ( parathion ) by a reconstituted mixed-function oxidase enzymesystem: studies of the covalent binding of the sulfur atom. Mol. Pharmacol., 12: 933-944.

Kamataki, T., Lee Lin, M. C. M., Belcher, D. H. y Neal, R. A., ( 1976 ). Studies of themetabolism of parathion with an apparently homogeneous preparation of rabbit livercytochrome P-450. Drug Metab. Dispos., 4: 180-189.
Katzung, B. G., ( 1994 ). Farmacología básica y clínica, 5ª ed., Manual Moderno, México.
Keizer, J. D., Agostino, G., Nagel, R., Gramenzi, F. y Vittozzi, L., ( 1993 ). Comparativediazinon toxicity in guppy and zebra fish: Different role of oxidative metabolism.Environ. Toxicol. Chem., 12: 1243-1250.
Khrer, J. P., ( 1993 ). Free radical as mediator of tissue injury and disease. Crit. Rev.Toxicol., 23: 21-48.
Koner, B. C., Banerjee, B. D. y Ray, A., ( 1998 ). Organochlorine pesticide-inducedoxidative stress and inmune suppression in rats. Indian J. Exp. Biol., 36: 395-398.
Kubitz, J. A. y Giesy, J. P., ( 1996 ). Changes in overlaying water quality and sedimenttoxicity to Hyalella azteca during a 10-day sediment bioassay: Static vs. renewalregimens. In: Munawar, M. y Dave, G. ( Eds. ). Development and Progress in SedimentQuality Assessment: Rationale, Challenges, Techniques and Strategies, SPB AcademicPublishing, Amsterdam, The Netherlands, 101-113.
Levine, M., Law, E. Y. W., Bandiera, S. M., Chang, T. K. H. y Bellward, G. D., ( 1998 ). Invivo cimetidine inhibits hepatic CYP2C6 and CYP2C11 but not CYP1A1 in adult malerats. J. Pharmacol. Exp. Ther., 284: 493-499.
Martin, D. W. Jr., Mayes, P. A., Rodwell, V. W. y Granner, D. K., ( 1985 ). Harper´sReview of Biochemistry, 20th ed., Lange Medical Publiclations, Los Altos, CA.
Merck, ( 1996 ). The Merck Index. An Encyclopedia of Chemicals, Drugs and Biologicals, 20th
ed., Merck &Co., Inc., Whitehouse Station, N. J., USA.
Metcalf, C. L. y Flint, W. P., ( 1984 ). Insectos destructivos e insectos útiles. Sus costumbres ycontrol. CECSA, México.
Miller, J. C. y Miller, J. N., ( 1993 ). Estadística para química analítica, 2ª ed., Addison-Wesley Iberoamericana, USA.
Mourelle, M., Giron, E., Amezcua, J. L. y Martínez-Tabche, L., ( 1986 ). Cimetidineenhances and phenobarbital decreases parathion toxicity, J. Appl. Toxicol., 6 ( 6 ): 401-404.

Neal, R. A. y Halpert, J., ( 1982 ). Toxicology of thiono-sulfur compounds. Ann. Rev.Pharmacol. Ther., 22: 321-339.
Ohhira, S., Matsui, H. y Watanabe, K., ( 2000 ). Effects on triphenyltin metabolism andtoxicity in mice. Toxicol. Lett., 117: 145-150.
O´Brien, S. K., ( 1991 ). Turbo Pascal 6: Manual de Referencia. Osborne Mc Graw-Hill,México.
Paolicchi, A., Tongiani, R., Tonarelli, P., Comporti, M. y Pompella, A., ( 1997 ).Gamma glutamil transpeptidase-dependent lipid peroxidation in isolatedhepatocytes and HepG2 hepatoma cells. Free Rad. Biol. Med., 22: 853-860.
Parrot, J. L., Chong-Kit, R. y Rokosh, D. A., ( 1999 ). EROD induction in fish: a tool tomeasure environmental exposure. In: Rao, S. S. ( Ed. ). Impact Assessment of HazardousAquatic Contaminants. Concepts and Approaches, CRC Press, 99-124.
Pennak, R. W., ( 1989 ). Freshwater Invertebrates of the Unites States. Johh Wiley and Sons,Inc., New York.
Phipps, G. L., Mattson, V. R. y Ankley, G. T., ( 1995 ). The relative sensitivity of threebenthic test species to 10 chemicals. Arch. Environ. Toxicol. Chem., 28: 281-286.
Reddy, M. S. y Rao, K. V., ( 1991 ). Phosphamidon, methylparathion and dichlorvosimpact on tissue oxidative metabolism in penaeid prawn, Metapenaeus monoceros.Biochem. Int., 23: 439-447.
Rendic, S., Kajfez, F., y Rulf, H. H., ( 1983 ). Characterization of cimetidine, ranitidineand related structures interaction with cytochrome P-450. Drug Metab. Dispos., 11: 137-142.
Scheuhammer, A. M. y Cherian, M. G., ( 1986 ). Quantification of metallothioneins by asilver-saturation method. Toxicol. Appl. Pharmacol., 82: 416-425.
Snyder, M. J., ( 2000 ). Cytochrome P450 enzymes in aquatic invertebrates: recentadvances and future directions. Aquat. Toxicol., 48: 529-547.
Spiegel, M. R., ( 1976 ). Teoría y problemas de probabilidad y estadística. Serie de compendiosSchaum. Mc Graw-Hill, México.
Stegeman, J. J. y Hahn, M. E., ( 1994 ). Biochemistry and molecular biology ofmonooxygenases: current perspectives on forms, functions and regulation of citochromeP450 in aquatic species. In: Malins, D. C., Ostrander, G. K. ( Eds. ). Aquatic Toxicology,

Molecular, Biochemical and Cellular Perspectives, Lewis Publishers, Boca Raton, FL, 87-206.
Straus, D. L., Schlenk, D. y Chambers, J. E., ( 2000 ). Hepatic microsomal desulfurationand dearilation of clorpyrifos and parathion in fingerling channel catfish: lack of effectfrom Aroclor 1254. Aquat. Toxicol., 50: 141-149.
Tate, S. S. y Meister, A., ( 1985 ). γ-Glutamyl transpeptidase from kidney. MethodsEnzymol., 113: 400-419.
Tukey, R. H. y Johnson, E. F., ( 1990 ). Molecular Aspects of Regulation and Estructureof the Drug-Metabolizing Enzymes. In: Pratt, W. B. y Taylor, P., ( Eds. ). Principles ofDrug Action. The Basis of Pharmacology, 3rd ed., Churchill Livingstone, UK, 423-467.
U. S. EPA, ( 1998 ). Methyl parathion . Environmental Fate and Effects. Preeliminary RiskAssessment. Office of Prevention, Pesticides and Toxic Substances. EnvironmentalProtection Agency, EPA 2460.
U. S. EPA, ( 2000 ). Methods for Measuring the Toxicity and Bioaccumulation of Sediment-associated Contaminants with Freshwater Invertebrates, 2nd ed., Office of Research andDevelopment, Office of Science and Technology, Office of Water and U. S.Environmental Protection Agency, EPA 20460.
Yarsan. E., Tanyuksel, M., Celik, A. y Aydin, A., ( 1999 ). Effects of aldicarb andmalathion on lipid peroxidation. Bull. Environ. Contam. Toxicol., 63: 575-581.





























