Embed Size (px)
Citation preview

UNIVERSIDADE ESTADUAL PAULISTA “JÚLIO DE MESQUITA FILHO”
FACULDADE DE CIÊNCIAS AGRONÔMICAS
CAMPUS DE BOTUCATU
“COMPLEXO DE PERCEVEJOS DA SOJA: REFLEXOS SOBRE A
PRODUTIVIDADE DE GENÓTIPOS E DETERMINAÇÃO DOS TIPOS
DE RESISTÊNCIA A Euschistus heros (FABRICIUS) (HEMIPTERA:
PENTATOMIDAE)”
EFRAIN DE SANTANA SOUZA
Tese apresentada à Faculdade de Ciências
Agronômicas da UNESP – Câmpus de
Botucatu, para obtenção do título de Doutor em
Agronomia (Proteção de Plantas)
BOTUCATU-SP
Outubro - 2013

UNIVERSIDADE ESTADUAL PAULISTA “JÚLIO DE MESQUITA FILHO”
FACULDADE DE CIÊNCIAS AGRONÔMICAS
CÂMPUS DE BOTUCATU
“COMPLEXO DE PERCEVEJOS DA SOJA: REFLEXOS SOBRE A
PRODUTIVIDADE DE GENÓTIPOS E DETERMINAÇÃO DOS
TIPOS DE RESISTÊNCIA A Euschistus heros (FABRICIUS)
(HEMIPTERA: PENTATOMIDAE)”
EFRAIN DE SANTANA SOUZA
Orientador: Prof. Dr. Edson Luiz Lopes Baldin
Co-orientador: Dr. André Luiz Lourenção
Tese apresentada à Faculdade de Ciências
Agronômicas da UNESP – Câmpus de
Botucatu, para obtenção do título de
Doutor em Agronomia (Proteção de
Plantas)
BOTUCATU-SP
Outubro - 2013

FICHA CATALOGRÁFICA ELABORADA PELA SEÇÃO TÉCNICA DE AQUISIÇÃO E TRATAMENTO DA INFORMAÇÃO – SERVIÇO TÉCNICO DE BIBLIOTECA E DOCUMENTAÇÃO - UNESP - FCA - LAGEADO - BOTUCATU (SP) Souza, Efrain de Santana, 1984- S731c Complexo de percevejos da soja: reflexos sobre a produ-
tividade de genótipos e determinação dos tipos de resistên-cia a Euschistus heros (Fabricius) (Hemiptera: Pentatomi
dae)/ Efrain de Santana Souza. – Botucatu : [s.n.], 2013 xiii, 106 f. : ils. color., grafs., tabs., fots. color. Tese (Doutorado) - Universidade Estadual Paulista, Fa- culdade de Ciências Agronômicas, Botucatu, 2013 Orientador: Edson Luiz Lopes Baldin Coorientador: André Luiz Lourenção Inclui bibliografia 1. Soja - Resistência a insetos. 2. Glycine max. 3. An-
tibiose. 4. Antixenose. 5. Percevejo. I. Baldin, Edson Luiz Lopes. II. Lourenção, André Luiz. III. Universidade Esta-dual Paulista “Júlio de Mesquita Filho” (Campus de Botuca-tu). Faculdade de Ciências Agronômicas. IV. Título.


II
BIBLIOGRAFIA DO AUTOR
EFRAIN DE SANTANA SOUZA, filho de Wanderley Rodrigues de Souza e
Maria Aparecida de Santana Souza, nasceu em General Salgado – SP, no dia 10 de
fevereiro de 1984.
Cursou o ensino fundamental na cidade natal e o ensino médio em Jales - SP,
onde também obteve o título de Técnico em Agricultura pelo Centro Paula Souza
(CETEPS) no ano de 2001.
No primeiro semestre do ano de 2003, ingressou no curso de Agronomia na
Universidade Camilo Castelo Branco, Campus de Fernandópolis. Nesse período, foi
bolsista de Iniciação Científica pela FAPESP e graduou-se em janeiro de 2007.
Iniciou o curso de Mestrado em Agronomia, área de concentração em Proteção
de Plantas, em agosto de 2007, na Faculdade de Ciências Agronômicas da Universidade
Estadual Paulista Júlio de Mesquita Filho, Campus de Botucatu, obtendo o título de
Mestre em fevereiro de 2010. Em março do mesmo ano, iniciou o curso de Doutorado
em Agronomia na mesma área e instituição.

III
A DEUS, fonte de toda a sabedoria, equilíbrio e amor,
...OFEREÇO
À minha família,
WANDERLEY RODRIGUES DE SOUZA (pai)
MARIA AP. DE SANTANA SOUZA (mãe)
GABRIEL DE SANTANA SOUZA (irmão),
Pelo dom da vida, pelo apoio e amor incondicional.
...DEDICO
Aos meus Avós,
ARMINDO
ENELZIRA
MOISÉS “in memorian”
INÊS,
Pelos incentivos e ensinamentos durante meus dias de vida,
...HOMENAGEIO
À minha namorada,
CAROLINE GERALDI PIEROZZI,
Pelos momentos juntos e pelo auxílio direto no desenvolvimento deste trabalho,
...DIVIDO

IV
Ao Dr. EDSON LUIZ LOPES BALDIN,
O eterno professor de Agronomia que me ensinou sempre alguma lição em cada
conversa, despertando meu interesse na área acadêmica. Minha eterna gratidão Baldin,
pelo ensino, orientação e amizade.
Se deres um peixe a um homem faminto, vai alimentá-lo por um dia. Se o ensinares a
pescar, vai alimentá-lo toda a vida.
Lao-Tsé

V
AGRADECIMENTOS
À Faculdade de Ciências Agronômicas – UNESP, Campus de Botucatu, por ter
oferecido condições para a realização deste trabalho.
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), pela
concessão da bolsa.
Aos Professores da FCA, em especial aos do Departamento de Proteção Vegetal da
FCA/UNESP, pelos ensinamentos transmitidos;
Ao meu co-orientador Dr. André Luiz Lourenção, do IAC de Campinas (Setor de
Entomologia), pelas sugestões e conhecimentos transmitidos durante o desenvolvimento
deste estudo;
À técnica Neide Zério do Laboratório de Biologia de Insetos da ESALQ-USP, pelos
ovos de E. heros e pelas informações que auxiliaram na metodologia de criação;
Ao doutorando em Estatística Lucas Santana da Cunha, pelo auxílio nas análises;
À Dra. Ana Carolina Firmino pela ajuda na obtenção das imagens de microscopia
eletrônica;
Aos estagiários e pós-graduandos do Departamento de Proteção Vegetal vinculados ao
Laboratório de Resistência de Plantas a Insetos e Plantas Inseticidas (LARESPI);
Ao meu amigo MSc. José Paulo Gonçalves Franco da Silva com quem tive o prazer de
trabalhar e desenvolver este trabalho;
Ao técnico laboratorial Sr. Domingos Paulossi, pela amizade e suporte em alguns
ensaios;

VI
Às funcionárias da Seção de Pós-Graduação da FCA/UNESP pela simpatia e
consideração com que sempre me atenderam;
Aos bons amigos que adquiri e cultivei durante minha permanência em Botucatu:
Maurício Andrião, Thaíse Dias, Renato Boreli, Ricardo Fujihara, Estelita Gourgel, Luiz
Panutti, José Paulo e Vinícius Canassa;
Aos insetos, animais fantásticos que frequentemente ensinam e estimulam a curiosidade
dos seres humanos.

VII
SUMÁRIO
Páginas
Lista de tabelas ............................................................................................. IX
Lista de figuras.................................................................................................. XII
1 Resumo.......................................................................................... 1
2 Summary......................................................................................... 3
3 Introdução........................................................................................ 5
4 Revisão de literatura....................................................................... 8
4.1 Cultura da soja.......................................................................... 8
4.2 Características do complexo de percevejos.............................. 9
4.3 Aspectos bioecológicos de Euschistus heros (Fabricius)........ 11
4.4 Danos associados ao complexo de percevejos da soja............. 13
4.5 Métodos de controle para o complexo de percevejos da soja..... 14
5 Material e Métodos.......................................................................... 20
5.1 Genótipos da soja utilizados..................................................... 20
5.2 Obtenção dos genótipos............................................................ 21
5.3 Criação estoque de E. heros........................................................ 23
5.4 Ensaios a campo....................................................................... 24
5.5 Ensaios em laboratório............................................................. 28
5.5.1 Atratividade e preferência alimentar........................... 28
5.5.2 Quantificação e comprimento de tricomas em vagens.... 30
5.5.3 Determinação da dureza das vagens............................. 31
5.5.4 Antibiose...................................................................... 32
5.6 Análises Estatísticas................................................................. 34
6 Resultados e Discussão................................................................... 35
6.1 Ensaios a campo....................................................................... 35
6.1.1 Flutuação populacional de percevejos pentatomídeos.... 35
6.1.2 Ocorrência de espécies de percevejos pentatomídeos...... 37
6.1.3 Atratividade, retenção foliar, produtividade e danos
ocasionados por percevejos pentatomídeos em
genótipos da soja.........................................................
38
6.1.3.1 Ciclo Precoce.................................................... 38
6.1.3.2 Ciclo Semiprecoce.................................................. 48

VIII
6.1.3.3 Ciclo Tardio............................................................ 57
6.2 Ensaios em laboratório......................................................... 66
6.2.1 Determinação de atratividade de Euschistus heros em
testes com chance de escolha....................................... 66
6.2.2 Determinação do número de picadas e tempo de
consumo de Euschistus heros em diferentes genótipos
de soja...................................................................... 68
6.2.3 Dureza, quantificação e comprimento de tricomas em
vagens R5/R6 de diferentes genótipos........................ 71
6.2.4 Avaliação de antibiose.................................................. 76
7 Conclusões............................................................................... 87
8 Referências bibliográficas........................................................... 89
Apêndices................................................................................................. 105

IX
LISTA DE TABELAS
Páginas
Tabela 1. Genealogia dos genótipos de soja de ciclos precoce, semiprecoce e
tardio utilizados no trabalho............................................................................... 21
Tabela 2. Número médio (±EP) de percevejos pentatomídeos (E. heros, P.
guildinii, N. viridula, E. meditabunda e Dichelops spp.) por pano de batida
obtidos em plantas de seis genótipos de soja de ciclo precoce durante dois
anos de ensaio a campo............................................................................. 40
Tabela 3. Retenção foliar média (±EP) ocasionada pelo ataque de percevejos
em seis genótipos de soja de ciclo precoce....................................................... 41
Tabela 4. Produtividade média (kg/ha-1
) (±EP) e percentagem de redução de
produção de seis genótipos de soja de ciclo precoce em parcelas infestadas e
não infestadas por percevejos.................................................................... 46
Tabela 5. Escore médio de dano nos grãos de seis genótipos de soja de
ciclo precoce em ensaios de campo, causados por percevejos
pentatomídeos........................................................................................ 47
Tabela 6. Número médio (±EP) de percevejos pentatomídeos (E. heros, P.
guildinii, N. viridula, E. meditabunda e Dichelops spp.) por pano de batita
obtido em plantas de seis genótipos de soja de ciclo semiprecoce durante dois
anos de ensaio a campo............................................................................ 49
Tabela 7. Retenção foliar média (±EP) ocasionada pelo ataque de percevejos
em seis genótipos de soja de ciclo semiprecoce........................................... 50
Tabela 8. Produtividade média (kg/ha-1
) (±EP) e percentagem de redução de
produção de seis genótipos de soja de ciclo semiprecoce em parcelas
infestadas e não infestadas por percevejos....................................................... 53
Tabela 9. Escores médios de dano nos grãos de seis genótipos de soja de
ciclo semiprecoce em ensaios de campo........................................................... 56
Tabela 10. Número médio (±EP) de percevejos pentatomídeos (E. heros, P.
guildinii, N. viridula, E. meditabunda e Dichelops spp.) por pano de batita
obtido em plantas de cinco genótipos se soja de ciclo tardio durante dois anos
de ensaio a campo...................................................................................... 59
Tabela 11. Retenção foliar média (±EP) ocasionada pelo ataque de
percevejos em cinco genótipos de soja de ciclo tardio...................................... 60

X
Tabela 12. Produtividade média (kg/ha-1
) (±EP) e percentagem de redução de
produção de cinco genótipos de soja de ciclo tardio em parcelas infestadas e
não infestadas com percevejos............................................................................ 62
Tabela 13. Escores médios de dano nos grãos causados por percevejos em
cinco genótipos de soja de ciclo tardio, em ensaios de campo.......................... 65
Tabela 14. Número médio (±EP) de adultos de Euschistus heros atraídos por
vagens R5/R6 de seis genótipos de soja de ciclo precoce, aos 15, 30, 45, 60,
120 e 180 minutos após a liberação (T= 26 2ºC, U.R.= 65 10% e
fotoperíodo=14h)............................................................................................... 66
Tabela 15. Número médio (±EP) de adultos de Euschistus heros atraídos por
vagens R5/R6 de seis genótipos de soja de ciclo semiprecoce, aos 15, 30, 45,
60, 120 e 180 minutos após a liberação dos insetos........................................... 67
Tabela 16. Número médio (±EP) de adultos de Euschistus heros atraídos por
vagens R5/R6 de cinco genótipos de soja de ciclo tardio, aos 15, 30, 45, 60,
120 e 180 minutos após a liberação dos insetos................................................ 68
Tabela 17. Número médio de picadas e tempo de picada por picada (±EP) de
adultos de Euschistus heros em vagens R5/R6 de seis genótipos de soja de
ciclo precoce...................................................................................................... 69
Tabela 18. Número médio de picadas e tempo de picada por picada (±EP) de
adultos de Euschistus heros em vagens R5/R6 de seis genótipos de soja de
ciclo semiprecoce............................................................................................... 70
Tabela 19. Número médio de picadas e tempo de picada por picada (±EP) de
adultos de Euschistus heros em vagens R5/R6 de cinco genótipos de soja de
ciclo tardio........................................................................................................... 71
Tabela 20. Densidade média de tricomas, comprimento de tricomas e dureza
de vagem (±EP)1 de seis genótipos de soja de ciclo precoce............................. 72
Tabela 21. Densidade média de tricomas, comprimento de tricoma e dureza
de vagem (±EP)1 de seis genótipos de soja de ciclo semiprecoce.................... 74
Tabela 22. Densidade média de tricomas, comprimento de tricoma e dureza
de vagem (±EP)1 de cinco genótipos de soja de ciclo tardio........................... 75
Tabela 23. Período médio (dias ± EP) de duração de ínstares ninfais e fase
ninfal total (dias ± EP) de Euschistus heros em 17 genótipos de soja (T= 26
2ºC, U.R.= 65 10% e fotoperíodo=14h)..........................................................
79

XI
Tabela 24. Longevidade média (dias ± EP) e peso de adultos recém-
emergidos (mg ± EP) de Euschistus heros obtidos em 17 genótipos de soja
(T= 26 2ºC, U.R.= 65 10% e fotoperíodo=14h).......................................... 84
Tabela 25. Viabilidade de ovos (± EP) de Euschistus heros confinados em 17
genótipos de soja (T= 26 2ºC, U.R.= 65 10% e
fotoperíodo=14h)............................................................................................... 85

XII
LISTA DE FIGURAS
Página
Figura 1. A - Área semeada com os genótipos de soja para a multiplicação de
sementes; B - sementes colhidas, selecionadas e acondicionadas
hermeticamente.................................................................................................. 22
Figura 2. A - Gaiola de criação de adultos de E. heros e placas contendo
ovos; B - vista interna da gaiola de criação; C - ninfas buscando água em
algodão embebido. (T= 26 2ºC, U.R.= 65 10% e fotoperíodo=14 h).......... 23
Figura 3. A - Vista geral da área experimental do primeiro ensaio realizado
em 2011-2012; B - área do segundo ensaio realizado em 2012-2013................ 25
Figura 4. A - Planta entrando em estádio reprodutivo R2; B - avaliação
populacional de percevejos pentatomídeos (E. heros, P. guildinii, N. viridula, E.
meditabunda e Dichelops spp.) com pano de batida........................................... 27
Figura 5. A - Colheita de quatro metros lineares centrais das parcelas; B -
Classificação do nível de dano nos grãos dos diferentes genótipos.................... 27
Figura 6. A - Percevejos mantidos sob jejum de 24 h; B - Vista geral de uma
arena; C - Adulto de E. heros se alimentando em vagem; D - Testes em
execução. (T=26 2ºC, U.R.=6510% e fotoperíodo=14h)................................ 29
Figura 7. A - Imagem dos tricomas de vagem obtida por microscopia
eletrônica; B - medição do comprimento dos tricomas pelo software EDN-2... 31
Figura 8. A - Placa de Petri contendo vagem e duas ninfas de E. heros; B -
Genótipo cultivado em casa-de-vegetação para o fornecimento de vagens; C -
Vista geral de parte do ensaio de antibiose. (T=262ºC, U.R.=6510% e
fotoperíodo=14h)............................................................................................... 33
Figura 9. Flutuação populacional de percevejos pentatomídeos (E. heros, P.
guildinii, N. viridula, E. meditabunda e Dichelops spp.) nos ensaios com os
diferentes genótipos de soja de ciclos precoce, semiprecoce e tardio................ 36
Figura 10. Ocorrência (%) de percevejos pentatomídeos (E. heros, P.
guildinii, N. viridula, E. meditabunda e Dichelops spp.) registrados em
ensaios de campo com soja................................................................................. 37
Figura 11. Número de grãos danificados por pentatomídeos de acordo com
quatro classes de danos em seis genótipos de soja de ciclo precoce em ensaios
de campo. Botucatu-SP, 2011-2012 e 2012-2013. A = grãos sem danos

XIII
visíveis; B = grãos com puncturas, mas sem deformação; C = grãos com
punctura e com deformação; D = grãos totalmente deformados.......................
46
Figura 12. Número de grãos danificados por pentatomídeos de acordo com
quatro classes de danos em seis genótipos de soja de ciclo semiprecoce em
ensaios de campo. Botucatu-SP, 2011-2012 e 2012-2013. A = grãos sem
danos visíveis; B = grãos com puncturas, mas sem deformação; C = grãos
com punctura e com deformação; D = grãos totalmente deformados................ 55
Figura 13. Número de grãos danificados por pentatomídeos de acordo com
quatro classes de danos em cinco genótipos de soja de ciclo tardio em ensaios
de campo. Botucatu-SP, 2011-2012 e 2012-2013. A = grãos sem danos
visíveis; B = grãos com puncturas, mas sem deformação; C = grãos com
punctura e com deformação; D = grãos totalmente deformados....................... 64
Figura 14. Correlação entre densidade de tricomas e atratividade de adultos
de Euschistus heros, e dureza de vagem, em teste com chance de escolha em
seis genótipos de soja de ciclo precoce.............................................................. 73
Figura 15. Correlação entre densidade de tricomas e atratividade de adultos
de Euschistus heros, e dureza de vagem, em teste com chance de escolha em
seis genótipos de soja de ciclo semiprecoce....................................................... 74
Figura 16. Correlação entre densidade de tricomas e atratividade de adultos
de Euschistus heros, e dureza de vagem, em teste com chance de escolha em
seis genótipos de soja de ciclo tardio.................................................................
75
Figura 17. Mortalidade (%) nos diferentes estádios ninfais e emergência de
adultos de Euschistus heros confinados em seis genótipos de soja de ciclo
precoce.......................................................................................................... 80
Figura 18. Mortalidade (%) nos diferentes estádios ninfais e emergência de
adultos de Euschistus heros confinados em seis genótipos de soja de ciclo
semiprecoce........................................................................................................ 81
Figura 19. Mortalidade (%) nos diferentes estádios ninfais e emergência de
adultos de E. heros confinados em cinco genótipos de soja de ciclo tardio....... 82
Figura 20. Ninfas de quinto ínstar de Euschistus heros que morreram presas à
exúvia no momento da ecdise................................................................ 82
Figura 21. Viabilidade ninfal (% ± EP) de Euschistus heros obtida em 17
genótipos de soja (T= 26 2ºC, U.R.= 65 10% e fotoperíodo=14h)............. 83

1
1 RESUMO
A soja, Glycine max (L.) Merrill, apresenta muitas espécies de
insetos, sendo os percevejos pentatomídeos classificados em pragas primárias, responsáveis
por reduções no rendimento e na qualidade das sementes de soja. Pela alta ocorrência e
elevados danos ocasionados, destacam-se dentro do complexo as espécies Nezara viridula
(Linneaus, 1758), Piezodorus guildinii (Westwood, 1837), Euschistus heros (Fabricius,
1798), Dichelops sp. e Edessa meditabunda (Fabricius, 1794). Considerando-se a carência de
métodos de controle eficientes e menos agresssivos ao ambiente, este trabalho avaliou 17
genótipos de soja, pertencentes a três grupos de maturação frente ao ataque do complexo de
percevejos em duas safras, estudando-se ainda possíveis mecanismos de resistência destes
genótipos em laboratório sobre o percevejo de maior ocorrência, E. heros. Nesse sentido, foi
avaliada a campo: infestação, retenção foliar, produtividade e os danos ocasionados nos
genótipos. Em laboratório, também foram avaliadas a preferência alimentar, características
físicas e morfológicas das vagens e aspectos biológicos de E. heros. Os genótipos de ciclo
precoce PI 171451 e PI 229358 foram menos atrativos e danificados pelo complexo de
percevejos, sendo que PI 171451 também revelou baixa retenção foliar e elevada
produtividade, juntamente com ‘IAC 17’. A alta densidade de tricomas de ‘IAC 17’ influencia
o comportamento de E. heros. Dentre os genótipos semiprecoces, PI 227687 e IAC 74-2832
foram menos infestados, sendo que PI 227687 apresentou baixa retenção foliar, e juntamente
com ‘IAC 100’ e IAC 78-2318 manteve boa produtividade sob ataque de percevejos,
indicando tolerância. PI 227687 e ‘IAC 100’ foram os menos danificados, reafirmando

2
antixenose e/ou tolerância. As linhagens tardias PI 274453 e PI 274454 foram menos
infestadas e danificadas por percevejos, apresentando ainda baixa retenção foliar, indicando
tolerância e/ou antixenose para alimentação. Vagens de PI 274454 também foram pouco
atrativas a E. heros em laboratório. L1-1-01, ‘IAC 100’, ‘IAC 23’ e ‘Coodetec 208’
prolongaram o desenvolvimento ninfal, indicando a ocorrência de antibiose e/ou antixenose,
sendo que PI 274454, ‘IAC 19’, PI 227687 e PI 229358 causaram elevada mortalidade ninfal,
também indicando antibiose e/ou antixenose. Em PI 274453 e PI 227687 houve redução no
peso de adultos e na viabilidade de ovos.
Palavras-chave: resistência de plantas a insetos; percevejo-marrom; antixenose; antibiose.

3
SOYBEAN STINK BUGS: REFLECTIONS ON THE PRODUCTIVITY OF
GENOTYPES AND DETERMINATION OF THE TYPES OF RESISTANCE ON
Euschistus heros (FABRICIUS) (HEMIPTERA: PENTATOMIDAE). Botucatu, 2013. 106
p. Tese (Doutorado em Agronomia/Proteção de Plantas). Faculdade de Ciências
Agronômicas. Universidade Estadual Paulista.
Author: EFRAIN DE SANTANA SOUZA
Adviser: EDSON LUIZ LOPES BALDIN
Co-adviser: ANDRÉ LUIZ LOURENÇÃO
2 SUMMARY
Soybean, Glycine max (L.) Merrill, has many species of insects,
stink bugs being classified into primary pests, responsible for reductions in yield and
quality of soybean seeds. For high frequency and high damage occasioned, stand out
within the complex species Nezara viridula (Linneaus, 1758), Piezodorus guildinii
(Westwood, 1837), Euschistus heros (Fabricius, 1798), Dichelops sp. and Edessa
meditabunda (Fabricius, 1794). Considering the lack of efficient control methods and less
agresssive for the environment, this study evaluated 17 genotypes belonging to different
maturity groups were against the attacks of the stink bug complex in two seasons, and the
possible mechanisms of resistance of these genotypes in the laboratory against the bug that
occurred most E. heros. Accordingly, were evaluated infestation in the field: infestation,
leaf retention, productivity and damage caused in genotypes. In laboratory, were also
evaluated feeding preference, physical and morphological characteristics of the pods and
biological aspects of E. heros. The genotypes of early cycle PI 171451 and PI 229358 were
less attractive and damaged by stink bug complex, and PI 171451 also showed high
productivity and low leaf retention, along with 'IAC 17'. The high density of trichomes in
‘IAC 17' likely to influence the behavior of E. heros. Among semiearly genotypes, PI
227687 and IAC 74-2832 were less infested, and PI 227687 showed low leaf retention,
where together with ‘IAC 100’ and IAC 78-2318, maintained good yield under attack by
stink bugs, indicating tolerance. PI 227687 and 'IAC 100' were the least damaged,
reaffirming antixenosis and/or tolerance. In late genotypes, the lines PI 274453 and PI
274454 were less damaged and infested by stink bugs, yet having low leaf retention,
indicating tolerance and/or antixenosis for feeding. Pods of PI 274454 were also

4
unattractive to E. heros in laboratory. L1 -1-01, ‘IAC 100’, ‘IAC 23' and 'Coodetec 208'
prolonged the nymphal development, indicating antibiosis and/or antixenosis, and PI
274454, 'IAC 19', PI 227687 and PI 229358 caused high nymphal mortality, also indicating
antibiosis and/or antixenosis. PI 274453 and PI 227687 provoked significantly reduced at
the weight of adults and eggs viability.
Keywords: host plant resistance; neotropical brown stink bug; nonpreference; antibiosis.

5
3 INTRODUÇÃO
O cultivo comercial da soja [Glycine max (L.) Merrill] no Brasil
revolucionou a prática da agricultura no país, tanto pela abertura de novas fronteiras
agrícolas, como pela ampla possibilidade de uso de diferentes métodos de controle de
pragas (HOFFMANN-CAMPO; CORRÊA-FERREIRA; MOSCARDI, 2012). A soja se
destaca por ser uma fonte alimentar saudável disponível em muitos produtos e ao mesmo
tempo gerar energia renovável, na forma de biodiesel (PRADO, 2007). Segundo projeções
do Departamento de Agricultura dos Estados Unidos (USDA) e da Companhia Nacional do
Abastecimento (CONAB, 2013), na safra de 2013/14, o Brasil produzirá 84 milhões de
toneladas, correspondentes ao cultivo de mais de 28 milhões de hectares, mantendo o país
como o quarto maior consumidor de soja e o segundo maior produtor mundial,
permanecendo atrás somente dos EUA (AGRIANUAL, 2013). Trata-se da principal
atividade agrícola do Brasil, respondendo pela geração de cerca de cinco milhões de
empregos diretos e indiretos (ROESSING; LAZZAROTTO, 2004).
Durante o processo de produção, alguns fatores implicam na
redução da produtividade dessa leguminosa ou limitam a expansão para novas áreas,
destacando-se entre esses fatores, as intempéries climáticas, plantas invasoras, doenças e
pragas. Embora as pragas tenham suas populações reduzidas por predadores, parasitoides e
entomopatógenos, elas frequentemente atingem níveis populacionais elevados, causando
perdas significativas e necessitando serem manejadas (EMBRAPA, 2011).
Na soja, são muitas as espécies de insetos e suas populações, sendo
os nocivos classificados em pragas primárias e secundárias, de acordo com a frequência em
que ocorrem e com o nível de dano que podem acarretar à cultura (MIYASAKA, 1982).

6
No primeiro grupo, são relacionados os percevejos fitófagos,
responsáveis por reduções no rendimento e na qualidade das sementes de soja, em
consequência do hábito alimentar na vagem e por se serem vetores de patógenos como o
fungo Nematospora coryli (Pegliom, 1901), que ocasiona a deterioração das sementes
(PANIZZI; SLANSKY, 1985; BELORTE et al., 2003; DEGRANDE; VIVAN, 2010).
Ainda sobre os danos decorrentes da alimentação do complexo de percevejos sobre as
vagens, somam-se o enrugamento e deformação dos grãos, a redução na germinação e no
vigor das sementes, alterações no teor de óleo e proteína, além de também ocasionar
retenção foliar, conhecida no Brasil como “soja-louca” (PANIZZI; SLANSKY JÚNIOR,
1985; COSTA; BORGES; VILELA, 1998; CORSO; GAZZONI, 1998; CORRÊA-
FERREIRA; PANIZZI, 1999). Por esses motivos alguns autores (TURNIPSEED;
SULLIVAN, 1976; KOGAN, 1976; LUSTOSA et al., 1999; BELORTE et al., 2003),
classificam os pentatomídeos como as pragas mais importantes na cultura da soja.
Pela alta ocorrência e elevados danos ocasionados às lavouras
brasileiras de soja, destacam-se as espécies Nezara viridula (Linneaus, 1758), Piezodorus
guildinii (Westwood, 1837), Euschistus heros (Fabricius, 1798) Dichelops sp. e Edessa
meditabunda (Fabricius, 1794) (JENSEN; NEWSON, 1972; GALLO et al., 2002; SOSA-
GÓMEZ; SILVA, 2010), sendo que a flutuação populacional de cada espécie e a
severidade dos danos estão relacionados respectivamente com a temperatura e o estádio
fenológico das plantas (SILVA; CORRÊA-FERREIRA; SOSA-GÓMEZ, 2006;
BELORTE et al., 2003).
Entre os percevejos, E. heros é o que ocorre com maior frequência
nas regiões produtoras de soja do Brasil. Sua ocorrência aumentou consideravelmente após
o avanço da cultura para o Centro-Oeste, provavelmente se adaptando melhor às elevadas
temperaturas dessa região (FERNANDES; CORRÊA; BORTOLI, 1990; CIVIDANES;
PARRA, 1994; CORRÊA-FERREIRA et al., 2010).
Para o manejo do complexo de percevejos da soja, recorre-se
geralmente ao uso de inseticidas sintéticos (DEGRANDE; VIVAN, 2010), que embora
muito eficazes e recomendados, frequentemente ocasionam problemas como desequilíbrio
ambiental, intoxicação humana, resíduos em alimentos e eliminação de inimigos naturais
(BYRNE et al., 2003). Além disso, alguns autores (PRABHAKER; COUDRIET; MEYER-
DRIK, 1985; STANSLY; SCHUSTER, 1992; SOSA-GÓMEZ; CORSO; MORALES,
2001; HART; PIMENTEL, 2002; SILVA et al., 2009) ressaltam que aplicações sucessivas
de um mesmo produto químico favorecem rápida seleção de indivíduos resistentes. Esse

7
fato foi estudado e relatado em diversos trabalhos de Sosa-Gómez et al. (2005, 2009,
2010), que descrevem a ocorrência de elevada resistência do percevejo E. heros a grupos
de inseticidas em algumas regiões brasileiras. Segundo Corrêa-Ferreira e Moscardi (1996),
para o manejo de percevejos, são usados anualmente no Brasil quatro milhões de litros de
inseticidas sintéticos.
O uso de genótipos resistentes às pragas pode ser uma alternativa
sinérgica, redutiva ou até mesmo substitutiva aos inseticidas nas culturas (KOGAN, 1989;
KEA; TURNIPSEED; CARNER, 1978; PANDA, 1979; SMITH, 2005), apresentando
ainda vantagens como acessibilidade; compatibilidade com o controle biológico, sem a
exigência de conhecimento prévio; redução nos custos de cultivo; controle das populações
das pragas a níveis que não causem danos econômicos sem interferência no ecossistema,
tornando a cultura mais lucrativa para o agricultor (PAINTER, 1951; ROSSETTO et al.,
1981; KOGAN, 1989; LARA, 1991, PANDA; KHUSH, 1995; SMITH, 2005).
Em função do potencial de danos que o complexo de percevejos
ocasiona à cultura da soja, aliado à necessidade do desenvolvimento de métodos menos
agressivos ao ambiente, realizou-se este trabalho, objetivando avaliar a possível resistência
de genótipos de soja de diferentes grupos de maturação sob ataque do complexo de
percevejos da soja a campo. Ensaios de laboratório também foram realizados com E. heros
a fim de caracterizar diferentes mecanismos de resistência ao inseto.

8
4 REVISÃO DE LITERATURA
4.1 Cultura da soja
A soja [Glycine max L. (Merrill)] é uma leguminosa exótica no
Brasil, tendo como centro de origem o leste da China, onde foi domesticada no século XI
a.C. e introduzida posteriormente em outras regiões do Oriente. Registros antigos reportam
que essa leguminosa representa a base alimentar do povo chinês há mais de 5.000 anos
(CÂMARA, 1998).
O germoplasma de soja possui grande diversidade quanto ao ciclo,
variando de 70 dias para as mais precoces até 200 dias para as mais tardias (BORÉM,
1999; SEDIYAMA, 2009). O ciclo total da planta pode ser dividido em duas fases:
vegetativa (emergência da plântula até a abertura das primeiras flores) e reprodutiva (início
da floração até a maturação). Van Schaik e Probst (1958), citado por Borém (1999), uma
planta pode emitir até 800 flores, porém a taxa de fertilização varia de 13 a 57 %,
dependendo do genótipo e das condições ambientais.
No Brasil, essa planta foi introduzida no ano de 1882 por Gustavo
Dutra no Estado da Bahia (VERNETTI, 1983; SEDIYAMA; PEREIRA; SEDIYAMA,
1985), chegando ao Estado de São Paulo em 1891 para testes como planta forrageira no
Instituto Agronômico de Campinas (MYASAKA; MEDINA, 1981). A partir de 1941
iniciou-se o cultivo comercial no sul do país, chegando o Estado do Paraná a produzir
60.000 toneladas de grãos na década de 60 (VERNETTI, 1983; KASTLER; QUEIROZ;
TERASAWA, 1981). Na década seguinte, a cultura evoluiu nos estados produtores,
expandindo-se também para os estados do Centro-Oeste do país (BORÉM, 1999).

9
A soja é considerada uma commodity nacional, e apresenta-se como
uma das principais fontes de proteína e óleo vegetal do mundo para nutrição humana e
animal, sendo rica em vitaminas do complexo B, potássio, zinco, dentre outros minerais
(SHAW; CHIN; PAN, 1995; MORAIS; SILVA, 1996). Além da importância nutricional, a
planta ainda possui inúmeras utilizações, tais como adubação verde, fabricação de tintas,
fibras, adesivos, tecidos e cosméticos (CÂMARA, 1998).
Mundialmente, destacam-se no ranking de produção de soja os
Estados Unidos, o Brasil e a Argentina, que juntos respondem por aproximadamente 80%
da produção mundial deste grão. No Brasil, os Estados de Mato Grosso, Paraná, Goiás e
Rio Grande do Sul são os maiores produtores, respondendo por 38,82; 16,49; 12,43 e
9,83% respectivamente, da produção nacional (CONAB, 2013, AGRIANUAL, 2013).
Com relação à área ocupada pela cultura no Brasil, na última safra
(2012/13), o país cultivou 27,72 milhões de hectares, passando a produzir 81,46 milhões de
toneladas devido ao incremento de 10 % de área cultivada em relação à safra anterior. As
razões que alavancaram a cultura estão relacionadas ao elevado nível das cotações da
oleaginosa no mercado internacional, suas repercussões no mercado interno e ao bom
desempenho da comercialização realizada de forma antecipada, que na última safra atingiu
níveis recordes (CONAB, 2013; AGRIANUAL, 2013). Sob condições normais de
semeadura, a produtividade média de soja na safra 2012/13 foi de 2.935 kg/ha, havendo
um acréscimo de 11,4 % em relação à safra anterior (IBGE, 2013).
4.2 Características do complexo de percevejos da soja
Os percevejos das vagens e sementes da soja são fitófagos
pertencentes à família Pentatomidae. Segundo Gillot (2005), trata-se de uma das maiores
famílias dentro da subordem Heteroptera, com mais de 5.000 espécies descritas. A maioria
dos pentatomídeos apresenta coloração distinta, podendo ser marrom, esverdeada ou muito
colorida (CARVER; GROSS; WOODWARD, 1991).
Dentre as principais características morfológicas presentes nos
pentatomídeos, destacam-se as antenas com cinco segmentos e escutelo grande e
triangular, porém menor que o cório e não atinge o ápice do abdome e tíbia, podendo ainda
ter ou não espinhos. Esses percevejos normalmente medem mais de 7 mm e são castanho-
acinzentado, verdes, mas com muitas espécies coloridas e brilhantes. Apesar de a maioria

10
destes percevejos ser fitófaga, existem algumas espécies predadoras (BORROR; WHITE,
1970; CARVER; GROSS; WOODWARD, 1991).
Segundo Costa Lima (1940), os pentatomídeos geralmente passam
por cinco estádios ninfais antes de atingirem a fase adulta, sendo ápteros nos dois
primeiros ínstares e apresentando tecas alares nos três ínstares seguintes. Ainda, segundo o
mesmo autor, neste grupo os insetos passam a ter três artículos tarsais na fase adulta,
diferindo das ninfas que apresentam somente dois. Após a eclosão, as ninfas ficam
agregadas, permanecendo sobre o córion e não se alimentam durante o primeiro ínstar.
Provavelmente, o agrupamento é uma tática contra dessecação (SILVA, 2013). Para o
percevejo N. viridula, a agregação ninfal também diminui o tempo de desenvolvimento e a
mortalidade dos indivíduos (LOCKWOOD; STORY, 1986).
Quando as ninfas de pentatomídeos atingem o segundo ínstar,
começam a se dispersar entre as plantas de soja, porém, esta dispesão limita-se a distâncias
relativamente pequenas (PANIZZI; BUENO; SILVA, 2012). A dispersão ninfal foi
descrita por Panizzi et al. (1980), que estudando ninfas de N. viridula e P. guildinii,
observaram até 12 m do ponto inicial em áreas de cultivo de soja. Estes autores relataram
ainda que, as ninfas se deslocam mais no sentido longitudinal do que no transversal das
linhas de plantio, sendo as de quarto e quinto ínstares capazes de percorrer maiores
distâncias. No complexo de percevejos da soja, a dispersão dos adultos para outras áreas,
ocorre geralmente antes que a cultura complete a maturação, sendo comum a dispersão a
partir da senescência das plantas (CORRÊA-FERREIRA; PANIZZI, 1999; PANIZZI;
BUENO; SILVA, 2012).
Durante as últimas safras, tem-se observado um número crescente
de percevejos nas lavouras de soja, principalmente nos cultivos mais tardios
(DEGRANDE; VIVAN, 2009). Segundo estes autores, em certos casos, a população
desses pentatomídeos supera mais de doze vezes o nível de ação definido para estas pragas.
Em termos de abundância de espécies, registros da safra de soja 2001/2002 em Balsas-MA,
revelaram que 89 % dos pentatomídeos coletados a campo pertenciam à espécie E. heros
(PANIZZI, 2002). Mais recentemente, em Londrina-PR, na safra 2007/08, 84 % do total
dos percevejos também foram dessa espécie (CORRÊA-FERREIRA; MACHADO;
HOFFMANN-CAMPO, 2010). Há grande dispersão deste percevejo pelo território
brasileiro com predominância sobre as demais espécies (CIVIDANES; PARRA, 1994;
DEGRANDE; VIVAN, 2010, CORRÊA-FERREIRA; MACHADO; HOFFMANN-
CAMPO, 2010), o que revela a necessidade de maiores estudos, visando aprofundar os

11
conhecimentos sobre sua biologia e comportamento, a fim de disponibilizar ferramentas
mais eficientes de manejo.
4.3 Aspectos bioecológicos de Euschistus heros (Fabricius)
O percevejo E. heros (Hemiptera: Pentatomidae), vulgarmente
descrito no Brasil como “percevejo-marrom”, é apontado como umas das mais importantes
pragas da soja, recebendo atenção destacada em função dos danos e perdas que ocasiona à
esta cultura (RAMIRO, 1982; KISHINO; ALVES, 1992; GALLO et al., 2002). Vários
estudos de ocorrência e aspectos biológicos foram realizados com esta espécie (COSTA
LIMA, 1940; VILLAS BÔAS; PANIZZI, 1980; BORGES et al., 1998; COSTA;
BORGES; VILELA, 1998; MOURÃO; PANIZZI, 2000), revelando grande expansão deste
percevejo nas lavouras de soja do país nos últimos anos (PANIZZI; ROSSI, 1991;
PANIZZI; BUERNO; SILVA, 2012).
Quanto às características morfológicas de E. heros, os adultos
apresentam coloração marrom-escura ou avermelhada e possuem dois prolongamentos
laterais no protórax em forma de espinhos pontiagudos, o que facilita a identificação.
Medem aproximadamente 13 mm de comprimento (11 a 15 mm) e possuem uma mancha
branca típica em forma de “meia-lua” na extremidade do escutelo. Os ovos podem ser
amarelos, verde-claros ou beges e são normalmente depositados em massas de 6 a 15
unidades, dispostas em duas ou três fileiras paralelas. As formas imaturas passam por cinco
estádios ninfais, os quais apresentam coloração marrom suave uniforme. Ninfas de
primeiro ínstar apresentam hábito gregário, permanecendo agrupadas em colônias sobre os
córions dos ovos e não causando danos à cultura, passando a sugar os grãos de soja
somente no terceiro ínstar (COSTA; BORGES; VILELA, 1998; MOURÃO; PANIZZI,
2000; GALLO et al., 2002; DEGRANDE; VIVAN, 2009).
Segundo Costa et al. (1998), o período médio de incubação dos
ovos para a espécie E. heros é de 7,1 dias. As fêmeas que acasalam mais de uma vez
colocam uma maior quantidade de ovos. A longevidade média dos adultos é de 116 dias e
o desempenho reprodutivo das fêmeas varia conforme o tipo de alimento ingerido e a
constância alimentar durante a fase ninfal (VILLAS-BÔAS; PANIZZI, 1980; PINTO;
PANIZZI, 1994).

12
O primeiro registro desse percevejo em soja no Brasil foi feito por
Williams et al. (1973) no Estado de São Paulo na década de 70. Posteriormente, a espécie
dispersou-se para outras áreas de soja, como Goiás, Minas Gerais, Mato Grosso
(KISHINO; ALVES, 1992), São Paulo, Mato Grosso do Sul, Paraná (PANIZZI;
SLANSKY, 1985) e Rio Grande do Sul (PEREIRA; SALVADORI, 2008), sendo também
registrado recentemente, pela primeira vez na Argentina (SALUSO et al., 2011).
O aumento da ocorrência dessa espécie no país parece estar
relacionado à uma melhor adaptação do inseto às regiões de temperaturas mais elevadas,
constituindo-se atualmente num grande problema na região central do Brasil (PANIZZI;
SLANSKY, 1985; FERNANDES; CORRÊA; BORTOLI, 1990; CIVIDANES; PARRA,
1994). Em adição, Corrêa-Ferreira et al. (2010), estudando parâmetros bioecológicos dessa
espécie, constataram que as altas densidades populacionais, inclusive na fase vegetativa da
soja, estão também relacionadas com as práticas inadequadas de manejo, principalmente
pelo uso abusivo e errôneo de agrotóxicos.
O percevejo-marrom é um pentatomídeo nativo da América
Tropical, que apesar de polífago é pouco conhecido em relação às suas plantas
hospedeiras, sendo considerado o menos polífago em relação aos demais percevejos do
complexo (PANIZZI; SLANSKY, 1985; DEGRANDE; VIVAN, 2010). De acordo com
Panizzi (1997), E. heros produz duas ou três gerações em leguminosas durante os meses de
primavera-verão, e quando a soja é colhida, no final do verão, podem eventualmente se
alimentar da planta daninha carrapicho-de-carneiro, Acanthospermum hispidium (DC.),
dispersando-se posteriormente para abrigos, sob folhas caídas e restos de cultura no solo,
permanecendo em diapausa até o próximo verão. Quando em diapausa, geralmente
apresentam os órgãos reprodutivos ainda imaturos com os espinhos pronotais menos
desenvolvidos e coloração do corpo marrom-escura, diferente dos maduros sexualmente,
que são marrom-avermelhados (MOURÃO; PANIZZI, 2000). Nesse período, de
aproximadamente sete meses, o percevejo não se alimenta, sobrevivendo das reservas
lipídicas armazenadas anteriormente (HOFFMANN-CAMPO et al., 2000).
Além da soja, E. heros alimenta-se de diversas espécies de
Brassicaceae, Compositae, Euphorbiaceae, Leguminosae e Solanaceae (LINK, 1979;
CORRÊA-FERREIRA; PANIZZI, 1982; LINK; GRAZIA, 1987; PINTO; PANIZZI,
1994). Como hospedeiros alternativos para este percevejo, alguns autores (MOURÃO;
PANIZZI, 2000; DEGRANDE; VIVAN, 2009; PANIZZI, 2003) também relataram as
plantas amendoim-bravo (Pterogyne nitens Tul.), algodoeiro (Gossypium hirsutum L.),

13
ligustro (Ligustrum sp.), mamona (Ricinus communis L.), feijão (Phaseolus vulgaris L.),
ervilha (Pisum sativum L.) e alfafa (Medicago sativa L.). Frequentemente, o percevejo-
marrom também tem sido relatado na cultura do milho (Zea mays L.), causando injúrias de
diferentes intensidades em plântulas ou em espigas (ROSA-GOMES, 2010). No Estado de
Mato Grosso do Sul, a espécie vem atacando e comprometendo plantações de algodão
(Gossypium hirsutum L.) (SORIA; DEGRANDE; PANIZZI, 2010).
Devido ao hábito sugador, esse percevejo afeta irreversivelmente as
sementes, grãos e hastes de soja, constituindo-se atualmente como uma das principais e
mais abundantes pragas da cultura no Brasil (CIVIDANES; PARRA, 1994; SOSA-
GÓMEZ; OMOTO, 2012). Segundo Soria et al. (2010), o aumento populacional e o ataque
desta espécie em soja, tornaram-se mais frequentes principalmente em áreas cultivadas
anteriormente com algodão-Bt, provavelmente devido à redução na quantidade de
inseticidas aplicados para o controle de pragas do algodoeiro.
4.4 Danos associados ao complexo de percevejos da soja
Ao atingir a fase reprodutiva ou na transição entre as fases
vegetativa e reprodutiva, geralmente surgem na cultura da soja os percevejos fitófagos da
família Pentatomidae (HOFFMANN-CAMPO et al., 2000). Dentro desse grupo destacam-
se um complexo de espécies, reconhecidos em ordem decrescente de importância
econômica e de ocorrência: o percevejo-marrom E. heros, o percevejo-verde-pequeno
P.guildinii, o percevejo-verde N. viridula e os percevejos-barriga-verde (Dichelops spp.).
Embora as espécies Chinavia sp. e E. meditabunda tenham aumentado consideravelmente
sua ocorrência nas lavouras de soja, ainda são consideradas de menor expressão
(PANIZZI; SLANSKY, 1985; VILLAS BÔAS et al., 1985; GALLO et al., 2002;
CORRÊA-FERREIRA; KRZYZANOWSKI; MINAMI, 2009; DEGRANDE; VIVAN,
2009, PANIZZI; BUENO; SILVA, 2012; SOSA-GÓMEZ; OMOTO, 2012).
Estes percevejos são sugadores que se alimentam durante todo o
período reprodutivo das plantas, tendo o pico populacional concentrado entre março e abril
(CORRÊA et al., 1977; CORRÊA-FERREIRA; PANIZZI, 1982; PANIZZI; SLANSKY,
1985). À medida que a soja amadurece, torna-se menos atrativa e adequada para a
alimentação e reprodução desses percevejos, ocorrendo então uma dispersão para as
cultivares tardias e/ou hospedeiros alternativos (CORRÊA-FERREIRA; PANIZZI, 1999;

14
PANIZZI; ROSSI, 1991; PANIZZI; NIVA 1994). Alguns autores (CIVIDANES; PARRA,
1994; MAGRINI et al., 1996) afirmam que é comum a variação populacional de espécies
de percevejos de região para região e de ano para ano, e essa variação está diretamente
relacionada com a cultivar semeada e com as condições climáticas locais.
Segundo Basso (2009), os percevejos podem sugar várias estruturas
da planta, mas se concentram nas vagens. Degrande e Vivan (2009) afirmam que o ataque
desses pentatomídeos às vagens pode ocasionar perdas superiores a 30 %, uma vez que as
mesmas tornam-se chochas (sem formação de grãos), secam e escurecem. Em ataques
intensos, as sementes podem perder mais de 50 % do poder germinativo, com redução de
vigor e alterações nos teores de óleo e proteínas, comprometendo a qualidade e o valor
final dos grãos e sementes (RAMIRO; MASSARIOL, 1977; VILLAS BOAS et al., 1990;
GALLO et al., 2002;). Como sinal visual do ataque de percevejos, observam-se puncturas
nos grãos e manchas escuras em áreas esbranquiçadas nas vagens, derivadas de espaços de
ar produzidos quando os insetos ingerem os conteúdos celulares (MINER, 1966; SOSA-
GÓMEZ et al., 2010).
Além dos danos diretos, a alimentação desses percevejos nas
vagens, favorece a transmissão do fungo Nematospora coryli (Peglio, 1901), causador da
doença “mancha-de-levedura” ou “mancha-fermento” (CORSO, 1984; GAZZONI;
YORINIORI, 1995; HORI, 2000). Também como dano secundário, durante a alimentação
ocorre a injeção de toxinas, frequentemente relacionadas ao distúrbio fisiológico conhecido
como “soja-louca” e caracterizado pela retenção anormal de folhas, maturação irregular de
vagens e presença de hastes verdes, obrigando o produtor recorrer a dessecantes para a
posterior colheita (VICENTINI; JIMENEZ, 1977; GALILEO; HEINRICHS, 1978; SOSA-
GÓMEZ; MOSCARDI, 1995; GALLO et al., 2002; DEGRANDE; VIVAN, 2010).
4.5 Métodos de controle para o complexo de percevejos da soja
Em função da ampla expansão da cultura da soja no Brasil e dentro
do que preconiza o Programa de Manejo Integrado de Pragas da soja, as decisões são
baseadas no nível de ataque, no número de insetos e no estádio de desenvolvimento da
soja, sendo as amostragens realizadas semanalmente utilizando-se o pano-de-batida, uma
vez que a simples observação visual sobre as plantas não permite dimensionar a população
real de percevejos presentes nas lavouras (DEGRANDE; VIVAN, 2009; PANIZZI;
BUENO; SILVA, 2012).

15
Durante a Reunião da Comissão de Soja em 2007, estabeleceu-se
um novo nível de ação para os percevejos da soja, iniciando-se o controle quando
observada uma média igual ou superior a dois percevejos adultos ou ninfas com mais de
0,5 cm por metro. Essa recomendação é reduzida pela metade em lavouras destinadas à
produção de sementes (EMBRAPA, 2011; CORRÊA-FERREIRA, 2012).
Dentre as táticas de controle recomendadas para as pragas da soja,
o método químico é o mais utilizado e tem trazido reconhecidos benefícios à cultura ao
longo dos anos, principalmente pela redução populacional de insetos-praga. De acordo
com informações do Ministério da Agricultura (AGROFIT, 2013), os principais inseticidas
registrados para o controle do complexo de percevejos da soja são: acefato, ciproconazol,
cipermetrina, beta-ciflutrina, imidacloprido, lambda-cialotrina, tiametoxam, parationa-
metílica, bifentrina, zeta-cipermetrina, malation, esfenvalerato, fenitrotiona, clorpirifós e
carbosulfano, com uso associado ou isolado desses ingredientes ativos dependendo da
formulação.
Entretanto, o uso abusivo de inseticidas de amplo espectro em
mistura com herbicidas, ou por ocasião das aplicações de fungicidas (“aproveitamento de
operações”), tem ocasionado grande desequilíbrio ambiental em algumas regiões, com
graves problemas relacionados a pragas. Como exemplo disso, observam-se a elevação dos
níveis populacionais das principais pragas e a mudança de status de insetos de importância
secundária, que sob condições especiais causam danos que justificam medidas de controle.
Somados a isso, a elevada ocorrência de populações do complexo de percevejos na fase
inicial do desenvolvimento da cultura, aliada à falta de conhecimentos por parte de
técnicos e produtores sobre os possíveis danos, leva ao uso inadequado de produtos
químicos sintéticos, fechando um ciclo de práticas inadequadas (CORRÊA-FERREIRA,
2005; EMBRAPA, 2010; ALEXANDRE, 2010; ROGGIA, 2010).
Dos problemas decorrentes do mau uso de inseticidas químicos
(ROSSETTO et al., 1984; PRABHAKER; COUDRIET; MEYER-DRIK, 1985; ELBERT;
NAUEN, 2000; BYRNE et al., 2003; AHMAD et al., 2002; BAUR et al., 2010), tem
destaque os desequilíbrios no ambiente, eliminação de insetos benéficos e frequentes
intoxicações ao homem, além da seleção de indivíduos resistentes.
No Brasil, alguns estudos foram realizados e constataram a
resistência de percevejos a inseticidas, sendo a espécie E. heros aquela que mais
apresentou-se resistência até o momento (SOSA-GÓMEZ; CORSO; MORALES, 2001;
SOSA-GÓMEZ; SILVA, 2010). A ineficiência de controle do percevejo-marrom na soja

16
utilizando inseticidas começou a ser detectada na década de 1990; porém, na época, as
falhas de controle foram atribuídas a problemas de formulação do endosulfam, um dos
inseticidas mais usados no período (SOSA GÓMEZ; OMOTO, 2012). Segundo os mesmos
autores, nos dias atuais, produtos como endosulfam foram retirados do mercado, restando
os piretroides e os neonicotinoides como os mais viáveis para o manejo de percevejos.
Algumas alternativas para a obtenção de melhores resultados no
manejo químico dos percevejos da soja foram divulgadas. Como exemplo cita-se o uso do
controle químico somente nas bordas da lavoura em início de infestação, sem aplicação na
área total, reduzindo os custos de produção. Outra alternativa é a adição de sal de cozinha
(0,5 % v/v) na calda inseticida, pois o sal age como arrestante sobre os percevejos. No
entanto, essa prática é polêmica, uma vez que o sal desencadeia reações de ferrugem nos
equipamentos de aplicação (CORSO, 1989; CORSO, 1990; NIVA; PANIZZI, 1996;
SOSA-GÓMEZ; OMOTO, 2012).
Com relação ao uso de agentes de controle biológico no manejo de
percevejos, existem exemplos de sucesso no país, onde os entomopatógenos, predadores e
principalmente os parasitoides de ovos têm sido relatados com frequência no campo
(BUENO et al., 2012). Ragsdale et al. (1981) e Thomazini e Gravena (1994) citaram o
percevejo Geocoris punctipes (Say, 1832) como um importante predador de ovos e ninfas
de percevejos pragas da soja. Esse gênero se caracteriza pela presença de olhos
proeminentes, que os levam a ser conhecidos como big-eyed bugs, sendo um predador de
ocorrência comum em todas as regiões produtoras de soja do Brasil (BUENO et al., 2012;
CIVIDANES; BARBOSA, 2001).
Medeiros et al. (1997) relataram vários agentes biológicos que
atuam no controle de ovos de pentatomídeos, com destaque para os parasitoides Trissolcus
basalis (Wollaston) e Telenomus podisi (Ashmead). Corrêa-Ferreira e Moscardi (1995)
reportaram 20 espécies de quatro famílias de micro-himenópteros parasitando ovos de
pentatomídeos em campos de soja, oito delas encontradas sobre ovos de E. heros, sendo
eles: Trissolcus basalis, Ttrissolcus brochymenae (Ashmead), Trissolcus urichi
(Crawford), Trissolcus sp., Telenomus podisi (Ashmead), Gryon obesum (Masner),
Ooencyrtus sp. e Neorileya ashmeadi (Crawford), com percentagem média de parasitismo
de 60 % na safra de 1989/1990 e de 59 % na de 1990/1991. Telenomus podisi é a espécie
mais comum de ocorrência no Brasil, associada ao percevejo sugador de sementes mais
abundante E. heros, seu principal hospedeiro (GODOY et al., 2005, 2007)

17
Como parasitoides de percevejos adultos, existem algumas espécies
representantes da família Tachinidae e alguns micro-himenópteros, podendo-se citar
Trichopoda giacomellii (Blanchard, 1966) parasitando principalmente N. viridula e o
encirtídeo Hexacladia smithii (Ashmead, 1891) (Hemiptera: Encyrtidae) parasitando E.
heros (CORRÊA-FERREIRA; PANIZZI, 1999; BUENO et al., 2012).
Segundo Corrêa-Ferreira e Panizzi (1999), o controle microbiano
exercido pelos fungos entomopatogênicos Beauveria bassiana (Vuill) e Metarhizium
anisopliae (Metsch) também contribuem para reduzir as populações de percevejos. No
entanto, a ocorrência desses fungos é baixa (controle de 0,5 % da população),
especialmente devido à resistência apresentada pelos percevejos à infecção e à necessidade
de condições climáticas favoráveis à sua multiplicação em campo (MOSCARDI, et al.,
1988). Sosa-Gómez et al. (1997) relataram que esta baixa infecção em condições naturais
se deve, entre outros fatores, à composição dos feromônios liberados pelos percevejos, que
são ricos em aldeídos [(E)-2-decenal] de ação fungistática, principalmente sobre M.
anisopliae.
Também como prática alternativa para o controle de pragas nas
lavouras, o uso de materiais resistentes pode ser uma estratégia valiosa (PAINTER, 1951;
KOGAN, 1986; LARA, 1991; SMITH, 2005). Dentre as principais vantagens desse
método sobre os demais, destacam-se o não acúmulo de resíduos nos alimentos e a não
poluição do ambiente, o benefício aos agricultores, a atuação contra qualquer nível de
infestação da praga, a não interferência em outras práticas agrícolas como a colheita (sem
carência), sendo de fácil acesso ao agricultor, a redução das populações de insetos-praga a
níveis que não causam danos econômicos, a não alteração do equilíbrio do
agroecossistema, a facilidaede de utilização, a compatibilidade com os demais métodos de
controle, a não exigência de conhecimentos específicos do agricultor, além de não onerar o
custo de produção (VENDRAMIM, 1990; LARA, 1991).
A resistência é classificada em três tipos: antixenose ou não-
preferência, antibiose e tolerância, sendo que esses fenômenos podem envolver alterações
comportamentais ou biológicas do inseto e, em outros casos, uma reação da própria planta,
que em nada afeta o inseto. A resistência por antixenose ocorre quando há uma alteração
no comportamento do inseto durante a seleção do hospedeiro, causada pela presença de
compostos aleloquímicos ou características morfológicas da planta, tornando-a menos
favorável à alimentação, abrigo e/ou oviposição pelos insetos. A antibiose é expressa por
um conjunto de caracteres da planta que afetam negativamente a biologia dos insetos que a

18
utilizaram como alimento. Geralmente, as causas da antibiose estão relacionadas à
produção de aleloquímicos pelas plantas, em que durante a alimentação, os insetos podem
ser afetados das seguntes formas: redução no potencial reprodutivo; elevada mortalidade
da fase imatura; baixa emergência de adultos; redução no tamanho e peso dos indivíduos;
alteração da longevidade, oviposição, mortalidade, fecundidade, entre outros. A tolerância
ocorre quando o genótipo mantém as características produtivas (quantitativa e
qualitativamente) sob o ataque da praga, sem afetar o comportamento ou a biologia da
mesma (PANDA, 1979; AUCLAIR, 1989; LARA, 1991; PANDA; SMITH, 2005;
VENDRAMIM; GUZZO, 2009).
Na cultura da soja, o uso de genótipos resistentes a insetos em
programas de melhoramento ganhou força após a identificação de três introduções (PI-
171451, PI-227687 e PI-229358), portadoras de resistência ao besouro-mexicano
Epilachna varivestis (Mulsant, 1850), nos EUA (DUYN; TURNIPSEED; MAXWELL,
1971). Posteriormente, foi observada resistência desses genótipos também contra
Helicoverpa zea (Boodie, 1850), Cerotoma trifurcata (Forster, 1771) (CLARK et al.,
1972), Plathypena scabra (Fabricius, 1798), Epicauta spp. e Trichoplusia ni (Hübner,
1803) (LUEDDERS; DICKERSON, 1977).
No Brasil, programas de melhoramento de soja têm sido realizados
há alguns anos pelo Instituto Agronômico de Campinas, SP (IAC), buscando a
incorporação de fatores de resistência, principalmente para insetos sugadores e
desfolhadores (MIRANDA; LOURENÇÃO, 2002). Nesses programas, linhagens como PI-
171451, PI-227687, PI-229358, PI-274453 e PI-274454 vêm sendo utilizadas como
genitores resistentes, pois manifestam resistência múltipla (KOGAN, 1989; MIRANDA;
LOURENÇÃO 2002). Outros materiais derivados dessas linhagens também foram
utilizados como fonte de genitores resistentes, como IAC 73-228, IAC 78-2318
(MIRANDA et al., 1979; LOURENÇÃO; MIRANDA, 1987) e D72-9601 (REZENDE;
MIRANDA; MASCARENHAS, 1980).
Em trabalho visando avaliar a resistência de diferentes genótipos de
soja sob ataque de percevejos, Rossetto et al. (1986) observaram que o genótipo ‘IAC-
80/4228’ apresentou tolerância em comparação com outros 35 genótipos. Entre possíveis
fatores associados à resistência de soja aos percevejos, Rossetto et al. (1995) citaram que
no caso da cultivar resistente ‘IAC-100’, pelo menos cinco fatores de resistência estão
envolvidos, sendo eles: menor período de enchimento de vagens; maior número de
sementes; aborto de vagens danificadas e substituição por novas vagens; senescência

19
normal com queda das folhas na maturação; e resistência à levedura N. coryli transmitida
por E. heros. Segundo Lourenção et al. (1997), as cultivares ‘IAC 100’ e ‘IAC 17’ são um
marco no melhoramento de soja no Brasil quanto à resistência a insetos, sendo os
primeiros materiais liberados para esta finalidade.
Avaliando a resistência de oito genótipos de soja à percevejos da
soja, Lourenção et al. (1987) constataram que, entre dez genótipos, as linhagens IAC 73-
228 e IAC-78-2318 foram os mais resistentes em condições de campo, após dois anos de
safra. Miranda et al. (2003) verificaram que a cultivar ‘IAC 24’ também apresenta
resistência a insetos nocivos da soja, com melhor desempenho agronômico em relação aos
demais genótipos testados. Fugi et al. (2005) ratificaram as características dessa cultivar e
também da ‘IAC 17’ ao verificarem antibiose para Anticarsia gemmatalis (Hübner,1818).
Recentemente, com a finalidade de avaliar as linhagens mais
produtivas e com maior resistência a danos causados pelo complexo de percevejos,
Lourenção et al. (2010) conduziram experimentos por duas safras consecutivas e
constataram que as linhagens IAC 98-4540, IAC 98-4576, IAC 98-3123, IAC 98-4136 e as
cultivares ‘IAC 18’ e IAC 98-4136 foram os genótipos mais promissores.
As pesquisas realizadas até o momento são de grande importância e
têm contribuído para os programas de melhoramento de soja, não só visando à obtenção de
genótipos com resistência ao complexo de percevejos, mas também para insetos
desfolhadores. Entretanto, considerando-se a importância crescente do complexo de
percevejos e em especial de E. heros nas lavouras de soja, estudos específicos são
necessários, visando ampliar o conhecimento sobre as interações deste inseto com a cultura
da soja. Em adição, a seleção de genótipos resistentes pode representar uma ferramenta
adicional para o manejo dessa praga nas lavouras.
Com base nisso, o presente trabalho avaliou a possível resistência
de genótipos de soja de diferentes grupos de maturação sob ataque do complexo de
percevejos da soja a campo. Ensaios de laboratório também foram realizados com E. heros
a fim de caracterizar diferentes mecanismos de resistência sobre o inseto.

20
5 MATERIAL E MÉTODOS
O presente trabalho foi realizado em condições de campo, na
Fazenda de Ensino, Pesquisa e Extensão (FEPE) e nos Laboratórios de Resistência de
Plantas a Insetos e Plantas Inseticidas (LARESPI) do Departamento de Proteção Vegetal,
da FCA-UNESP, Campus de Botucatu-SP, entre os anos de 2010 a 2013.
5.1 Genótipos de soja utilizados
Para a execução do trabalho, foram utilizados 17 genótipos de soja
(Tabela 1), divididos em três grupos de maturação (precoce, semiprecoce e tardio).
As cultivares ‘IAC 17’, ‘IAC 19’ e ‘IAC 24’, foram selecionadas
por apresentarem antixenose e antibiose para mosca-branca e lagartas desfolhadoras
(VALLE; LOURENÇÃO, 2002; VALLE et al., 2012) e para o percevejo-verde N. viridula
(LOURENÇÃO et al., 2000; MIRANDA et al., 2003). A cultivar ‘IAC 100’ foi incluída
por ser resistente a vários insetos, incluindo N. viridula (McPHERSON; BUSS;
ROBERTS, 2007). L1-1-01 é derivado deste material e portanto é possível que tenha
herdado estas características de resistência. As linhagens PI 171451, PI 229358, PI 227687,
PI 274453 e PI 274454 demonstraram resistência a vários insetos (VAN DUYN;
TURNIPSEED; MAXWELL, 1971, 1972; TURNIPSEED, 1977; KOGAN, 1989). As
linhagens D75-10169, IAC 78-2318 e IAC 74-2832 são derivados de PI 229358 e PI
274454 (MIRANDA et al., 1979; LOURENÇÃO; MIRANDA, 1987; KILEN;

21
LAMBERT, 1993). ‘Conquista’ e ‘ Coodetec 208’ são cultivares amplamente cultivadas
no Brasil, sendo incluídos como padrões de suscetibilidade.
Tabela 1. Genealogia dos genótipos de soja de ciclos precoce, semiprecoce e tardio
utilizados no trabalho.
Ciclo Genótipo Genealogia
Precoce
‘IAC-17’ D72-9601-1 x ‘IAC-8’
‘IAC-23’ ‘BR-6’ x IAC-83-23
PI-171451 Japão
PI-229358 Tóquio, Japão
D 75-10169 ‘Govan’ x (F4 ‘Bragg’ x PI 229358)
‘Coodetec-208’ OC-4 x ‘Williams 20’
Semiprecoce
‘IAC 18’ D72-9601 x ‘IAC-8’
‘IAC 24’ IAC80-1177 x IAC83-288
‘IAC 100’ 'IAC-12' x IAC-78-2318
IAC 74-2832 ‘Hill’ x PI 274454
IAC 78-2318 D72-96-1 x IAC 73-227
PI 227687 Okinawa, Japão
Tardio
‘IAC 19’ D72-9601-1 x ‘IAC-8’
PI 274453 Okinawa, Japão
PI 274454 Okinawa, Japão
L 1-1-01 ‘BR-6’ x ‘IAC-100’
‘Conquista’ Lo76-44842
x ‘Numbaíra’
5.2 Obtenção dos genótipos
Os genótipos foram fornecidos pelo Dr. André Luiz Lourenção do
Instituto Agronômico (IAC), Campinas, SP. Devido à baixa quantidade de sementes de
alguns materiais, foi necessário multiplicá-los a campo, visando obter quantidades
suficientes para todos os testes propostos (Figura 1A). A área de multiplicação de sementes
classifica-se como Latossolo Vermelho Distroférrico, segundo levantamento detalhado

22
realizado por Carvalho et al., (1983) e pelo Sistema Brasileiro de Classificação dos Solos
(EMBRAPA, 2006). Cerca de 50 dias antes da semeadura, fez-se a correção da acidez do
solo com utilização de calcário dolomítico no momento do preparo do solo, seguido por
aração e gradagem. A adubação foi realizada durante a semeadura, utilizando a fórmula
20-20-20 na base de 300 kg ha-1
, de acordo com análise de fertilidade do solo e as
recomendações para a cultura da soja (RAIJ; CANTARELLA; QUAGGIO, 1997).
Antes da semeadura, as sementes foram inoculadas com
Bradyrhizobium japonicum (Kirchner, 1896), na dosagem de 0,4 mL do produto comercial
(Masterfix L) para cada 100 g de semente, recebendo também tratamentos com fungicida
Vitavax-Thiram 200 SC (carboxina e tiram) e inseticida Cropstar (imidacloprid e
tiodicarbe) nas doses de 300 e 500 mL/100 kg de semente, respectivamente. Quando
necessário, as plantas foram irrigadas via aspersão. Realizaram-se também pulverizações
alternadas com os inseticidas Engeo Pleno (tiametoxam e lambda-cialotrina) e Akito (beta-
cipermetrina) e os fungicidas Ópera (piraclostrobina e epoxiconazol) e Priori Xtra
(azoxistrobina e ciproconazol) recomendados para a cultura, objetivando a proteção das
plantas de eventuais ataques de pragas e doenças.
As sementes foram colhidas manualmente quando apresentavam
umidade próxima a 14 % (Instrução Normativa n° 11 de 15/05/2007), sendo
posteriormente limpas, selecionadas, embaladas hermeticamente e acondicionadas em
câmara fria para posterior uso nos ensaios (Figura 1B).
Figura 1. A - Área semeada com os genótipos de soja para a multiplicação de sementes; B -
sementes colhidas, selecionadas e acondicionadas hermeticamente.
A B

23
5.3 Criação estoque de E. heros
Para início da criação de E. heros em laboratório, seguiu-se a
metodologia proposta por Silva et al. (2007), com T= 26 2ºC, U.R.= 65 10% e
fotoperíodo=14 h, obtendo-se massas de ovos junto ao Laboratório de Biologia de Insetos
da ESALQ/USP, Piracicaba, SP. Os ovos foram acondicionados em placas de Petri (6 cm
de diâmetro) contendo papel filtro ao fundo e uma pequena porção de algodão embebido
com água destilada, conforme proposto por Costa et al. (1998). Após a eclosão, as ninfas
foram mantidas na placa até atingirem o segundo ínstar, sendo posteriormente liberadas em
recipientes plásticos (40 x 22 x 14 cm) forrados com papel filtro ao fundo. Para a
alimentação, utilizou-se-se vagens de feijão (Phaseolus vulgaris L.) e amendoim (Arachis
hypogaea L.), mantendo-se também um recipiente plástico (50 mL) contendo algodão e
água destilada ao fundo, servindo como fonte de água e auxiliando na manutenção da
umidade do recipiente. Visando evitar a fuga, foi aplicada vaselina sólida na borda superior
do recipiente, acoplando-se também uma tampa adaptada com tecido voil, permitindo a
aeração (Figura 2).
Figura 2. A - Gaiola de criação de adultos de E. heros e placas contendo ovos; B - vista
interna da gaiola de criação; C - ninfas buscando água em algodão embebido.
(T= 26 2ºC, U.R.= 65 10% e fotoperíodo=14 h).
Visando favorecer e concentrar as posturas, foram adicionados
quatro pedaços de tecido de algodão cru no interior de cada gaiola de criação. As posturas
A B C

24
foram diariamente recolhidas, a fim de evitar o consumo dos ovos pelos próprios
percevejos, o que segundo Panizzi (1990), ocorre com frequência em criações
laboratoriais. Estes ovos foram separados do tecido e acondicionados em placas de Petri,
conforme citado anteriormente (Figura 2). A manutenção da criação foi feita a cada três
dias, conforme descrito por Souza (2010), colocando os insetos em gaiolas limpas com
alimento novo e água destilada, evitando-se assim, a proliferação de microrganismos.
Periodicamente, adultos de E. heros foram coletados em áreas
cultivadas com soja, com o auxílio de um pano de batida (1 m x 0,5 m) e introduzidos nas
gaiolas, visando manter o vigor da criação e evitar a degeneração resultante da seleção e
cruzamento entre irmãos. Durante as coletas a campo, optou-se por áreas cultivadas com
genótipos diferentes dos utilizados neste trabalho, a fim de se evitar o condicionamento
pré-imaginal (LARA, 1991). As coletas foram feitas em sua maioria no período da manhã,
até as 9:00h e no final da tarde após as 16:00h, evitando-se o sol forte, uma vez que,
segundo Panizzi (1990), na presença deste, os percevejos ficam menos expostos.
5.4 Ensaios a campo
Foram realizados dois ensaios de campo visando avaliar a
atratividade e colonização do complexo de percevejos sobre diferentes genótipos de soja
em condições normais de cultivo, a fim de caracterizar a possível ocorrência de resistência.
O primeiro ensaio foi realizado no ano agrícola 2011/2012 em uma
área experimental localizada a 22º82’48’’S e 48º42’80’’O, com altitude aproximada de 720
m. O segundo ensaio (2012/2013) foi instalado a 22°50’39’’S e 48°25’28’’O com altitude
de 780 m, sendo ambas as áreas pertencentes à Fazenda de Ensino, Pesquisa e Extensão
(FEPE) da FCA/UNESP, Campus de Botucatu, SP.
As áreas receberam preparo prévio, 20 dias antes da semeadura,
com dessecação via pulverização de herbicida Roundup WG (glifosato), sendo as parcelas
semeadas em sistema de plantio direto. A fertilização foi realizada no momento da
semeadura, baseada em análises de fertilidade de solo realizadas pelo Departamento de
Solos da FCA/UNESP (Anexos 1 e 2) e recomendações para a cultura da soja (RAIJ;
CANTARELLA; QUAGGIO, 1997).
Em cada um dos ensaios, uma área de aproximadamente 8.000 m2
foi dividida em três talhões e cada um deles abrigou um grupo de soja (precoce,

25
semiprecoce e tardio). Para correlacionar os genótipos com os danos ocasionados pelos
percevejos e os reflexos na produtividade, foram semeados quatro repetições de cada
material sem controle de percevejos, e outras quatro repetições onde foi realizado o
controle quando detectado o nível de ação (DEGRANDE; VIVAN, 2010, EMBRAPA,
2011). Ao todo foram instaladas 68 parcelas sem aplicação de inseticidas e 68 parcelas por
ensaio onde se realizaram aplicações, sendo as parcelas distribuídas em delineamento de
blocos casualizados (Figura 3).
As parcelas possuíam 6,75 m2 cada, contendo quatro linhas de
plantio espaçadas em 0,45 m (Figura 3). As dimensões foram 1,35 x 5,0 m e o
espaçamento entre as bordas das parcelas de 1,5 m, tanto frontal quanto lateralmente. Os
blocos tratados foram espaçados em dez metros daqueles onde não houve aplicações de
inseticidas, evitando o possível efeito de deriva das pulverizações nas parcelas não
tratadas.
Figura 3. A - Vista geral da área experimental do primeiro ensaio realizado em 2011-
2012; B - área do segundo ensaio realizado em 2012-2013.
Antes da semeadura, as sementes dos diferentes genótipos foram
tratadas com fungicida (Vitavax-Thiram 200 SC – carboxina e tiram) e inseticida (Cropstar
– imidacloprid e tiodicarbe) na dose de 300 mL e 500 mL para 100 kg de semente,
respectivamente. Posteriormente, estas sementes também foram inoculadas com B.
japonicum, na dose de 200 mL do produto comercial (Masterfix L) para cada 50 kg de
sementes, sendo homogeneizadas e secas à sombra. As sementes foram distribuídas
A B

26
manualmente na densidade de 20 sementes por metro linear e após 14 dias da emergência,
foi realizado o desbaste manual, mantendo-se uma densidade média de 16 plantas por
metro linear.
Quando os percevejos atingiam o nível de ação nas parcelas que
seriam tratadas (média de um percevejo por metro linear) era realizado o controle com
pulverizações de inseticida à base de lambda-cialotrina + thiamethoxam (Engeo Pleno) na
dose de 200 mL para 200 L de calda ha-1
. As aplicações foram realizadas com um
pulverizador costal elétrico modelo Ft-16 Yamaho, com capacidade para 16 L e bico
cônico regulável. As amostragens foram realizadas no momento das avaliações,
verificando-se a necessidade ou não do controle. Para o primeiro ensaio (2011/2012),
foram necessárias três aplicações nos genótipos de ciclo precoce e quatro nos demais, a
partir do início do florescimento da soja até a fase de colheita. Já para o segundo ensaio
(2012/2013) realizaram-se três aplicações para todos os genótipos.
Quando as plantas atingiram o período reprodutivo,
especificamente próximo ao estádio R2 (florescimento pleno), iniciaram-se as avaliações
semanais (sempre ao final da tarde) da infestação dos percevejos E. heros, P. guildinii, N.
viridula, E. meditabunda e Dichelops spp. nos genótipos de soja (Figura 4). Para as
avaliações foi utilizado um pano de batida padrão para soja, medindo 1m de comprimento
e 0,5m de largura, sustentado lateralmente por hastes de madeira (CORRÊA-FERREIRA;
PAVÃO, 2005). O pano de batida era cuidadosamente posicionado entre as linhas de
plantio, sobre o qual, na sequência, as plantas eram inclinadas e sacudidas alternadamente
(CORRÊA-FERREIRA; PANIZZI, 1999). Por meio desse método, foram amostrados dois
pontos por parcela sempre nas linhas centrais, contabilizando-se o número total de
percevejos (ninfas maiores de 0,5 cm e adultos) presentes no pano (Figura 4).
Foram realizadas avaliações semanais nas parcelas de cada grupo
de soja, totalizando oito avaliações. Após o término do período de avaliações, as plantas
das parcelas passaram a ser monitoradas quanto ao teor de umidade dos grãos com o
auxílio de uma caneca (modelo Multigrain), e quando estes apresentavam índices próximos
a 14%, foram colhidos manualmente quatro metros lineares das linhas centrais (Figura 5),
a fim de obter a produtividade média (kg.ha-1
) e nível de retenção foliar (%).
No momento da colheita, também foi avaliada a porcentagem de
retenção foliar e hastes verdes para posterior correlação com a presença dos percevejos.
Essa porcentagem foi obtida por meio de uma média por parcela da estimativa visual de
dois avaliadores (LOURENÇÃO et al., 2010).

27
Figura 4. A - Planta entrando em estádio reprodutivo R2; B - avaliação populacional de
percevejos pentatomídeos (E. heros, P. guildinii, N. viridula, E. meditabunda e
Dichelops spp.) com pano de batida.
A produtividade média dos genótipos cultivados nos dois ensaios
foi obtida por pesagem dos grãos em balança analítica modelo Marte AY 220 (0,0001g)
em laboratório. Para determinação desse parâmetro, adotou-se um arranjo fatorial (6 x 2
para os genótipos precoces e semiprecoces e 5 x 2 para os tardios) em delineamento de
blocos casualizados, com quatro repetições para as parcelas controladas e quatro para as
não controladas.
Figura 5. A - Colheita de quatro metros lineares centrais das parcelas; B - Classificação do
nível de dano nos grãos dos diferentes genótipos.
A classificação do nível de dano nos grãos foi determinada com
base na classificação proposta por JENSEN e NEWSON (1972), representada pelos níveis
A B
A B

28
1: grãos sem danos visíveis; 2: grãos com puncturas; 3: grãos com puncturas e com
deformação, e 4: grãos totalmente deformados. A classificação ocorreu retirando-se
amostras de 50 grãos de cada parcela, os quais foram classificados em seguida de acordo
com seus respectivos níveis de dano (Figura 5). Os dados obtidos foram analisados por
comparação de classe de dano e por uma abordagem de análise de dados categóricos
considerando os genótipos como uma variável nominal e a classe de dano como uma
variável ordinal. Para levar em conta o grau de dano sofrido pela semente, foram atribuídos
escores de 0 a 3, na ordem crescente aos danos causados pelos percevejos, ou seja, A = 0,
B = 1, C = 2, D = 3. Uma estatística Escore Médio que se aproxima de um Qui-quadrado
foi utilizada para verificar se há associação entre os genótipos e a classe de dano e qual(is)
genótipo(s) dentre os analisados classificam-se como mais resistentes ao ataque dos
percevejos.
5.5 Ensaios em laboratório
No laboratório (T= 26 2ºC, U.R.= 65 10% e fotoperíodo=14 h),
foram realizados testes apenas com o percevejo E. heros, visto ser a espécie de maior
predominância, com base nas avaliações de campo.
5.5.1 Atratividade e preferência alimentar
Para identificar os genótipos de soja menos atrativos e consumidos
por E. heros, foram realizados ensaios de atratividade e preferência alimentar com chance
de escolha, utilizando-se vagens verdes dos diferentes genótipos, analisados dentro de cada
grupo (precoce, semiprecoce e tardio). As vagens utilizadas foram coletadas de plantas
oriundas de casa de vegetação apresentando estágio fenológico R5, conforme descrição
proposta por Fehr e Caviness (1977). Os percevejos utilizados eram provenientes da
própria criação, tendo passado para o estágio adulto em no máximo 24 horas e
posteriomente mantidos em jejum por mais 24 horas antes do início dos ensaios (Figura 6).
Para esses testes, foram utilizadas arenas redondas de PVC (30 cm
de diâmetro x 8 cm de altura) forradas na base com papel filtro sobre isopor, onde foram
apoiadas duas vagens de cada genótipo e distribuídas equidistantemente ao acaso. Antes da

29
liberação dos percevejos, foi aplicada uma camada de vaselina sólida nas bordas superiores
da arena, evitando a fuga dos insetos. Liberaram-se posteriormente, no centro da arena,
dois percevejos adultos por genótipo (Figura 6).
Figura 6. A - Percevejos mantidos sob jejum de 24 h; B - Vista geral de uma arena; C
- Adulto de E. heros se alimentando em vagem; D - Testes em execução.
(T=26 2ºC, U.R.=6510% e fotoperíodo=14h).
Durante as avaliações anotou-se o número de insetos presentes em
cada genótipo aos 15, 30, 45, 60, 120 e 180 minutos após a liberação. Simultaneamente,
também foi observado o consumo dos percevejos sobre as vagens dos genótipos,
contabilizando-se o número e o tempo de picadas para alimentação, sendo possível calcular
esses valores ao final do ensaio (Figura 6). Para a determinação do número de picadas nas
vagens, realizou-se contagem a olho nu, conforme descrito por Rossetto et al. (1981) e
Souza (2010). O tempo de picada foi determinado com o auxílio de cronômetros digitais,
acionando-os ao início da alimentação de cada inseto e cessando a marcação quando o
percevejo retirava o estilete da vagem. Os ensaios foram gravados caso alguma repetição
A B
C D

30
necessitasse ser reavaliada. O registro foi feito por uma câmera digital modelo Sony
CyberShot W510 suspensa por tripé de alumínio e acoplada a uma placa de captura de
vídeo (EasyCap 2.0), sendo as filmagens armazenadas em um HD externo de computador.
Esses ensaios seguiram um delineamento experimental de blocos
casualizados, com dez repetições para cada grupo de maturação de soja estudada (precoce,
semiprecoce e tardio).
5.5.2 Quantificação e comprimento de tricomas em vagens
Para correlacionar a densidade e o comprimento dos tricomas das
vagens dos diferentes genótipos com a preferência alimentar de E. heros foi realizada
inicialmente a contagem do número de tricomas presentes em 0,25 cm2, baseando-se na
metodologia utilizada por Souza (2010) e Paron e Lara (2005). Para a contagem, foram
coletadas ao acaso, vagens em estágio fenológico R5 nos diferentes genótipos cultivados
de casa de vegetação. Essas vagens foram submetidas à observação em um
estereomicroscópio (Nikon SMZ-685) onde sobre o primeiro grão (região próxima ao
pedicelo) foi posicionado um delimitador de papel milimetrado, anotando-se
posteriormente o número total de tricomas (unciformes e aciculares) observados na área
delimitada. Escolheu-se o primeiro grão, próximo ao pedicelo da vagem pelo fato de ser a
região de maior preferência alimentar da espécie E. heros, segundo observações de Panizzi
et al. (1995).
Para a medição do comprimento dos tricomas, vagens também em
estádio R5 foram coletadas e enviadas ao Centro de Microscopia Eletrônica da Esalq-USP,
onde foram processadas e fotografadas num microscópio eletrônico de varredura modelo
Zeiss LEO435-VP. As melhores imagens obtidas foram analisadas no Laboratório de
Resistência de Plantas a Insetos e Plantas Inseticidas (LARESPI) do Departamento de
Produção Vegetal da FCA-UNESP, usando-se o software EDN-2 na medição dos tricomas
(Figura 7). Foram contabilizados os tricomas/0,25 cm2
em dez vagens de cada genótipo de
soja e medidos 20 tricomas em cada genótipo. Para análise, cada observação representou
uma repetição, seguindo um delineamento inteiramente casualizado.

31
Figura 7. A - Imagem dos tricomas de vagem obtida por
microscopia eletrônica; B - medição do
comprimento dos tricomas através do software
EDN-2.
5.5.3 Determinação da dureza das vagens
Para correlacionar a dureza das vagens dos genótipos de soja com o
consumo alimentar de E. heros foram realizadas medições de dureza da casca dessas
vagens, simulando a dificuldade do percevejo em atingir o grão. Estas medições foram
feitas em texturômetro, modelo CT3 Texture Analyzer, calibrado para penetração de 10
mm e velocidade de 2,0 mm seg-1
, utilizando-se o ponteiro TA 9/1000.
A
B

32
As avaliações foram padronizadas em vagens provenientes de
plantas em estádio fenológico R5, testando-se sempre a resistência de penetração do
primeiro grão da vagem (próximo ao pedicelo). Os resultados das medições foram
expressos em grama-força por centímetro quadrado (gf/cm2), e representam a força
máxima requerida para que uma parte do ponteiro penetrasse na polpa da vagem,
simulando a real situação em que o inseto encontra ao inserir seu aparelho bucal na vagem.
Avaliaram-se dez vagens de cada genótipo, sendo que, para análise, cada vagem
representou uma repetição em delineamento inteiramente casualizado.
5.5.4 Antibiose
O desempenho biológico do percevejo E. heros em diferentes
genótipos de soja foi avaliado em laboratório (T= 26 2ºC, U.R.= 65 10% e
fotoperíodo=14 h), baseado em metodologias descritas (COSTA; BORGES; VILELA,
1998; PIUBELLI et al., 2003). Durante o ensaio foram avaliados os seguintes parâmetros
biológicos:
a) Período de duração dos estádios ninfais (N2, N3, N4 e N5) e duração ninfal;
b) Viabilidade dos instares ninfais e do período nilfal;
c) Peso de adultos, com no máximo 24 h de idade;
d) Longevidade dos adultos;
e) Viabilidade de ovos.
Para iniciar o ensaio de antibiose, foram coletadas da criação em
laboratório massas de ovos recém ovipositados (com até 24h). Esses ovos foram
confinados em um recipiente forrado com papel filtro na base e algodão umidecido com
água destidada, visando suprir a necessidade hídrica das ninfas que mais tarde surgiriam.
Após a eclosão e no momento que atingiram o estádio N2, as ninfas foram acondicionadas
aos pares em placas de Petri (9 cm), também revestidas ao fundo com papel filtro, um
bastonete de algodão umedecido e uma vagem em fase de desenvolvimento R5/R6 de um
dos genótipos avaliados (Figura 8). Cada placa representou uma repetição, totalizando 25
repetições por genótipo, em delineamento inteiramente casualisado.

33
Figura 8. A - Placa de Petri contendo vagem e duas ninfas de E. heros; B - Genótipo
cultivado em casa-de-vegetação para o fornecimento de vagens; C - Vista
geral de parte do ensaio de antibiose. (T=262ºC, U.R.=6510% e
fotoperíodo=14h).
A opção por se iniciar o ensaio com ninfas de percevejo a partir do
segundo ínstar é justificada pela observação de elevada taxa de mortalidade natural com
ninfas de primeiro ínstar, fato que poderia comprometer a obtenção dos resultados. Outro
fator que também justifica a escolha das ninfas de segundo ínstar, deve-se ao hábito
gregário, característico das ninfas de primeiro ínstar de percevejos da família Pentatomidae
(GALLO et al., 2002; PANIZZI; SILVA, 2009).
A cada dois dias, realizou-se a troca das vagens e a porção de
algodão foi novamente umedecida, conforme a necessidade aparente ou trocada quando
necessário. A cada cinco dias, era trocado o papel filtro do fundo, pois frequentemente
estes ficavam com muitas excreções. Exúvias e insetos mortos foram retirados no
momento das avaliações, com auxílio de uma pinça metálica. Diariamente, as placas eram
inspecionadas, sempre ao final da tarde, avaliando-se o desenvolvimento biológico dos
insetos.
Ao atingir a fase adulta, dez insetos de cada genótipo foram
transferidos para placas de Petri (3 cm de diâmetro) e pesados individualmente em uma
balança analítica modelo Marte AY220 (0,0001g). Após a pesagem, os adultos foram
devolvidos para as placas onde se desenvolveram, visando acompanhar o desenvolvimento
e determinar a longevidade dos adultos em cada genótipo. O acompanhamento do peso de
adultos foi realizado somente nos genótipos que propiciaram essa fase, já que alguns
materiais não permitiram o completo desenvolvimento ninfal. Ao final das avaliações,
A C B

34
obteve-se uma média dos parâmetros biológicos obtidos com os indivíduos acondicionados
nas placas/genótipos.
Paralelamente, para a obtenção dos parâmetros de viabilidade de
ovos dos percevejos confinados aos diferentes genótipos, aproximadamente 120 ovos de
um dia de idade foram acondicionados em arenas plásticas (8 L, com 22 cm de diâmetro e
20 cm de altura), recobertas com tecido voil e tendo a superfície inferior revestida com
papel filtro e um bastão de algodão umidecido, apoiando dez vagens no estádio R5 dos
diferentres genótipos. A limpeza e troca de vagens era feita a cada dois dias, retirando as
exúvias, insetos mortos e vagens velhas que eram substituídas por outras recém-colhidas.
Quando os percevejos atingiram a fase adulta, foram adicionadas tiras de tecido de algodão
cru nas arenas para concentrar e estimular a oviposição. Todos os ovos depositados eram
retirados, contados e acondicionados em placas de 3 mm (idênticas às usadas para
pesagem), onde esperou-se a eclosão, obtendo-se os dados referentes à viabilidade de ovos.
Nesta etapa, foram selecionadas somente oito placas (oviposições) por genótipo,
correspondendo a oito repetições.
5.6 Análises estatísticas
Para todos os testes de hipóteses foi adotado um nível de
significância de 5%. Os dados obtidos foram submetidos ao teste F da análise de variância
em que se verificaram as pressuposições do teste pela normalidade dos erros, através do
teste de Shapiro-Wilk e a homogeneidade de variâncias pelo teste de Levene.
Para as análises onde houve significância do teste F, procedeu-se o
teste de comparação múltipla de Tukey, para distinguir as diferenças.
Nas análises de danos, utilizou-se uma abordagem de dados
categóricos, considerando genótipo como uma variável nominal e classe de dano como
uma variável ordinal, realizando uma análise de Escore Médio para apontar os genótipos
mais resistentes em relação às classes de dano.
Para auxiliar na obtenção das análises, utilizou-se o software
estatístico R-2.15.0.

35
6 RESULTADOS E DISCUSSÃO
6.1 Ensaios a campo
6.1.1 Flutuação populacional de percevejos pentatomídeos
No gráfico referente à flutuação populacional dos percevejos nos
dois anos agrícolas (2011-2012 e 2012-2013), verifica-se que em ambos os ensaios a
população atingiu o nível de dano econômico (NDE), que segundo Hoffmann-Campo et al.
(2000), é de quatro percevejos adultos ou ninfas com mais de 0,5 cm por batida de pano
(Figura 9). As espécies consideradas foram E. heros, P. guildinii, N. viridula, E.
meditabunda e Dichelops spp., sendo o NDE atingido na segunda avaliação (29/02) nos
materiais de ciclo precoce e na terceira avaliação (07/03) para os materiais de ciclo
semiprecoce e tardio no ensaio de 2011-2012 (Figura 9). Esse comportamento de
colonização já era esperado, uma vez que os materiais de ciclo precoce iniciaram a fase
reprodutiva com maior antecedência, atraindo os percevejos.

36
Figura 9. Flutuação populacional de percevejos pentatomídeos (E. heros,
P. guildinii, N. viridula, E. meditabunda e Dichelops spp.) nos
ensaios com os diferentes genótipos de soja de ciclos precoce,
semiprecoce e tardio.
No segundo ensaio (2012-2013), nota-se menor ocorrência
populacional de percevejos, quando comparado com o ensaio anterior (Tabela 10). O NDE
foi atingido somente após a sexta avaliação (01/03) em todos os materiais de diferentes
ciclos.
De modo geral, as avaliações populacionais de percevejos fitófagos
da soja ocorreram em períodos diferentes nos dois ensaios, iniciando-se em fevereiro na
safra de 2011-2012 e em janeiro na safra de 2012-2013. A antecipação do plantio na safra
2012-2013 permitiu que os genótipos entrassem no período reprodutivo quando as
populações de percevejos ainda estavam baixas (primeiras avaliações), pois segundo

37
Correia-Ferreira e Panizzi (1999), o pico populacional de pentatomídeos da soja concentra-
se entre os meses de março a abril, estando em crescimento nos meses de janeiro e
fevereiro. Isso sugere que a antecipação do plantio favorece uma assincronia fenológica
(LARA, 1991), reduzindo a necessidade de manejo dos percevejos.
6.1.2 Ocorrência de espécies de percevejos pentatomídeos
Com relação às espécies de percevejos encontrados nos ensaios,
observou-se predominância de E. heros, representando 42 e 34 % do total de espécies de
insetos registrados no primeiro e segundo ensaios respectivamente (Figura 10).
42%
29%
22%
4% 3%
34%
24%
29%
7% 6%
E. heros P. guildinii N. viridula Dichelops spp. E. meditabunda
% d
e p
erce
vejo
s
2011-2012
2012-2013
Figura 10. Ocorrência (%) de percevejos pentatomídeos (E. heros, P. guildinii,
N. viridula, E. meditabunda e Dichelops spp.) registrados em ensaios
de campo com soja.
As espécies P. guildinii e N. viridula ocorreram praticamente na
mesma proporção nos dois ensaios. A ocorrência de P. guildinii foi de 29 % no primeiro
ensaio e de 24 % no segundo; para N. viridula, a ocorrência foi de 22 e 29 % no primeiro e
segundo ensaio, respectivamente. Os percevejos Dichelops spp. e E. meditabunda foram os
que menos incidiram, com médias abaixo de 10 % em ambos os ensaios (Figura 10).
ssp.

38
Ramiro et al. (1988), realizando um levantamento de pragas em
quatro cultivares e duas linhagens de soja, já haviam relatado os percevejos E. heros e P.
guildinii como os mais abundantes no campo. Lourenção et al. (2010) reportaram que 58 %
de toda a população de pentatomídeos registrados em ensaio a campo em Assis-SP no ano
agrícola 2003-2004 eram E. heros. Estas observações corroboram os relatos na literatura,
de que as espécies E. heros, P. guildinii e N. viridula são as mais abundantes na cultura da
soja. Em razão desta elevada ocorrência e pelos danos que acometem, estas continuam a se
destacar como as principais espécies do complexo de percevejos da soja (PANIZZI;
SLANSKY JUNIOR, 1985; HOFFMANN-CAMPO et al., 2000; GALLO et al., 2002;
SOSA-GÓMEZ et al., 2010).
6.1.3 Atratividade, retenção foliar, produtividade e danos ocasionados
por percevejos pentatomídeos em genótipos de soja
6.1.3.1 Ciclo Precoce
As médias obtidas revelaram uma grande variação na ocorrência
destes percevejos nos genótipos ao longo das avaliações (Tabela 2).
No ensaio da safra 2011-2012, observou-se, já na primeira
avaliação dos genótipos precoces (22/fev.), preferência dos percevejos pela cultivar
‘Coodetec 208’, que diferiu dos demais tratamentos, apresentando uma média de dez
insetos/batida de pano. Na avaliação seguinte, verificou-se aumento geral nas médias de
percevejos por batida de pano em todos os genótipos, com destaque para ‘Coodetec 208’
que novamente se revelou o mais atrativo, diferindo de PI 171451, o menos atrativo no
período. Entre a terceira e quinta avaliações, não foi detectada diferença entre os materiais
de soja. Na sexta avaliação, observou-se menor média de percevejos em ‘IAC 23’ (5,6), PI
171451 (5,6) e ‘Coodetec 208’ (3,0). Na sétima avaliação não houve diferença entre os
genótipos. Já na última avaliação (12/abr.), ‘Coodetec 208’ foi o menos atrativo entre os
demais (Tabela 2).
As médias gerais de infestação de percevejos nos diferentes
genótipos de ciclo precoce durante o primeiro ensaio (2011-2012) mostram não haver
diferença entre os tratamentos, embora estes tenham apresentado médias acima do NDE
(quatro percevejos) (Tabela 2).

39
No segundo ensaio de atratividade, realizado no ano agrícola de
2012-2013 (Tabela 2), não houve diferença entre os genótipos na primeira avaliação
(25/jan.), sendo também possível verificar menor ocorrência de percevejos em relação ao
ano anterior. Na segunda e terceira avaliações, ‘Coodetec 208’ se revelou o mais atrativo,
diferindo dos demais genótipos. Na sexta avaliação (01/mar), os genótipos voltaram a
diferir, com destaque para PI 229358 (1,5) e D75-10169 (0,7) que foram os menos
atrativos, diferindo de ‘Coodetec 208’.
Considerando-se todas as avaliações do ensaio de 2012-2013, os
genótipos ‘IAC 17’, PI 171451, PI 229358 e D75-10169 foram os menos infestados pelos
percevejos fitófagos, enquanto que ‘Coodetec 208’ revelou-se o mais atrativo.
De modo geral, observa-se maior média de percevejos nas
avaliações do primeiro ensaio, revelando maior ocorrência populacional desses
pentatomídeos naquele ano agrícola (2011-2012) (Tabela 2). Porém, deve-se considerar
que as avaliações no primeiro ensaio iniciaram-se em fevereiro, enquanto que as do
segundo ensaio tiveram início em janeiro, onde as populações de percevejos ainda estavam
baixas (Figura 9).
Nos dois ensaios, as populações dos pentatomídeos começaram a
aumentar a partir do mês de março. Isso provavelmente está correlacionado com o período
em que as vagens normalmente se encontravam (R5/R6), favorecendo a alimentação dos
percevejos. Essas observações corroboram os dados de Costa e Link (1974) e Corrêa-
Ferreira e Panizzi (1999), que reportaram maior concentração de pentatomídeos da soja
nos meses de março e abril. Segundo os mesmos autores, após esse período, as populações
tendem a cair, principalmente pela senescência das plantas e dispersão pós a colheita.
Os resultados obtidos com as linhagens PI 171451 e PI 229358
corroboram os dados de Rossetto et al. (1986), que já reportaram estes materiais como
detentores de resistência a percevejos. Acredita-se que a possível presença de voláteis
indesejáveis nas vagens/folhas, possa estar associada à menor colonização dos percevejos
nestes genótipos.
‘Coodetec 208’ mostrou elevada suscetibilidade aos percevejos da
soja, confirmando os resultados obtidos por Lourenção et al. (2004), que compararam em
campo essa cultivar com outros genótipos de soja de ciclo precoce para resistência a
percevejos.

40
Tabela 2. Número médio (±EP) de percevejos pentatomídeos (E. heros, P. guildinii, N. viridula, E. meditabunda e Dichelops spp.) por pano
de batida obtidos em plantas de seis genótipos de soja de ciclo precoce durante dois anos de ensaio a campo. Botucatu-SP, 2011-
2012 e 2012-2013.
N° médio de insetos (2011 / 2012)1
Genótipo Avaliação 01 Avaliação 02 Avaliação 03 Avaliação 04 Avaliação 05 Avaliação 06 Avaliação 07 Avaliação 08 Média 22 fev. 28 fev. 07 mar. 14 mar. 22 mar. 29 mar. 05 abr 12 abr.
‘IAC 17’ 4,0 ± 1,2 b 4,0 ± 1,3 ab 6,8 ± 1,2 8,6 ± 1,7 8,6 ± 2,1 11,4 ± 1,7 a 6,2 ± 1,6 9,0 ± 1,1 a 7,3 ± 0,2
‘IAC 23’ 2,4 ± 0,7 b 5,6 ± 1,1 ab 10,4 ± 1,7 10,6 ± 1,7 6,8 ± 1,1 5,6 ± 1,0 b 4,6 ± 1,1 5,4 ± 0,9 ab 6,4 ± 0,6
PI 171451 2,4 ± 0,7 b 1,4 ± 0,4 b 6,0 ± 0,9 5,2 ± 1,8 5,4 ± 1,4 5,6 ± 0,5 b 4,2 ± 1,1 5,2 ± 0,6 ab 4,4 ± 0,5
PI 229358 1,0 ± 0,7 b 2,8 ± 1,2 ab 7,4 ± 1,0 8,8 ± 2,9 7,4 ± 2,2 8,2 ± 1,0 ab 3,6 ± 0,5 6,4 ± 0,5 ab 5,6 ± 0,2
D75-10169 2,2 ± 0,6 b 4,4 ± 0,7 ab 8,6 ± 1,9 9,6 ± 1,5 6,8 ± 1,6 6,4 ± 2,0 ab 6,2 ± 2,2 6,8 ± 2,1 ab 6,3 ± 1,1
‘Coodetec 208’ 10,0 ± 1,6 a 6,6 ± 1,1 a 12,2 ± 1,7 9,0 ± 2,2 4,2 ± 1,2 3,0 ± 0,7 b 3,2 ± 0,7 3,4 ± 0,5 b 6,4 ± 0,8
P <0,001 0,019 0,054 0,559 0,512 0,002 0,478 0,038 0,060
N° médio de insetos (2012 / 2013)1
Genótipo Avaliação 01 Avaliação 02 Avaliação 03 Avaliação 04 Avaliação 05 Avaliação 06 Avaliação 07 Avaliação 08 Média 25 jan. 01 fev. 08 fev. 15 fev. 22 fev. 01 mar. 08 mar. 15 mar.
‘IAC 17’ 0,7 ± 0,5 1,0 ± 1,0 b 0,0 ± 0,0 b 0,2 ± 0,2 0,2 ± 0,2 3,0 ± 0,7 ab 4,2 ± 1,3 4,7 ± 2,4 1,7 ± 0,3 b
‘IAC 23’ 1,0 ± 0,6 0,7 ± 0,2 b 1,2 ± 0,6 ab 0,5 ± 0,5 0,5 ± 0,5 6,5 ± 1,7 ab 4,5 ± 0,6 8,2 ± 4,1 2,9 ± 0,3 ab
PI 171451 0,0 ± 0,0 0,2 ± 0,2 b 0,0 ± 0,0 b 1,0 ± 0,6 1,0 ± 0,6 2,7 ± 1,5 ab 3,5 ± 1,0 3,0 ± 1,5 1,4 ± 0,3 b
PI 229358 0,2 ± 0,2 0,0 ± 0,0 b 1,0 ± 0,5 b 0,2 ± 0,2 0,2 ± 0,2 1,5 ± 0,5 b 4,7 ± 1,5 5,7 ± 2,9 1,7 ± 0,2 b
D75-10169 0,5 ± 0,5 0,5 ± 0,3 b 0,0 ± 0,0 b 0,2 ± 0,2 0,2 ± 0,2 0,7 ± 0,8 b 2,2 ± 0,5 7,0 ± 3,5 1,4 ± 0,2 b
‘Coodetec 208’ 1,2 ± 0,6 4,0 ± 0,9 a 2,7 ± 1,4 a 1,2 ± 0,5 1,0 ± 0,4 9,0 ± 3,0 a 6,5 ± 2,4 6,7 ± 3,4 4,0 ± 0,3 a
P 0,428 0,001 <0,001 0,370 0,522 0,013 0,423 0,493 <0,001
1/Médias seguidas pela mesma letra na coluna não diferem entre si pelo Teste de Tukey (α = 0,05). Dados originais.

41
Apesar de a cultivar ‘IAC 17’ ter se revelado como atrativa em
algumas avaliações no primeiro ensaio (2011-2012), a literatura descreve esta cultivar
como portadora de resistência do tipo tolerância (LOURENÇÃO et al., 2002;
LOURENÇÃO et al., 2004; LOURENÇÃO et al., 2010).
Os dados referentes à retenção foliar ocasionada pelos percevejos
nos genótipos de ciclo precoce revelaram menor média na linhagem PI 171451 (13,75%),
diferindo dos genótipos ‘IAC 17’ (60,5%), ‘IAC 23’ (75%), ‘Coodetec 208’ (70%) e D75-
10169. Já a linhagem PI 229358 apresentou média de retenção intermediária (47,50%), não
diferindo dos demais genótipos (Tabela 3).
Tabela 3. Retenção foliar média (±EP) ocasionada pelo ataque de
percevejos em seis genótipos de soja de ciclo precoce.
Botucatu, 2011-2012 e 2012-2013.
Genótipo Retenção (%)1
PI 171451 13,75 ± 3,61 b
PI 229358 47,50 ± 16,52 ab
‘IAC 17’ 60,50 ± 5,20 a
‘Coodetec 208’ 70,00 ± 10,00 a
‘IAC 23’ 75,00 ± 8,66 a
D75-10169 78,75 ± 4,27 a
P <0,001
1/Médias seguidas pela mesma letra na coluna não diferem entre si pelo teste de
Tukey (α = 0,05). Dados originais.
A retenção foliar apresentada pelas plantas pode ser decorrente do
ataque de algumas espécies de percevejos, onde as plantas tendem a florescer novamente e
produzir novas vagens, o que dificulta a colheita pelo retardamento da senescência das
plantas (ROSSETTO et al., 1986). Para este distúrbio fisiológico, também é comum a
denominação “soja-louca” (LUSTOSA et al. 1999).
Os dados obtidos nas avaliações de retenção foliar (Tabela 3) para
a cultivar ‘IAC 17’ contrastam com os obtidos em outros trabalhos realizados
anteriormente, que a classificam como resistente ao ataque de percevejos por apresentar
baixa retenção foliar. Lourenção et al. (2000), estudando esta cultivar, observaram uma

42
retenção foliar média de 7,1%, em relação a outros genótipos, e em estudo semelhante,
Fernandes et al. (1994) constataram uma retenção foliar média de 20,87 %. No presente
trabalho, levanta-se a hipótese de que este cultivar tenha apresentado alta retenção foliar
decorrente da elevada ocorrência de percevejos no primeiro ensaio, colocando em questão
a capacidade da expressão da resistência sob elevados níveis populacionais de percevejos.
No entanto, para confirmar esta hipótese, serão necessários estudos complementeares que
revelem essa limitação do cultivar ‘IAC 17’. A baixa retenção foliar (Tabela 3) observada
na linhagem PI 171451 (13,75%) confirma os resultados obtidos por Rossetto et al. (1986)
com este material que relatam uma retenção foliar média de 14,3% para essa linhagem,
comparando-a dentro de um grupo de 35 genótipos.
Quanto à produtividade dos genótipos de ciclo precoce, nota-se que
o uso de inseticidas possibilitou maior produtividade em todos os genótipos nos dois
ensaios, em relação às parcelas infestadas por percevejos (Tabela 4). No primeiro ensaio
(2011-2012), sob condições controladas, ‘Coodetec 208’ e ‘IAC 23’ foram os mais
produtivos, com produções médias de 3.708 e 3.609 kg ha-1
respectivamente, diferindo de
PI 229358, com produtividade média de 2.550 kg ha-1
(Tabela 4). O mesmo
comportamento de ‘Coodetec 208’ e ‘IAC 23’ foi observado no segundo ensaio (2012-
2013), apresentando-se novamente como os mais produtivos em condições controladas
(3.522 e 3.486 kg ha-1
respectivamente) e diferindo de PI 229358 e PI 171451 (Tabela 4).
Quando avaliada a produtividade dos genótipos nas parcelas
infestadas, observa-se que a presença dos percevejos reduziu as produtividades em todos
os materiais nos dois ensaios (Tabela 4). Essa redução foi mais acentuada para D75-10169
(91 %) e ‘Coodetec 208’ (86 %) no primeiro ensaio (2011-2012) e ‘Coodetec 208’ (79 %)
no segundo ensaio. Nas mesmas condições, os genótipos ‘IAC 17’ e PI 171451 foram mais
produtivos entre os tratamentos, com produtividades de 1.808 e 1.708 kg ha-1
(primeiro
ensaio) com redução na produtividade de 47 % e 38 % respectivamente. No segundo
ensaio esses genótipos se comportaram de modo semelhante ao primeiro, sendo a média de
‘IAC 17’ igual a 1.711 kg ha-1
e PI 171451 igual a 1.909 kg ha-1
, com reduções de 49 % e
33 % respectivamente. As baixas médias de redução na produtividade (%) em PI 171451
observada nos dois ensaios sugerem tolerância nesta linhagem ao ataque de percevejos
(Tabela 4).

43
Tabela 4. Produtividade média (kg/ha-1
) (±EP) e percentagem de redução de produção de seis genótipos de soja de ciclo
precoce em parcelas infestadas e não infestadas por percevejos. Botucatu, 2011-2012 e 2012-2013.
1/Médias seguidas pela mesma letra minúscula na coluna e maiúscula na linha não diferem entre si pelo teste de Tukey (α = 0,05). Dados originais.
Genótipo
Produtividade (kg/ha-1
)
Ensaio de 2011 - 2012 Ensaio de 2012 - 2013
Controle1 Infestada
1 % Redução Controle
1 Infestada
1 % Redução
‘IAC 17’ 3.417 ± 138 abA 1.808 ± 372 aB 47 3.376 ± 103 abA 1.711 ± 86 aB 49
‘IAC 23’ 3.609 ± 161 aA 914 ± 134 abB 75 3.486 ± 67 aA 1.076 ± 161 bB 69
PI 171451 2.796 ± 165 abA 1.708 ± 266 aB 38 2.872 ± 173 bcA 1.909 ± 186 aB 33
PI 229358 2.550 ± 163 bA 896 ± 164 abB 65 2.575 ± 171 cA 1.014 ± 84 bB 60
D75-10169 3.260 ± 357 abA 297 ± 23 bB 91 2.986 ± 225 abcA 1.014 ± 41 bB 66
‘Coodetec 208’ 3.708 ± 242 aA 523 ± 25 bB 86 3.522 ± 193 aA 737 ± 74 bB 79
P <0,001 --- <0,001 ---

44
As médias de produtividade apresentadas pela linhagem PI 171451
nos dois ensaios sob condições de infestação, se aproximam da produtividade obtida por
Rossetto et al. (1986), que avaliando este material frente ao ataque de percevejos, obteve
produtivade média de 1.895 kg ha-1
. É possível que a baixa infestação de percevejos em PI
171451 (Tabela 2) tenha promovido maior produtividade, caracterizando a expressão de
resistência por não-preferência.
Em trabalho de campo, Lourenção et al. (2002) verificaram que a
cultivar ‘IAC 17’ foi a mais produtiva em relação aos demais genótipos testados, com uma
média de produção igual a 1.733 kg ha-1
no município de Ribeirão Preto, SP. Este resultado
é semelhante aos do presente estudo, onde ‘IAC 17’, sob infestação, apresentou
produtividade média de 1.808 e 1.711 kg ha-1
nos ensaios de 2011-2012 e 2012-2013,
respectivamente (Tabela 4).
De acordo com os resultados desses ensaios, também se conclui
que ‘Coodetec 208’ é altamente produtiva; porém, em condições de infestação por
pentatomídeos, a cultivar demonstra alta suscetibilidade, comprometendo sua
produtividade (Tabela 4). Diante disso, sugere-se a escolha dessa cultivar quando for
possível a antecipação de plantio, possibilitando a colheita no começo do mês de fevereiro,
período este em que a população de percevejos geralmente se encontra abaixo do nível de
dano econômico (NDE), conforme descrito por Correia-Ferreira e Panizzi (1999).
Analisando-se os danos ocasionados nos grãos em decorrência da
alimentação dos percevejos, observa-se que no primeiro ensaio (2011-2012), PI 171451 e
D75-10169 apresentaram as maiores médias de grãos classificados na categoria A (grãos
sem danos visíveis), enquanto que os demais genótipos não possuíram classificação dentro
dessa categoria (Figura 11). Neste mesmo ensaio, PI 171451 também se destacou ao
apresentar a maior quantidade de grãos na categoria B (com puncturas, mas sem
deformação), indicando resistência. Ainda no primeiro ensaio, a linhagem D75-10169
apresentou o maior número de grãos classificados na categoria D, revelando alta
suscetibilidade de seus grãos ao ataque do complexo de percevejos (Figura 11).
No segundo ensaio (2012-2013), novamente PI 171451 e D75-
10169 se revelaram com o maior número de grãos classificados na categoria A (Figura 11).
A linhagem PI 229358 também apresentou grande quantidade de grãos nessa categoria.
Esses dados vêm confirmar a resistência dos grãos ao ataque dos percevejos da linhagem
PI 171451 e corroboram as observações de Rossetto et al. (1986), que estudando diferentes

45
genótipos de soja frente ao ataque de percevejos, também descreveram as linhagens PI
171451 e PI 229358 apresentando as maiores médias de grãos nas categorias A e B.
Com base na classificação dos danos em diferentes classes,
observa-se maior quantidade de sementes classificadas nas categorias A e B no segundo
ensaio (Figura 11). Este fato pode ser explicado pela menor ocorrência populacional de
percevejos no segundo ano agrícola (Figura 9), onde a população se manteve abaixo do
nível de dano econômico na maioria das avaliações.

46
0,00 1,50
20,75
27,75
IAC-17
A
B
C
D
0,00 3,25
21,2525,50
IAC-23
A
B
C
D
2,75
16,5
28,5
2,25
PI-171451
A
B
C
D
0,00 1,50
16,25
32,25
PI-229358
A
B
C
D
2,75
4,753,00
39,50
D75-10169
A
B
C
D
0,00 0,75
29,75
19,50
Coodetec 208
A
B
C
D
3,00
11,50
12,75
22,75
IAC-17
A
B
C
D
3,25
21,50
11,50
13,75
IAC-23
A
B
C
D
16,50
18,75
8,75
6,00
PI-171451
A
B
C
D
9,50
19,00
13,25
8,25
PI-229358
A
B
C
D
11,00
10,25
11,50
17,25
D75-10169
A
B
C
D
2,00
9,25
16,00
22,75
Coodetec 208
A
B
C
D Figura 11. Número de grãos danificados por pentatomídeos de acordo com
quatro classes de danos em seis genótipos de soja de ciclo precoce
em ensaios de campo. Botucatu-SP, 2011-2012 e 2012-2013. A =
grãos sem danos visíveis; B = grãos com puncturas, mas sem
deformação; C = grãos com punctura e com deformação; D =
grãos totalmente deformados.
Ensaio 2011-2012
Ensaio 2012-2013

47
Observando-se os dados de danos nos grãos dos genótipos de ciclo
precoce (Tabela 5), a linhagem PI 171451 apresenta o menor escore médio de dano nos
dois ensaios (1,60 e 1,08), confirmando-se como material resistente aos danos de
percevejos.
No segundo ensaio, observa-se também comportamento de
resistência da linhagem PI 229358, apresentando baixo escore de dano em grãos (1,40) e
também se apresentando como pouco danificada (Tabela 5). Porém, esse comportamento é
questionável, uma vez que no primeiro ensaio, sob maior infestação, esse genótipo
apresentou o maior escore médio de danos (2,61).
Tabela 5. Escore médio de dano nos grãos de seis genótipos de soja de ciclo precoce
em ensaios de campo, causados por percevejos pentatomídeos. Botucatu-SP,
2011-2012 e 2012-2013.
Genótipo Escore médio
-1
Ensaio 2011-2012 Ensaio 2012-2013
PI 229358 2,61 a 1,40 c
D75-10169 2,58 ab 1,70 bc
‘IAC 17’ 2,52 ab 2,10 a
‘IAC 23’ 2,44 ab 1,71 b
‘Coodetec 208’ 2,37 b 2,19 a
PI 171451 1,60 c 1,08 d
P <0,001 <0,001
1/Médias seguidas pela mesma letra na coluna não diferem entre si pelo Teste Qui-Quadrado (α = 0,05).
Dados transformados em escores médios.
A reduzida quantidade de danos nos grãos de PI 171451 e PI
229358 frente ao ataque do complexo de percevejos também foi relatado por
Suwanpornskul e Khadkao (1996) na Tailândia, que descreveram essas linhagens com as
menores porcentagens de vagens danificadas, classificando-as como resistentes aos
percevejos N. viridula, Riptortus spp. e Piezodorus hybneri. As linhagens PI 171451 e PI
229358 também foram descritas por Kogan (1989) como fontes de resistência a insetos
mastigadores, sendo que Rossetto et al. (1986) também verificaram que essas linhagens
foram pouco danificadas pelo complexo de percevejos da soja.

48
6.1.3.2 Ciclo Semiprecoce
Em relação à infestação dos genótipos de ciclo semiprecoce
(Tabela 6), observou-se a ausência de diferença entre os tratamentos nas três primeiras
avaliações (22/fev. a 07/mar.) do primeiro ensaio. Os dados da quarta (14/mar.) e quinta
(22/mar.) avaliações revelam que os genótipos PI 227687, ‘IAC 24’, IAC 74-2832 e IAC
78-2318 foram menos atrativos, apresentando as menores médias de percevejos. Na sexta
avaliação (29/mar.) novamente não houve diferença entre os genótipos. Nas duas últimas
avaliações (05/abr. e 12/abr.), a linhagem PI 227687 voltou a se destacar com baixa
colonização, diferindo principalmente da cultivar ‘IAC 24’, que foi a mais atrativa (Tabela
6). De acordo com a média geral do primeiro ensaio, PI 227687 e IAC 74-2832 foram os
menos atrativos (2,3 e 3,2 respectivamente), sugerindo a ocorrência de antixenose.
No segundo ensaio (2012-2013), verificou-se menor população de
percevejos a campo (Figura 9) em comparação com o primeiro. Não houve diferença de
colonização nos genótipos nas duas primeiras avaliações (25/jan. e 01/fev.). Na terceira e
quarta avaliações (08/fev. e 15/fev.) os genótipos com as menores médias de percevejos
foram ‘IAC 24’ e ‘IAC 100’, diferindo de ‘IAC 18’, que novamente se apresentou como o
mais suscetível aos pentatomídeos. Entre a quinta e a sétima avaliações, não houve
diferença entre os genótipos e na oitava avaliação a linhagem PI 227687 apresentou
ausência de infestação, confirmando a ocorrência de resistência por não-preferência
(Tabela 6).
De modo geral, analisando a atratividade nos dois ensaios, nota-se
a maior preferência dos percevejos pelas cultivares ‘IAC 18’ e ‘IAC 24’, que se revelam
altamente atrativas. De maneira oposta, a linhagem PI 227687 se apresentou pouco
atrativa, revelando-se portadora de resistência do tipo não-preferência aos pentatomídeos
(Tabela 6).
Analisando-se a atratividade dos percevejos na linhagem PI
227687, nota-se certa variação durante as avaliações nos dois ensaios (Tabela 6), havendo
uma redução dos valores médios com o passar do tempo. Este fato pode ser explicado
considerando-se a produção de flavonoides após injúrias ocasionadas pelos percevejos,
conferindo resistência induzida a este genótipo. Piubelli et al. (2003), estudando a
produção de flavonóides em genótipos de soja sob o ataque de percevejos, constataram que
PI 227687 foi o que mais produziu daidzina e genisteína após a infestação, aumentando
assim a resistência dessa cultivar.

49
Tabela 6. Número médio (±EP) de percevejos pentatomídeos (E. heros, P. guildinii, N. viridula, E. meditabunda e Dichelops spp.) por pano
de batita obtido em plantas de seis genótipos de soja de ciclo semiprecoce durante dois anos de ensaio a campo. Botucatu-SP,
2011-2012 e 2012-2013.
N° médio de insetos (2011 / 2012)1
Genótipo Avaliação 01 Avaliação 02 Avaliação 03 Avaliação 04 Avaliação 05 Avaliação 06 Avaliação 07 Avaliação 08 Média 22 fev. 28 fev. 07 mar. 14 mar. 22 mar. 29 mar. 05 abr 12 abr.
‘IAC 18’ 2,6 ± 0,5 a 1,6 ± 0,2 7,2 ± 1,7 10,8 ± 2,7 a 6,6 ± 0,8 ab 7,2 ± 1,4 6,2 ± 1,6 b 6,8 ± 3,0 ab 6,2 ± 0,8 a
‘IAC 24’ 1,2 ± 0,2 ab 1,6 ± 0,2 6,2 ± 1,7 6,6 ± 2,5 ab 4,4 ± 1,1 b 7,8 ± 1,1 12,8 ± 1,5 a 10,4 ± 4,7 a 6,4 ± 0,6 a
‘IAC 100’ 1,2 ± 0,4 ab 1,2 ± 0,5 6,2 ± 0,9 9,8 ± 1,3 ab 10,8 ± 1,4 a 6,8 ± 1,7 4,4 ± 1,5 ab 6,0 ± 2,7 ab 5,6 ± 0,3 a
IAC 74-2832 1,4 ± 0,5 ab 0,6 ± 0,2 3,0 ± 1,1 5,2 ± 1,0 ab 2,8 ± 0,7 b 5,0 ± 0,6 5,0 ± 1,1 ab 5,4 ± 2,4 ab 3,2 ± 0,4 b
IAC 78-2318 0,4 ± 0,2 b 1,8 ± 0,7 2,6 ± 0,9 6,0 ± 1,3 ab 3,6 ± 0,5 b 5,2 ± 1,2 7,8 ± 1,2 ab 6,8 ± 3,0 ab 4,2 ± 0,2 ab
PI 227687 0,6 ± 0,2 b 0,8 ± 0,4 5,4 ± 1,6 3,0 ± 0,7 b 5,4 ± 1,5 b 3,4 ± 1,8 0,0 ± 0,0 c 1,8 ± 0,8 c 2,3 ± 0,2 b
P 0,005 0,304 0,139 0,041 <0,001 0,239 <0,001 <0,001 <0,001
N° médio de insetos (2012 / 2013)1
Genótipo Avaliação 01 Avaliação 02 Avaliação 03 Avaliação 04 Avaliação 05 Avaliação 06 Avaliação 07 Avaliação 08 Média 25 jan. 01 fev. 08 fev. 15 fev. 22 fev. 01 mar. 08 mar. 15 mar.
‘IAC 18’ 0,2 ± 0,1 0,5 ± 0,3 1,2 ± 0,2 a 1,4 ± 0,4 a 2,7 ± 0,9 5,5 ± 1,3 6,5 ± 1,4 3,0 ± 1,5 ab 2,6 ± 0,9
‘IAC 24’ 0,5 ± 0,2 0,2 ± 0,2 0,0 ± 0,0 b 0,2 ± 0,2 b 3,5 ± 0,9 3,5 ± 0,5 4,7 ± 1,4 3,0 ± 1,5 ab 1,9 ± 0,7
‘IAC 100’ 0,0 ± 0,0 0,2 ± 0,2 0,0 ± 0,0 b 0,0 ± 0,0 b 2,2 ± 0,6 5,7 ± 0,9 6,5 ± 1,3 6,2 ± 3,1 a 2,6 ± 0,9
IAC 74-2832 0,7 ± 0,4 1,2 ± 0,2 0,2 ± 0,2 ab 0,3 ± 0,2 ab 1,2 ± 0,5 2,0 ± 0,7 2,2 ± 0,9 2,7 ± 1,4 ab 1,3 ± 0,5
IAC 78-2318 0,0 ± 0,0 0,5 ± 0,3 0,5 ± 0,5 ab 0,4 ± 0,3 ab 1,0 ± 0,4 2,7 ± 0,2 3,7 ± 0,9 5,0 ± 2,5 a 1,7 ± 0,6
PI 227687 1,5 ± 0,8 0,7 ± 0,2 0,5 ± 0,3 ab 0,5 ± 0,3 ab 1,5 ± 0,6 2,7 ± 1,8 7,2 ± 1,0 0,0 ± 0,0 b 1,8 ± 0,7
P 0,072 0,121 0,046 0,045 0,114 0,094 0,057 0,009 0,067
1/Médias seguidas pela mesma letra minúscula na coluna e maiúscula na linha não diferem entre si pelo Teste de Tukey (α = 0,05). Dados originais.

50
Os autores quantificaram a concentração de daidzina, que variou de
75 a 352,1 µg/g em plantas sem injúrias e injuriadas respectivamente, e genisteína variando
de 41,7 a 142,4 µg/g. Ramiro et al. (1988), estudando o ataque do complexo de percevejos
da soja, constataram baixas populações no genótipo IAC 78-2318, classificando-o como
resistente. Porém, esses autores fizeram a comparação com genótipos não estudados neste
trabalho, onde a linhagem PI 227687 se revelou mais resistente (Tabela 6). Estas
observações realçam a relatividade nos estudos de resistência de plantas, implicando
sempre na comparação entre genótipos (PAINTER, 1951; LARA, 1991).
Sabe-se que a constituição fitoquímica das plantas está intimamente
ligada à atratividade ou deterrência alimentar das pragas, sendo a quantidade de nitrogênio,
carboidratos solúveis, ácidos orgânicos e fitoesteroides variáveis de acordo com o genótipo
e a fase de desenvolvimento das plantas (TESTER, 1977; KOGAN, 1986). Em PI 227687,
Van Duyn et al. (1971) e Tester (1977) demonstraram que o teor total de nitrogênio e os
açúcares redutores foram consideravelmente menores quando comparados com variedades
comerciais, cuminando na resistência deste material frente a insetos-praga. Essa
constituição química de PI 227687 pode explicar a alta deterrência obtida com este
material nos ensaios a campo.
As menores médias de retenção foliar dentre os genótipos de ciclo
semiprecoce foram obtidas em PI 227687 (5,00 %) e ‘IAC 24’ (17,50 %), diferindo de
‘IAC 18’ (78,75 %), IAC 78-2318 (70,00 %), ‘IAC 100’ (47,50 %) e IAC 74-2832 (37,50
%) que apresentaram altas taxas de retenção (Tabela 7).
Tabela 7. Retenção foliar média (±EP) ocasionada pelo ataque de
percevejos em seis genótipos de soja de ciclo
semiprecoce. Botucatu, 2011-2012 e 2012-2013.
Genótipo Retenção (%)1
PI 227687 5,00 ± 0,00 e
‘IAC 24’ 17,50 ± 4,79 de
IAC 74-2832 37,50 ± 6,61 cd
‘IAC 100’ 47,50 ± 8,78 bc
IAC 78-2318 70,00 ± 7,07 ab
‘IAC 18’ 78,75 ± 8,00 a
P <0,001
1/Médias seguidas pela mesma letra na coluna não diferem entre si pelo Teste de
Tukey (α = 0,05). Dados originais.

51
Essas observações (Tabela 7) sugerem que a linhagem PI 227687
apresenta resistência ao ataque de pentatomídeos, confirmando os relatos de Rossetto et al.
(1986), que também apontaram essa linhagem como portadora de resistência por apresentar
ausência de retenção foliar em suas plantas. Essa característica possivelmente esteja
relacionada a baixa alimentação nesta linhagem, refletindo consequentemente na baixa
retenção foliar apresentada. Esses resultados sugerem a expressão de resistência do tipo
não-preferência para alimentação.
A alta retenção foliar apresentada pela cultivar ‘IAC 18’ neste
trabalho (Tabela 7) já foi documentada anteriormente por outros autores, como Lourenção
et al. (2000), que estimaram retenção foliar de aproximadamente 30 % nesta cultivar na
região de Ribeirão Preto-SP, em áreas onde não se realizou o controle de percevejos.
A cultivar ‘IAC 100’ se comportou de forma intermediária em
relação à retenção foliar (Tabela 7). Em estudos semelhantes, Godoi et al. (2002) avaliaram
a resistência de diferentes populações de soja obtidas a partir de cruzamentos entre alguns
genótipos, dentre eles ‘IAC 100’, constatando que as populações derivadas dessa cultivar
apresentaram menores valores médios de retenção foliar, sendo superiores quanto à essa
característica em relação aos demais tratamentos testados. Baixos índices de retenção foliar
para ‘IAC 100’ também foram reportados por outros autores (LOURENÇÃO et al., 2002;
LOURENÇÃO et al., 2004; LUSTOSA et al., 1999), em comparação a outros genótipos de
soja.
Analisando-se os dados de produtividade dos genótipos de ciclo
semiprecoce, novamente verifica-se que o controle dos percevejos com inseticidas resulta
em maior produtividade, havendo diferença no cultivo com condições controladas
(aplicação de inseticidas) e infestadas por percevejos (sem aplicação) (Tabela 8).
No primeiro ensaio (2011-2012), as parcelas controladas de ‘IAC
18’ e ‘IAC 24’ foram as mais produtivas, atingindo produções médias de 3.555 e 2.900 kg
ha-1
, respectivamente, e diferindo de PI 227687, com produtividade média de 1.264 kg ha-1
(Tabela 8). O mesmo comportamento foi observado no segundo ensaio (2012-2013) para
‘IAC 18’, apresentando-se novamente como a mais produtiva em áreas tratadas (4.206 kg
ha-1
), diferindo de PI 227687, IAC 78-2318, ‘IAC 100’ e ‘IAC 24’.
Quanto à produtividade nas parcelas infestadas, a presença dos
percevejos afetou a produtividade para a maioria dos genótipos nos dois ensaios. Porém,
no primeiro ensaio, ‘IAC 100’ e IAC 782318 tiveram médias de produção semelhantes,

52
independentemente da presença dos insetos, indicando a ocorrência de tolerância (Tabela
8). Apesar de PI 227687 apresentar a menor média de produtividade, esta linhagem
também revela uma baixa redução (18%) devido a presença dos percevejos. A tolerância
apresentada por estes genótipos não se manteve no segundo ensaio (2012-2013), porém,
estes foram os que novamente apresentaram as menores porcentagens de redução pelo
ataque dos percevejos (menos de 30 %). Em estudo semelhante, em que se avaliaram oito
genótipos de soja, as linhagens IAC 78-2318, IAC 73-228 e IAC 78-3258 revelaram
produtividades acima de 2.000 kg ha-1
, diferindo dos demais materiais sob infestação de
percevejos (LOURENÇÃO; MIRANDA; NAGAI, 1987). Os resultados da presente
pesquisa corroboram os dados de Lourenção et al. (1997), que também verificaram boa
produtividade de IAC 78-2318 em estudos conduzidos a campo.
Estudos envolvendo ‘IAC 18’ já mostraram estabilidade e a
elevada capacidade produtiva deste cultivar (RAMIRO et al., 2003; LOURENÇÃO et al.,
2010; LOURENÇÃO et al., 2000; SILVA, 2013b). Lourenção et al. (2010), avaliando
vários genótipos frente ao ataque de lagartas desfolhadoras e percevejos da soja, relataram
que apesar de a cultivar ‘IAC 18’ apresentar elevado índice de retenção foliar, este material
manteve-se produtivo mesmo na presença de percevejos, assim como no presente estudo.
Estes autores sugeriram ‘IAC 18’ como portador de resistência do tipo tolerância. No
entanto, esta hipótese torna-se duvidosa, uma vez que os autores não realizaram
comparações de produtividade entre áreas infestadas e manejadas com inseticidas, como no
presente estudo.
A tolerância apresentada pelo cultivar ‘IAC 100’ no primeiro
ensaio condiz com os frequentes relatos na literatura sobre a resistência deste cultivar.
Rossetto et al. (1995), discutindo a resistência de ‘IAC 100’, relataram vários mecanismos
de compensação sob o ataque de percevejos, sendo eles: menor período de enchimento de
vagens; maior número de sementes; rejeição de vagens novas danificadas e substituição
por novas vagens; senescência normal com queda de folhas na maturação e resistência à
levedura N. coryli transmitida por pentatomídeos. Estas características conferem tolerância
ao complexo de percevejos, de origem poligênica e aditiva (SOUZA e TOLEDO, 1995).
Pelos dados de atratividade e produtividade, obtidos neste trabalho, a tolerância dessa
cultivar está provavelmente ligada à capacidade de substituição de vagens novas
danificadas, visto que ‘IAC 100’ sofreu infestação intermediária nos dois ensaios a campo
(Tabelas 6 e 8).

53
Tabela 8. Produtividade média (kg/ha-1
) (±EP) e percentagem de redução de produção de seis genótipos de soja de ciclo
semiprecoce em parcelas infestadas e não infestadas por percevejos. Botucatu, 2011-2012 e 2012-2013.
1/Médias seguidas pela mesma letra minúscula na coluna e maiúscula na linha não diferem entre si pelo Teste de Tukey (α = 0,05). Dados
originais.
Genótipo
Produtividade (kg/ha-1
)
Ensaio de 2011 - 2012 Ensaio de 2012 - 2013
Controle1 Infestada
1 % Redução Controle
1 Infestada
1 % Redução
‘IAC 18’ 3.555 ± 92 aA 2.208 ± 526 aB 43 4.206 ± 151 aA 1.797 ± 201 abB 57
‘IAC 24’ 2.900 ± 503 aA 1.415 ± 291 aB 51 3.527 ± 58 bA 1.216 ± 78 cB 65
‘IAC 100’ 2.379 ± 167 abA 2.247 ± 227 aA 05 2.962 ± 183 cA 2.086 ± 134 aB 29
IAC 74-2832 2.484 ± 210 abA 1.167 ± 245 aB 53 3.720 ± 88 abA 1.468 ± 167 bcB 60
IAC 78-2318 2.298 ± 281 abA 2.239 ± 258 aA 02 2.597 ± 36 cA 1.972 ± 58 abB 24
PI 227687 1.264 ± 95 bA 1.032 ± 47 aA 18 1.741 ± 51 dA 1.292 ± 40 cB 25
P 0,012 --- <0,001 ---

54
Ainda com relação à cultivar ‘IAC 100’, estudos recentes com 65
genótipos de soja (todas com parentesco com esse material), foram conduzidos na busca
por materiais resistentes a N. viridula. Após avaliação do número de insetos por metro,
peso de grãos e danos causados pelos insetos, os autores identificaram quatro linhagens
promissoras, na qual a genealogia consiste de cruzamentos entre ‘IAC 100’ e ‘Hutcheson’
ou ‘IAC 100’ e V71-370, passíveis de uso no melhoramento contra percevejos
(McPHERSON; BUSS; ROBERTS, 2007).
Quanto aos danos ocasionados nos grãos, no primeiro ensaio
(2011-2012) a linhagem PI 227687 apresentou a maior média de grãos nas categorias A e
B (Figura 12), revelando novamente resistência. Ainda no primeiro ensaio, dentre os
genótipos que apresentaram mais grãos na categoria C, destacam-se ‘IAC 100’, seguido
por ‘IAC 18’, IAC 78-2318 e IAC 74-2832. ‘IAC 24’ foi o mais danificado.
No segundo ensaio (2012-2013), PI 227687 novamente apresenta
maior número de grãos nas categorias A e B, seguido por ‘IAC 100’ (Figura 12). ‘IAC 24’
repetiu as características de suscetibilidade, tendo seus grãos classificados em sua maioria
na classe D.
Lourenção et al. (1987), ao classificar os grãos danificados por
percevejos em diferentes genótipos, verificaram que IAC 78-2318 teve a menor quantidade
de grãos nas classes C e D; já neste trabalho, este material mostrou comportamento
intermediário. Trabalhos realizados (LOURENÇÃO et al., 1997) com ‘IAC 100’
destacaram o baixo índice de dano ocasionado pelos percevejos nas vagens deste material,
fortalecendo a hipótese de resistência desta cultivar contra os percevejos da soja.
Analisando a classe de danos nos grãos de um modo geral, verifica-
se que PI 227687 e ‘IAC-100’ apresentam poucos danos visuais nos grãos, indicando
resistência frente ao ataque dos percevejos.
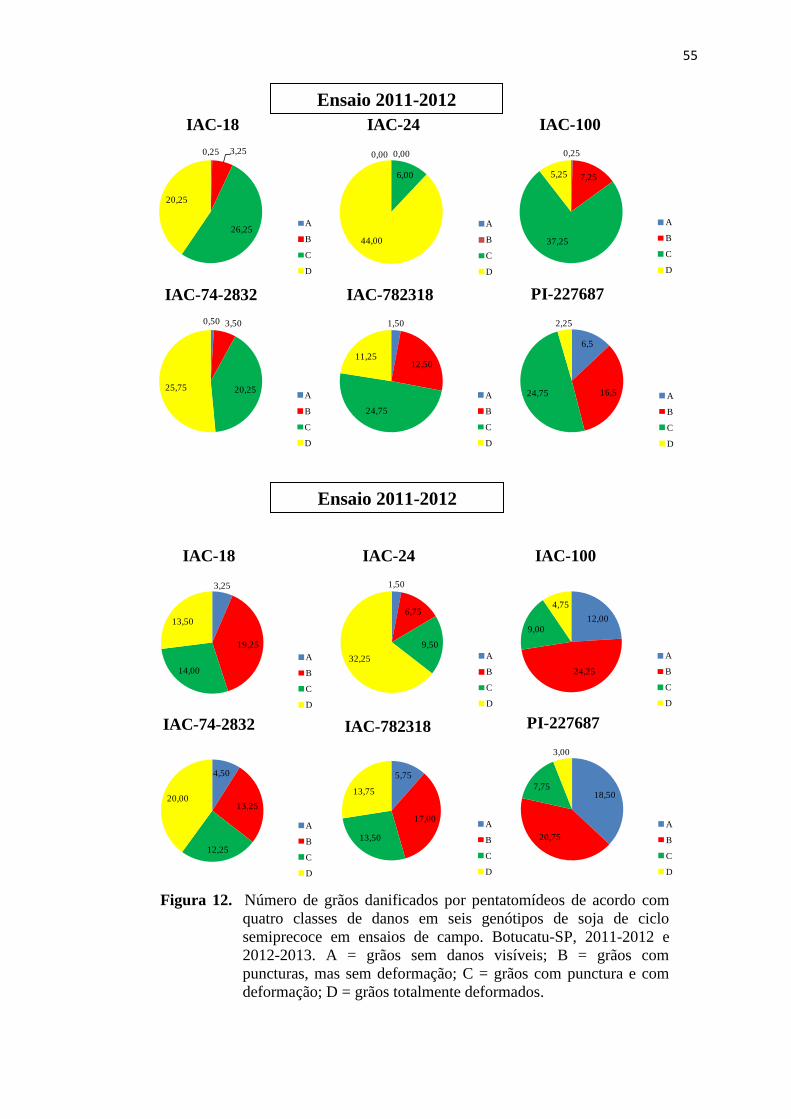
55
0,25 3,25
26,25
20,25
IAC-18
A
B
C
D
0,00 0,00
6,00
44,00
IAC-24
A
B
C
D
0,25
7,25
37,25
5,25
IAC-100
A
B
C
D
0,50 3,50
20,2525,75
IAC-74-2832
A
B
C
D
1,50
12,50
24,75
11,25
IAC-782318
A
B
C
D
6,5
16,524,75
2,25
PI-227687
A
B
C
D
3,25
19,25
14,00
13,50
IAC-18
A
B
C
D
1,50
6,75
9,50
32,25
IAC-24
A
B
C
D
12,00
24,25
9,00
4,75
IAC-100
A
B
C
D
4,50
13,25
12,25
20,00
IAC-74-2832
A
B
C
D
5,75
17,00
13,50
13,75
IAC-782318
A
B
C
D
18,50
20,75
7,75
3,00
PI-227687
A
B
C
D
Figura 12. Número de grãos danificados por pentatomídeos de acordo com
quatro classes de danos em seis genótipos de soja de ciclo
semiprecoce em ensaios de campo. Botucatu-SP, 2011-2012 e
2012-2013. A = grãos sem danos visíveis; B = grãos com
puncturas, mas sem deformação; C = grãos com punctura e com
deformação; D = grãos totalmente deformados.
Ensaio 2011-2012
Ensaio 2011-2012

56
A análise de danos nos grãos dos genótipos de ciclo semiprecoce
revela que a linhagem PI 227687 apresenta o menor escore médio nos dois ensaios (1,455 e
0,905), diferindo dos demais genótipos e confirmando sua resistência contra os percevejos
(Tabela 9).
Tabela 9. Escores médios de dano nos grãos de seis genótipos de soja de ciclo
semiprecoce em ensaios de campo. Botucatu-SP, 2011-2012 e 2012-2013.
Genótipo Escore médio
1
Ensaio 2011-2012 Ensaio 2012-2013
‘IAC 24’ 2,88 a 2,45 a
IAC 74-2832 2,42 b 1,95 b
‘IAC 18’ 2,33 b 1,75 b
‘IAC 100’ 1,95 c 1,13 c
IAC 78-2318 1,91 c 1,70 b
PI 227687 1,45 d 0,90 c
P <0,001 <0,001
1/Escores médios seguidos pela mesma letra na coluna não diferem entre si pelo Teste Qui-Quadrado (α
= 0,05).
A característica de produzir grãos pequenos em grandes
quantidades favorece genótipos pela capacidade de “diluição de danos” por percevejos,
sendo esse comportamento considerado um dos componentes de resistência da soja a esses
sugadores (ROSSETTO; LARA, 1991; PINHEIRO, 1993; GAZZONI, 1998). Somado a
isto, alguns genótipos considerados resistentes ainda apresentam elevado número de
vagens por planta, possibilitando maior produção de grãos livres de danos (MIRANDA et
al., 1979; LINK; ESTEFANEL, 1982; LOURENÇÃO; ROSSETTO; MIRANDA, 1985).
Segundo Suwanpornskul e Khadkao (1996), este comportamento de diluição de danos é
expresso por vários genótipos comprovadamente resistentes, dentre eles PI 227687. No
presente trabalho, a linhagem PI 227687 teve as menores médias de infestação (Tabela 6) e
danos (Tabela 9), apresentando características de resistência por diluição de danos (grande
quantidade de vagens e grãos diminutos). Esses dados confirmam os resultados obtidos por
Rossetto et al. (1986), que também relataram baixo percentual de dano em vagens desta
linhagem. A menor infestação a campo em PI 227687 (Tabela 6) também sugere a
ocorrência de antixenose e/ou antibiose, visto que este genótipo produz grandes

57
quantidades de flavonoides após sofrer injúrias ocasionadas por percevejos (PIUBELLI et
al., 2003).
Com base no baixo escore médio de danos nos grãos, a cultivar
‘IAC 100’ revela-se resistente, com médias de 1,95 no primeiro ensaio e 1,13 no segundo,
diferindo dos demais genótipos (Tabela 9). McPherson et al. (2007) também obtiveram
baixas taxas de danos em sementes de ‘IAC 100’ ocasionadas por percevejos durante
quatro anos de ensaios a campo. Estes autores atribuíram o fato à presença de resistência
do tipo tolerância à alimentação do complexo de percevejos. Pinheiro et al. (2005),
utilizando diversos genótipos em cruzamentos para estudar a herança de características de
resistência a insetos, concluíram que ‘IAC 100’ apresenta a melhor capacidade geral de
combinação, apresentando resistência principalmente quanto ao rendimento e danos nos
grãos. Estes autores sugerem o uso deste cultivar como genitor em programas de
melhoramento contra insetos. Ainda pela Tabela 13, observa-se o maior escore médio de
danos no cultivar ‘IAC 24’, confirmando sua elevada suscetibilidade aos danos dos
percevejos.
Em geral, os dados dos genótipos de ciclo semiprecoce obtidos em
dois anos de ensaios a campo, revelam que os genótipos PI 227687, IAC 78-2318 e ‘IAC
100’ apresentam características de resistência a percevejos, com baixa redução de
produtividade sob ataque e também com menores quantidades de grãos danificados. A
linhagem PI 227687 também apresenta menor atratividade e baixa retenção foliar,
características desejáveis em genótipos de soja (KOGAN, 1989).
6.1.3.3 Ciclo Tardio
Analisando-se a infestação dos percevejos nos genótipos de ciclo
tardio, notam-se elevadas médias de percevejos no primeiro ensaio (2011-2012), superando
na maioria das vezes o nível de dano econômico (Tabela 10). Na primeira avaliação
(22/fev.), a linhagem PI 274454 não apresentou insetos, diferindo de ‘IAC 19’. Na segunda
avaliação não foram verificadas diferenças entre os genótipos. Na terceira (07/mar.) e
quarta (14/mar.) avaliações, PI 274453 e PI 274454 foram as menos atrativas, enquanto
que na quinta avaliação (22/mar.) destacou-se PI 274453. Na sexta avaliação (29/mar.) não
se verificou diferença nas médias dos genótipos, porém estas foram superiores ao nível de
dano econômico (quatro/percevejos por batida de pano), exceto para a linhagem PI
264453, com média de 2,2 percevejos/pano. Na sétima (05/abr.) e oitava (12/abr.)

58
avaliações, verificou-se o mesmo padrão das amostragens anteriores, sendo ‘IAC 19’ e L1-
1-01 altamente atrativas, diferindo das PI’s. Neste ensaio, a cultivar ‘Conquista’ apresentou
infestação intermediária durante as avaliações (Tabela 10).
As médias gerais de atratividade do primeiro ensaio confirmam a
maior preferência dos percevejos em se alimentar no cultivar ‘IAC 19’ (8,0) e L1-1-01
(6,6), e menor preferência para as linhagens PI 274453 (1,4) e PI 274454 (3,6) (Tabela 10).
Para o segundo ensaio (2012-2013), verificaram-se menóres médias
de percevejos em relação ao primeiro ensaio. Não se constatou diferença entre os materiais
até a sexta avaliação (01/mar.) (Tabela 10). Esse fato se explica pela baixa ocorrência
populacional de percevejos nesse ensaio até 1º de março, atingindo níveis de dano
econômico somente a partir desta data (Figura 9). Na sétima avaliação (8/abr.), as
linhagens PI 274453 e PI 274454 se mostram pouco atrativas, apresentando médias de 2,6
e 2,7 percevejos por batida de pano, diferindo do cultivar ‘Conquista’ que apresentou
média de 11 percevejos. Na última avaliação (15/mar.), PI 274453 (1,0) se apresenta
novamente como o menos atrativo, diferindo de L1-1-01 (9,7).
No segundo ensaio, as maiores médias foram registradas nas
últimas avaliações, momento em que as plantas detinham as vagens sob enchimento de
grãos (R5) ou com grãos completamente cheios (R6), sendo que neste período ainda
coincide com os meses de maior ocorrência populacional de percevejos, realçando as
observações de Costa e Link (1974), Corrêa-Ferreira e Panizzi (1999) e Corrêa-Ferreira et
al. (2009), que descrevem a maior concentração de pentatomídeos nos meses de março a
abril na cultura da soja.
Considerando-se as médias de atratividade dos ensaios nos dois
anos agrícolas, verifica-se que PI 274453 e PI 274454 foram os menos atrativos ao
complexo de percevejos, indicando menor preferência dos insetos por estas linhagens. Em
contrapartida, a cultivar ‘IAC 19’ foi a mais atrativa, demonstrando maior suscetibilidade
ao ataque desses pentatomídeos (Tabela 10). Recentemente, Silva (2013b), estudando o
comportamento de diferentes genótipos de soja a P. guildinii, também detectou baixa
atratividade a campo das linhagens PI 274453 e PI 274454. As médias intermediárias de
atratividade do cultivar ‘Conquista’, obtidas nestes ensaios, corroboram os estudos de
Lourenção et al. (2010), que comprovaram certo nível de suscetibilidade ao ataque de
percevejos nesta cultivar.

59
Tabela 10. Número médio (±EP) de percevejos pentatomídeos (E. heros, P. guildinii, N. viridula, E. meditabunda e Dichelops spp.) por pano
de batita obtido em plantas de cinco genótipos se soja de ciclo tardio durante dois anos de ensaio a campo. Botucatu-SP, 2011-
2012 e 2012-2013.
Número médio de insetos (2011 / 2012)1
Genótipo Avaliação 01 Avaliação 02 Avaliação 03 Avaliação 04 Avaliação 05 Avaliação 06 Avaliação 07 Avaliação 08 Média 22 fev. 28 fev. 07 mar. 14 mar. 22 mar. 29 mar. 05 abr 12 abr.
‘IAC 19’ 2,2 ± 0,4 a 2,8 ± 1,1 12,8 ± 2,7 a 13,2 ± 2,8 a 7,6 ± 1,3 a 8,6 ± 1,7 8,0 ± 0,8 a 9,2 ± 4,1 a 8,0 ± 0,7 a
PI 274453 1,4 ± 0,5 ab 1, 2 ± 0,2 1,3 ± 0,3 c 1,0 ± 0,4 c 1,6 ± 0,4 b 2,2 ± 0,6 1,4 ± 0,2 c 1,6 ± 0,7 c 1,4 ± 0,2 c
PI 274454 0,0 ± 0,0 b 0,8 ± 0,4 4,4 ± 0,7 b 3,2 ± 1,2 bc 6,0 ± 1,1 ab 6,4 ± 1,7 3,6 ± 0,4 bc 4,0 ± 1,87 bc 3,6 ± 0,3 bc
L1-1-01 1,6 ± 0,5 ab 1,6 ± 0,5 7,0 ± 0,6 ab 14,4 ± 1,7 a 6,8 ± 0,7 a 8,6 ± 1,7 6,6 ± 0,4 a 7,6 ± 3,4 a 6,6 ± 0,3 a
‘Conquista’ 0,6 ± 0,2 ab 1,6 ± 0,4 6,8 ± 0,9 ab 11,8 ± 3,0 ab 5,6 ± 1,5 ab 6,6 ± 2,2 5,6 ± 1,0 ab 6,6 ± 3,0 ab 5,6 ± 0,8 ab
P 0,004 0,210 <0,001 <0,001 0,008 0,076 <0,001 <0,001 <0,001
Número médio de insetos (2012 / 2013)1
Genótipo Avaliação 01 Avaliação 02 Avaliação 03 Avaliação 04 Avaliação 05 Avaliação 06 Avaliação 07 Avaliação 08 Média 25 jan. 01 fev. 08 fev. 15 fev. 22 fev. 01 mar. 08 mar. 15 mar.
‘IAC 19’ 0,5 ± 0,3 0,2 ± 0,2 0,0 ± 0,0 0,7 ± 0,5 0,4 ± 0,2 2,5 ± 1,0 6,5 ± 1,3 ab 5,5 ± 2,8 ab 2,0 ± 0,7
PI 274453 0,0 ± 0,0 2,0 ± 0,9 0,6 ± 0,6 2,0 ± 0,5 1,5 ± 0,6 4,0 ± 1,3 2,6 ± 0,8 b 1,0 ± 0,5 b 1,7 ± 0,6
PI 274454 1,2 ± 0,5 0,7 ± 0,8 1,2 ± 0,8 1,7 ± 0,8 1,2 ± 0,8 1,2 ± 0,6 2,7 ± 0,9 b 5,5 ± 2,8 ab 2,0 ± 0,7
L1-1-01 0,0 ± 0,0 1,0 ± 0,7 0,2 ± 0,2 0,2 ± 0,2 0,6 ± 0,4 4,5 ± 1,3 8,5 ± 1,6 ab 9,7 ± 4,9 a 3,0 ± 1,1
‘Conquista’ 1,0 ± 0,4 0,2 ± 0,2 0,5 ± 0,5 0,5 ± 0,3 0,7 ± 0,2 5,5 ± 1,4 11,0 ± 2,2 a 2,7 ± 1,4 b 2,7 ± 1,0
P 0,057 0,356 0,464 0,108 0,122 0,138 0,005 0,013 0,197
1/Médias seguidas pela mesma letra na coluna não diferem entre si pelo Teste de Tukey (α = 0,05). Dados originais.

60
Os dados referentes à retenção foliar média dos genótipos de ciclo
tardio revelam maior retenção nas cultivares ‘IAC 19’ e ‘Conquista’, com 88,75 %,
diferindo das PI’s (274453 e 274454), que apresentaram retenções médias de 60 e 50 %
respectivamente (Tabela 11). Rossetto et al. (1986) também verificaram resistência em
seus estudos na linhagem PI 274454, a qual se caracterizou por apresenta baixa retenção
foliar em comparação com outros genótipos. Segundo Panizzi et al. (1986), as linhagens
PI’s foram introduzidas em diversos programas de melhoramento para repassarem essas
características de resistência a insetos-praga. Piubelli et al. (2003) destacam esta
característica de resistência de PI 274454 como sendo uma resistência induzida,
principalmente pela produção de daidzina e genisteína após o ataque dos insetos. Nota-se
também que as linhagens PI 274453 e PI 27445 foram pouco atrativas (Tabela 10),
indicando a ocorrência de não-preferência.
Tabela 11. Retenção foliar média (±EP) ocasionada pelo ataque de
percevejos em cinco genótipos de soja de ciclo tardio.
Botucatu, 2011-2012 e 2012-2013.
Genótipo Retenção (%)1
PI 274454 50,00 ± 5,40 b
PI 274453 60,00 ± 4,56 b
L1-1-01 70,00 ± 7,07 ab
Conquista 88,75 ± 3,15 a
IAC 19 88,75 ± 6,25 a
P <0,001
1/Médias seguidas pela mesma letra na coluna não diferem entre si pelo Teste de
Tukey (α = 0,05). Dados originais
Lourenção et al. (1985) também relataram a expressão de
resistência do tipo não-preferência nas linhagens PI 274453 e PI 274454, quando avaliaram
o comportamento desses genótipos frente ao ataque da desfolhadora Omiodes indicata
(Fabricius), verificando desfolhas na ordem de 4,6 e 4,1 % respectivamente e diferindo dos
demais genótipos testados, principalmente da cultivar ‘Paraná’, que apresentou 61,5 % de
desfolha.

61
Quanto à elevada média de retenção foliar apresentada pela cultivar
‘IAC 19’ (Tabela 11), os dados confirmam as conclusões de Lourenção et al. (1999), que
estudando o comportamento de diversos genótipos de ciclo tardio sob ataque de
percevejos, descreveram este material com o pior desempenho, com média de retenção
foliar de 42,5 %.
Os dados de produtividade dos genótipos de ciclo tardio em dois
ensaios a campo sob infestação de percevejos e em ambiente controlado, encontram-se na
Tabela 12. No primeiro ensaio (2011-2012), o genótipo L1-1-01 apresentou a maior média
de produtividade sob manejo de percevejos, seguido por ‘IAC 19’, PI 274453 e
‘Conquista’, diferindo de PI 274454, que apresentou média de 3.040 kg ha-1
. Quando não
realizado o manejo de percevejos, todos os genótipos apresentaram redução de
produtividade acima de 90 % (Tabela 12).
Já no segundo ensaio (2012-2013), o genótipo mais produtivo em
condições de controle de percevejos foi a cultivar ‘IAC 19’, com média de 3.829 kg ha-1
,
diferindo de PI 274454 (2.917 kg ha-1
), PI 274453 (3.212 kg ha-1
) e ‘Conquista’ (3.204 kg
ha-1
). Assim como no primeiro ensaio, sob infestação, todos os genótipos apresentaram
significante redução de produtividade, sem indicativo de tolerância (Tabela 12).
Observando-se a produtividade sob infestação de percevejos
pentatomídeos nos dois ensaios, notam-se menores médias no primeiro ensaio, com as
maiores porcentagens de redução em todos os genótipos (Tabela 12). Este fato está
provavelmente relacionado à maior ocorrência de percevejos no primeiro ensaio (Figura 9),
onde os níveis populacionais se mantiveram bem acima do nível de dano econômico
durante quase toda a fase reprodutiva dos genótipos.

62
Tabela 12. Produtividade média (kg/ha-1
) (±EP) e percentagem de redução de produção de cinco genótipos de soja de
ciclo tardio em parcelas infestadas e não infestadas com percevejos. Botucatu, 2011-2012 e 2012-2013.
1/Médias seguidas pela mesma letra minúscula na coluna e maiúscula na linha não diferem entre si pelo Teste de Tukey (α = 0,05). Dados
originais.
Genótipo
Produtividade (kg/ha-1
)
Ensaio de 2011 - 2012 Ensaio de 2012 - 2013
Controle1 Infestada
1 % Redução Controle
1 Infestada
1 % Redução
‘IAC 19’ 3.943 ± 301 abA 146 ± 18 abB 96 3.829 ± 100 aA 837 ± 202 bcB 70
PI 274453 3.653 ± 97 abA 367 ± 25 abB 90 3.212 ± 115 bcA 1.331 ± 59 abB 58
PI 274454 3.040 ± 48 bA 232 ± 40 bB 92 2.917 ± 52 cA 1.157 ± 77 abB 60
L1-1-01 4.057 ± 161 aA 308 ± 23 aB 92 3.634 ± 193 abA 1.762 ± 186 aB 51
‘Conquista’ 3.430 ± 348 abA 214 ± 36 abB 93 3.204 ± 116 bcA 484 ± 244 cB 84
P 0,040 --- <0,001 ---

63
Neste trabalho, a produtividade obtida no primeiro ensaio sob
condições controladas, com a linhagem PI 274453 (3.653 kg ha-1
) se aproxima da relatada
por Rossetto et al. (1986) na safra de 1983-1984, que foi de 3.675 kg ha-1
. Porém no
trabalho desses autores, não foi realizado qualquer tipo de manejo visando o controle de
percevejos. Apesar de esses autores não expressarem no trabalho a população de percevjos,
acredita-se na ocorrência de uma população muito abaixo do nível de controle (2
percevejos/batida de pano). De modo geral, ‘IAC 19’ e L1-1-01, se revelam como os
genótipos de ciclo tardio com maior capacidade produtiva, observando-se no cultivar ‘IAC
19’ maior estabilidade na comparação entre os dois anos (Tabela 12).
Considerando-se as classes de danos nos grãos dos genótipos de
ciclo tardio (Figura 13), no primeiro ensaio (2011-2012) verificou-se maior quantidade de
grãos comerciáveis (categorias A e B) nas linhagens PI 274453, PI 274453, PI 274454 e
nas cultivares ‘Conquista’ e ‘IAC-19’. Já os grãos de PI 274454 e L1-1-01 apresentam-se
predominantemente como não comerciáveis (categorias C e D), indicando maior
suscetibilidade aos percevejos em relação aos demais genótipos (Figura 13).

64
4,25
7,25
9,7528,75
IAC-19
A
B
C
D
12,50
17,00
13,25
7,25
PI-274453
A
B
C
D
7,50
16,7518,25
7,50
PI-274454
A
B
C
D
0,000,00 0,25
49,75
L1-1-01
A
B
C
D
4,50
8,00
13,25
24,25
Conquista
A
B
C
D
1,75
10,75
13,50
24,00
IAC-19
A
B
C
D
23,50
21,00
3,252,25
PI-274453
A
B
C
D
7,75
26,00
12,25
4,00
PI-274454
A
B
C
D
1,50
10,50
11,25
26,75
L1-1-01
A
B
C
D
1,25
4,50
11,75
32,50
Conquista
A
B
C
D
Figura 13. Número de grãos danificados por pentatomídeos de acordo com
quatro classes de danos em cinco genótipos de soja de ciclo tardio
em ensaios de campo. Botucatu-SP, 2011-2012 e 2012-2013. A =
grãos sem danos visíveis; B = grãos com puncturas, mas sem
deformação; C = grãos com punctura e com deformação; D = grãos
totalmente deformados.
Ensaio 2011-2012
Ensaio 2012-2013

65
Para o segundo ensaio (2012-2013), novamente verificou-se o bom
desempenho das linhagens PI 274453 e PI 274454, com grãos classificados em maioria nas
classes A e B (Figura 13), sugerindo resistência ao ataque dos percevejos sugadores.
Quanto ao escore médio de danos apresentados pelos diferentes
genótipos de ciclo tardio (Tabela 13), as linhagens PI 274453 e PI 274454 apresentaram as
menores médias de danos. A melhor qualidade dos grãos dessas linhagens em relação às
demais cultivares pode estar relacionada ao menor número de injúrias decorrentes da
alimentação dos pentatomídeos, sugerindo novamente a ocorrência de antixenose (Tabela 10).
‘IAC 19’ e L1-1-01 apresentaram os maiores escores, confirmando sua suscetibilidade
(Tabela 13).
Tabela 13. Escores médios de dano nos grãos causados por percevejos em cinco genótipos
de soja de ciclo tardio, em ensaios de campo. Botucatu-SP, 2011-2012 e 2012-
2013.
Genótipo Escore médio
1
Ensaio 2011-2012 Ensaio 2012-2013
PI 274453 1,30 d 0,68 d
PI 274454 1,83 c 1,25 c
‘IAC 19’ 2,26 b 2,19 b
‘Conquista’ 2,14 b 2,51 a
L1-1-01 2,99 a 2,26 ab
P <0,001 <0,001
1/Escores médios seguidos pela mesma letra na coluna não diferem entre si pelo Teste Qui-Quadrado (α =
0,05).
Genótipos de soja com a capacidade de produzir grãos pequenos são
geralmente mais resistentes, por possuírem um elevado número de grãos livres dos danos
causados pelos percevejos (LINK; ESTEFANEL, 1982; PANIZZI et al., 1986; GODOI et al.,
2002). Neste estudo, isso foi verificado nas linhagens PI 274454 e PI 274453, confirmando a
resistência ao complexo de percevejos.
Cabe destacar que, durante a execução dos ensaios a campo,
observou-se que PI 274453 foi o genótipo tardio que mais demorou a atingir o estágio
reprodutivo, apresentando vagens novas quando os demais genótipos já apresentavam vagens

66
entre R5 e R6. Nestas circunstâncias, a resistência deste material pode ser questionada, uma
vez que pode ter ocorrido uma evasão dos percevejos, devido à ausência de vagens verdes no
período de maior infestação.
6.2. Ensaios em laboratório
6.2.1. Determinação de atratividade de Euschistus heros em testes com
chance de escolha
Os dados de atratividade obtidos em diferentes períodos de avaliação
(15, 30, 45, 60, 120 e 180 minutos) não revelaram diferença entre os genótipos de ciclo
precoce em todas as avaliações (Tabela 14), com médias de adultos atraídos variando de 1,0
(‘IAC 17’) a 1,5 (PI 229358).
Tabela 14. Número médio (±EP) de adultos de Euschistus heros atraídos por vagens R5/R6 de
seis genótipos de soja de ciclo precoce, aos 15, 30, 45, 60, 120 e 180 minutos após a
liberação (T= 26 2ºC, U.R.= 65 10% e fotoperíodo=14h). Botucatu-SP, 2013.
Genótipo Número de adultos de E. heros/2 vagens
1
15 min 30 min 45 min 60 min 120 min 180 min Média
‘IAC 17’ 0,7 ± 0,4 0,8 ± 0,3 1,0 ± 0,5 1,1 ± 0,4 1,4 ± 0,5 1,6 ± 0,6 1,0 ± 0,5
‘IAC 23’ 0,9 ± 0,3 1,4 ± 0,4 1,4 ± 0,5 0,9 ± 0,3 2,0 ± 0,4 1,3 ± 0,4 1,3 ± 0,4
PI 171451 1,1 ± 0,4 1,1 ± 0,4 1,2 ± 0,6 1,1 ± 0,4 1,1 ± 0,4 1,2 ± 0,3 1,1 ± 0,4
PI 229358 1,1 ± 0,5 1,7 ± 0,6 1,7 ± 0,8 1,8 ± 0,5 1,2 ± 0,4 1,4 ± 0,3 1,5 ± 0,5
D75-10169 1,1 ± 0,3 1,1 ± 0,5 1,3 ± 0,8 1,3 ± 0,4 1,2 ± 0,4 1,4 ± 0,6 1,2 ± 0,5
‘Coodetec 208’ 1,0 ± 0,3 1,1 ± 0,5 0,9 ± 0,6 1,3 ± 0,4 1,3 ± 0,4 1,0 ± 0,5 1,1 ± 0,4
P 0,889 0,827 0,730 0,580 0,604 0,946 0,917
1/Dados originais.
A atratividade de E. heros avaliada em laboratório foi menor quando
comparada à obtida a campo ao complexo de percevejos (Tabela 2). No entanto, acredita-se
que a explicação mais plausível para esta diferença de atratividade seja a formação e acúmulo

67
de fitoalexinas em plantas que sofreram prévia herbivoria no campo, ocasionando
repelência/deterrência e ainda podendo intoxicar insetos-praga (SUTHERLAND et al., 1980;
KOGAN e PAXTON, 1983).
Quanto à atratividade com chance de escolha nos genótipos de ciclo
semiprecoce, também não se observou diferenças entre os tratamentos (Tabela 15).
Novamente, as mesmas hipóteses são levantadas, uma vez que as vagens usadas neste ensaio
também se encontravam em fase de transição entre R5 e R6 e também foram colhidas de
plantas que não sofreram nenhuma injúria prévia (casa-de-vegetação).
Tabela 15. Número médio (±EP) de adultos de Euschistus heros atraídos por vagens R5/R6
de seis genótipos de soja de ciclo semiprecoce, aos 15, 30, 45, 60, 120 e 180
minutos após a liberação dos insetos. Botucatu-SP, 2013.
Genótipo Número de adultos de E. heros/2 vagens
1
15 min 30 min 45 min 60 min 120 min 180 min Média
‘IAC 18’ 0,9 ± 0,3 1,1 ± 0,4 1,5 ± 0,6 2,2 ± 0,8 1,8 ± 0,5 1,7 ± 0,4 1,5 ± 0,5
‘IAC 24’ 1,1 ± 0,3 1,2 ± 0,4 1,5 ± 0,6 2,1 ± 0,5 1,8 ± 0,6 1,9 ± 0,5 1,6 ± 0,4
‘IAC 100’ 0,5 ± 0,3 0,7 ± 0,3 0,5 ± 0,2 0,6 ± 0,3 1,1 ± 0,4 1,0 ± 0,6 0,7 ± 0,3
IAC 74-2832 1,2 ± 0,5 0,8 ± 0,2 1,3 ± 0,6 1,1 ± 0,3 1,7 ± 0,3 1,5 ± 0,5 1,3 ± 0,4
IAC 78-2318 1,4 ± 0,5 1,4 ± 0,6 1,3 ± 0,8 1,0 ± 0,4 1,1 ± 0,4 1,1 ± 0,5 1,2 ± 0,5
PI 227687 0,5 ± 0,2 0,5 ± 0,2 0,5 ± 0,3 0,9 ± 0,3 1,0 ± 0,4 1,5 ± 0,6 0,8 ± 0,3
P 0,659 0,842 0,277 0,113 0,657 0,757 0,370
1/ Dados originais.
Nos ensaios de atratividade com vagens de ciclo tardio, observaram-se
diferenças em todos os períodos avaliados, exceto aos 60 minutos (Tabela 16).
Aos 15 minutos após a liberação dos insetos, a linhagem PI 274454 se
mostrou menos atrativa aos percevejos, diferindo de PI 274453, que apresentou a maior
média. Neste período, os genótipos ‘IAC 19’, ‘Conquista’ e L1-1-01 se apresentaram
mediamente atrativos. Nas avaliações de 30 e 45 minutos, novamente PI 274454 foi o menos
atrativo, com médias de 0,6 e 0,4 percevejos, respectivamente. Aos 60 minutos, não houve
diferença entre os tratamentos. Aos 120 e 180 minutos, PI 274454 se revela como o menos
atrativo e PI 274453 como o mais atrativo (Tabela 16).

68
Tabela 16. Número médio (±EP) de adultos de Euschistus heros atraídos por vagens R5/R6
de cinco genótipos de soja de ciclo tardio, aos 15, 30, 45, 60, 120 e 180 minutos
após a liberação dos insetos. Botucatu-SP, 2013.
Genótipo Número de adultos de E. heros/2 vagens
1
15 min 30 min 45 min 60 min 120 min 180 min Média
‘IAC 19’ 1,3 ± 0,4 ab 1,9 ± 0,5 ab 2,2± 0,7 a 1,9 ± 0,6 1,8 ± 0,7 a 1,9 ± 0,6 ab 1,6 ± 0,7 a
PI 274453 2,5 ± 0,7 a 2,7 ± 0,7 a 2,7± 0,6 a 2,5 ± 0,7 2,5 ± 0,6 a 2,5 ± 0,7 a 2,6 ± 0,8 a
PI 274454 0,6 ± 0,2 b 0,6 ± 0,2 b 0,4± 0,2 b 0,4 ± 0,2 0,2 ± 0,1 b 0,6 ± 0,3 b 0,5 ± 0,2 b
L1-1-01 0,7 ± 0,2 ab 0,9 ± 0,3 ab 1,2± 0,3 ab 1,2 ± 0,4 1,1 ± 0,2 ab 0,8 ± 0,2 ab 1,0 ± 0,3 ab
‘Conquista’ 1,2 ± 0,4 ab 1,4 ± 0,3 ab 1,6± 0,3 ab 1,4 ± 0,4 1,9 ± 0,4 a 2,5 ± 0,5 ab 1,8 ± 0,3 a
P 0,035 0,025 0,008 0,105 0,001 0,037 0,002
1/Médias seguidas pela mesma letra minúscula na coluna não diferem entre si pelo Teste de Tukey (α = 0,05).
Dados originais; para análise foram transformados em (x+0,5)1/2
.
Quanto à média geral das avaliações, conclui-se que a linhagem PI
274454 apresentou-se menos atrativa aos adultos de E. heros, sugerindo resistência do tipo
antixenose. Por outro lado, a linhagem PI 274453 revela elevada atratividade, contrariando as
médias obtidas a campo com o complexo de pentatomídeos (Tabela 10). Acredita-se que essa
diferença tenha se manifestado pelo fato das vagens não terem sofrido ataque, limitando a
produção de flavonoides, que caracterizam a resistência induzida neste material. Piubelli et al.
(2003) já haviam realçado esta habilidade em PI 274453, que após ataque, produz elevados
teores de daidzina e genisteína.
Neste sentido, seria interessante a realização de ensaios de atratividade
com vagens colhidas de plantas não atacadas e atacadas previamente, objetivando confirmar
as suspeitas sobre a indução de resistência nos materiais.
6.2.2 Determinação do número de picadas e tempo de consumo de E. heros
em diferentes genótipos de soja
Os resultados do número médio de picadas para alimentação e tempo
médio de alimentação em cada picada não revelaram diferenças entre as médias dos
genótipos de ciclo precoce (Tabela 17).
36

69
Tabela 17. Número médio de picadas e tempo de picada por
picada (±EP) de adultos de Euschistus heros em
vagens R5/R6 de seis genótipos de soja de ciclo
precoce. Botucatu-SP, 2013
Genótipo N° de picadas1 Tempo / picada (min)
1
PI-171451 4,7 ± 1,3 12,0 ± 3,6
‘IAC-17’ 4,9 ± 1,3 4,5 ± 1,9
‘Coodetec 208’ 5,0 ± 0,9 4,2 ± 0,7
‘IAC-23’ 6,5 ± 1,0 4,8 ± 1,5
D-75-10169 7,0 ± 2,3 4,2 ± 1,7
PI-229358 7,0 ± 2,0 27,0 ± 14,2
P 0,855 0,077
1/ Dados originais.
Em função da atratividade apresentada pelos genótipos de ciclo
precoce a campo (Tabela 2), havia a expectativa de um comportamento semelhante neste
teste. Silva (2013b), avaliando estes mesmos genótipos frente ao consumo de P. guildinii,
também não observou-se diferenças, sugerindo observações por um período maior, o que
talvez permita detectar diferenças significativas entre os materiais.
Vale destacar que, mesmo na ausência de diferença entre as médias de
consumo, PI 229358 apresentou média de sete picadas com duração de 27 minutos por picada,
tendendo a ser mais consumida. Para os genótipos menos consumidos, é possível que estes
apresentem compostos impalatáveis, resultando num comportamento supressante e inibidor da
alimentação, características da não-preferência alimentar (LARA, 1991). Souza (2010),
avaliando a alimentação de N. viridula em diferentes genótipos de soja, observou baixo
consumo na cultivar ‘IAC 17’, sugerindo a presença de compostos ligados à palatabilidade.
No presente trabalho, também se verificou baixo consumo de E. heros nessa cultivar (Tabela
17).
Quanto ao consumo nos genótipos de ciclo semiprecoce, novamente
não se detectaram diferenças quanto ao número de picadas e tempo médio por picada (Tabela
18), constatando-se novamente diferenças entre os resultados de campo e laboratório (Tabela
6) conforme discutido para os materiais de ciclo precoce. Apesar das médias de consumo de
E. heros em PI 227687 não diferir daquelas dos demais genótipos (Tabela 18), esta linhagem

70
se mostrou pouco consumida por N. viridula quando comparada dentro de um grupo de dez
materiais (SOUZA et al., 2012). Estes autores relataram um baixo número de picadas neste
material, que indicou a falta de estímulos para o percevejo-verde iniciar sua alimentação.
Neste sentido, pode-se inferir que os genótipos de soja podem afetar distintamente as
diferentes espécies de percevejos.
Tabela 18. Número médio de picadas e tempo de picada por
picada (±EP) de adultos de Euschistus heros em
vagens R5/R6 de seis genótipos de soja de ciclo
semiprecoce. Botucatu-SP, 2013.
Genótipo N° de picadas1
Tempo / picada (min)1
PI 227687 2,9 ± 0,8 10,7 ± 3,2
IAC 78-2318 3,1 ± 1,1 5,2 ± 1,6
‘IAC 100’ 3,5 ± 0,8 5,8 ± 1,3
IAC 74-2832 5,2 ± 1,3 8,2 ± 2,7
‘IAC 24’ 5,3 ± 1,9 19,3 ± 15,0
‘IAC 18’ 6,5 ± 1,7 8,2 ± 2,7
P 0,362 0,064
1/ Dados originais.
Também não foram observadas diferenças nas médias de alimentação
nos genótipos de ciclo tardio (Tabela 19), onde havia a expectativa de que os genótipos menos
atrativos a campo (PI 274453 e PI 274454) (Tabela 10) também fossem os menos consumidos
em laboratório.
Analisando os testes de número e tempo médio de picada de modo
geral (Tabelas 17, 18 e 19), associa-se a ausência de diferença entre as médias com fatores
comentados anteriormente, como a não produção de resistência induzida nas plantas, visto
que só foram utilizadas vagens de plantas não atacadas previamente. Para confirmar esta
hipótese, seriam necessárias avaliações em plantas, possibilitando a produção de metabólitos
secundários após o ataque dos percevejos o que consequentemente refletirá na alteração do
consumo destes insetos. No entanto, ensaios deste tipo se tornariam difíceis de avaliar
visualmente, uma vez que os percevejos ficam menos visíveis nas plantas.

71
Tabela 19. Número médio de picadas e tempo de picada por
picada (±EP) de adultos de Euschistus heros em
vagens R5/R6 de cinco genótipos de soja de ciclo
tardio. Botucatu-SP, 2013.
Genótipo N° de picadas1
Tempo / picada (min)1
PI 274454 3,3 ± 1,4 9,1 ± 2,5
L1-1-01 5,0 ± 0,9 5,4 ± 0,7
‘Conquista’ 5,3 ± 0,8 8,1 ± 1,2
‘IAC 19’ 6,4 ± 1,8 8,4 ± 1,1
PI 274453 8,1 ± 1,9 12,6 ± 4,2
P 0,181 0,306
1/ Dados originais.
Outro fato que também pode justificar a ausência de diferença entre as
médias de consumo, foi o tempo de observação (180 minutos). É possível que um tempo
maior de observações permita a distinção e caracterização dos genótipos quanto à preferência
alimentar do percevejo.
6.2.3 Dureza, quantificação e comprimento de tricomas em vagens de
diferentes genótipos
Com relação às características morfológicas das vagens verdes dos
genótipos de ciclo precoce (Tabela 20), a cultivar ‘IAC-17’ destacou-se com a maior média de
tricomas (109,8/0,25cm2), seguido de ‘Coodetec 208’ (96,2/0,25cm
2). PI-171451 e PI-229358
apresentaram as menores densidades, com 58,6 e 64,3 tricomas/0,25 cm2, respectivamente.
Até o momento, não foram publicados trabalhos relacionando
aspectos de tricomas com a preferência alimentar de percevejos, sendo proposto neste
trabalho a hipótese de uma correlação negativa entre esses fatores. Inicialmente, pela baixa
atratividade aos percevejos obtida a campo nos genótipos PI-171451, PI-229358, D75-10169
e ‘IAC 17’ (Tabela 2), acreditou-se na presença de grande quantidade de tricomas nestes
materiais, o que só foi constatado em ‘IAC 17’ (Tabela 20). Sendo assim, associa-se essa
característica morfológica à menor preferência dos percevejos por infestar ‘IAC 17’, não se
36

72
acreditando, porém, que esta seja a única característica envolvida na interação inseto-
hospedeiro.
Tabela 20. Densidade média de tricomas, comprimento de tricomas e dureza de vagem
(±EP)1 de seis genótipos de soja de ciclo precoce. Botucatu-SP, 2013.
Genótipo Densidade (0,25 cm2)
1 Comprimento (mm)
1 Dureza (g.f/cm
2)
1
PI 171451 58,6 ± 5,20 c 1,58 ± 0,1 272,0 ± 13,4 bc
PI 229358 64,3 ± 4,44 c 1,64 ± 0,1 233,2 ± 8,9 c
‘IAC 23’ 69,2 ± 6,51 bc 1,51 ± 0,1 302,6 ± 11,0 b
D75-10169 84,6 ± 5,55 abc 1,70 ± 0,1 283,6 ± 18,2 bc
‘Coodetec 208’ 96,2 ± 7,83 ab 1,93 ± 0,1 366,0 ± 10,5 a
‘IAC 17’ 109,8 ± 12,80 a 1,81 ± 0,1 276,8 ± 14,5 bc
P <0,001 0,058 <0,001
1/Médias seguidas pela mesma letra minúscula na coluna não diferem entre si pelo Teste de Tukey (P<0,05).
Dados originais.
O comprimento médio dos tricomas não diferiu entre os genótipos de
ciclo precoce, sendo as médias variando de 1,51 a 1,93 mm (Tabela 20). Nesse caso, conclui-
se que essas características são irrelevantes para a seleção hospedeira de adultos de E. heros
quando comparadas entre esses materiais.
A análise da dureza de vagens de soja precoce revelou maior
resistência à penetração no cultivar ‘Coodetec 208’, que diferiu dos demais genótipos (Tabela
20), e opostamente, a linhagem PI 229358 apresentou a menor média de dureza (233,2
g.f/cm2). A elevada dureza de ‘Coodetec 208’ e a maior atratividade de percevejos a campo
nessa cultivar rejeitam a hipótese de que a rigidez da epiderme das vagens influencie
percevejos adultos na seleção alimentar entre os genótipos de ciclo precoce avaliados (Tabela
17).
De acordo com Vendramim e Guzzo (2009), a não preferência
alimentar de insetos por plantas com epiderme mais rígida está diretamente relacionado com a
deposição de sílica ou lignina, que atuam como fatores de resistência a insetos em diversas
plantas cultivadas. No entanto, pelo porte do aparelho bucal bem desenvolvido dos adultos de
E. heros, descarta-se esta possibilidade nos presentes genótipos. Talvez, a maior rigidez
apresentada pela cultivar ‘Coodetec 208’ tenha influência sobre ninfas, inibindo ou limitando
a alimentação nas vagens; no entanto, isso não foi objeto de investigação do presente trabalho.

73
Figura 14. Correlação entre densidade de tricomas e atratividade de adultos de Euschistus
heros, e dureza de vagem, em teste com chance de escolha em seis genótipos de
soja de ciclo precoce.
Quanto à densidade de tricomas nos genótipos de soja de ciclo
semiprecoce, a cultivar ‘IAC 100’ foi a que apresentou a maior média, seguida por IAC 78-
2318 (Tabela 21). Os genótipos ‘IAC 18’ e IAC 74-2818 se destacaram como os de menor
densidade de tricomas, com menos de 98 estruturas em 25 mm2
de vagem.
Considerando-se o baixo número de tricomas na cultivar ‘IAC 18’,
juntamente com a elevada preferência dos percevejos em testes de atratividade a campo
(Tabela 6), existe uma relação negativa entre a densidade de tricomas e a preferência
alimentar. Porém, a alta densidade de tricomas em ‘IAC 100’ não inibiu a atratividade a
campo dos percevejos (Tabela 6). Neste sentido, coloca-se em dúvida novamente a influência
da densidade de tricomas de vagens sobre a seleção hospedeira de adultos de E. heros.
Quanto ao tamanho dos tricomas, também não se observou diferença
entre os genótipos de ciclo semiprecoce (Tabela 21). Nestes genótipos, o tamanho variou de
0,86 a 1,01 mm, valores inferiores quando comparados aos de ciclo precoce (Tabela 20). As
análises de dureza das vagens dos genótipos de ciclo semiprecoce revelam que o genótipo PI
227687 possui a maior resistência à penetração, diferindo dos demais (Tabela 21). É possível
que a elevada resistência à penetração em PI 227687 (434,0 g.f/cm2) pelos percevejos
represente uma barreira física à penetração dos estiletes, como parte dos mecanismos de
resistência deste genótipo.

74
Tabela 21. Densidade média de tricomas, comprimento de tricoma e dureza de vagem
(±EP)1 de seis genótipos de soja de ciclo semiprecoce. Botucatu-SP, 2013.
Genótipo Densidade (0,25 cm2)
1 Comprimento (mm)
1 Dureza (g.f/cm
2)
1
IAC 74-2832 91,0 ± 9,67 d 0,94 ± 0,06 323,6 ± 10,42 b
‘IAC 18’ 97,2 ± 10,40 d 0,95 ± 0,09 303,8 ± 14,94 b
‘IAC 24’ 113,7 ± 3,92 cd 0,91 ± 0,06 308,6 ± 14,24 b
PI 227687 130,6 ± 5,38 c 0,86 ± 0,04 434,0 ± 20,81 a
IAC 78-2318 173,5 ± 5,51 b 1,01 ± 0,05 338,3 ± 16,18 b
‘IAC 100’ 216,3 ± 10,03 a 0,96 ± 0,05 305,8 ± 21,19 b
P <0,001 0,699 <0,001
1/Médias seguidas pela mesma letra minúscula na coluna não diferem entre si pelo Teste de Tukey (P<0,05).
Dados originais.
y = -0,0053x + 1,908R² = 0,498
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
50 100 150 200 250
Den
sid
ad
e d
e E
. h
ero
s
Densidade de tricomas (25mm²)
y = -0,0037x + 2,4225R² = 0,2544
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
280 330 380 430 480
Densi
dad
e d
e E
. hero
s
Textura de vagem (g.f/cm²)
Figura 15. Correlação entre densidade de tricomas e atratividade de adultos de Euschistus
heros, e dureza de vagem, em teste com chance de escolha em seis genótipos de
soja de ciclo semiprecoce.
As avaliações morfológicas das vagens dos genótipos tardios mostrou
diferença entre o número de tricomas nos materiais (Tabela 22), sendo L1-1-01 e PI 274453
(191,5 e 162,7 unidades, respectivamente) os genótipos detentores das maiores médias,
diferindo de PI 227454, ‘Conquista’ e ‘IAC 19’, com 84,3, 96,6 e 103,4 estruturas presentes
por 25 mm2.
Apesar dessa diferença, é pouco provável que o elevado número de
tricomas em L1-1-01 e PI 274453 afete negativamente a escolha pelos percevejos, visto que o
genótipo L1-1-01 foi muito infestado a campo (Tabela 10).

75
Tabela 22. Densidade média de tricomas, comprimento de tricoma e dureza de vagem
(±EP)1 de cinco genótipos de soja de ciclo tardio. Botucatu-SP, 2013.
Genótipo Densidade (0,25 cm2)
1 Comprimento (mm)
1 Dureza (g.f/cm
2)
1
PI 274454 84,3 ± 5,29 b 1,00 ± 0,06 381,4 ± 11,67 a
‘Conquista’ 96,6 ± 3,70 b 1,21 ± 0,10 276,0 ± 24,05 b
‘IAC 19’ 103,4 ± 7,88 b 1,39 ± 0,16 374,0 ± 22,03 a
PI 274453 162,7± 6,20 a 1,13 ± 0,11 316,4 ± 18,29 ab
L1-1-01 191,5 ± 14,41 a 1,00 ± 0,08 385,5 ± 14,24 a
P <0,001 0,054 <0,001
1 Médias seguidas pela mesma letra minúscula na coluna não diferem entre si pelo Teste de Tukey (P<0,05).
Dados originais.
Quanto ao comprimento dos tricomas de vagens de soja de ciclo
tardio, também se verificam ausências de diferenças, com médias entre 1,00 e 1,39 mm
(Tabela 22). Os dados de dureza de vagens desses materiais apresentaram diferença
significativa, revelando menor média na cultivar ‘Conquista’ (Tabela 22). Esta baixa dureza
pode estar envolvida na suscetibilidade desta cultivar ao ataque de percevejos.
y = 0,0044x + 0,9411R² = 0,0654
0
0,5
1
1,5
2
2,5
3
50 100 150 200
Densi
dad
e d
e E
. hero
s
Densidade de tricomas (25mm²)
y = -0,0113x + 5,4177R² = 0,4676
0
0,5
1
1,5
2
2,5
3
230 280 330 380 430
Densi
dad
e d
e E
. hero
s
Textura de vagem (g.f/cm²)
Figura 16. Correlação entre densidade de tricomas e atratividade de adultos de Euschistus
heros, e dureza de vagem, em teste com chance de escolha em seis genótipos de
soja de ciclo tardio.
Em geral, acredita-se que as características morfológicas e físicas das
vagens de soja possam influenciar a seleção hospedeira de percevejos adultos, porém não

76
podem ser consideradas isoladamente como características que conferem resistência. É
possível que estas características atuem diferentemente sobre os genótipos de soja.
Na literatura, são poucos os trabalhos disponíveis sobre resistência
vinculados a características de vagens a sugadores, sendo mais comuns trabalhos com insetos
de menor porte, que geralmente apresentam maiores mudanças de comportamento frente a
estas características. Neste sentido, Valle e Lourenção (2002) demonstraram a correlação
positiva entre densidade de tricomas e atratividade e/ou oviposição de B. tabaci biótipo B, que
observaram elevadas médias de ovos/cm2 em genótipos com altas densidades de tricomas em
folhas, influenciando a permanência e oviposição do inseto. Em algodão e tomate também
foram observadas essas mesmas associações frente ao mesmo inseto (BERLINGER, 1986;
BUTTER; VIR, 1989; HEINZ; ZALOM, 1995).
As influências morfológicas e químicas das plantas também se
associam a outros parâmetros biológicos das pragas, afetando a atividade alimentar por
possuírem no tegumento da semente altos teores de lignina, o que garante sua proteção contra
herbívoros (BOESEWINKEL; BOUMAN, 1995). Segundo Panizzi e Silva (2009), o espaço
de ar entre a parede das vagens e as sementes também influencia de forma negativa a
alimentação de percevejos, entretanto este aspecto não foi investigado no presente trabalho.
Enfim, também deve-se considerar que as características morfológicas
avaliadas no presente trabalho poderiam se manifestar de forma mais contundente sobre
ninfas, que pelo menor porte, provavelmente se apresentariam mais suscetíveis.
6.2.4 Avaliação de antibiose
Para avaliar os parâmetros biológicos de E. heros confinados nos
diferentes genótipos, foram utilizadas ninfas a partir do segundo estádio, uma vez que as de
primeiro não se alimentam e são mais sensíveis à manipulação (OLIVERIA; PANIZZI,
2003). Apesar de não utilizar as ninfas de primeiro estádio nos bioensaios, verificou-se média
de duração de 3,1 dias nesta fase, sendo um valor muito próximo dos 3,3 dias observados por
Mourão e Panizzi (2000). Já o período médio de incubação observado foi de 6,4 dias, também
semelhante ao relatado por Villas Bôas e Panizzi (1980), de 6,8 e Mourão e Panizzi (2000), de
6,1 dias. Para o cálculo da duração de período ninfal, foi somada a média de 3,1 dias,
referente ao primeiro ínstar.

77
Os dados referentes à duração de cada ínstar e período ninfal de E.
heros, encontram-se na Tabela 23. No segundo ínstar, destacaram-se os genótipos ‘IAC 100’
e L1-1-01 por prolongarem o período de desenvolvimento nessa fase, com médias de 6,55 e
6,24 dias, respectivamente. De forma contrária, ‘IAC 18’ (3,19 dias) e as linhagens PI 274453
(3,34 dias) e PI 274454 (3,38 dias) apresentaram as menores médias de duração nesse estádio.
Avaliando o tempo de duração ninfal sob diferentes fotoperíodos,
Mourão e Panizzi (2000) concluíram que ninfas de segundo ínstar alimentadas com o cultivar
‘Paraná’ permaneceram neste ínstar por 6,5 dias quando submetidas a um fotoperíodo diário
de 10 horas, e 4,7 dias sob 14 horas, indicando que a velocidade do desenvolvimento ninfal
pode estar relacionada ao fotoperíodo. No presente trabalho, foi padronizado um fotoperíodo
de 14 horas diárias, sendo a duração de cada ínstar um reflexo apenas da alimentação nos
diferentes genótipos.
Quanto à duração do terceiro ínstar, observou-se que ‘IAC 19’ (9,88
dias), ‘Coodetec 208’ (9,0 dias), ‘Conquista’ (8,93 dias) e L1-1-01 (8,52 dias) foram os
genótipos que mais prolongaram a fase. Contrariamente, PI 274454 (3,55 dias), ‘IAC 17’
(4,21 dias), ‘IAC 24’ (4,79 dias), PI 274453 (4,48 dias), ‘IAC 18’ (4,37 dias) e mostraram-se
mais adequados às ninfas, permitindo um desenvolvimento mais rápido (Tabela 23).
Discutindo a duração do terceiro ínstar ninfal de E. heros, Pinto e Panizzi (1994) relataram
uma média de 4,7 dias em condições de fotoperíodo de 14 horas de luz. Este dado se encontra
muito próximo das médias apresentadas pelos genótipos mais favoráveis às ninfas.
Para o quarto ínstar ninfal, a linhagem PI 274453 apresentou a maior
média de duração (8,25 dias), seguida por ‘IAC 100’ (7,95 dias), PI 227687 (7,82 dias) e ‘IAC
23’ (7,79 dias) e diferindo da maioria dos materiais, principalmente de PI 274454, com média
de 4,57 dias (Tabela 23). No último ínstar, não foram verificadas diferenças entre os
tratamentos e as médias variaram de 6 a 11,50 dias. Nesta fase, não constam os dados da
linhagem PI 274454 que ocasionou 100% de mortalidade ainda no estádio N4 (Tabela 23).
Esta observação indica a ocorrência de elevado nível de antibiose nesta linhagem, conforme já
relatado para outros percevejos (PIUBELLI et al. 2003; ROSSETTO; LOURENÇÃO, 1981).
Em geral, a duração da última fase ninfal foi maior em relação às fases
anteriores, independentemente do genótipo utilizado como hospedeiro. Esta maior duração é
comum em pentatomídeos, visto que neste ínstar ocorrem muitas mudanças fisiológicas antes
de os insetos atingirem a fase adulta, além da maior quantidade de nutrientes necessários para

78
a realização da ecdise e formação de um adulto com bom potencial reprodutivo (VILLAS
BÔAS; PANIZZI, 1980; BIEHLER; McPHERSON, 1982; PANIZZI; PANIZZI, 1991).
Analisando-se o período ninfal total, observou-se grande variação
entre os genótipos, com médias variando de 24,10 a 34,18 dias (Tabela 23). Comparando as
médias de período ninfal característico de E. heros obtidos com aquela relatada por Costa et
al. (1998), alguns genótipos favoreceram o desenvolvimento ninfal, enquanto que outros
retardaram. Ninfas confinadas a L1-1-01, ‘IAC 100’, ‘IAC 23’ e ‘Coodetec 208’ prolongaram
o desenvolvimento do inseto, com médias de 34,48; 34,20; 31,27 e 30,80 dias
respectivamente, sugerindo a ocorrência de antibiose e/ou não preferência para alimentação.
PI 227687 (24,10 dias), ‘IAC 18’ (25,88 dias), ‘IAC 24’ (23,37 dias), IAC 78-2318 (26,60
dias), ‘IAC 17’ (23,77 dias), PI 274453 (26,98 dias) D75-10169 (27,33 dias) e IAC 74-2832
(27,54 dias) destacaram-se como os mais favoráveis, revelando suscetibilidade ao
desenvolvimento do percevejo. Não foi possível calcular a duração do período ninfal dos
insetos confinados em PI 274454, uma vez que no quarto ínstar, este genótipo já havia
provocado a morte de todas as ninfas (Tabela 23).
Vários fatores das plantas podem colaborar para o prolongamento do
período pós-embrionário de insetos em soja, com destaque para as características
morfológicas, compostos antinutricionais e baixo teor de proteínas, forçando o inseto a
permanecer por um período maior no estágio imaturo (CALHOUN; FUNDERBURK;
TEARE, 1988; LARA, 1991; PANIZZI, 1991).
Apesar de análises de constituição química não terem sido realizadas
nesse estudo, essa investigação seria interessante, objetivando a identificação de fatores de
resistência que atuam nesses genótipos frente ao ataque de E. heros.
Em função do prolongamento ninfal, a cultivar ‘IAC-100’ revela-se
portadora de resistência do tipo antibiose. Além da presença de posíveis fatores químicos e/ou
antinutricionais que caracterizam este tipo de resistência, é possível também a presença de
fatores morfológicos, uma vez que este material apresentou a maior densidade de tricomas
(Tabela 21). Esta elevada pilosidade, apesar de aparentemente não influenciar na alimentação
de adultos, provavelmente atua como barreira física aos primeiros estádios iniciais (segundo e
terceiro ínstares), os quais, devido ao menor porte, podem ter maior dificuldade na
alimentação.

79
Tabela 23. Período médio (dias ± EP) de duração de ínstares ninfais e fase ninfal total (dias ± EP) de Euschistus heros em 17 genótipos de
soja (T= 26 2ºC, U.R.= 65 10% e fotoperíodo=14h). Botucatu-SP, 2013.
Genótipo Duração (dias)1,2
N2 N3 N4 N5 Período Ninfal*
L1-1- 01 (n=29) 6,24 ± 0,67 a (n=23) 8,52 ± 0,65 ab (n=18) 6,22 ± 0,57 cde (n=10) 10,60 ± 1,33 (n=10) 34,38 ± 1,78 a
‘IAC 100’ (n=29) 6,55 ± 0,11 a (n=29) 7,50 ± 0,28 bc (n=22) 7,95 ± 0,15 ab (n=7) 9,10 ± 0,87 (n=7) 34,20 ± 1,20 a
‘IAC 23’ (n=30) 3,30 ± 0,11 de (n=29) 6,03 ± 0,27 cd (n=28) 7,79 ± 0,16 ab (n=6) 11,00 ± 1,13 (n=6) 31,27 ± 1,30 a
‘Coodetec 208’ (n=23) 5,17 ± 0,41 abc (n=13) 9,00 ± 0,53 ab (n=11) 5,27 ± 0,47 ef (n=9) 9,67 ± 0,44 (n=9) 30,80 ± 1,12 a
PI 229358 (n=49) 3,53 ± 0,11 de (n=27) 6,07 ± 0,41 cd (n=23) 7,74 ± 0,21 abc (n=2) 11,50 ± 0,50 (n=1) 30,10 ± 1,00 ab
PI 171451 (n=24) 5,71 ± 0,66 ab (n=17) 7,24 ± 0,55 bc (n=16) 7,31 ± 0,44 abcd (n=7) 9,86 ± 1,12 (n=7) 29,96 ± 1,39 ab
‘Conquista’ (n=27) 4,26 ± 0,18 bcde (n=14) 8,93 ± 0,66 ab (n=14) 6,14 ± 0,42 de (n=8) 9,00 ± 1,68 (n=8) 29,48 ± 1,69 ab
‘IAC 19’ (n=23) 4,48 ± 0,24 bcde (n=8) 9,88 ± 0,59 a (n=6) 6,67 ± 1,17 bcde (n=1) 8,00 ± 0,00 (n=1) 29,10 ± 0,00 ab
IAC 74 -2832 (n=23) 4,43 ± 0,22 bcde (n=23) 6,09 ± 0,30 cd (n=22) 6,64 ± 0,22 bcde (n=16) 7,06 ± 0,54 (n=16) 27,54 ± 0,88 b
D75-10169 (n=28) 4,21 ± 0,35 bcde (n=26) 5,88 ± 0,39 cd (n=20) 7,40 ± 0,22 abcd (n=8) 7,63 ± 0,50 (n=8) 27,33 ± 1.01 b
PI 274453 (n=29) 3,34 ± 0,18 e (n=21) 4,48 ± 0,19 de (n=16) 8,25 ± 0,30 a (n=8) 8,63 ± 0,78 (n=8) 26,98 ± 0,64 b
‘IAC 17’ (n=29) 3,30 ± 0,18 de (n=29) 4,21 ± 0,28 de (n=28) 7,86 ± 0,17 ab (n=18) 8,17 ± 0,20 (n=18) 23,77 ± 0,33 b
IAC 78 -2318 (n=30) 3,93 ± 0,19 cde (n=28) 5,71 ± 0,22 cde (n=21) 7,67 ± 0,27 abcd (n=6) 6,50 ± 0,72 (n=6) 26,60 ± 0,67 b
‘IAC 24’ (n=29) 3,38 ± 0,09 de (n=29) 4,79 ± 0,12 de (n=27) 7,59 ± 0,10 abcd (n=11) 8,00 ± 0,71 (n=11) 23,37 ± 0,71 b
‘IAC 18’ (n=27) 3,19 ± 0,23 e (n=27) 4,37 ± 0,37 de (n=27) 7,78 ± 0,19 abc (n=18) 7,83 ± 1,13 (n=18) 25,88 ± 0,57 b
PI 227687 (n=48) 4,77 ± 0,26 abcd (n=23) 6,30 ± 0,31 cd (n=11) 7,82 ± 0,18 ab (n=1) 6,00 ± 0,00 (n=1) 24,10 ± 0,00 b
PI 274454 (n=32) 3,38 ± 0,10 de (n=20) 3,55 ± 0,19 e (n=7) 4,57 ± 0,43 f --- ---
P <0,001 <0,001 <0,001 0,007 < 000,1
1/Médias seguidas pela mesma letra minúscula na coluna não diferem entre si pelo Teste de LSD (P<0,05). Dados originais;
2/n= número de insetos avaliados;
*Somado à média do primeiro ístar (N1=3,1).

80
A linhagem PI 227687 portadora de resistência múltipla a insetos da
soja (VAN DUYN; TURNIPSEED; MAXWELL, 1971; CLARK et al., 1972; SMITH, 1985;
BEACH; TODD, 1988), não prolongou a fase imatura do inseto neste estudo (24,10 dias).
Recentemente, Silva (2013b) também observou a mesma característica desta linhagem,
relatando o não prolongamento da fase ninfal em estudos de antibiose com P. guildinii. Estes
resultados envolvendo ninfas de percevejos também se assemelham aos obtidos por Lima e
Lara (2004), constatando desenvolvimento favorável de B. tabaci biótipo B nesta linhagem.
No entanto, a hipótese de suscetibilidade de PI 227687 ao desenvolvimento de E. heros não se
sustenta em função da elevada mortalidade ninfal, onde apenas 3 % dos indivíduos atingiram
a fase adulta (Figura 18). Piubelli et al. (2003), também relataram alta mortalidade ninfal em
percevejos que se alimentaram nesta linhagem, porém, no trabalho desses autores, a espécie
em questão era N. viridula. Autores como Carrão-Panizzi e Kitamura (1995) e Piubelli (2000)
também relataram a alta concentração de isoflavonas nesta linhagem associada com a elevada
mortalidade de N. viridula em comparação com outros genótipos.
3,3 0,0 3,3
33,360,0
IAC - 17
0,0
6,7
10,0
63,3
20,0
IAC - 23
20,0
23,3
3,3
30,0
23,3
PI - 171451
3,3
10,0
10,0
70,0
6,7
PI - 229358
6,76,7
20,0
40,0
26,7
D75 - 10169
23,3
33,36,7
6,7
30,0
Coodetec 208
10,0
43,3
0,0
20,0
26,7
Conquista
Ínstar 2 Ínstar 3 Ínstar 4 Ínstar 5 Emergência de adultos
Figura 17. Mortalidade (%) nos diferentes estádios ninfais e
emergência de adultos de Euschistus heros confinados
em seis genótipos de soja de ciclo precoce.

81
Outros genótipos que também merecem destaque são ‘IAC 19’ e PI
229358, que apesar de não diferirem dos demais genótipos quanto ao prolongamento ninfal
(Tabela 23), ocasionaram 96,7 e 93,3 % de mortalidade na fase jovem dos percevejos,
sugerindo antibiose (Figuras 18 e 20). Os índices de mortalidade ninfal provocados pelos
genótipos ‘IAC 17’ (Figura 17), PI 274453 (Figura 19), ‘IAC 18’ e ‘IAC 74-2832 (Tabela 19)
foram mais baixos em relação aos demais, permitindo emergências de adultos superiores a
50%.
Durante as avaliações de biologia, observou-se que os genótipos ‘IAC
23’ e PI 229358 além de prolongarem o quinto ínstar ninfal (Tabela 23), também ocasionaram
a maior taxa mortalidade nesta fase (Figura 17). Verificou-se que as ninfas de quinto ínstar
morreram em sua grande maioria no momento da ecdise, ficando presas à exúvia (Figura 20).
10,00,0
0,0
30,060,0
IAC - 18
0,0 0,0
6,9
55,2
37,9
IAC - 24
0,0 0,0
24,1
51,7
24,1
IAC - 100
23,3
0,03,3
20,0
53,3
IAC - 74 - 2832
0,0
6,7
23,3
50,0
20,0
IAC - 782318
7
17
40
33
3
PI - 227687
10,0
43,3
0,0
20,0
26,7
Conquista
Ínstar 2 Ínstar 3 Ínstar 4 Ínstar 5 Emergência de adultos
Figura 18. Mortalidade (%) nos diferentes estádios ninfais e
emergência de adultos de Euschistus heros confinados
em seis genótipos de soja de ciclo semiprecoce.

82
23,3
50,0
10,0
13,3
3,3
IAC - 19
10,0
6,7
10,0
20,0
53,3
PI - 274453
33,3
0,0
43,3
23,3
0,0
PI - 274454
3,3
20,0
16,7
26,7
33,3
L1 - 1 - 01
10,0
43,3
0,0
20,0
26,7
Conquista
10,0
43,3
0,0
20,0
26,7
Conquista
Ínstar 2 Ínstar 3 Ínstar 4 Ínstar 5 Emergência de adultos
Figura 19. Mortalidade (%) nos diferentes estádios ninfais e
emergência de adultos de Euschistus heros confinados
em cinco genótipos de soja de ciclo tardio.
Figura 20. Ninfas de quinto ínstar de Euschistus heros que morreram presas à
exúvia no momento da ecdise.

83
Os dados de viabilidade ninfal (Figura 21) revelam que, com exceção
de PI 274454, houve emergência de adultos em todos os materiais. Observa-se que os
genótipos ‘IAC 19’, PI-227687 e PI 229358 proporcionaram porcentagens menores que 10 %,
indicando a ocorrência de elevado nível de antibiose e/ou não-preferência para alimentação.
Possivelmente, as causas desta resistência estão associadas à existência de compostos
antinutricionais e/ou presença de compostos secundários, que afetam o desenvolvimento
ninfal.
ef ef
def
cde cde
cd cd
c
c c c
c
bc
ab
a a
0
10
20
30
40
50
60
70
Via
bili
dad
e N
infa
l (%
)
f
Figura 21. Viabilidade ninfal (% ± EP) de Euschistus heros obtida em 17
genótipos de soja (T= 26 2ºC, U.R.= 65 10% e
fotoperíodo=14h).
Quanto ao peso médio de adultos recém-emergidos, observou-se
grande variação entre os insetos desenvolvidos nos diferentes materiais (40,80 mg a 73,42 g)
(Tabela 24). PI 227687 foi o material que mais desfavoreceu o ganho de peso dos insetos,
seguido por PI 274453, com médias de 40,80 mg e 49,67 mg, respectivamente. Por outro lado,
PI 229358, ‘Coodetec 208’, ‘IAC 100’, ‘IAC 23’, PI 171 451, IAC 74-2832, IAC 78-2318 e
‘Conquista’, proporcionaram maiores ganhos de peso, sendo as médias, muito próximas das
apresentadas por Meneguim et al. (1989) (67 mg) e Pinto e Panizzi (1994) (68 mg) em
estudos de biologia de E. heros.

84
Tabela 24. Longevidade média (dias ± EP) e peso de adultos recém-emergidos (mg ± EP) de
Euschistus heros obtidos em 17 genótipos de soja (T= 26 2ºC, U.R.= 65 10% e
fotoperíodo=14h). Botucatu-SP, 2013.
Genótipo Ciclo Peso (mg)1,2
Longevidade (dias)¹
PI 229358 Precoce (n=6) 73,52 ± 2,32 a (n=6) 8,17 ± 2,89
‘Coodetec 208’ Precoce (n=9) 73,28 ± 1,74 a (n=9) 21,00 ± 2,97
‘IAC 100’ Semiprecoce (n=10) 72,87 ± 2,20 a (n=7) 17,71 ± 3,25
‘IAC 23’ Precoce (n=6) 72,20 ± 1,67 a (n=6) 10,00 ± 2,85
PI 171451 Precoce (n=7) 71,99 ± 3,13 a (n=7) 11,43 ± 2,16
IAC 74-2832 Semiprecoce (n=10) 71,61 ± 2,25 a (n=16) 20,69 ± 3,03
IAC 78-2318 Semiprecoce (n=6) 70,58 ± 3,18 a (n=6) 12,50 ± 3,12
‘Conquista’ Tardio (n=8) 69,96 ± 2,04 a (n=8) 12,38 ± 1,08
D75-10169 Precoce (n=8) 67,29 ± 3,02 ab (n=8) 17,38 ± 5,37
‘IAC 18’ Semiprecoce (n=10) 66,45 ± 3,55 ab (n=18) 16,56 ± 1,59
L1-1-01 Tardio (n=10) 66,21 ± 1,50 ab (n=10) 10,30 ± 1,86
‘IAC 24’ Semiprecoce (n=10) 64,15 ± 2,28 ab (n=11) 20,18 ± 3,30
‘IAC 19’ Tardio (n=1) 59,60 ± 0,00 ab (n=1) 12,00 ± 0,00
‘IAC 17’ Precoce (n=10) 59,06 ± 3,19 abc (n=18) 12,6 ± 2,22
PI 274453 Tardio (n=8) 49,67 ± 3,78 bc (n=8) 6,71 ± 1,51
PI 227687 Semiprecoce (n=1) 40,80 ± 0,00 c (n=1) 9,00 ± 0,00
PI 274454 Tardio * *
P --- <0,001 0,058
1/Médias seguidas pela mesma letra minúscula na coluna não diferem entre si pelo Teste de Fisher (P<0,05).
Dados originais; 2/n= número de insetos avaliados.
Piubelli et al. (2003), comparando o desempenho do percevejo N.
viridula em quatro genótipos de soja, também observaram baixo peso em adultos alimentados
em PI 227687, demonstrando que os percevejos acumulam menos lipídeos quando
alimentados nesta linhagem, sendo classificada pelos autores como inadequada ao
desenvolvimento de percevejos por apresentar efeitos deletérios. Panizzi et al. (1989)
demonstraram que, na falta de lipídeos nos alimentos, percevejos utilizam reservas
nutricionais acumuladas durante a fase ninfal, apresentando menor peso quando adultos.
Neste sentido, o baixo peso de insetos que se desenvolveram
alimentando-se em PI 227687 neste trabalho, reforça a expressão de resistência do tipo

85
antibiose neste material, tornando-o interessante para os programas de melhoramento de soja.
Nota-se que apesar dos diferentes genótipos influenciarem no peso dos adultos, não ocorreram
diferenças entre as médias de longevidade (Tabela 24), que variaram de 6,71 a 21,00 dias.
Quanto à viabilidade de ovos de insetos desenvolvidos nos diferentes
genótipos, observaram-se variações entre 65 e 95 % (Tabela 25), sendo as linhagens PI
274453 e PI 227687 as que afetaram negativamente essa viabilidade (65 e 71 %,
respectivamente).
Tabela 25. Viabilidade de ovos (± EP) de Euschistus heros
confinados em 17 genótipos de soja (T= 26 2ºC,
U.R.= 65 10% e fotoperíodo=14h). Botucatu-SP,
2013.
1/Médias seguidas pela mesma letra minúscula na coluna não diferem entre si pelo
Teste de LSD (P<0,05). Dados originais.
Genótipo
Ciclo Viabilidade (%)1
‘IAC 24’ Semiprecoce 95,0 ± 3,0 a
‘IAC 19’ Tardio 93,0 ± 3,0 a
IAC 74-2832 Semiprecoce 93,0 ± 4,0 a
‘IAC 23’ Precoce 92,0 ± 0,4 a
D75-10169 Precoce 91,0 ± 0,4 ab
IAC 78-2318 Semiprecoce 91,0 ± 4,0 ab
PI 274454 Tardio 91,0 ± 4,0 ab
PI 171451 Precoce 90,0 ± 4,0 ab
‘IAC 17’ Precoce 89,0 ± 4,0 ab
PI 229358 Precoce 89,0 ± 4,0 ab
L1-1-01 Tardio 88,0 ± 4,0 ab
‘Coodetec 208’ Precoce 85,0 ± 5,0 abc
‘IAC 18’ Semiprecoce 83,0 ± 5,0 abc
‘IAC 100’ Semiprecoce 81,0 ± 4,0 abc
‘Conquista’ Tardio 75,0 ± 7,0 bc
PI 227687 Semiprecoce 71,0 ± 6,0 c
PI 274453 Tardio 65,0 ± 1,2 c
P --- 0,003

86
Neste trabalho, estas linhagens já haviam se mostrado inadequadas à
alimentação de E. heros, ocasionando elevada mortalidade ninfal e baixo peso de insetos
(Tabelas 23 e 24), revelando-se resistentes.

87
7 CONCLUSÕES
Dentre os genótipos de ciclo precoce:
- Os genótipos PI 171451 e PI 229358 são menos atrativos ao complexo de percevejos em
condições de campo;
- PI 171451 apresenta baixa retenção foliar decorrente do ataque de pentatomídeos;
- PI 171451 e ‘IAC 17’ apresentam as maiores produtividades sob condições de infestações
em campo;
- Em campo, os grãos de PI 171451 e PI 229358 são menos danificados pelos percevejos;
- A alta densidade de tricomas em vagens de ‘IAC 17’ parece influenciar o comportamento
de E. heros;
Dentre os genótipos de ciclo semiprecoce:
- PI 227687 e IAC 74-2832 são menos infestados pelo complexo de percevejos em
condições de campo;
- PI 227687 apresenta baixa retenção foliar decorrente do ataque de pentatomídeos;
- PI 227687, ‘IAC 100’ e IAC 78-2318 mantêm os mesmos índices de produção em
parcelas infestadas e não infestadas em campo;
- PI 227687 e ‘IAC 100’ são os menos danificados pelos percevejos;
- PI 227687 possui a maior dureza de vagens;

88
Dentre os genótipos de ciclo tardio:
- As linhagens PI 274453 e PI 274454 são menos infestadas pelo complexo de percevejos
em campo, apresentando também baixa retenção foliar;
- Vagens de PI 274454 são menos atrativas em laboratório;
- Todos os genótipos apresentaram maiores produtividades sob condições controladas;
- As linhagens PI 274453 e PI 274454 são menos danificadas por percevejos;
Em relação aos ensaios de antibiose:
- L1-1-01, ‘IAC 100’, ‘IAC 23’ e ‘Coodetec 208’ prolongam o período de
desenvolvimento ninfal de E. heros;
- PI 274454, ‘IAC 19’, PI 227687 e PI 229358 causam elevada mortalidade ninfal;
- PI 274453 e PI 227687 ocasionam redução no peso de adultos e na viabilidade de ovos de E.
heros.

89
8 REFERÊNCIAS BIBLIOGRÁFICAS
AGRIANUAL 2013. Anuário Estatístico da Agricultura Brasileira. São Paulo: FNP –
Consultoria & Agroinformativos, 2013. p. 453-477.
AGROFIT, 2013 - Sistema de Agrotóxicos Fitossanitários. In: Ministério da Agricultura,
Pecuária e Abastecimento (MAPA). Disponível em:
<http://agrofit.agricultura.gov.br/agrofit_cons/principal_agrofit_cons>. Acesso em: 13 ago.
2013.
AHMAD, M.; ARIF, M. I.; AHMAD, Z.; DENHOLM, I. Cotton whitefly (Bemisia tabaci)
resistance to organophosphate and pyrethroid insecticides in Pakistan. Pest Management
Science, v. 58, n. 2, p. 203-208, 2002.
ALEXANDRE, T. M. Práticas de manejo de pragas utilizadas na soja e seu impacto
sobre a cultura. 2010. 104 f. Tese (Doutorado em Ciências) - Universidade Federal do
Paraná, Curitiba, 2010.
AUCLAIR, J. L. Host plant resistance. In: MINKS, A. K., HARREWIJN, P. (ed) Aphids –
their biology, natural enemies and control. Elsevier Science Publishers, Amsterdam, p.
225-265, 1989.
BASSO, C. J. Manual de identificação das principais pragas, doenças e algumas
deficiências nutricionais na cultura do algodoeiro, da soja e do milho. Frederico
Westphalen: Grafimax, 2009. 142 p.
BAUR, M. E.; SOSA-GOMEZ, D. R.; OTTEA, J.; LEONARD, B. R.; CORSO, I. C.; DA
SILVA, J. J.; TEMPLE, J.; BOETHEL, D. J. Susceptibility to insecticides used for control
of Piezodorus guildinii (Heteroptera: Pentatomidae) in the United States and Brazil.
Journal of Economic Entomology, v. 103, n. 3, p. 869-876, 2010.
BEACH, R. M.; TODD, J. W. Foliage consumption and developmental parameters of the
soybean looper and the velvetbean caterpillar (Lepidoptera: Noctuidae) reared on
susceptible and resistant soybean genotypes. Journal of Economic Entomology, v. 81, n.
1, p. 310-316, 1988.

90
BELORTE, L. C.; RAMIRO, Z. A.; FARIA, A. M.; MARINO, C. A. B. Danos causados
por percevejos (Hemiptera: Pentatomidae) em cinco cultivares de soja (Glycine max (L.)
Merrill, 1917) no município de Araçatuba, SP. Arquivos Instituto Biológico, v. 70, n. 2,
p. 169-175, 2003.
BERLINGER, M. J. Host plant resistance to Bemisia tabaci. Agriculture, Ecosystems &
Environment, v. 17, p. 69-82, 1986.
BIEHLER, J. A., McPHERSON, J. E. Life history and laboratory rearing of Galgupha
ovalis (Hemiptera: Corimelaenidae), with descriptions of immature stages. Annals of the
Entomological Society of America, v. 75, p. 465-470, 1982.
BOESEWINKEL, F. D.; BOUMAN, F. The seed: structure and function. In: KIGEL, J.;
GALILI, G, Seed development and germination. New York: Marcel Dekker, 1995, p. 1-
24.
BORÉM, A. Escape Gênico. Biotecnologia & Desenvolvimento (encarte especial). 1999.
p. 101-107.
BORÉM, A. Melhoramento de epécies cultivadas. Viçosa: UFV, 1999.
BORGES, M.; SCHMIDT, F. G. V.; SUJII, E. R.; MEDEIROS, M. A.; MORI, K.;
ZARBIN, P. H. G.; FERREIRA, J. T. B. Field responses of stink bugs to the natural and
synthetic pheromone of the Neotropical brown stink bug, Euschistus heros (Heteroptera:
Pentatomidae). Physiological Entomology, v. 23, p. 202-207,1998.
BORROR, J. D.; WHITE, E. R. Introduction to the study of insects. Boston: Houghton
Mifflin Company, 1970, 404 p.
BUENO, A. F.; SOSA-GÓMEZ, D. R.; CORRÊA-FERREIRA, B. S; MOSCARDI, F.;
BUENO, R. C. O. F. Inimigos naturais das pragas da soja. In: HOFFMANN-CAMPO, C.
B., CORRÊA-FERREIRA, B. S., MOSCARDI, F. Soja: manejo integrado de insetos e
outros artrópodes-praga. Embrapa, 2012, p. 491-629.
BUTTER, N. S.; VIR, B. K. Morphological basis of resistance in cotton to the whitefly
Bemisia tabaci. Phytoparasitica, v. 17, n. 4, p. 251-261, 1989.
BYRNE, F. J.; CASTLE, S.; PRABHAKER, N.; TOSCANO, N. C. Biochemical study of
resistence to imidacloprid in B biotype Bemisia tabaci from Guetemala. Pest
Management Science, v. 59, n. 3, p. 347-352, 2003.
CALHOUN, D. S.; FUNDERBURK, J. E.; TEARE, I. D. Soybean seed crude protein and
oil levels in relation to weight, developmental time, and survival of southern green stink
bug (Hemiptera: Pentatomidae). Environmental Entomology, v. 17, n. 4, p. 727-729,
1988.
CÂMARA, G. M. S. Soja: tecnologia da produção. Piracicaba: Publique, 1998. 577 p.

91
CARRÃO-PANIZZI, M. C.; KITAMURA, K. Isoflavone content in Brazilian soybean
cultivars. Breeding Science, v. 45, n. 3, p. 295-300, 1995.
CARVALHO, W. A.; ESPINDOLA, C. R.; PACCOLA, A. A. Levantamento de solos da
Fazenda Lageado – Estação Experimental “Presidente Médice”. Boletim Científico da
Faculdade de Ciências Agronômicas, Botucatu, 1983. 95 p.
CARVER, M.; GROSS, G. F.; WOODWARD, T. E. Hemiptera. In: CSIRO.
Commonwelth Scientific and Industrial Research Organization. Division of Entomology.
The insects of Australia: a textbook for students and research workers. Melbourne, 1991.
v. 1, p. 429-509.
CIVIDANES, F. J.; PARRA, J. R. P. Zoneamento ecológico de Nezara viridula (L.),
Piezodorus guildinii (West.) e Euschistus heros (Fabr.) (Heteroptera: Pentatomidae) em
quatro estados produtores de soja do Brasil. Anais da Sociedade Entomológica
Brasileira, v. 23, n. 2, p. 219-226, 1994.
CIVIDANES, F. J.; BARBOSA, J. C. Effects of no-tillage and of soybean-corn
intercropping on natural enemies and pests. Pesquisa Agropecuária Brasileira, v. 36, p.
235-241, 2001.
CLARK, W. J.; HARRIS, F. A.; MAXWELL, F. G.; HARTWIG, E. E. Resistance of
certain soybean cultivars to bean leaf beetle, striped blister beetle and bollworm. Journal
of Economic Entomology, v. 65, n. 6, p. 1669-1672, 1972.
CONAB – Companhia Nacional de Abastecimento. Acompanhamento da safa brasileira:
grãos, décimo levantamento, julho 2013. Brasília, 28p. Disponível em:
<http://www.conab.gov.br/OlalaCMS/uploads/arquivos/13_07_09_09_04_53_boletim_gra
os_junho__2013.pdf> Acesso em: 25 jul. 2013.
CORRÊA-FERREIRA, B. S. Amostragem de pragas da soja. In: HOFFMANN-CAMPO,
C. B., CORRÊA-FERREIRA, B. S., MOSCARDI, F. Soja: manejo integrado de insetos e
outros artrópodes-praga. Embrapa, 2012. 631-672 p.
CORRÊA-FERREIRA, B. S.; PANIZZI, A. R. Percevejos-praga da soja no norte do
Paraná: abundância em relação à fenologia da planta e hospedeiros intermediários. Na.
Semin. Nac. Pesq., 1982. 140-151 p.
CORRÊA-FERREIRA, B. S.; ALEXANDRE, T. M.; PELLIZZARO, E. C.; MOSCARDI,
F.; BUENO, A. F. Práticas de manejo de pragas utilizadas na soja e seu impacto sobre a
cultura. Londrina: Embrapa Soja, 2010. 16 p. (Embrapa Soja. Circular Técnica, 78).
CORRÊA-FERREIRA, B. S.; KRZYZANOWSKI, F. C.; MINAMI, C. A. Percevejos
e a qualidade da semente de soja – série sementes. Londrina: Embrapa Soja,
2009. 15 p. (Embrapa Soja. Circular Técnica, 67).
CORRÊA-FERREIRA, B.S.; MOSCARDI, F. Seasonal occurrence and host spectrum of
egg parasitoids associated with soybean stink bugs. Biological control, v. 5, p. 196-202,
1995.

92
CORRÊA-FERREIRA, B. S.; MACHADO, E. M.; HOFFMANN-CAMPO, C. B.
Sobrevivência e desempenho reprodutivo do percevejo marrom Euschistus heros (F.) na
entressafra de soja. In: Reunião de Pesquisa da Soja da Região Central do Brasil, 31.
2010b, Brasília, DF. Resumos Embrapa Soja, p. 81-83.
CORRÊA-FERREIRA, B. S.; MOSCARDI, F. Biological control of soybean stink busg by
inoculative releases of Trissolcus basalis. Entomologia Experimentalis et Applicata, v.
79, p. 1-7, 1996.
CORRÊA-FERREIRA, B. S.; PANIZZI, A. R. Percevejos da soja e seu manejo.
Londrina: Embrapa-CNPSo, 1999. 45 p. (Embrapa- CNPSo. Documentos, 24).
CORRÊA-FERREIRA, B. S.; PAVÃO, A. L. Monitoramento de percevejos da soja: maior
eficiência no uso do pano-de-batida. In: REUNIÃO DE PESQUISA DE SOJA DA
REGIÃO CENTRAL DO BRASIL, 27, 2005 Cornélio Procópio. Resumos... Londrina,
PR: Embrapa CNPSo, 2005. p. 152-153.
CORSO, I. C. Atratividade do sal de cozinha para espécimes de Nezara viridula (L., 1758).
In: EMBRAPA. Centro nacional de Pesquisa da Soja. Resultados de pesquisa de soja
1988/89. Londrina, 1989. p. 78-79. (EMBRAPA-CNPSo. Documentos, 43)
CORSO, I. C. Constatação do agente causal da mancha-de-levedura em percevejos que
atacam a soja no Paraná. In: SEMINÁRIO NACIONAL DE PESQUISA DE SOJA, 3.,
1984, Campinas. Anais... Londrina, PR: Embrapa CNPSo, 1984. p. 152-157.
CORSO, I. C. Uso de sal de cozinha na redução da dose de inseticida para o controle
de percevejos da soja. Londrina: EMBRAPA-CNPSo, 1990. 7 p. (EMBRAPA-CNPSo.
Comunicado Técnico, 45).
CORSO, I. C.; GAZZONI, D. L. Sodium chloride: an insecticide enhancer for controlling
Pentatomids on soybeans. Pesquisa Agropecuária Brasileira, v. 33, n. 10, p. 1563-1571,
1998.
COSTA LIMA, A. M. Insetos do Brasil: hemípteros. Rio de Janeiro: Escola Nacional de
Agronomia, 1940. t. 2. 351 p. (Série Didática,3).
COSTA, E. C.; LINK, D. Incidência de percevejos em soja. Revista do Centro de
Ciências Rurais, v. 4, n. 4, p. 397-400, 1974.
COSTA, M. L. M.; BORGES, M.; VILELA, E. F. Biologia reprodutiva de Euschistus
heros (F.) (Heteroptera: Pentatomidae). Anais da Sociedade Entomológica do Brasil,
Brasília, v. 27, n. 4, p. 559-568, 1998.
DEGRANDE, P. E.; VIVAN, L. M. Pragas da soja. In. YUYAMA, M. M.; SUZUKI, S.;
CAMACHO, S. A. Boletim de Pesquisa de Soja. Rondonópolis: Fundação MT, 2010. v.
14, 418 p.
DEGRANDE, P. E.; VIVAN, L. M., Pragas da soja. In. HIROMOTO, D. M.; CAJU, J.;
CAMACHO, S. A., Boletim de Pesquisa de Soja. Rondonópolis: Fundação MT, 2009. v.
13, 229-265 p.

93
DUYN, J. W. Van; TURNIPSEED, S. G.; MAXWELL, J. D. Resistance in soybeans to the
Mexican bean beetle. I. Sources of resistance. Crop Science, v. 11, n. 4, p. 572-573, 1971.
ELBERT, A.; NAUEN, R. Resistance of Bemisia tabaci (Homoptera: Aleyrodidae) to
insecticides in southern Spain with special reference to neonicotinoids. Pest Management
Science, v. 56, n. 1, p. 60-64, 2000.
EMBRAPA, 2010. Tecnologias de produção de soja região central do Brasil 2011.
Londrina: Embrapa Soja: Embrapa Cerrados: Embrapa Agropecuária Oeste, 2010. 255 p.
EMBRAPA, 2006. Sistema Brasileiro de Classificação de Solos. Brasília: Embrapa:
Centro Nacional de Pesquisa de Solos, 2006. 306 p.
EMBRAPA, 2011. Manejo de Insetos-Pragas. In: Tecnologias de produção de soja -
Região Central do Brasil. Londrina: Embrapa Soja (Sistemas de Produção), 2011. n. 15,
p. 261.
FEHR, W. R.; CAVINESS, C. E. Stages of soybean development. Ames: Iowa State
University, 1977, 12 p.
FERNANDES, F. M.; ATHAYDE, M. L. F.; LARA, F. M. Comportamento de cultivares
de soja no campo em relação ao ataque de percevejos. Pesquisa Agropecuária Brasileira,
Brasília, v. 29, n. 3, p. 363-367, 1994.
FERNANDES, O. A.; CORRÊA, A. C. B.; BORTOLI, S. A. Manejo integrado de
pragas e nematoides. Jaboticabal, FUNEP, 1990. 253 p.
FUGI, C. G. Q.; LOURENÇÃO, A. L.; PARRA, J. R. P. Biology of Anticarsia gemmatalis
on soybean genotypes with different degrees of resistance to insects. Scientia Agricola, v.
62, n. 1, p. 31-35, 2005.
GALILEO, M. H. M.; HEINRICHS, E. A. Retenção foliar em plantas de soja (Glycine
max (L.) Merrill) resultantes da ação de Piezodorus guildinii (Westwood, 1837)
(Hemiptera, Pentatomidae), em diferentes níveis e épocas de infestação. Anais da
Sociedade Entomológica do Brasil, 1978. v.7, p. 85-98.
GALLO, D.; NAKANO, O.; SILVEIRA NETO, S.; CARVALHO, R. P. L.; BATISTA, G.
C. de; BERTI FILHO, E.; PARRA, J. R. P.; ZUCCHI, R. A.; ALVES, S. B.;
VENDRAMIM, J. D.; MARCHINI, L. C.; LOPES, J. R. S.; OMOTO, C. Entomologia
agrícola. Piracicaba: FEALQ, 2002. 920 p.
GAZZONI, D. L. Efeito de populações de percevejos na produtividade, qualidade da
semente e características agronômicas da soja. Pesquisa Agropecuária Brasileira, v. 33,
n. 8, p. 1229-1237, 1998.
GAZZONI, D. L.; YORINORI, J. T. Manual de identificação de pragas e doenças da
soja. Brasília: Embrapa-SPI, 1995. 128 p.
GILLOT, C. Entomology. 3rd
Ed. Berlin: Springer, 2005, 835p.

94
GODOI, C. R. C.; PEREIRA, F. S.; UMENO, F.; ÁZARA, N. A.; LIMA, L. P. M. S.;
SILVA, R. P.; OLIVEIRA, A. B.; ARAÚJO, I. M.; ZUCCHI, M. I.; PINHEIRO, J. B.
Resistência a insetos em populações de soja com diferentes proporções gênicas de
genitores resistentes. Pesquisa Agropecuária Tropical, v. 32, n. 1, p. 47-55, 2002.
GODOY, K. B.; ÁVILA, C. J.; ARCE, C. C. M. Controle biológico de percevejos
fitófagos da soja na região de Dourados, MS. Dourados: Embrapa Agropecuária Oeste,
2007. 27 p. (Embrapa Agropecuária Oeste. Boletim de Pesquisa e Desenvolvimento, 40).
GODOY, K. B.; GALLLI, J. C.; ÁVILA, C. J.; CORRÊA-FERREIRA, B. S. Parasitismo
em ovos de percevejos da soja Euschistus heros (Fabricius) e Piezodorus guildinii
(Westwood) (Hemiptera: Pentatomidae), em São Gabriel do Oeste, MS. Ciência Rural, v.
35, p. 455-458, 2005.
HART, K. A.; PIMENTEL, D. Environmental and economic costs of pesticide use. In:
PIMENTEL, D. (Ed.). Encyclopaedia of pest management. New York: Marcel Dekker,
2002. p. 237-239.
HEINZ, K. M.; ZALOM, F. G. Variation in trichome-based resistance to Bemisia
argentifolii (Homoptera: Aleyrodidae) oviposition on tomato. Journal of Economic
Entomology, v. 88, n. 5, p. 1494-1502, 1995.
HOFFMANN-CAMPO, C. B.; MOSCARDI, F.; CORRÊA-FERREIRA, B. S.; SOSA-
GÓMEZ, D. R.; PANIZZI, A. R.; CORSO, I. C.; GAZZONI, D. L.; OLIVEIRA, E. B.
Pragas da soja no Brasil e seu manejo integrado. Londrina: Embrapa-CNPSo, 2000. 70
p. (Embrapa- CNPSo. Documentos, 30).
HORI, K. Z. Possible causes os disease symptoms resulting from the feeding of
phytophagous Heteroptera. In: SCHAEFER, C.W.; PANIZZI, A.R. (Ed.). Heteroptera of
economic importance. Boca Raton: CRC Press, 2000, p. 11-35.
HOFFMANN-CAMPO, C. B.; CORRÊA-FERREIRA, B. S.; MOSCARDI, F. Soja:
manejo integrado de insetos e outros artrópodes-praga. Embrapa, Brasília, 2012, v. 1, 859
p.
IBGE - INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA. Produção
Agrícola 2013. Disponível em: <http://www.ibge.gov.br/home/estatistica/indicadores/
agropecuaria/lspa/lspa_201306comentarios.pdf> Acesso em: 26 de jul. 2013.
JENSEN, R. L.; NEWSOM, L. D. Effect of stink-bug-damaged soybean seeds on
germination, emergence and yield. Journal of Economic Entomology, v. 65, n. 1, p. 261-
264, 1972.
KASTLER, M.; QUEIROZ, E. F.; TERASAWA, F. Introdução e evolução da soja no
Brasil. In: MYASAKA, S; MEDINA, J. C. (eds.). A soja no Brasil. Campinas: Editora
ITAL, 1981, p. 22-24.
KILEN, T. C.; LAMBERT, L. Registration of three glabrous and three dense pubescent
soybean germoplasm lines susceptible (D88-5320, D88-5295), moderately resistant (D88-

95
5328, D88-5272), ou resistant (D90-9216, D90-9220) to foliar-feeding insects. Crop
Science, v. 33, p. 215, 1993.
KISHINO, K. & ALVES, R. T. Biologia e manejo de insetos pragas da soja nos
Cerrados. Semin. EMBRAPA/ CAAG, 1992, p. 66-74.
KEA, W.C., S.G. TURNIPSEED & G.R. CARNER. Influence of resistant soybeans on the
susceptility of lepidopterous pests to insecticides. J. Econ. Entomol. p. 58-60, 1978.
KOGAN, M. Plant resistance in soybean insect control. In: A.J. PASCALE, World
Research Conference IV. Buenos Aires: Orientácion Gráfica Editora, S.R.L., 1989. p.
1519-1525.
KOGAN, M. Natural chemical in plant resistence to insects. Iowa State J. Res., 1986, p.
501-527.
KOGAN, M.; PAXTON, J. Natural inducers of plant resistence to insects. In: P. A.
HEDIN, Plant resistance to insects, Washington, ACS, D. C., 1983, p. 152-171.
KOGAN, M. Resistance in soybean to insect pests. In: Expanding the use of soybean.
Proc. Conference for Asia and Oceania. University of Illinois College of Agriculture, 1976.
p. 165-169.
LARA, F. M. Princípios de resistência de plantas a insetos. São Paulo: Ícone, 1991. v. 2,
336 p.
LIMA, A. C. S.; LARA, F. M. Resistance of soybean genotypes to the silverleaf whitefly
Bemisia tabaci (Genn.) biotype B (Hemiptera: Aleyrodidae). Neotropical Entomology, v.
33, n. 1, p. 1-75, 2004.
LINK, D.; ESTEFANEL, V. Influência do número de sementes por legume e de legumes
por planta sobre o nível de dano de pentatomídeos em soja. Revista do Centro de
Ciências Rurais, v. 12, n. 2, p.109-113, 1982.
LINK, D.; GRAZIA, J. Pentatomídeos da região central do Rio Grande do Sul
(Heteroptera). Anais da Sociedade Entomológica do Brasil, p. 116-129, 1987.
LOCKWOOD, J. A.; STORY, R. N. Adaptive functions of nymphal aggregation in the
southern green stink bug, Nezara viridula (L.) (Hemiptera: Pentatomidae). Environmental
Entomology, v. 15, n. 3, p. 739-749, 1986.
LOURENÇÃO, A. L.; BRAGA, N. R.; MIRANDA, M. A. C.; RECO, P. C.; VALLE, G.
E.; PEREIRA, J. C. V. N. A. Insect damage and powdery mildew severity in cultivars and
lines of soybean. Scientia Agricola, v. 61, n. 6, p. 584-592, 2004.
LOURENÇÃO, A. L., PEREIRA, J. C. V. N. A. ; MIRANDA, M. A. C.; AMBROSANO,
G. M. B. Danos de percevejos e de lagartas em cultivares e linhagens de soja de ciclos
médio e semi-tardio. Anais da Sociedade Entomológica do Brasil, v. 28, n. 1, 1999.

96
LOURENÇÃO, A. L.; BRAGA, N. R.; MIRANDA, M. A. C.; VALLE, G. E.; PEREIRA,
J. C. V. N. A.; RECO, P. C. Avaliação de danos de percevejos e de desfolhadores em
genótipos de soja de ciclos precoce, semiprecoce e médio. Neotropical Entomology, v.
31, n. 4, p. 623-630, 2002.
LOURENÇÃO, A. L.; MIRANDA, M. A. C. Resistência de soja a insetos: VIII. IAC 78-
2318, linhagem com resistência múltipla. Bragantia, v. 46, n. 1, p. 65-72. 1987.
LOURENÇÃO, A. L.; MIRANDA, M. A. C.; NAGAI, V. Resistência de soja a insetos:
VII. Avaliação de danos de percevejos em cultivares e linhagens. Bragantia, v. 46, n. 1, p.
45-57, 1987.
LOURENÇÃO, A. L.; MIRANDA, M. A. C.; PEREIRA, J. C. V. N. A.; AMBROSANO,
G. M. B. Resistência de soja a insetos: X. Comportamento de cultivares e linhagens em
relação a percevejos e desfolhadores. Anais da Sociedade Entomológica do Brasil, v. 26,
n. 3, p. 543-550, 1997.
LOURENÇÃO, A. L.; PEREIRA, J. C. V. N. A.; MIRANDA, M. A. C.; AMBROSANO,
G. M. B. Avaliação de danos causados por percevejos e por lagartas em genótipos de soja
de ciclo precoce e semiprecoce. Pesquisa Agropecuária Brasileira, Brasília, v. 35, n. 5,
p. 879-886, 2000.
LOURENÇÃO, A. L.; RECO, P. C.; BAGA, N. R.; DO VALLE, G. E.; PINHEIRO, J. B.
Produtividade de genótipos de soja sob infestação da lagarta-da-soja e de percevejos.
Neotopical Entomology, v. 39, n. 2, p. 275-281, 2010.
LOURENÇÃO, A. L.; ROSSETTO, C. J.; MIRANDA, M. A. C. Resistência de soja a
Insetos: IV. Comportamento de cultivares e linhagens em relação a Hedilepta indicata
(Fabr.). Bragantia, v. 44, n. 1, p. 149-157, 1985.
LUEDDERS, V. D.; DICKERSON, W. A. Resistance of selected soybean genotypes and
segregating populations to cabbage looper feeding. Crop Science, v. 17, n. 3, p. 395-397,
1977.
LUSTOSA, P. R.; ZANUNCIO, J. C.; LEITE, G. L. D.; PICANÇO, M. Qualidade da
semente e senescência de genótipos de soja sob dois níveis de infestação de percevejos
(Pentatomidae). Pesquisa Agropecuária Brasileira, v. 34, n. 8, p. 1347-1351, 1999.
MAGRINI, E. A.; SILVEIRA NETO, S.; BOTELHO, P. S. M.; NEGRIM, S. G.
Ocorrência de percevejos no município de Pirassununga-SP, associada com a fenologia da
soja. Revista de Agricultura, v. 71, n. 2, p. 187-193, 1996.
McPHERSON, R. M.; BUSS, G. R.; ROBERTS, P. M. Assessing stink bug resistance in
soybean breeding lines containing genes from germplasm IAC-100. Journal of Economic
Entomology, v. 100, n. 4, p. 1456-1463, 2007.
MEDEIROS, M. A.; SCHMIDT, F. V. G.; LOIÁCONO, M. S.; CARVALHO, F. V.;
BORGES, M. Parasitism and predation of Euschistus heros (Fabr.) (Heteroptera:
Pentatomidae) eggs in Federal District, Brazil. Anais da Sociedade Entomológica do
Brasil, p. 397-401, 1997.

97
MENEGUIM, A. M.; ROSSINI, M. G.; PANIZZI, A. R. Desenvolvimento de ninfas e
adultos de Euschistus heros (F.) (Hemiptera: Pentatomidae) em frutos de amendoim-bravo
Euphorbia heteroplhylla (Euphorbiaceae) e em sementes e vagens de soja. Resumos do
Congresso Brasileiro de Entomologia. 12, p. 43, 1989.
MINER, F. D. Biology and control of stink bugs on soybeans. Arkansas: Agricultural
Experiment Station, 1966. 40 p.
MIRANDA, M. A. C.; BRAGA, N. R.; LOURENÇÃO, A. L.; MIRANDA, F. T. S.;
UNEDA, S. H.; ITO, M. F. Descrição, produtividade e estabilidade da cultivar de soja
IAC-24, resistente a insetos. Bragantia, v. 62, n. 1, p. 29-37, 2003.
MIRANDA, M. A. C.; LOURENÇÃO, A. L. Melhoramento genético da soja para a
resistência a insetos: uma realidade para aumentar a eficiência do controle integrado de
pragas e viabilizar a soja orgânica. In CONGRESSO BRASILEIRO DE SOJA E
MERCOSOJA, 2. Londrina. Anais... Londrina: Embrapa-Soja, 2002, 52-60 p.
MIRANDA, M. A. C.; ROSSETTO, C. J.; ROSSETTO, D.; BRAGA, N. R.;
MASCARENHAS, H. A. A.; TEIXEIRA, J. P. F.; MASSARIOL, A. Resistência de soja a
Nezara viridula e Piezodorus guildinii em condições de campo. Bragantia, Campinas, v.
38, n. 1, p. 181-188, 1979.
MORAIS, A. A. C.; SILVA, A. L. Soja e suas aplicações. Rio de janeiro: Medsi, 1996.
259 p.
MOSCARDI, F.; CORRÊA-FERREIRA, B. S.; DINIZ, M. C.; BONO, I. L. S. Incidência
estacional de fungos entomógenos sobre populações de percevejos-pragas da soja. In:
EMBRAPA. Centro Nacional de Pesquisa de Soja. Resultados de Pesquisa de Soja
1986/87. Londrina, 1988. p. 73. (EMBRAPA-CNPSo. Documentos, 28).
MOURÃO, A. P. M. & PANIZZI, A. Diapausa e diferentes formas sazonais em Euschistus
heros (Fabr.) (Hemiptera: Pentatomidae) no Norte do Paraná. Anais da Sociedade
Entomológica do Brasil, v. 29, n. 2, p. 205-218, 2000.
MYASAKA, S. Soja no Brasil Central. Campinas: Fundação Cargil, 1982. 444 p.
MYASAKA, S.; MEDINA, J. C. A soja no Brasil. Campinas: Editora ITAL, 1981. 1062
p.
NIVA, C. C.; PANIZZI, A. R. Efeitos do cloreto de sódio no comportamento de Nezara
viridula (L.) (Heteroptera: Pentatomidae) em vagens de soja. Anais da Sociedade
Entomológica do Brasil, v. 25, p. 251-257, 1996.
OLIVEIRA, E. D. M.; PANIZZI, A. R. Performance of nymphs and adults of Piezodorus
guildinii (Westwood) (Heteroptera: Pentatomidae) on soybean pods at different
development stages. Brazilian Archives of Biology and Technology, v. 46, n. 2, p. 187-
192, 2003.

98
PAINTER, R. H. Insect resistence in crop plants. New York: McMillan, 1951. 520 p.
PANDA, N. Principles of host-plant resistance to insect pests. llanheld/Universe, New
York, 1979. 431 p.
PANDA, N.; KHUSH, G. S. Host plant resistance to insects. Wallingford: CAB
International / Marília: IRRI, 1995. 431 p.
PANIZZI, A. R. Ecologia nutricional de insetos e suas implicações no manejo de
pragas. Manole / Brasília, 1991. 359p.
PANIZZI, A. R.; NIVA, C. C. Overwintering strategy of the brown stink bug in Northern
Paraná. Pesquisa Agropecuária Brasileira, p. 509-511, 1994.
PANIZZI, A. R. A biodiversidade vegetal no manejo de percevejos. In: CORRÊA-
FERREIRA, B. S. (Org.). Soja orgânica: alternativas para o manejo dos insetos
pragas. Londrina: Embrapa Soja, 2003, p.47-55.
PANIZZI, A. R. Stink bugs on soybean in northeastern Brazil and a new record on the
southern green stink bug, Nezara viridula (L.) (Heteroptera: Pentatomidae). Neotropical
Entomology, v. 31, p. 331-332, 2002.
PANIZZI, A. R.; BUENO, A. F.; SILVA, F. A. C. Insetos que atacam vagens e grãos. In:
HOFFMANN-CAMPO, C. B., CORRÊA-FERREIRA, B. S., MOSCARDI, F. Soja:
manejo integrado de insetos e outros artrópodes-praga. Embrapa, 2012. 859 p.
PANIZZI, A. R.; MENEGUIM, A. M.; ROSSINI, M. C. Impacto da troca de alimento da
fase ninfal para a fase adulta e do estresse nutricional na fase adulta na biologia de N.
viridula (Hemiptera: Pentatomidae). Pesquisa Agropecuária Brasileira, v. 24, n. 8, p.
945-954, 1989.
PANIZZI, A. R.; NIVA, C. C.; HIROSE, E. Feeding preference by stink bugs
(Heteroptera: Pentatomidae) for seeds within soybean pods. Journal Entomology Science,
v.30, n. 3, p. 333-341, 1995.
PANIZZI, A. R.; PANIZZI, M. C. C.; BAYS, I. A.; ALMEIDA, L. A. Danos por
percevejos em genótipos de soja com semente pequena. Pesquisa Agropecuária
Brasileira, v. 21, n. 6, p. 571-577, 1986.
PANIZZI, A. R.; SILVA, F. A. C. Insetos sugadores de sementes (Heteroptera). In:
PANIZZI. A.R.; PARRA, J.R.P. Bioecologia e nutrição de insetos. Brasília: Embrapa,
2009, p. 465-522.
PANIZZI, A. R.; SLANSKY JUNIOR, F. Review of phytophagous pentatomids
(Hemiptera: Pentatomidae) associated with soybean in the Americas. Florida
Entomologist, v. 68, n. 1, p. 184-203, 1985.
PANIZZI, A. R. Wild hosts of pentatomids: ecological significance and role in their pest
status on crops. Annu. Rev. Entomol., p. 99–122, 1997.

99
PARON, M. J. F. O.; LARA, F. M. Relação entre tricomas foliares de genótipos de
feijoeiro comum, Phaseolus vulgaris L. e resistência a Diabrotica speciosa Germar, 1824
(Coleoptera: Chrysomelidae). Ciência e Agrotecnologia, v. 29, n. 4, p. 894-898, 2005.
PEREIRA, P. R. V. S.; SALVADORI, J. R. Aspectos populacionais de percevejos
fitófagos ocorrendo na cultura da soja (Hemiptera: Pentatomidae) em duas áreas do
norte do Rio Grande do Sul. Passo Fundo: Embrapa Trigo, 2008. 10 p. (Embrapa Trigo.
Comunicado Técnico, 253).
PINHEIRO, J. B. Dialelo parcial entre parentais de soja resistentes e suscetíveis a
insetos. 1993. 143 f. Dissertação (Mestrado em Genética e Melhoramento de Plantas) -
Escola Superior de Agricultura "Luiz de Queiroz", Universidade de São Paulo, Piracicaba,
São Paulo. 1993.
PINHEIRO, J. B.; VELLO, N. A.; ROSSETTO, C. J.; ZUCCHI, M. I. Potential of soybean
genotypes as insect resistance sources. Crop Breeding and Applied Biotecnology, v. 5, p.
293-300, 2005.
PINTO, S. B.; PANIZZI, A. R. Performance of nymphal and adult Euschistus heros (F.) on
milkweed and on soybean and effect of food switch on adult survivorship, reproduction
and weight gain. Anais da Sociedade Entomológica do Brasil, v.23, p. 549-555, 1994.
PIUBELLI, G. C. Ciclo biológico, preferência alimentar de Nezara viridula L.
(Hemiptera: Pentatomidae) e seu papel na produção de flavonóides induzidos em
genótipos de soja [Glycine max (L.) Merrill]. 2000. 89 f. Dissertação (Mestrado em
Entomologia Agrícola) - Faculdade de Ciências Agrárias e Veterinárias, Universidade
Estadual Paulista, Jaboticabal, São Paulo. 2000.
PIUBELLI, G. C.; HOFFMANN-CAMPO, C. B.; ARRUDA, I. C.; LARA, F. M. Nymphal
development, lipid content, growth and weight gain of Nezara viridula (L.) (Heteroptera:
Pentatomidae) fed on soybean genotypes. Neotropical Entomology, v. 32, n. 1, 2003.
PRABHAKER, N.; COUDRIET, D. L.; MEYER-DRIK, D. E. Insecticide resistance in the
sweetpotato-whitefly Bemisia tabaci (Homoptera: Aleyrodidae). Journal of Economic
Entomology, v. 78, p. 748-752, 1985.
PRADO, R. C. O. Soja é alimento e energia. Boletim de Pesquisa de Soja, Fundação MT,
2007. 11-14 p.
RAIJ, B.; CANTARELLA, H.; QUAGGIO, J. Recomendações de adubação e calagem
para o Estado de São Paulo. Campinas: Instituto Agronômico–Fundação IAC, 1997.
Boletim Técnico 100, 285 p.
RAGSDALE, D. W.;LAWSON, A. D.; NEWSOM, L. D. Quantitative assessment of the
predators of Nezara viridula eggs and nymphs within a soybean agroecosystem using an
ELISA. Environmental Entomology, v. 10, p. 402–405, 1981.
RAMIRO, L. C. B.; RAMIRO, Z. A.; FARIA, A. M.; MARINO, C. A. B. Danos causados
por percevejos (Hemiptera: Pentatomidae) em cinco cultivares de soja (Glycine max (L.)

100
Merrill, 1917) no município de Araçatuba, SP. Arquivos do Instituto Biológico, São
Paulo, v.20, n. 2, p. 169-175, 2003.
RAMIRO, Z. A. A soja no Brasil central. Campinas: Fundação Cargil, 1982, p. 215-243.
RAMIRO, Z. A.; MASSARIOL, A. A. manejo de insetos na cultura da soja. In: A soja no
Brasil central. Campinas: Fundação Cargil, p. 141-155, 1977.
RAMIRO, Z. A.; BATISTA FILHO, A; MACHADO, L. A.; SANTOS, J. C. C.; FARIA,
A. M. Levantamento de pragas em quatro cultivares e duas linhagens de soja no município
de Orlândia, SP. I - Percevejos. Anais da Sociedade Entomológica do Brasil, v. 17
(supl.), p. 05-17, 1988.
REZENDE, J. A. M; MIRANDA, M. A. C.; MASCARENHAS, H. A. A. Comportamento
de cultivares de soja em relação à área foliar comida por lagartas das folhas. Bragantia, v.
39, n. 1, p. 161-165, 1980.
ROESSING, A. C.; LAZZAROTTO, J. J. Criação de empregos pelo complexo
agroindustrial da soja. Londrina: Embrapa-CNPSo, 2004. 50 p. (Embrapa- CNPSo.
Documentos, 233).
ROGGIA, S. Caracterização de fatores determinantes dos aumentos populacionais de
ácaros tetraniquídeos em soja. 2010. 153 f. Tese (Doutorado em Entomologia) - Escola
Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba , 2010.
ROSA-GOMES, M. F. Avaliação de danos de quatro espécies de percevejos
(Heteroptera: Pentatomidae) em trigo, soja e milho. 2010. 93 f. Tese (Doutorado em
Agronomia) – Universidade de Passo Fundo, 2010.
ROSSETTO, C. J.; GALLO, P. B.; RAZERA, L. F.; BORTOLETTO, N.; IGUE, T.;
MEDINA, P. F.; TISSELI FILHO, O.; AQUILERA, V.; VEIGA, R. F. A.; PINHEIRO, J.
B. Mechanisms of resistance to stink bug complex in the soybean cultivar IAC-100. Anais
da Sociedade Entomológica do Brasil, 1995. v. 24, n. 3, 517-522 p.
ROSSETTO, C. J.; IGUE, T; MIRANDA, M. A. C.; LOURENÇÃO, A. L. Resistência de
soja a insetos: VI. Comportamento de genótipos em relação a percevejos. Bragantia, v. 45,
n. 2, p. 223-235, 1986.
ROSSETTO, C. J.; LARA, F. M. Diluição de dano, mecanismo da pseudo-resistência. In.
CONGRESSO BRASILEIRO DE ENTOMOLOGIA, 13., 1991, Recife. Resumos...
Recife, Pernambuco, 1991. 535 p.
ROSSETTO, C. J.; LOURENÇÃO, A. L.; IGUE, T.; MIRANDA, M. A. C. Picadas de
alimentação de Nezara viridula em cultivares e linhagens de soja de diferentes graus de
suscetibilidade. Bragantia, v. 40, n. 10, p. 109-114, 1981.
ROSSETTO, C. J.; LOURENÇÃO, A. L.; MIRANDA, M. A. C.; IGUE, T. Resistência de
soja a insetos. II. Teste de livre escolha entre a linhagem IAC 73/228 e o cultivar Paraná,
infestados por Nezara viridula (L.) em telado. Bragantia, v. 43, n. 1, p. 141-153, 1984.

101
SALUSO, A.; XAVIER, L.; SILVA, F. A. C.; PANIZZI, A. R. An invasive pentatomid
pest in Argentina: Neotropical brown stink bug, Euschistus heros (F.) (Hemiptera:
Pentatomidae). Neotropical Entomology, v. 40, p. 704-705, 2011.
SEDIYAMA, T., PEREIRA, M. G. & SEDIYAMA, C. S. Cultura da soja. Viçosa:
Universidade Federal de Viçosa, 1985. 95 p. (Boletim, 212)
SEDIYAMA, T. (Org.). Tecnologias de produção e usos da soja. Londrina, PR:
Mecenas, 2009, v. 1. 314 p.
SHAW, N.; CHIN, C.; PAN, W. A vegetarian diet rich in soybean products compromises
iron status in young students. Journal of Nutrition, v. 125, p. 212-219, 1995.
SILVA, C. C.; LAUMANN, R. A.; BLASSIOLI, M. C.; BORGES, M. Otimização da
técnica de criação de Euschistus heros para a multiplicação do parasitoide de ovos
Telenomus podisi. Boletim de Pesquisa e Desenvolvimento 195, Embrapa Recursos
Genéticos e Biotecnologia, 2007, 19 p.
SILVA, J. P. G. F. Resistência de genótipos de soja Glycine Max (L.) a Piezodorus
guildinii (Westewood) (Hemiptera: Pentatomidae). 2013. 100 f. Dissertação (Mestrado
em Agronomia) Universidade Estadual Paulista “Júlio de Mesquita Filho”, Botucatu,
2013b.
SILVA, G. V. Efeito de plantas Bt de soja e milho sobre pragas não-alvo e seus
inimigos naturais. 2013. 84 f. Dissertação (Mestrado em Ciências Biológicas)
Universidade Federal do Paraná, Curitiba, 2013.
SILVA, L. D.; OMOTO, C.; BLEICHER, E.; DOURADO, P. Monitoramento da
suscetibilidade a inseticidas em populações de Bemisia tabaci (Gennadius) (Hemiptera:
Aleyrodidae) no Brasil. Neotropical Entomology, v. 38, n. 1, p. 116-125, 2009.
SILVA, M. T. B.; CORRÊA-FERREIRA, B. S.; SOSA-GÓMEZ, D. R. Erro e resistência.
Revista Cultivar Grandes Culturas, Pelotas, v. 8, n. 82, p. 22-25, fev. 2006.
SMITH, C. M. Expression mechanisms and chemistry of resistance in soybean (Glycine
max L. Merr.) to soybean looper, Pseudoplusia includens (Walker). Insect Science and its
Application, v. 6, n. 3, p. 243-248, 1985.
SMITH, C. M. Plant resistance to arthropods. 2005, 423 p.
SORIA, M. F.; DEGRANDE, P. E.; PANIZZI, A. R. Algodoeiro invadido. Revista
Cultivar, v. 131, p. 18-20, 2010.
SOSA-GÓMEZ, D. R.; CORSO, I. C.; MORALES, L. C. Insecticide resistance to
endosulfan, monocrotophos and methamidophos in the Neotropical brown stink bug,
Euschistus heros (Fabr.). Neotropical Entomology, v. 30, n. 2, p. 317-320, 2001.
SOSA-GÓMEZ, D. R,; LOPES, I.; de O. N.; SILVA, J. J.; OLIVEIRA, M. C. N. de.
Resistência de pentatomídeos a inseticidas químicos e linhas base de suscetibilidade
determinadas mediante aplicação tópica. In: Reunião de pesquisa de soja da região central

102
do Brasil, 27., 2005, Cornélio Procópio. Resumos...Londrina: Embrapa Soja, 2005. 131-
132. (Embrapa Soja. Documentos, 257).
SOSA-GÓMEZ, D. R. & OMOTO, C. Resistência a inseticidas e outros agentes de
controle em artrópodes associado à cultura da soja. In: HOFFMANN-CAMPO, C. B.,
CORRÊA-FERREIRA, B. S., MOSCARDI, F. Soja: manejo integrado de insetos e outros
artrópodes-praga. Embrapa, 2012. 859 p.
SOSA-GÓMEZ, D. R.; BOUCIAS, D. G.; NATION, J.L. Attachment of Metarhizium
anisopliae to the Southern green stink bug Nezara viridula cuticle and fungistatic effect of
cuticular lipids and aldehydes. Journal of Invertebrate Pathology, v. 69, p. 31-39, 1997.
SOSA-GÓMEZ, D. R.; CORRÊA-FERREIRA, B. S.; HOFFMANN-CAMPO, C. B.;
CORSO, I. C.; OLIVEIRA, L. J.; MOSCARDI, F.; PANIZZI, A. R.; BUENO, A. F.;
HIROSE, E. Manual de identificação de insetos e outros invertebrados da cultura da
soja: 2. ed. Londrina: Embrapa Soja, 2010. 80 p.
SOSA-GÓMEZ, D. R.; MOSCARDI, F. Retenção foliar diferencial em soja provocada por
percevejos (Heteroptera: Pentatomidae). Anais da Sociedade Entomológica do Brasil,
1995. v. 24, n. 2, 401-404 p.
SOSA-GÓMEZ, D. R.; SILVA, J. J. 2010. Neotropical brown stink bug (Euschistus heros)
resistance to methamidophos in Paraná, Brasil. Pesquisa Agropecuária Brasileira, v.45,
p.767-769, 2010.
SOSA-GÓMEZ, D. R.; SILVA, J. J.; LOPES, I. O. N.; CORSO, I. C.; ALMEIDA, A. M.
R.; PIUBELLI DE MORAES, G. C.; BAUR, M. E. Insecticide susceptibility of Euschistus
heros (Heteroptera: Pentatomidae) in Brazil. Journal of Economic Entomology, v. 102, p.
1209-1216, 2009.
SOUZA, E. S. Resistência de genótipos de soja Glycine max (L.) a Nezara viridula (L.,
1758) (Hemiptera: Pentatomidae). 2010. 56 f. Dissertação (Mestrado em Proteção de
Plantas) – Faculdade de Ciências Agronômicas, Universidade Estadual Paulista, Botucatu,
São Paulo. 2010.
SOUZA, R. F.; TOLEDO, J. F. F. Genetic analysis of soybean resistance to stink
bug. Revista Brasileira de Genética, São Paulo, v.18, n.4, p. 593-598, 1995.
SOUZA, E. S.; BALDIN, E. E. L.; FANELLA, T. L. M. Desenvolvimento de Nezara
viridula (L., 1758) (Hemiptera: Pentatomidae) em genótipos de soja. Boletin de Sanidad
Vegetal Plagas, v. 38, p. 41-49, 2012.
STANSLY, P. A.; SCHUSTER, D. J. The sweet-potato whitefly and integrated pest
management of tomato. In: VAVRINA, C.S. Proceedings Florida Tomato Institute
Vegetal Crops Special Series. Gainesville: University of Florida, 1992. 54-74 p.
SUTHERLAND, O. R. W. RUSSELL, G. B.; BIGGS, D. R.; LANE, G. A. Insect feeding
deterrent activity of phytoalexin isoflavonoids. Biochemical Systematics and Ecology, v.
8, n. 1, p. 73-75, 1980.

103
SUWANPORNSKUL, R.; KHADKAO, S. Screening soybean to pod sucking bugs. TVIS
Newsletter, Chiangmai, Tailand, v.1, n. 1, p. 20-23, 1996.
TESTER, C. F. Constituents of soybean cultivars differing in insect resistance.
Phytochemistry, v.16, p. 1899-1901, 1977.
TOMAZINI, M. J.; GRAVENA, S. Espécies de Geocoris Fallen que ocorrem em algodão
e soja em Jaboticabal, SP. Anais da Sociedade Entomológica do Brasil, v. 23, p. 259-
264, 1994.
TURNIPSEED, S. G. Influence of trichome density on populations of small phytophagous
insects on soybean. Environmental Entomology, v. 6, p. 815-817, 1977.
TURNIPSEED, S. G.; SULLIVAN, M. J. Plant resistance in soybean insect management.
In: HILL, D. World Soybean Research. Interstate Printers and Publishers. Danville, 1976.
1073 p.
VALLE, G. E.; LOURENÇÃO, A. L. Resistência de genótipos de soja a Bemisia tabaci
(Genn.) biótipo B (Hemiptera: Aleyrodidae). Neotropical Entomology, v. 31, n. 1, p. 285-
295, 2002.
VAN DUYN, J. W.; TURNIPSEED, S. G.; MAXWELL, J. D. Resistance in soybeans to
the Mexican bean beetle. I. Sources of resistance. Crop Science, v. 11, n. 4, p. 572-573,
1971.
VAN DUYN, J. W.; TURNIPSEED, S. G.; MAXWELL, J. D. Resistance in soybeans to
the Mexican bean beetle: II. Ractions of the beetle to resistant plants. Crop Science, v. 12,
p. 561-562, 1972.
VAN SCHAIK, P. H. & PROBST, A. H. Effects of some enviromental factors on flower
production and reproductive efficiency in soybeans. Agronomy Journal, v.50, p.192-197,
1958.
VENDRAMIM, J. D. A resistência de plantas e o manejo de pragas. In: CROCOMO, W.B.
(Org.)., Manejo Integrado de Pragas. Botucatu: UNESP/CETESB, 1990, p. 177-197.
VENDRAMIM, J. D.; GUZZO, E. C. Resistência de plantas e a bioecologia e nutrição de
insetos. In: PANIZZI, A.R.; PARRA, J.R.P. Bioecologia e nutrição de insetos. Brasília:
Embrapa, 2009, p. 1107-1140.
VERNETTI, F. J. Soja – planta, clima, pragas, moléstias e invasoras – vol I, genética e
melhoramento. Campinas: Fundação Cargill, 1983. v. 2, 990 p.
VICENTINI, R.; JIMENEZ, H. A. El vaneo de los frutos en soja. Paraná, Argentina:
Instituto Nacional de Tecnología Agropecuaria; Estación Experimental Agropecuaria,
1977. 30 p.
VILLAS BÔAS, G. L.; PANIZZI, A. R. Biologia de Euchistus heros (Fabricius, 1798).
Anais da Sociedade Entomológica do Brasil, v. 9, n. 1, p. 105-113, 1980.

104
VILLAS BÔAS, G. L.; GAZZONI, D. L.; OLIVEIRA, M. C. N. de; COSTA, N. P.;
ROESSING, A. C.; FRANCA NETO, J. de B.; HENNING, A. A. Efeito de diferentes
populações de percevejos sobre o rendimento e seus componentes, características
agronômicas e qualidade de sementes de soja. Boletim de Pesquisa 1. Londrina:
Embrapa, 1990. 43 p.
VILLAS BÔAS, G. L.; MOSCARDI, F.; CORRÊA-FERREIRA, B. S.; HOFFMANN-
CAMPO, C. B.; CORSO, I. C., PANIZZI, A. R. Indicações do manejo de pragas para
percevejos. Londrina: Embrapa-CNPSo, 1985. 15 p. (Embrapa- CNPSo. Documentos, 9).
VILLAS-BÔAS, G. L.; PANIZZI, A. R. Biologia de Euschistus heros (Fabricius, 1789)
em soja (Glycine max L. Merrill). Anais da Sociedade Entomológica do Brasil, 1980.
v.9, 105-113 p.

105
APÊNDICES
APÊNDICE 1
APÊNDICE 2
LABORATÓRIO DE FERTILIDADE DO SOLO
# RESULTADOS DE ANÁLISE DE SOLO #
INTERESSADO: Efrain de Santana Souza / Prof. Edson Luiz Lopes Baldin ANO: 2011
PROCEDÊNCIA: Área 17 - Fazenda Pesquisa
AMOSTRA (S) pH M.O. Presina Al3+
H+Al K Ca Mg SB CTC V% S
Labor. Int. CaCl2 g/dm3 mg/dm
3
_ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ mmolc/dm
3 _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ mg/dm
3
UG 84 1 4,9 28 28 1 42 3,8 27 17 48 90 53 30
AMOSTRA (S) BORO COBRE FERRO MANGANÊS ZINCO
Labor. Int. _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _
mg/dm3 _ _ _ _ _ _ _ _ _ _ _ _ __ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _
UG 84 1 0,36 12,6 15 31,3 1,6

106
APÊNDICE 3
LABORATÓRIO DE FERTILIDADE DO SOLO
# RESULTADOS DE ANÁLISE DE SOLO #
INTERESSADO: Efrain de Santana Souza / Prof. Edson Luiz Lopes Baldin ANO: 2012
PROCEDÊNCIA: Área Experimental Didática do Dep. Agricultura
AMOSTRA (S) pH M.O. Presina Al3+
H+Al K Ca Mg SB CTC V% S
Labor. Int. CaCl2 g/dm3 mg/dm
3
_ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ mmolc/dm
3 _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ mg/dm
3
GT 79 1 4,7 20 35 --- 43 3,3 31 12 46 89 52 ---
APÊNDICE 4
LABORATÓRIO DE FÍSICA DE SOLO
# RESULTADOS DE ANÁLISE DE SOLO #
INTERESSADO: Prof. Edson Luiz Lopes Baldin / Efrain Santana Souza
PROCEDÊNCIA: Depto. Proteção de Plantas
Amostra(s) Areia
Argila Silte Umid. Textura Balão Vol. Torrão Imp.
Grossa Fina Total Densidade (g/cm3)
Labor. Int. (g/Kg) (%) do Solo Partículas Solo
1 1 ---- ---- 162 607 231 ---- Muito Arg. ---- ----
AMOSTRA (S) BORO COBRE FERRO MANGANÊS ZINCO
Labor. Int. _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _
mg/dm3 _ _ _ _ _ _ _ _ _ _ _ _ __ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _
GT 79 1 0,17 9,5 43 60,7 2,5





























