Embed Size (px)
Citation preview

UNIVERSIDADE ESTADUAL PAULISTA “JÚLIO DE MESQUITA FILHO”
FACULDADE DE CIÊNCIAS AGRONÔMICAS
CÂMPUS DE BOTUCATU
INFLUÊNCIA DO GLYPHOSATE NO PERFIL BIOQUÍMICO E
FISIOLÓGICO DE POPULAÇÕES DE AZEVÉM (Lolium multiflorum)
SUSCETÍVEIS E RESISTENTES AO HERBICIDA
GILMAR JOSÉ PICOLI JUNIOR
Tese apresentada à Faculdade de Ciências
Agronômicas da UNESP – Campus de
Botucatu para obtenção do título de
Doutor em Agronomia (Agricultura)
BOTUCATU – SP
Janeiro – 2016

UNIVERSIDADE ESTADUAL PAULISTA “JÚLIO DE MESQUITA FILHO”
FACULDADE DE CIÊNCIAS AGRONÔMICAS
CÂMPUS DE BOTUCATU
INFLUÊNCIA DO GLYPHOSATE NO PERFIL BIOQUÍMICO E
FISIOLÓGICO DE POPULAÇÕES DE AZEVÉM (Lolium multiflorum)
SUSCETÍVEIS E RESISTENTES AO HERBICIDA
GILMAR JOSÉ PICOLI JUNIOR
Orientador: Prof. Dr. Caio Antonio Carbonari
Co-Orientador: Prof. Dr. Edivaldo Domingues Velini
Tese apresentada à Faculdade de Ciências
Agronômicas da UNESP – Campus de
Botucatu para obtenção do título de
Doutor em Agronomia (Agricultura)
BOTUCATU – SP
Janeiro - 2016



III
“A tarefa não é tanto ver aquilo que ninguém viu, mas
pensar o que ninguém ainda pensou sobre aquilo que
todo mundo vê.” (Arthur Schopenhauer)
Aos meus queridos pais Giomar e Dayse, pelo o
AMOR incondicional e que apesar da distância,
sempre estiveram ao meu lado.
Ofereço e dedico.

IV
AGRADECIMENTOS
A Deus, por estar sempre ao meu lado nessa importante caminhada.
Aos meus pais, pela dedicação incondicional à minha educação e por acreditarem que
os meus sonhos são seus sonhos. Meus eternos agradecimentos.
Aos meus irmãos Andrei e Maria e cunhados Cesar e Elizabete pelo carinho, apoio e
incentivo.
Aos meus sobrinhos Yurik, Kalel, Davi e Arthur, por lembrarem que dentro de mim
mora uma criança com sorriso puro e olhar inocente quando estou com eles.
Ao meu orientador Prof. Dr. Caio Antonio Carbonari, por ter me aceitado como
orientado, pela paciência, amizade, pelo exemplo de profissionalismo e pelos conhecimentos
transmitidos. Muito obrigado!
Ao meu co-orientador Prof. Dr. Edivaldo Domingues Velini pela generosidade em
dividir sua sabedoria.
Aos meus indispensáveis e eternos “irmãos” da pós-graduação Ana Karollyna Alves
de Matos, Plínio Saulo Simões, Giovanna Larissa Gimenes Cotrick Gomes, Diego Belapart,
Leandro Tropaldi, Gabrielle de Castro Macedo, Edicarlos Batista de Castro, Renan Fonseca
Nascentes, Ronei Ben, Débora Latorre e Bruna Marchesi pela valiosa ajuda durante o período
dos experimentos e pelos bons momentos vividos.
Aos colegas de laboratório Rosilaine Araldi, Marcelo Girotto, Natália Corniani, Ilca
Puertas Freitas e Silva e Samir Paulo Jasper pelos bons momentos de convívio durante o curso
de pós-graduação.
Aos amigos e funcionários do Núcleo de Pesquisas Avançadas em Matologia José
Guilherme Cordeiro, José Roberto Marques Silva e Marcelo L. Siono pelas preciosas
colaborações durante a realização dos experimentos.
Á Prof. Dra. Giuseppina Pace Pereira Lima pela disponibilidade de seu laboratório e
pelos alegres momentos vividos em todos os dias de trabalho.
Aos colegas do Laboratório de Análises Bioquímicas Vegetais do Departamento de
Química e Bioquímica do Instituto de Biociências da UNESP de Botucatu pelas valiosas
contribuições.
Aos colegas Cleber Junior Jadoski e Luan Ormond pela ajuda com as análises do
IRGA e interpretação dos dados.

V
Aos meus queridos amigos Rosalvo Pires, Ivete Teixeira, Flaviane Lazarini, Carolina
Junqueira, Daiana Bampi e Ronaldo Araújo, que nesta caminhada, foram extremamente
importantes para que eu chegasse até aqui.
Ao Dr. Stephen B. Powles, pelo exemplo de pessoa, Dra. Qin Yu e Dr. Heping Han,
pela orientação, cuidado, e excelentes contribuições não só na área acadêmica como na vida
pessoal durante o período de doutorado sanduíche na Australia Herbicide Resistance
Iniciative (AHRI), University of Western Australia, Austrália.
Á Arysta LifeScience através de Angelo Stasievski pelo fornecimento das sementes.
Á Embrapa Trigo através do Prof. Dr. Leandro Vargas pelo fornecimento das
sementes.
Ao programa de pós-graduação em Agronomia/Agricultura e a Faculdade de Ciências
Agronômicas pela oportunidade e formação.
A Coordenação de Aperfeiçoamento de Pessoal de Ensino Superior (CAPES), pela
bolsa de estudos concedida.
A todos que de alguma forma contribuíram para que esse sonho se tornasse realidade.
Muito obrigado!

VI
SUMÁRIO
LISTA DE TABELAS...........................................................................................................VIII
LISTA DE FIGURAS...............................................................................................................IX
1. RESUMO ............................................................................................................................... 1
2. SUMMARY ........................................................................................................................... 3
3. INTRODUÇÃO ...................................................................................................................... 5
4. REVISÃO BIBLIOGRÁFICA ............................................................................................... 8
4.1 Azevém: morfologia, biologia e distribuição. ................................................................... 8
4.2 Azevém como planta daninha. .......................................................................................... 9
4.3 Glyphosate: importância, características químicas e modo de ação. .............................. 10
4.4 Efeitos do glyphosate na fotossíntese. ............................................................................ 18
4.5 Fenilalanina amônio liase (PAL)..................................................................................... 20
4.6 Resistência de plantas daninhas aos herbicidas............................................................... 22
4.7 Resistencia de plantas daninhas ao glyphosate. .............................................................. 26
4.8 Azevém (Lolium multiflorum) resistente ao glyphosate. ................................................ 30
5. MATERIAL E MÉTODOS .................................................................................................. 32
5.1 Curvas de dose-resposta das diferentes populações de azevém. ..................................... 32
5.2 Efeitos do glyphosate na enzima fenilalanina amônia liase nas diferentes populações...34
5.3 Efeitos do glyphosate na fotossíntese das diferentes populações. .................................. 37
5.4 Avaliação dos teores de glyphosate, AMPA e compostos da rota do ácido chiquímico
nas diferentes populações. ............................................................................................... 39
5.5 Análise dos dados ............................................................................................................ 42
5.5.1 Curvas de dose-resposta das diferentes populações de azevém ............................... 42
5.5.2 Efeitos do glyphosate na enzima fenilalanina amônia liase nas diferentes
populações................................................................................................................43
5.5.3 Efeitos do glyphosate sobre a fisiologia das diferentes populações ......................... 43
5.5.4 Avaliação dos teores de glyphosate, AMPA e compostos da rota do ácido
chiquímico nas diferentes populações ..................................................................... 43
6. RESULTADOS E DISCUSSÃO ......................................................................................... 44
6.1 Curvas de dose-resposta das diferentes populações de azevém. ..................................... 44
6.2 Efeitos do glyphosate na enzima fenilalanina amônia liase nas diferentes populações...48
6.3 Efeitos do glyphosate na fotossíntese das diferentes populações. .................................. 54
6.3.1 Suscetível (S) x Resistente (R2) ............................................................................... 54
6.3.2 Suscetível (S) x Resistente (R1) ............................................................................... 66
6.4 Avaliação dos teores de glyphosate, AMPA e compostos da rota do ácido chiquímico
nas diferentes populações. ............................................................................................... 73

VII
7. CONCLUSÕES .................................................................................................................... 97
8. REFERÊNCIAS ................................................................................................................... 98

VIII
LISTA DE TABELAS
Tabela 1. Doses de glyphosate utilizados no experimento. Botucatu - SP, 2014.. ................... 33
Tabela 2. Estimativas dos parâmetros a, b, c e o coeficiente de determinação (r²) do modelo
log-logístico para as populações de Lolium multiflorum em relação à porcentagem de controle
aos 21 DAA. Botucatu – SP... .................................................................................................. 47
Tabela 3. Estimativas dos parâmetros a, b, c e o coeficiente de determinação (r²) do modelo
log-logístico para as populações de Lolium multiflorum em relação à porcentagem de massa
seca aos 28 DAA. Botucatu – SP. ............................................................................................ 48
Tabela 4. Atividade da enzima fenilalanina amônia liase (PAL) em plantas de azevém
suscetível (S) e com suspeita de resistente (R1) coletadas as 12, 24 e 72 horas após a
aplicação do herbicida glyphosate. Botucatu - SP... ................................................................. 49
Tabela 5. Atividade da enzima fenilalanina amônia liase (PAL) em plantas de azevém
suscetível (S) e Resistente (R1) coletadas as 12, 24, 48 e 72 horas após a aplicação do
herbicida glyphosate. Botucatu - SP... ...................................................................................... 51
Tabela 6. Análise de variância e valores de diferença mínima significativa (DMS) para os
compostos analisados. Botucatu – SP................ ...................................................................... 73

IX
LISTA DE FIGURAS
Figura 1. Rotas de decomposição microbiológica do glyphosate. Fonte: Amarante Jr. et al.,
2002. ......................................................................................................................................... 11
Figura 2. Rota do ácido chiquímico. Fonte: Maeda e Dudareva, 2012. ................................... 14
Figura 3. Corismato em negrito como precursor do (1) prefenato, (2) antranilato, (3)
aminodeoxicorismato, (4) p-hidroxibenzoato, (5) isocorismato. As enzimas envolvidas são:
(1) corismato mutase, (2) antranilato sintase, (3) aminodeoxicorismato sintase, (4), corismato
piruvato liase, (5) isocorismato sintase. Fonte: Dosselaere e Vanderleyden,
2001...........................................................................................................................................15
5
Figura 4. Aminoácidos aromáticos e alguns compostos fenólicos. Fonte: Velini et al., 2012..16
Figura 5. Via biossintética fenilpropanóide. Fonte: Du et al., 2009. ........................................ 20
Figura 6. Estádio das plantas de azevém no momento da aplicação dos tratamentos Botucatu -
SP, 2014. ................................................................................................................................... 33
Figura 7. Pulverizador estacionário com as unidades experimentais. Botucatu - SP, 2014. .... 34
Figura 8. Procedimento de preparo de amostra para PAL. Botucatu - SP, 2014. .................... 36
Figura 9. Procedimento de extração e quantificação da PAL. Botucatu - SP, 2014. ............... 37
Figura 10. IRGA modelo LI-6400. Botucatu - SP, 2014. ......................................................... 38
Figura 11. Procedimentos da coleta das folhas. Botucatu - SP, 2014. ..................................... 40
Figura 12. Procedimentos para extração dos compostos. Botucatu - SP, 2014. ....................... 41
Figura 13. Cromatógrafo Líquido (Prominence UFLC) acoplado ao espectrômetro de massas
(3200 Q TRAP) – LC-MS/MS. ................................................................................................ 42
Figura 14. Intoxicação das plantas de azevém suscetível (S) e resistente (R1 e R2) após a
aplicação de diferentes doses do herbicida glyphosate aos 21 DAA. O gráfico se encontra em
escala logarítma. Botucatu – SP. .............................................................................................. 46
Figura 15. Massa seca da parte aérea das plantas de azevém suscetível (S) e resistente (R1 e
R2) após a aplicação de diferentes doses do herbicida glyphosate aos 28 DAA. O gráfico se
encontra em escala logarítma. Botucatu – SP. .......................................................................... 47
Figura 16. Taxa de assimilação de CO2 (μmolCO2 m-2
s-1
) em plantas de azevém suscetível e
resistente após a aplicação do herbicida glyphosate ao 1, 3, 7 e 28 DAA. Testemunha
suscetível (TS), testemunha resistente (TR), população suscetível submetida a 720 g e.a. ha-1
(S2), população suscetível submetida a 1080 g e.a. ha-1
(S3), população resistente

X
submetida a 720 g e.a. ha-1
(R2), população resistente submetida a 1080 g e.a. ha-1
(R3). As
barras verticais indicam o intervalo de confiança. Botucatu – SP. ........................................... 56
Figura 17. Condutância estomática (mmolH2O m-2
s-1
) em plantas de azevém suscetível e
resistente após a aplicação do herbicida glyphosate ao 1, 3, 7 e 28 DAA. Testemunha
suscetível (TS), testemunha resistente (TR), população suscetível submetida a 720 g e.a. ha-1
(S2), população suscetível submetida a 1080 g e.a. ha-1
(S3), população resistente submetida a
720 g e.a. ha-1
(R2), população resistente submetida a 1080 g e.a. ha-1
(R3). As barras
verticais indicam o intervalo de confiança. Botucatu – SP. ..................................................... 58
Figura 18. Concentração interna de CO2 na câmara subestomática (μmol(CO2) mol-1
ar) em
plantas de azevém suscetível e resistente após a aplicação do herbicida glyphosate ao 1, 3, 7 e
28 DAA. Testemunha suscetível (TS), testemunha resistente (TR), população suscetível
submetida a 720 g e.a. ha-1
(S2), população suscetível submetida a 1080 g e.a. ha-1
(S3),
população resistente submetida a 720 g e.a. ha-1
(R2), população resistente submetida a 1080
g e.a. ha-1
(R3). As barras verticais indicam o intervalo de confiança. Botucatu – SP. ........... 59
Figura 19. Taxa de transpiração (μmol(H2O) m-2
s-1
) em plantas de azevém suscetível e
resistente após a aplicação do herbicida glyphosate ao 1, 3, 7 e 28 DAA. Testemunha
suscetível (TS), testemunha resistente (TR), população suscetível submetida a 720 g e.a. ha-1
(S2), população suscetível submetida a 1080 g e.a. ha-1
(S3), população resistente submetida a
720 g e.a. ha-1
(R2), população resistente submetida a 1080 g e.a. ha-1
(R3). As barras
verticais indicam o intervalo de confiança. Botucatu – SP. ..................................................... 61
Figura 20. Eficiência do uso da água (μmol CO₂ mmol H₂O-1) em plantas de azevém
suscetível e resistente após a aplicação do herbicida glyphosate ao 1, 3, 7 e 28 DAA.
Testemunha suscetível (TS), testemunha resistente (TR), população suscetível submetida a
720 g e.a. ha-1
(S2), população suscetível submetida a 1080 g e.a. ha-1
(S3), população
resistente submetida a 720 g e.a. ha-1
(R2), população resistente submetida a 1080 g e.a. ha-1
(R3). As barras verticais indicam o intervalo de confiança. Botucatu – SP. ............................ 62
Figura 21. Eficiência de carboxilação (A/Ci) em plantas de azevém suscetível e resistente
após a aplicação do herbicida glyphosate ao 1, 3, 7 e 28 DAA. Testemunha suscetível (TS),
testemunha resistente (TR), população suscetível submetida a 720 g e.a. ha-1
(S2), população
suscetível submetida a 1080 g e.a. ha-1
(S3), população resistente submetida a 720 g e.a. ha-1
(R2), população resistente submetida a 1080 g e.a. ha-1
(R3). As barras verticais indicam o
intervalo de confiança. Botucatu – SP. ..................................................................................... 64
Figura 22. Taxa de assimilação de CO2 (μmolCO2 m-2
s-1
) em plantas de azevém suscetível e
resistente após a aplicação do herbicida glyphosate ao 1, 7 e 28 DAA. Testemunha suscetível
(TS), testemunha resistente (TR), população suscetível submetida a 720 g e.a. ha-1
(S2),
população suscetível submetida a 1080 g e.a. ha-1
(S3), população resistente submetida a 720
g e.a. ha-1
(R2), população resistente submetida a 1080 g e.a. ha-1
(R3). As barras verticais
indicam o intervalo de confiança. Botucatu – SP. .................................................................... 66
Figura 23. Condutância estomática (mmolH2O m-2
s-1
) em plantas de azevém suscetível e
resistente após a aplicação do herbicida glyphosate ao 1, 7 e 28 DAA. Testemunha suscetível
(TS), testemunha resistente (TR), população suscetível submetida a 720 g e.a. ha-1
(S2),
população suscetível submetida a 1080 g e.a. ha-1
(S3), população resistente

XI
submetida a 720 g e.a. ha-1
(R2), população resistente submetida a 1080 g e.a. ha-1
(R3). As
barras verticais indicam o intervalo de confiança. Botucatu – SP. ........................................... 67
Figura 24. Concentração interna de CO2 na câmara subestomática (μmol(CO2) mol-1
ar) em
plantas de azevém suscetível e resistente após a aplicação do herbicida glyphosate ao 1, 7 e 28
DAA. Testemunha suscetível (TS), testemunha resistente (TR), população suscetível
submetida a 720 g e.a. ha-1
(S2), população suscetível submetida a 1080 g e.a. ha-1
(S3),
população resistente submetida a 720 g e.a. ha-1
(R2), população resistente submetida a 1080
g e.a. ha-1
(R3). As barras verticais indicam o intervalo de confiança. Botucatu – SP. ........... 68
Figura 25. Taxa de transpiração (mmolH2O m-2
s-1
) em plantas de azevém suscetível e
resistente após a aplicação do herbicida glyphosate ao 1, 7 e 28 DAA. Testemunha suscetível
(TS), testemunha resistente (TR), população suscetível submetida a 720 g e.a. ha-1
(S2),
população suscetível submetida a 1080 g e.a. ha-1
(S3), população resistente submetida a 720
g e.a. ha-1
(R2), população resistente submetida a 1080 g e.a. ha-1
(R3). As barras verticais
indicam o intervalo de confiança. Botucatu – SP. .................................................................... 69
Figura 26. Eficiência do uso da água (μmol CO₂ mmol H₂O-1) em plantas de azevém
suscetível e resistente após a aplicação do herbicida glyphosate ao 1, 7 e 28 DAA.
Testemunha suscetível (TS), testemunha resistente (TR), população suscetível submetida a
720 g e.a. ha-1
(S2), população suscetível submetida a 1080 g e.a. ha-1
(S3), população
resistente submetida a 720 g e.a. ha-1
(R2), população resistente submetida a 1080 g e.a. ha-1
(R3). As barras verticais indicam o intervalo de confiança. Botucatu – SP. ............................ 70
Figura 27. Eficiência de carboxilação (A/Ci) em plantas de azevém suscetível e resistente
após a aplicação do herbicida glyphosate ao 1, 7 e 28 DAA. Testemunha suscetível (TS),
testemunha resistente (TR), população suscetível submetida a 720 g e.a. ha-1
(S2), população
suscetível submetida a 1080 g e.a. ha-1
(S3), população resistente submetida a 720 g e.a. ha-1
(R2), população resistente submetida a 1080 g e.a. ha-1
(R3). As barras verticais indicam o
intervalo de confiança. Botucatu – SP. ..................................................................................... 71
Figura 28. Níveis de glyphosate (μg g-1
) em plantas de azevém S, R1 e R2 aos 5, 11 e 28
DAA. Botucatu – SP. ................................................................................................................ 75
Figura 29. Níveis de Ampa (μg g-1
) em plantas de azevém S, R1 e R2 aos 5, 11 e 28 DAA.
Botucatu – SP... ........................................................................................................................ 77
Figura 30. Níveis de ácido chiquímico (μg g-1
) em plantas de azevém S, R1 e R2 aos 5, 11 e
28 DAA. Botucatu – SP. ........................................................................................................... 79
Figura 31. Níveis de ácido quínico (μg g-1
) em plantas de azevém S, R1 e R2 aos 5, 11 e 28
DAA. Botucatu – SP. ................................................................................................................ 81
Figura 32. Níveis de shiquimato-3-fosfato (μg g-1
) em plantas de azevém S, R1 e R2 aos 5, 11
e 28 DAA. Botucatu – SP. ........................................................................................................ 83
Figura 33. Níveis de Fenilalanina (μg g-1
) em plantas de azevém S, R1 e R2 aos 5, 11 e 28
DAA. Botucatu – SP. ................................................................................................................ 85

XII
Figura 34. Níveis de Tirosina (μg g-1
) em plantas de azevém S, R1 e R2 aos 5, 11 e 28 DAA.
Botucatu – SP..... ...................................................................................................................... 87
Figura 35. Níveis de Triptofano (μg g-1
) em plantas de azevém S, R1 e R2 aos 5, 11 e 28
DAA. Botucatu – SP. ................................................................................................................ 89
Figura 36. Níveis de Ácido Ferúlico (μg g-1
) em plantas de azevém S, R1 e R2 aos 5, 11 e 28
DAA. Botucatu – SP. ................................................................................................................ 91
Figura 37. Níveis de Ácido Coumárico (μg g-1
) em plantas de azevém S, R1 e R2 aos 5, 11 e
28 DAA. Botucatu – SP. ........................................................................................................... 92
Figura 38. Níveis de Ácido Cafeico (μg g-1
) em plantas de azevém S, R1 e R2 aos 5, 11 e 28
DAA. Botucatu – SP. ................................................................................................................ 95

1
1. RESUMO
No Brasil, o azevém (Lolium multiflorum) foi identificado como
resistente ao glyphosate se tornando um grande problema em determinadas lavouras. Dessa
forma, entender o comportamento a nível bioquímico e fisiológico desta planta daninha são
ferramentas que auxiliam num manejo eficiente. Com isso, o objetivo deste trabalho foi
comparar o perfil bioquímico e fisiológico de populações de azevém suscetíveis e resistentes
ao herbicida glyphosate aplicação do mesmo. Foram realizados quatro estudos em casa-de-
vegetação com delineamento experimental inteiramente casualizados com quatro repetições
sendo semeadas três populações de azevém (Lolium multiflorum) consideradas como
suscetível (S), com suspeita de resistência (R1) e resistente (R2) ao herbicida glyphosate. No
primeiro estudo foi obtido o controle aos 21 dias após a aplicação (DAA) e quantificada a
massa seca aos 28 DAA das três populações. Os tratamentos foram constituídos da aplicação
do herbicida glyphosate composto pelas doses: 0, 135, 270, 540, 1080, 2160, 4320, 8640 g
e.a. ha-1
. O segundo estudo teve como objetivo determinar a atividade da enzima fenilalanina
amônia liase (PAL) nas diferentes populações as 12, 24, 48 e 72 horas após a aplicação
(HAA). Os tratamentos foram compostos de duas doses (720 g e.a. ha-1
e 1080 g e.a. ha-1
)
mais uma testemunha sem aplicação. No terceiro estudo foram realizadas avaliações da
fotossíntese nas três populações ao 1, 3, 7 e 28 DAA. As variáveis analisadas foram: taxa de
assimilação líquida de CO2, condutância estomática, concentração interna de CO2,
transpiração, eficiência do uso da água e eficiência instantânea de carboxilação. Os
tratamentos foram compostos de duas doses (720 g e.a. ha-1
e 1080 g e.a. ha-1
) mais uma
testemunha sem aplicação. O quarto estudo teve o objetivo de quantificar compostos alterados
da rota do ácido chiquímico. Para isso, foram utilizados os mesmos tratamentos do primeiro
estudo e realizadas coletas das folhas aos 5, 11 e 28 DAA. Os compostos analisados foram:
glyphosate, AMPA (ácido aminometilfosfônico), ácido chiquímico, ácido quínico,
shiquimato-3-fosfato, os aminoácidos aromáticos fenilalanina, tirosina e triptofano, ácido
ferúlico, ácido coumárico e ácido cafeico. Na população considerada resistente, a atividade da
enzima fenilalanina amônia liase manteve-se alta após a aplicação do glyphosate. Todas as
variáveis fisiológicas foram afetadas após a aplicação do glyphosate nas três populações,
porém, R2 foi capaz de se recuperar apresentando valores semelhantes à testemunha. Os
níveis de ácido chiquímico e quínico apresentaram padrões semelhantes onde houve aumento
para as populações suscetíveis com o aumento da dose do herbicida enquanto que para a

2
resistente os valores se mantiveram semelhantes. Ocorreu aumento dos níveis de shiquimato-
3-fosfato para a população R2 se mantendo constante para as suscetíveis. Houve redução dos
aminoácidos aromáticos com a aplicação do glyphosate para as populações suscetíveis.
Palavras-chave: Resistência a herbicidas, ácido chiquímico, aminoácidos aromáticos,
fotossíntese, fenilalanina amônia liase, ácidos fenólicos, metabolismo secundário.

3
GLYPHOSATE INFLUENCE IN THE BIOCHEMICAL AND PHYSIOLOGICAL
PROFILE OF SUSCEPTIBLE AND RESISTANT RYEGRASS (Lolium multiflorum)
POPULATIONS TO HERBICIDE. Botucatu, 2016, 117p. Tese (Doutorado em
Agronomia/Agricultura) – Faculdade de Ciências Agronômicas, Universidade Estadual
Paulista.
Author: Gilmar José Picoli Junior
Adviser: Caio Antonio Carbonari
Co-adviser: Edivaldo Domingues Velini
2. SUMMARY
In Brazil, ryegrass (Lolium multiflorum) was identified as resistant to
glyphosate becoming a major problem in certain crops. Thus, understanding the behavior of
the biochemical and physiological level of this weed are tools that help in efficient
management. Thus, the aim of this study was to compare the biochemical and physiological
profile of ryegrass populations susceptible and resistant to glyphosate after spray it. Four
studies were carried out in greenhouse with experimental design completely randomized with
four replications being seeded three populations of ryegrass (Lolium multiflorum) considered
as susceptible (S), suspected of having resistance (R1) and resistant (R2) to the herbicide
glyphosate. In the first study was measured the control at 21 days after application (DAA) and
at 28 DAA, the dry mass the three populations. The treatments consisted of application of the
glyphosate composed of doses: 0, 135, 270, 540, 1080, 2160, 4320, 8640 g a.i. ha-1
. The
second study aimed to determine the phenylalanine ammonia lyase (PAL) activity in different
populations at 12, 24, 48 and 72 hours after application (HAA). The treatments consisted of
two doses (720 g a.i. ha-1
and 1080 g a.i. ha-1
) plus a control without application. In the third
study were carried out photosynthesis assessments at three populations at 1, 3, 7 and 28 DAA.
The variables analyzed were: CO2 net assimilation rate, stomatal conductance, CO2 internal
concentration, transpiration, water use efficiency and instantaneous carboxylation efficiency.
The treatments consisted of two doses (720 g a.i. ha-1
and 1080 g a.i. ha-1
) plus a control
without application. The fourth study aimed to quantify altered compounds of the shikimic
acid pathway. For this, the same treatments of the first experiment were used and made
collections of leaves at 5, 11, 28 DAA. The compounds analyzed were: glyphosate, AMPA
(aminomethylphosphonic acid), shikimic acid, quinic acid, shikimate 3-phosphate, the

4
aromatic amino acids phenylalanine, tyrosine and tryptophan, ferulic acid, coumaric acid and
caffeic acid. The phenylalanine ammonia lyase enzyme was not influenced by glyphosate in
resitant population. All physiological variables were affected after the application of
glyphosate at the three populations, but R2 was able to recover with values similar to the
control. The shikimic and quinic acid levels showed similar patterns where, there was an
increase for susceptible populations with increasing doses of the herbicide while in resistant,
the values remained similar. There was increase in levels of shikimate-3-phosphate to the R2
population, remaining constant for susceptible. There was a reduction of the aromatic amino
acids with the application of glyphosate for the susceptible populations.
Key-words: Herbicide resistance, shikimic acid, aromatic amino acids, photosynthesis,
phenylalanine ammonia lyase, phenolic acids, secondary metabolism.

5
3. INTRODUÇÃO
Pode-se dizer que existe uma relação direta entre o aumento da
produtividade das culturas e o aumento no uso de herbicidas, porém mesmo com o controle
químico, a infestação das plantas daninhas não tem diminuído. Na atualidade, os agricultores
depositam confiança excessiva no controle químico das plantas daninhas o que se deve,
principalmente, ao fato de que o controle químico tem sido muito eficiente e possui custo
atrativo, estando prontamente disponível e profissionalmente desenvolvido. Porém o uso
excessivo e indiscriminado dos mesmos ao longo dos anos propiciou a seleção de plantas
daninhas resistentes.
A resistência de plantas daninhas a herbicidas pode ser definida como a
capacidade adquirida de uma planta em sobreviver à dose de registro do herbicida que sob
condições normais, controlam os demais indivíduos da população (CHRISTOFFOLETI;
LÓPES-OVEJERO, 2008). Já uma planta suscetível sofre alterações em seu crescimento e
desenvolvimento levando-a a morte após a exposição do produto, ou seja, há variações no
perfil metabólico dessas plantas quando as mesmas são tratadas com herbicidas como o
glyphosate.
O glyphosate tem como mecanismo de ação a inibição da enzima
EPSPs, bloqueando a via do ácido chiquímico que produz aminoácidos aromáticos dos quais
não são somente utilizados para a produção de proteínas em plantas, mas também como
precursores de um grande número de metabólitos secundários (pigmentos, flavonóides,
auxinas, fitoalexinas, lignina e taninos) (VELINI et al., 2009a). Além disso, rotas
intermediárias podem servir como substrato para outras vias metabólicas, incluindo a

6
biossíntese de ácido quínico e produtos derivados, como o ácido clorogênico (HERMAN;
WEAVER, 1999). Sendo assim, as plantas submetidas ao glyphosate podem ter a síntese
desses compostos também alterada.
Herbicidas como o glyphosate pode afetar indiretamente a fotossíntese
através da inibição da biossíntese de carotenoides, clorofila, ácidos graxos (OLESEN;
CEDERGREEN, 2010; GOMES et al., 2014). Como um inibidor competitivo, o glyphosate
bloqueia a via do ácido chiquímico inibindo a biossíntese de metabólitos secundários nas
plantas, incluindo compostos relacionados com a fotossíntese como as quinonas (DEWICK,
1998).
Dentre as espécies em que foi registrada a resistência ao herbicida
glyphosate, tem-se o azevém (Lolium multiflorum), uma planta daninha utilizada como
forrageira e também no fornecimento de palha para o sistema de plantio direto, mas que se
constitui um grande problema para culturas como o trigo e pomares especialmente nos locais
de clima temperado como o Rio Grande do Sul e Santa Catarina. O aumento da presença desta
espécie nas lavouras comerciais associado à baixa perspectiva de lançamento de novas
moléculas herbicidas especificamente para o controle do azevém representa um grande
impacto econômico e técnico na agricultura brasileira (DIONISIO et al., 2013).
Diversos trabalhos têm sido feitos com o intuito de entender o
metabolismo das plantas após a aplicação de herbicidas. O conhecimento a nível bioquímico e
fisiológico das plantas daninhas, neste caso o azevém, é importante para um manejo eficiente,
pois pode ajudar na compreensão dos mecanismos de resistência ao glyphosate, assim como
compreender a intensidade dos efeitos nos biótipos suscetíveis e resistentes. Há poucas
explanações sobre os efeitos metabólicos e fisiológicos de plantas sensíveis e resistentes após
a inibição da EPSPs pela aplicação do herbicida glyphosate, sendo que a morte das plantas
não está apenas relacionada com o bloqueio da rota de biossíntese dos aminoácidos
aromáticos, mas também à falha na produção de um grande número de compostos
secundários, ao desregulamento no fluxo de carbono na planta e a redução da síntese de
proteínas.
Com isso, o objetivo geral do trabalho, foi comparar o perfil bioquímico
e fisiológico de populações de azevém suscetíveis e resistentes ao herbicida glyphosate após a
aplicação do mesmo. Como objetivos específicos: quantificar compostos alterados nas plantas
suscetíveis e resistentes; avaliar os níveis de intoxicação, alterações na fotossíntese e acúmulo

7
de matéria seca e investigar a influência do herbicida glyphosate sobre a enzima fenilalanina
amônia liase (PAL) em plantas resistentes.

8
4. REVISÃO BIBLIOGRÁFICA
4.1 Azevém: morfologia, biologia e distribuição
O gênero Lolium pertence a família Poaceae sendo composto por oito
espécies: L. temulentum L., L. persicum Boiss e Hohen, L. remotum Schrank, L. loliaceum
Bory e Chaubard (autógamas), L. canariense Steud (reprodução intermediária), L. perenne L.,
L. rigidum Gaundin e L. multiflorum Lam. (alógamas). Estas espécies podem ser pequenas
com 10 cm de altura, mas também grandes podendo chegar a 150 cm. São nativas da Europa,
Ásia e norte da África sendo introduzidas nos mais diversos países em todo o mundo,
inclusive no Brasil. Uma forma simples de verificar se uma planta pertence a este gênero é a
presença de duas glumas (POLOK, 2007).
Dentre essas espécies, L. multiflorum destaca-se por ser uma espécie
importante como forrageira e também para fornecimento de palha para o sistema de plantio
direto sendo cultivada em diversas regiões de clima temperado (LUBBERSTEDT;
SCHEJBEL; BACH, 2003). Pertence a subfamília Pooideae e tribo Poeae (WHEELER;
JACOBS; WHALLEY, 2002). Tem sua origem no mediterrâneo e provavelmente chegou ao
Brasil em 1985 trazida por imigrantes italianos. Ao passar dos anos, esta espécie sofreu
efeitos da seleção natural formando uma população bem adaptada às condições ecológicas do
sul do Brasil, onde no estado do Rio Grande do Sul é umas das gramíneas hibernais mais
cultivadas devido à boa capacidade de rebrote, perenização por ressemeadura natural,
resistência ao pisoteio, pastoreio e excesso de umidade, além da facilidade de implantação
(GONÇALVES, 1979; FREITAS; OLIVEIRA; CARVALHO, 2003; CARVALHO, 2004;
CASSOL et al., 2011).

9
É uma planta de ciclo anual ou bianual, morfologicamente muito
variável podendo atingir até 1,20 m de altura. Ereta, densamente perfilhada, glabra, de
inverno e herbácea. Suas folhas são finas e laminadas e sua inflorescência é em forma de
espiga do tipo dística com 15 a 20 cm de comprimento possuindo espiguetas onde as flores
estão inseridas. Cada flor possui duas glumas, uma pálea e um lema que envolve a cariopse.
Apresenta fácil dispersão propagando-se apenas por sementes, possui fecundação cruzada,
sistema radicular altamente ramificado e denso com muitas raízes adventícias e fibrosas
(POLOK, 2007; LORENZI, 2008; AGOSTINETTO; VARGAS, 2009).
O azevém é pouco exigente quanto ao tipo de solo, persistindo em uma
ampla gama de textura. Além disso, é tolerante a solos ácidos e alcalinos (pH 5,0 a 7,8) se
desenvolvendo melhor na faixa de pH entre 5,5 e 7,5. Possui metabolismo do tipo C3 e a
temperatura ótima para o seu desenvolvimento está entre 20 a 25°C sendo adaptado a climas
frios e úmidos (CARAMBULA, 1998; HANNAWAY et al., 1999).
4.2 Azevém como planta daninha
Planta daninha se enquadra como “toda e qualquer planta que ocorre
onde não é desejada” (OLIVEIRA JR; CONSTANTIN; INOUE, 2011). Porém Blanco (1972)
define como “toda e qualquer planta que germine espontaneamente em áreas de interesse
humano e que, de alguma forma, interfira prejudicialmente nas atividades agropecuárias do
homem”. Segundo Oerke (2006), o potencial de perda de produtividade devido à presença das
plantas daninhas é mais elevado do que outras pragas e doenças. Em média, 30 a 40% das
perdas em produtividade deve-se a interferência das infestantes (LORENZI, 2008).
Embora o azevém possa ser utilizado com uma espécie forrageira e
também para fornecimento de palha para o sistema de plantio direto, ele também pode se
constituir em uma planta daninha em culturas como o milho ou o trigo principalmente
(ROMAN et al., 2004), além de pomares especialmente no Rio Grande do Sul (VARGAS et
al., 2005). Dessa forma, o mesmo atua competindo com as culturas pelos recursos do meio
prejudicando assim suas produtividades. No caso do trigo, o grau de competição com o
azevém é intenso devido ambas as espécies serem morfologicamente e fisiologicamente
semelhantes e segundo Silva e Durigan (2006), quanto mais próximos forem essas
características, maior a competição pelos recursos do meio. Ainda, estudos demonstraram que

10
uma planta de azevém por metro quadrado, pode diminuir a produtividade da cultura do trigo
em 7,5 kg ha-1
(KISSMAN, 2007).
O florescimento e a frutificação das plantas de azevém ocorre no final
da primavera e segundo Piana et al. (1986), após a maturação fisiológica ocorre a abscisão das
sementes que, quando não colhidas, caem no solo permanecendo dormentes até o final do
verão quando iniciam a germinação. Esta característica de ressemeadura natural é útil para os
pecuaristas, porém pode ser ruim para os agricultores, pois ocasiona dificuldades no controle
desta espécie devido à mesma estar em diferentes estádios de desenvolvimento. Além disso,
são fontes de permanência em infestações futuras quando da utilização destas na prática de
rotação de cultura com cereais de inverno como cevada, centeio, trigo e triticale (ROMAN et
al., 2004).
O azevém é uma infestante da região sul do Brasil onde a época de
maior incidência ocorre entre as estações de outono-inverno não se constituindo em grande
problema nas culturas de verão. A dessecação ou o controle dessa espécie no sistema de
semeadura direta ou em pomares é feito de um modo geral com herbicidas não seletivos em
diferentes estádios de desenvolvimento e o herbicida glyphosate é o mais usado para este fim
sendo ele utilizado há longo tempo e repetidamente (CHRISTOFFOLETI; LÓPEZ-
OVEJERO, 2003).
4.3 Glyphosate: importância, características químicas e modo de ação
O glyphosate é o herbicida mais utilizado em todo o mundo e o seu
mercado tem crescido continuamente graças ao controle eficaz que exerce sobre as plantas
daninhas, aliado às características positivas quanto a aspectos toxicológicos, ecotoxicológicos,
facilidade de manuseio, custo relativamente baixo, aumento de produtividade, dentre outros
(GALLI, 2009; VELINI et al., 2009b). É um herbicida sistêmico, aplicado em pós-
emergência das plantas daninhas apresentando um amplo espectro de ação podendo controlar
76 das 78 infestantes mais agressivas (FRANZ, 1985; QUINN, 1993) e sendo registrado para
mais de vinte culturas (RODRIGUES; ALMEIDA, 2011).
Este herbicida é sintetizado a partir de um hidrogênio amínico do
aminoácido glicina por um radical éster fosfônico, o N-(fosfonometil) glicina. É muito pouco
solúvel em solventes orgânicos comuns e bastante solúvel em água. Além disso, possui um
coeficiente de adsorção elevado no solo e um coeficiente de partição octanol/água muito

11
baixo sugerindo que o glyphosate tem baixa mobilidade com pouca tendência para lixiviação
no solo (LINDERS et al., 1994; AMARANTE JR. et al., 2002). Apesar de ser bastante solúvel
em água (contribui em termos toxicológicos sendo rapidamente excretados pela urina e de
difícil acumulação em organismos vivos), há poucos riscos de contaminação de águas
subterrâneas devido à forte adsorção e rápida dissociação nos solos (AMARANTE JR. et al.,
2002; GIMSING; BORGGAARD; BANG, 2004; VELINI et al., 2012).
A degradação do glyphosate no solo pode ocorrer através de duas rotas
(Figura 1). A primeira consiste na transformação do glyphosate em sarcosina pela ação da
bactéria Agrobacterium radiobacter ou da Enterobacter aerogenes (enzima C-P liase). A
sarcosina entra no metabolismo destes microorganismos e em outros, degradando-se (DICK;
QUINN, 1995; FRANZ; MAO; SIKORSKI, 1997). Esta rota é pouco citada na literatura. A
segunda rota consiste na transformação do glyphosate em ácido aminometilfosfônico
(AMPA) sendo este mais persistente no solo do que o glyphosate, pois sua degradação é mais
lenta (COX, 1998).
Figura 1. Rotas de decomposição microbiológica do glyphosate. Fonte: Amarante Jr. et al.,
2002.

12
O glyphosate apresenta pouca volatilidade devido à sua baixa pressão
de vapor (7.5x10-8
mmHg) tendo como ponto de fusão 189,9°C. Devido ao grupamento ácido
fosfônico com uma ligação C-P bastante estável, este herbicida é muito resistente à hidrólise
química e também à fotodegradação em pH 5, 7 e 9. Sua meia-vida gira em torno de 35 dias
apresentando bastante estável na presença de luz, inclusive a temperaturas superiores a 60°C
(FRANZ; MAO; SIKORSKI, 1997).
Segundo Silva et al. (2005), a absorção de herbicidas envolve uma
rápida penetração inicial sobre a cutícula e em seguida ocorre absorção simplástica no qual
esse processo depende de diversos fatores tais como: idade da planta, condições ambientais,
entre outros. Logo que o herbicida é absorvido, deve ser translocado e atingir o sítio de ação
para que o mesmo atue prejudicando assim, o desenvolvimento da planta, ou seja, a
translocação é um processo vital para a eficácia do produto (WANAMARTA; PENNER,
1989; SATICHIVI et al., 2000). O glyphosate é lentamente absorvido pela cutícula onde há a
necessidade de um período de aproximadamente 6 horas sem chuva dependendo da
formulação utilizada para haver um controle satisfatório nas plantas suscetíveis
(RODRIGUES; ALMEIDA, 2011).
A penetração do glyphosate é mediada por proteínas de transporte de
fosfato presente na membrana (MERVOSH; BALKE, 1991). Proteínas transportadoras de
fosfato facilitaram a absorção de glyphosate nas membranas de Vicia faba (DENIS;
DELROT, 1993). Este herbicida se movimenta no floema seguindo a rota dos produtos
fotossintéticos partindo das folhas fotossinteticamente ativas para as partes em crescimento
(dreno), para a manutenção do metabolismo ou na formação de produtos de reserva como os
tubérculos (HETHERINGTON et al., 1999).
O mecanismo de ação do glyphosate é bastante singular porque ele é o
único herbicida capaz de inibir especificamente a enzima 5-enolpiruvilchiquimato 3-fosfato
sintase (EPSPs), que catalisa a reação de ligação do chiquimato-3-fosfato (S3P) com o
fosfoenolpiruvato (PEP), para formar 5-enolpiruvilchiquimato 3-fosfato (JAWORSKI, 1972;
ZABLOTOWICZ; REDDY, 2004). O glyphosate bloqueia então a via do ácido chiquímico,
que é a principal rota de produção dos aminoácidos aromáticos (fenilalanina, tirosina e
triptofano), afetando também o fluxo de carbono na planta, sendo que sob condições normais,
20% do carbono fixado pelas plantas são direcionados para esta rota (HASLAM, 1993). Com
a inibição da enzima EPSPs pelo glyphosate, é observado um grande acúmulo de ácido

13
chiquímico nas plantas, representando um forte dreno de carbono, desta forma, reduzindo a
fotossíntese (GOMES, 2011).
A via do ácido chiquímico está presente em bactérias, fungos, plantas e
parasitas do filo Apycomplexa e sintetiza metabólicos importantes para a vida desses
organismos (ARCURI et al., 2004). Consiste de sete enzimas que catalisam a conversão
sequencial de eritrose-4-fosfato (E4P) e fosfoenolpiruvato (PEP) em corismato, um precursor
dos aminoácidos aromáticos (CAMPBELL et al., 2004). Todos os intermediários da via
também podem ser considerados compostos com pontos de ramificações que podem servir
como substratos para outras vias metabólicas, além disso, a organização molecular e estrutural
das enzimas desta rota varia consideravelmente entre os grupos de microrganismos
(BENTLEY; HASLAM, 1990). A importância destas enzimas no metabolismo de organismos
mais simples e a sua ausência em mamíferos faz esta via um alvo potencial para o
desenvolvimento de herbicidas, neste caso o glyphosate (HERMAN; WEAVER, 1999).
A rota de formação do corismato (via do ácido chiquímico) está
representada na Figura 2. Inicialmente, ocorre uma condensação de dois metabólitos
fosforilados: fosfoenolpiruvato (PEP) e eritrose-4-fosfato (E4P). Através da enzima 3-deoxi-
d-arabino-heptulosonato 7-fosfato sintase (DAHPs), ocorre a formação do composto 3-deoxi-
d-arabino-heptulosonato 7-fosfato. A atividade de várias enzimas da via do chiquimato pode
mudar durante o crescimento e desenvolvimento normal das plantas. Os níveis de DAHPs
variaram durante o ciclo de desenvolvimento de células de cenoura aumentando durante a
formação de brotos bem como na germinação e formação de plântulas (WEAVER;
HERRMANN, 1997).
No segundo passo, ocorre a ação da enzima 3-desidroquinato sintase
(DHQs) formando o composto 3-desidroquinato (DHQ), resultando desta reação, a eliminação
do fosfato. Logo após, a desidroquinase desidratase catalisa a reação formando o metabólito
3-desidrochiquimato (DHS) e em seguida, a enzima chiquimato desidrogenase age formando
o chiquimato. No quinto passo, a chiquimato quinase catalisa a fosforilação do chiquimato
produzindo chiquimato-3-fosfato (S3P). Na próxima etapa, uma segunda PEP entra na rota, é
condensada com o S3P produzindo o composto 5-enolpiruvilchimato 3-fosfato (EPSP) e
fosfato inorgânico. Esta reação reversível é catalisada pela enzima EPSP sintase no qual o
glyphosate atua impedindo sua ação. Na última reação, ocorre a formação do corismato pela
ação da enzima corismato sintase onde a mesma elimina o fosfato da EPSP (ANDERSON;
JOHNSON, 1990; DEWICK, 1998; HERMAN; WEAVER, 1999).

14
Figura 2. Rota do ácido chiquímico. Fonte: Maeda e Dudareva, 2012.
A enzima que o glyphosate bloqueia é codificada no núcleo
desempenhando sua função no cloroplasto (WEAVER; HERRMANN, 1997; STAUFFER et
al., 2001), onde catalisa a ligação da reação dos compostos chiquimato-3-fosfato e o
fosfoenolpiruvato produzindo o enolpiruvilchiquimato-3-fosfato e fosfato inorgânico
(PETERSON et al., 1996). A mesma reage inicialmente com o S3P e depois com o PEP. O
glyphosate atua exatamente nesse ponto impedindo a ação da EPSPs prejudicando a formação
do corismato, além disso, a afinidade deste herbicida pelo complexo EPSPs-S3P, é setenta e
cinco vezes maior que a do PEP e sua velocidade de dissociação do sítio de ação é duas mil
vezes menor que a do PEP, ou seja, este herbicida é mais eficiente em se ligar e continuar
preso na reação (REAM et al., 1992). Outro fato que deve ser levado em consideração é que
as enzimas EPSPs podem diferir quanto à afinidade com o glyphosate de acordo com a
espécie envolvida. Fedtke e Duke (2004) encontraram valores diferentes em Pisium sativum e
em microorganismos.
O corismato, produto final da via do ácido chiquímico, é convertido por
cinco enzimas em prefenato, antranilato, aminodeoxicorismato, isocorismato e p-
hidroxibenzoato. Estes metabólitos compreendem os primeiros intermediários na biossíntese
de fenilalanina, tirosina, triptofano, folato, menaquinona, ubiquinona (Figura 3)
(DOSSELAERE; VANDERLEYDEN, 2001).

15
Figura 3. Corismato em destaque como precursor do (1) prefenato, (2) antranilato, (3)
aminodeoxicorismato, (4) p-hidroxibenzoato, (5) isocorismato. As enzimas
envolvidas são: (1) corismato mutase, (2) antranilato sintase, (3)
aminodeoxicorismato sintase, (4), corismato piruvato liase, (5) isocorismato
sintase. Fonte: Dosselaere e Vanderleyden, 2001.
De cada um dos aminoácidos aromáticos, são obtidos diversas
famílias de compostos envolvidos na regulação do crescimento ou na defesa da planta (Figura
4) tais como os taninos, antocianinas, ácido indol acético (AIA), lignina, entre outros que são
importantes para o desenvolvimento das mesmas, sendo assim, com a aplicação do
glyphosate, a formação desses produtos pode ser prejudicada (AMRHEIN et al., 1980;
ANDERSON; JOHNSON, 1990; HERMAN, 1995, VELINI et al., 2009a).

16
Figura 4. Aminoácidos aromáticos e alguns compostos fenólicos. Fonte: Velini et al., 2012.
Os taninos são compostos fenólicos que podem afetar o crescimento de
insetos, animais herbívoros e microrganismos através da precipitação de enzimas e proteínas e
são classificados em hidrolisáveis e condensados (BRYCE et al., 1993; SALMINEN et al.,
2001). Os taninos hidrolisáveis são formados a partir do chiquimato, já os condensados são
formados pela polimerização de unidade de flavonoides (OSSIPOV et al., 2003; MONTEIRO
et al., 2005; GARCÍA; CARRIL, 2009). Com o bloqueio da EPSPs causada pelo herbicida
glyphosate, ocorre um acúmulo de ácido chiquímico, um precursor dos taninos hidrolisáveis e

17
reduz a disponibilidade de fenilalanina, precursor dos taninos condensados. Nesse sentido,
não há como tirar conclusões gerais sobre o efeito do glyphosate sobre os taninos, os mesmos
devem ser analisados separadamente (VELINI, et al., 2009a).
A rota do ácido chiquímico é responsável pela maioria dos compostos
fenólicos nas plantas e a maioria desses compostos derivam do aminoácido aromático
fenilalanina. As antocianinas são flavonoides pigmentados responsáveis pela maioria das
cores das flores e frutos. Por isso são importantes na polinização e na dispersão de sementes
(GARCÍA; CARRIL, 2009).
O AIA é um importante regulador de crescimento e tem como um dos
seus precursores o aminoácido triptofano. Dentre suas várias funções, tem-se a elongação
celular, crescimento de caules e coleóptilos, dominância apical, formação de raízes laterais e
adventícias e produção de xilema (TAIZ; ZEIGER, 2006). A interferência de glyphosate na
síntese de AIA se dá quando se realiza a aplicação do glyphosate como maturador em cana-
de-açúcar, com quebra da dominância apical (morte ou não da gema apical) e brotação das
gemas laterais (VELINI et al., 2009a).
A lignina é um polímero altamente ramificado de fenilpropanóides.
Depois da celulose, é a substância orgânica mais abundante nas plantas. Encontra-se
covalentemente unida à celulose e a outros polissacarídeos da parece celular. É insolúvel em
água e na maioria dos solventes orgânicos sendo muito difícil de remover sem degradar.
Desempenha um papel estrutural fundamental, pois sua natureza química é a base da dureza
mecânica e rigidez que se manifesta nos troncos das árvores e caules lignificados. Ocorre na
parede celular de vários tecidos de suporte e de transporte, traqueídes e nos vasos do xilema.
Deposita-se principalmente na parede secundária, fortalece os caules e tecidos vasculares
permitindo o crescimento vertical e a condução de água e minerais através do xilema
(CABANÉ et al., 2004; GARCÍA; CARRIL, 2009).
Foram citados alguns exemplos de compostos que fazem parte da via do
ácido chiquímico no qual o herbicida glyphosate atua com o intuito de evidenciar a
importância da mesma. Segundo Velini et al. (2009a), os eventos pós inibição da EPSPs não
estão claramente estabelecidos e a morte das plantas pode não estar relacionados apenas com
o bloqueio dos aminoácidos aromáticos, pois a suplementação com esses aminoácidos não é
suficiente para reverter o processo. A morte das plantas pode estar relacionada à falha na
produção de vários compostos que derivam da rota; redução da síntese proteica e à
desregulação do fluxo de carbono.

18
4.4 Efeitos do glyphosate na fotossíntese
A fotossíntese é o principal processo bioquímico que ocorre em
organismos autotróficos e é conhecida por ser afetada por diversos fatores bióticos e
abióticos. Dentre os fatores bióticos pode-se citar os herbicidas. Alguns herbicidas afetam
diretamente a fotossíntese interrompendo o transporte de elétrons como o paraquat, um
inibidor do fotossistema I. Outros herbicidas como o glyphosate, afetam indiretamente através
da inibição da biossíntese de carotenoides, clorofila, ácidos graxos, etc. (OLESEN;
CEDERGREEN, 2010; GOMES et al., 2014). Como um inibidor competitivo, o glyphosate
bloqueia a via do ácido chiquímico inibindo a biossíntese de metabólitos secundários nas
plantas, incluindo compostos relacionados com a fotossíntese como as quinonas (DEWICK,
1998). No entanto, não está claro como o glyphosate leva às plantas a morte e hipóteses tais
como o esgotamento dos estoques de proteínas e drenagem de carbono a partir de outras vias
podem ser colocadas em questão (DUKE; POWLES, 2008). Vários trabalhos têm
demonstrado redução na taxa fotossintética após a aplicação do herbicida glyphosate
(MATEOS-NARANJO et al., 2009; YANNICCARU, et al., 2012; ZOBIOLE et al., 2012).
A clorofila e os carotenoides são os principais pigmentos relacionados à
eficiência fotossintética sendo responsáveis em absorver luz, fornecendo energia. O primeiro
passo da fotossíntese ocorre entre as membranas dos tilacóides através da excitação da luz
pelos pigmentos fotossintéticos. A partir daí, inicia o transporte de elétrons (TAIZ; ZEIGER,
2006). Alguns estudos têm demonstrado uma redução no teor de clorofila nas plantas após a
aplicação do glyphosate (ZOBIOLE et al., 2011; HUANG et al., 2012; KASPARY et al.,
2014). De acordo com Cakmak et al. (2009), o herbicida glyphosate pode impedir a
biossíntese de clorofila através da diminuição das concentrações de magnésio, que é
constituinte da mesma nas folhas reduzindo assim o teor de clorofila e a taxa fotossintética
(ZOBIOLE et al., 2012).
Reddy, Rimando e Duke, (2004) reportaram que os efeitos negativos do
herbicida glyphosate na biossíntese de clorofila depende da taxa de degradação do glyphosate
em AMPA, sendo ele, o responsável pelos efeitos negativos na biossíntese de clorofila,
porém, é um mecanismo pouco conhecido. No caso de plantas sem o contato com AMPA e
desprovida da glyphosate oxido-redutase (GOX) (enzima responsável pela degradação do
glyphosate em AMPA), o teor de clorofila pode ser reduzido pela ação do glyphosate pela
privação da assimilação de N que irá levar uma redução da produção de glutamato afetando a

19
biossíntese de clorofila e ALA (ácido aminolevulínico) (ZOBIOLE et al., 2011; SERRA et al.,
2013).
O glyphosate e o AMPA podem também afetar a fotossíntese
modificando o metabolismo do carbono nas plantas, assim, após a aplicação dos mesmos, a
capacidade de assimilação de CO2 é reduzida levando a um aumento da concentração
intracelular de CO2 reduzindo também a condutância estomática (MATEOS-NARANJO et
al., 2009; DING et al., 2011). Adicionalmente, a atividade da enzima Ribulose 1,5-bifosfato
carboxilase oxigenase (Rubisco) além dos níveis de Ribulose 1,5 bifosfato (RuBP) e do ácido
3-fosfoglicerato (PGA) podem ser reduzidos após a exposição ao glyphosate (SERVAITES;
TUCCI; GEIGER, 1987). De acordo com De María et al. (2006), houve redução de
aproximadamente 26% da atividade da rubisco em folhas de Lupinus albus sete dias após a
aplicação de 10mM de glyphosate.
O glyphosate também pode prejudicar o metabolismo do carbono
interferindo na translocação e metabolismo dos açúcares. Orcaray et al. (2012), encontraram
acúmulo de carboidratos em folhas e raízes de Pisum sativum tratadas com o herbicida
glyphosate. Como o crescimento foi interrompido, o acúmulo de carboidratos nas raízes foi
atribuído à falta de utilização dos açúcares disponíveis o que também causou acúmulo de
carboidratos solúveis nas folhas. Embora poucos estudos tem sido feitos sobre o efeito do
AMPA no metabolismo do carbono, mostrou-se uma redução no teor de açúcares solúveis em
plantas tratadas com este metabólito (SERRA et al., 2013).
Muitos estudos têm demonstrado uma relação do glyphosate com a
nutrição de N afetando a fixação ou assimilação do mesmo (ZABLOTOWICZ; REDDY,
2007; BELLALOULI et al., 2008; ZOBIOLE et al., 2012). Os efeitos do glyphosate no
metabolismo do nitrogênio têm sido estudados principalmente em soja, no qual a fixação
simbiótica de N representa de 40 a 70% da necessidade total de N pela planta
(ZABLOTOWICZ; REDDY, 2007). Este herbicida pode afetar através dos efeitos diretos nos
rizóbios simbiontes ou indiretamente afetando a fisiologia da planta hospedeira (ZOBIOLE et
al., 2010a).
Além das plantas, os microrganismos também possuem a enzima EPSPs
sendo assim, suscetíveis ao glyphosate (FISCHER et al., 1986). A inibição dessa enzima
desregula a rota o que resulta em um fluxo descontrolado de carbono e subsequentemente a
acumulação maciça de chiquimato e dos ácidos hidroxibenzóicos como o protocatecuico ou
gálico nas fontes como folhas e nódulos de leguminosas (SINGH; SHANER, 1998). O

20
acúmulo do ácido protocatecuico em nódulos de plantas afetadas pelo herbicida glyphosate
sugere uma possível translocação do herbicida para os nódulos. Esta hipótese foi reforçada
pela redução da atividade da enzima nitrogenase mostrada em Bradyrhizobium japonicum
(HERNANDEZ; GARCIA-PLAZAOLA; BECERRIL, 1999).
Os efeitos de AMPA na nodulação, fixação e assimilação de nitrogênio
ainda precisa ser mais estudado. Ding et al. (2011), não observaram qualquer efeito sobre os
parâmetros de nodulação como número de nódulos e peso seco ou na fixação de N. Isto
sugere que mesmo o AMPA sendo translocado para os nódulos, não causa toxidez aos
simbiontes envolvidos na fixação.
4.5 Fenilalanina amônia liase (PAL)
A Fenilalanina amônia liase (PAL) é uma das enzimas mais estudadas
em relação ao metabolismo secundário em plantas e um dos motivos, é que em um
determinado tecido, os níveis desta enzima podem variar significativamente em intervalos
relativamente curtos de tempo em resposta a uma grande variedade de estímulos (CAMM;
TOWERS, 1973). A primeira observação registrada do isolamento e reações da PAL foi
realizada por Koukol e Conn em 1961, demonstrando que a mesma catalisa uma reação que
converte a L-fenilalanina em ácido trans-cinâmico mais amônia:
L-fenilalanina ácido trans-cinâmico + NH3
Este processo é o primeiro passo da via do fenilpropanóide (Figura 5).
Figura 5. Via biossintética fenilpropanóide. Fonte: Du et al., 2009.

21
Os fenilpropanóides possuem como características um grupo fenol
sendo produzido pelo metabolismo secundário dos vegetais desempenhando várias funções
(TAIZ; ZAIGER, 2004). Dentre estas funções pode-se citar: suporte mecânico (lignina)
(WHETTEN; SEDEROFF, 1992), proteção contra estresses bióticos e abióticos
(antioxidantes) (DIXON; PAIVA, 1995), pigmentos como as antocianinas (HOLTON;
CORNISH, 1995), taninos (JONES, 1984), etc. Tem sido proposto que a PAL é uma enzima
chave na regulação da via biossintética que conduz a estes produtos finais (MACDONALD;
D’CUNHA, 2007). Estes compostos podem ser produzidos através de duas rotas
biogenéticas: a via do mevalonato, onde se inicia através da acetil-coenzima A e malonil-
coenzima ou pela via do ácido chiquímico (TAIZ; ZAIGER, 2004) no qual o glyphosate atua.
A PAL pode ser encontrada em plantas assim como em fungos e
leveduras e sua atividade é sensivelmente modificada em resposta a estímulos tais como
ferimentos, ataque de patógenos, irradiação ultravioleta, baixa temperatura, baixos níveis de
nitrogênio, fosfato, ferro, aplicação de herbicidas, etc (HAHLBROCK; GRISEBACH, 1979;
DUKE; HOAGLAND; ELMORE, 1980; DIXON; PAIVA, 1995).
Vários estudos têm demonstrado que o glyphosate afeta negativamente
o metabolismo secundário (HOAGLAND, 1980; LYDON; DUKE, 1989; ZOBIOLE et al.,
2010b) no qual a PAL tem função essencial. Como demonstrado na Figura 4, o glyphosate
inibe a enzima EPSPs prejudicando assim a formação dos compostos que virão
posteriormente na rota. Dentre estes compostos, estão os aminoácidos aromáticos no qual a
fenilalanina serve de substrato para a PAL. De acordo com as informações disponíveis, não
está totalmente definido como a regulação da via do ácido chiquímico ocorre e nem se os
sistemas são similares para todas as espécies de plantas.
Com a aplicação de glyphosate, ocorre a redução dos aminoácidos
aromáticos onde a fenilalanina serve de substrato para a PAL, consequentemente deveria
diminuir as concentrações desta enzima. Porém, Duke et al. (1980) observaram que a
aplicação de glyphosate reduziu as concentrações de tirosina e fenilalanina, mas elevou as
concentrações desta enzima. No mesmo experimento, a aplicação de inibidores da PAL
provocou redução dos níveis de trans-cinamato, porém aumentou os níveis de fenilalanina e
tirosina. Mobin et al. (2015), testando a atividade da PAL após a aplicação de glyphosate,
também concluíram o aumento da enzima. De acordo com estes autores, a indução da
atividade desta enzima em resposta a aplicação do glyphosate, pode ser vista como uma
tentativa para aumentar o fluxo de carbono nesta rota. Além disso, o excesso de amônia

22
observado em plantas tratadas com glyphosate pode ser devido ao aumento da atividade desta
enzima (HOWLES et al., 1996).
Esses resultados indicam que esta enzima é regulada não só pela
fenilalanina, mas também pelo produto da reação (ácido transcinâmico). De acordo com
Velini et al. (2009b), isto pode sugerir que nem sempre os produtos distantes do sítio de ação,
neste caso a enzima EPSPs, terão a sua produção bloqueada ou reduzida pelo glyphosate
devido a presença de sistemas de controle, que podem em parte, compensar a redução da
síntese de determinados compostos intermediários. Quanto menor o número de reações e
enzimas, no caso dos herbicidas, que separam um composto até o sítio de ação, maior a
possibilidade que os níveis deste produto sejam reduzidos devido à ação do produto.
Poucos autores atualmente tem demonstrado o efeito do glyphosate
nesta enzima tão importante na produção de compostos fenólicos e até o momento nenhum
deles comparou estes efeitos em plantas daninhas resistentes ao mesmo.
4.6 Resistência de plantas daninhas aos herbicidas
A utilização de herbicidas para o controle de plantas daninhas é um
método comum na agricultura possibilitando a obtenção de elevadas produtividades por
reduzir a interferência das infestantes nas culturas de interesse econômico. Atualmente, os
agricultores depositam muita confiança no controle químico das plantas daninhas, e essa
confiança deve-se, principalmente, ao fato de que este método tem sido muito eficiente
possuindo custo atrativo, estando prontamente disponível e profissionalmente desenvolvido
(AGOSTINETTO; VARGAS, 2009). Porém, o uso indiscriminado desses herbicidas
propiciou a seleção de muitos casos de resistência a tais compostos por diversas espécies
daninhas (BURNSIDE, 1992).
O fenômeno da resistência de plantas daninhas a herbicidas é conhecido
há vários anos e nos últimos tem gerado muitos interesses, pois houve introdução no mercado
de novos grupos químicos altamente eficientes, controlando as infestantes em baixas doses e
específicos quanto ao sítio de ação. Os principais efeitos deste fato são: restrição ou
inviabilização da utilização desses produtos, perdas de áreas de plantio, perdas de
rendimentos e qualidade dos produtos agrícolas, reaplicação de herbicidas, aumento de doses,
e com isso aumento nos custos e também um maior impacto ambiental (CHRISTOFFOLETI,
2008).

23
O primeiro relato de resistência de plantas daninhas aos herbicidas
ocorreu em 1957, quando foram identificados biótipos de Commelina difusa infestante na
cultura da cana-de-açúcar no Havaí resistente ao herbicida 2,4-D (HEAP, 2015). A partir daí,
os casos registrados vêm aumentando a cada ano. Atualmente existem 462 casos registrados
em todo o mundo composto por 248 espécies onde 144 são dicotiledôneas e 104
monocotiledôneas. Estes casos têm sido reportados em 86 culturas e em 66 países tendo a
cultura do trigo a maior quantidade de casos registrados (HEAP, 2015). Estima-se que, no
mundo, haja mais de cinco milhões de hectares de culturas com invasoras apresentando
alguma resistência (KISSMANN, 1996).
Segundo Maxwell e Mortimer (1994), o aparecimento das plantas
daninhas resistentes a herbicidas é um exemplo de evolução como consequência das
mudanças no ambiente provocada pelo homem e geralmente pode ser identificada quando
pelo menos 30% das plantas que se encontram, são resistentes. A teoria mais aceita para
explicar o desenvolvimento da resistência é a da seleção natural no qual diz que, em toda
população de planta daninha há biótipos resistentes preexistentes e devido a pressão de
seleção imposta pelo herbicida, os biótipos suscetíveis morrem permanecendo aqueles
resistentes que se reproduzem aumentando assim sua população (CHRISTOFFOLETI;
LÓPEZ-OVEJERO, 2004). O tempo para o surgimento de plantas resistentes varia de um
local para o outro, do herbicida utilizado e também da frequência de uso do mesmo
(MAXWELL; MORTIMER, 1994).
Uma planta é sensível a um herbicida quando o seu crescimento e
desenvolvimento são alterados como resultado da sua incapacidade em suportar a ação do
produto levando a mesma à morte. Já a resistência é a capacidade que um biótipo tem dentro
de uma determinada população em sobreviver a dose de registro do herbicida, onde em
condições normais controlaria os demais integrantes de uma população (CHRISTOFFOLETI,
2008; AGOSTINETTO; VARGAS, 2009). Sendo assim, há variações no perfil bioquímico e
fisiológico das plantas quando as mesmas são tratadas com herbicidas, neste caso o
glyphosate.
De acordo com Powles e Yu (2010), os fatores que afetam a evolução
da resistência podem ser divididos em genéticos (frequência do gene resistente, número de
genes resistentes e dominância do gene resistente); biológicos (tipo de fecundação,
capacidade de produção de sementes, longevidade do banco de sementes no solo e a
capacidade de movimentação da semente ou do pólen); herbicida (estrutura química, sítio de

24
ação e atividade residual) e operacional (dose do herbicida, habilidade do operador, fatores do
agroecossistemas). Porém, os três fatores principais que influenciam a resistência são: pressão
de seleção causada pelo herbicida, frequência inicial do gene resistente e a densidade da
planta daninha (CHRISTOFFOLETI; LÓPEZ-OVEJERO, 2008). Dentre os fatores, os
genéticos e os bioecológicos são de difícil manipulação pelo homem, porém de grande
importância na avalição do potencial de risco de resistência. Já os fatores agronômicos são
aqueles os quais podem ser manipulados pelo homem na implementação de estratégias de
manejo da resistência (CHRISTOFFOLETI; LÓPEZ-OVEJERO, 2008).
Dentro do conceito de resistência existem dois tipos: cruzada e
múltipla. A resistência cruzada se dá quando um biótipo é resistente a dois ou mais herbicidas
pertencentes a um mecanismo de ação enquanto que a resistência múltipla ocorre quando o
indivíduo é resistente a dois ou mais mecanismos de ação distintos. No caso da resistência
cruzada, dentro de um mecanismo de ação, um biótipo não necessariamente é resistente a
todos os grupos químicos pertencentes a este mecanismo podendo haver variação. Ainda,
pode haver variações no nível de resistência dentro de um mesmo grupo químico
(CHRISTOFFOLETI, 2008; AGOSTINETTO; VARGAS, 2009). Foram encontrados níveis
de resistência cruzada diferentes em Euphorbia heterophylla aos diferentes herbicidas
pertencentes ao grupo químico das imidazolinonas e sulfoniluréias, os quais fazem parte dos
herbicidas inibidores da ALS (XAVIER, 2014). Em relação a resistência múltipla existem
muitos trabalhos relatando a ocorrência deste fato sendo que o mesmo tem aumentado a cada
ano (HEAP, 2015). Neste caso, as alternativas de manejo são mais limitadas.
O surgimento de plantas daninhas resistentes é devido aos diferentes
mecanismos que conferem a resistência, sendo estes, relacionados a alteração ou não do local
de ação no qual o herbicida atua (POWLES; PRESTON, 2006). O mecanismo de resistência
causado pela alteração do local de ação pode ocorrer pela troca de um ou mais aminoácidos
na enzima alvo o que impede, o acoplamento adequado do herbicida, ou ainda, pode estar
relacionado com a super expressão da enzima alvo (POWLES; YU, 2010). Vários trabalhos
têm relatado este tipo de mecanismo (VILA-AIUB; NEVE; POWLES, 2005; TARDIF;
RACJAN; COSTEA, 2006; HOCHBERG; SIBONY; RUBIN, 2009; THIEL; KLUTH;
VARRELMANN, 2010). Quando o mecanismo não está relacionado a alteração do local de
ação, há uma dificuldade do herbicida em chegar em quantidades letais no ponto de inibição
impedindo sua ação completa. Neste caso, pode estar relacionados mecanismos como a
diminuição da absorção ou translocação, aumento da taxa de metabolismo ou sequestro do

25
herbicida (FENG et al., 2004; PEREZ-JONES et al., 2007; DINELLI et al., 2008; POWLES;
YU, 2010).
As espécies Lolium rigidum, Echinochloa crus-galli var. crus-galli e
Poa annua são as que apresentam o maior número de casos de resistência aos diferentes
herbicidas e seus sítios de ação com respectivamente 11, 10 e 9 casos registrados. Dentre os
sítios de ação, os inibidores da ALS, inibidores do PSII (triazinas) e inibidores da ACCase são
aqueles que apresentam o maior número de espécies de plantas daninhas resistentes com 151,
73 e 47 casos respectivamente. Os Estados Unidos da América é o país que mais apresenta
casos de plantas daninhas resistentes a herbicidas totalizando 154 (HEAP, 2015). O primeiro
caso de resistência de planta daninha a herbicida no Brasil foi registrado em 1993 no qual a
espécie Bidens pilosa mostrou-se resistente aos herbicidas inibidores da ALS e o primeiro
caso de resistência múltipla ocorreu em 2004 com a planta Euphorbia heterophylla resistente
aos inibidores da ALS e PPO. A partir daí, os casos registrados vêm aumentando sendo o
mais recente ocorrido em 2015 com a espécie Amaranthus palmeri resistente ao herbicida
glyphosate. Atualmente há no Brasil 35 biótipos com alguma resistência a herbicidas, dentre
estes, oito apresentam resistência múltipla a dois ou mais mecanismos de ação distintos.
Os principais custos da resistência de plantas daninhas a herbicidas
estão relacionados ao uso de herbicidas com mecanismos de ação diferentes e as perdas de
produtividade nas culturas de interesse econômico. A utilização de herbicidas alternativos
ocorre de acordo com o produtor variando assim seu preço, pois na maioria dos casos, existe
mais de uma opção de produto para o manejo das infestantes resistentes. De um modo geral, o
custo de controle em casos de resistência simples, pode variar entre US$ 1,56 a US$ 59,76 e
no caso de resistência múltipla US$ 7,81 a US$ 56,25 (VARGAS et al., 2013). Citando como
exemplo o estado do Rio Grande do Sul com uma área de aproximadamente 4 milhões de
hectares cultivados com soja e que 50% apresenta problemas com buva e azevém, os
prejuízos devido à resistência com a necessidade de utilizar herbicidas alternativos, estão
entre US$ 3,1 milhões e US$ 119,53 milhões por ano, sem contar o impacto ambiental
causado pela maior utilização de herbicidas (VARGAS et al., 2013).
Devido a não existir ou existir poucos herbicidas com mecanismos de
ação diferentes para o uso no controle de biótipos resistentes, o evento da resistência assume
grande importância a nível mundial. A ocorrência de resistência múltipla agrava ainda mais o
problema, pois são dois ou mais mecanismos que precisam ser substituídos. Dessa forma, o

26
controle de plantas resistentes fica seriamente comprometido, restringindo esta prática e
outros métodos menos eficientes (SILVA et al., 2007).
4.7 Resistência de plantas daninhas ao glyphosate
A biotecnologia moderna vem se mostrando primordial para assegurar a
competitividade dos países exportadores de alimentos em todo o mundo. O rápido
desenvolvimento de culturas geneticamente modificadas permitiu aos agricultores reduzir
seus custos elevando também os índices de produtividades. Antes da introdução dessa
tecnologia o herbicida glyphosate era aplicado uma vez ao ano aproximadamente na época de
pré-semeadura. A partir de 1996 com a introdução de culturas tolerantes ao glyphosate, o
mesmo, passou a ser utilizado mais vezes como no caso da cultura da soja sendo aplicado de
duas a três vezes por ciclo (VARGAS et al., 2007a). Isto leva a uma maior pressão de seleção
exercida pelo herbicida sobre as plantas daninhas favorecendo assim a ocorrência da
resistência. Este fato aliado à adoção de sistemas conservacionistas do solo além da
possiblidade de usar este herbicida em qualquer estádio fenológico das culturas contribui para
um elevado risco de aparecimento de espécies resistentes a este produto (CHRISTOFFOLETI
et al., 2009).
Os casos de plantas daninhas resistentes ao herbicida glyphosate podem
ser considerados recentes, pois até 1995 não existia registros no mundo sobre este fato
(HEAP, 2015). A partir de 1996 com o primeiro caso registrado por Jim Pratley e Christopher
Preston com Lolium rigidum resistente nas culturas de canola e cereais na Austrália, a
quantidade de espécies não controladas pelo mesmo vem aumentando. Atualmente no mundo,
existem 32 espécies resistentes ao herbicida glyphosate onde os EUA lideram com 15 casos
registrados (HEAP, 2015). No Brasil, o primeiro registro ocorreu em 2003 com Lolium
multiflorum resistente ao glyphosate em pomares e culturas anuais no estado do Rio Grande
do Sul (ROMAN et al., 2004), após isso, mais seis espécies foram registradas onde a última
foi o Amaranthus palmeri registrado em 2015 (HEAP, 2015).
Dentre os mecanismos básicos de resistência ao herbicida glyphosate
até o momento tem-se a alteração do local de ação. Este caso é tipicamente representado pela
substituição de um aminoácido afetando a interação do herbicida com a enzima alvo. O
primeiro caso reportado desse tipo de mecanismo ocorreu em biótipos de Eleusine indica na
Malásia devido à substituição do aminoácido prolina por uma serina na Pro-106 (Pro-106-Ser)

27
em uma região altamente conservada do gene EPSPs (LEE; NGIM, 2000). Baerson et al.
(2002), verificaram que a aplicação do glyphosate não induziu as plantas resistentes a
produzirem mais EPSPs pressupondo que o mecanismo de resistência relacionado fosse a
menor sensibilidade da enzima nessas populações. Posteriormente foram relatados
substituição pelos aminoácidos alanina e treonina na Pro-106 em biótipos de L. rigidum e E.
indica (NG et al., 2003; YU; CAIRNS; POWLES, 2007).
Estas substituições na Pro-106 conferem um modesto nível de
resistência ao herbicida glyphosate (POWLES; YU, 2010). Porém, substituições na Gly-96 ou
Thr-97, conferem alto nível de resistência reduzindo significativamente a afinidade da PEP
(ESCHENBURG et al., 2002; FRUNK et al., 2009).
A influência das mutações gênicas que conferem a resistência ao
glyphosate na adaptabilidade ecológica das plantas não está totalmente esclarecida. Existem
diversos estudos que evidenciam algumas diferenças (ISMAIL et al., 2002; PEDERSEN et
al., 2007; PRESTON et al., 2009) porém, nada evidente que comprove uma diminuição da
adaptabilidade ecológica dos biótipos resistentes.
Outro tipo de mecanismo que tem sido recentemente documentado está
relacionado à amplificação do gene. Este evento proporciona alta resistência ao herbicida
glyphosate e foi constatado em plantas de Amaranthus palmeri (GAINES et al., 2010). Neste
caso há um aumento no número de cópias da enzima EPSPs. Gaines et al. (2012),
demonstraram que pode haver hibridação interespecífica entre espécies do gênero
Amaranthus. Isto pode ter um grande impacto no manejo de plantas daninhas devido à
facilidade de dispersão do gene que codifica este tipo de resistência, podendo ser transferido
para várias espécies deste gênero dificultando assim o controle das mesmas.
Plantas com maior amplificação do gene EPSPs (53 vezes) mostraram
elevados níveis de resistência ao glyphosate enquanto àquelas que apresentaram menor
amplificação do gene EPSPs (21 vezes), dotaram de um nível mais baixo de resistência ao
glyphosate (VILA-AIUB et al., 2014).. Neste mesmo experimento, sem aplicação do
herbicida glyphosate, mas em condições competitivas, plantas exibindo até 76 vezes o gene
EPSPs amplificado, apresentaram altura e alocação da biomassa para os órgãos vegetativos e
reprodutivos semelhantes em comparação às plantas de A. palmeri sensíveis ao glyphosate
sem amplificação do gene (VILA-AIUB et al., 2014). Este fato denota uma semelhança na
adaptabilidade ecológica entre os biótipos resistentes e suscetíveis de A. palmeri. Esta é uma

28
informação muito valiosa no planejamento do manejo integrado de plantas daninhas
resistentes.
A translocação diferenciada do herbicida em determinadas plantas
também pode ser considerado um mecanismo de resistência. Em algumas espécies resistentes,
o glyphosate possui translocação restrita não permitindo assim, que o mesmo alcance o local
de sua ação. Desde o primeiro caso registrado de resistência ao herbicida glyphosate em
plantas de Lolium, a translocação restrita por toda a planta e raiz tem sido confirmada em
várias espécies de Lolium, Conyza, Sorghum halepense, etc. (WAKELIN; LORRAINE-
COLWILL; PRESTON, 2004; POWLES; PRESTON, 2006; YU et al., 2009; SHANER,
2009; GE et al., 2011; BOSTAMAM et al., 2012; VILA-AIUB et al., 2013; ADU-YEBOAH
et al., 2014; CARDINALI et al., 2015). Em experimento realizado na Austrália com
população de Lolium rigidum, a planta resistente mostrou padrão de translocação diferente da
suscetível. Neste caso, o glyphosate translocou-se mais para as raízes na população suscetível
enquanto que na resistente houve pouca translocação (LORRAINE-COLWILL et al., 2002).
Este tipo de mecanismo é mais comum em plantas do que a mutação na Pro-106 citada
anteriormente (POWLES; YU, 2010).
Experimentos usando ressonância nuclear magnética com o objetivo de
observar o glyphosate em células in vivo e compartimentos celulares identificaram pelo
menos dois tipos de mecanismos que resultaram em translocação restrita (GE et al., 2014). O
primeiro está relacionado ao sequestro do glyphosate acumulando-se no vacúolo, além disso,
o glyphosate não sai do vacúolo para o citoplasma pressupondo que o transporte através da
membrana seja unidirecional em todas as espécies estudadas até o momento (GE et al., 2013).
Neste mesmo experimento foi observado um acúmulo de 10 vezes mais glyphosate nos
vacúolos de biótipos resistentes de Conyza canadensis em relação ao biótipo suscetível.
Foi demonstrado que biótipos de C. canadensis resistentes após a
aplicação do glyphosate e em baixas temperaturas (8°C) tiveram o sequestro de glyphosate no
vacúolo inibido tornando-se suscetíveis ao mesmo (GE et al., 2011). Isto proporcionou uma
evidência inequívoca de que a resistência seria unicamente devido ao processo de sequestro
mostrando que as propriedades fisiológicas do mecanismo de resistência podem inverter a
resistência ao herbicida. Estudo realizado na Itália com L. multiflorum resistente em clima frio
e com aplicação de glyphosate, demonstrou que a maior parte da população resistente no qual
possuía também uma mutação na Pro-106, não se tornou sensível em baixa temperatura
(COLLAVO; SATTIN, 2012). Isto pressupõe que múltiplos mecanismos de resistência

29
podem contribuir para a magnitude global da resistência e nem todos os mecanismos são
sensíveis à temperatura (SAMMONS; GAINES, 2014).
Algumas espécies que apresentam translocação diferenciada como
mecanismo de resistência ao glyphosate possuem diferenças na adaptabilidade ecológica.
Biótipos de azevém sensíveis ao glyphosate acumularam maior quantidade de matéria seca,
maior quantidade de perfilhos por planta, maior número de inflorescências produzindo assim
maior quantidade de sementes. Outra questão interessante avaliada no mesmo experimento é
que o biótipo suscetível floresceu 19 dias antes do resistente completando o ciclo em média
25 dias antes (FERREIRA et al., 2009). Outros trabalhos também evidenciaram diferenças no
crescimento e desenvolvimento entre biótipos resistentes e suscetíveis de Lolium (PRESTON;
WAKELIN, 2008; PRESTON et al., 2009). Porém outros estudos não evidenciaram
diferenças ou apenas pequenas diferenças na adaptabilidade entre os biótipos resistentes e
suscetíveis (PEDERSEN et al., 2007; DAVIS et al., 2009; SHRESTHA et al., 2010).
De acordo com Michitte et al. (2007), há outros genes que podem
contribuir para uma menor penetração do herbicida na folha. Em biótipos de azevém
resistentes houve uma redução na penetração da folha após aplicação do glyphosate em torno
de 39% em relação aos biótipos suscetíveis (MICHITTE et al., 2007. Em experimento
realizado por Nandula et al. (2008), foi encontrado diferença de até 43% na redução da
penetração pelos biótipos resistentes.
Outro tipo de mecanismo envolvido está na diferença de metabolismo.
No solo e nas plantas, o principal metabólito do glyphosate é o ácido aminometilfosfônico
(AMPA) (REDDY et al., 2004). Em algumas plantas naturalmente tolerantes ao glyphosate
como Agropyron repens e Equisetum arvense este metabólito pode ser detectado
(COUPLAND, 1985; DYER, 1994). Na planta a metabolização do glyphosate em AMPA
pode ocorrer pela enzima GOX (glyphosate oxiredutase) onde o produto da reação pode ser
tóxico para aquelas que possuem EPSPs insensível a este herbicida (NANDULA et al., 2007).
O ácido aminometilfosfônico é considerado tóxico, mas menos ativo que o glyphosate
(REDDY; RIMANDO; DUKE, 2004). O mesmo foi encontrado em diversas culturas como
trigo, canola, ervilha, cevada, linho e soja tolerante ao glyphosate após a aplicação do mesmo
(CESSNA et al., 1994; 2000; 2002; DUKE et al., 2003). Arregui et al. (2003), detectaram
níveis de AMPA em folhas e sementes de soja tolerante ao glyphosate após a aplicação do
mesmo indicando a metabolização em seu produto.

30
Plantas resistentes ao glyphosate da espécie Ambrosia trifida tem
mostrado uma resposta diferente em relação aos mecanismos anteriormente citados. Folhas
maduras desta espécie, 12 horas após receber tratamento com o herbicida glyphosate,
apresentaram rápida necrose e morte das células (ROBERTSON, 2010). Neste caso a
translocação é bastante reduzida provavelmente devido a rápida necrose e a incapacidade do
glyphosate ser transportado a partir das folhas maduras. Ainda não se conhece geneticamente
e molecularmente o que exatamente causa este fenômeno e pesquisas futuras serão
importantes para elucidar tal fato.
Deve-se ressaltar que apesar dos mecanismos terem sido didaticamente
separados e esclarecidos, uma espécie pode possuir mais de um, e dentro desse contexto o
manejo se torna mais complexo pela dificuldade de alternativas no controle.
4.8 Azevém (Lolium multiflorum) resistente ao glyphosate.
O controle de azevém para formação de palha no plantio direto e em
pomares é comumente realizado com glyphosate. O número de aplicações em uma safra varia
de acordo com a cultura podendo ocorrer mais de cinco durante o ciclo produtivo no caso da
fruticultura (FERREIRA et al., 2009). Para culturas anuais existe maior número de herbicidas
com mecanismos de ação diferentes onde em geral os graminicidas pertencentes aos grupos
ariloxifenoxipropionatos e ciclohexanodionas controlam com eficiência o azevém, porém,
mesmo utilizando estes herbicidas para a dessecação, pode haver a necessidade de aplicar
glyphosate para o controle das dicotiledôneas.
A primeira constatação de azevém (Lolium multiflorum) resistente ao
herbicida glyphosate ocorreu em 2002 sendo identificado em pomares no Chile que recebiam
em média três aplicações por ciclo durante os últimos 10 anos (PERES; KOGAN, 2003).
Após isso, os casos vêm aumentando ao longo dos anos. Em relação ao Brasil, o primeiro
caso foi relatado por Roman et al. (2004), que identificaram biótipos de azevém resistente ao
glyphosate em lavouras de culturas anuais e em pomares.
O controle de azevém resistente ao glyphosate tem sido realizado com o
uso de graminicidas dependendo da cultura instalada sendo possível observar diferenças na
evolução dos sintomas e no período de controle tanto para os biótipos suscetíveis quanto para
os resistentes entre os diversos herbicidas utilizados sugerindo que o mecanismo que confere
a resistência ao glyphosate possa estar interferindo na atividade destes graminicidas

31
(VARGAS et al., 2005). Isto pode implicar em uma maior ou menor eficiência no controle de
acordo com o herbicida testado (HARKER; DEKKER, 1988). Powles et al. (1998), avaliaram
biótipos de Lolium rigidum resistente ao glyphosate e demonstraram que o mesmo apresentou
resistência cruzada ao herbicida diclofop (inibidor da ACCase) mostrando-se sensível ao
fluazifop e ao setoxydim (inibidores da ACCase).
Dentre os mecanismos de resistência identificados para Lolium
multiflorum podem-se citar a translocação diferenciada (FERREIRA et al., 2006a;
FERREIRA et al., 2008; GONZÁLES-TORRALVA et al., 2012a), mutações genéticas
(PEREZ-JONES et al., 2007; JASIENUIK et al., 2008; NANDULA et al., 2008) e
recentemente identificado, a amplificação do gene da EPSPs (SALAS et al., 2012; SALAS et
al., 2015).
Um fator que deve ser levado em consideração é que o azevém não só
adquiriu resistência ao glyphosate, mas também a outros herbicidas. Foram identificados
azevém com resistência múltipla ao glyphosate e aos inibidores da ACCase e ALS
(MARIANI et al., 2014). Este fato restringe o controle desta espécie pelo número reduzido de
herbicidas com mecanismos de ação distintos que podem ser menos eficientes e possuírem um
custo maior e ainda por cima serem mais fitotóxicos para as culturas. Os biótipos de azevém
resistentes ao herbicida glyphosate estão presentes em mais de 80% das lavouras de soja do
estado do Rio Grande do Sul e os que possuem resistência múltipla estão em mais de 30%. O
aumento da presença desta espécie nas lavouras comerciais associado à baixa perspectiva de
lançamento de novas moléculas herbicidas especificamente para o controle do azevém
representa um grande impacto econômico e técnico na agricultura brasileira (DIONISIO et al.,
2013).

32
5. MATERIAL E MÉTODOS
Foram realizados quatro estudos no Núcleo de Pesquisas Avançadas em
Matologia (NUPAM), pertencente ao Departamento de Produção e Melhoramento Vegetal da
Faculdade de Ciências Agronômicas (FCA) – Universidade Estadual Paulista “Júlio de
Mesquita Filho” (UNESP) – Campus de Botucatu - SP. Foram semeados 3 populações de
azevém (Lolium multiflorum) sendo consideradas como suscetível (S), com suspeita de
resistência (R1) e resistente (R2) ao herbicida glyphosate.
5.1 Curvas de dose-resposta das diferentes populações de azevém
Esse estudo teve como objetivo quantificar a massa seca e analisar a
fitointoxicação das três populações S, R1 e R2. A população suscetível foi adquirida da
empresa Agrocosmos situada na cidade de Engenheiro Coelho - SP, a população com suspeita
de resistência foi colhida no campo experimental da empresa Arysta LifeScience situada na
cidade de Pereiras-SP e a população resistente foi obtida da Embrapa Trigo situada na cidade
de Passo Fundo - RS. Para tal, foram semeados as três populações citadas anteriormente em
vasos com capacidade de 0,5 L em substrato Carolina (Turfa de Sphagno, vermiculita
expandida, resíduo orgânico casca de arroz torrefada, calcário dolomítico, gesso agrícola e
fertilizante NPK) com pH 5,5. A semeadura foi realizada no dia 10 de setembro de 2014 e
duas semanas após foi feito desbaste das mesmas deixando duas plantas por vaso. O herbicida
foi aplicado no dia 17 de outubro de 2014 aos 37 dias após a semeadura quando as plantas se
encontravam com aproximadamente seis perfilhos (Figura 6).

33
Figura 6. Estádio das plantas de azevém no momento da aplicação dos tratamentos Botucatu -
SP, 2014.
O delineamento experimental utilizado foi inteiramente casualisado
(DIC) com oito tratamentos e quatro repetições. O produto comercial utilizado foi o Roundup
Original (360g e.a. L-1
). Os tratamentos testados estão apresentados na Tabela 1. Foram
realizadas avaliações visuais de controle das plantas aos 21 dias após a aplicação (DAA),
segundo escala percentual de notas, onde “0” corresponde a nenhum controle e “100”
significa a morte das plantas, conforme Sociedade Brasileira da Ciência das Plantas Daninhas
(1995). Aos 28 DAA foi coletada a parte aérea das plantas e colocadas em sacos de papel e
levadas para estufa de circulação forçada de ar a 60°C por três dias até que o material
atingisse peso constante para então a massa seca ser pesada em balança de precisão (0,0001g).
Tabela 1: Doses de glyphosate utilizados no experimento. Botucatu - SP, 2014.
Tratamentos Doses de glyphosate
4
(g e.a. ha-1
)1
Doses de glyphosate
(%)2
1 0 0
2 135 12,5
3 270 25
4 540 50
5 10803
100
6 2160 200
7 4320 400
8 8640 800 1 gramas de equivalente ácido por hectare
2 dados em porcentagem da dose recomendada
3 dose recomendada (3 litros por hectare)
4 Roundup Ready (360 g e.a. L
-1)

34
Para a pulverização dos tratamentos descritos na Tabela 1 foi utilizado
um pulverizador estacionário (Figura 7), instalado no NUPAM, constituído por estrutura
metálica que suporta a barra de pulverização, com 1,5 metros de largura, que se desloca por
uma área útil de 6,0 m2 no sentido do seu comprimento. A barra é tracionada por um conjunto
de motor elétrico e modulador de frequência, tornando possível o controle da velocidade de
trabalho da barra. A barra foi equipada com quatro pontas XR 11002 VS, espaçadas em 0,5 m
entre si, e dispostas a 0,5 m de altura em relação às plantas. A pressão de trabalho utilizada
pelo equipamento foi de 2,0 kgf cm-2
, com velocidade de 3,6 km h-1
e consumo de calda de
200 L ha-1
. No momento da aplicação a temperatura estava em 34,1°C com umidade relativa
de 36.
Figura 7. Pulverizador estacionário com as unidades experimentais. Botucatu - SP, 2014.
5.2 Efeitos do glyphosate na enzima fenilalanina amônia liase nas diferentes
populações
O objetivo dessa etapa do trabalho foi determinar a atividade da enzima
fenilalanina amônia liase (PAL) nas diferentes populações após a aplicação do glyphosate.
Neste estudo, foram instalados dois experimentos em épocas diferentes contendo populações
diferentes, porém da mesma espécie. O delineamento experimental utilizado foi inteiramente
casualisado (DIC) com dois tratamentos (720 g e.a. ha-1
e 1080 g e.a. ha-1
) e quatro repetições

35
mais uma testemunha sem aplicação de cada população. O produto comercial utilizado foi o
Roundup Original (360 g e.a. L-1
).
No primeiro experimento utilizou-se sementes de populações
suscetíveis (S) e com suspeita de resistência ao herbicida glyphosate (R1). A semeadura foi
realizada no dia 05 de março de 2014 em vasos com capacidade de 1 litro contendo o mesmo
substrato descrito anteriormente. A aplicação do herbicida foi realizada no dia 17 de abril de
2014 com temperatura de 27,5°C e umidade relativa de 62%. O segundo experimento utilizou
sementes de populações suscetíveis (S) e com resistência ao herbicida glyphosate (R2). A
semeadura foi realizada no dia 9 de setembro de 2014 e a aplicação do herbicida foi feita no
dia 15 de outubro de 2014 com temperatura de 26,8°C e umidade relativa de 56%. Para ambos
os experimentos, duas semanas após a semeadura, foi realizado desbaste das plantas deixando
apenas quatro plantas por vaso. Após a aplicação dos tratamentos foram feitas coletas de todas
as folhas de cada unidade experimental as 12, 24, 48 e 72 horas após a aplicação (HAA).
Em cada coleta, as folhas foram lavadas com água destilada, após esse
processo foi retirado o excesso de água secando com papel toalha tomando o cuidado para não
romper as células ou danificar as folhas. Posteriormente, as folhas foram colocadas em sacos
plásticos, enroladas em papel alumínio, congeladas no nitrogênio líquido e armazenadas em
freezer -80°C para posterior análise da PAL (Figura 8).

36
Figura 8. Procedimento de preparo de amostra para PAL. Botucatu - SP, 2014.
Para a determinação da atividade da enzima PAL, foi utilizado o
método adaptado por Peixoto et al. (1999). Este trabalho foi realizado no Laboratório de
Análises Bioquímicas Vegetais do Departamento de Química e Bioquímica do Instituto de
Biociências, UNESP de Botucatu - SP. As amostras foram maceradas com nitrogênio líquido
no escuro e foram pesados 100 mg de cada unidade experimental. Posteriormente foram
adicionados 10 mL de tampão borato de sódio 0,1 M, pH 8,8 contendo 1,2 mL de β-
mercaptoetanol para cada 1000 mL de tampão borato e centrifugados a 18.836 g durante 25
minutos à 4°C. O processo de extração foi feito em triplicata, ou seja, para cada repetição
foram repetidos mais 3 vezes por amostra, totalizando 12 repetições por tratamento, sendo

37
assim, para cada amostra foram retirados 4 mL sendo colocado 1 mL por tubo de ensaio além
do branco da amostra. Foi acrescentado 1 mL de tampão borato 0,2 M, pH 8,8 e colocados em
banho-maria a 36°C por cinco minutos e acrescentado 1 mL de fenilalanina na concentração
de 1,652 gramas para cada 100 mL de água destilada. Ao final de 60 minutos, 100 μL de HCL
6N foram adicionados em cada tubo de ensaio com o objetivo de paralisar a reação e logo
após, feita a leitura de absorbância em espectrofotômetro no comprimento de onda de 290 nm
(Figura 9).
Figura 9. Procedimento de extração e quantificação da PAL. Botucatu - SP, 2014.
5.3 Efeitos do glyphosate na fotossíntese das diferentes populações
O objetivo desse estudo foi caracterizar alguns aspectos fisiológicos de
populações de azevém (Lolium multiflorum) suscetíveis e resistentes ao herbicida glyphosate
após a aplicação do mesmo. O delineamento experimental utilizado foi inteiramente
casualisado (DIC) com dois tratamentos (720 g e.a. ha-1
e 1080 g e.a. ha-1
) e quatro repetições

38
mais uma testemunha sem aplicação de cada população. O produto comercial utilizado foi o
Roundup Original (360 g e.a. L-1
).
Foram instalados dois experimentos em épocas diferentes contendo
populações diferentes, porém da mesma espécie. No primeiro experimento foi utilizado
sementes da população que se mostrou suscetível e população resistente ao herbicida
glyphosate. A semeadura foi realizada no dia 9 de setembro de 2014 em vasos com
capacidade de 1 litro contendo o mesmo substrato descrito anteriormente. Duas semanas após
a semeadura, foi realizado desbaste das plantas deixando apenas quatro plantas por vaso. A
aplicação do herbicida foi realizada no dia 15 de outubro de 2014 com temperatura a 26,8°C e
umidade relativa de 56%. Após a aplicação foram realizadas avaliações ao 1, 3, 7, e 28 DAA.
O segundo experimento foi instalado no dia 4 de dezembro de 2014 onde foram semeadas
sementes de azevém suscetíveis (S) e com suspeita de resistência (R1). Duas semanas após a
semeadura, foi realizado desbaste das plantas deixando apenas quatro plantas por vaso. A
aplicação do herbicida foi realizada no dia 9 de janeiro de 2015 com temperatura a 26,5°C e
umidade relativa de 74,1%. As avaliações foram realizadas ao 1, 7 e 28 DAA. Aos 3 DAA
não foi realizada avaliação devido ao tempo estar totalmente nublado impossibilitando este
tipo de avaliação.
Para as determinações fisiológicas foi utilizado um medidor portátil
IRGA, modelo LI-6400 xt (LI-COR) (Figura 10). As variáveis analisadas foram: taxa de
assimilação líquida de CO2 (fotossíntese), condutância estomática, concentração interna de
CO2 e transpiração. Com essas variáveis pôde-se calcular a eficiência do uso da água (taxa de
assimilação líquida de CO2/transpiração), eficiência instantânea de carboxilação (rubisco)
calculada pela taxa de assimilação líquida de CO2/concentração interna de CO2.
Figura 10. IRGA modelo LI-6400. Botucatu - SP, 2014.

39
5.4 Avaliação dos teores de glyphosate, AMPA e compostos da rota do ácido
chiquímico nas diferentes populações
O objetivo desse estudo foi quantificar compostos alterados em
populações de azevém (Lolium multiflorum) suscetíveis e resistentes ao herbicida glyphosate
após a aplicação do mesmo. Os compostos analisados foram: glyphosate, AMPA (ácido
aminometilfosfônico), ácido chiquímico, ácido quínico, shiquimato-3-fosfato, os aminoácidos
aromáticos fenilalanina, tirosina e triptofano, ácido ferúlico, ácido coumárico e ácido cafeico.
Nessa etapa foram instalados dois experimentos em épocas diferentes contendo populações
diferentes, porém da mesma espécie. O delineamento experimental utilizado foi inteiramente
casualisado (DIC) com oito tratamentos e quatro repetições. O produto utilizado foi o
Roundup Original (360 g e.a. L-1
). Os tratamentos testados estão apresentados na Tabela 1.
No primeiro experimento utilizou sementes das plantas suscetíveis (S) e
com suspeita de resistência ao herbicida glyphosate (R1). A semeadura foi realizada no dia 28
de novembro de 2013 em vasos com capacidade de 1 litro contendo o mesmo substrato citado
anteriormente. Duas semanas após a semeadura, foi realizado desbaste das plantas deixando
apenas quatro plantas por vaso. O herbicida foi aplicado no dia 9 de janeiro de 2014, quando
as plantas estavam no início do florescimento. No momento da aplicação a temperatura estava
em 26,4°C com umidade relativa de 69%. No segundo ensaio foi utilizado sementes da
população suscetível (S) e com resistência ao herbicida glyphosate (R2). A semeadura foi
realizada no dia 8 de setembro de 2014 e o herbicida foi aplicado no dia 29 de outubro de
2014, quando as plantas estavam no início do florescimento. No momento da aplicação a
temperatura estava em 24,8°C com umidade relativa de 52%.
Após a aplicação, foram realizadas coletas de todas as folhas
consideradas vivas (folhas verdes) de cada unidade experimental (vaso) aos 5, 11 e 28 dias
após a aplicação (Figura 11). As mesmas foram colocadas em sacos plásticos sendo lavadas
com 100 mL de água destilada por três vezes totalizando 300 mL com o intuito de remover o
glyphosate que permaneceu sobre a folha para analisar apenas o que foi absorvido e
metabolizado pela planta. Após a lavagem as folhas foram acondicionadas em sacos de papel
e colocadas em estufa de circulação forçada de ar, a 40°C, para que não ocorresse a
degradação dos compostos, por um período de 72 horas até que o material estivesse
totalmente seco.

40
Figura 11. Procedimentos da coleta das folhas. Botucatu - SP, 2014.
Após a secagem, as folhas foram maceradas com nitrogênio líquido e
colocadas em sacos de papel. A metodologia utilizada para extração dos compostos foi
proposta por Gomes et al. (2015). Após as amostras passarem pelos processos de secagem e
maceração, foi pesado 100 mg de cada amostra e colocadas em tubo “falcon” com capacidade
de 15 mL e adicionado10 mL de água acidificada a pH 2,5. Os tubos foram agitados para que
as amostras fossem misturadas com a água. Após esse processo, os tubos foram submetidos a
banho ultrassom durante 30 minutos a 45°C. Na sequência foram centrifugados a 10.595 g
durante 5 minutos a uma temperatura de 20°C e o sobrenadante coletado e filtrado em filtro
Millex HV (Millipore) 0,45 μm, com membrana durapore de 13 mm e acondicionados em vial
âmbar 9 mm (Flow Supply) com capacidade de 2 mL (Figura 12).

41
Figura 12. Procedimentos para extração dos compostos. Botucatu - SP, 2014.
Para a quantificação dos compostos foi utilizado um sistema LC-
MS/MS (Figura 13), composto por um Cromatógrafo Líquido de Alta Eficiência (HPLC),
Shimadzu, modelo Proeminence UFLC, que combina análise ultra-rápida e excelente
performance de separação, com alta confiabilidade de resultados. Os métodos de análise para
todos os compostos foi realizado de acordo com Gomes et al. (2015).

42
Figura 13. Cromatógrafo Líquido (Prominence UFLC) acoplado ao espectrômetro de
massas (3200 Q TRAP) – LC-MS/MS.
5.5 Análise dos dados.
5.5.1 Curvas de dose-resposta das diferentes populações de azevém
Os dados de massa seca e controle foram submetidos a análise de
variância e aplicação do teste F a 5% de probabilidade. Sendo os efeitos significativos, os
dados foram ajustados ao modelo de regressão não linear do tipo log-logístico proposto por
Streibig et al. (1988):
𝑦 =𝑎
[1 + (𝑥𝑏)𝑐
]
Onde: y = porcentagem de controle ou massa; x = dose do herbicida (g
e.a. ha-1
); a, b e c são parâmetros da equação, sendo que a = assíntota entre o ponto máximo e
mínimo da variável, b = dose que proporciona 50% da assíntota (corresponde ao C50 ou GR50)
e c = declividade da curva. Baseado na equação logística, as curvas foram elaboradas com
auxílio do software estatístico Sigmaplot 12.0.

43
5.5.2 Efeitos do glyphosate na enzima fenilalanina amônia liase nas diferentes
populações
Os dados obtidos foram avaliados estatisticamente pela análise de
variância. A magnitude do efeito dos tratamentos frente ao erro experimental foi testada
através do teste F. Os valores de F foram considerados significativos ao nível de significância
de 5% (p<0,05). Quando alcançada a significância estatística, as médias de cada nível dos
fatores foram comparadas entre si através do teste t (LSD), ao nível de significância de 5%.
5.5.3 Efeitos do glyphosate na fotossíntese das diferentes populações
Para os resultados das variáveis fisiológicas foi estabelecido o intervalo
de confiança pelo teste t a 10% de probabilidade. Para determinar o intervalo de confiança, foi
utilizada a seguinte equação:
IC = (t x desvpad) / raiz nr, onde:
IC = intervalo de confiança;
t = valor de t tabelado, ao nível de 10% de probabilidade;
desvpad = desvio padrão;
raiz nr = raiz quadrada do número de repetições.
5.5.4 Avaliação dos teores de glyphosate, AMPA e compostos da rota do ácido
chiquímico nas diferentes populações
Para os dados referentes aos níveis dos compostos nos tecidos foliares
foi realizada a análise de variância dos dados pelo teste F e as médias foram comparadas pelo
teste t a 5% de probabilidade. Foi calculado o erro padrão das médias de quatro repetições e
plotados nos gráficos como barras verticais.

44
6. RESULTADOS E DISCUSSÃO
6.1 Curvas de dose-resposta das diferentes populações de azevém
Os dados de porcentagem de controle bem como de massa seca da parte
aérea podem ser visualizados nas Figuras 14 e 15, respectivamente. De acordo com o
coeficiente de determinação (r2), os resultados mostraram ajustes próximos de 1 pelo modelo
proposto por Streibig et al. (1988). Observa-se que 25% da dose recomendada (270 g e.a. ha-1
)
foi suficiente para obter um controle acima de 80% na população S demonstrando assim, uma
grande suscetibilidade da mesma em relação ao herbicida glyphosate. Já para a população R1,
metade da dose recomendada (540 g e.a. ha-1
) foi capaz de proporcionar um controle acima de
80%. Apesar de estatisticamente as populações S e R1 serem diferentes, os valores tanto de
controle como de massa seca na dose recomendada (1080 g e.a. ha-1
) foram semelhantes, ou
seja, o glyphosate controlou ambas as populações eficientemente podendo as duas serem
consideradas suscetíveis ao mesmo.
Ao se obter o valor de C50 ou GR50, tem-se o fator de resistência
(F=R/S) no qual corresponde à quantidade de vezes em que a dose necessária para controlar
50% da população resistente é superior a que controla 50% da população suscetível
(CHRISTOFFOLETI; LÓPEZ-OVEJERO, 2008). De acordo com Saari et al. (1994), a
resistência pode ser confirmada quando o fator R/S for significativamente maior que 1,0.
Porém, a confirmação de um nível baixo de resistência é mais complexa. Quando ocorre um
caso deste tipo, é necessário fornecer um número maior de evidências. Este experimento foi
realizado em casa-de-vegetação e apesar de ter sido utilizado uma quantidade de doses

45
considerada ideal para obter resultados confiáveis, há a necessidade de fazer mais
experimentos principalmente no que se refere à condução em campo. Resultados semelhantes,
com herbicidas inibidores da ALS, foram encontrados por López-Ovejero et al. (2005), que
apesar do biótipo testado possuir um fator de resistência de 16,15, o mesmo foi
satisfatoriamente controlado na dose recomendada.
Quando uma população demonstra um elevado fator de resistência, ou
seja, doses dez vezes superiores à recomendada, não existem dúvidas de que a mesma é
resistente (CARVALHO; CHRISTOFFOLETI; LÓPEZ-OVEJERO, 2009). Porém, este caso
deve ser analisado de acordo com o herbicida em questão, pois o que pode ser considerado um
nível elevado de resistência para um, pode ser considerado baixo para outro. Os herbicidas
inibidores da ALS, ACCase e inibidores do fotossistema II, resultam de níveis extremamente
elevados de resistência, frequentemente maiores que 100 (GRESSEL, 2000), ou seja, são
considerados altos em relação ao herbicida glyphosate que possui fator de resistência
geralmente baixo apesar de YU et al. (2015), terem encontrado fator R/S de 182 em Eleusine
indica. Sendo assim, a população R2 pode ser considerada resistente ao herbicida glyphosate,
pois apresentou um elevado nível de resistência (Tabela 2).
Os resultados aqui mostrados corroboram com diversos autores
(ROMAN et al., 2004; VARGAS et al., 2005; FERREIRA et al., 2006b; SALAS et al., 2012).
As diferenças dos resultados entre os autores podem estar relacionadas com as condições
ambientais, diferentes formulações de glyphosate, condições de aplicação assim como o
crescimento das populações. O alto grau de resistência evidenciado na população R2 pode
indicar que o mecanismo de resistência envolvido pode ser resultante da ação de dois ou mais
mecanismos. Vários trabalhos têm reportado que a alteração na Pro106 juntamente com
translocação diferenciada ou outro mecanismo desconhecido, proporciona alto grau de
resistência que um mecanismo sozinho (KANDUM et al., 2011; BOSTAMAN et al., 2012;
NANDULA et al., 2013). Em outros casos, a duplicação de gene ou alteração no sítio de ação
foram detectados, mas não parece explicar totalmente os níveis mais elevados de resistência
(COLLAVO; SATTIN, 2012; BELL et al., 2013). A ocorrência de múltiplos mecanismos de
ação é comum, especialmente em espécies de polinização cruzada como a azevém,
produzindo maior nível de resistência em resposta à pressão de seleção imposta pela aplicação
constante do herbicida (SAMMONS; GAINES, 2014).
O manejo desta planta daninha deve ser fundamentado em herbicidas
com mecanismos de ação distintos, pois aumentar a dose de glyphosate não é

46
economicamente viável baseando-se na curva de dose-resposta, pois a aplicação de duas vezes
a dose recomendada, não foi suficiente para controlar esta população eficientemente.
A variável massa seca está em concordância com o que foi discutido
anteriormente. As populações S e R1 tiveram seus valores semelhantes na dose recomendada
sendo aproximadamente zero (Figura 15). Já a população R2 diferiu das mesmas provando
assim, seu grau de resistência (Tabela 3). Além disso, o dobro da dose não foi suficiente para
reduzir a massa seca desta população à zero. Apesar das populações S e R1 serem
consideradas suscetíveis, com o aumento da pressão de seleção ocasionado pelo uso constante
do herbicida glyphosate, a probabilidade de ocorrência da resistência é maior na população
R1 de acordo com os dados mostrados.
Doses de glyphosate (%)
10 100 1000
Cont
role
(%
)
0
20
40
60
80
100
120
R1
R2
S
Figura 14. Intoxicação das plantas de azevém suscetível (S) e resistente (R1 e R2) após a
aplicação de diferentes doses do herbicida glyphosate aos 21 DAA. O gráfico se
encontra em escala logarítma. Botucatu – SP.
Con
trole
(%
)

47
Tabela 2. Estimativas dos parâmetros a, b, c e o coeficiente de determinação (r²) do modelo
log-logístico para as populações de Lolium multiflorum em relação à porcentagem
de controle aos 21 DAA. Botucatu – SP.
Variável População a b (C50) c r² FR*
Controle (%)
R1 101,11 18,13
-2,04 0,99 1,99
R2 97,25 158,76 -3,74 0,99 17,39
S 100,01 9,13 -1,83 0,99
Equação do modelo: y=a/(1+((x/b)^c)) *Fator de Resistência=C50R/C50S
Doses de glyphosate (%)
10 100 1000
Mas
sa s
eca
(%)
0
20
40
60
80
100
120
140
R1
R2
S
Figura 15. Massa seca da parte aérea das plantas de azevém suscetível (S) e resistente (R1 e
R2) após a aplicação de diferentes doses do herbicida glyphosate aos 28 DAA. O
gráfico se encontra em escala logarítma. Botucatu – SP.

48
Tabela 3. Estimativas dos parâmetros a, b, c e o coeficiente de determinação (r²) do modelo
log-logístico para as populações de Lolium multiflorum em relação à porcentagem
de massa seca aos 28 DAA. Botucatu – SP.
Variável População a b (GR50) c r² FR*
Massa seca
(%)
R1 6,29 18,34 2,23 0,99 1,76
R2 6,05 150,42 2,84 0,96 14,46
S 6,55 10,40 2,88 0,99 Equação do modelo: y=a/(1+((x/b)^c))
*Fator de Resistência=GR50R/GR50S
6.2 Efeitos do glyphosate na enzima fenilalanina amônia liase nas diferentes
populações
No primeiro experimento, a atividade da enzima fenilalanina amônia
liase (PAL) apresentou interação significativa as 12, 24 e 72 HAA não sendo observado efeito
significativo as 48 HAA (Tabela 4). Doze horas após a aplicação houve redução da atividade
da PAL com o aumento da dose para a população S, porém o mesmo não foi identificado para
a população R1, pois na dose mais elevada (1080 g e.a. ha-1
), apresentou atividade
significativamente maior. Ao comparar as populações dentro de cada dose, nas doses zero e
720 g e.a. ha-1
a população S apresentou valores superiores em relação à população R1,
porém, na maior dose, houve efeito contrário. Vinte e quatro horas após a aplicação houve
aumento da atividade da enzima fenilalanina amônia liase na dose de 720 g e.a. ha-1
em
relação à testemunha para a população R1 seguida de uma redução na maior dose sendo que
para S, não foi observado o mesmo efeito. Apesar de ambas as populações serem
consideradas suscetíveis de acordo com a curva de dose-resposta, nas duas maiores doses
testadas, a atividade da enzima PAL foi mais elevada para R1 em relação à S. Já as 72 HAA
não houve alteração significativa com o aumento da dose para R1 enquanto que para S a
aplicação de 720 g e.a. ha-1
proporcionou aumento da atividade da enzima PAL em relação à
testemunha seguido de uma redução na maior dose.
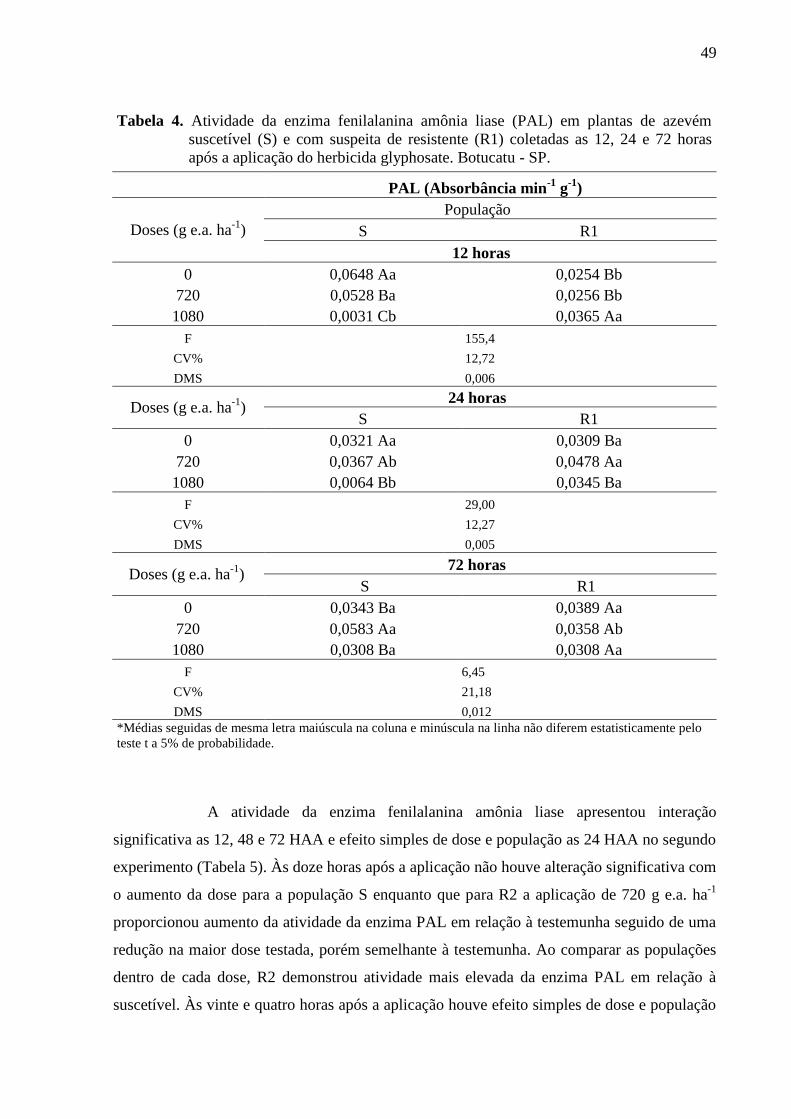
49
Tabela 4. Atividade da enzima fenilalanina amônia liase (PAL) em plantas de azevém
suscetível (S) e com suspeita de resistente (R1) coletadas as 12, 24 e 72 horas
após a aplicação do herbicida glyphosate. Botucatu - SP.
PAL (Absorbância min-1
g-1
)
Doses (g e.a. ha-1
)
População
S R1
12 horas
0 0,0648 Aa 0,0254 Bb
720 0,0528 Ba 0,0256 Bb
1080 0,0031 Cb 0,0365 Aa
F 155,4
CV% 12,72
DMS 0,006
Doses (g e.a. ha-1
) 24 horas
S R1
0 0,0321 Aa 0,0309 Ba
720 0,0367 Ab 0,0478 Aa
1080 0,0064 Bb 0,0345 Ba
F 29,00
CV% 12,27
DMS 0,005
Doses (g e.a. ha-1
) 72 horas
S R1
0 0,0343 Ba 0,0389 Aa
720 0,0583 Aa 0,0358 Ab
1080 0,0308 Ba 0,0308 Aa
F 6,45
CV% 21,18
DMS 0,012
*Médias seguidas de mesma letra maiúscula na coluna e minúscula na linha não diferem estatisticamente pelo
teste t a 5% de probabilidade.
A atividade da enzima fenilalanina amônia liase apresentou interação
significativa as 12, 48 e 72 HAA e efeito simples de dose e população as 24 HAA no segundo
experimento (Tabela 5). Às doze horas após a aplicação não houve alteração significativa com
o aumento da dose para a população S enquanto que para R2 a aplicação de 720 g e.a. ha-1
proporcionou aumento da atividade da enzima PAL em relação à testemunha seguido de uma
redução na maior dose testada, porém semelhante à testemunha. Ao comparar as populações
dentro de cada dose, R2 demonstrou atividade mais elevada da enzima PAL em relação à
suscetível. Às vinte e quatro horas após a aplicação houve efeito simples de dose e população

50
sendo que a suscetível apresentou menor atividade da enzima fenilalanina amônia liase em
relação à resistente e as doses de zero e 720 g e.a. ha-1
foram estatisticamente semelhantes e
superiores à maior dose testada. 48 HAA foi observado aumento da atividade da PAL para
ambas as populações principalmente na dose de 1080 g e.a. ha-1
e novamente como às 12
HAA, à 720 g e.a. ha-1
a resistente apresentou valor superior em relação à suscetível. Já as 72
horas após a aplicação, R2 reduziu a atividade da PAL nas duas maiores doses enquanto que
para a população suscetível, apenas na dose de 1080 g e.a. ha-1
este efeito foi observado. Ao
comparar as populações em cada dose, nota-se que S demonstrou valor superior à 720 g e.a.
ha-1
.

51
Tabela 5. Atividade da enzima fenilalanina amônia liase (PAL) em plantas de azevém
suscetível (S) e Resistente (R1) coletadas as 12, 24, 48 e 72 horas após a
aplicação do herbicida glyphosate. Botucatu - SP.
PAL (Absorbância min-1
g-1
)
Doses (g e.a. ha-1
)
População
S R2
12 horas
0 0,0749 Aa 0,0727 Ba
720 0,0723 Ab 0,0809 Aa
1080 0,0790 Aa 0,0734 Ba F 4,71
CV% 6,41
DMS 0,007
População 24 horas
S 0,0676 B
R2 0,0751 A F 4,74
CV% 11,76
DMS 0,007
Doses (g e.a. ha-1
) 24 horas
0 0,0696 AB
720 0,0778 A
1080 0,0666 B F 3,85
CV% 11,76
DMS 0,008
Doses (g e.a. ha-1
) 48 horas
S R2
0 0,0596 Ba 0,0650 Ba
720 0,0641 Bb 0,0854 Aa
1080 0,0813 Aa 0,0849 Aa F 8,5
CV% 6,46
DMS 0,007
Doses (g e.a. ha-1
) 72 horas
S R2
0 0,0802 Aa 0,0834 Aa
720 0,0807 Aa 0,0612 Bb
1080 0,0626 Ba 0,0578 Ba F 11,31
CV% 6,83
DMS 0,007
*Médias seguidas de mesma letra maiúscula na coluna e minúscula na linha não diferem estatisticamente pelo
teste t a 5% de probabilidade.
Apesar dos sintomas nas plantas tratadas com o herbicida glyphosate
aparecerem alguns dias após a aplicação, o mesmo não é válido quando se trata de enzimas.
Como pode ser observado nas Tabelas 4 e 5, doze horas após a aplicação, o herbicida

52
promoveu mudanças na atividade da enzima PAL. Apesar de ter sido utilizada a mesma
população considerada suscetível (S) para ambos os experimentos, as plantas não
apresentaram o mesmo comportamento após a aplicação do herbicida glyphosate, pois no
primeiro experimento houve redução da atividade da enzima fenilalanina amônia liase com o
aumento da dose, enquanto que no segundo, não foi observado diferenças entre as doses
aplicadas. Este fato pode ser considerado aceitável, pois, apesar de ser a mesma população, o
herbicida foi aplicado em épocas diferentes, ou seja, as plantas não estavam exatamente no
mesmo estádio fenológico. Além disso, como dito anteriormente, num determinado tecido, os
níveis da enzima PAL podem variar significativamente em intervalos relativamente curtos de
tempo em resposta a uma grande variedade de estímulos (CAMM; TOWERS, 1973).
Hoagland (1980), estudando a atividade desta enzima após a aplicação
de glyphosate em soja, demonstrou aumento da atividade entre 12 e 72 horas, porém, foi
utilizado apenas uma dose do herbicida e a soja não era tolerante ao glyphosate. Este
comportamento foi observado em outros trabalhos (DUKE; HOAGLAND, 1978; DUKE;
HOAGLAND; ELMORE, 1979; HOAGLAND; DUKE, 1982). O mesmo não ocorreu neste
experimento, pois ao longo do tempo, houve variação entre os valores de acordo com a dose
aplicada, época avaliada e população envolvida.
Em outro experimento, Hoagland, Duke e Elmore (1978), observaram
redução na atividade da enzima PAL 48 horas após a aplicação de glyphosate sendo esta
acompanhada pela redução das proteínas solúveis bem como os aminoácidos aromáticos
fenilalanina e tirosina em plântulas de milho. Porém quando se aplicou glyphosate em
Hemidesmus indicus, com o aumento da atividade da PAL, houve um amento no teor de
proteína (KUNDU; JAWALI; MITRA, 2012).
Em raízes de soja suscetíveis ao glyphosate, a atividade da PAL foi
significativamente diferente da testemunha com um aumento de aproximadamente 26%,
porém o mesmo não foi observado para a soja tolerante, neste caso, não houve diferença
significativa entre a testemunha e o tratamento (MARSHIOSI et al., 2009). Ainda neste
experimento, em associação com o aumento da atividade da PAL, ocorreu um acúmulo de
ácido chiquímico devido à inibição da EPSPs, resultando numa provável redução dos
aminoácidos aromáticos. De acordo com os autores, este aumento pode ser interpretado como
um efeito provavelmente relacionado com um distúrbio metabólico estressante causado pelo
herbicida ou pela depleção nos níveis dos produtos finais.

53
O aumento da atividade da enzima fenilalanina amônia liase tem sido
observado como um resultado do tratamento com glyphosate em soja (HOAGLAND; DUKE,
1983; HOAGLAND, 1990), porém com plantas consideradas suscetíveis ao herbicida. Tem
sido sugerido que este aumento é uma consequência secundária da redução da síntese de
certos compostos fenólicos que regulam esta enzima (HOLLANDER; AMRHEIN, 1980).
Estes resultados não se repetem para todas as espécies como reportado por outros autores
(HOAGLAND; DUKE; ELMORE, 1978; ISHIKURA; TAKESHIMA, 1984) nos quais
observaram inibição da PAL após exposição a este herbicida, resultados estes diferentes aos
aqui encontrado, pois tanto a população resistente como a suscetível, apresentaram variações
de acordo com a época avaliada e dose aplicada.
O excesso de amônia observado em plantas tratadas com o herbicida
glyphosate pode ser devido ao aumento da atividade da enzima PAL que catalisa a reação que
converte fenilalanina em ácido trans-cinâmico, liberando amônia (HOWLES et al., 1996). Em
experimentos com tabaco transgênico foi comprovado que a redução da quantidade de
aminoácidos aromáticos (efeito direto da ação do glyphosate) resultou num aumento do fluxo
através da via metabólica do chiquimato, o que levou um aumento na atividade da PAL bem
como uma duplicação dos níveis de ácido clorogênico, um composto polifenólico (GUILLET
et al., 2000). A interrupção do crescimento da planta pode ser causada não só pela redução
dos aminoácidos aromáticos, mas também pela toxicidade dos compostos fenólicos derivados
da rota ou pela toxicidade direta ocasionada pelo excesso de amônia (MICHALOWICZ;
DUDA, 2007).
Em experimento realizado por Mobin et al. (2015), foi observado que
com a aplicação de glyphosate, a atividade da PAL foi maior em relação à aplicação apenas
do aminoácido triptofano e em menor escala a fenilalanina, ou seja, o aumento do substrato
desta enzima (fenilalanina) não serviu para aumentar a atividade da mesma; o herbicida teve
uma capacidade maior em estimular tal efeito, fato este observado para a população R2. De
acordo com os autores, este resultado pode ser visto como uma tentativa para aumentar o
fluxo de carbono nesta via. Porém ainda há a necessidade de maiores estudos para tal
confirmação.
A indução de estresses em organismos vegetais promovidos por fatores
bióticos e abióticos resultam em resposta de defesa. A produção dos compostos de defesa é
desencadeada por enzimas no qual a PAL é a primeira e mais importante envolvida neste
processo (PASTÍROVÁ; REPCK; ELIASOVÁ, 2004; YAO; TIAN, 2005). Com a aplicação

54
de glyphosate a planta se encontra em estado de estresse, podendo este, estimular o aumento
da atividade desta enzima com o intuito de acionar os mecanismos de defesa da planta
protegendo assim a mesma. Este comportamento foi evidenciado principalmente para a
população R2. Além do glyphosate, o ataque de vírus em leguminosas promoveu uma rápida
elevação na atividade da PAL, estando esta atividade intimamente ligada ao estádio
fisiológico da planta (NUGROHO; VERBERNE; VERPOORTE, 2002).
Nota-se diferenças entre os valores encontrados por diversos autores no
comportamento da enzima PAL. Nem sempre ocorre um aumento linear da enzima após a
aplicação do glyphosate. Isto pode ser explicado pelas diferentes espécies utilizadas, estádio
fisiológico, momento de aplicação e quantidade do herbicida utilizado. Outro fator que pode
ainda contribuir nessa indefinição é a planta ser resistente ao herbicida. Foi demonstrado neste
experimento que mesmo a população sendo considerada resistente ao herbicida glyphosate,
pode ocorrer alterações significativas na atividade da enzima PAL podendo ser maior ou
menor em relação à testemunha de acordo com a dose aplicada e época avaliada. Porém, não
se pode afirmar se esse comportamento será o mesmo para as outras espécies consideradas
resistentes. Há a necessidade de maiores estudos.
6.3 Efeitos do glyphosate na fotossíntese das diferentes populações
6.3.1 Suscetível (S) x Resistente (R2)
No primeiro dia após a aplicação do herbicida glyphosate, a taxa de
assimilação de CO2 (A) (Figura 16) foi positiva para todos os tratamentos. Isto significa que
um dia não foi suficiente para que o glyphosate afetasse a fotossíntese independentemente da
planta ser sensível ou resistente ao mesmo. Este resultado é aceitável, pois este herbicida
possui uma ação lenta na planta apresentando sintomas dias após a aplicação. Imediatamente
após a sua absorção, o glyphosate é translocado juntamente com os fotoassimilados a partir do
local de aplicação (folhas) para os drenos (RODRIGUES; ALMEIDA, 2011), porém o tempo
necessário para que o mesmo comece a fazer efeitos significativos nas plantas depende de
diversos fatores como a retenção na folha, penetração e translocação para o sítio de ação
(KIRKWOOD; MACKAY, 1994).
Aos 3 DAA apenas as testemunhas (sem aplicação) permaneceram com
valores positivos, ou seja, além das plantas suscetíveis, as resistentes também apresentaram
valores negativos. Neste período, o glyphosate já começou a fazer efeito nas plantas. Este

55
caso indica que especificamente neste momento, as duas populações estavam respirando mais
do que fazendo fotossíntese reduzindo assim a capacidade em acumular massa. O glyphosate
pode afetar a fotossíntese reduzindo a atividade da rubisco e a síntese do ácido 3-
fosfoglicérico diminuindo a síntese de clorofila e também aumentar a taxa de respiração
celular (FLEXAS et al., 2005; AHSAN et al., 2008). A respiração é um processo importante
para as plantas, pois é partir desta reação que se produz energia na forma de ATP utilizando
os carboidratos produzidos na fotossíntese onde o mesmo pode ser utilizado na manutenção e
no desenvolvimento da planta (TAIZ; ZEIGER, 2006). O balanço entre a fotossíntese e a
respiração determina a quantidade de fotoassimilados necessários para o crescimento normal
da planta (GOMIDE; GOMIDE, 1999). Por isso, se algo prejudicar este balanço, pode haver
consequências negativas podendo levar à sua morte. De acordo com Yanniccari et al. (2012),
a redução na assimilação de CO2 é um bioindicador rápido e sensível ao efeito do glyphosate
comparado com outros como a fluorescência da clorofila.
Apesar de aos 3 DAA a população resistente estar respirando mais do
que fazendo a fotossíntese, aos 7 DAA a mesma se recuperou apresentando valores positivos.
Fato este não observado para a população considerada suscetível. Neste caso, a mesma
continuou a apresentar valores negativos. Em plantas suscetíveis, o glyphosate pode inibir a
síntese de clorofila e carotenoides considerados unidade básica fotossintética (ADRIANO et
al., 2013; KASPARY et al., 2014), reduz a assimilação de CO2 (DING et al., 2011) entre
outros. A consequência desses efeitos pode ser notada aos 28 DAA onde a população
suscetível foi completamente controlada pelo herbicida independentemente da dose aplicada
não sendo possível fazer medições. Já a população resistente permaneceu com valores
positivos semelhantes à testemunha resistente.
Resultados semelhantes foram encontrados por Santos et al. (2014), ao
avaliarem os efeitos do glyphosate na fotossíntese em buva resistente e suscetível a este
herbicida. Neste experimento, aos 3 DAA ambos os biótipos apresentaram redução na
assimilação de CO2 em relação à testemunha, resultado este que se estendeu até os 10 DAA,
porém aos 14 DAA apenas o resistente se recuperou. Um fato interessante é que neste
trabalho tanto os biótipos suscetíveis como os resistentes, não apresentaram valores negativos
independentemente da época de avaliação, diferentemente dos resultados aqui verificados.
A redução da assimilação de CO2 na população R2 mesmo que seja em
pouco tempo, pode ser considerada vantajosa em termos práticos, pois além de apresentar
uma redução na produção de carboidratos, ao se encontrar em competição com uma cultura, a

56
planta daninha pode ter seu crescimento reduzido. Dessa forma, a cultura poderá ter um maior
desenvolvimento sombreando assim a infestante dificultando seu crescimento. Silva et al.
(2014), constatou que houve redução na fotossíntese, condutância estomática e transpiração
em soja resistente ao glyphosate devido a interferência de buva resistente ao mesmo. Sendo
assim, qualquer manejo que possa dificultar o desenvolvimento da planta daninha, pode ser
benéfico para a cultura.
Figura 16. Taxa de assimilação de CO2 (μmolCO2 m-2
s-1
) em plantas de azevém suscetível e
resistente após a aplicação do herbicida glyphosate ao 1, 3, 7 e 28 DAA.
Testemunha suscetível (TS), testemunha resistente (TR), população suscetível
submetida a 720 g e.a. ha-1
(S2), população suscetível submetida a 1080 g e.a.
ha-1
(S3), população resistente submetida a 720 g e.a. ha-1
(R2), população
resistente submetida a 1080 g e.a. ha-1
(R3). As barras verticais indicam o
intervalo de confiança. Botucatu – SP.
*nd: não detectável.
Assim como na assimilação de CO2, a condutância estomática não sofreu
alterações significativas 1 DAA (Figura 17). Porém aos 3 DAA houve uma redução significativa
tanto para a população suscetível como a resistente se estendendo até 7 DAA. Yanniccari et al.
0
2
4
6
8
10
12
TS TR S2 S3 R2 R3
Taxa
de
ass
imila
ção
de
CO
₂ (μ
mo
lCO
₂.m
ˉ².s
ˉ¹)
Doses de glyphosate (L haˉ¹)
1 DAA
-20
-15
-10
-5
0
5
10
TS TR S2 S3 R2 R3
Taxa
de
ass
imila
ção
de
CO
₂ (μ
mo
lCO
₂.m
ˉ².s
ˉ¹)
Doses de glyphosate (L haˉ¹)
3 DAA
-5
0
5
10
15
TS TR S2 S3 R2 R3
Taxa
de
ass
imila
ção
de
CO
₂ (μ
mo
lCO
₂.m
ˉ².s
ˉ¹)
Doses de glypgosate (L haˉ¹)
7 DAA
0
2
4
6
8
10
TS TR S2 S3 R2 R3
Taxa
de
ass
imila
ção
de
CO
₂ (μ
mo
lCO
₂.m
ˉ².s
ˉ¹)
Doses de glyphosate (L haˉ¹)
28 DAA
nd* nd

57
(2012), notaram uma redução maior na condutância estomática em biótipos de L. rigidum
suscetíveis ao glyphosate em relação aos resistentes 7 dias após a aplicação do mesmo. A
fotossíntese e a respiração dependem do fluxo constante de CO2 e O2 entrando e saindo na
planta. Este fenômeno acontece através de estruturas denominadas estômatos no qual a abertura
e fechamento são essenciais para o balanço deste fluxo (MESSINGER; BUCKLEY; MOTT,
2006). Quando a planta se encontra em situação de estresse tende a fechar os estômatos como um
mecanismo de defesa contra a perda de água, aumentando assim a resistência, reduzindo a
condutância estomática (TAIZ; ZEIGER, 2006). Este fenômeno pode ser observado após a
aplicação de herbicidas. Alguns trabalhos têm evidenciado este efeito após a aplicação do
glyphosate (ZOBIOLE et al., 2010b; SILVA et al., 2014). Além disso, a condutância estomática
parece ser um biomarcador sensível para a resposta das plantas ao glyphosate (YANNICCARI et
al., 2012).
Zobiole et al. (2009), observaram que o fechamento estomático é um
fator importante que contribui para a assimilação de CO2, fato este observado neste
experimento. A redução da fotossíntese foi acompanhada pela redução na condutância
estomática. O CO2 é uma molécula essencial para que a fotossíntese aconteça, pois juntamente
com a água forma-se carboidratos que serão utilizados pela planta durante sua vida (TAIZ;
ZEIGER, 2006); qualquer fator que contribua para o decréscimo do mesmo afeta diretamente
a fotossíntese. A redução simultânea na condutância estomática e na assimilação de CO2 tem
sido observado por outros autores como Geiger et al. (1986), e Olesen e Cedergreen (2010).
Aos 28 DAA a população resistente apresentou condutância semelhante às testemunhas
demonstrando seu poder de recuperação.
A inibição da fotossíntese e a condutância estomática podem ser
causadas pela acumulação dos produtos finais da fotossíntese (YANNICCARI et al., 2012).
Orcaray et al. (2012), detectou acúmulo de glicose em folhas e raízes de Pisium sativum com
uma resposta rápida ao tratamento do glyphosate antes do declínio da fotossíntese.

58
Figura 17. Condutância estomática (mmolH2O m-2
s-1
) em plantas de azevém suscetível e
resistente após a aplicação do herbicida glyphosate ao 1, 3, 7 e 28 DAA.
Testemunha suscetível (TS), testemunha resistente (TR), população suscetível
submetida a 720 g e.a. ha-1
(S2), população suscetível submetida a 1080 g e.a. ha-1
(S3), população resistente submetida a 720 g e.a. ha-1
(R2), população resistente
submetida a 1080 g e.a. ha-1
(R3). As barras verticais indicam o intervalo de
confiança. Botucatu – SP.
*nd: não detectável.
Observou-se 3 dias após a aplicação de glyphosate que houve um
aumento da concentração interna de CO2 (Ci) nas duas populações onde na suscetível a 1080
g e.a. ha-1
apresentou o maior valor (Figura 18). Este resultado foi semelhante ao encontrado
aos 7 DAA. Os resultados aqui mostrados corroboram com os encontrados por Mateos-
Naranjo et al. (2009) e Ding et al. (2011), nos quais observaram redução da assimilação de
CO2 acompanhado ao aumento da concentração interna de CO2 após a aplicação do herbicida
glyphosate.
Em Abutilon theophrasti medikus, Fuchs et al. (2002), evidenciaram
que a redução da condutância estomática foi acompanhada da redução da Ci após a aplicação
0
0,05
0,1
0,15
0,2
TS TR S2 S3 R2 R3
Co
nd
utâ
nci
a e
sto
mát
ica
(mm
ol(
H2
O).
mˉ²
.sˉ¹
)
Doses de glyphosate (L haˉ¹)
1 DAA
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
TS TR S2 S3 R2 R3
Co
nd
utâ
nci
a e
sto
mát
ica
(mm
ol(
H2
O).
mˉ²
.sˉ¹
)
Doses de glyphosate (L haˉ¹)
3 DAA
0
0,1
0,2
0,3
0,4
TS TR S2 S3 R2 R3Co
nd
utâ
nci
a e
sto
mát
ica
(mm
ol(
H2
O).
mˉ²
.sˉ¹
)
Doses de glyphosate (L haˉ¹)
7 DAA
0
0,05
0,1
0,15
0,2
TS TR S2 S3 R2 R3Co
nd
utâ
nci
a e
sto
mát
ica
(mm
ol(
H2
O).
mˉ²
.sˉ¹
)
Doses de glyphosate (L haˉ¹)
28 DAA
nd* nd

59
de glyphosate onde aos 5 DAA chegou a zero, resultados contrários aos aqui observados. Em
plantas de feijão sob estresse hídrico, Jadoski (2012), mostrou uma correlação negativa entre
condutância estomática e concentração interna de CO2, ou seja, o comportamento dessas
variáveis depende do tipo de estresse no qual a planta sofre e também a espécie envolvida. A
concentração interna de CO2 na folha reflete a disponibilidade de substrato para a fotossíntese
onde durante as trocas gasosas os estômatos regulam essa concentração mantendo-a constante
(FARQUHAR; SHARKEY, 1982). Após sofrer estresse, a planta pode modificar o seu padrão
constante reduzindo ou amentando a Ci. Novamente aos 28 DAA, a população R2 apresentou
valores semelhantes à testemunha.
Figura 18. Concentração interna de CO2 na câmara subestomática (μmol(CO2) mol-1
ar) em
plantas de azevém suscetível e resistente após a aplicação do herbicida glyphosate
ao 1, 3, 7 e 28 DAA. Testemunha suscetível (TS), testemunha resistente (TR),
população suscetível submetida a 720 g e.a. ha-1
(S2), população suscetível
submetida a 1080 g e.a. ha-1
(S3), população resistente submetida a 720 g e.a. ha-1
(R2), população resistente submetida a 1080 g e.a. ha-1
(R3). As barras verticais
indicam o intervalo de confiança. Botucatu – SP.
*nd: não detectável.
0
100
200
300
400
TS TR S2 S3 R2 R3Co
nce
ntr
ação
inte
rna
de
C
O₂
(μm
ol(
CO
₂).m
olˉ
¹ar)
Doses de glyphosate (L haˉ¹)
1 DAA
0
200
400
600
800
TS TR S2 S3 R2 R3Co
nce
ntr
ação
inte
rna
de
C
O₂
(μm
ol(
CO
₂).m
olˉ
¹ar)
Doses de glyphosate (L haˉ¹)
3 DAA
0
200
400
600
TS TR S2 S3 R2 R3Co
nce
ntr
ação
inte
rna
de
C
O₂
(μm
ol(
CO
₂).m
olˉ
¹ar)
Doses de glyphosate (L haˉ¹)
7 DAA
0
100
200
300
400
TS TR S2 S3 R2 R3Co
nce
ntr
ação
inte
rna
de
C
O₂
(μm
ol(
CO
₂).m
olˉ
¹ar)
Doses de glyphosate (L haˉ¹)
28 DAA
nd* nd

60
O glyphosate reduziu drasticamente a taxa de transpiração na população
suscetível aos 3 DAA em relação à testemunha (Figura 19). Porém o mesmo não foi observado
para a população resistente. Este comportamento se manteve até os 7 DAA onde aos 28 DAA a
população R2 apresentou valores semelhantes à testemunha. A transpiração é o processo de
perda de água pelo vegetal na forma de vapor através dos poros estomáticos onde ao mesmo
tempo permitem a entrada de CO2 (VAVASSEUR; RAGHAVENDRA, 2005). Dependendo do
tipo de estresse, a planta fecha os estômatos com o intuito de evitar a perda de água. Como foi
observado na Figura 17, após a aplicação do glyphosate ocorreu redução da condutância
estomática, com isso, os estômatos fecharam, dificultando a saída de água reduzindo assim a
transpiração.
Resultados encontrados por Zobiole et al. (2010b), também mostraram
uma redução na taxa de transpiração juntamente com a redução na condutância estomática em
soja resistente ao glyphosate após a aplicação do mesmo. Santos et al. (2014) mostraram que a
buva resistente ao glyphosate apresentou redução na taxa de transpiração porém apresentou
maiores valores que a suscetível em todos os períodos avaliados. A redução na taxa de
transpiração após a aplicação de glyphosate também foi observada por outros autores
(SHANER, 1978; SHANER; LYON, 1980; VARGAS et al, 2014).

61
Figura 19. Taxa de transpiração (μmol(H2O) m-2
s-1
) em plantas de azevém suscetível e
resistente após a aplicação do herbicida glyphosate ao 1, 3, 7 e 28 DAA.
Testemunha suscetível (TS), testemunha resistente (TR), população suscetível
submetida a 720 g e.a. ha-1
(S2), população suscetível submetida a 1080 g e.a. ha-1
(S3), população resistente submetida a 720 g e.a. ha-1
(R2), população resistente
submetida a 1080 g e.a. ha-1
(R3). As barras verticais indicam o intervalo de
confiança. Botucatu – SP.
*nd: não detectável.
A eficiência do uso da água (EUA) teve um padrão semelhante à
assimilação de CO2 (A) após a aplicação de glyphosate para as duas populações e em todas as
épocas avaliadas (Figura 20). A EUA é a quantidade de carbono fixado no processo
fotossintético pela quantidade de água perdida no processo de transpiração, ou seja, matéria
seca produzida por grama de água transpirada. Este fato explica o porquê desta variável
apresentar valores negativos. Sob condições normais, a planta abre seus estômatos permitindo
a saída de água na forma de vapor e ao mesmo tempo ocorre a entrada de CO2 no qual será
utilizado para a formação de carboidratos. Porém foi mostrado anteriormente que após a
aplicação do glyphosate houve uma redução na assimilação de CO2 e transpiração para ambas
0
2
4
6
8
TS TR S2 S3 R2 R3
Taxa
de
tra
nsp
iraç
ão
(mm
ol(
H₂0
).m
ˉ².s
ˉ¹)
Doses de glyphosate (L haˉ¹)
1 DAA
0
2
4
6
8
10
TS TR S2 S3 R2 R3
Taxa
de
tra
nsp
iraç
ão
(mm
ol(
H₂O
).m
ˉ².s
ˉ¹)
Doses de glyphosate (L haˉ¹)
3 DAA
0
1
2
3
4
5
6
TS TR S2 S3 R2 R3
Taxa
de
tra
nsp
iraç
ão
(mm
ol(
H₂O
).m
ˉ².s
ˉ¹)
Doses de glyphosate (L haˉ¹)
7 DAA
00,5
11,5
22,5
33,5
4
TS TR S2 S3 R2 R3
Taxa
de
tra
nsp
iraç
ão
(mm
ol(
H₂O
).m
ˉ².s
ˉ¹)
Doses de glyphosate (L haˉ¹)
28 DAA
nd* nd

62
as populações, mas a resistente demonstrou seu poder de recuperação. Assim como o
esperado, as plantas consideradas suscetíveis foram mortas sendo que aos 3 DAA já
apresentaram valores negativos especialmente à 1080 g e.a. ha-1
. Já a população resistente
apesar de exibirem valores negativos aos 3 DAA, aos 7 DAA se recuperaram.
Vargas et al. (2014), não encontraram diferenças na EUA três dias após
a aplicação de glyphosate entre os biótipos de buva suscetíveis e resistentes ao mesmo. Porém
sete dias após houve uma grande diferença sendo que o biótipo resistente demonstrou maior
EUA mantendo valores significativamente iguais à testemunha até 14 dias após a aplicação.
Zobiole et al. (2010b), concluíram que soja resistente ao glyphosate reduziu a eficiência do
uso da água após a aplicação deste herbicida. Apesar de demonstrar redução da EUA, aos 28
DAA o azevém resistente apresentou valores semelhantes à testemunha. O uso eficiente da
água está diretamente relacionado ao tempo de abertura estomática, pois enquanto a planta
absorve CO2 a água é perdida por transpiração com intensidade variável seguindo uma
corrente de potenciais hídricos entre a folha e a atmosfera (PEREIRA-NETO, 2002). Ao
longo das avaliações, a população resistente assimilou CO2 e transpirou mais que a população
suscetível apresentando assim uma maior EUA.
Os dados apresentados neste experimento foram negativos para esta
variável apesar dos valores encontrados por outros autores terem sido positivos.
Provavelmente, este fato se deve ao grau de suscetibilidade e o tipo de espécie envolvida.
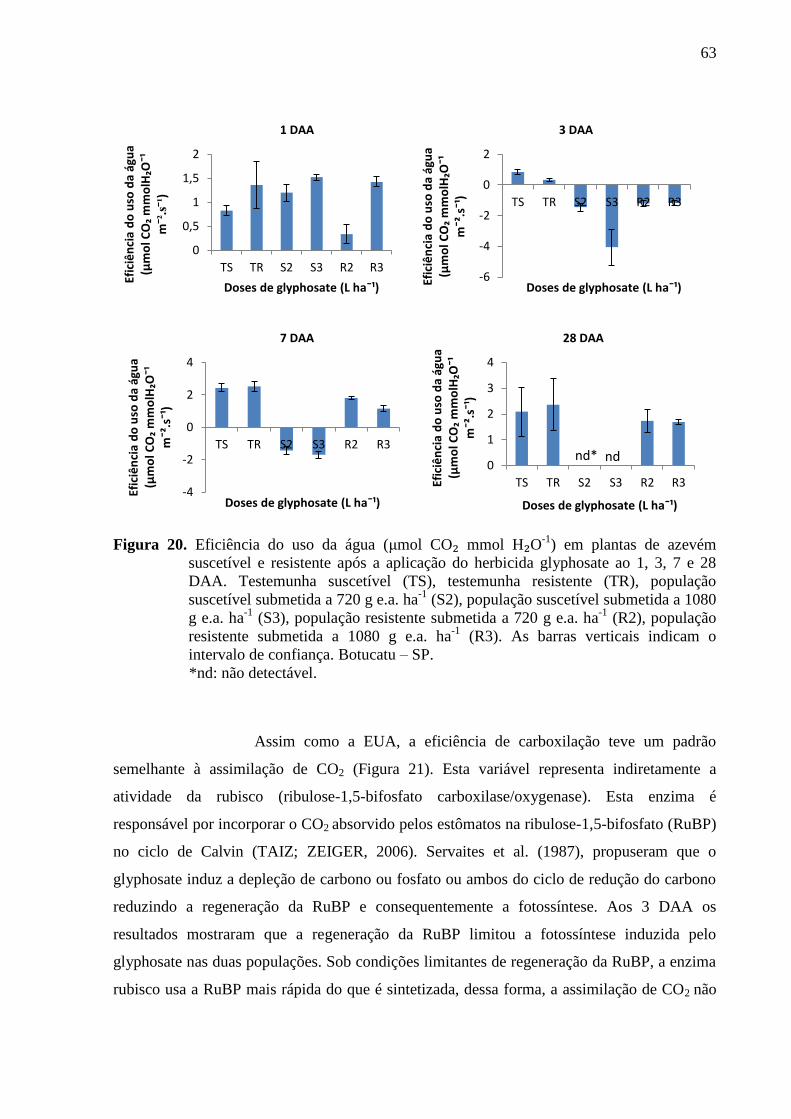
63
Figura 20. Eficiência do uso da água (μmol CO₂ mmol H₂O-1) em plantas de azevém
suscetível e resistente após a aplicação do herbicida glyphosate ao 1, 3, 7 e 28
DAA. Testemunha suscetível (TS), testemunha resistente (TR), população
suscetível submetida a 720 g e.a. ha-1
(S2), população suscetível submetida a 1080
g e.a. ha-1
(S3), população resistente submetida a 720 g e.a. ha-1
(R2), população
resistente submetida a 1080 g e.a. ha-1
(R3). As barras verticais indicam o
intervalo de confiança. Botucatu – SP.
*nd: não detectável.
Assim como a EUA, a eficiência de carboxilação teve um padrão
semelhante à assimilação de CO2 (Figura 21). Esta variável representa indiretamente a
atividade da rubisco (ribulose-1,5-bifosfato carboxilase/oxygenase). Esta enzima é
responsável por incorporar o CO2 absorvido pelos estômatos na ribulose-1,5-bifosfato (RuBP)
no ciclo de Calvin (TAIZ; ZEIGER, 2006). Servaites et al. (1987), propuseram que o
glyphosate induz a depleção de carbono ou fosfato ou ambos do ciclo de redução do carbono
reduzindo a regeneração da RuBP e consequentemente a fotossíntese. Aos 3 DAA os
resultados mostraram que a regeneração da RuBP limitou a fotossíntese induzida pelo
glyphosate nas duas populações. Sob condições limitantes de regeneração da RuBP, a enzima
rubisco usa a RuBP mais rápida do que é sintetizada, dessa forma, a assimilação de CO2 não
0
0,5
1
1,5
2
TS TR S2 S3 R2 R3
Efic
iên
cia
do
uso
da
águ
a (μ
mo
l CO
₂ m
mo
lH₂O
ˉ¹
mˉ².sˉ¹
)
Doses de glyphosate (L haˉ¹)
1 DAA
-6
-4
-2
0
2
TS TR S2 S3 R2 R3
Efic
iên
cia
do
uso
da
águ
a (μ
mo
l CO
₂ m
mo
lH₂O
ˉ¹
mˉ²
.sˉ¹
)
Doses de glyphosate (L haˉ¹)
3 DAA
-4
-2
0
2
4
TS TR S2 S3 R2 R3
Efic
iên
cia
do
uso
da
águ
a (μ
mo
l CO
₂ m
mo
lH₂O
ˉ¹
mˉ²
.sˉ¹
)
Doses de glyphosate (L haˉ¹)
7 DAA
0
1
2
3
4
TS TR S2 S3 R2 R3Efic
iên
cia
do
uso
da
águ
a (μ
mo
l CO
₂ m
mo
lH₂O
ˉ¹
mˉ²
.sˉ¹
)
Doses de glyphosate (L haˉ¹)
28 DAA
nd* nd

64
responde ao aumento da Ci (CAEMMERER; FARQUHAR, 1981). Este fato pode ser
observado neste experimento, pois, após a aplicação do herbicida glyphosate, houve um
amento da concentração interna de CO2 (Figura 18), porém a planta não aproveitou este
acontecimento sendo que A reduziu após a aplicação do mesmo (Figura 16).
Aos 7 DAA a população resistente voltou a ter valores positivos porém
bem menores que a testemunha especialmente a 1080 g e.a. ha-1
. Mesmo a população sendo
resistente, o glyphosate reduziu a eficiência de carboxilação. Resultados encontrados por De
Maria et al. (2006) em folhas de Lupinus Albus, mostraram uma redução de 26% na atividade
da rubisco cinco dias após a aplicação de glyphosate. Zobiole et al. (2010a), também
encontrou redução da atividade da rubisco em soja resistente ao glyphosate após a aplicação
do mesmo, com isso, houve redução da biomassa, pois a difusão do CO2 para o cloroplasto é
essencial para a fotossíntese. Aos 28 DAA novamente a população resistente mostrou
recuperação apresentando valores próximos à testemunha.
Uma das hipóteses que tem avançado para explicar o efeito do
glyphosate no metabolismo do carbono está relacionada ao fluxo de carbono para a via do
ácido chiquímico devido a desregulação da mesma. O efeito causado pelo glyphosate nesta
via conduz a uma acumulação de ácido chiquímico se tornando um dreno de carbono (DUKE;
POWLES, 2008). O arogenato é subproduto do corismato e inibidor da DAHP sintase,
primeira enzima da via do chiquimato (SIEHL, 1997). Segundo este autor, a inibição da
síntese de DAHP sintase pelo arogenato é a chave na regulação da via do chiquimato. Com a
inibição da enzima EPSPs causada pelo glyphosate há uma interferência na entrada do
carbono na via do chiquimato devido ao aumento da atividade da DAHP sintase (DEVINE;
BANDEEN; MCKERSIE, 1983). O aumento desta enzima se deve aos baixos níveis de
arogenato, pois a produção do seu precursor foi reduzida, assim a enzima continua atuando
provocando altos níveis de ácido chiquímico. A quantidade de ácido chiquímico acumulado
pela desregulação da rota representa um forte dreno de carbono devido ao desvio da eritrose-
4-fosfato que seria empregado na regeneração da ribulose-1,5-bifosfato reduzindo
drasticamente a fotossíntese (GEIGER et a., 1986; SERVAITES et al., 1987; SIEHL, 1997).

65
Figura 21. Eficiência de carboxilação (A/Ci) em plantas de azevém suscetível e resistente
após a aplicação do herbicida glyphosate ao 1, 3, 7 e 28 DAA. Testemunha
suscetível (TS), testemunha resistente (TR), população suscetível submetida a
720 g e.a. ha-1
(S2), população suscetível submetida a 1080 g e.a. ha-1
(S3),
população resistente submetida a 720 g e.a. ha-1
(R2), população resistente
submetida a 1080 g e.a. ha-1
(R3). As barras verticais indicam o intervalo de
confiança. Botucatu – SP.
*nd: não detectável.
Como esperado, o herbicida glyphosate causou danos irreversíveis à
população suscetível onde aos 28 DAA em todas as variáveis analisadas a mesma já se
encontrava morta impossibilitando assim sua análise, porém foi mostrado que os efeitos deste
herbicida foram intensificados a partir do terceiro dia após a aplicação. No caso da população
resistente, aos 3 DAA nas variáveis analisadas, sofreu efeitos significativos em relação à
testemunha mostrando que mesmo apresentando um elevado grau de resistência, o herbicida é
capaz de afetar seu metabolismo. Porém, a partir dos 7 DAA a mesma evidenciou seu poder
de recuperação onde aos 28 DAA apresentou valores semelhantes à testemunha, ou seja,
00,005
0,010,015
0,020,025
0,030,035
0,04
TS TR S2 S3 R2 R3
Efic
iên
cia
de
car
bo
xila
ção
(
A/C
i)
Doses de glyphosate (L haˉ¹)
1 DAA
-0,03
-0,02
-0,01
0
0,01
0,02
0,03
TS TR S2 S3 R2 R3
Efic
iên
cia
de
car
bo
xila
ção
(A
/Ci)
Doses de glyphosate (L haˉ¹)
3 DAA
-0,02
-0,01
0
0,01
0,02
0,03
0,04
0,05
TS TR S2 S3 R2 R3
Efic
iên
cia
de
car
bo
xila
ção
(A
/Ci)
Doses de glyphosate (L haˉ¹)
7 DAA
0
0,005
0,01
0,015
0,02
0,025
0,03
TS TR S2 S3 R2 R3
Efic
iên
cia
de
car
bo
xila
ção
(A
/Ci)
Doses de glyphosate (L haˉ¹)
28 DAA
nd* nd

66
mesmo após a aplicação do glyphosate, estas plantas foram capazes de retornar a um estado
fisiológico semelhante àquelas que não receberam o produto.
Os resultados aqui encontrados dependendo da variável corroboraram
com alguns autores, porém diferiram de outros. Fatores como espécie envolvida, momento de
aplicação, grau de suscetibilidade das plantas, quantidade do herbicida aplicado entre outros,
podem explicar tal fato.
6.3.2 Suscetível (S) x Resistente (R1)
Como foi observado no experimento de curva de dose-resposta, essas
duas populações foram consideradas suscetíveis ao herbicida glyphosate. Em todas as
variáveis fisiológicas analisadas, ambas apresentaram o mesmo comportamento comprovando
assim o seu grau de suscetibilidade sendo que aos 28 DAA todas as plantas estavam mortas
impossibilitando assim a sua análise (Figuras 22, 23, 24, 25, 26 e 27). Apesar das populações
S e R1 serem consideradas suscetíveis, com o aumento da pressão de seleção ocasionado pelo
uso constante do herbicida glyphosate, a probabilidade de ocorrência da resistência é maior na
população R1 de acordo com os dados mostrados no experimento de curva de dose-resposta.

67
Figura 22. Taxa de assimilação de CO2 (μmolCO2 m-2
s-1
) em plantas de azevém suscetível e
resistente após a aplicação do herbicida glyphosate ao 1, 7 e 28 DAA.
Testemunha suscetível (TS), testemunha resistente (TR), população suscetível
submetida a 720 g e.a. ha-1
(S2), população suscetível submetida a 1080 g e.a.
ha-1
(S3), população resistente submetida a 720 g e.a. ha-1
(R2), população
resistente submetida a 1080 g e.a. ha-1
(R3). As barras verticais indicam o
intervalo de confiança. Botucatu – SP.
*nd: não detectável.
0
5
10
15
20
TS TR S2 S3 R2 R3Taxa
de
ass
imila
ção
de
CO
₂ (μ
mo
lCO
₂ m
ˉ² s
ˉ¹)
Doses de glyphosate (L haˉ¹)
1 DAA
-20
-15
-10
-5
0
5
10
15
TS TR S2 S3 R2 R3
Taxa
de
ass
imila
ção
de
CO
₂ (μ
mo
lCO
₂ m
ˉ² s
ˉ¹)
Doses de glyphosate (L haˉ¹)
7 DAA
0
2
4
6
8
TS TR S2 S3 R2 R3
Taxa
de
ass
imila
ção
de
CO
₂ (μ
mo
lCO
₂ m
ˉ² s
ˉ¹)
Doses de glyphosate (L haˉ¹)
28 DAA
nd* nd nd nd

68
Figura 23. Condutância estomática (mmolH2O m-2
s-1
) em plantas de azevém suscetível e
resistente após a aplicação do herbicida glyphosate ao 1, 7 e 28 DAA. Testemunha
suscetível (TS), testemunha resistente (TR), população suscetível submetida a 720
g e.a. ha-1
(S2), população suscetível submetida a 1080 g e.a. ha-1
(S3), população
resistente submetida a 720 g e.a. ha-1
(R2), população resistente submetida a 1080
g e.a. ha-1
(R3). As barras verticais indicam o intervalo de confiança. Botucatu –
SP.
*nd: não detectável.
0
0,1
0,2
0,3
0,4
TS TR S2 S3 R2 R3
Co
nd
utâ
nci
a e
sto
mát
ica
(mm
ol(
H2
O)
mˉ²
.sˉ¹
)
Doses de glyphosate (L haˉ¹)
1 DAA
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
TS TR S2 S3 R2 R3
Co
nd
utâ
nci
a e
sto
mát
ica
(mm
ol(
H2
O)
mˉ²
.sˉ¹
)
Doses de glyphosate (L haˉ¹)
7 DAA
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
TS TR S2 S3 R2 R3
Co
nd
utâ
nci
a e
sto
mát
ica
(mm
ol(
H2
O)
mˉ²
.sˉ¹
)
Doses de glyphosate (L haˉ¹)
28 DAA
nd* nd nd nd

69
Figura 24. Concentração interna de CO2 na câmara subestomática (μmol(CO2) mol-1
ar) em
plantas de azevém suscetível e resistente após a aplicação do herbicida glyphosate
ao 1, 7 e 28 DAA. Testemunha suscetível (TS), testemunha resistente (TR),
população suscetível submetida a 720 g e.a. ha-1
(S2), população suscetível
submetida a 1080 g e.a. ha-1
(S3), população resistente submetida a 720 g e.a. ha-1
(R2), população resistente submetida a 1080 g e.a. ha-1
(R3). As barras verticais
indicam o intervalo de confiança. Botucatu – SP.
*nd: não detectável.
0
100
200
300
400
TS TR S2 S3 R2 R3Co
nce
ntr
ação
inte
rna
de
CO
₂ (μ
mo
l(C
O₂)
.mo
lˉ¹a
r)
Doses de glyphosate (L haˉ¹)
1 DAA
0
100
200
300
400
500
600
700
TS TR S2 S3 R2 R3
Co
nce
ntr
ação
inte
rna
de
CO
₂ (μ
mo
l(C
O₂)
.mo
lˉ¹a
r)
Doses de glyphosate (L haˉ¹)
7 DAA
0
100
200
300
400
500
TS TR S2 S3 R2 R3
Co
nce
ntr
ação
inte
rna
de
CO
₂ (μ
mo
l(C
O₂)
.mo
lˉ¹a
r)
Doses de glyphosate (L haˉ¹)
28 DAA
nd* nd nd nd

70
Figura 25. Taxa de transpiração (mmolH2O m-2
s-1
) em plantas de azevém suscetível e
resistente após a aplicação do herbicida glyphosate ao 1, 7 e 28 DAA. Testemunha
suscetível (TS), testemunha resistente (TR), população suscetível submetida a 720
g e.a. ha-1
(S2), população suscetível submetida a 1080 g e.a. ha-1
(S3), população
resistente submetida a 720 g e.a. ha-1
(R2), população resistente submetida a 1080
g e.a. ha-1
(R3). As barras verticais indicam o intervalo de confiança. Botucatu –
SP.
*nd: não detectável.
0
2
4
6
8
10
TS TR S2 S3 R2 R3
Taxa
de
tra
nsp
iraç
ão
(mm
ol(
H₂0
).m
ˉ².s
ˉ¹)
Doses de glyphosate (L haˉ¹)
1 DAA
0
2
4
6
8
10
TS TR S2 S3 R2 R3
Taxa
de
tra
nsp
iraç
ão
(mm
ol(
H₂0
).m
ˉ².s
ˉ¹)
Doses de glyphosate (L haˉ¹)
7 DAA
0
2
4
6
TS TR S2 S3 R2 R3
Taxa
de
tra
nsp
iraç
ão
(mm
ol(
H₂0
).m
ˉ².s
ˉ¹)
Doses de glyphosate (L haˉ¹)
28 DAA
nd* nd nd nd

71
Figura 26. Eficiência do uso da água (μmol CO₂ mmol H₂O-1) em plantas de azevém
suscetível e resistente após a aplicação do herbicida glyphosate ao 1, 7 e 28 DAA.
Testemunha suscetível (TS), testemunha resistente (TR), população suscetível
submetida a 720 g e.a. ha-1
(S2), população suscetível submetida a 1080 g e.a. ha-1
(S3), população resistente submetida a 720 g e.a. ha-1
(R2), população resistente
submetida a 1080 g e.a. ha-1
(R3). As barras verticais indicam o intervalo de
confiança. Botucatu – SP.
*nd: não detectável.
0
2
4
6
8
TS TR S2 S3 R2 R3Efic
iên
cia
do
uso
da
águ
a (μ
mo
l C
O₂
mm
olH
₂Oˉ¹
mˉ²
.sˉ¹
)
Doses de glyphosate (L haˉ¹)
1 DAA
-8
-6
-4
-2
0
2
4
TS TR S2 S3 R2 R3
Efic
iên
cia
do
uso
da
águ
a (μ
mo
l C
O₂
mm
olH
₂Oˉ¹
mˉ²
.sˉ¹
)
Doses de glyphosate (L haˉ¹)
7 DAA
0
1
2
3
TS TR S2 S3 R2 R3
Efic
iên
cia
do
uso
da
águ
a
(μm
ol C
O₂
mm
olH
₂Oˉ¹
mˉ²
.sˉ¹
)
Doses de glyphosate (L haˉ¹)
28 DAA
nd* nd nd nd

72
Figura 27. Eficiência de carboxilação (A/Ci) em plantas de azevém suscetível e resistente
após a aplicação do herbicida glyphosate ao 1, 7 e 28 DAA. Testemunha
suscetível (TS), testemunha resistente (TR), população suscetível submetida a
720 g e.a. ha-1
(S2), população suscetível submetida a 1080 g e.a. ha-1
(S3),
população resistente submetida a 720 g e.a. ha-1
(R2), população resistente
submetida a 1080 g e.a. ha-1
(R3). As barras verticais indicam o intervalo de
confiança. Botucatu – SP.
*nd: não detectável.
0
0,02
0,04
0,06
0,08
0,1
TS TR S2 S3 R2 R3
Efic
iên
cia
de
car
bo
xila
ção
(A
/Ci)
Doses de glyphosate (L haˉ¹)
1 DAA
-0,04
-0,02
0
0,02
0,04
0,06
TS TR S2 S3 R2 R3
Efic
iên
cia
de
car
bo
xila
ção
(A
/Ci)
Doses de glyphosate (L haˉ¹)
7 DAA
0
0,005
0,01
0,015
0,02
0,025
TS TR S2 S3 R2 R3
Efic
iên
cia
de
car
bo
xila
ção
(
A/C
i)
Doses de glyphosate (L haˉ¹)
28 DAA
nd* nd nd nd

73
6.4 Avaliação dos teores de glyphosate, AMPA e compostos da rota do ácido
chiquímico nas diferentes populações
Na Tabela 6 encontra-se a análise de variância dos dados e a diferença
mínima significativa entre as médias de cada um dos compostos analisados.
Tabela 6. Análise de variância e valores de diferença mínima significativa (DMS) para os
compostos analisados. Botucatu – SP.
5
DAA
11
DAA
28
DAA
5
DAA
11
DAA
28
DAA
5
DAA
11
DAA
28
DAA
Fenilalanina Tirosina Triptofano
CV (%) 18,72 10,08 15,91 14,95 14,24 14,72 14,57 12,44 16,17
F 9,84* 120,31* 74,18* 7,77* 36,63* 38,43* 4,00* 72,67* 43,85*
DMS 0,95 0,38 0,53 0,67 0,46 0,43 0,84 0,47 0,53
Glyphosate AMPA Ác. Chiquímico
CV (%) 34 133,38 55,53 50,18 203,30 134,44 10,33 21,78 16,37
F 3,12* 8,16* 37,49* 7,79* 9,12* 27.43* 8,34* 78,81* 168,7*
DMS 0,61 1,13 0,24 0,31 0,07 0,02 1,95 1,84 0,85
Ác. Quínico Ác. Ferúlico Ác. Cafeico
CV (%) 8,99 12,36 15,81 15,17 14,48 15,98 22,95 15,77 16,85
F 10,24* 209,73* 146,90* 11,30* 147,19* 187,95* 2,98* 30,57* 59,98*
DMS 1,65 1,05 0,89 0,52 0,46 0,41 0,28 0,10 0,11
Ác. Cumárico Shiquimato-3-fosfato Ác. Desidroshiquímico
CV (%) 20,27 43,26 63,31 28,36 40,42 27,35 41,91 13,93 16,26
F 2,86* 4,84* 2,50* 13,32* 12,14* 19,79* 3,43* 70,93* 68,96*
DMS 0,10 0,12 0,14 0,79 0,74 0,31 0,69 0,11 0,10
*Interação significativa.
Os níveis de glyphosate nas folhas apresentaram comportamento
semelhante nas três populações até a dose recomendada (100%) aos 5 DAA (Figura 28).
Observou-se um aumento dos teores de glyphosate entre as populações onde a R2 apresentou
maiores valores em relação à S e R1 nas doses subsequentes à recomendada. A absorção de
glyphosate nas plantas tem sido reportada em diversas espécies no qual geralmente é rápida
variando de 40% a 80% do total de herbicida aplicado dependendo da espécie envolvida
(LORRAINE-COLWILL et al., 2002). Hetherington et al. (1999), observaram que o milho

74
tolerante ao glyphosate absorveu 10% mais herbicida que o suscetível na maior dose testada.
Porém não houve diferença na absorção de glyphosate entre plantas de Beta vulgaris resistentes
e suscetíveis ao mesmo (GEIGER; SHIEH; FUCHS, 1999). Gomes (2014), testando várias
populações de Conyza spp., observou que a população considerada suscetível apresentou maiores
teores de glyphosate nas menores doses. Em Lolium rigidum Feng et al. (1999), não encontraram
diferenças na absorção entre os biótipos resistentes e suscetíveis ao glyphosate.
Aos 11 DAA na dose recomendada, a população S já estava morta não
sendo possível mensurar tal variável. Novamente, nesta dose não houve diferença entre as
populações R1 e R2, sendo que nas doses subsequentes devido ao seu grau de resistência,
xapenas na R2 foi possível medir os níveis de glyphosate. Observou-se que aos 28 DAA
somente para R2 foi possível detecta-lo nas folhas. Sob condições de temperatura mais altas,
no biótipo de buva resistente ao glyphosate, foi detectado 85% do total aplicado no vacúolo
enquanto que no suscetível foi de 15%, ou seja, o resistente foi mais eficiente em transportar o
herbicida através da membrana do tonoplasto, porém em temperaturas mais frias, houve
redução na eficiência do transporte para a população resistente (GE et al., 2011).
Ressalta-se que os valores de glyphosate mensurados aos 28 DAA
foram menores em relação às avaliações anteriores para a população R2. Possivelmente, a
planta já utilizou do seu mecanismo de resistência reduzindo assim a concentração de
glyphosate nas folhas.
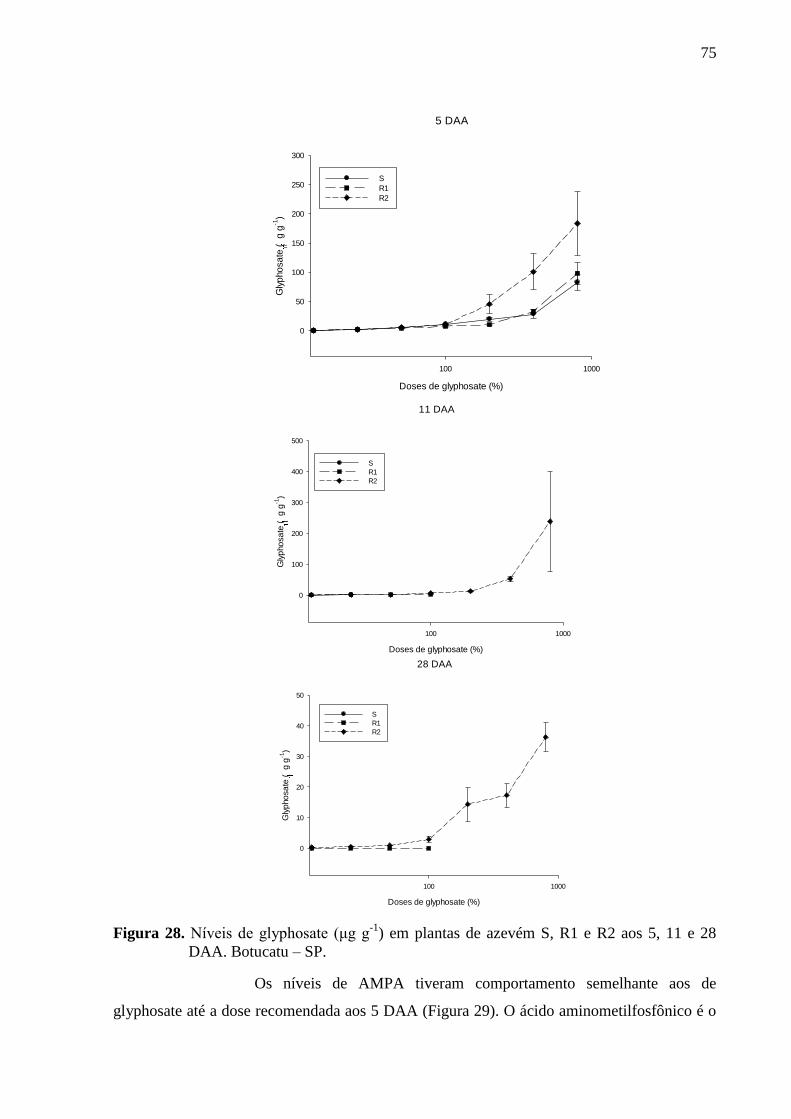
75
5 DAA
Doses de glyphosate (%)
100 1000
Gly
pho
sa
te (
g g
-1)
0
50
100
150
200
250
300
S
R1
R2
11 DAA
Doses de glyphosate (%)
100 1000
Gly
pho
sa
te (
g g
-1)
0
100
200
300
400
500
S
R1
R2
28 DAA
Doses de glyphosate (%)
100 1000
Gly
pho
sa
te (
g g
-1)
0
10
20
30
40
50
S
R1
R2
Figura 28. Níveis de glyphosate (μg g-1
) em plantas de azevém S, R1 e R2 aos 5, 11 e 28
DAA. Botucatu – SP.
Os níveis de AMPA tiveram comportamento semelhante aos de
glyphosate até a dose recomendada aos 5 DAA (Figura 29). O ácido aminometilfosfônico é o

76
principal metabólito do herbicida glyphosate sendo detectado naturalmente em plantas
tolerantes ao mesmo como Agropyron repens e Equisetum arvense (MONQUERO et al.,
2004). A reação que gera este produto, neste caso em folhas, pode ser catalisada pela enzima
glyphosate oxiredutase (GOX) podendo o mesmo ser tóxico inclusive para plantas que
possuem EPSPs insensível ao glyphosate (NANDULA et al., 2007).
Nas doses acima da recomendada a população R2 apresentou os
menores valores sendo próximos a zero. Este fato ajuda a explicar a resistência desta
população. O AMPA é um produto tóxico para a planta embora seja considerado menos ativo
que o glyphosate, quanto maior a sua quantidade, mais prejudicial é para a planta (REDDY et
al., 2004). A população R2, mesmo sendo considerada resistente, não metabolizou o
glyphosate em AMPA, ou seja, neste caso o mesmo permaneceu como molécula intacta,
porém, não houve morte das plantas. Provavelmente, o mecanismo de resistência envolvido,
não está relacionado à diferença no metabolismo, pois mesmo apresentando maior quantidade
de glyphosate em relação às populações suscetíveis, a população resistente não metabolizou
no seu produto.
Reddy et al. (2008) sete dias após a aplicação do herbicida glyphosate
observaram diferenças nos níveis de AMPA em soja tolerante ao glyphosate em relação à
suscetível onde esta apresentou os maiores valores. Porém, teve espécies no qual não foi
detectado este metabólito como o caso de Lolium multiflorum e também em milho tolerante e
suscetível ao glyphosate. Gonzales-Torralva et al. (2012b), observaram diferenças no
metabolismo do glyphosate entre biótipos de C. canadensis no qual detectaram AMPA 72
horas após a aplicação nos biótipos resistentes enquanto que nos suscetíveis não foi detectado,
ou seja, o resistente possuiu um metabolismo mais rápido reduzindo assim a quantidade de
glyphosate que alcançaria o sítio de ação.
Aos 11 e 28 DAA na dose recomendada, os níveis de AMPA
encontrados foi zero para as populações R1 e R2 sendo possível detectar o composto apenas
nas doses subsequentes a 100% para a população considerada resistente, porém em
quantidade muito inferior às visualizadas aos 5 DAA. Em algumas espécies o glyphosate é
metabolizado sendo em outras, encontrado como uma molécula intacta. A inabilidade em
detectar metabólitos nas plantas não significa que não houve metabolismo. A perda do
glyphosate não significa degradação, pois este herbicida é conhecido por ser exsudado das
raízes e folhas tratadas na forma de molécula intacta (RODRIGUES et al., 1982;
MONQUERO et al., 2004). As diferenças nos resultados descobertos por diversos autores
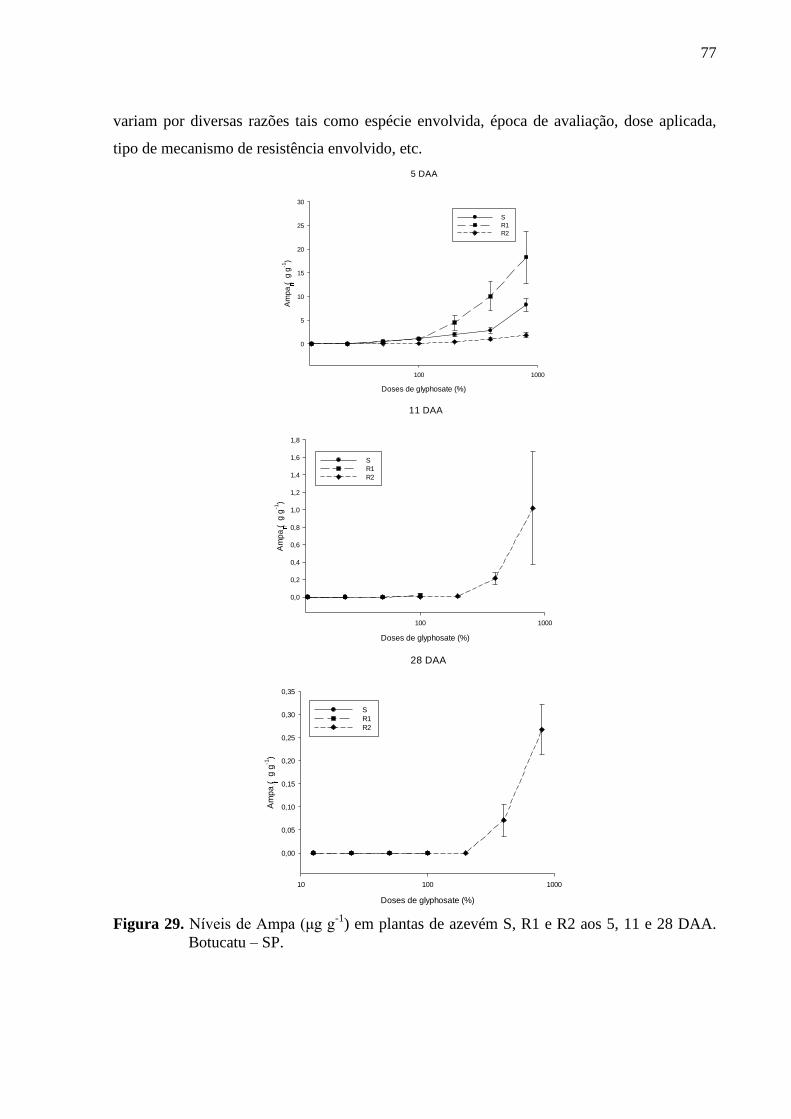
77
variam por diversas razões tais como espécie envolvida, época de avaliação, dose aplicada,
tipo de mecanismo de resistência envolvido, etc.
5 DAA
Doses de glyphosate (%)
100 1000
Am
pa
(g
g-1
)
0
5
10
15
20
25
30
S
R1
R2
11 DAA
Doses de glyphosate (%)
100 1000
Am
pa
(g
g-1
)
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
S
R1
R2
28 DAA
Doses de glyphosate (%)
10 100 1000
Am
pa
(g
g-1
)
0,00
0,05
0,10
0,15
0,20
0,25
0,30
0,35
S
R1
R2
Figura 29. Níveis de Ampa (μg g-1
) em plantas de azevém S, R1 e R2 aos 5, 11 e 28 DAA.
Botucatu – SP.

78
Os níveis de ácido chiquímico estão representados na Figura 30. Aos 5
DAA nas populações consideradas suscetíveis, houve aumento deste composto de acordo com
o aumento da dose. Já para a população resistente os valores permaneceram constantes. Esses
resultados foram semelhantes nas 3 épocas de avalição. Aos 11 e 28 DAA as plantas
suscetíveis não apresentaram o mesmo comportamento. Este resultado é esperado já que
neste momento as plantas suscetíveis já estavam com sintomas avançados do efeito do
herbicida onde a população S já havia morrido na dose recomendada. Talvez estas duas
épocas de avaliação não seja o momento ideal para medir tal variável. Santos et al. (2014),
avaliando o efeito do glyphosate sobre o acúmulo de ácido chiquímico em plantas de C.
sumatrensis observaram que até aos sete dias após o tratamento foi observado acúmulo deste
composto sendo que a partir deste momento houve redução dos valores até 14 DAA. O
acúmulo de ácido chiquímico em tecidos ocorre pela inibição competitiva da EPSPs pelo
glyphosate (BRESNAHAN et al., 2003).
Diversas literaturas citam o aumento da concentração de ácido
chiquímico após a aplicação do herbicida glyphosate em plantas suscetíveis, porém o mesmo
não é observado para as resistentes. Zelaya et al. (2011), observou acúmulo de ácido
chiquímico após a aplicação de glyphosate em soja suscetível ao mesmo. Conforme aumentou
a dose deste herbicida, houve um grande acúmulo deste ácido em Corydalis sempervirens
(AMRHEIN et al., 1983). Pline et al. (2002), observaram que as variedades suscetíveis de
algodão acumularam ácido chiquímico após a aplicação de glyphosate, porém nas variedades
resistentes o mesmo não foi verificado onde os níveis não se alteraram. Estes resultados
corroboram com os aqui encontrados reforçando a hipótese de que as populações S e R1 são
suscetíveis e R2 resistente. Resultados semelhantes foram encontrados por outros autores
(NANDULA at al., 2007; REDDY et al., 2008; CARVALHO; ALVES, 2012).
O acúmulo de ácido chiquímico em resposta ao glyphosate tem sido
utilizado como um biomarcador rápido e preciso na suscetibilidade das plantas ao mesmo
(MULLER et al., 2003; GONZALES-TORRALVA et al., 2010). Sendo assim, numa espécie
com reduzido acúmulo deste ácido, provavelmente haverá a necessidade de uma grande
quantidade deste herbicida para ser letal (GOMES, 2014). Novamente este fato reforça a
hipótese de R2 ser resistente, pois, oito vezes a dose recomendada de glyphosate não foi
suficiente para matar 100% desta população.

79
5 DAA
Doses de glyphosate (%)
100 1000
Ácid
o c
hiq
uím
ico
(g
g-1
)
0
1000
2000
3000
4000
5000
S
R1
R2
11 DAA
Doses de glyphosate (%)
100 1000
Ácid
o c
hiq
uím
ico
(g
g-1
)
0
500
1000
1500
2000
2500
3000
S
R1
R2
28 DAA
Doses de glyphosate (%)
100 1000
Ácid
o c
hiq
uím
ico
(g
g-1
)
0
200
400
600
800
1000
S
R1
R2
Figura 30. Níveis de ácido chiquímico (μg g
-1) em plantas de azevém S, R1 e R2 aos 5, 11
e 28 DAA. Botucatu – SP.

80
Os níveis de ácido quínico tiveram um padrão semelhante aos do ácido
chiquímico. Aos 5 DAA foi registrado um aumento nos valores deste composto com o
aumento das doses do herbicida para as populações S e R1 enquanto que para R2 foi
observado constância nos valores demonstrando um leve aumento nas doses subsequentes à
recomendada (Figura 31). Aos 11 DAA novamente foi notado aumento dos níveis de acordo
com o acréscimo das doses para as populações consideradas suscetíveis sendo observado um
leve aumento nas maiores doses para a população resistente onde aos 28 DAA este aumento
foi mais pronunciado.
Em experimento realizado por Gomes (2011), foi observado acúmulo
de ácido quínico após a aplicação de glyphosate em plantas de milho na maior dose testada.
Este aumento ocorreu até seis dias após a aplicação sendo notado decréscimo a partir dos 10
dias. No mesmo experimento, o autor encontrou alta correlação com o ácido chiquímico
sendo a concentração deste, quatro vezes superior à do ácido quínico, resultados diferente dos
aqui encontrados, pois os valores das variáveis em questão foram semelhantes.
O ácido quínico possui estrutura semelhante ao ácido chiquímico sendo
precursor dos hidroxibenzóicos. O acúmulo de ácido quínico após a aplicação de glyphosate
pode ser devido ao acúmulo de compostos acima da EPSPs na rota, como ocorre para os
ácidos chiquímicos, protocatecúico e gálico (ORCARAY et al., 2010). Como dito
anteriormente, o acúmulo de ácido chiquímico pode ser utilizado como biomarcador preciso e
rápido na suscetibilidade das plantas ao glyphosate, sendo assim, o ácido quínico também
pode ser considerado com a mesma função.

81
5 DAA
Doses de glyphosate (%)
100 1000
Ácid
o q
uín
ico
(g
g-1
)
0
1000
2000
3000
4000
5000
S
R1
R2
11 DAA
Doses de glyphosate (%)
100 1000
Ácid
o q
uín
ico
(g
g-1
)
0
500
1000
1500
2000
2500
S
R1
R2
28 DAA
Doses de glyphosate (%)
100 1000
Ácid
o q
uín
ico
(g
g-1
)
0
100
200
300
400
500
600
700
800
S
R1
R2
Figura 31. Níveis de ácido quínico (μg g-1
) em plantas de azevém S, R1 e R2 aos 5, 11 e 28
DAA. Botucatu – SP.

82
A concentração de shiquimato-3-fostato se manteve constante em todas
as doses aplicadas para as populações consideradas suscetíveis enquanto que para a resistente
foi observado aumento nas doses subsequentes à recomendada aos 5 DAA (Figura 32). O
mesmo comportamento ocorreu nas outras épocas de avaliação. Estes resultados são
interessantes devido ao fato de que para a população suscetível este composto deveria
aumentar com a aplicação de glyphosate, pois com a inibição da enzima EPSPs ocorre
acúmulo dos compostos acima da rota como o ácido chiquímico e shiquimato-3-fostato. Os
efeitos fitotóxicos causados por este herbicida é um resultado principalmente da acumulação
destes metabólitos (JENSEN, 1985; LORRAINE-COLWILL et al., 2002; ZABLOTOWICZ;
REDDY, 2004).

83
5 DAA
Doses de glyphosate (%)
100 1000
Chiq
uim
ato
3 fo
sfa
to (
g g
-1)
0
100
200
300
400
500
600
700
S
R1
R2
11 DAA
Doses de glyphosate (%)
100 1000
Chiq
uim
ato
3 fo
sfa
to (
g g
-1)
0
50
100
150
200
250
300
S
R1
R2
28 DAA
Doses de glyphosate (%)
100 1000
Chiq
uim
ato
3 fo
sfa
to (
g g
-1)
0
20
40
60
80
100
S
R1
R2
Figura 32. Níveis de chiquimato-3-fosfato (μg g-1
) em plantas de azevém S, R1 e R2 aos 5,
11 e 28 DAA. Botucatu – SP.

84
Na Figura 33, são encontrados os níveis de fenilalanina nas diferentes
populações. Nota-se que aos 5 DAA houve redução deste aminoácido conforme aumentou-se
as doses para S e R1. Porém, o mesmo não foi observado para a população considerada
resistente onde os níveis permaneceram constantes. A enzima EPSPs catalisa a condensação
do ácido chiquímico com o piruvato para formar o corismato, na síntese dos aminoácidos
aromáticos no qual a fenilalanina faz parte (VARGAS et al., 2007b). O glyphosate atua
especificamente nesta enzima prejudicando a produção deste composto que neste caso, foi
reduzido para as populações suscetíveis.
Resultados contrários foram encontrados por Gomes (2014a), no qual
observou um aumento deste aminoácido em populações de Conyza spp. conforme se
aumentou a dose para as populações resistentes onde na suscetível houve oscilações nos
resultados, porém, a mesma não apresentou redução significativa. Em Arabidopsis thaliana
foi observado um aumento de fenilalanina após a aplicação do herbicida glyphosate
(TRENKAMP et al., 2009). Este comportamento foi verificado em Brassica napus L., porém,
nas menores doses deste herbicida, pois nas maiores ocorreu redução deste composto
(PETERSEN et al., 2007).
Aos 11 DAA houve um pequeno aumento nos níveis deste composto na
população R2 até duas vezes a dose recomendada, porém, na maior dose testada os valores
foram semelhantes às menores doses. Observou redução do aminoácido com o aumento da
dose para as populações suscetíveis. Já aos 28 DAA o comportamento da população R2 foi
semelhante aos 5 DAA inclusive nos valores, ou seja, mesmo após a aplicação de 8 vezes a
dose recomendada, o glyphosate não foi capaz de causar efeito neste aminoácido.

85
5 DAA
Doses de glyphosate (%)
100 1000
Fe
nila
lanin
a (
g g
-1)
0
100
200
300
400
500
S
R1
R2
11 DAA
Doses de glyphosate (%)
100 1000
Fe
nila
lanin
a (
g g
-1)
40
60
80
100
120
140
S
R1
R2
28 DAA
Doses de glyphosate (%)
100 1000
Fe
nila
lanin
a (
g g
-1)
60
80
100
120
140
160
180
200
220
240
260
S
R1
R2
Figura 33. Níveis de Fenilalanina (μg g-1
) em plantas de azevém S, R1 e R2 aos 5, 11 e 28
DAA. Botucatu – SP.

86
Em relação aos níveis de tirosina, semelhante à fenilalanina aos 5 DAA,
foi observado redução dos valores de acordo com o aumento das doses de glyphosate para as
populações suscetíveis (Figura 34). Entretanto, a população R2 apresentou oscilações nos
resultados conforme se aumentou a dose do herbicida. Este comportamento também foi
verificado aos 11 e 28 DAA. Um fato que deve ser levado em consideração é que mesmo
tendo sido observado variações de acordo com o aumento das doses, R2 apresentou os
maiores valores em todas as épocas avaliadas e em todas as doses aplicadas com relação à S e
R1. Mesmo nas maiores doses, a população resistente apresentou maiores valores em relação
às menores doses nas plantas suscetíveis.
Gomes (2011) observou forte correlação entre os níveis de tirosina e
fenilalanina em milho após a aplicação do herbicida glyphosate. Esta mesma autora, em
trabalho realizado em 2014, demonstrou que os níveis de ambos os aminoácidos tiveram um
padrão semelhante após a aplicação do glyphosate para as plantas suscetíveis e resistentes.
Estes resultados são contrários aos aqui analisados, pois enquanto que os níveis de
fenilalanina se mantiveram constantes nas plantas resistentes, os de tirosina apresentou
variações. Petersen et al. (2007), encontrou reduções nos valores de tirosina nas maiores doses
do herbicida glyphosate em planta de B. napus.

87
5 DAA
Doses de glyphosate (%)
100 1000
Tir
osin
a (
g g
-1)
0
100
200
300
400
500
S
R1
R2
11 DAA
Doses de glyphosate (%)
100 1000
Tir
osin
a (
g g
-1)
20
40
60
80
100
120
140
160
180
200
S
R1
R2
28 DAA
Doses de glyphosate (%)
100 1000
Tir
osin
a (
g g
-1)
20
40
60
80
100
120
140
S
R1
R2
Figura 34. Níveis de Tirosina (μg g-1
) em plantas de azevém S, R1 e R2 aos 5, 11 e 28 DAA.
Botucatu – SP.

88
Na população S houve redução dos níveis de triptofano de acordo com
o aumento da dose de glyphosate aos 5 DAA (Figura 35). Comportamento semelhante foi
observado para a população R1, porém na maior dose foi observado um aumento em relação à
S. Novamente em R2 foi notado uma variação nos valores de acordo com o aumento da dose,
no entanto, essa oscilação foi menor em relação à tirosina. Aos 11 DAA foi observada uma
grande variação nos valores para as três populações. Porém, aos 28 DAA os níveis de
triptofano em R2 retornaram a um padrão mais constante, semelhante aos 5 DAA e para R1
foi observado redução de acordo com o aumento da dose.
Peterson et al. (2007) analisando o efeito do herbicida glyphosate sobre
Brassica napus L. não observou grandes variações nos níveis de triptofano com o aumento da
dose o que de acordo com o autor deveria reduzir. A fenilalanina e a tirosina possuem o
mesmo precursor diferentemente do triptofano. Dentre os aminoácidos aromáticos, ele é o
menos sensível ao efeito do glyphosate explicando em parte tal efeito (AMRHEIN et al.,
1980). Foi observado no presente experimento que para a população suscetível de um modo
geral houve redução deste aminoácido com o aumento da dose, efeito este, esperado após a
aplicação do herbicida. Porém o mesmo não foi verificado população considerada resistente.
Efeitos contrários foram observados por Gomes (2014), onde na população suscetível de
Conyza spp. houve grandes oscilações nos valores de triptofano com o aumento da dose de
glyphosate sendo que para duas populações consideradas resistentes os valores foram
constantes.

89
5 DAA
Doses de glyphosate (%)
100 1000
Tri
pto
fano
(g
g-1
)
0
50
100
150
200
250
300
350
S
R1
R2
11 DAA
Doses de glyphosate (%)
100 1000
Tri
pto
fano
(g
g-1
)
60
80
100
120
140
160
180
S
R1
R2
28 DAA
Doses de glyphosate (%)
100 1000
Tri
pto
fano
(g
g-1
)
40
60
80
100
120
140
160
180
S
R1
R2
Figura 35. Níveis de Triptofano (μg g
-1) em plantas de azevém S, R1 e R2 aos 5, 11 e 28
DAA. Botucatu – SP.

90
De acordo com a literatura, há um aumento geral dos aminoácidos após
a aplicação do glyphosate (COOLEY; FOY, 1992; PETERSEN et al., 2007; MOLDES et al.,
2008; GRAVENA et al., 2009). Essa alteração pode ser devido às regulações metabólicas na
biossíntese dos aminoácidos e hidrólise de proteínas. De acordo com Jaworski (1972), o
glyphosate causa uma desaceleração na síntese de proteínas e como consequência, a procura
por aminoácidos para a síntese proteica diminui, aumentando assim, os aminoácidos livres.
Essa pode ser uma explicação em determinadas situações em que há um aumento dos
aminoácidos aromáticos após exposição ao glyphosate.
Apesar da redução dos aminoácidos aromáticos ter sido reportada como
o primeiro passo da inibição causado pelo glyphosate na via do ácido chiquímico (COLE,
1985), foi demonstrado que esse comportamento não ocorre em todos os casos, ou seja, em
algumas espécies, tem aumentado ao invés de ser inibido, inclusive em plantas consideradas
suscetíveis ao herbicida. De acordo com os dados aqui analisados e de outros autores, não há
como tirar conclusões sobre o comportamento destes aminoácidos após a aplicação do
herbicida glyphosate para as espécies de um modo geral. Cada uma deve ser analisada
separadamente especialmente quando a resistência está envolvida.
Na figura 36 são observados os níveis de ácido ferúlico nas diferentes
populações. R2 demonstrou perfil constante nos valores de acordo com o aumento da dose em
todas as épocas de avaliação inclusive demonstrando números semelhantes nos três períodos
avaliados, ou seja, o herbicida não foi capaz de causar efeito significativo nesta variável ao
longo do experimento. Já para as populações consideradas suscetíveis, notou-se uma redução
até a dose recomendada aos 5 DAA se mantendo constante nas doses subsequentes. Aos 11 e
28 DAA conforme se aumentou a dose do herbicida houve redução dos níveis deste ácido.
Um fato a ser observado é que em todas as doses e épocas avaliadas, essas populações
demonstraram valores superiores à resistente.
O ácido ferúlico tem como seu precursor o aminoácido fenilalanina
(DU et al., 2009). Como demonstrado na figura 3, a população R2 apresentou padrão
constante ao longo das avaliações e doses do herbicida para este aminoácido. Isto pode
explicar em parte o comportamento semelhante do seu produto, neste caso, o ácido ferúlico. A
mesma conclusão pode ser estendida para S e R1, ou seja, ambas apresentaram padrão
semelhante ao seu precursor.

91
5 DAA
Doses de glyphosate (%)
100 1000
Ácid
o fe
rúlic
o (
g g
-1)
0
50
100
150
200
250
S
R1
R2
11 DAA
Doses de glyphosate (%)
100 1000
Ácid
o fe
rúlic
o (
g g
-1)
0
50
100
150
200
250
300
350
S
R1
R2
28 DAA
Doses de glyphosate (%)
100 1000
Ácid
o fe
rúlic
o (
g g
-1)
0
50
100
150
200
250
S
R1
R2
Figura 36. Níveis de Ácido Ferúlico (μg g
-1) em plantas de azevém S, R1 e R2 aos 5, 11 e 28
DAA. Botucatu – SP.

92
Os níveis de ácido coumárico mostraram oscilações de acordo com o
aumento das doses nas populações S e R2 aos 5 DAA, apesar de na suscetível haver uma
tendência de aumento (Figura 37). Aos 11 e 28 DAA a população resistente novamente
demonstrou variações em seus valores. Já para a R1 em todas as épocas avaliadas houve
constância nos resultados. Assim como o ácido ferúlico, o acido coumárico possui como
precursor a fenilalanina, porém não apresentou comportamento semelhante à mesma.

93
5 DAA
Doses de glyphosate (%)
100 1000
Acid
o c
oum
ári
co
(g
g-1
)
0,0
0,5
1,0
1,5
2,0
2,5
S
R1
R2
11 DAA
Doses de glyphosate (%)
100 1000
Ácid
o c
oum
ári
co
(g
g-1
)
0
1
2
3
4
5
S
R1
R2
28 DAA
Doses de glyphosate (%)
100 1000
Ácid
o c
oum
ári
co
(g
g-1
)
0
1
2
3
4
5
S
R1
R2
Figura 37. Níveis de Ácido Coumárico (μg g-1
) em plantas de azevém S, R1 e R2 aos 5, 11 e
28 DAA. Botucatu – SP.

94
Aos 5 DAA os níveis de ácido cafeico permaneceram constantes até a
dose recomendada para a população considerada resistente ocorrendo uma leve redução nas
doses subsequentes (Figura 38). Em S observou um aumento nos valores até 2 vezes a dose
recomendada havendo queda nas doses posteriores. A população R1 apresentou padrão
semelhante à S, porém a redução nos valores ocorreu a partir da dose recomendada
apresentando os menores resultados na maior dose testada. Aos 11 DAA R2 apresentou
comportamento diferente em relação aos 5 DAA havendo redução dos valores com o aumento
das doses até duas vezes a dose recomendada. Aos 28 DAA a mesma população demonstrou
comportamento semelhante à primeira avaliação.
O nível de ácido cafeico reduziu 1 dia após a aplicação de glyphosate
em células de Perilla, porém, se manteve constante até os cinco dias de avaliação sendo que
para a testemunha foi observado aumento deste composto nos mesmo períodos (ISHIKURA;
TAKESHIMA, 1984). Resultados semelhantes foram encontrado por Mobin et al. (2015),
onde com o aumento da dose do herbicida glyphosate, observou redução dos níveis de ácido
cafeico em Echinacea purpúrea (L.).

95
5 DAA
Doses de glyphosate (%)
100 1000
Ácid
o c
afe
ico
(g
g-1
)
0
2
4
6
8
10
12
14
16
18
20
S
R1
R2
11 DAA
Doses de glyphosate (%)
100 1000
Ácid
o c
afe
ico
(g
g-1
)
1
2
3
4
5
6
S
R1
R2
28 DAA
Doses de glyphosate (%)
100 1000
Ácid
o c
afe
ico
(g
g-1
)
2
3
4
5
6
7
8
9
10
S
R1
R2
Figura 38. Níveis de Ácido Cafeico (μg g
-1) em plantas de azevém S, R1 e R2 aos 5, 11 e 28
DAA. Botucatu – SP

96
Como pôde ser observado, o comportamento das variáveis analisadas
nem sempre seguiram aquilo que se esperava de acordo com pesquisas anteriormente feitas.
Há variações de acordo com a espécie envolvida, época de avaliação, dose do herbicida
aplicado, etc. Após a aplicação do glyphosate, os compostos que fazem parte da rota do qual o
mesmo atua não necessariamente vão sofrer seus efeitos. De acordo com Velini et al. (2009a),
isto pode sugerir que nem sempre os produtos distantes do sítio de ação, neste caso a enzima
EPSPs, terão a sua produção bloqueada ou reduzida pelo glyphosate devido a presença de
sistemas de controle, que podem em parte, compensar a redução da síntese de determinados
compostos intermediários. Quanto menor o número de reações e enzimas, no caso dos
herbicidas, que separam um composto até o sítio de ação, maior a possibilidade que os níveis
deste produto sejam reduzidos devido à ação do produto.

97
7. CONCLUSÕES
Na população considerada resistente, a atividade da enzima fenilalanina
amônia liase manteve-se alta após a aplicação do glyphosate;
Todas as variáveis fisiológicas foram afetadas após a aplicação do
glyphosate nas três populações, porém, R2 foi capaz de se recuperar apresentando valores
semelhantes à testemunha;
Os níveis de ácido chiquímico e quínico apresentaram padrões
semelhantes onde houve aumento para as populações suscetíveis com o aumento da dose do
herbicida enquanto que para a resistente os valores se mantiveram semelhantes;
Ocorreu aumento dos níveis de shiquimato-3-fosfato para a população
R2 se mantendo constante para as suscetíveis;
Houve redução dos aminoácidos aromáticos com a aplicação do
glyphosate para as populações suscetíveis.

98
8. REFERÊNCIAS
ADRIANO, R. C. et al. Phenotypic and biochemical responses of sugarcane cultivars to
glyphosate application. Sugar Tech, New Delhi, v. 15, n. 2, p. 127-135, 2013.
ADU-YEBOAH, P.et al. Reduced glyphosate in two glyphosate-resistant populations of rigid
ryegrass (Lolium rigidum) from fence in South Australia. Weed Science, Lawrence, v. 62, n.
1, p. 4-10, 2014.
AGOSTINETTO, D.; VARGAS, L. Resistência de plantas daninhas a herbicidas no
Brasil. Passo Fundo: Berthier, 2009. 352 p.
AMARANTE JR. O. P. et al. Glifosato: propriedades, toxicidade, usos e legislação. Química
Nova, São Paulo, v. 25, n. 4, p. 589-593, 2002.
AMRHEIN, N. et al. The site of the inhibition of the shikimate pathway by glyphosate. Plant
Physiology, Chicago, v. 66, p. 830-834, 1980.
AMRHEIN, N. et al. Biochemical basis for glyphosate-tolerance in a bacterium and a plant
tissue culture. Federation of European Biochemical Societies, New York, v. 157, p. 191-
196, 1983.
ANDERSON, K. S.; JOHNSON, H. A. Kinetic and structural analysis of enzyme
intermediates from EPSPs: lessons from EPSP synthase. Chemical Review, Urbana-
Champaign, v. 90, n. 7, p. 1131-1149, 1990.
AHSAN, N. et al. Glyphosate-induced oxidative stress in rice leaves revealed by proteomic
approach. Plant Physiology and Biochemistry, Paris, v. 46, n. 12, p. 1062-1070, 2008.
ARCURI, H. A. et al. Molecular models for shikimate pathway enzymes os Xylella fastidiosa.
Biochemical and Biophysical Research Communications, Amsterdam, v. 320, p. 979-991,
2004.

99
ARREGUI, M. C. et al. Monitoring glyphosate residues in transgenic glyphosate-resistant
soybean. Pest Management Science, London, v. 60, n. 2, p. 163-166, 2003.
BAERSON, S. R. et al. Glyphosate-resistant goosegrass. Identification of a mutation in the
target-site enzyme 5-enolpyruvylshikimate-3-phosphate synthase. Plant Physiology, Chicago,
v. 129, n. 3, p. 1265-1275, 2002.
BELL, M. S.; HAGER, A. G.; TRANEL, P. J. Multiple resistance to herbicides from four
site-of-action groups in waterhemp (Amaranthus tuberculatos). Weed Science, Lawrence, v.
61, n. 3, p. 460-468, 2013.
BELLALOULI, N. et al. Nitrogen metabolism and seed composition as influenced by
glyphosate application in glyphosate-resistant soybean. Journal of Agricultural and Food
Chemistry, Washington, v. 56, n. 8, p. 2765-2772, 2008.
BENTLEY, R; HASLAM, E. The shikimate pathway: a metabolic tree with many branches.
Critical Reviews Biochemistry and Molecular Biology, Madison, v. 25, n. 5, p. 307-384,
1990.
BLANCO, G. H. A importância dos estudos ecológicos nos programas de controle das plantas
daninhas. O Biológico, São Paulo, v. 38, n. 10, p. 343-350, 1972.
BOSTAMAM, Y. et al. Rigid ryegrass (Lolium rigidum) populations containing a target site
mutation in EPSPs and reduced glyphosate translocation are more resistant to glyphosate.
Weed Science, Lawrence, v. 60, n. 3, p. 474-479, 2012.
BRESNAHA, G. A. et al. Glyphosate applied preharvest induces shikimic acid accumulation
in hard red spring wheat (Triticum aestivum). Journal of Agriculture and Food Chemistry,
Washington, v. 51, n. 14, p. 4004-4007, 2003.
BRUNHARO, C. A. C. G. Resistência da planta daninha capim branco (Chloris
polidactyla) ao herbicida glyphosate. 2014. 153 f. Dissertação (Mestrado em Ciências -
Fitotecnia) - Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo,
Piracicaba, 2014.
BRYCE, J. H. et al. Plant biochemistry and molecular biology. Oxford: University Press,
1993. 312 p.
BURNSIDE, O. C. Rationale for developing herbicide-resistant crops. Weed Technology,
Champaign, v. 6, n. 3, p. 621-625, 1992.
CABANÉ, M. et al. Condensed lignins are synthesized in poplar leaves exposed to ozone.
Plant Physiology, Chicago, v. 134, n. 2, p. 586-594, 2004.
CAEMMERER, S. V.; FARQUHAR, G. D. Some relationships between the biochemistry of
photosynthesis and gas exchange pf leaves. Planta, New York, v. 153, n. 4, p. 376-387, 1981.
CAKMAK, I. et al. Glyphosate reduced seed and leaf concentrations of calcium, manganese,
magnesium, and iron in non-glyphosate resistant soybean. European Journal of Agronomy,
Amsterdam, v. 31, n. 3, p. 114-119, 2009.

100
CAMM, E. L.; TOWERS, G. H. N. Phenylalanine ammonia lyase. Phytochemistry, London,
v. 12, p. 961-973, 1973.
CAMPBELL, S. A. et al. A complete shikimate pathway in Toxoplasma gondii an accident
eukaryotic innovation. International Journal for Parasitology, New York, v. 34, p. 5-13,
2004.
CARAMBULA, M. Producción y manejo de pasturas sembradas. Montevideu: Hemisfério
Sur, 1998. 464 p.
CARDINALI, V. C. B. et al. Shikimate accumulation, glyphosate absorption and
translocation in Horseweed biotypes. Planta Daninha, Viçosa, v. 33, n. 1, p. 109-118, 2015.
CARVALHO, P. C. F. Manejando pastagens para ovinos. In: PEREIRA NETO, O. A. (Ed.).
Práticas em ovinocultura: ferramentas para o sucesso. Porto Alegre: Senar-RS, 2004. p. 15-
28.
CARVALHO, S. J. P.; CHRISTOFFOLETI, P. J.; LÓPEZ-OVEJERI, R. F. Métodos para a
comprovação da resistência de plantas daninhas a herbicidas. In: AGOSTINETTO D.;
VARGAS, L. Resistência de plantas daninhas a herbicidas no Brasil. Passo Fundo:
Berthier, 2009. p. 103-122.
CARVALHO, L. B.; ALVES, P. L. C. A. Pool of resistance mechanisms to glyphosate in
Digitaria insularis. Journal of Agriculture and Food Chemistry, Washington, v. 60, n. 2, p.
615-622, 2012.
CASSOL, L. C. et al. Produtividade e composição estrutural de aveia e azevém submetidos a
épocas de corte e adubação nitrogenada. Revista Ceres, Viçosa, v. 58, n. 4, p. 438-443, 2011.
CESSNA, A. J. et al. Residues of glyphosate and its metabolite AMPA in wheat seed and
foliage following preharvest applications. Canadian Journal of Plant Science, Ottawa, v.
74, n. 3, p. 653-661, 1994.
CESSNA, A. J. et al. Residues of glyphosate and metabolite AMPA in canola seed following
preharvest applications. Canadian Journal of Plant Science, Ottawa, v. 80, n. 2, p. 425-431,
2000.
CESSNA, A. J. et al. Residues of glyphosate and its metabolite AMPA in field pea, barley
and flax seed following preharvest applications. Canadian Journal of Plant Science,
Ottawa, v. 82, n. 2, p. 485-489, 2002.
CHRISTOFFOLETI, P. J.; LÓPEZ-OVEJERO, R. Principais aspectos da resistência de
plantas daninhas ao herbicida glyphosate. Planta Daninha, Viçosa, v. 21, n. 3, p. 507-515,
2003.
CHRISTOFFOLETI, P. J.; LÓPEZ-OVEJERO, R. Definições e situação da resistência de
plantas daninhas aos herbicidas no Brasil e no mundo. In: CHRISTOFFOLETI, P. J.; LÓPEZ-
OVEJERO, R.; CARVALHO, S. J. P. Aspectos de resistência de plantas daninhas a
herbicidas. Campinas: HRAC-BR, 2004. p. 3-22.

101
CHRISTOFFOLETI, P. J. Aspectos de resistência de plantas daninhas a herbicidas. 3. ed.
Piracicaba: HRAC-BR, 2008. 120 p.
CHRISTOFFOLETI, P. J.; LÓPEZ-OVEJERO, R. Resistência das plantas daninhas a
herbicidas: definições, bases e situação no Brasil e no mundo. In: CHRISTOFFOLETI, P. J.
Aspectos de resistência de plantas daninhas a herbicidas. Piracicaba: HRAC-BR, 2008.
120 p.
CHRISTOFFOLETI, P. J. et al. Resistência de plantas daninhas aos herbicidas inibidores da
EPSPs. In: VELINI, E. D. et al. Glyphosate. Botucatu: FEPAF, 2009. p. 309-355.
COLE, D. J. Mode of action of glyphosate: a literature analysis. In: GROSSBARD, E.;
ATKINSON, D. The herbicide glyphosate. London: Butterworths, 1985. p. 48-74.
COLLAVO, A.; SATTIN, M. Resistance to glyphosate in Lolium rigidum selected in Italian
perennial crops: bioevaluation, management and molecular bases of target-site resistance.
Weed Research, Oxford, v. 52, n. 1, p. 16-24, 2012.
COOLEY, W. E.; FOY, C. L. Effects of SC-0224 and glyphosate on free amino acids, soluble
protein, and protein synthesis in inflated duckweed (Lemma gibba). Weed Science,
Lawrence, v. 40, n. 3, p. 345-350, 1992.
COUPLAND, D. Metabolism of glyphosate in plants. In: GROSSBARD, E.; ATKINSON, D.
The herbicide glyphosate. London: Butterworth, 1985. p. 25-33.
COX, C. Glyphosate (Round up). Journal of Pesticide Reform. Eugene, Fall, v. 18, n. 3,
1998.
DAVIS, V. M. et al. Growth and seed production of horseweed (Conyza canadensis)
populations resistant to glyphosate, ALS-inhibiting, and multiple (glyphosate+ ALS-
inhibiting) herbicides. Weed Science, Lawrence, v. 57, n. 5, p. 494-504, 2009.
DE MARÍA, N. et al. New insights on glyphosate mode of action in nodular metabolism: role
of shikimate accumulation. Journal of Agricultural and Food Chemistry, Washington, v.
54, p. 2621-2628, 2006.
DENIS, M. H.; DELROT, S. Carrier-mediated uptake of glyphosate in broad bean (Vicia
faba) via transporter. Plant Physiology, Chicago, v. 87, p. 568-575, 1993.
DEVINE, M. D.; BANDEEN, J. D.; MCKERSIE, B. D. Temperature effects on glyphosate
absorption, translocation, and distribution in quackgrass (Agropyron repens). Weed Science,
Lawrence, v. 31, n. 4, p. 461-464, 1983.
DEWICK, P. M. The biosynthesis of shikimate metabolities. Natural Products Reports,
London, v. 10, p. 173-203, 1998.

102
DICK, R. E.; QUINN, J. P. Glyphosate-degrading environmental samples: occurrence and
pathways of degradation. Applied microbiology Biotechnology, Heidelberg, v. 43, n. 3, p.
545-550, 1995.
DINELLI, G. et al. Physiological and molecular bases of glyphosate resistance in Conyza
bonariensis biotypes from Spain. Weed Research, Oxford, v. 48, n. 3, p. 257-265, 2008.
DING, W. et al. Physiological responses of glyphosate-resistant and glyphosate-sensitive
soybean to aminomethylphosphonic acid, a metabolite of glyphosate. Chemosphere, Oxford,
v. 83, n. 4, p. 593-598, 2011.
DIONISIO, G. et al. Manejo de plantas daninhas resistentes ao glifosato no Brasil. In: RÍOS,
A. Viabilidad del glifosato em sistemas productivos sustentables. Montevideo: INIA.
2013. p. 111-118.
DIXON, R. A.; PAIVA, N. L. Stress-induced phenylpropanoid metabolism. The Plant Cell,
Waterbury, v. 7, n. 7, p. 1085-1097, 1995.
DOSSELAERE, F.; VANDERLEYDEN, J. A metabolic node in action: chorimate-utilizing
enzymes in microorganisms. Critical Reviews in Microbiology, Cleveland, v. 27, n. 2, p. 75-
131, 2001.
DU, H. et al. Biochemical and molecular characterization of plant MYB transcription factor
family. Biochemistry, Washington, v. 74, p. 1-11, 2009.
DUKE, S. O.; HOAGLAND, R. E. Effects of glyphosate on metabolism of phenolic
compounds. I. Induction of phenylalanine ammonia-lyase activity in dark-grown maize roots.
Plant Science Letters, Amsterdam,v. 11, n. 3/4, p. 185-190, 1978.
DUKE, S. O.; HOAGLAND, R. E.; ELMORE, C. D. Effects of glyphosate on metabolism of
phenolic compounds. IV. Phenylalanine ammonia-lyase activity, free amino acids, and
soluble hydroxyphenolic compounds in axes of light-grown soybeans. Physiologia
Plantarum, Copenhagen, v. 46, n. 4, p. 307-317, 1979.
DUKE, S. O.; HOAGLAND, R. E.; ELMORE, C. D. Effects of glyphosate on metabolism of
phenolic compounds. V. L-α-aminoox-β-phenylpropionic acid and glyphosate effects on
phenylalanine ammonia-lyase in soybean seedlings. Plant Physiology, Chicago, v. 65, p. 17-
21, 1980.
DUKE, S. O. et al. Isoflavone, glyphosate, and aminomethylphosphonic acid levels in seeds
of glyphosate-treated, glyphosate-resistant soybean. Journal of Agriculture and Food
Chemistry, Washington, v. 51, p. 340-344, 2003.
DUKE, S. O.; POWLES, D. O. Glyphosate: a once-in-a-century herbicide. Pest Management
Science, London, v. 64, p. 319-325, 2008.
DYER, W. E. Resistance to glyphosate. In: POWLES, S. B.; HOLTUM, J. A. M. Herbicide
resistance in plants. Boca Raton: CRC press, 1994. p. 229-241.

103
ESCHENBURG, S. et al. How the mutation glycine96 to alanine confers glyphosate
insensitivity to 5-enolpyruvyl shikimate-3-phosphate synthase from Escherichia coli. Planta,
New York, v. 216, p. 129-135, 2002.
FARQUHAR, G. D.; SHARKEY, T. D. Stomatal conductance and photosynthesis. Annual
Review of Plant Physiology, Palo Alto, v. 33, p. 317-345, 1982.
FEDTKE, C.; DUKE, S. O. Herbicides, In: HOCK, B.; ERICH, F. E. Plant toxicology. New
York: CRC Press, 2004. p. 247-330.
FENG, P. C. C.; PRATLEY, J. E.; BOHN, J. A. Resistance to glyphosate in Lolium rigidum.
II. Uptake, translocation, and metabolism. Weed Science, Lawrence, v. 47, p. 412-415, 1999.
FENG, P. C. C. et al. Investigations into glyphosate-resistant horseweed (Conyza
Canadensis): retention, uptake, translocation and metabolism. Weed Science, Lawrence, v.
52, n.4, p. 498-505, 2004.
FERREIRA, E. A. et al. Translocação do glyphosate em biótipos de azevém (Lolium
multiflorum). Planta Daninha, Viçosa, v. 24, n. 2, p. 365-370, 2006a.
FEREIRA, E. A. et al. Glyphosate no controle de biótipos de azevém e impacto na microbiota
do solo. Planta Daninha, Viçosa, v. 24, n. 3, p. 573-578, 2006b.
FERREIRA, E. A. et al. Distribuição de glyphosate e acúmulo de nutrientes em biótipos de
azevém. Planta Daninha, Viçosa, v. 26, n. 1, p. 165-173, 2008.
FERREIRA, E. A. et al. Resistência de Lolium multiflorum ao glyphosate. In:
AGOSTINETTO, D.; VARGAS, L. Resistência de plantas daninhas a herbicidas no
Brasil. Passo Fundo: Berthier, 2009. p. 271-289.
FISCHER, R. S. et al. Comparative action of glyphosate as a trigger of energy drain in
Eubacteria. Journal of Bacteriology, Washington, v. 168, n. 3, p. 1147-1154, 1986.
FLEXAS, J. et al. The effects of water stress on plant respiration. In: LAMBERS, H.; RIBAS-
CARDO, M. Plant respiration: from cell to ecosystem. Dordrecht: Springer, 2005. p. 85-93.
FRANZ, J. E. Discovery, development and chemistry of glyphosate. In: GROSSBARD, E.;
ATKINSON, D. The herbicide glyphosate. London: Butterworths, 1985. p. 3-17.
FRANZ, J. E.; MAO, M. K.; SIKORSKI, J. A. Glyphosate: a unique global herbicide.
Washington: American Chemical Society, 1997. 653 p.
FREITAS, F. A.; OLIVEIRA, A. C.; CARVALHO, F. I. F. Análise multivariada de
populações de azevém (Lolium multiflorum L.) em diferentes regimes de água. Revista
Brasileira de Agrociência, Pelotas, v. 9, n. 1, p. 17-23, 2003.
FRUNK, T. et al. Structural basis of glyphosate resistance resulting from the double mutation
thr97
→ lle and pro101
→ ser in 5-enolpyruvilshikimate-3-phosphate synthase from

104
Escherichia coli. The Journal of Biological Chemistry, Rockville, v. 284, n. 15, p. 9854-
9860, 2009.
FUCHS, M. A. et al. Mechanisms of glyphosate toxicity I velvetleaf (Abutilon theophrasti
medikus). Pesticide Biochemistry and Physiology, New York, v. 74, p. 27-39, 2002.
GAINES, T. A. et al. Gene amplification confers glyphosate resistance in Amaranthus
palmeri. Proceedings of the National Academy of Science, Washington, v. 107, n. 3, p.
1029-1034, 2010.
GAINES, T. A. et al. Interspecific hybridization transfers a previously unknown glyphosate
resistance mechanism in Amaranthus species. Evolutionary Applications, Malden, v. 5, p.
29-38, 2012.
GALLI, A. J. B. A molécula glyphosate e a agricultura brasileira. In: VELINI, E. D.
Glyphosate. Botucatu: FEPAF, 2009. p. 17.
GARCÍA, A. A.; CARRIL, E. P. U. Metabolismo secundario de plantas. Reduca, Madrid, v.
2, n. 3, p. 119-145, 2009.
GE, X. et al. Glyphosate-resistant horseweed made sensitive to glyphosate: low-temperature
suppression of glyphosate vacuolar sequestration revealed by 31
P NMR. Pest
Management Science, London, v. 67, n. 10, p. 1215-1221, 2011.
GE, X. et al. Application of 31
P-NMR spectroscopy to glyphosate studies in plants: insights
into cellular uptake and vacuole sequestration correlated to herbicide resistance. In:
KOBAYASHI, D.; WATANABE, E. Handbook on herbicides: biological activity,
classification and health & environment implications. New York: Nova Science Publishers,
2013. p. 55-84.
GE, X. et al. In vivo 31
P-nuclear magnetic resonance studies of glyphosate uptake, vacuolar
sequestration, and tonoplast pump activity in glyphosate-resistant horseweed. Plant
Physiology, Chicago, v. 166, n. 3, p. 1255-1268, 2014.
GEIGER, D. R.; KAPIRAN, S. W.; TUCCI, M. A. Glyphosate inhibits photosynthesis and
allocation of carbon to start in sugar beet leaves. Plant Physiology, Chicago, v. 82, n. 2, p.
468-472, 1986.
GEIGER, D. R.; SHIEH, W.; FUCHS, M. A. Causes of self-limited translocation of
glyphosate in Beta vulgaris plants. Pesticide Biochemistry and Physiology, New York, v.
64, n. 2, p. 124-133, 1999.
GIMSING, A. L.; BORGGAARD, O. K.; BANG, M. Influence of soil composition on
adsorption fo glyphosate and phosphate by contrasting Danish surface soils. European
Journal of Soil Science, Oxford, v. 55, p. 183-191, 2004.
GOMES, G. L. G. C. Alterações metabólicas de plantas de milho submetidas à aplicação
de glyphosate e fosfito. 2011. 97 f. Dissertação (Mestrado em Agronomia /Proteção de

105
Plantas) - Faculdade de Ciências Agronômicas, Universidade Estadual Paulista, Botucatu,
2011.
GOMES, G. L. G. C. Caracterização bioquímica e morfofisiológica de populações de
buva (Conyza spp.) resistentes ao glyphosate. 2014. 112 f. Tese (Doutorado em
Agronomia/Agricultura) - Faculdade de Ciências Agronômicas, Universidade Estadual
Paulista, Botucatu, 2014.
GOMES, M. P. et al. Alteration of plant physiology and its by-product
aminomethylphosphonic acid: an overview. Jounal of Experimental Botany, Oxford, v. 65,
n. 17, p. 4691-4703, 2014.
GOMES, G. L. G. C. et al. Extraction and simultaneous determination of glyphosate, AMPA
and compounds of the shikimic acid pathway in plants. Planta Daninha, Viçosa, v. 33, n. 2,
p. 295-304, 2015.
GOMIDE, C. A. M.; GOMIDE, J. A. Análise de crescimento de cultivares de Panicum
maximum Jacq. Revista Brasileira de Zootecnia, Viçosa, v. 28, n. 4, p. 675-680, 1999.
GONÇALVES, J. O. N. Nitrogênio e produção de matéria seca do azevém. Pesquisa
Agropecuária Brasileira, Brasília, v. 14, n. 1, p. 47-51, jan. 1979.
GONZÁLES-TORRALVA, F. et al. Differencial susceptibility to glyphosate among the
Conyza weeds species in Spain. Journal of Agriculture and Food Chemistry, Washington,
v. 58, n. 7, p. 4361-4366, 2010.
GONZÁLES-TORRALVA, F. et al. Target site and reduced translocation are present in a
glyphosate-resistant Lolium multiflorum Lam. biotype from Spain. Plant Physiology and
Biochemistry, Paris, v. 58, p. 16-22, 2012a.
GONZÁLES-TORRALVA, F. et al. Two non-target mechanisms are involved in glyphosate-
resistant horseweed (Conyza Canadensis L. Cronq.) biotypes. Journal of Plant Physiology,
Jena, v. 169, n. 17, p. 1673-1679, 2012b.
GRAVENA, R. et al. Low glyphosate rates do not affect Citrus limonia (L.) osbeck seedlings.
Pest Management Science, London, v. 65, n. 4, p. 420-425, 2009.
GRESSEL, J. Molecular biochemistry of resistance that have evolved in the field. In:
GRESSEL, J. Molecular biology of weed control. London: Taylor & Francis, 2000. p. 122-
218.
GUILLET, G. et al. Expression of tryptophan decarboxylase and tyrosine decarboxylase
genes in tobacco results in altered biochemical and physiological phenotypes. Plant
Physiology, Chicago, v. 122, n. 3, p. 933-944, 2000.
HAHLBROCK, K.; GRISEBACH, H. Enzymic controls in the biosynthesis of lignin and
flavonoids. Annual Review of Plant Physiology, Palo Alto, v. 30, p. 105-130, 1979.

106
HANNAWAY, D. et al. Annual ryegrass (Lolium multiflorum Lam.). Oregon State
University, PNW 501, 1999. Disponível em: <http://
ir.library.oregonstate.edu/xmlui/handle/1957/17440>. Acesso em 13 jan. 2015.
HARKER, K. N.; DEKKER, J. Effects of phenology on translocation patterns of several
herbicides in quackgrass (Agropyron repens). Weed Science, Lawrence, v. 36, n. 4, p. 463-
472, 1988.
HASLAM, E. Shikimic acid: metabolism and metabolites. Chinchester: John Wiley, 1993.
392 p.
HEAP, I. International survey of herbicide resistant weeds. Disponível em:
<http://www.weedscience.com/summary/home.aspx>. Acesso em: 07 fev. 2015.
HERMAN, K. M. The shikimate pathway as an entry to aromatic secondary metabolism.
Plant Physiology, Chicago, v. 107, p. 7-12, 1995.
HERMAN, K. M; WEAVER, L. M. The shikimate pathway. Annual Review of Plant
Physiology and Plant Molecular Biology, Palo Alto, v. 50, n. 3, p. 473-503, 1999.
HERNANDEZ, A.; GARCIA-PLAZAOLA, J. I.; BECERRIL, J. M. Glyphosate effects on
phenolic metabolism of nodulated soybean (Glycine max L. Merr.). Journal of Agricultural
and Food Chemistry, Washington, v. 47, n. 7, p. 2920-2925, 1999.
HETHERINGTON, D. R. REYNOLDS, T. L. MARSHALL, G. KIRKWOOD, R. C. The
absorption, translocation and distribution of the herbicide glyphosate in maize expressing the
CP-4 transgene. Journal of Experimental Botany, Oxford, v. 50, n. 339, p. 1567-1576,
1999.
HOAGLAND, R. E.; DUKE, S. O.; ELMORE, D. Effects of glyphosate on metabolism of
phenolic compounds. II. Influence on soluble hydroxyphenolic compound, free amino acid
and soluble protein levels in dark-grown maize roots. Plant Science Letters, Amsterdam, v.
13, n. 4, p. 291-299, 1978.
HOAGLAND, R. E. Effects of glyphosate on metabolism of phenolic compounds: VI. Effets
of glyphosine and glyphosate metabolites on phenylalanine ammonia-lyase activity, growth,
and protein, chlorophyll, and anthocyanin levels in soybean (Glycine max) seedlings. Weed
Science, Lawrence, v. 28, n. 4, p. 393-400, 1980.
HOAGLAND, R. E.; DUKE, S. O. Effects of glyphosate on metabolism of phenolic
compounds. VIII. Comparison of the effects of aminooxyacetate and glyphosate. Plant and
Cell Physiology, Oxford, v. 23, n. 6, p. 1081-1088, 1982.
HOAGLAND, R. E.; DUKE, S. Relationships between phenylalanine ammonia-lyase activity
and physiological responses of soybean (Glycine max) seedlings to herbicide. Weed Science,
Lawrence, v. 31, n. 6, p. 845-852, 1983.
HOAGLAND, R. E. Interaction of indolacetic acid and glyphosate on phenolic metabolism in
soybeans. Pesticide Biochemistry and Physiology, New York, v. 36, p. 68-75, 1990.

107
HOCHBERG, O.; SIBONY, M.; RUBIN, B. The response of ACCase-resistant Phalaris
paradoxa populations involves two diferente target site mutations. Weed Research, Oxford,
v. 49, p. 37-46, 2009.
HOLLANDER, H.; AMRHEIN, N. The site the inhibition of the shikimate pathway by
glyphosate. Plant Physiology, Chicago, v. 66, n. 5, p. 823-829, 1980.
HOLTON, T. A.; CORNISH, E. C. Genetics and biochemistry of anthocyanin biosynthesis.
The Plant Cell, Waterbury, v. 7, n. 7, p. 1071-1083, 1995.
HOWLES, P. A. et al. Overexpression of L-phenylalanine ammonia-lyase in transgenic
tobacco plants reveals control points for flux into phenylpropanoid biosynthesis. Plant
Physiology, Chicago, v. 112, n. 4, p. 1617-1624, 1996.
HUANG, J. et al. Effects of glyphosate on photosynthesis, chlorophyll fluorescence and
physiochemical properties of cogongrass (Imperata cylindrical L.). Plant Omics Journal,
Victoria, v. 5, n. 2, p. 177-183, 2012.
ISHIKURA, N.; TAKESHIMA, Y. Effects of glyphosate on caffeic acid metabolism in
Perilla cell suspensions cultures. Plant and Cell Physiology, Oxford, v. 25, p. 185-189,
1984.
ISMAIL, B. S. et al. Germination and seedling emergence of glyphosate-resistant and
susceptible of goosegrass (Eleusine indica [L.] Gaertn). Weed Biology and Management,
Fukui, v. 2, n. 4, p. 177-185, 2002.
JADOSKI, C. J. Efeitos fisiológicos da piraclostrobina em plantas de feijão (Phaseolus
vulgaris L.) condicionado sob diferentes tensões de água no solo. 2012. 80 f. Dissertação
(Mestrado em Agronomia/Agricultura) - Faculdade de Ciências Agronômicas, Universidade
Estadual Paulista, Botucatu, 2012.
JASIENIUK, M. et al. Glyphosate-resistant Italian ryegrass (Lolium multiflorum) in
California: distribution, response to glyphosate, and molecular evidence for an altered targe
enzyme. Weed Science, Lawrence, v. 56, n. 4, p. 496-502, 2008.
JAWORSKI, E. G. Mode of action of N-phosphonomethylglycine: inhibition of aromatic
amino acid biosynthesis. Journal of Agricultural and Food Chemistry, Washington, v. 20,
n. 6, p. 1195-1198, 1972.
JENSEN, R. A. The shikimate/arogenate pathway: link between carbohydrate metabolism and
secondary metabolism. Plant Physiology, Chicago, v. 66, p. 164-168, 1985.
JONES, D. H. Phenylalanine ammonia-lyase: regulation of its induction, an its role in plant
development. Phytochemistry, London, v. 23, n. 7, p. 1349-1359, 1984.
KANDUM, S. S. et al. A novel P106 mutation in EPSPs and an unknown mechanism(s) act
additively to confer resistance to glyphosate in a South African Lolium rigidum population.
Journal of Agriculture and Food Chemistry, Washington, v. 59, n. 7, p. 3227-3233, 2011.

108
KASPARY, T. E. et al. Pigmentos fotossintéticos em azevém suscetível e resistente ao
herbicida glyphosate. Ciência Rural, Santa Maria, v. 44, n. 11, p. 1901-1907, 2014.
KIRKWOOD, R. C.; MCKAY, I. Accumulation and elimination of herbicides in select crop
and weed species. Pesticide Science, Tokyo, v. 42, n. 3, p. 241-248, 1994.
KISSMANN, K. G. Resistência de plantas a herbicidas. São Paulo: Basf Brasileira, 1996.
33 p.
KISSMANN, K. G. Plantas infestante e nocivas. TOMO I. 3. ed. São Paulo: Basf Brasileira,
2007. 1 CD-ROM.
KOUKOL, J.; CONN, E. E. The metabolism of aromatic compounds in higher plants. IV.
Purification and properties of the phenylalanine deaminase of Hordeum vulgare. The Journal
of Biological Chemistry, Rockville, v. 236, n. 10, p. 2692-2693, 1961.
KUNDU, A.; JAWALI, N.; MITRA, A. Shikimate pathway modulates the elicitor-stimulated
accumulation of fragrant 2-hydroxy-4-methoxybenzaldehyde in Hemidesmus indicus roots.
Plant Physiology and Biochemistry, Paris, v. 56, p. 104-108, 2012.
LEE, L. J.; NGIM, J. A fisrt report of glyphosate-resistant goosegrass (Eleusine indica (L)
Gaertn) in Malaysia. Pest Management Science, London, v. 56, n. 4, p. 336-339, 2000.
LINDERS, J. B. H. J. et al. Pesticides: benefaction or Pandora’s box? A synopsis of the
environmental aspect of 243 pesticides. Research for Man and Environment. National
Institute of Public Health and Environment. Report 67101014, 1994.
LORENZI, H. Plantas daninhas do Brasil: terrestres, aquáticas, parasitas e tóxicas. 4. ed.
Nova Odessa: Instituto Plantarum, 2008. 640 p.
LÓPEZ-OVEJERO, R. F. et al. Resistência de populações de capim-colchão (Digitaria
ciliares) aos herbicidas inibidores da Acetil Co-A Carboxilase. Planta Daninha, Viçosa, v.
23, n. 3, p. 543-549, 2005.
LORRAINE-COLWILL, D. F. et al. Investigations into the mechanism of glyphosate
resistance in Lolium rigidum. Pesticide Biochemistry & Physiology, New York, v. 74, n. 2,
p. 62-72, 2002.
LUBBERSTEDT, T.; SCHEJBEL, A. B.; BACH, H. P. Development of ryegrass allelle-
specific (GRASP) markers sustainable grassland improvement: a new EU framework V
Project. Genetics and Plant Breeding, Praha, v. 39, p. 125-129, 2003. Special issue.
LYDON, J.; DUKE, S. O. Pesticides effects on secondary metabolism of higher plants.
Pesticide Science, Tokyo, v. 25, n. 4, p. 361-373, 1989.
MACDONALD, M. J.; D’CUNHA, G. B. A modern view of phenylalanine ammonia lyase.
Biochemistry and Cell Biology, Ottawa, v. 85, n. 3, p. 273-282, 2007.

109
MAEDA, H.; DUDAREVA, N. The shikimate pathway and aromatic amino acid biosynthesis
in plants. Annual Review of Plant Biology, Palo Alto, v. 63, p. 73-105, 2012.
MARCHIOSI, R. et al. Glyphosate-induced metabolic changes in susceptible and glyphosate-
resistant soybean (Glycine max) roots. Pesticide Biochemistry and Physiology, New York,
v. 93, p. 28-33, 2009.
MARIANI, F. et al. Novos horizontes no controle de plantas daninhas na cultura da soja. In:
REUNIÃO DE PESQUISA DE SOJA DA REGIÃO SUL, 40., 2014, Pelotas. Atas e
Resumos... Pelotas: Embrapa Clima Temperado, 2014. p. 31-39.
MATALLO, M. B. et al. Microwave-assisted solvent extraction and analysis of shikimic acid
from plant tissues. Planta Daninha, Viçosa, v. 27, p. 987-994, 2009.
MATALLO, M. B. et al. Glyphosate as a tool to produce shikimic acid in plants. Planta
Daninha, Viçosa, v. 32, n. 3, p. 601-608, 2014.
MATEOS-NARANJO, E. et al. Effectiveness of glyphosate and imazamox on the controlo f
the invasive cordgrass Spartina densiflora. Ecotoxicology and Environmental Safety,
Amsterdam, v. 72, p. 1694-1700, 2009.
MAXWELL, B. D.; MORTIMER, A. M. Selection from herbicide resistance. In: POWLES,
S. B. e HOLTUM, J. A. M. Herbicide resistance in plants: biology and biochemistry. Boca
Raton, 1994. p. 1-25.
MERVOSH, T. L.; BALKE, N. E. Effects of calcium, magnesium and phosphate on
glyphosate absorption by cultured plant cells. Weed Science, Lawrence, v. 39, n. 3, p. 347-
353, 1991.
MESSINGER, S. M.; BUCKLEY, T. N.; MOTT, K. A. Evidence for involvement of
photosynthetic process in the stomatal response to CO2. Plant Physiology, Chicago, v. 140, n.
2, p. 771-778, 2006.
MICHALOWICZ, J.; DUDA, W. Phenol-sources and toxicity. Polish Journal of
Environmental Studies, Olsztyn, v. 16, n. 3, p. 347-362, 2007.
MICHITTE, P. et al. Mechanisms of resistance to glyphosate in ryegrass (Lolium multiflorum)
biotype from Chile. Weed Science, Lawrence, v. 55, n. 5, p. 435-440, 2007.
MOBIN, M. et al. Studies on the glyphosate-induced amino acid starvation and addition of
precursors on caffeic acid accumulation and profiles in adventitious roots of Echinacea
purpurea (L.). Plant cell, tissue and organ culture, New York, v. 120, p. 291-301, 2015.
MOLDES, C. A. et al. Biochemical responses of glyphosate resistant and susceptible soybean
plants exposed to glyphosate. Acta Physiologiae Plantarum, Warszawa, v. 30, n. 4, p. 469-
479, 2008.

110
MONQUERO, P. A. et al. Absorção, translocação e metabolismo de glyphosate por plantas
tolerantes e suscetíveis a este herbicida. Planta Daninha, Viçosa, v. 22, n. 3, p. 445-451,
2004.
MONTEIRO, J. M. et al. Taninos: uma abordagem da química à ecologia. Química Nova,
São Paulo, v. 28, n. 5, p. 592-896, 2005.
MUELLER, T. C. et al. Shikimate accumulates in both glyphosate-sensitive and glyphosate-
resistant horseweed (Conyza Canadensis L. Cronq). Journal of Agriculture and Food
Chemistry, Washington, v. 51, n. 3, p. 680-684, 2003.
NANDULA, V. K. et al. Glyphosate-resistant and susceptible soybean (Glycine max) and
canola (Brassica napus) dose response and metabolism relationship with glyphosate. Journal
of Agriculture and Food Chemistry, Washington, v. 55, n. 9, p. 3540-3545, 2007.
NANDULA, V. K. et al. Glyphosate tolerance mechanism in Italian ryegrass (Lolium
multiflorum) from Mississipi. Weed Science, Lawrence, v. 56, n. 3, p. 344-349, 2008.
NANDULA, V. K. et al. Glyphosate resistance in tal waterhamp (Amaranthus tuberculatus)
from Mississipi is due to both target-site and nontarget-site mechanisms. Weed Science,
Lawrence, v. 61, n. 3, p. 374-383, 2013.
NG, C. H. et al. Gene polymorphisms in glyphosate-resistant and susceptible biotypes of
Eleusine indica from malaysia. Weed Reseach, Oxford, v. 43, n. 2, p. 108-115, 2003.
NUCROHO, L. H.; VERBERNE, M. V.; VERPOORTE, R. Activities of enzymes involved in
the phenylpropanoid pathway in constitutively salicylic acid-producing tobacco plants. Plant
Physiology and Biochemistry, Paris, v. 40, n. 9, p. 755-760, 2002.
OERKE, E. C. Crop loss to pests. Journal of Agricultural Science, Cambridge, v. 144, n. 1,
p. 31-43, 2006.
OLESEN, C. F.; CEDERGREEN, N. Glyphosate uncouples gas exchange and chlorophyl
fluorescence. Pest Managment Science, London, v. 66, n. 5, p. 536-542, 2010.
OLIVEIRA JR, R. S.; CONSTATIN, J.; INOUE, M. H. Biologia e manejo de plantas
daninhas. Curitiba: Omnipax, 2011. 348 p.
ORCARAY, L. et al. The possible role of quinate in the mode of action of glyphosate and
acetolactate synthase inhibitors. Pest Management Science, London, v. 66, n. 3, p. 262-269,
2010.
ORCARAY, L. et al. Impairment of carbon metabolism induced by the herbicide glyphosate.
Journal of Plant Physiology, Jena, v. 169, p. 27-33, 2012.
OSSIPOV, V. et al. Gallic acid and hydrolysable tannins are formedin birch leaves from an
intermediate compound of the shikimate pathway. Biochemical Systematics and Ecology,
Oxford, v. 31, p. 3-16, 2003.

111
PASTÍROVÁ, A.; REPCÁK, M.; ELIASOVÁ, A. Salicylic acid induces changes of coumarin
metabolites in Matricaria chamomilla L. Plant Science, North Brunswick, v. 167, n. 4, p.
819-824, 2004.
PEDERSEN, B. P. et al. Ecological fitness of a glyphosate-resistant Lolium rigidum
population: growth and seed production along a competition gradiente. Basic and Applied
Ecology, Amsterdam, v. 8, n. 3, p. 258-268, 2007.
PEIXOTO, P. H. P. et al. Aluminum effects on lipid peroxidation and on the activities on
enzimes of oxidative metabolism in sorghum. Revista Brasileira de Fisiologia Vegetal,
Lavras, v. 11, n. 3, p. 137-143, 1999.
PEREIRA-NETO, A. B. Crescimento e desenvolvimento. In: WACHOWICZ, C. M.;
CARVALHO, R. I. N. Fisiologia vegetal: produção e pós-colheita. Curitiba: Champagnat,
2002. p. 17-42.
PEREZ, A.; KOGAN, M. Glyphosate-resistant Lolium multiflorum in Chilean orchards.
Weed Research, Oxford, v. 43, n.1, p. 12-19, 2003.
PEREZ-JONES, A. et al. Investigating the mechanisms of glyphosate resistance in Lolium
multiflorum. Planta, New York, v. 226, n. 2, p. 395-404, 2007.
PETERSEN, I. L. et al. Metabolics effects in rapessed (Brassica napus L.) seedlings after
roots exposure to glyphosate. Pesticide Biochemistry and Physiology, New York, v. 89, n.
3, p. 220-229, 2007.
PETERSON, M. L. et ao. New simplified inhibitors of EPSP synthase: the importance of ring
size for recognition at the shikimate 3-phosphate site. Bioorganic & Medicinal Chemistry
Letters, Amsterdam, v. 6, n. 23, p. 2853-2858, 1996.
PIANA, Z.; CRISPIM, J. E.; NETO, J. A. Z. Superação da dormência de azevém anual
(Lolium multiflorum Lam.). Revista Brasileira de Sementes, Londrina, v. 8, n. 1, p. 67-71,
1986.
PLINE, W. A. et al. Tolerance and accumulation of shikimic acid in response to glyphosate
applications in glyphosate-resistant and nonglyphosate-resistant cotton (Gossypium hirsitum
L.). Journal of Agriculture and Food Chemistry, Washington, v. 50, n. 3, p. 506-512,
2002.
POLOK, K. Molecular evolution of the genus Lolium sp. Olsztyn: Studio Poligrafil
Komputerowej, 2007. 320 p.
POWLES, S. B. Evolved resistance to glyphosate in rigid ryegrass (Lolium rigidum) in
Australia. Weed Science, Lawrence, v. 46, n. 5, p. 604-607, 1998.
POWLES, S. B. PRESTON, C. Evolved glyphosate resistance in plants: biochemical and
genetic basis of resistance. Weed Technology, Lawrence, v. 20, n. 2, p. 282-289, 2006.

112
POWLES, S. B.; YU, Q. Evolution in Action: Plants Resistant to Herbicides. Annual Review
of Plant Biology, Palo Alto, v. 61, p. 317–347, 2010.
PRESTON, C.; WAKELIN, A. M. Resistance to glyphosate from altered herbicide
translocation patterns. Pest Management Science, London, v. 64, n. 4, p. 372-376, 2008.
PRESTON, C. et al. A decade of glyphosate-resistant Lolium around the World: mechanisms,
genes, fitness, and agronomic management. Weed Science, Lawrence, v. 57, n. 4, p. 435-441,
2009.
QUINN, J.P. Interactions of the herbicides glyphosate and glufosinate (phosphinothricin) with
the soil microflora. In: ALTMAN, J. Pesticides interactions in crop production. Beneficial
and deleterious effects. Boca Raton: CTC Press, 1993. p. 245-265.
REAM, J. E. et al. EPSP synthase: binding studies using isothermal titration microcalorimetry
and equilibrium dialysis and their implications for ligand recognition and kinetic mechanism.
Biochemistry, Washington, v. 31, p. 5528-5534, 1992.
REDDY, K. N.; RIMANDO, A. M.; DUKE, S. O. Aminomethylphosphonic acid, a
metabolite of glyphosate, causes injury in glyphosate-treated, glyphosate-resistant soybean.
Journal of Agriculture and Food Chemistry, Washington, v. 52, p. 5139-5143, 2004.
REDDY, K. N. et al. Aminomethylphosphonic acid accumulation in plant species treated with
glyphosate. Journal of Agriculture and Food Chemistry, Washington, v. 56, n. 6, p. 2125-
2130, 2008.
ROBERTSON, R. Physiological and biochemical characterization of glyphosate resistant
Ambrosia trifida L. 2010. 88 f. Dissertation (Master of Science) - Department of Horticulture
and Landscape Architecture, Purdue University, West Lafayette, 2010.
RODRIGUES, J. J. V.; WORSHAM, A. D.; CORBIN, F. T. Exudation of glyphosate from
wheat (Triticum aestivum) plants and its effects on interplanted corn (Zea mays) and soybeans
(Glycine max). Weed Science, Lawrence, v. 30, n. 3, p. 316-320, 1982.
RODRIGUES, B. N.; ALMEIDA, B. N. Guia de herbicidas. 5.ed. Londrina: Edição dos
Autores, 2011. 697 p.
ROMAN, E. S. et al. Resistência de azevém (Lolium multiflorum) ao herbicida glyphosate.
Planta Daninha, Viçosa, v. 22, n. 2, p. 301-306, 2004.
SAARI, L. L.; COTTERMAN, J. C.; THILL, D. C. Resistance to acetolactate synthase
inhibiting herbicides. In: POWLES, S. B.; HOLTUM, J. A. M. Herbicide resistance in
plants: biology and biochemistry. Boca Raton: CRC Press, 1994. 353 p.
SALAS, R. et al. EPSPs gene amplification in glyphosate-resistant Italian ryegrass (Lolium
perenne spp. multiflorum) from Arkansas. Pest Management Science, London, v. 68, n. 9, p.
1223-1230, 2012.

113
SALAS, R. et al. EPSPs gene amplification in glyphosate-resistant Italian ryegrass (Lolium
perenne spp. multiflorum) from Arkansas. Journal of Agricultural and Food Chemistry,
Washington, v. 63, n. 25, p. 5885-5893, 2015.
SALMINEN, J. P. et al. Seasonal variation in the content of hydrolysable tannins in leaves of
Betula pubescens. Phytochemistry, London, v. 57, p. 15-22, 2001.
SAMMONS, R. D.; GAINES, T. A. Glyphosate resistance: state of knowledge. Pest
Management Science, London, v. 70, n. 9, p. 1367-1377, 2014.
SANTOS, F. M. et al. Effects of glyphosate on the physiological parameters of horseweed.
Revista Brasileira de Ciências Agrárias, Recife, v. 9, n. 4, p. 519-525, 2014.
SATICHIVI, N. M. et al. Absorption and translocation of glyphosate isopropylamine and
trimethysulfonium salts in Abutilon theophrasti and Setaria faberi. Weed Science, Lawrence,
v. 48, p. 675-679, 2000.
SCHRUBBERS, L. C. et al. Glyphosate spray drift in Coffea arabica – sensitivity of coffee
plants and possible use of shikimic acid as a biomarker for glyphosate exposure. Pesticide
Biochemistry and Physiology, New York, v. 115, p. 15-22, 2014.
SERRA, A. A. et a. Low environmentally relevant levels of bioactive xenobiotics and
associated degradation cause cryptic perturbations of metabolism and molecular stress
responses in Arabidopsis thaliana. Journal of Experimental Botany, Oxford, v. 64, n. 10, p.
2753-2766, 2013.
SERVAITES, J. C.; TUCCI, M. A.; GEIGER, D. R. Glyphosate effects on carbon
assimilation, ribulose bisphosphate carboxylase activity, and metabolite levels in sugar beet
leaves. Plant Physiology, Chicago, v. 85, p. 370-374, 1987.
SIEHL, D. Inhibitors of EPSPs synthase, glutamine synthetase and histidine synthesis. In:
ROE, R.; BURTON, J.; KUHR, R. Herbicide activity: toxicology, biochemistry and
molecular biology. Amsterdam: IOS Press, 1997. p. 37-67.
SHANER, D. L. Effects of glyphosate on transpiration. Weed Science, Lawrence, v. 26, n. 5,
p. 513-516, 1978.
SHANER, D. L.; LYON, J. L. Interaction with aromatic amino acids on transpiration in
Phaseolus vulgaris. Weed Science, Lawrence, v. 28, p. 31-35, 1980.
SHANER, D. L. Role of translocation as a mechanism of resistance to glyphosate. Weed
Science, Lawrence, v. 57, n. 1, p. 118-123, 2009.
SHRESTHA, A. et al. Growth, phenology, and intraspecific competition between glyphosate-
resistant and glyphosate-susceptible horseweeds (Conyza canadensis) in the San Joaquin
Valley of California. Weed Science, Lawrence, v. 58, n. 2, p. 147-153, 2010.
SILVA, A. A.; FERREIRA, F. A.; FERREIRA, L. R. Controle de plantas daninhas.
Brasília, DF: ABEAS, 2005. 260 p.

114
SILVA, M. R. M.; DURIGAN, J. C. Períodos de interferência das plantas daninhas na cultura
do arroz de terras altas. I – Cultivar IAC 202. Planta Daninha, Viçosa, v. 24, n. 4, p. 685-
694, 2006.
SILVA, A. A.; VARGAS, L.; FERREIRA, E. A. Herbicidas: resistência de plantas. In:
SILVA, A. A.; SILVA, J. F. Tópicos em manejo de plantas daninhas. Viçosa: UFV, 2007.
p. 279-320.
SILVA, D. R. O. et al. Glyphosate-resistant hairy fleabane competition in RR soybean.
Bragantia, Campinas, v. 73, n. 4, p. 451-457, 2014.
SINGH, B. K.; SHANER, D. L. Rapid determination of glyphosate injury to plants and
identification of glyphosate-resistant plants. Weed Technology, Lawrence, v. 12, n. 3, p. 527-
530, 1998.
SOCIEDADE BRASILEIRA DA CIÊNCIA DAS PLANTAS DANINHAS. Procedimentos
para instalação, avaliação e análise de experimentos com herbicidas. Londrina: SBCPD,
1995. 42 p.
STAUFFER, M. E. et al. Chemical shift mapping of shikimate-3 phosphate binding to the
isolated N-terminal domain of 5-enolpyruvylshikimate-3-phosphate synthase. Federation of
European Biochemical Societies, New York, v. 499, n. 1-2, p. 182-186, 2001.
STREIBIG, J. C. Herbicide bioassay. Weed Research, Oxford, v. 28, n. 6, p. 479-484, 1988.
TAIZ, L.; ZEIGER, E. Fisiologia Vegetal. 3 ed. Porto Alegre: Artmed, 2004. 719 p.
TAIZ, L.; ZEIGER, E. Fisiologia Vegetal. 3 ed. Porto Alegre: Artmed, 2006. 719 p.
TARDIF, F. J.; RACJAN, I.; COSTEA, M. A mutation in the herbicide target site
acetohydroxyacid synthase produces morphological and structural alterations and reduces
fitness in Amaranthus poewllii. New Phytologist, London, v. 169, n. 2, p. 251-264, 2006.
THIEL, H.; KLUTH, C.; VARRELMANN, M. A new molecular method for the rapid
detection of a metamitron-resistant target site in Chenopodium album. Pest Management
Science, London, v. 66, n. 9, p. 1011-1017, 2010.
TRANKAMP, S. et al. Temporally resolved GC-MS-based metabolic profiling of herbicide
treated plants treated reveals that changes in polar primary metabolites alone can distinguish
herbicides of differing mode of action. Metabolomics, Boston, v. 5, n. 3, p. 277-291, 2009.
VARGAS, L. et al. Alteração das características biológicas dos biótipos de azevém (Lolium
multiflorum) ocasionada pela resistência ao herbicida glyphosate. Planta Daninha, Viçosa, v.
23, n. 1, p. 153-160, 2005.
VARGAS, L. et al. Buva (Conyza bonariensis) resistente ao glyphosate na região sul do
Brasil. Planta Daninha, Viçosa, v. 25, n. 3, p. 573-578, 2007a.

115
VARGAS, L.; MOREAES, R. M. A.; BERTO, C. M. Herança da resistência de azevém
(Lolium multiflorum) ao glyphosate. Planta Daninha, Viçosa, v. 25, n. 3, p. 567-571, 2007b.
VARGAS, L. et al. Resistência de plantas daninhas no Brasil: histórico, custo, e o desafio do
manejo no futuro. In: INIA. Viabilidad del glifosato em sistemas productivos sustentables.
Montevideo: Amalia Ríos, 2013. p. 99-110.
VARGAS, L. et al. Glyphosate influence on the physiological parameters of Conyza
bonariensis biotypes. Planta Daninha, Viçosa, v. 32, p. 151-159, 2014.
VAVASSEUR, A.; RAGHAVENDRA, A. S. Guard cell metabolism CO2 sensing. New
Phytologist, London, v. 165, n. 3, p. 665-682, 2005.
VELINI, E. D. et al. Modo de ação do glyphosate. In: VELINI, E. D. et al. Glyphosate.
Botucatu: FEPAF, 2009a. 496 p.
VELINI, E. D. et al. Glyphosate. Botucatu: FEPAF, 2009b. 496 p.
VELINI, E. D. et al. Modo de ação do glyphosate. In: VELINI, E. D. et al. Glyphosate: uso
sustentável. Botucatu: FEPAF, 2012. p. 39-57.
VILA-AIUB, M. M.; NEVE, P.; POWLES, S. B. Resistance cost os a cytochrome P540
herbicide metabolism mechanism but not an ACCase target site mutation in a multiple
resistant Lolium rigidum population. New Phytologist, London, v. 167, n. 3, p. 787-796,
2005.
VILA-AIUB, M. M. et al. Glyphosate resistance in Sorghum halepense and Lolium rigidum is
reduced at suboptimal growing temperatures. Pest Management Science, London, v. 69, n. 2,
p. 228-232, 2013.
VILA-AIUB, M. M. et al. No fitness cost of glyphosate resistance endowed by massive epsps
gene amplification in Amaranthus palmeri. Planta, New York, v. 239, n. 4, p. 793-801,
2014.
WAKELIN, A. M.; LORRAINE-COLWILL, D. F.; PRESTON, C. Glyphosate resistance in
four different populations of Lolium rigidum is associated with reduced translocation of
glyphosate to meristematic zone. Weed Research, Oxford, v. 44, n. 6, p. 453-459, 2004.
WANAMARTA, G. D.; PENNER, D. Foliar absorption of herbicides. Weed Science,
Lawrence, v. 4, p. 215-231, 1989.
WEAVER, L. M.; HERRMANN, K. M. Dynamics of the shikimate pathway in plants.
Trends in Plant Science, Cambridge, v. 2, n. 9, p. 346-351, 1997.
WHETTEN, R. W.; SEDEROFF, R. R. Phenylalanine ammonia lyase from loblolly pine:
Purification of the enzyme and isolation of complementary DNA clones. Plant Physiology,
Chicago, v. 98, p. 380-386, 1992.

116
WHEELER, D. J. B.; JACOBS, S. W. L.; WHALLEY, R. D. B. Grasses of New South
Wales. Armidale: University of New England Printery, 2002. 450 p.
XAVIER, E. Mecanismos de resistência e resposta aos herbicidas inibidores da ALS e da
PROTOX em Euphorbia heterophylla L. 2014. 211f. Dissertação (Mestrado em
Agronomia) – Universidade Tecnológica Federal do Paraná, Pato Branco, 2014.
YANNICCARI, M. et al. Glyphosate effects on gas exchange and chlorophyll fluorescence
responses of two Lolium perenne L. biotypes with differential herbicide sensitivity. Plant
Physiology and Biochemistry, Paris, v. 57, p. 210-217, 2012.
YAO, H.; TIAN, S. Effects of pre and post-harvest application of salicylic acid or methyl
jasmonate on inducing disease resistance of sweet cherry fruit in storage. Postharvest
Biology and Technology, Amsterdam, v. 35, n. 3, p. 253-262, 2005.
YU, Q.; CAIRNS, A.; POWLES, S. B. Glyphosate, paraquat and ACCase multiple herbicide
resistance evolved in a Lolium rigidum biotype. Planta, New York, v. 225, n. 2, p. 499-513,
2007.
YU, Q. et al. Distinct non-target site mechanism endow resistance to glyphosate, ACCase and
ALS-inhibiting herbicides in multiple herbicide-resistant Lolium rigidum. Planta, New York,
v. 230, n. 4, p. 713-723, 2009.
YU, Q. et al. Evolution of a double amino acid substitution in the 5-enolpyruvylshikimate-3-
phosphate synthase in Eleusine indica conferring high-level glyphosate resistance. Plant
Physiology, Chicago, v. 167, n. 4, p. 1440-1447, 2015.
ZABLOTOWICZ, R. M.; REDDY, K. N. Impact of glyphosate on the Bradyrhizobium
japonicum symbiosis with glyphosate-resistant transgenic soybean: a minireview. Journal of
Environmental Quality, Madison, v. 33, p. 825-831, 2004.
ZABLOTOWICZ, R. M.; REDDY, K. N. Nitrogenase activity, nitrogen content, and yield
responses to glyphosate in glyphosate-resistant soybean. Crop Protection, Amsterdam, v. 26,
n. 3, p. 370-376, 2007.
ZELAYA, I. A. et al. Evaluation of spectrophotometric and hplc methods for shikimic acid
determination in plants: models in glyphosate-resistant and susceptible crops. Journal of
Agriculture and Food Chemistry, Washington, v. 59, n. 6, p. 2202-2212, 2011.
ZOBIOLE, L. H. S. et al. Glyphosate reduces shoot concentrations of mineral nutrients in
glyphosate-resistant soybeans. Plant and Soil, Washington, v. 328, p. 57-69, 2009.
ZOBIOLE, L. H. S. et al. Effetcs of glyphosate on symbiotic N2 fixation and nickel
concentration in glyphosate-resistant soybeans. Applied Soil Ecology, New York, v. 44, n. 2,
p. 176-180, 2010a.
ZOBIOLE, L. H. S. et al. Water use efficiency and photosynthesis of glyphosate-resistant
soybean as affected by glyphosate. Pesticide Biochemistry and Physiology, New York, v.
97, n. 3, p. 182-193, 2010b.

117
ZOBIOLE, L. H. S. et al. Glyphosate affects chlorophyll, nodulation and nutrient
accumulation of “second generation” glyphosate-resistant soybean (Glycine max L.).
Pesticide Biochemistry and Physiology, New York, v. 99, p. 53-60, 2011.
ZOBIOLE, L. H. S. et al. Glyphosate effects on photosynthesis, nutrient accumulation, and
nodulation in glyphosate-resistant soybean. Journal of Plant Nutrition and Soil Science,
Weinheim, v. 175, n. 2, p. 319-330, 2012.





























