Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DE PELOTASPrograma de Pós-Graduação em Fitossanidade
Tese
Cochonilhas-farinhentas associadas à videira na Serra Gaúcha,
bioecologia e controle de Planococcus citri (Risso, 1813)
(Hemiptera: Pseudococcidae)
Wilson José Morandi Filho
Pelotas, 2008

Wilson José Morandi Filho
Cochonilhas-farinhentas associadas à videira na Serra Gaúcha,
bioecologia e controle de Planococcus citri (Risso, 1813)
(Hemiptera: Pseudococcidae)
Tese apresentada ao Programa de Pós-graduação em Fitossanidade da Universidade Federal de Pelotas, como requisito parcial à obtenção do título de Doutor em Ciências (Área do conhecimento: Entomologia)
Orientador: Dr. Anderson Dionei Grützmacher
Co-orientador: Dr. Marcos Botton
Pelotas, 2008

Dados de catalogação na fonte:( Marlene Cravo Castillo – CRB-10/744 )
M829c Morandi Filho, Wilson José Cochonilhas-farinhentas associadas à videira na Serra
Gaúcha, bioecologia e controle de Planococcus citri(Risso,1813) (Hemiptera: Pseudococcidae ) / Wilson José Morandi Filho. - Pelotas, 2008.
91f. : il.
Tese (Doutorado em Entomologia) – Programa de Pós-Graduação em Fitossanidade. Faculdade de Agronomia Eliseu Maciel. Universidade Federal de Pelotas. - Pelotas, 2008, Anderson Dionei Grützmacher, Orientador; co-orientador Marcos Botton.
. 1.Planococcus citri 2. Ciclo biológico 3. Constante
termica 4. Controle quimico 5. Graus-dia 6. Uva I Grützmacher, Anderson Dionei II .Título.
CDD 595.7

Banca Examinadora:
Dr. Anderson Dionei Grützmacher (Orientador)
Dra. Ana Paula Schneid Afonso
Dr. Dori Edson Nava
Dr. Régis Sivori Silva dos Santos
Dra. Vera Regina dos Santos Wolff

A minha mãe Vera Lúcia, meu pai Wilson, meu irmão Vinícius e minha
madrinha Sirlei pelo amor incondicional, carinho, exemplo, confiança e
auxílio para que meus sonhos sempre se tornassem realidade. Minha
eterna gratidão!
Dedico e Ofereço
A Deus pela saúde e disposição para superar obstáculos, oferecendo
sempre oportunidade de vivenciar momentos tão especiais.
Agradeço

AGRADECIMENTOS
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior
(CAPES) pela concessão da bolsa de estudos e pesquisa, fundamental para
realização do curso de Doutorado.
À Universidade Federal de Pelotas – UFPel, pela oportunidade de
realização do curso de Doutorado em Fitossanidade/Entomologia.
À Empresa Brasileira de Pesquisa Agropecuária (Embrapa) – Centro
Nacional de Pesquisa de Uva e Vinho (CNPUV), na pessoa do Chefe Geral Dr.
Alexandre Hoffmann, por ter permitido, através da parceria UFPel – Embrapa, a
realização do trabalho de conclusão do Curso de Doutorado, juntamente com as
atividades de pesquisa da Empresa.
Ao Dr. Anderson Dionei Grützmacher, Professor Associado do
Departamento de Fitossanidade da Faculdade de Agronomia “Eliseu Maciel”,
Universidade Federal de Pelotas (PPGFs/FAEM/UFPel) pela oportunidade de
realização do curso e orientação.
Ao pesquisador e professor Dr. Marcos Botton, da Embrapa Uva e Vinho,
pela co-orientação, pela idéia do trabalho a ser realizado, orientando em todos os
momentos, sempre disposto a indicar o melhor caminho. Agradeço por todo o
trabalho burocrático relacionado à aquisição de materiais, equipamentos e grupo
de auxílio, pelas oportunidades oferecidas durante este período como
participação em congressos, projetos, publicações e pelo fornecimento de
material bibliográfico ao longo do curso. Pelos ensinamentos, confiança,
paciência, verdadeira e sincera amizade, compreensão, estímulo e exemplo de
profissional a ser seguido.

Aos pesquisadores e funcionários da Embrapa Uva e Vinho pela acolhida,
ensinamentos, amizade e profissionalismo em especial: Dra. Ana Beatriz C.

Czermainski, Dr. Lucas da R. Garrido, Dra. Lucimara Antoniolli, Dra. Rosemary
Hoff, Dr. Saulo de Jesus Soria, Sandra de Souza Sebben e Viviane Zanella.
Aos professores Dr. Alci Enimar Loeck e Dr. Mauro Silveira Garcia do
Departamento de Fitossanidade, pelos ensinamentos e amizade durante a
realização do curso de Pós-Graduação.
À amiga, professora e pesquisadora Dra. Mari Inês Carissimi Boff, pela
amizade, estímulo e boa vontade em auxiliar-me, por ter despertado meu
interesse pela Entomologia. Mesmo estando distante, sempre esteve presente
nesta importante etapa de minha vida.
Aos pesquisadores-taxonomistas Dra. Maria Cristina Granara de Willink
(Conicet-Argentina) e Dr. Ernesto Prado (INIA/Chile), pelo auxilio na identificação
e confirmação dos exemplares de cochonilhas.
À amiga Aline Bertin, Bolsista de Iniciação Científica – CNPq, pela
amizade, estímulo, dedicação, revisão dos abstracts, pelo apoio para a realização
dos experimentos da tese.
À amiga e funcionária do Laboratório de Entomologia da Embrapa Uva e
Vinho, Vânia Maria Ambrosi Sganzerla, pelo auxilio, apoio, verdadeira amizade,
carinho, conselhos valiosos tanto na vida profissional como pessoal, convivência
e exemplo de profissionalismo.
Ao funcionário e amigo Léo Antonio Carollo, do Laboratório de Entomologia
da Embrapa Uva e Vinho, pela amizade e colaboração nos trabalhos de pesquisa.
Às bibliotecárias da Embrapa Uva e Vinho Kátia Midori Hiwatashi,
Fernanda Pazini e Claúdia Ana Reczko, pela amizade, auxílio nas revisões e
citações bibliográficas.
À amiga Aline Nondillo, pelo carinho, auxilio em todos os momentos,
palavras de incentivo, momentos de descontração e amizade incomparável.
Aos amigos que conquistei na casa dos estagiários da Embrapa Uva e
Vinho em especial Cristiano João Arioli, Cristiane Müller, Elisabete dos Santos
Barbosa, Fábio Nascimento da Silva, Jandora Severo Poli, Marcelo Zart, Magda
Andréia Tessmer, Patrik Luiz Pastori e Wagner da Roza Härter pela amizade,
convívio, apoio incondicional e festas durante a vivência na pousada.
Aos colegas e amigos do Departamento de Fitossanidade da FAEM/UFPel,
em especial Aldomário Santo Negrisoli Júnior, Carla Ruth de Carvalho Barbosa,

Cristine Ramos Zimmer, Fabrizio Pinheiro Giolo, Getúlio Jorge Stefanello Júnior e
Regina da Silva Borba, pela convivência, incentivo, companheirismo.
Aos colegas e amigos do Laboratório de Entomologia da Embrapa Uva e
Vinho, especialmente Aline Bertin, Cristiane Müller, Marcelo Zart, Patrícia
Poggere, Rafael Meirelles, Rodrigo Fornari, e Wilson Sampaio de Azevedo Filho,
pela convivência, incentivo, companheirismo, colaboração nos trabalhos de
pesquisa e compreensão nos momentos de mau humor.
A todos, que direta ou indiretamente, colaboraram para o êxito deste
trabalho, o meu sincero agradecimento.

“A razão por que os rios e os mares recebem a
homenagem de centenas de córregos das
montanhas, é que eles se acham abaixo dos
últimos. Por isso, o sábio desejando pairar acima
dos demais homens, coloca-se abaixo deles.”
Lao-Tsé

SUMÁRIO
LISTA DE TABELAS........................................................................................... 12
LISTA DE FIGURAS............................................................................................ 15
RESUMO.............................................................................................................. 16
ABSTRACT........................................….............................................................. 18
INTRODUÇÃO GERAL………………………...................………………………… 20
CAPÍTULO 1 - ESPÉCIES DE COCHONILHAS-FARINHENTAS
(HEMIPTERA: PSEUDOCOCCIDAE) ASSOCIADAS À CULTURA DA
VIDEIRA NA REGIÃO DA SERRA GAÚCHA..................................................... 23
1. Introdução........................................................................................................ 23
2. Material e Métodos........................................................................................... 25
3. Resultados e Discussão................................................................................... 26
4. Conclusões....................................................................................................... 33
CAPÍTULO 2 - BIOLOGIA E TABELA DE VIDA DE FERTILIDADE DE
Planococcus citri (RISSO, 1813) (HEMIPTERA: PSEUDOCOCCIDAE) EM
DIFERENTES ESTRUTURAS VEGETATIVAS E CULTIVARES DE
VIDEIRA............................................................................................................... 34
1. Introdução........................................................................................................ 34
2. Material e Métodos........................................................................................... 35
3. Resultados e Discussão................................................................................... 37
3.1 Duração e Viabilidade.................................................................................... 37

3.2 Longevidade e fecundidade........................................................................... 42
3.3 Tabela de Vida de Fertilidade........................................................................ 43
4. Conclusões....................................................................................................... 45
CAPÍTULO 3 - EXIGÊNCIAS TÉRMICAS E ESTIMATIVAS DO NÚMERO
ANUAL DE GERAÇÕES DE Planococcus citri (RISSO, 1813)
(HEMIPTERA: PSEUDOCOCCIDAE) NA CULTURA DA
VIDEIRA............................................................................................................... 46
1. Introdução........................................................................................................ 46
2. Material e Métodos........................................................................................... 47
3. Resultados e Discussão................................................................................... 48
4. Conclusões....................................................................................................... 56
CAPÍTULO 4 - EFEITO DE INSETICIDAS SOBRE A COCHONILHA-
FARINHENTA Planococcus citri (RISSO, 1813) (HEMIPTERA:
PSEUDOCOCCIDAE) NA CULTURA DA VIDEIRA........................................... 57
1. Introdução........................................................................................................ 57
2. Material e Métodos.......................................................................................... 59
2.1 Experimento em casa-de-vegetação.............................................................. 60
2.2 Experimento em vinhedo comercial............................................................... 60
2.3 Análise estatística.......................................................................................... 61
3. Resultados e Discussão................................................................................... 61
3.1 Experimento em casa-de-vegetação.............................................................. 61
3.2 Experimento em vinhedo comercial............................................................... 63
3.2.1 Aplicação via solo...................................................................................... 63
3.2.2 Aplicação via foliar..................................................................................... 67
4. Conclusões....................................................................................................... 69
CONCLUSÕES.................................................................................................... 70
REFERÊNCIAS.................................................................................................... 74

LISTA DE TABELAS
Tabela 1 - Data de coleta, variedade de uva, produtor e localização do
vinhedo com presença de cochonilhas-farinhentas em cachos de videira na
região de Bento Gonçalves. Bento Gonçalves, RS, 2007................................... 27
Tabela 2 - Data de coleta, variedade de uva, produtor e localização do
vinhedo com presença de cochonilhas-farinhentas em raízes de videira. Bento
Gonçalves, RS, 2006........................................................................................... 30
Tabela 3 - Duração (X EP, dias) das fases de desenvolvimento de
Planococcus citri em folhas de videira da cultivar Cabernet Sauvignon, Itália e
Isabel; raízes do porta enxerto 101-14, IAC-572 e cultivar Isabel; bagas da
cultivar Itália em laboratório. Temperatura de 25 2º C; UR: 70 10%;
Fotofase: 12 horas. Bento Gonçalves, RS, 2007................................................. 38
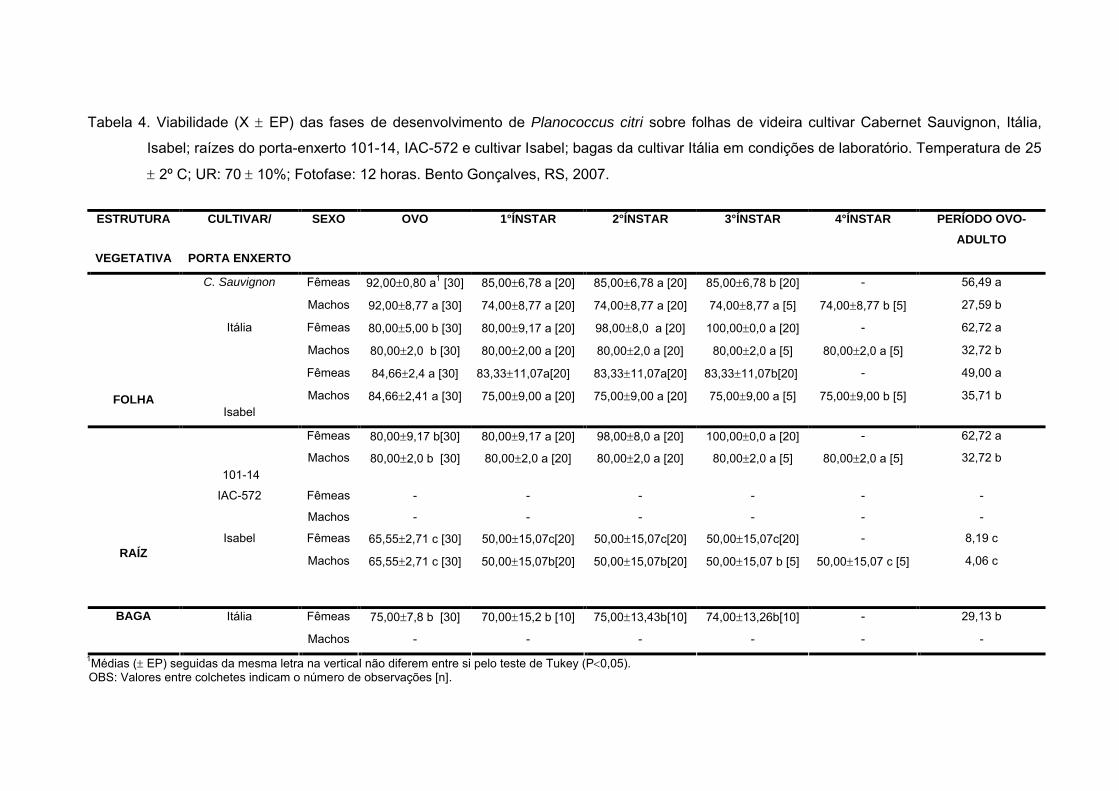
Tabela 4 - Viabilidade (X EP) das fases de desenvolvimento de Planococcus
citri sobre folhas de videira cultivar Cabernet Sauvignon, Itália, Isabel; raízes
do porta enxerto 101-14, IAC-572 e cultivar Isabel; bagas da cultivar Itália em
condições de laboratório. Temperatura de 25 2º C; UR: 70 10%; Fotofase:
12 horas. Bento Gonçalves, RS, 2007................................................................. 39
Tabela 5 - Longevidade (X EP) em dias de Planococcus citri e fecundidade
quando as fêmeas foram criadas sobre folhas de videira da cultivar Cabernet
Sauvignon, Itália, Isabel; raízes do porta enxerto 101-14, IAC-572 e cultivar
Isabel; bagas da cultivar Itália em condições de laboratório. Temperatura de
25 2º C; UR: 70 10%; Fotofase: 12 horas. Bento Gonçalves,
2007..................................................................................................................... 43

Tabela 6 - Duração média de uma geração (T), taxa líquida de reprodução
(Ro), razão infinitesimal de aumento (Rm) e razão finita de aumento () de
Planococcus citri sobre folhas de videira das cultivares Cabernet Sauvignon,
Itália e Isabel; raízes do porta enxerto 101-14, IAC-572 e cultivar Isabel; bagas
da cultivar Itália em condições de laboratório. Temperatura de 25 2º C; UR:
70 10%; Fotofase: 12 horas. Bento Gonçalves, RS, 2007................................ 44
Tabela 7 - Duração (média ± EP), da fase de ovo, 1º, 2º, 3ºinstar e ciclo
biológico (ovo-adulto) de Planococcus citri em folhas de videira da cultivar
Itália, nas temperaturas de 16, 19, 22, 25, 28 e 31 ºC (UR: 70 ± 10%; fotofase
12horas)............................................................................................................... 50
Tabela 8 - Viabilidade média (% ± EP), da fase de ovo, 1º, 2º, 3ºinstar e ciclo
biológico (ovo-adulto) de Planococcus citri mantidos em folhas de videira
cultivar Itália, nas temperaturas de 16, 19, 22, 25, 28 e 31 ºC (UR: 70 ± 10%;
fotofase 12 horas).................................................................................................. 51
Tabela 9 - Limiar térmico inferior de desenvolvimento (Tb), constante térmica
(K), em graus-dia (GD), equação linear da velocidade de desenvolvimento
(1/D) e coeficiente de determinação (R2) da fase de ovo, 1º instar, 2º instar e
3º instar e ciclo biológico (ovo-adulto) de P. citri mantidos em folhas de videira
cultivar Itália, nas temperaturas de 16, 19, 22, 25 e 28ºC (UR: 70 ± 10%;
fotofase 12 horas)................................................................................................ 53
Tabela 10 - Regiões produtoras de uva, Estados da Federação, temperaturas
médias anuais (ºC), graus-dias anuais acumulados (GD) e número provável
de gerações de P.citri por ano (Tb utilizada nos cálculos de 8,16 ºC para o
período de ovo-adulto)......................................................................................... 55
Tabela 11 - Inseticidas avaliados para o controle de Planococcus citri na
cultura da videira. Bento Gonçalves, RS, 2008................................................... 59

Tabela 12 - Número médio de ninfas vivas de primeiro ínstar (X EP) de
Planococcus citri vivas submetidas a diferentes tratamentos em casa-de-
vegetação em plantas novas de videira da cultivar Cabernet Sauvignon, via
solo. Bento Gonçalves, RS, 2007........................................................................ 64
Tabela 13 - Número médio de ninfas de primeiro ínstar (X EP) de
Planococcus citri vivas submetidas a diferentes tratamentos em vinhedo
comercial da cultivar Cabernet Sauvignon, via solo. Bento Gonçalves, RS,
2007..................................................................................................................... 65
Tabela 14 - Número médio de ninfas de primeiro ínstar (X EP) de
Planococcus citri vivas submetidas a diferentes tratamentos em vinhedo
comercial da cultivar Cabernet Sauvignon, via foliar. Bento Gonçalves, RS,
2008..................................................................................................................... 67

LISTA DE FIGURAS
Figura 1 - Porcentagem de vinhedos onde foram constatadas as diferentes
espécies de cochonilhas-farinhentas ocorrentes na Região de Bento
Gonçalves, RS..................................................................................................... 30
Figura 2 - Espécies de Pseudococcidae encontradas nos vinhedos infestados
na Região de Bento Gonçalves, RS e sua porcentagem de ocorrência,
RS........................................................................................................................ 30
Figura 3 - Estimativa do número de gerações/mês de P.citri para os
municípios de Petrolina (PE), São Lourenço (MG), Londrina (PR), Bagé (RS),
Jales (SP) e Bento Gonçalves (RS)..................................................................... 56
Figura 4 - Mortalidade de Planococcus citri após a aplicação via solo dos
inseticidas acetamiprid, imidacloprid e tiametoxam, em plantas de videira com
um ano e 15 anos de idade da cultivar Cabernet Sauvignon em casa-de-
vegetação (CV) e vinhedo comercial (VC). Bento Gonçalves, RS, 2007............ 62

Resumo
MORANDI FILHO, W.J. Cochonilhas-farinhentas associadas à videira na Serra Gaúcha, bioecologia e controle de Planococcus citri (Risso, 1813) (Hemiptera: Pseudococcidae). 2008. 91f. Tese (Doutorado) – Programa de Pós-Graduação em Fitossanidade. Universidade Federal de Pelotas, Pelotas.
Cochonilhas da família Pseudococcidae têm sido relatadas como causadoras de danos diretos (perdas na produção) e indiretos (transmissão de vírus) na cultura da videira. Poucas informações estão disponíveis sobre as espécies presentes nos vinhedos e a bioecologia das mesmas. Neste trabalho foram identificadas as espécies de Pseudococcidae associados à cultura da videira na região da Serra Gaúcha; estudado a biologia e determinado as exigências térmicas de Planococcus citri, estimado o número de gerações anuais nas diferentes regiões produtoras e avaliado o efeito de inseticidas alternativos aos fosforados para o controle da espécie. P. citri, Planococcus minor, Pseudococcus viburni, Pseudococcus maritimus, Dysmicoccus brevipes foram constatadas associadas à cultura da videira na região da Serra Gaúcha, sendo que P. citri foi a espécie mais abundante nos vinhedos. A biologia de P. citri foi estudada em folhas de videira das cultivares Cabernet Sauvignon, Itália e Isabel, bagas da cultivar Itália e raízes da cultivar Isabel e porta-enxertos 101-14 e IAC-572. Em raízes do porta-enxerto IAC-572 a cochonilha não completou o ciclo biológico. Em bagas de uva Itália, o inseto completou a fase de ninfa, porém os adultos foram inférteis. Em folhas de videira das diferentes cultivares, a duração média do período ovo a adulto dos machos foi de 24,630,10 dias com viabilidade de 32% enquanto que as fêmeas duraram 32,000,11 dias com viabilidade de 56%. Em raízes, a duração do ciclo biológico das fêmeas e machos foi de 32,450,19 e 29,500,51 dias, respectivamente. A menor viabilidade do ciclo ninfal foi verificada em raízes de Isabel, sendo de 8,1% para fêmeas e 4,0% para machos enquanto a maior foi obtida em folhas de Cabernet Sauvignon, sendo de 62,70% e 32,7% para fêmeas e machos, respectivamente. Em folhas a fecundidade foi de 67,2710,70; 53,331,68 e 66,094,68 ovos por fêmea nas cultivares Cabernet Sauvignon, Itália e Isabel, respectivamente. Enquanto em raízes, foi obtido 30,406,12 e 70,0010,5 ovos por fêmea no porta-enxerto 101-14 e Isabel, respectivamente. A velocidade de desenvolvimento de P. citri aumenta com a elevação da temperatura sendo a faixa de temperatura entre 25 ºC e 28 ºC a mais adequada para a espécie. A temperatura base e a constante térmica para o ciclo total (ovo-adulto) de P. citri em folhas de videira da cultivar Itália foi de 8,16 ºC e 574,71 graus-dia, respectivamente. Com base nas exigências térmicas estimou-se que o inseto pode completar 5,7 gerações anuais em Bento Gonçalves (RS), 6,2 em

Bagé (RS), 6,9 em São Lourenço (MG), 7,9 em Londrina (PR), 9,7 em Jales (SP) e 11,5 em Petrolina (PE). Na avaliação de inseticidas para o controle de P. citriverificou-se que os neonicotinóides acetamiprid (0,60g i.a./planta), imidacloprid (0,70g i.a./planta) e tiametoxam (0,75g i.a./planta) são eficazes no controle de P. citriem plantas de videira com um ano de idade. Entretanto, em plantas com 15 anos de idade de C. sauvignon os inseticidas neonicotinóides acetamiprid, imidacloprid e tiametoxam nas mesmas doses não foram eficazes para o controle da espécie. Ao estudar o efeito dos inseticidas neonicotinóides acetamiprid (6g i.a./100L), imidacloprid (7g i.a./100L), tiametoxam (7,5g i.a./100L) e os reguladores de crescimento buprofezin (25g i.a./100L) e pyriproxyfen (10g i.a./100L) aplicados via foliar observou-se que os mesmos não proporcionam mortalidade significativa de P. citri na cultura da videira após a pulverização nos vinhedos e avaliação da mortalidade em laboratório.
PALAVRAS-CHAVE: Planococcus citri, ciclo biológico, constante térmica, graus-dia, controle químico, uva.

Abstract
MORANDI FILHO, W.J. MEALYBUGS ASSOCIATED IN THE SERRA GAUCHA VINEYARDS, BIOECOLOGY AND CONTROL Planococcus citri (RISSO, 1813) (HEMIPTERA: PSEUDOCOCCIDAE). 2008. 91f. Tese (Doutorado) – Programa de Pós-Graduação em Fitossanidade. Universidade Federal de Pelotas, Pelotas.
Mealybugs of the Pseudococcidae family have been reported as causing direct damage (losses in production) and indirect (transmission of virus) in vineyards. Few information is available about the species occuring in the vineyards and the bioecology of pseudococidae in Brazil. In this work we identified the species of Pseudococcidae associated with vineyards in Serra Gaúcha region; studied the biology and determined the thermal requirements of Planococcus citri estimating the number of annual generations in different producing regions and, evaluated the effect of insecticides alternative to phosphorous for the control of the species. P. citri, Planococcus minor, Pseudococcus viburni, Pseudococcus maritimus, Dysmicoccus brevipes were found associated with the vineyards in the region. P. citri was the most abundant species. The biology of P. citri was studied in leaves of the vine cultivars Cabernet Sauvignon, Italy and Isabel, in fruits of cultivar Italy, in roots of cultivars Isabel and in 101-14 and IAC-572 rootstocks. On roots of the rootstock IAC-572, P. citri was unable to complete the life cycle. The average period from egg to adult males was 24.63 ± 0.10 days with viability of 32% while females lasted 32.00 ± 0.11 days with viability of 56% when fed on leaves of vine cultivars. In roots, the cycleduration of the females and males was 32.45±0.19 and 29.50±0.51 days, respectively. The lowest viability of the nymphal cycle was found in roots of Isabel, 8.1% for females and 4.0% for males while the high viability was obtained in leaves of Cabernet Sauvignon, 62.7% and 32.7% for females and males, respectively. The mealybug P. citri completed the life cycle in leaves of the vine cultivars Cabernet Sauvignon, Italy, Isabel; roots of the rootstocks 101-14 and Isabel. The Italy grape berries allowed the nymphal development of P. citri, however the adults were infertile. The roots of the door graft IAC-572 are immune to P. citri. The speed of development of P. citri increases with temperature elevation. The temperatures of 25°C and 28°C were the most appropriate for the insect development. The threshold temperature and thermal constant for the full cycle (egg-adult) of P. citri in vine leaves of the cultivar Italy was 8.16°C and 574.71 degree days, respectively. Based on the thermal requirements, we estimated that the insect may have 5.7 annual generations in Bento Gonçalves (RS), 6.2 in Bagé (RS), 6.9 in São Lourenço (MG), 7.9 in Londrina ( PR), 9.7 in Jales (SP) and 11.5 in Petrolina (PE). In the assessment of insecticides for the control of the insect, the neonicotinoids acetamiprid (0.60 g ai / plant), imidacloprid (0.70 g ai / plant) and thiamethoxam (0.75 g ai plant) were effective in the control of P. citri in vine plants with 1 year old. In adult plants (15 years) the same treatments and doses were not effective to pest control. Foliar spray of acetamiprid (6

20
ia/100L), imidacloprid (7 ia/100L), thiamethoxam (7.5 g ia/100L) and insect growth regulators buprofezin (25 ia/100L) and pyriproxyfen (10g ai 100L) were not effective to control P. citri.
KEY-WORDS: citrus mealybug, biological life cycle, thermal requirements, degree-day, chemical control, grape

INTRODUÇÃO GERAL
A produção de uvas no Brasil encontra-se principalmente nas regiões Sul,
Sudeste e Nordeste com destaque para os Estados do Rio Grande do Sul, Santa
Catarina, Paraná, São Paulo, Minas Gerais e Pernambuco (PROTAS et al., 2002). O
Rio Grande do Sul concentra aproximadamente 54% da área cultivada no Brasil que
é de 89.062 ha sendo o principal produtor de uvas para processamento (IBGE,
2007).
A vitivinicultura é uma atividade de grande importância econômica para o Rio
Grande do Sul, principalmente para os produtores localizados na região da Serra
Gaúcha, geograficamente conhecida como Encosta Superior da Serra do Sudeste e,
mais recentemente, nos municípios localizados na metade sul do Estado, onde a
atividade encontra-se em plena expansão (PROTAS et al., 2002).
Em todas as áreas vitivinícolas do mundo, as pragas e doenças constituem-se
num dos maiores obstáculos a expansão do cultivo da videira, afetando tanto a
quantidade como a qualidade do produto final, sendo consideradas limitantes ao
desenvolvimento da atividade (KUHN; NICKEL, 1998). Um dos fatores marcantes da
viticultura brasileira é a freqüente ocorrência de vírus nas diferentes regiões
produtoras o que resulta na degenerescência das plantas, podendo culminar com a
sua morte (REIS; MELO, 1984; REIS et al., 1998; FAJARDO et al., 2003; KISHINO
et al., 2007). Este tem sido um dos principais motivos para a importação de material
propagativo livre de vírus realizada pelo Brasil nos últimos anos (FAJARDO et al.,
2003, KUHN; FAJARDO, 2004). Mesmo com a introdução deste material “limpo”,
observações de campo têm indicado uma dispersão crescente de vírus nos
vinhedos, provavelmente associada à presença de insetos vetores.
Os insetos da família Pseudococcidae são conhecidos como cochonilhas-
farinhentas por apresentarem o corpo recoberto por uma secreção pulverulenta de
cera branca (RIPA; RODRIGUEZ, 1999). Estas cochonilhas têm sido as principais

22
responsáveis pela dispersão de vírus na cultura da videira em países como África do
Sul, Argentina, Austrália, Chile, Espanha, Estados Unidos, Itália, Nova Zelândia,
Portugal e Uruguai (GULLAN, 2000; GEIGER; DAANE, 2001; GONZÁLEZ;
VOLOSKY, 2004; CHARLES et al., 2006; CID et al., 2007). Especificamente no
Brasil, as cochonilhas desta família não têm sido relatadas como pragas que
causam danos diretos a produção (HICKEL, 1996, REIS et al., 1998, HAJI et al.,
2001; BOTTON et al., 2003b). Entretanto, as mesmas tem sido registradas como de
ocorrência frequente e prejudicial em uvas de mesa principalmente do grupo Itália
(BOTTON et al., 2003; MORANDI FILHO et al., 2007; EMBRAPA UVA E VINHO,
2007, KISHINO et al., 2007).
O dano de Pseudococcidae na cultura da videira decorre da sucção de seiva
e injeção de toxinas sendo sua presença facilmente notada, pois forma colônias de
coloração branca farinhosa nos cachos no período da colheita (FLAHERTY et al.,
1992; GONZÁLEZ, 2003). As secreções do inseto contêm açúcares que são
aproveitados por formigas doceiras, servindo também como substratos para o
desenvolvimento da fumagina (FOLDI; SORIA, 1989; ARTIGAS, 1994; GONZÁLEZ
et al., 2001; AFONSO, 2005). A presença da cochonilha nos rácimos aumenta
significativamente o descarte da fruta e, naquela destinada à exportação, caso seja
identificada a presença do inseto, pode haver restrições quarentenárias
(GONZÁLEZ; VOLOSKY, 2004).
Até o momento, não foi realizado um levantamento sistemático das espécies
de Pseudococcidae associadas aos vinhedos no Brasil. Dentre as espécies
relatadas para a cultura no Brasil destacam-se Pseudococcus maritimus (Ehrhorn,
1900), Pseudococcus longispinus (Targioni-Tozetti, 1867), Pseudococcus viburni
(Signoret, 1875), Planococcus citri (Risso, 1813) e Planococcus ficus (Signoret,
1875) (SILVA et al., 1968; WOERNER, 1983; FOLDI; SORIA, 1989; AFONSO, 2005;
KUNIYUKI et al., 2005; FOLDI; KOZÁR, 2006; KUNIYUKI et al., 2006).
As cochonilhas da família Pseudococcidae não têm sido relatadas como
pragas-chave causando danos diretos à produção (BOTTON et al., 2003a). A
importância da família na cultura diz respeito principalmente a transmissão de vírus
(GLRaV 3 e GVB) já comprovado para a espécie P. longispinus (KUNIYUKI et al.,
2005; KUNIYUKI et al., 2006).
Quando a população de cochonilhas-farinhentas nos parreirais é elevada, os
produtores geralmente aplicam inseticidas fosforados, que são altamente tóxicos,

23
pouco seletivos aos inimigos naturais e ainda possuem grande intervalo de carência
existindo a possibilidade de deixar resíduos nos frutos (GONZÁLEZ et al., 1995,
LARRIAN, 1999; WALTON; PRINGLE, 1999; BECERRA, 2001; BOTTON et al.,
2003a; EMATER, 2003; ERAZO, 2004). Tais fatos reforçam a necessidade de se
avaliar novas alternativas de controle, principalmente quando a videira é cultivada no
sistema de produção integrada e/ou orgânica.
O controle da cochonilha tem sido uma prática pouco realizada pelos
vitivinicultores devido ao hábito das colônias localizarem-se nas raízes e/ou sob a
casca, o que dificulta a visualização da praga. Nestes casos, o monitoramento da
cochonilha nos vinhedos seria fundamental para se detectar os focos de infestação e
o período de migração das ninfas do solo para a parte aérea das plantas, período
em que é indicado o controle (GONZÁLEZ, 2003).
As informações disponíveis no Brasil a respeito da taxonomia, bioecologia e
métodos de controle dessa família associada à cultura da videira são escassas. Os
produtores, não tendo uma orientação específica para o manejo da praga acabam
utilizando técnicas de manejo recomendadas para outras culturas, ou então não
adotam qualquer método de controle. A aplicação foliar de inseticidas também não
tem sido eficaz devido à localização da praga no solo ou sob a casca das plantas
(GEIGER; DAANE, 2001). Inseticidas neonicotinóides e reguladores de crescimento
já avaliados em outros países (GONZÁLEZ, 2002; BENTLEY et al., 2004) poderão
servir de alternativa aos inseticidas fosforados para o manejo da espécie, permitindo
um controle localizado nos vinhedos.
Os objetivos deste trabalho foram identificar as espécies de Pseudococcidae
associadas à cultura da videira na região da Serra Gaúcha; estudar a biologia e
determinar as exigências térmicas de P. citri, estimando o número de gerações
anuais nas diferentes regiões produtoras; e, conhecer o efeito de inseticidas
alternativos aos fosforados para o controle da espécie.

CAPÍTULO 1
ESPÉCIES DE COCHONILHAS-FARINHENTAS (HEMIPTERA:
PSEUDOCOCCIDAE) ASSOCIADAS À CULTURA DA VIDEIRA NA REGIÃO DA
SERRA GAÚCHA
1 Introdução
As cochonilhas-farinhentas (Hemiptera: Pseudococcidae) têm sido relatadas
na cultura da videira causando danos nos cachos durante a colheita, principalmente
nas uvas finas de mesa (MORANDI FILHO et al., 2007; KISHINO et al., 2007).
Nestes vinhedos, os prejuízos causados pelas cochonilhas são resultantes das
secreções que contêm açúcares e aminoácidos, os quais servem de substrato para
o desenvolvimento de fungos como a fumagina (Capnodium sp.), que depreciam os
cachos para comercialização (ARTIGAS, 1994; GONZÁLEZ et al., 2001; AFONSO,
2005). Além disso, a presença de Pseudococcidae nos racimos aumenta
significativamente o descarte da fruta e, naquela destinada à exportação, caso seja
identificada a presença da praga, pode haver restrições quarentenárias
(GONZALEZ; VOLOSKY, 2004).
A vitivinicultura é uma atividade de significativa importância econômica para o
Rio Grande do Sul, principalmente para os produtores localizados na região da Serra
Gaúcha, geograficamente conhecida como Encosta Superior da Serra do Sudeste
(PROTAS et al., 2002). Na região, a videira tem sido cultivada principalmente para
fins de processamento e neste caso, o ataque das cochonilhas-farinhentas não tem
sido relatado como responsáveis por prejuízos diretos a produção (BOTTON et al.,
2003a).
No entanto, a incidência de viroses com destaque para a causada pelo vírus
do enrolamento da videira GLRaV-3 (Grapevine leafroll associated virus 3) tem sido

25
constatado nas diferentes cultivares, afetando tanto a qualidade do produto final
como a longevidade dos vinhedos (KUHN; NICKEL, 1998; FAJARDO et al., 2003). A
principal estratégia para o manejo das viroses tem sido a implantação de novos
parreirais com material vegetativo livre de vírus (FAJARDO et al., 2003; KUHN;
FAJARDO, 2004). No entanto, mesmo com a introdução deste material “limpo”,
observações de campo têm indicado um aumento crescente na incidência de viroses
nos vinhedos, provavelmente associada a presença de insetos vetores (FAJARDO
et al., 2003; MORANDI FILHO et al., 2007).
As cochonilhas-farinhentas têm sido relatadas como as principais
responsáveis pela dispersão de vírus na cultura da videira em países como África do
Sul, Argentina, Austrália, Chile, Espanha, Estados Unidos, Itália, Nova Zelândia,
Portugal e Uruguai (CHARLES et al., 2006). No Brasil, as espécies de
Pseudococcidae relatadas para a cultura são P. citri (SILVA et al., 1968; WOERNER,
1983; FOLDI; SORIA, 1989); P. longispinus (SILVA et al., 1968; WOERNER, 1983;
FOLDI; SORIA, 1989; KUNIYUKI et al., 2005), P. viburni (FOLDI; SORIA, 1989;
AFONSO, 2005), P. ficus (SILVA et al., 1968; WOERNER, 1983; FOLDI; SORIA,
1989; FOLDI; KOZÁR, 2006) e P. maritimus (Ehrhorn, 1900) (FOLDI; SORIA, 1989).
Estas informações foram baseadas em coletas e identificações pontuais, sem
haver um inventário mais sistematizado sobre a incidência de Pseudococcidae nas
diferentes regiões produtoras. Além disso, a taxonomia do grupo é complexa
levando a problemas de identificações incorretas e sinonímias (SANTA-CECÍLIA et
al., 2002).
O conhecimento das espécies de Pseudococcidade associadas aos vinhedos
é fundamental para o manejo das viroses que ocorrem na cultura. No Brasil, o
controle das cochonilhas vetoras de vírus da videira não tem sido implementado
mesmo com a comprovação da transmissão de GLRaV-3 e GVB por P. longispinus
(KUNIYUKI et al., 2005; KUNIYUKI et al., 2006) e a constatação de que P. viburni
adquire os mesmos vírus, embora não tenha sido comprovada a transmissão
(AFONSO, 2005). Em outros países, entrentanto, o controle das cochonilhas vetoras
é prioritário visando reduzir a dispersão dos vírus na cultura (ROSCIGLIONE;
CASTELLANO, 1985; CABALEIRO; SEGURA, 1997a; GODINHO; FRANCO, 2001;
GEIGER; DAANE, 2001; GULLAN, 2000; GOLINO et al., 2002; CHARLES et al.,
2006; CID et al., 2007).

26
Este trabalho foi realizado com o objetivo de conhecer as espécies e a incidência
de cochonilhas-farinhentas associadas aos vinhedos na região da Serra Gaúcha
visando fornecer subsídios para futuros estudos de bioecologia, monitoramento e
controle.
2 Material e Métodos
As espécies de Pseudococcidae associadas à cultura da videira foram
avaliadas na safra 2006/07 durante a colheita (janeiro a março). A amostragem foi
realizada no momento da entrega da uva nas vinícolas selecionando-se produtores
da região da Serra Gaúcha. De cada vinhedo, foram coletados 50 cachos de uvas
por vinhedo num total de 131 produtores (Tab. 1), os quais foram triados na
plataforma de entrega das vinícolas coletando-se as fases de desenvolvimento das
cochonilhas presentes (ninfas e/ou fêmeas com ou sem ovissaco). As cochonilhas
encontradas foram levadas no próprio cacho para o Laboratório de Entomologia da
Embrapa Uva e Vinho, localizado em Bento Gonçalves, RS. No laboratório (25±1°C
de temperatura, 70±10% de umidade relativa e 14 horas de fotofase) as formas
coletadas foram individualizadas em abóboras (Cucurbita maxima Duchesne) da
variedade Cabotchá acondicionadas em bandejas plásticas cobertas com tecido voil
conforme a metodologia descrita por Lepage (1942). Este procedimento foi adotado,
pois é comum mais de uma espécie de Pseudococcidae estar associada ao mesmo
hospedeiro (SANTA-CECÍLIA et al., 2007). Ao atingir a fase adulta, as fêmeas foram
preparadas em lâminas e identificadas com base na chave de Williams; Willink
(1992) e confirmadas pela Drª. Maria Cristina Granara de Willink do Instituto Superior
de Entomologia, Facultad de Ciências Naturales e Instituto y Fundacion Miguel Lillo
de Tucumán, Argentina. As lâminas estão depositadas na coleção de referência no
laboratório de Entomologia da Embrapa Uva e Vinho (AZEVEDO-FILHO, 2007).
Paralelamente a este trabalho, conforme a demanda dos produtores foram
visitadas propriedades com relato da presença de cochonilhas nas raízes quando da
renovação dos vinhedos (Tab. 2). As espécies presentes foram coletadas e
identificadas seguindo o mesmo procedimento.

27
3 Resultados e Discussão
Do total de vinhedos amostrados (n=131), foi constatada a presença de
cochonilhas da família Pseudococidae em 35 propriedades ou 27% das amostras
(Tab. 1). As espécies encontradas e porcentagem de ocorrência foram: P. citri
31,43%, Dysmicoccus sp. 22,86%, Planococcus minor (Maskell, 1897) 11,43%,
Dysmicoccus brevipes (Cockerell, 1893) 5,71%, Pseudococcus sp. 17,14%, P.
viburni 8,57% e P. maritimus 2,86% (Fig. 1).
Em cada vinhedo amostrado com registro das cochonilhas, somente foi
encontrada uma espécie por cacho, geralmente com um número reduzido de
indivíduos. Da mesma forma, a porcentagem de cachos infestados por vinhedo foi
sempre igual ou inferior a 4% (Tab. 1) demonstrando uma baixa incidência das
espécies nos cachos nas uvas destinadas ao processamento, comprovando que
Pseudococcidae não é uma praga primária que cause danos diretos nos vinhedos
da região. O reduzido nível de infestação nos cachos também foi registrado por
Kuniyuki et al., (2006) na cultivar Niagara Branca em São Paulo e provavelmente
seja resultado da presença de inimigos naturais que conseguem manter a população
da cochonilha em níveis reduzidos (DE JONG; ALPHEN, 1989; ISLAM; COPLAND,
2000; DAANE et al., 2004; WALTON; PRINGLE, 2005; DAANE et al., 2006), da
baixa adaptação das espécies nas regiões produtoras e/ou da resistência das
cultivares.
Dentre as espécies encontradas, P. citri foi a mais abundante, representando
31,43% das espécies coletadas (Fig. 2). Esta espécie transmite os vírus GLRaV-3,
GVA e GVB (ROSCIGLIONE; CASTELLANO, 1985; CABALEIRO et al., 1997b; CID
et al., 2007). P.viburni, também encontrada neste inventário é vetora de GLRaV-3,
GVA e GVB (GOLINO et al., 2002) enquanto que P. maritimus transmite GLRaV-3
(SFORZA et al., 2000). Para as demais espécies não existem relatos de transmissão
de vírus na cultura, embora D. brevipes seja transmissora de vírus no abacaxi
(SETHER et al., 1998).
A espécie P. citri encontrada de forma mais abundante no inventário
normalmente esta associada à cultura dos citros (GULLAN, 2000), embora também
seja referida como praga do cafeeiro (SANTA CECÍLIA et al., 2007) e outras
frutíferas (WILLIAMS; WILLINK, 1992; WILLIAMS; MOGHADDAM, 2000).
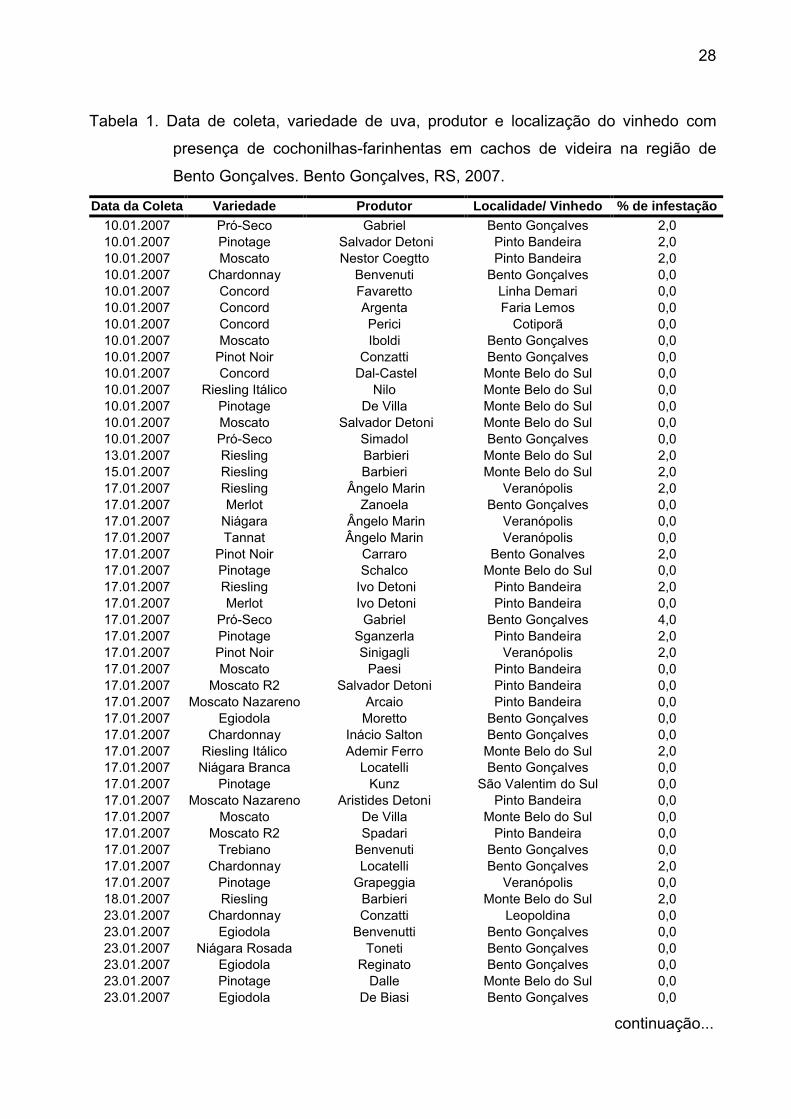
28
Tabela 1. Data de coleta, variedade de uva, produtor e localização do vinhedo com
presença de cochonilhas-farinhentas em cachos de videira na região de
Bento Gonçalves. Bento Gonçalves, RS, 2007.
Data da Coleta Variedade Produtor Localidade/ Vinhedo % de infestação
10.01.2007 Pró-Seco Gabriel Bento Gonçalves 2,010.01.2007 Pinotage Salvador Detoni Pinto Bandeira 2,010.01.2007 Moscato Nestor Coegtto Pinto Bandeira 2,010.01.2007 Chardonnay Benvenuti Bento Gonçalves 0,010.01.2007 Concord Favaretto Linha Demari 0,010.01.2007 Concord Argenta Faria Lemos 0,010.01.2007 Concord Perici Cotiporã 0,010.01.2007 Moscato Iboldi Bento Gonçalves 0,010.01.2007 Pinot Noir Conzatti Bento Gonçalves 0,010.01.2007 Concord Dal-Castel Monte Belo do Sul 0,010.01.2007 Riesling Itálico Nilo Monte Belo do Sul 0,010.01.2007 Pinotage De Villa Monte Belo do Sul 0,010.01.2007 Moscato Salvador Detoni Monte Belo do Sul 0,010.01.2007 Pró-Seco Simadol Bento Gonçalves 0,013.01.2007 Riesling Barbieri Monte Belo do Sul 2,015.01.2007 Riesling Barbieri Monte Belo do Sul 2,017.01.2007 Riesling Ângelo Marin Veranópolis 2,017.01.2007 Merlot Zanoela Bento Gonçalves 0,017.01.2007 Niágara Ângelo Marin Veranópolis 0,017.01.2007 Tannat Ângelo Marin Veranópolis 0,017.01.2007 Pinot Noir Carraro Bento Gonalves 2,017.01.2007 Pinotage Schalco Monte Belo do Sul 0,017.01.2007 Riesling Ivo Detoni Pinto Bandeira 2,017.01.2007 Merlot Ivo Detoni Pinto Bandeira 0,017.01.2007 Pró-Seco Gabriel Bento Gonçalves 4,017.01.2007 Pinotage Sganzerla Pinto Bandeira 2,017.01.2007 Pinot Noir Sinigagli Veranópolis 2,017.01.2007 Moscato Paesi Pinto Bandeira 0,017.01.2007 Moscato R2 Salvador Detoni Pinto Bandeira 0,017.01.2007 Moscato Nazareno Arcaio Pinto Bandeira 0,017.01.2007 Egiodola Moretto Bento Gonçalves 0,017.01.2007 Chardonnay Inácio Salton Bento Gonçalves 0,017.01.2007 Riesling Itálico Ademir Ferro Monte Belo do Sul 2,017.01.2007 Niágara Branca Locatelli Bento Gonçalves 0,017.01.2007 Pinotage Kunz São Valentim do Sul 0,017.01.2007 Moscato Nazareno Aristides Detoni Pinto Bandeira 0,017.01.2007 Moscato De Villa Monte Belo do Sul 0,017.01.2007 Moscato R2 Spadari Pinto Bandeira 0,017.01.2007 Trebiano Benvenuti Bento Gonçalves 0,017.01.2007 Chardonnay Locatelli Bento Gonçalves 2,017.01.2007 Pinotage Grapeggia Veranópolis 0,018.01.2007 Riesling Barbieri Monte Belo do Sul 2,023.01.2007 Chardonnay Conzatti Leopoldina 0,023.01.2007 Egiodola Benvenutti Bento Gonçalves 0,023.01.2007 Niágara Rosada Toneti Bento Gonçalves 0,023.01.2007 Egiodola Reginato Bento Gonçalves 0,023.01.2007 Pinotage Dalle Monte Belo do Sul 0,023.01.2007 Egiodola De Biasi Bento Gonçalves 0,0
continuação...
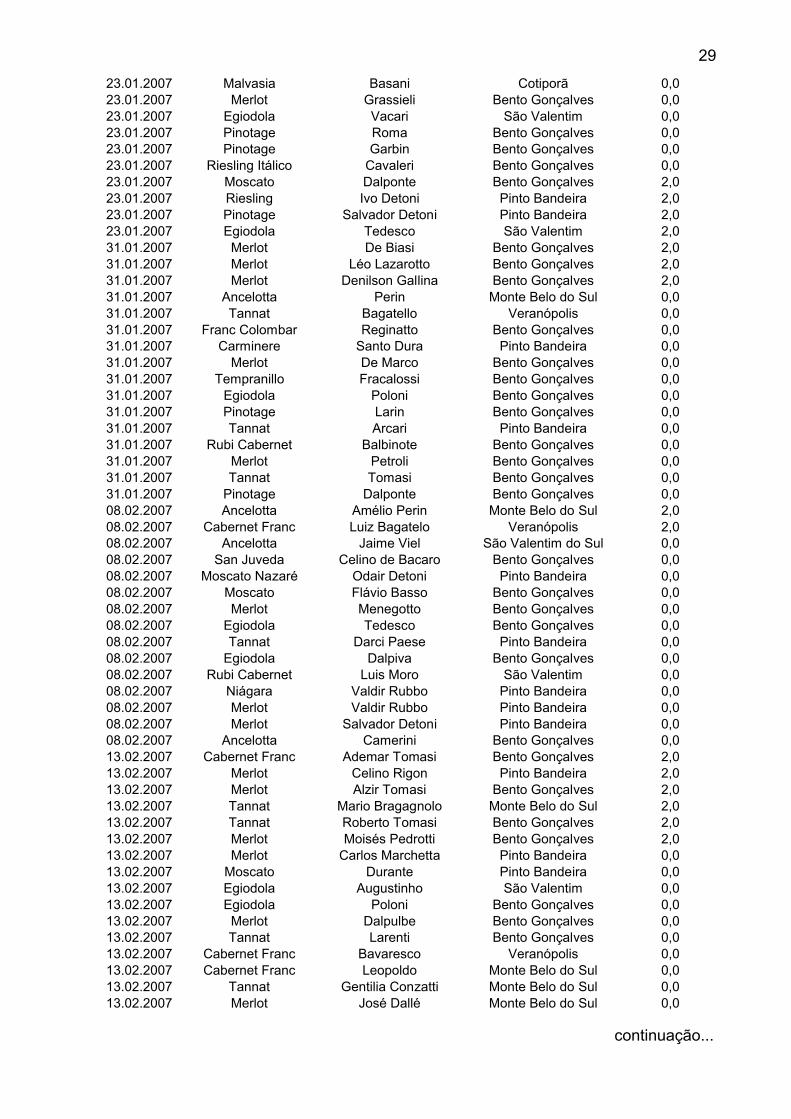
29
23.01.2007 Malvasia Basani Cotiporã 0,023.01.2007 Merlot Grassieli Bento Gonçalves 0,023.01.2007 Egiodola Vacari São Valentim 0,023.01.2007 Pinotage Roma Bento Gonçalves 0,023.01.2007 Pinotage Garbin Bento Gonçalves 0,023.01.2007 Riesling Itálico Cavaleri Bento Gonçalves 0,023.01.2007 Moscato Dalponte Bento Gonçalves 2,023.01.2007 Riesling Ivo Detoni Pinto Bandeira 2,023.01.2007 Pinotage Salvador Detoni Pinto Bandeira 2,023.01.2007 Egiodola Tedesco São Valentim 2,031.01.2007 Merlot De Biasi Bento Gonçalves 2,031.01.2007 Merlot Léo Lazarotto Bento Gonçalves 2,031.01.2007 Merlot Denilson Gallina Bento Gonçalves 2,031.01.2007 Ancelotta Perin Monte Belo do Sul 0,031.01.2007 Tannat Bagatello Veranópolis 0,031.01.2007 Franc Colombar Reginatto Bento Gonçalves 0,031.01.2007 Carminere Santo Dura Pinto Bandeira 0,031.01.2007 Merlot De Marco Bento Gonçalves 0,031.01.2007 Tempranillo Fracalossi Bento Gonçalves 0,031.01.2007 Egiodola Poloni Bento Gonçalves 0,031.01.2007 Pinotage Larin Bento Gonçalves 0,031.01.2007 Tannat Arcari Pinto Bandeira 0,031.01.2007 Rubi Cabernet Balbinote Bento Gonçalves 0,031.01.2007 Merlot Petroli Bento Gonçalves 0,031.01.2007 Tannat Tomasi Bento Gonçalves 0,031.01.2007 Pinotage Dalponte Bento Gonçalves 0,008.02.2007 Ancelotta Amélio Perin Monte Belo do Sul 2,008.02.2007 Cabernet Franc Luiz Bagatelo Veranópolis 2,008.02.2007 Ancelotta Jaime Viel São Valentim do Sul 0,008.02.2007 San Juveda Celino de Bacaro Bento Gonçalves 0,008.02.2007 Moscato Nazaré Odair Detoni Pinto Bandeira 0,008.02.2007 Moscato Flávio Basso Bento Gonçalves 0,008.02.2007 Merlot Menegotto Bento Gonçalves 0,008.02.2007 Egiodola Tedesco Bento Gonçalves 0,008.02.2007 Tannat Darci Paese Pinto Bandeira 0,008.02.2007 Egiodola Dalpiva Bento Gonçalves 0,008.02.2007 Rubi Cabernet Luis Moro São Valentim 0,008.02.2007 Niágara Valdir Rubbo Pinto Bandeira 0,008.02.2007 Merlot Valdir Rubbo Pinto Bandeira 0,008.02.2007 Merlot Salvador Detoni Pinto Bandeira 0,008.02.2007 Ancelotta Camerini Bento Gonçalves 0,013.02.2007 Cabernet Franc Ademar Tomasi Bento Gonçalves 2,013.02.2007 Merlot Celino Rigon Pinto Bandeira 2,013.02.2007 Merlot Alzir Tomasi Bento Gonçalves 2,013.02.2007 Tannat Mario Bragagnolo Monte Belo do Sul 2,013.02.2007 Tannat Roberto Tomasi Bento Gonçalves 2,013.02.2007 Merlot Moisés Pedrotti Bento Gonçalves 2,013.02.2007 Merlot Carlos Marchetta Pinto Bandeira 0,013.02.2007 Moscato Durante Pinto Bandeira 0,013.02.2007 Egiodola Augustinho São Valentim 0,013.02.2007 Egiodola Poloni Bento Gonçalves 0,013.02.2007 Merlot Dalpulbe Bento Gonçalves 0,013.02.2007 Tannat Larenti Bento Gonçalves 0,013.02.2007 Cabernet Franc Bavaresco Veranópolis 0,013.02.2007 Cabernet Franc Leopoldo Monte Belo do Sul 0,013.02.2007 Tannat Gentilia Conzatti Monte Belo do Sul 0,013.02.2007 Merlot José Dallé Monte Belo do Sul 0,0
continuação...

30
Na cultura da videira, a espécie normalmente não tem sido relatada em altas
populações (CABALEIRO; SEGURA, 1997a; CID et al., 2007). No entanto, P. citri é
muito próxima tanto biologicamente como morfologicamente a Planococcus ficus,
espécie invasora de grande importância como transmissora de vírus em vinhedos
dos Estados Unidos, África do Sul, Argentina e Uruguai a qual não foi encontrada
neste estudo (CHARLES et al., 2006) embora Foldi, Kozár (2006) tenham citado
para o município de Veranópolis, RS. As diferenças entre as duas espécies somente
são observadas com auxílio de especialistas o que tem levado ao desenvolvimento
de chaves moleculares para facilitar a identificação (DEMONTIS et al., 2007).
Quando foram coletadas cochonilhas presentes nas raízes, em duas
propriedades (Tab. 2) também foi encontrado a espécie P. citri, indicando que a
mesma pode também se desenvolver nesta estrutura vegetativa da videira. Numa
das propriedades a mesma foi encontrada juntamente com Dysmicoccus sp.
13.02.2007 Cabernet Franc Roberto Bono Pinto Bandeira 0,013.02.2007 Merlot Anzonet Veranópolis 0,013.02.2007 Rubi Cabernet Sérgio Tomasi Bento Gonçalves 0,013.02.2007 Tannat Marcolin Bento Gonçalves 0,013.02.2007 Rubi Cabernet Rossato Bento Gonçalves 0,002.03.2007 Cabernet
SauvignonCalisto Lerin Bento Gonçalves 2,0
02.03.2007 Cabernet Franc Lena Manfroi Bento Gonçalves 2,002.03.2007 Moscato Branco Vilmar Moroni Pinto Bandeira 0,002.03.2007 Cabernet
SauvignonEduardo Zanet Veranópolis 0,0
02.03.2007 Cabernet Sauvignon
Ulisses Tonho Pinto Bandeira 0,0
02.03.2007 Cabernet Sauvignon
Silo Perin Monte Belo do Sul 0,0
02.03.2007 Courdec 13 Marciano Fracalossi Bento Gonçalves 0,002.03.2007 Cabernet Franc Ciro Pavan Pinto Bandeira 0,002.03.2007 Moscato Branco Ildo Pozza Pinto Bandeira 0,002.03.2007 Cabernet Franc Valmor Dalpuber Bento Gonçalves 0,002.03.2007 Moscato Sganzerla Pinto Bandeira 0,002.03.2007 Cabernet
SauvignonOclides Arsego Bento Gonçalves 0,0
02.03.2007 Cabernet Sauvignon
Raimundo Zuchi Bento Gonçalves 0,0
09.03.2007 Courdec Mauri Moro Bento Gonçalves 2,009.03.2007 Courdec Argir Tomasi Bento Gonçalves 2,009.03.2007 Cabernet
SauvignonIvanir Rubbo Pinto Bandeira 2,0
09.03.2007 Cabernet Sauvignon
Fernando Cobalquini Bento Gonçalves 2,0
09.03.2007 Cabernet Sauvignon
Airton Frare Bento Gonçalves 0,0
09.03.2007 Moscato Híbrido Rudimar Moroni Pinto Bandeira 0,009.03.2007 Cabernet Franc Santo Duarte Pinto Bandeira 0,009.03.2007 Cabernet
SauvignonNatalino Paesi Pinto Bandeira 0,0
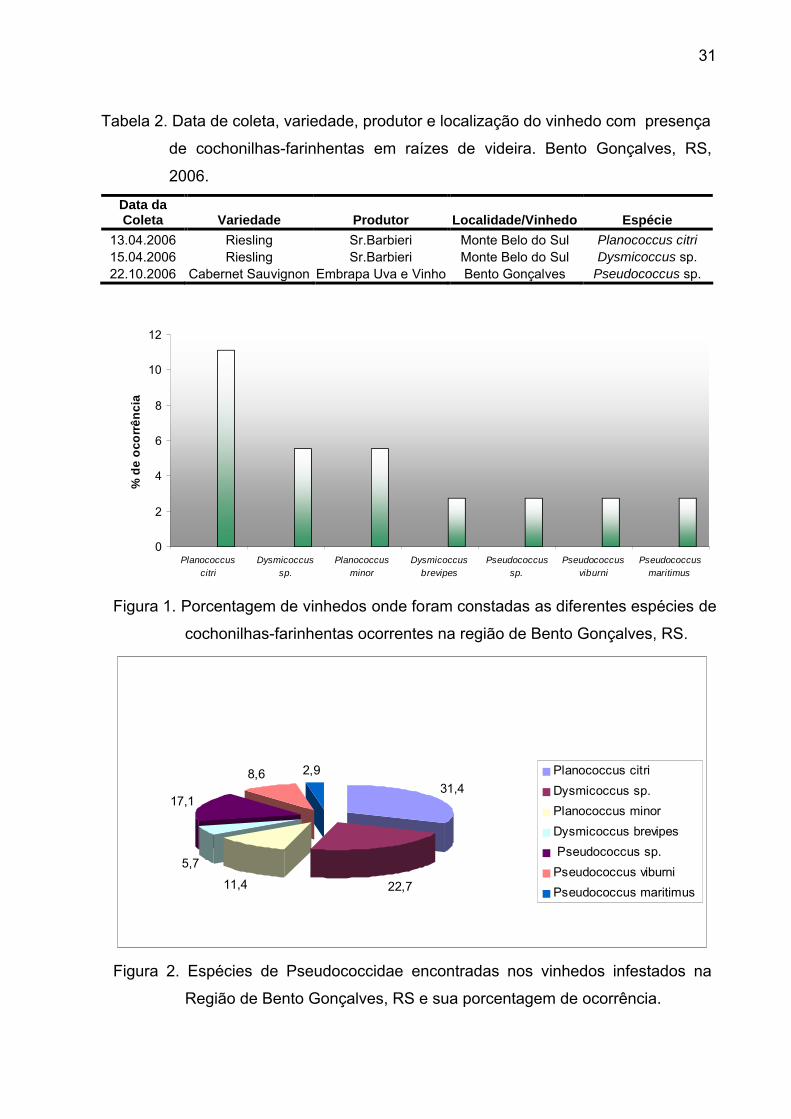
31
Tabela 2. Data de coleta, variedade, produtor e localização do vinhedo com presença
de cochonilhas-farinhentas em raízes de videira. Bento Gonçalves, RS,
2006.
Data da Coleta Variedade Produtor Localidade/Vinhedo Espécie
13.04.2006 Riesling Sr.Barbieri Monte Belo do Sul Planococcus citri 15.04.2006 Riesling Sr.Barbieri Monte Belo do Sul Dysmicoccus sp.22.10.2006 Cabernet Sauvignon Embrapa Uva e Vinho Bento Gonçalves Pseudococcus sp.
0
2
4
6
8
10
12
Planococcuscitri
Dysmicoccussp.
Planococcusminor
Dysmicoccusbrevipes
Pseudococcussp.
Pseudococcusviburni
Pseudococcusmaritimus
% d
e o
co
rrê
nc
ia
Figura 1. Porcentagem de vinhedos onde foram constadas as diferentes espécies de
cochonilhas-farinhentas ocorrentes na região de Bento Gonçalves, RS.
31,4
22,711,4
5,7
17,1
8,6 2,9 Planococcus citri
Dysmicoccus sp.
Planococcus minor
Dysmicoccus brevipes
Pseudococcus sp.
Pseudococcus viburni
Pseudococcus maritimus
Figura 2. Espécies de Pseudococcidae encontradas nos vinhedos infestados na
Região de Bento Gonçalves, RS e sua porcentagem de ocorrência.

32
A espécie P. minor também é muito similar a P. citri sendo facilmente
confundida (BASTOS et al., 2007). É uma praga importante causando danos em 250
culturas pertencentes a cerca de 80 famílias cultivadas na África, Austrália, Ásia e
regiões Neárticas, Neotropicais e Orientais (VENETTE; DAVIS, 2004). Entre as
culturas atacadas e de maior expressão econômica pode-se citar banana, batata,
beringela, cacau, café, citros, manga, milho, soja e uva (REDDY; SEETHARAMA
1997; LIT JR et al., 1998; REDDY et al., 1999; SANTA-CECÍLIA et al., 2002;
VENETTE; DAVIS, 2004). Em videira, a cochonilha já foi relatada em vários países
(WILLIAMS; WATSON, 1988; BEN-DOV, 1994; WILLINK et al., 1997). No Brasil, até
o momento, não existe relato desta espécie associada à cultura da videira.
Segundo Santa-Cecília (2007); Williams; Willink (1992) P. citri e P. minor podem
ocorrer na mesma planta hospedeira, além de causar danos similares atacando
tanto a parte aérea como as raízes. D. brevipes é considerada uma espécie polífaga
encontrada infestando raízes, frutos e folhas de diferentes culturas agrícolas com
destaque para o abacaxi (SANTA-CECÍLIA et al., 2007). Em videira, a espécie não
foi relatada no Brasil sendo encontrada com freqüência no Sul da Ásia (WILLIAMS,
2004). A espécie é transmissora do Pineapple mealybug wilt-associated virus
(PMWaV) na cultura do abacaxi (SETHER et al., 1998). Na videira, não existem
relatos que demonstram a transmissão de vírus pela cochonilha.
Pseudococcus viburni também é uma espécie cosmopolita e polífaga, com
capacidade de estabelecer-se sobre um amplo número de hospedeiros (BEN-DOV,
1994). Na cultura da videira, a mesma já foi relatada em zonas de clima temperado
do Hemisfério Norte, África do Sul, Austrália e Nova Zelândia, Itália, França,
Portugal, Espanha, como também desde o México até a Argentina incluindo o Brasil
(FOLDI; SORIA, 1989; GULLAN, 2000; GEIGER; DAANE, 2001; GODINHO;
FRANCO, 2001; GOLINO et al., 2002; GONZÁLEZ; VOLOSKY, 2004). Afonso
(2005) estudando esta espécie na cultura da videira, constatou que a mesma tem
capacidade de adquirir os vírus GLRaV-3 e GVB, no entanto, não foi comprovado a
transmissão.
Pseudococcus maritimus é uma espécie polífaga (MCKENZIE, 1967;
GONZÁLEZ, 1983; BEN-DOV, 1994). Na videira, P. maritimus pode ser localizada
em raízes, folhas e cachos (FLAHERTY et al., 1992; GONZÁLEZ, 2003). Segundo
González (2003), a espécie tem sido confundida com P. viburni, pois além de
estarem próximas taxonomicamente, muitas vezes coabitam a mesma região do

33
hospedeiro. A espécie é relatada como transmissora do vírus do enrolamento
GLRaV-3, GVA e GVB (GOLINO et al., 2002; CHARLES et al., 2006).
Em cachos de uva (Tab. 1) e em raízes (Tab. 2) foram coletados exemplares
do gênero Dysmicoccus os quais estão sendo consideradas como nova espécie
ainda não descrita (WILLINK, informação pessoal 2007). No caso de Pseudococcus
sp. coletada tanto em cachos (Tab. 1) como em raízes (Tab. 2) não foi possível
identificar os exemplares a nível de espécie devido a dificuldade para observar os
caracteres nas lâminas.
Em algumas coletas, foi registrada a presença de formigas junto às colônias
de cochonilha. Os exemplares foram identificados como pertencentes ao gênero
Linepithema pelo Dr. Odair Bueno (UNESP-Rio Claro), porém, devido a recente
revisão do gênero (WILD, 2007) não foi realizada a determinação específica. Em
outros países, a formiga argentina Linepithema humile (Mayr, 1868), é uma das
principais espécies associadas as cochonilhas farinhentas a qual se alimenta do
“honeydew” liberado pelo inseto. A presença de L. humile nos vinhedos é
considerado um indicativo da presença de populações de cochonilhas (DAANE et
al., 2006). Por outro lado, a implementação de um programa de manejo das
cochonilhas-farinhentas envolveria também o controle das formigas associadas, pois
estas além de auxiliarem na dispersão, também reduzirem o potencial de controle
biológico da praga (DAANE et al., 2007).
De maneira geral, foi registrado uma ampla gama de espécies de
Pseudococcidae associados aos vinhedos da região da Serra Gaúcha. A maioria
dos exemplares foram coletados nos frutos comprovando a multiplicação do inseto
na cultura. No entanto, devido a polifagia das espécies, os pseudococcídeos podem
estar também presentes em outros hospedeiros localizados dentro ou próximo ao
vinhedo incluindo plantas invasoras (BEN-DOV, 1994). Nem todas as espécies
encontradas já foram citadas como vetoras de vírus, porém, sabe-se que os
closterovírus (Família: Closteroviridae) a qual pertence o vírus do enrolamento da
folha da videira esta associado à Pseudococcidae como vetor (ROSCIGLIONE;
CASTELLANO, 1985; CABALEIRO, 1997a; FAJARDO et al., 2003; KUNIYUKI et al.,
2006; CID et al., 2007).
Neste caso, embora as cochonilhas não estejam causando danos diretos nas
uvas destinadas ao processamento, à presença das diferentes espécies em
aproximadamente 30% dos vinhedos é considerada significativa quando se trata de

34
vetores (DOUGLAS; KRÜGER, 2006). Os pseudococcídeos podem apresentar
várias gerações durante o ano além de possuírem uma ampla gama de hospedeiros
proporcionando a sua sobrevivência ao longo do ano (BEN DOV, 1994). No caso de
P. citri, também já foi comprovado que a transmissão do vírus pode ocorrer quando
somente um indivíduo se alimenta da planta contaminada (DOUGLAS; KRÜGER,
2008) além de haver uma relação cochonilha x vírus circulativa (CID et al., 2007).
Outro fator a ser considerado é que na região amostrada, as propriedades
são pequenas sendo freqüente o replantio dos vinhedos nas mesmas áreas. Nas
duas propriedades onde foram coletadas as cochonilhas nas raízes, a implantação
de novos vinhedos em áreas já contaminadas com vírus é uma prática que deve ser
evitada. Tal fato tem sido a origem de contaminação por vírus em novos plantios em
países como a África do Sul, devendo se adotar um manejo específico nestas
situações (PIETERSEN, 2008).
4 Conclusões
As espécies P. citri, P. minor, P. viburni, P. maritimus, D. brevipes,
Dysmicoccus sp. e Pseudococcus sp. estão associadas à cultura da videira na
região da Serra Gaúcha.
P. citri é a cochonilha mais abundante nos vinhedos da região da Serra
Gaúcha.

35
CAPÍTULO 2
BIOLOGIA E TABELA DE VIDA DE FERTILIDADE DE Planococcus citri (RISSO,
1813) (HEMIPTERA: PSEUDOCOCCIDAE) EM DIFERENTES ESTRUTURAS
VEGETATIVAS E CULTIVARES DE VIDEIRA
1 Introdução
A família Pseudococcidae inclui um grupo de insetos conhecidos como
cochonilhas-farinhentas por apresentarem o corpo recoberto por uma secreção
pulverulenta de cera branca (RIPA; RODRIGUEZ, 1999). Dentre os gêneros da
família Pseudococcidae, destaca-se o Planococcus, cujas espécies são
consideradas importantes pragas em vários agroecossistemas (CORREA, 2006). O
gênero Planococcus é originário do velho mundo e contém 43 espécies descritas
(BEN-DOV, 1994), sendo que seis ocorrem nas Américas Central e do Sul
(WILLIAMS; WILLINK, 1992) destacando-se P. citri conhecida como cochonilha-
branca, cochonilha-farinhenta ou cochonilha-dos-citros.
A espécie P. citri tem sido relatada como importante praga em diversas
plantas cultivadas, como por exemplo, abacaxizeiro, algodoeiro, bananeira, cafeeiro,
cana-de-açúcar, carambola, citros, coqueiro, figueira, goiabeira, mangueira,
macadâmia, soja e plantas ornamentais (BALACHOWSKY, 1935; SILVA et al., 1968;
DEAN et al., 1977; GULLAN, 2000; PARRELA, 2007). Na cultura da videira a
espécie também tem sido encontrada associada (MARTIN-MATEO, 1985;
MAROTTA, 1987; TEJERO, 1991; WILLIAMS; WILLINK, 1992; BEN-DOV, 1994;
WILLINK et al., 1997; UYGUN et al., 1998; WILLIAMS; MOGHADDAM, 2000;
GRAVENA, 2003; RAO et al., 2006; KISHINO et al., 2007, SANTA-CECÍLIA et al.,
2007).
No Brasil, P. citri é mencionada como uma das espécies de Pseudococcidae
associadas a cultura da videira juntamente com P. maritimus, P. longispinus, P.
viburni e P. ficus (SILVA et al., 1968; WOERNER, 1983; FOLDI; SORIA, 1989;

36
KUNIYUKI et al., 2005; FOLDI; KOZÁR, 2006; KISHINO et al., 2007). A espécie P.
citri é responsável pela transmissão do vírus do enrolamento da folha-da-videira
GLRaV-3, caneluras do tronco-da-videira (GVA) e intumescimento dos ramos (GVB),
importantes viroses que tem prejudicado de forma significativa a viticultura mundial
(ROSCIGLIONE; CASTELLANO, 1985; CABALEIRO; SEGURA, 1997a; CID et al.,
2007). A espécie pode se desenvolver em raízes, folhas e frutos da videira sendo
que neste último caso, ao se alimentar das bagas, sobre o “honeydew” excretado
desenvolve-se a fumagina que deprecia o produto comercial resultando em descarte
da fruta (GONZALEZ; VOLOSKY, 2004). Além disso, a presença do inseto nas
bagas impede a exportação devido às restrições quarentenárias impostas por alguns
países importadores (GONZALEZ et al., 1996; GONZALEZ; VOLOSKY, 2004).
No Brasil, existe uma diversidade de cultivares e porta-enxertos de videira os
quais, dependendo da combinação empregada, podem influenciar na maior ou
menor incidência da cochonilha nos vinhedos devido a resistência genética. Neste
trabalho, foi estudado a biologia e elaborando uma tabela de vida de fertilidade de P.
citri em diferentes estruturas vegetativas e cultivares de videira visando fornecer
informações para o manejo integrado da praga na cultura.
2 Material e Métodos
O trabalho foi conduzido no Laboratório de Entomologia da Embrapa Uva e
Vinho em câmara climatizada com temperatura de 25 2°C, 70 10% de umidade
relativa e 14 horas de fotofase. Fêmeas adultas de P. citri foram coletadas em raízes
e folhas da cultivar Riesling Itálico (Vitis vinifera) no mês de dezembro de 2006, no
município de Monte Belo do Sul (RS) (29º 09’ 46’’ S e 51º 37’ 54’’ O; 645 m de
altitude). Os insetos foram transportados para o laboratório e criados em abóbora
(Cucurbita maxima) da variedade Cabotchá acondicionadas em bandejas plásticas
cobertas com tecido “voile” conforme a metodologia descrita por Lepage (1942). A
cochonilha foi identificada pela Dra. Maria Cristina Granara de Willink do CONICET,
Superior Institute of Entomologia “Dr. A. Willink”, National University of Tucumán,
Argentina. Os exemplares estão disponíveis na coleção de referência no laboratório
de Entomologia da Embrapa Uva e Vinho (AZEVEDO-FILHO, 2007).

37
A biologia de P. citri foi estudada em folhas de videira Vitis vinifera das
cultivares Cabernet Sauvignon e Itália; Vitis labrusca cultivar Isabel, bagas da
cultivar Itália e raízes da cultivar Isabel e dos porta-enxertos 101-14 (Vitis riparia x
Vitis rupestris) e IAC-572 (Vitis caribaea x Vitis rupestris). As folhas foram colhidas
em vinhedos sem aplicação de inseticidas localizados na Embrapa Uva e Vinho,
sendo em seguida acondicionadas em placas de Petri contendo uma lâmina de 5
mm de ágar-água (15%). As raízes foram retiradas de mudas de videiras com um
ano de idade cultivadas em baldes plásticos (uma muda/balde) com capacidade de 5
litros. As bagas de uva foram adquiridas de produtores que não aplicam inseticidas
sendo individualizadas com o auxilio de tesoura, deixando-se o pedicelo. Logo após
a individualização, as bagas foram acondicionadas em recipientes de plástico (30
mL), sob uma camada de vermiculita esterilizada (0,5 cm), numerados e tampados.
Na tampa, foi aberto um orifício (1cm²) para a troca de gases com o ambiente e
posteriormente fechado com papel filtro e fita adesiva.
A cada cinco dias as folhas de videira e o ágar-água foram renovados,
substituindo-se as placas. A transferência da cochonilha foi feita cortando-se uma
pequena área foliar (3 mm) ao redor da mesma e, com o auxílio de uma pinça,
transferiu-se a área recortada para uma nova placa ocorrendo o deslocamento
natural do inseto, evitando-se danos nas peças bucais. Os discos foliares contendo
os casulos das ninfas macho foram transferidos para placas sem ágar, uma vez que,
nessa fase, o inseto não se alimenta (MALLESHAIAH et al., 2000).
Foram utilizadas 30 placas por repetição “inoculando-se” uma ninfa de
primeiro ínstar por placa no delineamento experimental inteiramente casualizado.
Após a “inoculação” das ninfas, as placas e os recipientes plásticos foram colocados
em câmara climatizada do tipo BOD (FANEM modelo 347-CDG).
A avaliação dos ínstares das fêmeas e machos foi realizada com base na
presença das exúvias liberadas pelas ninfas, analisando-as a cada dois dias, com
auxilio de uma lupa binocular (com aumento de 10 vezes) (CORREA et al., 2006).
Como não há diferença sexual evidente no início do desenvolvimento ninfal, as
repetições foram constituídas por indivíduos com sexo não conhecido, definindo-se o
sexo a partir do segundo instar quando os machos formam casulos para completar o
seu desenvolvimento (LOAEZA, 1958).
A fecundidade foi obtida através da contagem do número de ovos por saco
ovígero de 20 fêmeas e a viabilidade individualizando-se 20 ovos de cada fêmea em

38
placas de Petri de 9,5 cm de diâmetro, contendo papel filtro umedecido no fundo,
registrando-se o número de ninfas eclodidas.
Os seguintes parâmetros foram avaliados por sexo: duração e viabilidade dos
estágios de ovo e ninfa; fecundidade (em fêmeas) e longevidade. Com os dados
obtidos foi elaborada a tabela de vida de fertilidade para as diferentes cultivares
(SILVEIRA NETO et al., 1976) estimando-se a duração média de uma geração (T), a
taxa líquida de reprodução (Ro), a razão infinitesimal de aumento (Rm) e a razão
finita de aumento (). Os dados foram submetidos à análise de variância, sendo as
médias comparadas pelo teste de Tukey (p0,05) através do programa Genes
(CRUZ, 2001).
3 Resultados e Discussão
3.1 Duração e Viabilidade
Em raízes da cultivar IAC-572, as cochonilhas não se desenvolveram
indicando a inadequação deste porta enxerto ao desenvolvimento da espécie (Tab.
3). Nachtigal (2003) cita alguns porta-enxertos para a cultura da videira que são
resistentes a alguns insetos de solo dentre eles o IAC-572 que é resistente a filoxera
(Daktulosphaira vitifoliae) demonstrando a existência de componentes nas raízes
que conferem resistência. Este fato, pode auxiliar na explicação da imunidade
destas raízes ao dano do inseto, visto que o mesmo pode passar parte do ano nas
em raízes de videiras.
O desenvolvimento ninfal dos machos de P. citri, independente da cultivar e
da estrutura vegetativa em que o inseto se alimentou, foi de quatro ínstares, sendo
que no final do segundo, foi construído um casulo de filamentos cerosos (pupa), no
interior do qual as ninfas permaneceram durante o terceiro e quarto ínstares,
seguindo-se a emergência dos adultos (Tab. 3). As fêmeas apresentaram três
instares livres (sem presença de casulos).
A duração média do período embrionário foi semelhante em todas as
cultivares sendo em média de 4,00,14 dias com viabilidade de 79,9% (Tab. 4).
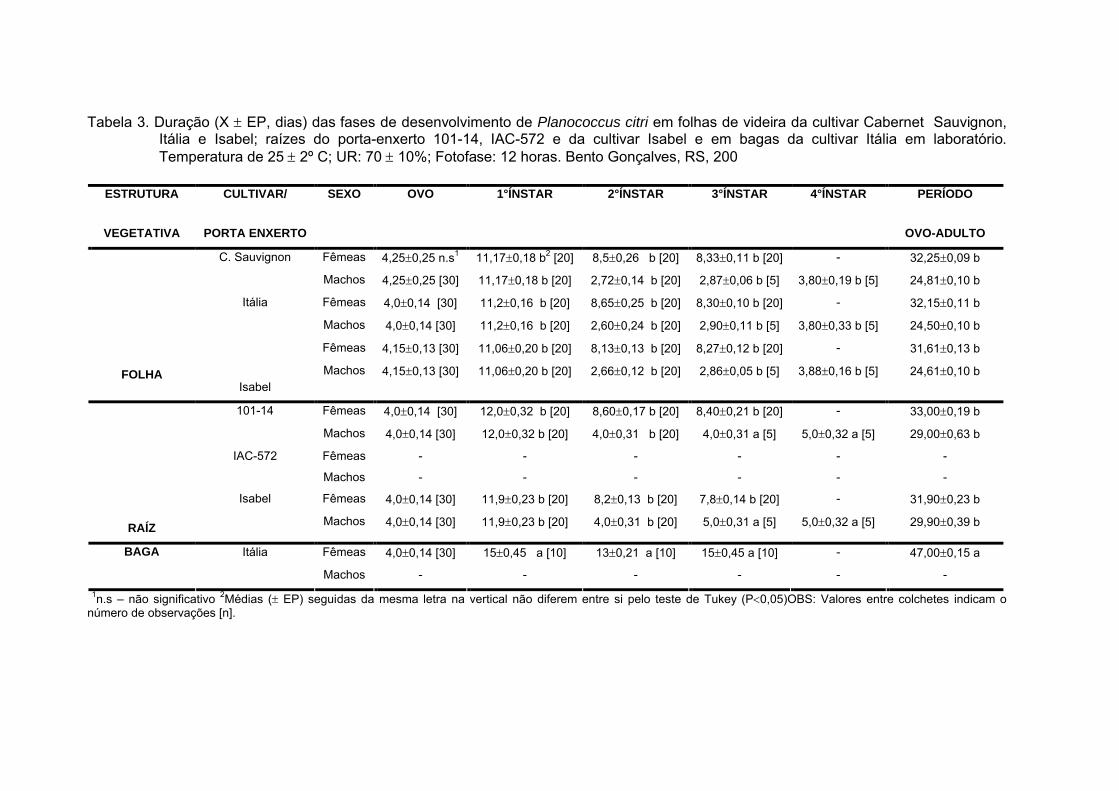
Tabela 3. Duração (X EP, dias) das fases de desenvolvimento de Planococcus citri em folhas de videira da cultivar Cabernet Sauvignon, Itália e Isabel; raízes do porta-enxerto 101-14, IAC-572 e da cultivar Isabel e em bagas da cultivar Itália em laboratório. Temperatura de 25 2º C; UR: 70 10%; Fotofase: 12 horas. Bento Gonçalves, RS, 200
1n.s – não significativo 2Médias ( EP) seguidas da mesma letra na vertical não diferem entre si pelo teste de Tukey (P0,05)OBS: Valores entre colchetes indicam o número de observações [n].
ESTRUTURA
VEGETATIVA
CULTIVAR/
PORTA ENXERTO
SEXO OVO 1°ÍNSTAR 2°ÍNSTAR 3°ÍNSTAR 4°ÍNSTAR PERÍODO
OVO-ADULTO
Fêmeas 4,250,25 n.s1 11,170,18 b2 [20] 8,50,26 b [20] 8,330,11 b [20] - 32,250,09 bC. Sauvignon
Machos 4,250,25 [30] 11,170,18 b [20] 2,720,14 b [20] 2,870,06 b [5] 3,800,19 b [5] 24,810,10 b
Fêmeas 4,00,14 [30] 11,20,16 b [20] 8,650,25 b [20] 8,300,10 b [20] - 32,150,11 bItália
Machos 4,00,14 [30] 11,20,16 b [20] 2,600,24 b [20] 2,900,11 b [5] 3,800,33 b [5] 24,500,10 b
Fêmeas 4,150,13 [30] 11,060,20 b [20] 8,130,13 b [20] 8,270,12 b [20] - 31,610,13 b
FOLHAIsabel
Machos 4,150,13 [30] 11,060,20 b [20] 2,660,12 b [20] 2,860,05 b [5] 3,880,16 b [5] 24,610,10 b
Fêmeas 4,00,14 [30] 12,00,32 b [20] 8,600,17 b [20] 8,400,21 b [20] - 33,000,19 b101-14
Machos 4,00,14 [30] 12,00,32 b [20] 4,00,31 b [20] 4,00,31 a [5] 5,00,32 a [5] 29,000,63 b
Fêmeas - - - - - -IAC-572
Machos - - - - - -
Fêmeas 4,00,14 [30] 11,90,23 b [20] 8,20,13 b [20] 7,80,14 b [20] - 31,900,23 b
RAÍZ
Isabel
Machos 4,00,14 [30] 11,90,23 b [20] 4,00,31 b [20] 5,00,31 a [5] 5,00,32 a [5] 29,900,39 b
Itália Fêmeas 4,00,14 [30] 150,45 a [10] 130,21 a [10] 150,45 a [10] - 47,000,15 aBAGA
Machos - - - - - -
40
Tabela 4. Viabilidade (X EP) das fases de desenvolvimento de Planococcus citri sobre folhas de videira cultivar Cabernet Sauvignon, Itália,
Isabel; raízes do porta-enxerto 101-14, IAC-572 e cultivar Isabel; bagas da cultivar Itália em condições de laboratório. Temperatura de 25
2º C; UR: 70 10%; Fotofase: 12 horas. Bento Gonçalves, RS, 2007.
ESTRUTURA
VEGETATIVA
CULTIVAR/
PORTA ENXERTO
SEXO OVO 1°ÍNSTAR 2°ÍNSTAR 3°ÍNSTAR 4°ÍNSTAR PERÍODO OVO-
ADULTO
Fêmeas 92,000,80 a1 [30] 85,006,78 a [20] 85,006,78 a [20] 85,006,78 b [20] - 56,49 aC. Sauvignon
Machos 92,008,77 a [30] 74,008,77 a [20] 74,008,77 a [20] 74,008,77 a [5] 74,008,77 b [5] 27,59 b
Fêmeas 80,005,00 b [30] 80,009,17 a [20] 98,008,0 a [20] 100,000,0 a [20] - 62,72 aItália
Machos 80,002,0 b [30] 80,002,00 a [20] 80,002,0 a [20] 80,002,0 a [5] 80,002,0 a [5] 32,72 b
Fêmeas 84,662,4 a [30] 83,3311,07a[20] 83,3311,07a[20] 83,3311,07b[20] - 49,00 a
FOLHAIsabel
Machos 84,662,41 a [30] 75,009,00 a [20] 75,009,00 a [20] 75,009,00 a [5] 75,009,00 b [5] 35,71 b
Fêmeas 80,009,17 b[30] 80,009,17 a [20] 98,008,0 a [20] 100,000,0 a [20] - 62,72 a
101-14
Machos 80,002,0 b [30] 80,002,0 a [20] 80,002,0 a [20] 80,002,0 a [5] 80,002,0 a [5] 32,72 b
Fêmeas - - - - - -IAC-572
Machos - - - - - -
Fêmeas 65,552,71 c [30] 50,0015,07c[20] 50,0015,07c[20] 50,0015,07c[20] - 8,19 cRAÍZ
Isabel
Machos 65,552,71 c [30] 50,0015,07b[20] 50,0015,07b[20] 50,0015,07 b [5] 50,0015,07 c [5] 4,06 c
Itália Fêmeas 75,007,8 b [30] 70,0015,2 b [10] 75,0013,43b[10] 74,0013,26b[10] - 29,13 bBAGA
Machos - - - - - -
1Médias ( EP) seguidas da mesma letra na vertical não diferem entre si pelo teste de Tukey (P0,05). OBS: Valores entre colchetes indicam o número de observações [n].

A duração do primeiro ínstar, tanto das fêmeas como dos machos, foram
semelhantes nas folhas e raízes, não havendo diferenças significativas entre as
mesmas, sendo em média 11,470,18 e 11,490,28 dias com viabilidade de 74,7% e
71,8% para fêmeas e machos, respectivamente (Tab. 3). Em bagas, foi observado
um prolongamento significativo desta fase quando comparado com as demais
estruturas, sendo que a duração foi de 150,45 dias com viabilidade de 70% (Tab.
3). Este instar apresentou maior duração, tanto para machos como para fêmeas,
assemelhando-se aos resultados encontrados por MALLESHAIAH et al. (2000), que
encontraram 10,91,93 dias para machos e 11,21,25 dias para fêmeas de P. citri
em frutos de abóbora na temperatura de 262°C. CORREA et al. (2005) também
verificaram em folhas de Citrus cinensis maior duração deste ínstar sendo 10,43,1
e 11,62,6 dias para machos e fêmeas, respectivamente, na temperaturas de
251°C.
O segundo instar dos machos e fêmeas alimentados em folhas foram
semelhantes, sendo de 8,40,21 dias com viabilidade de 81,5% para fêmeas e
2,800,16 dias com viabilidade de 71,8% para machos, não havendo diferença entre
os sexos. Os valores de duração deste ínstar (Tab. 3) divergiram daqueles
constatados por MALLESHAIAH et al. (2000), que foram de 2,50,60 e 8,60,75 dias
respectivamente, para machos e fêmeas (na temperatura de 262°C). Da mesma
maneira, CORREA et al. (2005), encontraram para o segundo instar a duração de
10,14,7 e 6,42,4 dias respectivamente para machos e fêmeas, nas temperaturas
de 251°C diferindo dos resultados encontrados neste trabalho. Quando
alimentados com raízes, o desenvolvimento dos machos foi mais prolongado, em
5,00,31 dias em comparação com os demais substratos (Tab. 3).
Para o terceiro instar, a duração média das ninfas fêmeas foi de 8,220,11
dias com viabilidade de 82,0%, não havendo diferenças entre as estruturas
vegetativas estudadas. Estes resultados foram semelhantes aos encontrados por
MALLESHAIAH et al. (2000) de 8,300,98 dias, para fêmeas, porém, foram
inferiores aos encontrados CORREA et al. (2005), que observaram 6,31,9 dias para
as fêmeas.
As diferenças ocorridas na duração de cada instar podem ser explicadas, por
diferenças de temperatura, hospedeiro e estruturas vegetativas das plantas
40

42
avaliadas neste experimento e os utilizados por MALLESHAIAH et al. (2000) em
abóbora e CORREA et al. (2005) em citros.
O período em que os machos passaram no interior do casulo abrangeu o
terceiro e quarto instares, correspondendo a um tempo de 6,700,23 dias e
viabilidade de 76,3% quando criados em folhas e de 9,50,32 dias e viabilidade de
65,0% em raízes. Estes resultados concordam com os encontrados por
MALLESHAIAH et al. (2000) no qual este período foi de 6,60,73 dias sem informar
a viabilidade da fase e discordam de CORREA et al. (2005) que obteram 10,72,7
dias com viabilidade de 100%.
A duração média do ciclo total (ovo-adulto) dos machos foi de 24,630,10
dias, e viabilidade do período de 32,0%, enquanto que as fêmeas duraram em média
320,11 dias com viabilidade de 56,0% quando alimentadas com folhas de videira
nas diferentes cultivares. Em raízes, o ciclo biológico da fêmea foi em média
32,450,19 dias enquanto a dos machos foi de 29,50,51 dias. A menor viabilidade
do ciclo ninfal foi verificada em raízes de Isabel, sendo de 8,1% para fêmeas e 4,0%
para machos. Foi encontrada a menor viabilidade do ciclo total em folhas e raízes de
Isabel. Estes resultados são semelhantes aos encontrados por MALLESHAIAH et al.
(2000) que verificaram uma duração do ciclo total das fêmeas de 31,450,96 dias e
dos machos de 23,400,97 dias, no entanto estes autores não discutem a
sobrevivência dos instares no experimento. Foi observado um prolongamento do
ciclo total em 15 dias quando a espécie se desenvolveu em bagas de uva da cultivar
Itália resultando num ciclo total de 47,000,15 dias com viabilidade de 29,1%
indicando que bagas de videira da cultivar Itália não são bons substratos para o
desenvolvimento da cochonilha (Tab. 3 e 4). Entretanto, em condições de campo, os
pseudococcídeos têm sido encontrados de maneira freqüente associados à
presença da fumagina (KISHINO et al., 2007). Em hipótese, a presença de P. citri
nas bagas é resultado do desenvolvimento das primeiras gerações da espécie em
folhas, migrando para os frutos próximo a colheita (ETCHEBARNE et al., 2003).
SALAZAR (1996), em estudos realizados em vinhedos da Espanha verificou
que sistemas de condução latada são ideais para o desenvolvimento do inseto, pois
as mesmas ficam protegidas da luminosidade, chuva e vento. O autor relata ainda
que a incidência da cochonilha é maior em videiras vigorosas, de maturação tardia e
racimos mais compactos fato corroborado por GONZÁLEZ (1983) no Chile. Este fato

43
pode auxiliar na confirmação das observações realizadas neste experimento onde
as cultivares Cabernet Sauvignon e Itália consideradas tardias e com rácimos
compactados foram mais adequadas ao desenvolvimento do inseto.
3.2 Longevidade e Fecundidade
A longevidade das fêmeas diferiu em função do substrato e cultivares na qual
a cochonilha foi alimentada nas fases imaturas (Tab. 5). Em folhas, a longevidade foi
de 56,416,4, 50,413,5 e 52,313,5 dias nas cultivares Cabernet Sauvignon, Itália e
Isabel, respectivamente (Tab. 5). Em raízes do porta-enxerto 101-14 e Isabel e em
bagas da cultivar Itália, a longevidade foi de 52,313,5 dias (Tab. 5). Estes valores
foram superiores àqueles obtidos por MALLESHAIAH et al. (2000), que constataram
uma duração de 45,1 dias para P. citri criada em frutos de abóbora sendo
semelhantes aos 56,0 dias encontrados por CORREA et al. (2005) em citrus.
Para os machos, a longevidade foi de 2,0 dias em todos os substratos
avaliados equivalentes ao observado por CORREA et al. (2005) em citros e
inferiores aos 4,4 dias verificados por MALLESHAIAH et al. (2000) em frutos de
abóbora, atribuindo-se as diferenças ao substrato empregado nos diferentes
trabalhos.
A fecundidade das fêmeas variou conforme a estrutura vegetativa em que os
insetos foram criados (Tab. 5). Em folhas de videira, a fecundidade foi de
67,2710,70; 53,331,68 e 66,094,68 ovos por fêmea nas cultivares Cabernet
Sauvignon, Itália e Isabel respectivamente. Em raízes, foi obtido 30,46,12 e
70,0010,5 ovos por fêmea no porta-enxerto 101-14 e Isabel respectivamente. Estes
resultados foram inferiores aos encontrados por MALLESHAIAH et al. (2000), que foi
de 232,5 ovos por fêmea, por MAYNE (1932) com 350-650 ovos, SOUTHERLAND
(1972) com 172 ovos, ENTWISTLE, (1972) com 150 a 300 e BODENTHEIMER
(1951) com 176 a 400 ovos por fêmeas em frutos de abóboras, café, cacau e citrus
respectivamente. No porta-enxerto IAC-572 e nas bagas de uva da cultivar Itália, não
foram obtidas posturas.
A espécie P. citri conseguiu se multiplicar em folhas de cultivares de Vitis
vinifera (Cabernet Sauvignon e Itália) e Vitis labrusca (Isabel) e em raízes dos porta
enxertos 101-14 e Isabel. O porta enxerto IAC-572 mostrou ser inadequado ao
desenvolvimento do inseto enquanto que em bagas da cultivar Itália, embora tenha

44
ocorrido o desenvolvimento das fases jovens, não originou descendentes, indicando
serem hospedeiros não adequados ao desenvolvimento da espécie.
Tabela 5. Longevidade (X EP) em dias de Planococcus citri e fecundidade quando
as fêmeas foram criadas sobre folhas de videira da cultivar Cabernet
Sauvignon, Itália, Isabel; raízes do porta enxerto 101-14, IAC-572 e
cultivar Isabel; bagas da cultivar Itália em condições de laboratório.
Temperatura de 25 2º C; UR: 70 10%; Fotofase: 12 horas. Bento
Gonçalves, RS, 2007.
ESTRUTURA
VEGETATIVA
CULTIVAR/
PORTA ENXERTO
LONGEVIDADE FECUNDIDADE
FÊMEA MACHO
C. Sauvignon 56,416,4 a1 [20] 2,00,5n.s2 [5] 67,2710,70 a [15]
Itália 50,413,5 b [20] 2,00,5 [5] 53,331,68 b [15]FOLHA Isabel 52,313,5 a [20] 2,0 0,5 [5] 66,094,68 a [10]
101-14 52,313,5 a [20] 2,00,5 [5] 30,46,12 c [15]
IAC-572 - - -
RAÍZ Isabel 52,313,5 a [20] 2,00,5 [5] 70,0010,51 a [10]
BAGA Itália 52,313,5 a [10] 2,00,5 [5] -
1Médias ( EP) seguidas da mesma letra na vertical não diferem entre si pelo teste de Tukey (P0,05).OBS: Valores entre colchetes indicam o número de observações [n].2n.s – não significativo.
3.3 Tabela de Vida de Fertilidade
Em raízes do porta-enxerto IAC-572 e em bagas de uva da cultivar Itália, não
foi calculado a tabela de vida de fertilidade devido a ausência de descendentes. Nos
demais substratos, a duração média de uma geração (T) de P. citri variou em função
da estrutura vegetativa e cultivar (Tab. 6). A menor duração de uma geração foi
verificada em folhas da cultivar Isabel (40,50 dias) sendo que na Cabernet
Sauvignon e Itália, o valor foi próximo de 41,00 dias em ambas as cultivares (Tab. 6).
Em raízes do porta-enxerto 101-14, a duração de uma geração foi de 42,00 dias,
enquanto que em raízes da cultivar Isabel foi de 41,00 dias.
A taxa líquida de reprodução Ro (número de vezes que a população aumenta
a cada geração), também apresentou diferença em função da estrutura vegetativa

45
avaliada ou da cultivar (Tab. 6). Em folhas e raízes da cultivar Isabel foi encontrada a
menor capacidade de reprodução sendo de 259,07 e 45,36 respectivamente.
Tabela 6. Duração média de uma geração (T), taxa líquida de reprodução (Ro), razão
infinitesimal de aumento (Rm) e razão finita de aumento () de Planococcus
citri sobre folhas de videira das cultivares Cabernet Sauvignon, Itália e Isabel;
raízes do porta enxerto 101-14, IAC-572 e cultivar Isabel; bagas da cultivar
Itália em condições de laboratório. Temperatura de 25 2º C; UR: 70 10%;
Fotofase: 12 horas. Bento Gonçalves, RS, 2007.
Em folhas de Cabernet Sauvignon, Itália e raízes de 101-14 os valores foram
de 301,37, 267,59 e 152,54 respectivamente.
A razão infinitesimal de aumento populacional (Rm) das estruturas
vegetativas nas diferentes cultivares de videira foram todas equivalentes sendo de
0,125 (Tab. 6).
A razão finita de aumento () que representa o número de fêmeas
adicionadas à população por fêmeas numa unidade de tempo foi semelhante em
todas as estruturas avaliadas apresentando maior valor (1,149) em folhas de videira
da cultivar Cabernet Sauvignon e menor (1,097) em raízes da cultivar Isabel (Tab.
6).
Através da utilização da tabela de vida de fertilidade, que é um critério de
avaliação do desempenho de uma dieta e ou hospedeiro na criação de insetos
ESTRUTURA
VEGETATIVA
CULTIVAR/
PORTA ENXERTO
T (dias) Ro Rm
Cabernet Sauvignon 41,00 301,37 0,1392 1,149
Itália 41,00 267,59 0,1363 1,146
FOLHAIsabel 40,50 259,07 0,1372 1,147
101-14 42,00 152,54 0,1196 1,1270
IAC-572 - - - -
RAÍZIsabel 41,00 45,36 0,093 1,097
BAGA Itália - - - -

46
(PARRA, 1996), verificou-se que diferentes estruturas vegetativas e cultivares
afetaram a duração em dias de uma geração, a taxa líquida de reprodução, a razão
infinitesimal de aumento populacional e a razão finita de aumento de P. citri. Desta
maneira, visando reduzir a infestação de P. citri nos vinhedos, recomenda-se o
emprego do porta-enxertos IAC-572 e/ou a cultivar Isabel que pode ser cultivada
tanto com pé-franco como utilizada como porta-enxerto. No caso de variedades
copa, pode se utilizar a cultivar Isabel e Cabernet Sauvignon.
4 Conclusões
A cochonilha P. citri completa o ciclo biológico em folhas de videira V. vinifera
das cultivares Cabernet Sauvignon e Itália; raízes porta enxerto 101-14 (V. riparia x
V. rupestris) e folhas e raízes de V. labrusca Isabel.
Bagas de uva da cultivar Itália permitem o desenvolvimento das fases ninfais
de P. citri mas os adultos são inférteis.
As raízes do porta enxerto IAC-572 não permitem o desenvolvimento de P.
citri.

47
CAPÍTULO 3
EXIGÊNCIAS TÉRMICAS E ESTIMATIVA DO NÚMERO ANUAL DE GERAÇÕES
DE Planococcus citri (RISSO, 1813) (HEMIPTERA: PSEUDOCOCCIDAE) NA
CULTURA DA VIDEIRA
1 Introdução
Na cultura da videira, as cochonilhas da família Pseudococcidae tem sido
relatadas causando danos diretos, principalmente em uvas de mesa da cultivar Itália
(MORANDI FILHO et al., 2007; KISHINO et al., 2007). Entretanto, a importância
desta família na cultura tem sido ampliada nos últimos anos, devido a capacidade
das espécies transmitirem vírus com destaque para o vírus do enrolamento GLRaV 3
(Grapevine leafroll associated virus 3), GVA (Grapevine trichovirus A) e GVB
(Grapevine trichovirus B) (ROSCIGLIONE; CASTELLANO, 1985; CABALEIRO;
SEGURA, 1997a, PETERSEN; CHARLES, 1997, SFORZA et al., 2003, GOLINO et
al., 2002, BORBÓN et al., 2004; KUNIYUKI et al., 2005, KUNIYUKI et al., 2006, CID
et al., 2007).
Na região da Serra Gaúcha, principal pólo produtor de uva para
processamento do Brasil (PROTAS et al., 2002) P. citri é mencionada como uma das
espécies de Pseudococcidade associadas a cultura da videira (SILVA et al., 1968;
WOERNER, 1983; FOLDI; SORIA, 1989; MORANDI FILHO et al., 2007). No entanto,
poucas informações estão disponíveis sobre a biologia da cochonilha na cultura e
principalmente em relação aos efeitos de fatores abióticos e bióticos no
desenvolvimento da espécie. Dentre os fatores abióticos que se destacam pelos
seus efeitos na biologia, ecologia e dinâmica populacional das espécies merece
destaque a temperatura (HONĔK, 1996, HADDAD et al., 1999, TRUDGILL et al.,
2005).
A influência da temperatura sobre a biologia de Pseudococcidae tem sido
pouco investigada. No caso de P. citri, existem informações quando a cochonilha foi

48
alimentada em batata, citros, roseira e cafeeiro (ISLAM et al., 1995, ARAI, 1996,
MARTINEZ-FERRER et al., 2003, LAFLIN; PARRELLA 2004, CORREA et al., 2008).
Na videira, não existem dados disponíveis sendo as informações geradas somente
para P. ficus (WALTON; PRINGLE 2005), espécie próxima a P. citri que tem ocorrido
com maior freqüência nos vinhedos de outros países (DEMONTIS et al., 2007).
O conhecimento das necessidades e dos limites térmicos para o
desenvolvimento de uma espécie é fundamental para definir o potencial de
adaptação e crescimento de uma praga nas diferentes regiões produtoras. No caso
da videira, este fator tem sido ainda mais importante visto a ampliação do cultivo
para diferentes regiões do Brasil incluindo as de clima tropical (PROTAS et al.,
2002).
Neste trabalho foi estudado o efeito da temperatura sobre a biologia de P. citri
em folhas de videira estimando-se o número de gerações anuais que o inseto pode
completar nas principais regiões produtoras de uva do Brasil.
2 Material e Métodos
O trabalho foi conduzido no Laboratório de Entomologia da Embrapa Uva e
Vinho em Bento Gonçalves, RS. A biologia de P. citri foi estudada em folhas de
videira Vitis vinifera da cultivar Itália, retiradas em vinhedos sem aplicação de
inseticidas. No laboratório, as folhas foram acondicionadas em placas de Petri (9,5
cm de diâmetro x 1,7 cm altura) contendo uma lâmina de 5 mm de ágar-água (15%)
e colocadas em câmaras climatizadas nas temperaturas de 16, 19, 22, 25, 28 e 31
ºC, com umidade relativa de 70 10% e 12 horas de fotofase. Os insetos utilizados
nos experimentos foram provenientes da criação de manutenção em abóboras
(Cucurbita maxima) da variedade Cabotchá conforme a metodologia descrita por
Lepage (1942). A cochonilha foi identificada pela Dra. Maria Cristina Granara de
Willink do CONICET, Instituto Superior de Entomologia “Dr. A. Willink”, Universidade
Nacional de Tucumán, Argentina. Os exemplares testemunha estão depositados na
coleção entomológica de referência da Embrapa Uva e Vinho (AZEVEDO-FILHO et
al., 2007).
Foram utilizadas 30 placas por repetição em cada temperatura inoculando-se
uma ninfa de primeiro ínstar com um dia de idade por placa no delineamento
experimental inteiramente casualizado. Para a verificação da temperatura base (Tb)

49
da fase de ovo, os mesmos foram provenientes das fêmeas existentes na criação de
manutenção em abóboras, utilizando-se 30 ovos em cada temperatura.
A avaliação do número de ínstares da cochonilha foi realizado com base na
presença das exúvias liberadas pelas ninfas, analisando-as diariamente, com auxilio
de uma lupa binocular (10 x) (CORREA, 2006).
As exigências térmicas das fases imaturas de P. citri foram estimadas pelo
método da hipérbole (HADDAD et al., 1999), calculando-se a temperatura base (Tb)
e a constante térmica (K) com o programa SAS (SAS INSTITUTE, 1998). A
significância das diferenças entre os períodos de duração foi comparada através do
teste de Tukey (p≤0,05) utilizando-se o programa Sisvar (FURTADO, 2007).
O valor de K foi obtido através da fórmula K= y (t - a), sendo K= constante
térmica em graus dia; y= tempo requerido, em dias, para completar o
desenvolvimento; t= temperatura ambiente em °C; a=limiar térmico inferior em °C
(SILVEIRA NETO et al., 1976).
O número provável de gerações (NG) anuais de P. citri foi calculado
utilizando-se as normais térmicas para Bento Gonçalves (RS), Bagé (RS), Londrina
(PR) Petrolina (PE), Jales (SP) e São Lourenço (MG) considerados importantes
pólos produtores de uva do Brasil (PROTAS et al., 2002) com base nas exigências
térmicas (CIVIDANES, 2000), utilizando-se as normais meteorológicas desses
municípios. Embora Londrina (PR) e São Lourenço (MG) não sejam tradicionais
produtores de uva, as normais térmicas destes municípios foram utilizadas, por
serem estes os mais próximos, e que apresentam altitude semelhante a de Marialva
(PR) e Andradas (MG), respectivamente.
As normais meteorológicas dos municípios foram obtidos no Departamento
Nacional de Meteorologia (BRASIL, 1992) e no Laboratório de Meteorologia da
Embrapa Uva e Vinho. O cálculo do NG foi efetuado por meio da equação: NG =
{T(Tm – Tb)/K}, onde: K = constante térmica, Tm = a temperatura média para cada
localidade estudada, Tb = temperatura base e T = o tempo em dias.
3 Resultados e Discussão
A duração das fases do ciclo biológico de P. citri apresentou relação inversa à
elevação da temperatura exceto quando os insetos foram mantidos a 31ºC, onde as
ninfas somente se desenvolveram até o segundo instar (Tab. 7). No período
embrionário, a maior duração foi registrada na temperatura de 16ºC (14,4 ± 0,10

50
dias) e a menor a 28ºC (3,9 ± 0,08 dias). Com exceção das temperaturas de 25 e
28ºC, os valores correspondentes a duração desta fase diferiram entre si (p≤0,05)
(Tab. 7). A viabilidade da fase de ovo não variou entre as temperaturas sendo em
todas de 100% (p≤0,05) (Tab. 8).
A influência da temperatura sobre a velocidade do desenvolvimento
embrionário de P. citri também foi relatada por Correa et al. (2007) para ovos
oriundos da criação de manutenção em laboratório em abóboras (Cucurbita maxima
Duchesne) em temperaturas que variaram de 15 a 35 ºC. Os valores médios da
duração dessa fase registrados pelos autores, nas temperaturas de 15 ºC (14,73 ±
0,26), 20 ºC (7,06 ± 0,04), 25 ºC (4,29 ± 0,13) e 30ºC (3,24 ± 0,09) são próximos aos
obtidos neste estudo, respectivamente, para as temperaturas de 16, 19, 25 e 28 ºC
(Tab. 7). A mesma tendência foi observada por Walton; Pringle (2005), para os ovos
de P. ficus em plantas de videira em temperaturas que variaram entre 18 (11,70 ±
0,12 dias) e 30 ºC (6,37 ± 0,2 dias), respectivamente.
A redução das durações médias para o estágio de ovo, também foi detectada
por Chong et al. (2003), para Phenacoccus madeirensis Green (Hemiptera:
Pseudococcidae) em crisântemo (Dendrathema grandiflora Tzevelev) nas
temperaturas entre 15 a 35 ºC. A duração média registrada pelos autores a 15 ºC
(19,9 ± 0,1), 20 ºC (13,5 ± 0,1), 25ºC (8,2 ± 0,1), 30ºC (6,3 ± 0,1) e 35 ºC (6,1 ±
0,02). Uma possível explicação, para o fato de alguns autores conseguirem
desenvolver a cochonilha a temperaturas acima de 30 ºC seria a adaptação da
população a temperaturas mais quentes.
O limiar térmico inferior de desenvolvimento (Tb) para o estágio de ovo de P.
citri foi de 10,62 °C e a constante térmica (K) de 64,60 graus-dia (GD) (Tab. 9). Um
valor de Tb (10,96) e GD (61,45) próximos a este foi registrado para os ovos da
mesma espécie de cochonilha em abóboras por Correa et al. (2007).
Cabe ressaltar que ocorreu um aumento na duração média da fase de ovo em
1,6 dias na temperatura de 31ºC, em relação à de 28ºC, indicando que a
temperatura base superior foi ultrapassada, resultando numa redução na velocidade
de desenvolvimento (Tab. 7). Este fato foi semelhante aos resultados de Correa et
al. (2007) que verificaram a mesma tendência quando inseto foi criado em abóbora.
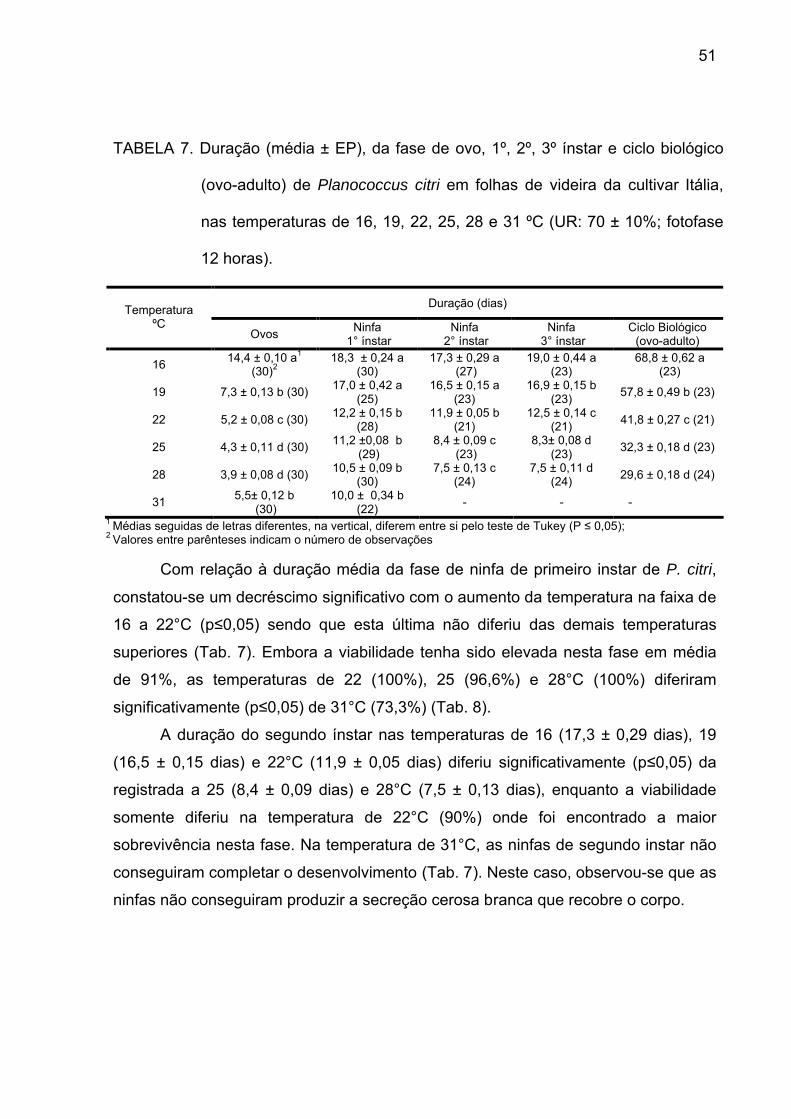
51
TABELA 7. Duração (média ± EP), da fase de ovo, 1º, 2º, 3º ínstar e ciclo biológico
(ovo-adulto) de Planococcus citri em folhas de videira da cultivar Itália,
nas temperaturas de 16, 19, 22, 25, 28 e 31 ºC (UR: 70 ± 10%; fotofase
12 horas).
Duração (dias)Temperatura
ºC Ovos
Ninfa 1° ínstar
Ninfa 2° ínstar
Ninfa3° ínstar
Ciclo Biológico (ovo-adulto)
1614,4 ± 0,10 a1
(30)218,3 ± 0,24 a
(30)17,3 ± 0,29 a
(27)19,0 ± 0,44 a
(23)68,8 ± 0,62 a
(23)
19 7,3 ± 0,13 b (30)17,0 ± 0,42 a
(25)16,5 ± 0,15 a
(23)16,9 ± 0,15 b
(23)57,8 ± 0,49 b (23)
22 5,2 ± 0,08 c (30)12,2 ± 0,15 b
(28)11,9 ± 0,05 b
(21)12,5 ± 0,14 c
(21)41,8 ± 0,27 c (21)
25 4,3 ± 0,11 d (30)11,2 ±0,08 b
(29)8,4 ± 0,09 c
(23)8,3± 0,08 d
(23)32,3 ± 0,18 d (23)
28 3,9 ± 0,08 d (30)10,5 ± 0,09 b
(30)7,5 ± 0,13 c
(24)7,5 ± 0,11 d
(24)29,6 ± 0,18 d (24)
315,5± 0,12 b
(30)10,0 ± 0,34 b
(22)- - -
1 Médias seguidas de letras diferentes, na vertical, diferem entre si pelo teste de Tukey (P ≤ 0,05); 2 Valores entre parênteses indicam o número de observações
Com relação à duração média da fase de ninfa de primeiro instar de P. citri,
constatou-se um decréscimo significativo com o aumento da temperatura na faixa de
16 a 22°C (p≤0,05) sendo que esta última não diferiu das demais temperaturas
superiores (Tab. 7). Embora a viabilidade tenha sido elevada nesta fase em média
de 91%, as temperaturas de 22 (100%), 25 (96,6%) e 28°C (100%) diferiram
significativamente (p≤0,05) de 31°C (73,3%) (Tab. 8).
A duração do segundo ínstar nas temperaturas de 16 (17,3 ± 0,29 dias), 19
(16,5 ± 0,15 dias) e 22°C (11,9 ± 0,05 dias) diferiu significativamente (p≤0,05) da
registrada a 25 (8,4 ± 0,09 dias) e 28°C (7,5 ± 0,13 dias), enquanto a viabilidade
somente diferiu na temperatura de 22°C (90%) onde foi encontrado a maior
sobrevivência nesta fase. Na temperatura de 31°C, as ninfas de segundo instar não
conseguiram completar o desenvolvimento (Tab. 7). Neste caso, observou-se que as
ninfas não conseguiram produzir a secreção cerosa branca que recobre o corpo.
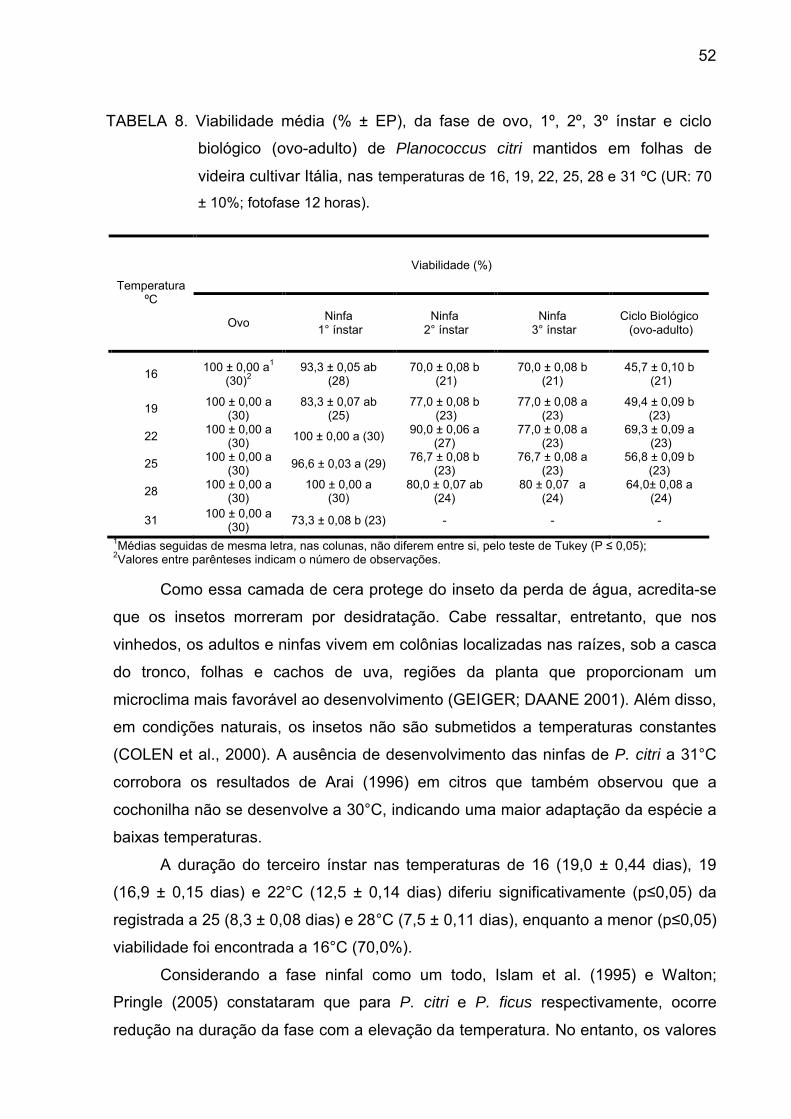
52
TABELA 8. Viabilidade média (% ± EP), da fase de ovo, 1º, 2º, 3º ínstar e ciclo
biológico (ovo-adulto) de Planococcus citri mantidos em folhas de
videira cultivar Itália, nas temperaturas de 16, 19, 22, 25, 28 e 31 ºC (UR: 70
± 10%; fotofase 12 horas).
1Médias seguidas de mesma letra, nas colunas, não diferem entre si, pelo teste de Tukey (P ≤ 0,05); 2Valores entre parênteses indicam o número de observações.
Como essa camada de cera protege do inseto da perda de água, acredita-se
que os insetos morreram por desidratação. Cabe ressaltar, entretanto, que nos
vinhedos, os adultos e ninfas vivem em colônias localizadas nas raízes, sob a casca
do tronco, folhas e cachos de uva, regiões da planta que proporcionam um
microclima mais favorável ao desenvolvimento (GEIGER; DAANE 2001). Além disso,
em condições naturais, os insetos não são submetidos a temperaturas constantes
(COLEN et al., 2000). A ausência de desenvolvimento das ninfas de P. citri a 31°C
corrobora os resultados de Arai (1996) em citros que também observou que a
cochonilha não se desenvolve a 30°C, indicando uma maior adaptação da espécie a
baixas temperaturas.
A duração do terceiro ínstar nas temperaturas de 16 (19,0 ± 0,44 dias), 19
(16,9 ± 0,15 dias) e 22°C (12,5 ± 0,14 dias) diferiu significativamente (p≤0,05) da
registrada a 25 (8,3 ± 0,08 dias) e 28°C (7,5 ± 0,11 dias), enquanto a menor (p≤0,05)
viabilidade foi encontrada a 16°C (70,0%).
Considerando a fase ninfal como um todo, Islam et al. (1995) e Walton;
Pringle (2005) constataram que para P. citri e P. ficus respectivamente, ocorre
redução na duração da fase com a elevação da temperatura. No entanto, os valores
Viabilidade (%)
TemperaturaºC
OvoNinfa
1° ínstarNinfa
2° ínstarNinfa
3° ínstarCiclo Biológico (ovo-adulto)
16100 ± 0,00 a1
(30)293,3 ± 0,05 ab
(28)70,0 ± 0,08 b
(21)70,0 ± 0,08 b
(21)45,7 ± 0,10 b
(21)
19100 ± 0,00 a
(30)83,3 ± 0,07 ab
(25)77,0 ± 0,08 b
(23)77,0 ± 0,08 a
(23)49,4 ± 0,09 b
(23)
22100 ± 0,00 a
(30)100 ± 0,00 a (30)
90,0 ± 0,06 a (27)
77,0 ± 0,08 a (23)
69,3 ± 0,09 a (23)
25100 ± 0,00 a
(30)96,6 ± 0,03 a (29)
76,7 ± 0,08 b (23)
76,7 ± 0,08 a (23)
56,8 ± 0,09 b (23)
28100 ± 0,00 a
(30)100 ± 0,00 a
(30)80,0 ± 0,07 ab
(24)80 ± 0,07 a
(24)64,0± 0,08 a
(24)
31100 ± 0,00 a
(30)73,3 ± 0,08 b (23) - - -

53
registrados por Islam et al. (1995), foram superiores a este trabalho, enquanto, os
encontrados por Walton; Pringle (2005) foram inferiores.
O limiar térmico inferior de desenvolvimento (Tb=8,16) para os instares ninfais
(1º, 2º e 3º) de P. citri foram inferiores ao encontrado por Correa et al. (2008) (Tb =
10,6) para a mesma fase na cultura do café. Laflin; Parrella (2004) encontraram para
a mesma espécie, tendo como hospedeiro roseira (Rosa hybrida L.) uma Tb = 11ºC
e uma constante térmica, inferior a este trabalho (327 GD). Atribui-se a variação
encontrada ao hospedeiro e população de P. citri com que foram conduzidos os
trabalhos visto que recursos alimentares de baixa qualidade nutricional podem exigir
um tempo maior de alimentação para garantir os nutrientes necessários para o
crescimento e desenvolvimento (STRONG et al., 1984).
O maior valor de Tb foi o obtido para a fase de ovo (10,62ºC) (Tab. 9),
concordando com Correa et al. (2007) que registraram uma Tb de 10,96 ºC e uma
constante térmica inferior de 61,45 GD similar aos 64,60 GD.
A duração média do ciclo biológico (ovo-adulto) de P. citri apresentou relação
inversa à elevação da temperatura (Tab. 9). Aproximadamente 98% do decréscimo
do tempo de desenvolvimento foi explicado pelo aumento na temperatura (R2 = 0,
98).
No intervalo de temperatura avaliado, foi observada diferença significativa
(p≤0,05) na duração do ciclo biológico, dos 16ºC (68,8 ± 0,62 dias) até 25ºC (32,3 ±
0,18 dias), sendo que a partir desta temperatura, os valores registrados não
diferiram (Tab. 7). O alongamento do ciclo biológico em função de baixas
temperaturas é comum entre os insetos, sendo esta uma das estratégias utilizadas
para as espécies sobreviverem em baixas temperaturas (LEATHER et al., 1993).
Islam et al. (1995), relataram uma duração do ciclo total de P. citri a 24ºC em
brotos de batata de 41,96 ± 0,67 dias, valores estes superiores aos 32,3 0,18 dias
encontrados neste experimento a 25ºC. Em abóboras, Malleshaiah et al. (2000),
constataram 31,45 0,96 dias a 26 2°C.
A menor viabilidade do ciclo biológico de P. citri foi de 45,7% (16 ºC) e a maior
69,3% (22 ºC), sendo significativa a diferença para os valores obtidos (p≤0,05). Com
base na duração do ciclo biológico e viabilidade (Tab. 7 e 8), verifica-se que
temperaturas entre 25 a 28ºC são as mais adequadas para o desenvolvimento do
inseto. Estes resultados corroboram as observações realizadas por Islam et al.
(1995) em brotos de batata, Arai et al. (1996) em citros e Laflin; Parrella (2004) em
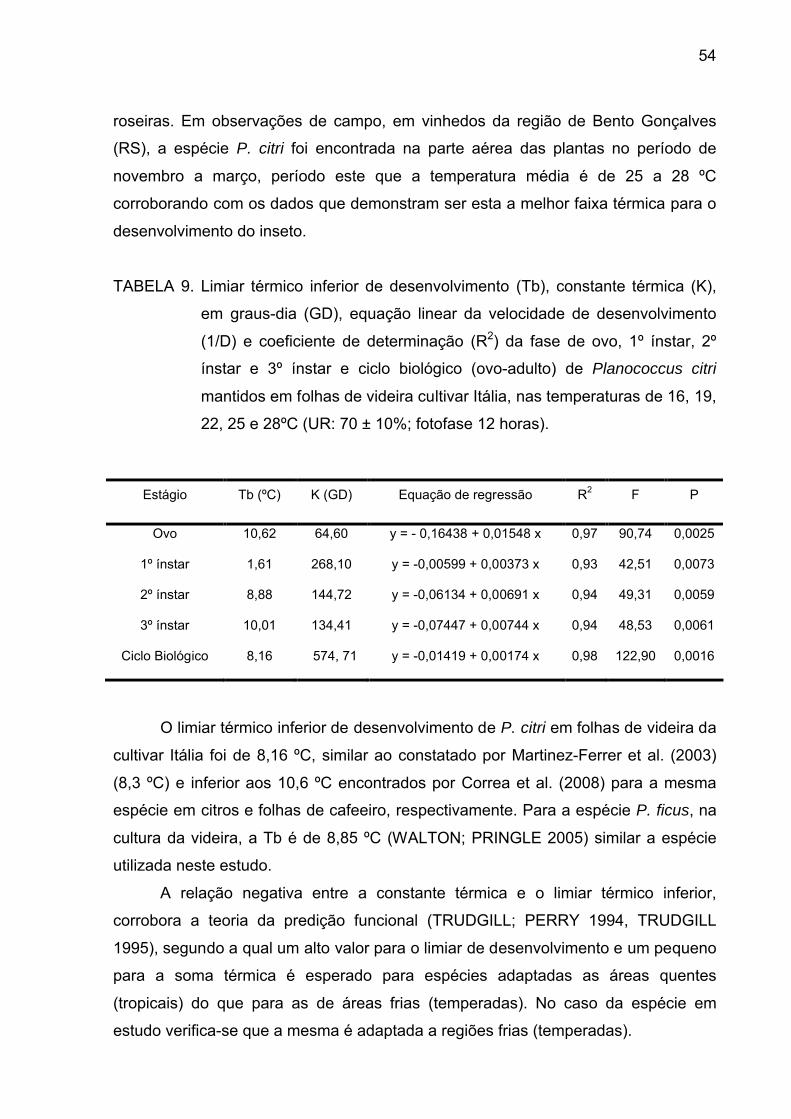
54
roseiras. Em observações de campo, em vinhedos da região de Bento Gonçalves
(RS), a espécie P. citri foi encontrada na parte aérea das plantas no período de
novembro a março, período este que a temperatura média é de 25 a 28 ºC
corroborando com os dados que demonstram ser esta a melhor faixa térmica para o
desenvolvimento do inseto.
TABELA 9. Limiar térmico inferior de desenvolvimento (Tb), constante térmica (K),
em graus-dia (GD), equação linear da velocidade de desenvolvimento
(1/D) e coeficiente de determinação (R2) da fase de ovo, 1º ínstar, 2º
ínstar e 3º ínstar e ciclo biológico (ovo-adulto) de Planococcus citri
mantidos em folhas de videira cultivar Itália, nas temperaturas de 16, 19,
22, 25 e 28ºC (UR: 70 ± 10%; fotofase 12 horas).
Estágio Tb (ºC) K (GD) Equação de regressão R2 F P
Ovo 10,62 64,60 y = - 0,16438 + 0,01548 x 0,97 90,74 0,0025
1º ínstar 1,61 268,10 y = -0,00599 + 0,00373 x 0,93 42,51 0,0073
2º ínstar 8,88 144,72 y = -0,06134 + 0,00691 x 0,94 49,31 0,0059
3º ínstar 10,01 134,41 y = -0,07447 + 0,00744 x 0,94 48,53 0,0061
Ciclo Biológico 8,16 574, 71 y = -0,01419 + 0,00174 x 0,98 122,90 0,0016
O limiar térmico inferior de desenvolvimento de P. citri em folhas de videira da
cultivar Itália foi de 8,16 ºC, similar ao constatado por Martinez-Ferrer et al. (2003)
(8,3 ºC) e inferior aos 10,6 ºC encontrados por Correa et al. (2008) para a mesma
espécie em citros e folhas de cafeeiro, respectivamente. Para a espécie P. ficus, na
cultura da videira, a Tb é de 8,85 ºC (WALTON; PRINGLE 2005) similar a espécie
utilizada neste estudo.
A relação negativa entre a constante térmica e o limiar térmico inferior,
corrobora a teoria da predição funcional (TRUDGILL; PERRY 1994, TRUDGILL
1995), segundo a qual um alto valor para o limiar de desenvolvimento e um pequeno
para a soma térmica é esperado para espécies adaptadas as áreas quentes
(tropicais) do que para as de áreas frias (temperadas). No caso da espécie em
estudo verifica-se que a mesma é adaptada a regiões frias (temperadas).

55
Segundo Honěk (1996), o limiar térmico inferior de desenvolvimento diminui
com o aumento da latitude. Espécies que vivem nos trópicos possuem uma
temperatura base maior (13,7 ºC) do que as que habitam regiões subtropicais (10,5
ºC) e temperadas (7,9 ºC). O valor do limiar térmico inferior calculado neste estudo
para o ciclo biológico (8,16 ºC) de P. citri demonstra que a população da espécie
esta adaptada a região de clima temperado.
No presente estudo, com base na Tb (8,16 ºC) constatou-se que P. citri
requer 574,71 GD para completar seu desenvolvimento. Valores inferiores foram
verificados para a mesma espécie por Correa et al. (2008) em folhas de cafeeiro
(334,90 GD), por Arai et al. (1996) em folhas de citros (401,00 GD) e por Laflin;
Parrella (2004) em roseiras (326,00 GD). Esta variação verificada nos valores
estimados para diferentes populações de P. citri se devem, segundo Laflin; Parrella
(2004) e Bergant; Trdan (2006) à qualidade nutricional da planta hospedeira, à
adaptação da população, às condições climáticas do local no qual vivem e ao
fotoperíodo.
O número estimado de gerações anuais de P.citri para os municípios
produtores de uva de Bento Gonçalves (RS), Bagé (RS), São Lourenço (MG),
Londrina (PR), Jales (SP) e Petrolina (PE) foram, respectivamente, de 5,7, 6,2, 6,9,
7,9, 9,7 e 11,5 (Tab. 10; Fig. 3). Desta maneira, um maior número de gerações é
esperado nas regiões mais quentes desde que as populações se adaptem as
condições climáticas de temperatura mais elevada.
Até o momento, P. citri tem sido relatada na região produtora da Serra
Gaúcha. Caso a espécie seja constatada nas demais regiões produtoras, os dados
de temperatura indicam que a mesma pode se adaptar e reproduzir nos vinhedos. A
determinação das exigências térmicas de P. citri em vinhedos poderá ser utilizada
como uma ferramenta para auxiliar na previsão de picos populacionais, indicando
quando as amostragens para monitoramento populacional devem ser iniciadas ou
intensificadas, e em que momento o controle deve ser implementado. Além disto,
estas informações são importantes para fins de estabelecimento de programas de
controle biológico, onde em determinadas épocas do ano as cochonilhas estão em
estágios de desenvolvimento com maior suscetibilidade ao parasitismo. Por
exemplo, Anagyrus pseudococci (Girault), Leptomastidea abnormis (Girault) e
Leptomastidea dactylopii (Howard) (Hymenoptera: Encyrtidae) são inimigos naturais
que parasitam fêmeas de cochonilhas da família Pseudococcidae. Desta maneira, os
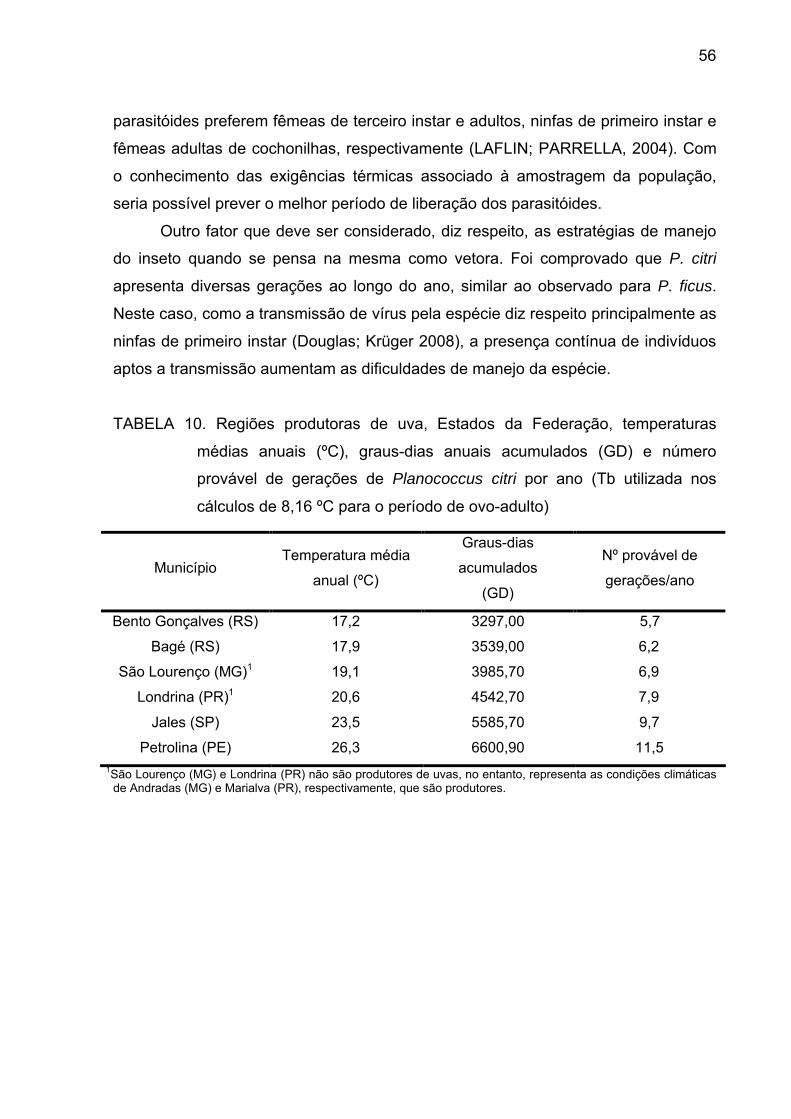
56
parasitóides preferem fêmeas de terceiro instar e adultos, ninfas de primeiro instar e
fêmeas adultas de cochonilhas, respectivamente (LAFLIN; PARRELLA, 2004). Com
o conhecimento das exigências térmicas associado à amostragem da população,
seria possível prever o melhor período de liberação dos parasitóides.
Outro fator que deve ser considerado, diz respeito, as estratégias de manejo
do inseto quando se pensa na mesma como vetora. Foi comprovado que P. citri
apresenta diversas gerações ao longo do ano, similar ao observado para P. ficus.
Neste caso, como a transmissão de vírus pela espécie diz respeito principalmente as
ninfas de primeiro instar (Douglas; Krüger 2008), a presença contínua de indivíduos
aptos a transmissão aumentam as dificuldades de manejo da espécie.
TABELA 10. Regiões produtoras de uva, Estados da Federação, temperaturas
médias anuais (ºC), graus-dias anuais acumulados (GD) e número
provável de gerações de Planococcus citri por ano (Tb utilizada nos
cálculos de 8,16 ºC para o período de ovo-adulto)
1São Lourenço (MG) e Londrina (PR) não são produtores de uvas, no entanto, representa as condições climáticas de Andradas (MG) e Marialva (PR), respectivamente, que são produtores.
MunicípioTemperatura média
anual (ºC)
Graus-dias
acumulados
(GD)
Nº provável de
gerações/ano
Bento Gonçalves (RS) 17,2 3297,00 5,7
Bagé (RS) 17,9 3539,00 6,2
São Lourenço (MG)1 19,1 3985,70 6,9
Londrina (PR)1 20,6 4542,70 7,9
Jales (SP) 23,5 5585,70 9,7
Petrolina (PE) 26,3 6600,90 11,5
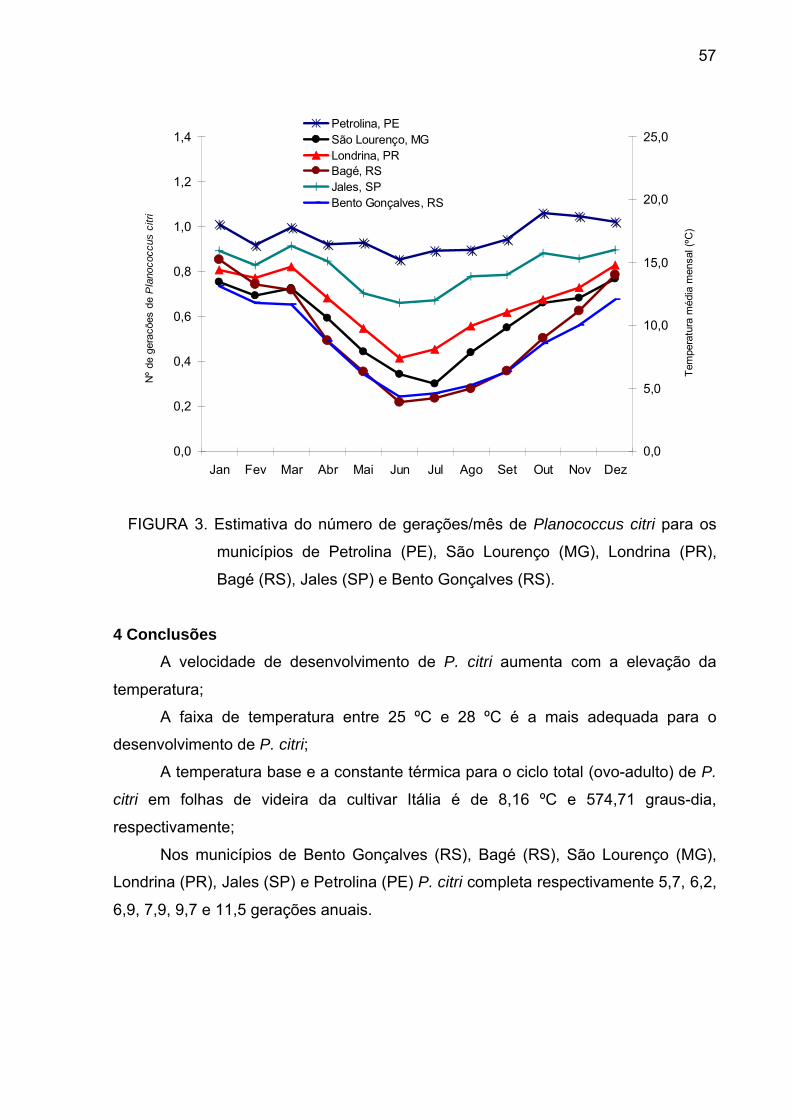
57
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
Jan Fev Mar Abr Mai Jun Jul Ago Set Out Nov Dez
Nº
de g
erac
ões
de P
lano
cocc
us c
itri
0,0
5,0
10,0
15,0
20,0
25,0
Tem
pera
tura
méd
ia m
ensa
l (ºC
)
Petrolina, PESão Lourenço, MGLondrina, PRBagé, RSJales, SPBento Gonçalves, RS
FIGURA 3. Estimativa do número de gerações/mês de Planococcus citri para os
municípios de Petrolina (PE), São Lourenço (MG), Londrina (PR),
Bagé (RS), Jales (SP) e Bento Gonçalves (RS).
4 Conclusões
A velocidade de desenvolvimento de P. citri aumenta com a elevação da
temperatura;
A faixa de temperatura entre 25 ºC e 28 ºC é a mais adequada para o
desenvolvimento de P. citri;
A temperatura base e a constante térmica para o ciclo total (ovo-adulto) de P.
citri em folhas de videira da cultivar Itália é de 8,16 ºC e 574,71 graus-dia,
respectivamente;
Nos municípios de Bento Gonçalves (RS), Bagé (RS), São Lourenço (MG),
Londrina (PR), Jales (SP) e Petrolina (PE) P. citri completa respectivamente 5,7, 6,2,
6,9, 7,9, 9,7 e 11,5 gerações anuais.

58
CAPÍTULO 4
EFEITO DE INSETICIDAS SOBRE A COCHONILHA-FARINHENTA Planococcus
citri (RISSO, 1813) (HEMIPTERA: PSEUDOCOCCIDAE) NA CULTURA DA
VIDEIRA
1 Introdução
Dentre as espécies de cochonilhas-farinhentas (Hemiptera: Pseudococcidae)
relatadas para a cultura da videira no Brasil destacam-se P. maritimus, P.
longispinus, P. viburni (Signoret, 1875), P. citri (Risso, 1813) e P. ficus (Signoret,
1875) (SILVA et al., 1968; WOERNER, 1983; FOLDI; SORIA, 1989; AFONSO, 2005;
KUNIYUKI et al., 2005; FOLDI; KOZÁR, 2006; KISHINO et al., 2007; MORANDI
FILHO, et al., 2007). As cochonilhas desta família, entretanto, não têm sido relatadas
como pragas-chave causando danos diretos à produção nas uvas destinadas ao
processamento (BOTTON et al., 2003a). Os principais prejuízos causados pelas
cochonilhas são observados em uvas de mesa da cultivar Itália (V. vinifera) onde
danificam as bagas, provocando o aparecimento da fumagina o que inviabiliza a
fruta para o comércio “in natura” e, em alguns casos, devido as restrições
quarentenárias, podem impedir a exportação (KISHINO et al., 2007; MORANDI
FILHO et al., 2007). Entretanto, a importância destas cochonilhas na cultura da
videira tem aumentado nos últimos anos principalmente após a comprovação da
transmissão de vírus GLRaV 3 (Grapevine leafroll associated virus 3), GVA
(Grapevine trichovirus A) e GVB (Grapevine trichovirus B) pelas espécies
(ROSCIGLIONE; CASTELLANO, 1985; ENGELBRECHT; KASDORF, 1990; GARAU
et al., 1995; CABALEIRO; SEGURA, 1997a; CABALEIRO; SEGURA, 1997b;
PETERSEN; CHARLES, 1997; SFORZA et al., 2003; GOLINO et al., 2002; BORBÓN
et al., 2004; CID et al., 2007).

59
No Brasil, este fato já foi comprovado para os vírus GLRaV 3 e GVB para P.
longispinus (KUNIYUKI et al., 2005; KUNIYUKI et al., 2006) sendo que no caso de P.
viburni, somente foi verificado a aquisição do vírus pela cochonilha, faltando a
comprovação da transmissão para plantas sadias (AFONSO, 2005).
O controle das cochonilhas-farinhentas tem sido uma prática pouco realizada
pelos viticultores devido ao desconhecimento da ação vetora e também, devido ao
hábito das cochonilhas localizarem-se nas raízes e/ou sob a casca, o que dificulta a
visualização da praga e o contato com inseticidas (GEIGER; DAANE, 2001; FU et al.,
2002). Quando a infestação é observada nos cachos, geralmente os produtores
aplicam inseticidas fosforados, os quais além de apresentarem elevada toxicidade,
são pouco seletivos aos inimigos naturais e apresentam grande período de carência
(BOTTON et al., 2003b). Além disso, as pulverizações, dependendo da fase em que
são realizadas apresentam reduzida eficácia devido à localização da praga no solo,
sob o ritidoma e/ou no interior dos cachos, limitando o contato com os ingredientes
ativos.
Considerando o hábito alimentar (sugadores de seiva do floema) e a
localização das cochonilhas na planta, os inseticidas neonicotinóides e reguladores
de crescimento poderão servir como alternativa aos inseticidas fosforados. Neste
sentido, resultados promissores já foram registrados em outros países como por
exemplo Alemanha, Argentina, Chile, Estados Unidos, México (DIESTRE; SAZO,
1990; ELBERT et al., 1991; MENDEL et al., 1991; BROEKSMA et al., 1993;
OROZCO et al., 1995; GONZALEZ et al., 1995; LARRAIN, 1999; CLOYD, 2003;
MUTHUKRISNAN et al., 2005; DAANE et al., 2006; GONZÁLEZ; VOLOSKY, 2006).
No caso dos neonicotinóides, os melhores resultados têm sido obtidos quando os
produtos são aplicados via solo, associado ao sistema de irrigação, permitindo
atingir os insetos via ingestão (ABBINK, 1991; ELBERT et al., 1991; PALUMBO et
al., 1996; LARRAIN, 1999; CAPELLA et al., 2004; ERAZO, 2004; CASTILLO et al.,
2004). Este tipo de aplicação permitiria um controle localizado nos vinhedos,
preservando os inimigos naturais além de atingir os insetos no inicio da infestação.
Outra estratégia importante para o emprego de inseticidas sistêmicos via solo seria o
uso em matrizeiros de plantas, onde o tratamento preventivo com os produtos
minimizaria o potencial de dispersão de vírus pelas cochonilhas vetoras antes que as
plantas apresentem os sintomas, permitindo eliminar as plantas viróticas, reduzindo
os focos de infestação (CHARLES et al., 2006). Em alguns casos, a aplicação via

60
solo tem sido preterida pelos produtores principalmente pela dificuldade de distribuir
o inseticida em locais onde não há sistema de irrigação além do tratamento ter que
ser realizado de maneira preventiva, em função do histórico de ocorrência da praga,
resultando num aumento do custo de produção. Nestes casos, a avaliação de
inseticidas alternativos aos fosforados para o controle do inseto via pulverização
foliar também deve ser pesquisado.
O objetivo do trabalho foi avaliar o efeito de inseticidas reguladores de
crescimento e neonicotinóides via solo e pulverização foliar visando o controle de P.
citri na cultura da videira.
2 Material e Métodos
Os experimentos foram conduzidos em casa-de-vegetação e em vinhedo
comercial da cultivar Cabernet Sauvignon. A metodologia utilizada teve como base o
trabalho de González et al. (1995). Os inseticidas (Tab. 11) foram aplicados nas
plantas e posteriormente, folhas foram oferecidas às ninfas de primeiro ínstar de P.
citri avaliando-se a mortalidade em laboratório. Os insetos utilizados nos
experimentos foram provenientes da criação de manutenção de P. citri mantida no
Laboratório de Entomologia da Embrapa Uva e Vinho conforme metodologia descrita
por Lepage (1942) utilizando abóboras (Cucurbita maxima) da variedade Cabotchá.
Tabela 11. Inseticidas avaliados para o controle de Planococcus citri na cultura da
videira. Bento Gonçalves, RS, 2008.
1Nomenclatura de acordo com ABAKERLI et al. (2003);2S= Sistêmico; C=Contato;3CT= Classe Toxicológica: I=extremamente tóxico; II=Altamente tóxico; III=Medianamente tóxico; IV=Pouco Tóxico.
Ingrediente
Ativo (i.a)1
Produto
Comercial (p.c.)
Formulação Concentração
g/L ou g/Kg
Modo de
Ação2
CT3 Grupo Químico
Tiametoxam Actara® 250 WG Granulado
dispersível
250 S III Neonicotinóide
Acetamiprid Mospilan® Pó solúvel 200 S III Neonicotinóide
Imidacloprid Confidor® 700
GRDA
Granulado
dispersível
700 S IV Neonicotinóide
Pyriproxyfen Tiger® 100 CE Concentrado
Emulsionável
100 C I Piridil Éter
Buprofezin Applaud® 250 Pó Molhável 250 C IV Thiadiazina

61
2.1 Experimento em casa-de-vegetação
Mudas enraizadas de videira com um ano de idade da cultivar Cabernet
Sauvignon enxertadas sobre o porta-enxerto Paulsen 1103 foram plantadas em
baldes (uma muda/balde) com capacidade de 5 litros em outubro de 2006. Os
inseticidas e doses avaliadas foram: acetamiprid (Mospilan, 0,6g i.a./planta),
imidacloprid (Confidor 700 GRDA, 0,7g i.a./planta) e tiametoxam (Actara 250 WG,
0,75g i.a./planta). Os inseticidas foram aplicados via solo 30 dias após o plantio,
utilizando 200 mL de calda por planta. Estas doses foram selecionadas de acordo
com a empregada para o controle de outros insetos pragas da videira (CASTILLO et
al., 2004; GONZÁLEZ; VOLOSKY, 2006). Como testemunha foram mantidas plantas
sem aplicação dos inseticidas.
O delineamento experimental foi inteiramente casualizado com dez
repetições, sendo cada planta considerada uma repetição. Aos 3, 7, 14, 21, 28, 35 e
60 dias após a aplicação (DAA) dos inseticidas, em cada repetição foi retirado uma
folha (da região mediana da planta) a qual foi levada ao laboratório e acondicionada
no interior de uma placa de Petri contendo ágar-água (15%) ao fundo para evitar o
ressecamento. Cada uma das placas foi infestada com 5 ninfas de primeiro instar
com um dia de idade num total de 10 repetições por tratamento. A avaliação da
mortalidade dos insetos foi realizada 24, 48, 72 e 96 horas após o fornecimento das
folhas as ninfas sendo considerado indivíduo “vivo” aquele que apresentou
movimentos perceptíveis ao toque de um pincel sob microscópio estereoscópico (20
X). As avaliações da mortalidade causada pelos produtos foram realizadas até o
momento em que o número de ninfas sobreviventes nos tratamentos com inseticidas
igualou-se a testemunha (sem controle).
2.2 Experimento em vinhedo comercial
No experimento em vinhedo comercial foram utilizadas plantas da cultivar
Cabernet Sauvignon, plantadas em 1993, no espaçamento 2,7m x 1,7m, conduzidas
em sistema latada. Os inseticidas e doses avaliadas via solo foram: acetamiprid
(Mospilan, 0,6g i.a./planta), imidacloprid (Confidor 700 GRDA, 0,7g i.a./planta) e
tiametoxam (Actara 250, WG 0,75g i.a./planta). Neste mesmo vinhedo foram
avaliados, via foliar os inseticidas e doses: acetamiprid (Mospilan, 6g i.a./100L),
buprofezin (Applaud 250, 25g i.a./100L), imidacloprid (Confidor 700 GRDA, 7g

62
i.a./100L), pyriproxyfen (Tiger 100CE, 10g i.a. mL/100L), tiametoxam (Actara 250
WG, 7,5g i.a./100L) e mantendo-se um tratamento testemunha (somente água).
Os produtos foram aplicados via solo em 28 de outubro de 2007 diluídos em 1
litro de calda por planta com auxílio de um becker graduado. Previamente a
aplicação dos tratamentos, procedeu-se a limpeza do solo ao redor do tronco das
plantas, num raio de 60 cm, retirando as plantas invasoras existentes. As
pulverizações via foliar, foram realizadas em 09 de janeiro de 2008 com pulverizador
costal manual, modelo PJH 20, com capacidade de 20 litros, equipado com bico de
cone vazio, modelo JA-1-5, num volume aproximado de 800 L/ha. No experimento
onde os inseticidas foram aplicados via solo, a metodologia e o período de avaliação
foram os mesmos do experimento conduzido em casa-de-vegetação. No caso do
experimento onde os produtos foram pulverizados via foliar, a mortalidade foi
avaliada aos 1, 5 e 10 (DAA).
2.3 Análise estatística
Os dados foram submetidos à análise de variância e as médias comparadas
pelo teste de Tukey (p≤0,05), com auxilio do programa Sisvar (Furtado, 2007) sendo
a porcentagem de mortalidade calculada pela fórmula de Abbott (1925). Os dados
não foram transformados, pois apresentaram homogeneidade da variância.
3 RESULTADOS E DISCUSSÃO
3.1 Experimento em casa-de-vegetação
Em todas as avaliações realizadas após a aplicação dos inseticidas, a
mortalidade observada às 24 horas após a infestação com as ninfas foi equivalente
(p≤0,05) a realizada às 96 horas. Por este motivo, todos os resultados serão
discutidos com base na mortalidade observada às 96 horas.
Na primeira avaliação realizada 3 DAA, foi observada uma redução
mortalidade das ninfas sendo de 46,67, 46,67 e 48,89% para os inseticidas,
tiametoxam, acetamiprid e imidacloprid, respectivamente (Tab. 12). Estes resultados,
entretanto, foram equivalentes (p≤0,05) ao tratamento testemunha (Tab. 12). Este
fato foi atribuído, provavelmente ao tempo necessário a absorção dos inseticidas
pelas raízes e deslocamento até as folhas onde foram ingeridos pela cochonilha.
Byrne; Toscano (2006) realizaram estudo em vinhedos irrigados na Califórnia
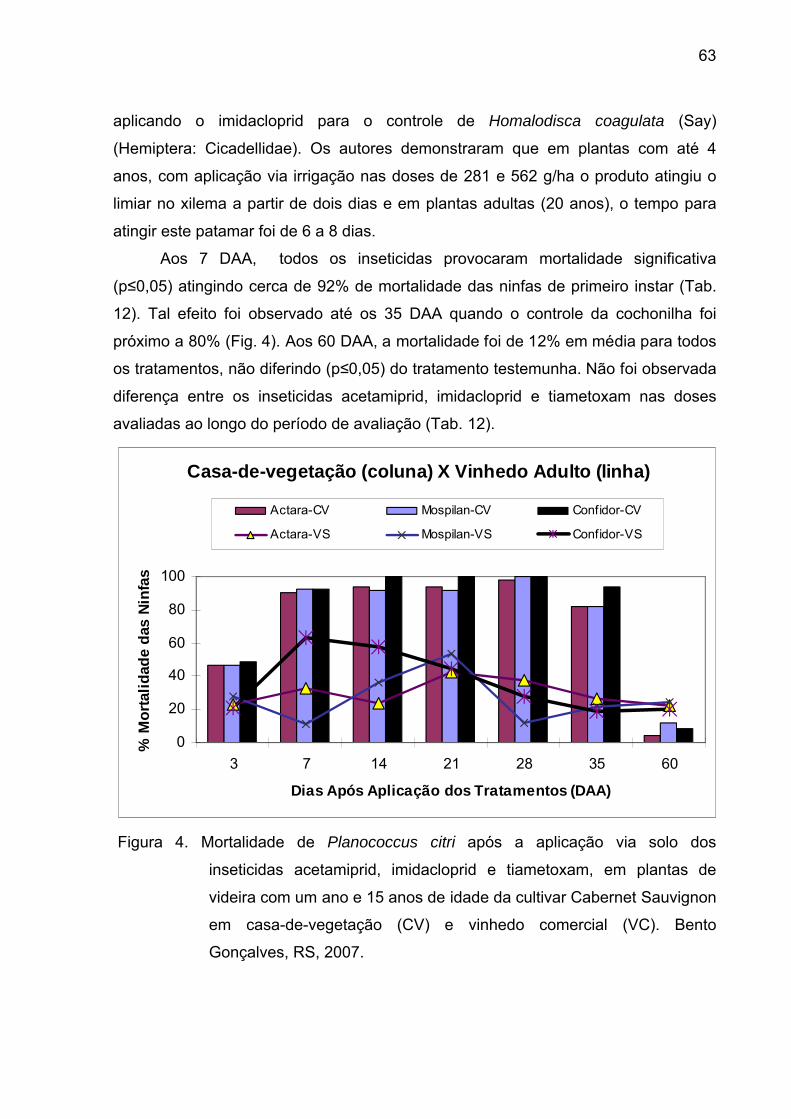
63
aplicando o imidacloprid para o controle de Homalodisca coagulata (Say)
(Hemiptera: Cicadellidae). Os autores demonstraram que em plantas com até 4
anos, com aplicação via irrigação nas doses de 281 e 562 g/ha o produto atingiu o
limiar no xilema a partir de dois dias e em plantas adultas (20 anos), o tempo para
atingir este patamar foi de 6 a 8 dias.
Aos 7 DAA, todos os inseticidas provocaram mortalidade significativa
(p≤0,05) atingindo cerca de 92% de mortalidade das ninfas de primeiro instar (Tab.
12). Tal efeito foi observado até os 35 DAA quando o controle da cochonilha foi
próximo a 80% (Fig. 4). Aos 60 DAA, a mortalidade foi de 12% em média para todos
os tratamentos, não diferindo (p≤0,05) do tratamento testemunha. Não foi observada
diferença entre os inseticidas acetamiprid, imidacloprid e tiametoxam nas doses
avaliadas ao longo do período de avaliação (Tab. 12).
Casa-de-vegetação (coluna) X Vinhedo Adulto (linha)
0
20
40
60
80
100
3 7 14 21 28 35 60
Dias Após Aplicação dos Tratamentos (DAA)
% M
ort
alid
ade
das
Nin
fas
Actara-CV Mospilan-CV Confidor-CV
Actara-VS Mospilan-VS Confidor-VS
Figura 4. Mortalidade de Planococcus citri após a aplicação via solo dos
inseticidas acetamiprid, imidacloprid e tiametoxam, em plantas de
videira com um ano e 15 anos de idade da cultivar Cabernet Sauvignon
em casa-de-vegetação (CV) e vinhedo comercial (VC). Bento
Gonçalves, RS, 2007.

64
3.2 Experimento em Vinhedo Comercial
3.2.1 Aplicação Via Solo
Quando os inseticidas foram aplicados via solo, foi observado que aos 3 DAA,
todos os inseticidas avaliados não apresentaram diferenças (p≤0,05) em relação à
testemunha (Tab. 13). Aos 7 DAA, o inseticida imidacloprid apresentou maior
mortalidade das ninfas de P. citri, reduzindo a população em 63,04%, não diferindo
(p≤0,05) do tiametoxam (32,61%), mas diferindo (p≤0,05) do acetamiprid (10,87%)
sendo que este último foi equivalente a testemunha sem controle (Fig. 4). Aos 14
DAA, o inseticida imidacloprid foi o que apresentou maior mortalidade das ninfas de
P. citri (57,45%), seguido pelo acetamiprid (36,17%) e o tiametoxam 23,40%. Tal
efeito foi verificado até os 21 DAA. Aos 28 DAA, a mortalidade média proporcionada
pelos inseticidas foi próxima a 20%. Aos 35 DAA todos os tratamentos igualaram-se
a testemunha, fato também observado até os 60 DAA (Fig. 4).
González (2003), em experimento realizado em vinhedos com 15 anos de
idade no Chile, também avaliou os inseticidas neonicotinóides (acetamiprid,
imidacloprid e tiametoxam) para controle de P. viburni, P. longispinus e
Pseudococcus calceolariae (Maskell, 1879) (Hemiptera: Pseudococcidae)
encontrando elevada mortalidade das três espécies. Resultados similares foram
obtidos por Larrain (1999) no Chile e Erazo (2004) na Argentina que estudaram o
efeito de imidacloprid, aplicado via solo, em plantas adultas para o controle de ninfas
e adultos de P.viburni e P. ficus, respectivamente. No caso de P. citri, Broesksma et
al. (1993) demonstraram que em pomares de citros da África do Sul, o inseticida
imidacloprid aplicado via solo foi eficaz no controle da praga por 80 dias.
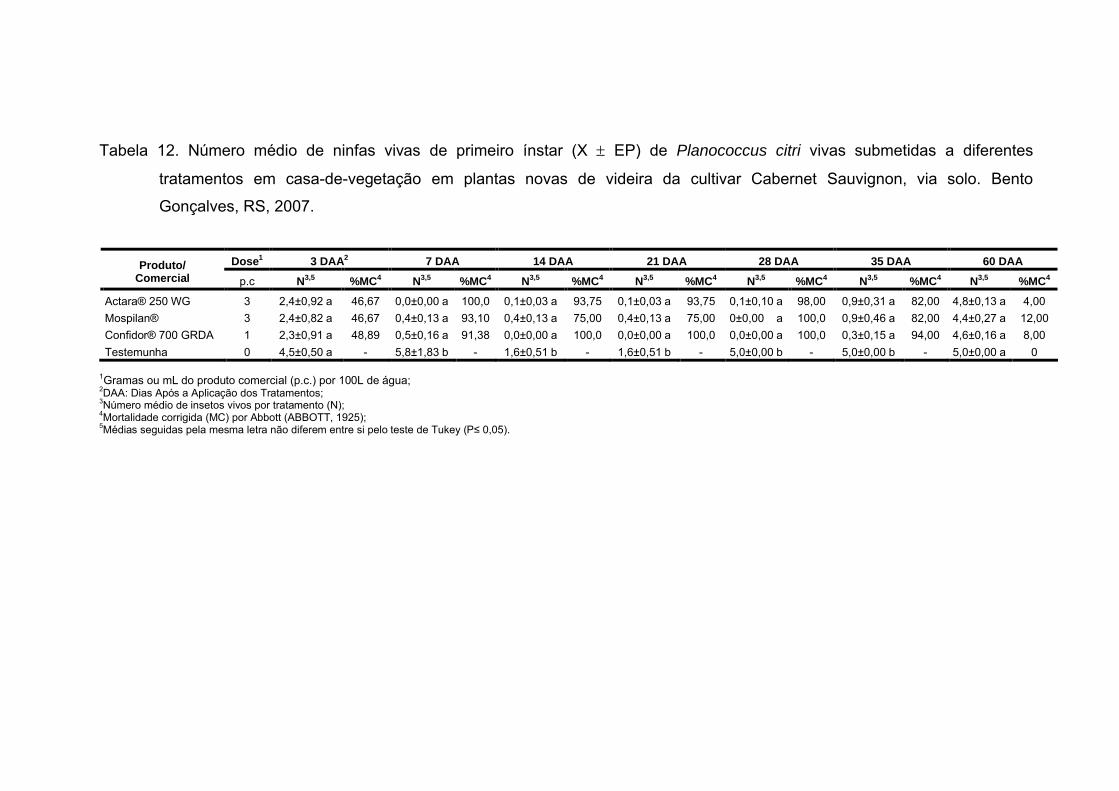
Tabela 12. Número médio de ninfas vivas de primeiro ínstar (X EP) de Planococcus citri vivas submetidas a diferentes
tratamentos em casa-de-vegetação em plantas novas de videira da cultivar Cabernet Sauvignon, via solo. Bento
Gonçalves, RS, 2007.
1Gramas ou mL do produto comercial (p.c.) por 100L de água;2DAA: Dias Após a Aplicação dos Tratamentos;3Número médio de insetos vivos por tratamento (N);4Mortalidade corrigida (MC) por Abbott (ABBOTT, 1925);5Médias seguidas pela mesma letra não diferem entre si pelo teste de Tukey (P≤ 0,05).
Dose1 3 DAA2 7 DAA 14 DAA 21 DAA 28 DAA 35 DAA 60 DAAProduto/Comercial p.c N3,5 %MC4 N3,5 %MC4 N3,5 %MC4 N3,5 %MC4 N3,5 %MC4 N3,5 %MC4 N3,5 %MC4
Actara® 250 WG 3 2,4±0,92 a 46,67 0,0±0,00 a 100,0 0,1±0,03 a 93,75 0,1±0,03 a 93,75 0,1±0,10 a 98,00 0,9±0,31 a 82,00 4,8±0,13 a 4,00
Mospilan® 3 2,4±0,82 a 46,67 0,4±0,13 a 93,10 0,4±0,13 a 75,00 0,4±0,13 a 75,00 0±0,00 a 100,0 0,9±0,46 a 82,00 4,4±0,27 a 12,00
Confidor® 700 GRDA 1 2,3±0,91 a 48,89 0,5±0,16 a 91,38 0,0±0,00 a 100,0 0,0±0,00 a 100,0 0,0±0,00 a 100,0 0,3±0,15 a 94,00 4,6±0,16 a 8,00
Testemunha 0 4,5±0,50 a - 5,8±1,83 b - 1,6±0,51 b - 1,6±0,51 b - 5,0±0,00 b - 5,0±0,00 b - 5,0±0,00 a 0
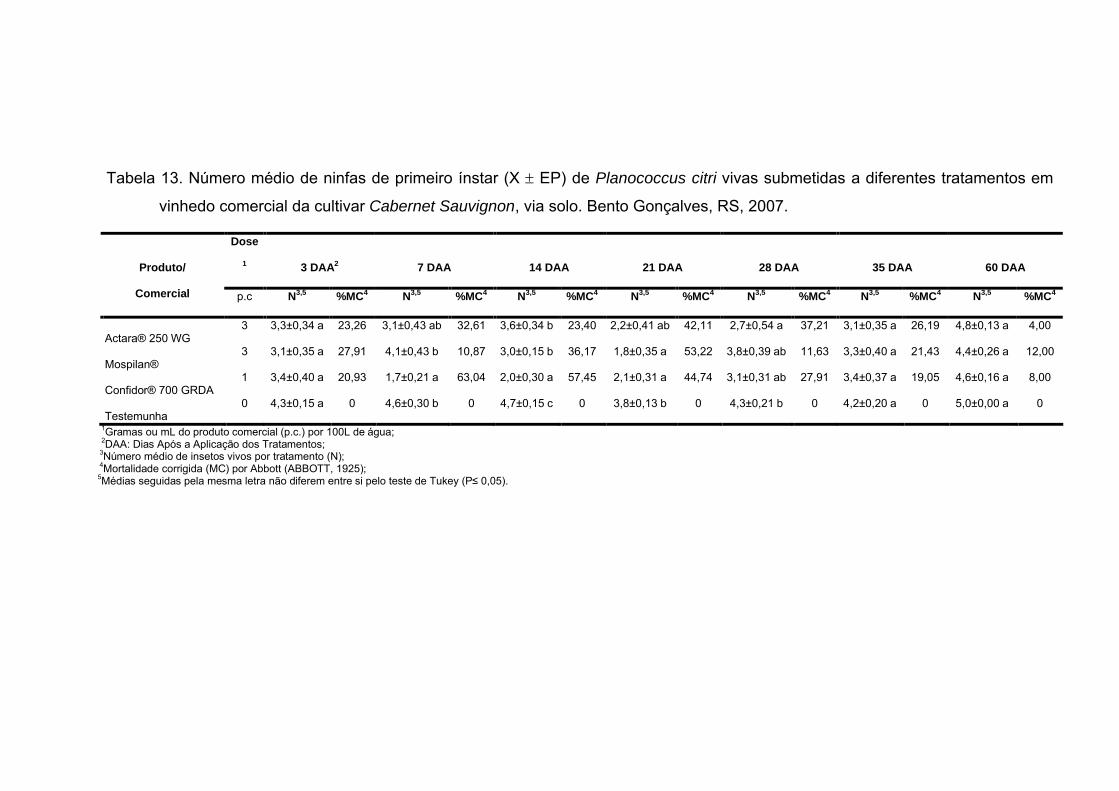
66
Tabela 13. Número médio de ninfas de primeiro ínstar (X EP) de Planococcus citri vivas submetidas a diferentes tratamentos em
vinhedo comercial da cultivar Cabernet Sauvignon, via solo. Bento Gonçalves, RS, 2007.
1Gramas ou mL do produto comercial (p.c.) por 100L de água; 2DAA: Dias Após a Aplicação dos Tratamentos; 3Número médio de insetos vivos por tratamento (N); 4Mortalidade corrigida (MC) por Abbott (ABBOTT, 1925); 5Médias seguidas pela mesma letra não diferem entre si pelo teste de Tukey (P≤ 0,05).
Dose
1 3 DAA2 7 DAA 14 DAA 21 DAA 28 DAA 35 DAA 60 DAAProduto/
Comercial p.c N3,5 %MC4 N3,5 %MC4 N3,5 %MC4 N3,5 %MC4 N3,5 %MC4 N3,5 %MC4 N3,5 %MC4
Actara® 250 WG3 3,3±0,34 a 23,26 3,1±0,43 ab 32,61 3,6±0,34 b 23,40 2,2±0,41 ab 42,11 2,7±0,54 a 37,21 3,1±0,35 a 26,19 4,8±0,13 a 4,00
Mospilan®3 3,1±0,35 a 27,91 4,1±0,43 b 10,87 3,0±0,15 b 36,17 1,8±0,35 a 53,22 3,8±0,39 ab 11,63 3,3±0,40 a 21,43 4,4±0,26 a 12,00
Confidor® 700 GRDA1 3,4±0,40 a 20,93 1,7±0,21 a 63,04 2,0±0,30 a 57,45 2,1±0,31 a 44,74 3,1±0,31 ab 27,91 3,4±0,37 a 19,05 4,6±0,16 a 8,00
Testemunha0 4,3±0,15 a 0 4,6±0,30 b 0 4,7±0,15 c 0 3,8±0,13 b 0 4,3±0,21 b 0 4,2±0,20 a 0 5,0±0,00 a 0

Por outro lado, Larrain (1999) verificou em vinhedos adultos da cultivar
Thompson Seedless no Chile, que o imidacloprid, 0,7 g i.a/planta (Confidor® 350
SC), quando utilizado para controlar P. viburni, foi absorvido pelas raízes ao ser
aplicado via solo, reduzindo a população de cochonilhas que estavam nos cachos
em 65,40%, neste caso, com resultados semelhantes aos 63% observados neste
experimento aos 14 DAA (Tab. 14). A mesma autora verificou que o inseticida foi
eficaz no controle da praga por um período de 40 dias após a aplicação.
Castillo et al. (2004) avaliaram no México, diferentes doses de imidacloprid
em plantas de videira com 20 anos de idade da cultivar Flame Seedless para o
controle de P. fícus. Os autores verificaram que doses de 0,15 e 0,12 g.ia/planta
reduziram a população do inseto em 87 e 86% por 77 dias. Na Argentina, Erazo
(2004) demonstrou efeito similar do imidacloprid via irrigação para o controle de P.
fícus, demonstrando que o produto reduziu os danos nos rácimos em 90% por um
período de 45 dias.
No presente trabalho, a mortalidade de P. citri obtida em plantas novas de
videira, demonstrou que os inseticidas acetamiprid, imidacloprid e thiamethoxam
foram equivalentes entre si. Em plantas adultas, entretanto, os melhores resultados
foram obtidos com o imidacloprid, mesmo assim com um máximo de controle de
63% aos 7 DAA. Neste caso, evidencia-se que os inseticidas neonicotinóides
acetamiprid, imidacloprid e tiametoxam apresentaram atividade biológica contra a
praga reduzindo a infestação na cultura da videira, porém, ajustes em relação à
dose empregada devem ser feitos de acordo com a idade das plantas. O melhor
desempenho dos inseticidas neonicotinóides verificados em outros países em
vinhedos comerciais adultos pode ser explicado, pelo fato que a maioria das
aplicações terem sido realizadas via irrigação, e em uva de mesa, o que permite
direcionar os tratamentos na zona onde as raízes estão ativas. Além disso, de
maneira geral, os solos em alguns países como Chile e Argentina onde a videira é
cultivada são arenosos e com baixa quantidade de matéria orgânica (1,2%)
(GONZÁLEZ et al., 1995, LARRAÍN, 1999; ERAZO, 2004) o que permite uma melhor
absorção deste tipo de composto (LARRAÍN, 1999; RAMAKRISHNAN et al., 2000;
BYRNE; TOSCANO, 2006).
66
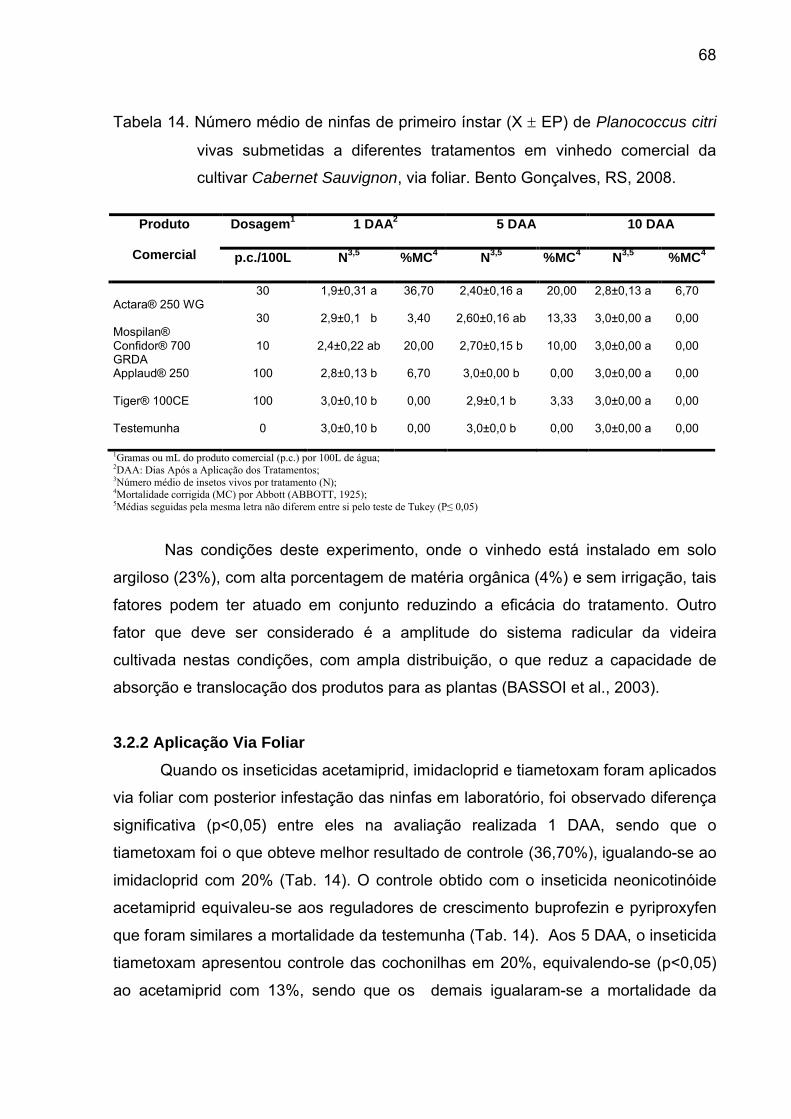
68
Tabela 14. Número médio de ninfas de primeiro ínstar (X EP) de Planococcus citri
vivas submetidas a diferentes tratamentos em vinhedo comercial da
cultivar Cabernet Sauvignon, via foliar. Bento Gonçalves, RS, 2008.
1Gramas ou mL do produto comercial (p.c.) por 100L de água;2DAA: Dias Após a Aplicação dos Tratamentos;3Número médio de insetos vivos por tratamento (N);4Mortalidade corrigida (MC) por Abbott (ABBOTT, 1925);5Médias seguidas pela mesma letra não diferem entre si pelo teste de Tukey (P≤ 0,05)
Nas condições deste experimento, onde o vinhedo está instalado em solo
argiloso (23%), com alta porcentagem de matéria orgânica (4%) e sem irrigação, tais
fatores podem ter atuado em conjunto reduzindo a eficácia do tratamento. Outro
fator que deve ser considerado é a amplitude do sistema radicular da videira
cultivada nestas condições, com ampla distribuição, o que reduz a capacidade de
absorção e translocação dos produtos para as plantas (BASSOI et al., 2003).
3.2.2 Aplicação Via Foliar
Quando os inseticidas acetamiprid, imidacloprid e tiametoxam foram aplicados
via foliar com posterior infestação das ninfas em laboratório, foi observado diferença
significativa (p<0,05) entre eles na avaliação realizada 1 DAA, sendo que o
tiametoxam foi o que obteve melhor resultado de controle (36,70%), igualando-se ao
imidacloprid com 20% (Tab. 14). O controle obtido com o inseticida neonicotinóide
acetamiprid equivaleu-se aos reguladores de crescimento buprofezin e pyriproxyfen
que foram similares a mortalidade da testemunha (Tab. 14). Aos 5 DAA, o inseticida
tiametoxam apresentou controle das cochonilhas em 20%, equivalendo-se (p<0,05)
ao acetamiprid com 13%, sendo que os demais igualaram-se a mortalidade da
Dosagem1 1 DAA2 5 DAA 10 DAAProduto
Comercial p.c./100L N3,5 %MC4 N3,5 %MC4 N3,5 %MC4
Actara® 250 WG30 1,9±0,31 a 36,70 2,40±0,16 a 20,00 2,8±0,13 a 6,70
Mospilan®30 2,9±0,1 b 3,40 2,60±0,16 ab 13,33 3,0±0,00 a 0,00
Confidor® 700 GRDA
10 2,4±0,22 ab 20,00 2,70±0,15 b 10,00 3,0±0,00 a 0,00
Applaud® 250 100 2,8±0,13 b 6,70 3,0±0,00 b 0,00 3,0±0,00 a 0,00
Tiger® 100CE 100 3,0±0,10 b 0,00 2,9±0,1 b 3,33 3,0±0,00 a 0,00
Testemunha 0 3,0±0,10 b 0,00 3,0±0,0 b 0,00 3,0±0,00 a 0,00

69
testemunha. Este fato não foi verificado aos 10 DAA, onde todos os tratamentos
igualaram-se a tratamento testemunha (Tab. 14).
Erazo (2004) avaliou o efeito de aplicações foliares de imidacloprid 20% SL
(20 g.i.a./100L), mostrando um controle efetivo de P. ficus em folhas e frutos de
videira. Resultados semelhantes foram verificados por Larrain (1999), em vinhedos
do Chile, avaliando o efeito de imidacloprid via aplicação foliar (21 g i.a./100L) para o
controle de P. viburni, resultando em presença do inseto a níveis inferiores a 20%
nos cachos. Os resultados obtidos neste trabalho, não corroboraram os verificados
em outros países que demonstraram uma eficácia dos inseticidas neonicotinóides
quando aplicados via foliar para o controle de diferentes espécies de
Pseudococcidae. Acredita-se que o principal fator para estas diferenças deve-se a
metodologia empregada, visto que nos outros trabalhos, quando os inseticidas foram
aplicados, atingiram também os insetos via contato. Além disso, os pesquisadores
avaliaram a presença/danos dos pseudococcídeos nos cachos de uva e não a
mortalidade direta causada nas cochonilhas. No presente trabalho os produtos não
entraram em contato direto com os insetos, o que pode ter resultado na redução da
eficiência de controle.
Mendel et al. (1991), avaliaram o efeito do regulador de crescimento
buprofezin, em condições de laboratório, sobre ninfas de primeiro e segundo ínstar e
fêmeas de P. citri, demonstrando que quando aplicado sobre as fêmeas, o mesmo
reduziu a fecundidade em 49%. Além disto, buprofezin 20 g.i.a/100L teve uma
elevada atividade ovicida (80%) para P. citri. No controle das ninfas, o inseticida foi
eficiente para ambas as espécies proporcionando uma mortalidade final acima de
90% em concentrações de 20 a 40g i.a/100L 28 (DAA). Neste trabalho, utilizando a
dose de 25g i.a./100L a mortalidade das ninfas foi de 6,7%. Ressalta-se que as
ninfas entraram em contato com os inseticidas via contato residual sem atingir
diretamente os insetos e no período de avaliação da mortalidade (96 horas), foi
possível observar a troca de instar, não observando-se efeito dos produtos durante a
ecdise.
Muthukrishnan et al. (2005), avaliaram em vinhedos o efeito do regulador de
crescimento buprofezin sobre ninfas da cochonilha Maconellicoccus hirsutus (Green,
1908) (Hemiptera: Pseudococcidae) aplicados via foliar nas concentrações de 56 e
75g i.a./100L, concluindo que após 3 dias de aplicação o produto não reduziu a
população do inseto. No entanto, foram feitas reaplicações dos tratamentos a cada

70
quinze dias resultando num controle de 76,70 e 79,00%, respectivamente. Estes
resultados contrastam com os encontrados neste trabalho provavelmente pela
diferença na metodologia e nas doses empregadas visto que o buprofezin tem sido
um dos produtos recomendados para o controle de P. ficus na Califórnia (DAANE et
al., 2006).
Comparando-se as formas de aplicação dos inseticidas neonicotinóides, foi
observado que as aplicações via solo apresentaram maior persistência biológica e
foram mais eficazes em relação as pulverizações foliares. O tratamento via solo teria
como vantagem adicional um menor efeito sobre os organismos benéficos
localizados na parte aérea das plantas (GUBLER et al., 1999). Além disso, os
inseticidas aplicados no solo permanecem protegidos da luz, reduzindo a
fotodegradação. No caso da pulverização foliar, embora em outros países tenham
sido observados resultados positivos com os neonicotinóides e reguladores de
crescimento, tais resultados não foram encontrados neste trabalho com a
metodologia empregada. Por isso, sugere-se que em trabalhos futuros os inseticidas
sejam empregados em doses maiores e/ou de maneira seqüencial permitindo atingir
diretamente a população de cochonilhas no vinhedo. Porém, para que isto ocorra,
trabalhos visando conhecer a estrutura etária da população ao longo do ano,
visando definir o melhor momento de controle, devem ser realizados.
4. Conclusões
Os inseticidas neonicotinóides acetamiprid (0,60g i.a./planta), imidacloprid
(0,70g i.a./planta) e tiametoxam (0,75g i.a./planta) são eficazes no controle de P. citri
em plantas de videira com 1 ano de idade.
Os inseticidas neonicotinóides acetamiprid, imidacloprid e tiametoxam nas
doses de 0,60g i.a./planta, 0,70g i.a./planta e 0,75g i.a./planta, respectivamente não
são eficazes para o controle de P. citri em plantas de videira com 15 anos de idade.
Os inseticidas neonicotinóides acetamiprid (6g i.a./100L), imidacloprid (7g
i.a./100L), tiametoxam (7,5g i.a./100L) e os reguladores de crescimento buprofezin
(25g i.a./100L), pyriproxyfen (10g i.a./100L) aplicados via foliar não proporcionam
mortalidade significativa de P. citri na cultura da videira.

71
CONCLUSÕES
Este trabalho apresentou pela primeira vez um inventário da presença de
cochonilhas da família Pseudococcidae na cultura da videira na Região da Serra
Gaúcha, principal pólo produtor de uvas para processamento do Brasil. As
informações disponíveis a respeito da taxonomia, bioecologia e métodos de controle
de pseudococcídeos associados à cultura da videira ainda são escassos no País. Os
estudos devem ser ampliados principalmente visando conhecer a ação da
cochonilha como vetora. Foi demonstrada a presença de cochonilhas da família
Pseudococcidae em 27% das propriedades amostradas. Isto é um fato importante se
for considerado que a amostragem não foi absoluta nos vinhedos. Das espécies
encontradas, Planococcus citri (Risso, 1813) 31,43%, Dysmicoccus sp. 22,86%,
Planococcus minor (Maskell, 1897) 11,43%, Dysmicoccus brevipes (Cockerell, 1893)
5,71%, Pseudococcus sp. 17,14%, Pseudococcus viburni (Signoret, 1875) 8,57% e
Pseudococcus maritimus (Ehrhorn, 1900) 2,86%, as mais freqüentes são
reconhecidamente transmissoras de vírus GLRaV-3, GVA e GVB na cultura.
Entretanto, como as diferenças taxonômicas entre as espécies somente podem ser
observadas com auxílio de especialistas, sugere-se a continuidade deste trabalho
visando elaborar chaves moleculares, para identificação, fato já realizado e de uso
rotineiro em diferentes paises.
Nos estudos de biologia conduzidos com a espécie P. citri, foi possível
observar que a mesma se desenvolve em diferentes variedades produtoras e de
porta-enxertos (Cabernet Sauvignon, Itália; Isabel, 101-14) bem como, sobre
diferentes estruturas vegetativas (raízes e folhas) o que demonstra o potencial de
adaptação da espécie. Outrossim, o não desenvolvimento da espécie em raízes do
porta-enxerto IAC-572 e a reduzida viabilidade em bagas da cultivar Itália, indicam o
potencial de emprego da resistência de plantas para reduzir a infestação da praga

72
e/ou, direcionar o tratamento a fases em que a cochonilha fica mais exposta como é
o caso das folhas.
Com base nas exigências térmicas, verifica-se que a cochonilha pode se
desenvolver nas principais regiões produtoras de uva do Brasil, completando de 5,7
a 11,5 gerações ao ano como é o caso de Bento Gonçalves (RS) e Petrolina (PE),
respectivamente. O grande número de gerações que a cochonilha pode completar
durante o ciclo de desenvolvimento da cultura é mais um fator que aumenta as
possibilidades de dispersão de vírus, reconhecidamente mais eficaz para as ninfas
de primeiro instar. A determinação das exigências térmicas de P. citri em vinhedos
poderá ser utilizada como ferramenta para auxiliar na previsão de picos
populacionais, indicando quando as amostragens devem ser iniciadas ou
intensificadas, e em que momento o controle deve ser implementado. Além disto,
estas informações são importantes para fins de estabelecimento de programas de
controle biológico, onde em determinadas épocas do ano as cochonilhas estão em
estágios de desenvolvimento com maior suscetibilidade ao parasitismo. Com o
conhecimento das exigências térmicas associado à amostragem da população, é
possível prever o melhor período de liberação dos parasitóides. Este tema, não
abordado no presente trabalho, deve ser um dos estudos prioritários para o futuro
visto o sucesso que o controle biológico com parasitóides tem atingido em outros
paises.
Outro fator que deve ser considerado diz respeito as estratégias de manejo do
inseto quando se pensa no mesmo como vetor. P. citri apresenta diversas gerações
ao longo do ano, similar ao observado para P. ficus que é a principal espécie vetora
de vírus nos EUA, África do Sul, Argentina e Uruguai. Neste caso, como a
transmissão de vírus pela espécie diz respeito principalmente as ninfas de primeiro
instar, a presença contínua de indivíduos aptos a transmissão aumentam as
dificuldades de manejo da espécie.
P. citri demonstrou ser sensível aos inseticidas neonicotinóides acetamiprid
(0,60g i.a./planta), imidacloprid (0,70g i.a./planta) e tiametoxam (0,75g i.a./planta) em
plantas de videira com 1 ano de idade. No entanto, em plantas com 15 anos de
idade, estes produtos utilizados nas mesmas doses não foram eficazes para o
controle de P. citri. A mortalidade de P. citri obtida em plantas novas de videira
demonstrou que os inseticidas acetamiprid, imidacloprid e thiametoxam foram
equivalentes entre si. Em plantas adultas, entretanto, os melhores resultados foram

73
obtidos com o imidacloprid, mesmo assim com um máximo de controle de 63%.
Neste caso, evidencia-se que os inseticidas neonicotinóides apresentaram atividade
biológica contra a praga reduzindo a infestação na cultura da videira, porém, ajustes
em relação à dose empregada devem ser feitos de acordo com a idade das plantas.
Quando aplicados via foliar, os inseticidas neonicotinóides acetamiprid (6g
i.a./100L), imidacloprid (7g i.a./100L), tiametoxam (7,5g i.a./100L) e os reguladores
de crescimento buprofezin (25g i.a./100L) e pyriproxyfen (10g i.a./100L) não
proporcionam mortalidade significativa de P. citri na cultura da videira. Comparando-
se as formas de aplicação dos inseticidas neonicotinóides, foi observado que as
aplicações via solo apresentaram maior persistência biológica e foram mais eficazes
em relação pulverização foliar. O tratamento via solo teria como vantagem adicional
um menor efeito sobre os organismos benéficos localizados na parte aérea das
plantas. Além disso, os inseticidas aplicados no subsolo permanecem protegidos da
luz, reduzindo a fotodegradação. No caso da pulverização foliar, embora em outros
países tenham sido observados resultados positivos com os neonicotinóides e
reguladores de crescimento, tais resultados não foram encontrados neste trabalho
com a metodologia empregada. Por isso, sugere-se que em trabalhos futuros, estes
inseticidas sejam empregados em doses maiores e/ou de maneira seqüencial,
permitindo atingir diretamente a população de cochonilhas no vinhedo. Além disso, a
mortalidade da praga deve ser avaliada diretamente no campo associado a redução
no dano nos cachos. Porém, para que isto ocorra, trabalhos visando conhecer a
estrutura etária da população ao longo do ano, visando definir o melhor momento de
controle, devem ser realizados.
Devido ao fato de diversas espécies de pseudococcídeos terem sido
encontradas no inventário, e algumas com capacidade de transmitir vírus nos
vinhedos, recomenda-se futuramente conduzir experimentos, objetivando conhecer a
biologia e a fenologia de ataque na cultura.
Em algumas coletas, foi registrada a presença de formigas junto às
cochonilhas. Em outros países, as formigas principalmente a argentina Linepithema
humile (Mayr, 1868), é uma das principais espécies associadas as cochonilhas
farinhentas a qual se alimenta do “honeydew” liberado pelo inseto. A presença de L.
humile nos vinhedos é considerada um indicativo da presença de populações de
cochonilhas. Por outro lado, a implementação de um programa de manejo das
cochonilhas-farinhentas envolveria também o controle das formigas associadas, pois

74
estas além de auxiliarem na dispersão, também reduzirem o potencial de controle
biológico da praga, sendo assim recomenda-se estudos posteriores visando o
controle de formigas dispersoras.

75
REFERÊNCIAS
ABAKERLI, R. B.; FAY, E. F.; REMBISCHEVSKI, P.; VEKIK, A. M.; GODOY, K.;
MAXIMIANO, A. A.; BONIFÁCIO, A. Regras para nomenclatura dos nomes comuns
dos agrotóxicos. Pesticidas: Revista de Ecotoxicologia e Meio Ambiente,
Curitiba, v.13, p. 29-36, 2003.
ABBINK, J. The biochemistry of imidacloprid. Pflanzenschutz Nachrichten Bayer,
Leverkusen, v.44, n.62, p.183-195, 1991.
ABBOTT, W. S. A method of computing the effectiveness of an insecticide. Journal
of Economic Entomologic, Lanham, v.18, p.265-267, 1925.
AFONSO, A. P. S. Biologia e controle de Parthenolecanium persicae (Fabricius,
1776) (Hemiptera: Coccidae) e transmissão de vírus por P. persicae e
Pseudococcus viburni (Signoret, 1875) (Hemiptera: Pseudococcidae) em
videira. 2005. 70f. Tese (Doutorado). Fruticultura de Clima Temperado. Faculdade
de Agronomia Eliseu Maciel, Universidade Federal de Pelotas, Pelotas.
ARAI, T. Temperature-dependent development rate of the three mealybug species,
Pseudococcus citriculus, Planococcus citri and Planococcus kraunhiae (Homoptera:
Pseudococcidae) on citrus. Japanese Journal of Applied Entomology and
Zoology, v.40, p.25-34, 1996.
ARTIGAS, J. Entomología económica: Insectos de interés agrícola, forestal,
médico y veterinario. Concepción: Universidad de Concepción, 1994. 2v. 943 p.

76
AZEVEDO-FILHO, W. S de; BOTTON, M.; SORIA, S. de J. Curadoria da Coleção
Entomológica da Embrapa Uva e Vinho. Bento Gonçalves: Embrapa Uva e Vinho,
2007. (Embrapa Uva e Vinho. Comunicado Técnico, 77). 10p.
BALACHOWSKY, A. M. Les cochenilles paleartiques de la Tribu Diaspini. Paris:
Inst. Pasteur, 1935. 450p.
BASSOI, L. H.; HOPMANS, J. W.; JORGE, L. A. C.; ALENCAR, C. M. de; SILVA, J.
A. M. Grapevine root distribution in drip and microsprinkler irrigation. Scientia
Agricola, Piracicaba, v.60, n.2, p.377-387, 2003.
BASTOS, C. S.; ALMEIDA, R. P. de.; VIDAL NETO, F. DAS. C.; ARAÚJO. G. P. de.
Ocorrência de Planococcus minor Maskell (Hemiptera: Pseudococcidae) em
Algodoeiro no Nordeste do Brasil. Neotropical Entomology, Londrina, v.36, n.4,
p.625-628, 2007.
BECERRA, V. Resíduos de pesticidas en el cultivo de la vid. Revista de
Información sobre investigación y desarollo agropecuario – idia XXI, Ed.
Instituto Nacional de Tecnologia Agropecuária, p.57-59, 2001.
BEN-DOV, Y. A systematic catalogue of the mealybugs of the world (Insecta:
Homoptera: Coccoidea: Pseudococcidae and Putoidae) with data on
geographical distribution, host plants, biology and economic importance.
Andover: Intercept, 1994. 686 p.
BENTLEY, W. J., ZALOM, F. G., GRANETT, J., SMITH, R. J., VARELA, L. G.;
PURCELL. A. H. University of California IPM Pest Management Guidelines:
Grape, Insects and Mites. Oakland: Univ. Calif., Agricul. Natr. Resc. Publ. 3448.,
2004.
BERGANT, K.; TRDAN, S. How reliable are thermal constants for insect
development when estimated from laboratory experiment? Entomologia
Experimentalis et Applicata, v.120, p.251-256, 2006.

77
BODENTHEIMER, F. S. Citrus entomology in the Middle east Asia. Hoitsema,
1951. p.396-423.
BORBÓN, C. M. de; GRACIA, O.; TALQUENCA, G. S. G. Mealybugs and grapevine
leafroll-Associated virus 3 in Vineyards of Mendoza, Argentina. American Journal
Enology and Viticulture, Davis, v.55, n.3, p.283-285, 2004.
BOTTON, M.; HICKEL, E. R.; SORIA, S. J. Pragas. In: FAJARDO, T. V. M. (Ed). Uva
para processamento: Fitossanidade. Bento Gonçalves: Embrapa Uva e Vinho,
Brasília: Embrapa Informação Tecnológica, 2003a. p. 82-105. (Frutas do Brasil, 35).
BOTTON, M.; AFONSO, A.P.S.; RINGENBERG, R. Manejo de pragas na cultura da
videira. In: SEMINÁRIO INTERESTADUAL DE FRUTICULTURA, 3., 2003b, Palmas.
Anais... Palmas, PR: FACIPAL, 2003b. p. 23-31.
BRASIL. Normais Climatológicas (1961-1990). Ministério da Agricultura e Reforma
Agrária – Secretaria Nacional de Irrigação. Departamento Nacional de Meteorologia
Brasília, 1992. 84p.
BROEKSMA, E.; ROBBERTSE, E.; SABA, F. Field trials with Confidor®
(imidacloprid) for the control of various insect species on citrus in the Republic of
South Africa. Pflanzenschutz Nachrichten Bayer, Leverkusen, v.46, n.64, p.5-31,
1993.
BYRNE, F. J.; TOSCANO, N. C. Uptake and persistence of imidacloprid in
grapevines treated by chemigation. Crop Protection, London, v.25, p.831-834,
2006.
CABALEIRO, C.; SEGURA, A. Some characteristics of the transmission of grapevine
leafroll associated virus 3 by Planococcus citri Risso. European Journal of Plant
Pathology, Dordrecht, v.103, p.373-378, 1997a.

78
CABALEIRO, C.; SEGURA, A. Field transmission of grapevine leafroll associated
virus 3 (GLRaV-3) by the mealybug Planococcus citri. Plant Disease, Saint Paul,
v.81, p.283-287, 1997b.
CAPELLA, A.; GUARNONE, A.; DOMENICHINI, P.; AIROLDI, M. Acetamipride
(EPIK), nuovo insetticida neonicotinoide per la difesa di fruttiferi, orticole, floreali e
ornamentali. Informatore Fitopalogico, Bologna, v.54, n.4, p.43-47, 2004.
CASTILLO, A. A. F.; BLANCO, J. L. M.; ACOSTA, G. O.; CARRILLO, J. L. Control
quimico de piojo harinoso Planococcus ficus Signoret (Homoptera: Pseudococcidae)
en vid de mesa. Agricultura Técnica en México, Hermosillo, v.30, n.1, p.101-105,
2004.
CHARLES, J. G.; COHEN, D.; WALKER, J. T. S.; FORGIE, S. A.; BELL, V. A.;
BREEN, K. C. A Review of Grapevine Leafroll associated Virus type 3 (GLRaV-3) for
the New Zealand wine industry. HortResearch Client Report, Havelock North, n.
18447, 2006. 83p.
CHONG, R. J., OETTING, R. D.; IERSEL, M. W. V. Temperature effects on the
development, survival, and reproduction of the Madeira mealybug Phenacoccus
madeirensis Green (Hemiptera: Pseudococcidae), on Chrysanthemum. Annals of
the Entomological Society of America, v.96, p.539-543, 2003.
CID, M.; PEREIRA, S.; CABALEIRO, C. Presence of grapevine leafroll-associated
virus 3 in primary salivary glands of the mealybug vector Planococcus citri suggests a
circulative transmission mechanism. European Journal of Plant Pathology,
Dordrecht, v.118, p.23-30, 2007.
CIVIDANES, F. J. Uso de graus-dia em entomologia: com particular referência ao
controle de percevejos pragas da soja. Jaboticabal: Funep, 2000. 31p.
CLOYD, R. A. Effect of insect growth regulators on citrus mealybug Planococcus citri
(Homoptera: Pseudococcidae) egg production. HortScience, East Lansig, v.38, n.7,
p. 1397-1399, 2003.

79
COLEN, K. G. F.; SANTA-CECÍLIA, L. V. C.; MORAES, J. C.; REIS, P. R. Efeitos de
diferentes temperaturas sobre a biologia da cochonilha pulverulenta Dysmicoccus
brevipes (Cockerell, 1893) (Hemiptera: Pseudococcidae). Revista Brasileira de
Fruticultura, v.22, p.248-252, 2000.
CORREA, L. R. B.; BONANI, J. P.; SANTA-CECÍLIA, L. V. C.; SOUZA, B. Aspectos
biológicos da cochonilha-branca Planococcus citri (Risso,1813) em Citros. Laranja,
Cordeirópolis, v.26, n.2, p.265-271, 2005.
CORREA, L. R. B. Efeitos de diferentes temperaturas e substratos alimentares
sobre o desenvolvimento da cochonilha-branca Planococcus citri (Risso, 1813)
(Hemiptera: Pseudococcidae). 2006. 33f. Monografia (Bacharelado) –
Universidade Federal de Lavras, Lavras.
CORREA, L. R. B.; SOUZA, B.; SANTA-CECÍLIA, L. V. C.; SOUZA, A. L. V.;
PRADO, E. Desenvolvimento e exigência térmica da fase de ovo de Planococcus
citri (Risso, 1813) (Hemiptera: Pseudococcidae). In: CONGRESSO DE PÓS-
GRADUANDOS DA UFLA, 16., 2007. Lavras. Resumos...Lavras, MG: [s.n.], 2007.
CORREA, L. R. B.; SANTA-CECÍLIA, L. V. C.; SOUZA, B.; CIVIDANES, F.J. Efeito
de diferentes temperaturas e exigências térmicas da cochonilha-branca Planococcus
citri (Risso,1813) (Hemiptera: Pseudococcidae) em Cafeeiro. Arquivos do Instituto
Biológico, São Paulo, v.75, n.1, p.53-58, 2008.
CRUZ, C. D. Programa Genes: aplicativo computacional em genética e estatística.
Viçosa: UFV, 2001. 648 p.
DAANE, K. M.; MALAKAR-KUENEN, R.; WALTON, V. M. Temperature development
of Anagyrus pseudococci (Hymenoptera: Encyrtidae) as a parasitoid of the vine
mealybug, Planococcus ficus (Homoptera: Pseudococcidae). Biological Control,
v.31, p.123-132, 2004.
DAANE, K. M.; BENTLEY, W. J.; WALTON, V. M.; MALAKAR-KUENEN, R.;
YOKOTA, G. Y.; MILLAR, J. G.; INGELS, C. A.; WEBER, E. A.; GISPERT, C.

80
Sustainable controls sought for the invasive vine mealybug. California Agriculture,
Berkeley, v.60, p.31-38, 2006.
DAANE, K. M.; SIME, K. R.; FALLON, J.; COOPER, M. L. Impacts of Argentine ants
on mealybugs and their natural enemies in California's coastal vineyards. Ecological
Entomology, v.32, p.583-596, 2007.
DEAN, H. A.; HART, W. G.; INGLE, S. Pest management considerations of the effect
of pesticides on Texas citrus pest and certain parasites. Journal Rio Grande Valley
Horticulture Society, Weslaco, v.31, p.37-47, 1977
.
DE JONG, P. W.; ALPHEN, J. J. M. Host stage selection and sex allocation in
Leptomastix dactylopii, a parasitoid of Planococcus citri. Entomologia
Experimentalis et Applicata, n.50, p.161-169, 1989.
DEMONTIS, M. A., ORTU, S.; COCCO, A.; LENTINI, A.; MIGHELI, Q. Diagnostic
markers for Planococcus ficus (Signoret) and Planococcus citri (Risso) by random
amplification of polymorphic DNA-polymerase chain reaction and species-specific
mitochondrial DNA primers. Journal of Applied Entomology, v.131, p.59-64, 2007.
DIESTRE, F.; SAZO, R. L. Control químico de Pseudococcus viburni (Homoptera:
Pseudococcidae). Ciencia e Investigación Agraria, Santiago, v.17, n.3, p.107-110,
1990.
DOUGLAS, N.; KRÜGER, K. Grapevine leafroll-associated virus 3 transmission
efficiency of Planococcus ficus and Pseudococcus longispinus (Hemiptera:
Pseudoccidae). In: 15th MEETING OF THE COUNCIL FOR THE STUDY OF VIRUS
AND VIRUS LIKE DISEASES OF THE GRAPEVINE (ICVG). 15., 2006,
Stellenbosch, South Africa. Extended Abstracts... Stellenbosch, 2006. p.191-192.
DOUGLAS, N.; KRÜGER, K. Transmission efficiency of Grapevine leafroll-associated
virus 3 (GLRaV-3) by de mealybugs Planococcus ficus and Pseudococcus
longispinus (Hemiptera: Pseudoccidae). European Journal of Plant Pathology.
Disponível em <http://www. springerlink.com/content/ g5412282

81
23up8085>. Acesso em: 5 mar. 2008.
ELBERT, A.; BECKER, B.; HARTWIG, J.; ERDELEN, C. Imidacloprid – a new
systemic insecticide. Pflanzenschutz Nachrichten Bayer, Leverkusen, v.44, n.62,
p. 113-136, 1991.
EMATER/RS. Recomendações para o manejo das doenças fúngicas e insetos
pragas da videira. Porto Alegre: EMATER/RS; Bento Gonçalves: Embrapa Uva e
Vinho, 2003. 72 p.
EMBRAPA UVA E VINHO. Pragas da Videira. Disponível em:
<http://sistemasdeproducao.cnptia.embrapa.br/FontesHTML/Uva/UvasViniferasRegi
õesClimaTemperadol>. Acesso em: 20 dez. 2007.
ENGELBRECHT, D. J.; KASDORF, G. G. F. Transmission of grapevine leafroll
disease and associated closteroviruses by the vine mealybug Planococcus ficus.
Phytophylactica, Pretoria, v.22, p.341-346, 1990.
ENTWISTLE, P. F. Pests of Cocoa. London: Longman, 1972.p. 132-53.
ERAZO, F .H. E. Aportes al conocimiento de la biología de la Cochinilla
Harinosa de la Vid, evaluación de la eficacia de pesticidas aplicados en
primavera para su control y determinación de curvas de degradación en uva de
mesa (Vitis vinifera L.) CV. Superior (San Juan-Argentina). 2004. 96 f.
Dissertação (Mestrado). Maestria de Viticultura y Enologia. Mendoza, Argentina.
ETCHEBARNE, F., STRAFILE, D.; BECERRA, V. Resultados preliminaries de
estudios sobre cochinilla nharinosa de la vid. Centro de Estúdios de Fitofarmacia,
Estácion Experimental Agropecuária Mendoza INTA. Revista el Vino & Su
Industria, Mendoza, n.12, p.27-35, 2003.
FAJARDO, T. V. M.; KUHN, G. B.; NICKEL, O. Doenças Virais. In: FAJARDO,
T.V.M. (Ed.). Uva para processamento: Fitossanidade. Brasília: Embrapa
Informação Tecnológica, 2003. p.45-62. (Frutas do Brasil, 35).

82
FLAHERTY, D. L.; PHILIPS, P. A.; LEGNER, E.F. PEACOCK, W.L.; BENTLEY, W. J.
Mealybugs. IN: FLAHERTY, D. L.; CHRISTENSEN, L. P.; LANINI, W. T.; MAROIS,
J.J.; PHILLIPS, P. A.; WILSON, L. T. (EDS.) Grape Pest Management. University
of California, Division of Agricultural and Natural Resources, Publication 3343,
Oakaland, CA, 1992, p.159-165.
FOLDI, I.; SORIA, S. J. Les cochonilles nuisibles a la vigne em Amérique du Sud
(Homoptera: Coccoidea). Annales de la Societè Entomologique de France, Paris,
v.24, n.25, p.411-430, 1989.
FOLDI, I.; KOZAR, F. New species of Cataenococcus and Puto from Brazil and
Venezuela, with data on others species (Hemiptera : Coccidea). Nouvelle Revue
d´Entomologie, n. 22, n.4, p. 305-312, 2006.
FU, C. A. A.; OSORIO, G. A.; MÁRQUEZ, C. A.; MIRANDA, B. J. L.; GRAGEDA, J.;
MARTÍNEZ, D. G. Manejo integrado del piojo harinoso de la vid. Instituto
Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Centro de
Investigación Regional del Noroeste, Campo Experimental Costa Hermosillo.
Hermosillo, Sonora, México. 16p. (Folleto Técnico, n.25), 2002.
FURTADO, D. Sisvar versão 4.3 (Build 4.5). Lavras: Universidade Federal de
Lavras, 2007. Disponível em: <http://www.ufl.br/>. Acesso em: 10 dez. 2007.
GARAU, R.; PROTA, V. A.; BOSCIA, D.; FIORI, M.; PROTA, U. Pseudococcus
affinis Mask., new vector of grapevine trichoviruses A and B. Vitis, Siebeldingen,
v.34, n.1, p.67-68, 1995.
GEIGER, C. A.; DAANE, K. M. Seasonal movement and sampling of the grape
mealybug, Pseudococcus maritimus (Ehrhorn) (Homoptera: Pseudococcidae) in San
Joaquin Valley vineyards. Journal of Economic Entomology, Lanham, v.94, p.291-
301, 2001.

83
GODINHO, M. A.; FRANCO, J. C. Survey on the pest status of mealybugs in
Portuguese vineyards. Integrated Control in Viticulture IOBC WPRS Bulletin,
v.24, n.7, p.221-225. 2001.
GOLINO, D. A.; SIM, S. T.; GILL, R.; ROWHANI, A. California mealybugs can spread
grapevine leafroll disease. California Agriculture, Bekerley, n.56, p.196-201, 2002.
GRANARA DE WILLINK, M. C. Identificacion de cochinillas (mensagem pessoal).
Mensagem recebida por [email protected] em 20 ago. 2007.
GRAVENA, S. Cochonilha Branca: descontrolada em 2001. Laranja, Cordeirópolis,
v.24, n.1, p.71-82, 2003.
GONZÁLEZ, R. H. Manejo de plagas de la vid. Santiago: Universidad de Chile-
Facultad de Ciencias Agrarias, Veterinarias e Florestales, 1983. 132p.
GONZÁLEZ, R. Degradación de residuos de plaguicidas en huertos frutales en
Chile. Universidad de Chile. Santiago de Chile: Serie Ciências Agronômicas, n.4,
2002. 163 p.
GONZÁLEZ, R. H. Chanchitos blancos de importancia agrícola y cuarentenaria en
huertos frutales de Chile. Revista Frutícola, Santiago, v.24, n.1, p.5-17, 2003.
GONZÁLEZ, R. H.; CURKOVIC, T. S.; BARRÍA, G. P. Control de Pseudococcus
affinis (Maskell) (Homoptera: Pseudococcidae) con diazinon, metidation e profenofos
en postcosecha de vides y ciruelos. Agricultura Técnica, Santiago, v.55, n.2, p.95-
98, 1995.
GONZÁLEZ, R. H.; CURKOVIC, T. S; BARRIA, G. Evaluacion de eficacia de
insecticidas sobre Chanchitos Blancos en Ciruelos y uva de mesa. Revista
Frutícola, Santiago, v.17, n.2, p.45-57, 1996.

84
GONZÁLEZ, R. H.; POBLETE, J.; BARRIA, G. El chanchito blanco de los frutales en
Chile, Pseudococcus viburni (Signoret, 1875) (Homoptera: Pseudococcidae).
Revista Frutícola, Santiago, v.22, n.1, p.17-26, 2001.
GONZÁLEZ, R. H.; VOLOSKY, C. Chanchitos blancos y Polillas de la fruta: problema
cuarentenarios de la fruta de exportación. Revista Frutícola, Santiago, v.25, n.2,
p.41-62, 2004.
GONZÁLEZ, R. H.; VOLOSKY, C. Desarrolo estacional y estrategias de manejo de
chanchitos blancos, Pseudococcus spp., (Hemiptera: Pseudococcidae) en
pomáceas, uva de mesa y vid vinífera. Revista Frutícola, Santiago, v.27, n.2, p.37-
47, 2006.
GUBLER, D. J.; STAPLETON, J.; LEAVITT, G.; PURCELL, A.; VARELA, L.; SMITH,
R. J. UC IPM pest management guidelines: grapes. Califórnia, USA. p.67-72,
1999.
GULLAN, P. J. Identificacion of the imature instars of mealybugs (Hemiptera:
Pseudococcidae) found on citrus in Austrália. Australian Journal of Entomology,
Victoria, v.39, n.3, p.160-166, 2000.
HADDAD, M. L., PARRA, J. R. P.; MORAES, R.C.B. Métodos para estimar os
limites térmicos inferior e superior de desenvolvimento de insetos. Piracicaba:
FEALQ, 1999. 29p.
HAJI, F. N. P.; ALENCAR, J. A.; BARBOSA, F. R. Pragas, 82-89. In: Leão, P. C. S. de
M. (Ed.). Uva de Mesa: produção. Brasília, Embrapa Informação Tecnológica, 2001.
128p.
HICKEL, E. R. Pragas da videira e seu controle no estado de Santa Catarina.
Florianópolis: EPAGRI, 1996. 52p.
HONEK, A. Geographical variation in thermal requirements for insect development.
European Journal of Entomology, v.93, p.303-312, 1996.

85
ISLAM, K. S.; COPLAND, M. J. W.; JAHAN, M. Laboratory studies on the
development of Planococcus citri (Risso) and Pseudococcus affinis (Mask.) at
different temperatures. Bangladesh Journal of Entomology, v.5, p.19-24, 1995.
ISLAM, K. S.; COPLAND, M. J. W. Influence of egg load and oviposition time interval
on the host discrimination and offspring survival of Anagyrus pseudococci
(Hymenoptera: Encyrtidae), a solitary endoparasitoid of citrus mealybug,
Planococcus citri (Hemiptera: Pseudococcidae). Bulletin of Entomological
Research, v.90, p.69-75, 2000.
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA - IBGE. Sistema IBGE
de recuperação automática. Disponível em:
<http://www.sidra.ibge.gov.br/bda/agric/default.asp>. Acesso em: 10 dez. 2007.
KISHINO, A. Y.; CARVALHO, S. L. C. de.; ROBERTO, S. R. Viticultura tropical o
sistema de produção do Paraná. Londrina: IAPAR, 2007. 366p.
KUHN, G. B.; NICKEL, O. Viroses e sua importância na viticultura brasileira. Informe
Agropecuário, Belo Horizonte, v.19, n.194, p.85-91, 1998.
KUHN, G B; FAJARDO, T. V. M. Doenças causadas por vírus. In: NACHTIGAL, J.
C.; CAMARGO, U. A.; CONCEIÇÃO, M. A. F. (Eds.) Uvas sem sementes -
cultivares BRS Morena, BRS Clara e BRS Linda. Bento Gonçalves: Embrapa Uva e
Vinho, 2004. p.67-76. (Sistemas de Produção)
KUNIYUKI, H.; REZENDE, J. A. M.; WILLINK, M. C. G.; NOVO, J. P. S.; YUKI, V. A.
Transmissão do “Grapevine leafroll-associated” vírus 3 pela cochonilha
Pseudococcus longispinus Targioni-Tozetti (Hemiptera: Pseudococcidae). Summa
Phytopathologica, Botucatu, v.31, n.1, p.65-68, 2005.
KUNIYUKI, H.; GIORIA, R.; REZENDE, J. A. M.; WILLINK, M. C. G.; NOVO, J. P. S.;
YUKI, V. A. Transmissão experimental do Grapevine virus B pela cochonilha
Pseudococcus longispinus Targioni-Tozzetti (Hemiptera: Pseudococcidae). Summa
Phytopathologica, Botucatu, v.32, n.2, p.151-155, 2006.

86
LAFLIN, H. M.; PARRELLA, M. P. Developmental biology of citrus mealybug under
conditions typical of Califórnia Rose production. Annals of the Entomological
Society of America, v.97, p.982-988, 2004.
LARRIAN, P. Efecto de la quimigación y el pintado con el imidacloprid (Confidor)
sobre la población de Pseudococcus viburni (Hemiptera: Pseudococcidae) en vides
de mesa. Agricultura Técnica, Santiago, v.59, n.1, p.13-25, 1999.
LEATHER, S. R., WALTERS, K. F. A.; BALE, J. S. The ecology of insect
overwintering. Cambridge: Cambridge University Press, 1993. 255p.
LEPAGE, H. S. Abóboras, cobaias para o estudo das pragas dos vegetais. O
Biológico, São Paulo, v.8, n.9, p.221-224, 1942.
LIT JR., I. L.; LIT, C. M.; CALILUNG, V. J. The mealybugs (Pseudococcidae,
Coccoidea, Hemiptera) in the Philippines. Philippine Entomologist, v.12, p.29-41,
1998.
LOAEZA, R. M. G. Contribucion al conocimiento de los Pseudococcidae de
México (Homoptera: Coccoidea). México: Instituto Politecnico Nacional – Escuela
Nal de Ciencias Biologicas, 1958. p.11-185. (Tesis Profesional).
MALLESHAIAH, B.; RAJAGOPAL, K.; GOWDA, K. N. M. Biology of citrus mealybug,
Planococcus citri (Risso) (Hemiptera: Pseudococcidae). Crop Research, Hisar, v.20,
n.1, p.130-133, 2000.
MARTINEZ-FERRER, M. T.; GARCIA-MARI, F.; RIPOLLES-MOLES, J.L. Population
dynamics of Planococcus citri (Risso) (Homopetra: Pseudococcidae) in citrus groves
in Spain. Integr. Contr. Fruit Crop, v.26, p.149-161, 2003.
MARTIN-MATEO, M. P. Inventario preliminar de los cóccidos de España. III.
Pseudococcidae, Ortheziidae y Margarodidae. Revista de Entomólogos Ibéricos,
Madrid, v.41, p.89-104, 1985.

87
MAROTTA, S. An annotated list of the Italian mealybugs. Bollettino del Laboratorio
di Entomologia Agraria 'Filippo Silvestri', Portici, v.43, suplemento, p.107-116,
1987.
MAYNE, W. Biology of Pseudococcus citri (Risso) on coffee. Annual Report of
Coffee Scientific Officer, p.32, 1932.
MCKENZIE, H. L. In: Mealybugs of California with taxonomy, biology, and
control of North American species (Homoptera: Coccoidea: Pseudococcidae).
Berkeley: Univ. Calif. Press, 1967. 526p.
MENDEL, Z.; BLUMBERG, D.; ISHAAYA, I. Effect of buprofezin on Icerya purchasi
and Planococcus citri. Phytoparasitica, Bet Dagam, v.19, n.2, p.103-112, 1991.
MORANDI FILHO, W. J.; BOTTON, M.; GRÜTZMACHER, A. D.; FAJARDO, T. V.
M.; PRADO, E. Vetor Encoberto, cochonilhas algodonosas em videira. Revista
Cultivar Hortaliças e Frutas, Pelotas, p.28-29, 2007.
MUTHUKRISHNAN, N.; MANOHARAN, T.; THEVAN, P. S. T.; ANBU, S. Evaluation
of buprofezin for the management of grape mealybug, Maconellicoccus hirsutus
(Green). Journal of Entomological Research, London, v.29, n.4, p.339-344, 2005.
NACHTIGAL, J. C. Produção de mudas de videiras em Regiões Tropicais e
Subtropicais. Bento Gonçalves: Embrapa Uva e Vinho, 2003. (Embrapa Uva e
Vinho. Circular Técnica, 46).
OROZCO, O. L.; ABELLA, F.; PINZÓN, C. Imidacloprid, herramienta de la química
del futuro dentro de un manejo integrado de la mosca blanca de los invernaderos.
Revista Colombiana de Entomologia, Bogotá, v.21, n.2, p.87-90, 1995.
PALUMBO, J. C.; KERNS, D. L.; ENGLE, C. E.; SANCHEZ, C. A.; WILCOX, M.
Imidacloprid formulation and soil placement effects on colonization by sweetpotato

88
whitefly (Homoptera: Aleyrodidae): head size and incidence of chlorosis in lettuce.
Horticultural Entomology, Tallahassee, v.89, n.3, p.735-742, 1996.
PARRA, J. R. P. Técnicas de criação de Anagasta kuehniella hospedeira alternativa
para produção de Trichogramma spp. In: PARRA, J. R. P.; ZUCCHI, R. A. (Org.).
Curso de controle biológico com Trichogramma. Piracicaba: ESALQ/USP, 1996.
p. 27-37.
PARRELA, M. P. The development and implementation of integrated pest
management strategies in floriculture. Davis: University of California, 1995.
Disponível em: <http://www.endowment.org/pr/p0594pr2.html>. Acesso em: 10 de
dez. 2007.
PETERSEN, C. L.; CHARLES, J. G. Transmission of grapevine leafroll-associated
closteroviruses by Pseudococcus longispinus and P. calceolariae. Plant Pathology,
Osney Mead, n.46, p.509-515, 1997.
PIETERSEN, G. Spread of grapevine leafroll disease in South África – a difficult,
but not insurmountable problem. Wynboer, Suider Paarl, 22 mar. 2008. Disponível
em: <http://www.wynboer.co.za/recentarticles/0406leaf.php3> Acesso em: 22 mar.
2008.
PROTAS, J. F. da S.; CAMARGO, U. A.; MELO, L. M. R. de. A vitivinicultura
brasileira: realidade e perspectivas. In: SIMPÓSIO MINEIRO DE VITICULTURA E
ENOLOGIA, 1., 2002, Andradas, MG. Viticultura e Enologia: atualizando conceitos.
Caldas, MG: Epamig, 2002. p.17-32.
RAMAKRISHNAN, R.; SUITER D. R.; NAKATSU C. H.; BENNETT, G. W. Feeding
inhibition and mortality in Reticulitermes flavipes (Isoptera: Rhinotermitidae) after
exposure to imidacloprid-treated soils. Journal of Economic Entomology, Lanham,
v.93, p.422-428, 2000.
RAO, C. N.; SHINVANKAR, V. J.; SINGH, S. Citrus mealybug (Planococcus citri)
(Risso) management – a review. Agricultural Reviews, v.27, n.2. p.142-146, 2006.

89
REDDY, K. B.; SEETHARAMA, H. G. Integrated management of mealybugs in
coffee. Indian Coffee. v.61, p.26-28. 1997.
REDDY, K. B.; BHAT, P. K.; NAIDU, R. Suppression of mealybugs and green scale
infesting coffee with natural enemies in Karnataka. Pest Manag. Econ. Zool. v.5, p.
119-121, 1999.
REIS, P. R.; MELO, L. A. S. Pragas da Videira. Informe Agropecuário, v.10, n.117,
p.68-72, 1984.
REIS, P. R.; SOUZA, J. C.; GONÇALVES, N. P. Pragas da Videira Tropical. Informe
Agropecuário, v.19, n.194, p.92-95, 1998.
RIPA, R.; RODRIGUEZ, F. Plagas de citros, sus enemigos naturales y manejo.
Santiago: Instituto de Investigaciones Agropecuárias, 1999. 151p. (INI, Libros, 3).
ROSCIGLIONE, B.; CASTELLANO, M. A. Further evidence that mealybugs can
transmit Grapevine virus A (GVA) to herbaceous hosts. Phytopathology
Mediterranean, Bologna, v.24, p.186-188, 1985.
SALAZAR, D. Melazos I. Unidad de Viticultura, Dep. Producción Vegetal,
Universidad Pontificia de Valencia, Valencia, 1996, 2623.
SANTA-CECÍLIA, L. V. C.; REIS, P. R.; SOUZA, J. C. Sobre a nomenclatura das
espécies de cochonilhas farinhentas do cafeeiro nos estados de Minas Gerais e
Espírito Santo. Neotropical Entomology, Londrina, v.31, n.2., p.333-334, 2002.
SANTA-CECÍLIA, L. V. C.; SOUZA, B.; SOUZA, J. C. de.; PRADO, E.; MOINO
JUNIOR, A.; FORNAZIER, M. J.; CARVALHO, G. A. Cochonilhas-farinhentas em
cafeeiros: bioecologia, danos e métodos de controle. Boletim Técnico, Belo
Horizonte, n.79, 48p, 2007.
SAS INSTITUTE. SAS/STAR® User's Guide, Version 8. Cary: SAS Institute, 1998.

90
SETHER, D. M.; ULLMAN, D. E.; HU, J. S. Transmission of pineapple mealybug
wilt-associated vírus by two of mealybug (Dysmicoccus spp.) Phytopathology, v.88,
p.1224-1230, 1998.
SFORZA, R., KOMAR, V.; GREIF, C. New scale insect vectors of grapevine
closteviruses. In: PROCEEDINGS OF THE 13TH INTERNATIONAL CONGRESS OF
VIRUS & VIRUS LIKE DISEASES OF GRAPEVINE. Adelaide, Australia. 12th-17th
2000, p.14-15.
SFORZA, R.; BOUDON-PADIEU, E.; GREIF, C. New mealybug species vectoring
grapevine leafroll associated viruses-1 and -3 (GLRaV-1 and -3). European Journal
of Plant Pathology, Dordrecht, n.109, p.975-981, 2003.
SILVA, A. G. A.; GONÇALVES, C. R.; GALVÃO, D. M.; GONÇALVES, A. J. L.;
GOMES, J.; SILVA, M. do N.; SIMONI, L. de. Quarto catálogo dos insetos que
vivem nas plantas do Brasil: seus parasitas e predadores. Rio de Janeiro:
Ministério da Agricultura, 1968. 622p.
SILVEIRA NETO, S.; NAKANO, O.; BARBIN, D.; VILLA NOVA, N.A. Manual de
ecologia dos insetos. São Paulo: Agronômica Ceres, 1976. 419 p.
SOUTHERLAND, J. R. G. Some observations on the common mealybug,
Pseudococcus citri (Risso). 23-24th. Annual Report of the Quebec Society for the
Protection of Plants. 1930-32. Canada, 105-18. 1972.
STRONG, D. R.; LAWTON, J.; SOUTHWOOD, R. Insects on the plants.
Cambridge: Harvard University Press, 1984. 313p.
TEJERO, J.C. Pseudococcus citri (Risso), en la zona de Jerez, 1990. Phytoma,
Valencia, n.27, p.45-51, 1991.

91
TRURDGILL, D. L. Why do tropical poikilothermic organisms tend to have higher
threshold temperature for development than temperate ones? Functional Ecology,
v.9, p.136-137, 1995.
TRURDGILL, D. L.; PERRY, J. N. Thermal time and ecological strategies – a unifying
hypothesis. Annals of Applied Biology, v.125, p.521-532, 1994.
TRURDGILL, D. L.; HONEK, A. D. L. I.; VAN STRAALEN, N. M. Thermal time –
concepts and utility. Annals of Applied Biology, v.146, p.1-14, 2005.
UYGUN, N.; SENGONCA, C.; ERKILIC, L.; SCHADE, M. The Coccoidea fauna and
their host plants in cultivated and non-cultivated areas in the East Mediterranean
region of Turkey. Acta Phytopathologica Academiae Scientiarum Hungaricae,
v.33, n.1-2, p.183-191, 1998.
VENNETE, R. C.; DAVIS, E. E. Mini risk assessment passionvine mealybug:
Planococcus minor (Maskell) Pseudococcidae: Hemiptera. Saint Paul: University of
Minnesota, 2004. 30p.
WALTON, V. M.; PRINGLE, K. L. Effect of pesticides used on table grapes on the
mealybug parasitod Coccidoxenoides peregrinus (Timberlake) (Hymenoptera:
Encyrtidae). South African Journal of Viticulture and Enology, v.25, p.23-25,
1999.
WALTON, V. M.; PRINGLE, K. L. Developmental biology of vine mealybug,
Planococcus ficus (Signoret) (Homoptera: Pseudococcidae), and its parasitoid
Coccidoxenoides perminutus (Timberlake) (Hymenoptera: Encyrtidae). African
Entomology, v.13, p.143-147, 2005.
WILD, A. L. Taxonomic Review of the ant genus Linepithema (Hymenoptera:
Formicidae). University of California, v.126, 2007. 162p.

92
WILLIAMS, D. J.; WATSON, G. W. In: The Scale Insects of the Tropical South Pacific
Region. Pt. 2: The Mealybugs (Pseudococcidae). London: CAB International Institute
of Entomology, 1988. 260 p.
WILLIAMS, D. J.; WILLINK, M. C. G. Mealybugs of Central and South America.
Wallingford: CAB International, 1992. 634 p.
WILLIAMS, D. J.; MOGHADDAM, M. Mealybug species of the genus Planococcus
Ferris in Iran (Homoptera: Coccoidea: Pseudococcidae) with a discussion of
Planococcus vovae (Nasonov). Journal of Entomological Society of Iran, v.1, n.1-
2, p.32-43, 2000.
WILLIAMS, D. J. In: Mealybugs of Southern Asia. The Natural History Museum,
Kuala Lumpur: Southdene SDN. BHD. 896 pp. 2004.
WILLINK, M. C. G; SCATONI, I. B. ; TERRA, A. L.; FRIONI, M. I. Cochinillas
harinosas (Homoptera, Pseudococcidae) que afectan plantas cultivadas y silvestres
en Uruguay. Agrociencia, Montevidéo, v.1, n.1, p.96-100, 1997.
WOERNER, R. P. Estudo sistematico dos Pseudococcideos (Hom.,
Pseudococcidae) no Rio Grande do Sul, Brasil. 1983. 128f. Dissertação
(Mestrado em Biociências - Zoologia) - Pontifícia Universidade Católica do Rio
Grande do Sul, Porto Alegre.





























