Embed Size (px)
Citation preview

MINISTÉRIO DA EDUCAÇÃO
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE CIÊNCIAS DA SAÚDE
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS DA SAÚDE
EFEITO IMUNOMODULADOR E ATIVIDADE ANTIMICROBIANA DE HETEROFUCANAS DE Sargassum filipendula
CINTHIA BEATRICE DA SILVA TELLES
NATAL/RN 2015

CINTHIA BEATRICE DA SILVA TELLES
EFEITO IMUNOMODULADOR E ATIVIDADE ANTIMICROBIANA DE HETEROFUCANAS DE Sargassum filipendula
Tese apresentada ao Programa de Pós-
Graduação em Ciências da Saúde da
Universidade Federal do Rio Grande do
Norte como requisito para a obtenção do
título de Doutor em Ciências da Saúde.
Orientador: Prof. Dr. Hugo Alexandre de O. Rocha
NATAL/RN 2015

ii

iii
MINISTÉRIO DA EDUCAÇÃO
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE CIÊNCIAS DA SAÚDE
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS DA SAÚDE
Coordenador do Programa de Pós-Graduação em Ciências da Saúde:
Prof. Dr. Eryvaldo Sócrates Tabosa do Egito

iv
CINTHIA BEATRICE DA SILVA TELLES
EFEITO IMUNOMODULADOR E ATIVIDADE ANTIMICROBIANA DE HETEROFUCANAS DE Sargassum filipendula
Aprovada em: 26/06/ 2015
Banca Examinadora:
Presidente da Banca:
Prof. Dr. Hugo Alexandre de Oliveira Rocha (UFRN)
Membros da Banca
Prof. José Veríssimo Fernandes
Prof. Valter Ferreira de Andrade Neto
Prof. Toshiyuki Nagashima Junior
Prof. Danilo José Ayres de Menezes

v
Dedico esta obra
A Deus.
Pela minha existência e por me oferecer condições para realizar essa conquista.
Ao meu filho, Davi.
Você é minha VIDA, tem me ensinado a cada dia o que é o amor incondicional, a
minha maior alegria é a sua felicidade. É o seu sorriso, a sua alegria de viver, seus
beijos e abraços apertados que me proporcionam força para continuar nessa
caminhada. Te amo muito.
Ao meu marido, Jean.
Não sei se somos almas gêmeas, afinal temos tantas diferenças, mas tenho a certeza
que nos completamos… você é a alegria que falta nas minhas manhãs, é a
sensibilidade dos meus dias… Obrigada por estar sempre ao meu lado.

vi
Dedico esta obra
Aos meus pais, Ana e Carlos.
Por serem exemplo de amor incondicional, presentes em todos os momentos da
minha vida, com certeza essa conquista foi construída graças a todo amor, apoio e
incentivo dedicado a mim durante todos esses anos. Espero poder retribuir todo o
investimento, e ainda proporcionar muito orgulho.
Ao meu irmão, Agusto.
Exemplo de homem… hoje tenho certeza que o que sou devo muito a você, meu
irmão mais novo, mas que sempre foi o mais maduro de nós dois… Agradeço o seu
amor, carinho, compreensão, apesar de não usarmos muito as palavras para
expressar todo o nosso amor, nos amamos muito.
A Hugo Rocha,
Amigo e orientador, exemplo de amor pela profissão… obrigada pela paciência, pelos
ensinamentos, pelos momentos de descontração e principalmente por estar ao meu
lado na conclusão desse trabalho, a caminhada foi longa, mas, com certeza, só
consegui realizar com o seu incentivo e confiança.

vii
Agradecimentos especiais
Ao meu marido. Jean Gouveia, meu amigo e companheiro. Obrigada por dividir a sua
vida comigo, que nosso amor supere todos os obstáculos.
Ao meu filho Davi, que Deus abençoe seus caminhos, e lhe dê muita saúde e
sabedoria, estarei sempre ao seu lado, lutando pela sua felicidade. Te amo muito.
À minha família, em especial, meus pais Ana Lúcia e Carlos Roberto e ao meu irmão Augusto César que me incentivaram em todos os momentos;
Ao amigo e orientador, Hugo Rocha, por estar ao meu lado nessa caminhada.
À UFRN, à Pós-graduação em Ciências da Saúde e ao Departamento de Bioquímica pela
oportunidade de concluir esse curso de Pós-graduação, assim como as agências Financiadoras CAPES e CNPq.
A todos os professores do CCS (UFRN) e do DBQ (UFRN), aos coordenadores do
programa de pós-graduação (PPGCSa) e às secretárias do programa.
Aos membros da banca de qualificação: Profa. Luciana Guimarães, que desde a qualificação do mestrado ofereceu valiosas sugestões para melhorar a minha pesquisa e a
e a Jailma Almeida uma profissional exemplar e sempre dedicada.
Aos meus amigos de laboratório pela colaboração e ajuda nos meus experimentos e pelos
momentos de descontração:
A Jadrilma minha amiga, madrinha, anjo… obrigada por fazer parte da minha vida
profissional e pessoal, devo muito da minha “transformação” ao seu carinho e presença na minha vida e na vida do meu filho… obrigada por tudo.
A Dayn exemplo de perseverança, mãe, mulher e profissional exemplar. Desejo muito que
o seu futuro profissional seja brilhante, tenho um grande carinho por você.
A Ruth e a Rafael, amigos que admiro muito, a Leandro (ou será rosa) hoje faz parte da família, você é um exemplo de profissional.
A Mariana ou Santana, amiga do lab, da turma do mestrado, hoje amiga para a vida, você
é um exemplo de dedicação, de amor pelo que faz obrigada pela ajuda sempre que precisei te adoro!
Agradeço muito à família BIOPOL, todos vocês são essenciais nessa trajetória: Gabriel,
Moacir, Joanna, Pablo, Raniere, Ivan, Leonardo Nobre (Leo), Letícia, Arthur, Vinicius, Max, Rony, Marília, Monique, Larisse, Mônica, Danielle, Almino Afonso, Profa.
Fabiana Lima, Ana Karina, Sara.

viii
Ao Prof. Marcelo Silva e a amiga Sônia Pestana do Instituto de Higiene e Medicina Tropical, Universidade Nova de Lisboa que durante os seis meses vividos em Lisboa,
Portugal, compartilharam comigo seus conhecimentos e proporcionaram meses maravilhosos;
A Profa. Tiana Tasca e todas as alunas da Faculdade de Farmácia da Universidade
Federal do Rio Grande do Sul, em especial a aluna Amanda Picolli, pelo acolhimento e
disposição na realização dos ensaios.
Agradeço a Carol pela ajuda na realização dos experimentos com Leishmania e todos do
Instituto de Medicina Tropical da UFRN que ajudaram na realização desse trabalho.
Agradeço, de forma especial, àqueles que contribuíram de forma diferencial para minha formação como pessoa, os meus amigos de SEMPRE: Tathiana, Raiane, Gabriela, Ana Paula, Isabel, Deborah, Nilmara e Isabelli. Sem o apoio de vocês tudo seria mais difícil
Agradeço aos meus sogros: Benigna e Gouveia. Sem a dedicação incondicional de vocês a criação do meu filho, nosso bebe, a realização dessa tese seria bem mais difícil. Muito
obrigada pela ajuda e pelo carinho. Também dedico esta tese a vocês!
Agradeço a todos os amigos que trabalham comigo na EAJ: Lígia, Welson, Ângela, André, Afrânio, Rose, etc. vocês são exemplos de profissionais, tenho aprendido muito
ao lado de vocês. E, em especial, agradeço as substitutas que são insubstituíveis, Maiara e Louize, uma das conquistas durante esse período na EAJ com certeza foi a amizade de
vocês.
Agradeço aos professores e amigos do IFRN campus Currais Novos, em especial, a
Andreilson, Itála, Jane, Jana, Dayana, Maura, Mayara, Lívia, Jonas, Marcelo, Danúbia,
Cris, dentre tantos outros que mesmo com uma palavra de incentivo me deram força para
a conclusão desse trabalho.
Obrigada a todos pelo apoio, pelo carinho, pelo consolo nas frustrações. Agradeço demais
a todos que contribuíram de forma direta ou indireta para a realização desse trabalho.

ix
“A vida é um sonho, torne-o real”.
(Madre Teresa)

x
RESUMO
Macroalgas marinhas constituem uma fonte extremamente rica de compostos
bioativos, dentre eles, polissacarídeos sulfatados. Sargassum filipendula (SF),
alga pertencente à ordem Phaeophycea, é fonte de heterofucanas conhecidas
pela capacidade de modular uma série de funções biológicas. Considerando a
necessidade de encontrar fármacos mais eficientes no combate a infecções
microbianas, as heterofucanas de SF foram avaliadas como agentes
imunomoduladores e antimicrobianos. As heterofucanas SF0.5V, SF0.7V e
SF1.0V apresentaram uma forte atividade imunomoduladora intensificando a
liberação de óxido nítrico (NO) por macrófagos murinos (RAW 264.7), bem
como, por macrófagos originados de monócitos primários humanos. Além
disso, quando macrófagos humanos foram infectados por Leishmania infantum
e tratados com SF0.5V, SF0.7V e SF1.0V ocorreu um aumento significativo na
liberação extracelular de NO. Como a citotoxicidadede de macrofágos contra a
forma intracelular de leishmanias é mediada pela produção de NO, avaliamos a
atividade leishmanicida sobre a forma amastigota intracelular de L. infantum e
observamos que macrófagos infectados e tratados com SF0.5V, SF0.7V e
SF1.0V se tornaram menos susceptíveis à infecção. As heterofucanas que se
mostraram com capacidade de induzir a atividade anti-leishmania também
apresentaram melhor taxa de produção de NO, porém os dados de correlação
levaram a observação que este não é o principal mecanismo de ação das
fucanas de SF no combate a esse protozoário. A heterofucana SF0.5V também
apresentou atividade inibitória da formação de biofilme (~ 50%) frente a
bactéria Staphylococcus epidermidis. Já SF0.7V e 1.0V inibiram quase que
totalmente a replicação do protozoário Trichomonas vaginalis. Resultados
como esse refletem o espectro de ação desses polissacarídeos sulfatados
obtidos de SF e mostram o seu potencial como agentes imunomoduladores e
microbicidas.
Palavras chaves: Imunomodulação; Oxido nítrico; Leishmania infatum,
Trichomonas vaginalis, antibiofilme; macrófagos; fucanas.

xi
LISTA DE ABREVIATURAS E SIGLAS
µL Microlitros
g Grama
h Hora
kDa Kilodalton
KH2PO4 Fosfato monopotássico
M Molar
mg Miligrama
Min. Minutos
mL Mililitros
mM Milimolar
mm Milímetros
NaCl Cloreto de sódio
NaOH Hidróxido de sódio
nm Nanômetros
PA Para análise
RAEC Linhagem de células endoteliais de aorta de coelho
Raw264.7 Linhagem de células de macrófagos murinos
SF0.5 Fração precipitada com 0,5 volumes de acetona
SF0.7 Fração precipitada com 0,7 volumes de acetona
SF1,5 Fração precipitada com 1,5 volumes de acetona
SF1.0 Fração precipitada com 1,0 volumes de acetona
SF2,0 Fração precipitada com 2,0 volumes de acetona

xii
LISTA DE FIGURAS Figura 1. Esquema representataivo da metodologia de extração e caracterização físico-
química e farmacológica das heterofucanas da alga S. filipendula....................... 25

xiii
SUMÁRIO
RESUMO................................................................................................................................... x LISTA DE ABREVIATURAS E SIGLAS..................................................................................... xi LISTA DE FIGURAS.................................................................................................................. xiii 1. INTRODUÇÃO....................................................................................................................... 15
1.1. SISTEMA IMUNOLÓGICO……………………………………………………………… 15 1.2. PARASITAS VERSUS SISTEMA IMUNOLÓGICO………………………………...... 16
1.2.1. Leihmania sp………………………………………………………………… 16 1.2.2. Trichomonas vaginalis……………………………………………………… 17 1.2.3. Staphylococcus epidermidis e Klebsiella pneumina…………………..... 18
1.3. MODIFICADORES DA RESPOSTA BIOLÓGICA……………………………………. 19 1.4. FUCANAS: POLISSACARÍDEOS SULFATADOS DE MACROALGAS
MARRONS………………………………………………………………………………... 19
2. JUSTIFICATIVA.................................................................................................................... 20 3. OBJETIVOS........................................................................................................................... 21
3.1. OBJETIVO GERAL................................................................................................. 21 3.2. OBJETIVOS ESPECÍFICOS.................................................................................. 21
4. MÉTODOS............................................................................................................................. 22 4.1. MATERIAIS BIOLÓGICOS..................................................................................... 22
4.1.1. Algas...................................................................................................... 22 4.2. OBTENÇÃO DAS HETEROFUCANAS DE Sargassum filipendula....................... 22
4.2.1. Obtenção do pó cetônico..................................................................... 22 4.2.2. Proteólise.............................................................................................. 22 4.2.3. Fracionamento do extrato bruto com concentrações crescentes
de acetona............................................................................................. 23
4.3. ANÁLISES QUÍMICAS........................................................................................... 23 4.3.1. Dosagem de açúcares totais............................................................... 23 4.3.2. Dosagem de sulfato……………………………………………................ 23 4.3.3. Dosagem de proteínas……….............................................................. 24 4.3.4. Determinação da composição monossacarídica……....................... 24
5. ARTIGOS PRODUZIDOS...................................................................................................... 26
5.1. ARTIGO 1 (SUBMETIDO)...................................................................................... 27 5.2. ARTIGO 2 (PUBLICADO)…................................................................................... 53
6. COMENTÁRIOS, CRÍTICAS E SUGESTÕES...................................................................... 72 7. REFERÊNCIAS..................................................................................................................... 74

15
Telles, C.B.S. PPGCSA/CCS
1. INTRODUÇÃO
1.1. Sistema imunológico
O sistema imunológico é uma rede interativa de órgãos linfóides, células,
fatores humorais, citocinas, quimiocinas, dentre outros elementos, que
trabalham em conjunto para defender o organismo do ataque desses
“invasores” [1]. A resposta imunológica é dividida em dois tipos de acordo com
a velocidade e especificidade da reação, respostas inatas e adaptativas. A
imunidade inata abrange os elementos do sistema imune (Neutrófilos,
monócitos, macrófagos, complemento, citocinas e proteínas de fase aguda)
que fornecem a defesa imediata do hospedeiro [2]. A adaptativa consiste de
reações específicas contra o antígeno através do recrutamento de linfócitos T e
linfócitos B [2].
Durante o processo infeccioso, ou seja, no momento da invasão e
multiplicação do patógenoé estabelecido um processo inflamatório que é a
resposta protetora do corpo contra a infecção [3]. O início do processo
inflamatório é a detecção pelo sistema imune inato de elementos que são
comuns várias classes de agentes infecciosos, conhecidos como padrões
moleculares associados a patógenos (PAMPs) como, por exemplo, o
lipopolissacarídeo bacteriano (LPS); ou pela identificação dos padrões
moleculares associados ao dano (DAMPs), moléculas endógenas que
sinalizam o dano do tecido às células do sistema imune inato [4]. Após o
reconhecimento de PAMPs ou DAMPs por receptores celulares, como os
receptores transmembrana Toll-like (TLR), expresso na superfície de células
fagocíticas, ocorre uma cascata de sinalização intracelular que promove a
expressão de citocinas pro-inflamatórias como interleucina 1 β (IL-1β),
interleucina 6 (IL-6) e 12 (IL-12), fator de necrose tumoral (TNF), interferon
(INF-γ)e óxido nítrico sintase induzível (iNOS) [5,6], que, justamente com
quimiocinas e moléculas coestimulatórias, induzem o recrutamento de células
efetoras da inflamação como os monócitos e neutrófilos para o sítio do dano. A
resposta imune adaptativa também participa do processo inflamatório,
principalmente através dos linfócitos T auxilares, T CD4+, que quando
estimulados por células apresentadoras de antígeno, podem se diferenciar em

16
Telles, C.B.S. PPGCSA/CCS
diferentes tipos de células efetoras: células Th1, pró-inflamatórias; Th2, anti-
inflamatórias; células T regulatórias, Tregs; e células Th17 [7].
As células Th1 são críticas durante a resposta pró-inflamatórias, induzem a
liberação de INF-γ, IL-2 e TNF-α, fudamentais na ativação de macrófagos. As
células Th2 regulam a resposta imune humoral, proliferação de células B,
respostas alérgicas e proteção contra infecções de helmintos, promovendo a
produção de citocinas anti-inflamatórias IL-4, IL-5, IL-10 e IL-13. As células
Treg promovem a supressão da ativação, proliferação e funções efetoras de
várias células imunes como as células T, células NK, células B e células
apresentadoras de antígeno [8]. Secretam citocinas imunossupressoras IL-10 e
TGF-β que inibem a proliferação da resposta Th1 e Th2 para minimizar o dano
tecidual [9]. As células Th17 estão envolvidas na resposta contra
microparasitas extracelulares que necessitam de uma forte resposta
inflamatória e que não são adequadamente tratadas pelas respostas Th1 ou
Th2 [10].
Apesar de o sistema imunológico atuar fielmente na defesa do
organismo, alguns agentes invasores podem transpor a resposta imune e, na
ausência de um sistema imunológico eficaz, mesmo infecções menores podem
se estabelecer e tornarem-se fatais.
1.2. Parasitas “versus” Sistema Imunológico
Os parasitas são um grupo altamente diversificado de organismos que
desenvolveram diferentes estratégias para infectar seus hospedeiros.
1.2.1. Leishmania sp.
As leishmanioses são antropozoonoses consideradas causa de
morbidade e mortalidade em áreas subtropicais e tropicais. É uma doença com
um amplo espectro de manifestações clínicas, que incluem as formas: cutânea,
mucocutânea e visceral [11]. O agente causador da leishmaniose pertence ao
gênero Leishmania [12], família Trypanosomatidae e infecta o homem através
da picada do vertebrado fêmea da subfamília Phlebotominae [13]. Este parasita
tem um ciclo de vida digenético, multiplicando-se no intestino médio de
flebotomíneos na forma promastigota flagelada e como amastigotas não

17
Telles, C.B.S. PPGCSA/CCS
flageladas dentro fagócitos de mamíferos [14], sendo considerado um parasita
intracelular obrigatório.
Os macrófagos, células alvo das formas amastigotas de Leishmania,
possuem diversas formas de combater microorganismos invasores, dentre
elas, a ativação da enzima NADPH oxidase que induz a formação de espécies
reativas do oxigênio; a alteração do pH das vesículas fagocíticas levando a
desnaturação proteica [15]; a ativação da enzima óxido nítrico sintase induzida
(iNOS), que quando ativada sintetiza óxido nítrico (NO), molécula efetora no
combate às infecções [16]. Porém, mesmo diante do arsenal imunológico
disponível contra microorganismos invasores, as espécies de Leishmania são
capazes de evadir a resposta imunológica do hospedeiro vertebrado desde o
momento da infecção. A primeira resposta evitada é a lise mediada pelo
sistema complemento, as metaloproteses de superfície majoritárias (MSP),
presente na superfície das formas promastigotas infectantes, atuam clivando o
componente C3 em seus subprodutos e convertendo C3b em sua forma inativa
C3bi, que permanecem aderidas a superfície do protozoário, opsonizando-as e
facilitando sua fagocitose através da ligação aos receptores CR1 e CR3 dos
macrófagos essa ligação leva à diminuição da expressão de INF-γ, IL-12 e,
aumento na secreção de IL-4, contribuindo para a sobrevivência das
promastigotas [17]. Os macrófagos, definem o curso da infecção dependendo
da maneira como são ativadas. As células T auxiliares apresentam papel
fundamental nessa ativação, a expansão de clones Th1 leva a um controle da
doença, enquanto a expansão de clones Th2 leva a uma piora no quadro da
doença [15].
O tratamento das leishmanioses é baseado em antimoniais
pentavalentes (PVAs), cuja ação curativa foi descoberta pelo brasileiro Gaspar
Vianna em 1912 [18]. No entanto, estes fármacos, que vêm sendo utilizadas há
mais de seis décadas, estão longe de serem satisfatórios devido aos seus altos
custos de produção, toxicidade, surgimento de resistência medicamentosa
frente a todas as espécies de leishimania em várias partes do mundo [11], além
de apresentarem reconhecidos efeitos colaterais [19].
1.2.2. Trichomonas vaginalis

18
Telles, C.B.S. PPGCSA/CCS
Trichomonas vaginalis é um protozoário flagelado parasita que causa
tricomoníase, a mais comum doença sexualmente transmissível não viral [20].
A aderência e a citotoxicidade exercidas pelo parasita sobre as células do
hospedeiro são controladas por fatores de virulência, como adesinas, cisteino-
proteinases, integrinas, cell-detaching factor (CDF) e glicosidases. A aderência
do parasita as células epiteliais modula a expressão gênica de proteínas
funcionais das células do hospedeiro, como aquelas associadas à manutenção
da estrutura da matriz extracelular, moléculas pró apoptóticas e pró-
inflamatória. Um mecanismo de escape do sistema de defesa do hospedeiro é
a capacidade de T. vaginalis se auto-revestir de proteínas presente no plasma
do hospedeiro, impedindo, assim a identificação deste pelo sistema imune [21].
Metronidazole e Tinidazole são os únicos fármacos recomendados para
o tratamento da infecção por T. vaginalis [22]. No entanto, alguns estudos vêm
mostrando a resistência do T. vaginalis a esses fármacos [23], o que dificulta
ainda mais o controle da infecção.
1.2.3. Staphylococcus epidermidis e Klebsiella pneumonia
As bactérias são os microorganismos que mais frequentemente causam
infecções no homem [24]. Estes microorganismos são capazes de se replicar
tanto no interior das células do hospedeiro como em ambientes extracelulares
[25]. As infecções causadas por bactérias extracelulares são as mais
frequentes. Nesses casos os mecanismos de defesa estão relacionados
principalmente com as barreiras naturais do hospedeiro, a resposta imune
inata, a ativação do complemento e a produção de anticorpos. Nas bactérias
intracelulares a característica principal é a capacidade de sobrevivência dentro
dos macrófagos, podendo estimular as células TCD4+ através da expressão de
antígeno associado ao MHC classe II, os quais produzem citocinas ativadoras
de TCD8+ que reconhecem e destroem as células infectadas que expressam
antígenos associados a moléculas do MHC classe I. A ativação de células
TCD4+ leva à secreção de IFN-γ, que ativa os macrófagos levando à produção
aumentada de NO e destruição da bactéria [24].
Independente do tipo de parasita causador da infecção, o sistema imune
é fundamental para manutenção da homeostase do organismo infectado,

19
Telles, C.B.S. PPGCSA/CCS
portanto, torna-se um alvo importante para o desenvolvimento de estratégias
de tratamento. Dentre as abordagens incluem estratégias como vacinação
terapêutica com o designer de adjuvantes para conduzir determinados tipos de
resposta imune [26] e a busca por moléculas imunomoduladoras [27].
1.3. Modificadores da resposta biológica
O crescimento dos conhecimentos de imunologia clínica tem revelado
que a fisiopatologia de doenças pode ser causada tanto por exacerbação como
imunodeficiências da resposta imune. A terapia imunológica moderna é dividida
em dois grupos de imunomoduladores: o dos imunoestimuladores, que
conduzem ao aumento da imunidade inata e adaptativa, e o dos
imunossupressores, que diminuem a atividade do sistema imune [28].
Modificadores da resposta biológica (BRMs) são moléculas que atuam
como ativadores ou supressores da resposta de células do sistema imune [29].
Os principais efeitos biológicos promovidos pelos BRMs são atividades
anticoagulante, antitumoral, antifúngica, antibacteriana e antiparasitária [30-35].
Polissacarídeos de diferentes origens e com características estruturais variadas
vêm sendo estudados quanto as suas potenciais aplicações biológicas [36-40].
1.4. Fucanas: Polissacarídeos sulfatados de macroalgas marrons
As fucanas são polímeros que têm como característica principal à
presença da L-fucose sulfatada na sua estrutura [39,41], são os principais
polissacarídeos sulfatados obtidos das Phaeophyceas ou algas marrons.
O gênero Sargassum C. Agardh (Sargassaceae) constitui um dos mais
representativos dentre os 41 gêneros da ordem Fucales (Phaeophyceae,
Heterokontophyta), é amplamente distribuído nas regiões tropicais e
subtropicais do globo e é considerado um importante componente da flora
marinha [42]. As espécies desse gênero são reconhecidas fontes de fucanas
com uma variedade de atividade biológica: Sargassum horneri [43]; S.
tenerrimum [44]; S. patens [45]; S. stenophyllum [46], S. wightii [47], S. vulgare
[48], S. siliquosum [49].A alga marrom Sargassum filipendula além de possuir
extratos ricos em nutrientes que são utilizados na preparação de produtos

20
Telles, C.B.S. PPGCSA/CCS
cosméticos [50], apresenta fucanas ampla capacidade antioxidante e antimural
[51,52].
2. JUSTIFICATIVA
Diante da patogenicidade das infecções causadas pelos microorganismos
avaliados no presente trabalho e, devido ao limitado arcenal terapêutico no
controle desses patógenos, além do efeito adverso dos fármacos disponíveis,
que possuem um alto custo e o surgimento de resistência medicamentosa, vem
sendo travada uma verdadeira batalha entre a criatividade humana na
produção de drogas antimicrobianas cada vez mais potentes e de amplo
espectro de ação.
Substâncias produzidas a partir de produtos naturais, como as macroalgas
marinhas, podem oferecer uma oportunidade para superar esses efeitos
adversos, pois apresenta como vantagens o suprimento sustentável e baixo
custo.
Macroalgas marinhas do grupo das Phaeophyceas são potentes fontes
de moléculas bioativas, dentre elas, destacam-se as fucanas. O gênero
Sargassum é reconhecido por apresentar na constituição das suas espécies
fucanas com ampla atividade biológica. Recentemente, o nosso grupo de
pesquisa demonstrou que a espécie Sargassum filipendula é fonte de cinco
heterofucanas com alta atividade antioxidante e antitumoral. Contudo, apesar
da forte atividade biológica encontrada para as fucanas obtidas desta alga
marron, as atividades imunomuladoras e antimicrobianas destes compostos
ainda não foram estudadas. Assim, este trabalho surge como o primeiro passo
para se entender a possível ação imunomuladora e antimicrobiana das fucanas
de Sargassum filipendula.

21
Telles, C.B.S. PPGCSA/CCS
3. OBJETIVOS
3.1. GERAL
Avaliar o potencial imunomudulador e antimicrobiano das heterofucanas
de S. filipendula frente aos microorganismos Leishmania infantum,
Trichomonas vaginalis, Staphylococcus epidermidis e Klebsiella pneumonia
(KPC).
3.2. ESPECÍFICOS
• Extrair polissacarídeos sulfatados da alga marrom Sargassum filipendula;
• Caracterizar quimicamente as heterofucanas obtidas;
• Avaliar o efeito das heterofucanas sobre macrófagos murinos (Raw 264.7)
quanto à produção de NO, Il-6 e TNF-α;
• Investigar a atividade anti-leishmania das heterofucanas sobre macrófagos
infectados com Leishmania infatum.
• Avaliar o efeito das heterofucanas nos ensaios anti-Trichomonas vaginalis;
• Investigar se as heterofucanas possuem efeito antibacteriano e antibiofilme
contra Staphylococcus epidermidis e Klebsiella pneumonia (KPC).

22
Telles, C.B.S. PPGCSA/CCS
4. MÉTODOS
Serão descritos a seguir apenas os métodos que necessitam de um
maior detalhamento para compreensão, os demais métodos encontram-se
claramente descritos nos artigos.
4.1. MATERIAIS BIOLÓGICOS
4.1.1. Algas
A alga marinha marrom Sargassum filipendulafoi coletada na Praia de
Búzios, município de Nísia Floresta (litoral sul do Rio Grande do Norte), em
marés baixas entre 0,0 a 0,2 metros a uma temperatura situada entre 28-30 °C.
Após serem coletadas, as algas foram trazidas ao laboratório de
Biotecnologia de Polímeros Naturais (BIOPOL) da Universidade Federal do Rio
Grande do Norte (UFRN), onde foram lavadas e retiradas as epífitas e
inclusões calcárias. Em seguida foram secadas em estufa a 45 ºC, trituradas e
guardadas em recipientes apropriados.
4.2. OBTENÇÃO DAS HETEROFUCANAS DA ALGA Sargassum filipendula
4.2.1. Obtenção do pó cetônico
A alga seca e pulverizada foi suspensa em dois volumes de acetona PA,
durante 24 horas para despigmentação e delipidação do material. A mistura foi
decantada e o resíduo colocado para secar a 45 °C sob aeração e denominado
de “pó cetônico”. Esse pó foi utilizado em seguida na proteólise.
4.2.2. Proteólise
A 100 gramas de pó cetônico foram adicionados dois volumes de NaCl a
0,25 M e o pH ajustado para 8,0 com NaOH. A esse material foi adicionado a
enzima proteolítica prozima (15 mg/g de pó cetônico). Essa suspensão

23
Telles, C.B.S. PPGCSA/CCS
permaneceu em banho-maria a 60 °C por 24 h. Depois, foi filtrado e o
sobrenadante submetido a uma centrifugação 10.000 x g por 15 minutos a
temperatura de 4 °C. Após a centrifugação, o sobrenadante foi denominado de
extrato bruto.
4.2.3. Fracionamento do extrato bruto com concentrações crescentes de
acetona
O extrato bruto obtido foi fracionado com volumes crescentes de
acetona, obtendo-se as frações polissacarídicas. Adicionou-se um volume de
acetona, sob agitação leve, necessário para que se visualizasse uma turvação
da solução, essa solução foi mantida em repouso a 4 ºC durante 18 h, o
precipitado foi coletado por centrifugação a 8.000 x g por 15 minutos a 4 ºC e
seco a pressão reduzida.
Em seguida, esse procedimento foi repetido até que não se visualizasse
mais a formação de precipitado [37]. As frações obtidas foram denominadas
conforme o volume de acetona no qual foram precipitadas (SF0.5V, SF0.7V,
SF1.0V, SF1.5V e SF2.0V) (Figura 01).
4.3. ANÁLISES QUÍMICAS
4.3.1. Dosagem de açúcares totais
Açúcares totais de cada extrato polissacarídico e de cada fração
polissacarídica obtida foram determinados pelo método do fenol/ácido sulfúrico,
utilizando-se como padrão galactose, sendo as leituras realizadas a 490 nm
[53].
4.3.2. Dosagem de sulfato
O teor de sulfato total de cada extrato polissacarídico e de cada fração
polissacarídica obtida foi quantificado, após uma hidrólise ácida com 4 N de
HCl por 6 horas à temperatura de 100 °C, por turbidimetria pelo método da
gelatina-bário, tendo-se como padrão o sulfato de sódio [54].

24
Telles, C.B.S. PPGCSA/CCS
4.3.3. Dosagem de proteínas
O teor de proteína correspondente de cada extrato polissacarídico e de
cada fração polissacarídica obtida foi determinado com o reagente de comassie
blue R 250 e a leitura realizada a 595 nm [55].
4.3.4. Determinação da composição monossacarídica
A composição monossacarídica dos polissacarídeos sulfatados das
algas Sargassum filipendula e Dyctiopteris delicatula foi determinada através
de cromatografia liquida de alta performance (HPLC) contendo um detector de
índice refrativo modelo L-2490. Utilizou-se uma coluna LichroCART® 250-4
(250 mm × 40 mm) contendo como pré-coluna a Lichrospher® 100 NH2 (5 μm).
Os polímeros foram hidrolisados (2 M HCl, 100 °C, 2 h) e posteriormente os
seus monossacarídeos foram analisados. Como referências, os seguintes
açucares foram utilizados como padrões: arabinose, galactose, glicose, fucose,
manose, ramnose, ácido glucurônico, ácido manurônico, N-acetil glicosamina e
xilose. A fase móvel consistiu de uma mistura de 0,1 mol/l de KH2PO4 (pH 10)-
acetonitrila (80:20). O fluxo foi de 1.0 mL/min e a temperatura da coluna foi de
80 °C.

25
Telles, C.B.S. PPGCSA/CCS
Figura 01. Esquema representataivo da metodologia de extração e caracterização físico-química e farmacológica das heterofucanas da alga S. filipendula.

26
Telles, C.B.S. PPGCSA/CCS
5. ARTIGOS PRODUZIDOS
5.1. Artigo 1 (SUBMETIDO)
Immunomodulatory effect and antimicrobial activity of Sargassum
filipendula heterofucanas
Periódico: Marine Drugs
Fator de impacto (2015): 3.512
ISSN: (Printed version)
ISSN: (Online version)
Qualis: Medicina II – A2
Indexada: PubMed – indexado por MEDLINE
5.2. Artigo 2 (PUBLICADO)
Methanolic Extracts from Brown Seaweeds Dictyota cilliolata and Dictyota
menstrualis Induce Apoptosis in Human Cervical Adenocarcinoma HeLa
Cells
Periódico: Molecules
Molecules. 2015 Apr 13;20(4):6573-91
Fator de impacto (2015): 2.095
ISSN: 1420-3049 (Printed version)
ISSN: 1420-3049 (Online version)
Qualis: Medicina II – A2
Indexada: PubMed – indexado por MEDLINE
5.3. Capítulo de livro
Chapter: Carrageenans
Periódico: Biochemistry and Molecular Biology in the Post Genomic Era
Binding: ebook Pub. Date: 2015 ISBN: 978-1-63483-002-7

27
Telles, C.B.S. PPGCSA/CCS
5.1. ARTIGO 1 (SUBMETIDO)
Mar. Drugs 2015, 13, 1-x manuscripts; doi:10.3390/md130x000x
marine drugs ISSN 1660-3397
www.mdpi.com/journal/marinedrugs
Article
Immunomodulator effects and antimicrobial activity of heterofucanas from Sargassum filipendula
Cinthia Beatrice Silva Telles 1,2
, Carolina Mendes-Aguiar 3, Gabriel Pereia Fidelis
1,
Amanda Piccoli Frasson4, Leandro Silva Costa
1,5, Tiana Tasca
3,; Selma Maria
Bezerra Jeronimo1,2
and Hugo Alexandre Oliveira Rocha 1,2,
*
1 Laboratório de Biotecnologia de Polímeros Naturais (BIOPOL), Departamento de
Bioquímica, Centro de Biociências, Universidade Federal do Rio Grande do Norte
(UFRN), Natal,
Rio Grande do Norte-RN 59078-970, Brasil; E-Mails: [email protected]
(C.B.S.T.); [email protected] (L.S.C.), [email protected]
(G.P.F.); [email protected] (H.A.O.R) 2 Programa de Pós-graduação em Ciências da Saúde, Universidade Federal do Rio
Grande do Norte (UFRN), Natal, Rio Grande do Norte-RN 59078-970, Brasil. 3 Instituto de Medicina Tropical do Rio Grande do Norte, Departamento de Bioqiímica
Universidade Federal do Rio Grande do Norte (UFRN), Natal, Rio Grande do Norte-
RN 59078-970, Brasil; E-Mail: [email protected] (C.M.A.);
[email protected] (S.M.B.J.) 4 Faculdade de Farmácia da Universidade Federal do Rio Grande do Sul, Av. Ipiranga,
2752, Porto Alegre, RS, 90610-000, Brasil. E-Mails: [email protected] (T.T);
[email protected] (A.P.F.) 5 Intituto Federal de Educação, Ciência e Tecnologia do Rio Grande do Norte (IFRN),
Santa Cruz, Rio Grande do Norte-RN 59200-000, Brasil.
* Author to whom correspondence should be addressed; E-Mail: [email protected];
Tel.: +55-84-3215-3416 (ext. 207); Fax: +55-84-3211-9208.
Academic Editor:
Received: / Accepted: / Published:
OPEN ACCESS

28
Telles, C.B.S. PPGCSA/CCS
Abstract: Fucans, sulphated polysaccharides that contain L-fucose in its
constitution, obtained from species of Phaeophyceae of the Sargassum kind,
display several biological activities. Heterofucans from Sargassum
filipendula are bioactive molecules that contain strong antiproliferative and
antioxidant activity. However, their immunomodulatory and antimicrobial
activities have not yet been examined. In this context, the aim of this
research was to evaluate the heterofucans as for their immunomodulatory
capacity and antimicrobial action against Leishmania infantum,
Trichomonas vaginalis, Staphylococcus epidermidis and Klebsiella
pneumonia (KPC). The five heterofucans obtained from S. filipendula show
activities that are distant as stimulants of the immune system and microbial
agent. The SF0.5V, SF0.7V amd SF1.0V heterofucans were capable of
acting in the activation of murine and human macrophages. In addition to
that, SF0.5V has shown antibiofilm activity of S. epidermides and SF0.7V
and 1.0V almost completely inhibited the survival of the protozoan T.
vaginalis. Results such as this one, reflect the broad range of action of the
sulphated polysaccharides obtained from seaweeds, especially from the
species S.filipendula
Keywords: Immunomodulation; Nitric oxide; Leishmania infatum,
Trichomonas vaginalis, antibiofilm; macrophages; fucans.
1. Introduction
The immune response performs a fundamental role in the defense against infectious
agents and it constitutes the most important prevention against the occurrence of
infection spreads that are normally associated with a high mortality rate [1]. The
establishment of an infection, in a vulnerable host, covers several mechanisms; one of
the most relevant is the way that the microorganism interacts with the immune system
and its response against the invader [2].
The parasites are highly diversified of organisms that developed different strategies
to infect their hosts. These infectious agents vary from single-celled organisms such as
bacteria and protozoans to multi-celled organisms, such as nematodes and helminths
(worms). The bacteria are microorganisms capable of replicating inside the host cells as
much as in extracellular environments, blood circulation, intestinal lumen, airways,
amongst others [2]. On the other hand, protozoans are infectious intracellular agents that
normally infect the host for long periods of time [1]. The immune mechanisms involved

29
Telles, C.B.S. PPGCSA/CCS
in fighting the bacterial infections and caused by protozoans vary; generally,
intracellular parasites are eliminated through mechanisms mediated by cell and the
extracellular ones via mechanisms that mainly involve the complement system and
antibodies [3, 4].
The drugs available for the treatment of parasite related diseases are far from
satisfactory due to their high production costs, toxicity, as well as the appearance of
resistance [5-8].
The increase in knowledge about clinical immunology has been revealing that the
physiopathology of illnesses can be caused by exacerbation, as well as by
immunodeficiencies of the immune response. The modern immunologic therapy is
divided in two basic groups of immunomodulators: the immunostimulants, which lead
to the increase of innate and adaptive immunity, and the immunosuppressives that
diminish the activity on the immune system [9].
Natural products are important sources of innovative therapeutic agents for infectious
diseases, cancer, lipid disorders and immunomodulation [10].
Sulphated polysaccharides are a complex group of bioactive macromolecules in
which some of the hydroxyl groups from sugar waste are replaced by sulphate groups.
Seaweeds are the main non-animal sources of obtainment of these anionic
polysaccharides; in this group, we find the Fucans, which are a family of
polysaccharides that contain L-fucose in its constitution [11].
Over the last few years, a variety of groups reported that sulphated polysaccharides
obtained from Phaeophytas species, from the genus Sargassun showcase various
biological activities: Sargassum horneri [12]; S. tenerrimum [13]; S. patens [14]; S.
stenophyllum [15], S. wightii [16], S. vulgare [17], S. siliquosum [18]. Our group
assessed the heterofucans from Sargassum filipendula, common seaweed along the
northeastern cost of Brazil, and demonstrated that these polymers are bioactive
molecules presenting strong antiproliferative and antioxidant activity. However, the
immunomodulator and antimicrobial purified activities of the sulphated polysaccharides
from S. filipendula have not yet been examined. In this context, the objective of this
study was to obtain sulphated polysaccharides from S. filipendula and evaluate their
immunomudulating and antimicrobial activities facing Leishmania infantum,
Trichomonas vaginalis, Staphylococcus epidermidis and Klebsiella pneumonia (KPC).
2. Results and Discussion
Chemical characteristics of heterofucanas SF0.5V, SF0.7V, SF1.0V, SF1.5V and
SF2.0V obtained from seaweed S. filipendula are presented in Table 1. The
heterofucans SF0.5V and SF2.0V were the ones that presented the lowest ratio
sugar/sulphate, indicating that the amount of sulphate by sugar residue is bigger when

30
Telles, C.B.S. PPGCSA/CCS
compared to the other heterofucans. The monosaccharides fucose, galactoses, glycoses
and xyloses were found in different amounts in all polysaccharides. Mannoses and
glucuronic acid are also present in almost all of the heterofucans, except for SF1.5V,
that does not have mannoses residues and SF2.0V that does not have glucuronic acid in
its structure. Comparing the data described on table 1 with the data published by Costa
and collaborators [19], it is possible to verify that both are very similar, which indicates
that the heterofucans extracted are the same obtained by Costa and collaborators [19].
These fucans present antioxidant and antitumor activities, however other activities have
not been assessed yet, therefore, we verified weather these fucans had
immunomodulator and antimicrobial effect.
Table 1. Chemical characteristics of the heterofucans from S.filipendula.
2.1. Imunomodulator effect of the sulphated heterofucans from S. filipendula in murine
macrophages (RAW 264.7)
Macrophages are essential for the maintenance of homeostasis and play a main role
in the defense of the host against pathogens. In this process, activated macrophages
release some immunomodulator factors, such as NO, IL-6, TNF-α, amongst others [21].
Therefore, initially, we evaluated the effect of heterofucans in murine macrophages
(RAW 264.7) in culture and isolation conditions.
2.1.1. Production of NO
The production of NO by RAW 264.7 cells, stimulated by the different heterofucans
(0.125, 0.25 and 0.5 mg/mL) is demonstrated on Figure 01. After 24 hours of treatment,
the level of NO in the cultures stimulated with the heterofucans SF1.5V and SF2.0V
was similar to that observed for the non-stimulated cells (control group). On the other
hand, it was observed that the amount of NO present on the supernatant of the
macrophages increased considerably when the cells were incubated with SF0.5V,
SF0.7V and SF1.0V, this effect presents a tendency to be dose-dependent. It is
important to stress that the RAW cells, in the presence of SF0.5V (0.5 mg/mL) made
the amount of NO in extracellular environment similar to the one found in the

31
Telles, C.B.S. PPGCSA/CCS
extracellular environment of the cells that were treated with positive control, which
suggests a strong immunostimulating action of the macrophages by this heterofucan.
Other groups also proved that different fucans have distinct potentials as
immunostimulating agents, e.g. fucans obtained from the weeds Ascophyllum nodosum
and Fucus vesiculosus increased the amount of NO released to the extracellular
environment when in contact with RAW cells. However, the Ascophyllum fucana was
six times more potent than Fucus [22]. The presented information clearly indicates that
the potency of stimulation of the RAW cells carried out by the fucans in order to release
NO is not the same and depends on the properties of each fucana, as was observed with
the heterofucans from S filipendula.
Amongst the properties featured as important for a fucan to interfere in the amount of
NO releases, it is important to mention the degree of sulphatation. Nevertheless, the
results obtained with the heterofucans (SF0.5V and SF2.0V) were intriguing; despite the
two present similar ratio sugar/sulphate, they presented different effects in the
immunostimulation of macrophages and releasing of NO, which means that one of them
is a stimulator (SF0.5v), whereas the other one (SF2.0v) does not affect the amount of
NO in the environment.
According to Leiro and collaborators [23] the sulphate groups are important points so
that a sulphated polysaccharide is able to stimulate RAW 264.7 cells to release NO to
the extracellular environment. These authors presented that sulphated polysaccharides
of the Ulva rígida weed were stimulating agents of the RAW cells, however when they
were disulphated, their activity was lost. Jiang and collaborators [24] also demonstrated
that when disulphating ascophylan, a homofucan obtained from the weed Ascophyllum
nodosum, its RAW macrophages stimulating activity was significantly decreased when
compared to the native ascoplylan.
Despite many authors presenting that the amount of sulphate groups present in the
polysaccharide is important to its action, the position in which these groups are
distributed in the molecule is a much more determining factor so that a sulphated
polysaccharide present higher or lower activity [25-28]. Therefore, we believe that this
would be the reason that makes SF0.5v and SF2.0V, that present similar ratio
sugar/sulphate, present distinct activities.

32
Telles, C.B.S. PPGCSA/CCS
Figure 1. Effect of the heterofucans from S. filipendula over the release of
NO by RAW264.7 cells. The data are presented as average ± standard
deviation (n = 3). The letters a, b indicate a significant difference (p <0,05)
between the concentration of the heterofucans. # Indicates the significant
difference (p <0,05) between the concentration of the heterofucana and
positive control. * Indicates significant difference (p <0,05) between the
concentration of the heterofucana e negative control. NC: Negatice control;
PC: Positive Control (LPS 2 µg/mL).
2.1.2. Production of the cytokines TNF-α and IL-6
On Figure 02 we observe that the cells RAW 264.7, when exposed to the
heterofucans SF1.5V and SF2.0V, did not promote alteration on the level of TNF-α and
IL-6 in the extracellular environment, proving, alongside with the previous data, that
these polymers possibly do not act as immunomodulator agents.
The heterofucana SF1.0V also did not induce the cells RAW 264.7 to produce and
release to the extracellular environment significant amount of TNF-α and IL-6. This
characteristic is not entirely of these fucans, other heterofucan, extracted from the
seaweed Dictyota menstrualis, was also not able to interfere on the production of these
two cytokines [29]. The presented data lead to the observation that the
immunomodulator mechanism of SF1.0V would be centred in its capacity to interfere
on the production and release of NO.
The fucans that altered the highest amount of TNF-α and IL-6 were SF0.5V and
SF0.7V, in both the cases the TNF-α was the cytokine that was most affected by the
presence of the fucans. As to SF0.5V, it was observed that the presence of 0.25 mg/mL

33
Telles, C.B.S. PPGCSA/CCS
of this fucan increased in about a thousand times the amount of TNF-α in the
extracellular environment in comparison with the amount of TNF-α found in the control
group. This result was very expressive, since other fucans of Fucus vesiculosus [22] and
Ascophyllum nodosum [30], in the concentration of 0.2 mg/mL, were only capable of
increasing the amount of TNF-α in the extracellular environment in 20 times.
Figure 2. Effect of the heterofucans from S. filipendula over the release of
TNF-α and IL-6 by RAW264.7 cells. The data are present as average ±
standard deviation (n = 3). The letters a, b indicate a significant difference
(p <0,05) between the concentration of the heterofucans. # Indicates
significant difference (p <0,05) between the effect of the heterofucana and
positive control. * Indicates significant difference (p <0,05) between the
effect of the heterofucan and negative control. NC: Negative control; PC:
Positive control (LPS 2µg/mL).
Do and collaborators [31], evaluating the effect of the homofucan extracted from the
brown weed Fucus vesiculosus in the induction of the production of NO by
macrophages (RAW 264.7 and primary peritoneal cells), observed that this fucan made
the cells that were being studied increase the release of NO to the extracellular
environment, as well as TNF-α. These authors suggest that TNF-α has a synergic effect
and increases the release of NO to the extracellular environment even more, which
would explain the effect of SF0.5V and SF0.7V, in other words, the way these fucans
stimulate the release of TNF-α induces a higher release of NO. And also explains why
SF1.0V stimulates the release of a smaller amount of NO, because that fucan does not
affect the release of TNF-α.
34
Telles, C.B.S. PPGCSA/CCS
Different groups that study fucans state that they are immunomodulators because
they induce the activation in vitro of murine macrophages (RAW 264.7) leading to the
increase of the production of NO and cytokines such as TNF-α and IL-6 [22,32-34].
Which leads us to propose that S. filipendula synthetizes imunomodulator fucans.
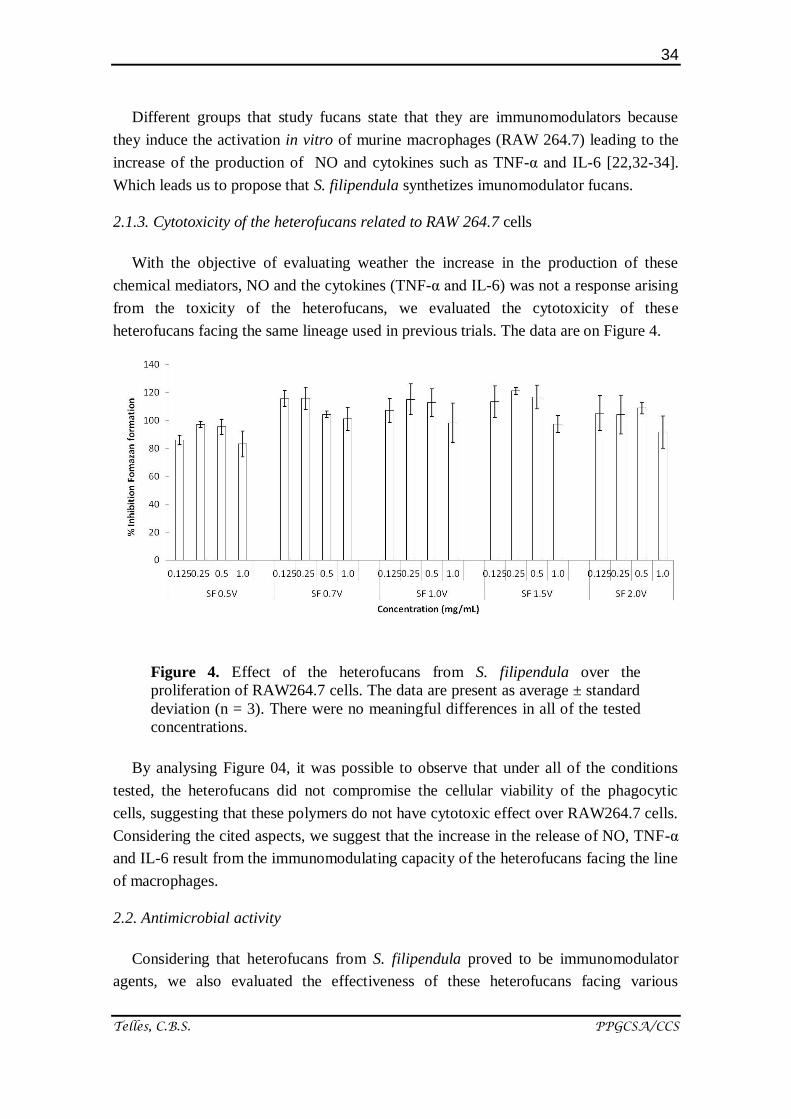
2.1.3. Cytotoxicity of the heterofucans related to RAW 264.7 cells
With the objective of evaluating weather the increase in the production of these
chemical mediators, NO and the cytokines (TNF-α and IL-6) was not a response arising
from the toxicity of the heterofucans, we evaluated the cytotoxicity of these
heterofucans facing the same lineage used in previous trials. The data are on Figure 4.
Figure 4. Effect of the heterofucans from S. filipendula over the
proliferation of RAW264.7 cells. The data are present as average ± standard
deviation (n = 3). There were no meaningful differences in all of the tested
concentrations.
By analysing Figure 04, it was possible to observe that under all of the conditions
tested, the heterofucans did not compromise the cellular viability of the phagocytic
cells, suggesting that these polymers do not have cytotoxic effect over RAW264.7 cells.
Considering the cited aspects, we suggest that the increase in the release of NO, TNF-α
and IL-6 result from the immunomodulating capacity of the heterofucans facing the line
of macrophages.
2.2. Antimicrobial activity
Considering that heterofucans from S. filipendula proved to be immunomodulator
agents, we also evaluated the effectiveness of these heterofucans facing various

35
Telles, C.B.S. PPGCSA/CCS
parasites, for instance: Leishmania infatum; trichomonas vaginalis; Klebsiella
pneumoniae (KPC) and Staphylococcus epidermidis.
2.2.1. Infection caused by Leishmania infatum in human macrophages
Considering that our group have not yet standardized the methodology for infection
of RAW cells with leishmania, we chose to work with macrophages originated from
primary human monocytes, as described in methods.
2.2.1.1. Production of NO
Macrophages originated from primary human monocytes exposed for 24 and 48
hours to the heterofucans from S. filipendula, in the concentration of 0.5 mg/mL,
increased distinctly the amount of NO in the extracellular environment (Figure 5A).
The heterofucans SF0.5V, SF0.7V and SF1.0V (Fig 5A) promoted meaningful increase
in the release of NO to the extracellular environment in all of the periods analysed when
compared to the untreated macrophages. In addition, the release of NO by macrophages
stimulated by SF0.5V and SF0.7V in the period of 48 hours of exposure was about 2
times higher than that presented by untreated macrophages. The other polysaccharides
(SF1.5V and SF2.0V) did not present meaningful effect over the production of NO by
macrophages. These data is similar to that obtained during the treatment of murine
macrophages (RAW 264) (Figure 02), that demonstrated the polysaccharides SF0.5V,
SF0.7V and SF1.0V presented an effective immunomodulating action.
A second cultivation of phagocytic human cells was carried out. However, this time
these cells were initially infected with the amastigote form (intracellular form of
leishmania) and posteriorly incubated with the heterofucans from S. filipendula with the
intent of evaluating the effect of the fucans in the release of NO by the infected cells.
On Figure 5B we can observe that the human macrophages infected and incubated
with the heterofucans from S. filipendula (0.5 mg/mL), also distinctly increased the
amount of NO in the extracellular environment. The production of NO after the
treatment for 24 and 48 hours with the heterofucana SF2.0V was not altered; it kept the
level of NO similar to that observed with the untreated observed cells. On the other
hand, all of the other heterofucans induced an increment in the production of NO
(Figure 5B). The heterofucans SF0.5V and SF0.7V were the best activators of the
immune response, and presented time depending effect. Within 48 h these fucans
elevated the amount of NO in about five times, comparing to the control.

36
Telles, C.B.S. PPGCSA/CCS

37
Telles, C.B.S. PPGCSA/CCS
Figure 5. Effect of the heterofucans from S. filipendula over the release of
NO by human macrophages. Production of NO by human macrophages after
treatment with the heterofucans (A); Production of NO by macrophages
infected by L. infatum and treated with the heterofucans (B). The data are
presented as averages ± standard deviation (n = 3). The letters a, b indicate
a significant difference (p <0,05) in the release of NO on different periods. *
Indicates significant difference (p <0,05) between different heterofucans and
the negative control.
2.2.1.2. Leishmanicidal activity
After the evaluation of the effects of the heterofucans over the production of NO, we
evaluated the leishmanicidal activity over the amastigote intracellular form of
Leishmania infantum.
The results on Figure 6 demonstrate that the highest percentage of cells infected by
amastigotes can be found in the control group cells. It is also possible to observe that
there is no significant difference between the percentage of cellular infection in the
control group and in the group treated with the heterofucan SF2.0V. The other fucans, at
least, in one of the conditions tested diminished significantly the rate of cellular
infection by amastigotes.

38
Telles, C.B.S. PPGCSA/CCS
Figure 6. Leishmanicidal activity of the heterofucans from S. filipendula.
The results are expressed in average ± standard deviation (n = 3) of the
percentages of the macrophages infected with L. infantum * Indicates
significant difference (p <0,05) between the concentration of the
heterofucan and the CN. CN: Negative control – Untreated macrophages
infected with L. infatum.
In a second experiment, the macrophages were plated over a cover slip and infected
in the presence or not of the heterofucans (0.5 mg/mL), posteriorly they were fixed and
coloured, and the number of amastigotes that were inside the cells was determined. The
data are on Figure 7. In the control group and in the group treated with SF2.0V, the
number of intracellular parasites was similar and corresponded to approximately 4
amastigotes per macrophage. During the treatment with the other heterofucans (SF0.5V,
SF0.7, SF1.0V and SF1.5V) the number of amastigotes was smaller, about 3
amastigotes/macrophages. (Figure 07A).
Figure 7. Leishmanicidal activity of the heterofucans from S. filipendula.
Number of amastigotes of L.infatum per infected macrophages (A).
Micrographs of the parasites internalized in the macrophages (B). The cover
slips were processed by optical microscopy, coloured with Panotic e
photographed using a 100X zoom. The arrows show internalized parasites.

39
Telles, C.B.S. PPGCSA/CCS
Figure 07B illustrates micrographs that represent the preparation of macrophages
infected by Leishmania (L.) infatum, treated or not with the heterofucans from S.
filipendula. On all of the preparations, we observed the presence of amastigotes in the
interior of the cells, indicated by arrows. We observed a slight decrease in the number
of amastigotes after treatment with the heterofucans when compared to the untreated
cells (control group)
Altogether, the presented data demonstrates that the fucans (mainly SF0.5V, SF0.7V
and SF1.0v) decreased in approximately 20% the percentage of macrophage infection
(Figure 06). This percentage is low if compared to the activity of fucoidan of F.
vesiculosus, which promotes a decrease of about 90% of the number of human
macrophages infected by Leishmania [35]. That indicates that the fucans from
Sargassum are not candidates for the treatment of leishmaniasis, but, due to their
immudomodulator capacity, they can act as supporting agents in the treatment of this
illness, and is raw material for the production of nanoparticles that carry
antileishmanicidal drugs.
2.2.2. Anti-trichomonas vaginalis activity
Heterofucans from the seaweed S. filipendula were also analysed against
trophozoites of T. vaginalis (Figure 08). Triage revealed that the heterofucans SF0.5V
and SF2.0V did not present inhibitory activity facing trophozoites. Whereas SF0.7V,
SF1.0V and SF1.5V presented anti-T. vaginalis activity after 24h of treatment.
Figure 8. Effect of different heterofucans from S. filipendula (2 mg /mL)
against isolated of T. vaginalis sensitive to metronidazol. Control:
Trophozoites in untreated culture with the heterofucans. The data represent
averages ± standard deviation of at least three experiments. . # Indicates
significant difference (p <0,05) between the concentration of the
heterofucan and control.

40
Telles, C.B.S. PPGCSA/CCS
The heterofucans SF0.7V and SF1.0V that presented best inhibitory capacity facing
trophozoites of T. vaginalis (approximately 90% of inhibition) present ratio
sugar/sulphate of approximately 4.5, in counterpart, SF0.5 and SF2.0V present smaller
values (4,06 and 3,72) did not have any activity in this essay, which leads us to suggest
that the degree of sulphatation should not be a preponderant factor to a satisfactory anti-
T. vaginalis activity.
The action of the heterofucans as strong inhibitory agents of the flagellate protozoan
Trichomonas vaginalis is of key importance since trichomoniasis, sexually transmitted
disease (STD), is an infirmity present all over the world [36] and its treatment is
essentially based in the use of the drug 5-nitroimidazole [37]. However, some reports
have demonstrated the appearance of resisting strains [38]. Therefore, it is necessary to
search for a new therapeutic arsenal. Besides, there is not a study that has demonstrated
the activity of polysaccharides in the fighting of T. vaginalis, so this is the first study
that brings up that polysaccharides obtained from seaweed (SF0.7V, SF1.0V and
Sf1.5V) have cytotoxic action against this pathogenic protozoan.
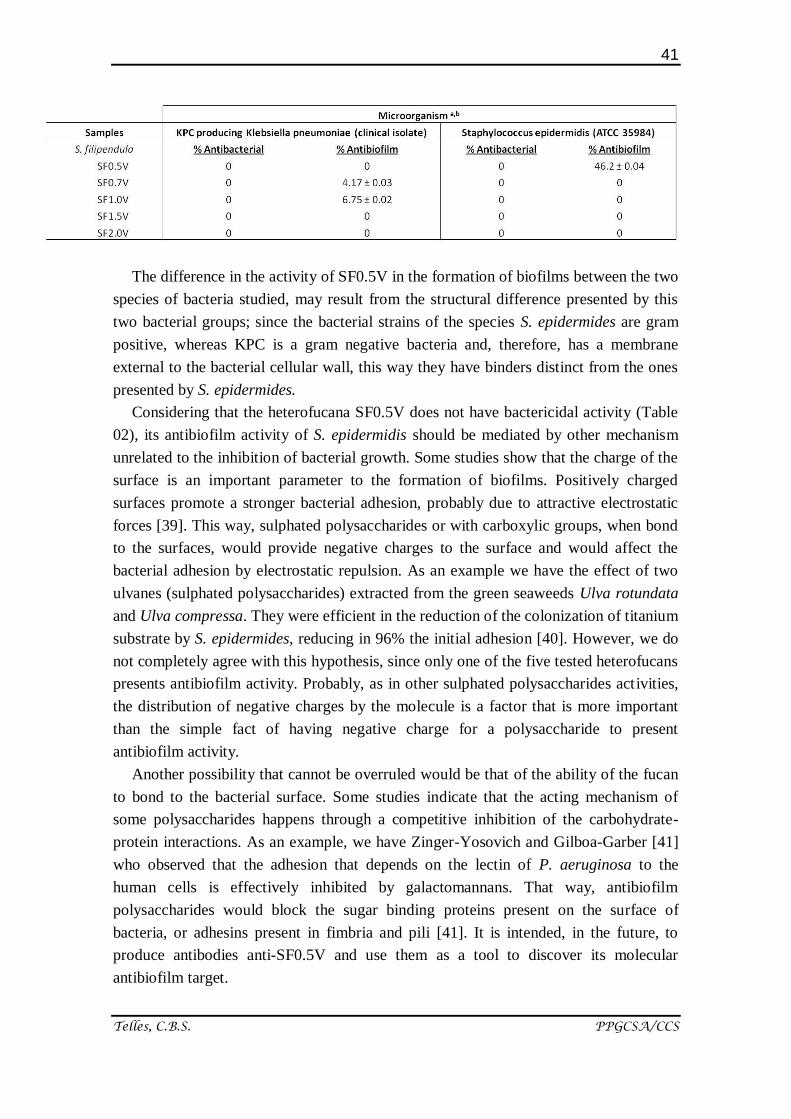
2.2.3. Antibacterial and Antibiofilm activity
All of the heterofucans from S. filipendula were evaluated as to their antibacterial
and inhibitory capacity concerning the formation of bacterial biofilms (Table 02). In this
study, no heterofucana of S. filipendula was proved effective in the fighting of bacterial
growth, also did not considerably inhibit the formation of biofilms, promoted by the
association of bacteria of the species Klebsiella pneumoniae (KPC). The heterofucans
also did not have antibacterial activity against Staphylococcus epidermidis. However,
SF0.5V presented antibiofilm activity, inhibiting about 50% of biofilm formation by
these bacteria.
Table 2. Antibacterial and antibiofilm activity of the heterofucans from S.filipendula. a
The data are the average value of three determinations ± DP; b
The results were
obtained in the concentration of 2 mg/mL, other tested concentrations did not have
significant activity; c rifampicin (Sigma-Aldrich Co., St. Louis, MO, USA) was used as
control antibiotic.
41
Telles, C.B.S. PPGCSA/CCS
The difference in the activity of SF0.5V in the formation of biofilms between the two
species of bacteria studied, may result from the structural difference presented by this
two bacterial groups; since the bacterial strains of the species S. epidermides are gram
positive, whereas KPC is a gram negative bacteria and, therefore, has a membrane
external to the bacterial cellular wall, this way they have binders distinct from the ones
presented by S. epidermides.
Considering that the heterofucana SF0.5V does not have bactericidal activity (Table
02), its antibiofilm activity of S. epidermidis should be mediated by other mechanism
unrelated to the inhibition of bacterial growth. Some studies show that the charge of the
surface is an important parameter to the formation of biofilms. Positively charged
surfaces promote a stronger bacterial adhesion, probably due to attractive electrostatic
forces [39]. This way, sulphated polysaccharides or with carboxylic groups, when bond
to the surfaces, would provide negative charges to the surface and would affect the
bacterial adhesion by electrostatic repulsion. As an example we have the effect of two
ulvanes (sulphated polysaccharides) extracted from the green seaweeds Ulva rotundata
and Ulva compressa. They were efficient in the reduction of the colonization of titanium
substrate by S. epidermides, reducing in 96% the initial adhesion [40]. However, we do
not completely agree with this hypothesis, since only one of the five tested heterofucans
presents antibiofilm activity. Probably, as in other sulphated polysaccharides activities,
the distribution of negative charges by the molecule is a factor that is more important
than the simple fact of having negative charge for a polysaccharide to present
antibiofilm activity.
Another possibility that cannot be overruled would be that of the ability of the fucan
to bond to the bacterial surface. Some studies indicate that the acting mechanism of
some polysaccharides happens through a competitive inhibition of the carbohydrate-
protein interactions. As an example, we have Zinger-Yosovich and Gilboa-Garber [41]
who observed that the adhesion that depends on the lectin of P. aeruginosa to the
human cells is effectively inhibited by galactomannans. That way, antibiofilm
polysaccharides would block the sugar binding proteins present on the surface of
bacteria, or adhesins present in fimbria and pili [41]. It is intended, in the future, to
produce antibodies anti-SF0.5V and use them as a tool to discover its molecular
antibiofilm target.

42
Telles, C.B.S. PPGCSA/CCS
This antibiofilm activity presented by the heterofucana SF05V is fundamentally
important since biofilms hamper the arrival of antimicrobial drugs and even phagocytic
cells to the infection site. It can be harmful to health, as in the case of bacterial pellicles
that develop on teeth - originating cavities - and other problems related to mouth, lungs,
urinary catheters and contact lenses, which can originate serious infections on tissues
(osteomyelitis and endocarditis) and rejection to prosthetic material[42,43].
2.3. Heterofucans from S.filipendula versus biological activity
In this study we analysed five heterofucans from the seaweed S. filipendula regarding
their immunomodulating and antimicrobial capacity. On table 3, we attributed positive
(+) and negative (-) scores to identify which heterofucan presented the best performance
on the trials carried out in this project. We observed that the heterofucan SF2.0V
followed by SF1.5 presented the worst responses on the tests performed. On
counterpart, SF0.5V and SF0.7V were the most promising. In addition, we noticed that
none of the fucans is effective in all kinds of tests because each polymer presents a
unique structure responsible for the specificity of a response during the different trials.
In the future, it is intended to structurally characterize these fucans and propose
correlations between their structures and the activities they perform.
Table 3. Biological activity of heterofucans from S. filipendula score.

43
Telles, C.B.S. PPGCSA/CCS
3. Experimental Section
3.1. Materials
Bromide of 3- (4,5-dimetiltiazol-2-il) -2-5-diphenyltetrazoliumbromide (MTT),
Griess reagent, Histopaque 1077, Methanol PA, MCSF (Stimulating factor of
macrophages colony), medium of bacterial cultivation Luria Bertani (LB), Serum AB
(SIGMA) and metronidazole were acquired from Sigma Chemical Company, St. Louis,
MO, USA. Medium of cellular culture (RPMI 1640) (Developed by Roswell Park
Memorial Institute) and DMEM (Dulbecco's Modified Eagle's Medium), trypsin and
Bovine Fetal Serum (BFS), were obtained from CULTILAB (Campinas, Brazil). L-
glutamine, gentamicin, penicillin, streptomycin, sodium bicarbonate, HEPES, sodium
pyruvate and saline solution tamponed with phosphate buffered saline (PBS) were
acquired from Invitrogen Corporation (Burlington, ON, USA). ELISA Kits (for the
TNF-α, and IL-6) were acquired from BD Biosciences. Ficoll-Hypaque was acquired
from GE Healthcare. Panoptic dye was acquired from NewProv. Entellan was acquired
from Merck. Half Schneider’s insect was obtained from Gibco. All of the other solvents
and chemical products were obtained from analytic degree.
3.2. Biological material
The cell line of murine macrophages RAW 264.7 (ATCC number TIB-71) was
donated by Dr. Carmen Ferreira (Biochemistry department, UNICAMP, Brazil). The
protozoan Trichomonas vaginalis (ATCC number 30236) and Leishmania infatum were
given, respectively, by Dr. Tiana Tasca (Clinical Analysis and Toxicology Laboratory,
Faculdade de Farmácia, UFRGS, Brazil) and by Dr. Selma Jerônimo (Institute of
Tropical Medicine of Rio Grande do Norte, Biochemistry Department, UFRN, Brazil).
The bacterial strains Staphylococcus epidermidis (ATCC 35984) and the clinical
isolate of Klebsiella pneumoniae were given by Dr. Alexandre Jose Macedo
(Biotecnology center and Faculdade de Farmácia, UFRGS, Brazil).
3.3. Maintenance of cell lineages
The lineage of murine macrophages (RAW 264.7) were cultivated in supplemented
DMEM with 10% of Bovine Fetal Serum (BFS) and antibiotics (100 U / mL of
penicillin and 100 µg / mL of streptomycin). The cells were maintained as cultures in
monolayers at a humidified atmosphere of 5% of CO2 at 37 ° C.
Tricomonas vaginalis were cultivated in vitro in the TYM (trypticase-yeast extract-
maltose) medium, pH 6.0, supplemented with 10% (v/v) of serum inactivated by heat,
and incubated at 37 ° C [44]. The organisms in logarithmic growing phase, displaying

44
Telles, C.B.S. PPGCSA/CCS
more than 95% of viability and normal morphology were retracted, centrifuged and
suspended again in medium TYM for utilizing in tests.
The promastigotes of Leishmania Infantum were kept at 25oC in Schneider’s insect
medium, supplemented with 10% BFS, 200 IU/mL of penicillin and 200 µg/mL of
streptomycin, and grew until stagnant phase.
Staphylococcus epidermidis and the clinic isolate of Klebsiella pneumoniae (KPC)
were utilized as biofilm former bacterial models. S. epidermidis and KPC were
cultivated in Luria Bertani medium (LB) at 37 ° C under agitation of 150 rpm (Shaker
Série Excella E25; New Brunswick Scientific) and adjusted to an OD600 equivalent to
108 CFU / mL for utilization in antibacterial and antibiofilm trials.
3.4. Obtainment of peripheral human blood mononuclear cells
For the obtainment of peripheral human blood mononuclear cells (PBMC), 15 mL of
venous blood from six donors were obtained aseptically in heparinized tube. The tubes
containing the blood were diluted in sterile saline solution (v/v). The PBMCs were
obtained through the Ficoll-Hypaque gradient and adjusted to the concentration of 2 x
106 cells/mL in RPMI 1640 complete (100 L/mL of gentamicin, L-glutamine 2mM,
30mM HEPES), containing 10% of AB Serum (Life technologies GIBCO BRL,
Gaithersburg, MD).
After that the PBMCs were incubated at 37 ° C, 5% CO2 for 4 h in 6 and 24 well
plates, for the adhesion of the monocytes. To the 6 and 24 well plates, were added,
respectively, 2 and 3 mL/well of cell suspension. The 6 well plates were prepared with 4
cover slips round/well, for posterior microscopic analysis of the cells. After 4 h of
incubation, the medium - with the non-adherent cells - was removed. The wells were
washed with RPMI medium at room temperature. The adherent cells were incubated in
complete RPMI medium (10% of serum AB, 5 ng/mL of MCSF (Macrophage Colony
Stimulating Factor) for 6 days at 37 ° C, 5% of CO2.
3.5. Extraction of sulphated polysaccharides (Heterofucans)
The Phaeophyta Sargassum filipendula was collected at Búzios beach, Nísia
Floresta, Rio Grande do Norte, Brazil. The heterofucans SF0.5V, SF0.7V, SF1.0V,
SF1.5V and SF2.0V of S. filipendula were obtained utilizing the methodology described
by Costa and collaborators. [19].
3.6. Production of nitric oxide (NO)
The production of NO was analysed through quantification of nitrite production by
Griess reaction [45]. To measure the production of nitrite, aliquots of 100 µL - obtained

45
Telles, C.B.S. PPGCSA/CCS
from supernatants of the cultures to be dosed - were incubated with equal volume of
Griess reagent and were incubated at room temperature for 10 minutes. The analysis
was carried out in the microplate reader Multiskan Ascent (Thermo Labsystems,
Franklin, MA, USA) with absorbance to 540 nm. Utilizing as standard curve the
generated per NaNO2.
3.7. Cytokine analysis
RAW 264.7 (3 × 105 cells/mL) cells treated with the different heterofucans in the
concentrations of 0.125, 0.25, 0.5 mg/mL, were cultivated in 24 well plates. After 24 h,
the supernatants of the culture were collected. Following, they were centrifuged at 4000
rpm for 5 min. The levels of TNF-α and IL-6 were determined utilizing specific ELISA
kits (immunoabsorbent enzymatic test), the negative control consists of untreated cells
with the heterofucans and positive control those in the presence of LPS (2µg/mL). The
plate was read at 450 nm, with remediation at 570 and 590 nm.
3.8. Cytotoxicity tests in macrophages
The cytotoxicity in RAW 264.7 cells were measured through the MTT test as
described previously by Telles and collaborators [26]. The cells were cultivated in 96
well plates to a density of 5 x 103 cells/well with the heterofucans in different
concentrations (0.125, 0.25, 0.5 and 1.0 mg/mL) for 24 hours at 37 ° C and 5% of CO2.
After the incubation, 100 uL of MTT were added to each well, incubated during 4 h at
37 ° C and 5% of CO2, in the dark. The product MTT-formazano, dissolved in 100 mL
of ethanol was estimated through the measurement of the absorbance to 570 nm
Multiskan Ascent (Thermo Labsystems, Franklin, MA, USA) microplate reader.
3.9. Leishmanicidal activity
Cultures of Leishmania infantum at stagnant phase were resuspended in 1 mL of
complete RPMI medium and the concentration was adjusted to 107
parasites/mL. 6 well
plates cultivated with human macrophages were incubated with 3 mL of suspension of
parasite/well at 37 ° C, 5% of CO2 for 2 h for the infection of macrophages. The
medium containing parasites that did not adhere was retracted and the wells were
washed with RPMI medium at room temperature. The different heterofucans (0,5
mg/mL) were added in supplemented RPMI medium (10% of AB serum) to the wells
with macrophages infected with the parasite and the treatment was held for 24h.
To evaluate the percentage of macrophages infected with amastigotes of L. infatum,
the covers slips were removed after 24h, washed with PBS, fixed in methanol PA for 3
min at room temperature and dyed with Panotic. The plates were set with Entellan on

46
Telles, C.B.S. PPGCSA/CCS
clean plate. 200 total macrophages were counted and the quantity of amastigotes per
infected macrophage was counted.
3.10. Anti-T. vaginalis assay
Heterofucans from S. filipendula were analysed against T. vaginalis trophozoites
(ATCC 30236). In 96 well microplates, 50 µL/well of solutions containing the different
heterofucans and150 µL/well of the suspension of trophozoites were added, resulting in
a final volume of 200 µL containing 2,5 x 105 trophozoite/mL and 2.0 mg/mL of the
heterofucan to be tested. In control cultures, heterofucans samples were substituted by
distilled water. The plates were incubated for 24 h at 37 ° C. After that period, 20 µL of
a solution of resaurzurin at 0.1 mg/mL in Phosphate Buffered Saline was added in each
well. After 1 hour of incubation at 37° C the fluorescence of each well was read in a
fluorescence spectrophotometer (Spectramax Gemini XS – Molecular Devices
Cooperation, Sunnyvale, CA, USA), the quantification of viable trophozoites was
carried out as described by Duarte and collaborators [46].
3.11. Antibiofilm assay
Antibiofilm activity was measured as described by Melo-Silveira and collaborators
[47]. 80 uL of bacterial suspension (Staphylococcus epidermidis ATCC-35984 and a
clinic isolate of KPC Klebsiella pneumonia type 174), 80 µl of the heterofucans (0,5;
1,0; 1,5; and 2,0 mg/mL) and 40 mL of tryptone soy broth (TSB) (Oxoid Ltd., England)
were added to the 96 well plate and incubated (37 °C for 24 h). The rest of the adhered
bacteria was fixed at 60 ° C for 1h. The biofilm formed was dyed with 0.4% of crystal
violet for 15 minutes at room temperature. The crystal violet bonded to the cells/biofilm
was solubilized with 99,5% of DMSO (Sigma-Aldrich Co., St. Louis, MO, USA) and
read at 570 nm (Spectramax M2e multimode Microplate Reader, Molecular Devices,
Sunnyvale, CA , USA). The controls were considered as 100% of the formation of
biofilm and the values obtained for the extract were the average of three experiments.
3.12. Bacterial inhibition assay
The experiment was carried out as described by Melo-Silveira e collaborators [47].
The bacterial growth of S. epidermidis (ATCC 35984) and a clinic isolate of KPC was
evaluated through the difference of absorbance measured at DO 600 nm in the end and
in the beginning of the incubation time, in 96 well polystyrene microtitration plates.
Different concentration of the heterofucans (0.5; 1.0; 1.5; and 2.0 mg/mL) were
incubated in the presence of each bacterial strain. The control with distilled water was
considered as 100% of bacterial growth. All of the experiments were carried out in
triplicate.

47
Telles, C.B.S. PPGCSA/CCS
3.13. Statistical analysis
All the data were expressed in average ± standard deviation. The analysis was carried
out by analysis of variance. Student-Newman-Keuls post-tests were done for multiple
comparison by group. In all the cases, statistical significance was established at p <0,05.
4. Conclusions
The heterofucans from S. filipendula presented distant activities as stimulators of the
immune system and antimicrobial agents. The heterofucans SF0.5V, SF0.7V and
SF1.0V were able to act in the activation of murine and human macrophages promoting
increase in the release of the chemical mediators that are important for fighting
intracellular parasites. In addition, SF0.5V presented antibiofilm activity facing the
strain of S. epidermides whereas SF0.7V and 1.0V inhibited almost completely the
survival of the protozoan T. vaginalis.
All five heterofucans obtained from the seaweed S. filipendula presented
different/specific levels of activities in the trials carried out, making again evident that
their biological activities depend on their structural characteristics. Results such as this
reflect the great spectrum of action of these sulphated polysaccharides, but also
intensify the need of posterior studies looking to elucidate the complete structure of
these polysaccharides, configuration of glycosidic bonds, their position as well as the
position of sulphate groups and ramification points.
Acknowledgments
Research was supported by Ministério de Ciência, Tecnologia e Informação
(MCTI), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)
and Conselho Nacional de Desenvolvimento Cientifico e Tecnologico (CNPq),
Brazil. Hugo A O Rocha is a CNPq fellowship honored researcher. Cinthia Telles
had a Ph.D. scholarship from CAPES.
Author Contributions
Conceived and designed the experiments: C.B.S.T, T.T, L.S.C and H.A.O.R.
Performed the experiments: C.B.S.T., C.M.A and A.P.F. Analyzed the data: C.B.S.T. and
H.A.O.R. Contributed reagents/materials/analysis tools: G.P.F, L.S.C., T.T., S.J., L.F.D.P
and H.A.O.R. Wrote the paper: C.B.S.T. and H.A.O.R.
Conflict of Interest
The authors declare no conflict of interest.

48
Telles, C.B.S. PPGCSA/CCS
References
1. Machado PRL, Carvalho L, Araújo MIAS, Carvalho EM. Immune response
mechanisms to infections. An bras Dermatol, Rio de Janeiro, 2004; 79(6):647-
664.
2. Coelho-Castelo AAM, Trombone APF, Rocha CD, Lorenzi JCC. Immune
response to infectious diseases. Medicina (Ribeirão Preto) 2009;42(2): 127-42.
3. Abbas A K, Lichtman A H, Pober JS. Imunologia celular e molecular. 5a ed.,
Rio de Janeiro, Elsevier, 2005. 580 p.
4. Janeway C A, Jr, Travers P, Walport M, Shlomchik M J. The Immune System in
Health and Disease. Immunobiology, 5th edition. 2001. New York: Garland
Science.
5. Chouhan G, Islamuddin M, Sahal D, Afrin F. Exploring the role of medicinal
plant-based immunomodulators for effective therapy of leishmaniasis. Front
Immunol. 2014; 5:193.
6. Helms D.J., Mosure D.J., Secor W.E., Workowski K.A. Management of
Trichomonas vaginalis in women with suspected metronidazole hypersensitivity.
Am. J. Obstet. Gynecol. 2008; 198: 370–7.
7. Innocente AM, Vieira Pde B, Frasson AP, Casanova BB, Gosmann G, Gnoatto
SC, Tasca T. Anti-Trichomonas vaginalis activity from triterpenoid derivatives.
Parasitol Res. 2014; 113:2933-40.
8. Limban C, Chifiriuc MC. Antibacterial activity of new dibenzoxepinone oximes
with fluorine and trifluoromethyl group substituents. Int J Mol Sci.
2011;12(10):6432-44.
9. Uthaisangsook S, Day NK, Bahna SL, Good RA, Haraguchi S. Innate immunity
and its role against infections. Ann Allergy Asthma Immunol. 2002
Mar;88(3):253-64; quiz 265-6, 318.
10. Altmann KH. Microtubule-stabilizing agents: a growing class of important
anticancer drugs. Curr Opin Chem Biol. 2001 5: 424-431.
11. Albuquerque I R, Queiroz K C, Alves LG, Santos EA, Leite EL, Rocha H A.
Heterofucans from Dictyota menstrualis have anticoagulant activity. Braz J Med
Biol Res. 2004, 37, 167-71.
12. Hoshino T, Hayashi T, Hayashi K, Hamada J, Lee JB. An antiviral active
sulfated polysaccharide from Sargassum horneri (Tuner) C. Agardh. Biol.
Pharm. Bull. 1998, 21, 730–734.
13. Sinha, S.; Astani, A.; Ghosh, T.; Schnitzler, P.; Ray, B. Polysaccharides from
Sargassum tenerrimum: structural features, chemical modification and anti-viral
activity. Phytochemistry 2010, 71, 235–242.

49
Telles, C.B.S. PPGCSA/CCS
14. Zhu, W.; Ooi, V.E.C; Chan, P.K.S.; Ang, P.O.J. Isolation and characterization of
a sulfated polysaccharide from the brown alga Sargassum patens and
determination of its anti-herpes activity. Biochem. Cell Biol. 2003, 81, 25–33.
15. Stevan, F.R.; Oliveira, M.B.; Bucchi, D.F.; Noseda, M.D.; Iacomini, M., Duarte,
M.E. Cytotoxic effects against HeLa cells of polysaccharides from seaweeds. J.
Submicrosc. Cytol. Pathol. 2001, 33, 477–484.
16. Josephine, A.; Veena, C.K.; Amudha, G.; Preetha, S.P.; Sundarapandian, R.;
Varalakshmi, P. Sulphated polysaccharides: new insight in the prevention of
cyclosporine A-induced glomerular injury. Basic Clin. Pharmacol. Toxicol.
2007, 101, 9–15.
17. Dore CM, das C Faustino Alves MG, Will LS, Costa TG, Sabry DA, de Souza
Rêgo LA, Accardo CM, Rocha HA, Filgueira LG, Leite EL. A sulfated
polysaccharide, fucans, isolated from brown algae Sargassum vulgare with
anticoagulant, antithrombotic, antioxidant and anti-inflammatory effects.
Carbohydr Polym. 2013 Jan 2;91(1):467-75.
18. Vasquez RD, Ruby SP, Garcia-Meim, Ramos JDA. Polysaccharide extracts from
sargassum siliquosum j.g. Agardh modulates production of pro-inflammatory
cytokines in lps-induced pbmc and delays coagulation time in-vitro. Jour.
Harmo. Res. Pharm., 2014, 3(3), 101-112
19. Costa LS, Fidelis GP, Telles CB, Dantas-Santos N, Camara RB, Cordeiro SL,
Costa MS, Almeida-Lima J, Melo-Silveira RF, Oliveira RM, Albuquerque IR,
Andrade GP, Rocha HA. Antioxidant and antiproliferative activities of
heterofucans from the seaweed Sargassum filipendula. Mar Drugs.
2011;9(6):952-66.
20. Costa LS, Telles CB, Oliveira RM, Nobre LT, Dantas-Santos N, Camara RB,
Costa MS, Almeida-Lima J, Melo-Silveira RF, Albuquerque IR, Leite EL,
Rocha HA. Heterofucan from Sargassum filipendula induces apoptosis in HeLa
cells. Mar Drugs. 2011;9(4):603-14.
21. Gamal-Eldeen AM, Amer H, Helmy WA, Talaat RM, Ragab H. Chemically-
modified polysaccharide extract derived from Leucaena leucocephala alters Raw
264.7 murine macrophage functions. Int Immunopharmacol. 2007 Jun;7(6):871-
8.
22. Jiang Z, Okimura T, Yamaguchi K, Oda T. The potent activity of sulfated
polysaccharide, ascophyllan, isolated from Ascophyllum nodosum to induce
nitric oxide and cytokine production from mouse macrophage RAW264.7 cells:
Comparison between ascophyllan and fucoidan. Nitric Oxide. 2011 Nov
30;25(4):407-15.
23. Leiro JM, Castro R, Arranz JA, Lamas J. Immunomodulating activities of acidic
sulphated polysaccharides obtained from the seaweed Ulva rigida C. Agardh. Int
Immunopharmacol. 2007 Jul;7(7):879-88.

50
Telles, C.B.S. PPGCSA/CCS
24. Jiang Z, Ueno M, Nishiguchi T, Abu R, Isaka S, Okimura T, Yamaguchi K, Oda
T. Importance of sulfate groups for the macrophage-stimulating activities of
ascophyllan isolated from the brown alga Ascophyllum nodosum. Carbohydr
Res. 2013 Oct 18;380:124-9.
25. Haroun-Bouhedja F, Ellouali M, Sinquin C, Boisson-Vidal C. Relationship
between sulfate groups and biological activities of fucans. Thromb Res. 2000
Dec 1;100(5):453-9.
26. Telles CBS, Sabry DA, Almeida-Lima J, Costa MSSP, Melo-Silveira RF,
Trindade ES, Sassaki GL, Wisbeck E, Furlan SA, Leite EL, Rocha HAO.
Sulfation of the extracellular polysaccharide produced by the edible
mushroomPleurotus sajor-caju alters its antioxidant, anticoagulant and
antiproliferativeproperties in vitro. Carbohydrate Polymers 2011 85, 514–521.
27. Barahona T, Encinas MV, Mansilla A, Matsuhiro B, Zúñiga EA. A sulfated
galactan with antioxidant capacity from the green variant of tetrasporic Gigartina
skottsbergii (Gigartinales, Rhodophyta). Carbohydr Res. 2012 Jan
10;347(1):114-20.
28. Liang W, Mao X, Peng X, Tang S. Effects of sulfate group in red seaweed
polysaccharides on anticoagulant activity and cytotoxicity. Carbohydr Polym.
2014 Jan 30;101:776-85.
29. Albuquerque IR, Cordeiro SL, Gomes DL, Dreyfuss JL, Filgueira LG, Leite EL,
Nader HB, Rocha HA. Evaluation of anti-nociceptive and anti-inflammatory
activities of a heterofucan from Dictyota menstrualis. Mar Drugs. 2013 Aug
2;11(8):2722-40.
30. Nakayasu S1, Soegima R, Yamaguchi K, Oda T. Biological activities of fucose-
containing polysaccharide ascophyllan isolated from the brown alga
Ascophyllum nodosum. Biosci Biotechnol Biochem. 2009 Apr 23;73(4):961-4.
31. Do H, Kang N-S, Pyo S, Billiar T R., Sohn E-H. Differential Regulation by
Fucoidan of IFN-g-Induced NO Production in Glial cells and Macrophages.
Journal of Cellular Biochemistry 2010, 111:1337–1345.
32. Nakamura T, Suzuki H, Wada Y, Kodama T, Doi T. Fucoidan induces nitric
oxide production via p38 mitogen-activated protein kinase and NF-kappaB-
dependent signaling pathways through macrophage scavenger receptors.
Biochem Biophys Res Commun. 2006 Apr 28;343(1):286-94.
33. Teruya T, Takeda S, Tamaki Y, Tako M. Fucoidan isolated from Laminaria
angustata var. longissima induced macrophage activation. Biosci Biotechnol
Biochem. 2010;74(9):1960-2.
34. Cho M, Lee DJ, Kim JK, You S. Molecular characterization and
immunomodulatory activity of sulfated fucans from Agarum cribrosum.
Carbohydr Polym. 2014 Nov 26;113:507-14.

51
Telles, C.B.S. PPGCSA/CCS
35. Kar S, Sharma G, Das PK. Fucoidan cures infection with both antimony-
susceptible and -resistant strains of Leishmania donovani through Th1 response
and macrophage-derived oxidants. J Antimicrob Chemother. 2011
Mar;66(3):618-25.
36. Sharafi SM, Yousefi M, Yousefi HA, Asghari G, Darani HY. In vitro effects of
various plants extracts on the growth of Trichomonas vaginalis. Infect Disord
Drug Targets. 2013 Oct;13(5):322-7.
37. Helms DJ, Mosure DJ, Secor WE, Workowski KA. Management of trichomonas
vaginalis in women with suspected metronidazole hypersensitivity. Am J Obstet
Gynecol. 2008 Apr;198(4):370.
38. Blaha C, Duchêne M, Aspöck H, Walochnik J. In vitro activity of
hexadecylphosphocholine (miltefosine) against metronidazole-resistant and -
susceptible strains of Trichomonas vaginalis. J Antimicrob Chemother. 2006
Feb;57(2):273-8.
39. Gottenbos B, van der Mei HC, Busscher HJ. Models for studying initial
adhesion and surface growth in biofilm formation on surfaces. Methods
Enzymol. 1999;310:523-34.
40. Gadenne V, Lebrun L, Jouenne T, Thebault P. Antiadhesive activity of ulvan
polysaccharides covalently immobilized onto titanium surface. Colloids Surf B
Biointerfaces. 2013 Dec 1;112:229-36.
41. Zinger-Yosovich KD, Gilboa-Garber N. Blocking of Pseudomonas aeruginosa
and Ralstonia solanacearum Lectins by plant and microbial branched
polysaccharides used as food additives. J Agric Food Chem. 2009 Aug
12;57(15):6908-13.
42. Bjarnsholt T. The role of bacterial biofilms in chronic infections. APMIS Suppl.
2013 May;(136):1-51.
43. Mohammadi Z, Mashouf RY, Jafarzadeh H, Heydari A, Shalavi S. Effect of
apically separated rotary instruments on bacterial leakage of obturated root
canals using resilon. N Y State Dent J. 2015 Mar;81(2):42-6.
44. Diamond LS. The establishment of various trichomonads of animals and man in
axenic cultures. J Parasitol. 1957 Aug;43(4):488-90.
45. Green LC, Wagner DA, Glogowski J, Skipper PL, Wishnok JS, Tannenbaum
SR. Analysis of nitrate, nitrite and nitrate in biological fluids. Anal Biochem
1982. 126: 131–138.
46. Duarte M, Giordani RB, De Carli GA, Zuanazzi JA, Macedo AJ, Tasca T. A
quantitative resazurin assay to determinate the viability of Trichomonas
vaginalis and the cytotoxicity of organic solvents and surfactant agents. Exp
Parasitol. 2009; 123: 195-8.
47. Melo-Silveira RF, Fidelis GP, Costa MS, Telles CB, Dantas-Santos N, de
Oliveira Elias S, Ribeiro VB, Barth AL, Macedo AJ, Leite EL, Rocha HA. In

52
Telles, C.B.S. PPGCSA/CCS
vitro antioxidant, anticoagulant and antimicrobial activity and in inhibition of
cancer cell proliferation by xylanextracted from corn cobs. Int J Mol Sci. 2012;
13: 409-26.

53
Telles, C.B.S. PPGCSA/CCS
5.1. ARTIGO 2 (PUBLICADO)

72
Almeida-Lima J. PPGCSA/CCS
6. COMENTÁRIOS, CRÍTICAS E SUGESTÕES
Há décadas nosso grupo de pesquisa tem realizado uma estensiva
busca de atividades biológicas/farmacológicas de polissacarídeos sulfatados
obtidos de macroalgas marinhas. Fucanas, obtidas de diferentes espécies de
algas marrons são objetivos de estudo de pesquisas do grupo, dentre elas, as
fucanas obtidas da macroalga marron Sargassum filipendula. No entando,
apesar de algumas atividades biolólogicas já terem sido descritas para esses
polímeros nenhum estudo sobre atividades antimicrobina foi realizado. Este
trabalho de pesquisa teve como ponto positivo a insersão, na gama de ensaios
biológicos realizados pelo grupo, novas metodologias a serem utilizadas no
laboratório (BIOPOL), permitindo a apliação da detecção de outros prováveis
efeitos biológicos das fucanas.
Um momento importante durante a realização do doutorado foi a
possibilidade de realizar doutorado-sanduíche durante seis meses na
Universidade Nova de Lisboa, Instituto de Higiene e Medicina Tropical, Unidade
Clínica de Doenças Tropicais. A orientação do Prof. Dr. Marcelo Sousa Silva e
os integrantes do seu grupo de pesquisa forneceram total apoio para o
desenvolvimento do estágio. No entanto, não adicionamos os resultados das
heterofucanas de S.filipendula frente as metaloproteinases de Trypanosoma
brucei, pois alguns empaces de cunho técnico durante o estabelecimento de
uma das metadologias não permitiu a confirmação dos resultados alcançados.
Ao longo do desenvolvimento desta pesquisa, o cronograma de
atividades não foi totalemente realizado, pois, inicialmente tinha-se como meta
o sub-fracionamento da heterofucana mais eficiente nos ensaios realizados,
mas diante de vários intercursos ao londo do período final do doutorado
(terceiro ano), como o nascimento do meu filho, sendo necessário o
afastamento durante o período de licença maternidade, e o início na carreira
profissional, assumindo o cargo de professora, não obtive tempo hábil para
cumprimento dessa meta.

73
Telles, C.B.S. PPGCSA/CCS
Durante o desenvolvimento deste estudo tive a oportunidade de
aprimorar bastante meus conhecimentos, pois tive a chance de aperfeiçoar das
técnicas laboratórias em diversos ramos da pesquisa científica, como:
imunologia, farmacologia, biologia celular, bioquímica, dentre tantos outros.
Além de interagir com vários grupos de pesquisas como o já citado em
Lisboa/Portugal sob a orientação do Professor Marcelo Sousa Silva, o grupo da
professora Tiana Tasca em Porto Alegre e, vários grupos de pesquisa
localizados na própria UFRN como os das professoras Selma Jerônimo e Katia
Scortecci.
Com relação à produção científica até o momento tenho 6 artigos
publicados em revista científicas, além de 10 resumos publicados em
congressos científicos durante o período do doutorado. Além disso, também fiz
parte na elaboração do capítulo de livro - Carragenan - e de uma banca
examinadora de trabalho de conclusão de curso (monografia).
Após a conclusão dessa etapa acadêmica, tenho como projeto para o
futuro, continuar com os estudos na mesma linha de pesquisa, em um futuro
pós-doutorado. Além de buscar a aprovação como Professora de Instituições
de Ensino Superior, engajando na tríade ensino/pesquisa/extensão.

74
Telles, C.B.S. PPGCSA/CCS
7. REFERÊNCIAS
1. Works HI. Understanding the Immune System How It Works referencia.
Disponível em: http://www.lupusny.org/sites/default/files/understanding-
immune-system.pdf. Accesso em: Apr 05, 2015.
2. Parkin J, Cohen B. An overview of the immune system. Lancet. 2001 Jun
2;357(9270):1777-89.
3. Lawrence T, Fong C. The resolution of inflammation: anti-inflammatory
roles for NF-kappaB. Int J Biochem Cell Biol. 2010 Apr;42(4):519-23.
4. Janeway C, Travers P, Walport M, Shlomchik M. 2005. Immunobiology:
The Immune System in Health and Disease. New York, New York:
Garland Science Publishing. p. 23, 80-84.
5. Hung ND, Kim MR, Sok D-E. Mechanisms for anti-inflammatory effects of
1-[15(S)-hydroxyeicosapentaenoyl] lysophosphatidylcholine,administered
intraperitoneally, in zymosan A-induced peritonitis. Br J Pharmacol. 2011
Mar; 162(5): 1119–1135.
6. Mueller M, Hobiger S, Jungbauer A. Anti-inflammatory activity of extracts
from fruits, herbs and spices. Food Chem. 2010; 122(4): 987-996.
7. Anthony RM, Rutitzky LI, Urban JF, Jr., Stadecker MJ, Gause WC.
Protective immune mechanisms in helminth infection. Nat. Rev.Immunol.
2007; 7: 975–987.
8. Sakaguchi S, Miyara M, Costantino CM, Hafler DA. FOXP3+ regulatory T
cells in the human immune system. Nat. Ver. Immunol. 2010; 10: 490-
500.
9. Suvas S., Azkur A. K., Kim B. S., Kumaraguru U., Rouse B.T. CD4+
CD25+ regulatory T cells control the severity of viral immunoinflammatory
lesions. J. Immunol. 2004; 172: 4123–32.
10. Korn T, Bettelli E, Oukka M, Kuchroo VK. IL-17 and Th17 Cells. Annu
Rev Immunol. 2009; 27: 485-517.
11. Chouhan G, Islamuddin M, Sahal D, Afrin F. Exploring the role of
medicinal plant-based immunomodulators for effective therapy of
leishmaniasis. Front Immunol. 2014; 5:193.
12. Desjeux P. Leishmaniasis. Nat Rev Microbiol. 2004 Sep; 2(9):692.

75
Telles, C.B.S. PPGCSA/CCS
13. Desjeux P. Leishmaniasis. Public health aspects and control. Clin
Dermatol. 1996; 14: 417-23.
14. Soong L, Henard CA, Melby PC. Immunopathogenesis of non-healing
American cutaneous leishmaniasis and progressive visceral
leishmaniasis. Semin Immunopathol. 2012; 34: 735-51.
15. Cunningham AC. Parasitic adaptive mechanisms in infection by
leishmania. Exp Mol Pathol. 2002; 72: 132-41.
16. Qadoumi M, Becker I, Donhauser N, Rollinghoff M, Bogdan C.
Expression of inducible nitric oxide synthase in skin lesiono f patients
with american cutaneous leishmaniasis. Infect Immun. 2002; 70: 4638-
42.
17. Sutterwala FS, Noel GJ, Clynes R, Mosser DM. Selective supression of
interleukin-12 induction after macrophage receptor ligation. J Exp Med.
1997; 185:1977-85.
18. Fiocruz. As Leishmanioses. 2010.
http://www.dbbm.fiocruz.br/tropical/leishman/leishext/html/tratamento.ht
m. Acesso em 08 de março.
19. Cruz A K, de Toledo JS, Falade M, Terrão MC, Kamchonwongpaisan S,
Kyle DE, Uthaipibull C. Current treatment and drug discovery against
Leishmania spp. and plasmodium spp.: A review. Curr Drug Targets, v.
10, p. 178–192, 2009.
20. Sharafi SM, Yousefi M, Yousefi HA, Asghari G, Darani HY. In vitro effects
of various plants extracts on the growth of Trichomonas vaginalis. Infect
Disord Drug Targets. 2013; 13:322-7.
21. Fichorova RN. Impact of T. vaginalis infection on innate immune
responses and reproductive outcome. J Reprod Immunol. 2009
Dec;83(1-2):185-9.
22. Helms D.J., Mosure D.J., Secor W.E., Workowski K.A. Management of
Trichomonas vaginalis in women with suspected metronidazole
hypersensitivity. Am. J. Obstet. Gynecol. 2008; 198: 370–7.
23. Innocente AM, Vieira Pde B, Frasson AP, Casanova BB, Gosmann G,
Gnoatto SC, Tasca T. Anti-Trichomonas vaginalis activity from
triterpenoid derivatives. Parasitol Res. 2014; 113:2933-40.

76
Telles, C.B.S. PPGCSA/CCS
24. Machado PRL, Carvalho L, Araújo MIAS, Carvalho EM. Immune
response mechanisms to infections. An bras Dermatol, Rio de Janeiro,
2004; 79(6):647-664.
25. Coelho-Castelo AAM, Trombone APF, Rocha CD, Lorenzi JCC. Immune
response to infectious diseases. Medicina (Ribeirão Preto) 2009;42(2):
127-42.
26. Sheikh NA, al-Shamisi M, Morrow WJ. Delivery systems for molecular
vaccination. Curr Opin Mol Ther. 2000;2:37–54.
27. Mohammed KA, Nasreen N, Ward MJ, Antony VB. Induction of acute
pleural inflammation by Staphylococcus aureus. I. CD4+ T cells play a
critical role in experimental empyema. J Infect Dis. 2000
May;181(5):1693-9.
28. Uthaisangsook S, Day NK, Bahna SL, Good RA, Haraguchi S. Innate
immunity and its role against infections. Ann Allergy Asthma Immunol.
2002 Mar;88(3):253-64; quiz 265-6, 318.
29. Abbas A K, Lichtman A H, Pober JS. Imunologia celular e molecular. 5a
ed., Rio de Janeiro, Elsevier, 2005. 580 p.
30. Feng N, Hongnan J, Wang M, Du C, Wrigth JA, Young AH. Antitumor
activity of Virulizin, a novel biological response modifier (BRM) in a panel
of human pancreatic cancer and melanoma xenografts. Cancer
Chemotherapy and Pharmacology, Berlin, v.51, p.247–255, 2003.
31. Poleshko A, Palagin I, Zhang R, Boimel P, Castagna C, Adams PD,
Skalka AM, Katz RA. Identification of cellular proteins that maintain
retroviral epigenetic silencing: evidence for an antiviral response. Journal
of Virology, Washington, v.82, n.5, p. 2313–2323, 2008.
32. El-Hasan A, Walker F, Schone J, Buchenauer H. Detection of
viridiofungin A and other antifungal metabolites excreted by Trichoderma
harzianumactive against different plant pathogens. European Journal of
Plant Pathology, Dordrecht, v.124, p.457–470, 2009.
33. Vazquéz-Lopez C, Armas-Serra C, Bernardina W, Rodriguez-Caabeiro
F. Oral inoculation with Gymnorhynchus gigas induces anti-parasite
anapyhylactic antibody production in both mice and rats and adverse

77
Telles, C.B.S. PPGCSA/CCS
reactions in challenge mice. International Journal of Food Microbiology,
v.64, p.307–315, 2001.
34. Hu J-F, Garo E, Hough GW, Goering MG, O`Neil-Johnson M, Eldridge
GR. Antibacterial, partially acetylated oligorhamnosides from
Cleistopholis patens. Journal of Natural Products, Cincinnati, v.69, p.585-
590, 2006.
35. Mendes SF, Santos JRO, Barbosa AM, Vasconcelosa AFD, Aranda-
Selverioa G, Monteiro NK, Dekkerc RFH, Pereira MS, Tovard AMF,
Mourão PAS, Silvam LC. Sulfonation and anticoagulant activity of
botryosphaeran from Botryosphaeriarhodina MAMB-05 grown on
fructose. International ournal of Biological Macromolecules, Guildford,
v.45, p.305-309, 2009.
36. Bohn J A, Bemiller J N. (13)β-d-glucans as biological response
modifiers: a review of structure-functional activity relationships.
Carbohydr. Polym. 1995; v. 28, p. 3–14.
37. Albuquerque I R, Queiroz K C, Alves LG, Santos EA, Leite EL, Rocha H
A. Heterofucans from Dictyota menstrualis have anticoagulant activity.
Braz J Med Biol Res, 37, 167-71 2004.
38. Costa LS, Fidelis GP, Cordeiro SL, Oliveira RM, Sabry DA, Câmara RB,
Nobre LT, Costa MS, Almeida-Lima J, Farias EH, Leite EL, Rocha HA.
Biological activities of sulfated polysaccharides from tropical seaweeds.
Biomed Pharmacother 2010; 64(1):21-8
39. Almeida-Lima, J.; Costa, L.S.; Silva, N.B.; Melo-Silveira, R.F.; Silva, F.V.;
Felipe, M.B.; Medeiros, S.R.; Leite, E.L.; Rocha, H.A. Evaluating the
possible genotoxic, mutagenic and tumor cell proliferation-inhibition
effects of a non-anticoagulant, but antithrombotic algal heterofucan. J
Appl Toxicol. 2010, 30, 708–715.
40. Melo-Silveira R F, Fidelis G P, Costa M S S P, Telles C B S, Dantas-
Santos N, Elias S O, Ribeiro V B, Barth A L, Macedo A J, Leite E L,
Rocha H A O. In Vitro Antioxidant, Anticoagulant and Antimicrobial
Activity and in Inhibition of Cancer Cell Proliferation by Xylan Extracted
from Corn Cobs. Int. J. Mol. Sci. 2012, 13, 409-426;

78
Telles, C.B.S. PPGCSA/CCS
41. Leite EL, Medeiros MGL, Rocha HAO, Farias GGM, Silva LF, Chavante
SF, et al. Structure of a new fucan from the algae Spatoglossum
schröederi. Plant Sci 1998; 132: 215-28.
42. Széchy MTM, Galliez M, Marconi M. Quantitative variables applied to
phenological studies of Sargassum vulgare C. Agardh (Phaeophyta-
Fucales) from Ilha Grande Bay, State of Rio de Janeiro. Revista
Brasileira de Botânica, 29 (1): 27-37.
43. Hoshino T, Hayashi T, Hayashi K, Hamada J, Lee JB. 1998. An antiviral
active sulfated polysaccharide from Sargassum horneri (Tuner) C.
Agardh. Biol. Pharm. Bull. 21, 730–734.
44. Sinha, S.; Astani, A.; Ghosh, T.; Schnitzler, P.; Ray, B. Polysaccharides
from Sargassum tenerrimum: structural features, chemical modification
and anti-viral activity. Phytochemistry 2010, 71, 235–242.
45. Zhu, W.; Ooi, V.E.C; Chan, P.K.S.; Ang, P.O.J. Isolation and
characterization of a sulfated polysaccharide from the brown alga
Sargassum patens and determination of its anti-herpes activity. Biochem.
Cell Biol. 2003, 81, 25–33.
46. Stevan, F.R.; Oliveira, M.B.; Bucchi, D.F.; Noseda, M.D.; Iacomini, M.,
Duarte, M.E. Cytotoxic effects against HeLa cells of polysaccharides
from seaweeds. J. Submicrosc. Cytol. Pathol. 2001, 33, 477–484.
47. Josephine, A.; Veena, C.K.; Amudha, G.; Preetha, S.P.;
Sundarapandian, R.; Varalakshmi, P. Sulphated polysaccharides: new
insight in the prevention of cyclosporine A-induced glomerular injury.
Basic Clin. Pharmacol. Toxicol. 2007, 101, 9–15.
48. Dore CM, das C Faustino Alves MG, Will LS, Costa TG, Sabry DA, de
Souza Rêgo LA, Accardo CM, Rocha HA, Filgueira LG, Leite EL. A
sulfated polysaccharide, fucans, isolated from brown algae Sargassum
vulgare with anticoagulant, antithrombotic, antioxidant and anti-
inflammatory effects. Carbohydr Polym. 2013 Jan 2;91(1):467-75.
49. Vasquez RD, Ruby SP, Garcia-Meim, Ramos JDA. Polysaccharide
extracts from sargassum siliquosum j.g. Agardh modulates production of
pro-inflammatory cytokines in lps-induced pbmc and delays coagulation
time in-vitro. Jour. Harmo. Res. Pharm., 2014, 3(3), 101-112

79
Telles, C.B.S. PPGCSA/CCS
50. Seaweed Industry Association. Disponível em:
https://seaweedindustry.com/. Acesso em: Abr, 10, 2014.
51. Costa LS, Fidelis GP, Telles CB, Dantas-Santos N, Camara RB,
Cordeiro SL, Costa MS, Almeida-Lima J, Melo-Silveira RF, Oliveira RM,
Albuquerque IR, Andrade GP, Rocha HA. Antioxidant and
antiproliferative activities of heterofucans from the seaweed Sargassum
filipendula. Mar Drugs. 2011;9(6):952-66.
52. Costa LS, Telles CB, Oliveira RM, Nobre LT, Dantas-Santos N, Camara
RB, Costa MS, Almeida-Lima J, Melo-Silveira RF, Albuquerque IR, Leite
EL, Rocha HA. Heterofucan from Sargassum filipendula induces
apoptosis in HeLa cells. Mar Drugs. 2011;9(4):603-14.
53. Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F.
Colorimetric method for determination of sugars, and related substances.
Anal Chem 1956; 28:350–356.
54. Dodgson K S, Price RG. A note on the determination of the ester
sulphate content of sulphated polysaccharides. Biochem J, 84, 106-10
1962.
55. Spector T. Refinement of the coomassie blue method of protein
quantitation. A simple and linear spectrophotometric assay for less than
or equal to 0.5 to 50 microgram of protein. Anal Biochem, 86, 142-6 1978.

80
Telles, C.B.S. PPGCSA/CCS





























