Embed Size (px)
Citation preview

Universidade Estadual de Montes Claros
Programa de Pós-Graduação Stricto Sensu em Ciências Biológicas
SUCESSO REPRODUTIVO DE Dimorphandra mollis (LEGUMINOSAE:
MIMOSOIDAE): O PAPEL DA INTERAÇÃO FORMIGA-HOMÓPTERA
KÁTIA CIPRIANA PEREIRA SILVA SANTOS
Montes Claros – Minas Gerais
2013

KÁTIA CIPRIANA PEREIRA SILVA SANTOS
SUCESSO REPRODUTIVO DE Dimorphandra mollis (LEGUMINOSAE:
MIMOSOIDAE): O PAPEL DA INTERAÇÃO FORMIGA-HOMÓPTERA
Dissertação apresentada ao Programa de Pós-Graduação Stricto Sensu em
Ciências Biológicas da Universidade Estadual de Montes Claros
Dissertação apresentada ao Programa de
Pós-Graduação Stricto Sensu em Ciências
Biológicas da Universidade Estadual de
Montes Claros, como requisito necessário
para conclusão do curso de Mestrado em
Ciências Biológicas.
Orientador: Prof. Dr. MARCÍLIO FAGUNDES
Montes Claros – Minas Gerais
2013

KÁTIA CIPRIANA PEREIRA SILVA SANTOS
SUCESSO REPRODUTIVO DE Dimorphandra mollis (LEGUMINOSAE:
MIMOSOIDAE): O PAPEL DA INTERAÇÃO FORMIGA-HOMÓPTERA
Dissertação apresentada ao Programa de
Pós-Graduação Stricto Sensu em Ciências
Biológicas da Universidade Estadual de
Montes Claros, como requisito necessário
para conclusão do curso de Mestrado em
Ciências Biológicas.
em 12 de setembro 2013
Dr. Maurício Lopes de Faria - UNIMONTES
Dr. André Rinaldo Senna Garraffoni - UFVJM
Orientador: Prof. Dr. MARCÍLIO FAGUNDES
Montes Claros – Minas Gerais
2013

A minha avó Adinê
DEDICO

AGRADECIMENTOS
Agradeço primeiramente a Deus, pelo dom da vida e por me dar forças a cada
dia, me fazendo acreditar que o sonho é possível.
Ao amor da minha vida, minha avó Adinê que infelizmente não está mais aqui,
mais que estará sempre presente em meu coração, obrigada por me amar, me ouvir,
cuidar de mim, me ensinar a importância de trabalhar, estudar, correr atrás, obrigada por
me ensinar a ter caráter e nunca passar por cima de ninguém, eu amo você
incondicionamente. Ao meu Pai Arlindo,avô, pai, amigo por me amar, por me criar, por
acretidar sempre em mim, na minha capacidade, por me ensinar os valores que sei hoje.
A minha amada mãe por ter me ensinado o valor do respeito, da dignidade, da
luta e de sempre ser perceverante, me mostrando que mesmo quando tudo parece
impossível é importante não desistir e arriscar. Ao meu pai Welligton que sempre me
acolheu e me ajudou na minha formação com seu carinho e amor. Aos meus irmãos por
me fazerem sorrir e me divertir sempre, recarregando minhas energias.
Obrigada ao meu querido tio e irmão Junior por me ajudar sempre nos
momentos difíceis, por me levar no campo toda semana para coletar, por me salvar de
TODAS as minhas enrrascadas. Aos meus outros tios por sempre estarem presentes em
minha vida.
Gostaria de agradecer também a família Oliveira em especial à Rosangela,
Rogério, Ana Paula e Mark que me acolheram como amiga, filha, participando
intensamente dessa jornada, obrigada pelos conselhos, pelo amor, pelas festinhas do
final de semanas, pelo direcionamento quando eu estava perdida.
É com muita gratidão que eu agradeço a Unimontes, ao PPGCB e ao laboratório
de Biologia da Conservação pelos seis anos que tive a oportunidade de estudar e
conhecer pessoas que levarei para o resto da vida. Muitissímo obrigada ao meu querido
orientador Marcílio Fagundes pelos ensinamentos, pela confiança em mim depositada e
pela oportunidade de desenvolver um trabalho ao seu lado.

Aos meus amigos da estufa os antigos e os atuais, pelas agradáveis horas no
café, pelas discussões, por me ajudarem com as análises, com os conselhos, com uma
palavra amiga de conforto, com nossas saídas divertidissímas. Os meus sinceros
agradecimentos à Luiz Falcão, Dudu, Lukete, Camila (Manu), Malzinha, Renata,
Gramps, Malu, Laurinha, Créu, Leozim, Medina, Serapas, Suellen, Hugo, Patty,
Lorenzo, Paulinho e em especial a Letícia minha grande companheira nesse trabalho,
que encarou esse trabalho com muita seriedade me ajudando em todas as etapas. A
Jacque (guiga) queria agradecer por me ouvir, por me ajudar sempre que preciso, por
salvar meu computador e a dissertação, obrigada por tudo. Gostaria de agradecer
também a toda minha turma do mestrado, vocês foram demais galera!!!
As minhas eternas amigas e companheiras Betty, Polly, Camila, Raissinha nossa
amizade é pra toda vida, obrigado por terem estado ao meu lado durante esses seis anos,
passando por tudo comigo, desde os momentos felizes, das horas no buraco aos
momentos tristes.
Por fim gostaria de agradecer a pessoa que mais me ajudou nessa etapa, obrigada
Mariana por tudo, por virar bióloga algumas vezes, coletando comigo, medindo
sementes. Obrigada por ser amiga quando eu precisei, por ser minha companheira
incondicional me colocando acima de tudo, quando pra mim era impossível ver uma luz
no fim do túnel, obrigada por arrumar minha vida pra que sempre desse tempo de eu
fazer minhas coisas, obrigada pelos fins de semanas, pelas histórias, pelas músicas
repetidas, pelo colo e por sempre me fazer sorrir.
Enfim obrigada a todos que participaram diretamente ou indiretamente dessa
jornada comigo. Amo todos vocês!

Pelo apoio e financiamento:

Resumo
As formigas podem estabelecer uma relação mutualística com os homóptera produtores
honeydew porque as formigas obtêm um recurso alimentar rico em nutrientes e em
contrapartida protegem os homóptera contra o ataque de predadores e parasitóides. A
ação destas formigas também pode estender seus efeitos para a planta hospedeira. As
formigas atuam como defesa biótica para planta afungentando insetos herbívoros. As
plantas geralmente não possuem recursos para investir simultaneamente em todas as
metabólicas, deste modo na presença das formigas, as plantas investem menos na
produção de compostos de defesa, alocando mais recursos para reprodução. Desde
modo, os efeitos dos homópteras, das formigas e da interação homóptera-formiga na
comunidade de insetos herbívoros associados as inflorescências e no sucesso
reprodutivo de Dimorphandra mollis (Fabaceae) foram testados em um experimento de
exclusão. O trabalho foi conduzido em uma área de cerrado sentido restrito, localizada
em Montes Claros, Minas Gerais. Foram marcadas 20 plantas, 3 inflorescências em
cada planta (t1=controle, t2=exclusão de homópteras, t3=exclusão de formigas).
Quatorze espécies de insetos (10 famílias) e seis espécies de formigas (2 famílias) foram
coletadas nas inflorescências de D. mollis. A exclusão do homóptera Membracis foliata
reduziu 270% da população de formigas. A exclusão do homóptera afetou
positivamente a abundância de mastigadores , por outro lado a exclusão das formigas
afetou positivamente a riqueza e abundância de abelhas . Os insetos sugadores não
foram afetados pelos tratamentos de exclusão. As flores sofreram maior ataque dos
herbívoros do que os botões florais. As inflorescências do tratamento que apresentava
interação entre homóptera-formigas (controle) produziram mais frutos, no entanto o
número de sementes não foi afetado pelos tratamento. No tratamento controle foi
observado frutos e sementes maiores, além disso a velocidade e a porcentagem de
germinação das sementes foram positivamente afetadas. As sementes pertencentes aos
tratamentos controle e exclusão de homóptera apresentaram plântulas mais vigorosas.
Portanto, os resultados encontrados nesse trabalho sugere que a interação homóptera-
formiga foi eficaz na redução da herbivoria e afetaram positivamente o sucesso
reprodutivo de D. mollis.
PALAVRAS-CHAVE: Alocação de recursos, Defesa biótica, Interação formiga-planta,
Germinação de sementes, Membracis foliata, Vigor de plântulas.

Sumário
Introdução 1
Metodologia 5
O Sistema Estudado…………………………………………………………………………...5
Área de Estudo……………………………………………………………………………….....6
Trabalho de campo……………………………………………………………………………...6
Fauna de insetos associados a inflorescência de Dimorphandra mollis………………..7
Produção de frutos e sementes em função dos tratamentos de exclusão………………..8
Germinação das sementes em função dos tratamentos de exclusão……………………...9
Resultados 11
Assembléia de formigas associada a inflorescência de Dimorphandra mollis ………11
Assembléia de insetos herbívoros associados a inflorescência de Dimorphandra
Mollis……………………………………………………………………………………………..12
Efeitos dos tratamentos de exclusão no tamanho e número de frutos…………………...18
Efeito dos tratamentos de exclusão no número e tamanho das sementes……………….20
Germinação de sementes e vigor de plântulas……………………………………………...22
Discussão 26
Referências 31

1
Introdução
As formigas (Hymenoptera: Formicidae) são os animais mais abundantes na
maioria dos ecossistemas terrestres (Bolton et al. 2007). As formigas desenvolveram
várias interações com diferentes grupos de animais e plantas que variam de negativas a
positivas (Fernandes et al. 1999). Por exemplo, algumas espécies de formigas podem
cultivar e se alimentar de fungos (Quilan & Cherrett 1977), enquanto outras são
predadoras de artrópodes e pequenos vertebrados (Hölldobler & Wilson 1990). As
formigas também podem se alimentar de fluidos açucarados secretados por nectários
extraflorais (Wäckers 2001; Lach 2005; Wäckers et al. 2008; Queiroz et al. 2013),
galhas (Fernandes et al.1999) ou do honeydew produzidos por borboletas, pulgões e
membracídeos (Buckley 1987; Del-Claro & Oliveira 1999; Oliveira & Del-Claro 2005;
Wäckers et al. 2008). O Honeydew é uma substância açucarada rica em açúcares,
aminoácidos, aminas e proteínas que representa uma fonte importante de alimento para
várias espécies de formigas (Auclair 1963; Mattson 1980).
As formigas podem estabelecer uma relação mutualística com os homóptera
produtores honeydew porque as formigas obtêm um recurso alimentar rico em
nutrientes e em contrapartida protegem os homóptera contra o ataque de predadores e
parasitóides (Buckley 1987; Cushman & Beattie 1991). A ação destas formigas também
pode estender seus efeitos para a planta hospedeira. De fato, as plantas podem se
beneficiar da interação formiga-homóptera porque as formigas que ordenham os afídeos
agem como defesa biótica, expulsando ou predando outros herbívoros presentes nas
plantas (Del-Claro & Oliveira 1999; Del-Claro & Oliveira 2000; Coley et al. 2005;
Fernandes et al. 2005, Fagundes et al. 2005; Queiroz et al. 2013). Além disto, alguns

2
estudos também sugerem que, na presença das formigas, as plantas investem menos
recurso na produção de compostos de defesa antiherbívoros (Whaterman et al. 1984).
Neste caso, estes recursos podem ser usados diretamente no desenvolvimento vegetativo
ou reprodutivo, aumentando o fitness da planta.
A interação planta-homóptera-formiga pode assumir um caráter mais complexo.
Por exemplo, se as formigas forem excluídas do sistema, a população de homóptera
pode aumentar (as formigas regulam a população dos homópteras eliminando
indivíduos de ramos altamente infestados) e afetar diretamente o desenvolvimento da
planta devido a elevada exploração da seiva (Fagundes et al. 2005). Além disto, quando
as formigas não ordenham os homópteras, ocorre um acúmulo de honeydew nos ramos
da planta que favorece o desenvolvimento de fungos e bactérias que prejudicam o
desenvolvimento vegetativo da planta (Neves et al. 2011). Contudo, este aspecto da
interação planta-homóptera-formiga ainda é pouco discutido na literatura.
Enquanto a maioria dos insetos herbívoros se alimentam de folhas (Coley &
Kursar 1996; Koptur et al. 2002; Queiroz et al. 2013), outros podem se alimentar de
partes reprodutivas como flores, frutos e sementes (Burgess 1991; Mccall & Irwin
2006). Os ataques podem ocorrer desde os botões florais até flores maduras (Burgess
1991; Fernandes et al. 2005). As flores apresentam vários atrativos como cores diversas,
produção de pólen e néctar para que funcionam como atração para polinizadores (Bawa
1990; Brugess 1991) mais podem também atrair outros insetos herbívoros (Fernandes et
al. 2005; Maccall & Irwin 2006; Raven et al. 2007).
Os homópteras produtores de honeydew normalmente formam colônias densas
em locais de intenso crescimento, especialmente próximos às inflorescências das plantas
(Fagundes et. al 2005; Del-Claro & Oliveira 1999; Neves et al. 2011). Neste caso, as
formigas que ordenham estes homópteras também patrulham as estruturas reprodutivas

3
das plantas e potencialmente podem proteger estes órgãos contra outros herbívoros (veja
Fernandes et al. 2005). Por outro lado, as formigas também podem afugentar possíveis
polinizadores, afetando negativamente a produção de sementes pelas plantas (Oliveira et
al. 1999). Neste cenário, o resultado da interação planta-homóptera também depende do
comportamento das formigas. Por exemplo, na ausência de formigas ocorre um aumento
na população de homópteras que afeta negativamente o desenvolvimento de frutos e
sementes pela planta (Buckley 1987). Por outro lado, quando as formigas estão
presentes, as plantas investem menos recursos na produção de compostos de defesa e os
recursos podem ser alocados para a produção de frutos e sementes (Whaterman et al.
1984). Novamente, estes aspectos da interação planta-homóptera-formiga ainda são
pobremente enfocados.
O tamanho das sementes (biomassa) afeta o tempo (Murali 1997), a
percentagem de germinação (Mölken 2005) e o vigor das plântulas (Yanlonget al.
2007). Sementes pequenas germinam mais rapidamente, sendo favorecidas em
ambientes transitórios (Baskin & Baskin 1998). Sementes grandes germinam mais
lentamente, mas normalmente apresentam maior percentagem de germinação, sendo
favorecidas em ambientes previsíveis (Geritz 1995; Ferreira & Borghetti 2004). Além
disto, sementes grandes produzem plântulas mais vigorosas e resistentes a injúrias
porque estas sementes acumulam maior quantidade de reservas (Yanlong et al. 2007).
Assim, seria esperado que variações nas caracterísitcas da interação planta-homóptera-
formiga afetem o sucesso reprodutivo das plantas.
Apesar de alguns estudos demonstrarem a importância das formigas na defesa de
inflorescências (Fagundes et al. 2005; Fernandes et al. 2005; Neves et al. 2011),
poucos trabalhos relatam e o efeito isolado do pulgão nas plantas (Fagundes et al. 2005;

4
Neves et al. 2011) e o efeito da interação homóptera-formiga no sucesso reprodutivo
das plantas (Messina 1981; Rico-Gray & Castro 1996).
Este estudo foca no papel da interação formiga-homóptera no sucesso
reprodutivo da planta hospedeira Dimorphandra mollis. O sistema é formado pela
planta D. mollis, o homóptera Membracis foliata e as formigas que ordenham os
homópteras. Assim, os objetivos deste trabalho foram avaliar a influência da interação
homóptera-formiga na defesa das inflorescências contra herbívoros e no sucesso
reprodutivo de Dimorphandra mollis, testando as seguintes hipóteses: (i) a presença de
homóptera afeta a atração de formigas para as inflorescências; (ii) a associação
homóptera-formiga confere as inflorescências defesa biótica contra insetos herbívoros e
(iii) a interação homóptera-formiga influencia no sucesso reprodutivo de D. mollis.

5
Metodologia
O Sistema Estudado
Dimorphandra mollis Benth. popularmente conhecida como fava d’anta, faveira,
favinha, falso barbatimão (Lorenzi 1992), a espécie pode ser encontrada em Minas
Gerais, Mato Grosso do Sul, Pará e São Paulo (Lorenzi 1992), Amazonas, Bahia,
Distrito Federal, Goiás, Maranhão, Piauí e Tocantins (Almeida et al. 1998). D. mollis é
importante na flora do cerrado brasileiro (Chaves & Usberti 2003), visto que a planta
serve de alimento para alguns vertebrados e invertebrados (Bizerril et al. 2005).
Contudo, D.mollis é considerada tóxica para várias espécies de Artropoda (Cintra et al.
2002), pois a planta produz flavonóides que minimizam a herbivoria (Costa et al. 2000).
As plantas apresentam homópteras associados com suas inflorescências que atraem
formigas. D. mollis apresenta inflorescências (500 flores) em espigas terminais com
flores hermafroditas 5-pétalas livres, 5-estames, ovário súpero, unilocular, com muitos
óvulos (Almeida et al.1998). As flores de D. mollis são polinizadas principalmente por
abelhas, mas suas flores podem ainda atrair outros polinizadores e insetos devido aos
flavonóides que podem atuar como sinais atrativos (Costa et al. 2000).
As plantas atingem de 4-8 metros de altura e diâmetro do tronco de 30-50
centímetros (Lorenzi 1992). A floração ocorre de setembro a novembro, com picos no
mês de outubro (Caldeira Junior et al. 2008). As inflorescências apresentam pequenos
botões florais e diversas flores amarelas organizadas em cachos. Os seus frutos crescem
em cachos e amadurecem na estação seca, geralmente de junho a agosto (Bizerril et al.
2005). O fruto é um legume semindeiscente, que se torna negro e rígido com o
amadurecimento (Bizerril et al. 2005). As sementes possuem tegumento rígido, que

6
dificulta a absorção de água pelas sementes, apresentando deste modo dormência física
(Rizzini 1965).
Área de Estudo
O presente estudo foi desenvolvido em uma área de cerrado strictu sensu
localizada na região norte da cidade de Montes Claros, Minas Gerais (16º 40'26" S e 43º
48'44" W). A área de estudo possui solo argiloso oligotrófico (Fagundes et al. 2011) e
estrato herbáceo-subarbustivo pouco desenvolvido que frequentemente é afetado pela
ação do fogo (Costa et al. 2011). Fisionomicamente, a região localiza-se entre os
domínios do Cerrado e da Caatinga (Rizzini 1997), apresentando clima semi-árido com
estações seca e chuvosa bem definidas. A temperatura média anual da região é de
aproximadamente 23°C e a média de precipitação é de cerca de 1.000 mm/ano,
concentrada principalmente nos meses de outubro a janeiro (Costa et al. 2011).
Trabalho de campo
Um total de vinte indivíduos de D. mollis que iniciavam a fase reprodutiva
(indivíduos em fase inicial de formação de botões florais) foi selecionado e marcado, na
área de estudo, durante o mês de outubro de 2011. Todos os indíviduos selecionados
apresentavam CAP (circunferência à altura do peito) ≥ 10 cm e altura entre dois e cinco
metros. Estas plantas distavam-se de pelo menos 50m e estavam distribuídas
aleatoriamente em uma área aproximada de cinco hectares.
Em cada um dos indivíduos marcados foram selecionadas três inflorescências
com flores em estágio de pré-antese e divididas em três tratamentos: (t1 = controle)
inflorescência com acesso livre para formigas e homópteras, (t2 = exclusão de formigas)
inflorescência com homópteras e sem acesso para formigas e (t3 = exclusão de
homópteras) inflorescência sem homópteras e com livre acesso para formigas. As

7
formigas foram excluídas do tratamento t2 através da aplicação de uma resina pegajosa
não tóxica (tanglefoot, Empresa uísque, Michigan, EUA) próxima a base da
inflorescência. Os homópteras foram excluídos do tratamento t2 manualmente com o
auxílio de um pincel. Os ramos que se encontravam próximos as inflorescências dos
tratamentos t2 e t3 foram cortados para evitar o acesso de formigas ou de homópteras,
respectivamente. Estas inflorescências foram monitoradas semanalmente durante todo o
experimento para se assegurar da eficiência dos tratamentos.
Fauna de insetos associados a inflorescência de Dimorphandra mollis
Para se avaliar os efeitos dos tratamentos de exclusão na fauna de insetos
associados a inflorescências de D. mollis cada inflorescência marcada foi monitorada
durante cinco minutos durante 10 semanas consecutivas, no período de outubro 2011 a
dezembro de 2012. Assim, todos os insetos presentes nas inflorescências (excluindo-se
membracis foliata) foram coletados e levados para o Laboratório de biologia da
Conservação para triagem e identificação. As amostragens foram realizadas no período
diurno entre 07h00min e 11h00min.
A análise dos dados foi realizada com o auxílio do software R versão 2.15.0 (R
Development Core Team 2011). Para testar o efeito da presença de homóptera na
riqueza e abundância de formigas, o efeito da fenologia e do tratamento na riqueza e
abundância dos insetos herbívoros e abelhas, foram construídos modelos lineares de
efeito misto (LME), onde a riqueza e a abundância de formigas, insetos herbívoros e
abelhas foram utilizados como variável resposta, enquanto o tratamento e a fenologia
foram utilizados como variáveis explicativas. Como os dados foram coletados nos
mesmos indivíduos ao longo da amostragem violando o pressuposto da independência
das amostras. Os modelos lineares de efeito misto são recomendados para minimizar os

8
efeitos da pseudorepetição temporal. Deste modo, com a utilização do LME a variável
resposta não representa uma medida individual, mas uma sequência de medidas em um
mesmo indivíduo durante a amostragem (Crawley 2007).
Produção de frutos e sementes em função dos tratamentos de exclusão
Para avaliar os efeitos dos tratamentos de exclusão na produção e
desenvolvimento dos frutos de D. mollis todas as inflorescências marcadas foram
monitoradas até o período de maturação dos frutos (julho 2012). Assim, o número
inicial de frutos produzidos em cada inflorescência foi determinado no final de janeiro
de 2012 através de contagem direta dos frutos imaturos. No início do mês de julho os
frutos maduros presentes em cada inflorescência foram contados novamente, coletados
e encaminhados ao laboratório de biologia da conservação para triagem. Assim, obteve-
se dados de comprimento, largura, espessura de cada fruto com uso de um paquímetro
digital.
Após estas medições os frutos foram quebrados manualmente para obtenção das
sementes. Todas as sementes foram medidas com um paquímetro digital, obtendo-se
dados biométricos de comprimento, largura e espessura de cada semente.
Posteriormente a retirada das sementes a material vegetal dos frutos foi acondicionado
individualmente em sacos plásticos para secagem em estufa a 70°C durante 72 horas
para obtenção do peso seco dos frutos.
Foram construídos modelos lineares generalizados (GLM) para avaliar a
influência do tratamento no sucesso reprodutivo de D. mollis. O número de frutos
produzidos, número de frutos maduros, número de sementes total, o tamanho dos frutos
(comprimento, largura e espessura) e o peso foram utilizados como variável resposta,

9
enquanto os tratamentos foram utilizados como variáveis explicativas. Para avaliar o
vigor das sementes em função do tratamento foram contruídos GLMs onde o tamanho
das sementes (comprimento, largura e espessura) e o peso foram utilizados como
variável resposta e os tratamento utilizados como variável explicativa. Os modelos
construídos foram utilizados com uma distribuição de erros adequada para cada variável
resposta, segundo a crítica do modelo (Crawley 2007).
Germinação das sementes em função dos tratamentos de exclusão
Um total de 256 sementes (t1: n = 110, t2: n = 110 e t3 n = 36) foram usadas
para avaliar os efeitos dos tratamentos de exclusão na germinabilidade das sementes de
D. mollis. Assim, estas sementes foram semeadas individualmente em bandejas de
germinação contendo vermiculita como substrato a aproximadamente 01cm de
profundidade.
Após a semeadura as bandejas foram acondicionadas em câmara de germinação
com fotoperíodo e temperatura controlados (12h/luz a 35°C e 12h/escuro a 30°C). Cada
semente foi irrigada diariamente com 1,5 mL de água diariamente como forma de se
manter a capacidade de campo do substrato. As sementes foram monitoradas
diariamente para determinar o percentual e a velocidade de germinação em cada
tratamento. Considerou-se germinada as sementes que emergiram do substrato expondo
os codilédones.
As plântulas que emergiram foram mantidas na câmara de germinação até a
queda dos cotilédones para se avaliar o vigor das plântulas. Após este período as
plântulas foram retiradas do substrato para se determinar o comprimento da raiz e da
parte aérea. Estas medidas foram realizadas com auxílio do paquímetro digital.
Finalmente, a parte aérea foi separada do sistema radicular de todas as plântulas. Este

10
material foi acondicionado em sacos de papel e secos em estufa a 70°C durante 72 horas
para obtenção do peso seco das plantulas.
Para avaliar o tempo de emergência entre os tratamentos foi utilizada a análise
de sobrevivência, visto que, o experimento apresentava pseudo-repetição temporal.
Foram consideradas como variável resposta a porcentagem de germinação entre
tratamentos e o tempo germinação como variável explicativa. Para avaliar a influência
do tratamento na porcentagem de germinação e no vigor das plantas foram contruídos
modelos lineares generalizados (GLM). A porcentagem de germinação, o comprimento
e o peso seco da parte áerea e do sistema radicular foram utilizados como variável
resposta e os tratamentos como variável explicativa. Todos os modelos foram
construídos utilizando a distribuição de erro adequada para cada variável resposta,
considerando a crítica do modelo (Crawley 2007). Cada modelo criado foi comparado
com o modelo nulo, o modelo mínimo adequado foi criado com a omissão dos termos
que não foram significativos. A adequação dos modelos foi testada a partir da análise de
resíduos

11
Resultados
Assembléia de formigas associada a inflorescência de Dimorphandra mollis
Foram coletados um total de 459 formicídeos pertencentes às subfamílias Formicinae e
Ectatomminae. As espécies mais abundantes foram Camponotus blandus (39,43%) e
Camponotus arbóreos (33,99%), enquanto Brachymyrmex sp. (0,22%) foi a menos
abundante no estudo (Tab. 1).
Tabela 1. Abundância de formicídeos amostrados nas inflorescências de Dimorphandra
mollis no período de Setembro à Novembro de 2012, Montes Claros, Minas Gerais.
Subfamília Espécies Abundância (%)
Formicinae Brachymyrmex sp. 1 0,22
Camponotus arboreos 156 33,99
Camponotus blandus 181 39,43
Camponotus crassus 58 12,64
Camponotus triceps 38 8,28
Ectatomminae Ectatoma turbeculato 25 5,44
Total 6 459
A riqueza dos formicídeos não variou em função da presença ou ausência de
Membracis foliata. Contudo, a abundância de formigas foi aproximadamente 270,16%
menor nos ramos onde os homópteras foram removidos (Tab. 2; Fig. 1).

12
Tabela 2 Análise de deviance dos modelos completos para avaliar o efeito dos
tratamentos de exclusão nas variáveis resposta (riqueza e abundância de formigas).
Variável Resposta Variável
Explicativa
F Distribuição
de erro
G.L G.L.R. P
Riqueza de formigas Tratamento 2.3207 Poisson 3 17 0.127
Abundância de formigas
Tratamento 65.268 Poisson 3 17 <0.001
Fig.1 Riqueza (A) e abundância (B) média de formigas nos tratamentos controle (presença de
homóptera e formigas) e exclusão de homóptera (presença apenas de formigas) em
inflorescências de Dimorphandra mollis. Barras verticais = erros padrão.
Assembléia de insetos herbívoros associados a inflorescência de Dimorphandra
mollis
Um total de 241 insetos herbívoros (excluindo Membracis foliata), distribuídas
em quatorze morfoespécies, dez famílias e três guildas alimentares foram observados
nas inflorescências de Dimorphandra mollis. A maioria das espécies de herbívoros
apresentou abundância baixa (Tab. 3). Contudo, deve-se destacar a elevada abundância
relativa de Pelidnota sp. (61%). Este inseto alimenta-se no interior da inflorescência
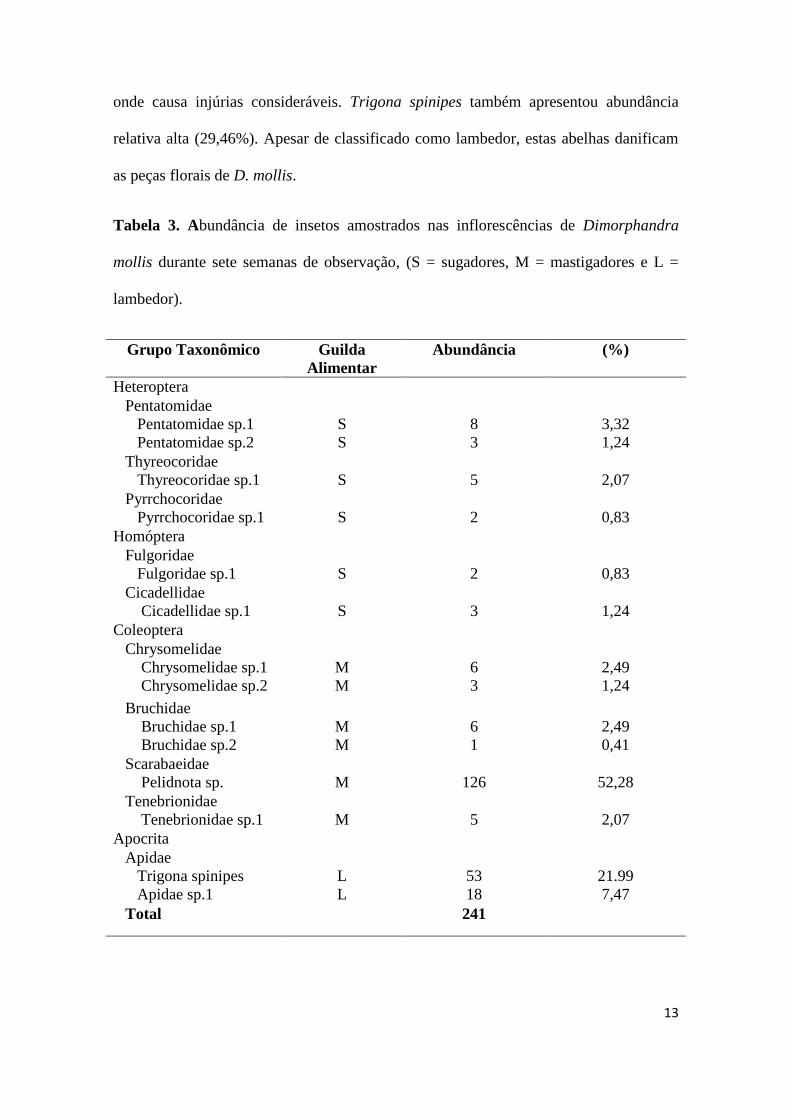
13
onde causa injúrias consideráveis. Trigona spinipes também apresentou abundância
relativa alta (29,46%). Apesar de classificado como lambedor, estas abelhas danificam
as peças florais de D. mollis.
Tabela 3. Abundância de insetos amostrados nas inflorescências de Dimorphandra
mollis durante sete semanas de observação, (S = sugadores, M = mastigadores e L =
lambedor).
Grupo Taxonômico Guilda
Alimentar
Abundância (%)
Heteroptera
Pentatomidae
Pentatomidae sp.1
Pentatomidae sp.2
S
S
8
3
3,32
1,24
Thyreocoridae
Thyreocoridae sp.1
S
5
2,07
Pyrrchocoridae
Pyrrchocoridae sp.1
S
2
0,83
Homóptera
Fulgoridae
Fulgoridae sp.1
S
2
0,83
Cicadellidae
Cicadellidae sp.1
S
3
1,24
Coleoptera
Chrysomelidae
Chrysomelidae sp.1
Chrysomelidae sp.2
M
M
6
3
2,49
1,24
Bruchidae
Bruchidae sp.1
Bruchidae sp.2
M
M
6
1
2,49
0,41
Scarabaeidae
Pelidnota sp.
M
126
52,28
Tenebrionidae
Tenebrionidae sp.1
M
5
2,07
Apocrita
Apidae
Trigona spinipes
Apidae sp.1
L
L
53
18
21.99
7,47
Total 241

14
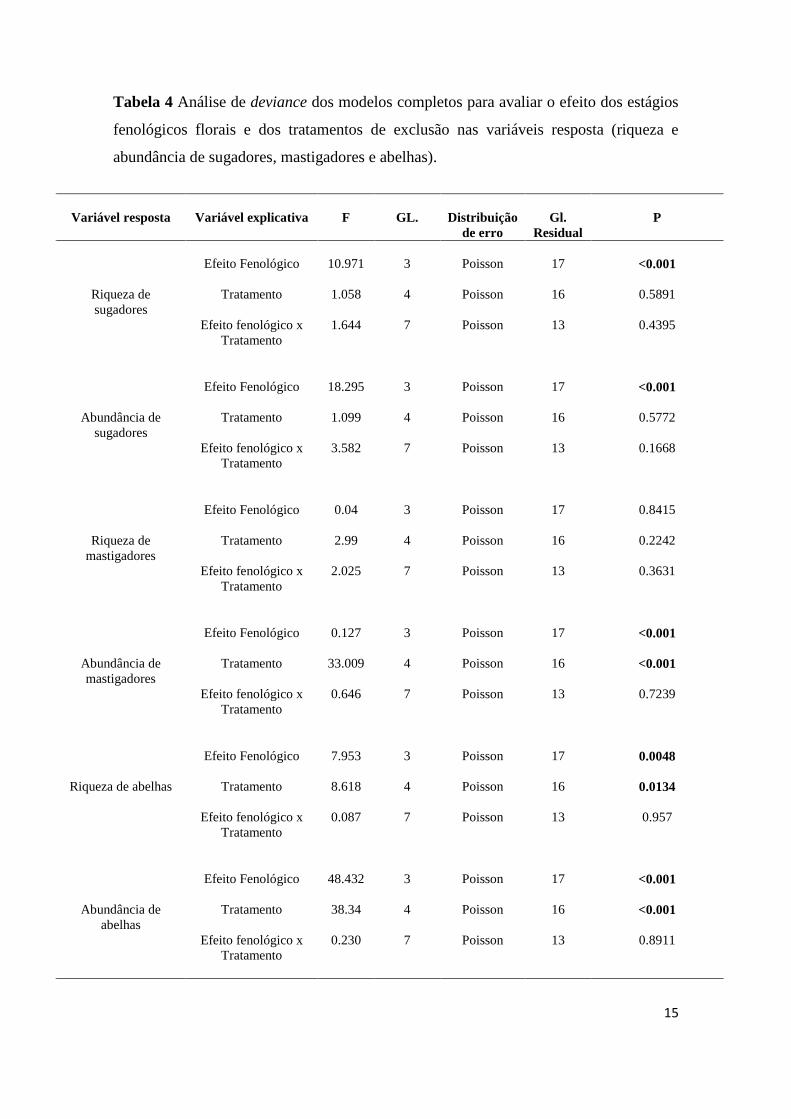
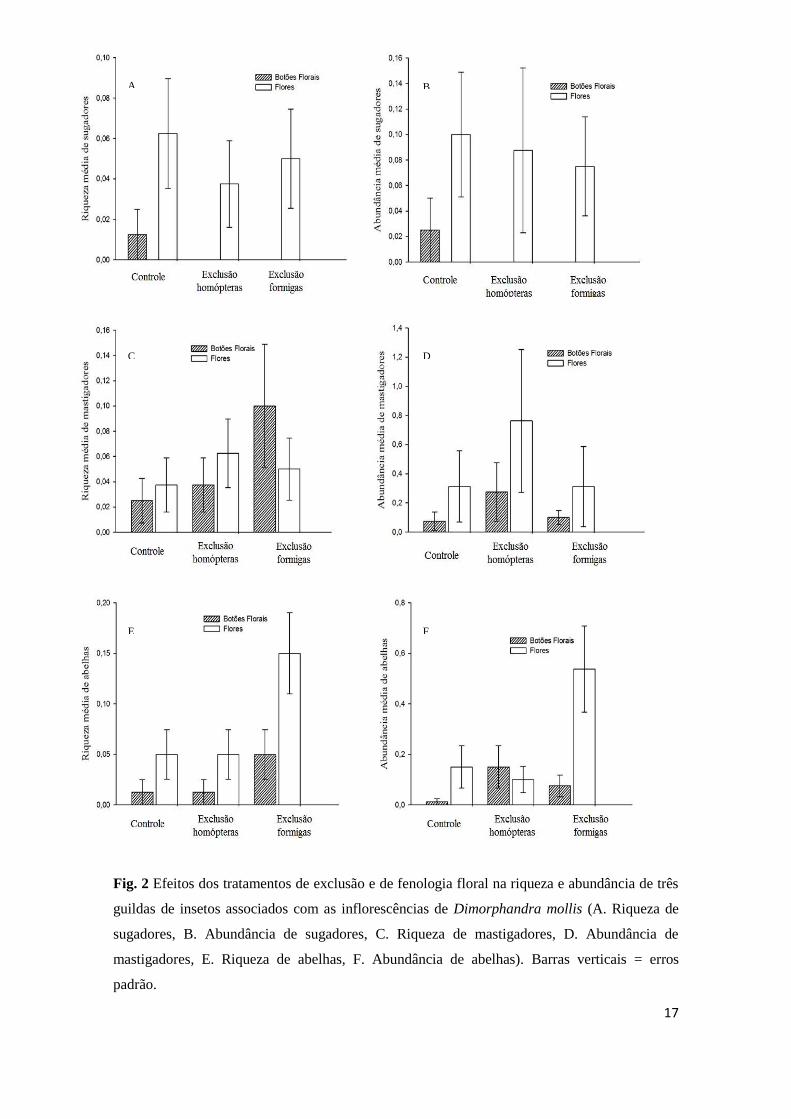
A riqueza e a abundância de insetos sugadores variaram entre os estágios
fenológicos das inflorescênciais de D. mollis. De fato, observou maior diversidade de
insetos sugadores nas inflorescências com flores em ântese. Contudo, os tratamentos de
exclusão e a interação tratamento x fenologia floral não afetaram a diversidade de
insetos sugadores (Tab. 4;Fig. 2a, b).
A riqueza de insetos mastigadores não foi afetada pelos tratamentos de exclusão,
pela fenologia floral ou pela interação tratamento x fenologia (Tab. 4). Contudo a
abundância de mastigadores foi maior nas inflorescências em antese e no tratamento de
exclusão de homóptera (Tab. 4;Fig. 2d ).
A riqueza e a abundância de abelhas responderam similarmente aos tratamentos.
Assim observou-se que à interação tratamento x fenologia floral não afetou a riqueza e a
abundância de abelhas (Tab. 4). Contudo, a riqueza e abundância de abelhas foram
maiores nas inflorescências em antese e no tratamento de exclusão de formigas (Tab. 4;
Fig. 2e, f).
15
Tabela 4 Análise de deviance dos modelos completos para avaliar o efeito dos estágios
fenológicos florais e dos tratamentos de exclusão nas variáveis resposta (riqueza e
abundância de sugadores, mastigadores e abelhas).
Variável resposta
Variável explicativa
F
GL.
Distribuição
de erro
Gl.
Residual
P
Riqueza de
sugadores
Efeito Fenológico
Tratamento
Efeito fenológico x
Tratamento
10.971
1.058
1.644
3
4
7
Poisson
Poisson
Poisson
17
16
13
<0.001
0.5891
0.4395
Abundância de
sugadores
Efeito Fenológico
Tratamento
Efeito fenológico x
Tratamento
18.295
1.099
3.582
3
4
7
Poisson
Poisson
Poisson
17
16
13
<0.001
0.5772
0.1668
Riqueza de
mastigadores
Efeito Fenológico
Tratamento
Efeito fenológico x
Tratamento
0.04
2.99
2.025
3
4
7
Poisson
Poisson
Poisson
17
16
13
0.8415
0.2242
0.3631
Abundância de
mastigadores
Efeito Fenológico
Tratamento
Efeito fenológico x
Tratamento
0.127
33.009
0.646
3
4
7
Poisson
Poisson
Poisson
17
16
13
<0.001
<0.001
0.7239
Riqueza de abelhas
Efeito Fenológico
Tratamento
Efeito fenológico x
Tratamento
7.953
8.618
0.087
3
4
7
Poisson
Poisson
Poisson
17
16
13
0.0048
0.0134
0.957
Abundância de
abelhas
Efeito Fenológico
Tratamento
Efeito fenológico x
Tratamento
48.432
38.34
0.230
3
4
7
Poisson
Poisson
Poisson
17
16
13
<0.001
<0.001
0.8911

16
A riqueza e a abundância de abelhas não diferiu entre os tratamentos controle e exclusão
de homóptera, sendo maior no tratamento de exclusão de formigas (Fig. 2).
17
Fig. 2 Efeitos dos tratamentos de exclusão e de fenologia floral na riqueza e abundância de três
guildas de insetos associados com as inflorescências de Dimorphandra mollis (A. Riqueza de
sugadores, B. Abundância de sugadores, C. Riqueza de mastigadores, D. Abundância de
mastigadores, E. Riqueza de abelhas, F. Abundância de abelhas). Barras verticais = erros
padrão.
A
C
B
D
E F
18
Efeitos dos tratamentos de exclusão no tamanho e número de frutos
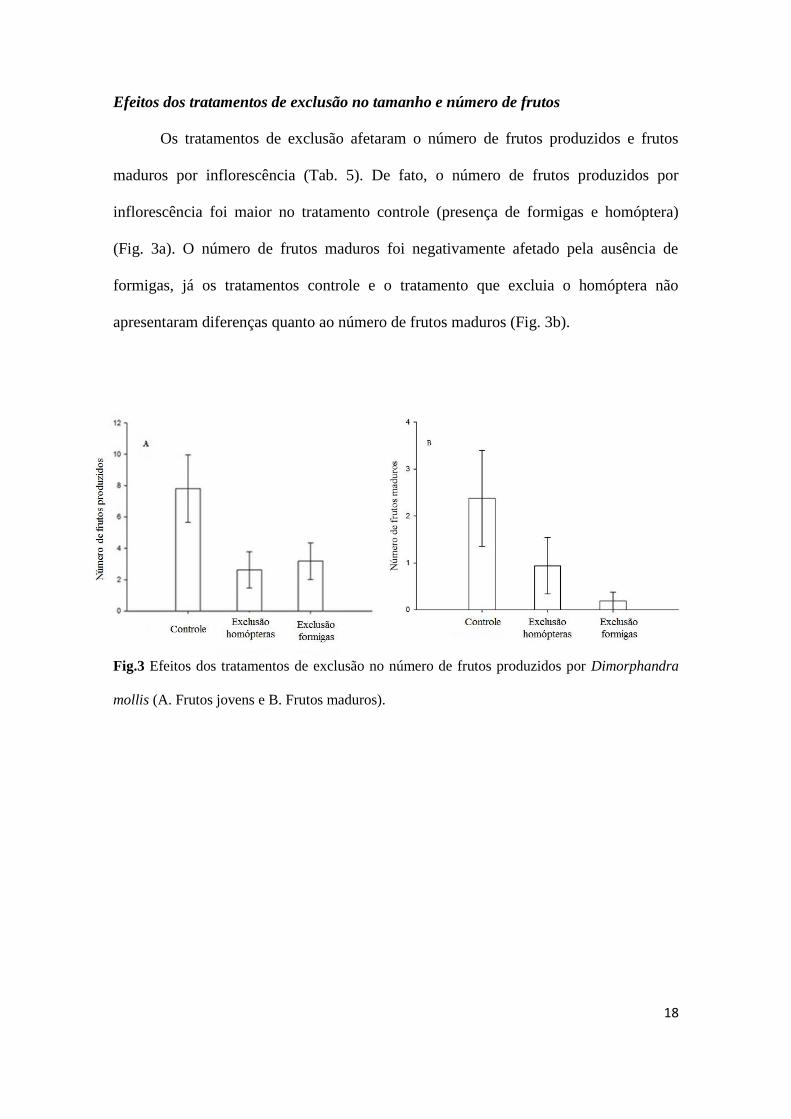
Os tratamentos de exclusão afetaram o número de frutos produzidos e frutos
maduros por inflorescência (Tab. 5). De fato, o número de frutos produzidos por
inflorescência foi maior no tratamento controle (presença de formigas e homóptera)
(Fig. 3a). O número de frutos maduros foi negativamente afetado pela ausência de
formigas, já os tratamentos controle e o tratamento que excluia o homóptera não
apresentaram diferenças quanto ao número de frutos maduros (Fig. 3b).
Fig.3 Efeitos dos tratamentos de exclusão no número de frutos produzidos por Dimorphandra
mollis (A. Frutos jovens e B. Frutos maduros).
19
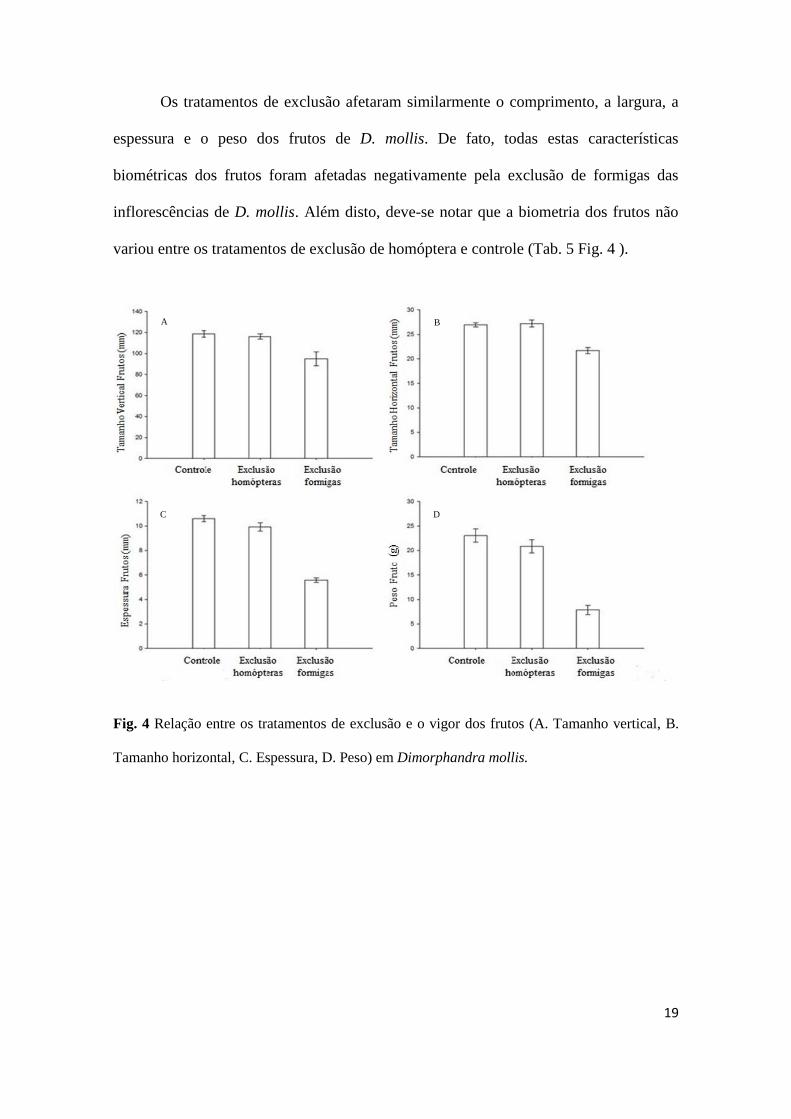
Os tratamentos de exclusão afetaram similarmente o comprimento, a largura, a
espessura e o peso dos frutos de D. mollis. De fato, todas estas características
biométricas dos frutos foram afetadas negativamente pela exclusão de formigas das
inflorescências de D. mollis. Além disto, deve-se notar que a biometria dos frutos não
variou entre os tratamentos de exclusão de homóptera e controle (Tab. 5 Fig. 4 ).
Fig. 4 Relação entre os tratamentos de exclusão e o vigor dos frutos (A. Tamanho vertical, B.
Tamanho horizontal, C. Espessura, D. Peso) em Dimorphandra mollis.
A B
C D

20
Tabela 5 Análise de deviance dos modelos completos para avaliar o efeito dos tratamentos de
exclusão nas variáveis resposta (tamanho vertical, horizontal, espessura, peso dos frutos,
número de frutos produzidos, número de frutos maduros).
Efeito dos tratamentos de exclusão no número e tamanho das sementes
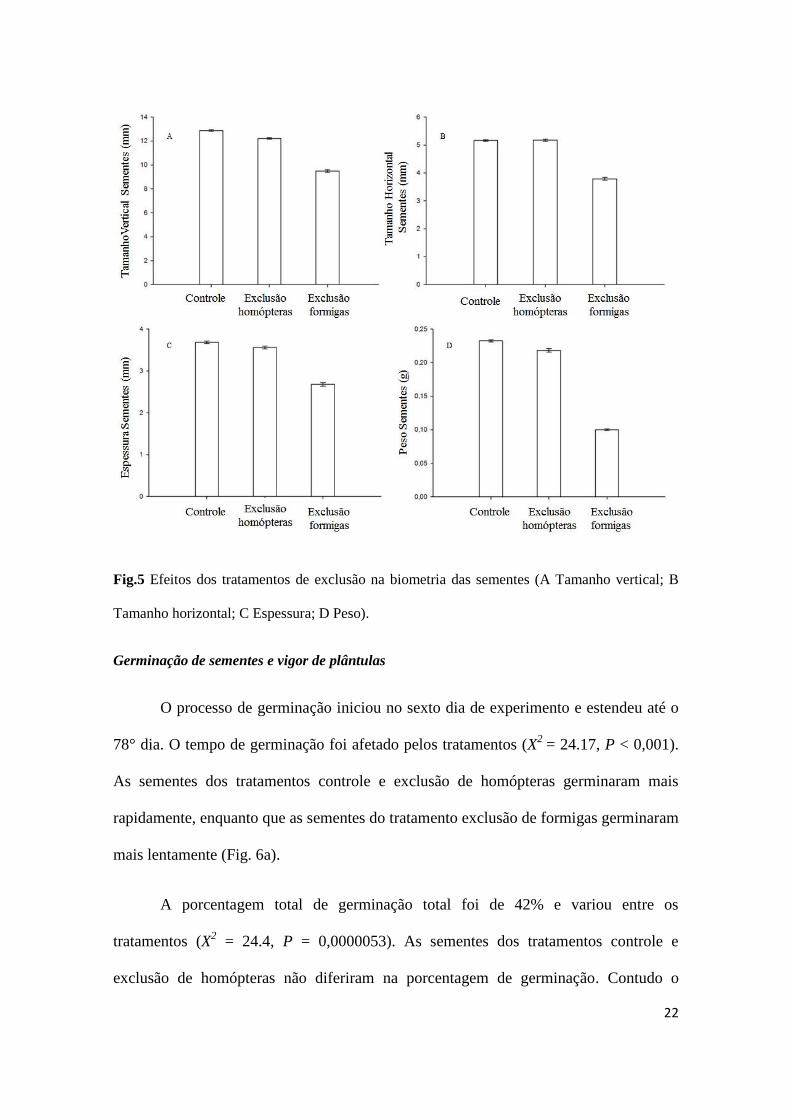
Os tratamentos de exclusão não afetaram o número de sementes por
inflorescência (Tab. 6). Contudo, o comprimento (Fig. 5a), a largura (Fig. 5b), a
espessura (Fig. 5c) e o peso das sementes variaram entre os tratamentos de exclusão
(Fig. 5d ). Além disto, a análise de contraste mostrou que as sementes do tratamento
exclusão de formigas apresentaram menores dimensões e peso comparativamente aos
demais tratamentos (Fig. 5).
Variável resposta
Variável
explicativa
F
Distribuição
de Erro
G.l.
Deviance
Gl.
Residual
Deviance
Residual
P
Tamanho vertical
frutos
Tratamento
3.0499
Gaussian
2
1558.8
49
12521
0.056
Tamanho
horizontal frutos
Tratamento
6.2481
Gaussian
2
80.344
49
315.04
0.004
Espessura frutos
Tratamento
17.424
Gaussian
2
70.41
49
99.003
<0.001
Peso frutos
Tratamento
6.2454
Gaussian
2
645.37
49
2531.7
0.004
Número de frutos
produzidos
Tratamento
53.444
Quasipoisson
2
53.444
45
306.64
0.038
Número de frutos
maduros
Tratamento
36.495
Quasipoisson
2
36.495
45
154.96
0.034
21
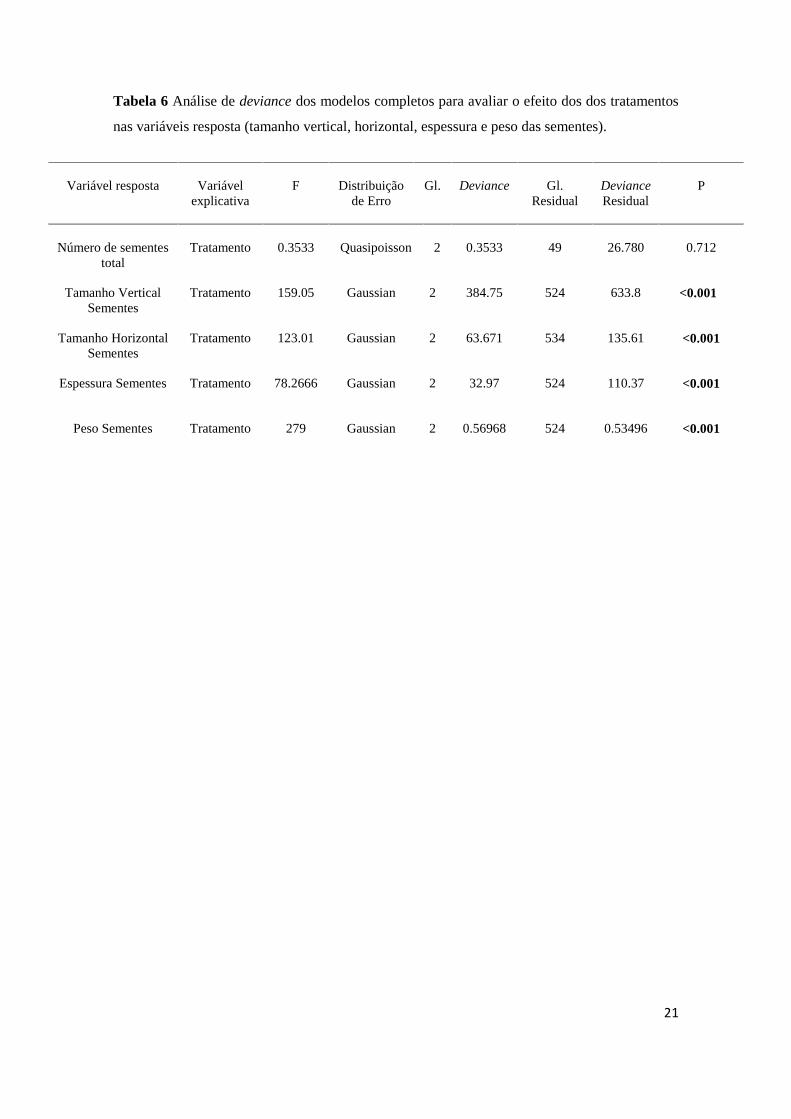
Tabela 6 Análise de deviance dos modelos completos para avaliar o efeito dos dos tratamentos
nas variáveis resposta (tamanho vertical, horizontal, espessura e peso das sementes).
Variável resposta
Variável
explicativa
F
Distribuição
de Erro
Gl.
Deviance
Gl.
Residual
Deviance
Residual
P
Número de sementes
total
Tratamento
0.3533
Quasipoisson
2
0.3533
49
26.780
0.712
Tamanho Vertical
Sementes
Tratamento
159.05
Gaussian
2
384.75
524
633.8
<0.001
Tamanho Horizontal
Sementes
Tratamento
123.01
Gaussian
2
63.671
534
135.61
<0.001
Espessura Sementes
Tratamento
78.2666
Gaussian
2
32.97
524
110.37
<0.001
Peso Sementes
Tratamento
279
Gaussian
2
0.56968
524
0.53496
<0.001
22
Fig.5 Efeitos dos tratamentos de exclusão na biometria das sementes (A Tamanho vertical; B
Tamanho horizontal; C Espessura; D Peso).
Germinação de sementes e vigor de plântulas
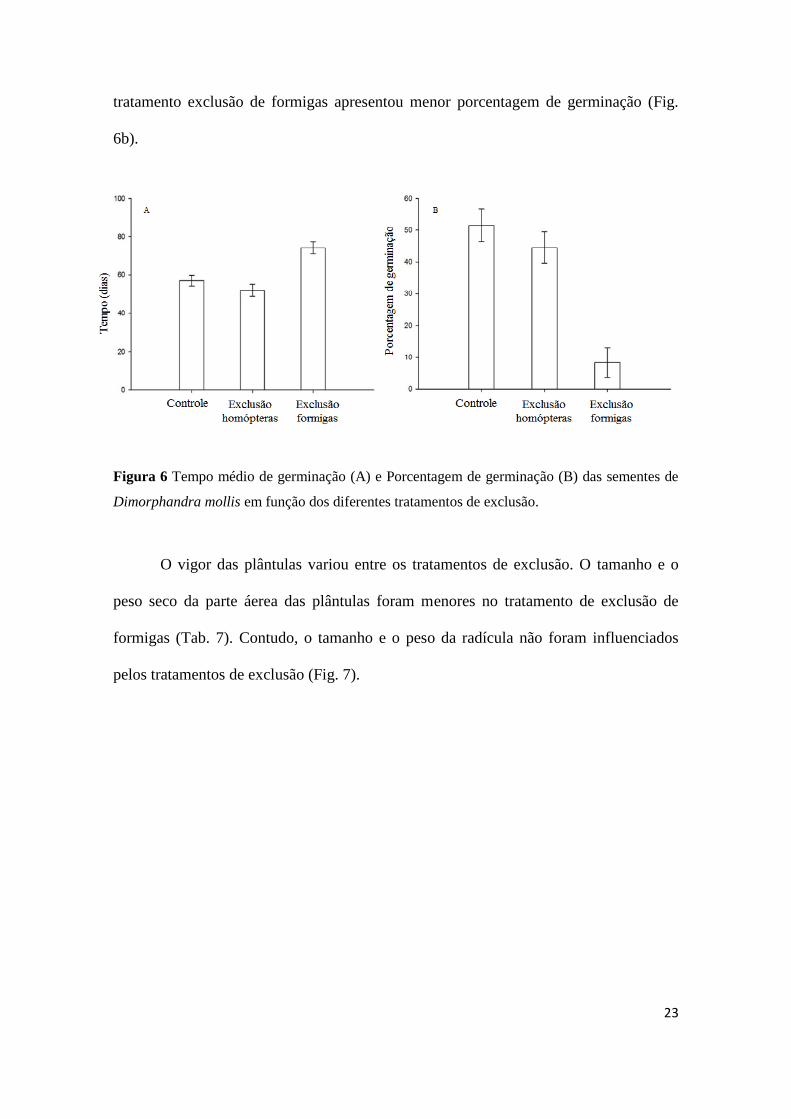
O processo de germinação iniciou no sexto dia de experimento e estendeu até o
78° dia. O tempo de germinação foi afetado pelos tratamentos (X2
= 24.17, P < 0,001).
As sementes dos tratamentos controle e exclusão de homópteras germinaram mais
rapidamente, enquanto que as sementes do tratamento exclusão de formigas germinaram
mais lentamente (Fig. 6a).
A porcentagem total de germinação total foi de 42% e variou entre os
tratamentos (X2 = 24.4, P = 0,0000053). As sementes dos tratamentos controle e
exclusão de homópteras não diferiram na porcentagem de germinação. Contudo o
23
tratamento exclusão de formigas apresentou menor porcentagem de germinação (Fig.
6b).
Figura 6 Tempo médio de germinação (A) e Porcentagem de germinação (B) das sementes de
Dimorphandra mollis em função dos diferentes tratamentos de exclusão.
O vigor das plântulas variou entre os tratamentos de exclusão. O tamanho e o
peso seco da parte áerea das plântulas foram menores no tratamento de exclusão de
formigas (Tab. 7). Contudo, o tamanho e o peso da radícula não foram influenciados
pelos tratamentos de exclusão (Fig. 7).

24
Tabela 7 Análise de deviance dos modelos completos para avaliar o efeito dos dos tratamentos
nas variáveis resposta (tamanho parte aérea, tamanho radícula, peso parte aérea, peso radícula).
Variável resposta
Variável
explicativa
F
Distribuição
de Erro
Gl.
Deviance
Gl.
Residual
Deviance
Residual
P
Tamanho parte
aérea
Tratamento
4.7295
Gaussian
2
1012.6
29
3104.5
0.0167
Tamanho radícula
Tratamento
2.8245
Gaussian
2
1018.4
29
5228.0
0.076.
Peso parte aérea
Tratamento
6.0942
Gaussian
2
0.010867
15
0.025856
0.006
Peso radícula
Tratamento
1.755
Gaussian
2
8.7422e-05
15
0.00072231
0.191

25
Figura 7 Efeitos dos tratamentos de exclusão no tamanho das plântulas de Dimorphandra
mollis (A Comprimento da parte aérea; B Comprimento da radícula; C Peso da parte aérea; D
Peso da parte aérea).

26
Discussão
Homópteras e formigas comumente são observados estabelecendo relação
mutualística em diferentes espécies de planta (e.g., Abe 1998; Fagundes et al. 2005;
Kaminski et al. 2010; Neves et al. 2011). Nesta associação as formigas atraídas pelo
honeydew protegem os homópetras contra parasitas e predadores (Bristow 1983;
Buckley 1987; Buckley & Gullan 1991; Del-Claro & Oliveira 2000) e muitas vezes
podem estender seus efeitos para outros níveis tróficos (Del Claro et al. 1996; Fagundes
et al. 2005; Tegelaar et al. 2012). Contudo, poucos estudos avaliam os efeitos isolados
de homópteras produtores de honeydew ou de formigas na interação planta-homóptera-
formiga (veja neves et al. 2011).
Comparativamente a outros estudos realizados no cerrado (e.g. Neves et al.
2012; Queiroz et al. 2013; Santos & Del-Claro 2001; Oliveira et al. 2002), pode-se
dizer que a riqueza de formigas observadas nas inflorescências de Dimorphandra mollis
foi baixa. Além disto, a presença de Membracis foliata não afetou a composição de
espécies de formigas nas inflorescências de D. mollis. Contudo, deve-se destacar que as
inflorescências colonizadas por M. foliata atraiu 2,71 vezes mais formigas do que as
inflorescênciais sem M. foliata. Estes resultados salientam a importância do honeydew
produzido por M. foliata para o recrutamento das formigas.
A riqueza de insetos herbívoros coletados nas inflorescências foi baixa,
provavelmente porque D. mollis apresenta alta concentração de flavonóides,
especialmente rutina nos seus tecidos (Chavez & Usberi 2003; Oliveira 2008). Além
disso, diversos trabalhos têm relatado que no período reprodutivo pode ser encontrado
na planta uma substância denominada Astilbin considerada tóxica para alguns insetos

27
(Cintra et al. 2002; Cintra et al. 2003; Cintra et al. 2005). Além disto, a fauna de insetos
herbívoros associada com as inflorescências de D. mollis caracterizou-se principalmente
pela baixa abundância da maioria das espécies. Resultados similares também foram
descritos por outros autores que trabalharam com insetos herbivoros de vida livre
associados com um hospedeiro particular (e.g. Baccharis dracunculifolia: Fagundes &
Fernandes 2012, Cariocar brasilienis: Neves et al. 2012, Copaifera langsdorffii:
Queiroz et al. 2013). Provavelmente esta raridade da fauna de herbívoros está associada
com a alta mobilidade, baixa especificidade e alta diversidade botânica encontrada nos
trópicos (Lewinsohn et al. 2005). Contudo, deve-se ressaltar a alta abundância de
Pelidnota sp. observado nas inflorescências de D. mollis. Este escarabaeídeo possui
hábito gregário e permanece por longos períodos se alimentando protegido no interior
das inflorescências de D. mollis.
Diversos estudos têm demonstrado que formigas atraídas pelo honeydew
produzidos por homópteros afetam negativamente a riqueza e a abundância de outros
insetos herbívoros (Neves et al. 2011; Flatt & Weisser 2000). Contudo, os resultados
desta interação variam no tempo e no espaço e também dependem das espécies de
formigas e da habilidade dos insetos de evitarem as formigas (Del-Claro 1998; Gaume
et al. 1997; Chamberlain & Holanda 2009; Rosumek et al. 2009). Por exemplo, insetos
sugadores geralmente são mais sensíveis a presença das formigas porque necessitam de
maior tempo para se alimentarem na planta (Oliveira et al. 1999, Mody & Linsenmair
2004). Além disto, a simples presença de homópteras na planta hospedeira pode afetar
negativamente outros herbívoros devido a mudanças na qualidade do recurso alimentar
(Fagundes et al. 2005; Stadler & Dixon 1999).
Contrariamente ao esperado, a riqueza e a abundância de insetos sugadores nas
inflorescências de D. mollis não variaram entre os tratamentos de exclusão. Contudo, a

28
grande maioria dos insetos sugadores ocorreu nas flores em antese provavelmente
devido a maior disponibilidade de recursos alimentares nesta fase do desenvolvimento
floral (Raven et al. 2007; Kirk et al. 1995). Por outro lado, a abundância de insetos
mastigadores foi maior no tratamento de exclusão de homóptera e nas inflorescências
em ântese. De acordo com Neves et al. (2011) a presença de homópteras pode alterar a
qualidade do recurso alimentar e afetar negativamente outros herbívoros na planta,
justificando a maior abundância de herbívoros nos ramos onde os homópteros foram
excluídos. Além disto, deve-se considerar que neste estudo 85,71% dos insetos
mastigadores pertence a espécie Pelidnota sp. que se alimenta protegido no interior das
inflorescências de D. mollis. Assim, este comportamento do escarabaeídeo poderia
inibir o possível papel protetor exercido pelas formigas enquanto a simples presença do
homóptera teria um efeito mais representativo para inibir o ataque deste herbívoro nas
inflorescências de D. mollis.
A hipótese que o comportamento de Pelidnota sp. funciona como um escape das
formigas pode ser reforçada quando se observa o papel das formigas na fauna de
abelhas. De fato, a riqueza e a abundância de abelhas nas inflorescências de D. mollis
foram maiores no tratamento de exclusão de formigas, indicando que as formigas
afugentaram as abelhas das inflorescências dos tratamentos controle e exclusão de
homóptera. As abelhas geralmente visitam as inflorescências procurando recursos da
planta (néctar, pólen, resinas, odores) e em troca promovem a fecundação das flores.
Contudo, observou-se que neste estudo Trigona spinipes também coleta honeydew
excretado por M. foliata (veja também Vieira et al. 2007) e danifica as flores de D.
mollis. Neste cenário, as formigas podem desempenhar dois papéis antagônicos na
reprodução de D. mollis: (1) as formigas inibem a ação das abelhas (prováveis agentes
polinizadores) e (2) as formigas reduzem os danos produzidos pelas abelhas nas peças

29
florais. Os resultados desta interação ainda merecem maior atenção em trabalhos
futuros.
Apesar de relativamente pouco documentado, as formigas que se alimentam nos
nectários extraflorais ou de exudados de homópteras também podem afetar
positivamente o número de frutos produzidos pelas plantas (e.g. Del-Claro et al. 1996;
Keeler 1981; Messina 1981; Fernandes et al. 2005;). Resultados similares foram
observados neste estudo porque D. molis produziu maior número de frutos (jovens e
maduros) nas inflorescências do tratamento controle (presença de formigas e
homóptera). Provavelmente este resultado está associado ao fato que as formigas podem
reduzir os danos a peças florais causado por abelhas enquanto que a presença do
homóptera inibe o ataque de Pelidinota sp., conforme discutido acima. Portanto, a
interação formiga-homóptera é importante para a produção de frutos por D. mollis.
Formigas que se alimentam de honeydew geralmente atuam como defesa biótica
das plantas (Koptur 1979; Messina 1981; Del-Claro & Oliveira 1999; Coley & Barone
2001; Coley et al. 2005; Fagundes et al. 2005). Além disto, alguns estudos sugerem que
na ausência desta defesa biótica as plantas investem maior quantidade de recursos na
produção de compostos de defesa anti-herbívoros (Jansen 1966; Rehr et al. 1973;
Seigler & Ebinger 1987; Whaterman et al. 1984). Contudo, as plantas geralmente não
dispõem de recursos para investir suficientemente em crescimento, reprodução e defesa
química, gerando uma demanda conflitante entre as diferentes rotas metabólicas (Kursar
& Coley 2003; Isagi et al. 1997; Obeso 2002; Stamp 2003; Imaji & Seiwa 2010; Bryant
et al. 1983; Herms & Mattson 1992). Os resultados deste estudo mostraram que na
ausência das formigas houve redução em todos os parâmetros biométricos dos frutos e
das sementes de D. mollis. Neste contexto nós hipotetizamos que na ausência das

30
formigas D. mollis alocou mais recursos para a produção de compostos de defesa em
detrimento da formação de frutos e sementes.
O tamanho das sementes é uma característica importante para a sobrevivência
das plântulas e organização das comunidades vegetais (Cordazzo 2002; Mölken 2005;
Yalong et al. 2007; Silveira et al. 2012). Geralmente sementes grandes germinam mais
lentamente e apresentam maior percentagem de germinação comparativamente a
sementes pequenas (Geritz 1995; Ferreira & Borghetti 2004). Além disto, sementes
grandes produzem plântulas mais vigorosas e resistentes que possuem vantagens
adaptativas especialmente em ambientes previsíveis (Geritz 1995; Ferreira & Borghetti
2004; Yanlong et al. 2007). Neste estudo, as sementes oriundas das inflorescências do
tratamento exclusão de formigas apresentaram menor percentagem de germinação e
produziram plântulas menos vigorosas, indicando que a presença de formigas na planta
hospedeira pode estender seus efeitos benéficos para a progênie da planta mãe.
De acordo com Fagundes et al. (2011) a distribuição das plantas de D. mollis
entre ambientes é afetada por características das sementes e qualidade do solo. Assim, o
menor tamanho das sementes e menor vigor das plântulas observado no tratamento de
exclusão de formigas e homópteras indicam que a interação formiga-homóptera também
pode ter amplos efeitos na sobrevivência das plântulas e distribuição geográfica de D.
mollis. Finalmente, baseado nos resultados deste estudo e nas evidências da literatura
pode-se concluir que a interação formiga-homóptera desempenha um importante papel
na reprodução, estabelecimento e colonização de D. mollis.

31
Referências
Abe, Y. 1988. Trophobiosis between the gall wasp Andricus symbioticus, and the
gallattending ant, Lasius niger. Apllied Entomological Zoology. 23: 41-44.
Almeida, S.P.; Proenca, C.E.B.; Sano, S.M.; Ribeiro, J.F. 1998. Cerrado: espécies
vegetais úteis. Planaltina: Embrapa-CPAC. 188pp.
Auclair, J. L. 1963. Aphid Feeding and Nutrition. Annual Review of Entomology. 8:
439-490.
Baskin, C. C. & Baskin, J. M. 1998. Seeds – ecology, biogeography, and evolution of
dormancy and germination. Academic Press. New York.
Bawa, K. S. 1990. Plant-Pollinator Interactions in Tropical Rain Forests. Annual
Review of Ecology and Systematics. 21: 399-422.
Bazzaz, F.A.;Chiariello, N. R.; Coley, P. D.; Pitelka, L. F. 1987. Allocating resources to
reproduction and defence. BioScience. 37: 58 – 67.
Bizerril, M.X.A.; Rodrigues, F.H.G.; Hass, A. 2005. Fruit consumption and seed
dispersal of Dimorphandra mollis Benth. (Leguminosae) by the lowland tapir in the
Cerrado of Central Brazil. Brazilian Journal of Biology.65: 407-413.
Bolton, B.; Alpert, G.; Ward, P.S.; Naskrecki, P. 2007. Bolton’s catalogue of ants of the
world, 1758-2005 [CD-ROM]. Cambridge (MA): Harvard University Press.
Burgess, K.H. 1991. Florivory: the ecology of flower feeding insects and their host
plants. PhD Thesis, Harvard University, Cambridge, MA.
Bristow, C. 1983.Treehoppers Transfer Parental Care to Ants: A New Benefit of
Mutualism. Science. 220:532-533.
Bryant, J.P; Chapin, F.S.; Klein, D.S. 1983. Carbon/Nutrient balance in boreal plants in
relation on vertebrate herbivory. Oikos. 40: 357-368.
Buckley, R. C. 1987. Interactions Involving Plants, Homóptera, and Ants. Annual
Review of Ecology and Systematics. 18: 111-135.
Buckley, R.C. & Gullan, P. 1991. More aggressive ant species (Hymenoptera:
Formicidae) provide better protection for soft scales and mealybugs (Homóptera:
Coccidae, Pseudococcidae). Biotropica. 23:282–286.
Caldeira Júnior, C. F.; Santos, A. M.; Queiroz, J. M. R.; De Paula, T. O. M.; Martins, E.
R.2008. Fenologia da fava-d’anta (Dimorphandra mollis Benth.) no norte de Minas
Gerais, Brasil. Revista Brasileira de Plantas Medicinais, Botucatu. 10: 18-28.

32
Cintra, P.; Malaspina, O.; Bueno, O. C. 2003.Toxicity of barbatimao to Apis mellifera
and Scaptotrigona postica, under laboratory conditions. Journal of apicultural research.
42:9-12.
Cintra, P.; Malaspina, O.; Petacci, F.; Fernandes, J. B.; Bueno, O. C.; Vieira, P. C.;
Silva, M. F. G. F. 2002. Toxicity of Dimorphandra mollis to Workers of Apis mellifera.
Journal of Brazilian Chemical Society.13: 115-118.
Cintra, P.; Malaspina, O.; Bueno, O. C.; Petacci, F.; Fernandes, J. B.; Vieira, P. C.;
Silva, M. F. G. F. 2005. Oral toxicity of chemical substances found in Dimorphandra
mollis (Caesalpiniaceae) against honeybees (Apis mellifera) (Hymenoptera: Apidae).
Sociobiology. 45:141-149.
Chamberlain, S. A. & Holland, N. 2009. Quantitative synthesis of context dependency
in ant–plant protection mutualisms. Ecology. 90:2384–2392.
Chaves, M.M. & Usberti, U. 2003. Previsão da longevidade de sementes de faveiro
(Dimorphandra mollis Benth.). Revista Brasileira de Botânica. 26: 557-564.
Coley, P.D. & Barone, J.A. 2001. Ecology of defenses. Encyclopedia of Biodiversity,
Academic Press, San Diego. 2:11–21.
Coley, P. D & Kursar, T. A. 1996. Anti-Herbivore Defenses of Young Tropical Leaves:
Physiological Constraints and Ecological Trade-offs. Tropical Forest Plant
Ecophysiology. 1:305-336.
Coley, P.D.; Lokvam, J.; Rudolph, K.; Bromberg, K.; Sackett, T.E.; Wright, L.; Brenes-
Arguedas, T.; Dvorett, S.; Ring, S.; Clark, A.; Baptiste, C.; Pennington, R.T.; Kursar,
T.A. 2005. Divergent defensive strategies of young leaves in two species of Inga.
Ecology 86:2633–2643.
Cordazzo, C. V. 2002. Effect of seed mass on germination and growth in three
dominant species in southern brazilian coastal dunes. Brazilian Journal of Biology. 62:
427-435.
Costa, F.V.; Fagundes, M.; Neves, F.S. 2010. Arquitetura da planta e diversidade de
galhas associadas à Copaifera langsdorffii (Fabaceae). Ecologìa Austral. 20:9–17.
Costa, F.V.; Neves, F.S.; Silva, J.O.; Fagundes, M. 2011. Relationship between plant
development, tannin concentration and insects associated with Copaifera langsdorffii
(Fabaceae). Arthropod-Plant Interact 5:9–18.
Crawley, M. J. 2007. The R Book. John Wiley & Sons.
Cushman, J. H. & Beattie, A. J. 1991. Mutualisms: assissing the benefits to hosts and
visitors. Trends Ecology and Evolution. 6 :193-195.

33
Del-Claro, K. 1998. A importância do comportamento de formigas em interações:
formigas e tripes em Peixotoa tomentosa (Malpighiaceae) no cerrado. Revista de
Etologia 1: 3–10.
Del-Claro, K.; Berto, V.; Réu, W. 1996. Effect of herbivore deterrence by ants on the
fruit set of an extrafloral nectary plant, Qualea multiflora (Vochysiaceae). Journal of
Tropical Ecology. 12:887–892.
Del-Claro, K.; Oliveira, P.S. 1999. Ant-Homóptera interactions in a neotropical savana:
the honeydew-producing treehopper Guauaquila xiphias (Membracidae) and its
associated ant fauna on Didymopanax vinosum (Araliaceae). Biotropica. 31:135–144.
Del-Claro, K.; Oliveira, P.S. 2000. Conditional outcomes in a neotropical treehoper-ant
association: temporal and species specific variation in ant protection and homópteran
fecundity. Oecologia. 124:156–165.
Fagundes, M.; Camargos, M. G.; Costa, F. V. 2011. A qualidade do solo afeta a
germinação das sementes e o desenvolvimento das plântulas de Dimorphandra mollis
Benth. (Leguminosae: Mimosoideae). Acta Botanica Brasilica. 25: 908-915.
Fagundes, M. & Fernandes, G.W. 2011. Insect herbivores associated with Baccharis
dracunculifolia (Asteraceae): responses of gall forming and free-feeding insect
herbivores to latitudinal variations. Revista de Biologia Tropical.59:76–89.
Fagundes, M.; Neves, F.S.; Fernandes, G.W. 2005. Direct and indirect interactions
involving ants, insect herbivores, parasitoids and the host plant Baccharis
dracunculifolia (Asteraceae). Ecological Entomology.30:28–35.
Fernandes, G. W.; Fagundes, M.; Grego, M. K. B.; Barbeitos, M. S.; Santos, J. C. 2005
Ants and their effects on an insect herbivore community associated with the
inflorescences of Byrsonima crassifolia (Linnaeus) H.B.K. (Malpighiaceae). Revista
Brasileira de Entomologia. 49: 264-269.
Fernandes, G. W.; Fagundes, M.; Woodman, R. L. & Price, P. W. 1999. Ant effects on
threetrpphic level interactions: plants, galls, and parasitoids. Ecological Entomology.
34:411-415.
Ferreira, A. G. & Borghetti, F. 2004. Germinação: do básico ao aplicado. Editora
Artmed. Porto Alegre.
Flatt, T.; Weisser, W.W. 2000. The effects of mutualistic ants on aphid life history
traits. Ecology 38:535–536.
Gaume, L.; McKey, D.; Anstett, M.C. 1997. Benefits conferred by ‘‘timid’’ ants: active
anti-herbivore protection of the rainforest tree Leonardoxa africana by the minute ant
Petalomyrmex phylax. Oecologia. 112:209–216.

34
Geritz, S. A. 1995. Evolutionarily stable seed polymorphism and small-scale spatial
variation in seedling density. The American Naturalist. 146: 685-707.
Herbivory. Ecology Letters. 9: 1351–1365.
Herms, D. A. & Mattson, W. J. 1992. The dilemma of the plants: To grow or to defend.
The Quarterly Review of Biology. 67: 283–335.
Hölldobler, B. & Wilson, E. O.1990. The ants. Harvard University Press, Mass.
Imaji, A. & Seiwa, K. 2010. Carbon allocation to defense, storage, and growth in
seedlings of two temperate broad-leaved tree species. Oecologia. 162: 273–281.
Isagi, Y.; Sugimura, K.; Sumida, A.; Ito, H. 1997. How does masting happen and
synchronize? Journal of Theoretical Biology. 187:231–39.
Janzen, D. H. 1966. Coevolution of mutualism between ants and acacias in Central
America. Evolution 20: 249–275.
Kaminski, L.A.; Freitas, A. V. L.; Oliveira, P. S. 2010. Interaction between mutualisms:
ant-tended butterflies exploit enemy-free space provided by ant-treehopper associations.
The American Naturalist. 176:322–334.
Keetler, K. H. 1981. Function of Mentzelia nuda (Loasaceae) Postfloral Nectaries in
Seed Defense. American Journal of Botany. 68: 295-299.
Kirk, W.D.; Ali, M.; Breadmore, K.N. 1995. The effects of pollen beetles on the
foraging behavior of honey bees. Journal of Apicultural Research. 34:15–22.
Koptur, S. 1979. Facultative mutualism between weedy vetches bearing extrafloral
nectaries and weedy ants in California. American Journal of Botany. 66: 1016-1020.
Koptur, A. S.; Rodriguez, M. C.; Oberbauer, S. F.; Weekley, C.; Herndon, Alan. 2002.
Herbivore-Free Time? Damage to New Leaves of Woody Plants after Hurricane. 34:
547-554.
Kursar, T. A.; Coley, P. D. 2003. Convergence in defense syndromes of young leaves in
tropical rainforests. Biochemical Systematics and Ecology. 31- 929–949.
Lach, L. 2005. Interference and exploitation competition of three nectar-thieving
invasive ant species. Insectes Sociaux. 52:257–262.
Lewinsohn, T. M.; Novotny, V.; Basset, Y. 2005. Insects on plants: Diversity of
Herbivore Assemblages Revisited. Annual Review of Ecology, Evolution, and
Systematics. 36:597–620.
Lorenzi, H. 1992. Árvores brasileira: manual de identificação e cultivo de plantas
arbóreas nativas do Brasil. São Paulo. 382pp.

35
Mattson, W. J. Jr. 1980. Herbivory in relation to plant nitrogen content. Annual Review
of Ecology and Systematics. 11: 119-161.
McCall, A & Irwin, R. E. 2006.Florivory: the intersection of pollination and herbivory.
Ecology Letters. 9: 1351–1365.
Messina, F. J. 1981. Plant protection as a consequence of an ant-membracid mutualism:
interactions on goldenrod (Solidago sp.). Ecology 62: 1433–1440.
Mody, K. & Linsenmair, K.E. 2004. Plant-attracted ants affect arthropod community
structure but not necessarily herbivory. Ecological Entomology.29:217–225.
Mölken, T.; Jorritsma-Wienk, L. D.; Hoek, P. H.; Kroon, W. H. 2005. Only seed size
matters for germination in different populations of the dimorphic pratensis subsp.
pratensis (Asteraceae). American Journal of Botany 92: 432-437.
Murali, K. S. 1997. Patterns of seed size, germination and seed viability of tropical tree
species in southern India. Biotropica. 29: 271-279.
Neves, F. S.; Braga, R. F.; Araújo, L. S.; Campos, R. I.; Fagundes, M. 2012.
Differential effects of land use on ant and herbivore insect communities associated with
Caryocar brasiliense (Caryocaraceae). Revista de Biología Tropical. 60: 1065-1073.
Neves, F.S.; Fagundes, M.; Sperber, C.F.; Fernandes, G.W. 2011. Tri-trophic level
interactions affect host plant development and abundance of insect herbivores.
Arthropod-Plant Interactions. 5:351–357.
Obeso, J. R. 2002. The costs of reproduction in plants. New Phytologist.155: 321-348.
Oliveira, P. S. & K. Del-Claro. 2005. Multitrophic interactions in a neotropical savanna:
Anthemipteran systems, associated insect herbivores, and a host plant. In: Burslem,
DFRP; Pinard, MA; Hartley, SE. (Org.). Biotic Interactions in the Tropics. 1: 414-438.
Oliveira, P.S.; Freitas, A.V.L.; Del-Claro, K. 2002. Ant foraging on plant foliage:
contrasting effects on the behavioral ecology of insect herbivores. In: Oliveira PS,
Marquis RJ (eds) The cerrados of Brazil: ecology and natural history of a neotropical
savanna. Columbia University Press, New York. 287–305
Oliveira, P.S.; Rico-Gray, V.; Diaz-Castelazo, C.; Castillo-Guevara, C.1999. Interaction
between ants, extraforal nectaries and insect herbivores in Neotropical coastal sand
dunes: herbivore deterrence by visiting ants increases fruit set en Opuntia stricta
(Cactaceae). Functional Ecology. 13:623–631
Queiroz, A. C. M.; Costa, F. V.; Neves, F. S.; Fagundes, M. 2013. Does leaf ontogeny
lead to changes in defensive strategies against insect herbivores? Arthropod-Plant
Interactions.7:99-107.
Quilan, R. J.; Cherrett, J. M. 1977.The role of substrate preparation in the symbiosis
between the leaf-cutting ant Acromyrmex octospinosus (Reich) and its food fungus.
Ecological Entomology. 2:161–170.

36
R Development Core Team. 2011. R: a language and environment for statistical
computing. R foundation for statistical computing, http://www.r-project.org. Accessed
10 february 2011.
Raven, P.H.; Evert, R.F.; Eichorn, S.E. 2007. Biologia Vegetal. 7a. edição. Rio de
Janeiro: Guanabara Koogan. 382-407 pp.
Rehr, S. S.; Feeny, P. P.; Janzen, D. H. 1973. Chemical defence in Central American
non-ant Acacias. – The Journal of Animal Ecology. 42: 405-416.
Rico-Gray, V. & Castro, G. 1996. Effect of an ant-aphid interaction on the reproductive
fitness of paullinia fuscecens (Sapindaceae). The southwestern naturalist. 41:434-440.
Rizzini, C. T. 1965. Estudos preliminares sobre o xilopódio e outros órgãos tuberosos
de plantas do cerrado. Anais da Academia Brasileira de Ciências. 37:87-113.
Rizzini, C. T. 1997. Tratado de Fitogeografia do Brasil: Aspectos Ecológicos,
Sociológicos e Florísticos. Rio de Janeiro: Âmbito Cultural. 747p.
Rosumek, F. B.; Silveira, F. A. O.; Neves, F. S.; Barbosa, N. P. U.; Diniz, L.; Oki, Y.;
Pezzini, F.; Fernandes, G. W.; Cornelissen, T. 2009. Ants on plants: a meta-analysis of
the role of ants as plant biotic defenses. Oecologia. 160:537–549.
Santos, J.C. & Del-Claro, K. 2001. Interação entre formigas, herbívoros e nectários
extraflorais em Tocoyena formosa (Cham. & Schlechtd.) K. Schum. (Rubiaceae) na
vegetação do cerrado. Revista Brasileira de Zoociências. 3:77-92.
Seigler, D. S. & Ebinger, J. E. 1987. Cyanogenic glycosides in ant-acacias of Mexico
and Central America. The Southwestern Naturalist. 32: 499–503.
Silveira, F. A. O.; Negreiros, D.; Araújo, L. M.; Fernandes, G. W. 2012. Does seed
germination contribute to ecological breadth and geographic range? A test wit sympatric
and Diplusodon (Lythraceae) species from rupestrian fields. Plant Species Biology,
27: 170–173.
Stadler, B. & Dixon, A. F. G. 1999. Ant attendance in aphids: why different degrees of
myrmecophily? Ecological Entomology. 24:363–369.
Stamp, S. 2003. Out of the Quagmire of Plant Defense Hypotheses. The Quarterly
Review of Biology. 78: 23-55.
Tegelaar, K.; Hagman, M.; Glinwood, R.; Pettersson, J.; Leimar, O. 2012. Ant–aphid
mutualism: the influence of ants on the aphid summer cycle. Oikos. 121:61–66.
Vieira, C. U.; Rodovalho, C. M.; Almeida, L. O.; Siquieroli, A. C. S.; Bonetti, A. M.
2007. Interação entre Trigona spinipes FABRICIUS, 1793 (Hymenoptera:Apidae) E
Aethalion reticulatum LINNAEUS, 1767 (Hemiptera: Aethalionidae) EM Mangifera
indica (Anacardiaceae). Bioscience Journal 23:10–13.

37
Wäckers, F. L. 2001. A comparison of nectar- and honeydew sugars with respect to their
utilization by the hymenopteran parasitoid Cotesia glomerata. Journal of Insect
Physiology. 47:1077–1084.
Wäckers, F. L.; Rijn, P. C. J. V.; Heimpel, G. E. 2008. Honeydew as a food source for
natural enemies: Making the best of a bad meal? Biological Control. 45:176–184.
Whatterman, P. G.; Ross, J. A. M.; MacKey, D. B. 1984. Factors affecting levels of
some phenolic compounds, digestibility, and nitrogen content of the mature leaves of
Barteria fistulosa (Passifloraceae). Journal of Chemical Ecology. 10: 387-401.
Yanlong, H.; Mantang, W.; Shujun, W.; Yanhui, Z.; Tao, M.; Guozhen, D. 2007. Seed
size effect on seedling growth under different light conditions in the clonal herb
Ligularia virgaurea in Qinghai- Tibet Plateau. Acta Ecologica Sinica. 27: 3091−3108.





























