Embed Size (px)
Citation preview

UNIVERSIDADE DO ESTADO DO AMAZONAS
FUNDAÇÃO DE MEDICINA TROPICAL DR. HEITOR VIEIRA DOURADO
PROGRAMA DE PÓS-GRADUAÇÃO EM MEDICINA TROPICAL
DOUTORADO EM DOENÇAS TROPICAIS E INFECCIOSAS
NÍVEIS DE EXPRESSÃO DE pvcrt-o E pvmdr-1 ESTÃO ASSOCIADOS COM A
RESISTÊNCIA À CLOROQUINA E MALÁRIA GRAVE POR PLASMODIUM VIVAX
EM PACIENTES DA AMAZÔNIA BRASILEIRA
GISELY CARDOSO DE MELO
MANAUS
2014

i
GISELY CARDOSO DE MELO
NÍVEIS DE EXPRESSÃO DE pvcrt-o E pvmdr-1 ESTÃO ASSOCIADOS COM A
RESISTÊNCIA À CLOROQUINA E MALÁRIA GRAVE POR PLASMODIUM VIVAX
EM PACIENTES DA AMAZÔNIA BRASILEIRA
Tese apresentada ao Programa de Pós-Graduação
em Medicina Tropical da Universidade do Estado
do Amazonas em Convênio com a Fundação de
Medicina Tropical Dr. Heitor Vieira Dourado, para
obtenção grau de Doutor em Doenças Tropicais e
Infecciosas.
Orientador: Dr. Marcus Vinícius Guimarães de Lacerda
Co-orientador: Dra. Maria del Carmen Fernandez-Becerra
MANAUS
2014

ii
M528n Melo, Gisely Cardoso de.
Níveis de expressão de pvcrt-o e pvmdr-1 estão associados com
a resistência à cloroquina e malária grave por Plasmodium vivax em
pacientes da Amazônia Brasileira. / Gisely Cardoso de Melo. --
Manaus: Universidade do Estado do Amazonas, Fundação de
Medicina Tropical, 2014.
xx, 158 f. : il.
Tese (Doutorado) apresentada ao Programa de Pós-Graduação
em Medicina Tropical da Universidade do Estado do Amazonas –
UEA/FMT, 2014.
Orientador: Marcus Vinícius Guimarães de Lacerda
Co-orientadora: Dra Maria del Carmen Fernandez-Becerra
1. Cloroquina. 2. Resistência 3. Malária 4. Plasmodium vivax 5.pvcrt-o
6.Pvmdr 1 I. Título.
CDU: 614.4(811ti)
Ficha Catalográfica elaborada pela Bibliotecária Maria Eliana N Silva, lotada na
Escola Superior de Ciências da Saúde - UEA

iii
FOLHA DE JULGAMENTO
Níveis de expressão de pvcrt-o e pvmdr-1 estão associados com a resistência
à cloroquina e malária grave por Plasmodium vivax em pacientes da
Amazônia Brasileira
GISELY CARDOSO DE MELO
“Esta Tese foi julgada adequada para obtenção do Título de Doutor em Doenças Tropicais e
Infecciosas, aprovada em sua forma final pelo Programa de Pós-Graduação em Medicina Tropical da
Universidade do Estado do Amazonas em convênio com a Fundação de Medicina Tropical Dr. Heitor
Vieira Dourado”.
Banca Julgadora:
______________________________________
Presidente
______________________________________
Membro
______________________________________
Membro
______________________________________
Membro

iv
DEDICATÓRIA
Desde o primeiro ano do curso farmácia na Universidade Estadual de Maringá, em 2000, me
aventurei ingenuamente como estagiária no Laboratório de Imunologia Clínica do
Departamento de Análises Clínicas. Neste ambiente, comecei a realizar meus primeiros
passos para o diagnóstico laboratorial das doenças infecciosas.
Mas não foi no estudo da malária que ingressei na pesquisa científica. De 2001 a 2003, fui
selecionada para o Programa Institucional de Bolsas de Iniciação Científica (PIBIC), sob
orientação do professora Ana Lúcia Falavigna Guilherme. Estudava a epidemiologia das
parasitoses intestinais no Estado do Paraná. Durante esse período, sempre envolvida em
projetos de outros alunos, fui monitora da disciplina de parasitologia, onde tive meu primeiro
contato mais diretamente com a malária. Após a conclusão do curso de farmácia em 2005, eu
e meu marido Wuelton mudamos para o Amazonas. Havíamos feito um concurso e éramos
funcionários da Secretaria Estadual de Saúde do Amazonas (SUSAM).
Em 2007, decidi ingressar no mestrado e fui selecionada para o Programa de Pós-graduação
em Medicina Tropical da Universidade do Estado do Amazonas e da Fundação de Medicina
Tropical do Amazonas. Precisava de um orientador, porém não conhecia ninguém. Fui
encaminhada para a malária pois precisavam de um aluno para desenvolver projeto de
mestrado. Foi assim, que desenvolvi minha dissertação de mestrado intitulado: “Proteção
contra a diminuição da hemoglobina em escolares com malária por Plasmodium vivax
coinfectados por helmintos intestinais”, a princípio, sob orientação da Profa. Dra. Maria das
Graças Costa Alecrim e co-orientação do Prof. Dr. Marcus Vinícius Guimarães de Lacerda.
Durante esse período, aprendi muita coisa com o Dr. Marcus Lacerda sobre malária,
pesquisas cientificas, epidemiologia, pois em cada viagem que fazíamos ao Careiro Castanho
para desenvolver o projeto discutíamos artigos e dados dos projetos, muitas vezes, com
diversos pesquisadores de outras partes do Brasil e do Mundo. Em 2009, quando defendi meu
mestrado, agora enfim sob orientação do Prof. Dr. Marcus, fui convidada para continuar no
seu grupo de pesquisa e em 2010 ingressei no doutorado.
Foram muitas as discussões sob o tema do meu projeto de doutorado. Nesse período conheci
a Prof. Dra. Carmen Fernandez-Becerra e Prof. Dr. Hernando del Portillo que haviam

v
verificado níveis aumentados de expressão gênica de pvcrt-o e pvmdr-1 em um paciente com
malária grave vivax em comparação com três não-graves. Decidimos incluir a princípios
mais pacientes graves e posteriormente pacientes resistentes à cloroquina. Nesse período,
passei pelo laboratório do Prof. Hernando e Profa. Carmen no CRESIB onde aprendi muita
coisa sobre biologia do parasita, métodos moleculares e onde muitas discussões aconteceram
sobre os meus dados e novas perspectivas. Ainda durante o doutorado, passei no concurso da
Universidade do Estado do Amazonas como professora de parasitologia.
Na tese desenvolvida na Gerência de Malária da Fundação de Medicina Tropical do
Amazonas apresento resultados que considero preliminares de uma pesquisa em malária na
Região Amazônica, na área da resistência à cloroquina e malária grave. Já que, poderíamos
prever a resistência a cloroquina e malária grave no momento do diagnóstico da malária
vivax.
Dedico este trabalho a todos que foram citados neste texto, e também a todos que
participaram direta ou indiretamente da história da minha vida acadêmica.

vi
AGRADECIMENTOS
A Deus, agradeço pela vida e por todas as oportunidades a mim oferecidas.
Aos meus pais, Gileno Cardoso de Melo e Maria Augusto de Melo (in memorian), porque
acreditaram em mim e investiram com amor na minha formação, sempre me incentivando.
Ao meu esposo, Wuelton Marcelo Monteiro, pelo amor, compreensão e presença, mesmo nos
momentos difíceis e atribulados de uma aluna de doutorado. Agradeço pelas contribuições
intelectuais para esta tese.
À minha filha Manuela que também foi um grande presente na fase final do meu doutorado.
Ao meu orientador, Prof. Dr. Marcus Vinicius Guimarães de Lacerda, pela atenção, dedicação
e incentivo durante a realização deste trabalho. Sua inteligência e competência foram suas
maiores lições para este aluno.
A minha co-orientadora, Profa. Dra. Maria del Carmen Fernandez-Becerra, pela presteza na
realização das técnicas moleculares e atenção dispensada durante todo esse período. Permitiu
que esta doutoranda pudesse aprender técnicas de produção de antígenos e marcadores
sorológicos no CRESIB.
Ao Prof. Dr. Hernando del Portillo, pela atenção e incentivo. Exemplo de pesquisador
inteligente e produtivo. Me permitiu desenvolver parte do doutorado em seu laboratório no
CRESIB cedendo material, reagentes, orientação e oportunidades de aprendizado.
‘À minha aluna de iniciação cientifica, Siuhelem Rocha da Silva, pela amizade, companhia e
auxílio na preparação dos materiais e experimentos.
Ao amigo André Machado de Siqueira, pela contribuição intelectual e análise estatística no
artigo cientifico.

vii
À gerente de Malária da FMT-HVD, Monica Regina Farias Costa, pelo apoio na realização do
projeto.
A todos os técnicos da Gerência de Malária pela ajuda e sabedoria. Maria Raimunda da Costa
e Wellington Silva pela auxilio na abordagem dos pacientes e coleta de amostras de sangue. A
José Eckner Alves Lessa pela revisão nas gotas espessas e contagem das parasitemias. À
Nélida Teresa Sanches pelo colaboração e auxilio nas extrações de DNA.
À Profa. Dra Maria das Graças Vale Barbosa, pela forma com que conduz a coordenação da
pós- graduação e agora como minha amiga de trabalho.
Aos amigos do Programa de Pós-graduação em Medicina Tropical, pela convivência
agradável.
Aos funcionários da Secretaria do Programa de Pós-Graduação em Medicina Tropical da
Universidade do Estado do Amazonas, pela disponibilidade e sempre bom atendimento.
A estes que nominei, e a todos os outros que contribuíram indiretamente neste trabalho.

viii
DECLARAÇÃO DAS AGÊNCIAS FINANCIADORAS
Este estudo foi financiado pelo Conselho Nacional de Desenvolvimento Científico e
Tecnológico (CNPq) (número de concessão 575788/2008-9). CFB recebeu bolsa de
pesquisador visitante do Programa Estratégico para a Ciência, Tecnologia e Inovação da
FAPEAM (PECTI-SAÚDE). MVGL é bolsista de produtividade nível 1 do Conselho
Nacional de Desenvolvimento Científico e Tecnológico (CNPq). A tese se insere no
Programa de Pós-Graduação em Medicina Tropical da Universidade do Estado do Amazonas,
em convênio com a Fundação de Medicina Tropical do Amazonas, beneficiando-se do auxílio
financeiro e da infraestrutura específica dessas instituições.

ix
EPÍGRAFE
“É muito melhor lançar-se em busca de
conquistas grandiosas, mesmo expondo-se ao
fracasso, do que alinhar-se com os pobres de
espírito, que nem gozam muito nem sofrem
muito, porque vivem numa penumbra cinzenta,
onde não conhecem nem vitória, nem derrota.”
Theodore Roosevelt

x
RESUMO
O estudo dos marcadores moleculares associados com o aumento da resistência à cloroquina e
a gravidade da doença em Plasmodium vivax são necessários. O objetivo deste estudo foi
avaliar os níveis de expressão de pvcrt-o e pvmdr-1 em um grupo de pacientes que
apresentaram resistência à cloroquina e pacientes que desenvolveram complicações graves
provocados exclusivamente pelo P. vivax. Dois grupos diferentes de pacientes foram
incluídos neste estudo realizado na Amazônia brasileira: 1) pacientes com malária por P.
vivax caracterizados clinicamente resistentes à cloroquina em comparação com pacientes
sensíveis em um estudo in vivo e 2) pacientes com malária vivax grave em comparação com
pacientes sem qualquer gravidade. Foi realizado PCR em Tempo Real Quantitativo Relativo
para comparar os níveis de transcrição de dois genes transportadores principais do P. vivax:
Gene de resistência do P. vivax à cloroquina (pvcrt-o) e gene de resistência do P. vivax à
múltiplas drogas (pvmdr-1). Doze casos resistentes à cloroquina e outros 15 isolados de casos
susceptíveis foram incluídos no primeiro grupo de pacientes. Para o segundo, foram incluídos
sete pacientes com malária vivax grave e 10 manifestações não graves. Parasitos de pacientes
com resistência à cloroquina apresentaram aumento na expressão genica de 6,1 (IC 95%: 3,8-
14,3) e 2,4 (IC 95%: 0,53-9,1) para pvcrt-o e pvmdr-1 em comparação com o grupo
susceptível . Parasitas com malária vivax grave apresentaram aumento de 2,9 (IC 95%: 1,1-
8,3) e 4,9 (IC 95%: 2,3-18,8) na expressão de pvcrt-o e pvmdr-1 em comparação com o grupo
controle com doença não grave. Estes resultados sugerem que a resistência à cloroquina e
gravidade clínica em infecções por P. vivax estão fortemente associados com aumento dos
níveis de expressão de pvcrt-o e pvmdr-1 genes provavelmente envolvidos na resistência à
cloroquina.
Palavras-chave: cloroquina, resistência, malária, Plasmodium vivax, pvcrt-o, pvmdr-1,
expressão gênica, marcadores moleculares.

xi
ABSTRACT
Molecular markers associated with the increase chloroquine resistance and disease severity in
Plasmodium vivax are needed. The objective of this study was to evaluate the expression
levels of pvcrt-o and pvmdr-1 genes in a group of patients presenting CQRPv and patients
who developed severe complications triggered exclusively by P. vivax infection. Two
different sets of patients were included to this comprehensive study performed in the
Brazilian Amazon: 1) patients with clinically characterized chloroquine-resistant P. vivax
compared with patients with susceptible parasites from in vivo studies and 2) patients with
severe vivax malaria compared with patients without severity. Quantitative real-time PCR
was performed to compare the transcript levels of two main transporters genes, P. vivax
chloroquine resistance transporter (pvcrt-o) and the P. vivax multidrug resistance transporter
(pvmdr-1). Twelve chloroquine resistant cases and other 15 isolates from susceptible cases
were included in the first set of patients. For the second set, seven patients with P. vivax-
attributed severe and 10 mild manifestations were included. Parasites from patients with
chloroquine resistance presented up to 6.1 (95%CI: 3.8-14.3) and 2.4 (95%CI: 0.53-9.1) fold
increase in pvcrt-o and pvmdr-1 expression levels, respectively, compared to the susceptible
group. Parasites from the severe vivax group had a 2.9 (95%CI: 1.1-8.3) and 4.9 (95%CI: 2.3-
18.8) fold increase in pvcrt-o and pvmdr-1 expression levels as compared to the control group
with mild disease. These findings suggest that chloroquine resistance and clinical severity in
P. vivax infections are strongly associated with increased expression levels of the pvcrt-o and
pvmdr-1 genes likely involved in chloroquine resistance.
Keywords: chloroquine, resistance, malaria, Plasmodium vivax, pvcrt-o, pvmdr-1, gene
expression, molecular markers.

xii
LISTA DE FIGURAS
Figura 1 Ilustração do ciclo biológico do plasmódio............................................. 5
Figura 2 Transmissão da malária por Plasmodium vivax no mundo .................... 6
Figura 3 Malária nas Américas por Índice Parasitário Anual (IPA), 2013............ 8
Figura 4 Estrutura molecular da cloroquina........................................................... 10
Figura 5 Distribuição mundial de falha terapêutica e resistência confirmada de
P. vivax à cloroquina................................................................................
11
Figura 6 Distribuição de estudos in vivo de eficácia da cloroquina para
Plasmodium vivax entre 2000 e 2013.......................................................
12
Figura 7 Estrutura do transportador PvCRT-O....................................................... 14
Figura 8 Estrutura do transportador PvMDR1....................................................... 15
Figura 9 Mapa do Estado do Amazonas indicando a localização do município
de origem dos isolados de Plasmodium vivax caracterizados neste
estudo.......................................................................................................
32
Figura 10 Fluxograma do estudo, descrevendo com detalhes os pacientes
elegíveis, excluídos e CQRPv e susceptível à CQ analisados.................
34
Figura 11 Fluxograma do estudo, descrevendo com detalhes os pacientes
elegíveis, excluídos e pacientes graves e não graves analisados..............
35
ARTIGO
Figure 1
Expression levels of chloroquine resistance genes in patients with
CQRPv parasites……………………………………………………...
61
Figure 2 Expression gene levels of pvcrt-o and pvmdr-1 in all the groups……… 62
Figure 3 Expression level of chloroquine resistance genes in severe patients…... 63
Figure 4 Correlation between expression gene levels pvcrt-o and pvmdr-1 genes
and hemoglobin concentration………………………………………….
64
Figure S1 Expression levels of chloroquine resistance genes in patients
susceptible to chloroquine treatment……………………………………
68
Figure S2 Expression level of chloroquine resistance genes in non-severe P.
vivax malaria……………………………………………………………
69

xiii
LISTA DE TABELAS
Tabela 1 Revisão dos estudos que relacionaram mecanismos de resistência
molecular de cloroquina com Plasmodium vivax...................................
17
Tabela 2 Critérios de malária grave por P. falciparum......................................... 24
Tabela 3 Primers usados para sequenciamento do DNA de genes ortólogos de
P.vivax....................................................................................................
39
Tabela 4 Oligonucleotideos usados na genotipagem do P. vivax......................... 40
ARTIGO
Table 1
Demographic and clinical characteristics of the study participants
admitted to a tertiary health center, Manaus, Amazonas, Brazil……...
65
Table 2 Frequency of polymorphisms in the pvmdr1 and pvcrt-o genes for the
different set of patients………………………………………………...
66
Table 3 Description of 7 patients presenting severe vivax malaria admitted to
a tertiary health center, Manaus, Amazonas, Brazil…………………..
67
Table S1 Oligonucleotide primers used for DNA sequencing of P. vivax
orthologs genes………………………………………………………..
70
Table S2 Oligonucleotides used for genotyping P. vivax parasites……………... 71
Table S3 Reproducibility and repeatability of measurements of Cts
amplification of targets pvcrt-o and pvmdr-1…………………………
72
Table S4 Gene expression and different intra-erythrocytic stages of chloroquine
resistance P. vivax parasites admitted to a tertiary health center,
Manaus, Amazonas, Brazil…………………………………….
73
Table S5 Different intra-erythrocytic stages of chloroquine-susceptible P. vivax
parasites admitted to a tertiary health center, Manaus, Amazon,
Brazil…………………………………………………………………...
74
Table S6 Gene expression and different intra-erythrocytic stages of patients
presenting severe vivax malaria admitted to a tertiary health center,
Manaus, Amazonas, Brazil…………………………………………….
75

xiv
LISTA DE ABREVIATURAS, SÍMBOLOS E UNIDADES DE MEDIDA
ACT Artesunato
AE Eluição
AL Lise
AMA-1 Antígeno I Membrana Apical
ARDS Síndrome do desconforto respiratório agudo
ATL Lise dos tecidos
AW1 Lavagem 1
AW2 Lavagem 2
C Celsius
C14H14ClN3S Corante Giemsa em pó
CH3OH Álcool metílico
CI Intervalo de confiança
CIVD Coagulação intravascular disseminada
CQ Cloroquina
CQR Resistência à cloroquina
CQRPv P. vivax resistente à cloroquina
CRESIB Barcelona Centre for International Health Research
CSP Proteína do circunsporozoíto
CV Coeficiente de variação
D0 Dia da admissão
DARC Antígeno Duffy/receptor para quimiocinas
DCQ Desetilcloroquina
dL Decilitros
DNA Ácido desoxirribonucleico
DR Dia de recrudescência
DTM Domínios transmembranas
FMT-HVD Fundação de Medicina Tropical Dr. Heitor Vieira Dourado

xv
g Grama
H2O Água
HAV Vírus da hepatite A
HbsAg Antígeno de superfície da hepatite B
HCV Vírus da hepatite C
HDV Vírus da hepatite D
He Heterozigosidade
HIV Vírus da imunodeficiência humana
HPLC Cromatografia líquida de alta eficiência
16H18ClN3S Azul de metileno medicinal em pó
IC Concentração inibitória
Ig G Imunoglobulina G
Ig M Imunoglobulina M
K2HPO4 Fosfato de potássio bibásico
Kg Quilograma
KH2PO4 Fosfato monobásico de potássio
Km Quilômetro
mg Miligrama
Mg Magnésio
MgCl2, Cloreto de magnésio
min Minutos
mL Mililitros
mm Milímetro
MQ Mefloquina
MSP Proteína de superfície do merozoíto
Na2HPO4 Fosfato bibásico de sódio
NaH2PO4 Fosfato de sódio monobásico
NBD Domínio de ligação a nucleotídeo
ng Nanograma

xvi
NIB Número de identificação permanente
nM Nanomolar
NS Casos não graves
OMS Organização Mundial da Saúde
PAD Proteínas ligação Duffy
pb Pares de bases
PBS Solução salina
PCR Reação de polimerase em cadeia
PDF Produtos de degradação da fibrina
PVCRT-O Transportador de resistência à cloroquina do P. vivax
pvcrt-o Gene de resistência do P. vivax à cloroquina
PVMDR1 Transportador de resistência a multidrogas tipo 1
pvmdr-1 Gene de resistência do P. vivax à múltiplas drogas
PvRBP Proteínas de ligação de reticulócitos
R Resistente
RNA Ácido ribonucleico
rpm Rotação por minuto
rRNA Ácido ribonucleico ribossômico
RT Transcrição reversa
S Susceptível
SD Desvio padrão
seg Segundos
Sev Casos graves
SNP Polimorfismo de nucleotídeo único
TAP Tempo de atividade de protrombina
TCLE Termo de consentimento livre e esclarecido
TM Temperatura anelamento
TRAP Proteíína adesiva relacionada à trombospondina
TTPA Tempo da tromboplastina parcial ativada

xvii
UTI Unidade de terapia intensiva
% Percentual
µL Microlitros
< Menor
α Alfa
≥ Maior ou igual

xviii
SUMÁRIO
1. INTRODUÇÃO........................................................................................................
01
1.1 Ciclo biológico......................................................................................................... 02
1.2 Malária no mundo.................................................................................................. 05
1.3 Malária na Amazônia brasileira......................................................................... 07
1.4 Resistência à cloroquina...................................................................................... 08
1.4.1 Histórico................................................................................................................ 10
1.4.2 Mecanismos moleculares de resistência à cloroquina........................................... 12
1.5 Malária grave.......................................................................................................... 22
1.6 Malária vivax grave e resistência à cloroquina................................................... 25
1.7 Justificativa e hipóteses..........................................................................................
27
2. OBJETIVOS.............................................................................................................
29
2.1 Objetivo geral......................................................................................................... 30
2.2 Objetivos específicos..............................................................................................
30
3. MATERIAL E MÉTODOS.....................................................................................
31
3.1 Tipo de estudo...................................................................................................... 32
3.2 Área de estudo………………………………………………………………… 32
3.3 Seleção dos pacientes…………………………………………………………... 33
3.3.1 Seleção de pacientes resistentes à cloroquina.................................................... 33
3.3.2 Seleção de pacientes com malária grave e não grave por P. vivax..................... 34
3.4 Diagnóstico de Plasmodium vivax por gota espessa.......................................... 35
3.5 Caracterização genética....................................................................................... 36
3.5.1 Extração do DNA................................................................................................ 36

xix
3.5.2 Realização de PCR.............................................................................................. 36
3.5.3 Extração do RNA................................................................................................ 37
3.5.4 Expressão gênica de pvcrt-o e pvmdr-1............................................................ 37
3.5.5 Sequenciamento dos genes pvcrt-o e pvmdr-1.................................................... 38
3.5.6 Análise em marcadores moleculares, msp1F3 e MS2 em pacientes resistentes à
cloroquina.................................................................................................................
39
3.6 Aspectos éticos...................................................................................................... 40
3.7 Análise estatística................................................................................................. 41
4 RESULTADOS.......................................................................................................
42
ARTIGO......................................................................................................................
43
5 DISCUSSÃO............................................................................................................
76
5.1 Contribuição para a patogênese molecular da resistência à cloroquina e malária grave
por Plasmodium vivax........................................................................................
77
5.2 Perspectivas futuras................................................................................................
80
6 CONCLUSÕES………………………………………………………………… 83
7 REFERÊNCIAS BIBLIOGRÁFICAS..................................................................
85
8 ANEXOS
98
8.1 Anexo 1
PROCEDIMENTOS OPERACIONAIS PADRÃO………………………………
99
8.2 Anexo 2
APROVAÇÃO DO COMITÊ DE ÉTICA EM PESQUISA COM SERES
HUMANOS...........................................................................................................................
144
8.3 Anexo 3

xx
TERMO DE CONSENTIMENTO LIVRE E ESCLARECIDO.....................................
8.4 Anexo 4
145
ARTIGOS PUBLICADOS NA VIGÊNCIA DO CURSO............................................... 149

1
1. INTRODUÇÃO

2
1.1 Ciclo biológico
Os agentes etiológicos da malária humana são protozoários pertencentes à classe Sporozoa,
família Plasmodiidae e ao gênero Plasmodium. São conhecidas cinco espécies: Plasmodium
malariae Laveran, 1881, Plasmodium falciparum Welch, 1897, Plasmodium vivax Grassi e
Feletti, 1890, Plasmodium ovale Stephens,1922 e Plasmodium knowlesi Knowles e Das.
Gupta, 1932 (1). P. falciparum é o responsável pelas formas mais graves e complicadas da
doença e é encontrado mais comumente nas regiões tropicais (2). Mais de 75 % dos episódios
clínicos causado pelo P. falciparum por ano concentra-se na África (3). O P. vivax é,
entretanto, o mais amplamente distribuído pelas zonas tropicais e subtropicais do globo (4).
Os transmissores da malária são culicídeos do gênero Anopheles, dos subgêneros:
Anopheles, Cellia, Nyssorhynchus e Kerteszia. Existem cerca de 500 espécies de mosquitos
do gênero Anopheles no mundo, mas somente em torno das 70 delas são vetores sob
condições naturais e destes 30 apresentam importância epidemiológica. No Brasil, as espécies
mais importantes na transmissão de plasmódios são: Anopheles (Nyssorhjncus) darlingi,
Anopheles (N.) albitarsis, Anopheles (N.) deaneorum, Anopheles (N.) aquasalis, Anopheles
(Kerteszia) cruzi e Anopheles (K.) bellator (5).
A biologia dos plasmódios inclui um ciclo de vida em um hospedeiro vertebrado
intermediário onde ocorre a fase assexuada e um hospedeiro definitivo invertebrado onde
ocorre a fase sexuada (Figura 1) (6). Inicialmente, os esporozoítos são transmitidos ao
hospedeiro humano, através da saliva da fêmea do anofelino durante o repasto sanguíneo. Na
picada são depositados aproximadamente de 15 a 200 esporozoítos, os quais podem
permanecer na área por várias horas até atingirem a corrente sanguínea. Aproximadamente,
70% dos esporozoítos alcançam a circulação sanguínea; 30% restante invadem o sistema
linfático e apesar de alguns conseguirem se desenvolver em formas similares às formas
exoeritrocíticas, são fagocitados por células dendríticas (7). Depois de atravessar os vasos
sanguíneos da pele, os esporozoítos são transportados pela corrente sanguínea para o fígado,
invadem os hepatócitos e se diferenciam em milhares de merozoítos (10.000 a 15.000 para P.
vivax e 40.000 para P. falciparum) (8). Este processo envolve proteínas de superfície do

3
esporozoíto e moléculas do hospedeiro humano e leva em torno de 4 semanas. Duas proteínas
principais da superfície dos esporozoítos, a proteína do circunsporozoíto (CSP) e a proteína
adesiva relacionada à trombospondina (TRAP) ligam-se à superfície dos hepatócitos através
da heparan-sulfato das proteoglicanas durante a invasão (9). Esse processo resulta na
formação de um vacúolo parasitóforo bem delimitado onde os esporozoítos se diferenciam em
trofozoítos que, após sofrerem várias divisões por esquizogonia, formam os esquizontes. Após
alguns dias, os esquizontes maduros liberam os merozoítos por um processo de brotamento
denominado merossomo (10). Ainda no fígado, P. vivax e P. ovale evoluem para uma forma
conhecida como hipnozoíto (11). Estas formas permanecerão “adormecidas” durante semanas,
meses ou mesmo anos, até aa sua reativação, quando completarão o ciclo hepático, dando
origem às recaídas (11). Os merozoítos são liberados para os sinusóides hepáticos envolvidos
por uma estrutura vesicular denominada merossomo, cuja superfície possui moléculas que
interagem com o endotélio dos sinusóides hepáticos. Têm a capacidade de não serem
reconhecidos pelo sistema imune do hospedeiro, permanecendo intactos por pelo menos uma
hora. Sem o reconhecimento pelas células fagocitárias, os merozoítos, após romperem a
membrana do merossomo, são libertados na circulação para invasão dos eritrócitos iniciando
o ciclo eritrocítico (12).
A invasão dos eritrócitos é um processo crucial à sobrevivência do parasita e envolve uma
sequência de eventos: ligação à superfície dos eritrócitos, reorientação, formação de uma
adesão fixa e finalmente a internalização (13). Para que os merozoítos invadam os eritrócitos
são também necessárias interações específicas envolvendo proteínas do parasito e receptores
presentes na superfície da hemácia (14). Os receptores para a invasão são encontrados nos
micronemas e nas roptrias, responsáveis pela mobilidade dos parasitos tanto no deslizamento
quanto na migração e invasão (15). Várias proteínas de superfície do merozoíto (MSPs) (11,
16) e duas proteínas de ligação ao reticulócitos (PvRBP-1 e PvRBP-2) (17) têm sido
caracterizadas no P. vivax. O parasito utiliza também o antígeno Duffy/Receptor para
quimiocinas (DARC) para penetrar nas hemácias através de proteínas ligação Duffy (PAD)
como ligantes (18). Este antígeno não ocorre na maioria dos seres humanos na África
Ocidental (fenótipo Fyab) e, como resultado P. vivax ocorre com menor frequência (19).
Quando os merozoítos invadem as hemácias sua extremidade apical contendo organelas
secretoras especializadas (micronemas, roptrias e grânulos densos) deve estar apontada para o
eritrócito. A interação do merozoíto com os eritrócitos leva à formação de uma junção de

4
células parasito-hospedeiro. Embora este processo ainda não seja bem compreendido, parece
haver a participação do antígeno I da membrana apical (AMA-1) (14, 20). Quando sua
entrada for concluída, o orifício de invaginação é fechado e a junção torna-se então parte do
vacúolo parasitóforo, criado pelo parasito, e que separa o citoplasma da célula hospedeira,
formando assim um ambiente favorável ao desenvolvimento (14, 20).
Os merozoítos liberados, provavelmente na forma de pequenas vesículas conhecidas como
merossomos, vão invadir as hemácias desenvolvendo-se em trofozoítos que, por divisão
celular, formam os esquizontes sanguíneos, as formas do plasmódio que incitam resposta
imunitária com maiores repercussões clínicas (11). Após algumas gerações de
desenvolvimento de merozoítos sanguíneos, alguns parasitos se diferenciam em estágios
sexuais - gametócitos masculinos e femininos – que ingeridos por mosquitos do gênero
Anopheles, inicia-se um processo dentro do lúmen do intestino médio de exflagelação que
resulta na libertação de quatro a oito microgametas nucleados. Ocorre fecundação do
macrogameta pelo microgameta com a consequente formação da única forma diplóide, o
zigoto (11). O zigoto resultante alonga-se e após divisão meiótica, por um período que varia
entre 18-24 horas dá origem a uma forma móvel e haplóide denominada oocineto que penetra
nas células epiteliais do intestino médio e se instala na superfície exterior junto à membrana
basal, onde se transforma em oocisto. Os oocistos multiplicam-se por um processo de divisão
assexuada, levando à liberação de esporozoítos que migram através do sistema hemolinfático
até alcançarem as glândulas salivares do mosquito para que esse, ao exercer o repasto
sanguíneo, reinicie um novo ciclo no hospedeiro vertebrado (11). Este processo de ruptura do
oocisto, migração dos esporozoítos através do hemocele, para as glândulas salivares do
mosquito, dura aproximadamente sete a oito dias (21).

5
Figura 1- Ilustração do ciclo biológico do plasmódio. Adaptada de Mueller et al., 2009 (22).
1.2 Malária no mundo
A malária continua sendo um grande problema de saúde pública em muitos países do
mundo, ocorrendo em mais de 104 países e territórios (23). Distribui-se por todos os
continentes, onde 40% da população mundial está exposta à infecção principalmente as
pessoas que vivem nas regiões tropicais e subtropicais. Há de 135 a 287 milhões de novos
casos de malária, no mundo, a cada ano, e estes resultam em 0,4 a 1,0 milhões de mortes,
principalmente entre crianças jovens, gestantes e adultos não-imunes (24). Em 2012, foram
registrados 207 milhões de episódios e 627.000 mortes por malária (23). Sendo o continente
africano o que apresenta o maior porcentual de mortalidade, associada à malária cerebral e
anemia grave, principalmente em crianças menores de cinco anos (19). Os países mais
afetados são os africanos, situados ao sul do deserto do Saara, os do Sudeste Asiático e os da
América Latina, particularmente os situados na região da Bacia Amazônica (25). Quase 60%

6
dos casos de malária nas Américas são notificados no Brasil e outros 40% são relatados da
Colômbia (14,2%), Peru (8,8%), Venezuela (5,4%), Bolívia (1,9%) e Equador (1,1%). Países
da América Central relataram a ocorrência de casos na Guatemala (3,8%), Panamá (0,4%) e
Honduras (1,5%) (26).
Atualmente, P. vivax é a espécie de plasmódio mais amplamente distribuída no mundo
(Figura 2). Estima-se que cerca de 2,85 bilhões de pessoas estão sob o risco de infecção nas
diferentes áreas endêmicas do planeta (19). As maiores prevalências são observadas no sul e
leste da Ásia (52%), leste do Mediterrâneo (15%) e América do Sul (13%). No leste e sul da
África, 5% das infecções por malária são atribuídas a essa espécie, que pode representar entre
seis e quinze milhões dos casos por ano (27, 28). Na América, a malária ocorre em 23 países,
com 20 % da população em risco. Em geral, P. vivax representou 80% de todos os casos
notificados em 2009 (29).
Água
P. vivax livre
P. vivax IPA < 0.1‰
P. vivax IPA ≥ 0.1‰

7
Figura 2- Transmissão da malária por Plasmodium vivax no mundo. Adaptada de Gething
(30).
1.3 Malária na Amazônia Brasileira
Nas Américas, a malária é endêmica em 21 países, acometendo quase três em cada 10
pessoas com diferentes graus de risco de transmissão. Em 2012, nas Américas foram
registrados 469.000 casos de malária e 108 mortes (24).O Brasil é o país com maior número
de casos, 99,7% oriundos da Região Amazônica, onde a doença acomete indistintamente
crianças, adolescentes e adultos. De 2000 a 2011, foram registrados em média de 422.858
casos de malária causada por essa espécie, sendo 78,7% causado pelo P. vivax (31).
No Brasil, a relação entre a incidência de P. falciparum e P. vivax, conhecida como
fórmula parasitária, tem se alterado ao longo dos anos, ora predominando uma, ora
predominando outra espécie, em função da maior ou menor efetividade das atividades de
controle executadas. De um modo geral, predominou P. vivax quando as ações de controle
foram mais efetivas (32). A Amazônia Legal, composta pelos estados Acre, Amapá,
Amazonas, Pará, Rondônia, Roraima, Maranhão, Mato Grosso e Tocantins, é a principal
região de ocorrência da malária. Em 2010, foram registrados 132.179 de casos nessa região,
sendo o Amazonas (33.948 casos), Pará (30.065 casos), Rondônia (16.401 casos) os
responsáveis por 61% deste total (33).
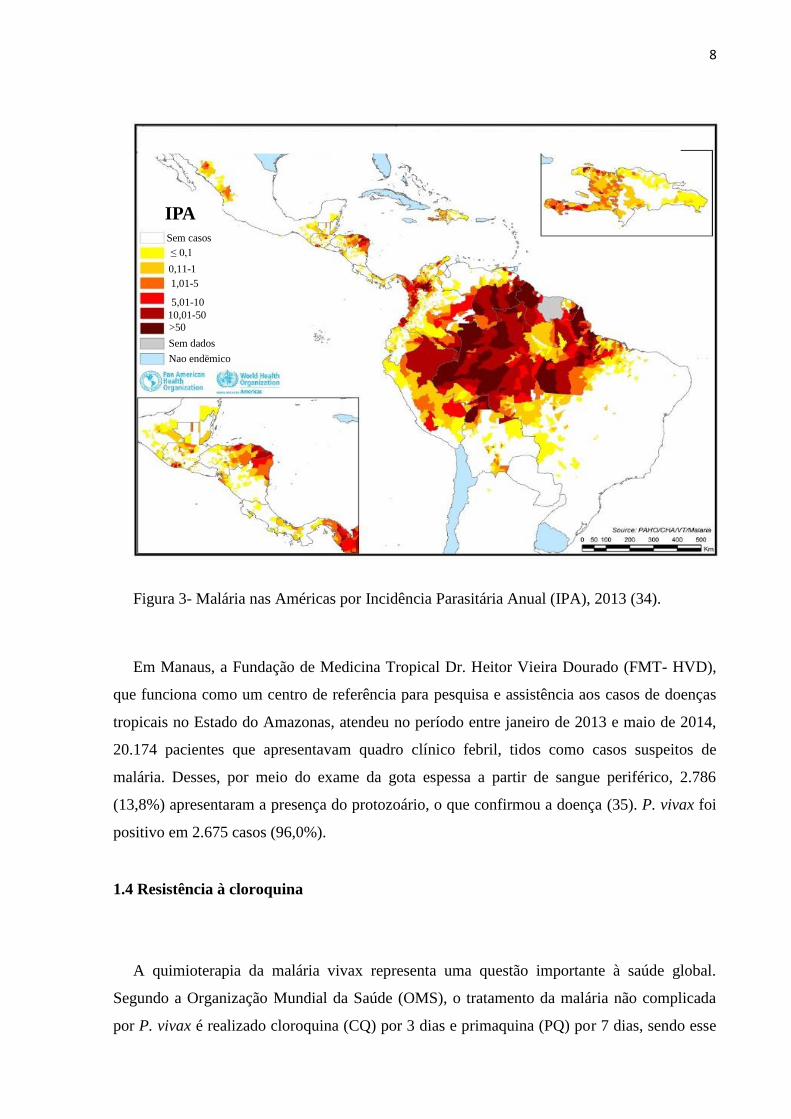
8
Figura 3- Malária nas Américas por Incidência Parasitária Anual (IPA), 2013 (34).
Em Manaus, a Fundação de Medicina Tropical Dr. Heitor Vieira Dourado (FMT- HVD),
que funciona como um centro de referência para pesquisa e assistência aos casos de doenças
tropicais no Estado do Amazonas, atendeu no período entre janeiro de 2013 e maio de 2014,
20.174 pacientes que apresentavam quadro clínico febril, tidos como casos suspeitos de
malária. Desses, por meio do exame da gota espessa a partir de sangue periférico, 2.786
(13,8%) apresentaram a presença do protozoário, o que confirmou a doença (35). P. vivax foi
positivo em 2.675 casos (96,0%).
1.4 Resistência à cloroquina
A quimioterapia da malária vivax representa uma questão importante à saúde global.
Segundo a Organização Mundial da Saúde (OMS), o tratamento da malária não complicada
por P. vivax é realizado cloroquina (CQ) por 3 dias e primaquina (PQ) por 7 dias, sendo esse
IPA
Sem casos
≤ 0,1
0,11-1
1,01-5
5,01-10
10,01-50
>50
Sem dados
Nao endëmico

9
o esquema terapêutico realizado no Brasil. Porém, em regiões onde a resistência à CQ é
extensa, o tratamento é realizado com arteméter/lumefantrine por 3 dias e PQ por 7 dias. No
caso da malária vivax grave é utilizado artesunato (ACT) ou arteméter por 7 dias
complementado com clindamicina. (36).
O ciclo de vida dos plasmódios define estratégias no uso dos quimioterápicos. Este
parasito passa por um ciclo de vida complexo marcado por morfologia distinta, função,
localização, consequência clínica e a susceptibilidade a agentes antimaláricos. A CQ age
contra formas sanguíneas de P. vivax e P. falciparum. Sua atividade como gametocitocida é
inexistente contra gametócitos de P. falciparum, mas potente para P. vivax, porém a CQ não
exerce função conhecida como esquizonticida tecidual. (37).
Um parasito é considerado resistente quando é capaz de sobreviver e de multiplicar-se, em
sua fase sanguínea assexuada, apesar da administração e absorção adequada de um regime
quimioterápico (38). Resistência confirmada in vivo à CQ é definida como a persistência de
estágios sanguíneos assexuados do P. vivax com níveis adequados de CQ e seu principal
metabólito ativo, desetilcloroquina (DCQ) no sangue total ou plasma (39). Falha terapêutica
foi definida como incapacidade de eliminar a parasitemia da malária ou diminuir os sintomas
clínicos da doença, apesar do uso de antimalárico. Muitos fatores podem contribuir para a
falha terapêutica: resistência ao medicamento, dosagem incorreta, falta de adesão do paciente,
má qualidade do medicamento, interações medicamentosas e má absorção do antimalárico
(40).
A metodologia para o diagnóstico de resistência à CQ lidou com o problema por ter que
distinguir recrudescência da recaída ou reinfecção. A identificação da resistência à CQ
envolve a dosagem da CQ e de seu metabólito, DCQ, no sangue do paciente no dia da
recrudescência. Para ser considerado como resistência, os níveis de CQ+DCQ devem
ultrapassar a concentração mínima efetiva de 100 ng/mL (39), concentração eficiente para
eliminar parasitas sensíveis, sejam originados de recrudescências, recaídas ou reinfecções. Na
recrudescência, os parasitas que sobrevivem ao tratamento da infecção são mantidos na

10
corrente sanguínea não detectáveis pela gota espessa e reaparecem dias depois na circulação
sanguínea.
1.4.1 Histórico
O químico alemão Hans Andersag desenvolveu e patenteou, em 1934, a molécula
conhecida como “resochin” e um derivado que chamou de sontochin. A sua estrutura química
possui uma base comum: 4-aminoquinoleína apontada como responsável pela ação
terapêutica na fase esquizonte do plasmódio (41). Durante a Segunda Guerra Mundial, a
molécula sintética resochin ganhou outro nome, CQ (Figura 4). O primeiro relato de
tratamento com CQ ocorreu em 1936 em quatro pacientes com sífilis na Alemanha, que
tinham P. vivax (42). No mesmo período, outra substância marcou fortemente a história dos
medicamentos antimaláricos, a PQ, uma 8-aminoquinoleína, desenvolvida por pesquisadores
norte-americanos. No entanto, prevaleceria o uso da CQ, em virtude de sua baixa toxicidade,
eficácia contra todas as espécies de plasmódios, poucos efeitos colaterais para grávidas e
crianças, grande eficácia em comparação com a PQ que causa efeitos colaterais indesejáveis
quando administrada em altas doses curativas e o baixo custo. A CQ foi formalmente
registrada em 1946 nos Estados Unidos e um mês depois Loeb et al.(43) publicou o trabalho
sobre a atividade da CQ contra P. falciparum e P. vivax. Tornou-se o fármaco mais popular e
extensivamente utilizado em praticamente todas as regiões endêmicas de malária.
Figura 4- Estrutura molecular da cloroquina (38).
Atualmente, a utilização de fármacos antimaláricos representa a solução mais eficaz no
controle da malária. No entanto, no decorrer das últimas décadas, o aparecimento e
propagação de parasitos resistentes a maior parte dos antimaláricos disponíveis têm revelado

11
como o principal obstáculo a uma eficiente contenção da malária. A resistência de P. vivax à
CQ foi relatada pela primeira vez em 1989 na Papua Nova Guiné (44) e Indonésia , em 1991.
A resistência à CQ tem sido relatada em muitos países (Figura 5), incluindo Índia (45),
Mianmar (46), América Central e do Sul (37). Um estudo clínico em isolados da fronteira da
Tailândia com Mianmar indicou que a CQR parecia estar surgindo nesta área (47). Foi
também relatada susceptibilidade reduzida à mefloquina (MQ) , amodiaquina e ACT (48). Na
Coréia, a CQ continua sendo a droga de escolha para o P. vivax (49), embora alguns casos
resistentes à CQ foram confirmados recentemente (50).
Figura 5- Distribuição mundial de falha terapêutica e resistência confirmada de P. vivax à
cloroquina. Adaptada de WHO, 2010. (40)
Na América latina, os primeiros casos de resistência à CQ foram registrados na Colômbia
(51) e Brasil (52) (Figura 6). No Brasil, em 1992, dois autores italianos afirmaram ter
identificado o primeiro caso de P. vivax resistente à CQ (52). Logo depois, no entanto, ficou
claro que não era um caso de recrudescência verdadeira (53). O primeiro caso confiável de P.
vivax resistente à CQ in vivo foi a partir de um paciente tratado na Fundação de Medicina
Tropical Dr. Heitor Vieira Dourado (FMT-HVD), em Manaus, na Amazônia Ocidental

12
Brasileira (54). Em um estudo recente, com acompanhamento clínico por 42 dias e dosagem
de CQ plasmática em 109 pacientes com diagnóstico de P. vivax, detectou-se resistência in
vivo à CQ em 10,1% dos casos (55). Em 2014, foi verificado que 5,2 % dos pacientes com
acompanhamento clínico por 28 dias apresentam resistência in vivo (56).
Figura 6- Distribuição de estudos in vivo de eficácia da cloroquina para Plasmodium vivax
entre 2000 e 2013 na América do Sul. Triângulos representam locais em que foi reportado
falha terapêutica. Quadrados representam locais em que não foi observado falha terapêutica
após 28 dias de acompanhamento (57).
1.4.2 Mecanismos moleculares de resistência à CQ
Os mecanismos moleculares de resistência à cloroquina permanecem desconhecidos, já
que a falta de cultivo in vitro para P. vivax tem dificultado os estudos. Para avaliar a
sensibilidade aos antimaláricos utilizam-se ensaios in vivo e in vitro. Os ensaios clínicos são
muito úteis, porém estão sujeitos a variações no estado imunológico dos pacientes,
metabolismo da droga e qualidade da droga (58). No entanto, os ensaios in vitro permitem

13
avaliar a atividade das drogas na ausência dos fatores de confusão do hospedeiro (59) mas
ainda não estão padronizados e validados. Por isso, o mecanismo de ação da CQ para P. vivax
permanece desconhecido.
Foi descrito, recentemente, que o gene pfcrt tem um papel central na resistência à CQ
pelo P. falciparum (60). Vários trabalhos têm sido realizados associando genes de resistência
do P. falciparum com genótipo de resistência (61-64). A PfCRT é uma proteína
transmembrana que está localizada no vacúolo digestivo do parasito, local de ação da CQ
(65). As mutações pontuais na proteína foram associadas com resistência in vitro na Ásia,
África e América do Sul. A importância destas mutações foi reforçada ainda mais pelos
resultados de transformação genética e por um estudo no oeste africano que mostrou que o
alelo mutante K76T do pfcrt estava presente em 100% dos pacientes com falha terapêutica,
em comparação com prevalência de 40% na população em geral (66).
A proteína PfMDR (transportador de resistência a multidrogas), codificada pelo gene
pfmdr1 tem sido associada com resistência a diversos antimaláricos. O primeiro estudo
verificou que mutantes de PfMDR1 associados ao fenótipo de resistência à CQ, produziam
mudanças no pH do lisossomo e diminuição no acúmulo da CQ, indicando que mutações no
gene pfmdr1 poderia alterar a função desta proteína (67, 68). Outros estudos verificaram que
polimorfismos neste gene pode alterar a função da proteína e modular a resistência à CQ em
P. falciparum (69, 70). Além disso, o aumento no número de cópias de pfmdr1 tem sido
relacionado com resistência à CQ (71) e MQ (72).
Os medicamentos para o tratamento da malária causada pelo P. vivax foram desenvolvidos
para o tratamento do P. falciparum, com base no pressuposto de que os mecanismos de ação e
de resistência eram semelhantes em ambas as espécies. Assim, os mecanismos moleculares
estudados até o momento baseiam-se em dois genes ortólogos do P. falciparum: pfmdr1 e
pfcrt.
Em 2001, Nomura et al. (73) descreveram o gene ortólogo de pfcrt em P. vivax,
denominado pvcrt-o. A proteína codificada por esse gene apresenta 10 domínios

14
transmembrana (DTM) e faz parte da família de transportadores de drogas (74) (Figura 7).
Alguns estudos não encontraram nenhuma relação entre mutações no gene pvcrt-o e fenótipo
de resistência à CQ (58, 73, 75). Nomura et al. (73) demonstraram que não existia associação
entre SNP de pvcrt-o e a resistência à CQ sugerindo que o mecanismo de resistência difere do
P. falciparum. Linhagens transgênicas de P. falciparum superexpressando pvcrt-o
apresentaram redução significativa de 2,2 vezes na sensibilidade à CQ (76). Níveis mais
elevados de expressão de pvcrt-o foram encontrados em paciente com malária vivax grave em
comparação a três pacientes não-graves (77). Apesar desse único paciente grave não ter sido
tratado com cloroquina, a maior expressão de pvcrt-o sugere que a gravidade pode estar
relacionada com a resistência à CQ tendo esse gene como marcador. Estes resultados sugerem
o envolvimento de outros mecanismos para explicar a resistência à CQ em P. vivax, sendo a
expressão gênica provável candidato (77).
Figura 7 - Estrutura do transportador PvCRT-O (60).
Em 2005, Brega et al. (78) descreveram o gene ortólogo de pfmdr1 em P. vivax,
denominado pvmdr1. Este gene assim como pfmdr1 possui um único éxon com 4395 pb que
codifica uma proteína de 465 aminoácidos. A proteína PvMDR1 possui 2 DTM hidrofóbicos
que atravessam a membrana várias vezes (Figura 8). Cada DTM consiste de 6 segmentos
transmembranas (α-hélices hipotéticas) com um total de 12 segmentos por molécula de
PvMDR1. Os outros dois domínios são hidrofóbicos e possuem um domínio de ligação a
nucleotídeo (NBD do inglês Nucleotide Binding Domain) que acopla a energia da hidrólise da
adenosina trifosfato (ATP) ao processo de transporte. Todas essas características estruturais
classificam a PvMDR1 como membro da superfamília de proteínas transportadoras ABC.

15
Figura 8 - Estrutura do transportador PvMDR1 (60).
Vários trabalhos têm procurado demonstrar a resistência à CQ pelo P. vivax (Tabela 1).
Foram estudados 10 isolados de diferentes regiões do mundo, onde se verificou que não
houve associação entre os polimorfismos achados e o fenótipo de resistência à CQ (79). A
mutação Y976F em pvmdr1 tem sido associada com CQR no Sudeste Asiático (58) e Papua
Nova Guiné (80), mas para outras regiões não foi observado o mesmo fenômeno. Suwanarusk
et al. (58) verificaram que 95% dos isolados da Indonésia tinham a mutação Y976F no gene
pvmdr1 em comparação com 25% dos isolados tailandeses, sendo o IC50 foi maior nos
isolados com mutação Y976F quando comparados com isolados do tipo selvagem. Foi
verificado que número de cópias de pvmdr1 estava aumentada em 21,0% dos isolados da
Tailândia e em nenhum da Indonésia. Em 2008, Imwong et al. (81) demonstraram que a
amplificação do pvmdr1 foi significativamente maior nas amostras do oeste da Tailândia onde
a MQ tem sido amplamente utilizada para o tratamento do P. falciparum.
No Brasil, a caracterização molecular de isolados de P. vivax também foi realizada (55, 60,
79, 82, 83). A análise completa dos códigos e sequências não-codificantes dos genes de
resistência pvcrt-o e pvmdr1 foi realizado em sete amostras e não foi encontrada nenhuma
associação com resistência à CQ (60). Gama et al. (82) identificaram uma potencial mutação
em pvmdr1 em 100% dos isolados examinados, sendo que contrasta com o relatado em 10%
das falhas no tratamento (55). Vargas-Rodrigues et al. (84) observaram presença de apenas 2

16
isolados com número de cópias aumentado em pvmdr1 onde CQ e MQ são usados no
tratamento da malária.

17
Tabela 1- Revisão dos estudos que relacionaram mecanismos de resistência molecular de Plasmodium vivax à cloroquina.
Referência Procedência Tipo de estudo Resultado encontrado
Picot et al. (85) Guiana Francesa Relato de caso Não foi encontrado mutação no gene
pvmdr1 para MQ.
Sa et al. (79) Brasil, El Salvador, Papua Nova
Guiné, Indonésia e Papua
Ensaios in vitro Não houve associação entre mutações
nos genes pvmdr1 e pvcrt-o e
resistência à CQ.
Sa et al. (76) Brasil Sistema de expressão heterólogo Linhagens transgênicas de P.
falciparum superexpressando pvcrt-o
com redução significativa de 2,2
vezes na sensibilidade à CQ.
Suwanarusk et al. (58) Papua e Tailândia Ensaio in vitro 95% dos isolados da Indonésia
tinham a mutação Y976F no gene
pvmdr1 em comparação com 25%
dos isolados tailandeses e o IC50 foi
maior nos isolados com mutação
Y976F quando comparados com
isolados do tipo selvagem.
Barnadas et al. (75) Madagascar Ensaio clínico Encontrado 5 mutações descritas em
estudos anteriores (G698S, M908L,

18
T958M, Y976F e F1076L) e cinco
novas mutações (F194Y, S510T
,S513R , I636T e A829V )
Suwanarusk et al. (86) Tailândia e Indonésia Ensaios in vitro Aumento do número de cópias de
pvmdr1 em 21% dos isolados da
Tailândia e nenhum da Indonésia. A
mutação Y976F de pvmdr1 foi
associada com diminuição da
sensibilidade à CQ.
Imwong et al. (81) Laos, Myanmar e Tailândia Estudo transversal Amplificação do gene pvmdr1 foi
significativamente mais comum na
Tailândia do que no resto do Sudeste
Asiático
Fernandez-Becerra et al. (77) Manaus, Brasil Relato de caso Níveis aumentado de expressão no
gene pvcrt-o e pvmdr1 foi encontrado
em paciente com malária grave por P.
vivax em comparação com pacientes
não graves.
Orjuela-Sanchez et al.(60) Manaus, Brasil Ensaio clínico Mutação nos genes pvmdr1 e pvcrt-o
revelaram nenhuma associação com
resistência à CQ.

19
Gama et al. (82) Pará, Brasil Estudo transversal A análise do gene pvmdr1 revelou a
existência de único (85,7%) e duplo
haplótipos mutantes (14,3%)
Dharia et al. (87) Iquitos, Peru Estudo transversal Foi encontrado mutações M908L e
T958M no gene pvmdr1
Lu et al. (88) Coréia, Tailândia, Myanmar e Papua
Nova Guiné
Estudo transversal e ensaio in vitro Mutações na posição de aminoácido
Y976F de pvmdr1 foram encontrados
somente nos isolados da Tailândia.
Número de cópias deste gene foi
único nos isolados da Coréia e duplo
em dois isolados tailandeses.
Jovel et al. (89) Honduras Estudo transversal Foi encontrado mutação Y976F no
gene pvmdr1 e presença de duas
cópias de pvmdr1 em 2,7 % amostras.
Rijken et al. (90) Tailândia Relato de caso Todos os parasitas tinham uma única
cópia de pvmdr1 e uma mutação
T958M e os parasitas dos D21, D49,
D126 e D143 mutação Y976F foi
observada.
Ranjitkar et al. (91) Nepal Estudo transversal Ocorreu baixa prevalência de
mutação Y976F no gene pvmdr1

20
(5%)
Lu et al. (92) China Estudo transversal Não foi encontrado nenhuma mutação
que confere resistência nos genes
pvmdr1 e pvcrt-o.
Vargas-Rodrigues et al. (84) Acre e Amazonas, Brasil Estudo transversal Foi observado que 2 amostras
apresentaram número de copias duplo
em pvmdr1.
Chehuan et al. (83) Manaus, Brasil Ensaio in vitro Nenhum marcador molecular
(pvmdr1 e pvmrp1) foi associado com
resistência à CQ.
Lin et al. (93) Tailândia e Camboja Estudo transversal Isolados do Camboja apresentaram a
mutação Y976F no gene pvmdr1
associada à resistência à CQ e
isolados da Tailândia apresentaram
aumento no número de cópias de
pvmdr1.
Marques et al. (56) Brasil Estudo in vivo Mutações genéticas no gene pvmdr1
não estavam relacionados com a
resistência in vivo.
Khattak et al. (94) Índia Estudo transversal 98% das amostras tinham a mutação

21
no códon F1076L e número de cópias
único do gene pvmdr1.
Ganguly et al. (95) Índia Ensaio clinico Não foi encontrada nenhuma mutação
no gene pvcrt-o. No gene pvmdr1
foram encontradas 8 mutações não
sinonímias (R88Q, A296V, E478G,
S513R, G698S, M908L, T958M e
F1076L).
Aguiar et al. (96) Brasil Ensaio in vitro A mutação Y976F não foi encontrada
no estudo no pvmdr1.
Kebede et al. (97) Etiópia Estudo transversal A mutação Y976F foi observada em
32,6% das amostras clínicas e o
F1076L não foi observada.
Shalini et al. (98) Índia Ensaio clínico e in vitro No estudo in vivo, 93,3% dos
pacientes apresentaram o haplótipo
mutante 958M/Y976/1076L e 6,7%
958M/Y976/F1076. No estudo in
vitro, 88,9% tinham o haplótipo
mutante 958M/Y976/1076L e 11,1%
o haplótipo 958M/Y976/F1076.

22
1.5 Malária grave
A maioria dos casos de malária grave é ocasionada pelo P. falciparum e o resultado é o
grande número de estudos realizados na África, onde o número de casos de gravidade por P.
falciparum é bem relatado e uma escassez de trabalhos focando a gravidade da malária por P.
vivax. No entanto, P. vivax representa a segunda espécie mais prevalente, responsável por 25-
40% dos casos clínicos da malária (99). Em regiões endêmicas, uma limitação em estudar
malária grave por P. vivax seria a inadequada exclusão de comorbidades e utilização de
métodos diagnósticos adequados. Diversos estudos têm demonstrado que a malária grave por
P. vivax tem sido relatada em diversas áreas endêmicas como Sudeste Asiático (100, 101),
América Latina (102, 103) e Índia (104, 105).
Por muitos anos, a malária grave foi retratada como duas síndromes principais: anemia
grave causada pela destruição das hemácias e malária cerebral causada pela obstrução de
pequenos vasos do cérebro por parasitos isolados. Uma ampla gama de manifestações graves
tem sido relatada, algumas das quais foram associados à morte subsequente. Manifestações
graves da malária vivax que já foram relatadas são: malária cerebral, anemia grave, ruptura do
baço, insuficiência renal aguda e síndrome da angústia respiratória aguda (104, 106).
A anemia e as complicações respiratórias são as manifestações clínicas mais comuns
relatadas em diferentes áreas endêmicas para P. vivax (104, 107, 108). Entre 1935 e 1947
foram relatados 900 casos fatais de malária por P. vivax na União Soviética (109). Um caso
de anemia, cardiomegalia, hepatomegalia e insuficiência renal em paciente com malária vivax
foi relatado em Los Angeles (110). Em 1996, Lakhkar et al. (111) relataram um caso de
coagulação intravascular disseminada em uma criança. Na Papua, Indonésia, a mortalidade
entre os internados com P. vivax foi de 0,8-1,6%, similar à observada em pacientes com P.
falciparum (1,6-2,2%) (100, 112). Vários estudos realizados na Índia têm demonstrado casos
de malária grave (104, 105, 113).

23
A ocorrência de casos complicados de infecção por P. vivax no Brasil tem sido relatada de
forma esporádica, contudo, crescente (114-120). Em Manaus, Djalma Batista descreveu uma
série de casos de malária com esplenomegalia e caquexia (121). Em 2000, em um estudo
realizado em Manaus, foi verificado que dos 73 pacientes internados, 90,5% apresentaram
plaquetopenia e destes 17,8% plaquetopenia grave, com níveis de plaquetas abaixo de
20.000/µL, e foi encontrado também um caso de coma por malária vivax (122). Em 2004, foi
verificado o caso de um paciente com púrpura trombocitopênica secundária à malária vivax
atendido na FMT-HVD (123). Em 2010, foram relatados icterícia e anemia grave (102).
Dados de nosso grupo demonstraram que crianças que apresentaram algum critério de
gravidade pela OMS tinham mais chance de serem internadas na Unidade de terapia intensiva
(UTI). Em 2012, em estudo realizado em autópsias, 13 das 17 mortes em que havia
diagnóstico antemortem de malária vivax, P. vivax estava relacionado ao desfecho fatal.
A fisiopatologia da malária grave tem sido melhor descrita para P. falciparum. A sua
virulência tem sido atribuída a altas parasitemias e à capacidade de citoaderência no endotélio
vascular ocasionando obstrução de capilares de importantes órgãos como cérebro, pulmão,
fígado e rins, cuja anóxia é a base fisiopatogênica do quadro clínico da malária grave (124).
Os eritrócitos parasitados podem também aderir a eritrócitos não-parasitados formando
rosetas, o que está associado à malária grave (125). Para P. vivax, as complicações clínicas
têm sido atribuídas a uma diversidade de fatores: intensidade da transmissão, presença de
comorbidades, fatores genéticos e resistência a drogas, porém a patogênese ainda continua
mal compreendida. Foi verificado que as hemácias infectadas apresentaram citoaderência in
vitro em receptores endoteliais conhecidos por mediar a citoaderência de P. falciparum (126,
127). Alguns estudos sugerem que as alterações pulmonares no P. vivax poderia ser causado
por sequestro dos parasitas dentro deste órgão (128). Foi verificado que o P. vivax tem a
capacidade de aderir a sulfato de condroitina A e o ácido hialurônico na placenta (126, 129,
130). Foi constatado que anticorpos policlonais contra proteínas recombinantes (VIRE4 e
VIRA5) inibiram a adesão ex vivo do P. vivax (126). Além disso, a formação de rosetas
poderia influenciar o fenômeno da citoadesão, porém nenhum trabalho demonstrou associação
entre formação de rosetas e gravidade da doença (131, 132). A grande dificuldade para
estudar a patogênese da malária grave por P. vivax seria a inexistência de um sistema de

24
cultivo contínuo, apesar dos avanços recentes (132), e não há biomarcadores específicos e
confiáveis para predição da gravidade.
Os critérios de gravidade para o P. falciparum foram estabelecidos pela OMS, em 1990,
sendo revisados e publicados novamente em 2012 (Tabela 2). Trata-se de um conjunto de
sinais e sintomas clínicos e resultados de exames laboratoriais que estão associados à maior
letalidade (17). Apesar desses critérios não terem sido originalmente validados para a infecção
por P. vivax, a maior parte das complicações observadas na literatura associadas a esse
parasito se encaixam em algum dos critérios.
Tabela 2 - Critérios de malária grave da OMS (133)
Forma da malária grave Manifestações clínicas Achados em exames
complementares
Malária cerebral Prostração, rebaixamento do
nível de consciência,
convulsões múltiplas ou
coma (escala coma Glasgow
<9 adultos ou Blantyre coma
score < 2 em crianças)
Tomografia computadorizada
de crânio normal ou com
edema cerebral difuso
Hipoglicemia Prostração, rebaixamento do
nível de consciência,
convulsões múltiplas ou
coma
Glicemia < 40 mg/dL
Anemia grave Intensa palidez cutâneo-
mucosa e astenia
Hemoglobina <7g/dL adultos
e <5g/dL crianças
Malária pulmonar Angústia respiratória com
crepitações à ausculta
Infiltrado alveolar difuso ou
imagem de condensação

25
pulmonar (inicialmente nas
bases)
difusa à radiografia de tórax
Acidose láctica Angústia respiratória com
respiração acidótica
Acidose à gasometria arterial
Hiperlactatemia
Malária álgida Síndrome do choque Pode haver hemocultura
positiva para bactérias Gram-
negativas
Malária renal Oligúria (< 400 mL) mesmo
após reidratação
Creatinina sérica >3,0 mg/mL
Coagulação intravascular
disseminada (CIVD)
Sangramento de grande
relevância
Plaquetopenia,
prolongamento de TAP e
TTPA, hipofibrinogenemia,
aumento dos produtos de
degradação da fibrina (PDF)
e dímeros-D
Febre hemoglobinúrica Colúria intensa Presença hemoglobinúrica
maciça ao exame de
sedimentação de urina
1.6 Malária vivax grave e resistência à cloroquina
A patogênese da malária grave envolve uma interação entre parasito e produtos de
membrana de eritrócitos, citocinas e receptores endoteliais, levando à ativação da inflamação
de plaquetas, alteração da hemostasia, um estado pró-coagulante, disfunção da
microcirculação e hipóxia tecidual, resultando em disfunções orgânicas diversas (134). Há
relatos de aumento de casos clínicos graves exclusivamente associados a infecções por P.
vivax, envolvendo anemia grave, insuficiência renal, icterícia, malária cerebral, convulsões,
insuficiência respiratória e morte (22, 102, 135). A anemia grave é a manifestação mais

26
comum e grave em Papua (67-80% dos casos) (100, 112) e foi responsável por cerca de um
terço dos casos graves na Índia (104) e na Amazônia brasileira (102).
Após o aparecimento da resistência à CQ em P. vivax, os relatos de gravidade clínica
exclusivamente associado a este parasito da malária humana começaram a ser descritos, o que
levou à hipótese de que a resistência à CQ pudesse de alguma forma estar relacionada à
gravidade clínica, especialmente para a anemia (100, 135, 136). Um tratamento ineficaz em
áreas onde parasitos resistentes estejam presentes, P. vivax produz uma doença febril aguda
com potencial para se tornar crônica, recorrente e grave, devido à parasitemia recorrente, em
especial a anemia grave (136). Infecções crônicas poderiam levar à diseritropoiese, que
desempenha um papel importante na patogênese na anemia (137). Portanto é necessária uma
avaliação aprofundada sobre a resistência à drogas e de seus marcadores genéticos em relação
à sua associação com complicações clínicas na malária vivax (138). A maior dificuldade para
se testar a hipótese, entretanto, está no fato de que em regiões onde existe muita resistência, já
não se utiliza mais a CQ de rotina (por exemplo, Papua e Indonésia), inviabilizando estudos
de associação in vivo. Além disso, não estão disponíveis bons testes de avaliação de
resistência in vitro. Assim, a identificação de marcadores moleculares associados à resistência
medicamentosa pode propiciar um método mais factível de demonstrar que parasitos
resistentes levam à maior gravidade clínica. Um estudo recente relatou níveis de expressão
aumentados em dois genes associados com multirresistência (pvcrt-o e pvmdr1) em um
paciente com malária grave de P. vivax (77), sendo esta a primeira evidência direta de que
esses dois genes podem ser utilizados como marcadores de gravidade associados com as
infecções por P. vivax.
O aumento da resistência à CQ tem sido associado com aumento de doença grave e
mortalidade (139). O uso de antimaláricos ineficazes pode resultar em persistência de
parasitas resistentes ocasionando proporção significativa de pacientes que desenvolvem mais
malária grave, especialmente anemia grave (140). A ocorrência simultânea da doença grave e
resistência à CQ em alguns países suscitou uma possível associação entre a gravidade e
resistência, especialmente para anemia em crianças (136).

27
1.7. Justificativa e hipóteses
Há alguns anos, o controle da malária deixou de ser enfatizado apenas na ação vetorial
onde a assistência aos pacientes era apenas ação complementar. Com isso, verificou-se que o
controle baseado em antimaláricos poderia ter importância fundamental. No entanto, com o
surgimento de resistência do P. vivax aos antimaláricos, essa dificuldade tornou-se um
obstáculo na estratégia de controle a partir das drogas. A compreensão dos mecanismos de
resistência à CQ é importante no entendimento da ação das drogas e na descoberta de
biomarcadores que servem como uma ferramenta no diagnóstico e na epidemiologia
molecular da resistência do parasito aos antimaláricos.
Apesar de todos estes avanços, é necessário buscar biomarcadores mais robustos e critérios
específicos envolvidos na patogênese da malária grave produzida pelo P. vivax para o
entendimento da malária e do embasamento para as futuras intervenções profiláticas e
terapêuticas. Recentemente, casos atípicos de malária vivax com maior morbidade e
mortalidade têm sido relatados no mundo e em particular no Brasil. É de fundamental
importância caracterizar tais casos e identificar ferramentas de predição do diagnóstico, além
de identificar possíveis alvos terapêuticos. A descrição de determinantes de proteção ou
gravidade é fundamental para guiar futuros esquemas de manejo dos pacientes e para estudos
epidemiológicos.
Esperamos que os resultados deste estudo permitam o conhecimento dos biomarcadores
moleculares relacionados com malária grave e que esses dados possam preencher lacunas na
falta de conhecimento nesta área. Os biomarcadores inovadores, tais como a expressão de
genes relacionados com a provável resistência da droga pode ser utilizado na prática clínica
para a monitorização da eficácia do tratamento, para o diagnóstico ou para prever o
prognóstico de malária grave ou resistência à CQ. Dessa forma, a expressão do pvcrt-o e
pvmdr1 pode funcionar como um marcador de resistência à CQ. O uso da expressão desses
genes como um biomarcador deve permitir a individualização do tratamento da malária,
ajudando a selecionar as drogas alternativas, como derivados de artemisinina em locais onde a
resistência à CQ foi registrada.

28
Diante do exposto, espera-se, neste trabalho, responder as seguintes questões: 1) A
expressão genica de pvcrt-o e pvmdr1 estará aumentada em pacientes graves? 2) A expressão
gênica de pvcrt-o e pvmdr1 estará influenciando algum parâmetro clínico ou laboratorial na
malária grave? e 3) A expressão gênica de pvcrt-o e pvmdr1 estará aumentada em pacientes
com malária resistente ao uso de cloroquina? A hipótese do trabalho seria que a expressão
gênica de pvcrt-o e pvmdr1 estaria aumentada tanto em pacientes com malária grave por P.
vivax quanto em pacientes resistentes à CQ.

29
2. OBJETIVOS

30
2.1 Objetivo geral
Avaliar se os níveis de expressão dos genes pvcrt-o e pvmdr1 estão associados com a
resistência à cloroquina e malária grave por Plasmodium vivax.
2.2 Objetivos específicos
2.2.1 Caracterizar o nível de expressão dos genes pvcrt-o e pvmdr1 em pacientes resistentes e
sensíveis à CQ;
2.2.2 Verificar se existe diferença na expressão gênica entre os pacientes resistentes e
sensíveis à CQ;
2.2.3 Caracterizar o nível de expressão dos genes pvcrt-o e pvmdr1 em pacientes graves e não
graves por Plasmodium vivax;
2.2.4 Verificar se existe diferença na expressão gênica entre os pacientes graves e não graves
por Plasmodium vivax;
2.2.5 Verificar se existe associação entre resistência à CQ e mutação nos genes pvcrt-o e
pvmdr1;
2.2.6 Correlacionar expressão desses genes com concentração de hemoglobina.

31
3. MATERIAL E MÉTODOS

32
3.1 Tipo de estudo
Foi realizado um estudo do tipo série de casos.
3.2 Área de estudo
O estado do Amazonas situa-se na porção ocidental da Região Norte do Brasil, entre os
meridianos de 56º05’ e 73º48’ longitude oeste e os paralelos de 2º01’ e 7º06’ latitude sul,
abrangendo 62 municípios, numa área de 1.559.159,148 km2. Possui uma população estimada
em 3.807.921 habitantes para o ano de 2013, com 80,9 % na zona urbana e 19,1 % na zona
rural. A capital Manaus tem uma população estimada em 1.982.177 habitantes, o que
representa 52,0% da população do estado (141) (Figura 9). O clima é do tipo equatorial
superúmido, com precipitações acima de 2.000 mm/ano e temperaturas médias anuais entre
26ºC e 28ºC, sem uma clara identificação de uma estação seca e outra chuvosa e com poucas
variações de temperatura na maior parte do estado.
Figura 9. Mapa do Estado do Amazonas indicando a localização do município de origem dos
isolados de Plasmodium vivax caracterizados neste estudo.

33
O estudo foi realizado entre junho de 2011 a dezembro de 2012 na Fundação de Medicina
Tropical Dr. Heitor Vieira Dourado (FMT-HVD), na cidade de Manaus (Estado do
Amazonas), localizada entre as coordenadas 2°57’ e 3°10’ e de latitude Sul e 59°53’ e 60°07’
de longitude oeste. A FMT-HVD é uma instituição de referência para o atendimento de
doenças infecciosas no Estado do Amazonas.
Durante esse período, no Estado do Amazonas, foram registrados 120.529 casos de malária,
sendo que 110.900 (92,0%) causados pelo P. vivax. Já, no município de Manaus, foram
registrados 19.690 casos de malária, sendo que 19.305 (98,0%) causada pelo P. Vivax. Na
FMT-HVD, foram atendidos 6.277 pacientes com malária, sendo que 6.090 casos por P. vivax
(97,0 %).
3.3 Seleção de pacientes
3.3.1 Seleção de pacientes resistentes à CQ
O estudo incluiu pacientes de ambos os sexos, entre 6 meses e 60 anos, apresentando peso
superior a 5 quilos, densidade parasitária entre 250 a 100.000 parasitas/mL e temperatura
axilar ≥ 37.5°C ou história de febre nas últimas 48 horas. Critérios de exclusão foram: uso de
antimaláricos nos últimos 30 dias, impossibilidade de ser acompanhado por 42 dias e qualquer
complicação clínica (36) (Figura 10). Pacientes receberam tratamento supervisionado com 25
mg/kg de fosfato de CQ por um período de 3 dias (10 mg/kg no dia 0 e 7,5 mg/kg nos dias 1 e
2). PQ foi prescrita no dia 42 por um período de 7 dias, na dosagem de 0,5 mg/kg por dia.
Pacientes que vomitaram 30 minutos após a ingestão do medicamento foram retratados com a
mesma dose. Pacientes foram avaliados nos dias 0, 1, 2, 3, 7, 14, 28 e 42 e, se apresentasse
qualquer sintoma durante o período de acompanhamento. No dia da admissão (D0) e no dia
de recrudescência (DR), 20 mL de sangue total foram coletados para armazenamento de DNA
e RNA. Nível plasmático de CQ e DCQ foi determinado somente em casos de falha
parasitológica (142), utilizando 3 alíquotas de 100 μL de sangue total armazenado em papel
de filtro para análise por cromatografia liquida de alta eficiência (HPLC) (143, 144). Paciente

34
resistente à CQ foi definido se apresentou: 1) recorrência do parasito com concentração
plasmática de CQ maior que 100 ng/dL ou 2) recorrência com concentração plasmática de CQ
menor que 100 ng/dL mas apresentando os mesmos alelos no D0 e DR. O grupo susceptível à
CQ consistiu de pacientes sem recorrência durante o período de acompanhamento do estudo.
Figura 10- Fluxograma do estudo, descrevendo com detalhes os pacientes elegíveis,
excluídos e PvCQR e susceptível à CQ analisados.
3.3.2 Seleção de pacientes com malária grave e não grave por P. vivax
Durante o mesmo período, pacientes foram admitidos e classificados como graves se
apresentassem algum critério de gravidade para malária descrita para P. falciparum (133)
(Figura 11). Na admissão, 20 mL de sangue total foi coletado para realização de hemograma
(aparelho automatizado PENTRA 120-Retic®, da ABX Diagnostics®), exames bioquímicos e

35
estocagem de RNA e DNA. Todos os pacientes realizaram ultrassom abdominal e raio X.
Para excluir pacientes com outras infecções, todos os pacientes realizaram hemocultura,
sorologia para leptospirose (IgM), HIV-1/HIV-2, hepatite A (anti-HAV IgM), hepatite B
(HBsAg), hepatite C (anti-HCV), hepatite D (total anti-HDV) e dengue (RT-PCR). Pacientes
que apresentaram qualquer critério de gravidade foram tratados com ACT intravenoso, como
recomendado pela OMS (36).
Figura 11- Fluxograma do estudo, descrevendo com detalhes os pacientes elegíveis,
excluídos e pacientes graves e não graves analisados.
3.4 Diagnóstico de Plasmodium vivax por gota espessa
O diagnóstico foi realizado através da gota espessa pela técnica de Walker (145) conforme
a descrição do POP_MAL_LB_001_v02D_PT (ANEXO 1) e avaliados por microscopistas

36
experientes (146). A densidade parasitária (parasitas/µL) foi calculada pela contagem do
número de parasitas em 500 leucócitos. Além disso, foi feita a contagem de formas
assexuadas (trofozoítos jovens, trofozoítos maduros e esquizontes) para assegurar que não
havia diferença entre os resistentes e sensíveis à CQ e entre graves e não graves.
3.5 Caracterização genética
3.5.1 Extração do DNA
Para confirmar a monoinfecção por P. vivax foi realizado Real time PCR. Para a extração
de DNA foi utilizado o kit QIAmp® Blood Mini kit (Qiagen, Hilden, Germany).
Resumidamente, para 200 µL de sangue com EDTA foram adicionados 20 µL de proteinase
K e 200 µL de tampão de lise. Após completa homogeneização em vórtex, a mistura foi
incubada em banho-maria a 56°C por 10 minutos. Ao lisado foram adicionados 200 µL de
etanol 96-100%, homogeneizando em vórtex por 5 segundos para a obtenção de uma solução
homogênea. Para a purificação do DNA, a solução foi aplicada em coluna, que posteriormente
foi lavada sucessivamente com os dois tampões de lavagem. O DNA obtido foi eluído com 50
µL do tampão de eluição, conforme descrita no POP_MAL_LB_002_v01D_PT (ANEXO 1)
que faz parte da rotina do laboratório.
3.5.2 Realização de PCR
Os primers e sondas usados para amplificar o gene 18S rRNA do P. vivax e P. falciparum
foram utilizados como descrito no POP_MAL_LB_003_v02D_PT (147) (Anexo 1) que faz
parte da rotina do laboratório. As PCRs foram realizadas em reações de 20 L contendo 300
nM de cada primer, 200 nM de cada sonda, 10 L de master mix TaqMan PCR (Applied®
Biosystems) e 3 L de DNA genômico. A PCR foi realizada em Applied® Biosystems 7500
Fast System nas seguintes condições: 2 min a 50°C, 10 min a 95°C para ativar AmpliTaq
Gold DNA polimerase, seguido de 45 ciclos de 15 seg a 95°C e 1 min a 60°C. Em cada

37
experimento, uma mistura sem DNA foi incluído como controle negativo e cada experimento
foi realizado em duplicata.
3.5.3 Extração do RNA
Para extração do RNA, as amostras foram tratadas com saponina e isoladas utilizando o
reagente Trizol® até a extração conforme descrito no POP_MAL_LB_004_v01D_PT (148)
(Anexo 1). Resumidamente, foram adicionados 200 µL de clorofórmio, homogeneizado,
incubado por 5-15 min e centrifugado a 12.000 g por 15 min a 4°C. O sobrenadante foi
transferido para um novo tubo com 500 µL de isopropanol e incubado por 10 min a
temperatura ambiente e centrifugado a 12.000 g por 10 min a 4°C. O sobrenadante foi
descartado e lavado com 1 mL de etanol 75 % e centrifugar a 7.500 g por 5 min 4°C. O pellet
foi ressuspenso em 20 µL de H2O DEPC e deixado em termobloco a 65C por 5 min, agitado
em um vórtex por 2 min e dar spin de vários segundos.
Para assegurar a qualidade do RNA foi realizado o gel de agarose a 2%. Para evitar que
contaminação por DNA genômico possa ter interferido nos resultados, todas as amostras de
RNA total foram tratadas com DNAse antes de serem submetidas ao RT-PCR (reação em
cadeia da polimerase e transcriptase reversa), conforme as instrução do fabricante, DNAse I –
Amplification Grade (Invitrogen®, EUA). Para a reação de transcrição reversa, foi utilizado o
kit SuperScript III (Invitrogen®, EUA), conforme descrito no POP MAL_LB_006_v01D_PT
(Anexo 1).
3.5.4 Expressão gênica de pvcrt-o e pvmdr-1
As reações de amplificação foram realizadas utilizando SYBR Green PCR Master Mix
(Applied Biosystems) e 45 ng de cDNA de cada amostra como descrito no
POP_MAL_LB_005_v01D_PT (Anexo 1). Os experimentos foram executados de forma
independente e em triplicata. Os produtos de PCR foram amplificados usando Applied
Biosystems® 7500 Fast System (77). Para analisar os níveis de transcrição relativa, o valor do
ciclo limite (Ct) de cada amostra foi utilizado para calcular e comparar o ∆Ct de cada amostra

38
do P. vivax do gene Sal I β-tubulina. O nível de expressão gênica foi calculado como N-fold =
2 - Δ Δ CT (149) para comparar o nível de transcrição de pvcrt-o e pvmdr1 em diferentes grupos
de pacientes incluídos no estudo versus pacientes susceptíveis à CQ. A PCR foi realizada nas
seguintes condições: desnaturação inicial a 95°C por 10 min, seguidos por 40 ciclos de 95°C
por 15 seg e 60°C for 1 minuto. Depois da PCR, uma etapa adicional de curva de dissociação
foi realizada e a presença de um único pico revelou a ausência de bandas não específicas, o
que confirmou a especificidade dos iniciadores de amplificação em tempo real PCR.
A reprodutibilidade do método (precisão inter-corrida) foi avaliada por meio de medidas
de Ct obtidos pela quantificação de cDNA de uma amostra analisada 10 vezes, em dois
ensaios que foram realizados em dias diferentes utilizando o mesmo equipamento. Os
resultados foram expressos como média, desvio padrão (SD) e coeficiente de variação
(CV%). A repetibilidade (precisão intra-corrida) do método foi avaliada em uma mesma
corrida por medidas de Cts obtidos pela quantificação de cDNA de uma amostra medido 10
vezes numa mesma corrida em dois ensaios. Os resultados foram expressos por média, desvio
padrão (SD) e coeficiente de variação (CV%).
3.5.5 Sequenciamento dos genes pvcrt-o e pvmdr1
Todos os grupos de pacientes que foram resistentes ou susceptível à CQ e graves ou não
graves, obtidos antes do tratamento (D0), foram sequenciados para pesquisar mutações
suspeitas de ser associada com CQR. Os primers utilizados e as condições da reação usadas
para amplificar as sequências de pvcrt-o e pvmdr-1 foi realizado como descrito (Tabela 3). As
PCRs foram realizadas em reações de 20 µL contendo: 2 μL de tampão 10X, 2,5 mM MgCl2,
0,2 mM de cada dNTP, 0,25 μM de cada primer, 0,5U de DNA polimerase e 3 µL de DNA
genômico. A PCR foi realizada em termociclador nas seguintes condições: desnaturação
inicial a 94°C por 10 min, acompanhado de 35 ciclos de desnaturação 94°C por 50 seg,
temperatura de anelamento dependendo de cada primer por 1 min, extensão a 72°C por 1 min.
Os produtos amplificados foram quantificados por NanoDrop® 2000 (Thermo Scientific®) e
enviados para sequenciar pela Macrogen® (Coreia do Sul). As sequências de aminoácidos

39
foram comparadas com sequências do Sal I (Salvador I) e alinhados e analisados usando
Mutation Surveyor®. As sequências dos genes pvcrt-o e pvmdr1 foram depositadas no
genebank com número de acesso KM016489 a KM016517 e KM016518 a KM016532,
respectivamente.
Tabela 3. Primers usados para sequenciamento do DNA de genes ortólogos de P.vivax.
Gene Cromossomo Sequência 5’ 3’ Pares
bases
Temp. anelamento
(°C)
Referências
pvcrt-o 1 AAGAGCCGTCTAGCCATCC 1.186 61 (92)
AGTTTCCCTCTACACCCG
pvmdr1 10 GGATAGTCATGCCCCAGGATTG 604 62 (92, 150)
CATCAACTTCCCGGCGTAGC
3.5.6 Análise em marcadores moleculares, msp1F3 e MS2 em pacientes resistentes à CQ
O MS2 foi escolhido devido a sua alta diversidade genética demonstrado em estudos
anteriores na América do Sul (151, 152) e o msp1F3 pois codifica uma região altamente
polimórfica da MSP1 e apresenta baixa ocorrência de artefatos na PCR (151, 153). A PCR foi
executada como descrito por Koepfli et al. (154), com pequenas modificações, sendo
realizado em reações de 20 L contendo 10 M de cada primer, 2 L de tampão 10X, 2,5
mM dNTP, 25 mM MgCl2, 1,5 U Taq DNA polimerase e 5 L DNA genômico (Tabela 4).
Foram diluídos 3 L do produto de PCR primário para ser utilizado como amostra na nested
PCR (msp1F3 e MS2). A reação foi realizada em termociclador nas seguintes condições:
desnaturação inicial a 95°C por 1 min, acompanhado de 29 ciclos (PCR primaria) ou 24 ciclos
(nested PCR) de desnaturação 95°C por 30 seg, anelamento a 59°C por 45 seg, extensão a
72°C por 1 min, acompanhada por extensão final a 72°C por 5 min. Em cada experimento,
uma mistura sem DNA foi incluído como controle negativo. Os produtos de PCR foram

40
estocados no escuro (embaladas em papel alumínio) a 4°C até ser enviada para Macrogen®
para realização de eletroforese capilar e foi analisada com GeneMarker® version 2.6.0.
Para assegurar a qualidade dos produtos de PCR foi realizado o gel de agarose a 2%. Os
perfis eletroforéticos foram analisados para cada amostra em relação ao comprimento do
amplicon e altura do pico. Os diferentes alelos detectados foram agrupados de acordo com o
respectivo comprimento de repetição (msp1F3 -3 pb, MS2 -4 pb). Os picos abaixo de 150 bp
não foram considerados, pois resultam de dímeros de primers residuais. O valor mínimo para
a altura do pico foi definido como 500 unidades de fluorescência relativa. De acordo com a
OMS, uma amostra foi classificada como recrudescente se pelo menos um alelo foi detectada
no D0 e DR (155, 156) e como reinfecção se todos os alelos para um gene são distintos no D0
e DR (155).
Tabela 4. Oligonucleotideos usados na genotipagem do P. vivax.
Marcador Cromossomo Tipo
marcador
Primer Sequencia 5' - 3' Dye
MS2 3 Microsatellite PF AGCACGACCAACAAGAGAGG 6-FAM
NF GAGCTAGCCAAAGGTTCAAA
NR TGGGGAGAGACTCCCTTTTC
msp1F3 7 Surface
antigen
PF GGAGAACATAAGCTACCTGTCC VIC
PR GTTGTTACTTGGTCTTCCTCCC
NF CAAGCCTACCAAGAATTGATCCCCAA
NR ATTACTTTGTCGTAGTCCTCGGCGTAGTCC
3.6 Aspectos éticos
O estudo foi aprovado pelo Comitê de Ética em Pesquisa da Fundação de Medicina
Tropical Heitor Vieira Dourado (FMT-HVD) (número de aprovação 343/2009). A coleta das
amostras sanguínea foi realizada após a exposição dos objetivos da pesquisa aos pacientes.
Participantes foram instruídos sobre os objetivos do estudo e assinaram o termo de

41
consentimento livre e esclarecido (TCLE). Em caso de menores de 18 anos, o TCLE foi
assinado pelo pai ou responsável. Pacientes diagnosticados com malária foram tratados de
acordo com o Ministério da Saúde (36).
3.7 Análise estatística
Os dados foram analisados usando SPSS® versão 16.0 for Windows (SPSS Inc® Chicago,
IL, USA). A distribuição normal de hemoglobina e expressão gênica foi testada com teste de
Kolmogorov-Smirnov. Utilizou-se regressão linear para correlacionar hemoglobina e
expressão gênica entre os pacientes hospitalizados; teste de correlação de Spearman's para
correlacionar concentração de CQ com expressão gênica no DR e para correlacionar
expressão gênica no D0 e DR com concentração de expressão RNA. O teste
de Wilcoxon pareado foi utilizado para comparar expressão gênica dos pacientes CQR em D0
e DR. Teste qui-quadrado ou de Fisher foi usado para testar diferenças na frequência de
polimorfismo a distribuição de estágios sanguíneos assexuados. A variação genética dos
marcadores moleculares (msp1F3 e MS2) foi mensurada calculando heterozigosidade (HE).
HE foi calculada em D0 e DR para cada locus como=[n/(n-1)][1-∑pi2] onde n é o número de
isolados e pi é a frequência do alelo i.

42
4 RESULTADOS

43
ARTIGO
Melo GC, Monteiro WM, Siqueira AM, Silva SR, Magalhães BML, Alencar ACC, Kuehn A,
del Portillo, HA,, Fernandez-Becerra C, Lacerda MVG. Expression levels of pvcrt-o and
pvmdr-1 are associated with chloroquine resistance and severe Plasmodium vivax malaria in
patients of the Brazilian Amazon. Artigo aceito para publicação na Revista Plos One.

44
Expression levels of pvcrt-o and pvmdr-1 are associated with chloroquine resistance and
severe Plasmodium vivax malaria in patients of the Brazilian Amazon
Gisely C. Melo1, Wuelton M. Monteiro1,2, André M. Siqueira1, Siuhelem R. Silva3, Belisa
M.L. Magalhães1 Aline C.C Alencar1, Andrea Kuehn2,4, Hernando A. del Portillo4,5,
Carmen Fernandez-Becerra4#, Marcus V.G. Lacerda1,2#
1 Universidade do Estado do Amazonas, Manaus, Amazonas, 69040-000 Brazil,
2 Fundação de Medicina Tropical Dr. Heitor Vieira Dourado, Manaus, Amazonas, 69040-000,
Brazil
3 Universidade Paulista UNIP, Manaus, Amazonas, 90050-030, Brazil.
4 Barcelona Centre for International Health Research (CRESIB, Hospital Clínic-Universitat de
Barcelona), Barcelona, 08036, Spain
5 Institució Catalana de Recerca i Estudis Avançats (ICREA), Barcelona, 08010, Spain
# Corresponding authors: Marcus VG Lacerda, Gerência de Malária, Fundação de Medicina
Tropical Dr. Heitor Vieira Dourado, Av. Pedro Teixeira, 25, Manaus 69040-000, Brazil, Tel:
+55 (92) 2127 3537 Fax: (+55) 92 2127 3537, E-mail: [email protected] and
Carmen Fernandez-Becerra, Barcelona Centre for International Health Research, E-mail:
[email protected], Tel. (+34) 932275400 (ext. 4276); Fax (+34) 932271850

45
ABSTRACT
Molecular markers associated with the increase chloroquine resistance and disease severity in
Plasmodium vivax are needed. The objective of this study was to evaluate the expression
levels of pvcrt-o and pvmdr-1 genes in a group of patients presenting CQRPv and patients
who developed severe complications triggered exclusively by P. vivax infection. Two
different sets of patients were included to this comprehensive study performed in the Brazilian
Amazon: 1) patients with clinically characterized chloroquine-resistant P. vivax compared
with patients with susceptible parasites from in vivo studies and 2) patients with severe vivax
malaria compared with patients without severity. Quantitative real-time PCR was performed
to compare the transcript levels of two main transporters genes, P. vivax chloroquine
resistance transporter (pvcrt-o) and the P. vivax multidrug resistance transporter (pvmdr-1).
Twelve chloroquine resistant cases and other 15 isolates from susceptible cases were included
in the first set of patients. For the second set, seven patients with P. vivax-attributed severe
and 10 mild manifestations were included. Parasites from patients with chloroquine resistance
presented up to 6.1 (95%CI: 3.8-14.3) and 2.4 (95%CI: 0.53-9.1) fold increase in pvcrt-o and
pvmdr-1 expression levels, respectively, compared to the susceptible group. Parasites from the
severe vivax group had a 2.9 (95%CI: 1.1-8.3) and 4.9 (95%CI: 2.3-18.8) fold increase in
pvcrt-o and pvmdr-1 expression levels as compared to the control group with mild disease.
These findings suggest that chloroquine resistance and clinical severity in P. vivax infections
are strongly associated with increased expression levels of the pvcrt-o and pvmdr-1 genes
likely involved in chloroquine resistance.
Keywords: chloroquine, resistance, malaria, Plasmodium vivax, pvcrt-o, pvmdr-1, gene
expression, molecular markers.

46
Introduction
Mechanisms underlying severe malaria triggered by Plasmodium vivax have been poorly
appreciated, mainly the likely higher and more-severe disease burden imposed by increasingly
chloroquine-resistant P. vivax parasites (CQRPv) (100, 157). Most common manifestations
of severe vivax malaria include severe anemia and respiratory distress, being particularly
associated to young age (100, 107, 158). Of interest, the majority of reports on severe vivax
malaria come from regions where drug-resistant P. vivax parasites significantly thread the
radical cure and control of this infection (136). The first reports of CQRPv were made in 1989
(44, 159) from Papua New Guinea and Indonesia. In Brazil, the first case of properly
ascertained in vivo CQRPv was from a patient treated in Manaus, in the Brazilian Amazon
(54). In this city, a subsequent trial assessed the efficacy of standard supervised CQ
monotherapy and the proportion of failures was 10.1% (55). In parallel to the emergence of
CQRPv in the Brazilian Amazon, reports of clinical severity exclusively associated with P.
vivax infection increased substantially (102, 103, 108, 158).
Based on observational studies, an association between CQRPv and disease severity in P.
vivax was proposed in the Indonesian Papua (100), although this has not been properly and
systematically evaluated. CQR in P. falciparum is associated with mutations in the pfcrt-o
and pfmdr-1 genes (160, 161) but similar studies with the orthologous genes of P. vivax,
pvcrt-o and pvmdr-1, failed to demonstrate a similar association (58, 73, 75). Interestingly, a
pioneer case of severe disease exclusively associated with P. vivax, demonstrated a significant
increase on expression levels, not mutations, of these genes (77). Altogether, these studies
suggest the possibility of a significant association of CQRPv and disease severity and that
expression levels of pvcrt-o and pvmdr-1 should be better explored as molecular markers of
both phenomena to further enhance the understanding of the clinico-epidemiological. The aim
of this study was to evaluate the expression levels of pvcrt-o and pvmdr-1 genes in a well-
characterized group of patients presenting CQRPv and in a group of patients who developed
severe complications triggered by P. vivax infection.

47
Methods
Ethics statement
The study was approved by the Ethics Review Board of the Fundação de Medicina Tropical
Heitor Vieira Dourado (FMT-HVD) (approval number 343/2009). Participants were
instructed about the objectives of the study and signed an informed consent. In the case of
under 18 years, the consent form was signed by the guardians. Patients diagnosed with
malaria were treated according to the guidelines of the Brazilian Ministry of Health (36).
Study area
The study was carried out from June 2011 to December 2012 in the FMT-HVD, an infectious
disease referral center located in Manaus, Western Brazilian Amazon.
Selection of patients for the CQRPv study
The study included patients with P. vivax malaria of both sexes, aged 6 months-60 years,
bodyweight greater than 5 kg, presenting blood parasite density from 250 to 100,000
parasites/ml and axillary temperature ≥ 37.5°C or history of fever in the last 48 hours.
Exclusion criteria were: use of antimalarials in the previous 30 days, refusal to be followed up
for 42 days and any clinical complication (36). Patients received supervised treatment with 25
mg/kg of chloroquine (CQ) phosphate over a 3-day period (10 mg/kg on days 0 and 7.5
mg/kg on days 1 and 2). Primaquine was prescribed at the end of follow-up or if disease
recurred for 7-day period in the dosage of 0.5 mg/kg per day. Patients who vomited the first
dose within 30 minutes after drug ingestion were re-treated with the same dose. Patients were
evaluated on days 0, 1, 2, 3, 7, 14, 28 and 42 and, if they felt ill, at any time during the follow-
up period. In all sample, blood smear, full blood counts and PCR were performed. On

48
admission (D0) and in the day of recrudescence (DR), 20 ml of intravenous whole blood was
also collected for DNA and RNA storage. CQ and desethylchloroquine (DCQ) plasmatic
levels were determined only in case of parasitological failure (142). Three aliquots of 100 μl
of whole blood from DR samples were spotted onto filter paper for later analysis by high
performance liquid chromatography (HPLC) to estimate the levels of CQ and DCQ as
previously described (143, 144). CQRPv was defined if: 1) recurrences of the parasite with
plasma concentrations of CQ higher than 100 ng/dl, or 2) recurrences with smaller
concentrations but presenting DR and D0 with the same alleles. The control group consisted
of patients with no parasitemia recurrence during the same follow-up period.
Selection of patients with severe P. vivax malaria and patients with mild symptoms
During the same period, admitted patients classified as severe if presenting any of the WHO
severity criteria (133), and patients with mild manifestations from the outpatient clinics. On
admission, 20 ml of intravenous whole blood was collected for complete blood count, blood
biochemistry and for DNA and RNA extraction. Other tests were requested at physician’s
discretion. To exclude patients with infection other than Plasmodium, all patients also had a
blood culture performed for aerobic bacteria and serological tests for leptospirosis (IgM),
HIV-1/HIV-2, hepatitis A (anti-HAV IgM), hepatitis B (HBsAg), hepatitis C (anti-HCV),
hepatitis D (total anti-HDV) and dengue (RT-PCR). Patients presenting any sign of severity
were treated with intravenous artesunate, as recommended by the WHO guidelines (36).
P. vivax malaria diagnosis
Thick blood smear was prepared as recommended by the Walker technique (145) and
evaluated by an experienced microscopist (146). Parasite densities (parasites/µl) were
calculated by counting the number of parasites per 500 leukocytes in high magnification
fields, and the number of leukocytes/µl per patient. In addition, differential counting of
asexual forms (ring-stage parasites, mature trophozoites and schizonts) was made to ensure
that there was no difference between groups of cases and controls.

49
Afterwards, real-time PCR was performed to confirm P. vivax mono-infection. The extraction
of total DNA from whole blood was performed using the QIAamp DNA Blood Mini Kit
(Qiagen®, USA), according to the manufacturer’s protocol. The DNA was amplified in an
Applied Biosystems® 7500 Fast System using primers and TaqMan fluorescence labeled
probes for real time PCR (162).
RNA extraction and expression of pvcrt-o and pvmdr-1 genes
For RNA extraction, 5 ml of whole blood were collected and processed using saponin. Total
RNA was purified using Trizol reagent (Invitrogen®) according to the manufacturer’s
instructions. Furthermore, 2% agarose gel electrophoresis was made to guarantee RNA
quality. Amplification reactions were performed using Power SYBR Green PCR Master Mix
(Applied Biosystems®) and 45 ng of template cDNA prepared from each sample. PCR
products were amplified and detected on a 7500 FAST (Applied Biosystems®). The primers
were
pvmdr-1F (AAGGATCAAAGGCAACCCA ),
pvmdr-1R (TCAGGTTGTTACTGCTGTTGCTATT),
pvcrt-oF (ATGTCCAAGATGTGCGACGAT),
pvcrt-oR (CTGGTCCCTGTATGCAACTGAC),
pvtubulinF (CCAAGAATATGATGTGTGCAAGTG),
pvtubulinR (GGCGCAGGCGGTTAGG) (77). Cycling parameters for PCR were an initial
denaturation step at 95°C for 10 minutes, followed by 40 cycles of 95°C for 15 seconds, and
60°C for 1 minute. Samples were run in triplicate. To analyze the relative transcript levels, the
threshold cycle value (Ct) of each sample was used to calculate and compare the ΔCt of each
sample to that of the P. vivax housekeeping gene Sal I β-tubulin; the ΔΔCt was also calculated
as N-fold = 2 - Δ Δ CT (149) to compare the transcript levels of pvcrt-o and pvmdr-1 in the
different groups of patients included in this study versus CQ-susceptible patients. After the

50
PCR reaction in real time, an additional step of decoupling the amplicon to generate a melting
curve was performed. The presence of only one peak in the dissociation curves reveals the
absence of unspecific bands, confirming the specificity of the amplification primers in real
time PCR.
Reproducibility of the method (accuracy inter-runs) was assessed through measures of Cts
obtained by cDNA quantification of one isolate analyzed 10 times in two runs that were
performed on different days, using the same equipment. The results were expressed as mean,
standard deviation (SD) and coefficient of variation (CV%). The repeatability of the method
(accuracy intra-run) was evaluated by measures of Cts obtained by cDNA quantification of
one isolate measured 10 times in the same run. The results were expressed by mean, standard
deviation (SD) and coefficient of variation (CV%).
Sequencing analysis of pvcrt-o and pvmdr-1 genes
All samples from patients presenting CQRPv or patients with severe symptoms and the
respective control groups, obtained prior to treatment (on D0), were sequenced for searching
mutations suspected to be associated to CQR. Extraction of whole DNA was carried out using
a QIAamp® DNA Mini Kit (QIAGEN®, Germany) according to the manufacturer’s protocol.
PCR primers and different reaction conditions used to amplify pvcrt-o and pvmdr-1 gene
sequences were made as previously described (Table S1). The amplification products were
quantified by NanoDrop® 2000 (Thermo Scientific®) and sent for sequencing to Macrogen®
(South Korea). Amino acid sequences were compared with sequences of Sal I (Salvador I):
GenBank accession nos. ADE74979.1 for pvcrt-o (73) and EU333979.1 for pvmdr-1 (138).
The deduced amino acid sequences were aligned and analyzed using Mutation Surveyor®.
The pvmdr-1 and pvcrt-o gene sequences were deposited in GenBank under the accession
numbers KM016489 to KM016517 and KM016518 to KM016532, respectively.
Polymorphism analysis of msp1F3 and MS2 molecular markers for patients with CQRPv

51
PCRs were performed in 20 l reactions containing 10 M of each primer, 2 l of 10 x buffer
B, 2.5 mM each dNTP, 25 mM MgCl2, 1.5 U Taq DNA polymerase and 5 l genomic DNA
(Table S2). 3 l of diluted primary PCR product was used as template for the nested PCR
(MS2 and msp1F3). These polymorphism markers were chosen based on preliminary data
from other patients from Manaus (unpublished data). PCRs were performed in a thermocycler
with conditions as follows: initial denaturation at 95°C for 1 min, followed by 29 cycles
(primary PCR) or 24 cycles (nested PCR) of denaturation at 95°C for 30 seconds, annealing at
59°C for 45 sec, elongation at 72°C for 1 min, followed by a final elongation at 72°C for 5
min. PCR products were stored in the dark (plates wrapped in foil) at 4°C until be sent to
Macrogen® for capillary electrophoresis-based sequencing and analyzed with GeneMarker®
version 2.6.0.
Statistical Analyses
Data were analyzed using SPSS® version 16.0 for Windows (SPSS Inc® Chicago, IL,
USA). Normal distribution of data of hemoglobin and gene expression was evaluated with the
Kolmogorov-Smirnov test. Linear regression was performed to correlate this variable with
hemoglobin and gene expression among hospitalized patients. Spearman's correlation test was
performed to correlate CQ concentration with gene expression in the DR and gene expression
at D0 and DR with concentration of RNA. The Wilcoxon rank pairs test was performed to
compare CQR patients had paired expression levels at D0 and DR. Chi-square or Fisher's test
was used to test differences in proportions of polymorphism and distribution of asexual blood
stages. The genetic variation for each microsatellite locus was measured by calculating the
expected heterozygosity (HE). HE was calculated using D0 and DR for each locus as =[n/(n-
1)][1-∑pi2] where n is the number of isolates sampled and pi is the frequency of allele i.

52
Results
Reproducibility, repeatability of measurements of Cts and specificity
For reproducibility, amplification of pvcrt-o and pvmdr-1 genes showed average Ct values
ranging from 35.14 to 37.93 (mean Ct value 36.82) and from 34.99 to 36.19 (mean Ct value
35.44), respectively. Among the twenty measured values, doing in to different runs, the
maximum coefficient of variation was 2.34 % and 1.12%, for pvcrt-o and pvmdr-1
respectively. For the repeatability, the coefficient of variation was 1.35% for the pvcrt-o and
0.75% for pvmdr-1 in the first run, and 2.77 % for the pvcrt-o and 1% for pvmdr-1 in the
second run (Table S3).
After the relative quantification, only one peak was observed in the dissociation curves,
confirming a good specificity of the primers in real time PCR.
Gene expression levels from CQR parasites
In this period, in a parallel study to evaluate efficacy of CQ with the 42-day follow-up, with
supervised drug administration, 135 patients were enrolled, and 16 were CQR (11.8%). Of
these, 12 with adequate RNA storage were included in the analysis. Clinical and laboratorial
characteristics of the 12 patients with CQRPv included in this study are presented in Table 1.
Out of this total, 83.3% patients were male, with a mean age of 29.0 years. The mean
hemoglobin was 13.1g/dl at D0 and 12.7g/dl in DR. The geometric mean parasitemia was
1781.7 parasites/l at D0 and 4206.5 parasites/l at DR. It was found that mean blood levels
of CQ plus DCQ were greater than 100 ng/ml on the DR in 10 of 12 (83.3%) patients with
recurring infection. Five patients with CQ-susceptible parasites and mild malaria were
selected to generate the relative expression of 12 CQRPv and 15 CQ-susceptible isolates.

53
In patients R1 to R7 gene expression was analysed at D0 and DR, while in patients R8 to R10
and R11 to R12, due to insufficient RNA amount, gene expression was measured only at D0
or DR, respectively. Patients with CQ-resistant P. vivax parasites presented a higher gene
expression of pvcrt-o and pvmdr-1 at D0 and DR when compared to the susceptible group
(Figure 1A and 1B). For the CQR patients, median gene expression values at D0 and DR,
presented 2.4 fold (95%CI: 0.96-7.1) and 6.1 fold (95%CI: 3.8-14.3) increase in pvcrt-o levels
(Figure 1A and 2A) compared to the susceptible patients at D0 with 0.12 fold (95%CI: 0.034-
0.324) (Figure S1A and 2A). Median gene expression for pvmdr-1 presented 2.0 fold (95%CI:
0.95-3.8) and 2.4 fold (95%CI: 0.53-9.1) increase levels at D0 and DR, for the CQR patients
(Figure 1B and 2B) versus 0.288 fold (95%CI: 0.068-0.497) for the susceptible patients at D0
(Figure S1B and 2B). Patients R2, R4, R6 and R7 showed a higher expression level of both
genes in relation to the control group at D0 and DR (Figure 1). It was not observed significant
differences in expression levels between D0 and DR for resistant parasites (p=0.375 for pvcrt-
o and p=0.844 for pvmdr-1). However, when we compared expression levels between
resistant and sensible parasites at D0 for both genes, values were significantly greater for
treatment failures (p<0.05 for both genes). No correlation was found between CQ
concentration and gene expression of DR for pvcrt-o (p=0.286) and pvmdr-1 (p=0.433) genes
and between gene expression of pvcrt-o and pvmdr-1 and concentration of RNA in D0 and
DR (p˃0.05).
To discard that difference in gene expression levels was not due to difference in the
distribution of asexual blood stages, parasites distribution was counted in both, CQRPv and
CQ-susceptible samples (Tables S4 and S5). Of notice, no significant differences were
observed between immature and mature asexual forms in both groups (p˃0.05). Moreover, it
was found that the pvcrt-o and pvmdr-1 gene mutations studied were not related to CQ-
resistance (p˃0.05) (Table 2).
In order to characterize the genetic variation for each individual population, two neutral
microsatellite loci (msp1F3 and MS2) were analyzed in D0 and DR samples. The analysis of
microsatellites showed at least one concordant allele (83.3 %) in primary infection and
recrudescence (Table 1). Patients which presented CQ concentration less than 100 ng/ml,
showed the same microsatellite allele in D0 and DR. The expected heterozygosity (HE) was

54
different in parasites from primary and recrudescence episodes (for msp1F3, primary:
HE=0.915 and recrudescence: HE=0.681; and for MS2, primary: HE=0.531 and recrudescence
HE=0.771).
Gene expression levels in severe vivax malaria parasites
In the period of study, 70 patients were hospitalized with P. vivax. However, due to many
comorbidities, and absence of any criterion of severity, only 16 could be characterized as
severe vivax malaria. Of these, 7 patients presenting mono-infection by P. vivax with any
criterion of severity were included. Of these patients, 4 had severe anemia, 1 had severe
anemia paralleled with acute respiratory distress syndrome (ARDS) and acute renal failure, 1
had acute renal failure and prostration and 1 had ARDS alone. Demographic and clinical
characteristics of the patients enrolled are presented in Table 3. Of these patients, 51.0% were
male, with a mean age of 38.6 years. The mean disease duration was 6.3 days, median
hemoglobin was 6.3 g/dl and mean parasitemia was 11,180 parasites/l.
Ten patients diagnosed with non-severe malaria were selected and included as controls. The
same 5 CQ-susceptible isolates from patients with mild malaria above cited were used to
generate the relative expression of seven severe malaria patients and 10 non-severe malaria
patients. The group of patients with severe vivax malaria had a 2.9 (95%CI: 1.1-8.3) and 4.9
(95%CI: 2.3-18.8) fold increase in pvcrt-o and pvmdr-1 expression level versus 0.7 (95%CI:
0.3-1.1) and 1.5 (95%CI: 0.7-2.4) for the control group with mild disease (p=0.007 and
p=0.01) (Figure 2, Figure 3 and Figure S2). To discard that difference in gene expression
levels was not due to difference in the distribution of asexual blood stages, parasites
distribution was counted in both, severe vivax malaria (Table S6) and non-severe. No
significant difference was found between immature and mature asexual forms among patients
with severe and non-severe malaria (p˃0.05) (Table S6). The pvcrt-o and pvmdr1 gene
mutations studied were not related to severity (p˃0.05) (Table 2).
In a linear regression analysis, it was found a negative correlation between hemoglobin values
(of severe and non-severe patients) and the log of the expression of the pvcrt-o (p<0.001) and

55
pvmdr-1 (p=0.007) genes among hospitalized patients (Figure 4).
Discussion
There have been recent reports on CQR from different regions of the world (37, 54, 55, 58,
82, 138, 163), including Brazil (102, 103, 158). Although the relevance of this problem, the
molecular mechanisms of CQRPv are poorly understood. In this work, expression levels of
pvcrt-o and pvmdr-1 genes were determined in patients with CQRPv from a well-
characterized cohort under supervised treatment, pointing to a link between in vivo CQR and
overexpression of both genes. Furthermore, these patients showed higher concentrations of
CQ and microsatellite revealed the presence of the same clonal nature at D0 and DR, bringing
evidence that the re-emergent infection is the same as the primary infection, therefore
indicating recrudescence.
It is likely that CQR contributes to the burden of severe vivax malaria in regions where
resistance is emerging (107, 164), but there is paucity of available data regarding this issue. In
this work, the group of patients with severe vivax malaria presented an increase in pvcrt-o and
pvmdr-1 expression levels, as compared to the control group with mild disease. These two
genes have been suggested as genetic markers of CQR in P. vivax (76, 78).
Our results are in concordance with those reported by Fernandez-Becerra et al. (77), where
high levels of pvcrt-o and pvmdr-1 gene expression were observed in a patient with severe
vivax malaria compared to three non-severe. A prospective study conducted in the Papua New
Guinea highlighted P. vivax as a major cause of severe malaria, particularly in settings with
established or emerging chloroquine resistance, supporting indirectly our findings (100). One
speculates that in areas in which CQR P. vivax circulates, acute febrile illness potentially
becomes chronic, recurrent and severe, due to recurrent parasitemia (136). Moreover, chronic
infections could lead to diserythropoiesis, as is seen in P. falciparum-induced anemia (137).

56
In this investigation, CQR and severe P. vivax malaria were observed in parallel with a high
expression of pvcrt-o and/or pvmdr-1 genes in patients seen at a reference center in the
Brazilian Amazon. Even though it is tempting to conclude about a causal association, a clear
limitation of this study is that CQR has not been evaluated in the same patients with severe
vivax malaria, especially because severe patients were in use of intravenous artesunate
(ACT), making it impossible to evaluate CQ-susceptibility in vivo phenotype. Actually,
patients with CQRPv were identified from following WHO criteria, excluding patients with
any clinical complication (36). Another clear limitation is the small number of samples used
in this study. Moreover, so far it is not possible to rule out a differential virulence concurring
with dissimilar drug resistance phenotypes. Studies on P. falciparum laboratory strains and
field isolates of P. vivax suggest that there is marked variability in growth rates of isolates ex
vivo with CQR isolates growing faster than CQ-susceptible isolates (59, 165). Since parasites
from patients with severe disease have greater ex vivo multiplication rates compared to those
from patients with non-severe disease (166), the possibility arises that the highly CQR isolates
may be more virulent.
Mefloquine (MQ) was used in the Brazilian Amazon until the beginning of the 2000s. Indeed,
CQ and MQ resistance parallel in this region (54). Thus, even after more than one decade of
non-exposure to MQ, we could not discard this effect in the present. Actually, increased
pvmdr-1 copy number was found in two isolates in the Western Brazilian Amazon (84).
Unfortunately, pvmdr-1 copy numbers were not measured in our samples.
In conclusion, our study suggests that CQR and severe P. vivax malaria are associated with
increased expression levels of the pvcrt-o and pvmdr-1 genes. CQRPv could be rising in the
Brazilian Amazon and probably this fact could be contributing to the simultaneous spread of
vivax malaria and clinical severity related to this parasite. Further studies are needed with
larger number of patients and using other molecular markers to confirm the underlying
pathogenesis of severe disease, and the degree to which this is related to the emergence of
multidrug resistant strains of P. vivax.

57
Acknowledgements
We acknowledge Omaira Veras for helpful discussion on microsatelite analysis, Nélida
Teresa Sánchez Rúa, Maria Raimunda Costa, Wellington Silva and Kim Machado for samples
collection and José Luiz Vieira and Marly Marques de Melo for drugs measurement.
Authors’ contributions
Conceived and designed the experiments: GCM CFB MVGL. Analyzed the data: GCM
WMM AMS AK HAP CFB. Wrote the paper: GCM WMM HAP CFB MVGL. Acquisition
of data: GCM AMS SRS BMLM ACCA. Critical revision of the manuscript: GCM WMM
AMS HAP CFB MVGL.
References
1. Baird JK (2011) Resistance to chloroquine unhinges vivax malaria therapeutics.
Antimicrob Agents Chemother 55: 1827–1830.
2. Tjitra E, Anstey NM, Sugiarto P, Warikar N, Kenangalem E, et al. (2008) Multidrug-
resistant Plasmodium vivax associated with severe and fatal malaria: a prospective
study in Papua, Indonesia. PLoS Med 5: e128.
3. Lanca EF, Magalhaes BM, Vitor-Silva S, Siqueira AM, Benzecry SG, et al. (2012) Risk
factors and characterization of Plasmodium vivax-associated admissions to pediatric
intensive care units in the Brazilian Amazon. PLoS One 7: e35406.
4. Genton B, D'Acremont V, Rare L, Baea K, Reeder JC, et al. (2008) Plasmodium vivax and
mixed infections are associated with severe malaria in children: a prospective cohort
study from Papua New Guinea. PLoS Med 5: e127.
5. Price RN, Douglas NM, Anstey NM (2009) New developments in Plasmodium vivax
malaria: severe disease and the rise of chloroquine resistance. Curr Opin Infect Dis 22:
430–435.
6. Rieckmann KH, Davis DR, Hutton DC (1989) Plasmodium vivax resistance to
chloroquine? Lancet 2: 1183–1184.

58
7. Baird JK, Basri H, Purnomo, Bangs MJ, Subianto B, et al. (1991) Resistance to chloroquine
by Plasmodium vivax in Irian Jaya, Indonesia. Am J Trop Med Hyg 44: 547–552.
8. Alecrim MdG, Alecrim W, Macedo V (1999) Plasmodium vivax resistance to chloroquine
(R2) and mefloquine (R3) in Brazilian Amazon region. Rev Soc Bras Med Trop 32:
67–68.
9. de Santana Filho FS, Arcanjo AR, Chehuan YM, Costa MR, Martinez-Espinosa FE, et al.
(2007) Chloroquine-resistant Plasmodium vivax, Brazilian Amazon. Emerg Infect Dis
13: 1125–1126.
10. Alexandre MA, Ferreira CO, Siqueira AM, Magalhaes BL, Mourao MP, et al. (2010)
Severe Plasmodium vivax malaria, Brazilian Amazon. Emerg Infect Dis 16: 1611–
1614.
11. Lacerda MV, Fragoso SC, Alecrim MG, Alexandre MA, Magalhaes BM, et al. (2012)
Postmortem Characterization of Patients With Clinical Diagnosis of Plasmodium
vivax Malaria: To What Extent Does This Parasite Kill? Clin Infect Dis 55: e67–74.
12. Lacerda MV, Mourao MP, Alexandre MA, Siqueira AM, Magalhaes BM, et al. (2012)
Understanding the clinical spectrum of complicated Plasmodium vivax malaria: a
systematic review on the contributions of the Brazilian literature. Malar J 11: 12.
13. Das S, Chakraborty SP, Hati AK, Roy S (2013) Association between prevalence of
chloroquine resistance and unusual mutation in pfmdr-I and pfcrt genes in India. Am J
Trop Med Hyg 88: 828–834.
14. Ngo T, Duraisingh M, Reed M, Hipgrave D, Biggs B, et al. (2003) Analysis of pfcrt,
pfmdr1, dhfr, and dhps mutations and drug sensitivities in Plasmodium falciparum
isolates from patients in Vietnam before and after treatment with artemisinin. Am J
Trop Med Hyg 68: 350–356.
15. Suwanarusk R, Russell B, Chavchich M, Chalfein F, Kenangalem E, et al. (2007)
Chloroquine resistant Plasmodium vivax: in vitro characterisation and association with
molecular polymorphisms. PLoS One 2: e1089.
16. Barnadas C, Ratsimbasoa A, Tichit M, Bouchier C, Jahevitra M, et al. (2008) Plasmodium
vivax resistance to chloroquine in Madagascar: clinical efficacy and polymorphisms in
pvmdr1 and pvcrt-o genes. Antimicrob Agents Chemother 52: 4233–4240.
17. Nomura T, Carlton JM, Baird JK, del Portillo HA, Fryauff DJ, et al. (2001) Evidence for
different mechanisms of chloroquine resistance in 2 Plasmodium species that cause
human malaria. J Infect Dis 183: 1653–1661.

59
18. Fernandez-Becerra C, Pinazo MJ, Gonzalez A, Alonso PL, del Portillo HA, et al. (2009)
Increased expression levels of the pvcrt-o and pvmdr1 genes in a patient with severe
Plasmodium vivax malaria. Malar J 8: 55.
19. WHO (2006) Guidelines for the treatment of malaria. Geneva.
20. Soto J, Toledo J, Gutierrez P, Luzz M, Llinas N, et al. (2001) Plasmodium vivax clinically
resistant to chloroquine in Colombia. Am J Trop Med Hyg 65: 90–93.
21. Ruebush TK, 2nd, Zegarra J, Cairo J, Andersen EM, Green M, et al. (2003) Chloroquine-
resistant Plasmodium vivax malaria in Peru. Am J Trop Med Hyg 69: 548–552.
22. Naing C, Aung K, Win DK, Wah MJ (2010) Efficacy and safety of chloroquine for
treatment in patients with uncomplicated Plasmodium vivax infections in endemic
countries. Trans R Soc Trop Med Hyg 104: 695–705.
23. WHO (2012) Management of severe malaria: a practical handbook Geneva
24. WHO (2010) Malaria Report: 2010.
25. MS (2009) Manual de Diagnóstico Laboratorial da Malária. In: Epidemiológica DdV,
editor. Brasilia.
26. Snounou G, Viriyakosol S, Zhu XP, Jarra W, Pinheiro L, et al. (1993) High sensitivity of
detection of human malaria parasites by the use of nested polymerase chain reaction.
Mol Biochem Parasitol 61: 315–320.
27. Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25: 402-408.
28. Orjuela-Sanchez P, de Santana Filho FS, Machado-Lima A, Chehuan YF, Costa MR, et
al. (2009) Analysis of single-nucleotide polymorphisms in the crt-o and mdr1 genes of
Plasmodium vivax among chloroquine-resistant isolates from the Brazilian Amazon
region. Antimicrob Agents Chemother 53: 3561–3564.
29. Gama BE, Oliveira NK, Souza JM, Daniel-Ribeiro CT, Ferreira-da-Cruz Mde F (2009)
Characterisation of pvmdr1 and pvdhfr genes associated with chemoresistance in
Brazilian Plasmodium vivax isolates. Mem Inst Oswaldo Cruz 104: 1009–1011.
30. Gama BE, Lacerda MV, Daniel-Ribeiro CT, Ferreira-da-Cruz Mde F (2011)
Chemoresistance of Plasmodium falciparum and Plasmodium vivax parasites in
Brazil: consequences on disease morbidity and control. Mem Inst Oswaldo Cruz 106
Suppl 1: 159–166.
31. Baird JK (2004) Chloroquine resistance in Plasmodium vivax. Antimicrob Agents
Chemother 48: 4075–4083.

60
32. Marfurt J, Mueller I, Sie A, Maku P, Goroti M, et al. (2007) Low efficacy of amodiaquine
or chloroquine plus sulfadoxine-pyrimethamine against Plasmodium falciparum and
P. vivax malaria in Papua New Guinea. Am J Trop Med Hyg 77: 947–954.
33. Sa JM, Yamamoto MM, Fernandez-Becerra C, de Azevedo MF, Papakrivos J, et al.
(2006) Expression and function of pvcrt-o, a Plasmodium vivax ortholog of pfcrt, in
Plasmodium falciparum and Dictyostelium discoideum. Mol Biochem Parasitol 150:
219–228.
34. Brega S, Meslin B, de Monbrison F, Severini C, Gradoni L, et al. (2005) Identification of
the Plasmodium vivax mdr-like gene (pvmdr1) and analysis of single-nucleotide
polymorphisms among isolates from different areas of endemicity. J Infect Dis 191:
272–277.
35. Menendez C, Fleming AF, Alonso PL (2000) Malaria-related anaemia. Parasitol Today
16: 469–476.
36. Reilly HB, Wang H, Steuter JA, Marx AM, Ferdig MT (2007) Quantitative dissection of
clone-specific growth rates in cultured malaria parasites. Int J Parasitol 37: 1599–
1607.
37. Russell B, Chalfein F, Prasetyorini B, Kenangalem E, Piera K, et al. (2008) Determinants
of in vitro drug susceptibility testing of Plasmodium vivax. Antimicrob Agents
Chemother 52: 1040–1045.
38. Chotivanich K, Udomsangpetch R, Simpson JA, Newton P, Pukrittayakamee S, et al.
(2000) Parasite multiplication potential and the severity of Falciparum malaria. J
Infect Dis 181: 1206–1209.
39. Vargas-Rodriguez RD, da Silva Bastos M, Menezes MJ, Orjuela-Sanchez P, Ferreira MU
(2012) Single-Nucleotide Polymorphism and Copy Number Variation of the
Multidrug Resistance-1 Locus of Plasmodium vivax: Local and Global Patterns. Am J
Trop Med Hyg.

61
Figure 1 - Expression levels of chloroquine resistance genes in patients with CQRPv
parasites. Relative quantification of pvcrt-o (A) and pvmdr1 (B) transcripts in total RNA
obtained from parasites from patients with chloroquine-resistant P. vivax vs a pool of total
RNA obtained from 5 patients with CQ-susceptible parasites. Chloroquine-resistant P. vivax
parasites (R). Chloroquine-susceptible P. vivax parasites (S). Day of admission (D0). Day
of recrudescence (DR). The error bars in Figures 1 and 3 reflect the average standard error of
the Ct.

62
Figure 2 - Expression gene levels of pvcrt-o and pvmdr-1 in all the groups. Relative
quantification of pvcrt-o (A) and pvmdr1 (B) transcript levels in total RNA obtained from
parasites from severe patients with chloroquine-resistant P. vivax, patients susceptible to CQ,
severe patients and patients without severity symptoms. Chloroquine-resistant P. vivax
parasites (R). Chloroquine-susceptible P. vivax parasites at D0 (S). Severe cases (Sev).
Non-severe cases (NS). *p<0.05

63
Figure 3 - Expression level of chloroquine resistance genes in severe patients.
Relative quantification of pvcrt-o (A) and pvmdr1 (B) transcript levels in total RNA
obtained from parasites from severe patients vs a pool of total RNA obtained from
parasites susceptible to CQ. Severe cases (S). Non-severe cases (NS). The error bars in
Figures 1 and 3 reflect the average standard error of the Ct.
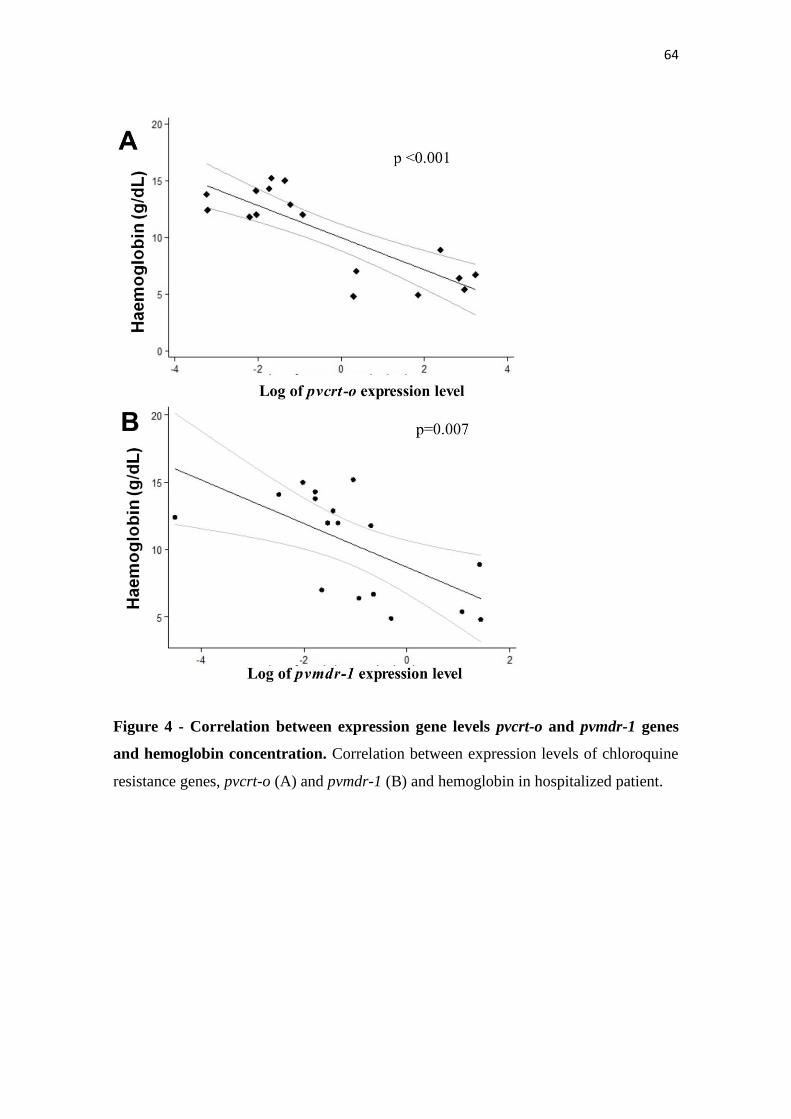
64
Figure 4 - Correlation between expression gene levels pvcrt-o and pvmdr-1 genes
and hemoglobin concentration. Correlation between expression levels of chloroquine
resistance genes, pvcrt-o (A) and pvmdr-1 (B) and hemoglobin in hospitalized patient.

65
Table 1. Demographic and clinical characteristics of the study participants admitted to a tertiary health center, Manaus, Amazonas,
Brazil.
Code
Sex
/Age
DR
Haemoglobin
level in D0
(g/dL)
Haemoglobin
level in DR
(g/dL)
Chloroquine
concentrations
(ng/dL)
Asexual malarial
parasites/ µl
in D0
Asexual malarial
parasites/µl
in DR
Day of
RNA
Isolation
Genotyping
R1 M/55 27 14 15.1 249 690.0 5695.0 D0/DR Same
R2 F/9 32 10.2 11.6 61 4347.0 6871.0 D0/DR Same
R3 M/23 34 13.4 12.6 68 297.0 3734.6 D0/DR Same
R4 M/42 36 14.2 13.5 256 686.0 436.8 D0/DR Same
R5 M/13 38 12.4 12.0 103 2582.1 2613.2 D0/DR Same
R6 M/25 25 15.2 15.6 533 1974.7 5385.6 D0/DR Same
R7 M/37 28 14.0 14.4 263 2548.0 50.0 D0/DR Same
R8 M/26 32 13.5 12.9 188 78.0 11096.3 D0 Same
R9 M/9 31 9.4 10.0 230 1136.8 4968.0 D0 Same
R10 M/47 31 12.9 12.0 130 5059.2 4160.0 D0 Different
R11 M/36 34 14.7 14.5 118 2105.6 2853.6 DR Different
R12 F/44 34 13.6 13.1 111 676.8 2613.2 DR Same
Day admission (D0). Day of recrudescence (DR).

66
Table 2. Frequency of polymorphisms in the pvmdr1 and pvcrt-o genes for the different set of patients.
Genotype
Number of samples (%)
P
Number of samples (%)
p
Chloroquine-resistant Chloroquine-susceptible Severe malária Non-severe malaria
pvmdr-1
Mutant L1076 codon 0/12 (0.0) 0/15 (0.0) NS 1/7 (14.3) 0/10 (0.0) NS
Wild-type Y976 codon 12/12 (100.0) 15/15 (100.0) NS 7/7 (100.0) 10/10 (100.0) NS
pvcrt-o
Wild-type without K10 insert 12/12 (100.0) 15/15 (100.0) NS 7/7 (100.0) 10/10 (100.0) NS
Not significant (NS)
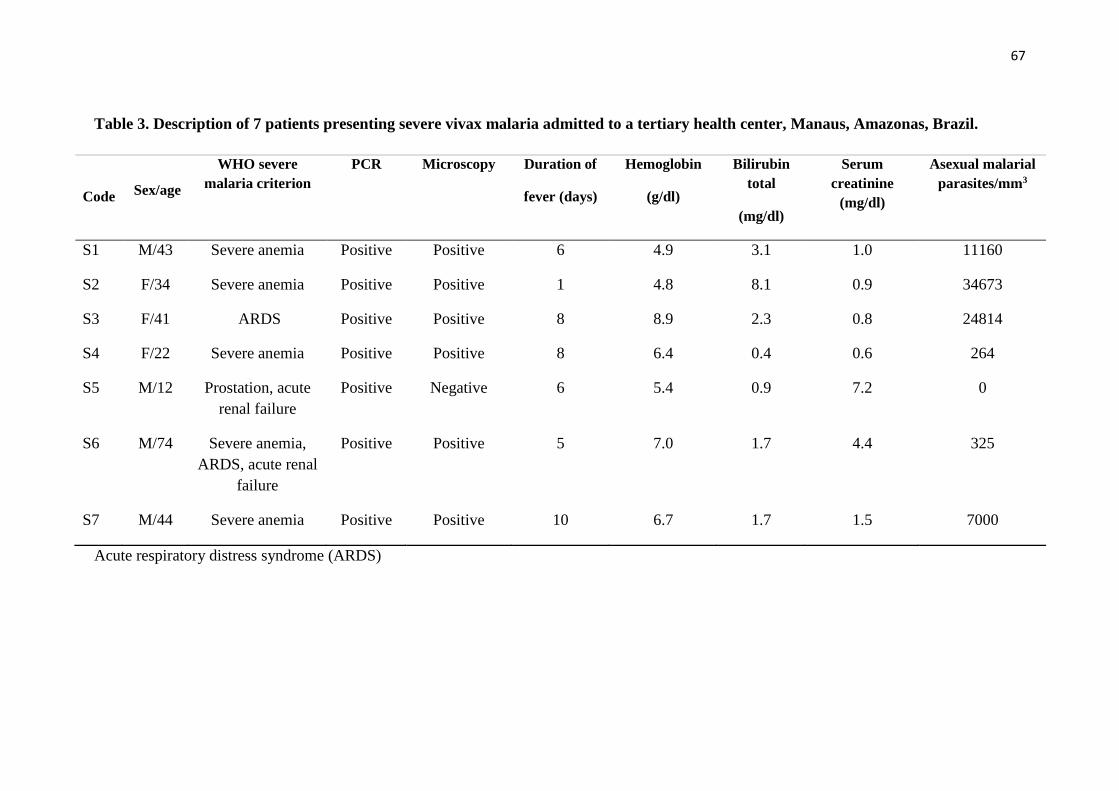
67
Table 3. Description of 7 patients presenting severe vivax malaria admitted to a tertiary health center, Manaus, Amazonas, Brazil.
Code Sex/age
WHO severe
malaria criterion
PCR Microscopy Duration of
fever (days)
Hemoglobin
(g/dl)
Bilirubin
total
(mg/dl)
Serum
creatinine
(mg/dl)
Asexual malarial
parasites/mm3
S1 M/43 Severe anemia Positive Positive 6 4.9 3.1 1.0 11160
S2 F/34 Severe anemia Positive Positive 1 4.8 8.1 0.9 34673
S3 F/41 ARDS Positive Positive 8 8.9 2.3 0.8 24814
S4 F/22 Severe anemia Positive Positive 8 6.4 0.4 0.6 264
S5 M/12 Prostation, acute
renal failure
Positive Negative 6 5.4 0.9 7.2 0
S6 M/74 Severe anemia,
ARDS, acute renal
failure
Positive Positive 5 7.0 1.7 4.4 325
S7 M/44 Severe anemia Positive Positive 10 6.7 1.7 1.5 7000
Acute respiratory distress syndrome (ARDS)
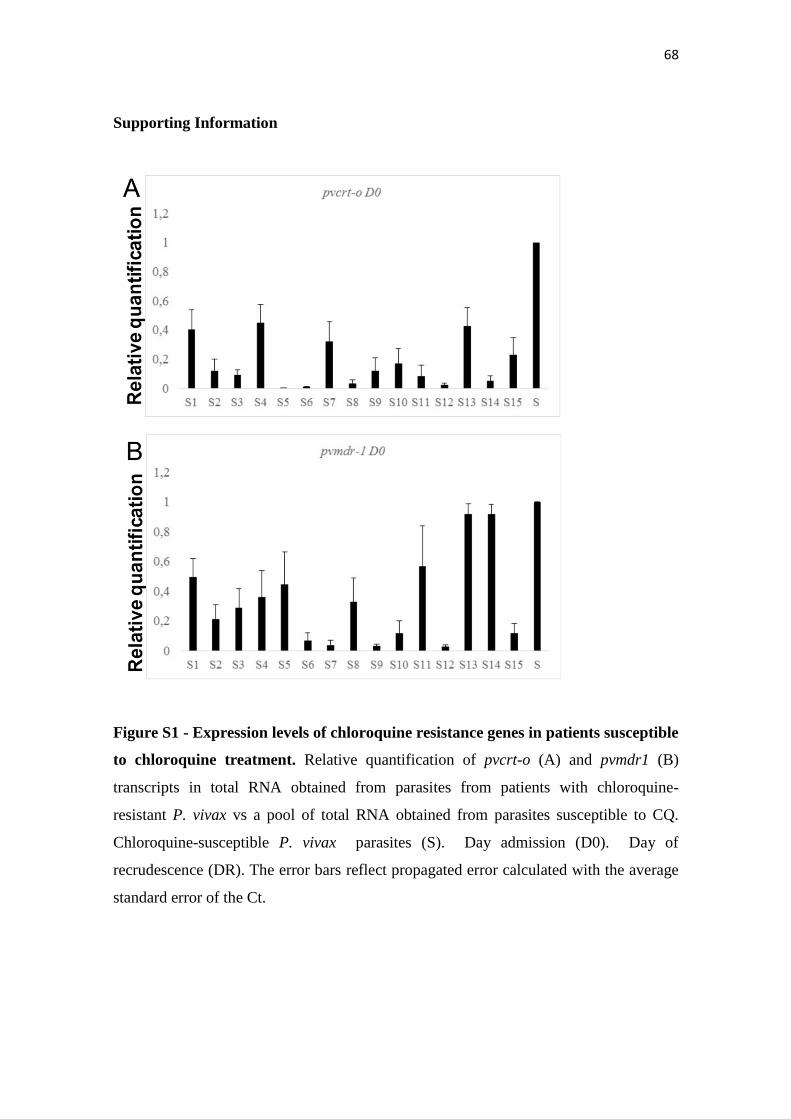
68
Supporting Information
Figure S1 - Expression levels of chloroquine resistance genes in patients susceptible
to chloroquine treatment. Relative quantification of pvcrt-o (A) and pvmdr1 (B)
transcripts in total RNA obtained from parasites from patients with chloroquine-
resistant P. vivax vs a pool of total RNA obtained from parasites susceptible to CQ.
Chloroquine-susceptible P. vivax parasites (S). Day admission (D0). Day of
recrudescence (DR). The error bars reflect propagated error calculated with the average
standard error of the Ct.

69
Figure S2 - Expression level of chloroquine resistance genes in non-severe P. vivax
malaria. Relative quantification of pvcrt-o (A) and pvmdr1 (B) transcript levels in total
RNA obtained from parasites from severe patients vs a pool of total RNA obtained from
parasites susceptible to CQ. Non-severe cases (NS). The error bars reflect propagated
error calculated with the average standard error of the Ct.

70
Table S1. Oligonucleotide primers used for DNA sequencing of P. vivax orthologs genes.
Gene Chromosome Sequence 5’ 3’ Base pairs Use References
pvcrt-o 1 AAGAGCCGTCTAGCCATCC 1,186 bps Sequencing (92)
AGTTTCCCTCTACACCCG pvmdr-1 10 GGATAGTCATGCCCCAGGATTG 604 bps Sequencing (92, 150)
CATCAACTTCCCGGCGTAGC

71
Table S2. Oligonucleotides used for genotyping P. vivax parasites.
Marker Cromossome Marker type Primer name Sequence 5' - 3' Fluorescent
dye
TM (ºC) Mg
MS2 3 Microsatellite primary forward AGCACGACCAACAAGAGAGG 6-FAM 59 25
nested forward GAGCTAGCCAAAGGTTCAAA
Nested reverse TGGGGAGAGACTCCCTTTTC
msp1F3 7 Surface
antigen
primary forward GGAGAACATAAGCTACCTGTCC VIC 59 25
primary reverse GTTGTTACTTGGTCTTCCTCCC
nested forward CAAGCCTACCAAGAATTGATCCCCAA
nested reverse ATTACTTTGTCGTAGTCCTCGGCGTAGTCC
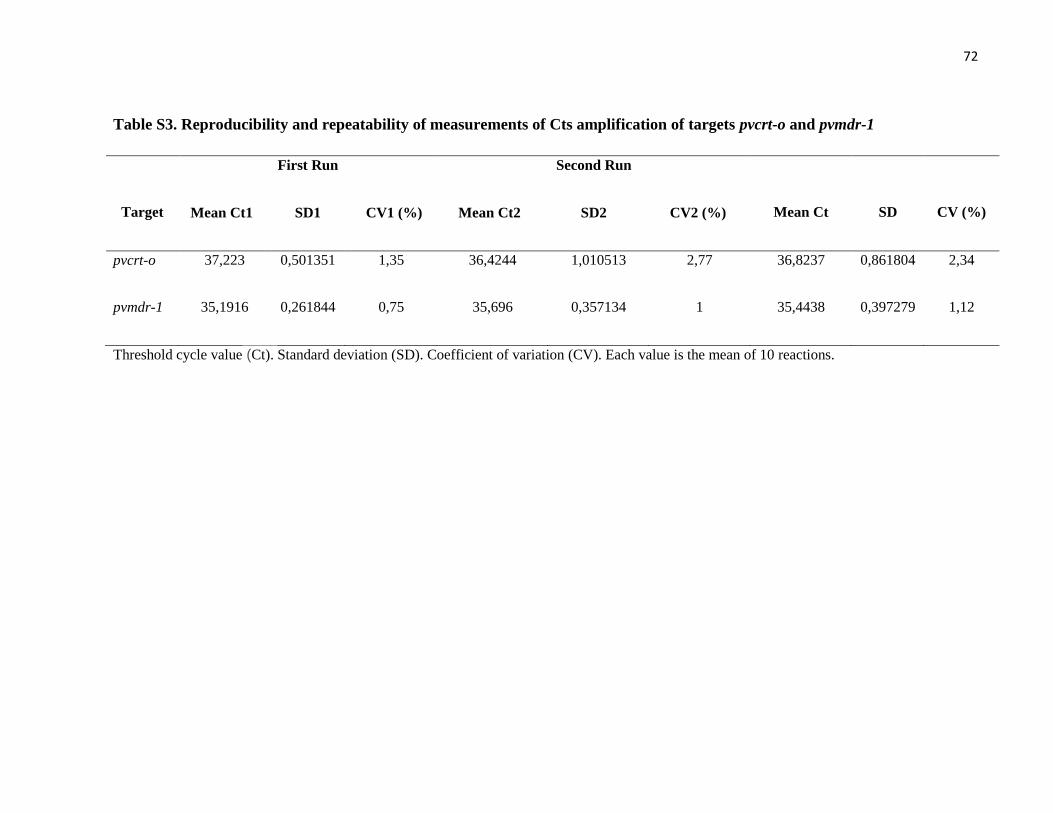
72
Table S3. Reproducibility and repeatability of measurements of Cts amplification of targets pvcrt-o and pvmdr-1
Target
First Run Second Run
Mean Ct
SD
CV (%) Mean Ct1 SD1 CV1 (%) Mean Ct2 SD2 CV2 (%)
pvcrt-o 37,223 0,501351 1,35 36,4244 1,010513 2,77 36,8237 0,861804 2,34
pvmdr-1 35,1916 0,261844 0,75 35,696 0,357134 1 35,4438 0,397279 1,12
Threshold cycle value (Ct). Standard deviation (SD). Coefficient of variation (CV). Each value is the mean of 10 reactions.
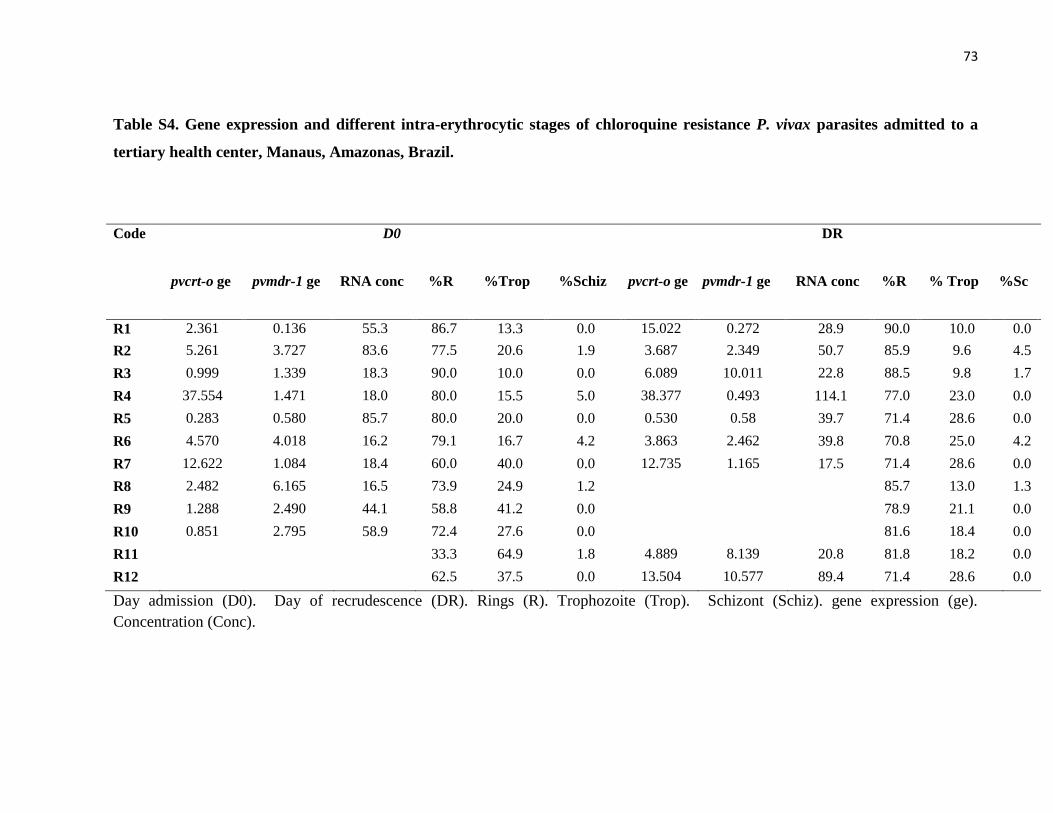
73
Table S4. Gene expression and different intra-erythrocytic stages of chloroquine resistance P. vivax parasites admitted to a
tertiary health center, Manaus, Amazonas, Brazil.
Code D0 DR
pvcrt-o ge pvmdr-1 ge RNA conc %R %Trop %Schiz pvcrt-o ge pvmdr-1 ge RNA conc %R % Trop %Sc
R1 2.361
0.136
55.3 86.7 13.3 0.0 15.022
0.272
28.9 90.0 10.0 0.0
R2 5.261
3.727
83.6 77.5 20.6 1.9 3.687
2.349
50.7 85.9 9.6 4.5
R3 0.999
1.339
18.3 90.0 10.0 0.0 6.089
10.011
22.8 88.5 9.8 1.7
R4 37.554
1.471
18.0 80.0 15.5 5.0 38.377
0.493
114.1 77.0 23.0 0.0
R5 0.283
0.580
85.7 80.0 20.0 0.0 0.530
0.58
39.7 71.4 28.6 0.0
R6 4.570
4.018
16.2 79.1
16.7 4.2 3.863
2.462
39.8 70.8 25.0 4.2
R7 12.622
1.084
18.4 60.0 40.0 0.0 12.735
1.165
17.5 71.4 28.6 0.0
R8 2.482
6.165
16.5 73.9 24.9 1.2 85.7 13.0 1.3
R9 1.288
2.490
44.1 58.8 41.2 0.0 78.9 21.1 0.0
R10 0.851
2.795
58.9 72.4 27.6 0.0 81.6 18.4 0.0
R11 33.3 64.9 1.8 4.889
8.139
20.8 81.8 18.2 0.0
R12 62.5 37.5 0.0 13.504
10.577
89.4 71.4 28.6 0.0
Day admission (D0). Day of recrudescence (DR). Rings (R). Trophozoite (Trop). Schizont (Schiz). gene expression (ge).
Concentration (Conc).

74
Table S5. Different intra-erythrocytic stages of chloroquine-susceptible P. vivax parasites admitted to a tertiary health center,
Manaus, Amazon, Brazil.
S1 S2 S3 S4 S5 S6 S7 S8 S9 S10 S11 S12 S13 S14 S15
% Rings 76.2
80.0 83.3
44.4 40.0 82.4
29.4 38.9 77.5 54.6 66.3 61.4 38.9 87.7 73.3
% Trophozoites 23.8 16.0 16.7 55.6 54.6 17.6 70.6 61.1 20.0 36.9 33.7 36.9 61.1 12.3 26.7
% Schizonts 0 4.0 0.0 0.0 5.4 0.0 0.0 0.0 2.5 9.0 0.0 1.7 0.0 0.0 0.0
Chloroquine-susceptible P. vivax parasites (S).

75
Table S6. Gene expression and different intra-erythrocytic stages of patients presenting severe vivax malaria admitted to a
tertiary health center, Manaus, Amazonas, Brazil.
Code pvcrt-o gene expression pvmdr-1 gene
expression
RNA Concentration
(ng/ml)
%Rings stages %Trophozoite %Schizont
S1 4.890 8.334 40.6 77.8 11.1 11.1
S2 2.310 0.572 91.1 73.0 18.0 9.0
S3 18.840
2.906 102.4 54.1 41.7 4.2
S4 4.890
5.294 106.3 80.0 20.0 0.0
S5 5.147
23.256 230.2 79.6 20.4 0.0
S6 1.067
1.469 79.7 75.0 25.0 0.0
S7 189.112 1.091 20.1 74.6 25.4 0.0

76
5 DISCUSSÃO

77
5.1 Contribuição para a patogênese da resistência a cloroquina e malária grave por
Plasmodium vivax
A CQ continua sendo a droga de escolha para o tratamento da malária não grave por P. vivax,
em muitas áreas endêmicas incluindo o Brasil, mas a resistência tem aumentado desde que foi
descrita pela primeira vez em Papua Nova Guiné (44) e Indonésia (159, 167). Diversos trabalhos
têm demonstrado que a CQR tem aumentado em todo o mundo. (37, 54, 55, 58, 82, 138, 163),
inclusive no Brasil (102, 103, 158). Apesar da relevância desse problema, o mecanismo
molecular ainda não foi totalmente elucidado. Neste presente trabalho, o nível de expressão
gênica de pvcrt-o e pvmdr1 foi determinado em pacientes com CQRPv em uma coorte bem
caracterizada em tratamento supervisionado, evidenciando uma associação entre CQR in vivo e
aumento na expressão desses genes. Além disso, esses pacientes apresentaram alta concentração
plasmática de CQ e microssatélites revelaram a presença do mesmo clone em D0 e DR indicando
que seja uma recrudescência.
Diversas pesquisas têm demonstrado que mutações nos genes pfcrt e pfmdr1 têm sido
associadas com resistência em P. falciparum (63, 160, 168). A falta de um sistema de cultivo
contínuo in vitro para os estágios sanguíneos do P. vivax tem dificultado o uso da genética para
estudar a função destes genes. No entanto, linhagens transgênicas de P. falciparum
superexpressando pvcrt-o apresentaram redução significativa de 2,2 vezes na sensibilidade à CQ
(76). Este resultado sugere que os níveis de expressão e não as mutações poderiam estar
associadas com resistência à CQ em P. vivax.
Verificamos que as mutações gênicas em pvcrt-o e pvmdr1 não foram associadas com
resistência à CQ, concordando com os resultados apresentados em outros trabalho (56, 60, 79).
Estes resultados justificam pesquisas futuras focalizando mutações genéticas nas regiões
promotoras desses genes alvo que poderiam estar influenciando na expressão desses genes.

78
Neste presente estudo foi verificado que a expressão gênica do gene pvcrt-o e pvmdr1 foi
significativamente maior tanto no D0 quanto no DR nos pacientes resistentes à CQ. Esses
resultados sugerem a expressão do gene pvcrt-o e pvmdr1 poderia funcionar como um marcador
de resistência à CQ, indicando que os biomarcadores inovadores, como expressão de genes
provavelmente relacionadas à resistência aos medicamentos poderia ser utilizada para mapear a
resistência em centros sentinela de referência.
É provável que CQR contribui para a carga da malária vivax grave em regiões onde a
resistência está surgindo (107, 164), mas há escassez de dados disponíveis. Neste trabalho, o
grupo de pacientes com malária vivax grave apresentaram um aumento na expressão gênica de
pvcrt-o e pvmdr1 em comparação com o grupo com malária não grave. Estes dois genes têm sido
sugeridos como marcadores genéticos de CQR em P. vivax (76, 78).
Nossos resultados estão em concordância com os relatados por Fernandez-Becerra et al. (77),
em que foram observados altos níveis de expressão gênica de pvcrt-o e pvmdr1 em um paciente
com malária grave vivax em comparação com três não-grave. Nossos resultados, portanto,
corroboram com um número maior de amostras. Um estudo prospectivo realizado em Papua-
Nova Guiné destacou que o P. vivax como uma das principais causas da malária grave,
particularmente em locais com resistência à CQ, apoiando indiretamente nossos resultados (100).
Especula-se que em áreas que CQRPv circula, doença febril aguda, potencialmente, torna-se
crônica, recorrente e grave, devido a parasitemia recorrente (136). Além disso, as infecções
crônicas podem levar a diseretropoiese, como é visto na anemia induzida por P. falciparum (137).
Neste trabalho, CQR e malária vivax grave foram observados em paralelo com uma alta
expressão gênica de pvcrt-o e/ou pvmdr1 em pacientes atendidos em um centro de referência na
Amazônia Brasileira. Mesmo que seja tentador concluir sobre uma associação causal, uma clara
limitação do estudo é que CQR não foi avaliada nos mesmos doentes com malária vivax,

79
especialmente porque os pacientes graves foram tratados com ACT intravenoso, tornando-se
impossível avaliar susceptibilidade a CQ in vivo. Na verdade, os pacientes com CQRPv foram
identificados a partir dos critérios da OMS, excluindo os pacientes com qualquer complicação
clínica (36). Outra limitação é o pequeno número de amostras usadas neste estudo. Além disso,
não é possível descartar uma virulência diferencial concordando com os fenótipos de resistência à
medicamentos diferentes. Estudos sobre a estirpes laboratoriais de P. falciparum e isolados de P.
vivax sugerem existir alta variabilidade nas taxas de crescimento dos isolados ex vivo, sendo que
isolados CQR crescem mais rápido do que os isolados CQ-susceptíveis (59, 165). Uma vez que
os parasitas de pacientes com doença grave têm maiores velocidades de multiplicação ex vivo em
comparação com os pacientes com doença não grave (166), surge a possibilidade de que a CQR
altamente isolados podem ser mais virulento.
Foi observado que dois pacientes CQRPv apresentaram concentração de CQ < 100 ng/mL,
porém foi observado que os marcadores moleculares mostraram alelos idênticos no D0 e DR.
Diversas linhas de pesquisa apoiaram suposições e ajudou a estimar a concentração mínima
eficaz de CQ e DCQ para classificar um pacientes em resistente e sensível em ensaio in vivo.
Alguns trabalhos observaram que a concentração foi de 120 ng/mL no DR (169, 170). Rombo et
al. observou um valor de 90 ng/mL no DR (171). Finalmente, foi observado que a concentração
se aproximava a 100 ng/mL no D35 que é quando começam a surgir as recaídas, sendo esse valor
estimado como concentração mínima eficaz de CQ e DCQ no sangue total. Apesar disso, esse
valor como concentração mínima não foi padronizado e validado para diversas regiões diferentes
do mundo e pode não ser que no Brasil seja diferente.
É de crucial relevância o desenvolvimento de um ensaio de cultivo in vitro padronizados e
validados para avaliar à susceptibilidade de isolados de P. vivax aos antimaláricos, para que
pudéssemos avaliar a expressão gênica de pvcrt-o e pvmdr1 em isolados in vitro. Existem
diversos fatores que podem influenciar o ensaio de susceptibilidade in vitro: parasitemia deve ser
superior a 0,5%; amostra deve ser processada em CF11 para retirar os leucócitos; os trofozoítos

80
maduros são insensível à CQ (59, 172) sendo importante que os trofozoítos jovens predominem
na amostra (90%).
Mefloquina foi usado na Amazônia Brasileira até o início dos anos 2000. Na verdade, CQ e
MQ apresentam resistência em paralelo na região (54). Assim, mesmo depois de mais de uma
década de não-exposição a MQ, não poderíamos descartar esse efeito. Foi demonstrado o
aumento do número de cópia de pvmdr1 em dois isolados na Amazônia Ocidental Brasileira
tratados com MQ. (84). Infelizmente, o número de cópias de pvmdr-1 não foi determinado em
nossas amostras, em função da limitação de volume de amostra coletada.
No presente trabalho, a PQ foi prescrita no dia 42 por um período de 7 dias. Em estudos para
avaliar a eficácia de CQ é importante que a PQ seja administrada no final do período de
acompanhamento pois a PQ + CQ no início do tratamento pode mascarar CQRPv e subestimar o
risco de falha terapêutica. Baird et al. observou que após um período de acompanhamento de 28
dias, a PQ age por sinergismo com CQ e foi mais eficaz para CQRPv (173). No Brasil,
demonstrou-se que a CQRPv foi maior quando tratada apenas com CQ (55) do que com CQ e PQ
(56).
5.2 Perspectivas futuras
A expressão gênica de pvcrt-o e pvmdr1 foi maior nos pacientes resistentes do que no grupo
susceptível à CQ desde o D0 e nos pacientes graves do que não graves sugerindo que este
biomarcador poderia ser utilizado como uma ferramenta inovadora para prever a resistência à CQ
e malária grave no momento do diagnóstico da malária vivax. No andamento deste projeto
evidenciou-se informações não conclusivas sobre os seguintes temas, que merecem ser
investigados futuramente:

81
1) Em recente estudo realizado em Manaus foi verificado que a prevalência de resistência à CQ
em estudos in vivo foi de 5,2% (174). Devido a frequência de resistência ser baixa a utilização de
maior número de amostras de Manaus e amostras procedentes de outras áreas com confirmadas
CQRPv permitirá conclusões mais confiáveis sobre a validação da expressão dos genes pvcrt-o e
pvmdr1 como biomarcador de resistência à CQ.
2) Trabalhos de ensaios ex vivo realizados na Tailândia sugerem que o aumento no número de
cópias de pvmdr1 está associado com aumento da susceptibilidade à MQ (93). MQ foi usado na
Amazônia brasileira até o início dos anos 2000. Estudos publicados mostram resistência à CQ e
MQ em paralelo na região (54). Em estudo realizado no Acre foi observado que somente 2
isolados apresentaram duas cópias do gene pvmdr1. Com isso, seria importante que fosse
realizado o número de cópias de pvmdr1 e verificar se a expressão gênica está associada com
resistência à CQ, independentemente do número de cópias de pvmdr1.
3) A definição de CQRPv seria a capacidade do parasita de crescer em altas concentração de
droga. Porém, no presente trabalho não verificamos correlação entre concentração de CQ e
expressão gênica de pvcrt-o e pvmdr1. Porém, a verificação se a pressão seletiva da CQ poderia
influenciar na expressão genica do P. vivax acrescentaria valiosa informação nas amostras
resistentes à CQ.
4) Estes resultados justificam pesquisas futuras focalizando mutações genéticas nas regiões
promotoras desses genes alvo que poderia estar influenciando na expressão desses genes.
5) Existe pouca informação sobre os mecanismos moleculares envolvidos na malária vivax grave
e resistência a cloroquina. Seria muito importante estudar outros genes candidatos como outros

82
genes transportadores de membrana que poderiam estar associado com resistência à CQ e malária
grave por P. vivax.

83
6 CONCLUSÕES

84
1) Parasitos resistentes à cloroquina apresentaram aumento na expressão dos genes pvcrt-o e
pvmdr1 tanto no dia de admissão quanto no dia de recrudescência.
2) Malária grave por P. vivax está associada com aumento na expressão desses genes.
3) Não foi observado diferença significativa na expressão gênica desses genes entre dia de
admissão e dia de recrudescência entre os parasitas resistentes à cloroquina.
4) Mutações gênicas em pvcrt-o e pvmdr-1 não foi associada com resistência à cloroquina.
5) Foi observado correlação negativa entre concentração de hemoglobina e expressão desses
genes entre os pacientes hospitalizados.

85
7 REFERÊNCIAS BIBLIOGRÁFICAS

86
1. Bruce-Chwatt LJ. Essential malariology. London: William Heinemann Medical
Books; 1980.
2. Gilles HM. Morbidity and mortality from parasitic disease in the year 2000. Ann
Parasitol Hum Comp. 1990;65 Suppl 1:43-4. Epub 1990/01/01.
3. Snow RW, Guerra CA, Noor AM, Myint HY, Hay SI. The global distribution of
clinical episodes of Plasmodium falciparum malaria. Nature. 2005;434(7030):214-7. Epub
2005/03/11.
4. Ferreira MS. Malária. In: Focaccia R, editor. Veronezi: tratado de infectologia. São
Paulo: Editora Atheneu; 2005. p. 1589-632.
5. Povoa MM, de Souza RT, Lacerda RN, Rosa ES, Galiza D, de Souza JR, et al. The
importance of Anopheles albitarsis E and An. darlingi in human malaria transmission in Boa
Vista, state of Roraima, Brazil. Mem Inst Oswaldo Cruz. 2006;101(2):163-8. Epub
2006/07/13.
6. Pessoa SB. Parasitologia Médica. 11 ed. ed. Rio de Janeiro: Guanabara Koogan; 1993.
7. Prudencio M, Rodriguez A, Mota MM. The silent path to thousands of: the merozoites
Plasmodium liver stage. Nat Rev Microbiol. 2006;4(11):849-56. Epub 2006/10/17.
8. Sturm A, Heussler V. Live and let die: manipulation of host hepatocytes by
exoerythrocytic Plasmodium parasites. Med Microbiol Immunol. 2007;196(3):127-33. Epub
2007/04/06.
9. Miller LH, Baruch DI, Marsh K, Doumbo OK. The pathogenic basis of malaria.
Nature. 2002;415(6872):673-9. Epub 2002/02/08.
10. Amino R, Thiberge S, Martin B, Celli S, Shorte S, Frischknecht F, et al. Quantitative
imaging of Plasmodium transmission from mosquito to mammal. Nat Med. 2006;12(2):220-4.
Epub 2006/01/24.
11. Barnwell JW, Galinski MR. Invasion of vertebrate cells: erythrocytes. In: Sherman
IW, editor. Malaria: Parasite Biology, Pathogenesis, and Protection. Washington, DC: ASM
Press; 1998. p. 93-120.
12. Sturm A, Amino R, van de Sand C, Regen T, Retzlaff S, Rennenberg A, et al.
Manipulation of host hepatocytes by the malaria parasite for delivery into liver sinusoids.
Science. 2006;313(5791):1287-90. Epub 2006/08/05.
13. Williams TN. Human red blood cell polymorphisms and malaria. Curr Opin
Microbiol. 2006;9(4):388-94. Epub 2006/07/04.
14. Gaur D, Mayer DC, Miller LH. Parasite ligand-host receptor interactions during
invasion of erythrocytes by Plasmodium merozoites. Int J Parasitol. 2004;34(13-14):1413-29.
Epub 2004/12/08.
15. Gaur D, Chitnis CE. Molecular interactions and signaling mechanisms during
erythrocyte invasion by malaria parasites. Curr Opin Microbiol. 2011. Epub 2011/08/02.
16. Garzon-Ospina D, Romero-Murillo L, Tobon LF, Patarroyo MA. Low genetic
polymorphism of merozoite surface proteins 7 and 10 in Colombian Plasmodium vivax
isolates. Infect Genet Evol. 2011;11(2):528-31. Epub 2010/12/25.
17. WHO. Severe falciparum malaria. World Health Organization, Communicable
Diseases Cluster. Trans R Soc Trop Med Hyg. 2000;94 Suppl 1:S1-90. Epub 2000/12/05.
18. Tuteja R. Malaria - an overview. FEBS J. 2007;274(18):4670-9. Epub 2007/09/11.

87
19. Guerra CA, Howes RE, Patil AP, Gething PW, Van Boeckel TP, Temperley WH, et
al. The international limits and population at risk of Plasmodium vivax transmission in 2009.
PLoS Negl Trop Dis. 2010;4(8):e774. Epub 2010/08/07.
20. Cowman AF, Crabb BS. Invasion of red blood cells by malaria parasites. Cell.
2006;124(4):755-66. Epub 2006/02/25.
21. Barillas-Mury C, Kumar S. Plasmodium-mosquito interactions: a tale of dangerous
liaisons. Cell Microbiol. 2005;7(11):1539-45. Epub 2005/10/07.
22. Mueller I, Galinski MR, Baird JK, Carlton JM, Kochar DK, Alonso PL, et al. Key
gaps in the knowledge of Plasmodium vivax, a neglected human malaria parasite. Lancet
Infect Dis. 2009;9(9):555-66. Epub 2009/08/22.
23. WHO. World Malaria: Report 2013. 2013.
24. WHO. World Malaria: Report 2012. 2012.
25. OPAS. Informe de la situación de los programas de malaria en las Américas. 2001.
26. WHO. World Malaria Report 2009. 2009.
27. Mendis K, Sina BJ, Marchesini P, Carter R. The neglected burden of Plasmodium
vivax malaria. Am J Trop Med Hyg. 2001;64(1-2 Suppl):97-106. Epub 2001/06/27.
28. Price RN, Tjitra E, Guerra CA, Yeung S, White NJ, Anstey NM. Vivax malaria:
neglected and not benign. Am J Trop Med Hyg. 2007;77(6 Suppl):79-87. Epub 2008/01/31.
29. WHO. World malaria report : 2010. 2010.
30. Gething PW, Elyazar IR, Moyes CL, Smith DL, Battle KE, Guerra CA, et al. A long
neglected world malaria map: Plasmodium vivax endemicity in 2010. PLoS Negl Trop Dis.
2012;6(9):e1814. Epub 2012/09/13.
31. MS. Situação epidemiológica da malária no Brasil, 2000 a 2011. 2013. p.
http://portalsaude.saude.gov.br/portalsaude/index.cfm?portal=pagina.visualizarTexto&codCo
nteudo=10252&codModuloArea=783&chamada=boletim-epidemiologico-da-malaria-_-2013.
32. Tauil PL. Avaliação de uma nova estratégia de controle da malária na Amazônia
Brasileira. Brasília: Universidade de Brasília; 2002.
33. Brasil MdS. Avaliação malaria_jan_maio_19_07_2010. 2010 [January 6, 2011];
Available from:
http://portal.saude.gov.br/portal/arquivos/pdf/avaliacao_malaria_jan_maio_19_07_2010.pdf
34. PAHO. Situation of Malaria in the Region of the Americas, 2000-2012. 2013.
35. FMT. Informe Epidemiológico Nº 12 - ANO V / 2010 da FMT. 2010 [updated
November 01, 2010]; Available from:
www.fmt.am.gov.br/informe/2010/MURAL%20MAL2007A2009.pdf
36. WHO. Guidelines for the treatment of malaria. Geneva2006.
37. Baird JK. Chloroquine resistance in Plasmodium vivax. Antimicrob Agents
Chemother. 2004;48(11):4075-83. Epub 2004/10/27.
38. Baird JK. Resistance to therapies for infection by Plasmodium vivax. Clin Microbiol
Rev. 2009;22(3):508-34. Epub 2009/07/15.
39. Baird JK, Leksana B, Masbar S, Suradi, Sutanihardja MA, Fryauff DJ, et al. Whole
blood chloroquine concentrations with Plasmodium vivax infection in Irian Jaya, Indonesia.
Am J Trop Med Hyg. 1997;56(6):618-20. Epub 1997/06/01.
40. WHO. Global report on antimalarial drug efficacy and drug resistance: 2000-2010.
Geneva2010.

88
41. Rosenthal PJ. Antimalarial chemotherapy: mechanisms of action, resistance, and new
directions in drug discovery. Totowa: MD Humana Press; 2001.
42. Coatney GR. Pitfalls in a discovery: the chronicle of chloroquine. Am J Trop Med
Hyg. 1963;12:121-8. Epub 1963/03/01.
43. Loeb RF, Clark WM, Coatney GR, Coggeshall LT, Dieuaide FR, Dochez AR, et al.
Activity of a new antimalarial agent, chloroquine (SN 7618). J Am Med Assoc.
1946;130:1069. Epub 1946/04/20.
44. Rieckmann KH, Davis DR, Hutton DC. Plasmodium vivax resistance to chloroquine?
Lancet. 1989;2(8673):1183-4. Epub 1989/11/18.
45. Dua VK, Kar PK, Sharma VP. Chloroquine resistant Plasmodium vivax malaria in
India. Trop Med Int Health. 1996;1(6):816-9. Epub 1996/12/01.
46. Myat Phone K, Myint O, Myint L, Thaw Z, Kyin Hla A, Nwe Nwe Y. Emergence of
chloroquine-resistant Plasmodium vivax in Myanmar (Burma). Trans R Soc Trop Med Hyg.
1993;87(6):687. Epub 1993/11/01.
47. Guthmann JP, Pittet A, Lesage A, Imwong M, Lindegardh N, Min Lwin M, et al.
Plasmodium vivax resistance to chloroquine in Dawei, southern Myanmar. Trop Med Int
Health. 2008;13(1):91-8. Epub 2008/02/23.
48. Chotivanich K, Udomsangpetch R, Chierakul W, Newton PN, Ruangveerayuth R,
Pukrittayakamee S, et al. In vitro efficacy of antimalarial drugs against Plasmodium vivax on
the western border of Thailand. Am J Trop Med Hyg. 2004;70(4):395-7. Epub 2004/04/22.
49. Han ET, Lee DH, Park KD, Seok WS, Kim YS, Tsuboi T, et al. Reemerging vivax
malaria: changing patterns of annual incidence and control programs in the Republic of
Korea. Korean J Parasitol. 2006;44(4):285-94. Epub 2006/12/16.
50. Lee KS, Kim TH, Kim ES, Lim HS, Yeom JS, Jun G, et al. Short report: chloroquine-
resistant Plasmodium vivax in the Republic of Korea. Am J Trop Med Hyg. 2009;80(2):215-7.
Epub 2009/02/05.
51. Arias AE, Corredor A. Low response of Colombian strains of Plasmodium vivax to
classical antimalarial therapy. Trop Med Parasitol. 1989;40(1):21-3. Epub 1989/03/01.
52. Garavelli PL, Corti E. Chloroquine resistance in Plasmodium vivax: the first case in
Brazil. Trans R Soc Trop Med Hyg. 1992;86(2):128. Epub 1992/03/01.
53. Loyola EG, Rodriguez MH. Chloroquine-resistant Plasmodium vivax in Brazil. Trans
R Soc Trop Med Hyg. 1992;86(5):570; author reply 1. Epub 1992/09/01.
54. Alecrim MdG, Alecrim W, Macedo V. Plasmodium vivax resistance to chloroquine
(R2) and mefloquine (R3) in Brazilian Amazon region. Rev Soc Bras Med Trop.
1999;32(1):67-8. Epub 1999/02/03.
55. de Santana Filho FS, Arcanjo AR, Chehuan YM, Costa MR, Martinez-Espinosa FE,
Vieira JL, et al. Chloroquine-resistant Plasmodium vivax, Brazilian Amazon. Emerg Infect
Dis. 2007;13(7):1125-6. Epub 2008/01/25.
56. Marques MM, Costa MR, Santana Filho FS, Vieira JL, Nascimento MT, Brasil LW, et
al. Plasmodium vivax chloroquine resistance and anemia in the western Brazilian Amazon.
Antimicrob Agents Chemother. 2014;58(1):342-7. Epub 2013/10/30.
57. Goncalves LA, Cravo P, Ferreira MU. Emerging Plasmodium vivax resistance to
chloroquine in South America: an overview. Mem Inst Oswaldo Cruz. 2014;0. Epub
2014/05/03.

89
58. Suwanarusk R, Russell B, Chavchich M, Chalfein F, Kenangalem E, Kosaisavee V, et
al. Chloroquine resistant Plasmodium vivax: in vitro characterisation and association with
molecular polymorphisms. PLoS One. 2007;2(10):e1089. Epub 2007/11/01.
59. Russell B, Chalfein F, Prasetyorini B, Kenangalem E, Piera K, Suwanarusk R, et al.
Determinants of in vitro drug susceptibility testing of Plasmodium vivax. Antimicrob Agents
Chemother. 2008;52(3):1040-5. Epub 2008/01/09.
60. Orjuela-Sanchez P, de Santana Filho FS, Machado-Lima A, Chehuan YF, Costa MR,
Alecrim MD, et al. Analysis of single-nucleotide polymorphisms in the crt-o and mdr1 genes
of Plasmodium vivax among chloroquine resistant isolates from the Brazilian Amazon region.
Antimicrob Agents Chemother. 2009.
61. Howard N, Durrani N, Sanda S, Beshir K, Hallett R, Rowland M. Clinical trial of
extended-dose chloroquine for treatment of resistant falciparum malaria among Afghan
refugees in Pakistan. Malar J. 2011;10:171. Epub 2011/06/28.
62. Bin Dajem SM, Al-Sheikh AA, Bohol MF, Alhawi M, Al-Ahdal MN, Al-Qahtani A.
Detecting mutations in PfCRT and PfMDR1 genes among Plasmodium falciparum isolates
from Saudi Arabia by pyrosequencing. Parasitol Res. 2011;109(2):291-6. Epub 2011/02/26.
63. Chinappi M, Via A, Marcatili P, Tramontano A. On the mechanism of chloroquine
resistance in Plasmodium falciparum. PLoS One. 2010;5(11):e14064. Epub 2010/12/03.
64. Ngassa Mbenda HG, Das A. Occurrence of multiple chloroquine-resistant Pfcrt
haplotypes and emergence of the S(agt)VMNT type in Cameroonian Plasmodium falciparum.
The Journal of antimicrobial chemotherapy. 2013. Epub 2013/10/05.
65. Alvarez G, Pineros JG, Tobon A, Rios A, Maestre A, Blair S, et al. Efficacy of three
chloroquine-primaquine regimens for treatment of Plasmodium vivax malaria in Colombia.
Am J Trop Med Hyg. 2006;75(4):605-9.
66. Krudsood S, Tangpukdee N, Wilairatana P, Phophak N, Baird JK, Brittenham GM, et
al. High-dose primaquine regimens against relapse of Plasmodium vivax malaria. Am J Trop
Med Hyg. 2008;78(5):736-40.
67. van Es HH, Karcz S, Chu F, Cowman AF, Vidal S, Gros P, et al. Expression of the
plasmodial pfmdr1 gene in mammalian cells is associated with increased susceptibility to
chloroquine. Mol Cell Biol. 1994;14(4):2419-28. Epub 1994/04/01.
68. van Es HH, Renkema H, Aerts H, Schurr E. Enhanced lysosomal acidification leads to
increased chloroquine accumulation in CHO cells expressing the pfmdr1 gene. Mol Biochem
Parasitol. 1994;68(2):209-19. Epub 1994/12/01.
69. Volkman SK, Cowman AF, Wirth DF. Functional complementation of the ste6 gene
of Saccharomyces cerevisiae with the pfmdr1 gene of Plasmodium falciparum. Proc Natl
Acad Sci U S A. 1995;92(19):8921-5. Epub 1995/09/12.
70. Peel SA. The ABC transporter genes of Plasmodium falciparum and drug resistance.
Drug Resist Updat. 2001;4(1):66-74. Epub 2001/08/22.
71. Duah NO, Matrevi SA, de Souza DK, Binnah DD, Tamakloe MM, Opoku VS, et al.
Increased pfmdr1 gene copy number and the decline in pfcrt and pfmdr1 resistance alleles in
Ghanaian Plasmodium falciparum isolates after the change of anti-malarial drug treatment
policy. Malar J. 2013;12:377. Epub 2013/11/01.

90
72. Price RN, Uhlemann AC, Brockman A, McGready R, Ashley E, Phaipun L, et al.
Mefloquine resistance in Plasmodium falciparum and increased pfmdr1 gene copy number.
Lancet. 2004;364(9432):438-47. Epub 2004/08/04.
73. Nomura T, Carlton JM, Baird JK, del Portillo HA, Fryauff DJ, Rathore D, et al.
Evidence for different mechanisms of chloroquine resistance in 2 Plasmodium species that
cause human malaria. J Infect Dis. 2001;183(11):1653-61. Epub 2001/05/09.
74. Bray PG, Martin RE, Tilley L, Ward SA, Kirk K, Fidock DA. Defining the role of
PfCRT in Plasmodium falciparum chloroquine resistance. Mol Microbiol. 2005;56(2):323-33.
Epub 2005/04/09.
75. Barnadas C, Ratsimbasoa A, Tichit M, Bouchier C, Jahevitra M, Picot S, et al.
Plasmodium vivax resistance to chloroquine in Madagascar: clinical efficacy and
polymorphisms in pvmdr1 and pvcrt-o genes. Antimicrob Agents Chemother.
2008;52(12):4233-40. Epub 2008/09/24.
76. Sa JM, Yamamoto MM, Fernandez-Becerra C, de Azevedo MF, Papakrivos J, Naude
B, et al. Expression and function of pvcrt-o, a Plasmodium vivax ortholog of pfcrt, in
Plasmodium falciparum and Dictyostelium discoideum. Mol Biochem Parasitol.
2006;150(2):219-28. Epub 2006/09/22.
77. Fernandez-Becerra C, Pinazo MJ, Gonzalez A, Alonso PL, del Portillo HA, Gascon J.
Increased expression levels of the pvcrt-o and pvmdr1 genes in a patient with severe
Plasmodium vivax malaria. Malar J. 2009;8:55. Epub 2009/04/04.
78. Brega S, Meslin B, de Monbrison F, Severini C, Gradoni L, Udomsangpetch R, et al.
Identification of the Plasmodium vivax mdr-like gene (pvmdr1) and analysis of single-
nucleotide polymorphisms among isolates from different areas of endemicity. J Infect Dis.
2005;191(2):272-7. Epub 2004/12/21.
79. Sa JM, Nomura T, Neves J, Baird JK, Wellems TE, del Portillo HA. Plasmodium
vivax: allele variants of the mdr1 gene do not associate with chloroquine resistance among
isolates from Brazil, Papua, and monkey-adapted strains. Exp Parasitol. 2005;109(4):256-9.
Epub 2005/03/10.
80. Marfurt J, de Monbrison F, Brega S, Barbollat L, Muller I, Sie A, et al. Molecular
markers of in vivo Plasmodium vivax resistance to amodiaquine plus sulfadoxine-
pyrimethamine: mutations in pvdhfr and pvmdr1. J Infect Dis. 2008;198(3):409-17. Epub
2008/06/28.
81. Imwong M, Pukrittayakamee S, Pongtavornpinyo W, Nakeesathit S, Nair S, Newton
P, et al. Gene amplification of the multidrug resistance 1 gene of Plasmodium vivax isolates
from Thailand, Laos, and Myanmar. Antimicrob Agents Chemother. 2008;52(7):2657-9.
Epub 2008/04/30.
82. Gama BE, Oliveira NK, Souza JM, Daniel-Ribeiro CT, Ferreira-da-Cruz Mde F.
Characterisation of pvmdr1 and pvdhfr genes associated with chemoresistance in Brazilian
Plasmodium vivax isolates. Mem Inst Oswaldo Cruz. 2009;104(7):1009-11. Epub 2009/12/23.
83. Chehuan YF, Costa MR, Costa JS, Alecrim MG, Nogueira F, Silveira H, et al. In vitro
chloroquine resistance for Plasmodium vivax isolates from the Western Brazilian Amazon.
Malar J. 2013;12:226. Epub 2013/07/04.
84. Vargas-Rodriguez RD, da Silva Bastos M, Menezes MJ, Orjuela-Sanchez P, Ferreira
MU. Single-Nucleotide Polymorphism and Copy Number Variation of the Multidrug

91
Resistance-1 Locus of Plasmodium vivax: Local and Global Patterns. Am J Trop Med Hyg.
2012. Epub 2012/09/06.
85. Picot S, Brega S, Gerome P, Velut G, de Monbrison F, Cheminel V, et al. Absence of
nucleotide polymorphism in a Plasmodium vivax multidrug resistance gene after failure of
mefloquine prophylaxis in French Guyana. Trans R Soc Trop Med Hyg. 2005;99(3):234-7.
Epub 2005/01/18.
86. Suwanarusk R, Chavchich M, Russell B, Jaidee A, Chalfein F, Barends M, et al.
Amplification of pvmdr1 associated with multidrug-resistant Plasmodium vivax. J Infect Dis.
2008;198(10):1558-64. Epub 2008/09/24.
87. Dharia NV, Bright AT, Westenberger SJ, Barnes SW, Batalov S, Kuhen K, et al.
Whole-genome sequencing and microarray analysis of ex vivo Plasmodium vivax reveal
selective pressure on putative drug resistance genes. Proc Natl Acad Sci U S A.
2010;107(46):20045-50. Epub 2010/11/03.
88. Lu F, Lim CS, Nam DH, Kim K, Lin K, Kim TS, et al. Genetic polymorphism in
pvmdr1 and pvcrt-o genes in relation to in vitro drug susceptibility of Plasmodium vivax
isolates from malaria-endemic countries. Acta Trop. 2011;117(2):69-75. Epub 2010/10/12.
89. Jovel IT, Mejia RE, Banegas E, Piedade R, Alger J, Fontecha G, et al. Drug resistance
associated genetic polymorphisms in Plasmodium falciparum and Plasmodium vivax
collected in Honduras, Central America. Malar J. 2011;10:376. Epub 2011/12/21.
90. Rijken MJ, Boel ME, Russell B, Imwong M, Leimanis ML, Phyo AP, et al.
Chloroquine resistant vivax malaria in a pregnant woman on the western border of Thailand.
Malar J. 2011;10:113. Epub 2011/05/07.
91. Ranjitkar S, Schousboe ML, Thomsen TT, Adhikari M, Kapel CM, Bygbjerg IC, et al.
Prevalence of molecular markers of anti-malarial drug resistance in Plasmodium vivax and
Plasmodium falciparum in two districts of Nepal. Malar J. 2011;10:75. Epub 2011/04/05.
92. Lu F, Wang B, Cao J, Sattabongkot J, Zhou H, Zhu G, et al. Prevalence of drug
resistance-associated gene mutations in Plasmodium vivax in Central China. Korean J
Parasitol. 2012;50(4):379-84. Epub 2012/12/12.
93. Lin JT, Patel JC, Kharabora O, Sattabongkot J, Muth S, Ubalee R, et al. Plasmodium
vivax isolates from Cambodia and Thailand show high genetic complexity and distinct
patterns of P. vivax multidrug resistance gene 1 (pvmdr1) polymorphisms. Am J Trop Med
Hyg. 2013;88(6):1116-23. Epub 2013/03/20.
94. Khattak AA, Venkatesan M, Khatoon L, Ouattara A, Kenefic LJ, Nadeem MF, et al.
Prevalence and patterns of antifolate and chloroquine drug resistance markers in Plasmodium
vivax across Pakistan. Malar J. 2013;12:310. Epub 2013/09/07.
95. Ganguly S, Saha P, Guha SK, Das S, Bera DK, Biswas A, et al. In vivo therapeutic
efficacy of chloroquine alone or in combination with primaquine against vivax malaria in
Kolkata, West Bengal, India, and polymorphism in pvmdr1 and pvcrt-o genes. Antimicrob
Agents Chemother. 2013;57(3):1246-51. Epub 2012/12/25.
96. Aguiar AC, Pereira DB, Amaral NS, De Marco L, Krettli AU. Plasmodium vivax and
Plasmodium falciparum ex vivo susceptibility to anti-malarials and gene characterization in
Rondonia, West Amazon, Brazil. Malar J. 2014;13:73. Epub 2014/03/04.

92
97. Kebede S, Medhin G, Berhe N, Teklehaymanot T, Gebru T, Clause R, et al. Return of
chloroquine-sensitive Plasmodium falciparum parasites and emergence of chloroquine-
resistant Plasmodium vivax in Ethiopia. Malar J. 2014;13(1):244. Epub 2014/06/27.
98. Shalini S, Chaudhuri S, Sutton PL, Mishra N, Srivastava N, David JK, et al.
Chloroquine efficacy studies confirm drug susceptibility of Plasmodium vivax in Chennai,
India. Malar J. 2014;13:129. Epub 2014/04/02.
99. Westenberger SJ, McClean CM, Chattopadhyay R, Dharia NV, Carlton JM, Barnwell
JW, et al. A systems-based analysis of Plasmodium vivax lifecycle transcription from human
to mosquito. PLoS Negl Trop Dis. 2010;4(4):e653. Epub 2010/04/14.
100. Tjitra E, Anstey NM, Sugiarto P, Warikar N, Kenangalem E, Karyana M, et al.
Multidrug-resistant Plasmodium vivax associated with severe and fatal malaria: a prospective
study in Papua, Indonesia. PLoS Med. 2008;5(6):e128. Epub 2008/06/20.
101. Zubairi AB, Nizami S, Raza A, Mehraj V, Rasheed AF, Ghanchi NK, et al. Severe
Plasmodium vivax malaria in Pakistan. Emerg Infect Dis. 2013;19(11):1851-4. Epub
2013/11/06.
102. Alexandre MA, Ferreira CO, Siqueira AM, Magalhaes BL, Mourao MP, Lacerda MV,
et al. Severe Plasmodium vivax malaria, Brazilian Amazon. Emerg Infect Dis.
2010;16(10):1611-4. Epub 2010/09/30.
103. Lacerda MV, Fragoso SC, Alecrim MG, Alexandre MA, Magalhaes BM, Siqueira
AM, et al. Postmortem Characterization of Patients With Clinical Diagnosis of Plasmodium
vivax Malaria: To What Extent Does This Parasite Kill? Clin Infect Dis. 2012;55(8):e67-74.
Epub 2012/07/10.
104. Kochar DK, Das A, Kochar SK, Saxena V, Sirohi P, Garg S, et al. Severe Plasmodium
vivax malaria: a report on serial cases from Bikaner in northwestern India. Am J Trop Med
Hyg. 2009;80(2):194-8. Epub 2009/02/05.
105. Nandwani S, Pande A, Saluja M. Clinical profile of severe malaria: study from a
tertiary care center in north India. Journal of parasitic diseases : official organ of the Indian
Society for Parasitology. 2014;38(1):11-5. Epub 2014/02/08.
106. Kochar DK, Saxena V, Singh N, Kochar SK, Kumar SV, Das A. Plasmodium vivax
malaria. Emerg Infect Dis. 2005;11(1):132-4. Epub 2005/02/12.
107. Genton B, D'Acremont V, Rare L, Baea K, Reeder JC, Alpers MP, et al. Plasmodium
vivax and mixed infections are associated with severe malaria in children: a prospective
cohort study from Papua New Guinea. PLoS Med. 2008;5(6):e127. Epub 2008/06/20.
108. Lacerda MV, Mourao MP, Alexandre MA, Siqueira AM, Magalhaes BM, Martinez-
Espinosa FE, et al. Understanding the clinical spectrum of complicated Plasmodium vivax
malaria: a systematic review on the contributions of the Brazilian literature. Malar J.
2012;11:12. Epub 2012/01/11.
109. Prokopenko LI. On epidemiology of fulminant P. vivax malaria. In: Beklemishev VN,
Rashina MG, editors. Transactions of the Moscow Martinovski Institute of Medical
Parasitology and Tropical Medicine. Moscow, Russia1959. p. 80-102.
110. Levin EB. P. vivax malaria; a case with anemia, cardiomegaly, hepatomegaly and
renal involvement. Calif Med. 1954;81(2):87-9. Epub 1954/08/01.
111. Lakhkar BB, Babu S, Shenoy V. DIC in vivax malaria. Indian Pediatr.
1996;33(11):971-2. Epub 1996/11/01.

93
112. Barcus MJ, Basri H, Picarima H, Manyakori C, Sekartuti, Elyazar I, et al.
Demographic risk factors for severe and fatal vivax and falciparum malaria among hospital
admissions in northeastern Indonesian Papua. Am J Trop Med Hyg. 2007;77(5):984-91. Epub
2007/11/07.
113. Nayak KC, Kumar S, Gupta BK, Gupta A, Prakash P, Kochar DK. Clinical and
histopathological profile of acute renal failure caused by falciparum and vivax monoinfection:
an observational study from Bikaner, northwest zone of Rajasthan, India. Journal of vector
borne diseases. 2014;51(1):40-6. Epub 2014/04/11.
114. Lacerda MVG, Oliveira SL, Alecrim MGC. Splenic hematoma in a patient with
Plasmodium vivax malaria. Rev Soc Bras Med Trop. 2007;40(1):96-7.
115. Lacerda MV, Alexandre MA, Santos PD, Arcanjo AR, Alecrim WD, Alecrim MGC.
Idiopathic thrombocytopenic purpura due to vivax malaria in the Brazilian Amazon. Acta
Trop. 2004;90(2):187-90.
116. Lacerda MV, Hipolito JR, Passos LN. Chronic Plasmodium vivax infection in a
patient with splenomegaly and severe thrombocytopenia. Rev Soc Bras Med Trop.
2008;41(5):522-3. Epub 2008/11/15.
117. Santos-Ciminera PD, Roberts DR, Alecrim MG, Costa MR, Quinnan GV, Jr. Malaria
diagnosis and hospitalization trends, Brazil. Emerg Infect Dis. 2007;13(10):1597-600. Epub
2008/02/09.
118. Alexandre MAA. Estudo clínico e epidemiológico dos casos graves de malária vivax
em pacientes atendidos na Fundação de Medicina Tropical do Amazonas, Brasil [dissertação].
Manaus: Universidade do Estado do Amazonas; 2004.
119. Alecrim MGC. Estudo clínico, resistência e polimorfismo parasitário na malária pelo
Plasmodium vivax, em Manaus (AM) [tese]. Brasília: Universidade de Brasília; 2000.
120. de Lacerda MV, de Oliveira SL, Alecrim MG. Splenic hematoma in a patient with
Plasmodium vivax malaria. Rev Soc Bras Med Trop. 2007;40(1):96-7. Epub 2007/05/09.
121. Batista D. The paludisme in the Amazon: a contribution to epidemiology,protozoology
and clinics, study about the billiary-hemoglobinuric fever. Rio de Janeiro: Imprensa Nacional;
1946.
122. Alecrim MGC. Estudo clínico, resistência e polimorfismo parasitário na malária pelo
Plasmodium vivax. [Tese de Doutorado]. Brasília: Universidade de Brasília; 2000.
123. Lacerda MV, Alexandre MA, Santos PD, Arcanjo AR, Alecrim WD, Alecrim MG.
Idiopathic thrombocytopenic purpura due to vivax malaria in the Brazilian Amazon. Acta
Trop. 2004;90(2):187-90. Epub 2004/06/05.
124. Silamut K, Phu NH, Whitty C, Turner GD, Louwrier K, Mai NT, et al. A quantitative
analysis of the microvascular sequestration of malaria parasites in the human brain. Am J
Pathol. 1999;155(2):395-410. Epub 1999/08/06.
125. Rowe JA, Moulds JM, Newbold CI, Miller LH. P. falciparum rosetting mediated by a
parasite-variant erythrocyte membrane protein and complement-receptor 1. Nature.
1997;388(6639):292-5. Epub 1997/07/17.
126. Carvalho BO, Lopes SC, Nogueira PA, Orlandi PP, Bargieri DY, Blanco YC, et al. On
the cytoadhesion of Plasmodium vivax-infected erythrocytes. J Infect Dis. 2010;202(4):638-
47. Epub 2010/07/14.

94
127. De las Salas B, Segura C, Pabon A, Lopes SC, Costa FT, Blair S. Adherence to human
lung microvascular endothelial cells (HMVEC-L) of Plasmodium vivax isolates from
Colombia. Malar J. 2013;12:347. Epub 2013/10/02.
128. Anstey NM, Handojo T, Pain MC, Kenangalem E, Tjitra E, Price RN, et al. Lung
injury in vivax malaria: pathophysiological evidence for pulmonary vascular sequestration
and posttreatment alveolar-capillary inflammation. J Infect Dis. 2007;195(4):589-96. Epub
2007/01/19.
129. Chotivanich K, Udomsangpetch R, Suwanarusk R, Pukrittayakamee S, Wilairatana P,
Beeson JG, et al. Plasmodium vivax adherence to placental glycosaminoglycans. PLoS One.
2012;7(4):e34509. Epub 2012/04/25.
130. Bernabeu M, Lopez FJ, Ferrer M, Martin-Jaular L, Razaname A, Corradin G, et al.
Functional analysis of Plasmodium vivax VIR proteins reveals different subcellular
localizations and cytoadherence to the ICAM-1 endothelial receptor. Cellular microbiology.
2012;14(3):386-400. Epub 2011/11/23.
131. Chotivanich KT, Pukrittayakamee S, Simpson JA, White NJ, Udomsangpetch R.
Characteristics of Plasmodium vivax-infected erythrocyte rosettes. Am J Trop Med Hyg.
1998;59(1):73-6. Epub 1998/07/31.
132. Russell B, Suwanarusk R, Borlon C, Costa FT, Chu CS, Rijken MJ, et al. A reliable ex
vivo invasion assay of human reticulocytes by Plasmodium vivax. Blood. 2011;118(13):e74-
81. Epub 2011/07/20.
133. WHO. Management of severe malaria: a practical handbook Geneva 2012.
134. van der Heyde HC, Nolan J, Combes V, Gramaglia I, Grau GE. A unified hypothesis
for the genesis of cerebral malaria: sequestration, inflammation and hemostasis leading to
microcirculatory dysfunction. Trends Parasitol. 2006;22(11):503-8. Epub 2006/09/19.
135. Baird JK. Neglect of Plasmodium vivax malaria. Trends Parasitol. 2007;23(11):533-9.
Epub 2007/10/16.
136. Price RN, Douglas NM, Anstey NM. New developments in Plasmodium vivax
malaria: severe disease and the rise of chloroquine resistance. Curr Opin Infect Dis.
2009;22(5):430-5. Epub 2009/07/03.
137. Menendez C, Fleming AF, Alonso PL. Malaria-related anaemia. Parasitol Today.
2000;16(11):469-76. Epub 2000/11/07.
138. Orjuela-Sanchez P, de Santana Filho FS, Machado-Lima A, Chehuan YF, Costa MR,
Alecrim MG, et al. Analysis of single-nucleotide polymorphisms in the crt-o and mdr1 genes
of Plasmodium vivax among chloroquine-resistant isolates from the Brazilian Amazon region.
Antimicrob Agents Chemother. 2009;53(8):3561-4. Epub 2009/05/20.
139. Trape JF. The public health impact of chloroquine resistance in Africa. Am J Trop
Med Hyg. 2001;64(1-2 Suppl):12-7. Epub 2001/06/27.
140. Meerman L, Ord R, Bousema JT, van Niekerk M, Osman E, Hallett R, et al. Carriage
of chloroquine-resistant parasites and delay of effective treatment increase the risk of severe
malaria in Gambian children. J Infect Dis. 2005;192(9):1651-7. Epub 2005/10/06.
141. Unidades da Federação: Amazonas [database on the Internet]. 2013 [cited
09/07/2014].

95
142. Soto J, Toledo J, Gutierrez P, Luzz M, Llinas N, Cedeno N, et al. Plasmodium vivax
clinically resistant to chloroquine in Colombia. Am J Trop Med Hyg. 2001;65(2):90-3. Epub
2001/08/18.
143. Ruebush TK, 2nd, Zegarra J, Cairo J, Andersen EM, Green M, Pillai DR, et al.
Chloroquine-resistant Plasmodium vivax malaria in Peru. Am J Trop Med Hyg.
2003;69(5):548-52. Epub 2003/12/26.
144. Naing C, Aung K, Win DK, Wah MJ. Efficacy and safety of chloroquine for treatment
in patients with uncomplicated Plasmodium vivax infections in endemic countries. Trans R
Soc Trop Med Hyg. 2010;104(11):695-705. Epub 2010/09/21.
145. WHO. Malaria Report: 2010. 2010.
146. MS. Manual de Diagnóstico Laboratorial da Malária. In: Epidemiológica DdV, editor.
Brasilia2009.
147. Perandin F, Manca N, Calderaro A, Piccolo G, Galati L, Ricci L, et al. Development
of a real-time PCR assay for detection of Plasmodium falciparum, Plasmodium vivax, and
Plasmodium ovale for routine clinical diagnosis. J Clin Microbiol. 2004;42(3):1214-9. Epub
2004/03/09.
148. Hummon AB, Lim SR, Difilippantonio MJ, Ried T. Isolation and solubilization of
proteins after TRIzol extraction of RNA and DNA from patient material following prolonged
storage. Biotechniques. 2007;42(4):467-70, 72. Epub 2007/05/11.
149. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25(4):402-8. Epub
2002/02/16.
150. Imwong M, Pukrittakayamee S, Looareesuwan S, Pasvol G, Poirreiz J, White NJ, et
al. Association of genetic mutations in Plasmodium vivax dhfr with resistance to sulfadoxine-
pyrimethamine: geographical and clinical correlates. Antimicrob Agents Chemother.
2001;45(11):3122-7. Epub 2001/10/16.
151. Marin-Menendez A, Bardaji A, Martinez-Espinosa FE, Botto-Menezes C, Lacerda
MV, Ortiz J, et al. Rosetting in Plasmodium vivax: a cytoadhesion phenotype associated with
anaemia. PLoS Negl Trop Dis. 2013;7(4):e2155. Epub 2013/04/18.
152. Chenet SM, Schneider KA, Villegas L, Escalante AA. Local population structure of
Plasmodium: impact on malaria control and elimination. Malar J. 2012;11:412. Epub
2012/12/13.
153. de Araujo FC, de Rezende AM, Fontes CJ, Carvalho LH, Alves de Brito CF. Multiple-
clone activation of hypnozoites is the leading cause of relapse in Plasmodium vivax infection.
PLoS One. 2012;7(11):e49871. Epub 2012/11/28.
154. Koepfli C, Mueller I, Marfurt J, Goroti M, Sie A, Oa O, et al. Evaluation of
Plasmodium vivax genotyping markers for molecular monitoring in clinical trials. J Infect Dis.
2009;199(7):1074-80. Epub 2009/03/12.
155. WHO. Methods and techniques for clinical trials on antimalarial drug efficacy :
genotyping to identify parasite populations. . 2008.
156. Barnadas C, Koepfli C, Karunajeewa HA, Siba PM, Davis TM, Mueller I.
Characterization of treatment failure in efficacy trials of drugs against Plasmodium vivax by
genotyping neutral and drug resistance-associated markers. Antimicrob Agents Chemother.
2011;55(9):4479-81. Epub 2011/06/29.

96
157. Baird JK. Resistance to chloroquine unhinges vivax malaria therapeutics. Antimicrob
Agents Chemother. 2011;55(5):1827-30. Epub 2011/03/09.
158. Lanca EF, Magalhaes BM, Vitor-Silva S, Siqueira AM, Benzecry SG, Alexandre MA,
et al. Risk factors and characterization of Plasmodium vivax-associated admissions to
pediatric intensive care units in the Brazilian Amazon. PLoS One. 2012;7(4):e35406. Epub
2012/04/24.
159. Baird JK, Basri H, Purnomo, Bangs MJ, Subianto B, Patchen LC, et al. Resistance to
chloroquine by Plasmodium vivax in Irian Jaya, Indonesia. Am J Trop Med Hyg.
1991;44(5):547-52. Epub 1991/05/01.
160. Das S, Chakraborty SP, Hati AK, Roy S. Association between prevalence of
chloroquine resistance and unusual mutation in pfmdr-I and pfcrt genes in India. Am J Trop
Med Hyg. 2013;88(5):828-34. Epub 2013/03/20.
161. Ngo T, Duraisingh M, Reed M, Hipgrave D, Biggs B, Cowman AF. Analysis of pfcrt,
pfmdr1, dhfr, and dhps mutations and drug sensitivities in Plasmodium falciparum isolates
from patients in Vietnam before and after treatment with artemisinin. Am J Trop Med Hyg.
2003;68(3):350-6. Epub 2003/04/11.
162. Snounou G, Viriyakosol S, Zhu XP, Jarra W, Pinheiro L, do Rosario VE, et al. High
sensitivity of detection of human malaria parasites by the use of nested polymerase chain
reaction. Mol Biochem Parasitol. 1993;61(2):315-20. Epub 1993/10/01.
163. Gama BE, Lacerda MV, Daniel-Ribeiro CT, Ferreira-da-Cruz Mde F.
Chemoresistance of Plasmodium falciparum and Plasmodium vivax parasites in Brazil:
consequences on disease morbidity and control. Mem Inst Oswaldo Cruz. 2011;106 Suppl
1:159-66. Epub 2011/09/09.
164. Marfurt J, Mueller I, Sie A, Maku P, Goroti M, Reeder JC, et al. Low efficacy of
amodiaquine or chloroquine plus sulfadoxine-pyrimethamine against Plasmodium falciparum
and P. vivax malaria in Papua New Guinea. Am J Trop Med Hyg. 2007;77(5):947-54. Epub
2007/11/07.
165. Reilly HB, Wang H, Steuter JA, Marx AM, Ferdig MT. Quantitative dissection of
clone-specific growth rates in cultured malaria parasites. Int J Parasitol. 2007;37(14):1599-
607. Epub 2007/06/26.
166. Chotivanich K, Udomsangpetch R, Simpson JA, Newton P, Pukrittayakamee S,
Looareesuwan S, et al. Parasite multiplication potential and the severity of Falciparum
malaria. J Infect Dis. 2000;181(3):1206-9. Epub 2000/03/18.
167. Schwartz IK, Lackritz EM, Patchen LC. Chloroquine-resistant Plasmodium vivax from
Indonesia. N Engl J Med. 1991;324(13):927. Epub 1991/03/28.
168. Fidock DA, Nomura T, Talley AK, Cooper RA, Dzekunov SM, Ferdig MT, et al.
Mutations in the P. falciparum digestive vacuole transmembrane protein PfCRT and evidence
for their role in chloroquine resistance. Mol Cell. 2000;6(4):861-71. Epub 2000/11/25.
169. Akintowa A, Meyer MC, Hwang PTR. Simultaneous determination of chloroquine
and desethylchloroquine in blood, plasma, and urine by high-performance liquid
chromatography. J Liq Chromatogr. 1983;6:1513–22.
170. Rombo L, Ericsson O, Alvan G, Lindstrom B, Gustafsson LL, Sjoqvist F. Chloroquine
and desethylchloroquine in plasma, serum, and whole blood: problems in assay and handling
of samples. Therapeutic drug monitoring. 1985;7(2):211-5. Epub 1985/01/01.

97
171. Rombo L, Bergqvist Y, Hellgren U. Chloroquine and desethylchloroquine
concentrations during regular long-term malaria prophylaxis. Bull World Health Organ.
1987;65(6):879-83. Epub 1987/01/01.
172. Sharrock WW, Suwanarusk R, Lek-Uthai U, Edstein MD, Kosaisavee V, Travers T, et
al. Plasmodium vivax trophozoites insensitive to chloroquine. Malar J. 2008;7:94. Epub
2008/05/29.
173. Baird JK, Basri H, Subianto B, Fryauff DJ, McElroy PD, Leksana B, et al. Treatment
of chloroquine-resistant Plasmodium vivax with chloroquine and primaquine or halofantrine.
J Infect Dis. 1995;171(6):1678-82. Epub 1995/06/01.

98
8 ANEXOS

99
8.1 Anexo 1
PROCEDIMENTOS OPERACIONAIS PADRÃO (POP)

100
PROCEDIMENTOS OPERACIONAIS PADRÃO (POP)
Pop de preparo e leitura de lâminas para detecção de Plasmodium spp
Código POP POP_MAL_LB_001_v02D_PT
Título Preparo e leitura de lâminas para detecção de Plasmodium spp.
Idioma da versão original PORTUGUÊS
Elaborado por:
Gisely Cardoso de Melo
Revisado por:
Aprovado por:
Wuelton Marcelo
Monteiro
Data de aplicação:
05 de julho de 2010
Data da próxima revisão:
Emenda Razão da emenda
.
OBJETIVOS
Descrever os procedimentos para preparo, coloração, leitura e registro de resultado de lâminas
para detecção de Plasmodium spp. utilizando gota espessa.
DEFINIÇÕES
Não se aplica
APLICÁVEL A
Todas as lâminas coletadas para detecção de Plasmodium spp. durante os estudo.
RESPONSABILIDADES
Pessoal encarregado da coleta das amostras, técnicos e microscopistas, chefia do laboratório.
Código: POP_MAL_LB_002_v01D_PT
Procedimento Operacional Padrão
Gerência de Malária
S

101
POPS RELACIONADOS
Seguimento dos participantes no estudo “Epidemiologia da Malária no Município do Careiro,
Amazonas” (versão atual do POP_MAL_TC_002).
Triagem e seguimento dos pacientes no estudo de malária grave por Plasmodium vivax
(versão atual do POP_MAL_HO_001).
PROCEDIMENTOS
6.1 Coleta e identificação das amostras
1 Dependendo do protocolo do estudo, em cada local (campo, unidade básica de saúde ou
hospital) serão coletadas:
- duas gotas espessas por indivíduo ou;
- quatro lâminas por pessoa - duas com duas gotas espessas na mesma lâmina e duas com
esfregaço.
2. Após limpar a polpa do dedo anelar com algodão embebido em álcool a 70%. Fazer a
punção digital com lanceta estéril. Comprimir o dedo suavemente.
3. Duas gotas espessas serão preparadas na mesma lâmina, alternativamente a uma gota
espessa e um esfregaço. Confeccionar uma gota espessa homogênea de 1,5 x 1,0 cm na
lâmina de vidro.
4. Uma etiqueta de identificação, contendo código de barras e números únicos, será colada na
margem da lâmina, como segue:

102
5. Os três formulários seguintes serão preenchidos no momento da coleta da lâmina no
hospital, na unidade básica de saúde ou no campo:
- Uma Ficha de amostras de laboratório;
- Cada estudo tem uma diferente ficha de amostras de laboratório, onde os tipos de amostras
coletadas de cada estudo são especificados;
- Uma etiqueta de identificação de amostra será pregada e o nome o número de identificação
no estudo de cada paciente será preenchido;
- Um formulário de resultado da primeira leitura de lâmina (versão atual do
POP_MAL_LB_001_A01) e de resultado da segunda leitura de lâmina (versão atual do
POP_MAL_LB_001_A02);
- Uma etiqueta de identificação será colada em cada formulário, que também será preenchido
com nome e número de identificação permanente (NIP) do paciente no estudo, além da data
de coleta da lâmina.
6.2 Recepção de amostras no laboratório
1. As lâminas serão levadas inicialmente para o laboratório onde serão devidamente coradas
(e será realizada a leitura rápida.

103
2. Para leitura de densidade parasitária, as lâminas serão enviadas ao laboratório da Gerência
de Malária acompanhadas de uma Ficha de amostras de laboratório para cada paciente,
acompanhada de:
- formulário de resultado da primeira leitura de lâmina (versão atual do
POP_MAL_LB_001_A01);
- formulário de resultado da segunda leitura de lâmina (versão atual do
POP_MAL_LB_001_A02).
3. Quando as amostras chegarem ao laboratório, a pessoa encarregada da recepção checará se
todas as amostras registradas na ficha de amostras de laboratório foram entregues.
4. A Ficha de amostras de laboratório será preenchida no local da coleta em ordem de data.
Cada estudo tem uma diferente
5. Após a recepção, caso não sejam realizada a leitura da densidade parasitária no mesmo
momento da chegada, estas serão armazenadas
6. Quando as amostras chegarem ao laboratório, a pessoa encarregada da recepção checará se
todas as amostras registradas na Ficha de amostras de laboratório foram entregues.
7. A Ficha de amostras de laboratório será preenchida no local da coleta em ordem de data.
8. Após a recepção, caso não sejam realizada a leitura da densidade parasitária no mesmo
momento da chegada, estas serão armazenadas conforme procedimento descrito na seção 8.
6.3 Coloração
6.3.1 Reagentes
Azul de metileno fosfatado
Solução alcoólica de Giemsa estoque

104
Solução alcoólica de Giemsa diluída 1:10
Água tamponada
6.3.1.1 Preparação
Azul de metileno fosfatado
Azul de metileno medicinal em pó 16H18ClN3S
200 mg
Fosfato de sódio monobásico NaH2PO4
600 mg
Fosfato de potássio bibásico K2HPO4
200 mg
Água destilada H2O
250 mL
Filtrar para retirar as impurezas
Solução alcoólica de Giemsa estoque
Corante Giemsa em pó C14H14ClN3S
750 mg
Álcool metílico PA CH3OH
65 mL

105
Glicerina PA CH2(OH)CH(OH)CH2OH
35 mL
Agitar bem (várias vezes por dia) em garrafa contendo pérolas de vidro.
Manter o recipiente tampado em forma de estoque.
Filtrar quando necessário.
Solução alcoólica de Giemsa diluída 1:10
Solução alcoólica de Giemsa estoque 1 gota
Água tamponada 1 mL
Água tamponada
Fosfato bibásico de sódio Na2HPO4 6 g
Fosfato monobásico de potássio KH2PO4 4 g
Misturar em gral de porcelana.
Diluir 1 g da mistura em 1000 mL de água destilada.
6.3.1.2 Armazenamento dos reagentes
Os reagentes utilizados nesse procedimento devem ser armazenados em temperatura
ambiente.

106
6.3.2 Procedimento
1ª fase: Desemoglobinização pela solução hipotônica de azul de metileno.
1. Aplicar a solução de azul de metileno fosfatado sobre a gota espessa de sangue, por dois
segundos.
2. Enxaguar a lâmina com água tamponada (sem jato forte).
2ª fase: Coloração pela solução de Giemsa para coloração da gota espessa.
1. As lâminas devem ser coradas dentro de 72 horas com uma solução de Giemsa a 10% com
água tamponada.
2. Colocar a lâmina com o lado da gota voltada para a superfície da placa de coloração.
3. Preparar uma solução de Giemsa na proporção de uma gota de corante para 1mL (1 gota)
de água tamponada. Homogeneizar
4. Aplicar esta solução na placa côncava de coloração, sob a lâmina invertida.
5. Deixar corar por 10 minutos.
6. Enxaguar com água tamponada (sem jato forte).
7. Secar ao calor suave ou sob ventilação.
3ª fase: Coloração do esfregaço método de Giemsa
1. Fixar o esfregaço com álcool metílico por um minuto.
2. Deixar secar.
3. Colocar a lâmina invertida sobre a placa de coloração.

107
4. Despejar a diluição do corante de Giemsa na proporção de uma gota do corante para 1mL
de água tamponada.
5. Deixar corar por 20 a 30 minutos.
6. Enxaguar com jato forte de água tamponada.
7. Secar ao calor suave ou sob ventilação.
4ª fase: Montagem das lâminas (no caso de a leitura da densidade parasitária não for realizada
no mesmo dia da coloração, realizar a montagem, como segue):
1. Pingar duas a três gotas de Entelan® em cada lâmina.
2. Colocar uma lamínula sobre as gotas.
3. Deixar secar sob temperatura ambiente.
6.4 Controle de qualidade da coloração de lâminas
1. A solução de Giemsa deve ser controlada antes do uso. Isto é realizado preparando a
solução de Giemsa 1:10 que será usada e corando uma lâmina sabidamente positiva. Não deve
haver coloração das hemácias.
2. Cada nova preparação de Giemsa deve ser submetida a controle de qualidade.
3. O tampão de pH deve ser checado utilizando pHmetro ou fita, estando o pH a 7,2. O pH
deve ser ajustado se necessário. O tampão deve ser analisado ao menos duas vezes por
semana se for preparado em grandes quantidades.

108
6.5 Leitura
1. Antes do início da contagem, o equivalente a 0.25 µL de sangue (aproximadamente 100
campos utilizando uma ocular de 10X e uma objetiva de 100X) deverá ser examinado para
determinar a espécie do parasito e estágios que possam estar presentes.
2. Coloque uma gota de óleo de imersão sobre a lamínula.
3. Ligue o microscópio. Cheque a presença de ocular de 10X.
4. Com o condensador elevado, a lâmina corada é colocada no suporte e a fonte de luz é
ajustada visibilizando pela ocular e pela objetiva de 100X.
5. Ao mover lentamente a objetiva de imersão, uma camada de óleo será formada entre a
lâmina e a lente. O ajuste fino é usado para focar o campo; a lente não deve tocar a lâmina.
6. O exame microscópico deve ser sistemático e padronizado. Ele deve iniciar pela
extremidade esquerda da lâmina. A leitura é iniciada da periferia do campo e termina no
centro. Quando o campo está lido, move-se a lâmina longitudinalmente para examinar os
campos adjacentes. Move-se a lâmina verticalmente para que outra fileira/largura seja lida. Há
cerca de 100 campos em um eixo de 2 cm da lâmina;
7. No início será realizada a leitura rápida das lâminas para guiar o tratamento e, após, será
feita a quantificação da densidade parasitária para determinar o resultado final.
6.5.1 Leitura rápida
1. As lâminas serão lidas imediatamente após a coleta para guiar o tratamento.
2. A leitura rápida será realizada conforme a prática padrão em cada centro.

109
3. Os resultados da leitura rápida serão transcritos no formulário apropriado de cada estudo
(versões atuais de POP_MAL_TC_002_A01, POP_MAL_TC_002_A02,
POP_MAL_HO_002_A01, POP_MAL_HO_002_A02)
6.5.2 Leitura da densidade parasitária: parasitos por μL
1. Uma vez coradas, as lâminas serão colocadas em caixas e distribuídas com o formulário de
primeira leitura de lâmina (versão atual de POP_MAL_LB_001_A01) ao microscopista que
realizará a primeira leitura.
2. Para contagem de parasitos e leucócitos separadamente, dois contadores devem ser usados,
um para as formas assexuadas do parasito e outro para os leucócitos. Caso detecte-se infecção
mista, outro contador deverá ser utilizado para a contagem de formas da outra espécie.
Determinando uma lâmina como negativa
1. O microscopista irá ler a lâmina até que 200 campos tenham sido contados.
2. A lâmina apenas será determinada negativa quando nenhum parasito for encontrado em 200
campos.
Caso sejam vistas formas assexuadas de Plasmodium spp.
1. O microscopista contará parasitos até que o número de 500 leucócitos ou 500 parasitos seja
alcançado.
2. Caso o microscopista já tenha contado 500 leucócitos ou mais quando o primeiro parasito
for visto, a leitura será interrompida.
3. A contagem de parasitos ou leucócitos não será interrompida até que o campo inteiro seja
lido.

110
4. Este método será usado tanto para infecções únicas ou mistas por Plasmodium. No segundo
caso, os parasitos de cada espécie deverão ser contados separadamente.
5. A contagem parasitária em relação à contagem de leucócitos pode ser convertida a parasitos
por μL usando a seguinte fórmula matemática:
Nº de parasitos X 8000 / Nº de leucócitos = parasitos por l
6. Está fórmula será calculada para cada espécie de parasito encontrada.
7. Caso o número de leucócitos para cada participante seja conhecido ( ex: estudo de malária
vivax grave), a densidade pode ser calculada mais precisamente com a seguinte fórmula:
Nº de parasitos X Nº de leucócitos / Nº de leucócitos = parasitos por μl
8. A contagem de gametócitos será feita de forma específica para cada espécie.
9. Este procedimento é semelhante aos métodos diagnósticos preconizados pelas diretrizes do
CLSI do CDC (contagem superior a 500 parasitos ou 1000 leucócitos) e também similar às
diretrizes da OMS (que para densidades < 10 parasitos por 200 leucócitos propõe contar
acima de 500 leucócitos, mas que para densidades superiores a 10 parasitos por 200 leucócitos
propõe parar a contagem ao atingir 200 leucócitos).
10. Após a primeira leitura, as lâminas serão mantidas na mesma ordem na bandeja ou caixa
para serem lidas posteriormente por outro microscopista. As lâminas serão entregues
conjuntamente com o formulário de segunda leitura de lâmina (versão atual de
POP_MAL_LB_001_A02) ao microscopista que realizará a segunda leitura, assegurando-se
que não seja a mesma pessoa que realizou a primeira leitura e que esta pessoa não tenha
acesso aos resultados no formulário de primeira leitura de lâmina.

111
6.6 Registro dos resultados
1. O microscopista registrará os resultados da leitura em μL nos formulários de primeira e
segunda leitura de lâmina. A primeira seção do formulário, incluindo a etiqueta de
identificação da amostra, já haverá sido preenchida no momento da coleta da amostra. A
seção de Resultados será preenchida pelo microscopista com a seguinte informação:
- Caso a lâmina for negativa, o número de campos examinados será registrado (deve ser 200)
e o número de parasitos deverá ser 0 para todas as espécies.
- Caso a lâmina for positiva, o número de leucócitos contados e o número de formas
assexuadas de parasitos contados para cada espécie deverão ser registrados.
- O número de gametócitos para cada espécie também deverá ser registrado.
- Caso a lâmina não possa ser lida, o motivo deverá ser registrado (não encontrada, quebrada,
má qualidade).
- O microscopista assinará e datará o formulário com a data da leitura e registrará seu código
do estudo.
- O microscopista que realizar a primeira leitura registrará os resultados no formulário de
primeira leitura de lâmina (versão atual de POP_MAL_LB_001_A01) e o que realizar a
segunda leitura registrará os resultados no formulário de segunda leitura de lâmina (versão
atual de POP_MAL_LB_001_A02).
- Os formulários resultado de primeira leitura e resultado de segunda leitura serão enviados
diariamente ao centro de registro de dados para serem inseridos nas bases de dados. Tais
formulários serão então armazenados pelos investigadores.

112
7 CÁLCULO DA DENSIDADE PARASITÁRIA E CONTROLE DE QUALIDADE
INTERNA
1. Todas as lâminas serão lidas duas vezes independentemente e os resultados registrados em
diferentes formulários de resultados de leitura de lâmina (primeira e segunda leituras), que
serão inseridos nas bases de dados.
2. As lâminas que NÃO caírem nos seguintes critérios deverão ser lidas uma terceira vez:
- Ambas as lâminas positivas, tendo ambas as leituras > 400 parasitos por μL e a proporção de
densidades de ambas as leituras (maior contagem / menor contagem) < 2.
- Ambas as lâminas positivas e uma ou ambas tem contagem < 400 parasitos por μl e a
maior contagem é menos que um log 10 superior à menor contagem.
- Ambas as lâminas negativas.
3. Um programa de computador gerará uma lista com as lâminas que devem ser lidas uma
terceira vez, baseado na proporção das densidades das duas primeiras leituras. No centro de
dados o programa produzirá impresso que será a ficha de Resultado de terceira leitura de
lâmina e incluirá o número de identificação de amostra, o número de identificação
permanente, a posição na caixa e os códigos dos microscopistas que realizaram as primeira e
segunda leituras.
4. No laboratório a lâmina será retirada da posição na caixa correspondente e será lida a
terceira vez por um microscopista.
5. O microscopista que realiza a terceira leitura registrará os resultados no impresso, que será
então devolvido ao centro de dados e inserido nas bases de dados.
6. O resultado definitivo (após a segunda leitura no caso de concordância ou após a terceira
leitura caso não haja concordância entre as duas primeiras) será calculado por um programa
de computador e levará em conta os seguintes critérios:

113
- Caso ambas as leituras sejam negativas, o resultado final será negativo.
- Caso as duas lâminas sejam positivas e haja concordância, o resultado final será a média
geométrica das duas leituras.
- Caso uma leitura seja positiva e a outra negativa e a terceira leitura seja positiva, o resultado
final será a média geométrica das duas densidades positivas. Caso a terceira leitura seja
negativa, o resultado final será negativo.
- Caso as três leituras sejam positivas, o resultado final será a média geométrica das duas
leituras com densidades mais próximas.
- Todos os resultados de leituras de lâminas serão armazenados pelos investigadores.
8 ARMAZENAMENTO DAS LÂMINAS
1. As lâminas serão levadas a um(a) técnico(a) de laboratório designado(a). Este deverá
conferir conjuntamente com o investigador que as trouxe a presença das lâminas e o correto
preenchimento dos formulários correspondentes.
2. No caso de a lâmina não haver sido montada com o Entelan® após a coloração, tal
procedimento deverá ser realizado, após a leitura do segundo microscopista para que a lâmina
seja armazenada.
3. Em seguida as lâminas deverão ser armazenadas nas caixas apropriadas, sendo o local de
cada lâmina registrado nos formulários de Resultado de primeira leitura de lâmina e Resultado
de segunda leitura de lâmina (versão atual de POP_MAL_LB_001_A01 e
POP_MAL_LB_001_A02).

114
4. As caixas serão numeradas à medida que forem preenchidas. As datas das lâminas que
ocuparem a primeira e a última posição em cada caixa deverão estar escritas no exterior da
caixa.
5. A partir do registro da posição das lâminas nas caixas, o microscopista retirará as lâminas à
medida que realizar as leituras de densidade parasitária.
6. O segundo microscopista deverá atentar para o número da caixa e a posição da lâmina
detalhados na ficha de Resultado de segunda leitura de lâmina (versão atual de
POP_MAL_LB_001_A02) para recolocar a lâmina na posição correta após a leitura.
7. Caso uma terceira leitura seja necessária, a lâmina será retirada da caixa e colocada
novamente na mesma posição quando a terceira leitura for encerrada.
8. As duplicatas das lâminas serão mantidas em uma caixa de lâminas distinta e armazenadas
no laboratório, sendo usadas apenas no caso de perda ou quebra da primeira lâmina. As datas
das lâminas que ocuparem a primeira e a última posição em cada caixa deverão estar escritas
no exterior da caixa.
9. As caixas de armazenamento devem ser mantidas num local trancado no laboratório, sob
condições apropriadas de temperatura e umidade e protegidas de luz direta.
9. BIOSSEGURANÇA
Todo o procedimento deve se realizar sob condições de biossegurança (1).
10 REFERÊNCIAS
1. Richmond JY, Mckinney RW. Biossegurança em laboratórios biomédicos e de
microbiologia. Brasília: Ministério da Saúde: Fundação Nacional de Saúde; 2001.
2. WHO manual for malaria diagnostic in developing countries.

115
3. Diagnoses and management of severe falciparum malaria. WHO/CDS/CPE/SMT/2000.4
4. Diagnostic procedures for blood specimens. Diagnostic for Parasitic Diseases, CDC,
Atlanta.
5. Manual de diagnóstico laboratorial da malária. Brasília: Secretaria de Vigilância em
Saúde/Ministério da Saúde; 2005

116
Pop para extração de DNA de Plasmodium spp. a partir de sangue total
Código POP POP_MAL_LB_003_v02D_PT
Título Procedimento para extração de DNA de Plasmodium spp. a partir de
sangue total
Idioma da versão original PORTUGUÊS
Elaborado por:
Gisely Cardoso de Melo
Revisado por:
Aprovado por:
Wuelton Marcelo
Monteiro
Data de aplicação:
05 de julho de 2010
Data da próxima revisão:
Emenda Razão da emenda
.
OBJETIVOS
Descrever o procedimento da extração de DNA de Plasmodium spp. a partir de sangue total.
DEFINIÇÕES
Não se aplica
APLICÁVEL A
Análise no laboratório de biologia molecular de amostras relacionadas à pesquisa.
Código: POP_MAL_LB_002_v01D_PT
Procedimento Operacional Padrão
Gerência de Malária
S

117
RESPONSABILIDADES
Pessoal encarregado da coleta das amostras da pesquisa.
Equipe do laboratório de biologia molecular.
POP'S RELACIONADOS
Seguimento dos participantes no estudo “Epidemiologia da Malária no Município do Careiro, Amazonas”
(versão atual do POP_MAL_TC_002).
Caracterização da Malária Grave por Plasmodium vivax na Fundação de Medicina Tropical Dr. Heitor
Vieira Dourado, Brasil (versão atual do POP_MAL_HO_001).
Procedimento para realização de PCR em tempo real para detecção de plasmódio (versão atual do
POP_MAL_LB_002).
PROCEDIMENTOS
6.1.2 Materiais
1. Pipetas automáticas de 10 µL, 100 µL e 1000 µL
2. Ponteiras com filtro para pipetas automáticas
3. Tubos para 1,5 e 2,0 mL
6.1.3 Equipamentos
1. Centrífuga
2. Vórtex
3. Banho Maria

118
6.1.4 Reagentes
Reagente Armazenamento
1. Tampão Lise dos tecidos (Tampão ATL) Temperatura ambiente
2. Proteinase K Temperatura ambiente
3. Tampão de Lise (Tampão AL) Temperatura ambiente
4. Etanol 95% PA Temperatura ambiente
5. Tampão de Lavagem 1 (Tampão AW1) Temperatura ambiente
6. Tampão de Lavagem 2 (Tampão AW2) Temperatura ambiente
7. Tampão de Eluição (Tampão AE) Temperatura ambiente
6.2 Obtenção de sangue total
6.2.1 Por punção endovenosa
1. Obter 5 mL de sangue venoso em tubo de EDTA identificado.
2. Centrifugar a 14000 rpm por 10 min.
3. Desprezar o soro e colocar a papa de hemácias em criotubo identificado.
4. Armazenar os criotubos a – 20ºC em caixa de plástico ou papelão identificada e numerada.
5. No laboratório de biologia molecular, os criotubos serão armazenados seguindo a mesma ordem que
as lâminas.

119
6.3 Extração do DNA.
Para a extração do DNA a partir de sangue total será usado o Protocolo de Extração do QIAMP® DNA
Mini Kit da Qiagen.
6.3.1 Procedimento para extração do DNA a partir de tubo
1. Pipetar 20 µL de Proteinase K (2) num tubo eppendorf de 1,5 mL.
2. Adicionar 200 µL de amostra ao tubo. Se não houver 200 µL de amostra, adicionar o volume
apropriado de PBS.
3. Adicionar 200 µL de Tampão AL (3) a amostra. Vortexar por 15 segundos.
4. Incubar por 56°C por 10 minutos.
5. Adicionar 200 µL de Etanol absoluto (96 a 100%) e agitar no vórtex imediantamente. Remover as
bolhas por centrifugação.
6. Cuidadosamente transferir o conteúdo do tubo para o tubo filtro. Tampar e centrifugar a 8000 rpm por
1 minuto. Descartar o filtrado com o tubo. Colocar o tubo filtro em um novo tubo coletor.
7. Cuidadosamente adicionar 500 µL de tampão AW1 e centrifugar a 8000 rpm por 1 minuto. Descartar o
filtrado com o tubo. Colocar em um novo tubo coletor.
8. Cuidadosamente adicionar 500 µL de tampão AW2 e centrifugar a 14000 rpm por 3 minutos.
9. Descartar o tubo coletor com o filtrado. Colocar o tubo filtro em novo tubo coletor e centrifugar a
14000 rpm por 1 minuto.

120
10. Colocar o tubo filtro em um tubo eppendorf de 1,5 mL novo. Descartar o filtrado e o tubo coletor.
Adicionar cuidadosamente 150 µL de tampão AE ou água destilada e incubar à temperatura ambiente por
1 minuto. Centrifugar a 8000 rpm por 1 minuto.
11. Armazenar por -20°C.
BIOSSEGURANÇA
Todo o procedimento deve se realizar sob condições de biossegurança (1).
7 REFERÊNCIAS
1. Richmond JY, Mckinney RW. Biossegurança em laboratórios biomédicos e de microbiologia. Brasília:
Ministério da Saúde: Fundação Nacional de Saúde; 2001.

121
Pop para realização de PCR em tempo real para detecção de plasmódio
Código POP POP_MAL_LB_003_v02D_PT
Título Procedimento para realização de PCR em tempo real para detecção de
plasmódio.
Idioma da versão original PORTUGUÊS
Elaborado por:
Gisely Cardoso de Melo
Revisado por:
Aprovado por:
Wuelton Marcelo
Monteiro
Data de aplicação:
19 de fevereiro de 2009
Data da próxima revisão:
Emenda Razão da emenda
V02 Revisão dos procedimentos de PCR em tempo real
.
OBJETIVOS
Descrever o procedimento de realização de PCR em tempo real para detecção de plasmódio.
DEFINIÇÕES
PCR: a reação em cadeia da polimerase é definida como uma técnica molecular que permite a
amplificação de pequenas quantidades de um DNA específico a partir de um molde do DNA,
usando uma reação enzimática simples, sem um organismo vivo. O limiar de detecção da
técnica é muito superior ao da gota espessa (técnica padrão utilizada no diagnóstico rotineiro
da malária): cerca de 0,004 parasitos/<L.
Código: POP_MAL_LB_003_v02D_PT
Procedimento Operacional Padrão
Gerência de Malária

122
PCR Tempo Real (PCR TR): Variação da técnica de PCR para detecção de sinal
fluorescente permitindo a quantificação de ácidos nucléicos em tempo real, assim como a
detecção qualitativa de sequências de ácidos nucléicos utilizando análises de curvas de
dissociação ou por meio de sondas marcadas. Não requer tratamento pós-PCR para
visualização do resultado.
Iniciador (primer): uma pequena sequência de DNA ou RNA a partir da qual pode começar
a replicação do DNA.
APLICÁVEL A
Análise no laboratório de biologia molecular de amostras relacionadas à pesquisa clínica.
RESPONSABILIDADES
Equipe do laboratório de biologia molecular envolvida na realização da PCR em tempo real.
POP'S RELACIONADOS
Seguimento dos participantes no estudo “Epidemiologia da Malária no Município do Careiro,
Amazonas” (versão atual do POP_MAL_TC_002).
Caracterização da Malária Grave por Plasmodium vivax na Fundação de Medicina Tropical
Dr. Heitor Vieira Dourado, Brasil (versão atual do POP_MAL_HO_001).
Procedimento para extração de DNA de Plasmodium spp. a partir de mosquitos inteiros ou
cabeça + tórax (versão atual de POP_MAL_LB_004).
PROCEDIMENTOS
Recursos necessários
6.1.2 Materiais
1. Pipetas automáticas de 10 µL, 100 µL e 1000 µL

123
2. Ponteiras com filtro para pipetas automáticas
3. Tubos de 1,5 e 2,0 mL
4. Microplacas de 0,1mL, com 96 poços, para sistema de PCR em Tempo Real
5. Filme óptico para vedar as microplacas
6.1.3 Equipamentos
1. Vórtex
2. Sistema de PCR em Tempo Real 7500 Fast Applied Biosystems
6.1.4 Reagentes
Reagente Armazenamento
Taqman Universal PCR Master Mix -20°C
Sondas (Tamra Taqman® Probe) -20°C
Iniciadores (primers) da reação -20°C
Água MiliQ Temperatura ambiente
Iniciadores:
Plasmodium vivax

124
VIV-F 5’ACGCTTCTAGCTTAATCCACATAACT
VIV-R 5’ATTTACTCAAAGTAACAAGGACTTCCAAGC
Plasmodium falciparum
FAL-F 5’CTTTTGAGAGGTTTTGTTACTTTGAGTAA
FAL-R 5’TATTCCATGCTGTAGTATTCAAACACAA
Sondas
FAL probe (FAM) 5_-TGTTCATAACAGACGGGTAGTCATGATTGAGTTCA 3' TAMRA
VIV probe (VIC) 5_-TTCGTATCGACTTTGTGCGCATTTTGC 3' TAMRA
6.2 PCR em Tempo Real
Para a detecção dos principais parasitos que infectam humanos, foi padronizada uma reação
de PCR em Tempo Real baseada no gene que codifica a menor subunidade do rRNA (1). Os
iniciadores (primers) usados para amplificação do P. vivax e P. falciparum flanqueiam regiões
espécie-específicas de 120pb e 396pb respectivamente.
6.2.1 Reação de PCR em Tempo Real
P. vivax
Extrato de DNA (50 a 100ng) 2µL
Taqman Master Mix 10 µL
VIV Probe 200nM
Iniciador F 300 nM

125
Iniciador R 300 nM
H2O para PCR 1µL
P. falciparum
Extrato de DNA (50 a 100ng) 2µL
Taqman Master Mix 2X 10 µL
FAL Probe 200 nM
Iniciador F 300 nM
Iniciador R 300 nM
H2O para PCR 1µL
Condições de amplificação:
1: 50ºC/2 minutos
2: 95ºC/10 minutos
3: 95ºC/15 segundos- total de 45 ciclos
4. 60 ºC/ 1 minuto
Modulo de corrida: Standard 7500.
6.2.2 Em cada reação de PCR, é realizado um controle negativo (água miliQ) e um controle
positivo contendo DNA de Plasmodium sp (P. falciparum - MR4, P. vivax – amostras de
cultivo in vitro e in vivo, respectivamente).
6.2.3 As amostras são testadas em duplicatas em cada experimento.

126
1 2 3 4 5 6 7 8 9 10 11 12
A
B
C
D
E
F
G
H CPV CPV CPF CPF CN CN
CPV: Controle positivo de Plasmodium vivax
CPF: Controle positivo de Plasmodium falciparum
CN: Controle negativo
6.2.4 As análises são realizadas pelo software distribuído pelo fabricante – Applied
Biosystems “7500 Fast System SDS Software”, e utiliza-se como ponto de corte Cts abaixo
de 40 ciclos.
6.2.5 Os valores do CT são comparadas às dos controles para verificar a especificidade da
reação e do produto gerado.

127
6.3 Restrições e limitações do procedimento
Como se faz a amplificação de DNA, a técnica não permite distinguir as diferentes formas
biológicas do parasito (trofozoíto, esquizonte, merozoíto ou gametócito). Existe a
possibilidade de contaminação da reação no momento da extração do DNA e no momento do
preparo da reação do PCR, dada a sensibilidade de amplificação da técnica.
6.4 Biossegurança
Todo o procedimento deve se realizar sob condições de biossegurança (2).
6.5 Registro dos resultados
O resultado da PCR será preenchido pelo responsável do procedimento no questionário
Resultado da PCR (POP_MAL_LB_002_A01_v01_PT), indicando resultado por espécie, o
motivo em caso de não ter realizado o exame, a ordem de armazenamento, o código e a
assinatura do membro da equipe do projeto e a data do resultado.
7 REFERÊNCIAS
1. De Monbrison F, Angei C, Staal A, Kaiser K, Picot S. Simultaneous identification of the
four human Plasmodium species and quantification of Plasmodium DNA load in human blood
by real-time polymerase chain reaction. Trans. R. Soc. Trop. Med. Hyg 2003;97:387–390.
2. Richmond JY, Mckinney RW. Biossegurança em laboratórios biomédicos e de
microbiologia. Brasília: Ministério da Saúde: Fundação Nacional de Saúde; 2001.

128
Pop para extração de RNA utilizando Trizol®
Código POP POP_MAL_LB_004_v01D_PT
Título Procedimento para extração de RNA utilizando Trizol®
Idioma da versão original PORTUGUÊS
Elaborado por:
Gisely Cardoso de Melo
Revisado por:
Gisely Cardoso de Melo
Aprovado por:
Wuelton Marcelo
Monteiro
Data de aplicação:
30 de setembro de 2011
Data da próxima
revisão:
Emenda Razão da emenda
1 OBJETIVOS
Descrever o procedimento da extração de RNA utilizando o trizol®.
2 DEFINIÇÕES
RT-PCR: é uma reação da transcriptase reversa, seguida de reação em cadeia da polimerase. Não
utiliza o DNA de cadeia dupla como molde e sim RNA de cadeia simples. A partir do RNA, a
enzima transcriptase reversa sintetiza uma cadeia de DNA complementar (chamado agora
de cDNA). Ao cDNA aplica-se a técnica de PCR.
3 APLICÁVEL A
Análise no laboratório de biologia molecular de amostras relacionadas à pesquisa.
4 RESPONSABILIDADES
Pessoal encarregado da coleta das amostras da pesquisa.
Código: POP_MAL_LB_004_v02D_PT
Procedimento Operacional Padrão
Gerência de Malária
S

129
Equipe do laboratório de biologia molecular.
5 POP'S RELACIONADOS
Procedimento para realização de RT-PCR (versão atual do POP_MAL_LB_005).
6 PROCEDIMENTOS
6.1 Recursos necessários
6.1.2 Materiais
1. Tubos cônicos de 15 mL RNAse free
2. Tubos de 1,5mL RNAse free
3. Saponina
4. RPMI
5. PBS 1x
6. Pipetas automáticas de 10 µL , 100 µL, 200 µL e 1000 µL.
7. Ponteiras com filtro para pipetas automáticas RNAse free
8. Clorofórmio
9. Isopropanol
10. Etanol
11. H2ODEPC
12. Formamida
13. Dodecil sulfato de sódio (SDS)
14. H2O RNAse free
15. Tris-borate-EDTA (TBE)
16. Agarose

130
17. Loading Buffer 2X
6.1.3 Equipamentos
1. Centrífuga refrigerada para tubos cônicos
2. Centrífuga refrigerada de eppendorf
3. Cuba de Eletroforese
4. Banho maria
5. Termobloco
6.2 Obtenção do RNA
6.2.1 Protocolo de tratamento do sangue com saponina
1. Coletar 5 mL de sangue em tubos a vácuo estéreis;
2. Centrifugar o sangue a 3000 rpm por 5 minutos para separar o plasma;
3. Retirar o plasma em tubos de 1,5 mL e guardar em freezer -80°C;
4. Separar a papa de hemácias remanescentes em dois tubos cônicos contendo 5 mL de RPMI
1640 e µL de saponina 1,5%.
5. Agitar o tubo e aguardar por 5 minutos;
6. Centrifugar a 3.000 rpm por 12 minutos a 20°C. Se o pellet permanecer escuro, repetir a etapa
4,5,6;
7. Retirar o sobrenadante e transferir o pellet para um tubo de 1,5 mL RNAse free contendo 1mL
de RPMI e homogeneizar bem;
8. Centrifugar a 4°C por 5 minutos a 13.000 rpm;

131
9. Repetir as etapas 7 e 8;
10. Retirar o sobrenadante e adicionar trizol® e armazenar a -80°C até a extração do RNA.
6.2.2 Protocolo de extração
1. Adicionar 200µL de clorofórmo e homogeneizar vigorosamente por 15s e incubar por 5-15
min a temperatura ambiente;
2. Centrifugar a 12.000g por 15 min a 4°C;
3. Transferir o sobrenadante para um novo tubo com 500µL de isopropanol e incubar por 10 min
a temperatura ambiente.
4. Centrifugar a 12.000g por 10 min a 4°C
5. Descartar o sobrenadante e lavar com 1mL de etanol 75 %
6. Centrifugar a 7500g por 5 min 4°C;
7. Ressuspender o pellet em um volume desejado de H2ODEPC ou Formamida e deixar em um
termobloco a 65C por 3-5 min posteriormente agitar em um vórtex por 2 min e dar un spin de
varios segundos.
8. Quantificar o RNA no Nanodrop utilizando 1ul da amostra
6.2.3 Análise da qualidade do RNA em gel de agarose
1. Deixar todo o material (Cuba, base e pentes) em SDS 1% por 1h, lavar com H2O RNAse free
(alternativamente também pode ser usada H2O MilliQ esteril já que se necessita de um grande
volume) e secar com etanol 70%.

132
2. Preparar um gel de agarose 1% em TBE 0,5 X (todas as solucões e reagentes devem ser RNase
free).
3. Colocar en cada novo tubo aproximadamente 10% do volume final da amostra diluida 1:1 com
Loading Buffer 2X especifico para RNA (Fermentas). A amostra diluida deve ser colocada a 70
C por 10 min, esfriada rapidamente em gelo posteriormente, dar um spin, aplicar todo o volume
no gel.
6.3 Biossegurança
Todo o procedimento deve se realizar sob condições de biossegurança (1)
7 REFERÊNCIAS
1. Chomczynski, P. (1993) A reagent for the single-step simultaneous isolation of RNA, DNA
and proteins from cell and tissue samples. BioTechniques 15, 532-537
2. Chomczynski, P., and Sacchi, N. (1987) Single Step Method of RNA Isolation by Acid
Guanidinium Thiocyanate-Phenol-Chloroform Extraction. Anal. Biochem. 162, 156-159
3. Hummon, A. B., Lim S. R., Difilippantonio, M. J., Ried, T. (2007) Isolation and solubilization
of proteins after TRIzol® extraction of RNA and DNA from patient material following
prolonged storage. BioTechniques 42, 467-472
1. REGISTRO DOS ANEXOS

133
TÍTULO DO ANEXO CÓDIGO DO ANEXO (Incluindo
versão/idioma):
NOME DO ANEXO:
CÓDIGO DO ANEXO (Incluindo
versão/idioma):
Novo
Criado por:
Data e assinatura
Aprovado por:
Data e assinatura
Revisão
Tradução
A versão atual deste POP foi traduzida a ____________________ e a
versão traduzida entra em efeito em ___/___/_____.
Assinatura do responsável: ______________________
dd mm aa

134
Pop para RT-PCR para expressão gênica de pvcrt-o e pvmdr-1.
Código POP POP_MAL_LB_005_v01D_PT
Título Procedimento para RT-PCR para expressão gênica de pvcrt-o e pvmdr-1.
Idioma da
versão
original
PORTUGUÊS
Elaborado por:
Gisely Cardoso de Melo
Revisado por:
Gisely Cardoso de Melo
Aprovado por:
Data de aplicação:
30 de setembro de 2011
Data da próxima
revisão:
Emenda Razão da emenda
1 OBJETIVOS
Descrever o procedimento da extração de RNA utilizando o trizol®.
2 DEFINIÇÕES
RT-PCR: é uma reação da transcriptase reversa, seguida de reação em cadeia da polimerase.
Não utiliza o DNA de cadeia dupla como molde e sim RNA de cadeia simples. A partir do RNA,
a enzima transcriptase reversa sintetiza uma cadeia de DNA complementar (chamado agora
de cDNA). Ao cDNA aplica-se a técnica de PCR.
Código: POP MAL_LB_005_v01D_PT
Procedimento Operacional Padrão
Gerência de Malária
S

135
PCR Tempo Real (PCR TR): Variação da técnica de PCR para detecção de sinal fluorescente
permitindo a quantificação de ácidos nucléicos em tempo real, assim como a detecção qualitativa
de seqüências de ácidos nucléicos utilizando análises de curvas de dissociação ou por meio de
sondas marcadas. Não requer tratamento pós-PCR para visualização do resultado.
3 APLICÁVEL A
Análise no laboratório de biologia molecular de amostras relacionadas à pesquisa.
4 RESPONSABILIDADES
Pessoal encarregado da coleta das amostras da pesquisa.
Equipe do laboratório de biologia molecular.
5 POP'S RELACIONADOS
Procedimento para extração de RNA utilizando o trizol® (versão atual do POP_MAL_LB_004).
6 PROCEDIMENTOS
6.1 Recursos necessários
6.1.1 Amostra obtida por extração de RNA seguindo POP_MAL_LB_004.
6.1.2 Materiais
1. Pipetas automáticas de 10 µL, 100 µL e 1000 µL
2. Ponteiras com filtro para pipetas automáticas RNAse free
3. Tubos de 0,2 mL
4. Microplacas de 0,1mL, com 96 poços, para sistema de PCR em Tempo Real

136
5. Filme óptico para vedar as microplacas
6.1.3 Equipamentos
1. Centrífuga refrigerada de eppendorf
2. Sistema de PCR em Tempo Real 7500 Fast Applied Biosystems
3. Banho Maria
6.1.4 Reagentes
Reagente Armazenamento
SuperScript™ III RT (200 U/µl) -20°C
5X First-Strand Buffer -20°C
0.1 M DTT -20°C
Random Primers -20°C
10 mM dNTP Mix -20°C
RNAseOUT™ Recombinant Ribonuclease Inhibitor (40 U/µl) -20°C
10X PCR Buffer -20°C
50 mM MgCl2 -20°C
Taq DNA polymerase (5 U/µl) -20°C

137
DNase I, Amp Grade -20°C
10X DNase I Reaction Buffer -20°C
25 mM EDTA (pH 8.0) -20°C
Iniciadores:
pvcrt-o
F-pvcrt-oRT 5'-ATGTCCAAGATGTGCGACGAT-3'
R-pvcrt-oRT 5'-CTGGTCCCTGTATGCAACTGAC-3'
Pvmdr-1
F-pvmdr1RT 5'-AAGGATCAAAGGCAACCCA-3'
R-pvmdr1RT5'-TCAGGTTGTTACTGCTGTTGCTATT-3'
Tubulina (controle interno)
F-pvtubulinRT 5' CCAAGAATATGATGTGTGCAAGTG 3'
R-pvtubulinRT 5' GGCGCAGGCGGTTAGG 3'
6.2 Reação de RT-PCR
6.2.1 Tratamento do RNA total com DNAse
RNA ? µL (1µg)

138
Buffer 10X 1 µL
DNase 1 µL
H2O RNAse free para 10 µl
Incubar 15 minutos a temperatura ambiente
Adicionar 2 µL de EDTA 25 mM.
Incubar por 10 minutos a 65 C.
Após esse procedimento, as amostras serão transferidas para o gelo e imediatamente submetidas
à reação de transcrição reversa.
6.2.2 Síntese de cDNA
1. Adicionar os componentes em um tubo de PCR RNAse free, sendo o volume final de 13 µL:
1.1 RNA tratado (~ 500 ng) 10 µL
1.2 Random Primers (300 ng/µl) 1 µL
1.3 10nM de mix de dNTPs 1 µL
1.4 H2O 1 µL
Incubar a 65 C por 5 min e depois no gelo por 1 minuto.
2. Preparo do mix:
2.1. Tampão 5X...................................................4 µL

139
2.2 DTT 0.1 M.....................................................1 µL
2.3 Rnase OUT...................................................1 µL
2.4 SuperScript™ III RT...................................... 1 µL
25 C por 5 minutos
50 C por 45 minutos
70 C por 15 minutos
4 C
6.2.3 Reação de PCR
10X PCR Buffer 2-4µL
50 mM MgCl2 10 µL
10 mM dNTP Mix) 0,2 µL
Sense primer (10 µM) 0,2 µL
Antisense primer (10 µM) 0,2 µL
Taq DNA polymerase (5 U/µl) 0.4 µL
cDNA (from first-strand reaction) 2 µL
Água para PCR para 50 µL
Condições de amplificação:

140
1: 94ºC/2 minutos
2. 15-40 ciclos
6.2.4 Protocolo de PCR tempo real
Extrato de cDNA 2-4µL
PowerSYBR Green PCR Master Mix 10 µL
Iniciador F (10 uM) 0,2 µL
Iniciador R (10 uM) 0,2 µL
H2O para PCR para 20 µL
Condições de amplificação:
1: 95ºC/10 minutos
2: 95ºC/15 segundos- total de 40 ciclos
3. 60 ºC/ 1 minuto
Modulo de corrida: Standard 7500
6.3 Biossegurança
Todo o procedimento deve se realizar sob condições de biossegurança (1)
7 REFERÊNCIAS

141
1. Kotewicz, M.L., D'Alessio, J.M., Driftmier, K.M., Blodgett, K.P., and Gerard, G.F. (1985)
Gene 35, 249.
2. Gerard, G.F., D'Alessio, J.M., Kotewicz, M.L., and Noon, M.C. (1986) DNA 5, 271.
3. Houts, G.E., Miyagi, M., Ellis, C., Beard, A., and Beard, J.W. (1979) J. Virol.29, 517.

142
8.2 Anexo 2
APROVAÇÃO DO COMITÊ DE ÉTICA EM PESQUISA COM SERES HUMANOS

143
8.3 Anexo 3
TERMO DE CONSENTIMENTO LIVRE E ESCLARECIDO

144
Você está sendo convidado para participar da pesquisa: Avaliação da expressão gênica de
pvcrt-o e pvmdr-1 de pacientes com malária grave atendidos na FMT-HVD. A qualquer
momento você pode desistir de participar e retirar seu consentimento. Sua recusa não trará
nenhum prejuízo em sua relação com o pesquisador ou com a instituição. O acompanhamento da
sua doença e tratamento continuará sendo realizado independentemente de você estar na pesquisa
ou não. Você será retirado do estudo caso você deixe de seguir as orientações para a participação
no estudo.
A Cidade de Manaus apresenta, anualmente, um grande número de pessoas acometidas
com malária. Para melhor entendermos a melhor forma da doença, o controle e tratamento, nós
estamos convidando você a considerar a participação deste estudo. Precisamos do maior número
possível de pessoas.
Trata-se de um estudo observacional sobre a expressão de genes de resistência a
cloroquina relacionada à gravidade pelo P. vivax em pacientes internados na PESCLIN ou
atendidos no ambulatório de malária da FMT-HVD e terá duração de 12 meses. Você não precisa
fazer qualquer coisa especial para participar desse projeto e receberá o mesmo tratamento e
acompanhamento estabelecido de acordo com as normas do Ministério da Saúde.
Nós coletaremos além de várias informações clínicas, uma amostra de sangue (15 mL)
através de uma agulha (venopunção) para fazer exames laboratoriais como sorologia,
hemograma e bioquímica.
Nós gostaríamos de sua permissão para fazer testes com as amostras de sangue e para
guardar o restante do sangue para ser usado em estudos sobre malária no futuro. Isto pode exigir
o armazenamento mais prolongado de seu sangue em laboratórios da FMT-HVD. Para isso, seu
sangue será guardado com um código e não deverá identificar seu nome.
Você pode sentir alguma dor por causa da coleta de amostra de sangue na veia do braço.
Mas qualquer dor deve durar apenas alguns instantes. Existe um risco muito pequeno de infecção
onde o sangue for coletado, mas qualquer infecção será monitorada e tratada pela equipe médica.
A amostra de sangue colhida é pequena e não representa nenhum risco à sua saúde.

145
O benefício em estar participando deste estudo será um aumento de informações sobre os
genes relacionados a resistência do P. vivax a cloroquina, mas você não será pago nem receberá
incentivo financeiro durante o acompanhamento.
Os participantes da pesquisa que vierem a sofrer qualquer tipo de dano previsto ou não
no termo de consentimento e resultante de sua participação, além do direito à assistência
integral, tem direito à indenização. Jamais poderá ser exigido do participante da pesquisa, sob
qualquer argumento, renúncia ao direito à indenização por dano.
Se você concordar em participar, todas as informações coletadas serão confidenciais,
usadas somente no estudo. Nós não compartilharemos suas informações, e um código será usado
para identificar você em vez de seu nome. Nós não tornaremos público qualquer detalhe sobre
você. No caso de algum pesquisador tirar uma foto sua, ele cuidará para que você não seja
identificado. Esta imagem sua será publicada apenas em revistas para médicos.
Pessoas para contato
Se você tiver qualquer pergunta ou preocupação sobre o estudo, por favor, vamos
esclarecer isso agora. Mesmo assim, se depois você desejar esclarecer suas dúvidas sobre a
pesquisa ou sobre ser um sujeito de pesquisa, por favor, sinta-se à vontade para contatar Gisely
Cardoso de Melo, pesquisador principal do projeto na Fundação de Medicina Tropical Dr.
Heitor Vieira Dourado. O endereço do hospital é Av. Pedro Teixeira, nº 25, Dom Pedro e o
número do telefone é (92) 2127-3443. O endereço do Comitê de Ética em Pesquisa da FMT-
HVD (que é um grupo de pessoas que avaliam este projeto e acompanham a pesquisa) é também
na Av. Pedro Teixeira, n° 25, Bairro Dom Pedro, e o telefone para contato é o (92) 2127 3432.
O presidente deste comitê é o Dr. Francisco Nailson Santos Pinto.

146
Eu,..............................................................................................................., participante
desse estudo ou responsável pelo paciente ......................................................................................,
entendi todas as informações dadas a mim sobre o estudo: Avaliação da expressão gênica de
pvcrt-o e pvmdr-1 de pacientes com malária grave atendidos na FMT-HVD, inclusive o seu
propósito e a forma como será executado. Eu tive a chance de fazer perguntas sobre o estudo e
concordo em participar do mesmo.
________________________________ Data ___/___/___
Assinatura do participante ou responsável
ou impressão do polegar direito
_________________________ ____________________ Data___/___/____
Nome do entrevistador Assinatura do entrevistador
_____________________________ ________________________ Data___/___/____
Nome do pesquisador responsável Assinatura do pesquisador responsável
Deve colocar a cópia da aprovação do CEP para seu estudo.
E os pacientes resistentes que usou do estudo do André? Existe alguma aprovação para isso? Foi
submetida alguma emenda ao protocolo dele ou você submeteu um estudo em separado.

147
8.4 Anexo 4
ARTIGOS COMPLETOS PUBLICADOS NA VIGÊNCIA DO CURSO

148
ARTIGOS PUBLICADOS
1. Monteiro WM, Franca GP, Melo GC, Queiroz AL, Brito M, Peixoto HM, et al. Clinical complications of G6PD deficiency in Latin American and Caribbean populations: systematic review and implications for malaria elimination programmes. Malar J. 2014;13:70. Epub 2014/02/27.

149
2. Coelho HC, Lopes SC, Pimentel JP, Nogueira PA, Costa FT, Siqueira AM, et al. Thrombocytopenia in Plasmodium vivax malaria is related to platelets phagocytosis. PLoS One. 2013;8(5):e63410. Epub 2013/06/01.

150
3. Chehuan YF, Costa MR, Costa JS, Alecrim MG, Nogueira F, Silveira H, et al. In vitro chloroquine resistance for Plasmodium vivax isolates from the Western Brazilian Amazon. Malar J. 2013;12:226. Epub 2013/07/04.

151
4. Versiani FG, Almeida ME, Melo GC, Versiani FO, Orlandi PP, Mariuba LA, et al. High levels of IgG3 anti ICB2-5 in Plasmodium vivax-infected individuals who did not develop symptoms. Malar J. 2013;12:294. Epub 2013/08/28.

152
5. Fabbri C, de Cassia Mascarenhas-Netto R, Lalwani P, Melo GC, Magalhaes BM, Alexandre MA, et al. Lipid peroxidation and antioxidant enzymes activity in Plasmodium vivax malaria patients evolving with cholestatic jaundice. Malar J. 2013;12:315. Epub 2013/09/12.

153
6. Brasil LW, Areas AL, Melo GC, Oliveira CM, Alecrim MG, Lacerda MV, et al. Pfatp6 molecular profile of Plasmodium falciparum isolates in the western Brazilian Amazon. Malar J. 2012;11:111. Epub 2012/04/11.

154
7. Martins-Campos KM, Pinheiro WD, Vitor-Silva S, Siqueira AM, Melo GC, Rodrigues IC, et al. Integrated vector management targeting Anopheles darlingi populations decreases malaria incidence in an unstable transmission area, in the rural Brazilian Amazon. Malar J. 2012;11:351. Epub 2012/10/24.

155
8. Magalhaes BM, Alexandre MA, Siqueira AM, Melo GC, Gimaque JB, Bastos MS, et al. Clinical Profile of Concurrent Dengue Fever and Plasmodium vivax Malaria in the Brazilian Amazon: Case Series of 11 Hospitalized Patients. Am J Trop Med Hyg. 2012. Epub 2012/10/04.

156
9. Machado Siqueira A, Lopes Magalhaes BM, Cardoso Melo G, Ferrer M, Castillo P, Martin-Jaular L, et al. Spleen rupture in a case of untreated Plasmodium vivax infection. PLoS Negl Trop Dis. 2012;6(12):e1934. Epub 2012/12/29.

157
10. Melo GC, Reyes-Lecca RC, Vitor-Silva S, Monteiro WM, Martins M, Benzecry SG, et al. Concurrent helminthic infection protects schoolchildren with Plasmodium vivax from anemia. PLoS One. 2010;5(6):e11206. Epub 2010/06/25.

158
ARTIGOS ACEITOS PARA PUBLICAÇAO
1. Gomes T, Mourao L, Melo GC, Monteiro WM, Braga E. Potential immune mechanisms
associated with anemia in Plasmodium vivax malaria: a puzzling question. Infection and
Immunity , 2014.
2. Monteiro WM, Val F, Siqueira AM, Franca GP, Sampaio V, Melo GC, et al. G6PD deficiency
in Latin America: systematic review on prevalence and variants. . Mem Inst Oswaldo Cruz. 2014.





























