Embed Size (px)
Citation preview

DÉBORA CHAOUI PIMENTA DOS SANTOS
FRUTAS TROPICAIS COMO AGENTES MITIGADORES DE METANO EM
RUMINANTES
ILHÉUS – BAHIA
DEZEMBRO DE 2013
UNIVERSIDADE ESTADUAL DE SANTA CRUZ
PROGRAMA DE PÓS - GRADUAÇÃO EM CIÊNCIAL ANIMAL

ii
DÉBORA CHAOUI PIMENTA DOS SANTOS
FRUTAS TROPICAIS COMO AGENTES MITIGADORES DE METANO EM
RUMINANTES
Dissertação apresentada à Universidade
Estadual de Santa Cruz como parte das
exigências para obtenção do título de
Mestre em Ciência Animal, Área de
Conhecimento em Nutrição de
Ruminantes
Orientador: Prof. Dr. José Augusto Gomes
Azevêdo.
ILHÉUS – BAHIA
DEZEMBRO DE 2013

iii
S231 Santos, Débora Chaoui Pimenta dos. Frutas tropicais como agentes mitigadores em ruminantes / Débora Chaoui Pimenta dos Santos. – Ilhéus, BA: UESC, 2013. xii, 58 f. : il. Orientador: José Augusto Gomes Azevêdo. Dissertação (mestrado) – Universidade Estadual de Santa Cruz. Programa de Pós-graduação em Ciência Animal. Inclui referências. 1. Ruminantes – Nutrição. 2. Nutrição animal. 3. Frutas tropicais. 4. Metano. I. Título. CDD 636.20852

iv
DÉBORA CHAOUI PIMENTA DOS SANTOS
FRUTAS TROPICAIS COMO AGENTES MITIGADORES DE METANO EM
RUMINANTES
Dissertação apresentada à Universidade
Estadual de Santa Cruz como parte das
exigências para obtenção do título de
Mestre em Ciência Animal, Área de
Conhecimento em Nutrição de
Ruminantes.
ILHÉUS-BA, 03/12/2013.
______________________________________ Prof. Dr. José Augusto Gomes Azevêdo
Universidade Estadual de Santa Cruz - UESC Orientador
________________________________________ Prof. Dr. Luiz Gustavo Ribeiro Pereira
Embrapa Gado de Leite
_______________________________________ Prof. Dr. Márcio dos Santos Pedreira
Universidade Estadual do Sudoeste da Bahia

iv
DEDICATÓRIA
Dedico esta obra ao meu marido Rafael, maior incentivador, parceiro e
responsável para a concretização do meu curso.

v
AGRADECIMENTOS
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior -
CAPES, pela concessão da bolsa de estudos, CNPq, FAPEMIG/PPM e Embrapa
pelo financiamento do projeto.
Ao secretário do Programa de Pós Graduação em Ciência Animal da
UESC, Eduardo por estar sempre disposto e tão bem informado para esclarecer
todas as dúvidas surgidas durante o curso.
À Coordenação do Programa de Pós Graduação em Ciência Animal da
UESC, por buscar melhorias para o curso aumentando o nosso conhecimento na
área.
Aos funcionários do laboratório de nutrição animal e análise de alimentos
da UESC, Pablo e Gerson.
A Deus por me fortalecer e dar condições de ir até o fim independente das
dificuldades, por estar presente em minha vida e na vida da minha família em
cada respirar, em cada abrir e fechar de olhos.
A minha família, minha base de sustentação e motivação de vida.
Meus filhos, João Luiz e Davi, que mesmo falando que meus estudos
atrapalhavam eles ficarem comigo, ficavam quietinhos para não incomodar a
mamãe.
Minha mãe Affif, por ser realmente avó, mãe e amiga dos meus filhos,
dividindo comigo todo o trabalho de criação.
Meu irmão Danilo, também veterinário, a quem recorro para tirar dúvidas
técnicas.
Meu sobrinho e afilhado Enzo, por deixar a minha vida mais feliz tendo
chegado bem no meio do meu mestrado
Minha sogra, cunhado e cunhadas por participarem ativamente da vida da
minha família.
Ao professor José Augusto Azêvedo, pela orientação, compreensão,
amizade e pela sabedoria demonstrada.
Ao professor Luiz Gustavo Pereira, por disponibilizar seu tempo,
conhecimentos e recursos para o desenvolvimento deste trabalho.

vi
Aos colegas Brena, Flávio, Ligia e Gisele por estar sempre à disposição
para ajudar.
Aos estagiários e bolsistas em especial a Jaci, Leandro e Valclei dispondo-
se a socorrer-me quando necessário.
A minha cunhada e amigas, Michelle, Gabi, Daiane, Renata, Alessandra,
Mércia, por virarem um pouco mãe dos meus filhos durante as minhas ausências
na escola, futebol...
.
Muito obrigada a todos!

vii
SANTOS, D. C. P. Tropical Fruit as Agents in Methane Mitigators Ruminantes. 2013.57 f. Dissertação (Mestrado em Ciência Animal) Universidade Estadual de Santa Cruz, Ilhéus, 2013.
RESUMO
Objetivou-se com este trabalho avaliar alterações nas características
fermentativas ruminais decorrentes da substituição do feno de Brachiaria
brizantha por frutas, (cacau, cupuaçu, graviola, jenipapo e a casca de
mangostão) buscando a identificação de potenciais agentes mitigadores de
metano entérico. Utilizou-se delineamento inteiramente casualizado onde testou-
se os níveis de substituição (0, 30, 40 e 50) através da técnica in vitro semi
automática de produção de gás para a avaliação de volume final de carboidratos
não fibrosos (VFCNF), taxa de degradação de carboidratos não fibrosos (kdCNF),
tempo de latência, volume final de carboidratos fibrosos (VFCF), taxa de
degradação de carboidratos fibrosos (kdCF), volume total (VT), digestibilidade in
vitro da matéria seca (DIVMS), volume de gases, gás garbônico (CO2), nitrogênio
amoniacal (NH3), ácidos graxos de cadeia curta (acético, butírico e propiônico) e
pH. O VFCF superou o VFCNF, independente da fruta utilizada. Na cinética de
fermentação ruminal, foi observado o comportamento da substituição da fruta
para o aproveitamento do feno pelo animal. O jenipapo, amêndoa de cacau,
cupuaçu seguidos da casca de cacau e graviola melhoraram o perfil da
fermentação ruminal in vitro, aonde todos mantiveram ou aumentaram as kdCF e
kdCNF já o mangostão não melhorou e piorou kdCF. No que diz respeito aos
produtos da fermentação, foram avaliados nitrogênio amoniacal (NH3), gás
carbônico (CO2), gás metano (CH4), volume de gases, ácidos graxos de cadeia
curta (AGCC), sendo eles, acético, propiônico e butírico, e pH. As frutas casca de
cacau, cupuaçu, graviola e jenipapo apresentaram aumento na produção do CH4
A amêndoa do cacau e a casca de mangostão no tempo de 24 horas de
incubação apresentaram comportamento diferente, já que para a amêndoa de
cacau não houve alteração (P>0,05) na produção de metano à medida que existiu
a substituição pelo feno; e a casca de mangostão houve redução de até 31,5%
na produção de metano quando comparado o nível de substituição de 50% do

viii
feno. Entre as frutas tropicais cacau, cupuaçu, graviola, jenipapo e a casca de
mangostão àquela com potencial mitigador de metano entérico in vitro é apenas a
casca do mangostão. A amêndoa do cacau não proporciona aumento na
produção de metano entérico in vitro.
Palavras–chave: Frutas Tropicais. Metano. Ruminante

ix
SANTOS, D. C. P. Tropical Fruit as Agents in Methane Mitigators Ruminantes. 2013.57 f. Dissertação (Mestrado em Ciência Animal) Universidade Estadual de Santa Cruz, Ilhéus, 2013.
ABSTRACT
The objective of this study was to evaluate changes in ruminal fermentation characteristics resulting from the replacement of a Brachiaria Brizantha by fruit (cacao , cupuaçu , soursop , mangosteen peel and genipap ) seeking to identify potential mitigating agents of enteric methane . We used a randomized design in which we tested the levels ( 0 , 30 , 40 and 50 ) through in vitro semi auto gas production to evaluate final volume of non-fiber carbohydrates ( VFCNF ) , rate degradation of non- fiber carbohydrates ( kdCNF ) , latency time , final volume of fiber ( VFCF ) , rate of degradation of fiber ( kdCF ) , total volume ( VT ) , in vitro digestibility of dry matter (DM ) , volume gases , carbônico dioxide (CO2 ) , ammonia nitrogen ( NH3 ) , short chain fatty acids (acetic , propionic and butyric acid ) and pH . The VFCF overcame VFCNF , regardless of the fruit used . On the kinetics of ruminal fermentation , the behavior of substitution of fruit for use by hay animal was observed . Genipap , cocoa beans , cupuaçu followed by cocoa husks and soursop improved the profile of ruminal fermentation in vitro , where all maintained or increased kdCF and kdCNF now the mangosteen has not improved and worsened kdCF . With respect to the fermentation products were evaluated ammoniacal nitrogen (NH3) , carbon dioxide ( CO2), methane ( CH4) gas volume , short chain fatty acids ( SCFA ) , namely , acetic , propionic and butyric and pH. The fruit peel cacao , cupuaçu , soursop and genipap showed increased production of CH4 Cocoa beans and coat mangosteen at 24 hours of incubation showed different behavior , since for cocoa beans did not change ( P > 0 05 ) in methane production as it existed at the replacement hay ; and mangosteen rind was up to 31.5 % reduction in methane production when compared to the replacement level of 50 % hay . Tropical fruit cacao, cupuaçu , soursop , mangosteen peel genipap and that containing enteric methane mitigation potential in vitro is only the rind of the mangosteen . Cocoa beans provides no increase in enteric methane production in vitro. Keywords: Tropical Fruits. Methane. Ruminant.

x
LISTA DE FIGURAS
Figura Pag.
Figura 1 Estimativa de produção de metano (CH4) (mL/ g MS incubada ou MS digerida), em função do nível de substituição do feno de Brachiaria brizantha pela amêndoa do cacau, por 12 e 24 horas de incubação in vitro.
43
Figura 2 Estimativa de produção de metano (CH4) (mL/ g MS incubada ou MS digerida), em função do nível de substituição do feno de Brachiaria brizantha pela casca de cacau, por 12 e 24 horas de incubação in vitro.
44
Figura 3 Estimativa de produção de metano (CH4) (mL/ g MS incubada ou MS digerida), em função do nível de substituição do feno de Brachiaria brizantha pelo cupuaçu, por 12 e 24 horas de incubação in vitro.
45
Figura 4 Estimativa de produção de metano (CH4) (mL/ g MS incubada ou MS digerida), em função do nível de substituição do feno de Brachiaria brizantha pela graviola, por 12 e 24 horas de incubação in vitro.
47
Figura 5 Estimativa de produção de metano (CH4) (mL/ g MS incubada ou MS digerida), em função do nível de substituição do feno de Brachiaria brizantha pelo jenipapo, por 12 e 24 horas de incubação in vitro.
48
Figura 6 Estimativa de produção de metano (CH4) (mL/ g MS incubada ou MS digerida), em função do nível de substituição do feno de Brachiaria brizantha pela casca de mangostão, por 12 e 24 horas de incubação in vitro.
49

xi
LISTA DE TABELAS
Tabela Pag.
Tabela 1. Composição bromatológica dos frutos e feno de Brachiaria brizantha. 26 Tabela 2. Estimativa dos parâmetros da cinética de fermentação ruminal in vitro
dos carboidratos em função do nível de substituição do feno de Brachiaria brizantha pelo jenipapo.
31
Tabela 3. Estimativa dos parâmetros da cinética de fermentação ruminal in vitro dos carboidratos em função do nível de substituição do feno de Brachiaria brizantha pela amêndoa de cacau.
31
Tabela 4. Estimativa dos parâmetros da cinética de fermentação ruminal in vitro dos carboidratos em função do nível de substituição do feno de Brachiaria brizantha pelo cupuaçu.
32
Tabela 5. Estimativa dos parâmetros da cinética de fermentação ruminal in vitro dos carboidratos em função do nível de substituição do feno de Brachiaria brizantha pela casca de cacau.
33
Tabela 6. Estimativa dos parâmetros da cinética de fermentação ruminal in vitro dos carboidratos em função do nível de substituição do feno de Brachiaria brizantha pela graviola.
34
Tabela 7. Estimativa dos parâmetros da cinética de fermentação ruminal in vitro dos carboidratos em função do nível de substituição do feno de Brachiaria brizantha pela casca de mangostão.
34
Tabela 8. Produtos da fermentação ruminal em função do nível de substituição do feno de Brachiaria brizantha pela casca do cacau, por 24 horas de incubação in vitro.
36
Tabela 9. Produtos da fermentação ruminal em função do nível de substituição do feno de Brachiaria brizantha pelo cupuaçu, por 24 horas de incubação in vitro.
37
Tabela 10. Produtos da fermentação ruminal em função do nível de substituição do feno de Brachiaria brizantha pela casca do mangostão, por 24 horas de incubação in vitro.
37
Tabela 11. Produtos da fermentação ruminal em função do nível de substituição do feno de Brachiaria brizantha pela amêndoa do cacau, por 24 horas de incubação in vitro.
39
Tabela 12. Produtos da fermentação ruminal em função do nível de substituição do feno de Brachiaria brizantha pelo jenipapo, por 24 horas de incubação in vitro.
40
Tabela 13. Produtos da fermentação ruminal em função do nível de substituição do feno de Brachiaria brizantha pela graviola, por 24 horas de incubação in vitro.
41

xii
SUMÁRIO
Pag.
1. INTRODUÇÃO 13
2. OBJETIVOS 14
2.1. Geral 14
2.2. Específicos 14
3. REVISÃO DE LITERATURA 15
3.1. Produção de Metano. 16
3.2. Estratégias de Mitigação. 18
3.3. Características de Alimentos com Potencial Mitigador de Metano. 19
3.4. Método in vitro para avaliação de metano entérico. 21
3.5. Frutas analisadas. 22
4. MATERIAL E MÉTODOS 24
4.1. Determinação da composição química. 25
4.2. Características fermentativas dos alimentos. 26
4.3. Parâmetros de cinética de fermentação e técnica semi-automática de produção de gases in vitro.
28
4.4. Análise estatística. 29
5. RESULTADOS E DISCUSSÃO 30
5.1. Cinética de fermentação ruminal in vitro dos carboidratos. 30
5.2. Produtos da fermentação ruminal in vitro. 36
5.3. Produção de metano entérico. 43
6. CONCLUSÕES 50
7. REFERÊNCIAS BIBLIOGRÁFICAS 51

13
1. INTRODUÇÃO
O CH4 é um GEE emitido significativamente pela pecuária, em especial, a
brasileira, o que à torna alvo de criticas. A produção de metano resulta da
fermentação anaeróbica da matéria orgânica em ambientes alagados, campos de
arroz cultivados por irrigação de inundação, fermentação entérica, tratamento
anaeróbico de resíduos animais e queima de biomassa (FAO, 2013).
A utilização de algumas práticas pode tornar a agropecuária mais
sustentável reduzindo a emissão desses gases. Existem várias formas de mitigar
GEE, destacando-se a melhora da qualidade das pastagens, utilização animais
geneticamente melhorados, introdução de alguns nutrientes à dieta que tenham
potencial mitigador de CH4, como por exemplo, lipídios e alimentos concentrados
que irão produzir mais ácido propiônico e menos ácido acético, diminuindo a
quantidade de H2 livre e conseqüentemente a metanogênese.
O Brasil destaca-se como grande produtor e exportador de frutas que por
serem produtos perecíveis existem altos índices de perdas. Recentemente tem
crescido o interesse pelo consumo de frutas tropicais e seus produtos na forma
industrializada, atrelado a isso, cresce o volume de resíduos gerados pela
industrialização destas frutas. Esse fato somado ao crescimento do poder
aquisitivo da população mundial e ao aumento no consumo de produtos de
origem animal tem levado os pesquisadores a buscar soluções para utilização
desses resíduos introduzindo-os na alimentação animal.
As pastagens são as fontes de alimentos mais utilizadas pelos animais,
porém, elas não estão disponíveis em quantidade e qualidade suficiente por todo
o ano, assim, existe a necessidade de suplementar os animais.
Acredita-se que além da viabilidade econômica, os subprodutos podem
garantir uma nutrição adequada aos animais que gerarão produtos de alto valor
nutricional e de forma sustentável, visto que, o excesso dos resíduos no solo pode
gerar gases de efeito estufa (GEE). Sendo os ruminantes um dos responsáveis
pela emissão desses GEE dentre eles destacando-se o metano (CH4), pesquisas
vêm sendo realizadas, a fim de conhecer a composição química desses
subprodutos descobrindo os benefícios dos mesmos tanto para nutrição animal
quanto para seu efeito mitigador de GEE.

14
Uma forma prática e economicamente viável para verificar a característica
dos alimentos, é a técnica de produção de gases in vitro, a qual, tem sido uma
alternativa adequada para mensuração de metano entérico, pois tem retratado
eficientemente os acontecimentos in vitro podendo ser utilizada para formação de
uma base de dados de produção potencial de metano de diversas dietas para
ruminantes.
O presente trabalho teve o objetivo de estudar cinco (05) frutas (cacau,
cupuaçu, graviola, jenipapo e mangostão), facilmente encontrados na região sul
da Bahia e descobrir através da incubação in vitro os seus potenciais de
mitigação de metano entérico sem prejuízos nutricionais para ruminantes.
2. OBJETIOS:
2.1. GERAL: Analisar alimentos a fim de verificar o potencial de mitigação
de metano entérico dos mesmos através da técnica de
incubação in vitro.
2.2. ESPECÍFICOS: Realizar análises laboratoriais de frutas para verificação da
composição químico-bromatológica das mesmas observando
a sua capacidade e qualidade nutricional para os animais,
potencial de digestibilidade e capacidade de produção de
gases;
Realizar incubação das frutas através da técnica de produção
de gases in vitro para mensurar o volume de metano
produzido pelas mesmas;
Avaliar frutas disponíveis no sul da Bahia com agentes
mitigadores de CH4 em ruminantes.

15
3. REVISÃO DE LITERATURA
Com o crescimento da população mundial e do poder aquisitivo da mesma
a demanda por alimentos de origem animal cresceu (FAO, 2013). De acordo com
dados do IBGE divulgados em novembro de 2010, estimou-se que o Brasil
possuía uma população de 190.732.694 pessoas que vivem em uma área com
superfície de 8.515.767,049 km2. A região Nordeste possui uma área de 1,56
milhões de km2 (18,2% do território nacional) e sua população foi estimada em
51.609.027 habitantes (28,05% da população brasileira).
Em relação a 2010, estimou-se que o rebanho bovino de 2011 teve um
crescimento nacional de 1,6%, totalizando cerca de 212,8 milhões de cabeça.
Após a Índia o Brasil ocupa a 2ª posição mundial em rebanho de gado bovino,
sendo que na Índia o rebanho não é comercial e inclui bubalinos. A distribuição do
rebanho no território brasileiro apresentava-se da seguinte forma: Região Centro
Oeste (34,1%), Norte (20,3%), Sudeste (18,5%), Nordeste (13,9%) e Sul (13,1%),
sendo Mato Grosso (13,8%) o estado de maior efetivo bovino (IBGE, 2011).
Contrapondo-se a esse crescimento surge um fator ambiental que leva a
pecuária brasileira a ser alvo de críticas pela emissão de gases de efeito estufa
(GEE). Na pecuária, o gás metano (CH4), formado a partir da fermentação
entérica dos carboidratos, é o principal responsável pelas emissões no setor. Isso
se agrava ainda mais devido ao modelo ineficiente de exploração pecuária,
baseada em pastagens degradadas que tem como conseqüência maior
quantidade de GEE por Kg de carne produzida (IPCC, 2007).
Os ruminantes possuem microrganismos no rúmen que vivem em simbiose
e são capazes de fermentar carboidratos da dieta e produzir ácidos graxos de
cadeia curta (AGCC) além de gases (dióxido de carbono (CO2) e CH4). O metano,
formado devido à redução do dióxido de carbono, não pode ser usado como um
substrato energético e é eructado no ar contribuindo com gases de efeito estufa
(GEE), resultando em poluição do ambiente (MARTIN et al., 2009).
Segundo Thorpe (2009) o Brasil é o maior emissor de metano entérico
através de bovinos (9,6 Tg de CH4 /ano). A produção de metano entérico pelos
ruminantes contribui 22% da produção mundial desse gás que representa 3,3%
do total dos GEE (USEPA, 2000). Como o aquecimento global vem sendo foco de
preocupação da sociedade e considerado que o CH4 é 25 vezes maior que o CO2

16
em potencial de aquecimento global (IPCC, 2006) e seu tempo de vida na
atmosfera é estimado em 9 a 15 anos, ações de pesquisa apresentam um
importante papel no sentido de desenvolver estratégias para uma produção de
alimentos de origem animal, ambientalmente seguros e sustentáveis.
A elaboração de dietas e sistemas de manejo que minimizem a produção
de metano sem afetar negativamente à eficiência produtiva animal tem sido um
desafio (MACHADO et al., 2011). Para que fiquem claras as formas de mitigar a
produção de metano, faz-se necessário conhecer como se dá essa produção.
3.1. Produção de Metano
A produção de metano resulta da fermentação anaeróbica da matéria
orgânica em ambientes alagados, campos de arroz cultivados por irrigação de
inundação, fermentação entérica, tratamento anaeróbico de resíduos animais e
queima de biomassa (MACHADO et al., 2011). Devido ao fato do CH4 poder ser
produzido através da matéria orgânica, pode ser considerado um biogás.
Segundo Harper et al. (1999), a produção de arroz em terrenos alagados, a
fermentação entérica e a fermentação de dejetos contribui respectivamente com
11%, 16% e 17% do total de CH4 liberado para a atmosfera.
Uma das principais fontes antrópicas de metano, é o cultivo de arroz
irrigado por inundação. De acordo com o Painel Intergovernamental de Mudança
Climática - IPCC (1996), a emissão global desse gás nos campos de arroz
irrigado corresponde a 16% do total de emissão de todas as fontes, estimando-se
de 20 a 100 teragramas, média de 60 Tg (Tg= Teragrama 1012 grama) por ano.
Durante a queima da biomassa de resíduos de colheita e de culturas
agrícolas na pré-colheita, leva à produção de alguns gases, entre eles, metano,
óxido nitroso (N2O), óxidos de nitrogênio (NOx) e monóxido de carbono (CO),
além do CO2. Durante a combustão, o fogo libera o carbono da biomassa e
diretamente do solo. Um exemplo de queima de cultura no Brasil é a da cana de
açúcar, porém, embora ocorra liberação de CO2 durante a queima, as emissões
deste gás não são consideradas como uma emissão líquida ao longo do tempo
por esses sistemas, pois, o CO2 emitido será reabsorvido no ciclo seguinte desta
cultura (IPCC, 1996).

17
Já a pecuária, objeto de interesse deste estudo, contribui para as emissões
de metano por duas vias: fermentação entérica e dejetos animais.
Vários fatores influenciam a quantidade de CH4 produzido por um animal,
destacando-se os fatores nutricionais, tipo de carboidrato na dieta, o nível de
consumo de ração, o nível de produção (produção anual de leite e carne e seus
derivados), taxa de passagem da dieta, a presença de ionóforos, o grau de
saturação dos lipídios na dieta, fatores ambientais, tais como a temperatura
(McALLISTER et al., 1996), e fatores genéticos, como a eficiência de conversão
alimentar (NKRUMAH et al., 2006).
A produção de metano a partir dos dejetos animais acontece quando estes
são manipulados na forma líquida, em condições de anaerobiose. Na pecuária
nacional em 2005, 6% do metano emitido foi de origem no esterco (MCT, 2010).
De acordo com o IPCC, 1996, essa condição se dá geralmente em criações
grandes de animais confinados.
As emissões globais de metano através da fermentação entérica
acontecem em herbívoros ruminantes, como bovinos, ovinos, bubalinos e
caprinos, e são estimadas em cerca de 80 milhões de toneladas anuais
(U.S.E.P.A, 2000). De acordo com MCT (2010), em 2005, 94% do metano emitido
pela pecuária nacional foi de origem entérica.
Durante o processo oxidativo da fermentação, co-fatores reduzidos (NADH,
NADPH e FADH) são formados e devem ser reoxidados a (NAD+, NADP+ e
FAD+) a fim de que o processo oxidativo não seja paralisado. Durante o processo
de reoxidação, hidrogênios são liberados no rúmen através de reações de
desidrogenação. O hidrogênio livre é utilizado pelas Archaea metanogênicas para
a produção de metano (metanogênese), sendo este processo importante para o
ecossistema ruminal, pois evita o acúmulo do hidrogênio no rúmen (McALLISTER
e NEWBOLD, 2008).
Durante o metabolismo dos carboidratos, (CHO) ocorre fermentação onde
esses CHO são transformados em ácidos graxos de cadeia curta (sendo o ácido
acético (C2), propiônico (C3) e butírico (C4), os principais) CO2 e CH4
(BERCHIELLI et al., 2003). A proporção de cada um desses ácidos dependerá do
tipo da dieta (grãos ou volumosos), durante a fermentação dos carboidratos
fibrosos, produz-se mais acetato e butirato, resultando em liberação líquida de

18
hidrogênio (H2) e favorecendo a metanogênese. Já a formação de propionato,
favorecido por dietas ricas em grãos, é uma via competitiva de utilização de H2 no
rúmen, reduzindo a disponibilidade de substrato para a metanogênese. Assim, a
produção de metano, que depende do balanço de H2 no rúmen, é influenciada
pelas taxas de produção de acetato e propionato (OWENS e GOETSCH, 1988;
HEGARTY, 2001).
3.2. Estratégias de Mitigação
Segundo Machado et al. (2011), a rotulação dos bovinos como os grandes
vilões das mudanças climáticas que vem sendo construída e muitas vezes sem
embasamento técnico científico, pode ser futuramente um pretexto para a criação
de barreiras não tarifárias à exportação de produtos pecuários brasileiros.
Este fato ligado ainda as pressões ambientais indicam que a redução da
emissão de CH4 de origem pecuária, é um dos principais fatores para nortear as
pesquisas com a produção de ruminantes (MACHMÜLLER, 2006).
Estando a produção de metano entérico relacionado à presença de H2 livre,
pesquisadores buscam avaliar alguns alimentos e seu potencial de mitigação de
CH4. Kebreab et al. (2010), demonstraram que o volume de metano produzido
depende da quantidade do alimento e de suas características e composição, visto
que a composição química dos alimentos determinará a quantidade de cada ácido
graxo de cadeia curta e a quantidade de hidrogênio produzidos.
De acordo com Machado et al. (2011), a substituição de alimentos com
maiores concentrações de carboidratos fibrosos (CF) (forragens) por carboidratos
não fibrosos (CNF) (amido e açucares), implica no desenvolvimento de bactérias
amilolíticas e, por conseguinte em mudanças na produção dos AGCC,
promovendo aumento na proporção do propionato e redução do acetato. Esta
condição provoca diminuição na produção de metano devido à redução da
disponibilidade de H2 no rúmen.
Martin et al. (2009), afirmam que a baixa relação acetato:propionato nem
sempre ocorre em animais alimentados com dietas ricas em concentrado. Nessas
situações a redução da emissão de metano observada pode ser explicada pela
redução do pH ruminal e declínio do número de protozoários. O baixo pH ruminal

19
também pode inibir o crescimento e/ou atividade das metanogênicas e bactérias
celulolíticas.
Alguns pesquisadores, como Pedreira et al. (2005), sugerem a melhoria da
qualidade da dieta, das pastagens e a suplementação como forma de diminuir a
emissão do CH4 por conseqüência da redução do ciclo de produção, aumentando
a eficiência alimentar e reduzindo a produção de metano por unidade de produto
(carne, leite). Existem ainda aqueles que afirmam o maior desempenho dos
animais também ajuda a diminuir o CH4 em função da redução do número de
animais no sistema de produção (MOSS e GIVENS, 2002). Segundo Guimarães
et al. (2010), melhorando a eficiência do sistema de produção a emissão de
metano será proporcionalmente reduzida, uma vez que mais produto (carne, leite,
lã, etc.) será produzido em relação aos recursos utilizados. As pesquisas mostram
que, mesmo que a emissão de N2O possa ser aumentada pelo uso de fertilizantes
nitrogenados, intensificando-se os sistemas de produção pode-se reduzir a
emissão entérica de CH4 por unidade de produto. (MONTEIRO, 2009). De acordo
com Berndt (2011), esta redução está principalmente relacionada ao melhor
aproveitamento do alimento e à redução da idade de abate.
3.3. Características de Alimentos com Potencial Mitigador de Metano
As forragens são o principal alimento do rebanho bovino brasileiro, visto
que, a maior parte do rebanho é criado a pasto.
O uso de grãos e alimentos concentrados, processamento e conservação
de forragens, leguminosas, taninos e saponinas, óleos e gorduras saturadas e
insaturadas, ionóforos, nitrato, leveduras, malato e fumarato, óleos essenciais e
extratos vegetais, têm sido estudado com o objetivo de reduzir as emissões de
metano (Berndt, 2010).
A substituição de alimentos com maiores concentrações de carboidratos
fibrosos por carboidratos não fibrosos promove aumento na proporção do
propionato e redução do acetato. Esta condição provoca diminuição na produção
de metano devido à redução da disponibilidade de H2 no rúmen (MACHADO et
al., 2011). Todavia, a utilização de grande percentual de carboidratos não fibrosos

20
na dieta de ruminantes, além de aumentar o custo de produção, pode prejudicar o
metabolismo animal e influenciar diretamente na produção.
A utilização de ionóforos, como a monensina, na dieta de animais
ruminantes também pode influenciar a produção de CH4, isto porque os ionóforos
selecionam as bactérias gram-negativas produtoras de propionato, enquanto as
gram-positivas são produtoras de acetato e butirato (geram CO2 e H2 para a
síntese de CH4) e diminuem a população de Archaeas metanogênicas (MOOS,
1993; SAUER et al., 1998).
Porém, os ionóforos não sustentam a metanogênese por um longo período,
provavelmente devido à habilidade de adaptação da microflora ruminal. De acordo
com Sauer et al. (1998), vacas em lactação que nunca haviam sido
suplementadas com monensina, tiveram redução na produção de metano de 10%
a 21% ao longo de 65 dias de suplementação com monensina. Após 161 dias
sem suplementação, os mesmos animais receberam novamente monensina e
observaram-se pequenos efeitos na produção de metano, de +1% a -11%. Além
disso, surge a preocupação de que o uso de antibióticos na produção animal pode
originar fatores de resistência transmissíveis, que podem comprometer a
eficiência terapêutica dos antibióticos no tratamento de infecções que acometem
os humanos, porém, é questionável (CASEWELL et al., 2003). Em medida de
precaução, a legislação da União Européia proíbe o uso de antibióticos
promotores de crescimento, como os ionóforos, na alimentação de animais
(REGULAMENTO 1831/2003, 2003). Portanto, nenhum produto de animais que
receberam essas substâncias pode ter ingresso no mercado da União Europeia.
Atualmente, o aumento do teor de lipídios da dieta de ruminantes é
considerado a forma mais eficaz de metanogênese ruminal consistentemente
deprimente (GRAINGER e BEAUCHEMIN, 2011).
A depender do grau de saturação da gordura e a quantidade do
fornecimento de gorduras insaturadas na dieta dos ruminantes, estas podem
promover a redução de CH4 por ação deletéria sobre as metanogênicas e
consumo do H2 no processo de biohidrogenação (MACHMÜLLER et al., 1998;
FIEVEZ et al., 2003). Em trabalho avaliando o efeito do fornecimento de ácido
graxo de cadeia média, Machmüller (2006) encontrou que mesmo em

21
concentrações inferiores a 3% da dieta, a suplementação com os ácidos graxos
C12:0 e C14:0 é responsável pela redução de 50% na produção de CH4.
No entanto o uso da suplementação lipídica em altos níveis tende a reduzir
a digestibilidade dos nutrientes no rúmen, reduzindo assim o ganho de peso e a
produtividade do sistema.
Sendo assim, é difícil encontrar uma única estratégia alimentar que seja
eficiente no processo de mitigação de metano entérico. Para a obtenção de um
melhor resultado na redução da emissão do CH4, além da utilização de aditivos
alimentares com potencial de mitigar metano, podem-se adotar várias medidas
que em conjunto tornam-se mais eficientes. De acordo com O´hara et al. (2003), o
fornecimento de forragens de melhor qualidade, o melhoramento genético dos
animais priorizando o desempenho produtivo são os principais recursos
conhecidos disponíveis.
3.4. Método in vitro para avaliação de metano entérico
Para que se desenvolver estratégias de mitigação de metano, faz se
necessário a mensuração entérica desse gás. Existem algumas técnicas capazes
de realizar essa mensuração, entre elas está a técnica in vitro de produção de
gases (MACHADO et al., 2011).
A técnica de produção de gases in vitro tem sido uma alternativa adequada
para mensuração de metano entérico, pois tem refletido os acontecimentos in vivo
com acurácia, portanto, são adequadas para a formação de uma base de dados
de produção potencial de metano de diversas dietas para ruminantes e análise de
estratégias para mitigar as emissões (GETACHEW et al., 2005; BHATTA et al.,
2006).
Essa técnica também é viável quando consideramos o custo da realização
de mensuração in vivo, já que a manutenção de animais para a realização desta
avaliação representam custos elevados.
Nas técnicas in vitro, o ambiente ruminal é simulado através de uma
amostra de líquido ruminal em cultura contínua ou em frascos de fermentação. A
produção de metano pode então ser calculada por cromatografia gasosa
realizando a mensuração da produção total de gases, amostragem dos gases

22
produzidos e análise de sua composição. A medição do volume de gases pode
ser realizada com o auxílio de um transdutor de pressão ou voltímetros, conforme
Maurício et al. (2003) e Schofield e Pell (1995).
Alguns autores como Hristov et al. (2012) afirmam que apesar de os
sistemas in vitro, serem convenientes para o rastreio de um grande número de
tratamentos, existem alguns fatores onde eles não têm representatividade da in
vivo rúmen e geralmente não abordam a principal questão de adaptação do
ecossistema ao rúmen a prática de mitigação.
Dessa forma, o ideal seria utilizar a priori a técnica in vitro e após os
resultados, sendo estes positivos, testá-los na técnica in vivo de produção de
gases.
3.5. Frutas analisadas
Cacau
O nome científico da planta do cacau é Theobroma cacao L. O cacau é
uma planta originaria da América do Sul e Central e seu cultivo tem se espalhado
por todo mundo (ROSS, 2001). Um fruto pode conter de 30 a 40 sementes
envolvidas pela polpa do fruto. Proteínas, gorduras, fibras, cinzas, cálcio, ferro,
tiamina, teobromina, cafeína, niacina são compostos encontrados no cacau. O
grão cru (cacau) possui 1-2% de teobromina e 0,1% de cafeína (DUKE, 1983).
Segundo dados do IBGE, o Brasil produziu 218,48 mil toneladas de cacau
(amêndoa) em 2009, sendo as Regiões Norte/ Nordeste responsável por mais de
96% dessa produção. Nesse mesmo ano, o Nordeste produziu 137,92 mil
toneladas da amêndoa do fruto, respondendo a Bahia por 100% da produção
regional e cerca de 64% da nacional (SENA, 2011).
No arraçoamento de animais, a casca pode ser considerada como um
volumoso de boa qualidade e um excelente substituto do capim de corte, tanto
pela sua composição quanto pela sua palatabilidade (SILVA NETO et al., 2001).

23
Cupuaçu
O cupuaçuzeiro (Theobroma grandiflorum, Schum) é uma planta frutífera
encontrada na parte sul e sudeste da Amazônia Oriental (VENTURIERI et al.,
1985). Segundo Carvalho (2004), o estado do Pará é o maior produtor nacional
desta fruta, seguido pelos estados do Amazonas, Rondônia e Acre. Foi
introduzida no sul da Bahia em 1930, na antiga Estação Experimental de Água
Preta, no município de Uruçuca.
O produto mais comercializado deste fruto é a polpa (ARAGÃO, 1992). A
torta do cupuaçu é o resíduo da extração do óleo da semente seca, livre de
qualquer resíduo da polpa (CARVALHO, 2004).
Graviola
A gravioleira (Annona muricata) é originária da América Central, sendo
cultivada no Brasil, Colômbia, México, Havaí e algumas regiões da África e Ásia,
desenvolve-se bem em regiões de clima tropical e subtropical. Nutricionalmente, a
fruta é rica em carboidratos, especialmente a frutose (SACRAMENTO, 2000).
Jenipapo
O jenipapo (Genipa americana L.) é uma fruta nativa da América central,
porém, bastante disseminada no norte e nordeste brasileiro e possui excelentes
características relacionadas ao sabor, aroma e aparência (HANSEN et al. 2008).
Os frutos são do tipo baga sub-globosa, de 8 a 10cm de comprimento e 6 a
7cm de diâmetro, casca mole, parda ou pardacento-amarelada, membranosa, fina
e enrugada (Correa, 1969). O jenipapeiro adapta-se muito bem ao clima tropical,
não existindo restrições quanto a altas temperaturas (Xavier & Xavier, 1976).
Os ácidos orgânicos constituem uma excelente reserva energética dos
frutos, através de sua oxidação no ciclo de Krebs (Brody, 1996).
Mangostão
O mangostão (Garcinia mangostana L.) é nativo do sudeste da Ásia e
considerado a fruta mais saborosa do trópico asiático (Sacramento et al., 2007).
Considerando a sua região de origem o mangostanzeiro é cultivado em áreas

24
onde o clima é quente e úmido com chuvas bem distribuídas durante o ano, o que
favorece o cultivo na região sul da Bahia.
A casca é dura e libera uma espécie de resina amarela quando cortada. A
casca do mangostão representa expressivo teor de resíduo agroindustrial não
aproveitado e pode ser benéfica à saúde, pois apresenta elevado teor de fibras.
A casca de mangostão contém de taninos e saponinas, os quais exercem
um específico efeito contra protozoários do rúmen e reduziu emissões de gás
metano, enquanto que o resto do rúmen a biomassa permanece inalterado
(NGAMSAENG et al. , 2006).
De acordo com Guo et al. (2008) compostos de plantas tais como taninos
condensados e saponinas têm sido mostrados para diminuir atividade
metanogênica de protozoários.
4. MATERIAL E MÉTODOS
Foram usados no teste de screening frutos disponíveis na região sul da
Bahia (graviola, cupuaçu, jenipapo, mangostão e cacau (casca e amêndoa).
Os frutos foram coletados em árvores na zona rural do município de Ilhéus-
Ba, no distrito do Japú.
Do cacau colhido à mão em estágio de amadurecimento pronto para o
consumo, utilizou-se a amêndoa inteira do cacau e separadamente a casca
externa da fruta. Esses dois constituintes da fruta foram analisados
separadamente.
Do cupuaçu também coletado à mãe direto da árvore e já amadurecido,
utilizou-se a parte interna, polpa e sementes, sendo retirada apenas a casca.
A graviola foi colhida ainda verde e utilizada inteira, incluído a casca, polpa,
sementes e talos, não havendo seleção prévia existindo, portanto variação de
tamanhos e grau de amadurecimento.
O jenipapo maduro foi coletado à mão e utilizado inteiro, incluído a casca,
polpa e sementes, e assim como na graviola, não houve seleção prévia existindo,
portanto variação de tamanhos e grau de amadurecimento.
Do mangostão, coletado na árvore, foi utilizada apenas a casca da fruta
com todos os seus constituintes.

25
4.1. Determinação da composição química
Cerca de 15 kg de amostra de cada fruto foram pré-secas em estufa de
ventilação forçada a 60-65ºC, por 72 horas e, depois processadas em moinho de
facas tipo Willey, com peneira de 1 milímetro e em seguida foram armazenadas
em recipientes de polietileno, para posteriores análises de matéria seca (MS),
compostos nitrogenados (N), matéria mineral (MM), extrato etéreo (EE) e fibra em
detergente ácido (FDA) conforme AOAC (1990).
Nas análises de fibra em detergente neutro (FDN), as amostras foram
tratadas com alfa-amilase termo-estável, sem o uso de sulfito de sódio, e
corrigidas para nitrogênio e cinzas residuais (Mertens, 1992). A determinação dos
conteúdos de compostos nitrogenados insolúveis nos detergentes neutro (NIDN)
e ácido (NIDA) foram realizados conforme Licitra et al. (1996).
As frações que compõem os carboidratos totais (CT) foram estimadas
conforme Sniffen et al. (1992), obtidas pela fórmula: %CT= 100– (%PB + %EE +
%MM), em que: %PB corresponde à concentração de proteína bruta da amostra,
%EE corresponde à concentração de extrato etéreo e %MM à concentração de
cinzas. A concentração dos carboidratos não fibrosos (CNF), expressos em % na
matéria seca, foi estimada de acordo com Hall (2003):
%CNF = 100 - (%PB + %FDNcp + %EE + %MM)
Para determinação da energia bruta (EB) empregou-se uma bomba
calorimétrica, modelo Parr 1108, com unidade de ignição Parr 2901, calorímetro
Parr 1341, fabricada pela Parr Instrument Company (Moline, IL, USA). Cadinhos
de aço inox, fornecidos com a bomba, foram usados para receber a amostra. Fios
metálicos, também vendidos pelo mesmo fornecedor, serviram para a condução
da corrente elétrica entre os eletrodos e a amostra.
Os parâmetros de cinética de fermentação e da composição química foram
realizadas no laboratório de Nutrição Animal, do Departamento de Ciências
Agrárias e Ambientais da Universidade Estadual de Santa Cruz, localizado no
município de Ilhéus, Bahia, e os resultados da composição química podem ser
visualizados na Tabela 2. As características fermentativas dos alimentos foram
realizadas na Embrapa gado de leite de Coronel Pacheco.

26
Tabela 1. Composição bromatológica dos frutos e feno de Brachiaria brizantha.
ACA CAC CUP FENO GRA JEN MAG
MS % 33,53 12,28 22,91 - 15,00 24,45 34,83
MM %MS 3,30 8,90 4,59 10,78 4,74 2,90 3,26
EE
35,43 2,16 32,13 0,88 4,21 3,80 1,43
PB
5,71 5,12 8,84 11,7 8,79 3,27 3,36
PIDN
1,90 3,65 1,74 3,85 2,39 1,19 1,47
CIDN
1,93 2,69 2,10 5,16 2,16 1,91 1,60
FDNcp
25,65 50,49 26,65 62,48 40,90 25,25 49,26
PIDA
0,93 3,60 1,98 0,22 0,97 0,86 2,62
CIDA
1,82 2,21 2,13 6,60 2,28 2,50 2,52
FDAcp
16,66 39,9 18,48 28,67 32,25 21,64 44,66
CHT
55,55 83,82 54,44 76,64 82,26 90,03 91,96
CNFcp
29,90 33,33 27,79 14,16 41,36 64,78 42,70
EBKcal 6,76 4,50 6,34 4,52 4,92 4,51 4,95
ACA = amêndoa de cacau, CAC = casca de cacau, CUP = cupuaçu, GRA= graviola. JEN = jenipapo, MAG = mangostão, MS = matéria seca, MM = matéria mineral, EE=extrato etéreo, PB=proteína bruta, PIDN= proteína insolúvel em detergente neutro, CIDN = cinzas insolúvel em detergente neutro, FDNcp = fibra em detergente neutro corrigido para cinzas e proteína, NIDA = nitrogênio insolúvel em detergente ácido, CIDA = cinzas insolúvel em detergente ácido, FDAcp = fibra em detergente ácido corrigido para cinzas e proteína, CHT = carboidratos totais, CNFcp = carboidratos não fibrosos corrigido para cinzas e proteínas, EB = energia bruta.
4.2. Características fermentativas dos alimentos
Foram conduzidos dois ensaios: o primeiro para avaliação da cinética de
fermentação ruminal e o segundo para avaliar o efeito na produção de gases, pH
e metabólitos.
Foi utilizada a técnica de produção de gases in vitro para mensuração de
metano entérico onde o ambiente ruminal é simulado através de uma amostra de
líquido ruminal em cultura contínua ou em frascos de fermentação
Na determinação de N-NH3liq (%), 5mL do líquido a partir de cada frasco
da incubação foram transferidos para frascos de coleta para análises laboratorial,
acrescido de 6 gotas de ácido sulfúrico 50% (H2SO4). Destes, 1,6 mL a partir de
cada frasco foi transferido para tubos de 2 mL de micro- centrifugação e
centrifugada a 14.000 xg durante 10 min a 4º C (Spectrafuse 16M , National
Labnet Co. , Edison, NJ, EUA ) para obtenção de partículas em suspensão e
precipitado de proteínas. O sobrenadante foi transferido para tubos de 2 mL de
micro-centrifugação e analisado para o N-NH3liq.
A análise de ácidos graxos de cadeia curta (AGCC) foi realizada com 5mL
do liquido utilizado na incubação acrescido de 1mL de ácido metafosfórico (HPO3)

27
a 20%, uma sub-amostra de 1,5 mL foi recolhida, e centrifugada como
anteriormente descrito para a análise N-NH3liq. O sobrenadante foi congelado a -
20º C até serem analisadas para as concentrações de AGCC (Holtshausen et al. ,
2009). A determinação dos ácidos acéticos, butírico e propiônico foram realizadas
por meio de cromatografia líquida de alto desempenho (HPLC) Waters 2695
equipado com Detector PAD 2998 (Photodiode Array Detector), com sistema de
separação constituído de pré-coluna e coluna de fase reversa ambas C18 ODS
80A (150 x 4,6 mm x 5 µm). As condições de análise foram: Fase móvel com
mistura isocrática constituída de 90% de solução aquosa ácida pH 2,45-2,55 e 10
% de metanol, temperatura do forno de 30 °C, volume de injeção de amostra de
20 microlitros, corrida com 12 minutos e detector com comprimento de onda de
excitação em 210 nm.
A proporção de metano no gás produzido pela fermentação das amostras
foi conduzida na Embrapa Gado de Leite- Juiz de Fora – MG, mensurada para os
tempos 12 e 24 horas de incubação. Foi utilizada a técnica in vitro semi-
automática de produção de gases, segundo Maurício et al. (1999). A medição dos
gases foi realizada com transdutor de pressão (DPI 705) e a conversão dos
valores de libra por polegada quadrada (PSI) em volume foi realizada a partir da
equação Volume = -0,0171pressão2 + 3,0926pressão + 0,0078; R² = 0,9877,
estabelecida para as condições locais do laboratório. Nos tempos de 12 e 24 hs
foram coletadas amostras de gases em exetaineres para a determinação da
concentração de metano em cromatógrafo a gás. O equipamento foi equipado
com duas válvulas de seis vias, sendo uma utilizada para o sistema amostrador
interligada a um loop de 0,5 mL. A outra, como seletora, permitindo aos
constituintes passarem ou não pela segunda coluna.
Injetor tipo split-splitless utilizado no modo split a 50:1 a uma temperatura
de 120 °C. O sistema de separação e constituído de duas colunas: uma HP-
Plot/Q 30m x 0,530 mm x 40,0 µm; a outra HP-Molesieve 30m x 0,530mm x 25,0
µm, utilizando H2 como gás de arraste a um fluxo de 7mL/min. O sistema de
deteção foi constituído por: Detector ECD – condições: 200°C aquecimento; 25
mL/min fluxo de referência; 3 mL/min fluxo de complementação(H2); 10 mL/min
coluna+complementação constante. Detector FID – Condições: 300°C
aquecimento; 10 mL/min fluxo H2; 400 mL/min fluxo ar sintético; 20 mL/min fluxo

28
complementação. Metanador a 375 °C aquecimento.
A temperatura do forno foi mantida a 50 °C por 4,5 minutos tempo
necessário para a eluição dos constituintes desejados. Pós análise, iniciou-se
uma rampa de aquecimento a 60 °C/min. até 250°C. Pós corrida a temperatura foi
mantida a 250°C por 2 minutos.
A calibração do cromatógrafo foi realizada com padrões de referência,
certificados pela Linde nas concentrações CH4 : 5,05; 10,2; 14,7; 20,1 e CO2:
20,2; 39,7; 58,3; 79,9 %.
4.3. Parâmetros de cinética de fermentação e técnica semi-automática de produção de gases in vitro
Para a obtenção do inoculo da incubação in vitro foram utilizados dois
bovinos machos mestiços, castrados, canulados no rúmen, com peso médio de
400 kg. Os animais receberam dieta contendo 70% de volumoso (capim
Brachiaria decumbens) e 30% de concentrado. O líquido ruminal foi retirado
manualmente pela manhã, antes do fornecimento da dieta, de várias partes do
rúmen, e armazenado em garrafa térmica previamente aquecida (39ºC) e
imediatamente levada para o laboratório. O líquido ruminal foi filtrado, passando
por duas camadas de gazes de algodão. Posteriormente, foi adicionado a
solução tampão e o líquido de rúmen respeitando a relação 2:1 (v/v), sob injeção
contínua de CO2;
Para as incubações foram utilizados 300 mg de amostra combinada
(volumoso de brachiaria brizanta e substrato (fruto)) e frascos de vidro (50 mL) de
acordo com procedimento descrito por Schofield et al., (1994). A solução tampão,
macro e micromineral, descrita por Menke e Steingass (1988), foram preparados
duas horas antes à incubação, sendo mantida aquecida a 39ºC, sob gaseificação
contínua com CO2 utilizando um termômetro imerso na solução, e
homogeneização com agitador do tipo “bailarina”. O pH da solução foi monitorado
constantemente até atingir valor de 6,8. A cada f r a sco foram adicionados
28,125 mL de solução tampão e 3,125 mL do inoculo ruminal.
A pressão (psi= pressão por polegada quadrada) originada dos gases
acumulados na parte superior dos frascos foi medida utilizando um transdutor de

29
pressão tipo “Press DATA 800” nos tempos 1, 2, 3, 4, 6, 8,10, 12, 14, 17, 20, 24,
28, 34, 48, 72, 96, 120 horas após a inoculação. Os resultados foram corrigidos
para o branco (frasco contendo fluido ruminal com meio de cultura, sem a
presença de amostra) e comparados ao padrão (feno de capim Brachiaria
brizantha), para validação do procedimento metodológico.
A equação de regressão utilizada para a conversão de pressão (P) em
volume foi: (V) (mL)= 0,04755 + 1,9754P + 0,01407P2, (R2 = 0,99), em que:
V= volume de gases produzido e; P= pressão em psi.
Esta equação foi estabelecida conforme Santos et al. (2010) para as
condições do laboratório de nutrição animal da UESC, seguindo metodologia
proposta por Maurício et al. (2003). As variáveis relacionadas à cinética de
fermentação ruminal in vitro foram estimadas pelo modelo bicompartimental
proposto por Schofield et al., (1994):
V = VFCNF / (1 + exp (2 - 4*kdCNF*(T - L))) + VFCF / (1 + exp(2 - 4*kdCF*(T -
L))),
em que: VFCNF equivale ao volume máximo dos gases da fração dos
CNF; kdCNF, à taxa de degradação (h-1) desta mesma fração (CNF); VFCF, ao
volume máximo de gás da fração dos CF; kdCF, à taxa de degradação (h-1) dos
CF; e T e L, aos tempos de incubação (horas) e à latência (horas),
respectivamente.
4.4. Análise estatística
Utilizou-se o delineamento experimental inteiramente casualizado (DIC),
com cada alimento testado isoladamente. Os dados foram analisados pelos
procedimentos da análise de variância e de regressão. Realizou-se o estudo para
verificar se as pressuposições de distribuição normal de aditividade e de
homocedasticidade dos dados foram atendidas.
Quanto aos fatores quantitativos (níveis de substituição do feno pelo
alimento alternativo testado), procedeu-se à análise de regressão polinomial.
Testou-se o modelo de regressão linear simples (Ŷ= βo + β1x) e, em seguida, o
modelo de segundo grau (Ŷ= βo + β1x + β2x2), em que se considerou “Ŷ” como

30
variável dependente e “x” como variável independente, além de “βo, β1 e β2”,
como coeficientes médio, de efeito linear e de efeito quadrático, respectivamente.
Diante da semelhança estatística de dois modelos, optou-se pelo de menor
ordem e àqueles cuja significância das variáveis que compunham o modelo foi
significativa. Como medida de precisão do modelo, utilizou-se o coeficiente de
determinação (R2/r2). Foram empregados como ferramenta de auxílio os
procedimentos PROC GLM e PROC REG, do programa SAS (Statistical Analysis
System, 2008, versão 9.2.). Em todos os procedimentos estatísticos, adotou-se
0,05 como nível crítico de probabilidade para o erro tipo I.
5. RESULTADOS E DISCUSSÃO
5.1. Cinética de fermentação ruminal in vitro dos carboidratos
Entre as frutas tropicais utilizadas, apenas o jenipapo proporcionou
aumento (P<0,05) no volume final dos CNF (VFCNF) (Tabela 3), representando até
134,2% de aumento em relação aos valores observados entre o feno e o nível de
50% de substituição, conseqüência da maior concentração de CHT e CNF desta
fruta (Tabela 2). Isto indicou que durante o processo fermentativo, à medida que
aumentava os níveis de jenipapo, aumentou-se também as fontes de rápida
disponibilidade de energia para os microrganismos ruminais, sendo esta fração a
que suporta o crescimento inicial de microorganismos.
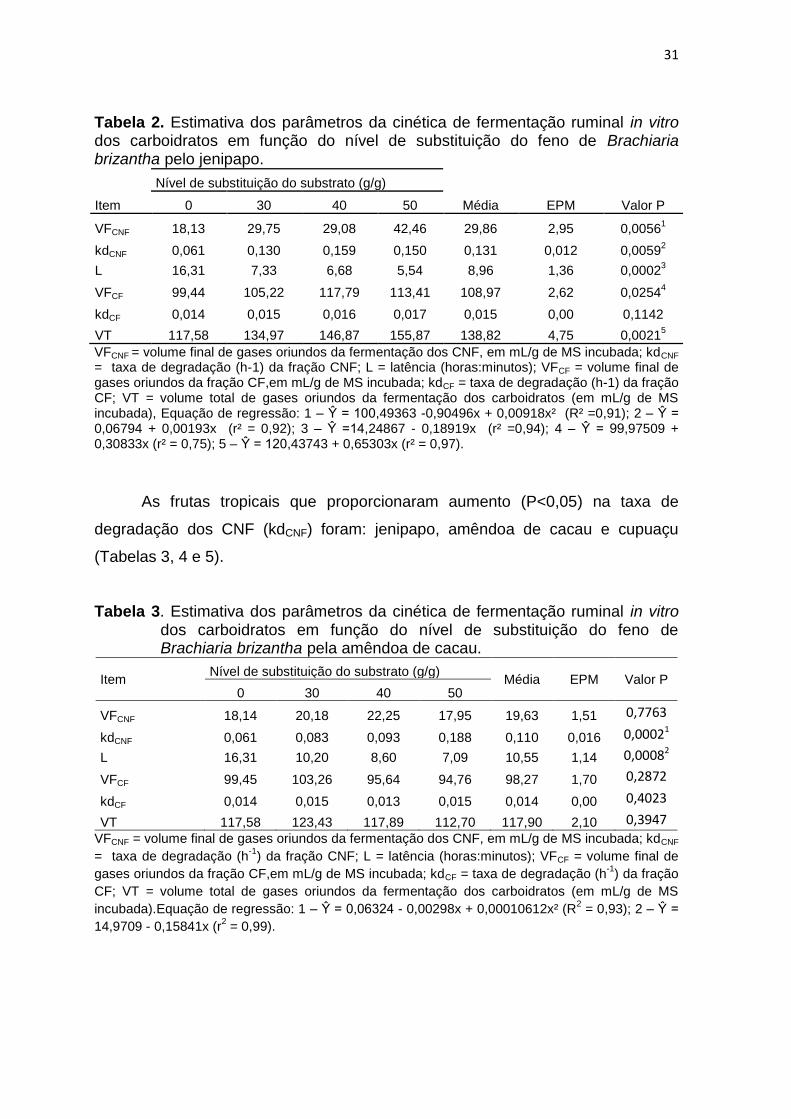
31
Tabela 2. Estimativa dos parâmetros da cinética de fermentação ruminal in vitro dos carboidratos em função do nível de substituição do feno de Brachiaria brizantha pelo jenipapo.
Item
Nível de substituição do substrato (g/g)
Média EPM Valor P 0 30 40 50
VFCNF 18,13 29,75 29,08 42,46 29,86 2,95 0,00561
kdCNF 0,061 0,130 0,159 0,150 0,131 0,012 0,00592
L 16,31 7,33 6,68 5,54 8,96 1,36 0,00023
VFCF 99,44 105,22 117,79 113,41 108,97 2,62 0,02544
kdCF 0,014 0,015 0,016 0,017 0,015 0,00 0,1142
VT 117,58 134,97 146,87 155,87 138,82 4,75 0,00215
VFCNF = volume final de gases oriundos da fermentação dos CNF, em mL/g de MS incubada; kdCNF = taxa de degradação (h-1) da fração CNF; L = latência (horas:minutos); VFCF = volume final de gases oriundos da fração CF,em mL/g de MS incubada; kdCF = taxa de degradação (h-1) da fração CF; VT = volume total de gases oriundos da fermentação dos carboidratos (em mL/g de MS incubada), Equação de regressão: 1 – Ŷ = 100,49363 -0,90496x + 0,00918x² (R² =0,91); 2 – Ŷ = 0,06794 + 0,00193x (r² = 0,92); 3 – Ŷ =14,24867 - 0,18919x (r² =0,94); 4 – Ŷ = 99,97509 + 0,30833x (r² = 0,75); 5 – Ŷ = 120,43743 + 0,65303x (r² = 0,97).
As frutas tropicais que proporcionaram aumento (P<0,05) na taxa de
degradação dos CNF (kdCNF) foram: jenipapo, amêndoa de cacau e cupuaçu
(Tabelas 3, 4 e 5).
Tabela 3. Estimativa dos parâmetros da cinética de fermentação ruminal in vitro dos carboidratos em função do nível de substituição do feno de Brachiaria brizantha pela amêndoa de cacau.
Item Nível de substituição do substrato (g/g)
Média EPM Valor P 0 30 40 50
VFCNF 18,14 20,18 22,25 17,95 19,63 1,51 0,7763
kdCNF 0,061 0,083 0,093 0,188 0,110 0,016 0,00021
L 16,31 10,20 8,60 7,09 10,55 1,14 0,00082
VFCF 99,45 103,26 95,64 94,76 98,27 1,70 0,2872
kdCF 0,014 0,015 0,013 0,015 0,014 0,00 0,4023
VT 117,58 123,43 117,89 112,70 117,90 2,10 0,3947
VFCNF = volume final de gases oriundos da fermentação dos CNF, em mL/g de MS incubada; kdCNF
= taxa de degradação (h-1
) da fração CNF; L = latência (horas:minutos); VFCF = volume final de
gases oriundos da fração CF,em mL/g de MS incubada; kdCF = taxa de degradação (h-1
) da fração
CF; VT = volume total de gases oriundos da fermentação dos carboidratos (em mL/g de MS
incubada).Equação de regressão: 1 – Ŷ = 0,06324 - 0,00298x + 0,00010612x² (R2 = 0,93); 2 – Ŷ =
14,9709 - 0,15841x (r2 = 0,99).
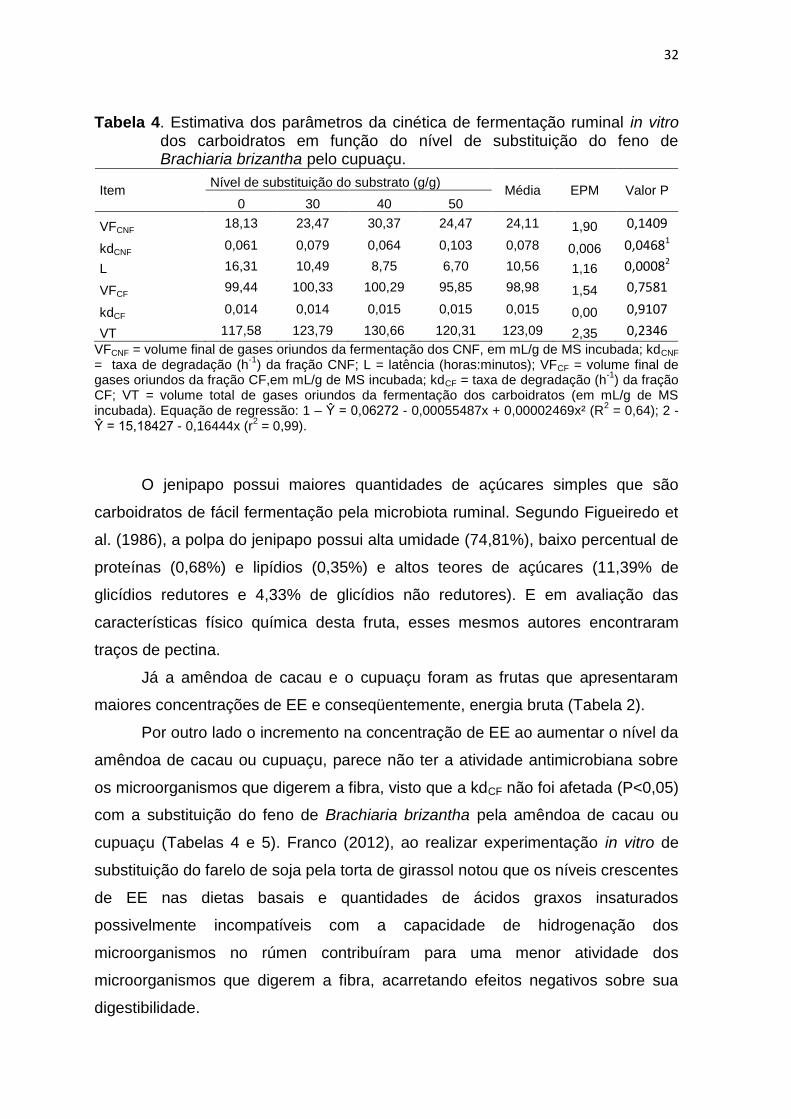
32
Tabela 4. Estimativa dos parâmetros da cinética de fermentação ruminal in vitro dos carboidratos em função do nível de substituição do feno de Brachiaria brizantha pelo cupuaçu.
Item Nível de substituição do substrato (g/g)
Média EPM Valor P 0 30 40 50
VFCNF 18,13 23,47 30,37 24,47 24,11 1,90 0,1409
kdCNF 0,061 0,079 0,064 0,103 0,078 0,006 0,04681
L 16,31 10,49 8,75 6,70 10,56 1,16 0,00082
VFCF 99,44 100,33 100,29 95,85 98,98 1,54 0,7581
kdCF 0,014 0,014 0,015 0,015 0,015 0,00 0,9107
VT 117,58 123,79 130,66 120,31 123,09 2,35 0,2346
VFCNF = volume final de gases oriundos da fermentação dos CNF, em mL/g de MS incubada; kdCNF = taxa de degradação (h
-1) da fração CNF; L = latência (horas:minutos); VFCF = volume final de
gases oriundos da fração CF,em mL/g de MS incubada; kdCF = taxa de degradação (h-1
) da fração CF; VT = volume total de gases oriundos da fermentação dos carboidratos (em mL/g de MS incubada). Equação de regressão: 1 – Ŷ = 0,06272 - 0,00055487x + 0,00002469x² (R
2 = 0,64); 2 -
Ŷ = 15,18427 - 0,16444x (r2 = 0,99).
O jenipapo possui maiores quantidades de açúcares simples que são
carboidratos de fácil fermentação pela microbiota ruminal. Segundo Figueiredo et
al. (1986), a polpa do jenipapo possui alta umidade (74,81%), baixo percentual de
proteínas (0,68%) e lipídios (0,35%) e altos teores de açúcares (11,39% de
glicídios redutores e 4,33% de glicídios não redutores). E em avaliação das
características físico química desta fruta, esses mesmos autores encontraram
traços de pectina.
Já a amêndoa de cacau e o cupuaçu foram as frutas que apresentaram
maiores concentrações de EE e conseqüentemente, energia bruta (Tabela 2).
Por outro lado o incremento na concentração de EE ao aumentar o nível da
amêndoa de cacau ou cupuaçu, parece não ter a atividade antimicrobiana sobre
os microorganismos que digerem a fibra, visto que a kdCF não foi afetada (P<0,05)
com a substituição do feno de Brachiaria brizantha pela amêndoa de cacau ou
cupuaçu (Tabelas 4 e 5). Franco (2012), ao realizar experimentação in vitro de
substituição do farelo de soja pela torta de girassol notou que os níveis crescentes
de EE nas dietas basais e quantidades de ácidos graxos insaturados
possivelmente incompatíveis com a capacidade de hidrogenação dos
microorganismos no rúmen contribuíram para uma menor atividade dos
microorganismos que digerem a fibra, acarretando efeitos negativos sobre sua
digestibilidade.
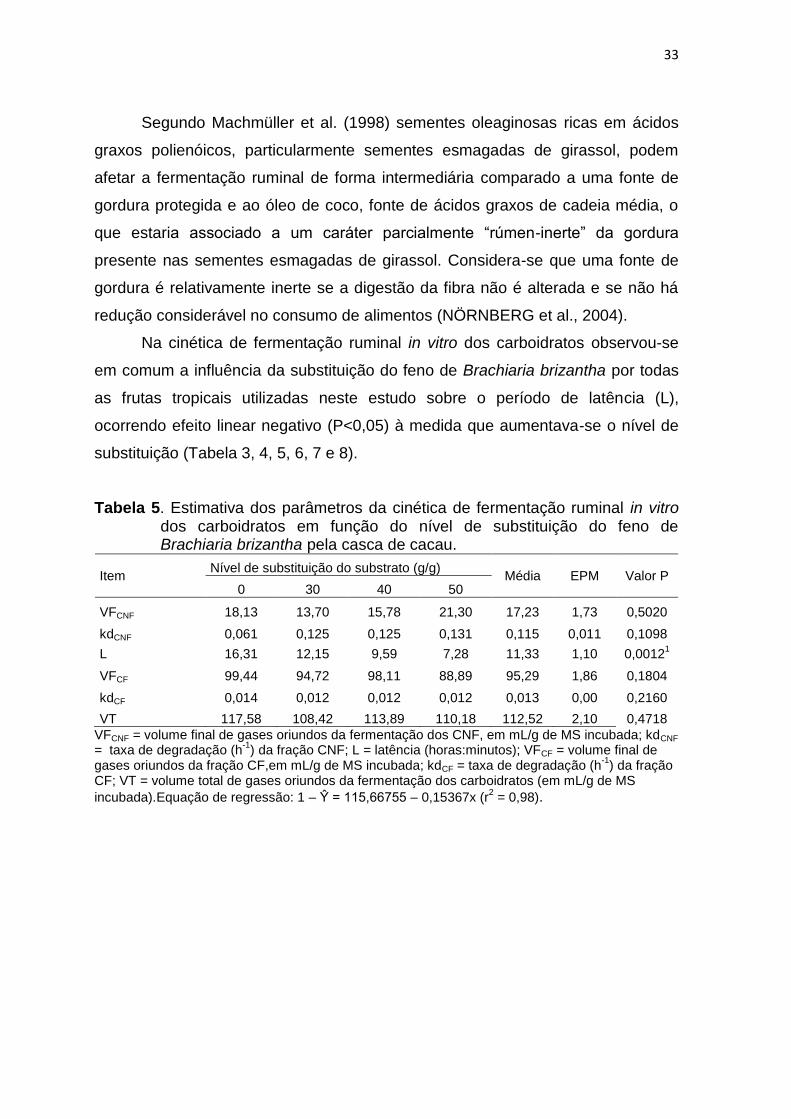
33
Segundo Machmüller et al. (1998) sementes oleaginosas ricas em ácidos
graxos polienóicos, particularmente sementes esmagadas de girassol, podem
afetar a fermentação ruminal de forma intermediária comparado a uma fonte de
gordura protegida e ao óleo de coco, fonte de ácidos graxos de cadeia média, o
que estaria associado a um caráter parcialmente “rúmen-inerte” da gordura
presente nas sementes esmagadas de girassol. Considera-se que uma fonte de
gordura é relativamente inerte se a digestão da fibra não é alterada e se não há
redução considerável no consumo de alimentos (NÖRNBERG et al., 2004).
Na cinética de fermentação ruminal in vitro dos carboidratos observou-se
em comum a influência da substituição do feno de Brachiaria brizantha por todas
as frutas tropicais utilizadas neste estudo sobre o período de latência (L),
ocorrendo efeito linear negativo (P<0,05) à medida que aumentava-se o nível de
substituição (Tabela 3, 4, 5, 6, 7 e 8).
Tabela 5. Estimativa dos parâmetros da cinética de fermentação ruminal in vitro dos carboidratos em função do nível de substituição do feno de Brachiaria brizantha pela casca de cacau.
Item Nível de substituição do substrato (g/g)
Média EPM Valor P 0 30 40 50
VFCNF 18,13 13,70 15,78 21,30 17,23 1,73 0,5020
kdCNF 0,061 0,125 0,125 0,131 0,115 0,011 0,1098
L 16,31 12,15 9,59 7,28 11,33 1,10 0,00121
VFCF 99,44 94,72 98,11 88,89 95,29 1,86 0,1804
kdCF 0,014 0,012 0,012 0,012 0,013 0,00 0,2160
VT 117,58 108,42 113,89 110,18 112,52 2,10 0,4718
VFCNF = volume final de gases oriundos da fermentação dos CNF, em mL/g de MS incubada; kdCNF = taxa de degradação (h
-1) da fração CNF; L = latência (horas:minutos); VFCF = volume final de
gases oriundos da fração CF,em mL/g de MS incubada; kdCF = taxa de degradação (h-1
) da fração CF; VT = volume total de gases oriundos da fermentação dos carboidratos (em mL/g de MS
incubada).Equação de regressão: 1 – Ŷ = 115,66755 – 0,15367x (r2 = 0,98).
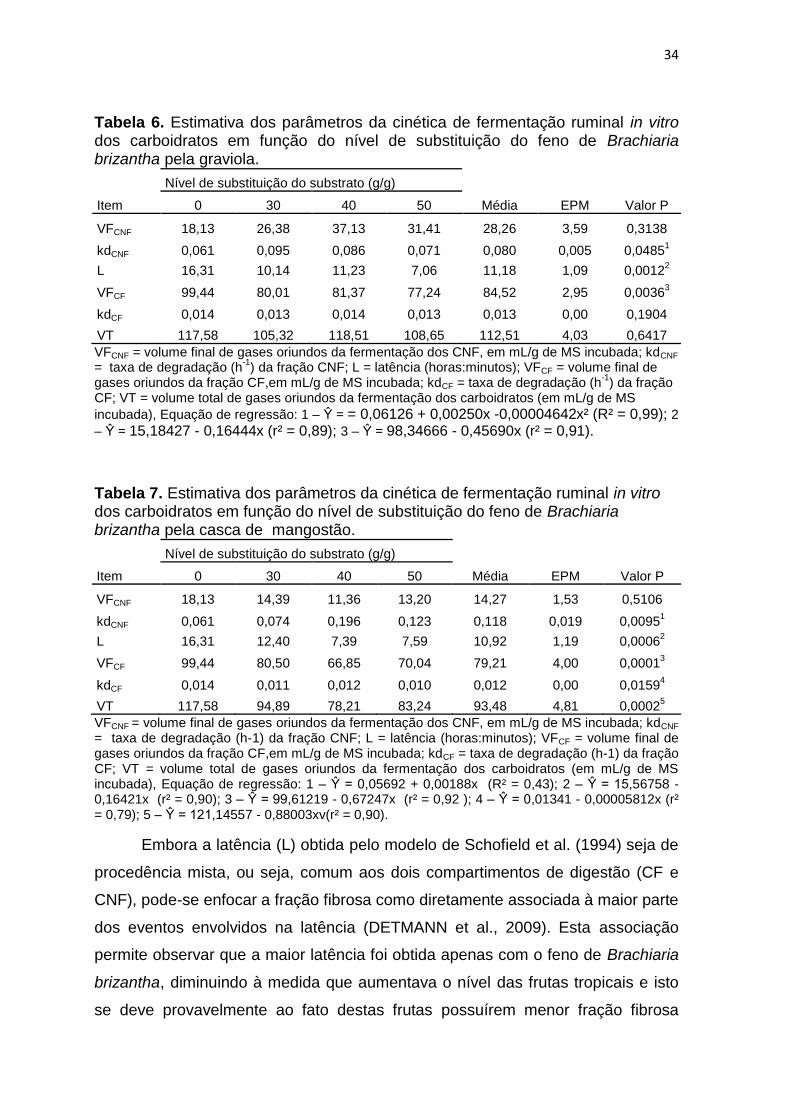
34
Tabela 6. Estimativa dos parâmetros da cinética de fermentação ruminal in vitro dos carboidratos em função do nível de substituição do feno de Brachiaria brizantha pela graviola.
Item
Nível de substituição do substrato (g/g)
Média EPM Valor P 0 30 40 50
VFCNF 18,13 26,38 37,13 31,41 28,26 3,59 0,3138
kdCNF 0,061 0,095 0,086 0,071 0,080 0,005 0,04851
L 16,31 10,14 11,23 7,06 11,18 1,09 0,00122
VFCF 99,44 80,01 81,37 77,24 84,52 2,95 0,00363
kdCF 0,014 0,013 0,014 0,013 0,013 0,00 0,1904
VT 117,58 105,32 118,51 108,65 112,51 4,03 0,6417
VFCNF = volume final de gases oriundos da fermentação dos CNF, em mL/g de MS incubada; kdCNF = taxa de degradação (h
-1) da fração CNF; L = latência (horas:minutos); VFCF = volume final de
gases oriundos da fração CF,em mL/g de MS incubada; kdCF = taxa de degradação (h-1
) da fração CF; VT = volume total de gases oriundos da fermentação dos carboidratos (em mL/g de MS
incubada), Equação de regressão: 1 – Ŷ = = 0,06126 + 0,00250x -0,00004642x² (R² = 0,99); 2
– Ŷ = 15,18427 - 0,16444x (r² = 0,89); 3 – Ŷ = 98,34666 - 0,45690x (r² = 0,91).
Tabela 7. Estimativa dos parâmetros da cinética de fermentação ruminal in vitro dos carboidratos em função do nível de substituição do feno de Brachiaria brizantha pela casca de mangostão.
Item
Nível de substituição do substrato (g/g)
Média EPM Valor P 0 30 40 50
VFCNF 18,13 14,39 11,36 13,20 14,27 1,53 0,5106
kdCNF 0,061 0,074 0,196 0,123 0,118 0,019 0,00951
L 16,31 12,40 7,39 7,59 10,92 1,19 0,00062
VFCF 99,44 80,50 66,85 70,04 79,21 4,00 0,00013
kdCF 0,014 0,011 0,012 0,010 0,012 0,00 0,01594
VT 117,58 94,89 78,21 83,24 93,48 4,81 0,00025
VFCNF = volume final de gases oriundos da fermentação dos CNF, em mL/g de MS incubada; kdCNF = taxa de degradação (h-1) da fração CNF; L = latência (horas:minutos); VFCF = volume final de gases oriundos da fração CF,em mL/g de MS incubada; kdCF = taxa de degradação (h-1) da fração CF; VT = volume total de gases oriundos da fermentação dos carboidratos (em mL/g de MS incubada), Equação de regressão: 1 – Ŷ = 0,05692 + 0,00188x (R² = 0,43); 2 – Ŷ = 15,56758 - 0,16421x (r² = 0,90); 3 – Ŷ = 99,61219 - 0,67247x (r² = 0,92 ); 4 – Ŷ = 0,01341 - 0,00005812x (r² = 0,79); 5 – Ŷ = 121,14557 - 0,88003xv(r² = 0,90).
Embora a latência (L) obtida pelo modelo de Schofield et al. (1994) seja de
procedência mista, ou seja, comum aos dois compartimentos de digestão (CF e
CNF), pode-se enfocar a fração fibrosa como diretamente associada à maior parte
dos eventos envolvidos na latência (DETMANN et al., 2009). Esta associação
permite observar que a maior latência foi obtida apenas com o feno de Brachiaria
brizantha, diminuindo à medida que aumentava o nível das frutas tropicais e isto
se deve provavelmente ao fato destas frutas possuírem menor fração fibrosa

35
comparativamente ao feno (Tabela 2). Entre as frutas a que proporcionou maior e
menor redução foram o jenipapo e casca do cacau, respectivamente, as quais
para cada nível de substituição reduziu 0,189 e 0,153 horas, respectivamente, da
latência.
Como o volume final dos CF (VFcf) superou à do VFcnf, independente da
fruta utilizada, esta foi a fonte de maior disponibilidade de energia à população
microbiana ruminal. Isto porque mesmo no jenipapo que foi a fruta com maior
concentração de CNF o nível de 50% de substituição não proporcionou
quantitativamente superioridade entre a concentração de CNF (39,47% MS) e CF
(43,86% MS). O jenipapo foi a fruta que influenciou (P<0,05) no aumento do
volume de gases oriundo dos CF (Tabela 3), indicando possível efeito associativo
positivo, enquanto a graviola (Tabela 7) e o mangostão (Tabela 8) proporcionaram
efeito associativo negativo (P<0,05) à medida que existiu substituição do feno
pela fruta.
Tanto a graviola (32,25% MS) quanto a casca do mangostão (44,66% MS)
apresentaram maior concentração de FDAcp quando comparado ao feno (28,67%
MS), indicando existir fatores limitantes como a lignina, no aproveitamento da
fração fibrosa pela população microbiana.
Para o mangostão os impactos sobre a redução no VFcf foram maiores já
que além de ter sido a fruta de maior concentração de FDAcp, a combinação
existente entre baixa concentração de proteína (3,26% na MS) aliado ao alto valor
de PIDA nesta proteína (78% da PB) pode ter limitado a disponibilidade de N-NH3
à população de bactérias celulolíticas ao longo do tempo e resultou em
interferência negativa (P<0,05) no volume e na taxa de degradação dos CF (kdcf).
Os microorganismos fibrolíticos utilizam amônia como fonte de N para a síntese
de proteína microbiana (RUSSELL, 1992), no entanto, perdas de amônia podem
acontecer quando não existe compatibilidade da degradação ruminal entre fontes
de proteína e carboidratos (NOCEK; RUSSEL, 1998).
Diante das características apresentadas pela estimativa dos parâmetros da
cinética de fermentação ruminal in vitro, o jenipapo foi a única fruta que
possibilitou maior (P<0,05) volume final total (VFT), em conseqüência do aumento
na kdcnf, VFcnf e VFcf. No geral, as frutas melhoraram a fermentação ruminal e
estas melhoras podem interferir positivamente no desempenho e ter impactos

36
positivos para a mitigação de metano em relação à quantidade de produto (carne
e leite).
5.2. Produtos da fermentação ruminal in vitro
Para os produtos da fermentação ruminal in vitro, observou-se
comportamento semelhante entre as frutas: casca do cacau, cupuaçu e
mangostão (Tabelas 9, 10 e 11). Estas frutas interferiram negativamente (P<0,05)
na digestibilidade da MS (DIVMS). A casca de cacau e o mangostão
apresentaram maiores concentrações de FDAcp do que o feno e isto indica maior
presença de lignina que é um componente indigestível. Pires et al. (2009)
encontraram baixa DIVMS na casca de cacau e atribuíram a maiores valores de
lignina em relação aos outros produtos testado. Do mesmo modo, Carvalho et al.
(2007) verificaram redução na DIVMS de silagens de capim elefante acrescidas
de casca de cacau (0, 7, 14, 21 e 28%).
Tabela 9. Produtos da fermentação ruminal em função do nível de substituição do
feno de Brachiaria brizantha pela casca do cacau, por 24 horas de
incubação in vitro.
Item Nível de substituição do substrato (g/g)
Média EPM Valor P 0 30 40 50
DIVMS, g/g 41,57 32,33 31,56 31,19 36,63 1,38 0,00011
Volume de gases, mL/g MS 67,55 70,54 66,51 79,90 69,93 1,95 0,1191
CO2, mL/ g MS 32,88 35,39 32,57 34,99 33,60 1,21 0,8555
NH3, mL/g MS 18,51 26,13 24,73 31,97 23,06 2,06 0,0967
Acético, mL/g MS 46,12 51,41 58,58 48,94 49,55 1,80 0,0879
Propiônico, mL/g MS 21,57 25,42 23,35 22,27 22,62 0,57 0,0927
Butírico, mL/g MS 8,81 8,28 6,76 7,32 8,13 0,38 0,4039
A:P, mL/g MS 2,16 2,02 2,51 2,20 2,20 0,08 0,3755
pH 6,66 6,67 6,70 6,65 6,67 0,01 0,5377
EPM – Erro padrão da média; DIVMS – Digestibilidade in vitro da matéria seca, A:P – Proporção acético: propiônico, P < 0,05, Equações de regressão: 1 – Ŷ = 41,56587- 0,45051x + 0,00489x² (R² = 0,999).
37
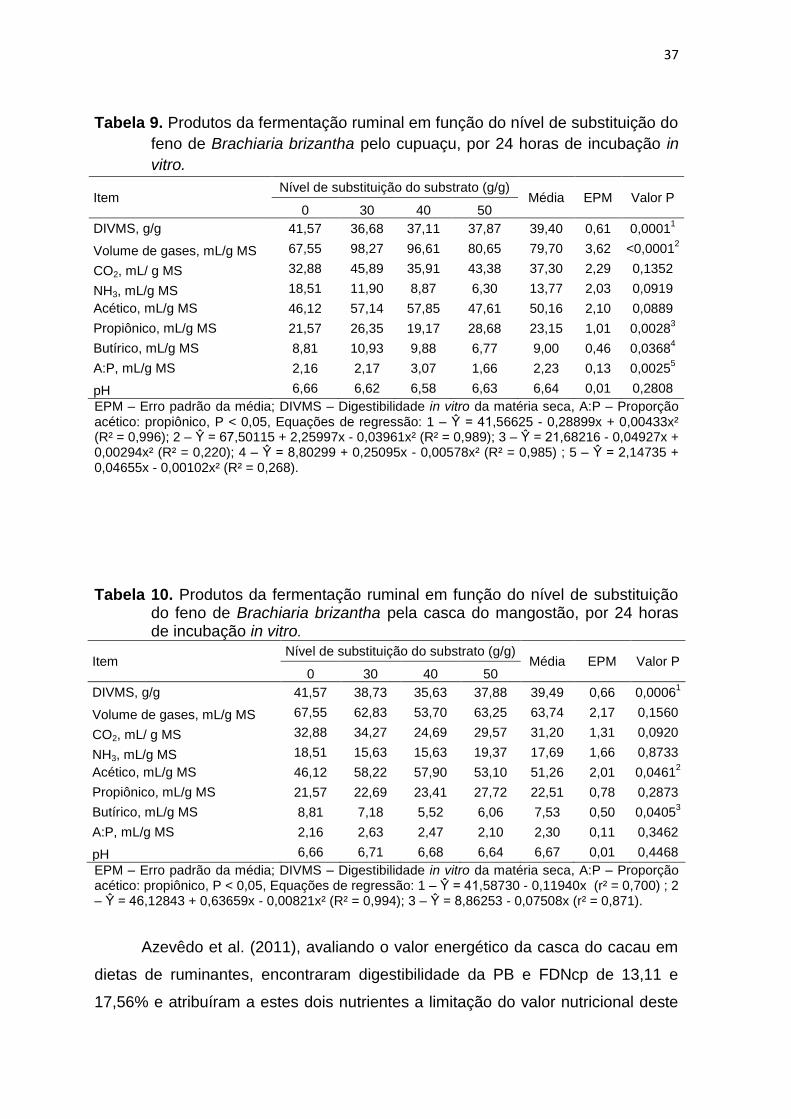
Tabela 9. Produtos da fermentação ruminal em função do nível de substituição do
feno de Brachiaria brizantha pelo cupuaçu, por 24 horas de incubação in
vitro.
Item Nível de substituição do substrato (g/g)
Média EPM Valor P 0 30 40 50
DIVMS, g/g 41,57 36,68 37,11 37,87 39,40 0,61 0,00011
Volume de gases, mL/g MS 67,55 98,27 96,61 80,65 79,70 3,62 <0,00012
CO2, mL/ g MS 32,88 45,89 35,91 43,38 37,30 2,29 0,1352
NH3, mL/g MS 18,51 11,90 8,87 6,30 13,77 2,03 0,0919
Acético, mL/g MS 46,12 57,14 57,85 47,61 50,16 2,10 0,0889
Propiônico, mL/g MS 21,57 26,35 19,17 28,68 23,15 1,01 0,00283
Butírico, mL/g MS 8,81 10,93 9,88 6,77 9,00 0,46 0,03684
A:P, mL/g MS 2,16 2,17 3,07 1,66 2,23 0,13 0,00255
pH 6,66 6,62 6,58 6,63 6,64 0,01 0,2808
EPM – Erro padrão da média; DIVMS – Digestibilidade in vitro da matéria seca, A:P – Proporção acético: propiônico, P < 0,05, Equações de regressão: 1 – Ŷ = 41,56625 - 0,28899x + 0,00433x² (R² = 0,996); 2 – Ŷ = 67,50115 + 2,25997x - 0,03961x² (R² = 0,989); 3 – Ŷ = 21,68216 - 0,04927x + 0,00294x² (R² = 0,220); 4 – Ŷ = 8,80299 + 0,25095x - 0,00578x² (R² = 0,985) ; 5 – Ŷ = 2,14735 + 0,04655x - 0,00102x² (R² = 0,268).
Tabela 10. Produtos da fermentação ruminal em função do nível de substituição do feno de Brachiaria brizantha pela casca do mangostão, por 24 horas de incubação in vitro.
Item Nível de substituição do substrato (g/g)
Média EPM Valor P 0 30 40 50
DIVMS, g/g 41,57 38,73 35,63 37,88 39,49 0,66 0,00061
Volume de gases, mL/g MS 67,55 62,83 53,70 63,25 63,74 2,17 0,1560
CO2, mL/ g MS 32,88 34,27 24,69 29,57 31,20 1,31 0,0920
NH3, mL/g MS 18,51 15,63 15,63 19,37 17,69 1,66 0,8733
Acético, mL/g MS 46,12 58,22 57,90 53,10 51,26 2,01 0,04612
Propiônico, mL/g MS 21,57 22,69 23,41 27,72 22,51 0,78 0,2873
Butírico, mL/g MS 8,81 7,18 5,52 6,06 7,53 0,50 0,04053
A:P, mL/g MS 2,16 2,63 2,47 2,10 2,30 0,11 0,3462
pH 6,66 6,71 6,68 6,64 6,67 0,01 0,4468
EPM – Erro padrão da média; DIVMS – Digestibilidade in vitro da matéria seca, A:P – Proporção acético: propiônico, P < 0,05, Equações de regressão: 1 – Ŷ = 41,58730 - 0,11940x (r² = 0,700) ; 2 – Ŷ = 46,12843 + 0,63659x - 0,00821x² (R² = 0,994); 3 – Ŷ = 8,86253 - 0,07508x (r² = 0,871).
Azevêdo et al. (2011), avaliando o valor energético da casca do cacau em
dietas de ruminantes, encontraram digestibilidade da PB e FDNcp de 13,11 e
17,56% e atribuíram a estes dois nutrientes a limitação do valor nutricional deste

38
alimento. Além disso, estes autores como também Silva et al. (2007) e Silva et al.
(2005) observaram baixa preferência e sobras no cocho ao fornecer este alimento
aos animais ruminantes. Marcondes et al. (2009) encontraram baixa
digestibilidade da PB na casca de cacau durante experimento de avaliação da
degradação ruminal e intestinal da proteína bruta em bovinos.
A casca de cacau e o cupuaçu possuem teobromina que possui ação
diurética e apresenta-se como um fator antinutricional podendo levar a intoxicação
e lesões cutâneas nos animais. Pires et al. (2005) observaram redução no
consumo de MS de novilhas alimentadas com inclusão de 50% de farelo de cacau
no concentrado e constataram lesões na pele, podendo ser conseqüência da
teobromina. Azevêdo et al. (2011) verificaram que os animais que receberam pela
primeira vez dietas com casca de cacau urinaram 38,4% a mais que os demais
animais. Pereira (2009) sugeriu que a teobromina possua algum efeito bactericida
ou bacteriostático sobre a população microbiana, a exemplo de outros compostos
presentes nos vegetais, comprometendo a digestão dos componentes da dieta.
Suchitra e Wanapat (2008), afirmam que a casca de mangostão é rica em
taninos condensados e saponinas (16 e 10% de MS, respectivamente). Dessa
forma, o prejuízo à DIVMS pode estar associado ao alto nível de inclusão da
casca de mangostão e conseqüentemente de taninos condensados na incubação.
Este resultado foi consistente com Ngamseng et al. (2006) e Poungchompu et al.
(2009) que relataram que a alimentação dietética com taninos e saponinas
provocam a diminuição da digestibilidade aparente.
De acordo com Thanh et al. (2012), taninos condensados são benéficos
para a fermentação no rúmen, quando presente em quantidade moderada (4 a
6% do total) nas dietas, contudo, as concentrações dietéticas elevadas (6-12%
MS) podem deprimir o consumo voluntário, a eficiência digestiva e produtividade
animal.
A amêndoa de cacau e o jenipapo apresentaram semelhanças de
comportamento para DIVMS, volume de gases, CO2, relação acetato:propionato
(A:P) (Tabelas 12 e 13).
39
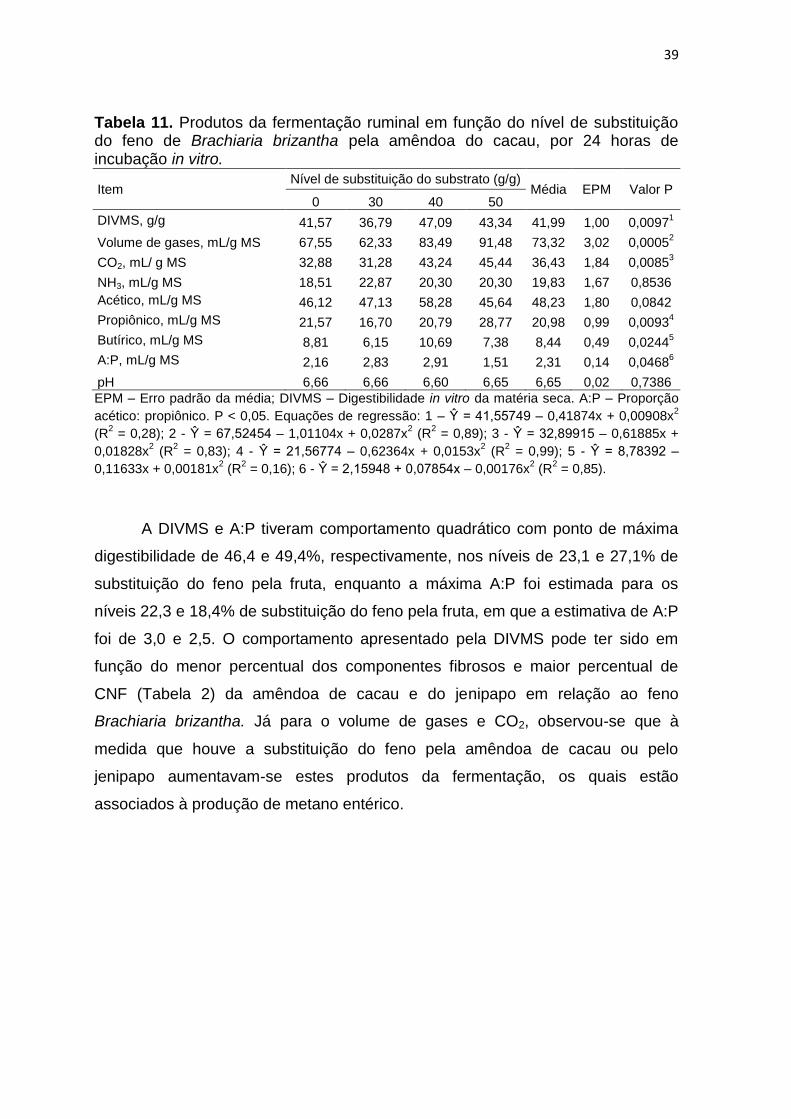
Tabela 11. Produtos da fermentação ruminal em função do nível de substituição do feno de Brachiaria brizantha pela amêndoa do cacau, por 24 horas de incubação in vitro.
Item Nível de substituição do substrato (g/g)
Média EPM Valor P 0 30 40 50
DIVMS, g/g 41,57 36,79 47,09 43,34 41,99 1,00 0,00971
Volume de gases, mL/g MS 67,55 62,33 83,49 91,48 73,32 3,02 0,00052
CO2, mL/ g MS 32,88 31,28 43,24 45,44 36,43 1,84 0,00853
NH3, mL/g MS 18,51 22,87 20,30 20,30 19,83 1,67 0,8536
Acético, mL/g MS 46,12 47,13 58,28 45,64 48,23 1,80 0,0842
Propiônico, mL/g MS 21,57 16,70 20,79 28,77 20,98 0,99 0,00934
Butírico, mL/g MS 8,81 6,15 10,69 7,38 8,44 0,49 0,02445
A:P, mL/g MS 2,16 2,83 2,91 1,51 2,31 0,14 0,04686
pH 6,66 6,66 6,60 6,65 6,65 0,02 0,7386
EPM – Erro padrão da média; DIVMS – Digestibilidade in vitro da matéria seca. A:P – Proporção
acético: propiônico. P < 0,05. Equações de regressão: 1 – Ŷ = 41,55749 – 0,41874x + 0,00908x2
(R2 = 0,28); 2 - Ŷ = 67,52454 – 1,01104x + 0,0287x
2 (R
2 = 0,89); 3 - Ŷ = 32,89915 – 0,61885x +
0,01828x2 (R
2 = 0,83); 4 - Ŷ = 21,56774 – 0,62364x + 0,0153x
2 (R
2 = 0,99); 5 - Ŷ = 8,78392 –
0,11633x + 0,00181x2 (R
2 = 0,16); 6 - Ŷ = 2,15948 + 0,07854x – 0,00176x
2 (R
2 = 0,85).
A DIVMS e A:P tiveram comportamento quadrático com ponto de máxima
digestibilidade de 46,4 e 49,4%, respectivamente, nos níveis de 23,1 e 27,1% de
substituição do feno pela fruta, enquanto a máxima A:P foi estimada para os
níveis 22,3 e 18,4% de substituição do feno pela fruta, em que a estimativa de A:P
foi de 3,0 e 2,5. O comportamento apresentado pela DIVMS pode ter sido em
função do menor percentual dos componentes fibrosos e maior percentual de
CNF (Tabela 2) da amêndoa de cacau e do jenipapo em relação ao feno
Brachiaria brizantha. Já para o volume de gases e CO2, observou-se que à
medida que houve a substituição do feno pela amêndoa de cacau ou pelo
jenipapo aumentavam-se estes produtos da fermentação, os quais estão
associados à produção de metano entérico.
40
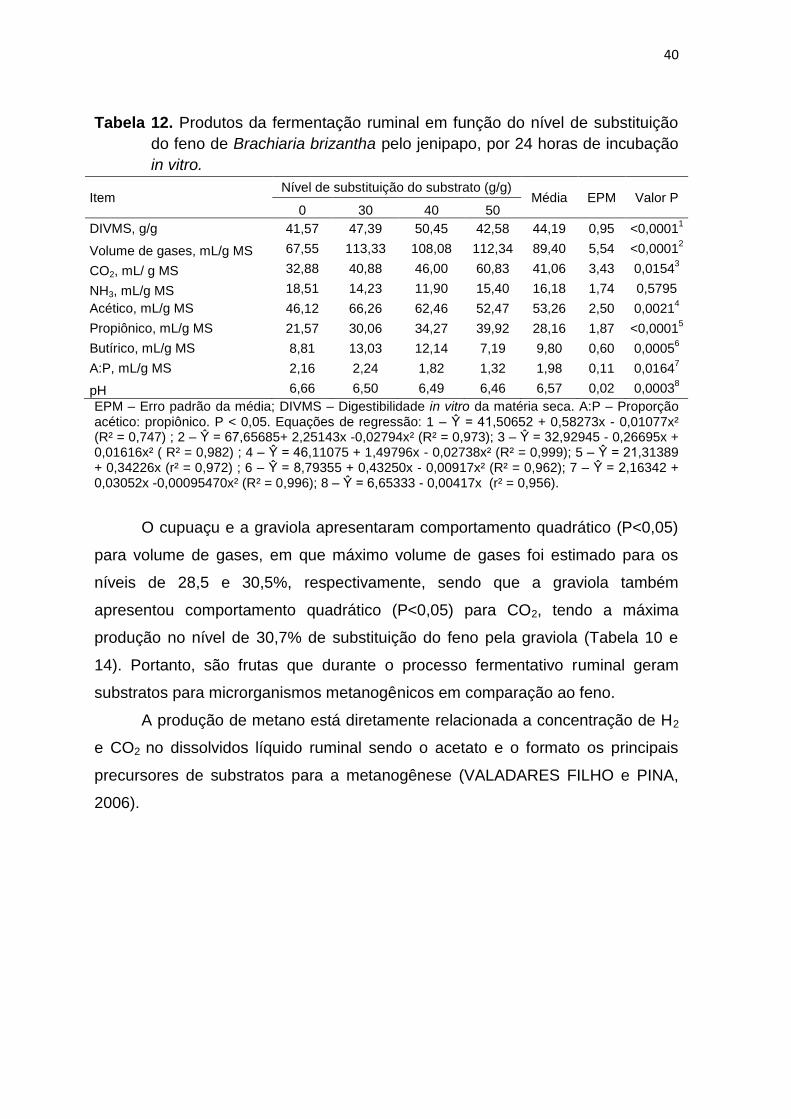
Tabela 12. Produtos da fermentação ruminal em função do nível de substituição
do feno de Brachiaria brizantha pelo jenipapo, por 24 horas de incubação
in vitro.
Item Nível de substituição do substrato (g/g)
Média EPM Valor P 0 30 40 50
DIVMS, g/g 41,57 47,39 50,45 42,58 44,19 0,95 <0,00011
Volume de gases, mL/g MS 67,55 113,33 108,08 112,34 89,40 5,54 <0,00012
CO2, mL/ g MS 32,88 40,88 46,00 60,83 41,06 3,43 0,01543
NH3, mL/g MS 18,51 14,23 11,90 15,40 16,18 1,74 0,5795
Acético, mL/g MS 46,12 66,26 62,46 52,47 53,26 2,50 0,00214
Propiônico, mL/g MS 21,57 30,06 34,27 39,92 28,16 1,87 <0,00015
Butírico, mL/g MS 8,81 13,03 12,14 7,19 9,80 0,60 0,00056
A:P, mL/g MS 2,16 2,24 1,82 1,32 1,98 0,11 0,01647
pH 6,66 6,50 6,49 6,46 6,57 0,02 0,00038
EPM – Erro padrão da média; DIVMS – Digestibilidade in vitro da matéria seca. A:P – Proporção acético: propiônico. P < 0,05. Equações de regressão: 1 – Ŷ = 41,50652 + 0,58273x - 0,01077x² (R² = 0,747) ; 2 – Ŷ = 67,65685+ 2,25143x -0,02794x² (R² = 0,973); 3 – Ŷ = 32,92945 - 0,26695x + 0,01616x² ( R² = 0,982) ; 4 – Ŷ = 46,11075 + 1,49796x - 0,02738x² (R² = 0,999); 5 – Ŷ = 21,31389 + 0,34226x (r² = 0,972) ; 6 – Ŷ = 8,79355 + 0,43250x - 0,00917x² (R² = 0,962); 7 – Ŷ = 2,16342 + 0,03052x -0,00095470x² (R² = 0,996); 8 – Ŷ = 6,65333 - 0,00417x (r² = 0,956).
O cupuaçu e a graviola apresentaram comportamento quadrático (P<0,05)
para volume de gases, em que máximo volume de gases foi estimado para os
níveis de 28,5 e 30,5%, respectivamente, sendo que a graviola também
apresentou comportamento quadrático (P<0,05) para CO2, tendo a máxima
produção no nível de 30,7% de substituição do feno pela graviola (Tabela 10 e
14). Portanto, são frutas que durante o processo fermentativo ruminal geram
substratos para microrganismos metanogênicos em comparação ao feno.
A produção de metano está diretamente relacionada a concentração de H2
e CO2 no dissolvidos líquido ruminal sendo o acetato e o formato os principais
precursores de substratos para a metanogênese (VALADARES FILHO e PINA,
2006).
41
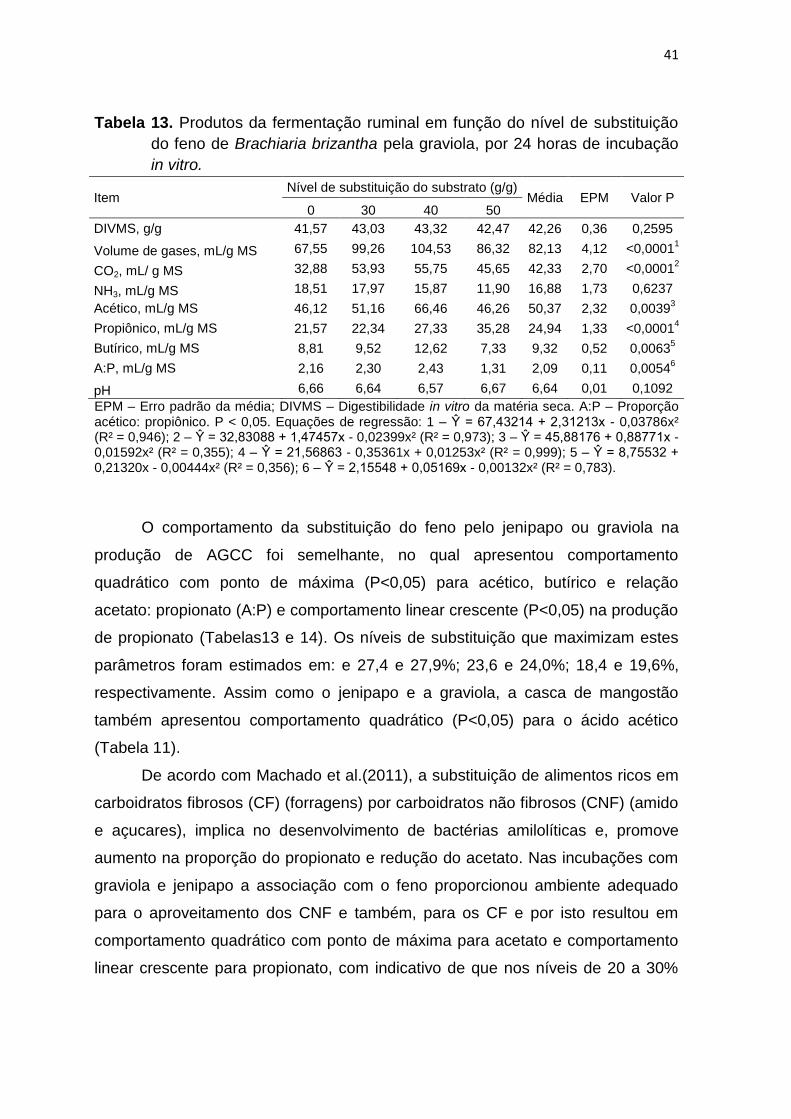
Tabela 13. Produtos da fermentação ruminal em função do nível de substituição
do feno de Brachiaria brizantha pela graviola, por 24 horas de incubação
in vitro.
Item Nível de substituição do substrato (g/g)
Média EPM Valor P 0 30 40 50
DIVMS, g/g 41,57 43,03 43,32 42,47 42,26 0,36 0,2595
Volume de gases, mL/g MS 67,55 99,26 104,53 86,32 82,13 4,12 <0,00011
CO2, mL/ g MS 32,88 53,93 55,75 45,65 42,33 2,70 <0,00012
NH3, mL/g MS 18,51 17,97 15,87 11,90 16,88 1,73 0,6237
Acético, mL/g MS 46,12 51,16 66,46 46,26 50,37 2,32 0,00393
Propiônico, mL/g MS 21,57 22,34 27,33 35,28 24,94 1,33 <0,00014
Butírico, mL/g MS 8,81 9,52 12,62 7,33 9,32 0,52 0,00635
A:P, mL/g MS 2,16 2,30 2,43 1,31 2,09 0,11 0,00546
pH 6,66 6,64 6,57 6,67 6,64 0,01 0,1092
EPM – Erro padrão da média; DIVMS – Digestibilidade in vitro da matéria seca. A:P – Proporção acético: propiônico. P < 0,05. Equações de regressão: 1 – Ŷ = 67,43214 + 2,31213x - 0,03786x² (R² = 0,946); 2 – Ŷ = 32,83088 + 1,47457x - 0,02399x² (R² = 0,973); 3 – Ŷ = 45,88176 + 0,88771x -0,01592x² (R² = 0,355); 4 – Ŷ = 21,56863 - 0,35361x + 0,01253x² (R² = 0,999); 5 – Ŷ = 8,75532 + 0,21320x - 0,00444x² (R² = 0,356); 6 – Ŷ = 2,15548 + 0,05169x - 0,00132x² (R² = 0,783).
O comportamento da substituição do feno pelo jenipapo ou graviola na
produção de AGCC foi semelhante, no qual apresentou comportamento
quadrático com ponto de máxima (P<0,05) para acético, butírico e relação
acetato: propionato (A:P) e comportamento linear crescente (P<0,05) na produção
de propionato (Tabelas13 e 14). Os níveis de substituição que maximizam estes
parâmetros foram estimados em: e 27,4 e 27,9%; 23,6 e 24,0%; 18,4 e 19,6%,
respectivamente. Assim como o jenipapo e a graviola, a casca de mangostão
também apresentou comportamento quadrático (P<0,05) para o ácido acético
(Tabela 11).
De acordo com Machado et al.(2011), a substituição de alimentos ricos em
carboidratos fibrosos (CF) (forragens) por carboidratos não fibrosos (CNF) (amido
e açucares), implica no desenvolvimento de bactérias amilolíticas e, promove
aumento na proporção do propionato e redução do acetato. Nas incubações com
graviola e jenipapo a associação com o feno proporcionou ambiente adequado
para o aproveitamento dos CNF e também, para os CF e por isto resultou em
comportamento quadrático com ponto de máxima para acetato e comportamento
linear crescente para propionato, com indicativo de que nos níveis de 20 a 30%

42
de substituição ocorreria a máxima disponibilização de substrato para formação
de metano.
Para a substituição do feno pelas frutas cupuaçu e mangostão observou-se
diminuição linear (P<0,05) na produção de ácido butírico (Tabelas 10 e 11),
enquanto para a amêndoa do cacau observou-se comportamento quadrático
(Tabela12) com ponto de mínima produção no nível de substituição de 32,1%.
Manasri et al. (2012), também observaram aumento de ácido propiônico
em experimento in vivo com casca de mangostão.
Suchitra e Wanapat (2008), em experimento in vivo com vacas leiteiras,
utilizaram a casca de mangostão com níveis de inclusão de 100 e 200g/d com e
sem adição de óleo de girassol e de côco e os seus resultados foram
semelhantes aos encontrados neste trabalho, com aumento de ácido propiônico
pra qualquer nível de inclusão da casca de mangostão.
A substituição do feno pela amêndoa de cacau e cupuaçu apresentaram
comportamento semelhante para A:P com comportamento quadrático com ponto
de máximo 22,3 e 22,8%, respectivamente, porém, no que diz respeito ao ácido
propiônico, a amêndoa de cacau apresentou comportamento quadrático com
ponto de mínimo 20,4% e o cupuaçu comportamento sigmóide.

43
CH4, mL/g MS digestível
4 6 8 10 12 14 16 18
Nív
el d
e su
bst
ituiç
ão,
g/g
0
30
40
50
12 h
24 h
CH4, mL/ g MS digestível
4 6 8 10 12 14 16 18
Nív
el d
e su
bst
ituiç
ão,
g/g
0
10
20
30
40
50
12 h
24 h
p/ 12 h (R2 = 0,66)
p/ 24 h (y = x)
CH4, mL/ g MS incubada
0 2 4 6 8
Nív
el d
e su
bst
itu
ição
, g
/g
0
10
20
30
40
50
12 h
24 h
p/ 12 h (r2 = 0,44)
p/ 24 h (y = x)
CH4, mL/g MS incubada
0 2 4 6 8
Nív
el d
e su
bst
itu
ição
, g
/g
0
30
40
50
12 h
24 h
5.3. Produção de metano entérico
Na substituição do feno pela amêndoa de cacau, cupuaçu e mangostão
observou-se interação (P<0,05) entre o tempo de incubação e os níveis de
substituição para produção de metano (mL/ g de MS incubada e mL/ g de MS
digestível) (Figura 1, 2 e 3).
ŷ12 = 2,142 + 0,033x (r² = 0,44) ŷ 12 = 5,405 + 0,253x – 0,004x² (R² = 0,77)
ŷ 24 = 5,1443 ŷ 24 = 12,1731
Figura 1. Estimativa de produção de metano (CH4) (mL/ g MS incubada ou MS digerida),
em função do nível de substituição do feno de Brachiaria brizantha pela
amêndoa do cacau, por 12 e 24 horas de incubação in vitro. Letras diferentes
entre os tempos, para cada nível, diferem entre si (P<0,05).
a
b
a b
a
b
a b
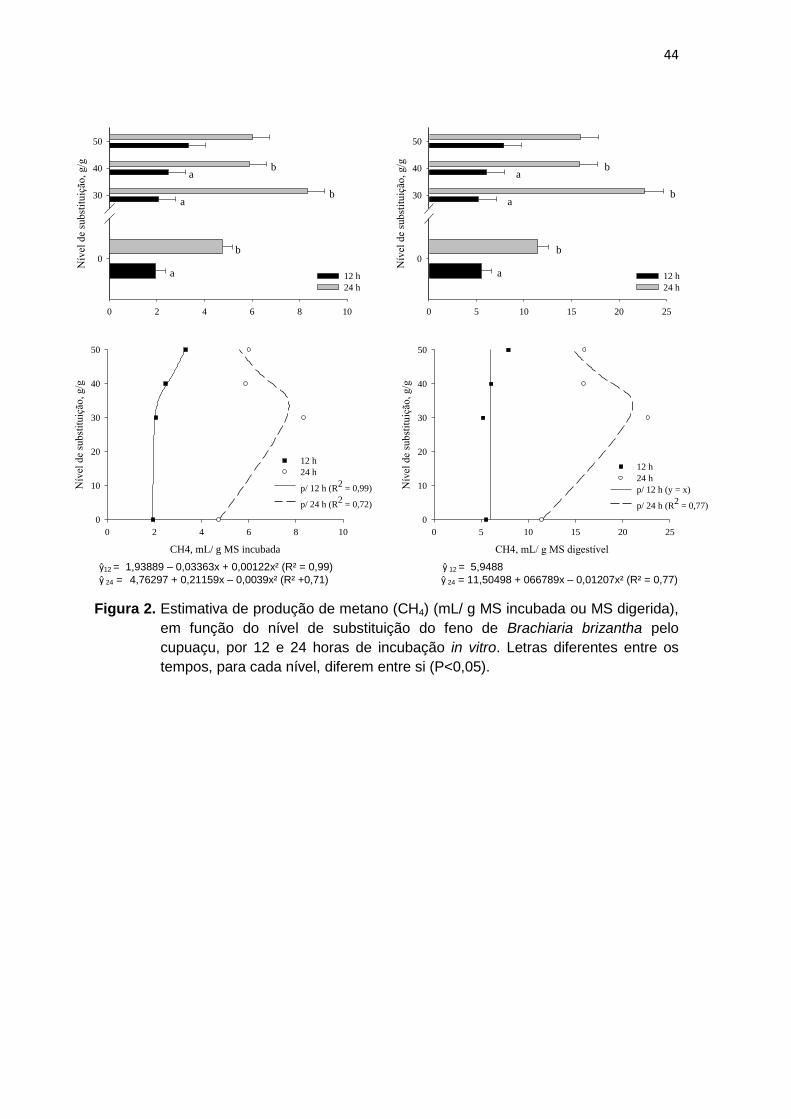
44
CH4, mL/g MS incubada
0 2 4 6 8 10
Nív
el d
e su
bst
itu
ição
, g
/g
0
30
40
50
12 h
24 h
CH4, mL/g MS digestível
0 5 10 15 20 25
Nív
el d
e su
bst
itu
ição
, g
/g
0
30
40
50
12 h
24 h
CH4, mL/ g MS digestível
0 5 10 15 20 25
Nív
el d
e su
bst
ituiç
ão,
g/g
0
10
20
30
40
50
12 h
24 h
p/ 12 h (y = x)
p/ 24 h (R2
= 0,77)
CH4, mL/ g MS incubada
0 2 4 6 8 10
Nív
el d
e su
bst
itu
ição
, g
/g
0
10
20
30
40
50
12 h
24 h
p/ 12 h (R2 = 0,99)
p/ 24 h (R2 = 0,72)
ŷ12 = 1,93889 – 0,03363x + 0,00122x² (R² = 0,99) ŷ 12 = 5,9488
ŷ 24 = 4,76297 + 0,21159x – 0,0039x² (R² +0,71) ŷ 24 = 11,50498 + 066789x – 0,01207x² (R² = 0,77)
Figura 2. Estimativa de produção de metano (CH4) (mL/ g MS incubada ou MS digerida),
em função do nível de substituição do feno de Brachiaria brizantha pelo
cupuaçu, por 12 e 24 horas de incubação in vitro. Letras diferentes entre os
tempos, para cada nível, diferem entre si (P<0,05).
a
a
a
b
b
b
a
a
a
a
b
b
b
a
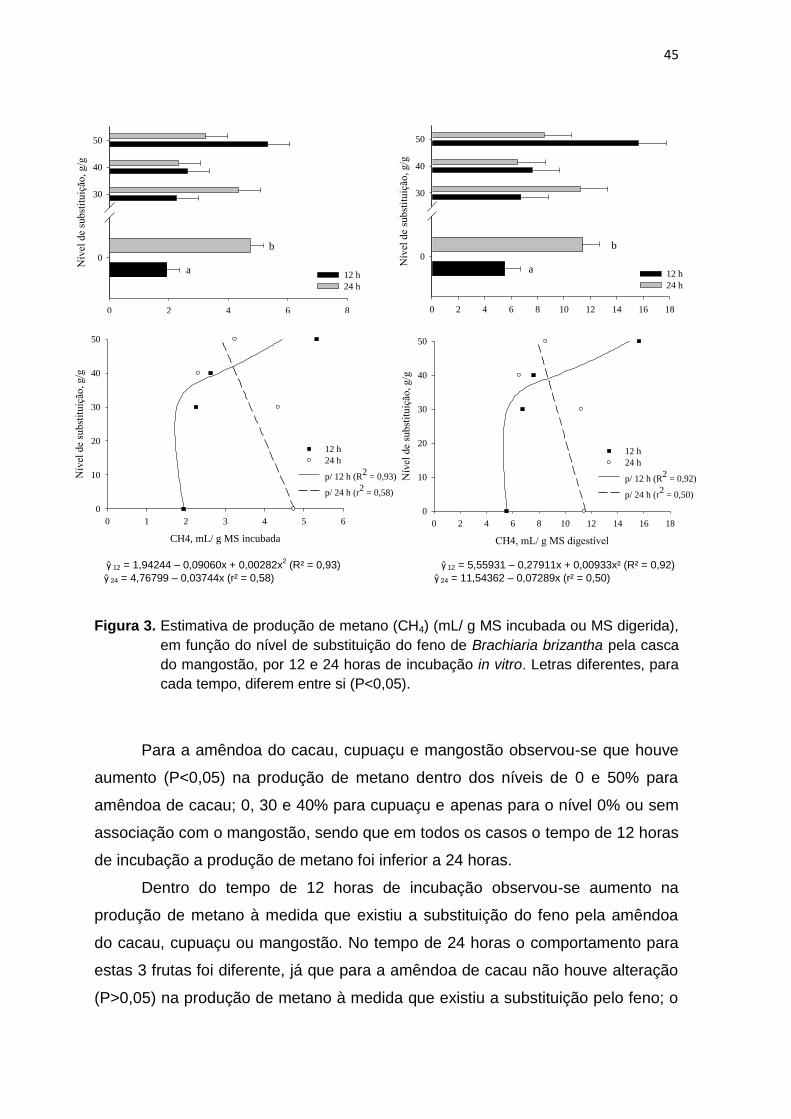
45
CH4, mL/g MS incubada
0 2 4 6 8
Nív
el d
e su
bst
itu
ição
, g
/g
0
30
40
50
12 h
24 h
CH4, mL/g MS digestível
0 2 4 6 8 10 12 14 16 18
Nív
el d
e su
bst
itu
ição
, g
/g
0
30
40
50
12 h
24 h
CH4, mL/ g MS incubada
0 1 2 3 4 5 6
Nív
el d
e su
bst
ituiç
ão,
g/g
0
10
20
30
40
50
12 h
24 h
p/ 12 h (R2
= 0,93)
p/ 24 h (r2
= 0,58)
CH4, mL/ g MS digestível
0 2 4 6 8 10 12 14 16 18
Nív
el d
e su
bst
itu
ição
, g
/g
0
10
20
30
40
50
12 h
24 h
p/ 12 h (R2
= 0,92)
p/ 24 h (r2 = 0,50)
ŷ 12 = 1,94244 – 0,09060x + 0,00282x
2 (R² = 0,93) ŷ 12 = 5,55931 – 0,27911x + 0,00933x² (R² = 0,92)
ŷ 24 = 4,76799 – 0,03744x (r² = 0,58) ŷ 24 = 11,54362 – 0,07289x (r² = 0,50)
Figura 3. Estimativa de produção de metano (CH4) (mL/ g MS incubada ou MS digerida),
em função do nível de substituição do feno de Brachiaria brizantha pela casca
do mangostão, por 12 e 24 horas de incubação in vitro. Letras diferentes, para
cada tempo, diferem entre si (P<0,05).
Para a amêndoa do cacau, cupuaçu e mangostão observou-se que houve
aumento (P<0,05) na produção de metano dentro dos níveis de 0 e 50% para
amêndoa de cacau; 0, 30 e 40% para cupuaçu e apenas para o nível 0% ou sem
associação com o mangostão, sendo que em todos os casos o tempo de 12 horas
de incubação a produção de metano foi inferior a 24 horas.
Dentro do tempo de 12 horas de incubação observou-se aumento na
produção de metano à medida que existiu a substituição do feno pela amêndoa
do cacau, cupuaçu ou mangostão. No tempo de 24 horas o comportamento para
estas 3 frutas foi diferente, já que para a amêndoa de cacau não houve alteração
(P>0,05) na produção de metano à medida que existiu a substituição pelo feno; o
a
b
a
b

46
cupuaçu proporcionou aumento na produção de metano e o mangostão houve
redução de até 31,5% na produção de metano quando comparado o nível de
substituição de 50% do feno.
Thanh (2012) observou redução do metano de 11,4-5,5 mL/g de substrato
fermentado após 21h de incubação (uma queda de 51%) com a utilização da
casca de mangostão. Manasri et al. (2012) encontraram redução de metano com
a adição da casca de mangostão em experimento in vivo corroborando com
Suchitra e Wanapat (2008), que também encontraram valores médios de metano
(CH4), no rúmen significativamente menores para a suplementação com a casca
de mangostão, sem e com a suplementação de óleo vegetal em comparação com
o grupo de controle.
No entanto, a de se considerar que esta fruta proporcionou redução
(P<0,05) na DIVMS, enquanto que apesar da amêndoa de cacau não reduzir a
produção de metano em relação ao feno, proporcionou maior DIVMS no nível de
23,1% de substituição, a qual possibilitou melhor aproveitamento dos nutrientes e
pode elevar a produtividade animal, o que reduziria a produção de metano por
produto gerado. Embora pesquisadores (Abdalla et al., 2008), afirmem o potencial
mitigador de CH4 de alimentos com alto teor de gordura, a amêndoa de cacau e o
cupuaçu com concentração de EE de 35,43 e 32,13% na MS não apresentaram
esta característica.
Para as frutas casca do cacau, graviola e jenipapo não houve interação
(P>0,05) entre o tempo de incubação e os níveis de substituição (Figura 4, 5 e 6).
Porém, independente do nível de substituição existiu maior (P<0,05) produção de
metano no tempo de 24 horas em relação ao tempo de 12 horas de incubação.
Também à medida que foi substituindo o feno por estas frutas houve aumento
(P<0,05) na produção de metano, independente do tempo de incubação.
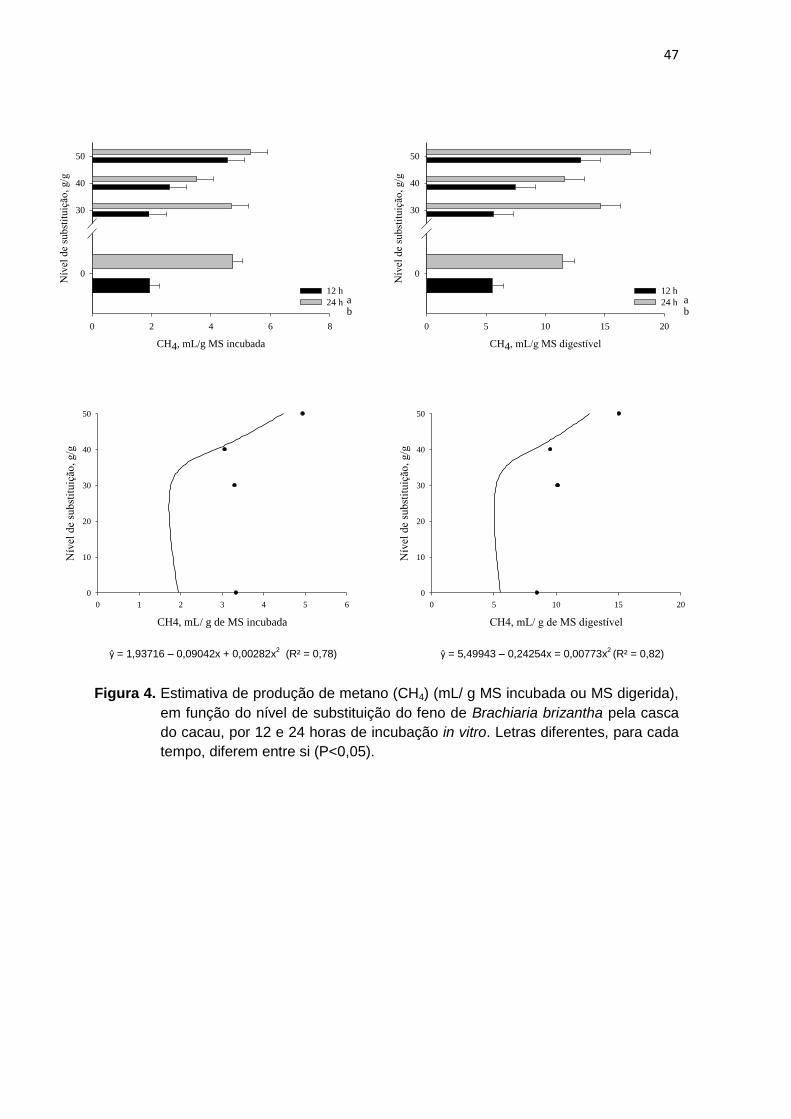
47
CH4, mL/g MS digestível
0 5 10 15 20
Nív
el d
e su
bst
itu
ição
, g
/g
0
30
40
50
12 h
24 h
CH4, mL/g MS incubada
0 2 4 6 8
Nív
el d
e su
bst
itu
ição
, g
/g
0
30
40
50
12 h
24 h
CH4, mL/ g de MS digestível
0 5 10 15 20
Nív
el d
e su
bst
ituiç
ão,
g/g
0
10
20
30
40
50
CH4, mL/ g de MS incubada
0 1 2 3 4 5 6
Nív
el d
e su
bst
ituiç
ão,
g/g
0
10
20
30
40
50
ŷ = 1,93716 – 0,09042x + 0,00282x
2 (R² = 0,78) ŷ = 5,49943 – 0,24254x = 0,00773x
2 (R² = 0,82)
Figura 4. Estimativa de produção de metano (CH4) (mL/ g MS incubada ou MS digerida),
em função do nível de substituição do feno de Brachiaria brizantha pela casca
do cacau, por 12 e 24 horas de incubação in vitro. Letras diferentes, para cada
tempo, diferem entre si (P<0,05).
a a b b
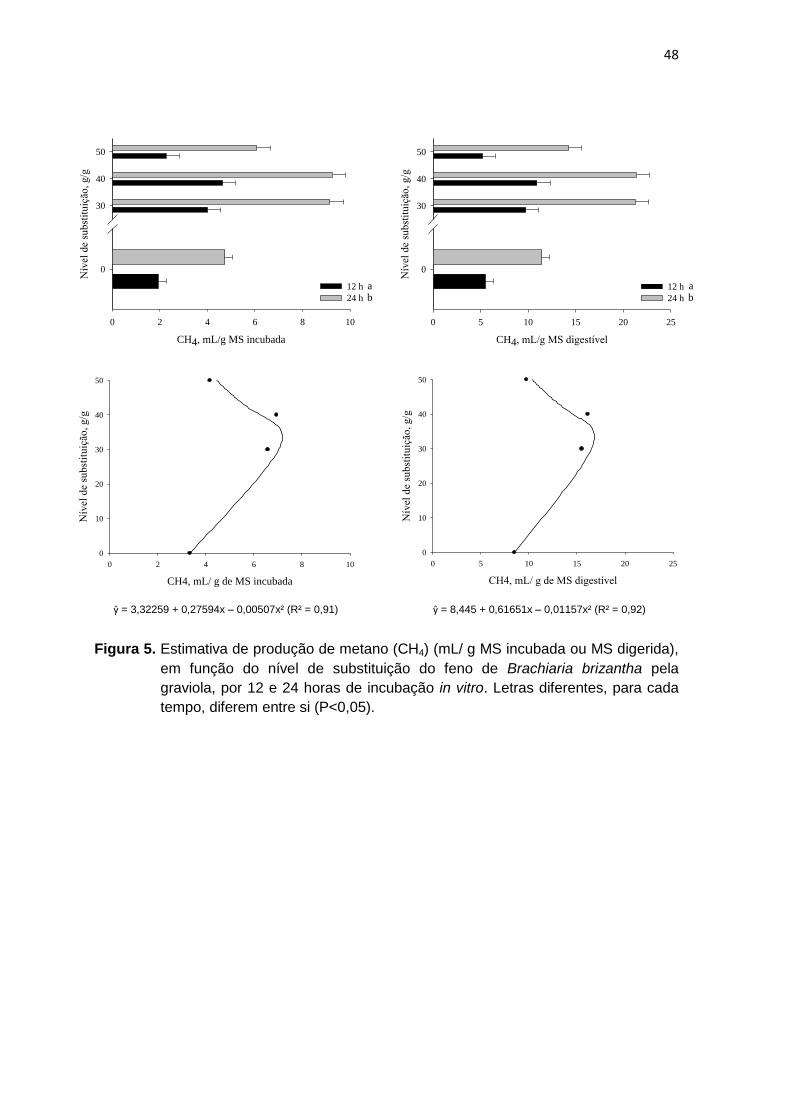
48
CH4, mL/g MS incubada
0 2 4 6 8 10
Nív
el d
e su
bst
itu
ição
, g
/g
0
30
40
50
12 h
24 h
CH4, mL/g MS digestível
0 5 10 15 20 25
Nív
el d
e su
bst
itu
ição
, g
/g
0
30
40
50
12 h
24 h
CH4, mL/ g de MS incubada
0 2 4 6 8 10
Nív
el d
e su
bst
ituiç
ão,
g/g
0
10
20
30
40
50
CH4, mL/ g de MS digestível
0 5 10 15 20 25
Nív
el d
e su
bst
ituiç
ão,
g/g
0
10
20
30
40
50
ŷ = 3,32259 + 0,27594x – 0,00507x² (R² = 0,91) ŷ = 8,445 + 0,61651x – 0,01157x² (R² = 0,92)
Figura 5. Estimativa de produção de metano (CH4) (mL/ g MS incubada ou MS digerida),
em função do nível de substituição do feno de Brachiaria brizantha pela
graviola, por 12 e 24 horas de incubação in vitro. Letras diferentes, para cada
tempo, diferem entre si (P<0,05).
a b
a b

49
CH4, mL/g MS incubada
0 2 4 6 8 10
Nív
el d
e su
bst
itu
ição
, g
/g
0
30
40
50
12 h
24 h
CH4, mL/g MS digestível
0 5 10 15 20
Nív
el d
e su
bst
itu
ição
, g
/g
0
30
40
50
12 h
24 h
CH4, mL/ g de MS incubada
0 2 4 6 8 10
Nív
el d
e su
bst
ituiç
ão,
g/g
0
10
20
30
40
50
CH4, mL/ g de MS digestível
0 5 10 15 20
Nív
el d
e su
bst
ituiç
ão,
g/g
0
10
20
30
40
50
ŷ = 3,39377 + 0,05732x (r² = 0,77) ŷ = 8,53576 + 0,08916x (r² = 0,,58)
Figura 6. Estimativa de produção de metano (CH4) (mL/ g MS incubada ou MS digerida), em função do nível de substituição do feno de Brachiaria brizantha pelo jenipapo, por 12 e 24 horas de incubação in vitro. Letras diferentes, para cada tempo, diferem entre si (P<0,05).
De acordo com Gonzales et al. (2013), a casca do fruto do cacaueiro pode
ser empregada na produção biogás em digestores, devido as sua característica
composição química, rica em lipídios, fibras e carboidratos, na produção de gás
metano. A casca de cacau contém açúcares disponíveis acelerando a produção
de CH4 que pára quando esses açúcares se esgotam (HARWOOD, 1980).
a b
a b

50
6. CONCLUSÕES
A substituição do feno de Brachiaria brizantha pelas frutas jenipapo,
amêndoa de cacau, cupuaçu, seguidos de graviola e casaca de cacau, melhoram
a cinética de fermentação ruminal e podem contribuir para um melhor
desempenho animal, estudos in vivo devem ser realizados para ver se o aumento
no desempenho tem potencial de mitigar metano por Kg de produto (leite/ carne)
produzido.
A amêndoa de cacau não altera na produção de metano, mas melhora a
fermentação ruminal enquanto que a casca do mangostão reduz em 31,5% a
produção de metano entérico, porém, piorou a fermentação dos carboidratos
fibrosos.
Estudos in vivo devem ser realizados para afirmar os resultados
encontrados in vitro e confirmar o melhor nível inclusão na dieta dos animais
ruminantes.

51
7. REFERÊNCIA BIBLIOGRÁFICAS
ABDALLA, A.L.; FILHO, J. C. S.; GODOI, A. R.; CARMO, C. A.; EDUARDO, J. L. P. Utilização de subprodutos da indústria de biodiesel na alimentação de ruminantes. Revista Brasileira de Zootecnia vol.37, Viçosa, July 2008. AOAC. Association Of Official Analytical Chemistry - Official Methods of Analysis, AOAC International 15th ed. , Arlington, 1990. ARAGÃO, C.G. Mudanças físicas e químicas da semente de cupuaçu (Theobroma grandiflorum Shum.) durante o processo fermentativo. Dissertação (Mestrado em Ciência dos Alimentos). Instituto Nacional de Pesquisa da Amazônia, 115p. Manaus, 1992. AZÊVEDO, J.A.G.; VALADARES FILHO,S.C.; PINA, D.S.; VALADARES, R.F.D.; DETMANN, E.; PAULINO, M.F. ; DINIZ, L.L. ; FERNANDES, H.J. Consumo, digestibilidade total, produção de proteína microbiana e balanço de nitrogênio em dietas para ruminantes de subprodutos Arq. Bras. Med. Vet. Zootec. vol.63 n°.1, Belo Horizonte, Fev., 2011. BERCHIELLI, T.T.; PEDREIRA, M.S.; OLIVEIRA, S.G; PRIMAVESI, O.; LIMA, M.; FRIGUETO, R.T.S. Determinação da produção de metano e pH ruminal em bovinos de corte alimentados com diferentes relações volumoso: concentrado. Anais Reunião anual da sociedade brasileira de zootecnia. Santa Maria: SBZ, 2003. BERNDT, A. Estratégias nutricionais para redução de metano. In: IV Congresso Latino Americano de Nutrição Animal - CLANA, 2010, Estância de São Pedro, SP. Colégio Brasileiro de Nutrição Animal (Eds.). p. 295 – 306, 2010. BERNDT, A. Impavto Da Pecuária De Corte Brasileira Sobre Os Gases Do Efeito Estufa. In: VII Simpósio de Produção de Gado de Corte, III Simpósio Internacional de Produção de Gado de Corte, Viçosa-MG, 2011. BHATTA, R.; TAJIMA, K.; TAKUSARI, N.; HIGUCHI, K.; ENISHI, O.; KURIHARA, M. Comparison of sulfur hexafluoride tracer technique, rumen simulation technique and in vitro gas production techniques for methane production from ruminant feeds. International Congress Series, v. 1293, p. 58–61, 2006. BRODY, A.L. Envasado de alimentos en atmosferas controladas, modificadas y a vacio. Zaragoza: Acribia, 1996. 220p. CARVALHO, G.G.P.; GARCIA, R.; PIRES, A.J.V. et al. Valor nutritivo de silagens de capim-elefante emurchecido ou com adição de farelo de cacau. Revista Brasileira de Zootecnia, v.36, n.5, p.1495-1501, 2007.

52
CARVALHO, J.E.D; MÜLLER, C.H.; ALVES. R.M.; R.A. NAZARÉ. Cupuaçuzeiro. Comunicado Técnico Embrapa N. 115 p. .1-4, 2004. CASEWELL, M.; FRIIS, C.; MARCO, E.; McMULLIN, P.; PHILLIPS, I. The European ban on growth-promoting antibiotics and emerging consequences for human and animal health. Journal Antimicrobials Chemotheraphy, 52 159–161, 2003. CORREA, M.P. Dicionário das plantas úteis do Brasil e das exóticas cultivadas. Rio de Janeiro: IBDF, v.4, p.515-519, 1969. DETMANN, E.; DA SILVA, J.F.C.; VÁSQUEZ, H.M.; HENRIQUES, L.T.; HADDADE, I.R. Cinética da degradação ruminal dos carboidratos de quatro gramíneas tropicais em diferentes idades de corte e doses de adubação nitrogenada: técnica de produção de gases. Revista Brasileira de Zootecnia, v. 38, n. 1, p. 149-158, 2009. DUKE, J. A.. Theobroma Cacao L. Handbook of Energy Crops, 1983. Disponível em: www.hort.purdue.edu/newcrop/duke_energy/Theobroma_cacao.html. Acesso em 29 de outubro de 2013. FAO. World Food and Agriculture Organization of the United Nations. Fao Statistical Yearbook 2013, Rome, 2013. Disponível em: http://www.fao.org/docrep/018/i3107e/i3107e.pdf. Acesso em 29 de outubro de 2013. FIEVEZ, V., DOHME, F., DANNEELS, M., RAES, K., DEMEYER, D. Fish oils as potent rumen methane inhibitors and associated effects on rumen fermentation in vitro and in vivo. Animal Feed Science and Technology, Amsterdam, v.104, p.41-58, 2003. FIGUEIREDO, R.W.; MAIA, G.A.; HOLANDA, L.F.F.; MONTEIRO, J.C. Características físicas e químicas do jenipapo. Pesquisa Agropecuária Brasileira, v.21, n.4, p.421-428, 1986. FRANCO, A. L. C. Fermentação ruminal e emissões de metano entérico in vitro de dietas contendo co-produtos da cadeia do biodiesel. Dissertação (Mestrado em Bioenergia) - Universidade Estadual de Londrina – Londrina, 87 f, 2012. GETACHEW, G.; ROBINSON, P.H.; DEPETERS, E.J.; TAYLOR, S.J.; GISI, D.D.; HIGGINBOTHAM, G.E.; RIORDAN, T.J. Methane production from commercial dairy rations estimated using an in vitro gas technique. Animal Feed Science and Technology, v. 123–124, n. 1, p. 391-403, 2005. GONZALES, A. D. F.; LIMA, J. M. ; VITAL, A. V. D. V.; RODRIGUES, M. B. S. Desenvolvimento sustentável para o resgate da cultura do cacau baseado no aproveitamento de resíduos. Interfaces Científicas -Saúde e Ambiente - Aracaju V.1 N.2 p. 41-52 fev., 2013.

53
GRAINGER, C., and BEAUCHEMIN, K. A. Can enteric methane emissions from ruminants be lowered without lowering their production Animal Feed Science and Technology , 166-167:308-320, 2011. GUIMARÃES JÚNIOR, R.; MARCHAO, R. L.; VILELA, L.; PEREIRA, L. G. R. Produção animal na integração lavoura-pecuária. In: Simpósio Mineiro de Nutrição de Gado de Leite. Anais... Belo Horizonte: UFMG, p. 111-123, 2010. GUO, Y.Q.; LIU, J.-X.; LU, Y.; ZHU, W.Y.; S.E. Denman, and C.S. McSweeney. Effect of tea saponin on methanogenesis, microbial community structure and expression of mcrA gene, in cultures of rumen micro-organisms. Letters in. Applied. Microbiology. 47, 421-426, 2008. HALL, M.B. Neutral Detergent-Soluble Carbohydrates. Nutritional Relevance and Analysis. University of Florida, Gainesville, 2003. HANSEN, D. S. et al. Caracterização química de frutos de jenipapeiros nativos do recôncavo baiano visando ao consumo natural e industrialização. Revista Brasileira de Fruticultura, Jaboticabal, v. 30, n. 4, p.964-969, 2008. HARPER, L.A.; DENMEAD, O.T.; FRENEY, J.R.; BYERS, F.M. Direct measurements of methane emissions from grazing and feedlot cattle. Journal of Animal Science, Savoy, v.77, p.1392-1401, 1999. HARWOOD, J. H. Pesquisa para produção de biogás na Amazônia. ACTA Amazônica 10 (2): 403-409, 1980. HEGARTY, R. S. Greenhouse gas emissions from the Australian livestock sector what do weknow, what can we do? Canberra, NSW Agriculture Australian Greenhouse Office, 35p., 2001. HOLTSHAUSEN, L.; CHAVES, A. V.; BEAUCHEMIN, K. A.; MCGINN, S. M.; MACALLISTER, T. A.; CHEEKE, P. R.; BENCHAAR, C. Feeding saponin-containing Yucca schidigera and Quillaja saponaria to decrease enteric methane production in dairy cows. Journal of Dairy Science. 92:2809-2821, 2009. HRISTOV, A. N. Historic, pre-European settlement, and presentday contribution of wild ruminants to enteric methane emissions in the United States. Journal of Animal Science 90 :1371–1375, 2012. IBGE - Instituto Brasileiro de Geografia e Estatística. Produção da Pecuária Municipal vol. 39. Brasil, 2011. IPCC. Intergovernamental Panel on Climate Change. IPCC Guidelines for national greenhouse gas inventories: reference manual, 1996. IPCC.Intergovernamental Panel on Climate Change. Emissions from livestock and manure management. In: Eggleston, H. S.; Buendia, L.; Miwa, K.; Ngara, T.;

54
Tabane, K. (eds). IPCC Guideliness for nacional greenhouse gas inventories. Hayama: IGES, chap. 10, p. 747- 846, 2006. IPCC. Intergovernamental Panel on Climate Change. Mudança do Clima 2007: A Base das Ciências Físicas. Geneva 2, Suíça, 2007. KEBREAB, E.; STRATHE, A.; FADEL, J.; MORAES, L. FRANCE, J. Impact of dietary manipulation on nutrient flows and greenhouse gas emissions in cattle. Revista Brasileira de Zootecnia, v. 39, suplemento especial, p. 458-464, 2010. LICITRA, G.; HERNANDEZ, T.M.; VAN SOEST, P.J. Standardization of procedures for nitrogen fractionation of ruminant feeds. Animal Feed Science and Technology, v.57, p.347-358, 1996. MACHADO, F. S., PEREIRA, L. G. R. P., JÚNIOR, R. G. J., LOPES, F. C. F., CHAVES, A. V. C., CAMPOS, M. M., MORENZ, M. J. F. Emissões de metano na pecuária: conceitos, métodos de avaliação e estratégias de mitigação. Embrapa Gado de Leite, Documento 147, 92 p., 2011. MACHMÜLLER, A. Medium-chain fatty acids and their potential to reduce methanogenesis in domestic ruminants. Agriculture, Ecosystems and Environment, Amsterdam, v.112, p.107-114, 2006. MACHMÜLLER, A.; OSSOWSKI, D.A.; WANNER, M.; KREUZER, M. Potential of various fatty feeds to reduce methane release from rumen fermentation in vitro (Rusitec). Animal Feed Science and Technology, v. 71, n. 1, p. 117–130, 1998. MANASRI, N.; WANAPAT, M.; NAVANUKRAW, C. Effect of mangosteen peel-garlic powder pellet supplementation on rumen ecology and methane gas production in beef cattle. Khonkaen Agr. J.40 Supplment p.154-159, 2012. MARCONDES, M. I.; VALADARES FILHO, S. C.; DETMANN, E.; VALADARES, R.F.D.; SILVA, L. F. C.; FONSECA, M. A. Degradação ruminal e digestibilidade intestinal da proteína bruta de alimentos para bovinos. Revista Brasileira de Zootecnia., v.38, n.11, p.2247-2257, 2009. MARTIN, C.; MORGAVI, D.P.; DOREAU, M. Methane mitigation in ruminants: from microbes to the farm scale. Animal, 4:3, p. 351-365, 2009. MAURICIO, R.M.; MOULD, F.L.; DHANOA, M.S.; OWEN, E.; CHANNA, K.S.; THEODOROU, M.K. A semi-automated in vitro gas production technique for ruminants feedstuff evaluation. Animal Feed Science and Technology, v.79, p.321-330, 1999. MAURICIO, R.M.; PEREIRA, L.G.R.; GONÇALVES, L.C.; RODRIGUEZ, N.M. Relação entre pressão e volume para implantação da técnica in vitro semi-automática de produção de gases na avaliação de forrageiras tropicais. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v.55, p.216-219, 2003.

55
McALLISTER, T. A.; OKINE, E. K.; MATHISON, G. W. and CHENG, K. J. Dietary, environmental and microbiological aspects of methane production in ruminants. Canadian Journal of Animal Science. 76: 231-243, 1996. McALLISTER, T. A. and NEWBOLD, C.J. Redirecting rumen fermentation to reduce methanogenesis. Australian Journal of Experimental Agriculture 48, 7-13, 2008. MCT - Ministério da Ciência e Tecnologia. Segunda comunicação nacional do Brasil à convenção-quadro das Nações Unidas sobre mudança do clima. Brasília, 280p, 2010. MENKE, K.H.; STEINGASS, H. Estimation of the energetic feed value obtained from chemical analysis and gas production using rumen fluid. Animal Research Development, v. 28, p. 7-55, 1988. MERTENS, D.R. Analysis of fiber in feeds and its use in feed evaluation and ration formulation. In: Simpósio Internacional de Ruminantes, Anais… Lavras: Sociedade Brasileira de Zootecnia, p.1-32, 1992. MONTEIRO, R.B.N.C. Desenvolvimento de um modelo para estimativas da produção de gases de efeito estufa em diferentes sistemas de produção de bovinos de corte. Dissertação (Mestrado) – Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, 75p., 2009. MOSS, A.R. Methane: global warming and production by animals. Kingston: Chalcombe Publications, United Kingdom, 105p., 1993. MOSS, A.R.; GIVENS, D.I. The effect of supplementing grass silage with soya bean meal on digestibility, in sacco degradability, rumen fermentation an methane production in sheep. Animal Feed Science and Technology, Amsterdam, v.97, p.127-143, 2002. NGAMSAENG, A., M. WANAPAT, and KHAMPA, S. Effects of mangosteen peel (Garcinia mangostana) supplementation on rumen ecology, microbial protein synthesis, digestibility and voluntary feed intake in cattle. Pakistan Journal of Nutrition. 5, 445-452, 2006. NKRUMAH, J. D., OKINE, E. K., MATHISON, G. W., SCHMID, K., LI, C., BASARAB, J. A., PRICE, M. A., WANG, Z. & MOORE, S. S. Relationships of feedlot feed efficiency, performance, and feeding behavior with metabolic rate, methane production, and energy partitioning in beef cattle. Journal of Animal Science. 84: 145-153, 2006. NOCEK, J.; RUSSEL, J.B. Protein and carbohydrate as an integrating system. Relationship of ruminal availability to microbial contribution and milk production. Journal of Dairy Science, v. 71, n. 5, p. 2070-2075, 1998.

56
NÖRNBERG, J.L.; STUMPF JR., W.; LÓPEZ, J.; COSTA, P.B. Valor do farelo de arroz integral como fonte de gordura na dieta de vacas Jersey na fase inicial da lactação: digestibilidade aparente de nutrientes. Revista Brasileira de Zootecnia, v. 33, n. 6, p. 2412-2421, 2004. O´HARA, P.; FRENEY, J.; ULYATT, M. Abatement of agricultural non-carbon dioxide greenhouse gas emissions: a study of research requirements. Report prepared for the Ministry of Agriculture and Forestry on Behalf of the Convenor, Ministerial Group on Climate Change, the Minister of Agriculture and the Primary Industries Council. New Zealand: Crown Copyright - Ministry of Agriculture and Forestry, 170p., 2003. Disponível em: http://maxa.maf.govt.nz/mafnet/rural-nz/sustainable-resource use/climate/abatement-of-agricultural-greenhouse-gas-emissions/greenhouse-gas-emissions.pdf Acesso em 25/10/2013. OWENS, F. N.; GOETSCH, A. L. Ruminal fermentation. In: Church, D. C. (Ed). The ruminant animal: digestive physiology and nutrition. Waveland Press, p.145-171, 1988. PEDREIRA, M.S.; OLIVEIRA, S.G.; BERCHIELLI, T.T.; PRIMAVESI, O.. Aspectos relacionados com a emissão de metano de origem ruminal em sistenas de produção de bovinos. (Ruminal methane emission related aspects in cattle production systems). Archives of Veterinary Science, v. 10, n. 3, Printed in Brazil ISSN: 1517-784X, p. 24-32, 2005. PEREIRA, E. M. O. Torta de cupuaçu (Theobroma grandiflorum) na alimentação de ovinos. Tese doutorado em zootecnia (Produção Animal) Jaboticabal - São Paulo – Brasil, 2009. PIRES, A.J.V.; CARVALHO, G.G.P.; GARCIA, R.; JUNIOR, J. N. C.; RIBEIRO, L. S. O.; CHAGAS, D. M. T. Capim-elefante ensilado com casca de café, farelo de cacau ou farelo de mandioca R. Bras. Zootec. vol.38 no.1, Viçosa Jan., 2009. PIRES, A.J.V.; VIEIRA, V.F.; SILVA, F.F. et al. Níveis de farelo de cacau (Theobroma cacao) na alimentação de bovinos. Revista Electronica da Veterinária, v.6, p.1-10, 2005. POUNGCHOMPU, O.; WANAPAT, M.; WACHIRAPAKORN, C.; WANAPAT, S. an CHERDTHONG, A. Manipulation of ruminal fermentation and methane production by dietary saponins and tannins from mangosteen peel and soapberry fruit. Archives of Animal Nutrition 5, 389-400, 2009. REGULAMENTO (CE) N.o 1831/2003 DO PARLAMENTO EUROPEU E DO CONSELHO relativo aos aditivos destinados à alimentação animal. Jornal Oficial da União Europeia L 268/29, setembro de 2003.

57
ROSS, Jessica. Cocoa and Chocolate as Functinal Foods. March 2001. Disponível em: http://www.uoguelph.ca/nhptc/Jessica1.html. Acesso em 25/10/2013. RUSSEL, J.B.; O'CONNOR, J.D.; FOX, D.G.; VAN SOEST, P.J.; SNIFFEN, C.J. A net carbohydrate and protein system for evaluating cattle diets: I. Ruminal fermentation. Journal of Animal Science, v. 70, n. 11, p. 3551-3561, 1992. SACRAMENTO, C. K. Graviola. Transcrito do Jornal CEPLAC Notícias - julho 2000. Disponível em: http://www.ceplac.gov.br/radar/graviola.htm. Acesso em 05 de novembro de 2013. SACRAMENTO, C.K.; COELHO JUNIOR, E.; CARVALHO, J.E.U.; MÜLLER, C.H.; NASCIMENTO, W.M.O. Cultivo do Mangostão no Brasil. Revista Brasileira de Fruticultura, Jaboticabal, v. 29, n. 1, p. 195-203, 2007. SANTOS, M.G.M.F; AZEVÊDO, J.A.G; PEREIRA, L.G.R.; SILVA, T.C.; CABRAL, I.S.; FARIAS, D.S. Relação entre pressão e volume para implantação da técnica in vitro de produção de gases no trópico úmido. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA Anais. 47ª. Salvador, 2010. SAUER, F. D., FELLNER, V., KINSMAN, R., KRAMER, J.K.G., JACKSON, H.A., et al. Methane output and lactation response in Holstein cattle with monensina or unsaturated fat added to the diet. Journal of Animal Science 76, 906–914, 1998. SCHOFIELD, P.; PELL, A. N. Measurement and kinetic analysis of the neutral detergent-soluble carbohydrate fraction of legumes and grasses. Journal of Animal Science, v. 73, p. 3455-3463, 1995. SCHOFIELD, P.; PITT, R.E.; PELL, A.N. Kinetic of fiber digestion from in vitro gas production. Journal of Animal Science, v. 72, n. 11, p. 2980-2991, 1994. SENA, J. V. C. Informe Rural Etene. Banco do Nordeste. Ano V nº 02– Fevereiro de 2011. Disponível em: http://www.bnb.gov.br/content/aplicacao/etene/etene/docs/ire_ano5_n2.v Acessado em 05/11/2013. SILVA, H.G.O.; PIRES, A.J.V.; CUNHA NETO, P. A. et al. Digestibilidade de dietas contendo silagem de capim-elefante amonizado farelo de cacau ou torta de dendê em ovinos. Pesquisa Agropecuária Brasileira, v.36, p.499-506, 2007. SILVA, H.G.O.; PIRES, A.J.V.; SILVA, F.F. et al. Digestibilidade aparente de dietas contendo farelo de cacau ou torta de dendê em cabras lactantes. Pesquisa Agropecuária Brasileira, v.40, n.4, p.405-411, 2005. SILVA NETO, P. J.; MATOS, P. G. G.; MARTINS, A. C. S.; SILVA, A. P. Sistema de produção de cacau para a Amazónia brasileira. Belém, CEPLAC, 125p., 2001.

58
SNIFFEN, C.J.; O'CONNOR, J.D.; VAN SOEST, P.J.; FOX, D.G.; RUSSELL, J.B. A net carbohydrate and protein system for evaluating cattle diets: II– Carbohydrate and protein availability. Journal of Animal Science, v.70, n.12, p.3562-3577, 1992.
SUCHITRA, K. and WANAPAT, M. Effects of mangosteen (Garcinia mangostana) peel and sunflower and coconut oil supplementation on rumen fermentation, milk yield and milk composition in lactating dairy cows. Livestock Research for Rural Development. Volume 20, supplement, 2008 . THANH, V. D.; PRESTON, T. R. and LENG, R. A. Effect of NPN source and mangosteen peel (Garcinia mangostana) on methane production in vitro incubations and on growth performance of Phan Rang sheep in the Mekong delta of Vietnam. Master of science thesis in agricultural sciences animal husbandry Code Number: 60 - 62 – 40. Can Tho City, Viet Nam – 2012. THORPE, A. Enteric fermentation and ruminant eructation: the role (and control?) of methane in the climate change debate. Climatic change, v. 93, p. 407-431, 2009. USEPA - United States Environmental Protection Agency . Evaluating Ruminant Livestock Efficiency Projects and Programs In: Peer Review Draft. Washington, D.C, 48p., 2000. VALADARES FILHO, S.C.; PINA, D.S. Fermentação Ruminal. In: BERCHIELLI, T.T.; PIRES, A.V.; OLIVEIRA, S.G. (Eds). Nutrição de Ruminantes. p.151-182, 2006. VENTURIERI, G.A. et al. O cultivo do cupuaçuzeiro. Informativo SBF, Campinas, v.4, n.1, p.15-17, 1985. XAVIER, M.; XAVIER, A.T.T.N. Jenipapo: uma espécie indígena para reflorestar. Cerrado, Brasília, v.B, n.34, p.20-23, 1976.





























