Embed Size (px)
Citation preview

UNIVERSITA’ DEGLI STUDI DI PERUGIA
FACOLTA’ DI MEDICINA VETERINARIA
DIPARTIMENTO DI PATOLOGIA DIAGNOSTICA E CLINICA
VETERINARIA
SEZIONE DI SCIENZE SPERIMENTALI E BIOTECNOLOGIE APPLICATE
CORSO DI LAUREA MAGISTRALE IN MEDICINA VETERINARIA
TESI DI LAUREA
PESTE DEI PICCOLI RUMINANT I(PPR) NEL SAHARA
OCCIDENTALE: indagine epidemiologica nelle Wilaias e nei “Territori Liberati”della Repubblica Democratica Araba
Saharawi (RASD).
PESTE DES PETITS RUMINANTS (PPR) IN WESTERN
SAHARA: a epidemiological survey among Wilaias and “Liberated Territories” of the Saharawi Arab Democratic
Rebublic (SARD).
Settore Scientifico Disciplinare VET 05
LAUREANDO RELATORE
Sabatini Laura Prof. Passamonti Fabrizio
ANNO ACCADEMICO 2008-2009

INDICE
ABSTRACT pag. 4
RIASSUNTO pag. 6
1. INTRODUZIONE pag. 8
2. CARATTERISTICHE GENERALI DELLA PPR pag. 11
2.1 EZIOLOGIA pag. 11
2.2 EPIDEMIOLOGIA E TRASMISSIONE pag. 16
2.3 SEGNI CLINICI E LESIONI ANATOMOPATOLOGICHE pag. 20
2.4 DIAGNOSI pag. 24
2.4.1 DIAGNOSI DIFFERENZIALE pag. 25
2.5 PREVENZIONE E CONTROLLO pag. 28
3. PARTE SPERIMENTALE pag. 31
3.1 AREA DI STUDIO pag. 31
3.1.1 STORIA E SITUAZIONE POLITICA pag. 32
3.1.2 IL POPOLO SAHARAWI E LA SUA STORIA pag. 34
3.1.3 I CAMPI PROFUGHI SAHARAWI OGGI pag. 39
3.2 POPOLAZIONE ED ECONOMIA pag. 42
3.2.1 TECNICHE DI ALLEVAMENTO pag. 46
3.2.2 I SERVIZI VETERINARI SAHARAWI pag. 52
3.3 ASPETTI CLIMATICI E TERRITORIALI pag. 53
3.4 MATERIALI E METODI pag. 58
3.4.1 PIANO DI CAMPIONAMENTO pag. 59
2

3.4.2 ELABORAZIONE STATISTICA DEL CAMPIONE DI
STUDIO pag. 61
3.4.3 SCREENING ANTICORPALE NELLA RASD pag. 64
3.4.4 PROTOCOLLO ELISA–COMPETITIVA pag. 65
4. RISULTATI pag. 70
4.1 ANALISI DI SIEROPREVALENZA pag. 71
4.1.1 SIEROPREVALENZA GENERALE pag. 71
4.1.2 SIEROPREVALENZA PER SINGOLE REGIONI pag. 73
4.1.3 SIEROPREVALENZA PER CLUSTER pag. 75
4.2 ANALISI DI CORRELAZIONE pag. 78
4.2.1 CORRELAZIONE PER SPECIE ANIMALE pag. 78
4.2.2 CORRELAZIONE PER ETA’ DEI SOGGETTI pag. 79
4.2.3 CORRELAZIONE PER SESSO DEI SOGGETTI pag. 80
4.2.4 FUNZIONE DI CORRELAZIONE pag. 80
4.3 ANALISI DI MODELLAZIONE SPAZIALE pag. 82
5. CONCLUSIONI pag. 84
BIBLIOGRAFIA pag. 86
3

ABSTRACT
Peste des Petitis Ruminants (PPR) is an acute viral disease affecting small
ruminants and characterized by high morbidity and mortality. Up to today it is
considered a tropical disease because of its major diffusion in sub-saharian Africa,
Middle East and south of Asia, but ecological and environmental changes that
have taken place in these last years and the intensification of the animal trade
could favour the spread of this disease even in Europe.
This study aims to perform an epidemiological survey of PPR in the Saharawi
Arab Democratic Republic (SARD), assessing the potential presence of the virus,
since no reference data had been reported, but also its prevalence and the
distribution in these territories.
This work, conducted with the collaboration of the Veterinary Direction of the
Pubblic Helth Ministery, is based on census and collection of sheep and goat
population and camels living on this territories, from which blood samples were
collected to undergo serological tests.
Blood samples were collected in March and April 2008 and the sampling
methodologyperformed has been allowed to identify 11 sites of sampling for a
total of 976 tested animals.
The results of these tests have evidenced a serological positive prevalence in
about 30% of the tested animals, even though during the collection no animal
presented clinical signs related to the subjected disease. A major number of
4

positive animals was revealed in goats with higher prevalence in subjects over 36
moths of age. Some positive cases were reported also in camels, which normally
aren’t considered a susceptible species to this disease.
5

RIASSUNTO
La Peste dei Piccoli Ruminanti (PPR) è una malattia virale acuta che colpisce gli
ovi-caprini, caratterizzata da elevata morbilità e mortalità. A tutt’oggi viene
considerata una malattia tropicale in quanto presenta la sua maggiore diffusione in
Africa sub-sahariana, Medio Oriente e Asia meridionale, ma i cambiamenti di
ordine ecologico-ambientale a cui si assiste negli ultimi anni e l’intensificarsi del
commercio animale potrebbero favorire la diffusione di questa malattia anche in
Europa.
Lo scopo di questo studio è stato quello di eseguire un’indagine epidemiologica
sulla PPR nei Campi Profughi Saharawi e nei Territori Liberati della Repubblica
Democratica Araba Saharawi (RASD), per valutarne sia l’eventuale presenza,
dato che non esistonosono dati bibliografici in merito, sia la prevalenza e
distribuzione.
Lo studio, svolto in collaborazione con la Direzione di Veterinaria del Ministero
della Salute Pubblica della RASD, si è basato sul censimento e campionamento
della popolazione ovi-caprina e dei dromedari presenti in queste regioni dai quali
sono stati eseguiti prelievi di sangue da sottoporre a test sierologici.
La raccolta dei campioni di ematici è stata effettuata nei mesi di Marzo e Aprile
2008 utilizzando una specifica metodica di campionamento che ha permesso di
individuare 11 aree di campionamento per un totale di 976 animali testati.
I risultati degli esami eseguiti hanno messo in evidenza una prevalenza sierologica
in circa il 30% dei campioni testati, anche se al momento del campionamento
6

nessun soggetto presentava segni clinici riconducibili alla malattia. Una maggiore
sieropositività è stata riscontrata nei caprini con maggior prevalenza negli animali
di età superiore a 36 mesi. Alcune positività sono state evidenziate anche nei
dromedari, i quali normalmente non rappresentano una specie sensibile a questa
malattia .
7

1. INTRODUZIONE
La Peste dei Piccoli Ruminati (PPR) è una malattia infettiva degli ovini e dei
caprini caratterizzata da alta morbilità e mortalità. E’ una malattia soggetta ad
obbligo di notifica, tenuta sotto stretta sorveglianza dall’Organizzazione Mondiale
per la Salute Animale (OIE) e un tempo inserita nella lista “A”, ossia tra le
malattie ad elevato impatto economico. La PPR risulta capillarmente distribuita in
tutta l’Africa Sub-Sahariana, nel Medio Oriente e nell’Asia Meridionale (fig.1),
dove causa gravi perdite economiche (Taylor, 1984a; Shaila et al., 1989; Diallo, 2003) e si
presenta come un forte ostacolo allo lo sviluppo dell’allevamento dei piccoli
ruminanti in questi Paesi (Couacy-Hymann et al., 2007).
Figura 1: Distribuzione mondiale della PPR (dati OIE, 2004).
Malattia riportata presente
Malattia riportata assente Dati non disponibili o assenti
8

Questa malattia fu descritta per la prima volta in Costa d’Avorio nel 1942
(Gargadennec & Lalanne, 1942) e fu denominata “Peste dei Piccoli Ruminanti” per
favorire la sua differenziazione dalla Peste Bovina con cui presenta numerose
correlazioni dal punto di vista eziologico, clinico ed anatomo-patologico. E’ anche
nota come complesso stomatite-pneumoenterite che meglio descrive i segni clinici
associati alla malattia (Rowland et al., 1969, 1970). In seguito la PPR fu segnalata anche
in alcune regioni francesi dell’Africa Occidentale ed in Senegal (Gilbert e Monnier,
1962) con conseguente diffusione in gran parte del continente africano, in particolar
modo nelle regioni al nord della zona sub-sahariana, in Sudan (El Hag Ali e Taylor,
1984; Taylor, 1984), in Africa Orientale, Kenya e Uganda (Wamwayi et al, 1995) ed
Etiopia (Roeder et al., 1994). In Tanzania non sono stati mai segnalati focolai attivi
nonostante la presenza della malattia nei paesi confinanti quali Kenya ed Uganda
(P.N. Wambura, 2000).
A tutt’oggi è quindi possibile affermare che la PPR è presente in molti paesi
africani che si trovano tra l’Oceano Atlantico e il Mar Rosso. L’area principalmente
interessata è quella che si estende dal nord dell’Egitto al sud del Kenia e da est a
ovest del Gabon; mancano segnalazioni nel resto del continente africano (fig.2).
Il 23 luglio 2008 le autorità sanitarie del Marocco hanno segnalato la presenza di
due focolai nella parte centrale del Paese dove la malattia si è rapidamente diffusa a
sud e a nord-est ed il 14 agosto 2008 sette focolai sono stati riportati al confine con
l’Algeria. Attualmente più di un centinaio di focolai sono stati denunciati su tutto il
territorio marocchino (CESME).
9

Al momento la situazione in Africa sta diventando sempre più preoccupante in
quanto si stanno evidenziando nuovi focolai d’infezione.
Figura 2: Distribuzione della PPR nel continente africano.
Per quanto riguarda la Penisola Arabica la PPR è endemica in Oman (Taylor et al.,
1990) ed è stata anche riscontrata in Arabia Saudita (Abu Elzein et al., 1990) mentre in
Medio Oriente è presente in Libano, Giordania (Lefefre et al., 1991) ed Israele (OIE,
1993).
In Asia fu segnalata per la prima volta in India nel 1987 (Shaila et al., 1989) dove da
allora si presenta a carattere endemico (Nanda et al., 1996) e da cui si è estesa sino al
Nepal, Bangladesh, Tajikistan, Pakistan ed Afganistan (AVIS).
Nel 2007 nuovi focolai sono stati notificati in Cina (Tibet e Nepal), dove la malattia
è tutt’ora presente.
In Europa, dove la PPR non viene segnalata, il rischio maggiore è rappresentato
dall’introduzione di animali recettivi e infetti (piccoli ruminanti) provenienti da
Paesi endemici ed è per questo motivo che la legislazione veterinaria europea vieta
il commercio di animali sensibili alla PPR con paesi non indenni (CESME).
10

La distribuzione geografica degli ultimi 50 anni probabilmente è riconducibile ad
una maggiore consapevolezza della malattia dovuta ad una più alta disponibilità di
mezzi diagnostici e ad una maggiore conoscenza del virus che in natura potrebbe
avere subito mutazioni (AVIS).
La PPR è una malattia contagiosa a rapida diffusione che colpisce gli ovi-caprini,
animali che per le popolazioni africane rappresentano la maggiore fonte di
sostentamento. Dal commercio del bestiame, infatti, deriva buona parte del reddito
di queste popolazioni e quindi la presenza della PPR implica un grande impatto
socio-economico.
Studi epidemiologici risultano essere, di conseguenza, particolarmente significativi
non solo nelle aree dove la malattia è endemica ma anche in quelle a rischio di
introduzione come ad esempio l’Europa dove in questo momento la PPR è assente,
sebbene alcuni paesi europei, in particolare quelli più a sud come la Spagna ed a est
come la Grecia, a causa della loro posizione geografica, risultano essere
particolarmente esposti all’introduzione della malattia. Occorre inoltre ricordare che
un ulteriore rischio per il continente europeo è rappresentato dalla presenza della
malattia in Turchia, dove l’infezione è presente fin dal 1999 (CESME).
2. CARATTERISTICHE GENERALI DELLA PPR
2.1 EZIOLOGIA
Il virus della Peste dei Piccoli Ruminanti (PPRV) appartiene alla famiglia
Paramyxoviridae, genere Morbillivirus, antigenicamente correlato al virus della
Peste Bovina, del Cimurro del cane, del Morbillo umano, e ai virus che colpiscono i
11

mammiferi marini (Phocine distemper delle foche, Morbillivirus dei Cetacei isolati
da delfini e focene), anch’essi inclusi nello stesso genere (Gibbs et al., 1979; Barrett et
al., 1993).
Gli appartenenti alla famiglia dei Paramyxoviridae sono
virus a RNA molto grandi (150-300 nm di diametro),
particolarmente pleomorfi. Il nucleocapside, a simmetria
elicoidale, contiene un filamento di ssRNA (–) di 15-16
kb, non segmentato, ed avvolto dall’envelope (fig.3). Figura 3: struttura del PPRV (AVIS).
La resistenza delle particelle virali al calore, all’acidità e ai comuni disinfettanti è
molto scarsa in tutti i membri della famiglia, con l’eccezione del virus della malattia
di Newcastle o pseudo-peste aviare, che oltre ad essere termoresistente, sopravvive
per mesi a temperatura ambiente e a 4°C, nei tessuti degli animali infetti e nelle
uova.
Il virus della PPR può resistere a temperatura di 60°C per 60 minuti, è stabile a pH
compreso tra 4 e 10, è sensibile ad alcool, etere ed ai comuni disinfettanti e può
sopravvivere al congelamento e alla refrigerazione per lunghi periodi.
I membri della famiglia Paramyxoviridae possiedono due tipi di proteine del
nucleocapside, strutturali ed ausiliarie: le strutturali sono proteine che vengono
denominate in modo diverso a seconda del genere (NP, NC o N). Sono proteine
costituite da due domini: amino-terminale, che comprende i due terzi della molecola
e che interagisce direttamente con l’RNA, e carbossi-terminale, che si pone sulla
superficie esterna del nucleocapside, rendendolo particolarmente flessibile.
12

Le proteine ausiliari sono invece implicate nel processo di trascrizione e
replicazione del genoma. Si differenziano in proteina L (large), molto grande (che
verosimilmente rappresenta la RNA polimerasi) e proteina P, fosforilata
(polymerase associated), più piccola. Insieme possiedono l’attività enzimatica
richiesta per la trascrizione (RNA polimerasi) e per le modificazioni post-
trascrizionali dell’RNAm: poliadenilazione all’estremità 3’ e metilazione al 5’.
Le glicoproteine dell’envelope hanno funzioni sia di attacco che di fusione. Le
proteine di attacco sono diverse nei diversi generi: H (emoagglutinina) ed N
(neuroaminidasi) nel genere Paramyxovirus sono in grado di riconoscere i recettori
cellulari specifici che presentano acido sialico, per i quali hanno un’affinità molto
elevata; sono in grado di agglutinare gli eritrociti dei mammiferi e dei uccelli e,
contemporaneamente, presentano attività neuraminidasica che si esplica al
momento del rilascio delle particelle. La proteina H nel genere Morbillivirus è
deputata all’ adsorbimento ed è emoagglutinante, ma non ha attività
neuraminidasica. La proteina G nel genere Pneumovirus possiede la sola funzione
di adsorbimento e manca di attività emoagglutinante e neuraminidasica. La proteina
5F, di fusione, è presente in tutti i generi e garantisce la fusione dell’envelope con la
membrana cellulare, permettendo il rilascio del nucleocapside nel citoplasma. Le
proteine F vengono sintetizzate come precursori inattivi (F0), e sono attivate
dall’azione di un enzima cellulare tripsino-simile. Questo scinde la F0 in F1, la cui
parte idrofobica si inserisce nella membrana cellulare, e in F2, sub-unità più piccola
che rimane ancorata alla F1 mediante ponti disolfuro. Grazie a questo meccanismo,
l’entrata del nucleocapside avviene a pH neutro. Il fatto che l’ingresso del virus
13

nelle cellule sia legato alla modificazione enzimatica della F0 ha due importanti
conseguenze: 1) il tropismo del virus è ristretto ai tipi cellulari che possiedono le
adatte proteasi; 2) la maggiore o minore sensibilità delle diverse F0 all’azione
enzimatica è direttamente collegata a diversi gradi di patogenicità.
La proteina F agisce anche come fattore di fusione tra cellule contigue, permettendo
la diffusione del virus persino in presenza di anticorpi. Dal momento che la proteina
F è fondamentale per la penetrazione, come lo sono le proteine di adesione, gli
anticorpi anti-F contribuiscono, insieme a quelli diretti contro le proteine di
adesione, a conferire immunità nei confronti del virus.
Dal punto di vista antigenico, le proteine superficiali dei Paramyxovirus sono molto
più stabili rispetto a quelle degli Orthomyxovirus. Sono stati infatti definiti alcuni
“sierotipi” nel genere Paramyxovirus (che forse sono da intendere come specie
virali diverse), ma nell’ambito dello stesso sierotipo le variazioni tra isolati diversi
sono molto scarse ed evidenziabili solo mediante anticorpi monoclonali. Nel genere
Morbillivirus sono state messe in evidenza estese cross-reazioni tra gli antigeni
superficiali F, ma non con H. All’interno di una singola specie virale la
configurazione antigenica è molto uniforme.
Sempre nell’ambito delle proteine di attacco dell’envelope ritroviamo anche le
proteine M, di matrice. Queste si allineano all’interno dell’envelope e sono deputate
al riconoscimento del nucleocapside durante l’assemblaggio e verosimilmente
partecipano anche alla formazione dell’envelope stesso.
14

Il processo di replicazione si esplica con lisi cellulare con formazione di sincizi e di
corpi inclusi acidofili. Pur essendo la replicazione completamente a carico del
citoplasma, corpi inclusi acidofili si formano anche nel nucleo.
La replicazione inizia con la trascrizione primaria dell’RNA gonomico, ad opera
della RNA polimerasi-RNA dipendente, in 6-10 (secondo i generi) filamenti di
RNAm monocistronici, che utilizzano la stessa sequenza leader e sono sintetizzati
nello stesso ordine in cui i rispettivi geni sono situati nel genoma. Deve quindi
esistere, all’inizio e alla fine di ogni gene, un meccanismo che controlla l’inizio e la
fine della trascrizione: in effetti, alle estremità dei geni, esistono delle sequenze
nucleotidiche omologhe alle sequenze coinvolte nei segnali di inizio/fine
trascrizione del DNA cellulare. Inoltre, esiste una fine regolazione nella “quantità”
di RNAm trascritto per ciascun gene, che dipende dalla posizione del gene stesso
nel genoma. L’efficienza della trascrizione, infatti, decresce con l’aumentare della
distanza del gene stesso dall’estremità 3’ del genoma. Il motivo di tale regolazione
è che il virus necessita di molte copie delle proteine strutturali (come ad esempio la
NP) e solo di alcune copie delle proteine di attività enzimatica (come ad esempio la
L).
Contemporaneamente alla trascrizione degli RNAm si producono alcuni filamenti
(+) a lunghezza genomica che fungono da intermedi replicativi per la sintesi
dell’RNA progenie. Ovviamente, la trascrizione dell’intero RNA (–) genomico in
un singolo filamento (+) comporta che, in questo caso, la trascrizione stessa
avvenga ignorando tutti i segnali di inizio/fine. Non è ancora del tutto chiaro come
il medesimo enzima (RNA polimerasi) possa comportarsi in due modi così diversi.
15

Potrebbe essere dovuto al fatto che l’interazione dell’enzima con le proteine del
nucleocapside in via di formazione sia in grado di influenzare l’attività in questo
senso.
La maturazione e il rilascio prevedono l’incorporazione delle proteine strutturali
glicosilate [H e N (N o G) e F] nella membrana plasmatica della cellula ospite,
l’associazione della proteina M alla superficie interna della stessa membrana,
l’allineamento del nucleocapside (RNA+NP+L+P) al di sotto della proteina M e,
infine, il rilascio, mediante budding, dei virioni maturi (Poli).
In natura è recettiva al virus della PPR la capra, in minor misura la pecora ed il
bovino che, se sottoposto ad infezione sperimentale, presenta soltanto febbre di
modesta entità. Nel maiale è riproducibile soltanto la forma subclinica, seguita da
risposta anticorpale, non associata ad escrezione del virus mentre nel cervo,
l’infezione sperimentale, può provocare sintomi di entità molto variabile con
possibile acquisizione di infezione subclinica o di infezione acuta o letale.
Su base sierologica, con l’impiego di anticorpi policlonali, è possibile dimostrare
strette correlazioni antigeniche nei confronti degli altri membri del genere
Morbillivirus, che risultano particolarmente marcate per il virus della Peste Bovina
andando a giustificare la presenza di reazioni sierologiche crociate in pecore e
bovini inoculati con i rispettivi virus. Comunque l’analisi del genoma con sonde
molecolari derivate dal gene N (nucleoproteina), oppure l’impiego di antisieri
monoclonali attivi sulle proteine di superficie consentono di riconoscere
l’appartenenza dei virus a specie distinte (Farina & Scatozza 1998).
16

2.2 EPIDEMIOLOGIA E TRASMISSIONE
La Peste dei Piccoli Ruminanti (PPR) è una infezione virale acuta, altamente
contagiosa che colpisce i piccoli ruminanti e occasionalmente anche alcuni
ruminanti selvatici (Gazella dorcas, Capra ibex nubiana, Oryx gazella, Ovis
orientalis laristanica), caratterizzata da febbre alta, scolo nasale e oculare,
polmonite, necrosi, infiammazione ed ulcerazione delle mucose e delle membrane
del tratto gastro-intestinale che portano a grave diarrea (Radostits et al., 2000).
A causa dell’elevato tasso di mortalità, soprattutto tra i giovani animali, la PPR
riveste una grande importanza economica anche per le misure restrittive che porta
al commercio del bestiame (Al-Majali, A.M., et al., 2008).
Quando la malattia si presenta per la prima volta in una zona sono possibili
manifestazioni caratterizzate da febbre molto alta e forte depressione che spesso
esitano nella morte dei soggetti colpiti ancor prima dell’evidenziazione di ogni
altro tipico segno clinico. Un quadro più comune tuttavia, si appalesa con una
rapida diffusione della malattia con improvvisa comparsa di depressione,
secrezioni nasali e oculari, respirazione anomala, tosse, diarrea e morte (fig.4).
Figura 4: gregge affetto da PPR (P.L. Roeder).
17

Sebbene sia le capre che le pecore siano suscettibili all’infezione e possano
manifestare segni clinici di malattia, a volte tale specie non vengono colpite
simultaneamente. Nel continente africano ad esempio sono più comunemente
colpiti i caprini, mentre in Asia meridionale ed occidentale principalmente gli
ovini (AVIS).
Le secrezioni oculari e nasali tipiche della PPR, come pure le feci, contengono
grandi quantità di virus. Piccolissime particelle aerosoliche, che si liberano in
seguito a tosse e starnuti, favoriscono la diffusione del virus tra gli animali e lo
stretto contatto quindi rappresenta il più importante fattore di rischio ai fini della
trasmissione. Secrezioni ed escrezioni infette possono anche contaminare acque di
bevanda, alimenti o le lettiere, trasformandoli in ulteriori fonti di infezione anche
se quest’ultimi sono relativamente importanti in quanto il virus della PPR non è in
grado di sopravvivere a lungo al di fuori dell’ospite (Recognizing Peste des Petits
Ruminants, a field manual FAO).
La localizzazione primaria del virus è rappresentata dalla mucosa delle prime vie
respiratorie a cui segue replicazione nelle tonsille palatine e nei linfonodi faringei
e mandibolari. Il successivo adsorbimento alla superficie dei mononucleati è
responsabile della viremia che precede di 1-2 giorni la comparsa dei sintomi. Le
localizzazioni secondarie sono rappresentate dagli organi linfoidi, nonchè dalle
mucose degli apparati digerente e respiratorio. Nella fase prodromica, la
replicazione virale è molto efficiente ma si arresta successivamente alla comparsa
di febbre e di anticorpi specifici nel torrente circolatorio. La viremia persiste a
18

lungo in presenza di stipiti virali ad elevata virulenza e/o di soggetti ad elevata
recettività, ma di norma alla guarigione non segue lo stato di portatore anche se,
sia pure occasionalmente, il virus è stato isolato a distanza di settimane o mesi
dalla regressione dei sintomi.
Casi clinici di PPR possono essere associati a movimentazione degli animali,
all’abitudine di costituire gruppi di animali di specie ed età diverse,
all’introduzione di nuovi soggetti, al commercio animale ed, in particolare, nei
mercati dove greggi provenienti da differenti regioni vengono poste a stretto
contatto o, infine, con greggi nomadi attraverso la condivisione di pascoli e punti
di abbeverata. Anche i cambiamenti climatici possono svolgere un ruolo
importante, episodi di malattia infatti si registrano più frequentemente all’inizio
della stagione delle piogge (caldo e umido), o l’inizio della stagione secca. Nelle
aree endemiche la maggior parte degli animali malati o morti risultano quelli di
età compresa tra i quattro mesi e i due anni (AVIS).
La trasmissione avviene quindi per contatto diretto per via inalatoria, mentre il
contatto indiretto, pur non potendo essere escluso, assume rilevanza modesta. La
comparsa di PPR in territori riconosciuti indenni è costantemente riferibile
all’importazione di animali infetti che presentano una forma subclinica o
asintomatica.
Nelle aree indenni, quindi, in presenza di animali ad alta recettività, la PPR
evolve in forma grave e letale anche dopo contatto con stipiti a bassa virulenza al
contrario di quanto si verifica invece nelle aree endemiche. La morbilità può
raggiungere anche il 100% in una popolazione recettiva, mentre la mortalità varia
19

dal 50 all’80%. Queste percentuali sono in genere più basse nelle zone endemiche
dove gli animali entrano frequentemente in contatto con il virus.
2.3 SEGNI CLINICI E LESIONI ANATOMOPATOLOGICHE
I segni clinici, in seguito ad infezione naturale, compaiono in media dopo due/sei
giorni dall’avvenuto contatto, con l’insorgenza improvvisa di febbre alta, 40-
41°C, anoressia e ottundimento del sensorio. Inizialmente si può osservare
un’essudazione oculo-nasale sierosa che si trasforma poi in muco-purulenta in
seguito ad infezioni batteriche secondarie che si sviluppano soprattutto a carico
dell’apparato respiratorio (Pasteurella sp.) (fig. 5 e 6). A breve distanza di tempo
compaiono erosioni ed aree necrotiche sulle mucose del cavo orale associate a
diarrea persistente.
Figura 5: secrezione oculare sierosa (FAO). Figura 6: secrezione oculo-nasale muco- Purulenta (FAO).
Uno o due giorni dopo l’insorgenza della febbre le membrane delle mucose
buccali e oculari diventano molto congeste. La necrosi epiteliale porta a delle
piccole aree tondeggianti di colore grigio sulle gengive, sul palato, sulle labbra,
all’interno delle guance e sulla superficie superiore delle lingua che tendono a
crescere di numero e di dimensioni fino a confluire (fig.7). La cavità buccale
20

assume un aspetto drasticamente modificato, assume un colorito pallido ed è
ricoperta di cellule morte che in alcuni casi possono formare una vera e propria
pellicola di materiale spesso ed appiccicoso al di sotto del quale si possono
trovare ulteriori erosioni superficiali (fig.8). In alcuni casi le lesioni possono non
essere facilmente notate e richiedono un attendo esame visivo. Con il semplice
sfregamento della gengiva e del palato è possibile asportare materiale
maleodorante contenente brandelli di tessuto epiteliale. Stesse modificazioni si
posso riscontrare anche nelle mucose del naso, della vulva e della vagina dove le
labbra tendono a gonfiarsi e a ricoprirsi di croste.
Figura 7: tipica lesione buccale in corso Figura 8: tipico aspetto della mucosa di PPR (CESME). Buccale in corso di PPR (FAO).
Con il progredire della malattia viene prodotto un caratteristico essudato
maleodorante dalla bocca e, i soggetti colpiti, a causa del dolore non riescono ad
aprire la bocca e quindi a nutrirsi.
La diarrea compare generalmente dopo due o tre giorni dall’insorgenza della
febbre, anche se, in alcuni casi può non essere evidente. Le feci sono inizialmente
morbide e poi acquose, maleodoranti e possono contenere tracce di sangue e pezzi
di tessuto necrotico. Quando la diarrea non è evidente, l’inserimento di un
21

tampone di cotone idrofilo nel retto può rilevare la presenza di feci molli spesso
macchiate di sangue.
Gli animali colpiti presentano una respirazione anomala, a volte talmente veloce
da presentare discordanza toraco-addominale ad ogni atto respiratorio. Nei casi
più gravi si può avere difficoltà nella respirazione con intensi rumori respiratori,
che l’animale esprime estendendo testa e collo, dilatando le narici, protendendo la
lingua e con una tosse non secca ma dolorosa, tipici segni di una polmonite.
Questi soggetti possono andare incontro a forte disidratazione visibile dagli occhi
notevolmente incavati. La morte può subentrare dai sette ai dieci giorni dalla
comparsa dei primi segni clinici di malattia. Alcuni animali, a seguito di una
lunga convalescenza, possono invece recuperare un buono stato di salute.
Superata la fase acuta della malattia frequentemente si possono riscontrare piccole
lesioni nodulari a livello cutaneo, sulla parte esterna delle labbra ed intorno al
muso. L’eziologia di queste lesioni non è ben nota ma viene ipotizzata
un’infezione da Dermatophilus sp. o la riattivazione del virus dell’ectima
contagioso (AVIS).
La carcassa di un animale colpito generalmente risulta essere emaciata, con i
quarti posteriori sporchi e ricoperti di feci morbide o acquose e con i bulbi oculari
estremamente incavati in seguito alla imponente disidratazione. Gli occhi e il naso
risultano secchi o coperti da essudazioni.
A livello della bocca si riscontrano erosioni biancastre sulle gengive, sul palato
molle e su quello duro, sulla lingua, nelle guance fino a tutto l’esofago. Le labbra
22

sono gonfie e possono presentare anch’esse erosioni ed eventualmente croste e
noduli.
Il rivestimento della cavità nasale è congesto con essudazione di colore chiara o
gialla cremosa e presenza di erosioni.
I polmoni presentano aree di colore rosso scuro o porpora, duri al tatto,
specialmente nei lobi anteriore e cardiaco, come riscontro di polmonite lobare
acuta è indicativa di complicanze secondarie (fig. 9 e 10).
Figura 9: riscontro di polmonite lobare acuta Figura 10: tipiche lesioni polmonari in corso in corso di PPR (FAO). di PPR (FAO).
I linfonodi, a livello polmonare ed intestinale, sono morbidi ed aumentati di
volume. L’abomaso è congesto e talvolta emorragico. Anche il piccolo intestino
appare congesto, emorragico con presenza di erosioni e le placche del Peyer
presentano deplezione linfocitaria indicativa del coinvolgimento delle strutture
linfoidi. Il grosso intestino (cieco, colon e retto) presenta piccole emorragie
rossastre lungo le pieghe della mucosa che tendono a confluire con il tempo
diventando più scure fino a risultare verdi-nerastre nelle carcasse più vecchie (fig
11).
23

Figura 11: tipiche lesioni “a zebra” del colon (FAO).
2.4 DIAGNOSI
L’elevata diffusibilità della PPR ed il relativo comportamento su base anatomo-
clinico, che si possono riscontrare nelle aree endemiche, danno fondamento al
sospetto diagnostico anche se i caratteri della malattia possono variare ampiamente
in relazione alla recettività degli animali e alla virulenza degli stipiti virali presenti
nei singoli focolai. Supporto significativo nell’indagine anamnestica è sicuramente
quello riguardante la movimentazione degli animali con attenzione
all’introduzione di nuovi soggetti provenienti dalle zone infette e/o aree
endemiche.
La comparsa improvvisa di secrezioni oculo-nasali, diarrea e morte associata a
problemi respiratori, soprattutto in soggetti giovani, deve suscitare il sospetto di
malattia così come i rilievi dell’indagine post-mortem nelle carcasse colpite.
La sola diagnosi “provvisoria” non è però sufficiente perché molte altre gravi
malattie che colpiscono gli ovi-caprini possono presentare segni clinici
riconducibili alla PPR; per questo è sempre necessaria anche una diagnosi di
laboratorio per individuare la presenza del virus o di anticorpi specifici.
24

2.4.1 DIAGNOSI DIFFERENZIALE
La peste dei Piccoli Ruminati può facilmente essere confusa con altri quadri
morbosi, quindi nell’emissione di un sospetto clinico di PPR occorre tenere
presente diverse patologie caratterizzate da una sintomatologia e da lesioni
sovrapponibili. Tra queste possono essere citate:
- la Peste Bovina che può colpire anche gli ovicaprini determinando quadri
clinici molto simili.
- la Febbre Catarrale degli ovini (Bluetongue) che può essere confusa con la
PPR per le lesioni orali e i sintomi respiratori, anche se tuttavia la
bluetongue ha un andamento per lo più stagionale e si differenzia per la
presenza di zoppia legata alle lesioni erosive a livello del cercine
coronario, l’edema della testa, della lingua e del musello. Inoltre, nel caso
della bluetongue, l’incidenza attesa di animali infetti nel gregge è
inferiore.
- l’Ectima contagioso: le lesioni orali sono pustolose e crostose, mancano la
diarrea e la polmonite a differenza della PPR.
- il Vaiolo ovicaprimo: mancano i sintomi gastroenterici tipici della PPR.
- l’Afta epizootica: nella PPR non sono presenti zoppia e le tipiche
vescicole dell’afta.
- la Pasteurellosi: mancano le lesioni necrotiche e la diarrea tipiche della
PPR.
25

- la Pleuropolmonite contagiosa caprina (CCPP): può essere confusa per i
sintomi respiratori, tuttavia mancano la diarrea e le lesioni delle mucose
tipiche della PPR.
- l’Idropericardite dei ruminanti: si differenzia per la presenza di sintomi
neurologici.
- la Coccidiosi, Salmonellosi ed Elmintiasi: possono essere confuse per i
sintomi gastroenterici, tuttavia mancano le lesioni orali e i sintomi
respiratori.
- gli avvelenamenti da sostanze di origine minerale e vegetale: i sintomi
variano in relazione alla sostanza responsabile, pertanto l’anamnesi ed un
attento esame clinico, in aggiunta ad esami tossicologici, possono
permettere la differenziazione dalla PPR.
Per l’isolamento virale se eseguito intra vitam è consigliato l’utilizzo di sangue
defribinato, secreto oculare e raschiato delle lesioni del cavo orale; nell’esame
post mortem, possibilmente su eseguito su carcasse di soggetti sacrificati non oltre
4-5 giorni dall’esordio della sintomatologia, sono da preferire linfonodi, milza e
tonsille. Il virus può essere isolato da colture primarie di rene di diverse specie
animali (pecora, capra, bovino, scimmia), da testicolo di capra ed amnios umano,
fra le linee cellulari continue sono utilizzabili le BHK 21, Hep-2, MDBK e Vero.
L’identificazione virale può essere eseguita mediante sieroneutralizzazione,
immunofluorescenza diretta, precipitazione in gel di agar (AGID) e fissazione del
complemento. L’isolamento e la successiva identificazione però, pur auspicabili
26

per sensibilità ed affidabilità, richiedono tempi di esecuzione relativamente
lunghi. Inoltre l’individuazione degli antigeni virali tramite AGID, seppur utile
come prova iniziale, non permette la differenziazione tra la PPR e la Peste Bovina
(PB).
La diagnosi sierologica su campionamento seriale è ugualmente affidabile ma non
trova applicazione soprattutto nelle aree indenni in quanto richiede 2-3 settimane
di attesa fra il prelievo di fase acuta e quello effettuato in fase di remissione dei
sintomi. Esistono, per tale motivo, anche test immunoenzimatici (ELISA) che
permettono di svelare la presenza di IgM specifiche che compaiono precocemente.
Anche il ricorso all’istologia potrebbe fornire un valido supporto diagnostico
attraverso l’osservazione di necrosi focale delle mucose dell’apparato digerente e
dei linfociti, mentre nel polmone si evidenziano cellule multinucleate contenenti
inclusi eosinofili a sede nucleare e citoplasmatica. Questa tecnica associata con
l’utilizzo di anticorpi monoclonali specifici (immunoistochimica) permette anche
la differenziazione del virus della PPR dalla PB.
Nell’ultimo decennio, grazie all’utilizzo di tecniche di biologia molecolare molti
dei problemi sopracitati sono stati superati attraverso la validazione di RT-PCR
(reverse transcriptase polymerase chain reaction) (Jingyue Bao. Et al., 2008; E. Couacy-
Hymann et al., 2005), attualmente caratterizzati da una notevole rapidità di
esecuzione, sensibilità e precisione e che permettono l’individuazione anche di
minime tracce dell’acido nucleico virale. Questi, coadiuvati da test ELISA,
rappresentano le prove di laboratorio a cui, ad oggi, più frequentemente si fa
ricorso.
27

2.5 PREVENZIONE E CONTROLLO
Un’ importante caratteristica dell’infezione causata dal PPRV è rappresentata,
come per altri Morbillivirus, da un’intensa ma transitoria immunosoppressione
che il virus provoca nell’ospite, con il conseguente aumento di suscettibilità nei
confronti di infezioni da germi di irruzione secondaria e quindi aumento della
mortalità. Questo effetto immunosoppressivo non è solamente la conseguenza
diretta della moltiplicazione del virus nelle cellule linfoidi, ma anche delle diverse
strategie che i Morbillivirus hanno elaborato per eludere il sistema immunitario
degli ospiti (Rajak et al., 2005; Schneider-Schaulies et al., 2001). Tuttavia, l’effetto è di
tipo transitorio ed il recupero dalla malattia è di solito seguito da una risposta
immunitaria, da parte dell’ospite, specifica e a lungo termine (Servet-Delprat et el.,
2003; Cosby et al., 2005).
Subito dopo il primo isolamento del virus in colture tissutali, negli anni ’60,
furono eseguiti diversi tentativi per sviluppare dei vaccini attenuati, ma senza
successo (Gilbert Y. e Monnier J., 1962; Benazet B., 1973). A quei tempi era disponibile
un vaccino vivo attenuato, molto valido contro la Peste Bovina (Plowright W., Ferris
RD., 1962) e vista la stretta correlazione antigenica tra i due virus fu utilizzato
questo vaccino per immunizzare le capre nei confronti della PPR . Le valutazioni
della risposta immunitaria in questi animali dimostrarono la presenza di anticorpi
neutralizzanti contro il PBV, ma non contro il PPRV, se non in alcuni casi (Taylor
WP., 1979). Tuttavia, in tutti gli animali, in seguito a challenge, fu evidenziato un
certo grado di resistenza all’azione patogena del virus, accompagnata da un
innalzamento nell’attività degli anticorpi neutralizzanti contro il PPRV.
28

Nonostante non proteggesse nei confronti dell’infezione questo vaccino è stato per
anni utilizzato con successo, in quanto conferiva comunque un’immunità di lunga
durata (Rossiter PB., Taylor WP., 1994; Bourdin P., Rioche M., Laurent A., 1970; Taylor WP. Et
al., 1990).
Un altro vaccino molto utilizzato in passato è stato il TCRV (tissue-culture
rinderpest), estratto da cellule tissutali di animali infetti da Peste Bovina. Secondo
Gibbs et al. (1977) il TCRV conferiva un’immunità completa per le capre senza
trasferire la malattia ad altri animali e secondo Adu e Nawathe (1981) anche gli
animali gravidi erano ben tolleranti a questo vaccino. Al contrario, Abegunde
(1983) specificò che se gli animali risultavano già infetti prima della vaccinazione,
magari in forma subclinica, a seguito di questa potevano mostrare segni di
malattia ed inoltre in quelli gravidi si poteva registrare aborto.
Nel 1989 si riuscì con successo ad attenuare il PPRV attraverso dei passaggi
seriali su cellule Vero. I soggetti vaccinati con tale vaccino attenuato non
trasmettevano il virus agli altri animali con cui erano a stretto contatto e gli
anticorpi prodotti persistevano per almeno tre anni, ossia l’effettiva vita
economica degli animali. Il vaccino omologo attenuato è attualmente l’unico
permesso nelle pecore e nelle capre per garantire un’efficace immunizzazione nei
confronti dell’infezione da PPRV (Diallo A. et al., 2007) anche se un inconveniente
nell’uso è rappresentato dal fatto che, essendo allestito con un virus appartenente
alla famiglia delle Paramyxoviridae, è molto sensibile al calore e quindi non
adatto all’utilizzo nelle aree endemiche caratterizzate da un clima estremamente
29

caldo dove talvolta mancano le strutture per garantire la conservazione del
vaccino stesso.
Questo problema è stato risolto da Worwall et al. attraverso l’allestimento di un
vaccino liofilizzato termoresistente congelato con un crioprotettore contenente
trealosio (Worwall et al., 2001).
L’utilizzo di questi vaccini però non permette la differenziazione tra la risposta
immunitaria vaccinale e l’infezione naturale, rendendo difficile il controllo della
malattia nelle aree endemiche. Per questo motivo sono allo studio dei vaccini
ricombinanti che prevedono l’utilizzo delle due glicoproteine dell’envelope virale
capaci di sviluppare una risposta anticorpale, rappresentate dalla proteina di
fusione (F) e l’emoagglutinina (H). Le proteine F ed H del PPRV sono state
quindi inserite separatamente nel genoma di un Capripox virus (Berhe G. et al., 2003)
ed il conseguente virus ricombinato è stato testato nelle capre dimostrandosi
efficiente nel proteggere gli animali nei confronti della PPR. Entrambi i vaccini
hanno dimostrato la loro utilità anche perché possiedono caratteristiche DIVA,
ossia danno la possibilità di distinguere, in sede di diagnosi laboratoristica, gli
animali vaccinati da quelli che sono stati esposti naturalmente al virus (Diallo A. et
al., 2002).
Per l’Italia e i Paesi Europei la PPR è considerata una malattia esotica, quindi si
applicano i dispositivi sanitari previsti nei piani di emergenza per le malattie della
ex lista “A” dell’ OIE.
E’ importante sapere che per i Paesi indenni è previsto il blocco dell’importazione
di animali vivi e prodotti di origine animale provenienti da zone infette ed, in caso
30

di comparsa di un focolaio, l’obbligo di denuncia alle autorità sanitari competenti
con notifica all’OIE. E’ inoltre obbligatorio lo stamping-out (abbattimento coatto
degli animali infetti e sospetti di infezione o contaminazione) con distruzione
delle carcasse mediante incenerimento o sotterramento, creazione di una zona di
protezione e di sorveglianza, ed infine può essere prevista una vaccinazione di
massa nelle zone adiacenti al focolaio.
Per i Paesi endemici invece il protocollo sanitario in caso di comparsa di nuovi
focolai di malattia prevede l’immediata notifica all’OIE, lo stretto controllo delle
movimentazioni dei capi animali, una rigorosa sorveglianza epidemiologica e la
vaccinazione (CESME).
3. PARTE SPERIMENTALE
3.1 AREA DI STUDIO
Lo studio oggetto di questa tesi ha avuto come scopo l’indagine epidemiologica
della PPR nella Repubblica Democratica Araba Saharawi (RASD).
Tale ricerca è stata parte integrante del progetto “Soutien à l’élevage de bétail
dans les Camps de Réfugiés Sahraoui” (ONG-PVD/2006/131-812), finanziato
dalla Commissione Europea e coordinato da Africa ’70, una Organizzazione non
Governativa Italiana (ONG).
Le caratteristiche, l’estensione e la gravità di una malattia quale la PPR,
dipendono ovviamente dalla sua epidemiologia, ma anche dall’ambiente e dalla
popolazione in cui si sviluppa. Per questo è necessario conoscere l’area
31

considerata dalla studio, la sua storia, le sue caratteristiche biologiche e
climatiche, la popolazione umana residente ed il bestiame allevato.
3.1.1 STORIA E SITUAZIONE POLITICA
Il Sahara Occidentale è una regione del Nord Africa confinante a nord con il
Marocco, a nord-est con l’Algeria, a sud e ad est con la Mauritana e ad ovest con
l’Oceano Atlantico.
Questo territorio è stato senza un’autorità governativa autonoma fin dal 1960
quando fu colonizzato dalla Spagna (prendendo il nome di Rio de Oro o Sahara
Spagnolo). Successivamente al ritiro coloniale spagnolo (1975) il Sahara
Occidentale venne invaso dal Marocco e dalla Maurtania, mentre il Fronte
Polisario (Fronte Popolare per la Liberazione di Saguia-el-Hamra e del Rio de
Oro) rivendicava la sovranità su tutto il territorio.
Il conflitto creatosi ha condotto alla proclamazione, il 27 Febbraio 1976, della
Repubblica Democratica Araba Saharawi (RASD), in seguito al ritiro della
Mauritania pattuito con il Fronte Polisario (Shelley, 2004). Da quell’anno il Marocco
ha costantemente mantenuto l’occupazione del territorio, mentre il Polisario ha
continuato la sua campagna militare contro le armate marocchine con lo scopo di
ottenere la liberazione. Nel 1991 le Nazioni Unite, attraverso la MINURSO
(Missione delle Nazioni Unite per il Referendum del Sahara Occidentale), quale
parte del programma di risoluzione del conflitto iniziato con il cessate il fuoco,
organizzò un referendum di autodeterminazione libero ed equo per mezzo del
32

quale veniva data la possibilità al Popolo Saharawi di scegliere fra l’integrazione
con il Marocco e l’indipendenza.
Attualmente la RASD controlla circa il 20% del territorio del Sahara Occidentale.
Questa regione, definita “Territori Liberati”, comprende la porzione orientale del
Saquia-el-Hamra e del Rio de Oro. Il Marocco invece controlla il resto del
territorio che gestisce come sue “Province Meridionali”, mentre vengono
considerate dalla RASD come “Territori Occupati”. Queste due zone sono tuttora
divise fra loro dal Berm, una struttura difensiva lunga più di 2.700 km che
rappresenta una “zona tampone” fra le due parti. E’ controllata militarmente dalle
forze armate marocchine ed è formata essenzialmente da un muro di sabbia il cui
perimetro è costellato interamente da mine antiuomo (Jesen, 2005). Il muro si
estende dal sud del Marocco fino al nord della Mauritania, attraversando tutto il
Sahara Occidentale, e separa fisicamente la parte orientale del Sahara Occidentale
sotto il controllo del Polisario dalla parte occidentale occupata dal Marocco (San
Martín, 2004; Loewenberg, 2005).
La RASD è attualmente riconosciuta come autorità governativa del Sahara
Occidentale da 43 Stati e anche se non ha alcun rappresentante presso le Nazioni
Unite, la repubblica è un membro a pieno titolo dell’ Unione Africana.
33

3.1.2 IL POPOLO SAHARAWI E LA SUA STORIA
Saharawi, letteralmente “gente del deserto”, è il nome dato a quelle tribù di
pastori nomadi che storicamente abitavano la zona costiera del Nord Africa che
corrisponde all’attuale Sahara Occidentale.
Questo popolo origina dalla fusione tra il gruppo arabo Bani Hasan, proveniente
dallo Yemen e giunto in Africa tra l’ XI e il XIII secolo A.C., e quello berbero
Sahjaha, originario del Sahara Occidentale. Attraverso un lento processo di
integrazione tra i migranti e la popolazione berbera autoctona, avvenuto fino al
XVI secolo, si è costituito il Popolo Saharawi. Questo popolo è costituito
essenzialmente da nomadi, di religione mussulmana sunnita, dediti
all’allevamento di dromedari, capre e pecore, che basano la propria alimentazione
principalmente sulla carne ed il latte di dromedario, datteri, zucchero e qualche
cereale e legume (OXFAM, 1995). La loro struttura sociale è di tipo tribale ed è
organizzata suddividendo gli individui in gruppi gerarchici patrilinei che tuttavia
si sono sempre trovati in competizione, tra violenze e negoziati, per avere accesso
alle limitate risorse locali (Caratini, 2000).
Il processo storico che ha condotto le diverse popolazioni di questa regione a
formare un insieme relativamente unito ed omogeneo, quale il Popolo Saharawi,
comprende tre tappe: la prima è quella che precede la colonizzazione, ovvero il
Sahara non ancora compreso da confini precisi; la seconda è quella della
colonizzazione spagnola, che ha assegnato le frontiere al territorio, ha riunito la
popolazione in un quadro ben definito e ha permesso al popolazione di divenire
34

un popolo unito con una sua propria identità; infine, la terza tappa che corrisponde
al periodo contemporaneo, che è caratterizzata dalla lotta per la liberazione.
Durante il periodo pre-coloniale la regione del Sahara Occidentale era
principalmente popolata da tribù berbere venute dal nord, rappresentate da due
gruppi tradizionalmente rivali: i Sanhaja e gli Zeneti. A partire dal VII secolo e
fino all’inizio del XII, l’infiltrazione di genti arabo-yemenite, i Moquil, favorì il
mescolamento dei gruppi nomadi berberi e anche l’adozione della religione
islamica e della lingua araba. Si formarono così tribù di differente origine e
importanza che vivevano in luoghi distinti, ma che avevano numerosi tratti
comuni dovuti alla loro storia, ai loro contatti, alla natura del territorio di origine e
alle sue condizioni climatiche. Infatti, queste tribù, possedevano uno stile di vita
pressoché identico, gli stessi costumi, le stesse feste, la stessa lingua (l’Hassanya,
un dialetto arabo) e la stessa religione.
Ogni tribù aveva una sua organizzazione e struttura sociale specifica che
comprendeva delle frazioni e delle sub-frazioni ripartite a loro volta in diversi
accampamenti che contavano varie decine di famiglie; l’autorità era esercitata
dagli shekh e dalle assemblee.
Le tribù vivevano relazionandosi con le altre nei periodi di nomadismo ed
esistevano tra loro anche dei rapporti gerarchici, poiché alcune erano sottomesse
ad altre. Carattere comune e predominante, inoltre, era l’attaccamento alla loro
indipendenza e alla loro libertà. A nord mantenevano e difendevano la loro
indipendenza nei confronti del Sultano del Marocco, mentre a sud respingevano
le incursioni degli europei che provenivano dal mare.
35

Il Periodo Coloniale ebbe inizio già verso la fine del ‘400, ma la colonizzazione
spagnola vera e propria iniziò solo nel 1884 con la conferenza di Berlino che
riconobbe la sovranità spagnola sul Rio de Oro. Infatti gli spagnoli cominciarono
ad avere forte interesse nel territorio del Sahara Occidentale dopo la scoperta
dell’America, sollecitati dall’avanzata francese in Algeria, Marocco e Mauritania.
Il dominio spagnolo ha giocato un ruolo fondamentale nella formazione del
Popolo Saharawi apportando delle trasformazioni profonde che hanno lasciato
tracce indelebili fino ai nostri giorni.
La Spagna, innanzitutto, ha delimitato il territorio dandogli, per la prima volta,
delle frontiere ben determinate, fissate da convenzioni internazionali (Parigi nel
1900 e 1904, Madrid nel 1912) e basate sui meridiani e i paralleli.
Questi confini vennero tracciati però senza riflettere sulle realtà sociali e
geografiche e basandosi esclusivamente sui rapporti tra le potenze coloniali
presenti nella regione, cioè la Spagna e la Francia. Il territorio, definito dalla
Spagna all’inizio del secolo, in accordo con la Francia, comprendeva due zone a
statuto differente: da una parte, la regione di Tarfaya, tra il Wadi Draa, il
meridiano 11° e il parallelo 27°40’ (oggi facente parte del Marocco) e con statuto
di protettorato; dall’altra parte, più a sud, la Saguia el Hamra e il Rio de Oro con
statuto di colonia e formanti il Sahara Spagnolo.
Solo a partire dal 1934 gli spagnoli cominciarono a spingersi all’interno del
territorio installando postazioni militari e strutture amministrative. Nel 1934
l’amministrazione spagnola attribuì alla popolazione uno stato civile e un
documento di identità con l’introduzione di un visto obbligatorio per la
36

transumanza in territori francesi. Contemporaneamente inizia la formazione di una
resistenza da parte dei Saharawi contro lo sfruttamento e i soprusi coloniali. Nel
1961 furono creati degli organismi amministrativi locali, ed El-Aaiun divenne la
capitale del territorio.
Nel 1967 si istituì un’Assemblea Generale eletta dai Saharawi allo scopo di
rappresentarli. Tutte queste istituzioni contribuirono a formare un insieme
omogeneo che prese coscienza della sua unità e identità. Nel 1974 un censimento
effettuato stimava il totale della popolazione Saharawi di 73.497 persone, la
maggior parte delle quali vivevano nelle tre città principali: El-Aaiun, Smara e
Villa Cisneros. Le professioni più esercitate erano quelle del pastore e del
manovale.
E’ solo durante il Periodo Contemporaneo, alla fine degli anni ’50, che il
Popolo Saharawi cominciò ad affermare la sua indipendenza. Nel 1957-58 si ebbe
una prima grande sommossa che, a causa della repressione spagnola, spinse parte
della popolazione Saharawi a rifugiarsi nel sud del Marocco ed in Mauritania.
Una decina d’anni più tardi, dopo l’indipendenza dei paesi limitrofi, cominciarono
ad apparire vari movimenti nazionalisti (tra cui il Movimento di Liberazione del
Sahara con a capo Mohamed Bassiri) con l’intento di opporsi alla colonizzazione
spagnola. Ma nel 1970 ci fu una nuova durissima repressione con morti e
centinaia di arresti, tra cui lo stesso Bassiri, determinando un sentimento sempre
più crescente di identità nazionale che portò nel 1973 alla creazione del FRONTE
POLISARIO (Fronte Popolare di Liberazione della Saguia el Hamra e Rio de
Oro).
37

Nel 1956 il Marocco, dopo aver ottenuto l’indipendenza dalla Francia, rivendicò
un territorio che comprendeva il Sahara Occidentale, la Mauritania, parte
dell’Algeria e del Mali. La Mauritania dopo l’indipendenza (1960), pretese
anch’essa il dominio sul Sahara Occidentale. Questa situazione di conflitto
interruppe il processo di decolonizzazione iniziato dall’ONU e culminò con la
decisione della Spagna di cedere il Sahara Occidentale al Marocco e alla
Mauritania (Accordo di Madrid, 14.11.1975). Al momento del ritiro definitivo
della Spagna dai territori, il Fronte Polisario proclamò la Repubblica Democratica
Araba Saharawi (RASD., 27.02.1976), a cui seguì l’invasione del Sahara
Occidentale da parte del Marocco e della Mauritania che provocò un forte esodo
di rifugiati nei paesi vicini, soprattutto in Algeria (Hammada de Tindouf), e
l’intensificarsi dei movimenti armati del Fronte Polisario, tanto che la Mauritania
si ritirò definitivamente dai territori nel 1979.
Nel 1980 la RASD venne ammessa come stato membro dell’OUA
(Organizzazione per l’Africa Unita) e ottenne così il riconoscimento di Stato
Rifugiato da più di ottanta paesi. Nello stesso anno il Polisario recuperò una parte
di territorio dai marocchini costringendo Hassan II (regnante dal 1961 fino al
1999) a iniziare la costruzione dei “muri di sabbia” lungo gran parte del territorio.
Nell’agosto 1988 il Segretario Generale dell’ONU ottenne dal Marocco e dal
Fronte Polisario un accordo per un piano di pace. Il piano prevedeva il cessate il
fuoco, il dispiegamento di forze dell’ONU e un Referendum di
autodeterminazione, per sancire l’indipendenza o l’integrazione al Marocco.
38

Il 29 aprile 1991 il Consiglio di Sicurezza dell’ONU approvò la risoluzione
690/91 che stabiliva il piano di pace, mettendo in moto il meccanismo del
referendum e l’istituzione della missione di pace della MINURSO (Missione delle
Nazioni Unite per il Referendum nel Sahara Occidentale).
Il Referendum, inizialmente fissato per il settembre del 1992, viene
continuamente rimandato a causa dei continui ostracismi marocchini che ne
impediscono tuttora l’attuazione.
Il Marocco è sostenuto dalla Francia e dagli Stati Uniti, da cui viene considerato
un alleato storico e leale nella lotta al terrorismo e con cui ha firmato accordi di
sfruttamento delle risorse petrolifere. Il Polisario invece è appoggiato
dall’Algeria, la quale ha dichiarato di non accettare che la sovranità del Marocco
si estenda a tutto il Sahara Occidentale, e dalla Spagna che ha ribadito il sostegno
ad una soluzione del conflitto che favorisca la libera scelta della popolazione
Saharawi.
3.1.3 I CAMPI PROFUGHI SAHARAWI OGGI
Gli accampamenti dei rifugiati sono situati nella parte occidentale del deserto
algerino (Hammada de Tindouf), vicino al confine con il Sahara Occidentale, in
un’area concessa dal governo algerino che si estende per circa 10.000 Kmq2. E’
qui che a partire dal 1976, dopo un vero e proprio esodo attraverso il deserto, si
rifugiò parte della popolazione Saharawi a causa dell’invasione marocchina. Le
popolazioni arrivate fin qui si riunirono inizialmente attorno alle prime fonti
39

d’acqua del deserto algerino (a Rabouni) e successivamente si divisero in quattro
grandi tendopoli per evitare il rischio di epidemie.
Oggi la divisione amministrativa della RASD è composta dai Campi Profughi,
localizzati in territorio algerino, e dai Territori Liberati, la parte orientale del
Sahara occidentale, divisa in sei regioni militari.
Figura 12: tipica wilaia dei Campi Profughi (Sabatini L., 2008).
I Campi Profughi si sviluppano intorno a Rabouni, centro amministrativo e
governativo, sede dei ministeri e dell’ospedale nazionale ed anche centro di
accoglienza e di residenza delle associazioni umanitarie operanti nel territorio.
I Campi sono organizzati in 4 grandi tendopoli chiamate wilaia (“provincie”) (fig.
12) a cui è stato dato il nome delle città tuttora occupate dai marocchini: El-Aaiun,
ex capitale del Sahara Occidentale, Awserd, Smara e Dakhla, un tempo città
costiera. Negli ultimi anni si è formata un quinto centro abitato chiamato 27
Febrero che si può considerare ormai come un quinta wilaia, di dimensioni più
piccole, situata a circa 10 km da Rabouni dove è presente una scuola professionale
per le donne. Ogni wilaia è suddivisa fisicamente ed amministrativamente in 6-7
daira (“comuni”). Awserd e Smara comprendono oggi sei daira. Dakhla ed El-
40

Aaiun ne possiedono sette. Il 27 Febrero è costituito da un'unica daira. Ogni daira
è a sua volta suddivisa in baladiat (“distretti”).
Le tendopoli distano da Rabouni circa 35-50 Km (fa eccezione Dakhla che si trova
a 160 Km). Da Rabouni la strada asfaltata raggiunge solo Smara e il 27 Febrero,
mentre per raggiungere le altre wilaia bisogna seguire piste nel deserto
estremamente dissestate.
Le sei regioni militari presenti nei Territori Liberati sono numerate da 1 a 6 e
chiamate con i nomi delle principale città presenti: Bir Lehlou, Tifariti, Mehaires,
Mijek, Agwanit, e Dougaj. Le singole regioni sono amministrate dai rispettivi
comandanti militari del Fronte
Polisario (fig. 13).
Oggi l’intera popolazione
Saharawi è suddivisa tra i Campi
Profughi in Algeria (circa
165.000 abitanti), la parte
occidentale del Sahara
Occidentale sotto il controllo del
Marocco (circa 65.000 – 90.000
abitanti), la parte orientale del
Sahara Occidentale controllata
dal Polisario (circa 10.000 - 30.000 abitanti) e la Mauritania (circa 25.000
abitanti), mentre una piccola minoranza si divide tra Spagna, le Isole Canarie,
l’Algeria e il sud del Marocco (Arkell, 1991; UNHCR, 2002, 2005).
Figura 13: mappa del Western Sahara (UN, 2004).
41

Nei Campi la gente vive in tende di tela e case di fango e presentano numerose
difficoltà nell’approvvigionamento di cibo e acqua. L’unione Europea insieme a
donatori bilaterali, agenzie delle Nazioni Unite e gruppi di solidarietà europei,
forniscono loro cibo, alloggi e altre necessità (Loewenberg, 2005). Il contesto in cui si
trovano attualmente i Saharawi, costretti ad abbandonare le loro tradizioni e a
dipendere quasi esclusivamente dagli aiuti umanitari, li ha portati a lottare
attraverso la resistenza militare, sociale e culturale, per la loro identità che si sta
perdendo (Seddon, 2000; Shelley, 2004; Volpato et al., 2007).
3.2 POPOLAZIONE ED ECONOMIA
Attualmente la maggior parte della popolazione Saharawi vive tra i Campi
Profughi (165.000 abitanti) e i Territori Liberati (10.000-30.000 abitanti).
La lingua ufficiale è l’Arabo ma la lingua più parlata è l’Hassanya, un tipo di
dialetto diffuso anche in Mauritania. Come conseguenza del periodo coloniale, la
lingua spagnola è parlata dalla maggioranza della popolazione La religione
ufficiale è l’Islam, di dottrina sunnita.
I campi sono per la maggior parte abitati da donne, anziani e bambini, visto che la
maggioranza degli uomini sono impegnati al fronte o emigrati all’estero per
lavorare. Proprio per questo motivo la donna saharawi, a differenza del resto del
mondo arabo, ha assunto un ruolo importante all’interno dell’organizzazione
sociale.
42

Le tende degli accampamenti non sono le tradizionali jaimas di lana di
dromedario, ma tende militari in tela. Ognuna ospita un nucleo familiare e vicino
alla tenda ogni famiglia ha costruito inoltre delle strutture in mattoni di fango,
adibite a cucina e abitazione per i mesi più freddi.
L’illuminazione è fornita da lampade al neon, alimentate da batterie collegate a
piccoli pannelli solari, o da lampade a gas. L’acqua manca nelle wilaia di Awserd
e Smara e del 27 Febrero: per questo viene prelevata giornalmente dai pozzi di
Rabouni e trasportata con autobotti per riempire le cisterne presenti nei baladiat.
Nelle altre due wilaia di El-Aaiun e Dakhla l’acqua è reperibile in pozzi poco
profondi disseminati nelle varie daira.
La maggior parte dei fabbisogni della popolazione sono soddisfatti dagli aiuti
internazionali che sono gestiti e finanziati dall’UNHCR (Alto Commissariato
delle Nazioni Unite per i Rifugiati), dal PAM (Programma Alimentare Mondiale)
e da ECHO (Organismo per gli Aiuti Umanitari della Commissione Europea).
Solamente una minima parte dei fabbisogni alimentari è assicurata
dall’allevamento familiare (soprattutto ovi-caprini e dromedari) e dall’orticoltura.
Infatti, nonostante la scarsa fertilità del terreno e l’elevata salinità dell’acqua, si
coltivano cipolle, patate, legumi e qualche albero da frutto in orti comuni costruiti
all’interno delle wilaia.
Per anni il senso di provvisorietà ha prevalso tra la popolazione Saharawi, in
quanto il loro obbiettivo finale era quello di poter tornare in patria. Questo ha
influito negativamente sullo sviluppo della zootecnia, dell’agricoltura e
dell’economia in generale.
43

Oggi, dopo il fallimento da parte dell’ONU nell’istituzione del Referendum per il
Sahara Occidentale, le cose stanno cambiando: le abitazioni in mattoni di cemento
stanno lentamente sostituendo le tende ed i mattoni di fango e all’interno dei
centri abitati nascono un po’ ovunque piccole attività e negozi.
Nonostante una solida struttura sociale i Saharawi si trovano costantemente a
dover far fronte a grandi problemi, in particolar modo quelli sanitari. Le cattive
condizioni di salute interessano sia la popolazione che il sistema di allevamento
del bestiame e sono estremamente condizionate dall’insufficiente conoscenza in
materia e dalla mancanza di un sistema di prevenzione e controllo delle principali
malattie. Infatti i Saharawi hanno sempre basato la medicina su rimedi naturali
tradizionali sia per la salute umana che animale, spesso anche efficaci, ma non
sicuramente in grado di risolvere le carenze e combattere efficacemente le
malattie più diffuse (Volpato G., et al., 2007). Anche se l’instaurarsi di un sistema
sanitario basato su un modello di tipo occidentale ha portato, negli ultimi anni, al
miglioramento del sistema sanitario, resta il fatto che le condizioni di salute
estremamente precarie risultano essere un problema ancora attuale. Proprio per
questo motivo la popolazione è ancora fortemente legata alle organizzazioni
internazionali e al sostegno delle Organizzazioni Non Governative (ONG) che
agiscono sul territorio attraverso programmi di emergenza e di sviluppo (Olmi,
1998; Lopriore e Branca, 2001).
L’economia del Popolo Saharawi si basa principalmente sull'allevamento e sul
commercio del bestiame, spesso con i paesi vicini come la Mauritania, il Mali e
l’Algeria (fig. 14 e 15). Tuttavia le condizioni climatiche, la poca acqua e spesso
44

la carenza del foraggio a disposizione rendono molto difficile lo sviluppo di un
sistema forte ed indipendente.
Il commercio si effettua nei mercati presenti in tutte le wilaia, dove gli animali
vengono importati ed esportati (fig. 16 e 17). Spesso i Campi Profughi
rappresentano una vera e propria via di transito del bestiame proveniente da sud
(Mauritania, Mali e Territori Liberati) e diretto a nord (Algeria). Questo aspetto si
rivela di particolare importanza dal punto di vista epidemiologico.
Figura 14: animali entranti dalla Mauritania Figura 15 : mercato del bestiame di Smara (Rossi D., 2008). (Rossi D., 2008).
Figura 16: trasporto degli ovi-caprini Figura 17: trasporto dei dromedari
(Rossi D., 2008). (Rossi D., 2008).
45

Da sempre la principale fonte di sussistenza dei nomadi saharawi è rappresentata
dalla pastorizia. Dopo la guerra la maggior parte della popolazione ha perso le
proprie mandrie, dovendo così abbandonare la pastorizia. Dopo un primo periodo
di emergenza i rifugiati saharawi hanno ripreso le attività pastorali, nonostante i
problemi dovuti alla condizione sedentaria e l’assenza di pascoli nei Campi
Profughi. Da questa situazione ha preso vita un regime di semi-nomadismo per cui
le popolazioni si spostano verso i Territori Liberati durante la stagione delle
piogge, alla ricerca di pascoli per il bestiame, permettendo così la conservazione
delle tradizioni del Popolo Saharawi (Broglia A. e Volpato G., 2008).
3.2.1 TECNICHE DI ALLEVAMENTO
Nella RASD esistono due tipi di allevamento: l’allevamento di stato e quello
privato.
L'allevamento di stato comprende alcune grosse mandrie di dromedari (in totale
circa 11.000 capi dal censimento 2007 della Direcciòn de Veterinaria Saharawi)
allevate nei Territori Liberati ed oltre il confine con la Mauritania. Ogni mandria è
formata da un numero che varia tra gli 80 e i 100 capi: un maschio intero, il
gruppo delle femmine ed i giovani. Di solito un maschio è sufficiente per 100
femmine; gli altri vengono castrati all’età di 2-3 anni secondo la metodica
tradizionale a scroto aperto che prevede un taglio netto del funicolo spermatico e
l’utilizzo della sabbia per tamponare l’emorragia.
46

Il sistema di allevamento è quello nomade: uomini e mandrie si spostano
periodicamente con l’alternanza delle stagioni, alla ricerca di fonti d’acqua e di
pascolo, assicurando un equilibrio tra le risorse pastorali ed il bestiame.
L’intera struttura del Polisario e dei singoli ministeri possiede queste mandrie che
vengono utilizzate come fonte di autofinanziamento. Vi sono inoltre alcune
piccole mandrie stanziali di dromedari allevate nelle tendopoli che servono a
fornire latte a strutture sanitarie adiacenti (ospedali, scuole, un centro per
tubercolotici, un centro per bambini con ritardo di crescita).
L’allevamento di stato nei Territori Liberati comprende anche ovini e caprini
allevati in gruppi di 50-200 animali con il tradizionale sistema nomade, come le
mandrie di dromedari (in totale circa 30.000 capi dal censimento della Direccòn
de Veterinaria Saharawi).
A Rabouni, inoltre, esiste un allevamento industriale di galline ovaiole gestito dal
Ministero della Cooperazione Saharawi. Questo consta di tre capannoni ognuno
dei quali può contenere fino a 25.000 animali, ma attualmente solo uno è in
funzione.
L’allevamento privato è principalmente costituito da ovini, caprini e dromedari.
Per quanto riguarda l’allevamento ovi-caprino l’ultimo censimento del 2007 ha
identificato più di 60.000 capi di cui il 51% è rappresentato da ovini ed il 49% dai
caprini (Direciòn de Veterinaria, 2007).
Questi animali durante il giorno vengono lasciati liberi in greggi comuni alla
ricerca dello scarso pascolo che questa regione desertica può offrire. Dopo il
47

tramonto vengono rinchiusi in recinti di fortuna che ospitano in media dai 6 ai 10
capi. Questi recinti, chiamati corrales, sono collocati alla periferia delle daira per
motivi igienico-sanitari e sono costruiti con materiali di recupero tra i più
disparati: pezzi di lamiera, reti metalliche e rottami di auto e camion che
conferiscono a queste strutture un’ottima funzionalità. Essi, infatti, servono a dare
un riparo alle greggi nelle ore più calde e a rinchiuderli durante la notte e all’ora
della mungitura (fig. 18).
Figura 18: tipici corrales alla periferia delle wilaia (Sabatini L., 2008).
Il pascolo è incapace di coprire i fabbisogni alimentari delle greggi. Perciò
l’alimentazione degli ovi-caprini è costituita essenzialmente dai residui
dell’alimentazione umana (pane, pasta, riso, farine, legumi, vegetali, fondi ti tè
ecc.). Per integrare la carenza di fibra grezza indispensabile per la ruminazione
(questo tipo di alimentazione rende di fatto questi animali dei monogastrici)
vengono forniti anche pezzi di cartone o qualsiasi altro materiale cartaceo. Spesso
48

gli animali sono costretti a cercare il cibo tra le immondizie, dove ingeriscono
stracci, plastica e altri rifiuti, causando gravissimi problemi digestivi (fig. 19 e
20): questi materiali cadono nel rumine e qui vi si accumulano nel tempo,
formando grosse matasse che esitano in atrofia delle papille ruminali, indigestioni,
stenosi, occlusioni e gravi ruminiti. Tra gli ovi-caprini è segnalata anche un’alta
frequenza di coprofagia che può essere spiegata da un punto di vista dietetico-
carenziale.
Figura 19: pecora in cerca di cibo Figura 20: gregge tra l’immondizia (Rossi D., 2008). (Sabatini L., 2008).
Le razze allevate sono tutte di tipo saheliano. I caprini appartengono a razze
Canaria, Arabe, Busghender e Sika; gli ovini a razze Tadamant e Akarran. Gli
animali che vengono acquistati provengono dal Senegal, dalla Mauritania e
dall’Algeria. Le razze saheliane sono il risultato di un lungo processo di selezione
e di adattamento a queste dure condizioni ambientali che le ha rese
particolarmente resistenti, di moderate necessità nutrizionali e insostituibili
nonostante la loro scarsa produttività.
Spesso gli animali vengono acquistati nei paesi limitrofi quali Mali, Mauritania e
a volte persino in Senegal. Una volta giunti nelle wilaia, gli animali devono
49

ricevere il controllo sanitario da parte dei servizi veterinari saharawi, il quale
consta attualmente di una visita clinica e del Test Rosa Bengala per la diagnosi di
brucellosi la quale attualmente è assente. Spesso però alcuni animali appartenenti
a commercianti non rispettosi delle leggi sfuggono a questo controllo.
Gli ovini vengono allevati principalmente per la produzione di carne, mentre i
caprini per la produzione del latte. Raramente il latte viene consumato fresco: man
mano che viene munto si raccoglie in sacche di pelle e lasciato acidificare
naturalmente all’interno della tenda. Completata l’acidificazione viene separato il
burro mediante sbattimento, mentre il latte viene posto in grandi tazze e
consumato dopo l’aggiunta di acqua fresca e zucchero.
La macellazione degli ovi-caprini non è frequente. Avviene soprattutto in
concomitanza con le feste e ad essere sacrificati sono i giovani maschi in quanto
le femmine sono troppo importanti sia per la riproduzione che per il latte.
Le macellazioni, sia che si tratti di quelle domestiche che di quelle commerciali,
cioè di animali la cui carne è destinata alla vendita, avvengono sempre secondo il
rito islamico. Questo è regolamentato da precise leggi coraniche che devono
essere rigorosamente rispettate affinchè la carne possa essere definita halal,
ovvero “lecita”.
Normalmente il sacrificio avviene lontano dalle aree destinate ai corrales. La
macellazione avviene direttamente sul campo e le parti non utilizzate (pelle,
estremità degli arti, coda, organi genitali, vescica, cistifellea, contenuto gastrico ed
intestinale) vengono normalmente abbandonate nel luogo del sacrificio (fig. 21).
50

Figura 21: area destinata alla macellazione nella periferia di Dakhla (Sabatini L., 2008).
Per quanto riguarda la macellazione commerciale, l’unica carne ad essere venduta
è quella di dromedario. La maggior parte degli animali proviene dai Territori
Liberati, dalla Mauritania, dal Mali e dall’Algeria ed a volte da paesi sahariani e
saheliani molto più lontani.
Prima della macellazione è prevista per ogni animale una visita clinica ante-
mortem da parte di un tecnico veterinario.
Terminata la macellazione la carne viene caricata su delle Land-Rover e portata al
Dipartimento di Veterinaria della wilaia competente, dove viene eseguita
l’ispezione delle carni.
Nei Campi Profughi l'ispezione veterinaria non è obbligatoria per le macellazioni
familiari che vengono condotte secondo la maniera tradizionale, spesso mettendo
la popolazione a serio rischio di zoonosi. I visceri sono spesso abbandonati e
costituiscono cibo per cani randagi ed uccelli con il risultato di una potenziale
rapida diffusione delle malattie infettive (Africa ’70, 2007).
Le macellerie dove viene venduta la carne sono piccole costruzioni in mattoni di
fango essiccato con il tetto in lamiera ed una piccola finestra da dove la carne
51

viene esposta e venduta. Il livello di igiene è insufficiente, anche se, grazie al
lavoro svolto da Africa '70 e dalla Dirección de Veterinaria Saharawi attraverso
corsi di formazione ai macellai, interventi di ristrutturazione delle macellerie e
campagne di sensibilizzazione della popolazione, si sono ottenuti negli ultimi anni
notevoli miglioramenti nel sistema di vendita. Nonostante questo però ancora oggi
i risultati non sono del tutto soddisfacenti.
3.2.2 I SERVIZI VETERINARI SAHARAWI
Nella RASD i servizi veterinari sono organizzati nella Dirección de Veterinaria
(DV) la quale è inquadrata all'interno del Ministero della Sanità. La sede centrale
della DV è ubicata a Rabouni, vicino all'Ospedale Nazionale, mentre in ciascuna
wilaia è presente un Dipartimento di Veterinaria dove lavorano medici, tecnici ed
ausiliari veterinari. Medici e tecnici si sono diplomati in università cubane grazie
ad accordi esistenti tra il governo cubano ed il Fronte Polisario. Gli ausiliari sono
stati formati in loco durante l'ultimo progetto coordinato da Africa '70. Attraverso
le attività di cooperazione di Africa '70 presente in loco fin dal 1999, la DV è stata
formata, organizzata, potenziata ed equipaggiata. Le è stata conferita una struttura
piramidale con la presenza di un Direttore Centrale e di Direttori di Dipartimento
che coordinano le attività dei vari tecnici, ausiliari e donne coadiuvanti veterinarie
presenti in ogni daira e baladiat. Questo personale è stato dislocato sul territorio
in maniera uniforme in modo tale da garantire un’assistenza adeguata a tutti gli
allevatori delle wilaia.
Le attività della Direzione di Veterinaria prevedono:
52

- ispezione ante e post-mortem degli animali le cui carni sono destinate alla
vendita;
- vigilanza periodica delle macellerie per valutarne le condizioni igienico-
sanitarie;
- attività di sensibilizzazione e formazione degli addetti al settore;
- attività clinica di appoggio ai problemi sanitari del bestiame allevato;
- educazione sanitaria della popolazione soprattutto per quanto riguarda le
zoonosi ed il rischio sanitario connesso;
- attività di consulenza per gli allevatori al fine di migliorare le tecniche di
allevamento;
- periodici studi epidemiologici sulla situazione sanitaria del bestiame.
3.3 ASPETTI CLIMATICI E TERRITORIALI
Il Sahara Occidentale è un’area di 284.000 Km2 situata tra il 20° e il 30° parallelo
vicina al tropico del Cancro, che può essere divisa in tre ecosistemi distinti: il
primo a nord-est si estende dalla catena montuosa dell’Atlas alle colline dello
Zemmour. Si tratta di un deserto roccioso (Hammada) caratterizzato da altopiani
brulli e pochi punti d’acqua. E’ la zona più simile al territorio dove sorgono i
Campi Profughi. Il secondo è costituito dai fiumi Wadi Draa a nord e il Jat a
ovest. Qui l’acqua si raccoglie nella breve stagione delle piogge, ma, a causa
dell’elevata temperatura, evapora rapidamente. Questa zona è attraversata dal
Saguiat el-Hamra che dà il nome alla regione e che garantisce una sufficiente
53

vegetazione per il pascolo ed un terreno favorevole alla coltivazione del
frumento. L’ultimo ecosistema è quello del Rio de Oro. Questo si trova all’interno
e si estende fino a sud. E’ formato da pianure e dune di sabbia. Qui la terra è
troppo permeabile per trattenere l’acqua e troppo piana per permetterne il suo
scorrimento, per cui l’acqua viene ad accumularsi in falde freatiche presenti nel
sottosuolo formando numerosi pozzi, ma avendo un’elevata salinità non risulta
essere potabile ed è quindi di difficile utilizzo agricolo.
Nel Sahara Occidentale l’ecosistema è molto variabile. L’ambiente cambia da
regione a regione ed è possibile trovare caratteristiche differenti in ogni territorio.
Le wilaia si trovano in uno dei deserti più inospitali della terra, l’Hammada,
anche noto come il “giardino del diavolo”. Si tratta di un altopiano desertico a
circa 500 metri di altitudine, composto interamente da rocce e sabbia (fig. 22).
Figura 22: deserto dell’Hammada o “giardino del diavolo” (SaharaMet.org).
54

Qui la temperatura può oscillare dai -5°C in inverno fino ad oltre i 50°C in estate.
I venti, quasi sempre presenti, determinano molto spesso la produzione di
tempeste di sabbia. Le piogge sono rare e spesso, durante gli episodi piovosi di
forte intensità si vengono a creare delle vere e proprie inondazioni, come
registrato ad El-Aaiun nel 1993 dove circa 4.000 persone rimasero senza riparo e
come si è ripetuto nei campi di Smara ed El-Aaiun nel 2006 dove le aree rimasero
allagate per diversi mesi (fig. 23 e 24).
Figura 23: alluvione a Smara 2006 (Rossi, 2006). Figura 24: alluvione a El-Aiun 2006 (Rossi,2006)
A causa della scarsità di piogge, la vegetazione è quasi inesistente, tranne per
alcuni alberi di acacia (Acacia spp.) ed alcune specie di alofite, come lo
Zygophyllum gaetulum.
L’acqua è reperibile a breve profondità (2-6 metri), ma avendo un’elevata salinità
non è potabile e di difficile uso agricolo.
La regione del Sahara Occidentale comprendente i Territori Liberati è
caratterizzata, invece, da un clima desertico di tipo sub-oceanico dove la
mancanza di piogge è parzialmente compensata dall’elevata percentuale di
55

umidità dell’aria. Tale proprietà determina lo sviluppo di differenti tipologie
vegetative, come piante annuali (es. Diplotaxis pitiridana, Cotulea cinerea,
Asphodelus tenuifolius, Astragalus spp.), alcune specie di alofite della famiglia
delle Chenopodiacee (es. Trangum nudatum, Salsola spp., Cornulacea
moncantha, Nucularia perrinii) ed alberi di acacia (Ozenda, 1991). Inoltre, i
Territori Liberati sono caratterizzati da una forte biodiversità, con un ecosistema
che risulta differenziarsi drasticamente da nord a sud.
La regione di Bir Lehlou (situata a nord-est, la più vicina ai Campi) è quasi
paragonabile a quella delle wilaia, con distese di sabbia battuta e pianure rocciose,
dove è presente una vegetazione più diffusa, tanto che dopo le piogge il paesaggio
assume le caratteristiche di una savana (fig. 25 e 26).
Figura 25: regione di Bir-Lehlou. Figura 26: ecosistema dopo le piogge. (Rossi D., 2008). (Rossi D., 2008).
Le regioni di Tifariti e Mehaires (situate a nord-ovest) si trovano ad altitudini più
elevate e sono caratterizzate da un terreno collinare il cui paesaggio nei pressi
dell’altopiano di Tifariti può diventare anche molto ricco di vegetazione, tipico
esempio di paesaggio saheliano. Qui infatti le regioni sono più umide e anche se
56

le piogge sono comunque variabili, il tasso di precipitazione è al di sopra della
media (fig 27 e 28).
Figura 27: regione di Tifariti Figura 28: ecosistema della regione di Tifariti (Sabatini L., 2008). (Rossi D., 2008).
Questo tipo di ecosistema è presente anche nella parte settentrionale della regione
di Mijek (regione centrale dei territori Liberati), ma gradualmente si differenzia
per la presenza di banchi di sabbia nella zona più meridionale.
La regione di Agwanit (regione centro-orientale) è molto più arida ed è
caratterizzata da pianure di sabbia e di roccia dalle quali si innalzano isolate
formazioni di granito (fig. 29).
Figura 29: tipicho ecosistema della regione di Agwanit (Sabatini L., 2008).
57

Le uniche dune di sabbia presenti nei Territori Liberati si trovano nei pressi di Zug
che fa parte della regione militare di Dougaj (ultima regione all’estremo sud) (fig.
30).
Figura 30: dune di sabbia presenti nella regione di Dougaj (Rossi D., 2008).
3.4 MATERIALI E METODI
Il monitoraggio epidemiologico di una data popolazione animale e la valutazione
dei rischi legati alla diffusione di specifiche malattie infettive vengono
“idealmente” condotti da autorità sanitarie di competenza nazionale e delineati
all’interno di programmi di sorveglianza e controllo. Purtroppo, l’assenza di tali
sistemi di controllo governativi nella RASD, come in altri paesi in via di sviluppo
del continente africano, ha costituito un grosso ostacolo nell’approccio allo studio
e nell’indagine epidemiologica oggetto di questa tesi. Oltre alla mancanza di
informazioni storiche sulla prevalenza e diffusione della PPR nel territorio di
studio, un altro importante ostacolo è stato la presenza di campi minati, che hanno
impedito l’accesso ad alcune zone che, conseguentemente, non sono state inserite
58

nell’indagine. Inoltre il sistema di allevamento nomade ha reso laboriosa la scelta
del metodo di campionamento ed il calcolo statistico del campione oggetto di
studio, che è stato, per tanto, accuratamente elaborato per il caso specifico (Di
Nardo, 2008).
3.4.1 PIANO DI CAMPIONAMENTO
Lo studio del livello di rischio che la Peste dei Piccoli Ruminanti può
rappresentare per l’intera popolazione animale suscettibile presente nella RASD,
ha richiesto l’elaborazione di un protocollo d’indagine epidemiologica specifico al
fine di valutare la presenza, e nel caso, la prevalenza e la distribuzione di tale
malattia nella suddetta popolazione. Tuttavia a causa della tipologia di
allevamento presente in questa area i classici metodi di campionamento per
randomizzazione, necessari per estrarre valide conclusioni circa la popolazione,
non sono risultati essere applicabili ed inoltre l’utilizzazione su tutto il territorio
della RASD sarebbe risultata dispendiosa sia in termini di tempo che di costi.
Anche la mancanza di una completa lista delle unità campionarie presente
nell’area geografica ed il caratteristico allevamento di tipo nomade, che non
permette la precisa localizzazione delle mandrie, avrebbero reso tale approccio
inadatto (Di Nardo, 2008).
Per questi motivi, è stato necessario sviluppare un metodo di campionamento
ragionato ed adatto al contesto, in grado di far fronte alle difficoltà descritte che
permettesse di ottenere risultati validi.
59

Conseguentemente, nella selezione del campione oggetto di studio è stata
utilizzata una metodica di campionamento “a cluster” secondo la tecnica a due
stadi (Thrusfield, 2007), considerata come la più indicata per le indagini
epidemiologiche condotte in paesi in via di sviluppo (Henderson e Sundaresan, 1982;
Bennet et al., 1991; Otte e Gumm, 1997). Tale metodica consiste nel selezionare un certo
numero di gruppi, mutuamente esclusivi, chiamati cluster (nella maggioranza dei
casi tali cluster vengono ad essere definiti su base spaziale), e all’interno di questi
scegliere degli individui per randomizzazione. Di conseguenza i cluster
rappresentano le unità del primo stadio e gli elementi estratti da questi (individui)
quelli del secondo stadio (Thrusfield, 2007). Nello studio, è stato identificato come
cluster ogni accampamento, punto di abbeveraggio o area di pascolo, dove la
probabilità di reperire mandrie risultava essere più elevata. Inoltre, tali cluster
sono stati selezionati anche in base alle differenze nell’ecosistema, nella struttura
della popolazione animale, e nel sistema di allevamento presenti tra le wilaia e i
Territori Liberati (Di Nardo et al., 2009).
Figura 31: Localizzazione dei singoli cluster riferiti alle singole Wilaia e Regioni Militari. (Di Nardo et al., 2009)
60

Successivamente, sono state identificante 11 aree rispondenti ai requisiti sopra
citati: un cluster per ogni wilaia (n=5) e 18 per le 6 Regioni Militarei, dei Territori
Liberati, per un totale di 23 siti di campionamento (Fig. 31).
Poiché nella rappresentazione spaziale, i cluster vengono definiti come singoli
punti (identificati mediante coordinate geografiche e raggiungibili attraverso
l’utilizzo di apparecchiature GPS), è stato necessario definire un raggio fisso per
ciascun zona, in modo tale da determinare l’area geografica in cui gli animali
venivano ad essere campionati ed aumentarne, di conseguenza, la probabilità di
reperimento del campione. Nello specifico, poiché il raggio viene calcolato in
dipendenza dalla densità della popolazione animale, sono stati prestabiliti un
raggio di 5km per ogni cluster delle wilaia ed uno di 20km per quelli delle
Regioni Militari (Di Nardo et al., 2009).
3.4.2 ELABORAZIONE STATISTICA DEL CAMPIONE DI STUDIO
In assenza di dati epidemiologici riferiti alla PPR nel territorio del Sahara
Occidentale precedenti allo studio condotto, per il calcolo del campione è stato
utilizzato un approccio di natura conservativa, considerando il paese come
endemico a bassa prevalenza (10-20%) (Di Nardo, comunicazione personale, 2009).
Come discusso in dettaglio, il calcolo è stato effettuato mediante utilizzo della
seguente formula, fissando il livello di confidenza al 95% (Thrusfield, 2007):
61

dove: g = numero di cluster da campionare
Pexp = prevalenza attesa
d = precisione assoluta desiderata
Vc = varianza tra cluster
Nel caso specifico, la Pexp è stata fissata al 18%, la d pari a 0.05 (ossia del 5%) e
la Vc con valore di 0.002635 (stimata da dati OIE riferiti alla Mauritania per
l’anno 2003). La popolazione animale oggetto di studio comprende capre, pecore
e dromedari presenti nelle singole wilaia e nelle singole Regioni Militari dei
Territori Liberati. Nella selezione del campione sono stati presi in considerazione
anche i dromedari, in quanto anche questi animali sembrano essere recettivi al
virus dalla PPR: studi condotti in Etiopia hanno dimostrato infatti come questa
specie animale possa svolgere un ruolo importante nell’epidemiologia di tale
malattia (Roger et al., 2001) e, anche se il decorso appare sempre in forma
subclinica, la presenza di anticorpi attesta un avvenuto contatto con il virus e
quindi un possibile stato di portatore.
1.962gP (1-P ) exp exp_________________
gd2 – 1.962Vc
62

Dromedario Pecora Capra TOT
WILAIA El Aaiun 140 7424 7657 15221Awserd 89 4675 5035 979927 Febrero 12 355 378 745Dakhla 125 3729 3518 7372Smara 92 7628 7537 15257TOT 458 23811 24125 48394
REGIONI MILITARI (Territori Liberati) Dougaj 1715 1395 1014 4124Tifariti 5090 9940 11500 26530Mijek 12000 18000 8000 38000Mehaires 3000 5700 3800 12500Bir Lehlou 1052 845 600 2497Agwanit 2860 2990 2610 8460TOT 25717 38870 27524 92111
Popolazione animale totale 26175
62681
51649 140505 Tabella 1: popolazione animale presente nel territorio della RASD filtrata per specie (NVD-SARD 2007)
La grandezza del campione è stata calcolata per ogni specie animale, tenendo in
considerazione i dati provenienti dal censimento del 2007 (NVD-SADR, 2007) (Tab.
1) ed utilizzando la formula di cui sopra.
In tabella 2 è riportata la grandezza del campione ottenuta (n = 1086.35),
suddivisa per specie animale (pecore, n = 360.99; capre, n = 361.44; dromedari, n
= 363.92) e per singolo cluster di campionamento (n = ~33).
Pecora Capra Dromedario Popolazione totale 62681 51649 26175 Cluster 11 11 11 Grandezza campione
360.99 (32.82/cluster)
361.44 (32.86/cluster)
363.92 (33.08/cluster)
Totale campione 1086.35 Tabella 2: Grandezza del campione di studio calcolata per singola specie animale
63

Successivamente, la validità del calcolo del campione è stata analizzata attraverso
CSurvey 2.0 (UCLA, Los Angeles, CA, USA) (Farid e Frerichs, 2007), valutando i limiti di
confidenza e l’errore standard come parametri testati in funzione del teorema
centrale limite (distribuzione Normale). Tutti i parametri computati dal
programma sono risultati in accordo con il campione ottenuto, determinandone la
correttezza nell’approccio al calcolo (Di Nardo, comunicazione personale, 2009) e ogni
animale è stato campionato in modalità assolutamente casuale, attraverso
generazione di tabelle di randomizzazione in Survey Toolbox 1.0b (ACIAR,
Camberra, AU) (Cameron, 1999). Tale metodo risulta necessario per evitare una
selezione viziata in fase di campionamento che porti ad errori sistematici
(campionario e non-campionario), per cui il campione risulterebbe non
rappresentativo o distorto (non corrispondente alla vera caratteristica di
popolazione) (Di Nardo et al., 2009).
3.4.3 SCREENING ANTICORPALE NELLA RASD
Il campionamento è stato svolto tra Marzo ed Aprile 2008, coprendo l’intero
territorio della RASD, ossia le wilaia ed i Territori Liberati, in base a quanto
dettagliato nel protocollo d’indagine. Tale esercizio ha coinvolto le associazioni
veterinarie della RASD, sotto il coordinamento del Dipartimento Veterinario del
Ministero della Salute Pubblica e con la supervisione di Africa ’70 (ONG italiana
finanziata dalla Commissione Europea).
I campioni ematici sono stati raccolti dalla vena giugulare utilizzando aghi sterili e
provette Venoject® e correttamente conservati ad una temperatura di
64

refrigerazione (~4°C). In seguito è stata effettuata la separazione del siero
mediante utilizzo di una centrifuga portatile da campo e i sieri ottenuti sono stati
prontamente congelati a -21°C nelle basi regionali della MINURSO, garantendo il
mantenimento della catena del freddo. Alla fine del periodo di campionamento, i
sieri sono stati trasferiti in Italia presso l’Istituto Zooprofilattico Sperimentale
(IZS) dell’Abruzzo e del Molise “G. Caporale”, dove sono stati analizzati
mediante ELISA competitiva (test di referenza OIE).
3.4.4 PROTOCOLLO ELISA–COMPETITIVA
Per testare la presenza di anticorpi contro la PPR nei campioni raccolti è stato
utilizzato il test ELISA, sviluppato e validato dal CIRAD (Centre de coopération
Internationale en Recherche Agronomique pour le Développement) e prodotto e
distribuito come kit dalla BDSL-2000 (Biological Diagnostic Supplies Limited,
UK), seguendo il protocollo fornito (Libeau et al, 1995). Nello specifico si tratta di
un’ELISA di tipo competitivo; tale metodologia è basata sull’abilità di inibire il
legame dell’anticorpo monoclonale (mAb) ad un epitopo PPR-specifico in
presenza di siero positivo. Tale inibizione viene ad essere rilevata come una
riduzione nel valore di densità ottica (OD), ottenuta mediante aggiunta del
coniugato e del substrato di attivazione enzimatica.
Il kit fornisce i seguenti reagenti liofilizzati:
- Antigene ricombinante N-PPRV, prodotto in cellule di insetto.
- Anticorpo monoclonale anti-PPRV (mAb anti-PPRV).
65

- Anticorpo anti-IgG di topo coniugato con perossidasi.
- Soluzione di stop [Acido solforico (H2SO4) 1 M].
- Siero di controllo fortemente positivo (C++).
- Siero di controllo positivo (C+).
- Siero di controllo negativo (C−).
Non sono invece compresi:
- Piastre ELISA Immuno MaxiSorp (ditta NUNC)
- Substrato = Tetrametilbenzidina (TMB) (ditta SIGMA)
- Tween-20 (ditta SIGMA)
Il protocollo prevede le seguenti fasi:
1. Fase di coating (o adsorbimento in fase solida): un volume di
50µl/pozzetto di Ag N-PPRV, diluito 1:3000 in PBS, viene utilizzato per
l’adsorbimento passivo dell’Ag alle pareti del pozzetto, mediante
incubazione a 37°C per 1h.
2. Fase di lavaggio: effetuata 3 volte con soluzione di lavaggio (PBS diluito
1:5 in acqua distillata/deionizzata + 0,05% Tween-20).
3. Fase di blocco: il legame Ag-fase solida viene mantenuto saldo attraverso
l’aggiunta di 45µl/pozzetto di soluzione di blocco (PBS diluito 1:5 in
acqua distillta/deionizzata + 0,05% Tween-20 + 0,5% C−).
Allestimento dei pozzetti di controllo (Tab. 3):
- 4 pozzetti (F1, F2, G1 e G2) per il controllo del monoclonale (Cm):
5µl/pozzetto di soluzione di blocco.
66

- 2 pozzetti (H1 e H2) per il controllo del coniugato (Cc): 50µl/pozzetto di
soluzione di blocco.
- 4 pozzetti (A1, A2, B1 e B2) per il controllo fortemente positivo (C++):
5µl/pozzetto di siero di controllo.
- 4 pozzetti (C1, C2, D1 e D2) per il controllo positivo (C+): 5µl/pozzetto di siero
di controllo.
- 2 pozzetti (E1 ed E2) per il controllo negativo (C-): 5 µl.
Successivamente, i sieri da testare vengono aggiunti in duplicato (A-H 3-12) in
ragione di un volume pari a 5µl/pozzetto.
1 2 3 4 5 6 7 8 9 10 11 12
A Cc Cc 1 1 9 9 17 17 25 25 33 33 B C++ C++ 2 2 10 10 18 18 26 26 34 34 C C++ C++ 3 3 11 11 19 19 27 27 35 35 D C+ C+ 4 4 12 12 20 20 28 28 36 36 E C+ C+ 5 5 13 13 21 21 29 29 37 37 F Cm Cm 6 6 14 14 22 22 30 30 38 38 G Cm Cm 7 7 15 15 23 23 31 31 39 39 H C- C- 8 8 16 16 24 24 32 32 40 40
Tabella 3: Layout piastra microtitre nell’ELISA-CIRAD
4. Fase di aggiunta dell’Ab marcato: un volume di 50/µl di mAb anti-PPRV,
diluito 1:50 in soluzione di blocco, viene aggiunto in tutti i pozzetti ad
eccezione di quelli per il controllo del coniugato.
5. Successivamente le piastre vengono lasciate ad incubare a 37°C per 1h.
6. Fase di lavaggio: effettuata 3 volte con soluzione di lavaggio.
7. Fase di aggiunta dell’enzima coniugato: in piastra viene aggiunto un
volume di 50µl/pozzetto di Ab-anti-IgG di topo coniugato con perossidasi
67

(HRPO) diluito 1:1000 in soluzione tampone, lasciando incubare a 37°C
per 1h.
8. Fase di lavaggio: effettuata 3 volte con soluzione di lavaggio.
9. Fase di aggiunta del substrato per l’enzima: un volume di 100µl/pozzetto
di TMB viene addizionato alla piastra, lasciata incubare per 20min al buio
ed a temperatura ambiente.
10. Fase di arresto della reazione enzimatica: al termine dei 20min viene
aggiunto un volume di 100µl/pozzetto di soluzione di stop (50µl di H2SO4
1M).
11. Fase di lettura: la lettura viene effettuta mediante spettrofotometro ad una
lunghezza d’onda di 492 nm.
L’inibizione del legame del mAb all’antigene PPRV, visualizzata come riduzione
nella misura della densità ottica, indica la presenza di anticorpi specifici nei sieri e
viene espressa come percentuale di inibizione (PI) del controllo del monoclonale
secondo la seguente formula:
PI = 100 − [(net-OD del campione / net-OD del Cm) x 100]
Il protocollo per l’ELISA-CIRAD possiede dati di controllo qualità (QA) al fine
di validare correttamente l’analisi diagnostica (Tab. 4) (Libeau et al., 1995). In tali
range di dati rientrano i limiti nei valori del net-OD e delle PI riferiti ai sieri di
controllo.
68

Internal Quality Control
(IQC) Upper Control Limit (UCL) Lower Control Limit (LCL)
Cm (OD) 1.500 0.500
Cc (PI) +105 +95
Cm (PI) +20 -19
C++ (PI) 90 81
C+ (PI) 80 51
C- (PI) 30 5
Tabella 4: dati di controllo qualità riferiti all’ELISA CIRAD (il dato in rosso è riferito al valoredi cut-off (Libeau et al., 1995)
Quando il valore di controllo del C+ non supera il valore limite inferiore di 50,
vengono considerati come dubbi tutti quei campioni che presentano valori di PI
compresi tra quello del C+ e 50. Il valore di cut-off (o valore soglia
positivo/negativo) viene definito dal 50% d’inibizione del Cm (50PI),
conseguentemente un valore di PI>50 definisce un risultato come positivo, mentre
un valore di PI<50 come negativo. Inoltre, essendo il test eseguito in duplicato per
ogni singolo campione, entrambi i valori di PI, riferiti a ciascun siero, devono
risultare al di sopra o al di sotto della soglia di cut-off, in caso contrario il test non
è da considerarsi attendibile e deve essere quindi ripetuto.
69

4. RISULTATI
Conformemente al protocollo d’indagine, l’area definita per ogni cluster ha
permesso il campionamento all’interno del sistema di allevamento nomade
presente nella RASD per un totale di 23 siti di raccolta suddivisi in base a quanto
precedentemente descritto. (Fig. 32).
14
13
9
1 2
3 4
23
5
6
7 8
10
11 12
15 16
17
18
19 20
21
22
Figura 32 - Localizzazione dei siti di campionamento nell’area di studio (Di Nardo, 2008)
Dei 1086 campioni totali calcolati, ne sono stati effettivamente raccolti solo 976.
Questa riduzione nel numero totale dei campioni è stata essenzialmente dovuta
all’estrema diffidenza dei pastori nomadi, soprattutto proprietari dei dromedari, nel
permettere l’esecuzione dei prelievi, ed alla difficoltà nella metodica di
separazione dei sieri che è stata condotta in condizione molto disagiate.
70

Dai risultati ottenuti attraverso analisi ELISA, è stata elaborata un’indagine
statistico-analitica specifica al fine di definire accuratamente la situazione
epidemiologica riferita alla Peste dei Piccoli Ruminanti nella RASD. L’analisi è
stata rivolta ad evidenziare i risultati di sieroprevalenza, la correlazione esistente
tra sieroprevalenza riscontrata e dati epidemiologici riferiti alla popolazione
animale presente ed, infine, alle caratteristiche spaziali di sieroprevealenza
analizzando la dimensione dei cluster. L’elaborazione dei dati è stata eseguita
attraverso l’utilizzo di EpiInfo 3.5.1 (CDC, Atlanata, GA, USA) (Burton et al., 1990) e R
2.9.0 (www.R-project.org) per l’analisi di sieroprevalenza e di correlazione, mentre
l’elaborazione dell’analisi spaziale è stata eseguita in SaTScan 8.0 (Kulldorff, 2009).
4.1 ANALISI DI SIEROPREVALENZA
I risultati sono stati analizzati come sieroprevalenza generale, sieroprevalenza per
singole regioni e sieroprevalenza per cluster, filtrando, inoltre, ciascun dato per
singole specie animali.
4.1.1 SIEROPREVALENZA GENERALE
Dei 976 campioni testati 264 (29.3%; 95% IC 28.9% ; 29.7%) sono risultati essere
positivi per IgG nei confronti del PPRV (Tab. 5).
71

N/N totale campionato Sieroprevalenza (95% IC)
Positivi 264/976 29.3% (28.9% ; 29.7%)
Negativi 712/976 70.7% (70.3% ; 71.1%)
Tabella 5: Sieroprevalenza generale
Durante la raccolta dei campioni non è stato rilevato alcun segno clinico di
malattia.
Analizzando il dato per specie animale, 141 capre su 461 campionate sono
risultate positive (29.8%; 95% IC 29.2% ; 30.4%) così come 121 pecore su 457
(30.0%; 95% IC 29.4% ; 30.6%) e 2 dromedari su 58 (2.6%; 95% IC 1.7% ;
3.8%)(Tab. 6).
Capra Pecora Dromedario
Npos/Ntot
Sieroprevalenza (95% IC)
Npos/Ntot
Sieroprevalenza (95% IC)
Npos/Ntot
Sieroprevalenza (95% IC)
Positivi 141/ 461
29.8% (29.2% ;30.4%)
121/457
30.0% (29.4%a30.6%) 2/58 2.6%
(1.7% ; 3.8%)
Negativ 320/461
70.2% (69.6% ; 70.8%)
336/457
70.0% (69.4% ; 70.6%) 56/58 97.4%
(96.2% ; 98.3%) Tabella 6: Sieroprevalenza generale filtrata per specie animale.
72

4.1.2 SIEROPREVALENZA PER SINGOLE REGIONI
Le regioni, come stato detto, sono state suddivise in base ad un criterio geografico
e valutando i differenti ecosistemi presenti. Di conseguenza sono state considerate
come unica regione le Wilaia di El Aaiun, Awserd, 27 Febrero e Smara presenti in
territorio algerino, mentre la Wilaia di Dakhla e le sei regioni militari dei Territori
Liberati (Bir Lehlou, Tifariti, Mehaires, Mijek, Agwanit e Dougaj) come singole
regioni.
La sieroprevalenza più elevata è stata riscontrata tra le wilaia con 108 positivi su
354 (30.1%; 95% IC 29.7% ; 30.6%), mentre il dato per le regioni militari risulta
essere inferiore, osservando generalmente un simile livello di prevalenza in Bir
Lehlou (33.5%; 95% IC 30.3% a 36.9%), in Tifariti (45.2%; 95% IC 42.0% a
48.5%) e in Agwanit (31.0%; 95% IC 28.4% ; 33.7%).
73
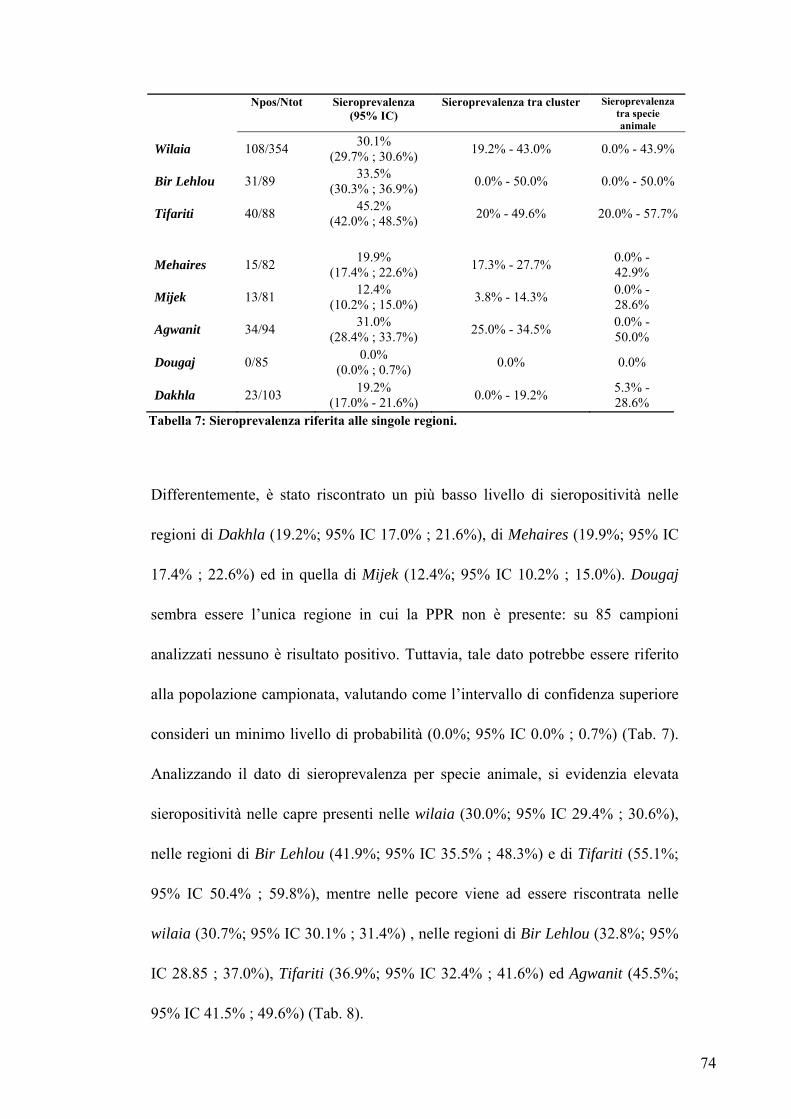
Npos/Ntot Sieroprevalenza (95% IC)
Sieroprevalenza tra cluster Sieroprevalenza tra specie animale
Wilaia 108/354 30.1% (29.7% ; 30.6%) 19.2% - 43.0% 0.0% - 43.9%
Bir Lehlou 31/89 33.5% (30.3% ; 36.9%) 0.0% - 50.0% 0.0% - 50.0%
Tifariti 40/88 45.2% (42.0% ; 48.5%) 20% - 49.6% 20.0% - 57.7%
Mehaires 15/82 19.9%
(17.4% ; 22.6%) 17.3% - 27.7% 0.0% - 42.9%
Mijek 13/81 12.4% (10.2% ; 15.0%) 3.8% - 14.3% 0.0% -
28.6%
Agwanit 34/94 31.0% (28.4% ; 33.7%) 25.0% - 34.5% 0.0% -
50.0%
Dougaj 0/85 0.0% (0.0% ; 0.7%) 0.0% 0.0%
Dakhla 23/103 19.2% (17.0% - 21.6%) 0.0% - 19.2% 5.3% -
28.6% Tabella 7: Sieroprevalenza riferita alle singole regioni.
Differentemente, è stato riscontrato un più basso livello di sieropositività nelle
regioni di Dakhla (19.2%; 95% IC 17.0% ; 21.6%), di Mehaires (19.9%; 95% IC
17.4% ; 22.6%) ed in quella di Mijek (12.4%; 95% IC 10.2% ; 15.0%). Dougaj
sembra essere l’unica regione in cui la PPR non è presente: su 85 campioni
analizzati nessuno è risultato positivo. Tuttavia, tale dato potrebbe essere riferito
alla popolazione campionata, valutando come l’intervallo di confidenza superiore
consideri un minimo livello di probabilità (0.0%; 95% IC 0.0% ; 0.7%) (Tab. 7).
Analizzando il dato di sieroprevalenza per specie animale, si evidenzia elevata
sieropositività nelle capre presenti nelle wilaia (30.0%; 95% IC 29.4% ; 30.6%),
nelle regioni di Bir Lehlou (41.9%; 95% IC 35.5% ; 48.3%) e di Tifariti (55.1%;
95% IC 50.4% ; 59.8%), mentre nelle pecore viene ad essere riscontrata nelle
wilaia (30.7%; 95% IC 30.1% ; 31.4%) , nelle regioni di Bir Lehlou (32.8%; 95%
IC 28.85 ; 37.0%), Tifariti (36.9%; 95% IC 32.4% ; 41.6%) ed Agwanit (45.5%;
95% IC 41.5% ; 49.6%) (Tab. 8).
74

Capra Pecora Dromedario N+/Ntot Sieroprev.
(95% IC) N+/Ntot Sieroprev.
(95%IC) N+/Ntot Sieroprev.
(95% IC) Wilaia 57/166 30%
(29.4%;30.6%) 51/166 30.7%
(30.1%;31.4%) 0/22 0.0%
(0.0%;1.3%)
Bir Lehlou
18/43 41.9% (35.5%;48.3%)
13/40 32.8% (28.8%;37.0%)
0/6 0.0% (0.0%;7.1%)
Tifariti 23/42 55.1% (50.4%;59.8%)
16/41 36.9% (32.4%;41.6%)
1/5 20.0% (7.7%;38.6%)
Mehaires 8/41 20.1% (16.4%;24.4%)
7/41 19.6% (16.3%;23.4%)
???? ?????
Mijek 12/42 28.6% (23.8%;33.9%)
1/37 1.4% (0.5%;3.3%)
0/2 0.0% (0.0%;5.1%)
Agwanit 11/42 26.2% (21.9%;31.0%)
23/48 45.5% (41.5%;49.6%)
0/4 0.0% (0.0%;1.7%)
Dougaj 0/43 0.0% (0.0%;1.1%)
0/42 0.0% (0.0%;1.3%)
???? ?????
Dakhla 12/42 28.6% (24.2%;33.4%)
10/42 23.8% (19.7%;28.4%)
1/19 5.3% (3.3%;8.1%)
Tabella 8: Sieroprevalenza riferita alle singole regioni e filtrata per specie animale.
4.1.3 SIEROPREVALENZA PER CLUSTER
Come riportato in tabella 9, si evidenzia una sostanziale sieroperevalenza che
viene ad essere osservata nella maggioranza dei cluster, con elevati livelli di
sieropositività riferiti alle singole wilaia e le regioni di Bir Lehlou, Tifariti, ed
Agwanit. Per quanto riguarda le wilaia, si nota un picco in 27 Febrero-1 (34.5%;
95% IC 31.1% ; 38.05), Awserd-3 (40.0%; 95% IC 42.0% ; 44.0%) ed El Aaiun-4
(32.7%; 95% IC 32.0% ; 33.5%), mentre nelle Regioni Militari elevati valori
vengono riferiti ai cluster di Bir Lehlou-5 (33.3%; 95% IC 29.2% ; 37.7%), Bir
Lehlou-8 (50.0%; 95% IC 42.5% ; 57.5%), Tifariti-9 (39.3%; 95% IC 33.8% ;
45.1%), Tifariti-10 (49.6%; 95% IC 45.5% ; 53.7%), e di Agwanit-20 (34.5%;
95% IC 31.2% ; 38.0%). Il dato filtrato per specie animale mette in evidenza una
maggior incidenza di PPR tra le capre, considerando elevati livelli di
sieropositività in 27 Febrero-1 (43.9%; 95% IC 38.9 ; 49.1%), Bir Lehlou-5
(41.9%; 95% IC 35.5% ; 48.3%), Bir Lehlou-8 (50.0%; 95% IC 42.5% ; 57.5%),
75

Tifariti-9 (50.0%; 95% IC 41.7% ; 58.3%), Tifariti-10 (57.7%; 95% IC 51.9% ;
63.3%) e Mehaires-14 (42.9%; 95% IC 30.4% ; 56.5%). Tra le pecore, dati
significativi vengono riportati nei cluster di Awserd-3 (42.9%; 95% IC 41.4% ;
44.3%), Tifariti-10 (41.2%; 95% IC 35.5% ; 47.1%) ed Agwanit-20 (42.9%; 95%
IC 37.8% ; 48.0%). Nessun dato di positività viene riportato per i cluster di
Dougaj anche se, come considerato precedentemente, l’intervallo di confidenza
superiore riporti un livello di probabilità di ~1.3%.
76

Cluster Npos/Ntot Sieroprevalenza
(95% IC) Specie Sieroprevalenza (95% IC)
27 Febrero 1 29/87 34.5% (31.1% ; 38.0%)
Dromedario Pecora Capra
0.0% (0.0% ; 26.5%) 25.6% (21.2% ; 30.5%) 43.9% (38.9% ; 49.1%)
Smara 2 16/92 19.2% (18.5% ; 19.8%) Dromedario Pecora Capra
0.0% (0.0% ; 3.9%) 19.5% (18.6% ; 20.4%) 19.0% (18.2% ; 20.0%)
Awserd 3 36/86 40.0% (42.0% ; 44.0%) Dromedario Pecora Capra
0.0% (0.0% ; 4.1%) 42.9% (41.4% ; 44.3%) 43.9% (42.5% ; 45.3%)
El Aaiun 4 27/89 32.7% (32.0% ; 33.5%) Dromedario Pecora Capra
0.0% (0.0% ; 4.5%) 35.2% (33.9% ; 36.1%) 31.0% (29.9% ; 32.0%)
Bir Lehlou 5 22/59 33.3% (29.2% ; 37.7%) Pecora
Capra 25.0% (19.9% ; 30.7%) 41.9% (35.5% ; 48.3%)
Bir Lehlou 6 0/6 0.0% (0.0% ; 7.1%) Dromedario 0.0% (0.0% ; 7.1%)
Bir Lehlou 7 2/10 20.0% (12.3% ; 29.8) Pecora 20.0% (12.3% ; 29.8%)
Bir Lehlou 8 7/14 50.0% (42.5% ; 57.5%) Pecora 50.0% (42.5% ; 57.5%)
Tifariti 9 10/23 39.3% (33.8% ; 45.1%) Pecora Capra
28.6% (21.6% ; 36.6%) 50.0% (41.7% ; 58.3%)
Tifariti 10 29/60 49.6% (45.5% ; 53.7) Pecora Capra
41.2% (35.5% ; 47.1%) 57.7% (51.9% ; 63.3%)
Mehaires 12 1/16 17.3% (12.6% ; 22.8%) Pecora Capra
33.3% (25% ; 42.5%) 0.0% (0.0% ; 3.3%)
Mehaires 13 4/21 17.9% (13.9% ; 22.6%) Pecora Capra
14.3% (9.3% ; 20.8%) 21.4% (15.2% ; 28.5%)
Mehaires 14 4/15 27.7% (19.8% ; 36.5%) Pecora Capra
12.5% (4.9% ; 24.4%) 42.9% (30.4% ; 56.5%)
Mehaires 15 4/19 22.9% (16.4% ; 30.2%) Pecora Capra
16.7% (8.7% ; 26.8%) 28.6% (19.1% ; 39.9%)
Mehaires 16 2/11 18.2% (11.1% ; 27.0%) Pecora 18.2% (11.1% ; 27.0%)
Mijek 17 1/15 3.8% (1.2% ; 8.2%) Dromedario Pecora
0.0% (0.0% ; 5.1%) 7.7% (2.4% ; 16.0%)
Mijek 18 12/66 14.3% (11.7% ; 17.3%) Pecora Capra
0.0% (0.0% ; 1.5%) 28.6% (23.8% ; 33.9%)
Agwanit 19 17/38 25.0% (21.1% ; 29.4) Dromedario Pecora
0.0% (0.0% ; 1.7%) 50.0% (43.2% ; 56.8%)
Dougaj 21 0/42 0.0% (0.0% ; 1.3%) Pecora 0.0% (0.0% ; 1.3%) Dougaj 22 0/43 0.0% (0.0% ; 1.1%) Capra 0.0% (0.0% ; 1.1%)
Dakhla 23 23/103 19.2% (17.0% ; 21.6%) Dromedario Pecora Capra
5.3% (3.3% ; 8.1%) 23.8% (19.7% ; 28.4%) 28.6% (24.2% ; 33.4%)
Tabella 9: Sieroprevalenza riferita ai singoli cluster e filtrata per specie animale.
77

4.2 ANALISI DI CORRELAZIONE
Nella pratica d’indagine epidemiologica è frequente la necessità di verificare se
esiste una specifica concordanza e/o relazione tra i dati osservati, sia di natura
qualitativa che quantitativa. Tale calcolo risulta particolarmente utile nella fase
iniziale dell’analisi statistica, quando vengono ricercate le variabili più indicative
e le relazioni di associazione esistenti (Thrusfield, 2007). Nel caso specifico sono
stati utilizzati due differenti test, ossia il test χ2 (Pearson’s χ2 test) ed il test Esatto
di Fisher, computati attraverso l’utilizzo di tabelle di contingenza 2xn. Tali test
vengono utilizzati frequentemente ed hanno valore nella verifica dell’associazione
tra le varie modalità di due o più caratteri qualitativi al dato quantitativo osservato
(Thrusfield, 2007). Il livello di significatività dei test è dato dal valore di probabilità
p (o p-value), che indica il minimo livello di significatività del test per il quale si
rifiuterebbe l’ipotesi nulla, in questo caso la mancanza di associazione tra caratteri
qualitativi e quantitativi (Kirkwood e Sterne, 2006). Il valore di cut-off riferito al p-
value è fissato, convenzionalmente, pari a 0.05 (p=0.05), determinando come
valori inferiori indichino proporzionalmente il livello di significatività nell’analisi
(Kirkwood e Sterne, 2006). L’analisi di associazione è stata eseguita tra
sieroprevalenza riscontrata e caratteristiche di specie, età e sesso della
popolazione animale campionata.
4.2.1 CORRELAZIONE PER SPECIE ANIMALE
Dai risultati dell’analisi risulta evidente come la sieropositività riscontrata è
massima per capre e pecore (χ2=19.3697; p<0.0001), minima per dromedari, e
78

con elevata frequenza nelle capre (53.4%) (Tab. 10). Tale dato indica come le
capre in particolar modo, ed in percentuale rilevante anche le pecore, sembrano
rivestire un ruolo predominante nell’epidemiologia della PPR all’interno
dell’ecosistema presente nella RASD. Inoltre, tale dato risulta essere in linea con
le caratteristiche epidemiologiche della PPR che indicano nelle capre una
maggiore suscettibilità all’infezione rispetto alle pecore (Aklaku, 1980; Bourdin,
1973).
Specie
Capra Pecora Dromedario TOT Positivi 141 121 2 264 Negativi 320 336 56 712 TOT 461 457 58 976
Tabella 10: Correlazione tra specie animale e prevalenza riscontrata (χ2 19.3697, p<0.0001)
4.2.2 CORRELAZIONE PER ETA’ DEI SOGGETTI
Come riportato in tabella 11, il livello di sieroprevalenza risulta aumentare
proporzionalmente con l’età dei soggetti (χ2 17.6462, p<0.0001), considerando
come il valore più elevato sia riscontrato nei soggetti di età superiore ai 36 mesi
(58.7%). Tale dato risulta interessante nella valutazione della situazione
epidemiologica della RASD riferita alla PPR, per cui tale territorio potrebbe
essere riferito come endemico in considerazione del fatto che: la prevalenza
riscontrata risulta essere pari a ~30%; nel siero è stata riscontrata la presenza di
IgG, quindi un’infezione non recente; il 37.1% degli animali risultati positivi
possiede età superiore ai 48 mesi ed, infine, che nel corso dell’indagine di campo
non sono stati evidenziati segni clinici riferibili alla PPR.
79

ETÀ in mesi
<12 12-36 >36 TOT Positivi 41 68 155 264 Negativi 126 272 314 712 TOT 167 340 469 976
Tabella 11: Correlazione tra età dei sogetti e sieroprevalenza riscontrata (χ2 17.6462, p<0.0001)
4.2.3 CORRELAZIONE PER SESSO DEI SOGGETTI
Il dato di associazione tra sieroprevalenza e sesso degli animali campionati risulta
non significativo (p=0.2), anche se il riscontro di positività nelle femmine risulta
avere una probabilità del 130% superiore rispetto ai soggetti di sesso maschile
(OR = 1.309) (Tab 12).
Sesso
Femmine Maschi TOT
Positivi 228 36 264 Negativi 590 122 712 TOT 818 158 976
Tabella 12: Correlazione tra sesso dei sogetti e sieroprevalenza riscontrata (Test esatto di Fisher, p=0.2)
4.2.4 FUNZIONE DI CORRELAZIONE
In ultima analisi, la relazione tra le caratteristiche epidemiologiche della
popolazione animale (specie, sesso ed età dei soggetti) ed il risultato di
sieroprevalenza è stata analizzata mediante regressione logistica binaria. Tale
metodologia statistica si propone di studiare e quantificare questa relazione per
cui la variabile dipendente è di natura dicotomica, ossia riconducibile a due valori
(in questo caso valore di sieroprevalenza positivo o negativo) (Thrusfield, 2007). Nel
caso specifico, la regressione logistica non predice la variabile così com’è, ma la
80

trasforma in funzione logit, di conseguenza si predice il logaritmo del rapporto tra
la probabilità di appartenere ad un gruppo (positivo) pittosto che ad un altro
(negativo), definiti dalla variabile dipendente (Kirkwood and Sterne, 2006). La misura
dell’associazione viene ad essere basata sul calcolo dell’Odds Ratio (OR), o anche
definito come “rapporto incrociato” (Thrusfield, 2007). Tale dato definisce il
rapporto esistente tra le frequenze osservate tra le categorie o gruppi definiti nella
funzione di regressione. Nel caso specifico, un valore di OR = 1 indica assenza di
associazione tra sieropositività e caratteristiche di popolazione, dove valori <1
indicano un’associazione negativa, mentre valori >1 un’associazione positiva. Il
dato viene ad essere validato dal test di significatività statistica (p-value) per
escludere che l’associazione sia dovuta al caso (Kirkwood and Sterne, 2006).
Dai risultati dell’analisi di regressione logistica binaria (Tab. 13) risulta evidente
come la sieropositività riportata nella RASD sia in relazione significativa con la
specie (p<0.0001) e l’età dei soggetti (p=0.003) della popolazione campionata,
mentre il sesso dell’individuo non ne influenza la relazione (p=0.713).
Ulteriormente, si evidenzia come il rapporto di probabilità di infezione da PPR
aumenta di 1.57 volte in animali di specie caprina e di 1.43 volte in soggetti di età
superiore ai 36 mesi.
Sesso
Coef SE Coef Z P OR 95% IC Constante -310.610 0.533301 -5.82 0.000 Specie 0.450666 0.124983 0,1673611 0.000 1.57 1.23 ;
2.00 0.078069 0.212399 0.37 0,4951389 1.08 0.71 ;
1.64 Età 0.358125 0.119170 3.01 0.003 1.43 1.13 ;
1.81 Tabella 13 : Risultati della funzione di regressione logistica binaria (p<0.0001).
81

4.3 ANALISI DI MODELLAZIONE SPAZIALE
Il verificarsi di manifestazioni epizootiche e/o la distribuzione endemica di
malattia, generalmente, vengono ad essere rappresentati in dimensione spaziale
come loci puntiformi d’infezione, ossia localizzazione di eventi di malattia. Tale
proprietà viene ad essere spesso associata con particolari caratteristiche
geografiche, per cui l’epidemiologia di diverse malattie infettive risulta essere
influenzata da tali fattori, mostrando come la loro diffusione si basi su determinati
modelli ambientali (Relman et al., 2008). In questo caso l’ambiente viene ad essere
considerato non solo come ecologico, ma soprattutto come insieme di specifiche
caratteristiche di popolazione animale (es. conformazione dei gruppi, densità,
movimentazioni). Di conseguenza, metodi di statistica spaziale risultano fornire
un valore aggiuntivo nell’analisi delle dinamiche di prevalenza di malattia e come
parte integrante dell’indagine epidemiologica. In ambito epidemiologico l’analisi
spaziale viene designata allo scopo di: individuare loci d’infezione su base
spaziale e temporo-spaziale nella pratica di sorveglianza; testare se l’infezione è
distribuita su base casuale o viene ad essere definita in base a caratteristiche
spaziali, temporali o temporo-spaziali; valutare il significato statistico dei loci
d’infezione; modellare, su base geografica, la distribuzione di malattia in paesi
endemici (Carpenter, 2001). Metodi di modellazione spaziale vengono ad essere
maggiormente indirizzati nell’individuazione e valutazione di loci d’infezione, sia
su base puramente spaziale che temporale o considerandone la caratteristica di bi-
dimensionalità (temporo-spaziale) (Kulldroff, 2009). Tale analisi viene ad essere
effettuata attraverso la scansione di “finestre geografiche” (definite su basi
dimensionali di spazio-tempo) valutando il rapporto tra casi osservati e casi attesi
82

in ciascuno dei loci d’infezione: la “finestra geografica” con elevata probabilità
statistica viene considerata come uno dei loci (cluster) d’infezione primaria. Il
modello di elaborazione spaziale utilizzato nell’analisi dei dati per la PPR nella
RASD è il discrete Poisson permutation model (Kulldroff, 1997) computato
attraverso il programma SaTscan (Kulldroff, 2009), considerandone 9999 iterazioni:
tale metodologia considera che i casi d’infezione all’interno di una determinata
area geografica siano distribuiti secondo la distribuzione statistica di Poisson; il
totale di casi attesi in ciascun cluster viene ad essere considerato come
proporzionale alla sua popolazione totale (Kulldroff, 1997). Dai risultati (Tab. 14,
Fig. 32), viene evidenziato un cluster significativo (p=0.0283) comprendente i siti
di campionamento di Bir Lehlou-8, Tifariti-9, Tifariti-10, Tifariti-11, ed avente
come centro la regione di Tifariti. Inoltre, tale cluster evidenzia un rischio relativo
(RR) pari a 1.753, considerando come in tale area geografica vi sia un 175% di
rischio maggiore nella diffusione e sviluppo di focolai di PPR rispetto ad altre
regioni della RASD.
Location ID Coordinate
(radius) N° casi/N° attesi
(ratio) RR P
Bir Lehlou-8 Tifariti-9 Tifariti-10 Tifariti-11
26.266389 N 10.319722 E (27.01 Km)
47/29.3 (1.619)
1.753 0.0283
Tabella 14 - Risultati dell’analisi di modellazione spaziale (p=0.0283).
83

5. CONCLUSIONI
La PPR rappresenta un’importante malattia dei piccoli ruminanti che sta mettendo
a serio rischio questa popolazione animale in Africa, nel Medio Oriente e nell’
Asia meridionale.
L’importanza economica di tale malattia è stata messa in evidenza attraverso una
relazione internazionale eseguita da Perry et al. (2002) in cui viene dato risalto al
fatto che la PPR rappresenta una malattia prioritaria da tenere sotto controllo per
ridurre il problema della povertà soprattutto in Africa Occidentale e nell’Asia
meridionale. Nella stessa relazione è stato inoltre sottolineato come la PPR sia
ancora una malattia poco conosciuta, soprattutto per quanto riguarda le
caratteristiche epidemiologiche e le dinamiche di trasmissione. Nonostante questo,
risulta comunque evidente che la presenza della malattia è legata principalmente
alla movimentazione ed al commercio degli animali recettivi ed infetti, provenienti
da paesi confinanti, come la Mauritania, il Mali ed il Marocco, che non vengono
adeguatamente controllati al momento dell’ingresso nel paese. Nel nostro studio
infatti la maggiore positività sierologica riscontrata nelle wilaia è dovuta proprio al
fatto che in queste zone c’è una maggiore concentrazione di animali provenienti da
paesi limitrofi per la presenza di fiere e mercati. Il riscontro di positività anche in
specie animali normalmente non sensibili all’infezione, come i dromedari, fa
supporre che altre specie animali, diverse dai piccoli ruminanti, potrebbero avere
un ruolo dal punto di vista epidemiologico nella diffusione della malattia, pur non
manifestando mai una sintomatologia clinica. In quest’ottica risulta importante,
84

quindi, il monitoraggio di altre specie animali soprattutto di quelle che vivono a
stretto contatto con pecore e capre che risultano altamente suscettibili.
Questo lavoro è stato il primo studio epidemiologico sulla PPR svolto nell’area
geografica decritta da cui è emerso, dall’analisi dei risultati ottenuti, l’effettiva
presenza della malattia nel territorio con una sieroprevalenza di ~30%. La
segnalazione di continui nuovi focolai, come quelli registrati di recente in Marocco
e Turchia, sta ad indicare la costante circolazione virale nel continente africano e
quindi l’importanza di una stretta sorveglianza, al fine di evitare l’eventuale
introduzione della malattia in paesi indenni come ad esempio il nostro.
85

BIBLIOGRAFIA
- Abegunde A. 1983. Problems connected with TCRV vaccination of sheep and
goats. In: Hill D H (ed), Peste des petite ruminants (PPR) in sheep and goats.
Proceedings of the international workshop held at IITA, Ibadan, Nigeria, 24-26
September 1980. lLCA (International Livestock Centre for Africa), Addis Ababa,
Ethiopia. pp. 79-81.
- Abraham G., Sintayehu A., Libeau G., Albina E., Roger F., Laekemariam Y.,
Abayneh D., Awoke K.M., 2005. Antibody seroprevalences against peste des
petits ruminants (PPR) virus in camels, cattle, goats and sheep in Ethiopia.
Preventive Veterinary Medicine 70 (2005) 51–57.
- Abu Elzein, E.M.E., Hassanien, M.M., Al-Afaleq, A.I., Abd Elhadi, M.A.,
Housawi, F.M.T., 1990. Isolation of peste des petits ruminants from goats in
Saudi Arabia. Vet. Record 127, 309-310.
- Adu F D and Nawathe D R. 1981. Safety of tissue culture rinderpest vaccine in
pregnant goats. Research note. Tropical Animal Health and Production 13:166.
- Africa ’70, 2007. Soutien à l’élevage de bétail dans les Camps de Réfugiés
Sahraoui. First report. Monza: Africa ’70, ONG-PVD/2006/131-812.
86

- Ahmad M.Al-Majali, Nazmi O. Hussain, Nadim M. Amarin, Aggrey A. Majok,
2008 Seroprevalence of, and risk factors for, peste des petits ruminants in sheep
and goats in Northern Jordan, Prev. Vet. Med. (2008), doi:
10.1016/j.prevetmed.2008.01.002
- Aklaku I.K. (1980). Principal causes of mortality in small ruminants in Ghana
Bulletin de l’Office International des Epizooties, 92: 1227-1231
- Arkell T., 1991. The decline of pastoral nomadism in the Western Sahara.
Geography, 76(331): 162-166.
- AVIS. Peste des petits ruminants. Working partnership between the Institute of
Animal Helth, UK; The Food and Agriculture Organisation (FAO), Rome;
L’Office International des Epizooties, Paris and Telos ALEFF Ltd., UK.
- Barrett, T., Visser, I.K.G., Mamaev, L., Goatley, L., Bressem, M.-F., Van
Osterhaus, A.D.M., 1993. Dolphin and porpoise morbilliviruses are genetically
distinct from phocine distemper virus. Virology 193, 1010–1012.
- Benazet B. La peste des petits ruminants: ´etude exp´erimentale de la vaccination.
Th`ese Doctorat V´et´erinaire, Universit´e de Toulouse, 1973.
87

- Bennett, S., Woods, T., Liyanage, W.M. and Smith, D.L., 1991. A simplified
general method for cluster-sample surveys of health in developing countries. Worl
Health Statistic Quarterly, 44 (3), p. 98-106.
- Berhe G, Minet C, Le Goff C, Barrett T, Ngangnou A, Grillet C, et al.
Development of a dual recombinant vaccine to protect small ruminants against
peste-des-petits-ruminants virus and capripoxvirus infections. J Virol
2003;77:1571–7.
- Bourdin P. (1973) Las peste des petits ruminants et sa prophylaxie au Sénégal et
en Afrique de l’Ouest. Revue d’Élevage et de Médicine Vétérinaire des Pays
Tropicaux, 26: 71-74.
- Bourdin P, Rioche M, Laurent A. Emploi d’un vaccin anti-bovipestique produit
sur cultures cellulaires dans la prophylaxie de la peste des petits ruminants au
Dahomey. Rev Elev M´ed V´et Pays Trop 1970;23:295–300.
- Broglia A., Volpato G., 2008. Pastoralism and displacement: strategies and
limitations in livestock raising by Sahrawi refugees after thirty years of exile.
Journal of Agriculture and Environment for International Development 2008, 102
(1/2): 105-122.
88

- Burton A.H., Dean J.A., Dean A.G. (1990). Software for data management and
analysis in epidemiology. World Health Forum, 11: 75-77.
- Cameron, A., 1999. Survey Toolbox for livestock disease - A practical manual
and software package for active surveillance in developing countries.
Canberra:Australian Centre for International Agricultural Research Monograph
No.54 Available from: http://www.aciar.gov.au/publication/MN054 [Accessed 7
August 2008].
- Caratini S., 2000. Système de parenté Sahraoui. L’impact de la révolution.
L’Homme, 154-155: 431-456.
- Carpenter T.E., (2001). Methods to investigate spatial and temporal clustering in
veterinari epidemiology. Preventive Veterinary Medicine, 48: 303-320.
- Cauacy-Hymann E., Bodjo S.C., Danho T., Koffi M.Y., Libeau G., Diallo A.
Early detection of viral excretion from experimentally infected goats with peste-
des-petits-ruminants virus. Prev. Vet. Med. 78 (2007) 85-88.
- Couacy-Hymann E., Bodjo C., Danho T., Libeau G. e Diallo A., 2005.
Surveillance of wildlife as a tool for monitoring rinderpest and peste des petits
ruminants in West Africa. Rev. sci. tech. Off. int. Epiz., 2005, 24 (3), 869-877.
89

- CESME. Peste dei Piccoli Ruminanti. Centro di Referenza Nazionale per le
Malattie Esotiche, Istituto Zooprofilattico Sperimentale dell’Abruzzo e del Molise
“G. Caporale”, Teramo.
- Cosby SL, Kai C, Yamanouchi K. Immunology of rinderpest- an
immunosuppression but a lifelong vaccine protection. In: Barrett T, Pastoret P-P,
TaylorWP, editors. Rinderpest and peste des petits ruminants. Virus plagues of
large and small ruminants. Amsterdam: Academic Press, Elsevier; 2005. p. 197–
221.
- Diallo A., Minet C., Le Goff C., Berhe G., Albina E., Libeau G., Barrett T. The
threat of peste des petits ruminants: progress in vaccine development for disease
control. Vaccine 25 (2007) 5591–5597.
- Diallo A, Minet C, Berhe G, Le Goff C, Black DN, Fleming M, et al. Goat
immune response to capripox vaccine expressing the haemaglutinin protein of
peste des petits ruminants. Ann NY Acad Sci 2002;969:88–91.
- Diallo, A., 2003. Control of PPR: classical and newgeneration of vaccine. Dev.
Biol. Basel, Karger 114, 85-91.
- Di Nardo A., 2008. Rift Valley fever (RVF) in Western Sahara: a
seroepidemiological survey among Wilayas and “Liberated Territories” of the
90

Saharawi Arab Democratic Republic (SADR). MSc Thesis, College of Medicine
and Veterinary Medicine, University of Edinburgh.
- Di Nardo A. Rossi D., Mohammed Lamin Saleh S., Mohammed Lejlifa S., Hatri
Hamdi S., Di Gennaro A., Savini G., Thrusfield M.V. (2009). Seroprevalence of
Rift Valley fever in wilayas and ‘liberated territories’ regions of the Sahrawi Arab
Democratic Republic, Western Sahara. Preventive Veterinary Medicine. In
Stampa.
- El Hag Ali, B., Taylor, W.P., 1984. Isolation of peste des petits ruminants virus
from Sudan. Res. Vet. Sci. 36, 1–4.
- Farid, M.N. and Frerichs, R.R., 2007. CSurvey software and manual version 2.0.
Los Angeles, CA: Department of Epidemiology, University of California.
Avaliable from: http://www.ph.ucla.edu/epi/programs/csurvey2_manual.pdf
[Accessed 3 August 2008].
- Farina R., Scatozza F. Trattato di malattie infettive degli animali. UTET, 1998.
- Gargadennec, L., Lalanne, A., 1942. La peste des petits ruminants. Bulletin des
Services Zoo Techniques et des Epizooties de I’Afrique Occidentale Francaise 5,
16–21.
91

- Gibbs E P J. Taylor W P and Lawman M J P. 1977. The isolation of adenoviruses
from goats affected with peste des petite ruminants (kata and stomatitis
pneumoenteritis complex) in Nigeria. Research in Veterinary Science 23(3): 331-
335.
- Gibbs, E.P.J., Taylor, W.P., Bryant, J., 1979. Classification of peste des petits
ruminants virus as the fourth member of genus Morbillivirus. Intervirology 11,
268–274.
- Gilbert Y, Monnier J. Adaptation du virus de la peste des petits ruminants aux
cultures cellulaires. Rev Elev Med Vet Pays Trop 1962;15:321–35.
- Gilbert, Y., Monnier, J., 1962. Adaptation des virus de la peste des petits
ruminants aux cultures cellularaires.Revue d’Elevage et de Medecine Veterinaire
des Pays Tropicaux 15, 321–335.
- Henderson R.H. and Sundaresan, T., 1982. Cluster sampling to assess
immunization coverage: a review of experience with a simplified sampling
method. Bulletin of the World Health Organization, 60 (2), p. 253-260.
- Jensen, E., 2005. Western Sahara: anatomy of a stalemate. Boulder, CO: Lynne
Rienner Publishers.
92

- Jingyue Bao, Lin Li, Zhiliang Wang, Tom Barrett , Longciren Suo, Wenji Zhao,
Yutian Liu , Chunju Liu , Jinming Li, 2008. Development of one-step real-time
RT-PCR assay for detection and quantitation of peste des petits ruminants virus.
Journal of Virological Methods 148 (2008) 232–236.
- Kirkwood B.R. and Sterne J.A.C. (2006). Medical statistics. Blackwell
Publishing Ltd., Oxford, 502 pp.
- Kulldorff M (1997). A spatial scan statistic. Communication in Statistics: Theory
and Methods, 26: 1481-1496.
- Kulldorff M. (2009). SaTScanTM v8.0: software for the spatial and space-time
scan statistics. Kulldorr M. and Information Management Services Inc.
http://www.satscan.org/.
- Lefefre P.C., Diallo A., Schenkel F., Hussein S., Staak G., 1991. Serological
evidence of peste des petits ruminants in Jordan. Vet. Record 128, 110.
- Libeau, G., Prehaud, C., Lancelot, R., Colas, F., Guerre, L., Bishop, D.H.L., 1995.
Development of a competitive ELISA for detecting antibodies to peste des petits
ruminants virus using a recombinant nucleoprotein. Research in Veterinary
Science 58, 50–55.
93

- Loewenberg S., 2005. Displacement is permanent for the Sahrawi refugees. The
Lancet, 365: 1295-1296.
- Lopriore, C. and Branca, F., 2001. Strategies to fight anaemia and growth
retardation in Sahrawi refugee children. Rome: Italian Nutrition Institute.
- Nanda, Y.P., Chatterjee, A., Purohit, A.K., Diallo, A., Inui, K., Sharma, R.N.,
Libeau, G., Thevasagayam, J., Bruning, A., Kitching, P., Anderson, J., Barrett, T.,
Taylor, W.P., 1996. The isolation of peste des petits ruminants virus from
northern India. Vet. Microbiol. 51, 207–216.
- NVD-SADR (2007). Censo 2007. Dirección Nacional de Veterinaria, Ministerio
de Salud Pública Saharui, República Árabe Democrática Saharaui, Rabouni.
- Office International des Epizooties (OIE) (1993) Outbreaks occurring during the
month of January; list A diseases. Bullettin 105-1, 7-10.
- Olmi G., 1998. Sahara Occidentale, appunti di viaggio. Rome: Edizione
Associate.
- Otte M.J. and Gumm I.D., 1997. Intra-cluster coefficients of 20 infections
calculated from the results of cluster-sample surveys. Preventive Veterinary
Medicine, 31 (1-2), p. 147-150.
94

- OXFAM, 1995. Western Sahara. OXFAM Belgium and Comité belge de soutien
au peuple Sahraoui, Bruxelles.
- Ozenda P., 1991. Flore et vegetation du Sahara. 3rd ed., CNRS Éditions, Paris,
662 pp.
- Perry BD, Randolph TF, McDermott JJ, Sones KR, Thornton PK. Investing in
animal health research to alleviate poverty. Nairobi, Kenya: ILRI (International
Livestock Research Institute); 2002. pp. 148.
- Plowright W., Ferris RD. Studies with rinderpest virus in tissue culture. The use
of attenuated culture virus as a vaccine for cattle. Res Vet Sci, 1962;3:172–82.
- Poli G., Cocilovo A. Microbiologia e immunologia veterinaria. UTET, 2006.
- Radostits O.M., Blood D.C., Gay C.C., 2000. Veterinary Medicine, ninth ed. WB
Saunders Co., Philadelphia.
- Rajak KK, Sreenivasa BP, Hosamani M, et al. Experimental studies on
immunosuppressive effects of peste des petits ruminants (PPR) virusin goats.
Comp Immunol Microbiol Infect Dis 2005;28:287–96.
95

- Relman D.A., Hamburg M.A., Choffnes E.R., Mack A., (2008). Global climate
change and extreme weather events: understanding the contribution to infectious
diseases emergence. Workshop summary. Forum on Microbial Threats, Board on
Global Health, Intitute of Medicine of the National Academies. The National
Academies Press, Washington D.C, 304 pp.
- Roeder P.L., Abraham G., Kenfe G., Barrett T., 1994. Peste des petits ruminants
in Ethiopian goats. Trop. Anim. Hlth. Prod. 26, 69–73.
- Roger F., Guebre Yesus M., Libeau G., Diallo A., Yigezu L.M. e Yilma T., 2001.
Detection of antibodies of rinderpest and peste des petits ruminants viruses
(Paramyxoviridae, Morbillivirus) during a new epizootic disease in Ethiopian
camels (Camelus dromedarius). Revue Méd. Vét., 2001, 152, 3, 265-268.
- Rossiter PB, Taylor WP. Peste des petits ruminants. In: Coetzer AW, Thomson
GR, Tustin RC, editors. Infectious diseases of livestock, vol. II. 1994. p. 758-65.
- Rowland AC, Bourdin P. The histological relationship between peste des petits
ruminants and kata in West Africa. Rev Elev M´ed V´et Pays Trop 1970;23:301–
307.
- Rowland AC, Scott GR, Hill DH. The pathology of an erosive stomatitis and
enteritis in West African dwarf goats. J Pathol 1969;98:83–7.
96

- San Martín P., 2004. Briefing: Western Sahara: road to perdition? African Affairs,
103/413: 651-660.
- Schneider-Schaulies S, Niewiesk S, Schneider-Schaulies J, Volker ter Meulen V.
Measles virus induced immunosuppression: targets and effector mechanisms.
Curr Molec Med 2001;1:163–81.
- Seddon D., 2000. Western Sahara – point of no return? The Review of African
Political Economy, 27(84): 338-340.
- Servet-Delprat C, Vidalain PO, Valentin H, Rabourdin-Combe C. Measles virus
and dendritic cell functions: how specific response cohabits with
immunosuppression. Curr Top Microbiol Immunol 2003;276:103–23.
- Shaila M.S., Purushothaman V., Bhavasar D., Venugopal K., Venkatesan R.A.,
1989. Peste des petits ruminants in India. Vet. Rec. 125, 602.
- Shelley, T., 2004. Endgame in Western Sahara: What future for Africa’s last
colony? London: Zed Books Ltd.
- Taylor WP. Protection of goats against peste des petits ruminants with attenuated
rinderpest virus. Res Vet Sci 1979;27:321–4.
97

- Taylor W.P., 1984a. The distribution and epidemiology of peste des petits
ruminants. Prev. Vet. Med. 2,157-166.
- Taylor W.P., Al Busaidy S., Barett T., 1990. The epidemiology of peste des petits
ruminants in the Sultanate of Oman. Vet Microbiology 22, 341-352.
- Thrusfield, M., 2007. Veterinary epidemiology. 3rd ed. Oxford: Blackwell
Publishing.
- UNHCR, 2005. 2004 Global Refugee Trends. Population and Geographical Data
Section, Division of Operational Support, UNHCR, Geneva. Online at:
http://www.unhcr.ch/statistics.
- UNHCR. 2002. 2002 Global Appeal. Geneva: UNHCR.
- Volpato G., Ahmadi Emhamed A., Lamin Saleh S.M., Broglia A., Di Lello S.,
2007. Procurement of traditional remedies and transmission of medicinal
knowledge among Sahrawi people displaced in south-western Algeria refugee
camps. In Pieroni A., Vandebroek I. (Eds), Traveling Cultures and Plants: the
Ethnobiology and Ethnopharmacy of Migrations, Berghahn Books, Oxford, pp.
355-384.
98

- Wambura P.N., 2000. Serological evidence of the absence of peste des petits
ruminants in Tanzania. Vet. Record 146, 473-474.
- Wamwayi H.M., Rossiter P.B., Kariuki D.P., Wafula J.S., Barrett T., Anderson J.,
1995. Peste des petits ruminants antibodies in east Africa. Vet. Rec. 136, 199–
200.
- Worrwall EE, Litamoi JK, Seck BM, Ayelet G. Xerovac. An ultra rapid method
for the dehydration and preservation of live attenuated rinderpest and peste des
petits ruminants vaccines. Vaccine 2001;19:834–9.
- Wosu L.O., Okiri J.E. and Enwezor P.A. Optimal time for vaccination against
peste des petits ruminants (PPR) disease in goats in the humid tropical zone in
southern Nigeria - moment optimum de vaccination des caprins contre la peste
des petits ruminants (PPR) dans la zone tropicale humide du sud du Nigéria. FAO
Corporate Document Repository.
99

100
RINGRAZIAMENTI
Questo lavoro, ma soprattutto questa esperienza umana e lavorativa che mi porto
dietro, non sarebbe stata possibile senza la presenza del grande capo Davide
Rossi, mio mentore e ormai caro amico, che ringrazio infinitamente.
L’appoggio e la pazienza del professore Fabrizio Passamonti sono stati preziosi e
senza dubbio indispensabili, come il grande aiuto del Dott. Antonello Di Nardo
che mi ha aiutata con tutti quei numerini da analizzare.
Un ringraziamento speciale va a Saleh e Sidumu e a tutti i saharawi che hanno
reso il mio soggiorno nei Campi Profughi un’esperienza indimenticabile.
Per quanto riguarda i miei lunghi anni universitari non posso che ringraziare
l’estrema pazienza e fiducia che hanno avuto i miei genitori e mio fratello, che mi
hanno sempre appoggiata in tutte le mie scelte, dato la possibilità di fare
esperienze preziose e che sono e saranno sempre le persone più speciali della mia
vita, a cui devo tutto ciò che sono.
Ringrazio tutte Las Amigas che con la loro costante presenza riempiono
quotidianamente la mia vita, in particolar modo Ciop per esserci sempre, la Vanex
e Chiara compagne di avventure, Laura un’amica preziosa, Giulia cugina ma
anche sorella e Lauretta la saggia.
Ed in fine ringrazio me stessa per non aver mollato mai!





























