Embed Size (px)
Citation preview

Untersuchung von Ras/Effektor-
Komplexen mit gezielt veränderten
elektrostatischen Eigenschaften
Dissertation
zur Erlangung des akademischen Grades
des Doktors der Naturwissenschaften
der Fakultät für Chemie
an der Ruhr-Universität Bochum
vorgelegt von
Christina Kiel aus Bünde
Bochum 2003

Die vorliegende Arbeit wurde in der Zeit von Januar 2000 bis März 2003 in der Abteilung
Strukturelle Biologie des Max-Planck-Instituts für molekulare Physiologie in Dortmund unter
der Anleitung von Prof. Dr. Alfred Wittinghofer und Prof. Dr. Christian Herrmann
angefertigt.
Erster Gutachter: Prof. Dr. Alfred Wittinghofer
Zweiter Gutachter: Prof. Dr. Rolf Heumann
Dritter Prüfer: Prof. Dr. Martin Feigel

Dieses Baumes Blatt, der von Osten meinem Garten anvertraut,
gibt geheimen Sinn zu kosten, wie`s den Wissenden erbaut.
Ist es ein lebendig Wesen, das sich in sich selbst getrennt? Sind es zwei, die sich erlesen, daß man sie als eines kennt?
Solche Fragen zu erwidern, fand ich wohl den rechten Sinn.
Fühlst du nicht an meinen Liedern, daß ich eins und doppelt bin?
(Johann Wolfgang von Goethe „Suleika“)

INHALT
I EINLEITUNG 1 1.1 GTP-bindende Proteine: Zentrale Rolle in Signalübertragungs-
prozessen der Zelle 1
1.2 Ras als molekularer Schalter 31.3 Ras Effektoren: Proteine mit unterschiedlichen biologischen
Funktionen 6
1.4 Die Ras Bindungsdomäne als gemeinsames Strukturelement aller Effektoren
9
1.5 Die Signalweiterleitung über Ras als Angriffspunkt zur Entwicklung von möglichen Medikamenten in der Krebstherapie
11
1.6 Analyse von Bindungsenergien in der Kontaktfläche zwischen Proteinen und Liganden
13
1.7 Zielsetzung der Arbeit 16 II MATERIALIEN UND METHODEN 18 2.1 Materialien 182.1.1 Chemikalien 182.1.2 DNA-Konstrukte 182.1.3 Enzyme, Proteine und Antikörper 182.1.4 Geräte 182.1.5 Lösungen 192.1.6 Materialien für die Chromatographie 192.1.7 Mikroorganismen und Zelllinien 192.1.8 Nährmedien 192.1.9 Puffer-Lösungen 19 2.2 Methoden 202.2.1 Molekularbiologische Methoden 20
2.2.1.1 PCR-Mutagenese 202.2.1.2 Restriktion von Plasmid-DNA 212.2.1.3 Agarose-Gelelektrophorese und Gelelution 212.2.1.4 Ligation von DNA-Fragmenten 212.2.1.5 Herstellung kompetenter Bakterien 212.2.1.6 Transformation von Bakterien 212.2.1.7 Plasmidpräparation 222.2.1.8 DNA-Sequenzierung 22
2.2.2 Proteinchemische Methoden 222.2.2.1 Glyzerindauerkulturen, Zellanzucht und Proteinsynthese 222.2.2.2 Zellaufschluss durch Ultraschall 232.2.2.3 SDS-Polyacrylamid-Gelelektrophorese 232.2.2.4 Bestimmung der Proteinkonzentration nach Bradford 232.2.2.5 Reinigung von RalGDS- und Raf-RBD Wildtyp und Mutanten 242.2.2.6 Reinigung von Ras und Rap Wildtyp und Mutanten 24

2.2.2.7 Nukleotidaustausch an Ras und Rap Proteinen 252.2.2.8 Kristallographie von Proteinen 25
2.2.3 Biophysikalische Methoden 252.2.3.1 Stopped-Flow Messungen 252.2.3.2 Isotherme Titrationskalorimetrie 262.2.3.3 Differential Scanning Kalorimetrie 262.2.3.4 Phosphor-NMR 26
2.2.4 Zellbiologische Methoden 272.2.4.1 Ral-Beladungs Assay 272.2.4.2 Luziferase-Reportergen Assay 28
III ERGEBNISSE 29 3.1 Kinetische und thermodynamische Eigenschaften der
Wechselwirkung von Ras Proteinen mit Effektor-RBDs 29
3.1.1 Kinetische Eigenschaften von Ras/Effektor-RBD-Komplexen 293.1.2 Thermodynamik der Bindung von Ras Proteinen an Effektoren-RBDs 363.1.3 Vergleich der Affinitäten ermittelt durch Stopped-Flow und ITC 41 3.2 Rolle der Elektrostatik für die kinetischen Eigenschaften der
Wechselwirkung von RalGDS- und Raf-RBD mit Ras Proteinen 42
3.2.1 Kinetik der Bindung von Mutanten-Komplexen aus RalGDS-RBD und Ras/Rap⋅mGppNHp
42
3.2.2 Kinetik der Bindung von Raf-RBD-Mutanten an Ras⋅mGppNHp 45 3.3 Gezielte Manipulation von RalGDS- und Raf-RBD zur
Veränderung der elektrostatischen Eigenschaften 48
3.3.1 Manipulation von RalGDS-RBD und kinetische Analyse der Mutanten im Komplex mit Ras⋅mGppNHp
49
3.3.2 Design und kinetische Analyse von Raf-RBD-Mutanten im Komplex mit Ras⋅mGppNHp
54
3.3.3 Kinetische Analyse von Raf-RBDWT und Mutanten im Komplex mit Ras⋅mGDP 59 3.4 Vergleich von experimentellen und berechneten kon-Werten 633.4.1 Bindung von RalGDS-RBD-Mutanten an Ras⋅mGppNHp 633.4.2 Bindung von Raf-RBD-Mutanten an Ras⋅mGppNHp 643.4.3 Bindung von Raf-RBD-Mutanten an Ras⋅mGDP 65 3.5 Thermodynamik der Bindung von Ras/RBD-Komplexen mit
veränderten elektrostatischen Eigenschaften 66
3.5.1 Assoziation von RalGDS-RBD-Mutanten an Ras⋅GppNHp 663.5.2 Thermodynamik der Bindung von Raf-RBD-Mutanten an Ras⋅GppNHp 693.5.3 Thermodynamik der Assoziation von Raf-RBDWT und Mutanten an Ras⋅GDP 74 3.6 Aktivierungsenergien der Assoziation von Mutanten Komplexen
mit veränderter Elektrostatik 78
3.6.1 Bindung von RalGDS-RBDWT und Mutanten an Ras⋅mGppNHp 783.6.2 Bindung von Raf-RBDWT und Mutanten an Ras⋅mGppNHp 803.6.3 Bindung von Raf-RBDWT und Mutanten an Ras⋅mGDP 82

3.7 Bindung von Raf-RBDWT und Mutanten an Ras⋅(m)GppNHpT35A
und Ras⋅mGppNHpT35S 83
3.7.1 Kinetik der Bindung von Raf-RBDWT und Mutanten an Ras⋅mGppNHpT35A und Ras⋅mGppNHpT35S
83
3.7.2 Thermodynamik der Bindung von Raf-RBDWT und Mutanten an Ras⋅GppNHpT35A 85 3.8 Strukturelle Analyse der Bindung von Raf-RBDA85K an Ras⋅GDP
und Ras⋅GppNHpT35A 86
3.8.1 31P-NMR-Messungen 863.8.1.1 Bindung von Raf-RBDA85K an Ras⋅GppNHpT35A 873.8.1.2 Bindung von Raf-RBDWT und Raf-RBDA85K an Ras⋅GDP 90
3.8.2 Kristallisationsversuche von Raf-RBDA85K im Komplex mit Ras⋅GDP und Raps⋅GDP 933.8.3 Doppel-Mutanten Analyse zur Ermittlung der energetischen Beiträge einzelner
Aminosäure-Kontakte 95
3.8.3.1 Kopplungsenergie zwischen Lys85 von Raf-RBD und Glu31 von Ras⋅GDP 953.8.3.2 Kopplungsenergie zwischen Lys85 von Raf-RBD und Glu31 von
Ras⋅GppNHp 96
3.8.3.3 Kopplungsenergie zwischen Lys85 von Raf-RBD und dem γ-Phosphat von Ras⋅GppNHp
97
3.9 Zelluläre Signalweiterleitung von Ras/Effektor-Mutanten-
Komplexen mit veränderter Elektrostatik 99
3.9.1 Der Ral-Beladungs-Assay zur Quantifizierung der Signalweiterleitung über RalGDS 993.9.2 Der Luziferase-Reportergen-Assay zur Quantifizierung der Signalweiterleitung über
Raf 99
3.9.2.1 Transfektion von RasG12V und Raf-Mutanten 1013.9.2.2 Transfektion von RasWT und Raf-Mutanten 1043.9.2.3 Transfektion von RasG12V,T35S bzw. von RasG12V,T35A und Raf-Mutanten 105
3.10 Vergleich der Aktivierung in vivo mit den Ergebnissen aus kinetischen und thermodynamischen Messungen
106
3.10.1 Affinitäten (Kd) und Luziferase-Aktivitäten 1073.10.2 Enthalpieänderungen (∆H°) und Luziferase-Aktivitäten 1093.10.3 Entropieänderungen (T∆S°) und Luziferase-Aktivitäten 1093.10.4 Assoziationsratenkonstanten (kon) und Luziferase-Aktivitäten 1103.10.5 Dissoziationsratenkonstanten (koff) und Luziferase-Aktivitäten 111 IV DISKUSSION 113 4.1 Die Oberflächeneigenschaften von Ras und Effektor-RBDs und
thermodynamische und kinetische Parameter bei der Bindung 113
4.1.1 Die thermodynamischen Parameter der Bindung von Effektor-RBDs an Ras Proteine geben Hinweis auf die Art der Bindung
113
4.1.2 Komplementäre elektrostatische Ladungen in der Kontaktfläche von Ras/Effektor-RBD-Komplexen beschleunigen die Assoziation
115
4.1.3 Es werden unterschiedliche thermodynamische und kinetische Hot-Spots in Ras/Effektor-RBD-Komplexen gefunden
116
4.1.4 Ergebnisse aus Doppel-Mutanten-Analysen können die Bindungsspezifitäten der Bindung von RalGDS- und Raf-RBD an Ras und Rap quantifizieren
120

4.2 Die gezielte Veränderung der Elektrostatik der Oberflächen von RalGDS- und Raf-RBD führt zu veränderten kinetischen Eigenschaften
122
4.2.1 Die kinetischen Parameter werden in Komplexen von RalGDS- und Raf-RBD unterschiedlich beeinflusst
123
4.2.2 Die Korrelation von experimentellen und berechneten Werten für kon ist für Komplexe mit RalGDS-RBD-Mutanten am besten
124
4.2.3 Günstige elektrostatische Wechselwirkungen können die Aktivierungsbarriere durch eine höhere Affinität des Encounter-Komplexes erniedrigen
126
4.2.4 Enthalpie- und Entropie-Beiträge in Mutanten-Komplexen mit veränderten elektrostatischen Eigenschaften
128
4.3 Bindung von Raf-RBDWT und Mutanten an Ras⋅GDP und
Ras⋅GppNHpT35A 132
4.3.1 Zusammenhang von kinetischen Parametern, Sättigungskinetik und Aufspaltung der Phosphor-Resonanzen
133
4.3.2 Affinitäten und Wärmekapazitäten 1364.3.3 Mögliche Rolle der Mutation Lys85 in Raf-RBD 137 4.4 Biologische Bedeutung der Bindungskinetik 1384.6.1 Ist die Signalweiterleitung in vivo nur abhängig von der Affinität oder auch von der
Assoziationsratenkonstante? 139
4.4.2 Für die Signalweiterleitung über Ras⋅GDP sind nicht die Assoziations-ratenkonstanten, sondern die Lebenszeiten der Komplexe entscheidend
140
4.4.3 Vergleich der Effizienz der Signalweiterleitung über Ras⋅GTP und Ras⋅GDP 1404.4.4 Besteht eine Korrelation mit thermodynamischen Parametern und den Luziferase-
Aktivitäten? 141
4.4.5 Signalweiterleitung über RasT35A und RasT35S 141 V ZUSAMMENFASSUNG 142 VI ANHANG 144 6.1 Abkürzungen 1446.2 Oligonukleotide 1456.3 Klonierungen 1466.4 Zusätzliche Ergebnis-Tabellen 1486.5 Lebenslauf 1516.6 Vorträge und Konferenzen 152 VII LITERATUR 154

EINLEITUNG
1
I EINLEITUNG
In einem vielzelligen Organismus wird jede Zelle während ihrer Entwicklung darauf programmiert, auf spezifische Signale zu antworten. Einzelne oder koordinierte Signale beeinflussen das Verhalten der Zelle und regulieren dadurch wichtige Prozesse wie die Zellteilung, die Zelldifferenzierung und den Stoffwechsel. Am häufigsten finden parakrine Signalübertragungen statt, bei denen Signalmoleküle schnell und lokal aufgenommen werden und nur auf unmittelbar benachbarte Zellen wirken können. Zusätzlich erfolgen zentrale Kontrollen über endokrine und synaptische Signalübertragungen: die von endokrinen Zellen freigesetzten Hormone werden über das Blut zu allen Zielzellen im Körper transportiert; die von Nervenzellen hergestellten Neurotransmitter wirken lokal auf die postsynaptischen Zellen, mit denen ihre Axone in Verbindung stehen. Zelluläre Signalübertragungen erfolgen über extrazelluläre Signalmoleküle, die an Rezeptoren einer Zelle binden. Kleine, hydrophobe Moleküle, wie Steroidhormone oder lösliche Gase können durch die Plasmamembran diffundieren und aktivieren intrazelluläre Rezeptoren. Die meisten extrazellulären Signalmoleküle sind jedoch hydrophil und können nur an Oberflächen-Rezeptoren binden. In beiden Fällen erfolgt nach Aktivierung der entsprechenden Rezeptoren die intrazelluläre Signalweiterleitung über niedermolekulare Botenstoffe (Second Messenger) und Proteine. Hervorzuheben unter den Proteinen sind Proteinkinasen und -phosphatasen, die über ihren Phosphorylierungszustand reguliert werden, bzw. häufig auch andere Proteine regulieren. Des Weiteren stellen Adaptorproteine wichtige Verbindungsglieder in einer Signalkette dar, indem sie über Proteinmodule, die bestimmte Bindungs-Sequenzen enthalten, mit anderen Proteinen wechselwirken. Zu der größten Proteinfamilie gehören – zusammen mit den Kinasen - die regulatorischen GTP-bindenden Proteine (GTPasen): diese treten in vielseitigen Formen auf, werden aber weitgehend nach denselben Prinzipien aktiviert und reguliert. Die meisten Signale werden auf diese Weise von der Zelloberfläche zum Zellkern weitergeleitet, wodurch die Expression von Genen verändert wird.
1.1 GTP-bindende Proteine: Zentrale Rolle in Signal-
übertragungsprozessen der Zelle GTP-bindende Proteine, auch als GNBPs bezeichnet (guanine nucleotide binding proteins) wirken als molekulare Schalter: je nach gebundenem Nukleotid können sie in einem „AN“- (GTP-gebundenen) und „AUS“- (GDP-gebundenen) Zustand vorliegen (Abbildung 1.1). GNBPs werden aktiviert, wenn Protein gebundenes GDP freigesetzt wird und stattdessen GTP bindet, welches im Überschuss in der Zelle vorliegt. Dieser Austausch wird durch Guaninnukleotid Austauschfaktoren, GEFs (guanine nucleotide exchange factor) aktiviert. Im GTP Zustand binden sie an Effektoren als nachgeschaltete Proteine, welche als Folge mit

EINLEITUNG
2
anderen Proteinen kommunizieren. Die Wechselwirkung mit Effektoren wird durch Hydrolyse des gebundenen GTPs zu GDP beendet. Diese intrinsische GTPase-Reaktion ist langsam, kann aber durch GAPs (GTPase aktivierende Proteine) stark beschleunigt werden. Eine Säugerzelle enthält ungefähr 100 bis 150 verschiedene GTP-bindende Proteine. Sie regulieren viele biologische Prozesse wie z.B. Zellwachstum und -differenzierung, die Proteinbiosynthese und den Transport durch Membranen. Entsprechend ihrer Funktion lassen sie sich in mehrere Superfamilien einteilen (Bourne et al., 1990; Bourne et al., 1991): Die α-Untereinheiten der heterotrimeren G-Proteine (Gs, Gi, Gq, G12), die Translationsfaktoren der Proteinbiosynthese (IF-2, EF-Tu, EF-G, RF-3), die Familie der Dynamin verwandten großen GTPasen (Dynamin, Mx, GBP), das Signalerkennungspartikel und sein Rezeptor (SRP54, FtsY) sowie die Ras verwandten GTP-bindenden Proteine (siehe Kapitel 1.2).
Die gemeinsame Eigenschaft der GNBPs ist die Mg2+-abhängige Bindung von Guaninnukleotiden und die Fähigkeit GTP zu GDP zu hydrolysieren. Alle GNBPs besitzen als Strukturelement eine ca. 20-kD große G- (Guaninnukleotid) Domäne welche essentiell für die Nukleotidbindung und -hydrolyse ist. Die Struktur der G-Domäne ist für alle GNBPs konserviert und besteht aus einem gemischten sechssträngigen ß-Faltblatt sowie fünf Helices auf beiden Seiten. Alle bisher aufgelösten dreidimensionalen Strukturen von GTP-bindenden Proteinen zeigen eine hoch konservierte G-Domäne und unterscheiden sich durch die Struktur und relative Orientierung ihrer Extra-Domänen (Vetter & Wittinghofer, 2001).
ABBILDUNG 1.1: Funktion und Regulation von GTP-bindenden Proteinen. Die GNBPs liegen entweder im inaktiven GDP gebundenen Zustand oder gebunden an GTP vor. Die Dissoziation der Nukleotide sowie die intrinsische GTP Hydrolyse sind sehr langsam. Durch GEFs wird der Austausch von GDP zu GTP katalysiert. In der aktiven, GTP-gebundenen Form ist die Wechselwirkung mit Effektoren möglich, was zu einer zellulären Antwort führt. Durch GAPs, die die Hydrolyse des GTPs zu GDP katalysieren, wird das GNBP wieder in den inaktiven Zustand überführt.

EINLEITUNG
3
Innerhalb der G-Domäne gibt es vier bis fünf konservierte Sequenz-Elemente, die sich in der Nähe der Nukleotid-Bindungsstelle befinden: Das GxxxxGKS/T Motiv (x bedeutet beliebige Aminosäure) wird als P- (Phosphatbindungs-) Schleife (Saraste et al., 1990) oder Walker A-Motiv bezeichnet und tritt auch in anderen nukleotidbindenden Proteinen wie, Myosin und anderen ATP-abhängigen Enzymen auf (Walker et al., 1982). Die P-Schleife macht Kontakte zum β- und γ- Phosphat des Nukleotids. Zusammen mit dem N/TKxD Motiv, welches Kontakte zur Base des Nukleotids macht, sind dies die wichtigsten Beiträge zur Nukleotidbindung. Die Spezifität für Guaninenukleotide erfolgt durch die Seitenkette des Asp vom N/TKxD Motiv, welches Wasserstoff-Brücken zur Base ausbildet. Der Vergleich von Strukturen von GNBPs in der GDP- und in der GDP- gebundenen Form zeigt, dass die Konformationsänderungen insgesamt nur gering sind; sie sind hauptsächlich zwei Bereichen zuzuordnen, die als Schalter I und Schalter II bezeichnet werden. Diese Änderungen können jedoch teilweise sehr große Auswirkungen auf die Wechselwirkung mit anderen Proteinen und somit auf die Signalweiterleitung haben.
1.2 Ras als molekularer Schalter Die Ras verwandte Superfamilie der GTP-bindenden Proteine besitzt Molmassen im Bereich von 20 bis 25 g. Die Mitglieder lassen sich durch Vergleich ihrer Aminosäuresequenzen in die Ras-, Rho-, Rab-, Arf- und Ran-Unterfamilien einteilen (Valencia et al., 1991; Buguski & Mc Cormick, 1993). Die Funktionen der Proteine der einzelnen Unterfamilien reichen von der Proliferation und Differenzierung (Ras-Unterfamillie, Vojtek & Der, 1998), Organisation des Zytoskeletts (Rho-Unterfamilie, MacKay, 1998), Steuerung von Intermembrantransport (Rab-Unterfamilie, Schimmöller et al., 1997), Vesikeltransport zwischen ER und Golgi-Apparat (Arf-Unterfamilie, Moss & Vougham, 1998) bis zur Regulation des Kernimports und -exports (Ran, Moore, 1998). Weitere Mitglieder der Ras-Superfamilie sind die Rad-, Rheb-, Rit- und Rag-Unterfamilie, deren Funktionen aber noch weitgehend unbekannt sind. Die Proteine der Rit-Unterfamilie sind möglicherweise an calciumabhängigen Signalwegen beteiligt (Lee et al., 1996) und über Rad ist bekannt, dass es in Muskelzellen von Diabetes Typ II überexprimiert wird (Garvey et al., 1997, Bilan et al., 1998).
Anfang der 80er Jahre wurde erstmals die Bedeutung des Ras Proteins erkannt, als das ras-Gen bei Retroviren gefunden wurde, die in Ratten Tumore vom Sarkomatyp (Ras wird abgeleitet von Ratten Sarkoma) auslösen können (Barbacid, 1987). Es gibt drei verschiedene humane Ras Gene: H-, N- und K-ras (mit den zwei Splice-Varianten K-ras4A und K-ras4B). Die Proteine H-, N- und K-Ras, die von diesen Genen kodiert werden, besitzen Molmassen von 21 kDa. H-Ras ist strukturell und biochemisch am besten charakterisiert und wird nachfolgend als Ras bezeichnet.

EINLEITUNG
4
Die Primärsequenzen von Ras Proteinen werden aufgrund ihrer Homologie in mehrere Bereiche unterteilt (Abbildung 1.2-B): N-terminal sind die Aminosäuren 4-31 (Nummerierung für H-Ras) variabel. Es folgt die ca. 160 Aminosäuren lange G-Domäne, in der die fünf konservierten Nukleotidbindungsmotive liegen. Der C-terminale Bereich ist variabel und umfaßt 13-49 Aminosäuren. Am C-Terminus befindet sich die CaaX-Box (C bedeutet Cystein, a aliphatische Aminosäure und X beliebige Aminosäure). Das CaaX-Motiv dient als Erkennungssequenz für Enzyme, die posttranslationale Modifizierungen durch Lipidanker katalysieren, was zur Membranverankerung dient und notwendig für die biologische Aktivität von Ras ist (Hancock et al., 1990). Die Strukturen von Ras konnten sowohl im GDP- als auch im GppNHp- (GppNHp ist ein GTP analoges, nicht hydrolysierbares Nukleotid) gebundenen Zustand aufgelöst werden (Pai, et al., 1989; Pai et al., 1990; Milburn et al., 1990; Prive et al., 1992). Die Röntgenstruktur von C-terminal verkürztem Ras gebunden an GppNHp ist in Abbildung 1.2-A dargestellt (Pai et al., 1989).
ABBILDUNG 1.2: Tertiärstruktur von Ras und Bereichsdiagramm der Bindungsmotive. (A) Kristallstruktur von C-terminal verkürztem Ras im Komplex mit GppNHp mit den konservierten Sequenzelementen der Nukleotid-Bindungsstelle (orange) und den Schalter Regionen I und II („switch I“ und „switch II“, blau), in denen die größten strukturellen Änderungen beim Wechsel zwischen GTP- und GDP- gebundenen Zustand erfolgen. (B) Schematische Darstellung von Ras mit den konservierten Sequenzelementen (orange) und Schalter Regionen (blau).

EINLEITUNG
5
Die Struktur des gesamten Proteins konnte bis jetzt nicht gelöst werden, aber die biochemischen Eigenschaften von verkürztem und vollständigem Ras unterscheiden sich nicht (John et al., 1990). Das Protein setzt sich aus einem sechs-strängigen gemischten β-Faltblatt umgeben von fünf α-Helices zusammen (G-Domänen Faltung). 10 Schleifen verbinden diese α-Helices und β-Stränge. Die Schleife 1 (P-Schleife) umfasst die Aminosäuren 10-15 und bindet zusammen mit L2 und L4 die γ-Phosphatgruppe. L2 (AS 26-36, Schalter I) ist an der Bindung von Effektormolekülen und L4 (AS 59-64, Schalter II) an der Wechselwirkung mit GAPs beteiligt. Die konservierten Sequenzmotive PM1 bis PM3 (beteiligt an Phosphat und Mg2+ Bindung) und G1 bis G3 (beteiligt an Bindung zur Guaninbase) befinden sich hauptsächlich in diesen Schleifen. Der Vergleich der aktiven GppNHp-Form mit der GDP-Form zeigt – wie bei allen GNBPs - vor allem Positionsänderungen von Schalter I und II (blau dargestellt in Abbildung 1.2).
Wie alle GTP-bindenden Proteine ist Ras aktiv im GTP-gebundenen und inaktiv im GDP-gebundenen Zustand (Wittinghofer et al., 1993; Schweins & Wittinghofer, 1994; Wiesmüller & Wittinghofer, 1994). Die Nukleotidbindung ist Mg2+-abhängig und sowohl GDP als auch GTP werden mit hoher Affinität gebunden (Die Dissoziationskonstanten liegen im Bereich von 10-11 und 10-12 M, John et al., 1990). Der Austausch von GDP zu GTP erfolgt daher nur katalytisch durch GEFs, wie z.B. Sos und Cdc25, welche die nukleotidfreie Form stabilisieren. In der aktiven Form kann die Wechselwirkung mit Effektormolekülen erfolgen wodurch bestimmte Signalwege eingeschaltet werden (siehe Kapitel 1.3 und 1.4). Die anschließende Überführung in die GDP-Form erfolgt durch Hydrolyse von GTP zu GDP, wobei die intrinsische Hydrolyse sehr langsam verläuft (kcat = 0.028 min-1); sie kann jedoch durch GAPs, wie z.B. NF1 und p120GAP ca. 105-fach beschleunigt werden (Gideon et al., 1992).
ABBILDUNG 1.3: Signalwege zur Aktivierung von Ras. Die Aktivierung von Ras erfolgt an der Membran über Rezeptor-Tyrosin-Kinasen (RTK) oder G-Protein gekoppelte Rezeptoren (GPCR). Vom aktiven Ras⋅GTP verzweigt sich die Signalweiterleitung durch Wechselwirkung mit einer Vielzahl von Effektoren. Abbildung verändert aus Wittinghofer & Waldmann, 2000.

EINLEITUNG
6
Die Aktivierung von Ras kann durch Wachstumsfaktoren und über die Aktivierung von G-Protein gekoppelten Rezeptoren erfolgen (Abbildung 1.3). So führt z.B. die Bindung des Wachstumsfaktors EGF (epidermal growth factor) an den EGF-Rezeptor zu Phosphorylierung von Tyrosinresten im intrazellulären Teil des Rezeptors, welche als Bindungsstellen für Adaptorproteine mit SH2 (Src-homology-2) Domänen, wie z.B. Grb2 (growth factor receptor bound protein 2) dienen. Grb2 bindet durch seine SH3 (Src-homology-3) Domäne fest an die prolinreiche Region des Austauschfaktors Sos, was zu dessen Membranlokalisation führt und dort als GEF zur Aktivierung von Ras dient. Ebenso führt die Aktivierung über G-Protein gekoppelte Rezeptoren zur Stimulation von Austauschfaktoren für Ras. Vom aktiven Ras⋅GTP aus wird das Signal mit Hilfe einer Vielzahl verschiedener Effektoren weitergeleitet und daher erfolgt hier eine Verzweigung in verschiedene Signalwege.
1.3 Ras Effektoren: Proteine mit unterschiedlichen
biologischen Funktionen In den vergangenen Jahren wurde eine große Anzahl möglicher Effektormoleküle gefunden und analysiert (Übersichtsartikel: Wittinghofer & Herrmann, 1995; Wittinghofer & Nassar, 1996; Herrmann & Nassar, 1996; Marshall, 1996; Herrmann, 2003). Der erste charakterisierte Ras Effektor aus Säugerzellen ist die Serine/Threonin Kinase Raf (rapid fibrosarcosis); ursprünglich wurde sie als Onkogen in Retroviren identifiziert (Rapp et al., 1983). Von den Raf-Kinasen existieren drei Isoformen (raf-1, A-Raf und B-Raf). Die Raf-Kinase wirkt im Signalweg von vielen Rezeptor-Tyrosinkinasen und ist somit wichtig für Zellwachstum und -differenzierung (Übersicht: Rapp et al., 1988; Daum et al., 1994; Avruch et al., 1994). Der Mechanismus der Aktivierung der Raf-Kinase und insbesondere die Rolle von Ras dabei sind noch nicht endgültig verstanden. Es werden sowohl Modelle diskutiert, nach denen die alleinige Funktion von Ras die Rekrutierung von Raf zur Membran ist und dort weitere Faktoren für die Aktivierung verantwortlich sind (Leevers et al., 1994; Stokoe et al., 1994), als auch Modelle nach denen Ras zusätzlich direkte Einflüsse auf die Kinaseaktivität hat (Stokoe & McCormick, 1997; Tamada et al., 1997). Die Signalweiterleitung verläuft ausgehend vom aktivierten Raf Protein mit Hilfe von Proteinkinasen, die in Form einer Kaskade von Kinasen hintereinander geschaltet sind. Diese Kinasen werden aufgrund ihrer Regulierbarkeit durch extrazelluläre, häufig mitogen wirkende Liganden als MAPK (mitogen aktivierte Proteinkinasen) oder als ERK (extrazellulär regulierte Kinasen) bezeichnet. Aktiviertes Raf phosphoryliert und aktiviert die Proteinkinasen MEK1 und MEK2 (MEK = MAP/ERK-Kinase), diese wiederum aktivieren ERK1 und ERK2. Die Substrate von MEK1 und MEK2 sind MAPK1 und MAPK2, die nach Phosphorylierung in den Zellkern gelangen. Dort erfolgt die Aktivierung der Genexpression über die Phosphorylierung des Transkriptionsfaktors Elk-1. Dieser bindet zusammen mit dem SRF (serum response faktor)

EINLEITUNG
7
an das SRE (serum response element), das man als regulatorische Sequenz von verschiedenen Genen (z.B. c-fos) findet. Zusätzlich phosphoryliert ERK1/2 c-Jun, was zur Aktivierung des AP1 Transkriptionsfaktors, bestehend aus den c-Fos und c-Jun Heterodimer, führt. Als Folge dieses Transkriptionsaktivators werden regulatorische Proteine des Zellzyklus, wie z.B. Cyclin D exprimiert, welches das Fortschreiten im Zellzyklus fördert.
Als weiterer Effektor wurde die PI3- (Phosphatidylinositol-3) Kinase gefunden, die in viele Signaltransduktionswege involviert ist, welche ihren Effekt über die Produktion von Second Messengern ausüben (Stephens et al., 1993; Fry, 1994). Es wurde eine direkte Wechselwirkung der katalytischen p110 Domäne der PI3-Kinase mit Ras nachgewiesen (Kodaki et al., 1994; Rodriguez-Viciana et al., 1994), was zur Aktivierung der PI3-Kinase als Folge ihrer Translokation zur Membran und struktureller Änderungen führt. Die PI3-Kinase phosphoryliert Phosphatidylinositol-4,5-bisphosphat (PtdIns (4,5) P2) zu Phosphatidylinositol-3,4,5-trisphosphat (PtdIns (3,4,5) P3), welches ein Second Messenger ist und an viele Proteine über PH (Pleckstrin Homologie) Domänen binden kann. Dadurch kontrolliert die PI3-Kinase viele Protein Aktivitäten, wobei die Aktivierungen der Kinasen PDK1 (3-phosphoinoside-dependent protein kinase-1) und Akt (auch als PKB bezeichnet) detailliert untersucht worden sind. PDK1 ist wichtig für die Aktivierung einer großen Anzahl von weiteren Kinasen, einschließlich Akt/PKB, einige PKCs (Protein Kinase C), p70S6K (p70 ribosomale Protein S6 Kinase) und RSK (Glykogen Synthase Kinase). Die Wirkung von Akt/PKB ist anti-apoptotisch durch die Phosphorylierung verschiedener Moleküle, wie z.B. Bad (Datta et al., 1999). Des Weiteren führt die Aktivierung der PI3-Kinase zur Stimulierung von Rac, wodurch die Regulation des Zytoskeletts beeinflusst wird.
Ein dritter, gut untersuchter Effektor ist RalGDS (Ral guanosinenucleotide dissociation stimulator), auch bekannt als RalGEF (Ral guanosinenucleotide exchange factor), der ursprünglich als Austauschfaktor für Ral beschrieben wurde (Albright et al., 1993). Zwei zu RalGDS homologe Effektoren, Rgl (RalGDS like) und Rlf (Rgl2, RalGDS like 2) sind ebenfalls beschrieben worden (Wolthuis et al, 1996). Als Folge der Ral spezifischen Austauschfunktion von RalGDS, Rgl und Rlf aktiviert Ral⋅GTP die Phospholipase D1 sowie das Ral Bindungsprotein 1 (RalBP1), was zu Änderungen in der Zell-Morphologie führt.
Als weitere Effektoren wurden Byr2, AF6, Rin, NORE1 sowie PLCε identifiziert. Byr2 ist ein Effektor von Ras in S. pombe und ist der MEK-Kinase in Säugerzellen homolog (Lange-Carter et al., 1993). AF6 (ALL-fused protein 6) wurde als Fusionsprotein von ALL-1 (acute lymphoblastic leukemia 1) bei akuter myeloider Leukämie entdeckt (Prasad et al., 1993). AF6 bindet an Rap1A und es wurde eine Bindung an das Actin Zytoskelett Regulator Protein Profilin beschrieben (Boettner et al., 2000). Rin1 und 2 (Ras-interacting) kompetitieren möglicherweise mit Raf um die Bindung an Ras (Han & Colicelli, 1995; Wang et al., 2002). NORE1 (novel Ras effector 1) (Vavvas et al., 1998) und das NORE Homolog RASSF1 binden im Komplex mit MST1 an Ras⋅GTP sowie an andere Proteine der Ras Familie und es

EINLEITUNG
8
konnte gezeigt werden, dass RasG12V durch die Bindung an den NORE1/MST1 Komplex einen apoptotischen Effekt ausübt (Khokhlatchev et al., 2002). Die Phospholipase C Domäne von PLCε (Phospholipase Cε) katalysiert die Hydrolyse von PtdIns (4,5) P2 zu Diacylglycerol und Inositol-1,4,5-Trisphosphat und somit verbindet PLCε die Ras Aktivierung mit der Aktivierung von PKC und der Calcium vermittelten Signalweiterleitung (Kelley et al., 2001).
Eine Übersicht über die wichtigsten bekannten Signalwege über Ras ist in Abbildung 1.4 dargestellt. Es wird angenommen, dass die verschiedenen Signalwege in Kombination wirken (Signal Integration) und so die entsprechende Antwort in der Zielzelle hervorgerufen wird.
Ras⋅GTP
PI3KNORE1 Raf RalGDS PLCε
Ral Akt PKCPDK1
RalBP1
Rac/Cdc42
Ca2+
Vesikel TransportZellzyklus
Signalweiterleitungüber Calcium
p70S6K
Anti-ApoptoseZytoskelett
PLD
Mst1 MEK
ERK
Transkription Zellzyklus
Apoptose
ABBILDUNG 1.4: Wechselwirkung von Ras mit Effektoren. Vom aktiven Ras⋅GTP verzweigt sich die Signalweiterleitung durch Wechselwirkung mit einer Vielzahl von Effektoren. Legende: NORE1 (novel Ras effector 1), Raf (rapid fibrosarcosis), RalGDS (Ral guanosinenucleotide dissociation stimulator), PI3K (Phosphatidylinositol-3 Kinase), PLCε (Phospholipase Cε), Mst1 (Proapoptotische GC Protein Kinase der Gruppe2), MEK (mitogen aktivierte Protein Kinase, MAPK), ERK (extrazellulär regulierte Kinasen), PLD (Phospholipase D), Ral (Mitglied der Ras Unterfamilie), Akt (Protein Kinase, auch als PKB bekannt), PDK1 (Phosphatidylinositol triphosphate-dependent kinase 1), PKC (Protein Kinase C), RalBP1 (Ral Bindungsprotein 1), p70S6K (p70 ribosomal protein S6 kinase) und Rac/Cdc42 (Mitglied der Rho/Rac Unterfamilie der Ras Familie ).

EINLEITUNG
9
Wichtig ist, dass auch andere Proteine der Ras Familie, wie z.B. Rap und R-Ras teilweise mit den genannten Effektoren wechselwirken können, da sie in ihrer Effektor Bindungsregion sehr homolog sind. Dadurch wird die Komplexität der Signalweiterleitung noch größer, die Bedeutung ist aber im Einzelnen nicht verstanden. Möglicherweise wird die Spezifität der Effektoren trotzdem durch die Lokalisation von Ras Proteinen in unterschiedlichen zellulären Kompartimenten gewährleistet (Prior et al., 2001).
1.4 Die Ras Bindungsdomäne als gemeinsames
Strukturelement aller Effektoren
Die Aminosäuresequenz der verschiedenen Effektoren weist keine Homologie auf und entsprechend ihrer unterschiedlichen Funktionen in der Zelle unterscheiden sich auch die Größen und die Domänenstrukturen dieser Proteine. Als gemeinsame Eigenschaft besitzen jedoch alle Effektoren eine Ras Bindungsdomäne (RBD) (Abbildung 1.5). Die RBD ist ein Strukturbereich, über den die Bindung an den Effektorbereich von Ras im GTP-Zustand erfolgt. Auch die Sequenzhomologie unter den RBDs ist nur sehr niedrig, trotzdem besitzen alle bisher strukturell charakterisierten RBDs (von Raf, RalGDS, Rgl, Rlf, Byr2 und PI3Kγ) dasselbe Faltungsmuster (ßßαßßαß), welches als Ubiquitin Faltung bezeichnet wird (Nassar et al., 1995; Geyer et al., 1997; Huang et al., 1997; Kigawa et al., 1998; Esser et al., 1998; Walker et al., 1999; Gronwald et al, 2001).
c-Raf RBD C1 Kinase Aktivität
Byr2 SAM RBD Kinase Aktivität
RalGDS GEFN GEF RBD
NORE1 C1 RBD coil
AF6 RBD coil RA FHA PDZ
ABBILDUNG 1.5: Domänenstrukturen von Ras Effektoren. Dargestellt sind die Domänenstrukturen der Effektoren c-Raf (648 Aminosäuren = as), Byr2 (658 as), RalGDS (914 as), NORE1 (413 as) und AF6 (1612as). Alle Effektoren haben als gemeinsames Strukturelement eine Ras Bindungsdomäne (RBD). Abkürzungen: C1 (protein kinase conserved region 1), SAM (sterile alpha motif), GEF (guaninenucleotide exchange factor), coil (coil-coil domain), FHA (forkhead associated domain), RA (ras associating domain), PDZ (post-synaptic density proteins 95 domain)

EINLEITUNG
10
Die Kristallstrukturen von Rap1A⋅GppNHp und der Mutante Rap1A⋅GppNHp E30D, K31E (Mutationen in der Effektor-Bindungsregion, die Rap Ras-ähnlich machen) wurden jeweils im Komplex mit Raf-RBD gelöst (Abbildung 1.6; Nassar et al., 1995; Nassar et al., 1996). Außerdem wurden die Strukturen von Ras⋅GppNHp und Ras⋅GppNHpE31K in Komplex mit RalGDS-RBD (Huang et al., 1998; Vetter et al., 1999), Ras⋅GppNHp in Komplex mit der gesamten PI3 Kinase (Pacold et al., 2000) und die Komplexstruktur von Ras⋅GppNHp mit Byr2-RBD (Scheffzek et al., 2001) gelöst. In allen Strukturen erfolgt die Bindung hauptsächlich über ein intermolekulares ß-Faltblatt, bestehend aus ß2 und ß3 von Ras und ß1 und ß2 von Raf-RBD; beispielhaft ist die Struktur von Raf-RBD in Komplex mit Raps⋅GppNHp in Abbildung 1.6 dargestellt. Die Bindung wird in allen Kontaktflächen der Komplexe hauptsächlich über polare und ionische Aminosäure Seitenketten und nur zu einem geringen Teil über hydrophobe Wechselwirkungen vermittelt. Die Oberfläche der Kontaktfläche ist groß (1300 Å2) und relativ flach und die Kontakte erfolgen teilweise über Wassermoleküle. Trotz der Ähnlichkeiten der bisher bekannten Strukturen von Ras Proteinen in Komplex mit verschiedenen RBDs, unterscheiden sich die Affinitäten der Komplexe deutlich, wie in biochemischen Untersuchungen gezeigt werden konnte (Hermann et al., 1996; Linnemann et al., 1999; Gronwald et al., 2001; Scheffzek et al., 2001). Besonders beim Vergleich von Ras und Rap fallen deutliche Unterschiede auf: So bindet RalGDS-RBD mit 100fach höherer Affinität an Rap als an Ras, während umgekehrt Raf-RBD mit 60fach höherer Affinität an Ras als an Rap bindet (Nassar et al., 1996). Dies ist insbesondere erstaunlich, wenn man die Sequenzen der Effektor-Bindungsregion von Ras Proteinen vergleicht: Die Regionen weisen eine sehr hohe Homologie auf. Dies lässt vermuten, dass die spezifischen Wechselwirkungen von einzelnen Aminosäuren für die Bindungsspezifitäten verantwortlich sind.
ABBILDUNG 1.6: Komplexstruktur von Raf-RBD und Rap1A. Die Struktur von Raf-RBD im Komplex mit Raps (Rap1A E30D, K31E) wurde mit einer Auflösung von 2.0 Å gelöst (Nassar et al., 1996). Abbildung aus Wittinghofer & Waldmann, 2000.

EINLEITUNG
11
Basierend auf Sequenz-Homologien ist eine Familie von homologen Domänen, bezeichnet als RA- (Ras associating) Domänen mit ca. 20 Mitgliedern identifiziert worden, wobei erwartet wird, dass die RA-Domänen ebenfalls eine Ubiquitin-Faltung aufweisen (Ponting & Benjamin, 1996). Die Proteine, die RA Domänen enthalten, sind zum einen einige der oben beschriebenen typischen Ras Effektoren, wie RalGDS, AF6, PLCε, Rin und NORE1. In diesen Proteinen entspricht die RA-Domäne somit der RBD. Die RBDs von Raf und Byr2 dagegen wurden nicht als RA-Domänen gefunden. Andere Proteine, die RA-Domänen enthalten, sind z.B. Epac1, Krit1, Repac1, Tiam und PDZGEF. Dabei ist aber noch nicht untersucht, ob diese Proteine auch an Ras binden; dadurch könnte die Signalweiterleitung noch komplexer werden.
Kinetische Untersuchungen haben gezeigt, dass die Bindung von RBDs an Effektoren sehr dynamisch ist, d.h. sowohl die Assoziations- als auch die Dissoziationsratenkonstanten sind sehr schnell (Hermann et al., 1996; Sydor et al., 1998; Gronwald et al., 2001; Linnemann et al., 2002). Damit sind die mittleren Lebenszeiten der Komplexe nur sehr kurz (z. B. für die Bindung von Raf-RBD an Ras ist sie 0.14 s bei 25 °C, Sydor et al., 1998), was eine Bedeutung für regulative Prozesse haben könnte, z.B. für die Ausübung der Funktionen der GAPs. Des Weiteren konnte gezeigt werden, dass die Assoziation von RBDs an Ras Proteine bei höheren Konzentrationen gesättigt werden kann, was einen Bindungsmechanismus mit mindestens zwei Schritten anzeigt (Sydor et al., 1998; Linnemann et al, 1999; Linnemann et al., 2002). Da der geschwindigkeitsbestimmende Schritt für alle RBD/Ras Protein Komplexe ähnlich ist, wird vermutet, dass die Ursache in den Konformationsänderungen von Schalter I und Schalter II liegen könnte. Diese Hypothese wird dadurch unterstützt, dass mit Hilfe von Phosphor-NMR zwei Populationen der chemischen Verschiebung des ß-Phosphats gefunden wurden, was den zwei Schalter Zuständen zugeordnet wurde (Geyer et al., 1996). Bei Titration von Effektor RBDs wird dabei ein Zustand stärker populiert.
1.5 Die Signalweiterleitung über Ras als Angriffspunkt zur
Entwicklung von möglichen Medikamenten in der Krebstherapie
Das normale Verhalten jeder einzelnen Zelle in einem vielzelligen Organismus wird streng kontrolliert durch ein Netzwerk verschiedener Signalwege, in denen die verschiedenen Ras Proteine eine Schlüsselfunktion besitzen. Diese Kontrolle stellt sicher, dass die Zellen sich nur dann vermehren, wenn es erwünscht ist, wie z.B. während der Entwicklung oder bei der Wundheilung. Es kommt zur Krebsentstehung, wenn die normale Wachstumskontrolle nicht mehr funktioniert, z.B. aufgrund von Defekten innerhalb der beteiligten Signalwege (Übersicht: Downward, 2003). Die gestörte Signalweiterleitung in Krebszellen ist die Folge

EINLEITUNG
12
von verschiedenen Mutationen – am häufigsten wurden Mutationen des Ras Gens gefunden: 20 % aller menschlichen Tumore besitzen aktivierende Punktmutationen in ras (fast alle im Codon 12, 13 oder 61, Bos, 1989), am häufigsten in K-ras (ca. 85 %), gefolgt von N-ras (ca. 15 %) und H-ras (weniger als 1 %). Diese Mutationen verhindern die GAP katalysierte GTP Hydrolyse, so dass Ras permanent in der aktivierten GTP-gebundenen Form vorliegt. Ras bedingte Tumore können aber auch durch das Fehlen von GAPs verursacht werden. Das bekannteste Beispiel hierfür ist Neurofibromin, welches von dem nf1 Gen kodiert wird (Weiss et al., 1999). Der Verlust eines Alleles führt zur Typ 1 Neurofibromatose, der Verlust beider Allele zur permanenten Aktivierung von Ras. Außerdem können Signalweiterleitungen über Ras auch in Tumoren aktiviert werden als Folge der Überexpression von Rezeptor-Tyrosin-Kinasen oder bei Expression von extrazellulär verkürzten Rezeptoren. Beispiele hierfür sind der EGF- und der ERBB2- Rezeptor, welche häufig in Tumoren der Brust, der Eierstöcke und des Magens überexprimiert sind (Mendelsohn & Baselga, 2000).
Medikamente gegen Krebs können somit viele Angriffspunkte in den verschiedenen Signalwegen haben. Vier Strategien, die direkt auf die gestörte Signalweiterleitung von onkogenem Ras abzielen, werden nachfolgend diskutiert (Übersicht: Wittinghofer & Waldmann, 2000). Eine Möglichkeit ist die Inhibierung von posttranslationalen Modifikationen von Ras, da die Lokalisation an der Plasmamembran notwendig für die biologische Funktion ist (Hancock et al., 1990). Der erste Schritt dieser Modifizierungen erfolgt durch das Enzym Farnesyltransferase (FT) und daher zielen viele Strategien auf die Inhibierung dieses Enzyms ab. Es wurden einige sehr effiziente FT-Inhibitoren gefunden (Übersicht: Cox & Der, 2002) und bereits an transgenen Mäusen viel versprechend getestet (Kohl et al., 1995).
Eine weitere Möglichkeit zur Verhinderung der Sinalweiterleitung über Ras ist die Inhibierung des Nukleotid-Austausches von GDP zu GTP. Dies ist aber wahrscheinlich nur sinnvoll, wenn die Signalweiterleitung oberhalb von Ras betroffen ist, da onkogenes Ras fast ausschließlich an GTP gebunden vorliegt (Trahey & McCormick, 1987). Es konnten kompetitive Inhibitoren gefunden werden, die den Nukleotid-Austausch inhibieren (mit IC50-Werten im mikromolaren Bereich), aber nicht mit der Bindung um GDP konkurrieren (Taveras et al., 1997). Die Entwicklung verschiedener GTP- Analoga mit Veränderungen im Riboseteil dagegen führte zu einer hohen Affinität zu normalem und onkogenem Ras - bis zu 2 fach höher als GDP (Noonan et al., 1991). Einige davon zeigten in vitro eine, wenn auch nur mittelmäßige Inhibierung des Nukleotid-Austausches (Wolin et al., 1996).
Da GAPs ihre Wirkung an onkogenem Ras nicht ausüben können und somit Ras nicht mehr in den inaktiven Zustand überführt werden kann, ist eine weitere Strategie Moleküle zu finden, die die GTPase Aktivität von onkogenem Ras stimulieren können. Hierbei ist das Molekül DABT-GTP von besonderem Interesse: Ursprünglich geplant als ein nicht-hydrolysierbares GTP-Analog, wurde es entgegen den Erwartungen durch die Gα-

EINLEITUNG
13
Untereinheiten von heterotrimeren G-Proteinen (Zor et al., 1997; Zor et al., 1998) und auch durch Ras sowie allen onkogenen Ras Mutanten (einschließlich G12V) sehr effizient hydrolysiert (Ahmadian et al., 1999). Dies zeigt, dass die GTP-Hydrolyse auch an onkogenem Ras prinzipiell möglich ist.
Zur Inhibierung der Wechselwirkung zwischen Ras und Effektoren wurden Peptide entwickelt, basierend auf der Sequenz der Effektor-Bindungs-Region von Ras. Auf diese Weise wurden Peptide gefunden, die die Bindung von Raf an Ras⋅GTP mit mikromolaren IC50-Werten inhibieren (Barnard et al., 1995). Vielversprechender ist die Inhibierung der Ras-Raf Wechselwirkung durch Sulindacsulfid und Derivaten davon: Diese wurden in mehreren Studien getestet und scheinen die Signalweiterleitung über Ras in vivo zu blockieren (Thompson et al., 1997, Herrmann et al., 1998, Karaguni et al, 2002); in vitro wurde eine Inhibierung der Ras-Raf Wechselwirkung nachgewiesen (Herrmann et al., 1998).
1.6 Analyse von Bindungsenergien in der Kontaktfläche
zwischen Proteinen und Liganden Die spezifischen Wechselwirkungen zwischen Proteinen haben eine große Bedeutung in fast allen Signalweiterleitungs-Prozessen der Zelle, die wie ein kompliziertes Netzwerk miteinander verbunden sind. In ebensolchem Maße verursachen Störungen in diesen streng kontrollierten Netzwerken viele Krankheiten. Als mögliche Medikamente kommen dann Moleküle in Betracht, die gezielt unerwünschte Wechselwirkungen inhibieren (siehe Kapitel 1.5). Solche Inhibitoren wurden bisher fast ausschließlich durch das Testen einer sehr großen Anzahl chemischer Substanzen (HTS, high throughput screening) - häufig auch Abwandlungen von natürlichen Inhibitoren gefunden. Da Moleküle zur gezielten Blockierung von Protein-Protein Interaktionen hoch spezifisch sein und sehr fest binden sollten, ist die Methode des HTS sehr zeitaufwendig und häufig mit Glück verbunden. Besser wäre eine rationale Molekülentwicklung. Aber selbst wenn die Kristallstrukturen von einem Proteinkomplex vorliegen, und mögliche Inhibitor Substanzen durch das Anpassen an die Protein-Kontaktfläche („modeling“) entwickelt werden, ist häufig die Bindung solcher Stoffe zu schwach. Der Grund dafür ist, dass immer noch nicht ausreichend verstanden ist, mit welchen energetischen Beiträgen einzelne Aminosäuren wechselwirken. Beispielsweise trägt die Aminosäure Arg89 von Raf-RBD im Komplex mit Ras mindestens die Hälfte zur gesamten Bindungsenergie bei, und auch die Mutation von Arg89 zu der sehr ähnlichen Aminosäure Lys reduziert die Affinität drastisch. Andere Arginine in der Kontaktfläche dagegen haben nur eine geringe Auswirkung auf die Bindungsenergie.
Die Affinität eines Protein Komplexes mit der Dissoziationskonstanten (Kd) ist eine Funktion der Assoziations- und der Dissoziationsratenkonstanten (kon und koff), wobei die Beziehung

EINLEITUNG
14
Kd = koff/kon gilt. Man kann deutlich zwischen den Kräften die zu kon und koff beitragen unterscheiden (Selzer & Schreiber, 2001). So hängt die Dissoziationsratenkonstante hauptsächlich von der Stärke der kurz-reichenden („short-range“) Wechselwirkungen zwischen den Proteinen, wie van-der Waals Wechselwirkungen, Wasserstoff-brückenbindungen, hydrophobe Wechselwirkungen und Salzbrücken, ab (Selzer et al., 2000). Im Gegensatz dazu ist die Rate der Assoziation durch Diffusion getrieben und kann durch günstige elektrostatische weit-reichende („long-range“) Wechselwirkungen beschleunigt werden (Shoup & Szabo, 1982; Berg & van Hippel, 1985; Schreiber & Fersht, 1996).
Basierend auf der Beziehung Kd = koff/kon können höhere Affinitäten zwischen Proteinen prinzipiell sowohl mit langsameren Dissoziations- als auch mit schnelleren Assoziationsratenkonstanten erreicht werden. Um Protein Komplexe mit langsameren Dissoziationsratenkonstanten zu entwerfen ist das genaue Verständnis der energetischen Beiträge zwischen Aminosäurekontakten nötig, aber dies ist zur Zeit noch schwierig, da nur wenige allgemeine Gesetzmäßigkeiten hierüber bekannt sind (Shi, 1993). In den vergangenen Jahren erfolgten viele theoretische und empirische Untersuchungen um die Affinität von Protein-Komplexen – basierend auf den dreidimensionalen Strukturen - vorherzusagen (Übersicht: Gohlke & Klebe, 2002). Ziel ist die Entwicklung von Energie Funktionen, die verschiedene Energie-Terme enthalten, welche zur Komplexstabilität beitragen. Man unterscheidet dabei zwischen drei verschiedenen Ansätzen für die Entwicklung von Energie-Funktionen: (i) rein physikalische Ansätze (PEEF, physical effective energy function), (ii) statistische Ansätze (SEEF, statistical EEF), die auf Potentialen basierend auf Atom Kontakten aus einer Protein-Datenbank bestehen (Lazaidis & Karplus, 2000) und (iii) empirische Ansätze (EEEF, empirical EEF), die eine Kombination aus einer physikalischen Beschreibung und der Information aus Messungen einer großen Anzahl von Protein Wechselwirkungen sind. Insbesondere die EEEF-Ansätze sind vielversprechend und darauf basierend entwickelte die Arbeitsgruppe von Dr. Luis Serrano vom EMBL in Heidelberg die Energie Funktion FOLDEF (FOLD-X energy function) welche Korrelationskoeffizienten von 0.8 für den Vergleich von experimentellen und berechneten Werten für die freien Enthalpieänderungen ∆G° bei der Komplexbildung liefert (Guerois et al., 2002).
Einfacher sollte es sein die Assoziationsrate zu beschleunigen, indem die elektrostatische Komplementarität zwischen den Proteinen erhöht wird. Dieses Prinzip wird in der Natur häufig gefunden für Wechselwirkungen zwischen Protein Paaren, bei denen die Geschwindigkeit entscheidend für die biologische Funktion ist. Beispiele hierfür sind die schnelle Assoziation zwischen Barnase und dem zellulären Inhibitor Barstar (Schreiber et al., 1994), zwischen RNase und seinem Inhibitor RI (Lee et al., 1989) und für den Proteinkomplex zwischen Acetylcholinesterase und Fasciculin (Harel et al., 1995; Radic et al., 1997). Die Assoziation zweier Moleküle beginnt mit der zufälligen Annäherung gefolgt von dem genauen Andocken der zwei Kontaktflächen. Die Rate dieser zufälligen Annäherung wird durch die Brownsche Bewegung bestimmt und führt nach der Smoluchowski-Einstein

EINLEITUNG
15
Gleichung (Smoluchowski, 1917) zu einer Kollisionsrate von ca. 7* 109 µM-1 s-1, abhängig vom Diffusionskoeffizienten und damit von der Größe der Moleküle. Diese zufällige Kollision führt aber nicht automatisch zu einer Komplexbildung, denn hierfür ist zusätzlich die richtige Orientierung der Moleküle zueinander von entscheidender Bedeutung. Dadurch wird die Assoziationsratenkonstante der Reaktion um drei bis fünf Größenordnungen reduziert. Schnellere Ratenkonstanten, die dennoch für einige Reaktionen beobachtet werden, können durch günstige elektrostatische Kräfte erklärt werden. Dieses wird als elektrostatische Lenkung („elektostatic steering“) bezeichnet. In Abbildung 1.7 ist schematisch das Energieprofil der freien Enthalpie für die Bildung eines Proteinkomplexes AB aus den freien Proteinen A und B dargestellt. Die Proteine A und B diffundieren zuerst frei in der Lösung, bis sie einen nur gering stabilen Komplex AB* („encounter complex“) bilden, welcher zur Dissoziation neigt (k-1 >>k2). Der Komplex AB* verläuft über den Übergangszustand [AB]#, welches schon in solcher Orientierung vorliegt, dass der Endkomplex AB gebildet werden kann. In Anwesenheit von günstigen elektrostatischen Wechselwirkungen zwischen den Proteinen A und B wird die Bildung des Komplexes AB* begünstigt und somit die Assoziationsratenkonstante schneller.
„steering region“
[AB]#
k2
k-2 k-1
k1 G
AB* „encounter complex“
A+B freie Proteine
AB Komplex
Reaktionskoordinate
ABBILDUNG 1.7: Bildung eines Protein Komplexes. Schematisches Energieprofil der freien Enthalpie (G) bei der Bildung eines Proteinkomplexes AB aus den freien Proteinen A und B über den „encounter complex“ AB* und den Übergangszustand AB#.
Zur theoretischen Beschreibung der Assoziationsratenkonstanten wurden mehrere Methoden verwendet. Die Gruppe von Dr. Gideon Schreiber vom Weizmann-Institut in Israel entwickelte das Computer Programm PARE (Protein Association Rate Enhancement) zur Berechnung von Assoziationsraten von Mutanten Komplexen. Das Programm berechnet kon aus der Debye Hückel Energie des Mutanten Komplexes und einem experimentell ermittelten

EINLEITUNG
16
Wert für kon für die Assoziation des Wildtyp Protein Komplexes. Die Fähigkeit von PARE zur Berechnung von Assoziationsratenkonstanten wurde am Protein Komplex für TEM und BLIP getestet und liefert Korrelationskoeffizienten von 0.95 (Selzer et al., 2000). Für die Affinität und Dynamik der Bindung der Effektor RBDs an Ras Proteine wird ebenfalls angenommen, dass elektrostatische Wechselwirkungen eine zentrale Rolle haben, und es wäre wichtig, diese Bedeutung in Hinsicht auf die zelluläre Signalweiterleitung zu analysieren.
1.7 Zielsetzung der Arbeit Die verschiedenen Proteine der Ras Familie sind wichtige molekulare Schalter der Zelle, indem sie die Signale von Membran-Rezeptoren zum Zellkern weiterleiten. Ein zentraler Punkt ist hierbei die Wechselwirkung von Ras mit Effektor Proteinen, wobei gezeigt werden konnte, dass die zelluläre Signalübertragung mit der Affinität der Effektor-RBD Bindung an Ras korreliert (Block et al., 1996; Jaitner et al., 1997).
Ziel dieser Arbeit ist es die Rolle der elektrostatischen Wechselwirkungen zwischen Ras und Effektoren für die Signalübertragung in der Zelle zu analysieren. Wie bereits gezeigt wurde, besteht eine gute Korrelation zwischen der elektrostatischen Komplementarität in einem Protein-Komplex und der Größe der Assoziationsratenkonstanten, was zur Entwicklung von Methoden zur theoretischen Beschreibung von Assoziationsratenkonstanten genutzt wurde (siehe Kapitel 1.6; Selzer et al., 2000). Es sollte getestet werden, ob auf diese Weise die Dynamik von Ras/Effektor RBD Komplexen gezielt verändert werden kann. Da Ras in der Effektor-Bindungsregion hauptsächlich negativ geladen ist, und die RBDs der Effektoren hauptsächlich positiv geladen sind, wird die Strategie verfolgt, Aminosäuren am Rand der Kontaktfläche der Effektor RBDs zu positiv geladenen Aminosäuren zu mutieren, um die elektrostatische Komplementarität zu erhöhen. Es wird erwartet, dass die Dissoziationsratenkonstanten dadurch nicht verändert werden, da keine kurz-reichenden Wechselwirkungen in der Kontaktfläche zerstört werden und somit sollten, da die Assoziationsratenkonstante größer wird, bessere Affinitäten vorliegen.
Die hergestellten Mutanten Komplexe sollen detailliert kinetisch untersucht werden, insbesondere auch der Mechanismus der Bindung, d.h. wie die einzelnen Teilschritte beeinflusst werden. Der Vergleich von experimentellen und berechneten Assoziations-ratenkonstanten dient zur Feststellung, ob die theoretischen Berechnungen auch auf das Ras/Effektor System anwendbar sind oder ob eventuell Veränderungen des theoretischen Modells sinnvoll sind. In zellulären Assays soll der Einfluss von Mutanten Komplexen mit veränderten elektrostatischen Eigenschaften auf die Signalweiterleitung in der Zelle analysiert werden. Interessant sind dabei auch solche Mutationen, bei denen Aminosäuren in der

EINLEITUNG
17
Kontaktfläche mutiert werden: hier erwartet man neben einer größeren Assoziationsratenkonstanten auch eine Beschleunigung der Dissoziation, wobei die Affinitäten gleich bleiben könnten. Es wird interessant sein zu sehen, wie der Effekt von solchen Mutanten Komplexen in der Zelle aussieht, d.h. ob die Signalübertragung nur eine Frage der Affinität oder auch eine Frage der Dynamik ist.
Neben der speziellen Betrachtung der Bedeutung der elektrostatischen Wechselwirkungen anhand des Ras/Effektor Systems sollen aber auch die Prinzipien von Protein Wechselwirkungen im Allgemeinen besser verstanden werden. Daher sollen auch die thermodynamischen Parameter von den Mutanten Komplexen untersucht werden, d.h. die Beiträge der Enthalpie- und Entropieänderungen zur Affinität. Zum einen hat man dadurch eine zusätzliche Kontrolle der Affinitätsmessung – neben den kinetischen Messungen also auch eine Methode der Affinitätsbestimmung im Gleichgewicht. Zum anderen wäre es interessant zu sehen, ob man kinetische und thermodynamische Parameter korrelieren kann.
Eine weitere Fragestellung ist die Bedeutung der GTP-gebundenen Form für die Wechselwirkung mit Effektoren. Wie gezeigt werden konnte, ist die Affinität wesentlich höher bei der Bindung von Effektoren an Ras im GTP- als im GDP-Zustand. Von Interesse wäre, ob man durch Veränderung der Elektrostatik auch Komplexe von Effektor RBDs mit Ras⋅GDP herstellen kann, die eine bessere Affinität zeigen. Hier wären wiederum die Auswirkungen in der Zelle sehr interessant zu analysieren. In diesem Zusammenhang ist auch wissenswert, welche Rolle die Schalteränderungen für die Bindung an Effektoren haben und für die Signalübertragung in der Zelle haben.
Von Mutanten Komplexen, die bedeutsame kinetische oder thermodynamische Effekte zeigen oder die eine ungewöhnliche Signalweiterleitung in der Zelle zeigen, sind auch strukturelle Analysen wichtig, wie z.B. Kristallstrukturen oder NMR- Experimente.
Die Ergebnisse dieser Arbeit könnten die Basis für die Entwicklung von neuen Prinzipien in der Wirkstoffentwicklung sein, die nicht darauf abzielen, spezifisch die Kontakte zwischen Proteinen zu inhibieren, sondern die eine Abstimmung der Dynamik von verschiedenen Wechselwirkungen mit einbeziehen. Eventuell könnten auf diese differenzierte Weise verschiedene Signalwege unterschiedlich aktiviert und beeinflusst werden. Möglicherweise wäre dies ein neuer Ansatz zur Entwicklung von Medikamenten bei der gestörten Ras-Signalweiterleitung in Tumoren: Da sich ausgehend vom Ras Protein die Signalweiterleitung über die Effektoren stark verzweigt und die verschiedensten Reaktionen in der Zelle hervorruft, wäre eine differenzierte Abstimmung der gestörten Signalweiterleitung wünschenswert.

MATERIALIEN UND METHODEN
18
II MATERIALIEN UND METHODEN
2.1 Materialien 2.1.1 Chemikalien Alle Chemikalien wurden von einer der folgenden Firmen bezogen: Aldrich (Steinheim), Baker (Deventer, Holland), Fluka (Neu-Ulm), GERBU (Gaiberg), Merck (Darmstadt), Roth (Karlsruhe), Serva (Heidelberg) und Sigma (Deisenhofen).
2.1.2 DNA-Konstrukte Ras und Rap Konstrukte: ptac:H-RasWT, ptac:H-RasE31A, ptac:H-RasE31K, ptac:H-RasD33A, ptac:H-RasT35A, ptac:H-RasT35S, ptac:H-RasE37A, ptac:H-RasD38A, ptac:H-RasE62A, ptac:H-RasE63A, ptac:rapC`WT+pGROE; ptac:rapC`E30D,K31E jeweils in CK600K (D. Kühlmann, Dortmund). RalGDS-RBD Konstrukte: pGEX4T3:RalGDS-RBDR16A, pGEX4T3:RalGDS-RBDK28A, pGEX4T3:RalGDS-RBDK44A, pGEX4T3:RalGDS-RBDD47A, pGEX4T3:RalGDS-RBDD47K, pGEX4T3:RalGDS-RBDK48A, pGEX4T3:RalGDS-RBDE52K, pGEX4T3:RalGDS-RBDE53A, jeweils in BL21, (T. Linnemann, Dortmund); pGEX2T:RalGDS-RBDWT, pGEX2T:RalGDS-RBDN50K jeweils in BL21 (S. Wohlgemuth, Dortmund). Raf-RBD Konstrukte: pGEX2T:Raf-RBDR59A, pGEX2T:Raf-RBDN64D, pGEX2T:Raf-RBDK65A, pGEX2T:Raf-RBDK65E, pGEX2T:Raf-RBDR67A, pGEX2T:Raf-RBDK84A, pGEX2T:Raf-RBDK84E jeweils in BL21 (B. Voss, Dortmund), pGEX2T:Raf-RBDM83K, pGEX2T:Raf-RBDM83D jeweils in BL21 (S. Wohlgemuth, Dortmund). Konstrukte für Reportergen-Assay: pCDNA3, pCDNA3:Raf fl.WT, pSVK3:Ras fl. G12V, pSVK3:Rasfl.WT, E743-tk80-luc (Reportergen), pEQ176 (ß-Galaktosidase Expressions-konstrukt), p44 Map-kinaseERK1), pSGER81 (Ets-Transkriptionsfaktor) (B. Voss, Dortmund). 2.1.3 Enzyme, Proteine und Antikörper Alkalische Phosphatase (Boehringer, Mannheim); Antikörper Maus �-HA: (12CA5) Roche; Pfu-Polymerase (Stratagene, Heidelberg); Proteinstandard SDS 7, MG:14.2, 20, 24, 29, 36, 45 und 66 kDa (Sigma, Deisenhofen); Restriktionsendonukleasen (Boehringer, Mannheim), Thrombin (Serva, Heidelberg) und BSA (Pierce, Illinois, USA). 2.1.4 Geräte Differential Scanning Kalorimeter (MicroCal, Northampton,USA); Elektrophorese (BioRad, München); Fluoreszenzspektrometer (Luminescence Spectrometer LS50B, Perking Elmer, Düsseldorf); FPLC-Anlage (GradiFrac System, Pharmacia, Freiburg); Gelscanner (Bioprofil 5.0, Fröbel, Lindau); Heizblock (Thermomixer 5436, Eppendorf, Hamburg); HPLC-Anlage

MATERIALIEN UND METHODEN
19
(HPLC NEC System Gold, Beckmann, München); Mikrokalorimeter (MCS ITC, MicroCal, Northampton,USA); NMR-Spektrometer (AMX500, Bruker, Karlsruhe); PCR-Thermocycler (DNA Trio-Thermoblock, Biometra, Göttingen); pH-Meter (Calimatic 765, Knick, Berlin); Photometer (Pharmacia Ultrospec III (Pharmacia, Freiburg); Phospho-Imager (Phospho Analyst, Biorad, München); Stopped-Flow Apparatur (Applied Photophysics, Leatherhed, UK) und Ultraschallgerät (Sonifier 450, Branson Ultrasonics, Danbury, USA). 2.1.5 Lösungen Agarose-Lösung (0.5 bis 2 g Agarose/100 ml TAE-Puffer); Entfärbe-Lösung (40 % Ethanol, 10 % Essigsäure); Färbe-Lösung (0.25 % Coomassie-Brilliantblau R250, 40 % Ethanol, 10 % Essigsäure); Bradford-Reagenz (Coomassie Protein Assay Plus Reagenz, Pierce, Illinois, USA); PMSF-Lösung (0.1 M PMSF in Isopropanol); Superfect (Qiagen, Hilden); Trypsin-Lösung (0.5 g/l Trypsin, 0.2 g/l EDTA, Gibco, Eggenstein) und FCS-Serum (Gibco, Eggenstein). 2.1.6 Materialien für die Chromatographie GSH-Sepharose, DEAE-Sepharose, Q-Sepharose und Superdex G75 Gelfiltration: Pharmacia, Uppsala, Schweden. 2.1.7 Mikroorganismen und Zelllinien E-coli-Stämme (BL21, Studier & Moffat, 1986; CK600K; TG1, Gibson, 1984) und Zelllinien (COS-7, Nierenfibroblasten der grünen Meerkatze; RK13, Zelllinie aus Kaninchennieren). 2.1.8 Nährmedien DMEM (mit Pyridoxin, mit Glutamax-1, Gibco, Eggenstein); LB-Medium (10 g Bacto Trypton, 5 g Hefextrakt, 10 g NaCl, 5 mM NaOH, ad 1 l H2O); LB-Amp-Medium (LB-Medium + 50 µg/ml Ampicillin); MEM (Modified Eagle`s Medium, mit Glutamax-1 und Earle`s Dalzen, Gibco, Eggenstein); LB-Agar (10 g Bacto Trypton, 5 g Hefeextrakt, 10 g NaCl, 15 g Bacto Agar, ad 1 l H2O); Standard-I-Medium (25 g Nährboullion, 5 g Thiamin, 0.011 g Thymin, ad 1 l H2O); TB-Medium (12 g Bacto Trypton, 24 g Hefeextrakt, 4 ml Glycerol, 0.17 mM KH2PO4, 0.072 mM K2HPO4, ad 1 l H2O) 2.1.9 Puffer-Lösungen Anodenpuffer: 0.025 M Tris/HCl, pH 8.9. DNA-Auftragspuffer (6fach): 0.25 % Bromphenolblau, 0.25 % Xylencyanolblau, 30 % Glycerol. Elutionspuffer (Ral-Assay): 4mM EDTA, 0.1 % SDS, 2mM DTE, 0.5 mM GTP, 0.5mM GDP Gelpuffer: 3 M Tris/HCl (pH 8.45), 0.3 % SDS, 0.001 mM NaN3.

MATERIALIEN UND METHODEN
20
HEBS-Puffer (Reportergen-Assay) (50 mM Hepes (pH 7.05), 140 mM NaCl, 1.5 mM Na2HPO4. HPLC-Puffer: 100 mM K2HPO4/KH2PO4 (pH 6.5), 10 mM Tetrabutylammoniumbromid, 7.5 % bzw. 20 % (v/v) Acetonitril. Kathodenpuffer: 0.1 M Tris/HCl (pH 8.25), 0.1 M Tricin, 0.1 % SDS, 0.1 mM NaN3. Luciferase-Assay Reagenz: 20 mM Tricine, 1.07 mM (MgCO3)4Mg(OH)2, 2.67 mM MgSO4, 0.1 mM EDTA (pH 8.0), 33.3 mM DTE, 270 µM Coenzym A, 470 µM Luziferin, 530 µM ATP, mit 1 M Tris auf pH 7.8. Lysispuffer (Ral-Assay): 50 mM Tris/HCl (pH 7.4), 140 mM NaCl, 1 % Nonidet P40, 1 % Triton X-100, 2 mM MgCl2, 1 mM KCl, 1 mM KPO4, 1 mM PMSF. Lysis-Puffer (Reportergen-Assay): (25 mM Tris/HCl (pH 7.8), 2 mM EDTA, 10 % Glycerin, 1 % Triton X-100, 2 mM DTE. Nukleotid-Austauschpuffer: 20 mM Tris/HCl (pH 7.5), 5 mM MgCl2, 1 mM DTE, 0.2 M (NH4)2SO4, 0.1 µM ZnCl2, 10 mM EDTA. PBS-Puffer: 10.2 mM Na2HPO4, 1.7 mM KH2PO4 (pH 7.45), 137 mM NaCl, 2.7 mM KCl SDS-Probenpuffer (5fach): 10 % Glycerin, 5 % β-Mercaptoethanol, 3 % SDS, 0.0625 M Tris/HCl (pH 8.8), 0.1% Bromphenolblau. TAE-Puffer: 40 mM Tris/Acetat, 1 mM EDTA. Waschpuffer (Ral-Assay): 50 mM Tris/HCl (pH 7.5), 300 mM NaCl, 5 mM MgCl2, 0.1 % Triton X-100, 0.01% SDS. TM-Puffer (Ral-Assay): 10 mM Tris/HCl (pH 7.5), 2 mM MgCl2. Z-Puffer (Reportergen-Assay): (60 mM Na2HPO4, 40 mM KH2PO4 (pH 7), 10 mM KCl, 1 mM MgSO4, 2 mM DTE, 1 mg/ml o-Nitrophenyl-ß-D-galactosid.
2.2 Methoden 2.2.1 Molekularbiologische Methoden
2.2.1.1 PCR-Mutagenese Zur Einführung von Mutationen in DNA-Sequenzen wurde der QuikChangeTM Site-Directed Mutagenesis Kit verwendet. Das Prinzip dieser Methode beruht auf der Verwendung eines Vektors als Template und zweier Oligonukleotid-Primer, die die gewünschte Mutation enthalten. Dabei wird das gesamte Plasmid mit den Primern einer PCR-Reaktion unterzogen und anschließend die Wildtyp-DNA mit der methylierungsabhängigen Restriktions-endonuclease DpnI gespalten. Der 50 µl PCR-Ansatz enthielt 5 µl 10fach Pfu-Puffer, 5 µl DMSO, 5 bis 50 ng Template DNA, 0.5 µl dNTPs, jeweils 125 ng des jeweiligen Oligonukleotid-Primers und 1 µl Pfu-Polymerase. Zur Einführung von Mutationen in RalGDS-RBD, Raf-RBD und Ras wurde die PCR-Reaktion mit folgenden Schritten durchgeführt: 94 °C (5 min)/Start Zyklus 1/92 °C (1 min)/50 °C (2 min)/68 °C (4 min)/25

MATERIALIEN UND METHODEN
21
mal/4 °C. Für Mutationen in Raf fl. wurden folgende Schritte durchgeführt: 94 °C (5 min)/Start Zyklus 1/92 °C (1 min)/52 °C (2 min)/68 °C (10 min)/25 mal/4 °C. Anschließend wurde der PCR-Ansatz mit 2 µl Dpn1 für 2 Stunden bei 37 °C inkubiert. Im Anhang sind für alle Mutagenesen die jeweiligen verwendeten Template-Vektoren und Oligonukleotid-Paare gelistet. Der Entwurf der Länge der Oligonukleotide erfolgte so, dass die Summe aus AT-Gehalt mal 2 und CG-Gehalt mal 4 zwischen 80 und 90 lag. 2.2.1.2 Restriktion von Plasmid-DNA Die Spaltung von doppelsträngiger DNA wurde unter den vom Hersteller angegebenen Puffer- und Temperaturbedingungen durchgeführt. Anschließend wurden die Restriktionsansätze mittels Agarose-Gelelektrophorese gereinigt. 2.2.1.3 Agarose-Gelelektrophorese und Gelelution Die Auftrennung von DNA-Fragmenten erfolgte durch Gelelektrophorese. Entsprechend der Größe der Fragmente wurden Agarosekonzentrationen von 0.5 % bis 1 % in TAE-Puffer verwendet. Die Proben wurden vor dem Auftragen mit DNA-Probenpuffer versetzt. Als Größenstandard wurde Lambda-DNA verwendet. Die Elektrophorese erfolgte bei 90 Volt. Anschließend erfolgte eine Inkubation des Gels in Ethidiumbromid-Lösung. Die Isolierung der DNA-Fragmente erfolgte mit dem Gelelutions-Kit der Firma Qiagen nach Angaben des Herstellers. 2.2.1.4 Ligation von DNA-Fragmenten Die Ligation von geschnittenem Vektor und Insert erfolgte in einen 20 µl-Ansatz mit 1 U T4-DNA-Ligase, 2 µl 10fach Ligase-Puffer und einem 5-fachen Überschuss von Insert- gegenüber Vektor-DNA. Dieser Ansatz wurde für vier Stunden bei 30 °C inkubiert. 2.2.1.5 Herstellung kompetenter Bakterien Kompetente Bakterien wurden nach der Methode von Chuang et al. 1989 hergestellt. Dazu wurden Bakterien der Stämme TG1 oder BL21 als Einzelklon in 5 ml LB-Medium über Nacht bei 37 °C inkubiert. Mit dieser Vorkultur wurden 100 ml LB-Medium 1:100 angeimpft und bei 37 °C bis zu einer OD600 von 0.5 inkubiert. Die Bakterien wurden auf Eis abgekühlt und mit 4000 rpm bei 4 °C zentrifugiert. Anschließend erfolgte die Resuspendierung in LB-Medium mit 10 % w/v PEG 8000, 5 % v/v DMSO und 50 mM MgCl2. Nach Zugabe von 15 % v/v Glycerin wurde die Suspension auf Eis aliquotiert, in flüssigem Stickstoff schockgefroren und bei -80 °C gelagert. 2.2.1.6 Transformation von Bakterien Zur Transformation wurden 100µl kompetente Bakterien auf Eis aufgetaut, mit 1 bis 10 µl einer Plasmid-Lösung vorsichtig gemischt und für 45 Minuten auf Eis inkubiert.

MATERIALIEN UND METHODEN
22
Anschließend erfolgte ein Hitzeschock bei 42 °C für 60 Sekunden mit nachfolgender Regeneration auf Eis. Nach Zugabe von 800 µl LB-Medium wurden 100 µl auf LB-Amp Platten ausgestrichen. Die restlichen Bakterien wurden in einer Tischzentrifuge sedimentiert, der Überstand bis auf 100 µl abgegossen, das Sediment resuspendiert und auf einer weiteren LB-Amp Platte verteilt. Die Platten wurden bei 37 °C über Nacht inkubiert. Am folgenden Tag wurden Klone mit einer sterilen Pipettenspitze gepickt und in 2 ml LB-Amp-Medium bei 37 °C inkubiert. 2.2.1.7 Plasmidpräparation Die Isolierung von Plasmid-DNA erfolgte durch Ionenaustausch-Chromatographie in unterschiedlichem Maßstab unter Verwendung der Mini-, Midi- und Maxi-Präp-Kits von Qiagen nach entsprechenden Protokollen. Die Isolierung beruht auf dem Prinzip der alkalischen Lyse mit anschließender Fällung der Proteine durch Kalium-Dodecylsulfat. Die Konzentration der DNA wurde durch Messung der Extinktion bei 260 nm bestimmt (A260), wobei für eine A260-Einheit 50 µg/ml DNA angenommen wurde. 2.2.1.8 DNA-Sequenzierung Die Sequenzierreaktion erfolgte nach der Kettenabbruch-Methode nach Sanger (Sanger et al., 1977) mit fluoreszenzmarkierten Didesoxynukleotiden. Dazu wurde folgender 20 µl PCR-Ansatz pipettiert: 4 µl Terminator-Mix, 5 µl Plasmid-DNA (aus Minipräparation) und jeweils 1 µl des entsprechenden Sequenzier-Oligonukleotids der Konzentration 10 pmol/µl. Die PCR-Reaktion wurde wie folgt durchgeführt: 96 °C (30 s)/Start Zyklus 1/96 °C (30 s)/50 °C (15 s)/60 °C (4 min)/25 mal/ 4 °C. Zur Fällung der DNA wurde der PCR-Ansatz mit H2O auf 100 µl aufgefüllt und mit 1 µl Dextranblau-Lösung (20 µg/µl in H2O), 10 µl 3 M Na-Acetat (pH 4.8) und 250 µl 100 % Ethanol versetzt und für 20 Minuten mit 14000 rpm zentrifugiert. Direkt danach wurde der Überstand mit einer Pipette abgenommen, 400 µl 70 % Ethanol zugefügt und 5 Minuten mit 14000 rpm zentrifugiert. Der Überstand wurde abgenommen, das Sediment luftgetrocknet und zur Sequenzierung in der zentralen Einrichtung für Biotechnologie im Haus abgegeben.
2.2.2 Proteinchemische Methoden
2.2.2.1 Glycerindauerkulturen, Zellanzucht und Proteinsynthese Zur Herstellung einer Glycerindauerkultur wurden 0.5 ml einer 2 ml Bakterienkultur in ein 1.25 ml Plastikschraubgefäß gegeben, mit 100 % Glycerin aufgefüllt, gemischt und bei -80 °C gelagert. Um 5 l Nährmedium mit Bakterien anzuimpfen, wurde jeweils am Vortag etwas aus einer Glycerindauerkultur entnommen und in einen Erlenmeyerkolben mit 50 ml Nährmedium gegeben. Diese Vorkultur wurde über Nacht bei 37 °C inkubiert. Am folgenden Tag wurden 5 l Medium auf zwei 5-l-Erlenmeyerkolben verteilt und mit jeweils 25 ml der Vorkultur versetzt. Anschließend wurden die Kolben bei 37 °C geschüttelt, so dass die Zellen

MATERIALIEN UND METHODEN
23
bis zu einer optischen Dichte (600 nm) von 0.8 heranwachsen konnten. Die Induktion der Proteinüberexpression erfolgte durch Zugabe von 1 ml 250 mM IPTG-Lösung. Die Temperatur wurde entsprechend den jeweiligen Expressionsbedingungen verändert und die Zellen über Nacht weiter geschüttelt. Die Bakterienkultur wurde zentrifugiert, das Sediment mit dem jeweiligen Aufschlusspuffer gewaschen und bei -80 °C in 50-ml-Falcon-Gefäßen gelagert. 2.2.2.2 Zellaufschluss durch Ultraschall Zum Zellaufschluss wurden die Zellen mit dem doppelten Volumen Aufschlusspuffer auf Eis aufgetaut. Die Suspension wurde mit folgenden Reagenzien 5 mM EDTA (pH 8),1 mM PMSF, 0,5 mM Pefabloc, 1 % Triton, 5 mM DTE und 100 µM GDP (für Ras-und RapC`-Präparationen) versetzt. Der Zellaufschluss erfolgte am Ultraschallgerät mit den Intensitäten Limit 6 und Duty Cycle 60 % für 5 mal 2 Minuten. Anschließend wurde die Suspension bei 18000 rpm zentrifugiert und der Überstand zur weiteren Proteinreinigung weiter verwendet. 2.2.2.3 SDS-Polyacrylamid Gelelektrophorese Die qualitative Analyse von Proteingemischen erfolgte durch ihre Auftrennung nach der Größe in einem denaturierenden Polyacrylamidgel nach Schägger und Jagow. Hierzu wurde ein diskontinuierliches Gel verwendet, welches aus einem Sammelgel (für 2 Gele: 480 µl Acrylamid, 1.8 ml H20, 400 µl Gelpuffer, 48 µl APS (10 %) und 4.8 µl TEMED) mit niedriger (4 %) und einem Trenngel (für 2 Gele: 3.8 ml Acrylamid, 2.2 ml H20, 2.2 ml Gelpuffer, 100 µl APS (10 %) und 10 µl TEMED) mit höherer (14 %) Acrylamid-konzentration bestand. Die Proteinproben wurden mit 5fachem SDS-Probenpuffer versetzt und als Proteinstandard wurde ein SDS-7 Marker verwendet. Nach Befüllung der Elektrophoresekammern mit Anoden- bzw. Kathodenpuffer erfolgte der Elektrophoreselauf mit 45 mA pro Gel. Anschließend wurde das Gel in Färbelösung unter Schwenken für 20 Minuten angefärbt. Entfärbt wurde durch mehrmaligen Wechsel der Entfärbelösung. 2.2.2.4 Bestimmung der Proteinkonzentration nach Bradford Die Ermittlung von Proteinkonzentrationen nach der Bradford-Methode (Bradford, 1976) beruht auf der Bindung des Farbstoffs Coomassie Brilliant-Blau G-250 an Proteine. Dabei verschiebt sich das Absorptionsmaximum von 465 nach 595 nm. Zur Konzentrationsermittlung wurden 1 ml des Bradford-Reagenzes in eine Plastikküvette gegeben und einige Mikroliter der Proteinlösung bis zur leichten Blaufärbung der Lösung zugegeben. Die Absorption dieser Mischung wurde bei einer Wellenlänge von 595 nm gegen Bradford-Reagenz als Leerwert bestimmt. Mit Hilfe einer Eichgeraden, bei der BSA als Standard diente, wurden die entsprechenden Proteinkonzentrationen ermittelt.

MATERIALIEN UND METHODEN
24
2.2.2.5 Reinigung von RalGDS- und Raf-RBD Wildtyp und Mutanten RalGDS- und Raf-RBD wurden als GST-Fusionsproteine exprimiert und über eine GSH-Säule gereinigt. GST-Fusionsproteine sind N-terminal gebunden an den C-terminalen Teil der Gluthation-S-Transferase aus Schistosoma japanicum. Diese Proteine binden hochspezifisch an das Gluthation des GSH-Sepharose 4B Säulenmaterials. Nach Abspaltung des GST-Teils durch Thrombin kann das Protein von der Säule eluiert werden. Die Zellanzucht erfolgte für RalGDS- und Raf-RBD in TB-Medium mit 50 µg/µl Ampicillin. Nach Induktion der Proteinüberexpression mit IPTG erfolgte die Inkubation der Zellen über Nacht bei 25 °C. Am nächsten Tag wurden die Zellen zentrifugiert. Als Aufschlusspuffer wurde 50 mM Tris/HCl (pH 7.4), 5 mM MgCl2 und 300 mM NaCl verwendet. Nach dem Ultraschallaufschluss wurde die Suspension bei 18000 rpm bei 4 °C für 45 Minuten zentrifugiert. Der Überstand wurde auf die mit Puffer (50 mM Tris/HCl (pH 7.4), 5 mM MgCl2 und 300 mM NaCl) equilibrierte GSH-Säule aufgetragen. Anschließend wurde die beladene Säule mit demselben Puffer gewaschen. Danach wurden 150 U Thrombin aufgetragen und über Nacht bei 4 °C (für Raf-RBD) bzw. bei Raumtemperatur (für RalGDS-RBD) inkubiert. Am folgenden Tag konnte das vom GST abgespaltene Fusionsprotein eluiert und die Protein-Lösung mit Hilfe eines Vivaspins konzentriert werden. Der zweite Reinigungsschritt erfolgte durch eine Gelfiltration mit einer Superdex-75-Säule in dem gewünschten Puffer (15 mM Hepes (pH 7.4), 5 mM MgCl2). Bei der Präparation von Raf-RBD enthielt der Puffer zusätzlich 300 mM NaCl. 2.2.2.6 Reinigung von Ras und Rap Wildtyp und Mutanten Die Reinigung von Ras erfolgte im ersten Schritt über eine DEAE-Chromatographiesäule. Hierbei handelt es sich um einen Anionenaustauscher, so dass sich die negativ geladenen Proteine an die positiv geladenen Diethylaminoethan-Gruppen der Säule binden. Die Elution der gebundenen Proteine erfolgt durch einen steigenden Salz-Gradienten. Die ptac-Ras enthaltenen Bakterienzellen CK600K wurden in Standard-I-Medium mit 100 µg/ml Ampicilin und 50 µg/ml Kanamycin kultiviert. Die Proteinexpression erfolgte über Nacht bei 30 °C. Die Zellen wurden in Aufschlusspuffer (30 mM Tris/HCl (pH 7.4), 5 mM MgCl2) gewaschen, mit Ultraschall aufgeschlossen und die Suspension bei 18000 rpm bei 4 °C für 45 Minuten zentrifugiert. Der Überstand wurde auf die mit Puffer (30 mM Tris/HCl (pH 7.4), 5 mM MgCl2) equilibrierte DEAE-Säule aufgetragen. Zur Elution des Ras-Proteingemisches wurde ein Salzgradient von 0 bis 300 mM NaCl verwendet. Ras-Proteinlösungen konnten durch Fällung mit Ammoniumsulfat konzentriert werden. Dazu wurde die Ras enthaltende Proteinlösung durch portionsweise Zugabe von Ammoniumsulfat auf eine 2.4 M Ammoniumsulfat-Lösung eingestellt. Nach einer weiteren Inkubation für 10 Minuten bei 4 °C unter Rühren wurde das Protein durch Zentrifugation bei 13000 rpm sedimentiert. Anschließend wurde das Protein in dem gewünschten Puffer wieder gelöst. Der zweite Reinigunsschritt erfolgte ebenfalls durch eine Gelfiltration mit einer Superdex-75 Säule in dem gewünschten Puffer. Das Rap enthaltene Proteingemisch wurde im ersten

MATERIALIEN UND METHODEN
25
Reinigungsschritt auf eine Q-Sepharose-Säule aufgetragen. Diese Säule ist ebenfalls ein Anionenaustauscher, der quartäre Amine enthält. Das Reinigungsprinzip ist dasselbe wie bei der DEAE-Säule. 2.2.2.7 Nukleotidaustausch an Ras und Rap Proteinen Der Nukleotidaustausch an Ras und Rap gegen GppHNp und mGppNHp erfolgte mit Hilfe von alkalischer Phosphatase in Gegenwart eines 3.5 fachen Überschusses an GppNHp/mGppNHp. GDP wird dabei durch das Enzym zu Guanosin hydrolysiert, welches nur mit geringer Affinität an Ras-Proteine bindet und somit gegen GppNHp/mGppNHp ausgetauscht wird. Zum Austausch wurde die gewünschte Menge an Ras mit 5 mM EDTA (pH 8.6), einem 3.5 fachen molaren Überschuß an GppNHp/mGppNHp, 200 mM (NH4)2SO4, 0.1 µM ZnCl2 und 1 U alkalische Phosphatase pro Milligramm Protein versetzt. Dieser Austauschansatz wurde über Nacht bei 4 °C oder für 4 Stunden bei Raumtemperatur inkubiert. Durch eine anschließende Gelfiltration wurden Salze, überschüssige Nukleotide und Guanosin abgetrennt. Die Konzentrationsbestimmung der Nukleotidlösung erfolgte durch Absorptionsmessung bei 252 nm nach Auftrennung mittels reverse-phase HPLC (high pressure liquid chromatography). Hierbei diente als stationäre Phase eine reverse-phase Säule, auf der sich eine hydrophobe C18-Matrix befindet. Die mobile Phase war eine Tetrabutylammoniumbromid-Lösung (HPLC-Puffer), welche zur Auftrennung von Nukleotiden 7.5 % Acetonitril und für mant-Nukleotide 20 % Acetonitril enthält. Durch Eichung des HPLC-Durchflussphotometers und Einsatz eines Integrators konnte die Konzentration der Nukleotide bestimmt werden. 2.2.2.8 Kristallographie von Proteinen Zur Kristallisation von Raps⋅GDP bzw. Ras⋅GDP im Komplex mit Raf-RBD wurde die Methode des hängenden Tropfens benutzt. Zur Komplexbildung wurden die jeweiligen Proteine in äquimolaren Mengen gemischt und die Abtrennung überschüssiger Moleküle vom Komplex erfolgte durch Gelfiltration. Bei der Methode des hängenden Tropfens wurden Limbro-Platten mit 24 Vertiefungen verwendet und mit 700 µl eines entsprechenden Puffer- Fällungsmittel Gemisches gefüllt. Anschließend wurde jeweils 2 µl aus dem 700 µl Reservoir auf ein Glasplättchen pipettiert und mit 2 µl der Proteinkomplexlösung vermischt. Das Plättchen wurde kopfüber auf eine Vertiefung gelegt und mit Silikonöl luftdicht verschlossen.
2.2.3 Biophysikalische Methoden
2.2.3.1 Stopped-Flow Messungen Zur Analyse der Geschwindigkeit schneller Reaktionen wurde die Stopped-Flow Methode angewendet. Das Prinzip dieser Methode ist, dass die Lösungen zweier Spritzen, die die Bindungspartner enthalten, schlagartig gemischt werden. Dies führt zu einer schnellen

MATERIALIEN UND METHODEN
26
Durchmischung der Lösungen in einer Mischkammer und die Reaktionslösung wird in eine Auffangspritze geführt. Der Reaktionsverlauf kann spektroskopisch verfolgt werden und die Zeitabhängigkeit der Reaktion wird von einem Oszillographen aufgezeichnet. Zur Messung der Assoziationsratenkonstanten (kon) der Bindung von Ras Proteinen an Effektor RBDs enthielt eine Spritze Ras gebunden an mGppNHp (0.5 µM), die andere die RBD Lösung (5 bis 200 µM). Nach schnellem Mischen kommt es zu einer Abnahme der Fluoreszenz, da sich die Umgebung des Fluorophors ändert. Die Anregung der mant-Fluoreszenz erfolgte bei einer Wellenlänge von 366 nm und die Emission wurde durch einen Kantenfilter oberhalb von 405 nm gemessen. Zur Messung der Dissoziationsratenkonstanten (koff) der Reaktion enthielt eine Spritze 0.5 µM Ras⋅mGppNHp im Komplex mit einer ca. 20 mikromolaren RBD-Lösung, die andere Spritze enthielt 50 µM Ras⋅GppNHp (ohne Fluoreszenzmarkierung). Beim Mischen der Lösung kommt es zu einer Zunahme der Fluoreszenz, da Ras⋅mGppNHp durch Ras⋅GppNHp aus dem Komplex mit der Effektor RBD verdrängt wird. 2.2.3.2 Isotherme Titrationskalorimetrie Kalorimetrische Titrationen wurden mit einem Microcal MCS ITC Titrationskalorimeter durchgeführt. Zur Messung der Bindung von Ras Proteinen an Effektor RBDs wurden jeweils 2 ml einer 50 bis 100 µM RBD-Lösung in die Zelle gefüllt und die Spritze mit einer 10 fachen Konzentration an Ras Protein gebunden an GppNHp gefüllt. Die Reinigung der Ras Proteine nach dem Nukleotidaustausch erfolgte mittels Gelfiltration und es wurde der jeweilig verwendete Puffer für die Messung genommen. Dies ist wichtig, da bereits geringfügige Unterschiede in der Pufferzusammensetzung zu Wärmeänderungen führen können. Die Titration erfolgte mit 6 bis 8 µl Injektionsvolumen und die Zeit zwischen zwei Injektionen betrug 4 Minuten. Die Auswertung erfolgte mit dem Microcal Origin 2.9 Programm für ITC. 2.2.3.3 Differential Scanning Kalorimetrie Die Analyse der thermischen Entfaltung von RalGDS- und Raf-RBD WT sowie Mutanten wurde mit einem Microcal VP-DSC Mikrokalorimeter untersucht. Die Probenzelle enthielt zwischen 20 und 200 µM der zu untersuchenden RBD Lösung und die Messung erfolgte mit einer Heizrate von 1°C pro Minute. Die Messungen wurden mit dem Mikrocal Origin 4.1 Programm für DSC ausgewertet. Dabei wurde jeweils die Referenz-Schmelzkurve (Puffer gegen Puffer) von der Schmelzkurve der Proteinprobe abgezogen. 2.2.3.4 Phospor-NMR 31P-NMR Spektren wurden mit einem Bruker DMX-500 Spektrometer bei der Phosphor-Resonanzfrequenz von 202 MHz aufgenommen. Die Proteinproben wurden in Puffer mit 40 mM Hepes (pH 7.4), 5 mM MgCl2, 1 mM DTE, 150 mM NaCl, 5 % D2O und 0.1 mM DSS aufgenommen. Die Referenzierung wurde indirekt über das DSS Signal bestimmt. Hierzu wurde vor jeder 31P-Messung ein 1H-Spektrum aufgenommen und ein Ξ-Wert von

MATERIALIEN UND METHODEN
27
0.4048073561 eingesetzt, der 85 % iger Phosphorsäure in Glaskügelchen entspricht (Maurer & Kalbitzer, 1996). Die Messungen wurden in einem 10 mm Probenkopf in 8 mm Shigemi-Probenröhrchen durchgeführt. Die Volumina der Proben betrugen zwischen 0.7 und 1.2 ml bei Proteinkonzentrationen von 0.8 bis 1.6 mM. Bei der Datenaufnahme wurden die Protonen während der Datenaufnahme durch eine GARP Sequenz entkoppelt (Shaka et al., 1985). Es wurden 60 bis 70° Pulse mit einer Wiederholungszeit von 4 bis 7 Sekunden eingestrahlt. Die digitale Auflösung der Spektren war dabei besser als 0.4 Hz pro Datenpunkt.
2.2.4 Zellbiologische Methoden
2.2.4.1 Ral-Beladungs Assay COS-7 Zellen wurden bei 37 °C und 5.2 % CO2-Atmosphäre in Kunststoff-Kulturflaschen in DMEM versetzt mit 10 % FCS und 100 U/ml Penicillin G sowie 100 µg/ml Streptomycin kultiviert. Die Ablösung der Zellen bei der Passage erfolgte nach dem Waschen mit PBS durch Inkubation mit 1.5 ml Trypsin-Lösung. Auf diese Weise wurden die Zellen ca. alle vier Tage 1:10 verdünnt passagiert. Die Transfektion erfolgte 24 Stunden nach der Passage bei einer Konfluenz von 20-30 % in 6-cm Kulturschalen unter Verwendung der Superfect-Methode. Es wurden 5 µg DNA mit 125 µl DMEM ohne Antibiotika und Serum gemischt und 15 Minuten bei Raumtemperatur inkubiert. Danach wurde 1 ml DMEM mit Antibiotika und Serum zugefügt und die gesamte Lösung auf den – zuvor mit PBS gewaschenen - Zellen verteilt. Nach Inkubation von 3 Stunden im Brutschrank wurden die Zellen zweimal mit PBS gewaschen und mit frischem DMEM versorgt. Am darauf folgenden Tag wurden die Zellen zweimal mit DMEM ohne Phosphat und ohne Serum gewaschen und mit 0.3 mCi 32P-Orthophosphat in 1.5 ml DMEM ohne Phosphat und Serum für 5 Stunden bei 37 °C und 5.2 % C02 im Isotopenlabor inkubiert. Anschließend wurden die Zellen zweimal mit eiskaltem PBS gewaschen, auf Eis mit 700 µl Lysispuffer lysiert und mit einem Zellschaber in ein Eppendorf-Gefäß überführt. Nach Zentrifugation für 5 Minuten bei 4 °C mit 14000 rpm wurde der Überstand zu 10 µl Protein-G-Sepharose gegeben, an welche 4 µg 12CA5α Antikörper gekoppelt war. Mit einer HA-Markierung versehenes RalA wurde auf diese Weise innerhalb von 45 Minuten bei 4 °C immunpräzipitiert. Die Protein-G-Sepharose wurde danach fünfmal mit Lysispuffer, dreimal mit Waschpuffer und zweimal mit TM-Puffer gewaschen. Anschließend wurde das immunpräzipitierte RalA mit 35 µl Elutionspuffer inkubiert. Nach kurzer Zentrifugation wurden 10 µl des Überstandes auf PEI-Cellulose aufgetragen und das Dünnschichtchromatogramm in 0.8 M HCl mit 1.2 M Ammoniumformiat entwickelt. 32P-markiertes GDP/GTP wurde mit einem Phospho-Imager detektiert und densitometrisch quantifiziert. Dabei wurden die GDP- und GTP-Werte entsprechend ihres Phosphorgehaltes mit 1/2 bzw. 1/3 multipliziert. Aus diesen Werten konnte der Anteil des an RalA gebundenen GTP bestimmt werden.

MATERIALIEN UND METHODEN
28
2.2.4.2 Luziferase-Reportergen Assay Die Kultivierung der RK13-Zellen erfolgte in 50-ml-Gewebekulturflaschen mit 10 ml Kulturmedium (MEM, 10 % FCS, je 5000 U/ml Penicillin und Streptomycin) bei 37 °C und 7.5 % CO2. Die Passage der Zellen erfolgte ca. alle vier Tage. Dazu wurden die Zellen mit PBS gewaschen und mit 1.5 ml Trypsin-Lösung etwa fünf Minuten bei 37 °C inkubiert. Nach dem Ablösen der Zellen von der Unterlage wurden diese mit Medium aufgenommen und je nach Bedarf verdünnt oder ausplattiert. Die Durchführung des Luziferase-Reportergen-Assays erstreckt sich über vier Tage. Am ersten Tag wurden 385000 RK13-Zellen pro 6 cm-Petrischale mit 4 ml Medium ausplattiert. Am zweiten Tag wurden die Zellen bei einer 25 %-igen Konfluenz nach der Calzium-Phosphat Kopräzipitationsmethode (Sambrock et al., 1989) transfiziert. Jeder Transfektionsansatz (Doppelbestimmung) enthielt 80 ng pSVK3:RasG12V (oder Mutanten), 1.5 µg pCDNA3:Raf fl. (oder Mutanten), 1.5 µg p44MAPK, 1.5 µg pSGER81, 2 µg pE743-tk80-luc, 0.5 µg pEQ176, 20 µl CaCl2 (2.5 M). Dieser Ansatz wurde mit pCDNA3 auf 10 µg DNA und auf 200 µl Gesamtvolumen mit Wasser eingestellt. In 10-ml Plastikröhrchen wurde jeweils 200 µl HEBS-Puffer vorgelegt und unter ständigem Schütteln wurde der jeweilige Transfektionsansatz zugetropft, wodurch sich ein möglichst feines Kalziumphosphat-DNA-Präzipitat bildete. Das Gemisch wurde innerhalb von einer Minute auf die Zellen gegeben und unter leichtem Schwenken mit dem Medium vermischt. Nach einer Inkubation von sechs Stunden im Brutschrank wurden die Zellen dreimal mit PBS gewaschen und anschließend mit frischem Kulturmedium versetzt. Am vierten Tag wurden die transfizierten Zellen dreimal mit PBS gewaschen und mit 300 µl Lysispuffer versetzt. Die Zellen wurden abgeschabt, jeweils in ein 1.5-ml-Eppendorfgefäß überführt und für 10 Minuten bei 4 °C zentrifugiert. Der Überstand wurde in ein neues Eppendorfgefäß pipettiert. Zur Messung der Luziferase-Aktivität wurden jeweils 20 µl Zelllysat in 5-ml-Kunststoff-Röhrchen gegeben (Doppelbestimmung) und anschließend im Luminometer automatisch mit 100 µl Luziferase-Assay-Reagenz versetzt. Die Messung des entstandenen Lichtes erfolgte für 15 Sekunden und ist proportional zur Luziferase-Aktivität. Da die Luziferase-Aktivität von der Transfektionseffizienz abhängig ist, wurde diese mit Hilfe des ß-Galaktosidase-Aktivitätstest ermittelt. Dazu wurden je 20 µl Zelllysat auf eine Mikrotiterplatte gegeben (Doppelbestimmung) und anschließend zeitgleich (Multipipette) mit 200 µl Z-Puffer versetzt. Die Absorption bei 405 nm wurde nach leichter Gelbfärbung (ca. 5 bis 10 min) in einem ELISA-Photometer gemessen. Durch Division der Luziferaseaktivität durch die ß-Galaktosidaseaktivität wurde die normalisierte Luziferase-Aktivität erhalten. Die relative Luziferase-Aktivität wurde jeweils als Mehrfaches der Aktivität der pCDNA3-Leervektor-Kontrolle (auf 1 normiert) berechnet. Um verschiedene Versuche miteinander vergleichen zu können, wurde die Luziferase-Aktivität für die Transfektion des Raf WT mit RasG12V auf 100 % normiert.

ERGEBNISSE
29
III ERGEBNISSE Anhand der Untersuchungen von Ras und Effektor Proteinen soll zum einen das Verständnis von Protein-Wechselwirkungen im Allgemeinen vertieft werden. Zum anderen möchte man speziell an diesem System die Rolle von elektrostatischen Wechselwirkungen für die kinetischen und thermodynamischen Eigenschaften in vitro und auch für die Signalweiterleitung in vivo verstehen. Die Ras Proteine und die RBDs der Effektoren sind gut löslich, in relativ großen Mengen durch Produktion in E. coli Zellen zu erhalten und daher gut für biochemische Messungen geeignet. Zur Analyse der Signalweiterleitung in vivo gibt es mit dem Ral-Beladungs- und dem Luziferase-Reportergen-Assay zwei etablierte Systeme zur Quantifizierung der Ras/RalGDS- und der Ras/Raf-Wechselwirkungen in der Zelle.
3.1 Kinetische und thermodynamische Eigenschaften der
Wechselwirkung von Ras Proteinen mit Effektor-RBDs
Aus vielen biochemischen Untersuchungen der bisher untersuchten Ras/Effektor-RBD-Komplexe sind detaillierte kinetische und thermodynamische Parameter bekannt. Jedoch erfolgten diese Messungen unter variierenden Puffer- sowie Temperaturbedingungen. Zur besseren Übersicht und zum Vergleich ist es aber besser, alle Messungen unter denselben Bedingungen durchzuführen. Insbesondere für die Berechnungen der Assoziations-ratenkonstanten ist die exakte Bestimmung dieses Wertes für die Bindung der Wildtyp Proteine notwendig. Daher wurden zu Beginn dieser Arbeit die Wechselwirkungen von vier verschiedenen RBDs mit Ras Proteinen im Detail kinetisch und thermodynamisch analysiert. Die Messungen wurden zum einen in Hepes-Puffer ohne Zusatz von NaCl durchgeführt, um die elektrostatischen Wechselwirkungen in der Kontaktfläche nicht zu beeinflussen. Zum anderen erfolgten die Messungen in Hepes-Puffer mit zusätzlich 125 mM NaCl, welches einer physiologischen Ionenstärke entspricht. Messungen unter diesen Bedingungen sind wichtig, um Rückschlüsse und Vergleiche mit der Situation in vivo machen zu können.
3.1.1 Kinetische Eigenschaften von Ras/Effektor-RBD-Komplexen
Die Messung von Assoziations- und Dissoziationsratenkonstanten erfolgte mit der Stopped-Flow-Methode. Zur Messung enthielt eine Spritze Ras im Komplex mit GppNHp gebunden an eine mant-Gruppe als Fluoreszenzmarkierung (Ras⋅mGppNHp). Bei schnellem Mischen mit einer RBD-Lösung aus der anderen Spritze kommt es zur Assoziation der Proteine, was eine Fluoreszenzabnahme bewirkt. Dieses Verhalten wurde für die Bindung sämtlicher RBDs an Ras Proteine beobachtet. Unter Bedingungen pseudo-erster Ordnung, d.h., wenn die Effektor-RBD Konzentration mindestens 10-fach höher ist als die von Ras⋅mGppNHp, können die beobachteten zeitlichen Änderungen mit einer einfach exponentiellen Gleichung

ERGEBNISSE
30
RBD]/[K1
kkk1
22-obs
++=
angepasst werden. Diese Ratenkonstanten (kobs) werden zunächst mit steigender RBD Konzentration größer, während bei hohen Konzentrationen eine Sättigung zu beobachten ist (Abbildung 3.2). Diese Sättigungskinetik wurde für alle bisher analysierten Ras/Effektor-Komplexe gefunden (Sydor et al., 1998; Linnemann et al., 1999; Linnemann et al., 2001) und zeigt, dass die Bindung einem Mechanismus mit mindestens zwei Schritten folgt (Abbildung 3.1). ABBILDUNG 3.1: Zweistufen-Mechanismus der Bindung von Effektor -RBDs an Ras Proteine. Die Assoziation von Ras und RBD verläuft über ein Zwischenprodukt [Ras:RBD] mit niedriger Affinität (K1) („Encounter Komplex“). Der Endkomplex Ras:RBD wird mit der Ratenkonstante k2 + k-2 gebildet.
Dabei kommt es zuerst zur Bildung eines Komplexes mit niedriger Affinität („Encounter Komplex“) mit der Dissoziationskonstante K1 = k-1/k1. Anschließend wird durch strukturelle Umlagerungen der hoch affine Endkomplex gebildet. Basierend auf diesem Zweistufen-Mechanismus kann die Ratenkonstante (kobs) der Fluoreszenzänderung folgendermaßen beschrieben werden: (3.1)
Eine Auftragung von kobs gegen die RalGDS-RBD Konzentration wie in Abbildung 3.2 mit nachfolgender Anpassung der Daten entsprechend Gleichung 3.1 zeigt, dass die Affinität des vorgelagerten Gleichgewichtes mit 134 µM sehr niedrig ist. Der Wert für k2 wurde zu 459 s-1 bestimmt. Daraus wird eine Assoziationsratenkonstante nach der Beziehung kon = k2/K1 von 3.4 µM -1s-1 berechnet.
ABBILDUNG 3.2: Assoziationskinetik der Bindung von Ral GDS-RBDWT an Ras⋅mGppNHp. Auftragung der Ratenkonstanten der Fluoreszenzänderungen (kobs) gegen die RalGDS-RBD Konzentration. Die Messungen erfolgten bei 10 °C in 15 mM Hepes (pH 7.4) und 5 mM MgCl2. Die Daten wurden nach Gleichung 3.1 angepasst, was folgende Werte liefert: K1 = 134 µM, k2 = 459 s-1. Daraus berechnet sich die Assoziationsratenkonstante mit kon = k2/K1 zu 3.4 µM-1s -1.
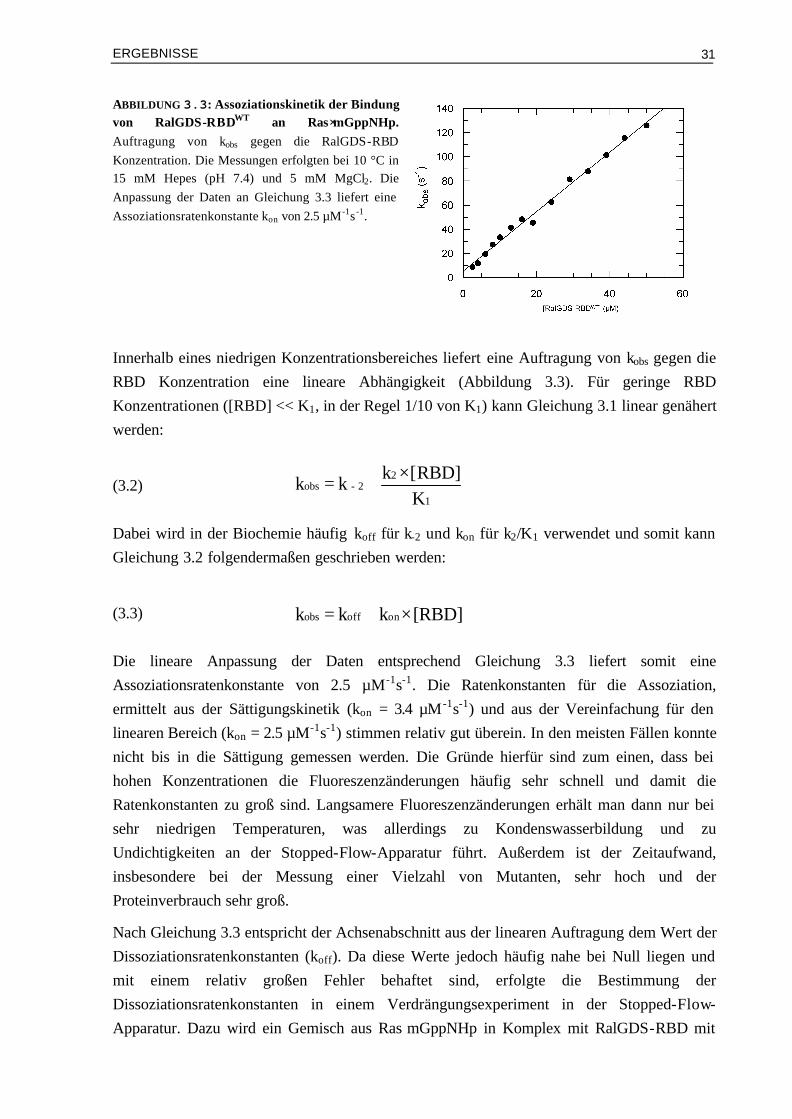
ERGEBNISSE
31
KRBD][kkk
1
22-obs
×+=
RBD][kkk onoffobs ×+=
ABBILDUNG 3.3: Assoziationskinetik der Bindung von RalGDS-RBDWT an Ras⋅mGppNHp. Auftragung von kobs gegen die RalGDS-RBD Konzentration. Die Messungen erfolgten bei 10 °C in 15 mM Hepes (pH 7.4) und 5 mM MgCl2. Die Anpassung der Daten an Gleichung 3.3 liefert eine Assoziationsratenkonstante kon von 2.5 µM-1s -1.
Innerhalb eines niedrigen Konzentrationsbereiches liefert eine Auftragung von kobs gegen die RBD Konzentration eine lineare Abhängigkeit (Abbildung 3.3). Für geringe RBD Konzentrationen ([RBD] << K1, in der Regel 1/10 von K1) kann Gleichung 3.1 linear genähert werden:
(3.2) Dabei wird in der Biochemie häufig koff für k-2 und kon für k2/K1 verwendet und somit kann Gleichung 3.2 folgendermaßen geschrieben werden:
(3.3) Die lineare Anpassung der Daten entsprechend Gleichung 3.3 liefert somit eine Assoziationsratenkonstante von 2.5 µM-1s-1. Die Ratenkonstanten für die Assoziation, ermittelt aus der Sättigungskinetik (kon = 3.4 µM -1s-1) und aus der Vereinfachung für den linearen Bereich (kon = 2.5 µM-1s-1) stimmen relativ gut überein. In den meisten Fällen konnte nicht bis in die Sättigung gemessen werden. Die Gründe hierfür sind zum einen, dass bei hohen Konzentrationen die Fluoreszenzänderungen häufig sehr schnell und damit die Ratenkonstanten zu groß sind. Langsamere Fluoreszenzänderungen erhält man dann nur bei sehr niedrigen Temperaturen, was allerdings zu Kondenswasserbildung und zu Undichtigkeiten an der Stopped-Flow-Apparatur führt. Außerdem ist der Zeitaufwand, insbesondere bei der Messung einer Vielzahl von Mutanten, sehr hoch und der Proteinverbrauch sehr groß.
Nach Gleichung 3.3 entspricht der Achsenabschnitt aus der linearen Auftragung dem Wert der Dissoziationsratenkonstanten (koff). Da diese Werte jedoch häufig nahe bei Null liegen und mit einem relativ großen Fehler behaftet sind, erfolgte die Bestimmung der Dissoziationsratenkonstanten in einem Verdrängungsexperiment in der Stopped-Flow-Apparatur. Dazu wird ein Gemisch aus Ras⋅mGppNHp in Komplex mit RalGDS-RBD mit

ERGEBNISSE
32
kkK
on
offd =
RTlnKG a−=°∆
K1
Kd
a =
einem Überschuss an Ras⋅GppNHp ohne Fluoreszenzmarkierung gemischt. Dies führt zu einem Anstieg der Fluoreszenzänderung, da Ras⋅mGppNHp aus dem Komplex mit RalGDS-RBD durch Ras⋅GppNHp verdrängt wird. Da die Dissoziationsreaktion einer Kinetik erster Ordnung folgt, kann die Fluoreszenzzunahme ebenfalls mit einer einfach exponentiellen Gleichung angepasst werden und die Ratenkonstante entspricht der Dissoziationsraten-konstanten koff der Reaktion. Für die Dissoziationskonstante Kd, als Maß für die Affinität des Komplexes gilt die folgende Beziehung: (3.4) Da meistens jedoch die Komplexbildung behandelt wird und zum besseren Vergleich mit kalorimetrischen Daten, wird z. T. auch die Assoziationskonstante Ka verwendet: (3.5) Für die Änderung der freien Enthalpie, oder Gibbs-Energie, ∆G° gilt dann Gleichung 3.6, mit R als Gaskonstante und T der Temperatur in Kelvin. (3.6)
In Tabelle 3.1 sind die Ergebnisse der Stopped-Flow-Messungen für die Bindung von vier verschiedenen RBDs an Ras⋅mGppNHp, Raps⋅mGppNHp und Rap⋅mGppNHp gemessen bei 25 °C in Puffer mit niedriger Ionenstärke (15 mM Hepes, 5 mM MgCl2) und in Puffer mit physiologischer Ionenstärke (15 mM Hepes, 5 mM MgCl2, 125 mM NaCl) dargestellt. Für die Bindung der RBDs an Ras⋅mGppNHp in Puffer mit niedriger Ionenstärke ist zu erkennen, dass die Unterschiede in den Affinitäten hauptsächlich eine Folge von Änderungen der Assoziationsratenkonstanten sind, während die Dissoziationsratenkonstanten in einem ähnlichen Bereich liegen. Besonders deutlich unterschieden sich die Assoziationsratenkonstanten von RalGDS- und Raf-RBD, was anhand der Elektrostatik der Kontaktflächen erklärt werden kann. Während die Oberflächen von Ras und Rap im Effektor Bindungsbereich überwiegend negative Ladungen enthalten, sind die Oberflächen der RBDs von RalGDS und Raf hauptsächlich positiv geladen (Nassar et al., 1996). Insbesondere zwischen der Kontaktfläche von Raf-RBD und Ras besteht eine sehr starke elektrostatische Komplementarität, was den sehr hohen Wert für kon als Folge von elektrostatischer Lenkung erklären kann (siehe Kapitel 1.6). Im Gegensatz dazu ist die Kontaktfläche zwischen RalGDS-RBD und Ras nicht so stark elektrostatisch komplementär geladen, was eine mögliche Erklärung für den niedrigeren kon-Wert sein könnte.
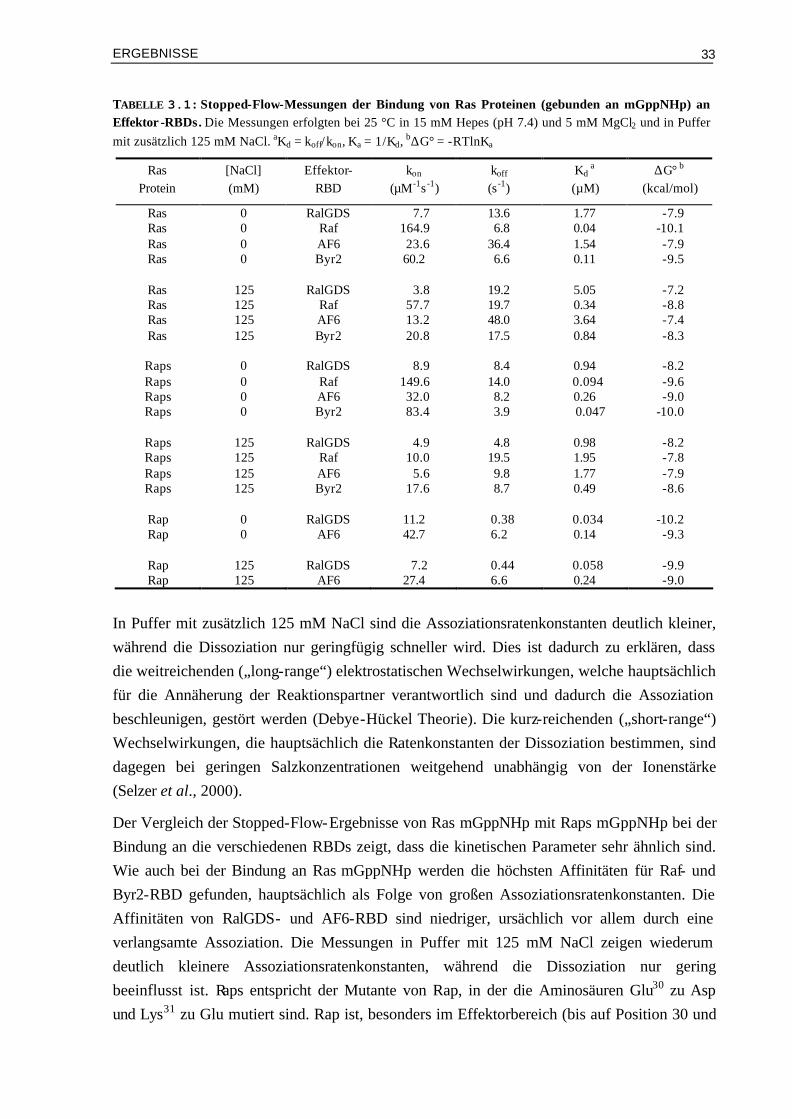
ERGEBNISSE
33
TABELLE 3.1: Stopped-Flow-Messungen der Bindung von Ras Proteinen (gebunden an mGppNHp) an Effektor -RBDs. Die Messungen erfolgten bei 25 °C in 15 mM Hepes (pH 7.4) und 5 mM MgCl2 und in Puffer mit zusätzlich 125 mM NaCl. aKd = koff/kon, Ka = 1/Kd, b∆G° = -RTlnKa
In Puffer mit zusätzlich 125 mM NaCl sind die Assoziationsratenkonstanten deutlich kleiner, während die Dissoziation nur geringfügig schneller wird. Dies ist dadurch zu erklären, dass die weitreichenden („long-range“) elektrostatischen Wechselwirkungen, welche hauptsächlich für die Annäherung der Reaktionspartner verantwortlich sind und dadurch die Assoziation beschleunigen, gestört werden (Debye-Hückel Theorie). Die kurz-reichenden („short-range“) Wechselwirkungen, die hauptsächlich die Ratenkonstanten der Dissoziation bestimmen, sind dagegen bei geringen Salzkonzentrationen weitgehend unabhängig von der Ionenstärke (Selzer et al., 2000).
Der Vergleich der Stopped-Flow-Ergebnisse von Ras⋅mGppNHp mit Raps⋅mGppNHp bei der Bindung an die verschiedenen RBDs zeigt, dass die kinetischen Parameter sehr ähnlich sind. Wie auch bei der Bindung an Ras⋅mGppNHp werden die höchsten Affinitäten für Raf- und Byr2-RBD gefunden, hauptsächlich als Folge von großen Assoziationsratenkonstanten. Die Affinitäten von RalGDS- und AF6-RBD sind niedriger, ursächlich vor allem durch eine verlangsamte Assoziation. Die Messungen in Puffer mit 125 mM NaCl zeigen wiederum deutlich kleinere Assoziationsratenkonstanten, während die Dissoziation nur gering beeinflusst ist. Raps entspricht der Mutante von Rap, in der die Aminosäuren Glu30 zu Asp und Lys31 zu Glu mutiert sind. Rap ist, besonders im Effektorbereich (bis auf Position 30 und
Ras Protein
[NaCl] (mM)
Effektor- RBD
kon (µM-1s -1)
koff (s-1)
Kd a
(µM) ∆G° b
(kcal/mol)
Ras 0 RalGDS 7.7 13.6 1.77 -7.9 Ras 0 Raf 164.9 6.8 0.04 -10.1 Ras 0 AF6 23.6 36.4 1.54 -7.9 Ras 0 Byr2 60.2 6.6 0.11 -9.5
Ras 125 RalGDS 3.8 19.2 5.05 -7.2 Ras 125 Raf 57.7 19.7 0.34 -8.8 Ras 125 AF6 13.2 48.0 3.64 -7.4 Ras 125 Byr2 20.8 17.5 0.84 -8.3
Raps 0 RalGDS 8.9 8.4 0.94 -8.2 Raps 0 Raf 149.6 14.0 0.094 -9.6 Raps 0 AF6 32.0 8.2 0.26 -9.0 Raps 0 Byr2 83.4 3.9 0.047 -10.0
Raps 125 RalGDS 4.9 4.8 0.98 -8.2 Raps 125 Raf 10.0 19.5 1.95 -7.8 Raps 125 AF6 5.6 9.8 1.77 -7.9 Raps 125 Byr2 17.6 8.7 0.49 -8.6
Rap 0 RalGDS 11.2 0.38 0.034 -10.2 Rap 0 AF6 42.7 6.2 0.14 -9.3
Rap 125 RalGDS 7.2 0.44 0.058 -9.9 Rap 125 AF6 27.4 6.6 0.24 -9.0

ERGEBNISSE
34
31) sehr homolog zu Ras und daher führt die Mutation an Position 30 und 31 von Rap zu den entsprechenden Aminosäuren in Ras dazu, dass die biochemischen Eigenschaften von Raps (Der Name wird hergeleitet aus Rap zu Ras) und Ras sehr ähnlich sind. Der Vergleich von biochemischen Messungen von Raf-RBD/Ras Komplexen mit Strukturdaten erfolgt vor allem mit der Struktur von Raps in Komplex mit Raf-RBD (Nassar et al, 1995), da bisher keine Komplexstruktur von Raf-RBD und Ras gelöst werden konnte.
Für die Komplexe von RBDs und Rap⋅mGppNHp konnten nur für RalGDS- und AF6-RBD kinetische Parameter ermittelt werden, da die Bindungen von Raf- und Byr2-RBD an Rap⋅mGppNHp unter keinen Bedingungen (bei 10, 15 und 25 °C und in Puffer mit und ohne NaCl) zu einer Änderung des Fluoreszenzsignals führten. Die Affinität der Bindung von RalGDS-RBD an Rap⋅mGppNHp ist wesentlich höher als an Ras⋅mGppNHp. Interessanterweise ist dies nicht die Folge einer größeren Assoziationsratenkonstante (diese liegt in derselben Größenordung wie bei der Bindung an Ras⋅mGppNHp), sondern hier ist die Dissoziation extrem langsam. Im Gegensatz dazu sind die Affinität sowie die Ratenkonstanten von Assoziation und Dissoziation für die Bindung von AF6-RBD an Rap⋅mGppNHp in derselben Größenordnung wie bei der Bindung an Ras⋅mGppNHp.
Um den Effekt von höheren Salzkonzentrationen auf kon und koff zu analysieren, wurden Stopped-Flow-Messungen für die Bindung von Raf-RBD an Ras⋅mGppNHp in Puffer mit Salzkonzentrationen von 0 bis 1000 mM NaCl durchgeführt (Tabelle 3.2 und Abbildung 3.4).
TABELLE 3.2: Stopped-Flow-Messungen der Bindung von Ras ⋅mGppNHp an Raf-RBD in Puffer mit steigenden Salzkonzentrationen. Die Messungen erfolgten bei 25 °C in 50 mM Tris (pH 7.4) und 5 mM MgCl2 mit steigenden Konzentrationen an NaCl. aKd = koff/kon, Ka = 1/Kd, b∆G° = -RTlnKa
In Abbildung 3.4 sind die Werte für kon, koff und Kd – nach steigender Assoziationsratenkonstante geordnet – für die Bindung von Raf-RBD an Ras⋅mGppNHp dargestellt. Da die drei ermittelten Parameter unterschiedliche Einheiten haben, und ihre Zahlenwerte in unterschiedlichen Größenordnungen liegen, wurden in diesem und in den folgenden Diagrammen dieser Art relative Werte dargestellt. D.h. kon, koff und Kd wurden jeweils auf einen Bezugswert normiert, so dass die Ergebnisse sinnvoll erkennbar in einem Diagramm dargestellt werden können. Die absoluten Werte sind in den entsprechenden Tabellen zu finden.
[NaCl] (mM)
kon (µM-1s -1)
koff (s-1)
Kd a
(µM) ∆G° b
(kcal/mol)
0 60.0 4.4 0.073 -9.7 100 24.0 5.7 0.24 -9.0 200 17.3 7.7 0.44 -8.7 500 9.5 12.4 1.3 -8.0 800 7.4 18.9 2.6 -7.6 1000 3.0 27.9 9.2 -6.9
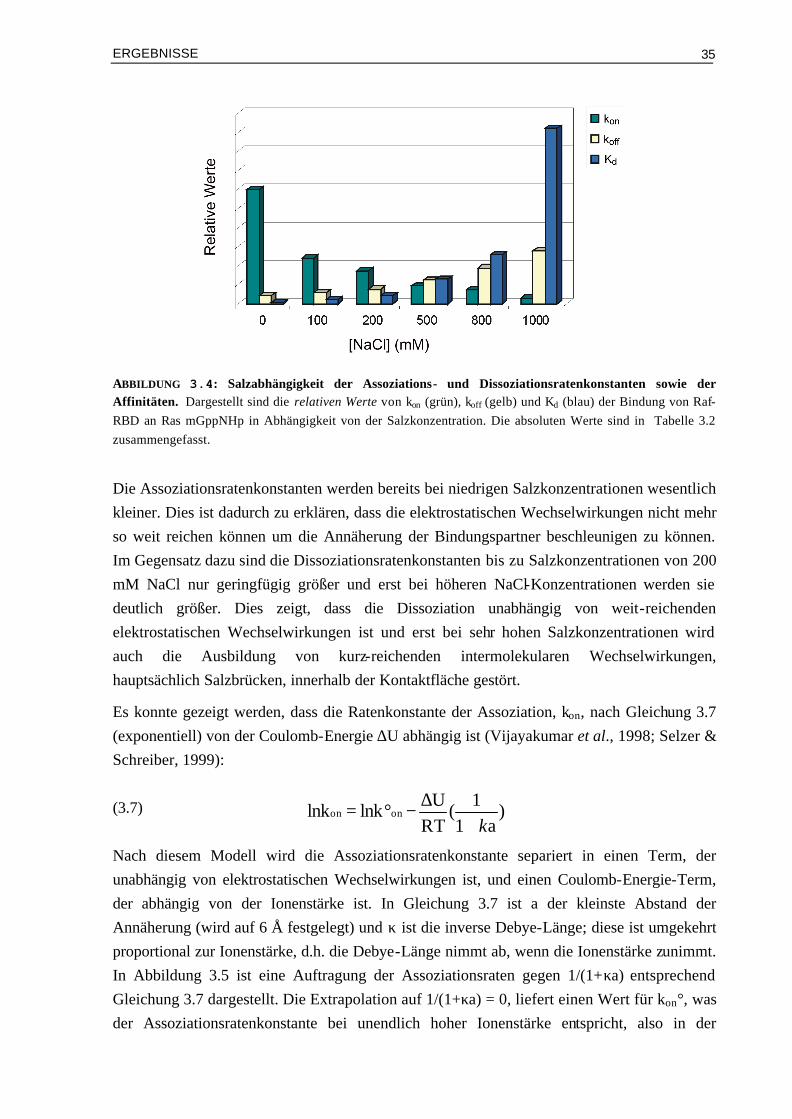
ERGEBNISSE
35
)a1
1(RT
Ulnklnk ononκ+
∆−°=
ABBILDUNG 3.4: Salzabhängigkeit der Assoziations- und Dissoziationsratenkonstanten sowie der Affinitäten. Dargestellt sind die relativen Werte von kon (grün), koff (gelb) und Kd (blau) der Bindung von Raf-RBD an Ras⋅mGppNHp in Abhängigkeit von der Salzkonzentration. Die absoluten Werte sind in Tabelle 3.2 zusammengefasst.
Die Assoziationsratenkonstanten werden bereits bei niedrigen Salzkonzentrationen wesentlich kleiner. Dies ist dadurch zu erklären, dass die elektrostatischen Wechselwirkungen nicht mehr so weit reichen können um die Annäherung der Bindungspartner beschleunigen zu können. Im Gegensatz dazu sind die Dissoziationsratenkonstanten bis zu Salzkonzentrationen von 200 mM NaCl nur geringfügig größer und erst bei höheren NaCl-Konzentrationen werden sie deutlich größer. Dies zeigt, dass die Dissoziation unabhängig von weit-reichenden elektrostatischen Wechselwirkungen ist und erst bei sehr hohen Salzkonzentrationen wird auch die Ausbildung von kurz-reichenden intermolekularen Wechselwirkungen, hauptsächlich Salzbrücken, innerhalb der Kontaktfläche gestört.
Es konnte gezeigt werden, dass die Ratenkonstante der Assoziation, kon, nach Gleichung 3.7 (exponentiell) von der Coulomb-Energie ∆U abhängig ist (Vijayakumar et al., 1998; Selzer & Schreiber, 1999): (3.7) Nach diesem Modell wird die Assoziationsratenkonstante separiert in einen Term, der unabhängig von elektrostatischen Wechselwirkungen ist, und einen Coulomb-Energie-Term, der abhängig von der Ionenstärke ist. In Gleichung 3.7 ist a der kleinste Abstand der Annäherung (wird auf 6 Å festgelegt) und κ ist die inverse Debye-Länge; diese ist umgekehrt proportional zur Ionenstärke, d.h. die Debye-Länge nimmt ab, wenn die Ionenstärke zunimmt. In Abbildung 3.5 ist eine Auftragung der Assoziationsraten gegen 1/(1+κa) entsprechend Gleichung 3.7 dargestellt. Die Extrapolation auf 1/(1+κa) = 0, liefert einen Wert für kon°, was der Assoziationsratenkonstante bei unendlich hoher Ionenstärke entspricht, also in der

ERGEBNISSE
36
Abwesenheit von elektrostatischen Wechselwirkungen. Für diese sog. Basalratenkonstante der Assoziation (kon°) wurde für die Bindung von Raf-RBD an Ras⋅mGppNHp ein Wert von 4.2*105 M-1s-1 ermittelt (Abbildung 3.5). Dieser Wert liegt in derselben Größenordnung wie für viele andere Protein Komplexe gefunden wurde (Selzer & Schreiber, 1999; Selzer et al., 2000).
ABBILDUNG 3.5: Extrapolation der Assoziationsratenkonstante bei unendlich hoher Ionenstärke. Auftragung von kon gegen 1/(1+κa) für die Bindung von Raf-RBD an Ras⋅mGppNHp entsprechend Tabelle 3.2. Die Extrapolation bei 1/(1+κa) = 0 liefert nach Gleichung 3.7 den Wert für die Assoziationsratenkonstante in Abwesenheit von elektrostatischen Wechselwirkungen von kon° = 4.2*105 M-1s -1.
3.1.2 Thermodynamik der Bindung von Ras Proteinen an Effektor-RBDs
Die Analyse der Komplexbildung zwischen Ras Proteinen und Effektor-RBDs erfolgte durch die Isotherme Titrationskalorimetrie (ITC). Dies ist eine Methode zur vollständigen thermodynamischen Charakterisierung einer Reaktion (Wisemann et al., 1989, Ladbury & Chowdry, 1996), d.h. man erhält neben den Assoziationskonstanten (Ka), als Maß für die Affinität der Bindung, auch Informationen über die Enthalpie- und Entropieänderungen (∆H° und T∆S°) bei der Komplexbildung. In einem ITC-Experiment wird die Wärmeenergie der Assoziation eines Proteins direkt mit einem dazu titrierten Liganden gemessen. Das ITC-Gerät enthält zwei identische Zellen (Probenzelle und Referenzzelle) umgeben von einem thermostatisierten Mantel. Dieser wird gekühlt, so dass Wärmeenergie nötig ist, um die beiden Zellen auf konstanter Temperatur zu halten. Die Referenzzelle ist mit Puffer gefüllt und die Probenzelle enthält die eine Komponente der zu messenden Reaktion. Wird nun die zweite Komponente innerhalb einer Serie von mehreren Injektionen der Probenzelle zugeführt, kommt es zu einer Enthalpieänderung. Gemessen wird die Heizleistung, die der Probenzelle zugeführt werden muss, damit sie immer die gleiche (auf ca. 0.001 °C genaue) Temperaturdifferenz zur Referenzzelle aufweist. In Abbildung 3.6 ist ein typisches ITC-Experiment der Assoziation von RalGDS-RBD an Ras⋅GppNHp dargestellt. In diesem Experiment wurden 900 µM Ras⋅GppNHp aus der Spritze zu 45 µM RalGDS-RBD in der Probenzelle in einer Serie von 19 Injektionen titriert. Im oberen Teil von Abbildung 3.6 ist eine Auftragung der Heizleistung gegen die Zeit aufgetragen.
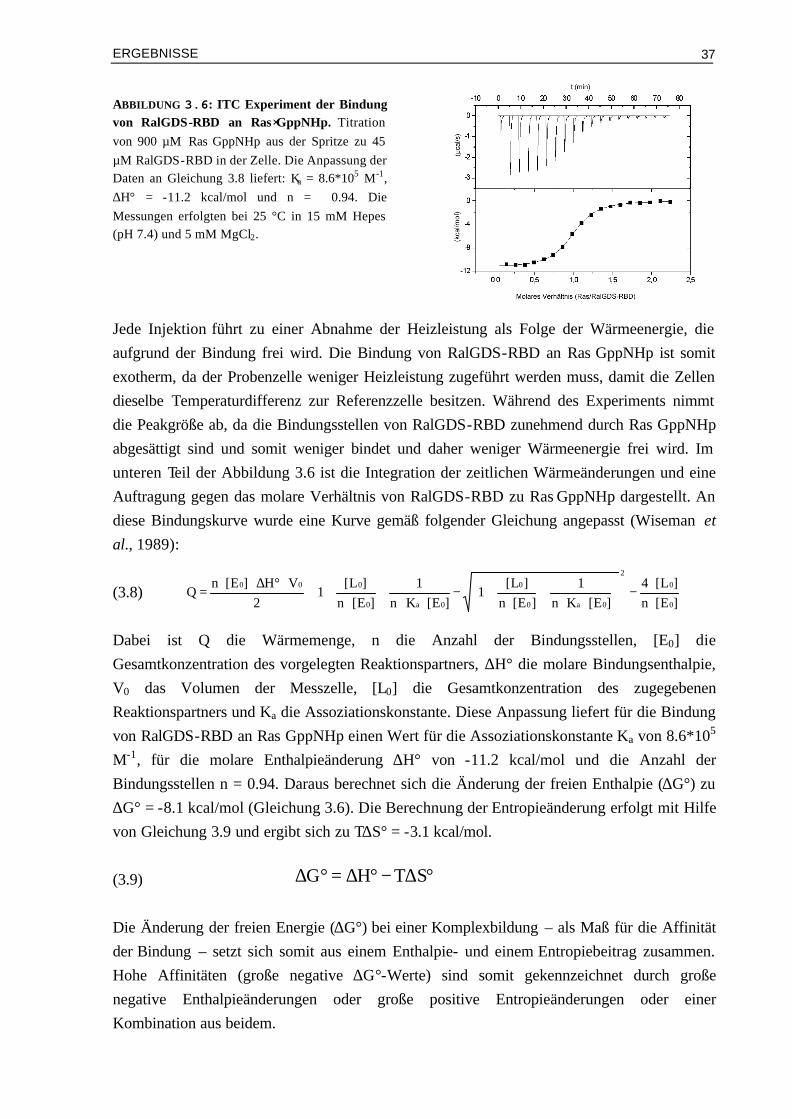
ERGEBNISSE
37
STHG °∆−°∆=°∆
]E[n]L[4
]E[Kn1
]E[n]L[
1]E[Kn
1]E[n
]L[1
2VH]E[n
Q0
02
0a0
0
0a0
000
⋅⋅
−
⋅⋅
+⋅
+−⋅⋅
+⋅
+⋅⋅°∆⋅⋅
=
ABBILDUNG 3.6: ITC Experiment der Bindung von RalGDS-RBD an Ras⋅GppNHp. Titration von 900 µM Ras⋅GppNHp aus der Spritze zu 45 µM RalGDS-RBD in der Zelle. Die Anpassung der Daten an Gleichung 3.8 liefert: Ka = 8.6*105 M-1, ∆H° = -11.2 kcal/mol und n = 0.94. Die Messungen erfolgten bei 25 °C in 15 mM Hepes (pH 7.4) und 5 mM MgCl2. Jede Injektion führt zu einer Abnahme der Heizleistung als Folge der Wärmeenergie, die aufgrund der Bindung frei wird. Die Bindung von RalGDS-RBD an Ras⋅GppNHp ist somit exotherm, da der Probenzelle weniger Heizleistung zugeführt werden muss, damit die Zellen dieselbe Temperaturdifferenz zur Referenzzelle besitzen. Während des Experiments nimmt die Peakgröße ab, da die Bindungsstellen von RalGDS-RBD zunehmend durch Ras⋅GppNHp abgesättigt sind und somit weniger bindet und daher weniger Wärmeenergie frei wird. Im unteren Teil der Abbildung 3.6 ist die Integration der zeitlichen Wärmeänderungen und eine Auftragung gegen das molare Verhältnis von RalGDS-RBD zu Ras⋅GppNHp dargestellt. An diese Bindungskurve wurde eine Kurve gemäß folgender Gleichung angepasst (Wiseman et al., 1989): (3.8) Dabei ist Q die Wärmemenge, n die Anzahl der Bindungsstellen, [E0] die Gesamtkonzentration des vorgelegten Reaktionspartners, ∆H° die molare Bindungsenthalpie, V0 das Volumen der Messzelle, [L0] die Gesamtkonzentration des zugegebenen Reaktionspartners und Ka die Assoziationskonstante. Diese Anpassung liefert für die Bindung von RalGDS-RBD an Ras⋅GppNHp einen Wert für die Assoziationskonstante Ka von 8.6*105 M-1, für die molare Enthalpieänderung ∆H° von -11.2 kcal/mol und die Anzahl der Bindungsstellen n = 0.94. Daraus berechnet sich die Änderung der freien Enthalpie (∆G°) zu ∆G° = -8.1 kcal/mol (Gleichung 3.6). Die Berechnung der Entropieänderung erfolgt mit Hilfe von Gleichung 3.9 und ergibt sich zu T∆S° = -3.1 kcal/mol. (3.9) Die Änderung der freien Energie (∆G°) bei einer Komplexbildung – als Maß für die Affinität der Bindung – setzt sich somit aus einem Enthalpie- und einem Entropiebeitrag zusammen. Hohe Affinitäten (große negative ∆G°-Werte) sind somit gekennzeichnet durch große negative Enthalpieänderungen oder große positive Entropieänderungen oder einer Kombination aus beidem.
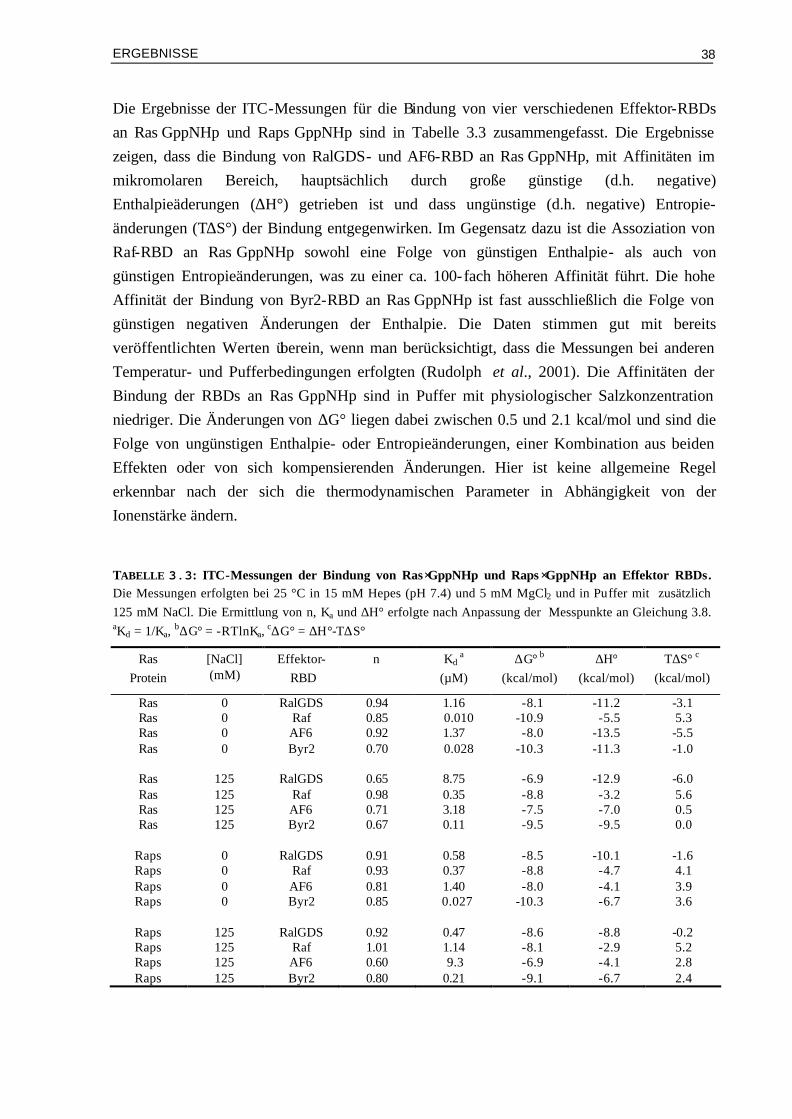
ERGEBNISSE
38
Die Ergebnisse der ITC-Messungen für die Bindung von vier verschiedenen Effektor-RBDs an Ras⋅GppNHp und Raps⋅GppNHp sind in Tabelle 3.3 zusammengefasst. Die Ergebnisse zeigen, dass die Bindung von RalGDS- und AF6-RBD an Ras⋅GppNHp, mit Affinitäten im mikromolaren Bereich, hauptsächlich durch große günstige (d.h. negative) Enthalpieäderungen (∆H°) getrieben ist und dass ungünstige (d.h. negative) Entropie-änderungen (T∆S°) der Bindung entgegenwirken. Im Gegensatz dazu ist die Assoziation von Raf-RBD an Ras⋅GppNHp sowohl eine Folge von günstigen Enthalpie- als auch von günstigen Entropieänderungen, was zu einer ca. 100-fach höheren Affinität führt. Die hohe Affinität der Bindung von Byr2-RBD an Ras⋅GppNHp ist fast ausschließlich die Folge von günstigen negativen Änderungen der Enthalpie. Die Daten stimmen gut mit bereits veröffentlichten Werten überein, wenn man berücksichtigt, dass die Messungen bei anderen Temperatur- und Pufferbedingungen erfolgten (Rudolph et al., 2001). Die Affinitäten der Bindung der RBDs an Ras⋅GppNHp sind in Puffer mit physiologischer Salzkonzentration niedriger. Die Änderungen von ∆G° liegen dabei zwischen 0.5 und 2.1 kcal/mol und sind die Folge von ungünstigen Enthalpie- oder Entropieänderungen, einer Kombination aus beiden Effekten oder von sich kompensierenden Änderungen. Hier ist keine allgemeine Regel erkennbar nach der sich die thermodynamischen Parameter in Abhängigkeit von der Ionenstärke ändern.
TABELLE 3.3: ITC-Messungen der Bindung von Ras ⋅GppNHp und Raps ⋅ GppNHp an Effektor RBDs. Die Messungen erfolgten bei 25 °C in 15 mM Hepes (pH 7.4) und 5 mM MgCl2 und in Puffer mit zusätzlich 125 mM NaCl. Die Ermittlung von n, Ka und ∆H° erfolgte nach Anpassung der Messpunkte an Gleichung 3.8. aKd = 1/Ka,
b∆G° = -RTlnKa, c∆G° = ∆H°-T∆S°
Ras Protein
[NaCl] (mM)
Effektor- RBD
n Kd a
(µM) ∆G° b
(kcal/mol) ∆H°
(kcal/mol) T∆S° c
(kcal/mol)
Ras 0 RalGDS 0.94 1.16 -8.1 -11.2 -3.1 Ras 0 Raf 0.85 0.010 -10.9 -5.5 5.3 Ras 0 AF6 0.92 1.37 -8.0 -13.5 -5.5 Ras 0 Byr2 0.70 0.028 -10.3 -11.3 -1.0
Ras 125 RalGDS 0.65 8.75 -6.9 -12.9 -6.0 Ras 125 Raf 0.98 0.35 -8.8 -3.2 5.6 Ras 125 AF6 0.71 3.18 -7.5 -7.0 0.5 Ras 125 Byr2 0.67 0.11 -9.5 -9.5 0.0
Raps 0 RalGDS 0.91 0.58 -8.5 -10.1 -1.6 Raps 0 Raf 0.93 0.37 -8.8 -4.7 4.1 Raps 0 AF6 0.81 1.40 -8.0 -4.1 3.9 Raps 0 Byr2 0.85 0.027 -10.3 -6.7 3.6
Raps 125 RalGDS 0.92 0.47 -8.6 -8.8 -0.2 Raps 125 Raf 1.01 1.14 -8.1 -2.9 5.2 Raps 125 AF6 0.60 9.3 -6.9 -4.1 2.8 Raps 125 Byr2 0.80 0.21 -9.1 -6.7 2.4
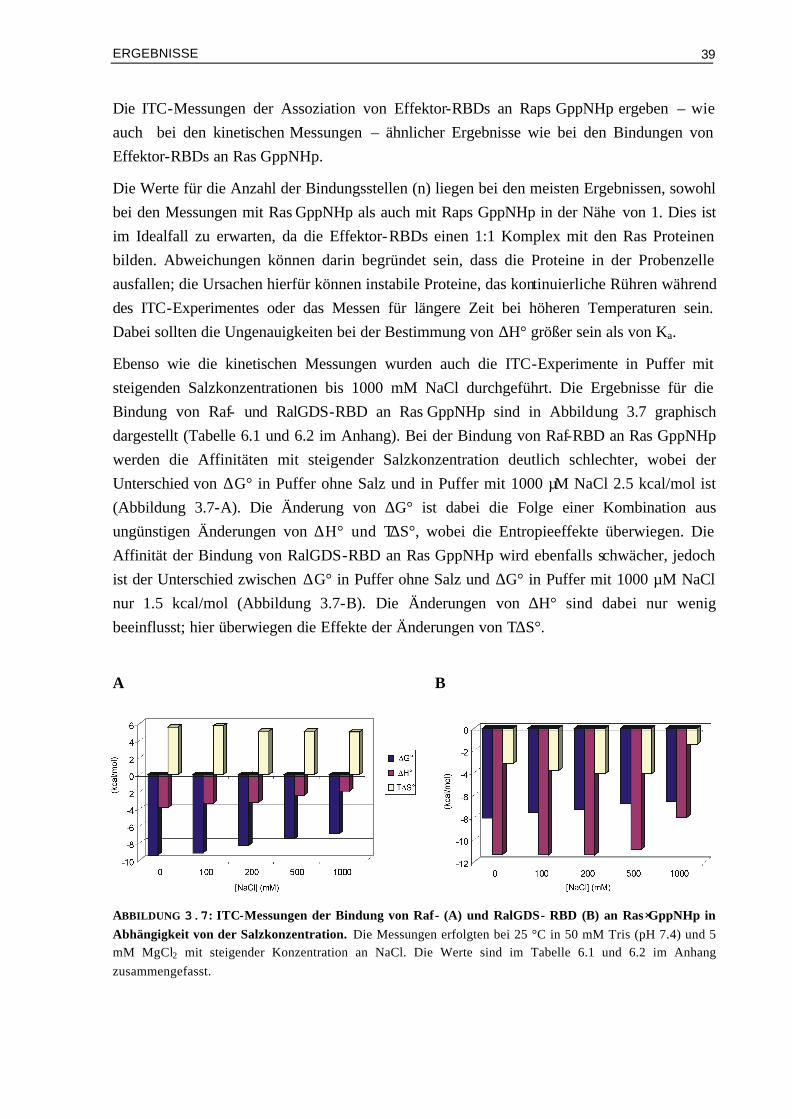
ERGEBNISSE
39
Die ITC-Messungen der Assoziation von Effektor-RBDs an Raps⋅GppNHp ergeben – wie auch bei den kinetischen Messungen – ähnlicher Ergebnisse wie bei den Bindungen von Effektor-RBDs an Ras⋅GppNHp.
Die Werte für die Anzahl der Bindungsstellen (n) liegen bei den meisten Ergebnissen, sowohl bei den Messungen mit Ras⋅GppNHp als auch mit Raps⋅GppNHp in der Nähe von 1. Dies ist im Idealfall zu erwarten, da die Effektor-RBDs einen 1:1 Komplex mit den Ras Proteinen bilden. Abweichungen können darin begründet sein, dass die Proteine in der Probenzelle ausfallen; die Ursachen hierfür können instabile Proteine, das kontinuierliche Rühren während des ITC-Experimentes oder das Messen für längere Zeit bei höheren Temperaturen sein. Dabei sollten die Ungenauigkeiten bei der Bestimmung von ∆H° größer sein als von Ka.
Ebenso wie die kinetischen Messungen wurden auch die ITC-Experimente in Puffer mit steigenden Salzkonzentrationen bis 1000 mM NaCl durchgeführt. Die Ergebnisse für die Bindung von Raf- und RalGDS-RBD an Ras⋅GppNHp sind in Abbildung 3.7 graphisch dargestellt (Tabelle 6.1 und 6.2 im Anhang). Bei der Bindung von Raf-RBD an Ras⋅GppNHp werden die Affinitäten mit steigender Salzkonzentration deutlich schlechter, wobei der Unterschied von ∆G° in Puffer ohne Salz und in Puffer mit 1000 µM NaCl 2.5 kcal/mol ist (Abbildung 3.7-A). Die Änderung von ∆G° ist dabei die Folge einer Kombination aus ungünstigen Änderungen von ∆H° und T∆S°, wobei die Entropieeffekte überwiegen. Die Affinität der Bindung von RalGDS-RBD an Ras⋅GppNHp wird ebenfalls schwächer, jedoch ist der Unterschied zwischen ∆G° in Puffer ohne Salz und ∆G° in Puffer mit 1000 µM NaCl nur 1.5 kcal/mol (Abbildung 3.7-B). Die Änderungen von ∆H° sind dabei nur wenig beeinflusst; hier überwiegen die Effekte der Änderungen von T∆S°. A B
ABBILDUNG 3.7: ITC-Messungen der Bindung von Raf- (A) und RalGDS- RBD (B) an Ras⋅GppNHp in Abhängigkeit von der Salzkonzentration. Die Messungen erfolgten bei 25 °C in 50 mM Tris (pH 7.4) und 5 mM MgCl2 mit steigender Konzentration an NaCl. Die Werte sind im Tabelle 6.1 und 6.2 im Anhang zusammengefasst.
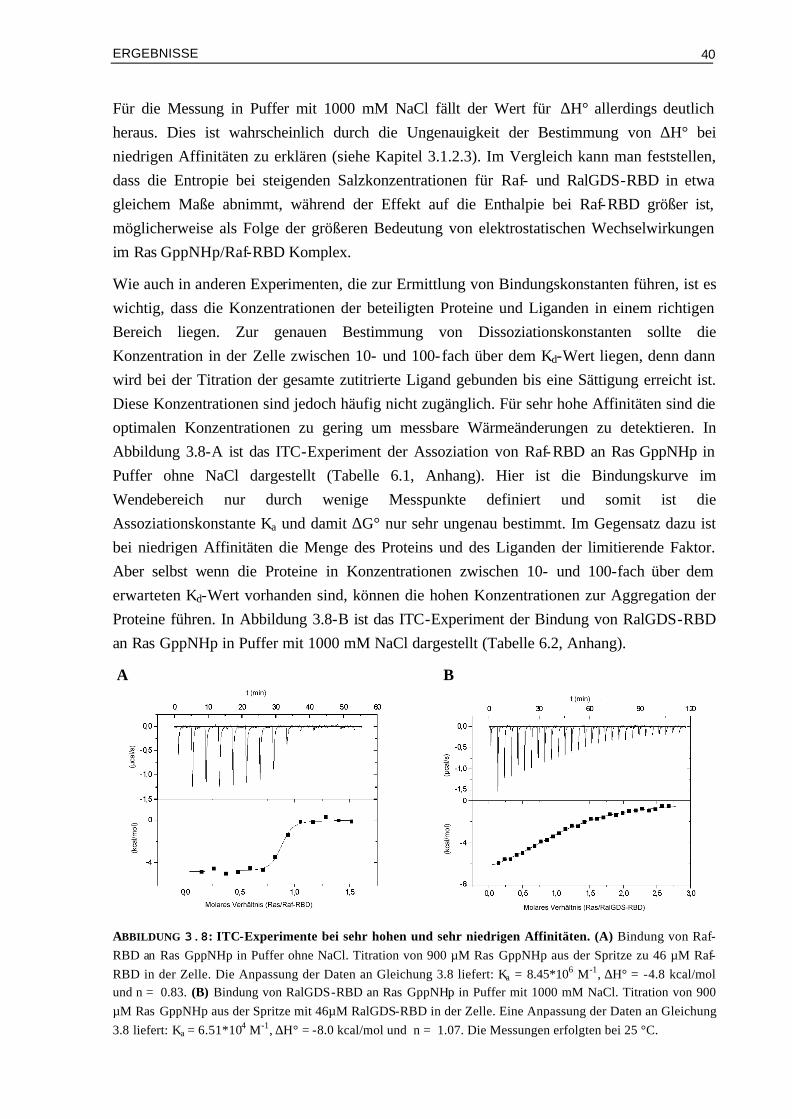
ERGEBNISSE
40
Für die Messung in Puffer mit 1000 mM NaCl fällt der Wert für ∆H° allerdings deutlich heraus. Dies ist wahrscheinlich durch die Ungenauigkeit der Bestimmung von ∆H° bei niedrigen Affinitäten zu erklären (siehe Kapitel 3.1.2.3). Im Vergleich kann man feststellen, dass die Entropie bei steigenden Salzkonzentrationen für Raf- und RalGDS-RBD in etwa gleichem Maße abnimmt, während der Effekt auf die Enthalpie bei Raf-RBD größer ist, möglicherweise als Folge der größeren Bedeutung von elektrostatischen Wechselwirkungen im Ras⋅GppNHp/Raf-RBD Komplex.
Wie auch in anderen Experimenten, die zur Ermittlung von Bindungskonstanten führen, ist es wichtig, dass die Konzentrationen der beteiligten Proteine und Liganden in einem richtigen Bereich liegen. Zur genauen Bestimmung von Dissoziationskonstanten sollte die Konzentration in der Zelle zwischen 10- und 100-fach über dem Kd-Wert liegen, denn dann wird bei der Titration der gesamte zutitrierte Ligand gebunden bis eine Sättigung erreicht ist. Diese Konzentrationen sind jedoch häufig nicht zugänglich. Für sehr hohe Affinitäten sind die optimalen Konzentrationen zu gering um messbare Wärmeänderungen zu detektieren. In Abbildung 3.8-A ist das ITC-Experiment der Assoziation von Raf-RBD an Ras⋅GppNHp in Puffer ohne NaCl dargestellt (Tabelle 6.1, Anhang). Hier ist die Bindungskurve im Wendebereich nur durch wenige Messpunkte definiert und somit ist die Assoziationskonstante Ka und damit ∆G° nur sehr ungenau bestimmt. Im Gegensatz dazu ist bei niedrigen Affinitäten die Menge des Proteins und des Liganden der limitierende Faktor. Aber selbst wenn die Proteine in Konzentrationen zwischen 10- und 100-fach über dem erwarteten Kd-Wert vorhanden sind, können die hohen Konzentrationen zur Aggregation der Proteine führen. In Abbildung 3.8-B ist das ITC-Experiment der Bindung von RalGDS-RBD an Ras⋅GppNHp in Puffer mit 1000 mM NaCl dargestellt (Tabelle 6.2, Anhang).
A B
ABBILDUNG 3.8: ITC-Experimente bei sehr hohen und sehr niedrigen Affinitäten. (A) Bindung von Raf-RBD an Ras ⋅GppNHp in Puffer ohne NaCl. Titration von 900 µM Ras ⋅GppNHp aus der Spritze zu 46 µM Raf-RBD in der Zelle. Die Anpassung der Daten an Gleichung 3.8 liefert: Ka = 8.45*106 M-1, ∆H° = -4.8 kcal/mol und n = 0.83. (B) Bindung von RalGDS-RBD an Ras⋅GppNHp in Puffer mit 1000 mM NaCl. Titration von 900 µM Ras⋅GppNHp aus der Spritze mit 46µM RalGDS-RBD in der Zelle. Eine Anpassung der Daten an Gleichung 3.8 liefert: Ka = 6.51*104 M-1, ∆H° = -8.0 kcal/mol und n = 1.07. Die Messungen erfolgten bei 25 °C.
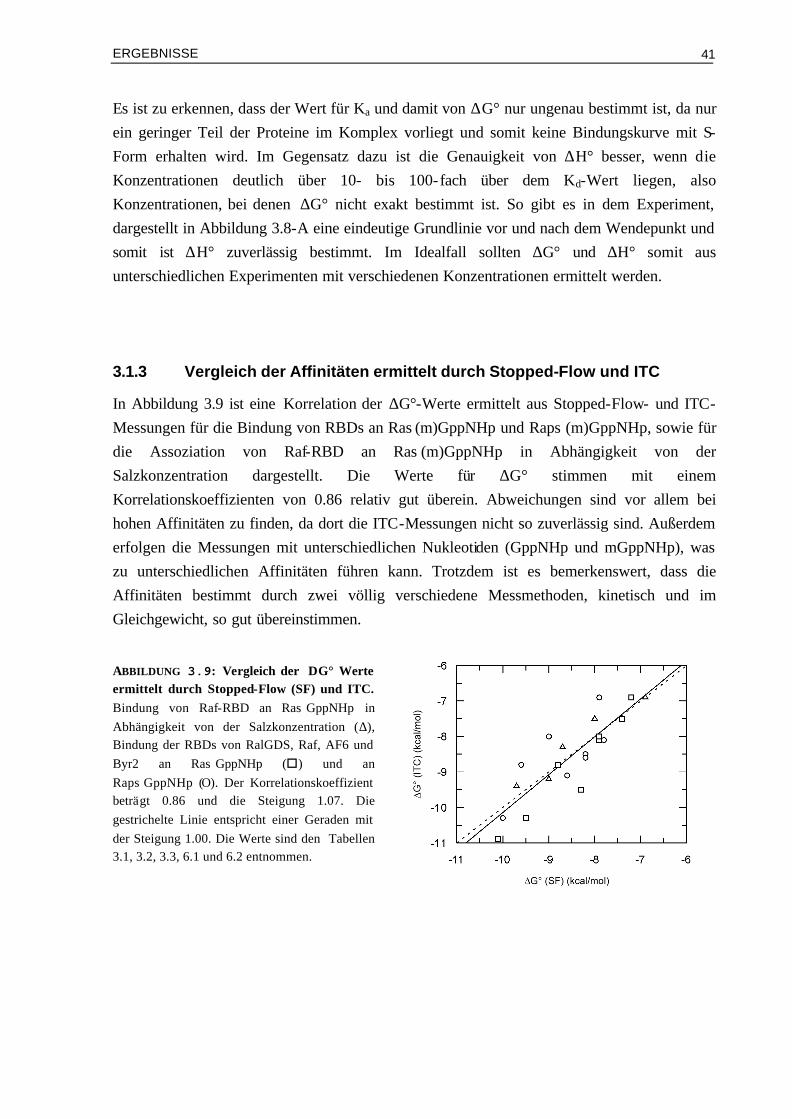
ERGEBNISSE
41
Es ist zu erkennen, dass der Wert für Ka und damit von ∆G° nur ungenau bestimmt ist, da nur ein geringer Teil der Proteine im Komplex vorliegt und somit keine Bindungskurve mit S-Form erhalten wird. Im Gegensatz dazu ist die Genauigkeit von ∆H° besser, wenn die Konzentrationen deutlich über 10- bis 100-fach über dem Kd-Wert liegen, also Konzentrationen, bei denen ∆G° nicht exakt bestimmt ist. So gibt es in dem Experiment, dargestellt in Abbildung 3.8-A eine eindeutige Grundlinie vor und nach dem Wendepunkt und somit ist ∆H° zuverlässig bestimmt. Im Idealfall sollten ∆G° und ∆H° somit aus unterschiedlichen Experimenten mit verschiedenen Konzentrationen ermittelt werden.
3.1.3 Vergleich der Affinitäten ermittelt durch Stopped-Flow und ITC
In Abbildung 3.9 ist eine Korrelation der ∆G°-Werte ermittelt aus Stopped-Flow- und ITC-Messungen für die Bindung von RBDs an Ras⋅(m)GppNHp und Raps⋅(m)GppNHp, sowie für die Assoziation von Raf-RBD an Ras⋅(m)GppNHp in Abhängigkeit von der Salzkonzentration dargestellt. Die Werte für ∆G° stimmen mit einem Korrelationskoeffizienten von 0.86 relativ gut überein. Abweichungen sind vor allem bei hohen Affinitäten zu finden, da dort die ITC-Messungen nicht so zuverlässig sind. Außerdem erfolgen die Messungen mit unterschiedlichen Nukleotiden (GppNHp und mGppNHp), was zu unterschiedlichen Affinitäten führen kann. Trotzdem ist es bemerkenswert, dass die Affinitäten bestimmt durch zwei völlig verschiedene Messmethoden, kinetisch und im Gleichgewicht, so gut übereinstimmen.
ABBILDUNG 3.9: Vergleich der ∆ G° Werte ermittelt durch Stopped-Flow (SF) und ITC. Bindung von Raf-RBD an Ras ⋅GppNHp in Abhängigkeit von der Salzkonzentration (∆), Bindung der RBDs von RalGDS, Raf, AF6 und Byr2 an Ras ⋅GppNHp (o) und an Raps⋅GppNHp (Ο). Der Korrelationskoeffizient beträgt 0.86 und die Steigung 1.07. Die gestrichelte Linie entspricht einer Geraden mit der Steigung 1.00. Die Werte sind den Tabellen 3.1, 3.2, 3.3, 6.1 und 6.2 entnommen.

ERGEBNISSE
42
3.2 Rolle der Elektrostatik für die kinetischen Eigenschaften der Wechselwirkung von RalGDS- und Raf-RBD mit Ras Proteinen
Für viele Protein-Komplexe konnte gezeigt werden, dass die Assoziationsratenkonstante größer ist, je höher die elektrostatische Komplementarität in der Kontaktfläche ist. So sind die RBDs von Raf und RalGDS hauptsächlich positiv geladen und Ras und Rap sind, insbesondere in der Effektor-Bindungsregion negativ geladen. Die Rolle der Elektrostatik für die kinetischen Eigenschaften (kon und koff) der Wechselwirkungen von Ras Proteinen mit Effektor-RBDs sollte zunächst an RalGDS-RBD/Ras- bzw. Rap- Komplexen sowie an Raf-RBD/Ras-Komplexen mit bereits aus Mutationsstudien vorhandenen Protein-Mutanten analysiert werden. Dabei wurden nur Mutanten untersucht, die durch Veränderung von geladenen Aminosäureseitenketten die Elektrostatik in den Komplexen beeinflussen.
3.2.1 Kinetik der Bindung von Mutanten-Komplexen aus RalGDS-RBD und Ras/Rap⋅mGppNHp
Bei den analysierten Mutanten von RalGDS-RBD handelt es sich um Mutationen in der Kontaktfläche, die mit der Effektor-Region von Ras wechselwirken. Für RalGDS-RBD sind dies die Aminosäuren aus ß1, ß2 sowie Helix 1 (Vetter et al., 1999). Es wurden nur Proteine mit Mutationen von oder zu geladenen Aminosäuren analysiert (R16A, K28A, K44A, D47A, D47K, K48A, N50K, E52A, E53A und E53K). Auf der Seite von Ras wurden Mutanten von geladenen Aminosäure-Resten im Effektor-Bereich (E31A, E31K, D33A, E37A, R41A, E62A und E63A) verwendet. Es wurden die Assoziations- und Dissoziationsratenkonstanten der Bindung von RalGDS-RBDWT und Mutanten an Ras⋅mGppNHp und Rap⋅mGppNHp sowie an Ras⋅mGppNHp-Mutanten im Stopped-Flow gemessen. Die Assoziations-ratenkonstanten wurden aus der linearen Abhängigkeit für geringe RBD-Konzentrationen ermittelt, und die Dissoziationsratenkonstanten jeweils aus einem Verdrängungsexperiment bestimmt. Beispielhaft ist die Assoziationskinetik der Bindung von RalGDS-RBDWT sowie den Mutanten RalGDS-RBDE53A und RalGDS-RBDK44A an Ras⋅mGppNHp in Abbildung 3.10 dargestellt. Die Ergebnisse aller Messungen sind in Tabelle 3.4 zusammengefasst. (Die Tabelle enthält auch berechnete Assoziationsratenkonstanten. Das Prinzip der Berechnungen sowie die Beschreibung der Ergebnisse erfolgt allerdings erst in Kapitel 3.3 und 3.4.)
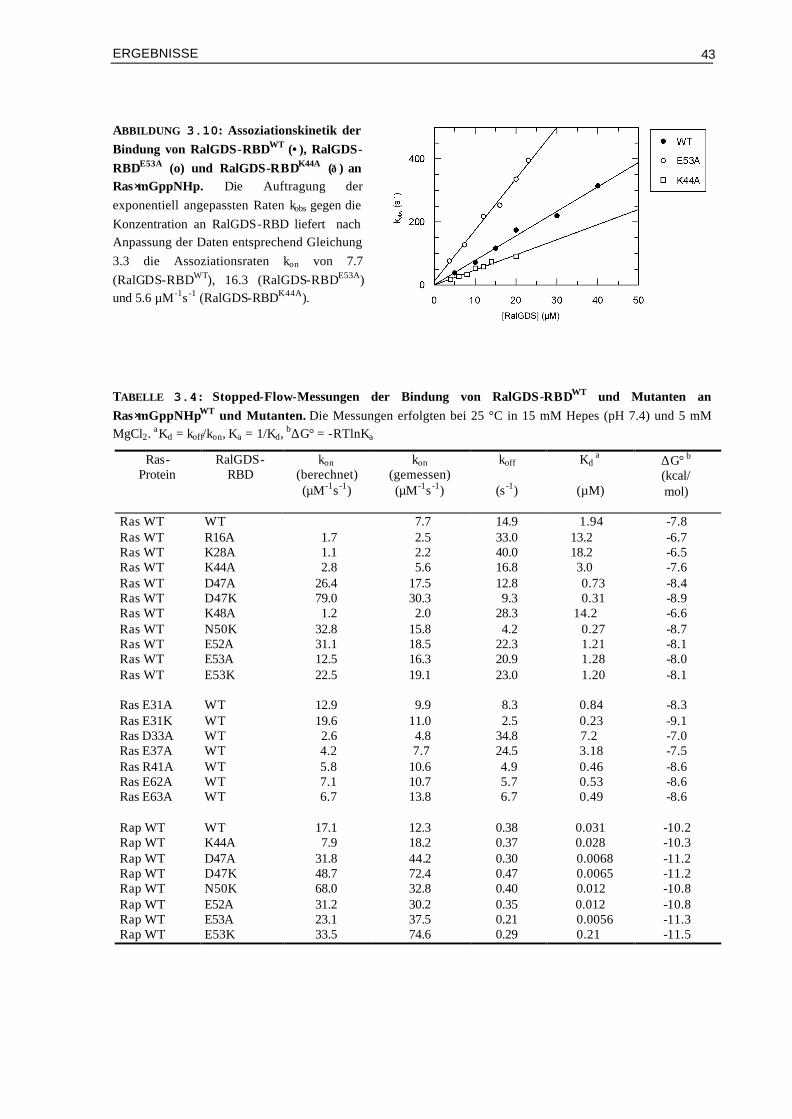
ERGEBNISSE
43
ABBILDUNG 3.10: Assoziationskinetik der Bindung von RalGDS-RBDWT (•), RalGDS-RBDE53A (o) und RalGDS-RBDK44A (ð ) an Ras⋅mGppNHp. Die Auftragung der exponentiell angepassten Raten kobs gegen die Konzentration an RalGDS-RBD liefert nach Anpassung der Daten entsprechend Gleichung 3.3 die Assoziationsraten kon von 7.7 (RalGDS-RBDWT), 16.3 (RalGDS-RBDE53A) und 5.6 µM -1s -1 (RalGDS-RBDK44A).
TABELLE 3.4: Stopped-Flow-Messungen der Bindung von RalGDS-RBDWT und Mutanten an Ras⋅mGppNHpWT und Mutanten. Die Messungen erfolgten bei 25 °C in 15 mM Hepes (pH 7.4) und 5 mM MgCl2. aKd = koff/kon, Ka = 1/Kd, b∆G° = -RTlnKa
Ras- Protein
RalGDS-RBD
kon (berechnet) (µM-1s -1)
kon (gemessen) (µM-1s -1)
koff
(s-1)
Kd a
(µM)
∆G° b (kcal/ mol)
Ras WT WT 7.7 14.9 1.94 -7.8 Ras WT R16A 1.7 2.5 33.0 13.2 -6.7 Ras WT K28A 1.1 2.2 40.0 18.2 -6.5 Ras WT K44A 2.8 5.6 16.8 3.0 -7.6 Ras WT D47A 26.4 17.5 12.8 0.73 -8.4 Ras WT D47K 79.0 30.3 9.3 0.31 -8.9 Ras WT K48A 1.2 2.0 28.3 14.2 -6.6 Ras WT N50K 32.8 15.8 4.2 0.27 -8.7 Ras WT E52A 31.1 18.5 22.3 1.21 -8.1 Ras WT E53A 12.5 16.3 20.9 1.28 -8.0 Ras WT E53K 22.5 19.1 23.0 1.20 -8.1 Ras E31A WT 12.9 9.9 8.3 0.84 -8.3 Ras E31K WT 19.6 11.0 2.5 0.23 -9.1 Ras D33A WT 2.6 4.8 34.8 7.2 -7.0 Ras E37A WT 4.2 7.7 24.5 3.18 -7.5 Ras R41A WT 5.8 10.6 4.9 0.46 -8.6 Ras E62A WT 7.1 10.7 5.7 0.53 -8.6 Ras E63A WT 6.7 13.8 6.7 0.49 -8.6 Rap WT WT 17.1 12.3 0.38 0.031 -10.2 Rap WT K44A 7.9 18.2 0.37 0.028 -10.3 Rap WT D47A 31.8 44.2 0.30 0.0068 -11.2 Rap WT D47K 48.7 72.4 0.47 0.0065 -11.2 Rap WT N50K 68.0 32.8 0.40 0.012 -10.8 Rap WT E52A 31.2 30.2 0.35 0.012 -10.8 Rap WT E53A 23.1 37.5 0.21 0.0056 -11.3 Rap WT E53K 33.5 74.6 0.29 0.21 -11.5
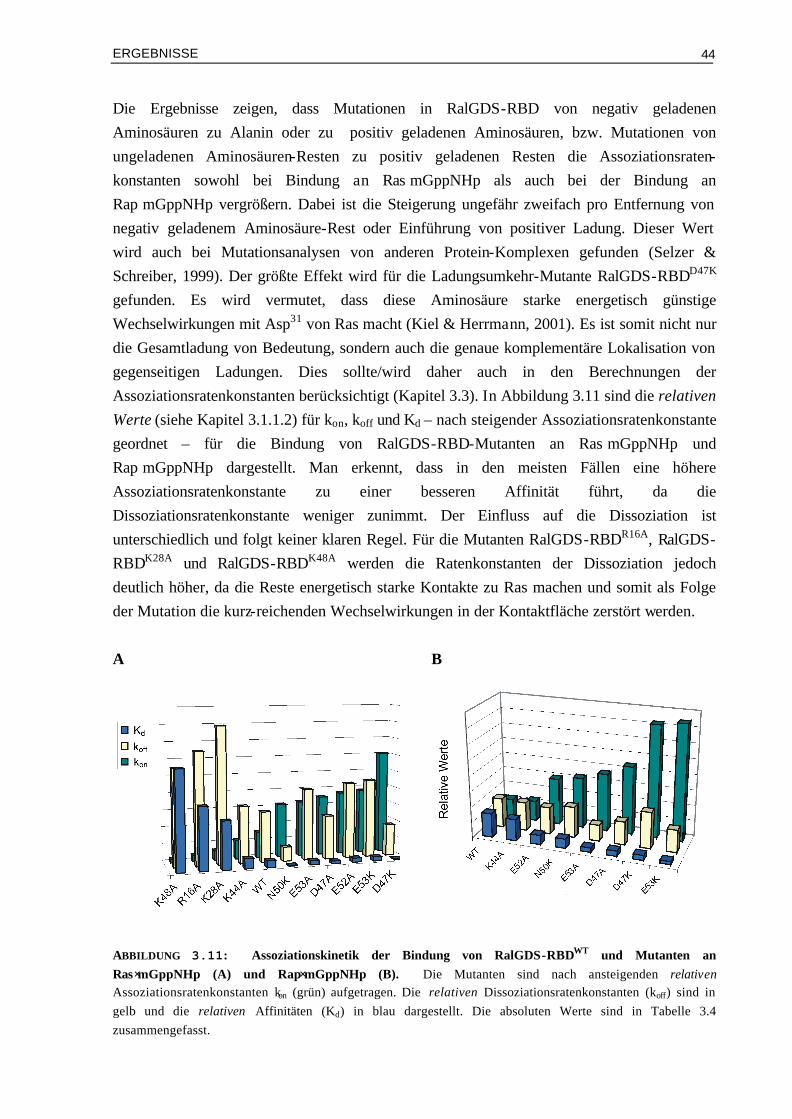
ERGEBNISSE
44
Die Ergebnisse zeigen, dass Mutationen in RalGDS-RBD von negativ geladenen Aminosäuren zu Alanin oder zu positiv geladenen Aminosäuren, bzw. Mutationen von ungeladenen Aminosäuren-Resten zu positiv geladenen Resten die Assoziationsraten-konstanten sowohl bei Bindung an Ras⋅mGppNHp als auch bei der Bindung an Rap⋅mGppNHp vergrößern. Dabei ist die Steigerung ungefähr zweifach pro Entfernung von negativ geladenem Aminosäure-Rest oder Einführung von positiver Ladung. Dieser Wert wird auch bei Mutationsanalysen von anderen Protein-Komplexen gefunden (Selzer & Schreiber, 1999). Der größte Effekt wird für die Ladungsumkehr-Mutante RalGDS-RBDD47K gefunden. Es wird vermutet, dass diese Aminosäure starke energetisch günstige Wechselwirkungen mit Asp31 von Ras macht (Kiel & Herrmann, 2001). Es ist somit nicht nur die Gesamtladung von Bedeutung, sondern auch die genaue komplementäre Lokalisation von gegenseitigen Ladungen. Dies sollte/wird daher auch in den Berechnungen der Assoziationsratenkonstanten berücksichtigt (Kapitel 3.3). In Abbildung 3.11 sind die relativen Werte (siehe Kapitel 3.1.1.2) für kon, koff und Kd – nach steigender Assoziationsratenkonstante geordnet – für die Bindung von RalGDS-RBD-Mutanten an Ras⋅mGppNHp und Rap⋅mGppNHp dargestellt. Man erkennt, dass in den meisten Fällen eine höhere Assoziationsratenkonstante zu einer besseren Affinität führt, da die Dissoziationsratenkonstante weniger zunimmt. Der Einfluss auf die Dissoziation ist unterschiedlich und folgt keiner klaren Regel. Für die Mutanten RalGDS-RBDR16A, RalGDS-RBDK28A und RalGDS-RBDK48A werden die Ratenkonstanten der Dissoziation jedoch deutlich höher, da die Reste energetisch starke Kontakte zu Ras machen und somit als Folge der Mutation die kurz-reichenden Wechselwirkungen in der Kontaktfläche zerstört werden. A B
ABBILDUNG 3.11: Assoziationskinetik der Bindung von RalGDS-RBDWT und Mutanten an Ras⋅mGppNHp (A) und Rap⋅ mGppNHp (B). Die Mutanten sind nach ansteigenden relativen Assoziationsratenkonstanten kon (grün) aufgetragen. Die relativen Dissoziationsratenkonstanten (koff) sind in gelb und die relativen Affinitäten (Kd) in blau dargestellt. Die absoluten Werte sind in Tabelle 3.4 zusammengefasst.

ERGEBNISSE
45
Die Mutation von negativ geladenen Aminosäuren von Ras (Asp33 und Glu37) zu Alanin führen zu einem niedrigeren Wert für kon, was den Erwartungen entspricht, da die elektrostatische Komplementarität geringer wird; die Werte für koff werden größer. Im Gegensatz dazu führen die Mutationen der ebenfall negativ geladenen Aminosäuren Glu31, Glu62 und Glu63 zu Alanin zu einer höheren Assoziationsratenkonstante. Hier sind die Werte für kon somit höher, obwohl die elektrostatische Komplementarität verringert wird. Diese Ergebnisse können möglicherweise durch günstige kurz-reichende intermolekulare Wechselwirkungen erklärt werden, denn für diese Mutanten wird eine langsamere Dissoziation gemessen. Die Mutanten Ras⋅mGppNHpE31K und Ras⋅mGppNHp R41A führen zu schnelleren Assoziationen mit einer verlangsamten Dissoziation. Der höhere kon-Wert für Ras⋅mGppNHpE31K im Komplex mit RalGDS-RBD entspricht hinsichtlich der niedrigeren elektrostatischen Komplementarität nicht den Erwartungen. Eine sehr nahe liegende Erklärung hierfür ist, dass der zweite Schritt des Bindungsmechanismus verändert ist, aufgrund energetisch günstiger Wechselwirkungen zwischen Lys31 von RalGDS-RBD und Glu31 von Ras.
3.2.2 Kinetik der Bindung von Raf-RBD-Mutanten an Ras⋅mGppNHp
Die Rolle der Elektrostatik wurde auch durch Analyse der Bindung von Raf-RBD-Mutanten im Komplex mit Ras⋅mGppNHp untersucht. Analysiert wurden wie auch bei RalGDS-RBD, Mutationen von geladenen Aminosäuren (R59A, N64D, K65A, K65E, R67A, K84A und K84E), die an Kontakten zur Effektor-Bindungsregion von Ras beteiligt sind (Nassar et al, 1995, Nassar et al, 1996). Die Messungen der Mutanten wurden bei 25 °C ohne Salz und bei 15 °C in Puffer mit zusätzlich 125 mM NaCl durchgeführt. Die Messungen bei physiologischer Kochsalzkonzentration sind wichtig für spätere Vergleiche mit der Signalweiterleitung in vivo. Außerdem sind die Ratenkonstanten der Assoziation in Puffer ohne NaCl sehr groß, ebenso wie die kobs-Werte, so dass schon bei niedrigen Raf-RBD Konzentrationen die Grenze der Auflösung mittels Stopped-Flow erreicht ist. Des Weiteren kommt es bei Raf-RBD in Puffer ohne NaCl teilweise zu Stabilitätsproblemen der Raf-RBD Proteine.
Die Assoziationsratenkonstanten wurden aus dem linearen Bereich ermittelt und eine Auftragung der exponentiell angepassten Ratenkonstanten, kobs, gegen die Raf-RBD Konzentration ist für WT und drei Mutanten in Abbildung 3.12 dargestellt. Da die Achsenabschnitte auch hier sehr dicht bei Null liegen oder teilweise sogar negativ sind, wurden die Dissoziationsratenkonstanten wiederum aus Verdrängungsexperimenten bestimmt. Alle Ergebnisse für die Assoziations-, Dissoziationsratenkonstanten und für die Affinitäten (sowie für die berechneten Assoziationsratenkonstanten) sind in Tabelle 3.5 zusammengefasst.
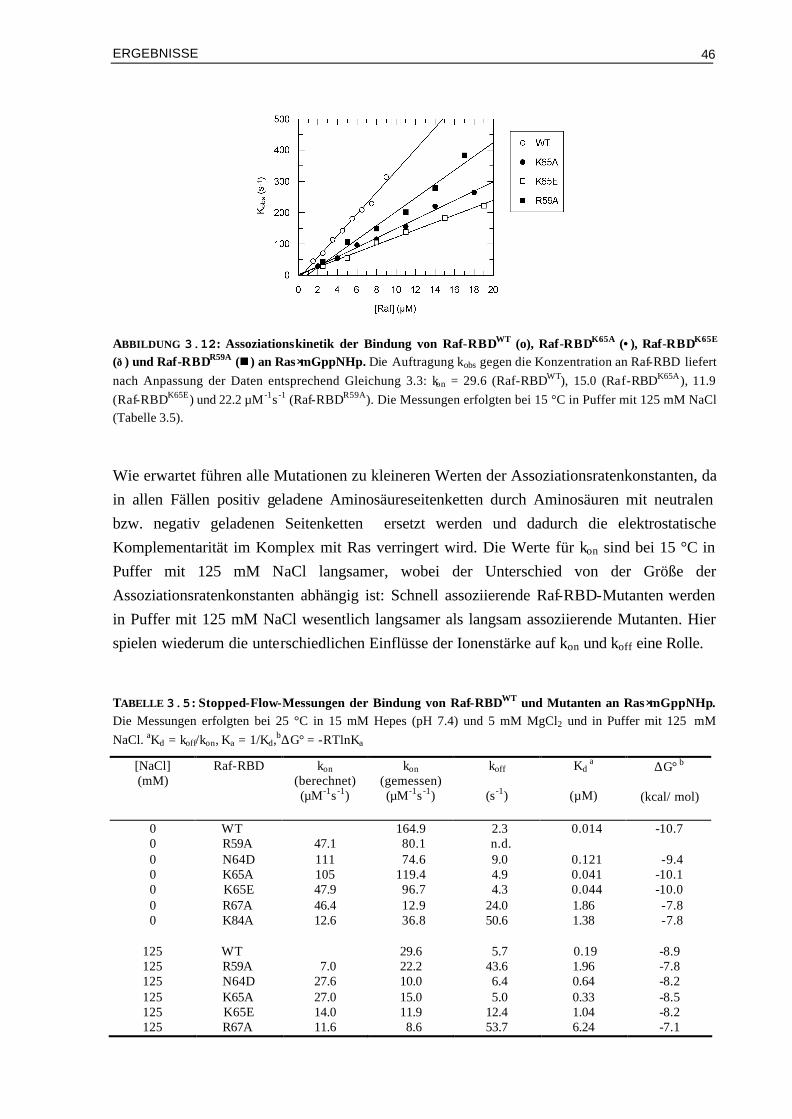
ERGEBNISSE
46
ABBILDUNG 3.12: Assoziationskinetik der Bindung von Raf-RBDWT (o), Raf-RBDK65A (•), Raf-RBDK65E (ð ) und Raf-RBDR59A (n) an Ras⋅mGppNHp. Die Auftragung kobs gegen die Konzentration an Raf-RBD liefert nach Anpassung der Daten entsprechend Gleichung 3.3: kon = 29.6 (Raf-RBDWT), 15.0 (Raf-RBDK65A), 11.9 (Raf-RBDK65E) und 22.2 µM -1s -1 (Raf-RBDR59A). Die Messungen erfolgten bei 15 °C in Puffer mit 125 mM NaCl (Tabelle 3.5).
Wie erwartet führen alle Mutationen zu kleineren Werten der Assoziationsratenkonstanten, da in allen Fällen positiv geladene Aminosäureseitenketten durch Aminosäuren mit neutralen bzw. negativ geladenen Seitenketten ersetzt werden und dadurch die elektrostatische Komplementarität im Komplex mit Ras verringert wird. Die Werte für kon sind bei 15 °C in Puffer mit 125 mM NaCl langsamer, wobei der Unterschied von der Größe der Assoziationsratenkonstanten abhängig ist: Schnell assoziierende Raf-RBD-Mutanten werden in Puffer mit 125 mM NaCl wesentlich langsamer als langsam assoziierende Mutanten. Hier spielen wiederum die unterschiedlichen Einflüsse der Ionenstärke auf kon und koff eine Rolle.
TABELLE 3.5: Stopped-Flow-Messungen der Bindung von Raf-RBDWT und Mutanten an Ras⋅mGppNHp. Die Messungen erfolgten bei 25 °C in 15 mM Hepes (pH 7.4) und 5 mM MgCl2 und in Puffer mit 125 mM NaCl. aKd = koff/kon, Ka = 1/Kd,b∆G° = -RTlnKa
[NaCl] (mM)
Raf-RBD kon (berechnet) (µM-1s -1)
kon (gemessen) (µM-1s -1)
koff
(s-1)
Kd a
(µM)
∆G° b
(kcal/ mol)
0 WT 164.9 2.3 0.014 -10.7 0 R59A 47.1 80.1 n.d. 0 N64D 111 74.6 9.0 0.121 -9.4 0 K65A 105 119.4 4.9 0.041 -10.1 0 K65E 47.9 96.7 4.3 0.044 -10.0 0 R67A 46.4 12.9 24.0 1.86 -7.8 0 K84A 12.6 36.8 50.6 1.38 -7.8
125 WT 29.6 5.7 0.19 -8.9 125 R59A 7.0 22.2 43.6 1.96 -7.8 125 N64D 27.6 10.0 6.4 0.64 -8.2 125 K65A 27.0 15.0 5.0 0.33 -8.5 125 K65E 14.0 11.9 12.4 1.04 -8.2 125 R67A 11.6 8.6 53.7 6.24 -7.1
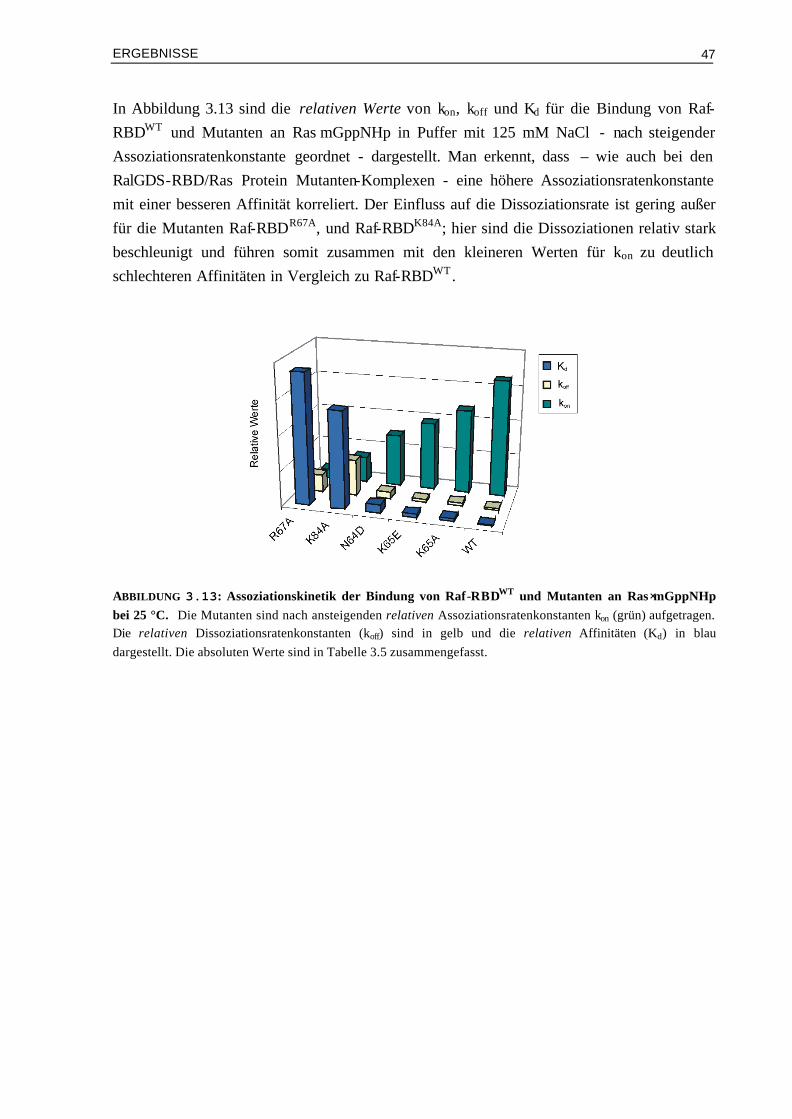
ERGEBNISSE
47
In Abbildung 3.13 sind die relativen Werte von kon, koff und Kd für die Bindung von Raf-RBDWT und Mutanten an Ras⋅mGppNHp in Puffer mit 125 mM NaCl - nach steigender Assoziationsratenkonstante geordnet - dargestellt. Man erkennt, dass – wie auch bei den RalGDS-RBD/Ras Protein Mutanten-Komplexen - eine höhere Assoziationsratenkonstante mit einer besseren Affinität korreliert. Der Einfluss auf die Dissoziationsrate ist gering außer für die Mutanten Raf-RBDR67A, und Raf-RBDK84A; hier sind die Dissoziationen relativ stark beschleunigt und führen somit zusammen mit den kleineren Werten für kon zu deutlich schlechteren Affinitäten in Vergleich zu Raf-RBDWT .
ABBILDUNG 3.13: Assoziationskinetik der Bindung von Raf-RBDWT und Mutanten an Ras ⋅mGppNHp bei 25 °C. Die Mutanten sind nach ansteigenden relativen Assoziationsratenkonstanten kon (grün) aufgetragen. Die relativen Dissoziationsratenkonstanten (koff) sind in gelb und die relativen Affinitäten (Kd) in blau dargestellt. Die absoluten Werte sind in Tabelle 3.5 zusammengefasst.

ERGEBNISSE
48
)a1
1(RT
Ulnklnk ononκ+
∆−°=
3.3 Gezielte Manipulation von RalGDS- und Raf-RBD zur Veränderung der elektrostatischen Eigenschaften
Anhand der Untersuchung von vielen Protein-Komplexen und auch bei der Wechselwirkung von Ras mit Raf-RBD (Kapitel 3.1.1) konnte gezeigt werden, dass die Assoziationsratenkonstante eine Funktion von einer basalen Ratenkonstante kon° (in Abwesenheit von elektrostatischen Wechselwirkungen) und der Coulomb-Energie, berechnet aus den elektrostatischen Wechselwirkungen der Proteine, ist (Gleichung 3.7).
In Kooperation mit Prof. Gideon Schreiber vom Weizmann-Institut in Israel wurde das dort entwickelte Computerprogramm PARE (Protein Association Rate Enhancement, Selzer et al., 2000) zur Berechnung von Assoziationsratenkonstanten von Mutanten-Komplexen verwendet. Ziel war die gezielte Manipulation von Aminosäuren der Proteine RalGDS- und Raf-RBD um Veränderungen der Elektrostatik zu erzielen, welche im Komplex mit Ras zu veränderten kinetischen Eigenschaften führen sollten. Die Betrachtung der Oberflächenpotentiale von RalGDS- und Raf-RBD sowie Ras zeigen, dass Ras hauptsächlich negativ geladen ist und die RBDs positiv. Es wurde versucht durch gezielte Einführung von positiv geladenen Aminosäuren in die RBDs von RalGDS und Raf die Ladungs-komplementarität zu erhöhen und folglich die Assoziationsratenkonstante bei der Bindung an Ras⋅mGppNHp zu erhöhen.
Zum Entwurf von Mutanten wurden als erstes Aminosäuren von RalGDS- und Raf-RBD identifiziert, die exponiert zur Oberfläche sind und sich in der Nähe der Proteinkontaktfläche befinden, aber nicht an kurz-reichenden Wechselwirkungen in der Kontaktfläche beteiligt sind. Diese Reste wurden mit Hilfe des Strukturprogramms Swiss-PDB Viewer zu Lysinen mutiert, um so die elektrostatische Komplementarität im Komplex mit Ras zu erhöhen. Nach Energieminimierung der Orientierung der eingeführten Seitenkette wurde die erwartete Assoziationsratenkonstante mit Hilfe des Computerprogramms PARE berechnet. Die Berechnungen beruhen auf Gleichung 3.7, wobei die Coulomb-Energie für die einzelnen Proteine nach folgender Gleichung berechnet wird (3.10):
(3.10) Dabei sind i und j die geladenen Atome in dem Protein, r der Abstand der geladenen Atome, ε? die relative Dielektrizitätskonstante (für Wasser 80), a ist der minimale Abstand der Annäherung (wird auf 6 Å festgesetzt, Selzer & Schreiber, 1999), und κ ist die inverse Debye-Länge (wird auf 0.488*109 m-1 bei einer Ionenstärke I von 0.022 M festgesetzt). Die
a1
arer4
qq21
U)ij(
ij ijo
ji
κ
κ
επε +=
−−
∑

ERGEBNISSE
49
elektrostatische Wechselwirkungsenergie (∆U) zwischen zwei Proteinen wird dann nach folgender Gleichung berechnet: (3.11) Dabei werden UKomplex und UProtein nach Gleichung 3.10 berechnet. Die Beziehung zwischen kon und ∆U bei einer Ionenstärke von 0.022 M ist dann durch folgende Gleichung gegeben: (3.12) Dabei führt der Wert für I = 0.022 zum Wert von 0.9 im Nenner. Mutationen, die die Assoziationsratenkonstante beschleunigen, wurden innerhalb eines Proteins kombiniert und kon erneut berechnet. Glycine und Proline wurden nicht mutiert um zu verhindern, dass die Protein-Rückgrad Konformationen verändert werden.
3.3.1 Manipulation von RalGDS-RBD und kinetische Analyse der Mutanten
im Komplex mit Ras⋅mGppNHp
Die Bindung von RalGDS-RBDWT an Ras⋅mGppNHp ist mit einer Affinität im mikromolaren Bereich relativ schwach. Dabei ist die Assoziationsratenkonstante mit 7.7 µM-1s-1 relativ langsam und die Dissoziationsratenkonstante mit 14.9 s-1 relativ schnell. Es sollten Mutanten von RalGDS-RBD entworfen werden, die zu einer Beschleunigung der Assoziationsratenkonstante bei Bindung an Ras⋅mGppNHp führen. Unter der Annahme, dass diese Mutationen die Dissoziation nicht beschleunigen, da keine Reste innerhalb der Kontaktfläche mutiert werden, sollten somit besser bindende RalGDS-RBD/Ras-Komplexe erhalten werden. Die Ergebnisse der berechneten Assoziationsratenkonstanten sind in Abbildung 3.14 dargestellt und in Tabelle 3.6 zusammengefasst. In Abbildung 3.14 wurden die berechneten Werte für kon auf kon (Wildtyp) normiert und daher als Beschleunigungen im Vergleich zu RalGDS-RBDWT bezeichnet. Während die Oberfläche von Ras – insbesondere die Effektor-Region - hauptsächlich negativ geladen ist, besitzt die Oberfläche von RalGDS-RBD positiv geladene Bereiche. Alle zusätzlich positiv geladenen eingeführten Lysine sollten die Assoziation somit beschleunigen.
Die Berechnungen liefern für Einzelmutationen von ungeladenen Aminosäuren (S18K, L19K, N23K, M26K, L51K, N88K und Y89K) Beschleunigungen für kon zwischen 1.7- und 4.5-fach. Für Mutationen von negativ geladenen Resten (E54K, D90K) werden 3.0- bzw. 5.5-fach höhere Werte für kon im Vergleich zu Raf-RBDWT berechnet.
UUUU Protein2Protein1Komplex −−=∆
on(wt)muwt
)022.0on(mut)(I lnk9.0
UUlnk +
∆−∆==
t
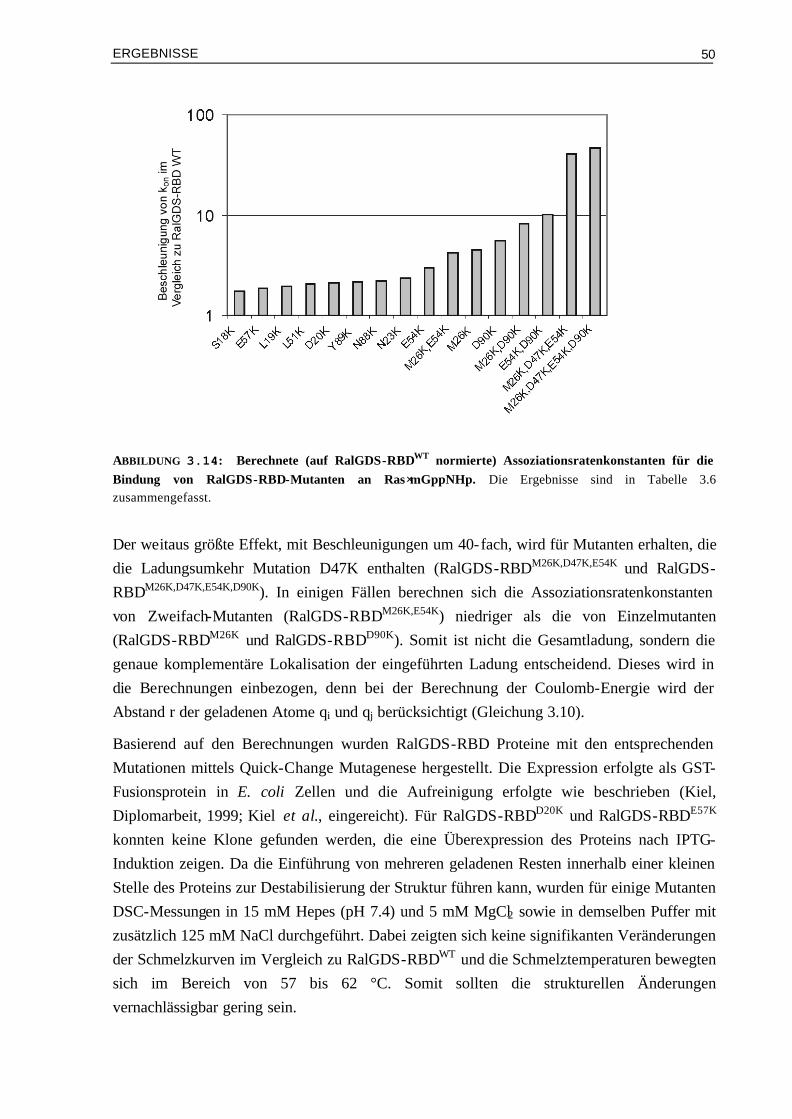
ERGEBNISSE
50
ABBILDUNG 3.14: Berechnete (auf RalGDS-RBDWT normierte) Assoziationsratenkonstanten für die Bindung von RalGDS-RBD-Mutanten an Ras⋅mGppNHp. Die Ergebnisse sind in Tabelle 3.6 zusammengefasst.
Der weitaus größte Effekt, mit Beschleunigungen um 40-fach, wird für Mutanten erhalten, die die Ladungsumkehr Mutation D47K enthalten (RalGDS-RBDM26K,D47K,E54K und RalGDS-RBDM26K,D47K,E54K,D90K). In einigen Fällen berechnen sich die Assoziationsratenkonstanten von Zweifach-Mutanten (RalGDS-RBDM26K,E54K) niedriger als die von Einzelmutanten (RalGDS-RBDM26K und RalGDS-RBDD90K). Somit ist nicht die Gesamtladung, sondern die genaue komplementäre Lokalisation der eingeführten Ladung entscheidend. Dieses wird in die Berechnungen einbezogen, denn bei der Berechnung der Coulomb-Energie wird der Abstand r der geladenen Atome qi und qj berücksichtigt (Gleichung 3.10).
Basierend auf den Berechnungen wurden RalGDS-RBD Proteine mit den entsprechenden Mutationen mittels Quick-Change Mutagenese hergestellt. Die Expression erfolgte als GST-Fusionsprotein in E. coli Zellen und die Aufreinigung erfolgte wie beschrieben (Kiel, Diplomarbeit, 1999; Kiel et al., eingereicht). Für RalGDS-RBDD20K und RalGDS-RBDE57K konnten keine Klone gefunden werden, die eine Überexpression des Proteins nach IPTG-Induktion zeigen. Da die Einführung von mehreren geladenen Resten innerhalb einer kleinen Stelle des Proteins zur Destabilisierung der Struktur führen kann, wurden für einige Mutanten DSC-Messungen in 15 mM Hepes (pH 7.4) und 5 mM MgCl2 sowie in demselben Puffer mit zusätzlich 125 mM NaCl durchgeführt. Dabei zeigten sich keine signifikanten Veränderungen der Schmelzkurven im Vergleich zu RalGDS-RBDWT und die Schmelztemperaturen bewegten sich im Bereich von 57 bis 62 °C. Somit sollten die strukturellen Änderungen vernachlässigbar gering sein.
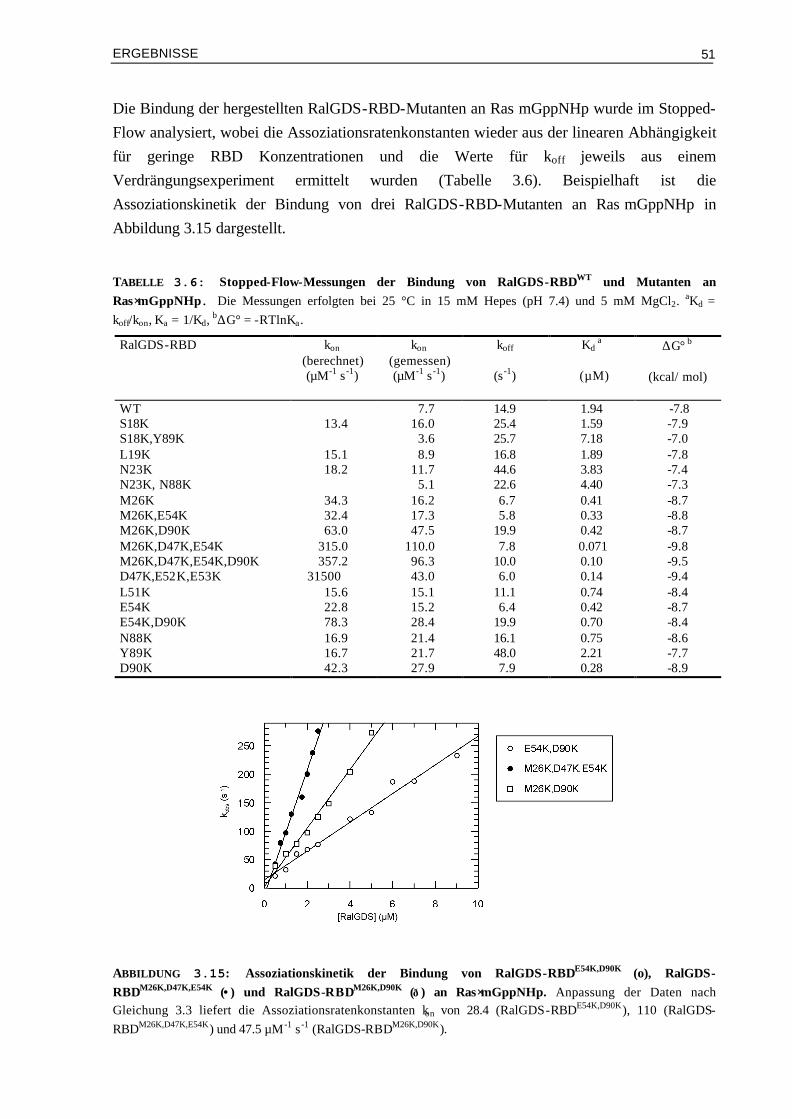
ERGEBNISSE
51
Die Bindung der hergestellten RalGDS-RBD-Mutanten an Ras⋅mGppNHp wurde im Stopped-Flow analysiert, wobei die Assoziationsratenkonstanten wieder aus der linearen Abhängigkeit für geringe RBD Konzentrationen und die Werte für koff jeweils aus einem Verdrängungsexperiment ermittelt wurden (Tabelle 3.6). Beispielhaft ist die Assoziationskinetik der Bindung von drei RalGDS-RBD-Mutanten an Ras⋅mGppNHp in Abbildung 3.15 dargestellt.
TABELLE 3.6: Stopped-Flow-Messungen der Bindung von RalGDS-RBDWT und Mutanten an Ras⋅mGppNHp. Die Messungen erfolgten bei 25 °C in 15 mM Hepes (pH 7.4) und 5 mM MgCl2. aKd = koff/kon, Ka = 1/Kd, b∆G° = -RTlnKa .
ABBILDUNG 3.15: Assoziationskinetik der Bindung von RalGDS-RBDE54K,D90K (o), RalGDS-RBDM26K,D47K,E54K (•) und RalGDS-RBDM26K,D90K (ð ) an Ras⋅mGppNHp. Anpassung der Daten nach Gleichung 3.3 liefert die Assoziationsratenkonstanten kon von 28.4 (RalGDS-RBDE54K,D90K), 110 (RalGDS-RBDM26K,D47K,E54K) und 47.5 µM -1 s -1 (RalGDS-RBDM26K,D90K).
RalGDS-RBD
kon (berechnet) (µM-1 s -1)
kon
(gemessen) (µM-1 s -1)
koff
(s-1)
Kd a
(µM)
∆G° b
(kcal/ mol)
WT 7.7 14.9 1.94 -7.8 S18K 13.4 16.0 25.4 1.59 -7.9 S18K,Y89K 3.6 25.7 7.18 -7.0 L19K 15.1 8.9 16.8 1.89 -7.8 N23K 18.2 11.7 44.6 3.83 -7.4 N23K, N88K 5.1 22.6 4.40 -7.3 M26K 34.3 16.2 6.7 0.41 -8.7 M26K,E54K 32.4 17.3 5.8 0.33 -8.8 M26K,D90K 63.0 47.5 19.9 0.42 -8.7 M26K,D47K,E54K 315.0 110.0 7.8 0.071 -9.8 M26K,D47K,E54K,D90K 357.2 96.3 10.0 0.10 -9.5 D47K,E52K,E53K 31500 43.0 6.0 0.14 -9.4 L51K 15.6 15.1 11.1 0.74 -8.4 E54K 22.8 15.2 6.4 0.42 -8.7 E54K,D90K 78.3 28.4 19.9 0.70 -8.4 N88K 16.9 21.4 16.1 0.75 -8.6 Y89K 16.7 21.7 48.0 2.21 -7.7 D90K 42.3 27.9 7.9 0.28 -8.9
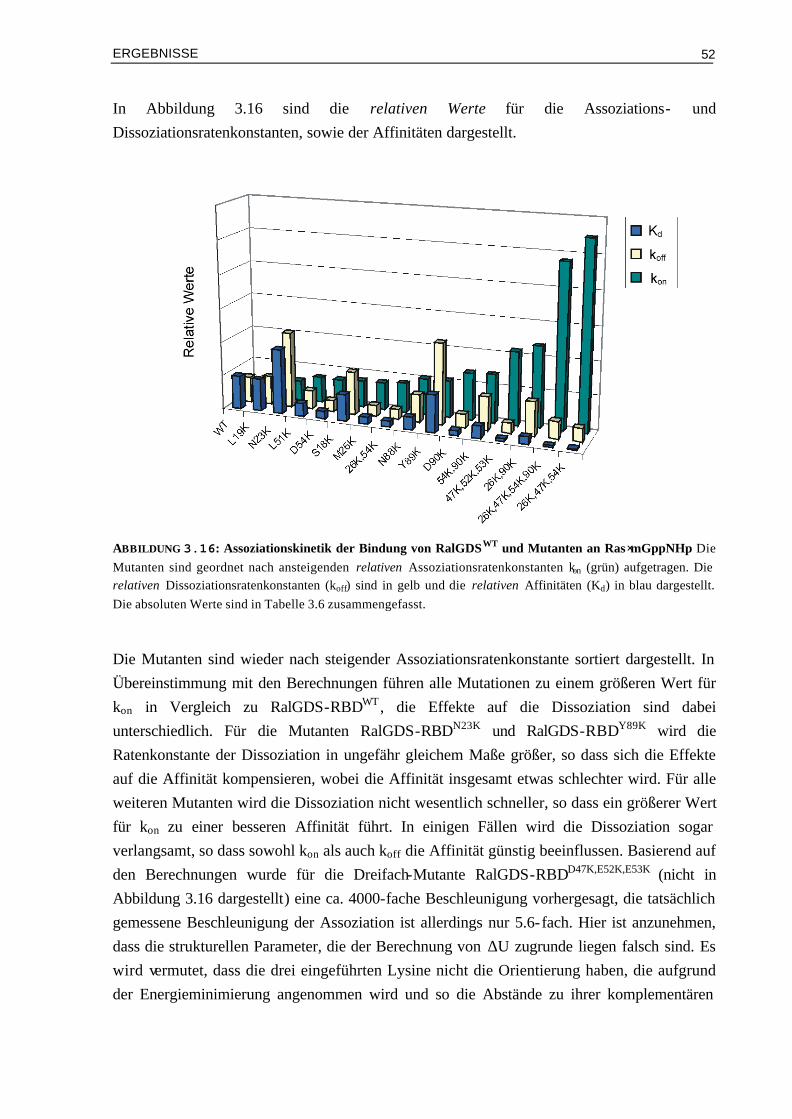
ERGEBNISSE
52
In Abbildung 3.16 sind die relativen Werte für die Assoziations- und Dissoziationsratenkonstanten, sowie der Affinitäten dargestellt.
ABBILDUNG 3.16: Assoziationskinetik der Bindung von RalGDSWT und Mutanten an Ras ⋅mGppNHp Die Mutanten sind geordnet nach ansteigenden relativen Assoziationsratenkonstanten kon (grün) aufgetragen. Die relativen Dissoziationsratenkonstanten (koff) sind in gelb und die relativen Affinitäten (Kd) in blau dargestellt. Die absoluten Werte sind in Tabelle 3.6 zusammengefasst.
Die Mutanten sind wieder nach steigender Assoziationsratenkonstante sortiert dargestellt. In Übereinstimmung mit den Berechnungen führen alle Mutationen zu einem größeren Wert für kon in Vergleich zu RalGDS-RBDWT , die Effekte auf die Dissoziation sind dabei unterschiedlich. Für die Mutanten RalGDS-RBDN23K und RalGDS-RBDY89K wird die Ratenkonstante der Dissoziation in ungefähr gleichem Maße größer, so dass sich die Effekte auf die Affinität kompensieren, wobei die Affinität insgesamt etwas schlechter wird. Für alle weiteren Mutanten wird die Dissoziation nicht wesentlich schneller, so dass ein größerer Wert für kon zu einer besseren Affinität führt. In einigen Fällen wird die Dissoziation sogar verlangsamt, so dass sowohl kon als auch koff die Affinität günstig beeinflussen. Basierend auf den Berechnungen wurde für die Dreifach-Mutante RalGDS-RBDD47K,E52K,E53K (nicht in Abbildung 3.16 dargestellt) eine ca. 4000-fache Beschleunigung vorhergesagt, die tatsächlich gemessene Beschleunigung der Assoziation ist allerdings nur 5.6-fach. Hier ist anzunehmen, dass die strukturellen Parameter, die der Berechnung von ∆U zugrunde liegen falsch sind. Es wird vermutet, dass die drei eingeführten Lysine nicht die Orientierung haben, die aufgrund der Energieminimierung angenommen wird und so die Abstände zu ihrer komplementären
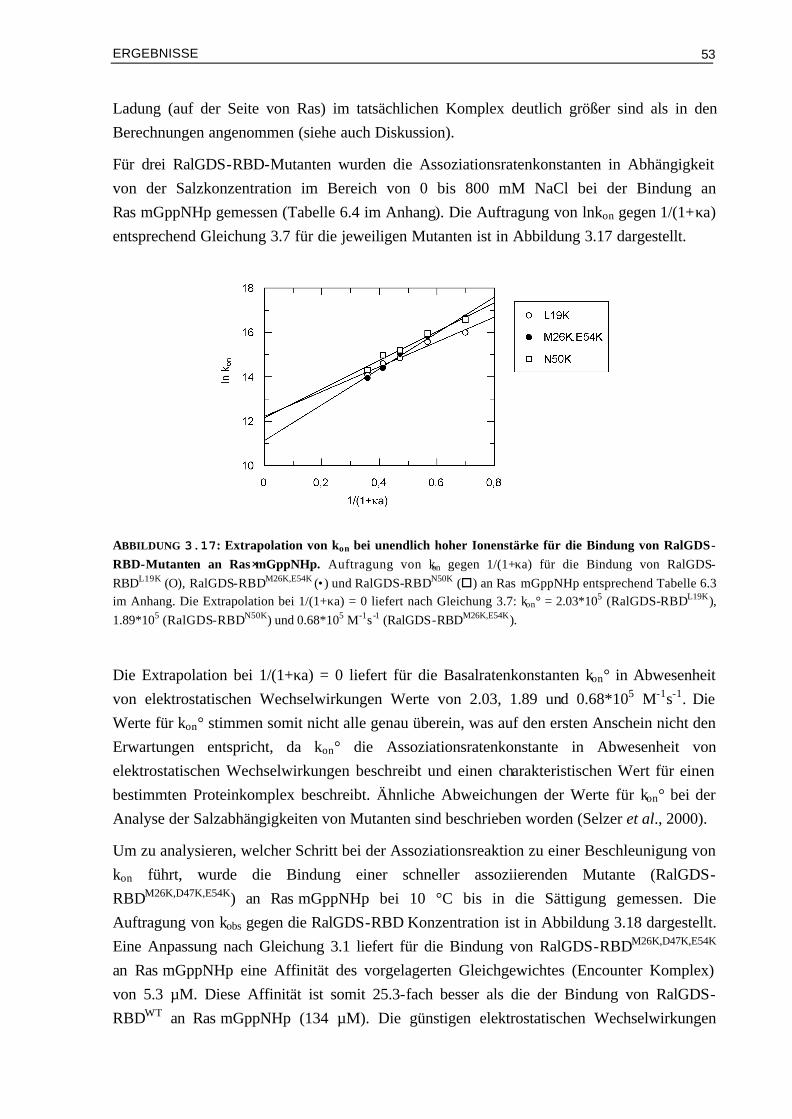
ERGEBNISSE
53
Ladung (auf der Seite von Ras) im tatsächlichen Komplex deutlich größer sind als in den Berechnungen angenommen (siehe auch Diskussion).
Für drei RalGDS-RBD-Mutanten wurden die Assoziationsratenkonstanten in Abhängigkeit von der Salzkonzentration im Bereich von 0 bis 800 mM NaCl bei der Bindung an Ras⋅mGppNHp gemessen (Tabelle 6.4 im Anhang). Die Auftragung von lnkon gegen 1/(1+κa) entsprechend Gleichung 3.7 für die jeweiligen Mutanten ist in Abbildung 3.17 dargestellt. ABBILDUNG 3.17: Extrapolation von kon bei unendlich hoher Ionenstärke für die Bindung von RalGDS-RBD-Mutanten an Ras ⋅mGppNHp. Auftragung von kon gegen 1/(1+κa) für die Bindung von RalGDS-RBDL19K (Ο), RalGDS-RBDM26K,E54K (•) und RalGDS-RBDN50K (o) an Ras ⋅mGppNHp entsprechend Tabelle 6.3 im Anhang. Die Extrapolation bei 1/(1+κa) = 0 liefert nach Gleichung 3.7: kon° = 2.03*105 (RalGDS-RBDL19K), 1.89*105 (RalGDS-RBDN50K) und 0.68*105 M-1s -1 (RalGDS-RBDM26K,E54K).
Die Extrapolation bei 1/(1+κa) = 0 liefert für die Basalratenkonstanten kon° in Abwesenheit von elektrostatischen Wechselwirkungen Werte von 2.03, 1.89 und 0.68*105 M-1s-1. Die Werte für kon° stimmen somit nicht alle genau überein, was auf den ersten Anschein nicht den Erwartungen entspricht, da kon° die Assoziationsratenkonstante in Abwesenheit von elektrostatischen Wechselwirkungen beschreibt und einen charakteristischen Wert für einen bestimmten Proteinkomplex beschreibt. Ähnliche Abweichungen der Werte für kon° bei der Analyse der Salzabhängigkeiten von Mutanten sind beschrieben worden (Selzer et al., 2000).
Um zu analysieren, welcher Schritt bei der Assoziationsreaktion zu einer Beschleunigung von kon führt, wurde die Bindung einer schneller assoziierenden Mutante (RalGDS-RBDM26K,D47K,E54K) an Ras⋅mGppNHp bei 10 °C bis in die Sättigung gemessen. Die Auftragung von kobs gegen die RalGDS-RBD Konzentration ist in Abbildung 3.18 dargestellt. Eine Anpassung nach Gleichung 3.1 liefert für die Bindung von RalGDS-RBDM26K,D47K,E54K
an Ras⋅mGppNHp eine Affinität des vorgelagerten Gleichgewichtes (Encounter Komplex) von 5.3 µM. Diese Affinität ist somit 25.3-fach besser als die der Bindung von RalGDS-RBDWT an Ras⋅mGppNHp (134 µM). Die günstigen elektrostatischen Wechselwirkungen
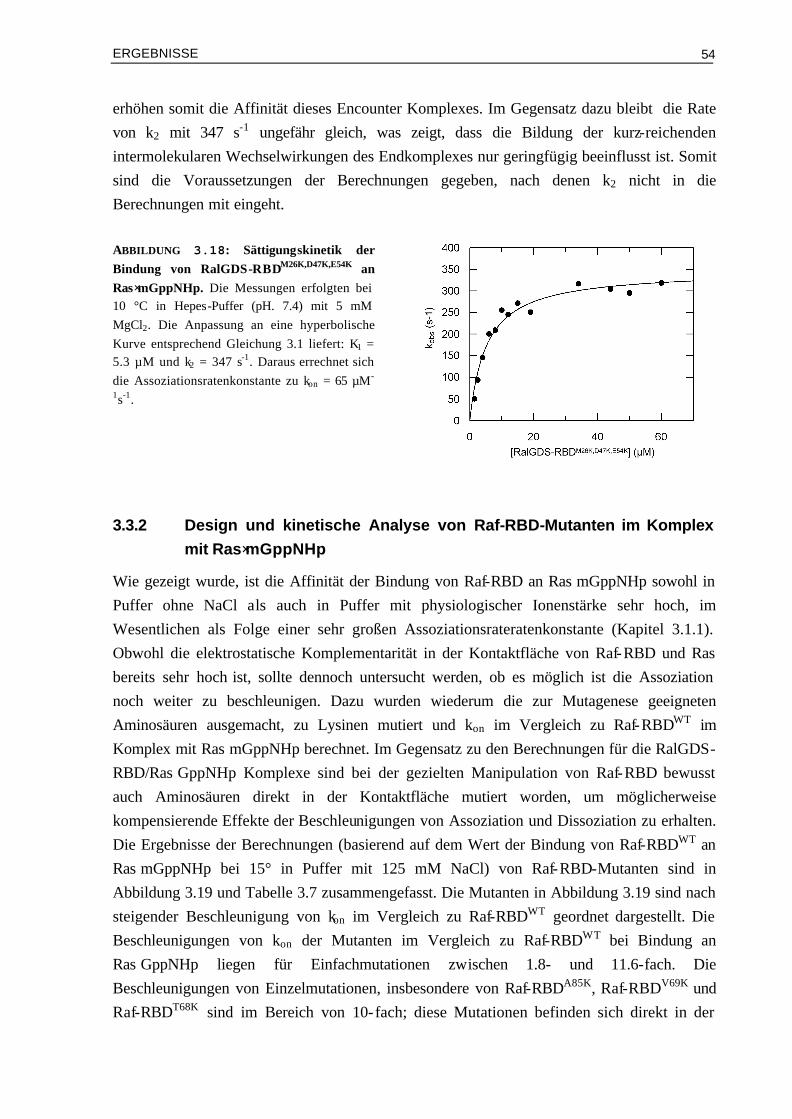
ERGEBNISSE
54
erhöhen somit die Affinität dieses Encounter Komplexes. Im Gegensatz dazu bleibt die Rate von k2 mit 347 s-1 ungefähr gleich, was zeigt, dass die Bildung der kurz-reichenden intermolekularen Wechselwirkungen des Endkomplexes nur geringfügig beeinflusst ist. Somit sind die Voraussetzungen der Berechnungen gegeben, nach denen k2 nicht in die Berechnungen mit eingeht.
ABBILDUNG 3.18: Sättigungskinetik der Bindung von RalGDS-RBDM26K,D47K,E54K an Ras⋅mGppNHp. Die Messungen erfolgten bei 10 °C in Hepes-Puffer (pH. 7.4) mit 5 mM MgCl2. Die Anpassung an eine hyperbolische Kurve entsprechend Gleichung 3.1 liefert: K1 = 5.3 µM und k2 = 347 s-1. Daraus errechnet sich die Assoziationsratenkonstante zu kon = 65 µM-
1s-1.
3.3.2 Design und kinetische Analyse von Raf-RBD-Mutanten im Komplex
mit Ras⋅mGppNHp
Wie gezeigt wurde, ist die Affinität der Bindung von Raf-RBD an Ras⋅mGppNHp sowohl in Puffer ohne NaCl als auch in Puffer mit physiologischer Ionenstärke sehr hoch, im Wesentlichen als Folge einer sehr großen Assoziationsrateratenkonstante (Kapitel 3.1.1). Obwohl die elektrostatische Komplementarität in der Kontaktfläche von Raf-RBD und Ras bereits sehr hoch ist, sollte dennoch untersucht werden, ob es möglich ist die Assoziation noch weiter zu beschleunigen. Dazu wurden wiederum die zur Mutagenese geeigneten Aminosäuren ausgemacht, zu Lysinen mutiert und kon im Vergleich zu Raf-RBDWT im Komplex mit Ras⋅mGppNHp berechnet. Im Gegensatz zu den Berechnungen für die RalGDS-RBD/Ras⋅GppNHp Komplexe sind bei der gezielten Manipulation von Raf-RBD bewusst auch Aminosäuren direkt in der Kontaktfläche mutiert worden, um möglicherweise kompensierende Effekte der Beschleunigungen von Assoziation und Dissoziation zu erhalten. Die Ergebnisse der Berechnungen (basierend auf dem Wert der Bindung von Raf-RBDWT an Ras⋅mGppNHp bei 15° in Puffer mit 125 mM NaCl) von Raf-RBD-Mutanten sind in Abbildung 3.19 und Tabelle 3.7 zusammengefasst. Die Mutanten in Abbildung 3.19 sind nach steigender Beschleunigung von kon im Vergleich zu Raf-RBDWT geordnet dargestellt. Die Beschleunigungen von kon der Mutanten im Vergleich zu Raf-RBDWT bei Bindung an Ras⋅GppNHp liegen für Einfachmutationen zwischen 1.8- und 11.6-fach. Die Beschleunigungen von Einzelmutationen, insbesondere von Raf-RBDA85K, Raf-RBDV69K und Raf-RBDT68K sind im Bereich von 10-fach; diese Mutationen befinden sich direkt in der
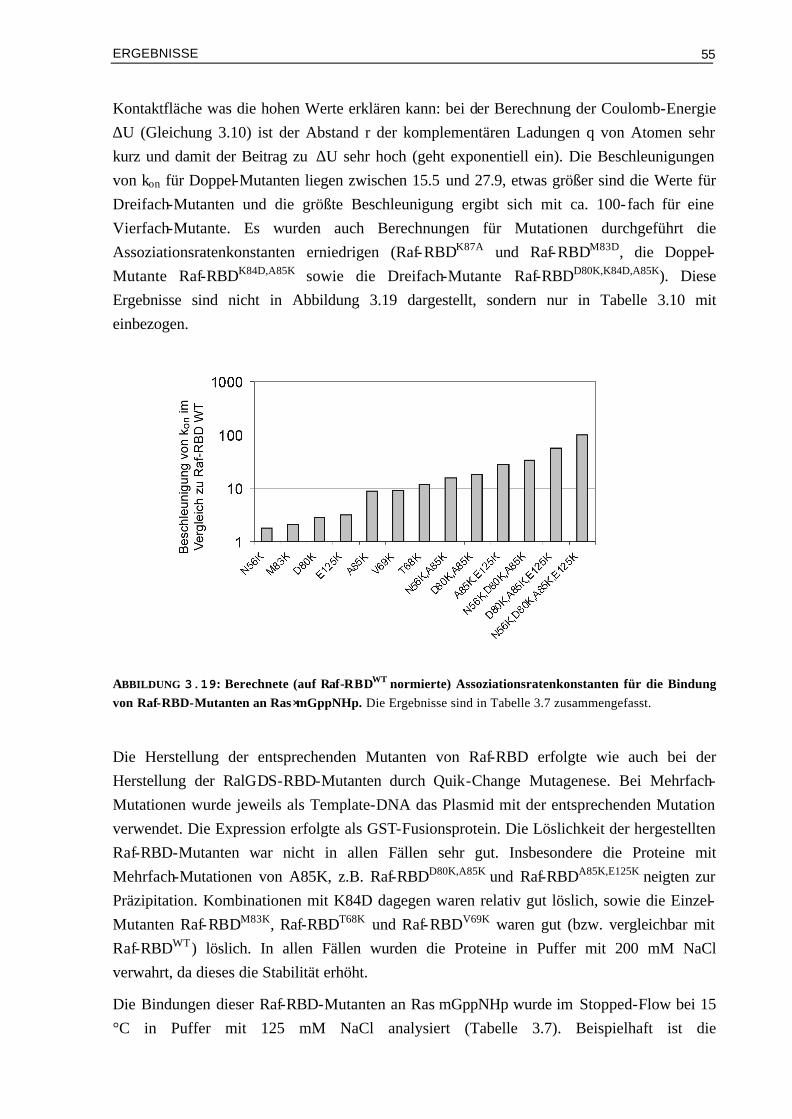
ERGEBNISSE
55
Kontaktfläche was die hohen Werte erklären kann: bei der Berechnung der Coulomb-Energie ∆U (Gleichung 3.10) ist der Abstand r der komplementären Ladungen q von Atomen sehr kurz und damit der Beitrag zu ∆U sehr hoch (geht exponentiell ein). Die Beschleunigungen von kon für Doppel-Mutanten liegen zwischen 15.5 und 27.9, etwas größer sind die Werte für Dreifach-Mutanten und die größte Beschleunigung ergibt sich mit ca. 100-fach für eine Vierfach-Mutante. Es wurden auch Berechnungen für Mutationen durchgeführt die Assoziationsratenkonstanten erniedrigen (Raf-RBDK87A und Raf-RBDM83D, die Doppel-Mutante Raf-RBDK84D,A85K sowie die Dreifach-Mutante Raf-RBDD80K,K84D,A85K). Diese Ergebnisse sind nicht in Abbildung 3.19 dargestellt, sondern nur in Tabelle 3.10 mit einbezogen.
ABBILDUNG 3.19: Berechnete (auf Raf-RBDWT normierte) Assoziationsratenkonstanten für die Bindung von Raf-RBD-Mutanten an Ras ⋅mGppNHp. Die Ergebnisse sind in Tabelle 3.7 zusammengefasst.
Die Herstellung der entsprechenden Mutanten von Raf-RBD erfolgte wie auch bei der Herstellung der RalGDS-RBD-Mutanten durch Quik-Change Mutagenese. Bei Mehrfach-Mutationen wurde jeweils als Template-DNA das Plasmid mit der entsprechenden Mutation verwendet. Die Expression erfolgte als GST-Fusionsprotein. Die Löslichkeit der hergestellten Raf-RBD-Mutanten war nicht in allen Fällen sehr gut. Insbesondere die Proteine mit Mehrfach-Mutationen von A85K, z.B. Raf-RBDD80K,A85K und Raf-RBDA85K,E125K neigten zur Präzipitation. Kombinationen mit K84D dagegen waren relativ gut löslich, sowie die Einzel-Mutanten Raf-RBDM83K, Raf-RBDT68K und Raf-RBDV69K waren gut (bzw. vergleichbar mit Raf-RBDWT) löslich. In allen Fällen wurden die Proteine in Puffer mit 200 mM NaCl verwahrt, da dieses die Stabilität erhöht.
Die Bindungen dieser Raf-RBD-Mutanten an Ras⋅mGppNHp wurde im Stopped-Flow bei 15 °C in Puffer mit 125 mM NaCl analysiert (Tabelle 3.7). Beispielhaft ist die
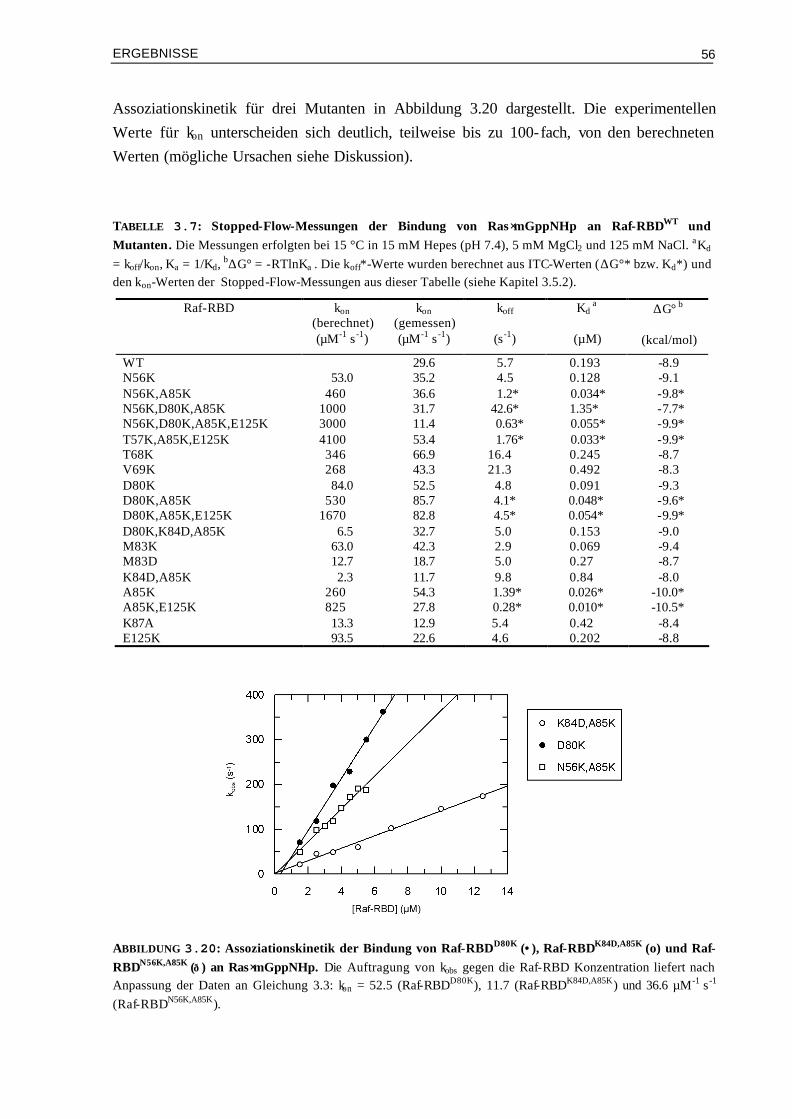
ERGEBNISSE
56
Assoziationskinetik für drei Mutanten in Abbildung 3.20 dargestellt. Die experimentellen Werte für kon unterscheiden sich deutlich, teilweise bis zu 100-fach, von den berechneten Werten (mögliche Ursachen siehe Diskussion).
TABELLE 3.7: Stopped-Flow-Messungen der Bindung von Ras ⋅mGppNHp an Raf-RBDWT und Mutanten. Die Messungen erfolgten bei 15 °C in 15 mM Hepes (pH 7.4), 5 mM MgCl2 und 125 mM NaCl. aKd = koff/kon, Ka = 1/Kd, b∆G° = -RTlnKa . Die koff*-Werte wurden berechnet aus ITC-Werten (∆G°* bzw. Kd*) und den kon-Werten der Stopped-Flow-Messungen aus dieser Tabelle (siehe Kapitel 3.5.2).
ABBILDUNG 3.20: Assoziationskinetik der Bindung von Raf-RBDD80K (•), Raf-RBDK84D,A85K (o) und Raf-RBDN56K,A85K (ð ) an Ras⋅mGppNHp. Die Auftragung von kobs gegen die Raf-RBD Konzentration liefert nach Anpassung der Daten an Gleichung 3.3: kon = 52.5 (Raf-RBDD80K), 11.7 (Raf-RBDK84D,A85K) und 36.6 µM-1 s -1 (Raf-RBDN56K,A85K).
Raf-RBD kon (berechnet) (µM-1 s -1)
kon (gemessen) (µM-1 s -1)
koff
(s-1)
Kd a
(µM)
∆G° b
(kcal/mol)
WT 29.6 5.7 0.193 -8.9 N56K 53.0 35.2 4.5 0.128 -9.1 N56K,A85K 460 36.6 1.2* 0.034* -9.8* N56K,D80K,A85K 1000 31.7 42.6* 1.35* -7.7* N56K,D80K,A85K,E125K 3000 11.4 0.63* 0.055* -9.9* T57K,A85K,E125K 4100 53.4 1.76* 0.033* -9.9* T68K 346 66.9 16.4 0.245 -8.7 V69K 268 43.3 21.3 0.492 -8.3 D80K 84.0 52.5 4.8 0.091 -9.3 D80K,A85K 530 85.7 4.1* 0.048* -9.6* D80K,A85K,E125K 1670 82.8 4.5* 0.054* -9.9* D80K,K84D,A85K 6.5 32.7 5.0 0.153 -9.0 M83K 63.0 42.3 2.9 0.069 -9.4 M83D 12.7 18.7 5.0 0.27 -8.7 K84D,A85K 2.3 11.7 9.8 0.84 -8.0 A85K 260 54.3 1.39* 0.026* -10.0* A85K,E125K 825 27.8 0.28* 0.010* -10.5* K87A 13.3 12.9 5.4 0.42 -8.4 E125K 93.5 22.6 4.6 0.202 -8.8
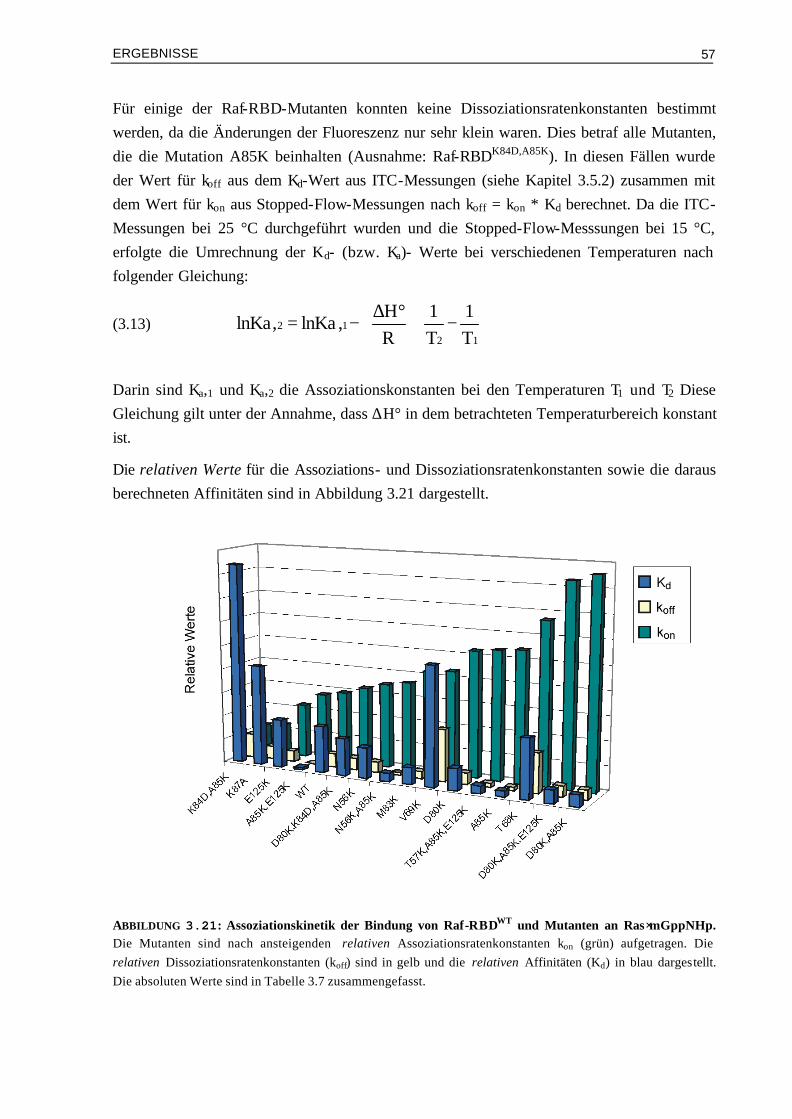
ERGEBNISSE
57
T1
T1
RH
,lnKa,lnKa12
12
−
°∆
−=
Für einige der Raf-RBD-Mutanten konnten keine Dissoziationsratenkonstanten bestimmt werden, da die Änderungen der Fluoreszenz nur sehr klein waren. Dies betraf alle Mutanten, die die Mutation A85K beinhalten (Ausnahme: Raf-RBDK84D,A85K). In diesen Fällen wurde der Wert für koff aus dem Kd-Wert aus ITC-Messungen (siehe Kapitel 3.5.2) zusammen mit dem Wert für kon aus Stopped-Flow-Messungen nach koff = kon * Kd berechnet. Da die ITC-Messungen bei 25 °C durchgeführt wurden und die Stopped-Flow-Messsungen bei 15 °C, erfolgte die Umrechnung der Kd- (bzw. Ka)- Werte bei verschiedenen Temperaturen nach folgender Gleichung: (3.13) Darin sind Ka,1 und Ka,2 die Assoziationskonstanten bei den Temperaturen T1 und T2 Diese Gleichung gilt unter der Annahme, dass ∆H° in dem betrachteten Temperaturbereich konstant ist.
Die relativen Werte für die Assoziations- und Dissoziationsratenkonstanten sowie die daraus berechneten Affinitäten sind in Abbildung 3.21 dargestellt.
ABBILDUNG 3.21: Assoziationskinetik der Bindung von Raf-RBDWT und Mutanten an Ras⋅mGppNHp. Die Mutanten sind nach ansteigenden relativen Assoziationsratenkonstanten kon (grün) aufgetragen. Die relativen Dissoziationsratenkonstanten (koff) sind in gelb und die relativen Affinitäten (Kd) in blau dargestellt. Die absoluten Werte sind in Tabelle 3.7 zusammengefasst.

ERGEBNISSE
58
Man erkennt, dass eine schnellere Assoziation mit einer besseren Affinität korreliert. Ausnahmen sind die Bindung von Raf-RBDT68K und Raf-RBDV69K, bei denen die schnellere Dissoziation die höhere Assoziationsratenkonstante kompensiert, wobei insgesamt die Affinität schlechter im Vergleich zu Raf-RBDWT wird. Die Mutanten Raf-RBDT68K und Raf-RBDV69K sind somit ein gutes Beispiel für sich kompensierende Werte für kon und koff und hier wird es interessant sein, wie die zelluläre Signalweiterleitung für diese Mutanten aussieht. Für die Mutanten Raf-RBDD80K und Raf-RBDM83K bewirkt eine größere Assoziationsratenkonstante zusammen mit einer geringfügig langsameren Dissoziation eine Verbesserung der Affinität. Raf-RBDA85K sowie Mehrfach-Mutationen in Kombination mit A85K zeigen eine sehr drastische Verlangsamung der Dissoziation im Bereich von 5- bis 20- fach im Vergleich zu Raf-RBDWT . Die Mutante Raf-RBDE125K sowie die Doppel-Mutante Raf-RBDA85K,E125K zeigen entgegen den Erwartungen basierend auf den Berechnungen einen kleineren Wert für kon.
In Tabelle 3.8 sind die Stopped-Flow-Ergebnisse der Bindung von einigen Raf-RBD-Mutanten an Ras⋅mGppNHpE31K dargestellt. Diese Messungen wurden im Wesentlichen deshalb durchgeführt, um Vergleiche mit der Situation in vivo machen zu können. (Daher sind auch keine Berechnungen von kon-Werten in der Tabelle angegeben).
TABELLE 3.8: Stopped-Flow-Messungen der Bindung von Ras ⋅mGppNHpWT und Ras ⋅mGppNHpE31K an Raf-RBDWT und Mutanten. Die Messungen erfolgten bei 15 °C in 15 mM Hepes (pH 7.4), 5mM MgCl2, und 125 mM NaCl. aKd = koff/kon, Ka = 1/Kd, b∆G° = -RTlnKa.
Ziel war es, eventuell Komplexe mit sehr langsamen Werten für koff zu erhalten. Wie in Kapitel 3.1.1 dargestellt, ist die Assoziationsratenkonstante der Bindung von Rap⋅mGppNHp an RalGDS-RBDWT sehr niedrig und die sehr hohe Affinität (vergleichbar mit der von Ras⋅mGppNHp an Raf-RBDWT) ist die Folge einer sehr langsamen Dissoziation des Komplexes. Es sollte versucht werden, Mutanten-Komplexe zwischen Raf-RBD und Ras zu finden, die ähnliche kinetische Parameter wie der Rap/RalGDS-RBD Komplex besitzen. Dazu wurde zum einen die Mutante von Ras Glu31 zu Lys gewählt, die Ras Rap-ähnlich macht. Zum anderen wurden die Mutanten M83D, K84D und A85K von Raf-RBD gewählt. Aufgrund von Strukturvergleichen könnte die Position Asp47/Lys48 von RalGDS-RBD den Aminosäuren Met83/Lys84 oder der Position Lys84/Ala85 entsprechen. Wie die Ergebnisse
Ras⋅mGppNHp
Raf-RBD
kon (µM-1s -1)
koff (s-1)
Kd a
(µM) ∆G° b
(kcal/mol)
WT WT 29.6 5.7 0.19 -8.9 WT M83D 18.7 5.0 0.27 -8.7 E31K WT 17.4 31.9 1.83 -7.8 E31K N56K,A85K 51.3 4.9 0.096 -9.3 E31K M83D 25.9 25.4 0.98 -7.9 E31K K84D,A85K 15.5 10.5 0.68 -8.4 E31K A85K 22.7 6.5 0.29 -8.9
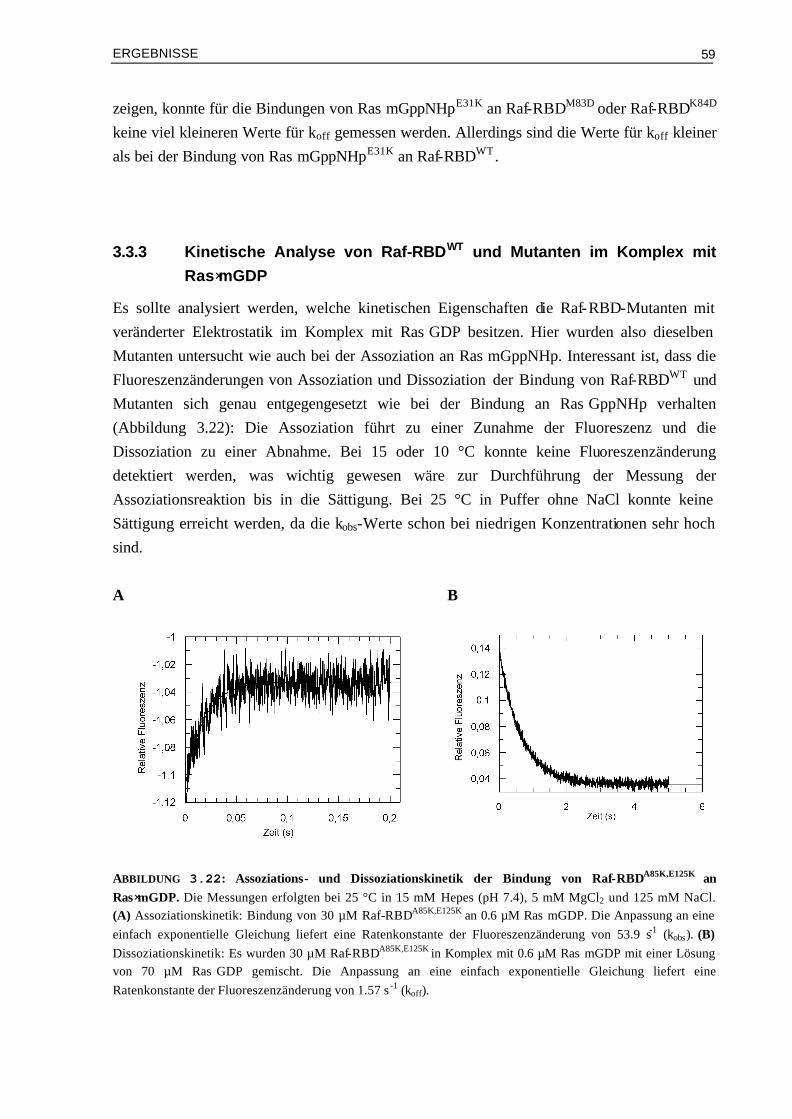
ERGEBNISSE
59
zeigen, konnte für die Bindungen von Ras⋅mGppNHpE31K an Raf-RBDM83D oder Raf-RBDK84D keine viel kleineren Werte für koff gemessen werden. Allerdings sind die Werte für koff kleiner als bei der Bindung von Ras⋅mGppNHpE31K an Raf-RBDWT .
3.3.3 Kinetische Analyse von Raf-RBDWT und Mutanten im Komplex mit
Ras⋅mGDP
Es sollte analysiert werden, welche kinetischen Eigenschaften die Raf-RBD-Mutanten mit veränderter Elektrostatik im Komplex mit Ras⋅GDP besitzen. Hier wurden also dieselben Mutanten untersucht wie auch bei der Assoziation an Ras⋅mGppNHp. Interessant ist, dass die Fluoreszenzänderungen von Assoziation und Dissoziation der Bindung von Raf-RBDWT und Mutanten sich genau entgegengesetzt wie bei der Bindung an Ras⋅GppNHp verhalten (Abbildung 3.22): Die Assoziation führt zu einer Zunahme der Fluoreszenz und die Dissoziation zu einer Abnahme. Bei 15 oder 10 °C konnte keine Fluoreszenzänderung detektiert werden, was wichtig gewesen wäre zur Durchführung der Messung der Assoziationsreaktion bis in die Sättigung. Bei 25 °C in Puffer ohne NaCl konnte keine Sättigung erreicht werden, da die kobs-Werte schon bei niedrigen Konzentrationen sehr hoch sind.
A B
ABBILDUNG 3.22: Assoziations- und Dissoziationskinetik der Bindung von Raf-RBDA85K,E125K an Ras⋅mGDP. Die Messungen erfolgten bei 25 °C in 15 mM Hepes (pH 7.4), 5 mM MgCl2 und 125 mM NaCl. (A) Assoziationskinetik: Bindung von 30 µM Raf-RBDA85K,E125K an 0.6 µM Ras⋅mGDP. Die Anpassung an eine einfach exponentielle Gleichung liefert eine Ratenkonstante der Fluoreszenzänderung von 53.9 s-1 (kobs). (B) Dissoziationskinetik: Es wurden 30 µM Raf-RBDA85K,E125K in Komplex mit 0.6 µM Ras⋅mGDP mit einer Lösung von 70 µM Ras ⋅GDP gemischt. Die Anpassung an eine einfach exponentielle Gleichung liefert eine Ratenkonstante der Fluoreszenzänderung von 1.57 s -1 (koff).
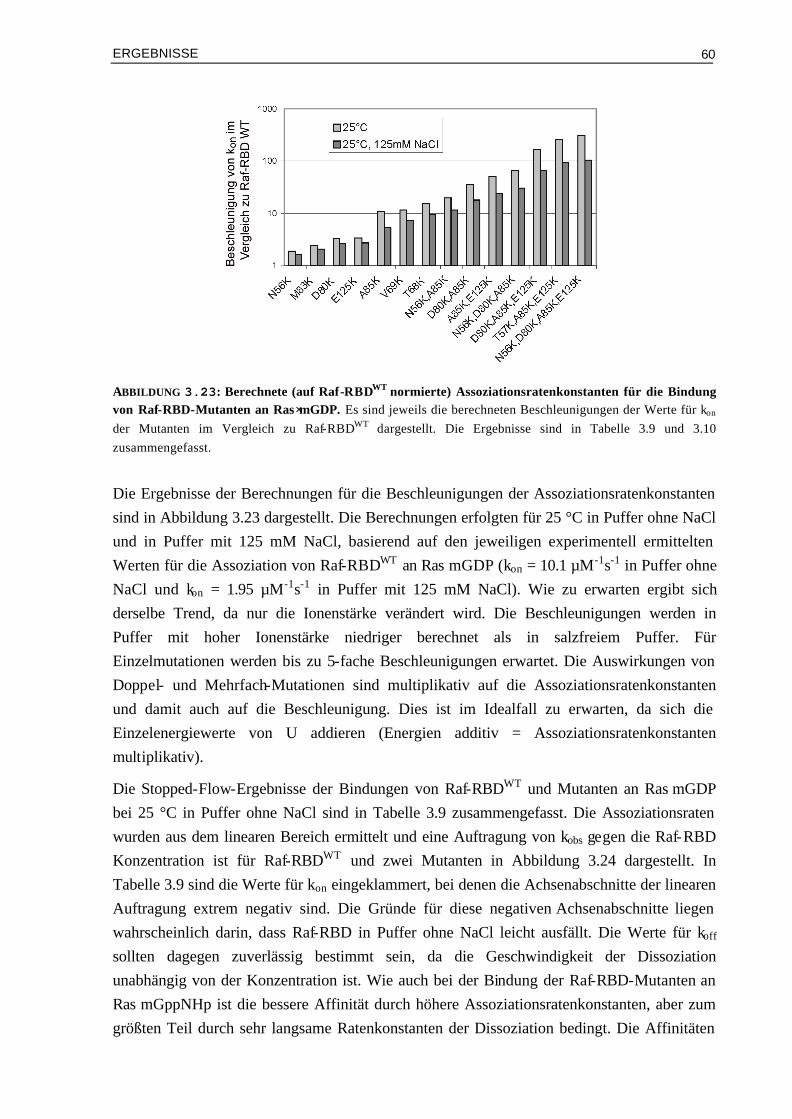
ERGEBNISSE
60
ABBILDUNG 3.23: Berechnete (auf Raf-RBDWT normierte) Assoziationsratenkonstanten für die Bindung von Raf-RBD-Mutanten an Ras ⋅mGDP. Es sind jeweils die berechneten Beschleunigungen der Werte für kon der Mutanten im Vergleich zu Raf-RBDWT dargestellt. Die Ergebnisse sind in Tabelle 3.9 und 3.10 zusammengefasst.
Die Ergebnisse der Berechnungen für die Beschleunigungen der Assoziationsratenkonstanten sind in Abbildung 3.23 dargestellt. Die Berechnungen erfolgten für 25 °C in Puffer ohne NaCl und in Puffer mit 125 mM NaCl, basierend auf den jeweiligen experimentell ermittelten Werten für die Assoziation von Raf-RBDWT an Ras⋅mGDP (kon = 10.1 µM-1s-1 in Puffer ohne NaCl und kon = 1.95 µM-1s-1 in Puffer mit 125 mM NaCl). Wie zu erwarten ergibt sich derselbe Trend, da nur die Ionenstärke verändert wird. Die Beschleunigungen werden in Puffer mit hoher Ionenstärke niedriger berechnet als in salzfreiem Puffer. Für Einzelmutationen werden bis zu 5-fache Beschleunigungen erwartet. Die Auswirkungen von Doppel- und Mehrfach-Mutationen sind multiplikativ auf die Assoziationsratenkonstanten und damit auch auf die Beschleunigung. Dies ist im Idealfall zu erwarten, da sich die Einzelenergiewerte von U addieren (Energien additiv = Assoziationsratenkonstanten multiplikativ).
Die Stopped-Flow-Ergebnisse der Bindungen von Raf-RBDWT und Mutanten an Ras⋅mGDP bei 25 °C in Puffer ohne NaCl sind in Tabelle 3.9 zusammengefasst. Die Assoziationsraten wurden aus dem linearen Bereich ermittelt und eine Auftragung von kobs gegen die Raf-RBD Konzentration ist für Raf-RBDWT und zwei Mutanten in Abbildung 3.24 dargestellt. In Tabelle 3.9 sind die Werte für kon eingeklammert, bei denen die Achsenabschnitte der linearen Auftragung extrem negativ sind. Die Gründe für diese negativen Achsenabschnitte liegen wahrscheinlich darin, dass Raf-RBD in Puffer ohne NaCl leicht ausfällt. Die Werte für koff sollten dagegen zuverlässig bestimmt sein, da die Geschwindigkeit der Dissoziation unabhängig von der Konzentration ist. Wie auch bei der Bindung der Raf-RBD-Mutanten an Ras⋅mGppNHp ist die bessere Affinität durch höhere Assoziationsratenkonstanten, aber zum größten Teil durch sehr langsame Ratenkonstanten der Dissoziation bedingt. Die Affinitäten
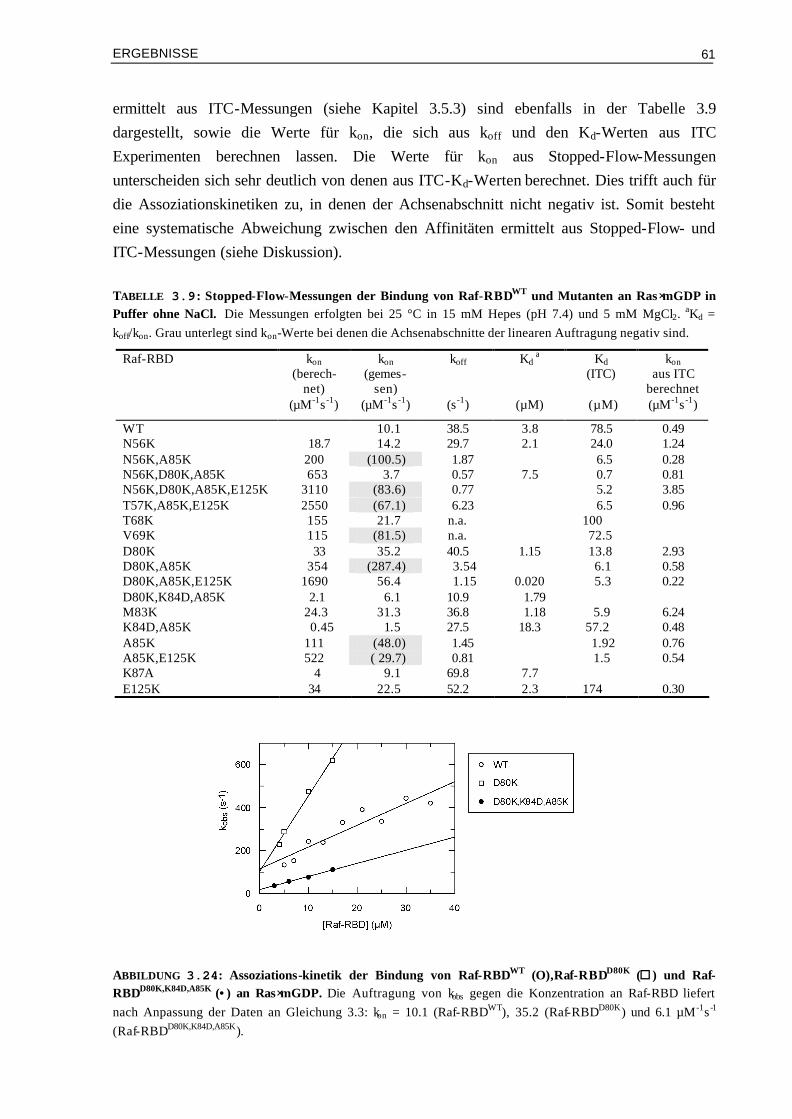
ERGEBNISSE
61
ermittelt aus ITC-Messungen (siehe Kapitel 3.5.3) sind ebenfalls in der Tabelle 3.9 dargestellt, sowie die Werte für kon, die sich aus koff und den Kd-Werten aus ITC Experimenten berechnen lassen. Die Werte für kon aus Stopped-Flow-Messungen unterscheiden sich sehr deutlich von denen aus ITC-Kd-Werten berechnet. Dies trifft auch für die Assoziationskinetiken zu, in denen der Achsenabschnitt nicht negativ ist. Somit besteht eine systematische Abweichung zwischen den Affinitäten ermittelt aus Stopped-Flow- und ITC-Messungen (siehe Diskussion). TABELLE 3.9: Stopped-Flow-Messungen der Bindung von Raf-RBDWT und Mutanten an Ras ⋅mGDP in Puffer ohne NaCl. Die Messungen erfolgten bei 25 °C in 15 mM Hepes (pH 7.4) und 5 mM MgCl2. aKd = koff/kon. Grau unterlegt sind kon-Werte bei denen die Achsenabschnitte der linearen Auftragung negativ sind.
ABBILDUNG 3.24: Assoziations-kinetik der Bindung von Raf-RBDWT (Ο),Raf-RBDD80K (o ) und Raf-RBDD80K,K84D,A85K (•) an Ras⋅mGDP. Die Auftragung von kobs gegen die Konzentration an Raf-RBD liefert nach Anpassung der Daten an Gleichung 3.3: kon = 10.1 (Raf-RBDWT), 35.2 (Raf-RBDD80K) und 6.1 µM-1s -1 (Raf-RBDD80K,K84D,A85K).
Raf-RBD kon (berech-
net) (µM-1s -1)
kon (gemes-
sen) (µM-1s -1)
koff
(s-1)
Kd a
(µM)
Kd (ITC)
(µM)
kon aus ITC
berechnet (µM-1s -1)
WT 10.1 38.5 3.8 78.5 0.49 N56K 18.7 14.2 29.7 2.1 24.0 1.24 N56K,A85K 200 (100.5) 1.87 6.5 0.28 N56K,D80K,A85K 653 3.7 0.57 7.5 0.7 0.81 N56K,D80K,A85K,E125K 3110 (83.6) 0.77 5.2 3.85 T57K,A85K,E125K 2550 (67.1) 6.23 6.5 0.96 T68K 155 21.7 n.a. 100 V69K 115 (81.5) n.a. 72.5 D80K 33 35.2 40.5 1.15 13.8 2.93 D80K,A85K 354 (287.4) 3.54 6.1 0.58 D80K,A85K,E125K 1690 56.4 1.15 0.020 5.3 0.22 D80K,K84D,A85K 2.1 6.1 10.9 1.79 M83K 24.3 31.3 36.8 1.18 5.9 6.24 K84D,A85K 0.45 1.5 27.5 18.3 57.2 0.48 A85K 111 (48.0) 1.45 1.92 0.76 A85K,E125K 522 ( 29.7) 0.81 1.5 0.54 K87A 4 9.1 69.8 7.7 E125K 34 22.5 52.2 2.3 174 0.30
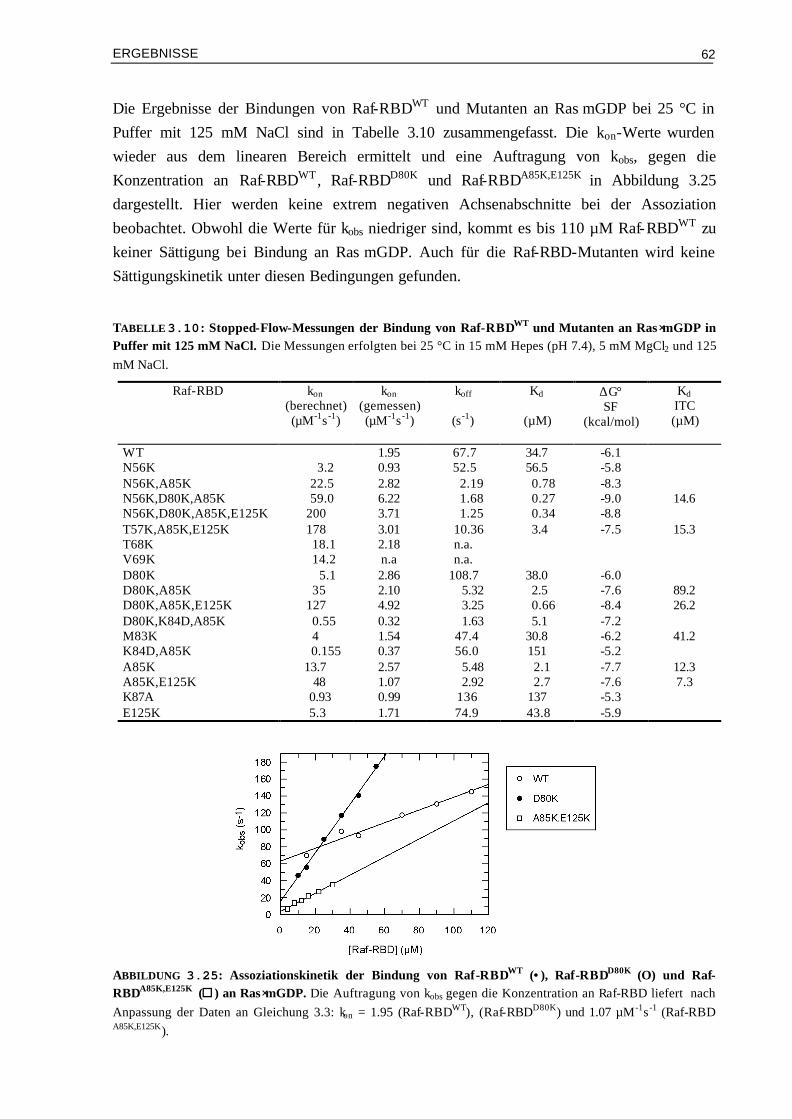
ERGEBNISSE
62
Die Ergebnisse der Bindungen von Raf-RBDWT und Mutanten an Ras⋅mGDP bei 25 °C in Puffer mit 125 mM NaCl sind in Tabelle 3.10 zusammengefasst. Die kon-Werte wurden wieder aus dem linearen Bereich ermittelt und eine Auftragung von kobs, gegen die Konzentration an Raf-RBDWT , Raf-RBDD80K und Raf-RBDA85K,E125K in Abbildung 3.25 dargestellt. Hier werden keine extrem negativen Achsenabschnitte bei der Assoziation beobachtet. Obwohl die Werte für kobs niedriger sind, kommt es bis 110 µM Raf-RBDWT zu keiner Sättigung bei Bindung an Ras⋅mGDP. Auch für die Raf-RBD-Mutanten wird keine Sättigungskinetik unter diesen Bedingungen gefunden.
TABELLE 3.10: Stopped-Flow-Messungen der Bindung von Raf-RBDWT und Mutanten an Ras ⋅mGDP in Puffer mit 125 mM NaCl. Die Messungen erfolgten bei 25 °C in 15 mM Hepes (pH 7.4), 5 mM MgCl2 und 125 mM NaCl.
ABBILDUNG 3.25: Assoziationskinetik der Bindung von Raf-RBDWT (•), Raf-RBDD80K (Ο) und Raf-RBDA85K,E125K (o ) an Ras⋅mGDP. Die Auftragung von kobs gegen die Konzentration an Raf-RBD liefert nach Anpassung der Daten an Gleichung 3.3: kon = 1.95 (Raf-RBDWT), (Raf-RBDD80K) und 1.07 µM-1s -1 (Raf-RBD A85K,E125K).
Raf-RBD kon (berechnet) (µM-1s -1)
kon (gemessen) (µM-1s -1)
koff
(s-1)
Kd
(µM)
∆G° SF
(kcal/mol)
Kd ITC (µM)
WT 1.95 67.7 34.7 -6.1 N56K 3.2 0.93 52.5 56.5 -5.8 N56K,A85K 22.5 2.82 2.19 0.78 -8.3 N56K,D80K,A85K 59.0 6.22 1.68 0.27 -9.0 14.6 N56K,D80K,A85K,E125K 200 3.71 1.25 0.34 -8.8 T57K,A85K,E125K 178 3.01 10.36 3.4 -7.5 15.3 T68K 18.1 2.18 n.a. V69K 14.2 n.a n.a. D80K 5.1 2.86 108.7 38.0 -6.0 D80K,A85K 35 2.10 5.32 2.5 -7.6 89.2 D80K,A85K,E125K 127 4.92 3.25 0.66 -8.4 26.2 D80K,K84D,A85K 0.55 0.32 1.63 5.1 -7.2 M83K 4 1.54 47.4 30.8 -6.2 41.2 K84D,A85K 0.155 0.37 56.0 151 -5.2 A85K 13.7 2.57 5.48 2.1 -7.7 12.3 A85K,E125K 48 1.07 2.92 2.7 -7.6 7.3 K87A 0.93 0.99 136 137 -5.3 E125K 5.3 1.71 74.9 43.8 -5.9

ERGEBNISSE
63
Wie auch bei der Bindung der Mutanten in Puffer ohne NaCl ist die bessere Affinität hauptsächlich die Folge von sehr kleinen Dissoziationsratenkonstanten. Der Vergleich mit Kd-Werten aus ITC-Messungen zeigt sehr deutliche Abweichungen von den Kd-Werten aus Stopped-Flow-Messungen. Es besteht keine systematische Abweichung, sondern diese sind abhängig von der jeweiligen Mutante und unterscheiden sind bis zu 50-fach. Für manche Mutanten (Raf-RBDM83K, Raf-RBDA85K und Raf-RBDA85K,E125K) stimmen die Werte allerdings gut überein und unterscheiden sich nur ca. 2-fach.
3.4 Vergleich von experimentellen und berechneten kon-Werten
Der Vergleich von experimentellen und berechneten Werten für kon dient zur Analyse inwieweit die Berechnungen, basierend auf dem Comupterprogramm PARE, auch anwendbar auf das Ras-Effektor System sind. Dazu wurden die logarithmierten kon-Werte, erhalten aus Berechnungen und aus Stopped-Flow-Experimenten, gegeneinander aufgetragen. Anschließend wurden die Daten einer linearen Gleichung angepasst und daraus die Steigung sowie der Korrelationskoeffizient als Maß für die Übereinstimmung erhalten. Die Auftragung von logarithmierten Werten für die Assoziationsratenkonstanten ist sinnvoll, da die Berechnungen auf der Bestimmung von Energiewerten beruhen und sich Assoziationsratenkonstanten exponentiell zu Energiewerten verhalten.
3.4.1 Bindung von RalGDS-RBD an Ras⋅mGppNHp
In Abbildung 3.26 ist eine Korrelation der experimentellen und berechneten Assoziationsratenkonstanten dargestellt von RalGDS-RBDWT und Mutanten bei der Bindung an Ras und Rap⋅mGppNHp und Mutanten. In Abbildung 3.26-A handelt es sich dabei um die Mutanten von RalGDS-RBD und Ras, die bereits aus früheren Mutationsstudien vorhanden waren und hauptsächlich in der Kontaktfläche lokalisiert sind (siehe Kapitel 3.2.1). Die Übereinstimmung von experimentellen und berechneten Assoziationsraten ist mit einem Korrelationskoeffizienten von 0.9 sehr gut. Allerdings beträgt die Steigung nur 0.71. Es ist ein Trend zu erkennen, bei dem langsame Werte für kon zu schnelleren kon-Werten führen als die Berechnungen vorhersagen und schnellere kon-Werte werden langsamer gemessen als die berechneten Raten. Die Beziehung zwischen berechneten und experimentell ermittelten kon-Werten für neu hergestellte Mutanten von RalGDS-RBD, die sich hauptsächlich am Rand der Kontaktfläche befinden, ist in Abbildung 3.26-B dargestellt (siehe Kapitel 3.3.1). Es besteht eine sehr gute Übereinstimmung mit einem Korrelationskoeffizienten von 0.91. Dabei weichen ebenfalls hauptsächlich zwei und mehrfach Mutanten zum Teil stark von den berechneten Daten ab. Dabei ist die Steigung wiederum kleiner als 1 (0.66). Die kon-Wert für RalGDS-RBDD47K,E52K,E53K wurden nicht mit in die Korrelation einbezogen.

ERGEBNISSE
64
A B
ABBILDUNG 3.26: Bindung von RalGDS-RBDWT und Mutanten an Ras ⋅mGppNHp: Korrelation von log kon (berechnet) und log kon (experimentell). (A) Assoziation von RalGDS-RBDWT und Mutanten an Ras⋅mGppNHp und Mutanten (n) und an Rap⋅mGppNHp (o). Der Korrelationskoeffizient beträgt 0.90 und die Steigung 0.71. Die Werte wurden Tabelle 3.4 entnommen. (B) Bindung von RalGDS-RBDWT und Mutanten an Ras⋅mGppNHp entsprechend Tabelle 3.6. Der Korrelationskoeffizient beträgt 0.91 und die Steigung 0.66. Die gestrichelte Linie entspricht einer Geraden mit der Steigung 1.
3.4.2 Bindung von Raf-RBD-Mutanten an Ras⋅mGppNHp
In Abbildung 3.27-A ist eine Korrelation der experimentellen und berechneten Assoziationsraten für die Bindung von Raf-RBDWT und Mutanten an Ras⋅mGppNHp dargestellt. Hierbei handelt es sich um die Ergebnisse der Mutanten, die bereits aus Mutationsstudien vorhanden waren und hauptsächlich Mutationen von postiv geladenen Resten zu neutralen und negativen Seitenketten sind (siehe Kapitel 3.2.2). Die Übereinstimmung ist mit einem Korrelationkoeffizienten von 0.73 nicht so gut wie für die Bindung von RalGDS-RBD-Mutanten an Ras⋅mGppNHp. Es ist kein Unterschied für die Übereinstimmung zwischen experimentellen und berechneten Werten für kon bei 25 °C in Puffer ohne Salz und bei 15 °C in Puffer mit 125 mM NaCl zu erkennen. In Abbildung 3.27-B ist die Korrelation der experimentellen und berechneten Assoziationsratenkonstanten für die Bindung von Raf-RBDWT und neu hergestellte Mutanten dargestellt (siehe Kapitel 3.3.2). Der Korrelationskoeffizient ist mit 0.68 schlecht, insbesondere beträgt die Steigung aber nur 0.19. Der schwarz markierte Datenpunkt wurde nicht mit in den Vergleich einbezogen. Während die experimentellen Werte für kon der Raf-RBD-Mutanten mit Einzelmutationen relativ gut übereinstimmen, ergeben sich deutliche Abweichungen für Raf-RBD-Mutanten Komplex mit Zwei- und Mehrfach-Mutanten, insbesondere bei Raf-RBD-Mutanten die die Mutation A85K beinhalten (Raf-RBDN56K,A85K , Raf-RBDN56K,A85K, Raf-RBDN56K,D80K,A85K, Raf-RBDN56K,D80K,A85K,E125K, Raf-RBDT57K,A85K,E125K, Raf-RBDD80K,A85K, Raf-RBDD80K, Raf-
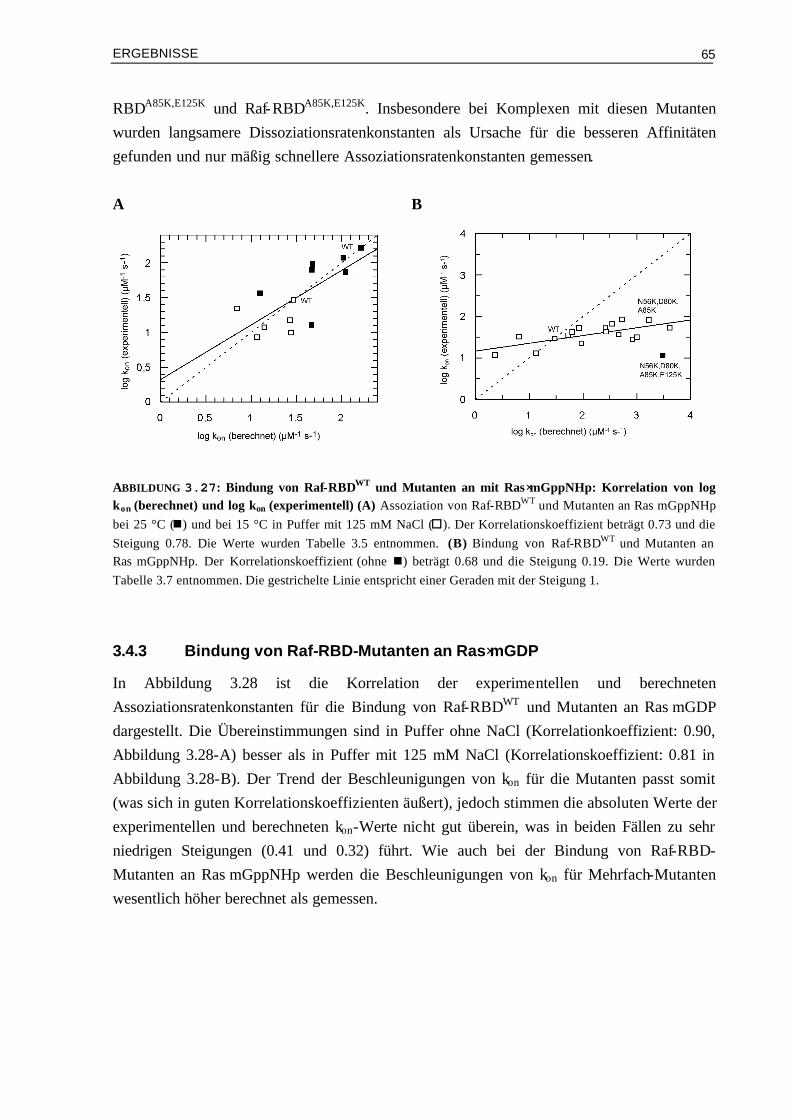
ERGEBNISSE
65
RBDA85K,E125K und Raf-RBDA85K,E125K. Insbesondere bei Komplexen mit diesen Mutanten wurden langsamere Dissoziationsratenkonstanten als Ursache für die besseren Affinitäten gefunden und nur mäßig schnellere Assoziationsratenkonstanten gemessen. A B
ABBILDUNG 3.27: Bindung von Raf-RBDWT und Mutanten an mit Ras⋅mGppNHp: Korrelation von log kon (berechnet) und log kon (experimentell) (A) Assoziation von Raf-RBDWT und Mutanten an Ras⋅mGppNHp bei 25 °C (n) und bei 15 °C in Puffer mit 125 mM NaCl (o). Der Korrelationskoeffizient beträgt 0.73 und die Steigung 0.78. Die Werte wurden Tabelle 3.5 entnommen. (B) Bindung von Raf-RBDWT und Mutanten an Ras⋅mGppNHp. Der Korrelationskoeffizient (ohne n) beträgt 0.68 und die Steigung 0.19. Die Werte wurden Tabelle 3.7 entnommen. Die gestrichelte Linie entspricht einer Geraden mit der Steigung 1.
3.4.3 Bindung von Raf-RBD-Mutanten an Ras⋅mGDP
In Abbildung 3.28 ist die Korrelation der experimentellen und berechneten Assoziationsratenkonstanten für die Bindung von Raf-RBDWT und Mutanten an Ras⋅mGDP dargestellt. Die Übereinstimmungen sind in Puffer ohne NaCl (Korrelationkoeffizient: 0.90, Abbildung 3.28-A) besser als in Puffer mit 125 mM NaCl (Korrelationskoeffizient: 0.81 in Abbildung 3.28-B). Der Trend der Beschleunigungen von kon für die Mutanten passt somit (was sich in guten Korrelationskoeffizienten äußert), jedoch stimmen die absoluten Werte der experimentellen und berechneten kon-Werte nicht gut überein, was in beiden Fällen zu sehr niedrigen Steigungen (0.41 und 0.32) führt. Wie auch bei der Bindung von Raf-RBD-Mutanten an Ras⋅mGppNHp werden die Beschleunigungen von kon für Mehrfach-Mutanten wesentlich höher berechnet als gemessen.
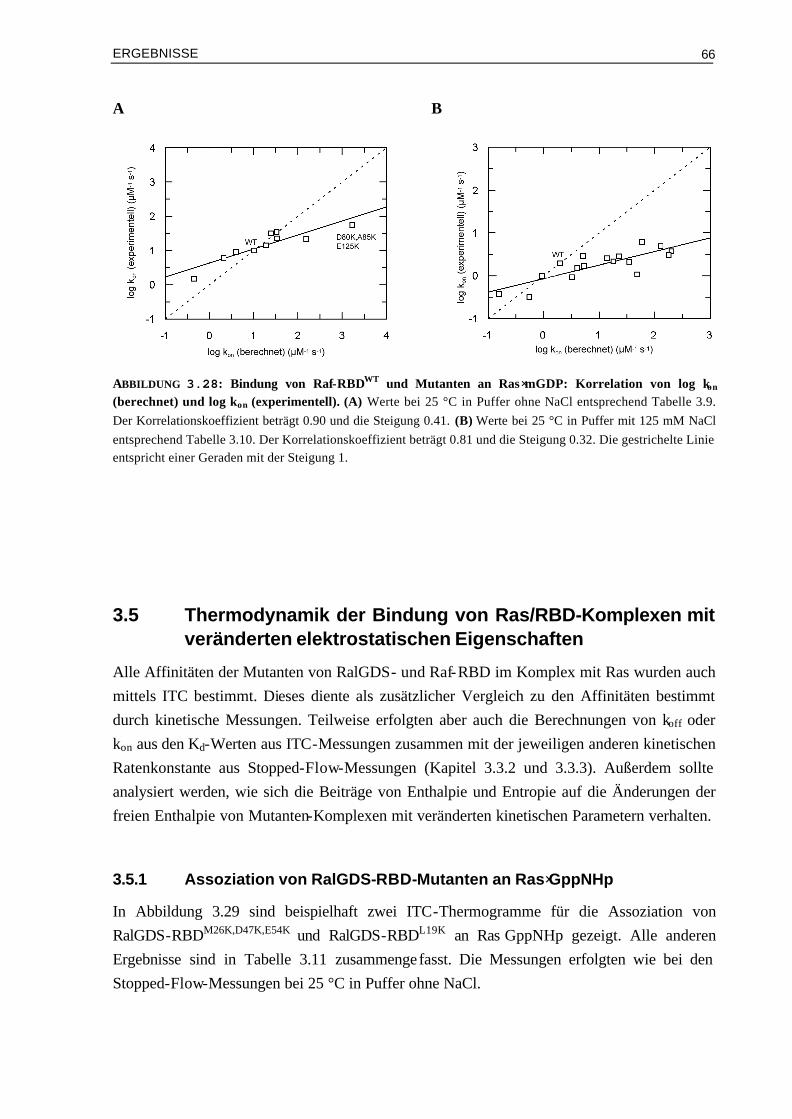
ERGEBNISSE
66
A B
ABBILDUNG 3.28: Bindung von Raf-RBDWT und Mutanten an Ras ⋅mGDP: Korrelation von log kon (berechnet) und log kon (experimentell). (A) Werte bei 25 °C in Puffer ohne NaCl entsprechend Tabelle 3.9. Der Korrelationskoeffizient beträgt 0.90 und die Steigung 0.41. (B) Werte bei 25 °C in Puffer mit 125 mM NaCl entsprechend Tabelle 3.10. Der Korrelationskoeffizient beträgt 0.81 und die Steigung 0.32. Die gestrichelte Linie entspricht einer Geraden mit der Steigung 1.
3.5 Thermodynamik der Bindung von Ras/RBD-Komplexen mit veränderten elektrostatischen Eigenschaften
Alle Affinitäten der Mutanten von RalGDS- und Raf-RBD im Komplex mit Ras wurden auch mittels ITC bestimmt. Dieses diente als zusätzlicher Vergleich zu den Affinitäten bestimmt durch kinetische Messungen. Teilweise erfolgten aber auch die Berechnungen von koff oder kon aus den Kd-Werten aus ITC-Messungen zusammen mit der jeweiligen anderen kinetischen Ratenkonstante aus Stopped-Flow-Messungen (Kapitel 3.3.2 und 3.3.3). Außerdem sollte analysiert werden, wie sich die Beiträge von Enthalpie und Entropie auf die Änderungen der freien Enthalpie von Mutanten-Komplexen mit veränderten kinetischen Parametern verhalten.
3.5.1 Assoziation von RalGDS-RBD-Mutanten an Ras⋅GppNHp
In Abbildung 3.29 sind beispielhaft zwei ITC-Thermogramme für die Assoziation von RalGDS-RBDM26K,D47K,E54K und RalGDS-RBDL19K an Ras⋅GppNHp gezeigt. Alle anderen Ergebnisse sind in Tabelle 3.11 zusammengefasst. Die Messungen erfolgten wie bei den Stopped-Flow-Messungen bei 25 °C in Puffer ohne NaCl.
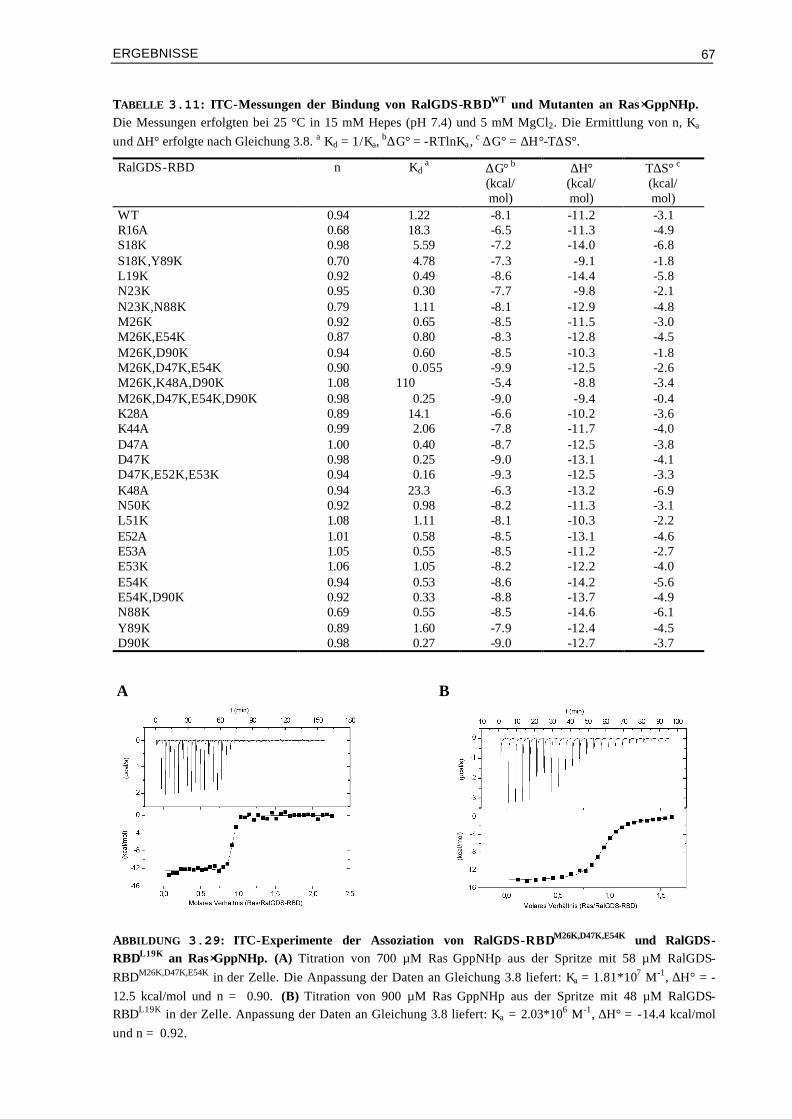
ERGEBNISSE
67
TABELLE 3.11: ITC-Messungen der Bindung von RalGDS-RBDWT und Mutanten an Ras ⋅GppNHp. Die Messungen erfolgten bei 25 °C in 15 mM Hepes (pH 7.4) und 5 mM MgCl2. Die Ermittlung von n, Ka und ∆H° erfolgte nach Gleichung 3.8. a Kd = 1/Ka,
b∆G° = -RTlnKa , c ∆G° = ∆H°-T∆S°.
A B
ABBILDUNG 3.29: ITC-Experimente der Assoziation von RalGDS-RBDM26K,D47K,E54K und RalGDS-RBDL19K an Ras⋅GppNHp. (A) Titration von 700 µM Ras ⋅GppNHp aus der Spritze mit 58 µM RalGDS-RBDM26K,D47K,E54K in der Zelle. Die Anpassung der Daten an Gleichung 3.8 liefert: Ka = 1.81*107 M-1, ∆H° = -12.5 kcal/mol und n = 0.90. (B) Titration von 900 µM Ras ⋅GppNHp aus der Spritze mit 48 µM RalGDS-RBDL19K in der Zelle. Anpassung der Daten an Gleichung 3.8 liefert: Ka = 2.03*106 M-1, ∆H° = -14.4 kcal/mol und n = 0.92.
RalGDS-RBD n Kd a
∆G° b (kcal/ mol)
∆H° (kcal/ mol)
T∆S° c (kcal/ mol)
WT 0.94 1.22 -8.1 -11.2 -3.1 R16A 0.68 18.3 -6.5 -11.3 -4.9 S18K 0.98 5.59 -7.2 -14.0 -6.8 S18K,Y89K 0.70 4.78 -7.3 -9.1 -1.8 L19K 0.92 0.49 -8.6 -14.4 -5.8 N23K 0.95 0.30 -7.7 -9.8 -2.1 N23K,N88K 0.79 1.11 -8.1 -12.9 -4.8 M26K 0.92 0.65 -8.5 -11.5 -3.0 M26K,E54K 0.87 0.80 -8.3 -12.8 -4.5 M26K,D90K 0.94 0.60 -8.5 -10.3 -1.8 M26K,D47K,E54K 0.90 0.055 -9.9 -12.5 -2.6 M26K,K48A,D90K 1.08 110 -5.4 -8.8 -3.4 M26K,D47K,E54K,D90K 0.98 0.25 -9.0 -9.4 -0.4 K28A 0.89 14.1 -6.6 -10.2 -3.6 K44A 0.99 2.06 -7.8 -11.7 -4.0 D47A 1.00 0.40 -8.7 -12.5 -3.8 D47K 0.98 0.25 -9.0 -13.1 -4.1 D47K,E52K,E53K 0.94 0.16 -9.3 -12.5 -3.3 K48A 0.94 23.3 -6.3 -13.2 -6.9 N50K 0.92 0.98 -8.2 -11.3 -3.1 L51K 1.08 1.11 -8.1 -10.3 -2.2 E52A 1.01 0.58 -8.5 -13.1 -4.6 E53A 1.05 0.55 -8.5 -11.2 -2.7 E53K 1.06 1.05 -8.2 -12.2 -4.0 E54K 0.94 0.53 -8.6 -14.2 -5.6 E54K,D90K 0.92 0.33 -8.8 -13.7 -4.9 N88K 0.69 0.55 -8.5 -14.6 -6.1 Y89K 0.89 1.60 -7.9 -12.4 -4.5 D90K 0.98 0.27 -9.0 -12.7 -3.7
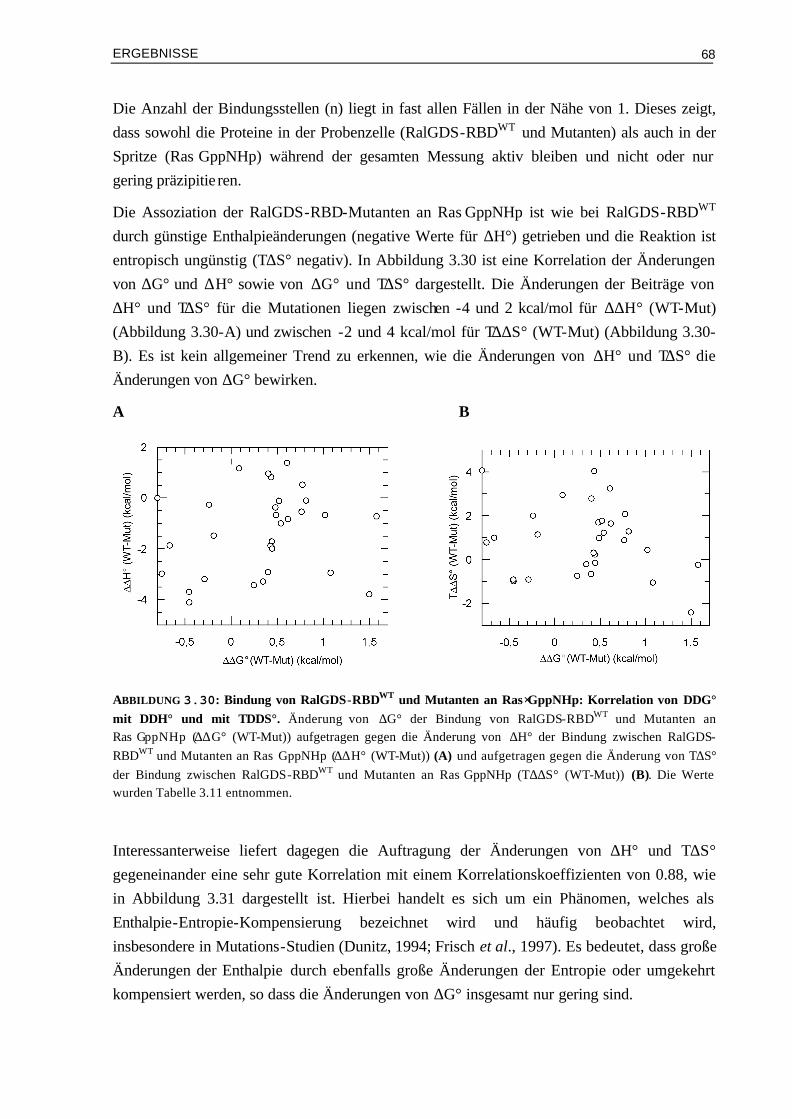
ERGEBNISSE
68
Die Anzahl der Bindungsstellen (n) liegt in fast allen Fällen in der Nähe von 1. Dieses zeigt, dass sowohl die Proteine in der Probenzelle (RalGDS-RBDWT und Mutanten) als auch in der Spritze (Ras⋅GppNHp) während der gesamten Messung aktiv bleiben und nicht oder nur gering präzipitie ren.
Die Assoziation der RalGDS-RBD-Mutanten an Ras⋅GppNHp ist wie bei RalGDS-RBDWT durch günstige Enthalpieänderungen (negative Werte für ∆H°) getrieben und die Reaktion ist entropisch ungünstig (T∆S° negativ). In Abbildung 3.30 ist eine Korrelation der Änderungen von ∆G° und ∆H° sowie von ∆G° und T∆S° dargestellt. Die Änderungen der Beiträge von ∆H° und T∆S° für die Mutationen liegen zwischen -4 und 2 kcal/mol für ∆∆H° (WT-Mut) (Abbildung 3.30-A) und zwischen -2 und 4 kcal/mol für T∆∆S° (WT-Mut) (Abbildung 3.30-B). Es ist kein allgemeiner Trend zu erkennen, wie die Änderungen von ∆H° und T∆S° die Änderungen von ∆G° bewirken.
A B
ABBILDUNG 3.30: Bindung von RalGDS-RBDWT und Mutanten an Ras⋅GppNHp: Korrelation von ∆∆G° mit ∆∆H° und mit T∆∆S°. Änderung von ∆G° der Bindung von RalGDS-RBDWT und Mutanten an Ras⋅GppNHp (∆∆G° (WT-Mut)) aufgetragen gegen die Änderung von ∆H° der Bindung zwischen RalGDS-RBDWT und Mutanten an Ras ⋅GppNHp (∆∆H° (WT-Mut)) (A) und aufgetragen gegen die Änderung von T∆S° der Bindung zwischen RalGDS-RBDWT und Mutanten an Ras ⋅GppNHp (T∆∆S° (WT-Mut)) (B). Die Werte wurden Tabelle 3.11 entnommen.
Interessanterweise liefert dagegen die Auftragung der Änderungen von ∆H° und T∆S° gegeneinander eine sehr gute Korrelation mit einem Korrelationskoeffizienten von 0.88, wie in Abbildung 3.31 dargestellt ist. Hierbei handelt es sich um ein Phänomen, welches als Enthalpie-Entropie-Kompensierung bezeichnet wird und häufig beobachtet wird, insbesondere in Mutations-Studien (Dunitz, 1994; Frisch et al., 1997). Es bedeutet, dass große Änderungen der Enthalpie durch ebenfalls große Änderungen der Entropie oder umgekehrt kompensiert werden, so dass die Änderungen von ∆G° insgesamt nur gering sind.
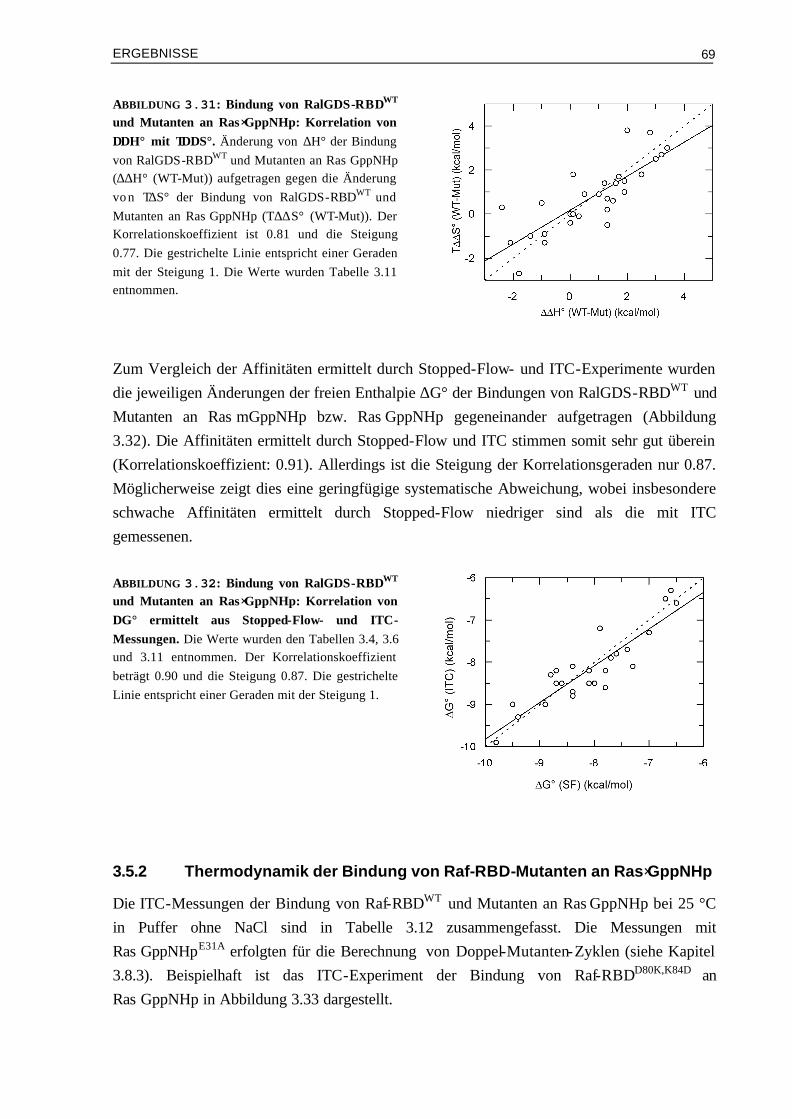
ERGEBNISSE
69
ABBILDUNG 3.31: Bindung von RalGDS-RBDWT und Mutanten an Ras⋅GppNHp: Korrelation von ∆∆H° mit T∆∆S°. Änderung von ∆H° der Bindung von RalGDS-RBDWT und Mutanten an Ras ⋅GppNHp (∆∆H° (WT-Mut)) aufgetragen gegen die Änderung vo n T∆S° der Bindung von RalGDS-RBDWT und Mutanten an Ras ⋅GppNHp (T∆∆S° (WT-Mut)). Der Korrelationskoeffizient ist 0.81 und die Steigung 0.77. Die gestrichelte Linie entspricht einer Geraden mit der Steigung 1. Die Werte wurden Tabelle 3.11 entnommen. Zum Vergleich der Affinitäten ermittelt durch Stopped-Flow- und ITC-Experimente wurden die jeweiligen Änderungen der freien Enthalpie ∆G° der Bindungen von RalGDS-RBDWT und Mutanten an Ras⋅mGppNHp bzw. Ras⋅GppNHp gegeneinander aufgetragen (Abbildung 3.32). Die Affinitäten ermittelt durch Stopped-Flow und ITC stimmen somit sehr gut überein (Korrelationskoeffizient: 0.91). Allerdings ist die Steigung der Korrelationsgeraden nur 0.87. Möglicherweise zeigt dies eine geringfügige systematische Abweichung, wobei insbesondere schwache Affinitäten ermittelt durch Stopped-Flow niedriger sind als die mit ITC gemessenen.
ABBILDUNG 3.32: Bindung von RalGDS-RBDWT und Mutanten an Ras⋅GppNHp: Korrelation von ∆G° ermittelt aus Stopped-Flow- und ITC-Messungen. Die Werte wurden den Tabellen 3.4, 3.6 und 3.11 entnommen. Der Korrelationskoeffizient beträgt 0.90 und die Steigung 0.87. Die gestrichelte Linie entspricht einer Geraden mit der Steigung 1.
3.5.2 Thermodynamik der Bindung von Raf-RBD-Mutanten an Ras⋅GppNHp
Die ITC-Messungen der Bindung von Raf-RBDWT und Mutanten an Ras⋅GppNHp bei 25 °C in Puffer ohne NaCl sind in Tabelle 3.12 zusammengefasst. Die Messungen mit Ras⋅GppNHpE31A erfolgten für die Berechnung von Doppel-Mutanten-Zyklen (siehe Kapitel 3.8.3). Beispielhaft ist das ITC-Experiment der Bindung von Raf-RBDD80K,K84D an Ras⋅GppNHp in Abbildung 3.33 dargestellt.
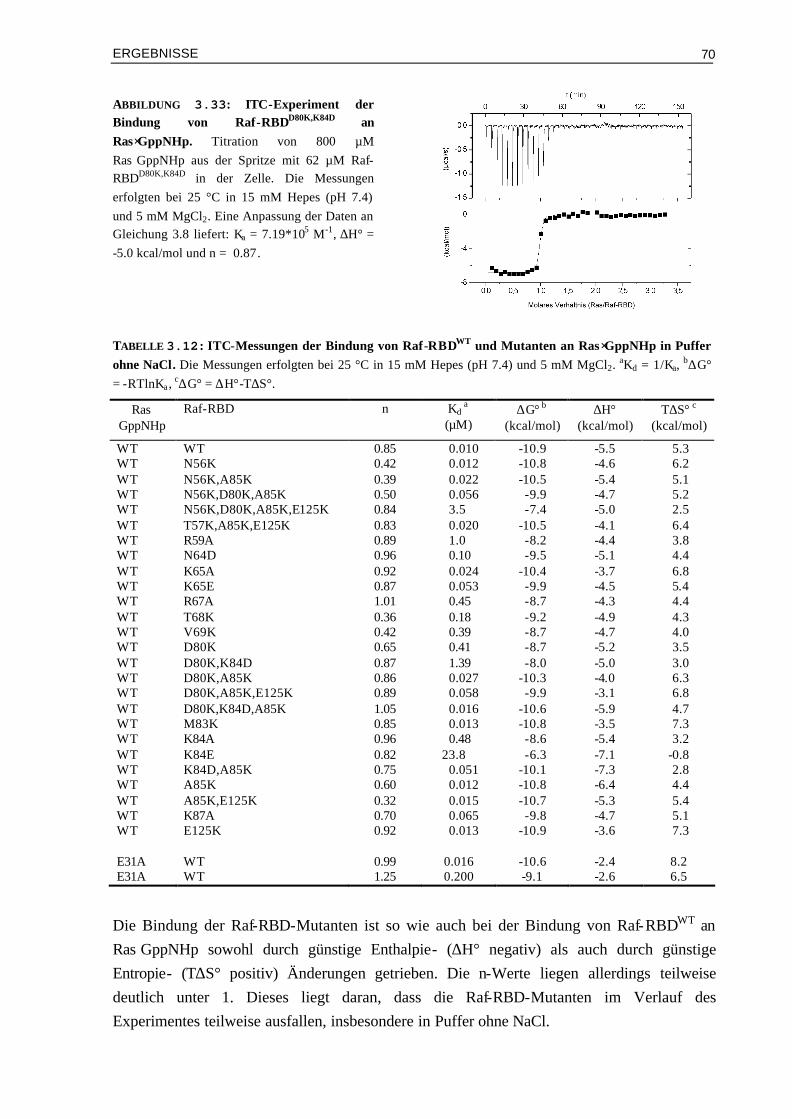
ERGEBNISSE
70
ABBILDUNG 3.33: ITC-Experiment der Bindung von Raf-RBDD80K,K84D an Ras⋅GppNHp. Titration von 800 µM Ras⋅GppNHp aus der Spritze mit 62 µM Raf-RBDD80K,K84D in der Zelle. Die Messungen erfolgten bei 25 °C in 15 mM Hepes (pH 7.4) und 5 mM MgCl2. Eine Anpassung der Daten an Gleichung 3.8 liefert: Ka = 7.19*105 M-1, ∆H° = -5.0 kcal/mol und n = 0.87.
TABELLE 3.12: ITC-Messungen der Bindung von Raf-RBDWT und Mutanten an Ras ⋅GppNHp in Puffer ohne NaCl. Die Messungen erfolgten bei 25 °C in 15 mM Hepes (pH 7.4) und 5 mM MgCl2. aKd = 1/Ka,
b∆G° = -RTlnKa , c∆G° = ∆H°-T∆S°.
Die Bindung der Raf-RBD-Mutanten ist so wie auch bei der Bindung von Raf-RBDWT an Ras⋅GppNHp sowohl durch günstige Enthalpie- (∆H° negativ) als auch durch günstige Entropie- (T∆S° positiv) Änderungen getrieben. Die n-Werte liegen allerdings teilweise deutlich unter 1. Dieses liegt daran, dass die Raf-RBD-Mutanten im Verlauf des Experimentes teilweise ausfallen, insbesondere in Puffer ohne NaCl.
Ras⋅ GppNHp
Raf-RBD n Kd a
(µM) ∆G° b
(kcal/mol) ∆H°
(kcal/mol) T∆S° c
(kcal/mol)
WT WT 0.85 0.010 -10.9 -5.5 5.3 WT N56K 0.42 0.012 -10.8 -4.6 6.2 WT N56K,A85K 0.39 0.022 -10.5 -5.4 5.1 WT N56K,D80K,A85K 0.50 0.056 -9.9 -4.7 5.2 WT N56K,D80K,A85K,E125K 0.84 3.5 -7.4 -5.0 2.5 WT T57K,A85K,E125K 0.83 0.020 -10.5 -4.1 6.4 WT R59A 0.89 1.0 -8.2 -4.4 3.8 WT N64D 0.96 0.10 -9.5 -5.1 4.4 WT K65A 0.92 0.024 -10.4 -3.7 6.8 WT K65E 0.87 0.053 -9.9 -4.5 5.4 WT R67A 1.01 0.45 -8.7 -4.3 4.4 WT T68K 0.36 0.18 -9.2 -4.9 4.3 WT V69K 0.42 0.39 -8.7 -4.7 4.0 WT D80K 0.65 0.41 -8.7 -5.2 3.5 WT D80K,K84D 0.87 1.39 -8.0 -5.0 3.0 WT D80K,A85K 0.86 0.027 -10.3 -4.0 6.3 WT D80K,A85K,E125K 0.89 0.058 -9.9 -3.1 6.8 WT D80K,K84D,A85K 1.05 0.016 -10.6 -5.9 4.7 WT M83K 0.85 0.013 -10.8 -3.5 7.3 WT K84A 0.96 0.48 -8.6 -5.4 3.2 WT K84E 0.82 23.8 -6.3 -7.1 -0.8 WT K84D,A85K 0.75 0.051 -10.1 -7.3 2.8 WT A85K 0.60 0.012 -10.8 -6.4 4.4 WT A85K,E125K 0.32 0.015 -10.7 -5.3 5.4 WT K87A 0.70 0.065 -9.8 -4.7 5.1 WT E125K 0.92 0.013 -10.9 -3.6 7.3 E31A WT 0.99 0.016 -10.6 -2.4 8.2 E31A WT 1.25 0.200 -9.1 -2.6 6.5
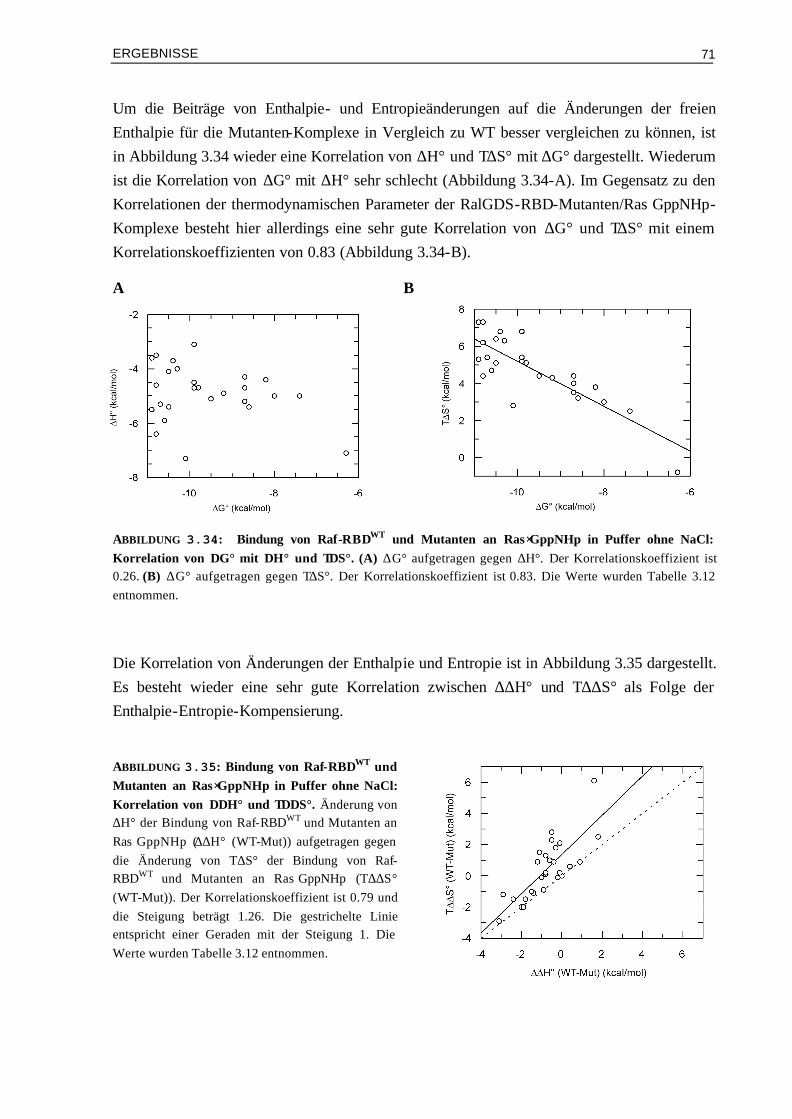
ERGEBNISSE
71
Um die Beiträge von Enthalpie- und Entropieänderungen auf die Änderungen der freien Enthalpie für die Mutanten-Komplexe in Vergleich zu WT besser vergleichen zu können, ist in Abbildung 3.34 wieder eine Korrelation von ∆H° und T∆S° mit ∆G° dargestellt. Wiederum ist die Korrelation von ∆G° mit ∆H° sehr schlecht (Abbildung 3.34-A). Im Gegensatz zu den Korrelationen der thermodynamischen Parameter der RalGDS-RBD-Mutanten/Ras⋅GppNHp-Komplexe besteht hier allerdings eine sehr gute Korrelation von ∆G° und T∆S° mit einem Korrelationskoeffizienten von 0.83 (Abbildung 3.34-B). A B
ABBILDUNG 3.34: Bindung von Raf-RBDWT und Mutanten an Ras ⋅GppNHp in Puffer ohne NaCl: Korrelation von ∆G° mit ∆H° und T∆S°. (A) ∆G° aufgetragen gegen ∆H°. Der Korrelationskoeffizient ist 0.26. (B) ∆G° aufgetragen gegen T∆S°. Der Korrelationskoeffizient ist 0.83. Die Werte wurden Tabelle 3.12 entnommen.
Die Korrelation von Änderungen der Enthalpie und Entropie ist in Abbildung 3.35 dargestellt. Es besteht wieder eine sehr gute Korrelation zwischen ∆∆H° und T∆∆S° als Folge der Enthalpie-Entropie-Kompensierung. ABBILDUNG 3.35: Bindung von Raf-RBDWT und Mutanten an Ras⋅GppNHp in Puffer ohne NaCl: Korrelation von ∆∆H° und T∆∆S°. Änderung von ∆H° der Bindung von Raf-RBDWT und Mutanten an Ras⋅GppNHp (∆∆H° (WT-Mut)) aufgetragen gegen die Änderung von T∆S° der Bindung von Raf-RBDWT und Mutanten an Ras ⋅GppNHp (T∆∆S° (WT-Mut)). Der Korrelationskoeffizient ist 0.79 und die Steigung beträgt 1.26. Die gestrichelte Linie entspricht einer Geraden mit der Steigung 1. Die Werte wurden Tabelle 3.12 entnommen.
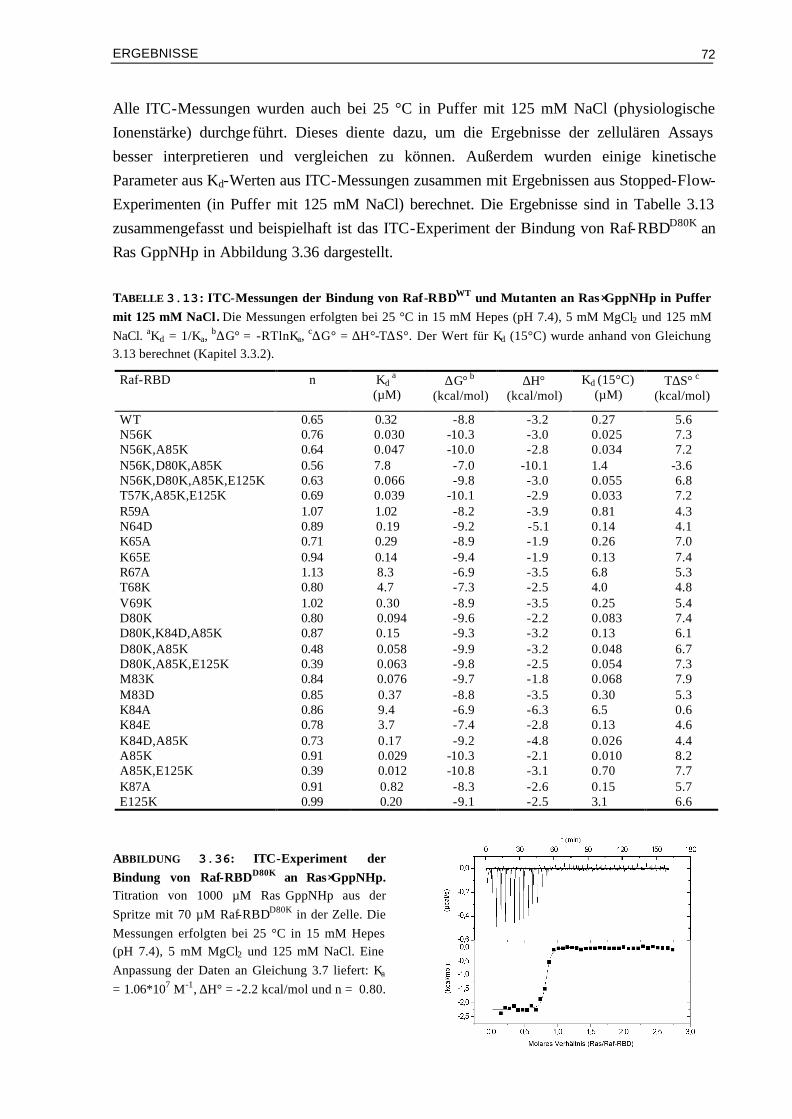
ERGEBNISSE
72
Alle ITC-Messungen wurden auch bei 25 °C in Puffer mit 125 mM NaCl (physiologische Ionenstärke) durchge führt. Dieses diente dazu, um die Ergebnisse der zellulären Assays besser interpretieren und vergleichen zu können. Außerdem wurden einige kinetische Parameter aus Kd-Werten aus ITC-Messungen zusammen mit Ergebnissen aus Stopped-Flow-Experimenten (in Puffer mit 125 mM NaCl) berechnet. Die Ergebnisse sind in Tabelle 3.13 zusammengefasst und beispielhaft ist das ITC-Experiment der Bindung von Raf-RBDD80K an Ras⋅GppNHp in Abbildung 3.36 dargestellt.
TABELLE 3.13: ITC-Messungen der Bindung von Raf-RBDWT und Mutanten an Ras ⋅GppNHp in Puffer mit 125 mM NaCl. Die Messungen erfolgten bei 25 °C in 15 mM Hepes (pH 7.4), 5 mM MgCl2 und 125 mM NaCl. aKd = 1/Ka,
b∆G° = -RTlnKa, c∆G° = ∆H°-T∆S°. Der Wert für Kd (15°C) wurde anhand von Gleichung
3.13 berechnet (Kapitel 3.3.2).
ABBILDUNG 3.36: ITC-Experiment der Bindung von Raf-RBDD80K an Ras⋅GppNHp. Titration von 1000 µM Ras ⋅GppNHp aus der Spritze mit 70 µM Raf-RBDD80K in der Zelle. Die Messungen erfolgten bei 25 °C in 15 mM Hepes (pH 7.4), 5 mM MgCl2 und 125 mM NaCl. Eine Anpassung der Daten an Gleichung 3.7 liefert: Ka = 1.06*107 M-1, ∆H° = -2.2 kcal/mol und n = 0.80.
Raf-RBD n Kd a
(µM) ∆G° b
(kcal/mol) ∆H°
(kcal/mol) Kd (15°C)
(µM) T∆S° c
(kcal/mol)
WT 0.65 0.32 -8.8 -3.2 0.27 5.6 N56K 0.76 0.030 -10.3 -3.0 0.025 7.3 N56K,A85K 0.64 0.047 -10.0 -2.8 0.034 7.2 N56K,D80K,A85K 0.56 7.8 -7.0 -10.1 1.4 -3.6 N56K,D80K,A85K,E125K 0.63 0.066 -9.8 -3.0 0.055 6.8 T57K,A85K,E125K 0.69 0.039 -10.1 -2.9 0.033 7.2 R59A 1.07 1.02 -8.2 -3.9 0.81 4.3 N64D 0.89 0.19 -9.2 -5.1 0.14 4.1 K65A 0.71 0.29 -8.9 -1.9 0.26 7.0 K65E 0.94 0.14 -9.4 -1.9 0.13 7.4 R67A 1.13 8.3 -6.9 -3.5 6.8 5.3 T68K 0.80 4.7 -7.3 -2.5 4.0 4.8 V69K 1.02 0.30 -8.9 -3.5 0.25 5.4 D80K 0.80 0.094 -9.6 -2.2 0.083 7.4 D80K,K84D,A85K 0.87 0.15 -9.3 -3.2 0.13 6.1 D80K,A85K 0.48 0.058 -9.9 -3.2 0.048 6.7 D80K,A85K,E125K 0.39 0.063 -9.8 -2.5 0.054 7.3 M83K 0.84 0.076 -9.7 -1.8 0.068 7.9 M83D 0.85 0.37 -8.8 -3.5 0.30 5.3 K84A 0.86 9.4 -6.9 -6.3 6.5 0.6 K84E 0.78 3.7 -7.4 -2.8 0.13 4.6 K84D,A85K 0.73 0.17 -9.2 -4.8 0.026 4.4 A85K 0.91 0.029 -10.3 -2.1 0.010 8.2 A85K,E125K 0.39 0.012 -10.8 -3.1 0.70 7.7 K87A 0.91 0.82 -8.3 -2.6 0.15 5.7 E125K 0.99 0.20 -9.1 -2.5 3.1 6.6
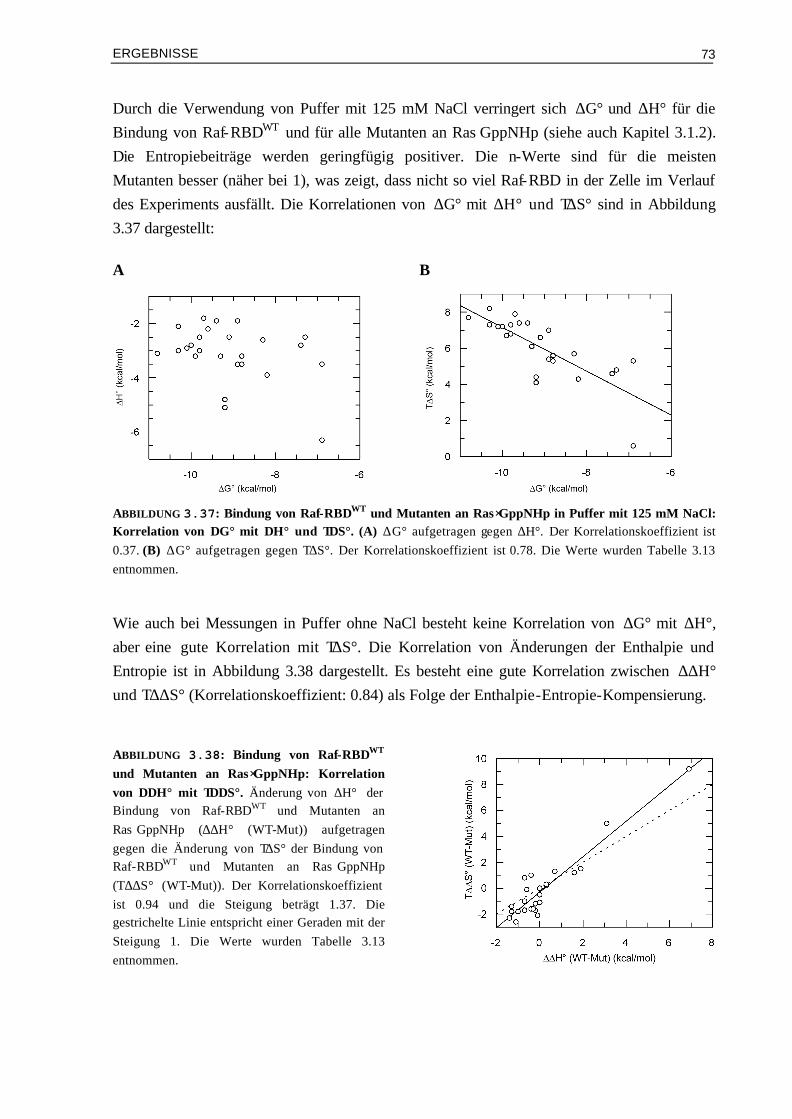
ERGEBNISSE
73
Durch die Verwendung von Puffer mit 125 mM NaCl verringert sich ∆G° und ∆H° für die Bindung von Raf-RBDWT und für alle Mutanten an Ras⋅GppNHp (siehe auch Kapitel 3.1.2). Die Entropiebeiträge werden geringfügig positiver. Die n-Werte sind für die meisten Mutanten besser (näher bei 1), was zeigt, dass nicht so viel Raf-RBD in der Zelle im Verlauf des Experiments ausfällt. Die Korrelationen von ∆G° mit ∆H° und T∆S° sind in Abbildung 3.37 dargestellt: A B
ABBILDUNG 3.37: Bindung von Raf-RBDWT und Mutanten an Ras⋅GppNHp in Puffer mit 125 mM NaCl: Korrelation von ∆G° mit ∆H° und T∆S°. (A) ∆G° aufgetragen gegen ∆H°. Der Korrelationskoeffizient ist 0.37. (B) ∆G° aufgetragen gegen T∆S°. Der Korrelationskoeffizient ist 0.78. Die Werte wurden Tabelle 3.13 entnommen.
Wie auch bei Messungen in Puffer ohne NaCl besteht keine Korrelation von ∆G° mit ∆H°, aber eine gute Korrelation mit T∆S°. Die Korrelation von Änderungen der Enthalpie und Entropie ist in Abbildung 3.38 dargestellt. Es besteht eine gute Korrelation zwischen ∆∆H° und T∆∆S° (Korrelationskoeffizient: 0.84) als Folge der Enthalpie-Entropie-Kompensierung.
ABBILDUNG 3.38: Bindung von Raf-RBDWT und Mutanten an Ras⋅GppNHp: Korrelation von ∆∆H° mit T∆∆S°. Änderung von ∆H° der Bindung von Raf-RBDWT und Mutanten an Ras⋅GppNHp (∆∆H° (WT-Mut)) aufgetragen gegen die Änderung von T∆S° der Bindung von Raf-RBDWT und Mutanten an Ras ⋅GppNHp (T∆∆S° (WT-Mut)). Der Korrelationskoeffizient ist 0.94 und die Steigung beträgt 1.37. Die gestrichelte Linie entspricht einer Geraden mit der Steigung 1. Die Werte wurden Tabelle 3.13 entnommen.
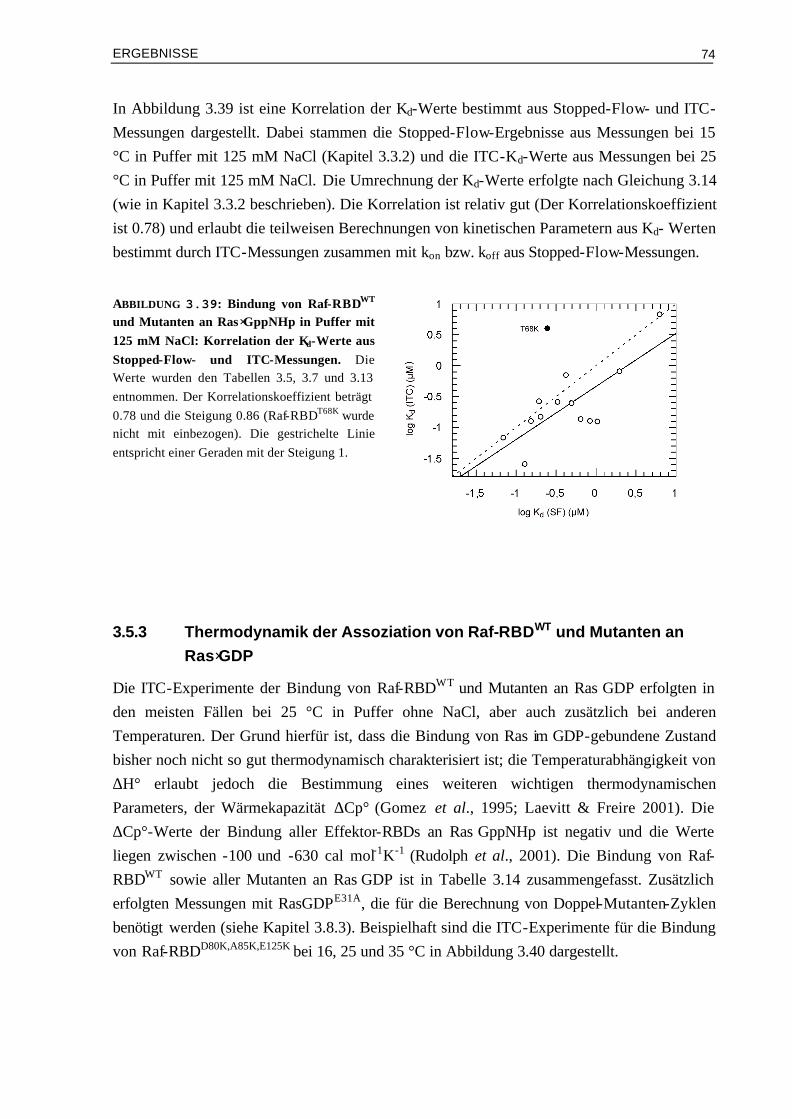
ERGEBNISSE
74
In Abbildung 3.39 ist eine Korrelation der Kd-Werte bestimmt aus Stopped-Flow- und ITC-Messungen dargestellt. Dabei stammen die Stopped-Flow-Ergebnisse aus Messungen bei 15 °C in Puffer mit 125 mM NaCl (Kapitel 3.3.2) und die ITC-Kd-Werte aus Messungen bei 25 °C in Puffer mit 125 mM NaCl. Die Umrechnung der Kd-Werte erfolgte nach Gleichung 3.14 (wie in Kapitel 3.3.2 beschrieben). Die Korrelation ist relativ gut (Der Korrelationskoeffizient ist 0.78) und erlaubt die teilweisen Berechnungen von kinetischen Parametern aus Kd- Werten bestimmt durch ITC-Messungen zusammen mit kon bzw. koff aus Stopped-Flow-Messungen.
ABBILDUNG 3.39: Bindung von Raf-RBDWT und Mutanten an Ras ⋅GppNHp in Puffer mit 125 mM NaCl: Korrelation der Kd-Werte aus Stopped-Flow- und ITC-Messungen. Die Werte wurden den Tabellen 3.5, 3.7 und 3.13 entnommen. Der Korrelationskoeffizient beträgt 0.78 und die Steigung 0.86 (Raf-RBDT68K wurde nicht mit einbezogen). Die gestrichelte Linie entspricht einer Geraden mit der Steigung 1.
3.5.3 Thermodynamik der Assoziation von Raf-RBDWT und Mutanten an
Ras⋅GDP
Die ITC-Experimente der Bindung von Raf-RBDWT und Mutanten an Ras⋅GDP erfolgten in den meisten Fällen bei 25 °C in Puffer ohne NaCl, aber auch zusätzlich bei anderen Temperaturen. Der Grund hierfür ist, dass die Bindung von Ras im GDP-gebundene Zustand bisher noch nicht so gut thermodynamisch charakterisiert ist; die Temperaturabhängigkeit von ∆H° erlaubt jedoch die Bestimmung eines weiteren wichtigen thermodynamischen Parameters, der Wärmekapazität ∆Cp° (Gomez et al., 1995; Laevitt & Freire 2001). Die ∆Cp°-Werte der Bindung aller Effektor-RBDs an Ras⋅GppNHp ist negativ und die Werte liegen zwischen -100 und -630 cal mol-1K-1 (Rudolph et al., 2001). Die Bindung von Raf-RBDWT sowie aller Mutanten an Ras⋅GDP ist in Tabelle 3.14 zusammengefasst. Zusätzlich erfolgten Messungen mit RasGDPE31A, die für die Berechnung von Doppel-Mutanten-Zyklen benötigt werden (siehe Kapitel 3.8.3). Beispielhaft sind die ITC-Experimente für die Bindung von Raf-RBDD80K,A85K,E125K bei 16, 25 und 35 °C in Abbildung 3.40 dargestellt.
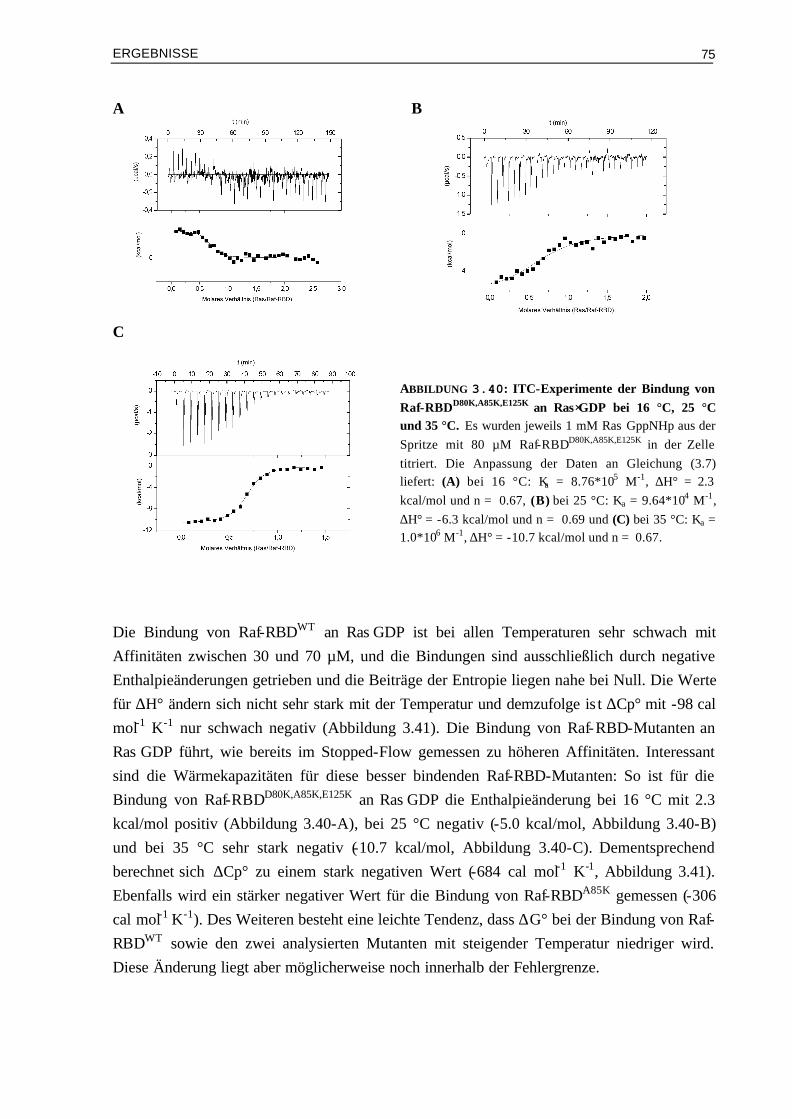
ERGEBNISSE
75
A B
C ABBILDUNG 3.40: ITC-Experimente der Bindung von Raf-RBDD80K,A85K,E125K an Ras⋅GDP bei 16 °C, 25 °C und 35 °C. Es wurden jeweils 1 mM Ras ⋅GppNHp aus der Spritze mit 80 µM Raf-RBDD80K,A85K,E125K in der Zelle titriert. Die Anpassung der Daten an Gleichung (3.7) liefert: (A) bei 16 °C: Ka = 8.76*105 M-1, ∆H° = 2.3 kcal/mol und n = 0.67, (B) bei 25 °C: Ka = 9.64*104 M-1, ∆H° = -6.3 kcal/mol und n = 0.69 und (C) bei 35 °C: Ka = 1.0*106 M-1, ∆H° = -10.7 kcal/mol und n = 0.67.
Die Bindung von Raf-RBDWT an Ras⋅GDP ist bei allen Temperaturen sehr schwach mit Affinitäten zwischen 30 und 70 µM, und die Bindungen sind ausschließlich durch negative Enthalpieänderungen getrieben und die Beiträge der Entropie liegen nahe bei Null. Die Werte für ∆H° ändern sich nicht sehr stark mit der Temperatur und demzufolge is t ∆Cp° mit -98 cal mol-1 K-1 nur schwach negativ (Abbildung 3.41). Die Bindung von Raf-RBD-Mutanten an Ras⋅GDP führt, wie bereits im Stopped-Flow gemessen zu höheren Affinitäten. Interessant sind die Wärmekapazitäten für diese besser bindenden Raf-RBD-Mutanten: So ist für die Bindung von Raf-RBDD80K,A85K,E125K an Ras⋅GDP die Enthalpieänderung bei 16 °C mit 2.3 kcal/mol positiv (Abbildung 3.40-A), bei 25 °C negativ (-5.0 kcal/mol, Abbildung 3.40-B) und bei 35 °C sehr stark negativ (-10.7 kcal/mol, Abbildung 3.40-C). Dementsprechend berechnet sich ∆Cp° zu einem stark negativen Wert (-684 cal mol-1 K-1, Abbildung 3.41). Ebenfalls wird ein stärker negativer Wert für die Bindung von Raf-RBDA85K gemessen (-306 cal mol-1 K-1). Des Weiteren besteht eine leichte Tendenz, dass ∆G° bei der Bindung von Raf-RBDWT sowie den zwei analysierten Mutanten mit steigender Temperatur niedriger wird. Diese Änderung liegt aber möglicherweise noch innerhalb der Fehlergrenze.
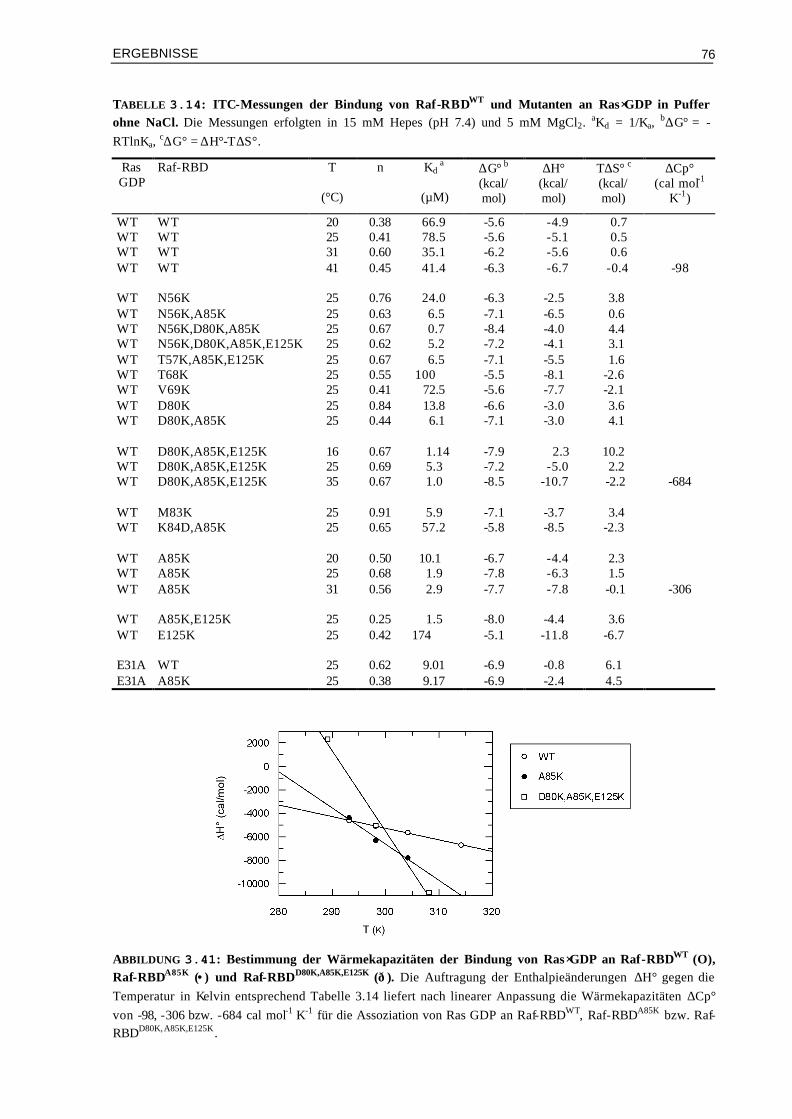
ERGEBNISSE
76
TABELLE 3.14: ITC-Messungen der Bindung von Raf-RBDWT und Mutanten an Ras⋅GDP in Puffer ohne NaCl. Die Messungen erfolgten in 15 mM Hepes (pH 7.4) und 5 mM MgCl2. aKd = 1/Ka,
b∆G° = -RTlnKa,
c∆G° = ∆H°-T∆S°.
ABBILDUNG 3.41: Bestimmung der Wärmekapazitäten der Bindung von Ras ⋅GDP an Raf-RBDWT (O), Raf-RBDA85K (•) und Raf-RBDD80K,A85K,E125K (ð). Die Auftragung der Enthalpieänderungen ∆H° gegen die Temperatur in Kelvin entsprechend Tabelle 3.14 liefert nach linearer Anpassung die Wärmekapazitäten ∆Cp° von -98, -306 bzw. -684 cal mol-1 K-1 für die Assoziation von Ras ⋅GDP an Raf-RBDWT, Raf-RBDA85K bzw. Raf-RBDD80K, A85K,E125K .
Ras GDP
Raf-RBD T
(°C)
n Kd a
(µM)
∆G° b (kcal/ mol)
∆H° (kcal/ mol)
T∆S° c (kcal/ mol)
∆Cp° (cal mol-1
K-1)
WT WT 20 0.38 66.9 -5.6 -4.9 0.7 WT WT 25 0.41 78.5 -5.6 -5.1 0.5 WT WT 31 0.60 35.1 -6.2 -5.6 0.6 WT WT 41 0.45 41.4 -6.3 -6.7 -0.4 -98 WT N56K 25 0.76 24.0 -6.3 -2.5 3.8 WT N56K,A85K 25 0.63 6.5 -7.1 -6.5 0.6 WT N56K,D80K,A85K 25 0.67 0.7 -8.4 -4.0 4.4 WT N56K,D80K,A85K,E125K 25 0.62 5.2 -7.2 -4.1 3.1 WT T57K,A85K,E125K 25 0.67 6.5 -7.1 -5.5 1.6 WT T68K 25 0.55 100 -5.5 -8.1 -2.6 WT V69K 25 0.41 72.5 -5.6 -7.7 -2.1 WT D80K 25 0.84 13.8 -6.6 -3.0 3.6 WT D80K,A85K 25 0.44 6.1 -7.1 -3.0 4.1 WT D80K,A85K,E125K 16 0.67 1.14 -7.9 2.3 10.2 WT D80K,A85K,E125K 25 0.69 5.3 -7.2 -5.0 2.2 WT D80K,A85K,E125K 35 0.67 1.0 -8.5 -10.7 -2.2 -684 WT M83K 25 0.91 5.9 -7.1 -3.7 3.4 WT K84D,A85K 25 0.65 57.2 -5.8 -8.5 -2.3 WT A85K 20 0.50 10.1 -6.7 -4.4 2.3 WT A85K 25 0.68 1.9 -7.8 -6.3 1.5 WT A85K 31 0.56 2.9 -7.7 -7.8 -0.1 -306 WT A85K,E125K 25 0.25 1.5 -8.0 -4.4 3.6 WT E125K 25 0.42 174 -5.1 -11.8 -6.7 E31A WT 25 0.62 9.01 -6.9 -0.8 6.1 E31A A85K 25 0.38 9.17 -6.9 -2.4 4.5
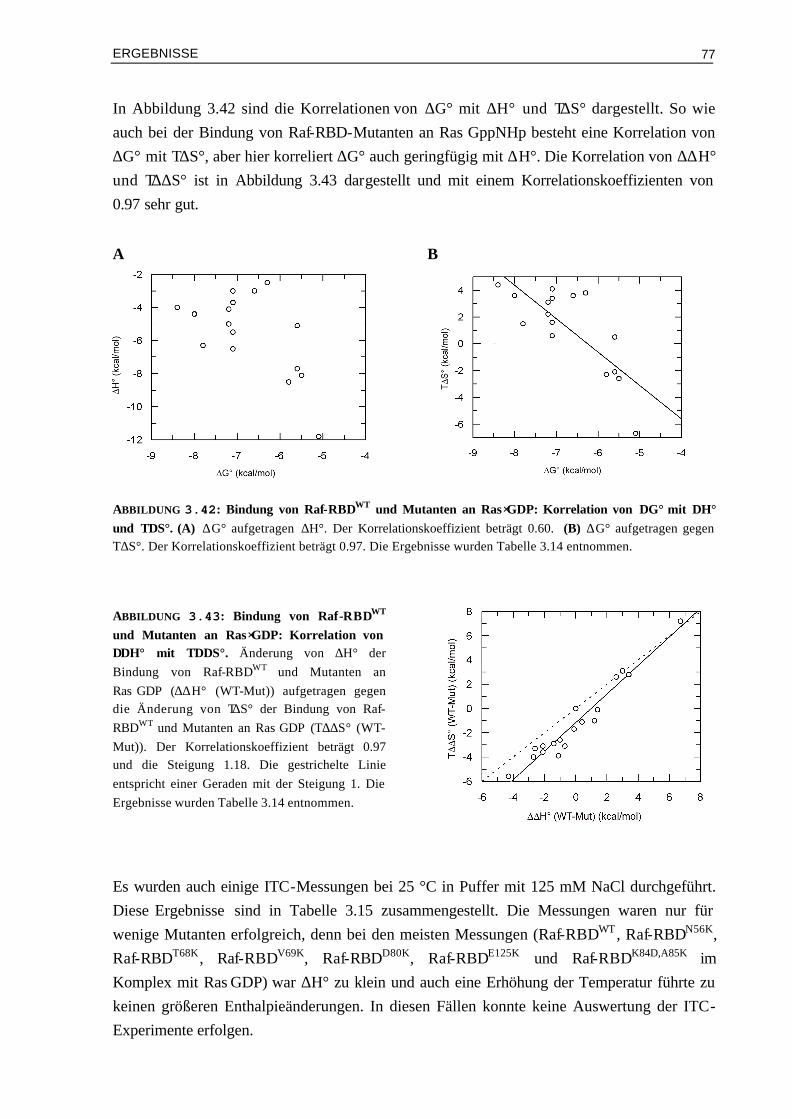
ERGEBNISSE
77
In Abbildung 3.42 sind die Korrelationen von ∆G° mit ∆H° und T∆S° dargestellt. So wie auch bei der Bindung von Raf-RBD-Mutanten an Ras⋅GppNHp besteht eine Korrelation von ∆G° mit T∆S°, aber hier korreliert ∆G° auch geringfügig mit ∆H°. Die Korrelation von ∆∆H° und T∆∆S° ist in Abbildung 3.43 dargestellt und mit einem Korrelationskoeffizienten von 0.97 sehr gut.
A B
ABBILDUNG 3.42: Bindung von Raf-RBDWT und Mutanten an Ras⋅GDP: Korrelation von ∆G° mit ∆H° und T∆S°. (A) ∆G° aufgetragen ∆H°. Der Korrelationskoeffizient beträgt 0.60. (B) ∆G° aufgetragen gegen T∆S°. Der Korrelationskoeffizient beträgt 0.97. Die Ergebnisse wurden Tabelle 3.14 entnommen.
ABBILDUNG 3.43: Bindung von Raf-RBDWT und Mutanten an Ras⋅GDP: Korrelation von ∆∆H° mit T∆∆S°. Änderung von ∆H° der Bindung von Raf-RBDWT und Mutanten an Ras⋅GDP (∆∆H° (WT-Mut)) aufgetragen gegen die Änderung von T∆S° der Bindung von Raf-RBDWT und Mutanten an Ras ⋅GDP (T∆∆S° (WT-Mut)). Der Korrelationskoeffizient beträgt 0.97 und die Steigung 1.18. Die gestrichelte Linie entspricht einer Geraden mit der Steigung 1. Die Ergebnisse wurden Tabelle 3.14 entnommen. Es wurden auch einige ITC-Messungen bei 25 °C in Puffer mit 125 mM NaCl durchgeführt. Diese Ergebnisse sind in Tabelle 3.15 zusammengestellt. Die Messungen waren nur für wenige Mutanten erfolgreich, denn bei den meisten Messungen (Raf-RBDWT , Raf-RBDN56K, Raf-RBDT68K, Raf-RBDV69K, Raf-RBDD80K, Raf-RBDE125K und Raf-RBDK84D,A85K im Komplex mit Ras⋅GDP) war ∆H° zu klein und auch eine Erhöhung der Temperatur führte zu keinen größeren Enthalpieänderungen. In diesen Fällen konnte keine Auswertung der ITC-Experimente erfolgen.

ERGEBNISSE
78
TkhkRTlnG
b
on#
××=∆
TABELLE 3.15: ITC-Messungen der Bindung von Raf-RBDWT und Mutanten an Ras⋅GDP. Die Messungen erfolgten bei 25 °C in 15 mM Hepes (pH 7.4), 5 mM MgCl2 und 125 mM NaCl. aKd = 1/Ka,
b∆G° = -RTlnKa , c∆G° = ∆H°-T∆S°.
3.6 Aktivierungsenergien der Assoziation von Mutanten-Komplexen mit veränderter Elektrostatik
Die Assoziationsratenkonstante kon und die Differenz der freien Enthalpie zwischen den ungebundenen Proteinen (A + B) und dem Übergangszustand ([A:B]#) (siehe Kapitel 1.6) stehen nach der Theorie des Übergangszustandes in folgender Beziehung (Frisch et al, 2001): (3.14) Dabei ist ∆G# die Aktivierungsenergie, h die Planck-Konstante, kb die Boltzmann-Konstante und kon die Assoziationsratenkonstante. Diese Gleichung gilt streng genommen nur für mikroskopische Ratenkonstanten für einzelne Schritte (Glasstone et al., 1941) und nicht für zusammengesetzte Ratenkonstanten wie kon. Daher ist die Betrachtung von absoluten Werten problematisch, aber Änderungen von ∆G# können interpretiert werden, da diese mit einem quasi- thermodynamischen Gleichgewicht zwischen dem Grundzustand und dem Übergangszustand der Assoziationsreaktion verbunden sind (Oliveberg et al., 1995; Frisch et al, 2001). Im Folgenden soll analysiert werden, in welcher Beziehung die Änderungen der Aktivierungsenergien (∆G#) und die Änderungen der freien Enthalpien (∆G°) bei der Komplexbildung zueinander stehen.
3.6.1 Bindung von RalGDS-RBDWT und Mutanten an Ras⋅mGppNHp
Die Ergebnisse für die Aktivierungsenergien sowie die Werte für ∆G° aus Stopped-Flow- und ITC-Messungen sind in Tabelle 3.16 zusammengefasst. Es ist zu erkennen, dass sowohl für die Bindung von RalGDS-RBDWT als auch für alle Mutanten die Aktivierungsenergien mit Werten zwischen 6 und 9 kcal/mol relativ groß sind. Es besteht somit eine hohe
Raf-RBD n Kd a
(µM)
∆G° b (kcal/ mol)
∆H° (kcal/ mol)
T∆S° c (kcal/ mol)
N56K,D80K,A85K 0.65 14.6 -6.6 -3.4 3.2 T57K,A85K,E125K 0.34 15.3 -6.6 -2.9 3.6 D80K,A85K 0.22 89.2 -5.5 -8.4 -2.9 D80K,A85K,E125K 0.52 26.2 -6.3 -4.3 2.0 M83K 0.84 41.2 -6.0 -2.0 4.0 A85K 0.76 12.3 -6.7 -4.5 2.2 A85K,E125K 0.71 7.3 -7.0 -2.2 4.8
ERGEBNISSE
79
Energiebarriere zwischen den Ausgangsproteinen und den Endkomplexen. Als Grund für diese hohen Energiebarrieren wird neben strukturellen Umlagerungen vor allem der Energieverlust durch Desolvatation, insbesondere von positiv geladenen Resten, was thermodynamisch sehr ungünstig ist, diskutiert (Schreiber, 2002).
Der Vergleich der Bindung von RalGDS-RBDWT mit einer schneller assoziierenden Mutante (RalGDS-RBDM26K,D47K,E54K,D90K) zeigt, dass die höhere Affinität mit ∆∆G° (WT-Mut) = 1.7 kcal/mol hauptsächlich die Folge einer um 1.5 kcal/mol niedrigeren Aktivierungsenergie ist. Um zu analysieren, ob eine Beziehung zwischen den Änderungen der Aktivierungsenergien zwischen Wildtyp und Mutante (∆∆G# (WT-Mut)) und den Änderungen der freien Enthalpie ∆∆G° (WT-Mut) besteht, wurden die ent sprechenden Änderungen in einem Diagramm gegeneinander aufgetragen (Abbildung 3.44). TABELLE 3.16: Aktivierungsenergien und Änderungen der freien Enthalpie der Bindung von RalGDS-RBDWT und Mutanten an Ras⋅(m)GppNHp. Die Werte für ∆G# wurden aus den kon-Werten aus den Tabellen 3.4 und 3.6 entsprechend Gleichung 3.14 berechnet. Die ∆G°-Werte aus ITC- und Stopped-Flow- (SF-) Messungen entstammen den Tabellen 3.4, 3.6. und 3.11.
RalGDS-RBD kon
(µM-1
s -1)
∆G#
(kcal/ mol)
∆∆G#
(kcal/ mol)
∆G° SF
(kcal/ mol)
∆∆G° SF
(kcal/ mol)
∆G° ITC
(kcal/ mol)
∆∆G° ITC
(kcal/ mol)
WT 7.7 8.06 -7.8 -8.1 R16A 2.5 8.72 -0.67 -6.5 -1.1 -6.5 -1.6 S18K 16.0 7.62 0.44 -7.9 0.1 -7.2 -0.9 S18K,Y89K 3.6 8.51 -0.45 -7.0 -0.8 -7.3 -0.8 L19K 8.9 7.97 0.09 -7.8 0.0 -8.6 0.5 N23K 11.7 7.81 0.25 -7.4 -0.4 -7.7 -0.4 N23K,N88K 5.1 8.30 -0.24 -7.3 -0.5 -8.1 0.0 M26K 16.2 7.62 0.44 -8.7 0.9 -8.5 0.4 M26K,E54K 17.3 7.58 0.48 -8.8 1.0 -8.3 0.2 M26K,D90K 47.5 6.98 1.08 -8.7 0.9 -8.5 0.4 M26K,D47K,E54K 110.0 6.48 1.56 -9.8 2.0 -9.9 1.8 M26K,D47K,E54K,D90K 96.3 6.56 1.50 -9.5 1.7 -9.0 0.9 K28A 2.2 8.81 -0.75 -6.6 -1.3 -6.6 -1.5 K44A 5.6 8.25 -0.19 -7.6 -0.2 -7.8 -0.3 D47A 17.5 7.57 0.49 -8.4 0.6 -8.7 0.6 D47K 30.3 7.25 0.81 -8.9 1.1 -9.0 0.9 D47K,E52K,E53K 43.0 7.04 1.02 -9.4 1.6 -9.3 1.2 K48A 1.2 9.15 -1.09 -6.3 -1.2 -6.3 -1.8 N50K 15.8 7.63 0.43 -8.7 0.9 -8.2 0.1 L51K 15.1 7.66 0.40 -8.4 0.6 -8.1 0.0 E52A 18.5 7.54 0.52 -8.1 0.3 -8.5 0.4 E53A 16.3 7.61 0.44 -8.0 0.2 -8.5 0.4 E53K 19.1 7.52 0.54 -8.1 0.3 -8.2 0.1 E54K 15.2 7.65 0.40 -8.7 0.9 -8.6 0.5 E54K,D90K 28.4 7.28 0.77 -8.4 0.6 -8.8 0.7 N88K 21.4 7.45 0.61 -8.6 0.8 -8.5 0.4 Y89K 21.7 7.44 0.61 -7.7 -0.1 -7.9 -0.2 D90K 27.9 7.30 0.76 -8.9 1.1 -9.0 0.9

ERGEBNISSE
80
Es besteht eine sehr gute Korrelation zwischen der Änderung der Aktivierungsenergie und der freien Enthalpie für die Bindung an Ras⋅(m)GppNHp von RalGDS-RBDWT und Mutanten. Der Korrelationskoeffizient für die Auftragung von ∆∆G# (WT-Mut) gegen ∆∆G° (WT-Mut) aus Stopped-Flow-Messungen ist dabei mit 0.92 besser (Abbildung 3.44-A) als bei der Auftragung gegen ∆∆G° (WT-Mut) aus ITC-Messungen (Korrelationskoeffizient 0.88). Dieses entspricht den Erwartungen, da ∆G# aus den kon-Werten aus Stopped-Flow-Messungen ermittelt wird. Die Affinitäten der analysierten Mutanten korrelieren somit mit den Aktivierungsenergien. A B
ABBILDUNG 3.44: Korrelation von ∆∆G# mit ∆∆G° der Bindung von RalGDS-RBDWT und Mutanten an Ras⋅mGppNHp aus Stopped-Flow- und ITC-Experimenten. Aufgetragen ist die Änderung der Aktivierungsenergie (∆∆G# (WT-Mut)) gegen die Änderung der freien Enthalpie (∆∆G° (WT-Mut)). (A) Stopped-Flow-Messungen: Der Korrelationskoeffizient ist 0.92 und die Steigung 1.28. (B) ITC-Messungen: Der Korrelationskoeffizient ist 0.88 und die Steigung 1.19. Die gestrichelte Linie entspricht einer Geraden mit der Steigung 1. Die Werte wurden der Tabelle 3.16 entnommen.
3.6.2 Bindung von Raf-RBDWT und Mutanten an Ras⋅mGppNHp
Die Aktivierungsenergien für die Bindung von Raf-RBDWT und Mutanten an Ras⋅mGppNHp sind in Tabelle 3.17 zusammengefasst. Die Werte für ∆G# liegen wie bei der Bindung von RalGDS-RBD an Ras⋅mGppNHp im Bereich von 6 bis 8 kcal/mol. Die Korrelation von ∆∆G# (WT-Mut) mit ∆∆G° (WT-Mut) ist in Abbildung 3.45 dargestellt. Es besteht eine gute Korrelation, aber die Steigung ist 1.72. Das bedeutet, dass neben der Änderung der Aktivierungsenergie auch der zweite Schritt (k2) beeinflusst wird; dieses passt gut, da bei der Bindung von Raf-RBD-Mutanten an Ras⋅mGppNHp die Dissoziation stärker beeinflusst wird.
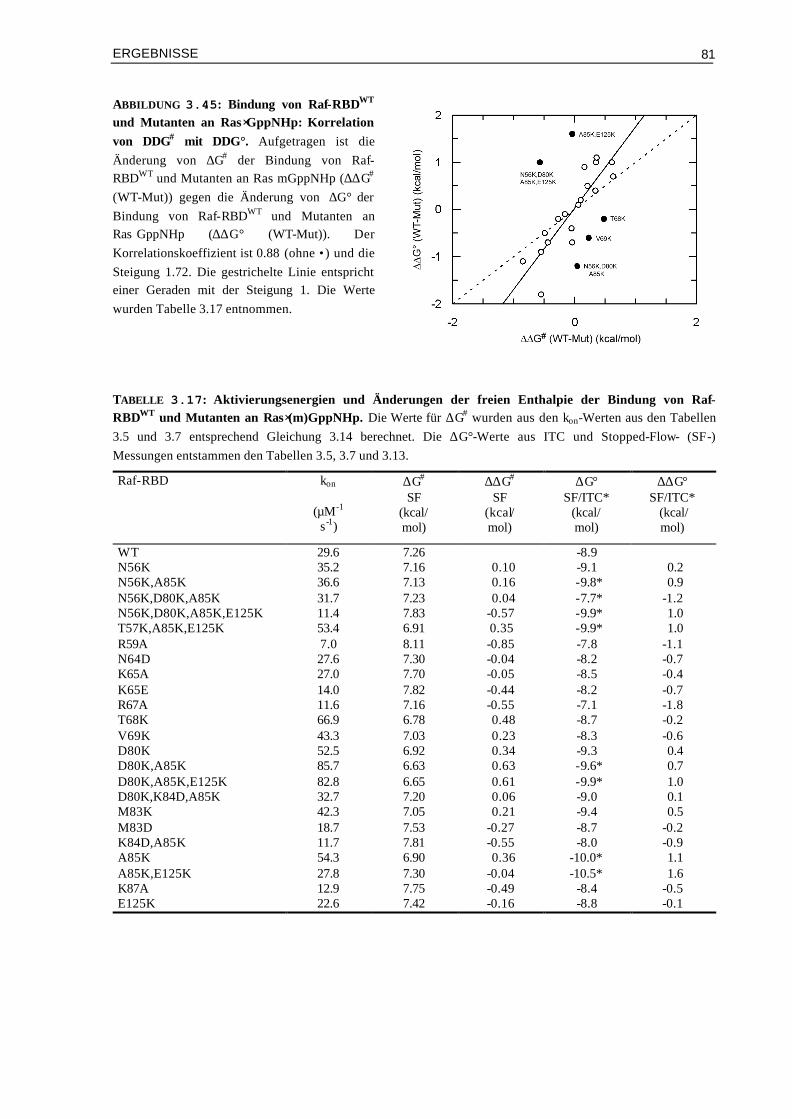
ERGEBNISSE
81
ABBILDUNG 3.45: Bindung von Raf-RBDWT und Mutanten an Ras ⋅GppNHp: Korrelation von ∆∆G# mit ∆∆G°. Aufgetragen ist die Änderung von ∆G# der Bindung von Raf-RBDWT und Mutanten an Ras ⋅mGppNHp (∆∆G# (WT-Mut)) gegen die Änderung von ∆G° der Bindung von Raf-RBDWT und Mutanten an Ras⋅GppNHp (∆∆G° (WT-Mut)). Der Korrelationskoeffizient ist 0.88 (ohne •) und die Steigung 1.72. Die gestrichelte Linie entspricht einer Geraden mit der Steigung 1. Die Werte wurden Tabelle 3.17 entnommen.
TABELLE 3.17: Aktivierungsenergien und Änderungen der freien Enthalpie der Bindung von Raf-RBDWT und Mutanten an Ras⋅(m)GppNHp. Die Werte für ∆G# wurden aus den kon-Werten aus den Tabellen 3.5 und 3.7 entsprechend Gleichung 3.14 berechnet. Die ∆G°-Werte aus ITC und Stopped-Flow- (SF-) Messungen entstammen den Tabellen 3.5, 3.7 und 3.13.
Raf-RBD kon
(µM-1
s -1)
∆G# SF
(kcal/ mol)
∆∆G# SF
(kcal/ mol)
∆G° SF/ITC*
(kcal/ mol)
∆∆G° SF/ITC*
(kcal/ mol)
WT 29.6 7.26 -8.9 N56K 35.2 7.16 0.10 -9.1 0.2 N56K,A85K 36.6 7.13 0.16 -9.8* 0.9 N56K,D80K,A85K 31.7 7.23 0.04 -7.7* -1.2 N56K,D80K,A85K,E125K 11.4 7.83 -0.57 -9.9* 1.0 T57K,A85K,E125K 53.4 6.91 0.35 -9.9* 1.0 R59A 7.0 8.11 -0.85 -7.8 -1.1 N64D 27.6 7.30 -0.04 -8.2 -0.7 K65A 27.0 7.70 -0.05 -8.5 -0.4 K65E 14.0 7.82 -0.44 -8.2 -0.7 R67A 11.6 7.16 -0.55 -7.1 -1.8 T68K 66.9 6.78 0.48 -8.7 -0.2 V69K 43.3 7.03 0.23 -8.3 -0.6 D80K 52.5 6.92 0.34 -9.3 0.4 D80K,A85K 85.7 6.63 0.63 -9.6* 0.7 D80K,A85K,E125K 82.8 6.65 0.61 -9.9* 1.0 D80K,K84D,A85K 32.7 7.20 0.06 -9.0 0.1 M83K 42.3 7.05 0.21 -9.4 0.5 M83D 18.7 7.53 -0.27 -8.7 -0.2 K84D,A85K 11.7 7.81 -0.55 -8.0 -0.9 A85K 54.3 6.90 0.36 -10.0* 1.1 A85K,E125K 27.8 7.30 -0.04 -10.5* 1.6 K87A 12.9 7.75 -0.49 -8.4 -0.5 E125K 22.6 7.42 -0.16 -8.8 -0.1
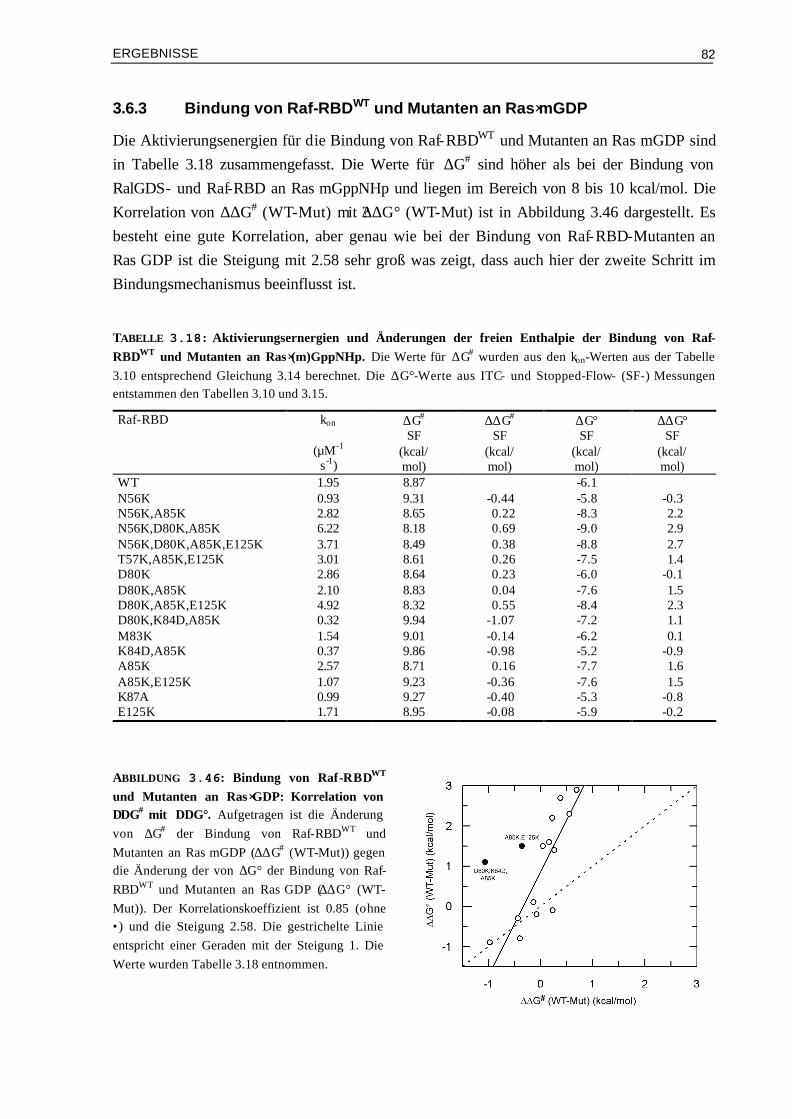
ERGEBNISSE
82
3.6.3 Bindung von Raf-RBDWT und Mutanten an Ras⋅mGDP
Die Aktivierungsenergien für die Bindung von Raf-RBDWT und Mutanten an Ras⋅mGDP sind in Tabelle 3.18 zusammengefasst. Die Werte für ∆G# sind höher als bei der Bindung von RalGDS- und Raf-RBD an Ras⋅mGppNHp und liegen im Bereich von 8 bis 10 kcal/mol. Die Korrelation von ∆∆G# (WT-Mut) mit ?∆∆G° (WT-Mut) ist in Abbildung 3.46 dargestellt. Es besteht eine gute Korrelation, aber genau wie bei der Bindung von Raf-RBD-Mutanten an Ras⋅GDP ist die Steigung mit 2.58 sehr groß was zeigt, dass auch hier der zweite Schritt im Bindungsmechanismus beeinflusst ist.
TABELLE 3.18: Aktivierungsernergien und Änderungen der freien Enthalpie der Bindung von Raf-RBDWT und Mutanten an Ras⋅(m)GppNHp. Die Werte für ∆G# wurden aus den kon-Werten aus der Tabelle 3.10 entsprechend Gleichung 3.14 berechnet. Die ∆G°-Werte aus ITC- und Stopped-Flow- (SF-) Messungen entstammen den Tabellen 3.10 und 3.15.
ABBILDUNG 3.46: Bindung von Raf-RBDWT und Mutanten an Ras ⋅GDP: Korrelation von ∆∆G# mit ∆∆G°. Aufgetragen ist die Änderung von ∆G# der Bindung von Raf-RBDWT und Mutanten an Ras ⋅mGDP (∆∆G# (WT-Mut)) gegen die Änderung der von ∆G° der Bindung von Raf-RBDWT und Mutanten an Ras ⋅GDP (∆∆G° (WT-Mut)). Der Korrelationskoeffizient ist 0.85 (ohne •) und die Steigung 2.58. Die gestrichelte Linie entspricht einer Geraden mit der Steigung 1. Die Werte wurden Tabelle 3.18 entnommen.
Raf-RBD kon
(µM-1 s -1)
∆G# SF
(kcal/ mol)
∆∆G# SF
(kcal/ mol)
∆G° SF
(kcal/ mol)
∆∆G° SF
(kcal/ mol)
WT 1.95 8.87 -6.1 N56K 0.93 9.31 -0.44 -5.8 -0.3 N56K,A85K 2.82 8.65 0.22 -8.3 2.2 N56K,D80K,A85K 6.22 8.18 0.69 -9.0 2.9 N56K,D80K,A85K,E125K 3.71 8.49 0.38 -8.8 2.7 T57K,A85K,E125K 3.01 8.61 0.26 -7.5 1.4 D80K 2.86 8.64 0.23 -6.0 -0.1 D80K,A85K 2.10 8.83 0.04 -7.6 1.5 D80K,A85K,E125K 4.92 8.32 0.55 -8.4 2.3 D80K,K84D,A85K 0.32 9.94 -1.07 -7.2 1.1 M83K 1.54 9.01 -0.14 -6.2 0.1 K84D,A85K 0.37 9.86 -0.98 -5.2 -0.9 A85K 2.57 8.71 0.16 -7.7 1.6 A85K,E125K 1.07 9.23 -0.36 -7.6 1.5 K87A 0.99 9.27 -0.40 -5.3 -0.8 E125K 1.71 8.95 -0.08 -5.9 -0.2

ERGEBNISSE
83
3.7 Bindung von Raf-RBDWT und Mutanten an Ras⋅(m)GppNHpT35A und Ras⋅mGppNHpT35S
Die Bindung von Raf-RBD an Ras⋅GppNHpT35A weist möglicherweise Parallelen zu der Bindung an Ras⋅GDP auf. So ist die Amplitude der Fluoreszenzänderung von Raf-RBD an Ras⋅mGppNHpT35A genauso wie bei der Bindung an Ras⋅mGDP: Bei der Assoziation kommt es zu einer Zunahme der Fluoreszenz und bei der Dissoziation zu einer Abnahme, also genau entgegengesetzt wie bei der Bindung aller Effektor-RBDs an Ras⋅mGppNHp. Es sollte analysiert werden, ob die Raf-RBD-Mutanten mit veränderter Elektrostatik auch besser an Ras⋅(m)GppNHpT35A binden können und durch welche kinetischen und thermodynamischen Parameter die Bindung charakterisiert ist. Zusätzlich sollte die Bindung der Raf-RBD-Mutanten im Komplex mit Ras⋅mGppNHpT35S untersucht werden.
3.7.1 Kinetik der Bindung von Raf-RBDWT und Mutanten an
Ras⋅mGppNHpT35A und Ras⋅mGppNHpT35S
Die Ergebnisse der Stopped-Flow-Messungen der Bindungen von Raf-RBDWT und Mutanten an Ras⋅mGppNHpT35A und Ras⋅mGppNHpT35S sind in Tabelle 3.19 zusammengefasst. Beispielhaft ist die Assoziationskinetik der Bindung von Raf-RBDWT und Raf-RBDA85K an Ras⋅mGppNHpT35A gemessen bei 15 °C in Puffer mit 125 mM NaCl in Abbildung 3.47 dargestellt.
TABELLE 3.19: Stopped-Flow-Messungen der Bindung von Raf-RBDWT und Mutanten an Ras⋅mGppNHpT35A und Ras⋅mGppNHpT35S. Die Messungen erfolgten in 15 mM Hepes (pH 7.4), 5 mM MgCl2 und teilweise mit zusätzlich 125 mM NaCl. aKd = koff/kon, Ka = 1/Kd ,b∆G° = -RTlnKa, n.d.: nicht auswertbar, da stark negativer Achsenabschnitt.
[NaCl] (mM)
Ras⋅ m⋅GppNHp
Raf-RBD T (°C)
kon
(µM-1s -1) koff (s-1)
Kd a
(µM) ∆G° b (kcal/ mol)
125 WT WT 15 29.6 5.7 0.193 -8.9
125 T35S WT 15 6.0 9.0 1.51 -7.7 125 T35S A85K 15 13.5 0.59 0.044 -9.7 125 T35S N56K,A85K 15 19.4 0.72 0.037 -9.8
125 T35A WT 15 2.29 8.9 3.90 -7.1 125 T35A A85K 15 14.7 16.4 1.12 -7.8
125 T35A WT 25 3.3 26.3 7.97 -6.7 125 T35A D80K,A85K 25 14.0 17.4 1.24 -8.1 125 T35A A85K 25 21.3 35.8 1.68 -7.9
0 T35A WT 25 n.d. 15.4 0 T35A N56K,A85K 25 n.d. 12.3 0 T35A N56K,D80K,A85K 25 n.d. 10.6 0 T35A A85K 25 n.d. 16.8
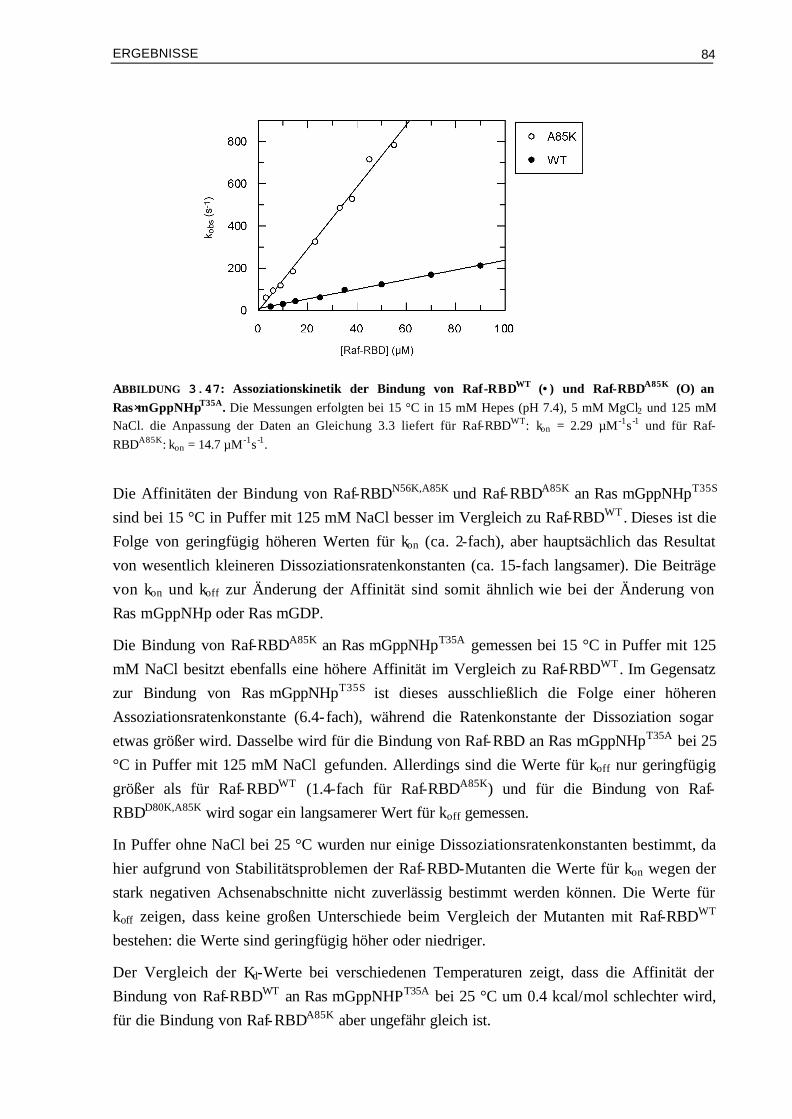
ERGEBNISSE
84
ABBILDUNG 3.47: Assoziationskinetik der Bindung von Raf-RBDWT (•) und Raf-RBDA85K (Ο) an Ras⋅mGppNHpT35A. Die Messungen erfolgten bei 15 °C in 15 mM Hepes (pH 7.4), 5 mM MgCl2 und 125 mM NaCl. die Anpassung der Daten an Gleichung 3.3 liefert für Raf-RBDWT: kon = 2.29 µM-1s -1 und für Raf-RBDA85K: kon = 14.7 µM-1s -1.
Die Affinitäten der Bindung von Raf-RBDN56K,A85K und Raf-RBDA85K an Ras⋅mGppNHpT35S sind bei 15 °C in Puffer mit 125 mM NaCl besser im Vergleich zu Raf-RBDWT . Dieses ist die Folge von geringfügig höheren Werten für kon (ca. 2-fach), aber hauptsächlich das Resultat von wesentlich kleineren Dissoziationsratenkonstanten (ca. 15-fach langsamer). Die Beiträge von kon und koff zur Änderung der Affinität sind somit ähnlich wie bei der Änderung von Ras⋅mGppNHp oder Ras⋅mGDP.
Die Bindung von Raf-RBDA85K an Ras⋅mGppNHpT35A gemessen bei 15 °C in Puffer mit 125 mM NaCl besitzt ebenfalls eine höhere Affinität im Vergleich zu Raf-RBDWT . Im Gegensatz zur Bindung von Ras⋅mGppNHpT35S ist dieses ausschließlich die Folge einer höheren Assoziationsratenkonstante (6.4-fach), während die Ratenkonstante der Dissoziation sogar etwas größer wird. Dasselbe wird für die Bindung von Raf-RBD an Ras⋅mGppNHpT35A bei 25 °C in Puffer mit 125 mM NaCl gefunden. Allerdings sind die Werte für koff nur geringfügig größer als für Raf-RBDWT (1.4-fach für Raf-RBDA85K) und für die Bindung von Raf-RBDD80K,A85K wird sogar ein langsamerer Wert für koff gemessen.
In Puffer ohne NaCl bei 25 °C wurden nur einige Dissoziationsratenkonstanten bestimmt, da hier aufgrund von Stabilitätsproblemen der Raf-RBD-Mutanten die Werte für kon wegen der stark negativen Achsenabschnitte nicht zuverlässig bestimmt werden können. Die Werte für koff zeigen, dass keine großen Unterschiede beim Vergleich der Mutanten mit Raf-RBDWT bestehen: die Werte sind geringfügig höher oder niedriger.
Der Vergleich der Kd-Werte bei verschiedenen Temperaturen zeigt, dass die Affinität der Bindung von Raf-RBDWT an Ras⋅mGppNHPT35A bei 25 °C um 0.4 kcal/mol schlechter wird, für die Bindung von Raf-RBDA85K aber ungefähr gleich ist.

ERGEBNISSE
85
3.7.2 Thermodynamik der Bindung von Raf-RBDWT und Mutanten an Ras⋅GppNHpT35A
Die ITC-Ergebnisse der Bindung von Raf-RBDWT und Raf-RBDN56K,A85K an Ras⋅GppNHpT35A sind in Tabelle 3.20 dargestellt. Die Messungen erfolgten in Puffer ohne NaCl, da sonst die Enthalpieänderung zu klein war. Die Affinität der Bindung von Raf-RBDWT an Ras⋅GppNHpT35A wird geringfügig besser mit steigender Temperatur. Ebenso wird die Enthalpieänderung etwas negativer bei Temperaturerhöhung, woraus die Wärmekapazität zu -55 cal mol-1K-1 bestimmt wird. Dieser Wert für ∆Cp° liegt also in derselben Größenordnung wie bei der Bindung von Raf-RBDWT an Ras⋅GDP. Die Messungen für die Raf-RBD-Mutanten an RasGppNHpT35A waren zumeist sehr schlecht auswertbar, da die Enthalpieänderung sehr klein ist. Die höher gemessene Affinität von Raf-RBDN56K,A85K im Vergleich zu Raf-RBDWT (∆∆G° (WT-Mut) ist ungefähr 0.7 kcal/mol) konnte jedoch die Stopped-Flow-Ergebnisse bestätigen.
Die Bindung von Raf-RBDN56K,A85K an Ras⋅GppNHpT35A ist bei 15 °C durch eine positive Enthalpieänderung charakterisiert, allerdings konnte der Ka-Wert nicht zuverlässig ausgewertet werden. Bei 25 °C konnte ebenfalls nur eine sehr gering negative Enthalpieänderung detektiert werden. Die Werte für ∆H° bei 15 °C und bei 30 °C erlauben jedoch eine Abschätzung für die Wärmekapazität ∆Cp° von ca. -250 cal mol-1 K-1. Dieses ist vergleichbar mit den Ergebnissen für ∆Cp° bei der Bindung von Raf-RBD-Mutanten an Ras⋅GDP, die ebenfalls deutlich negativer sind als für die Bindung von Raf-RBDWT . TABELLE 3.20: ITC-Messungen der Bindung von Raf-RBDWT und Mutanten an Ras⋅GppNHpT35A. Die Messungen erfolgten in 15 mM Hepes (pH 7.4) und 5 mM MgCl2. aKd = 1/Ka , b∆G° = -RTlnKa , c∆G° = ∆H°-T∆S°.
Raf-RBD T
(°C)
n Kd a
(µM)
∆G° b (kcal/ mol)
∆H° (kcal/ mol)
T∆S° c (kcal/ mol)
∆Cp° (cal
mol-1k-1) WT 15 0.96 4.52 -7.05 -2.83 4.22 25 0.85 3.46 -7.45 -3.26 4.19 35 0.85 2.14 -7.99 -3.93 4.09 -55 N56K,A85K 15 n.a. 1.88 30 0.60 0.51 -8.73 -1.50 7.23 ca. -250

ERGEBNISSE
86
3.8 Strukturelle Analyse der Bindung von Raf-RBDA85K an Ras⋅GDP und Ras⋅GppNHpT35A
Die Raf-RBD-Mutanten, die zu einer höheren elektrostatischen Komplementarität im Komplex mit Ras führen, binden mit deutlich höheren Affinitäten an Ras⋅GDP und auch an Ras⋅GppNHpT35A als Raf-RBDWT . Es konnte gezeigt werden, dass die bessere Bindung von Raf-RBD-Mutanten an Ras⋅mGDP nur zu einem kleinen Teil die Folge einer schnelleren Assoziation ist, aber zum größten Teil aus einer langsameren Dissoziation resultiert. Für die Bindung von Raf-RBDA85K und Raf-RBDN56K,A85K an Ras⋅mGppNHpT35A dagegen ist die bessere Affinität hauptsächlich eine Folge von höheren Werten für kon, während die Werte für koff etwas größer werden. Es sollte analysiert werden, was die strukturellen Ursachen der veränderten Affinitäten der Bindung von Raf-RBD-Mutanten an Ras⋅GDP und Ras⋅GppNHpT35A sind. Die Veränderungen der Schalter-Regionen von Ras im GTP/GppNHp- und im GDP-gebundenen Zustand wurden für die unterschiedlichen Affinitäten der Bindung von Effektor-RBDs an Ras⋅GppNHp und Ras⋅GDP verantwortlich gemacht. Es sollte daher untersucht werden, wie der Effektor-Bereich und die Schalter-Regionen von Ras⋅GDP im Komplex mit den besser bindenden Raf-RBD-Mutanten aussehen, d.h. ob die Mutanten Ras⋅GDP in eine GTP-ähnliche Konformation drängen können. Als besser bindende Mutante von Raf-RBD wurde für die verschiedenen Experimente jeweils Raf-RBDA85K gewählt, da diese die Einfach-Mutante mit dem größten Effekt auf die Affinität im Komplex mit Ras⋅GDP und Ras⋅GppNHpT35A ist.
3.8.1 31P-NMR-Messungen
Es konnte gezeigt werden, dass Ras gebunden an GppNHp in zwei Zuständen vorliegt, die durch unterschiedliche Konformationen gekennzeichnet sind (Abbildung 3.48). Dies wird durch eine Aufspaltung der Resonanzfrequenzen am ß- und geringfügig auch am α-Phosphat in 31P-NMR-Experimenten sichtbar (Geyer et al., 1996). Die Aufspaltung (bezeichnet als Zustand 1 und Zustand 2) wurde durch die Mobilität der Schalter-Regionen gedeutet, wobei Tyr32 von Ras in die Nähe des γ-Phosphates gelangt und dadurch der π-Elektronen-Ringstrom-Effekt des aromatischen Rings die beobachtete Hochfeldverschiebung der Resonanzlinien (Zustand 2) verursacht. Bei Zugabe eines Effektors wird dieser Zustand (2) stabilisiert (Geyer et al., 1996, Linnemann et al., 1999, Spoerner et al., 2001). Als Gründe hierfür wurden im Fall der Bindung von Raf-RBD an Ras⋅GppNHp Kontakte bei der Bindung von Raf-RBD an die Aminosäuren Glu31 und Asp33 von Ras diskutiert, wodurch Tyr32 von Ras wegklappt (Wittinghofer & Nassar, 1996). Dieses führt zu einer Stabilisierung durch Ausbildung einer Wasserstoffbrücke von Tyr32 zum γ-Phosphat und daraus resultierend die Aufspaltung der Signale am ß-Phosphat. Für die Ras-Mutanten Ras⋅GppNHpT35A und Ras⋅GppNHpT35S konnte keine Aufspaltung der Resonanzlinien am ß-Phosphat gezeigt werden (Geyer et al., 1996, Spoerner et al., 2001) und die chemische Verschiebung entspricht
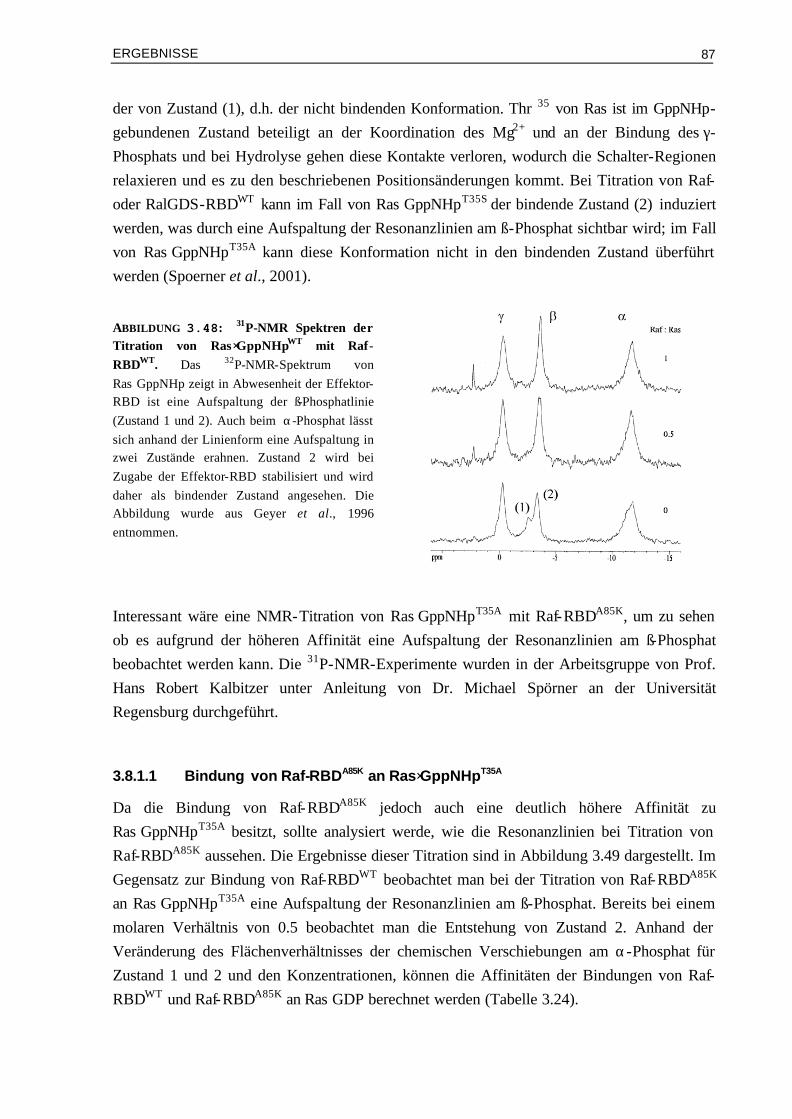
ERGEBNISSE
87
der von Zustand (1), d.h. der nicht bindenden Konformation. Thr 35 von Ras ist im GppNHp-gebundenen Zustand beteiligt an der Koordination des Mg2+ und an der Bindung des γ-Phosphats und bei Hydrolyse gehen diese Kontakte verloren, wodurch die Schalter-Regionen relaxieren und es zu den beschriebenen Positionsänderungen kommt. Bei Titration von Raf- oder RalGDS-RBDWT kann im Fall von Ras⋅GppNHpT35S der bindende Zustand (2) induziert werden, was durch eine Aufspaltung der Resonanzlinien am ß-Phosphat sichtbar wird; im Fall von Ras⋅GppNHpT35A kann diese Konformation nicht in den bindenden Zustand überführt werden (Spoerner et al., 2001).
ABBILDUNG 3.48: 31P-NMR Spektren der Titration von Ras⋅GppNHpWT mit Raf-RBDWT. Das 32P-NMR-Spektrum von Ras⋅GppNHp zeigt in Abwesenheit der Effektor-RBD ist eine Aufspaltung der ß-Phosphatlinie (Zustand 1 und 2). Auch beim α-Phosphat lässt sich anhand der Linienform eine Aufspaltung in zwei Zustände erahnen. Zustand 2 wird bei Zugabe der Effektor-RBD stabilisiert und wird daher als bindender Zustand angesehen. Die Abbildung wurde aus Geyer et al., 1996 entnommen.
Interessant wäre eine NMR-Titration von Ras⋅GppNHpT35A mit Raf-RBDA85K, um zu sehen ob es aufgrund der höheren Affinität eine Aufspaltung der Resonanzlinien am ß-Phosphat beobachtet werden kann. Die 31P-NMR-Experimente wurden in der Arbeitsgruppe von Prof. Hans Robert Kalbitzer unter Anleitung von Dr. Michael Spörner an der Universität Regensburg durchgeführt.
3.8.1.1 Bindung von Raf-RBDA85K an Ras⋅GppNHpT35A
Da die Bindung von Raf-RBDA85K jedoch auch eine deutlich höhere Affinität zu Ras⋅GppNHpT35A besitzt, sollte analysiert werde, wie die Resonanzlinien bei Titration von Raf-RBDA85K aussehen. Die Ergebnisse dieser Titration sind in Abbildung 3.49 dargestellt. Im Gegensatz zur Bindung von Raf-RBDWT beobachtet man bei der Titration von Raf-RBDA85K an Ras⋅GppNHpT35A eine Aufspaltung der Resonanzlinien am ß-Phosphat. Bereits bei einem molaren Verhältnis von 0.5 beobachtet man die Entstehung von Zustand 2. Anhand der Veränderung des Flächenverhältnisses der chemischen Verschiebungen am α-Phosphat für Zustand 1 und 2 und den Konzentrationen, können die Affinitäten der Bindungen von Raf-RBDWT und Raf-RBDA85K an Ras⋅GDP berechnet werden (Tabelle 3.24).
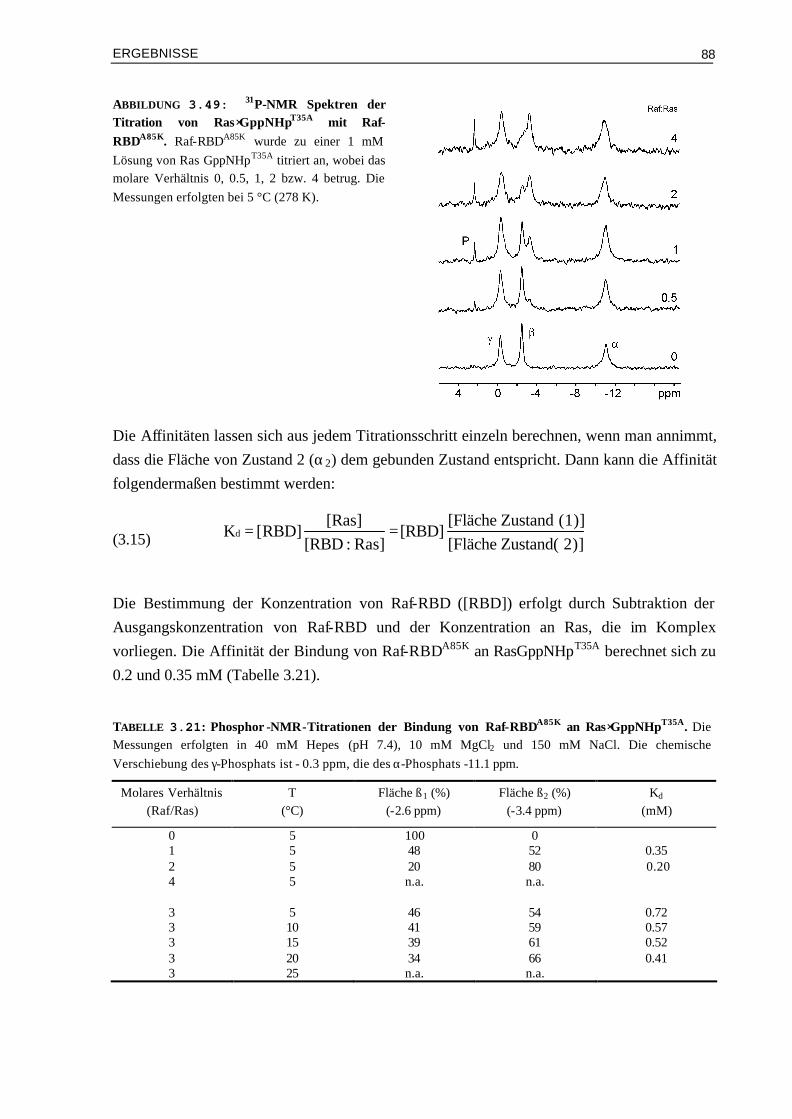
ERGEBNISSE
88
2)] Zustand([Fläche
(1)] Zustand[Fläche [RBD]
Ras]:[RBD[Ras]
]RBD[Kd ==
ABBILDUNG 3.49: 31P-NMR Spektren der Titration von Ras⋅GppNHpT35A mit Raf-RBDA85K. Raf-RBDA85K wurde zu einer 1 mM Lösung von Ras ⋅GppNHpT35A titriert an, wobei das molare Verhältnis 0, 0.5, 1, 2 bzw. 4 betrug. Die Messungen erfolgten bei 5 °C (278 K). Die Affinitäten lassen sich aus jedem Titrationsschritt einzeln berechnen, wenn man annimmt, dass die Fläche von Zustand 2 (α2) dem gebunden Zustand entspricht. Dann kann die Affinität folgendermaßen bestimmt werden:
(3.15)
Die Bestimmung der Konzentration von Raf-RBD ([RBD]) erfolgt durch Subtraktion der Ausgangskonzentration von Raf-RBD und der Konzentration an Ras, die im Komplex vorliegen. Die Affinität der Bindung von Raf-RBDA85K an RasGppNHpT35A berechnet sich zu 0.2 und 0.35 mM (Tabelle 3.21).
TABELLE 3.21: Phosphor -NMR-Titrationen der Bindung von Raf-RBDA85K an Ras⋅GppNHpT35A. Die Messungen erfolgten in 40 mM Hepes (pH 7.4), 10 mM MgCl2 und 150 mM NaCl. Die chemische Verschiebung des γ-Phosphats ist - 0.3 ppm, die des α-Phosphats -11.1 ppm.
Molares Verhältnis (Raf/Ras)
T (°C)
Fläche ß1 (%) (-2.6 ppm)
Fläche ß2 (%) (-3.4 ppm)
Kd (mM)
0 5 100 0 1 5 48 52 0.35 2 5 20 80 0.20 4 5 n.a. n.a. 3 5 46 54 0.72 3 10 41 59 0.57 3 15 39 61 0.52 3 20 34 66 0.41 3 25 n.a. n.a.
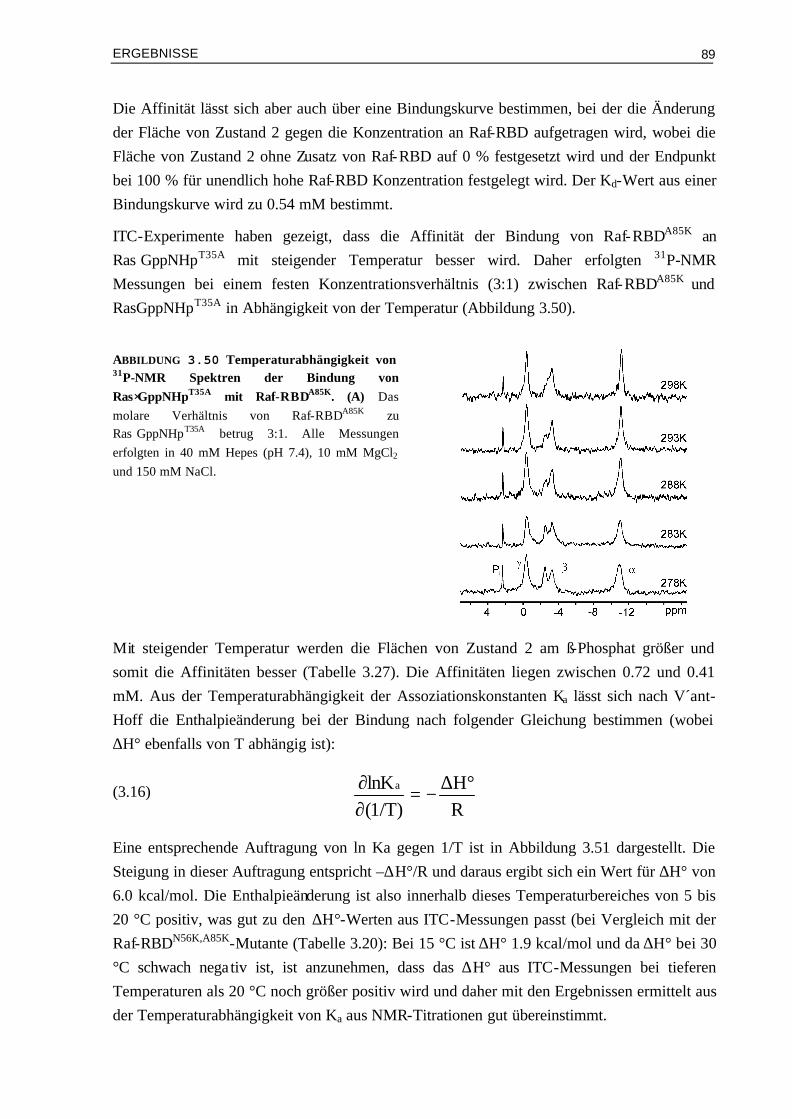
ERGEBNISSE
89
RH
1/T)(lnKa °∆
−=∂∂
Die Affinität lässt sich aber auch über eine Bindungskurve bestimmen, bei der die Änderung der Fläche von Zustand 2 gegen die Konzentration an Raf-RBD aufgetragen wird, wobei die Fläche von Zustand 2 ohne Zusatz von Raf-RBD auf 0 % festgesetzt wird und der Endpunkt bei 100 % für unendlich hohe Raf-RBD Konzentration festgelegt wird. Der Kd-Wert aus einer Bindungskurve wird zu 0.54 mM bestimmt.
ITC-Experimente haben gezeigt, dass die Affinität der Bindung von Raf-RBDA85K an Ras⋅GppNHpT35A mit steigender Temperatur besser wird. Daher erfolgten 31P-NMR Messungen bei einem festen Konzentrationsverhältnis (3:1) zwischen Raf-RBDA85K und RasGppNHpT35A in Abhängigkeit von der Temperatur (Abbildung 3.50).
ABBILDUNG 3.50 Temperaturabhängigkeit von 31P-NMR Spektren der Bindung von Ras⋅GppNHpT35A mit Raf-RBDA85K. (A) Das molare Verhältnis von Raf-RBDA85K zu Ras⋅GppNHpT35A betrug 3:1. Alle Messungen erfolgten in 40 mM Hepes (pH 7.4), 10 mM MgCl2 und 150 mM NaCl.
Mit steigender Temperatur werden die Flächen von Zustand 2 am ß-Phosphat größer und somit die Affinitäten besser (Tabelle 3.27). Die Affinitäten liegen zwischen 0.72 und 0.41 mM. Aus der Temperaturabhängigkeit der Assoziationskonstanten Ka lässt sich nach V´ant-Hoff die Enthalpieänderung bei der Bindung nach folgender Gleichung bestimmen (wobei ∆H° ebenfalls von T abhängig ist): (3.16)
Eine entsprechende Auftragung von ln Ka gegen 1/T ist in Abbildung 3.51 dargestellt. Die Steigung in dieser Auftragung entspricht –∆H°/R und daraus ergibt sich ein Wert für ∆H° von 6.0 kcal/mol. Die Enthalpieänderung ist also innerhalb dieses Temperaturbereiches von 5 bis 20 °C positiv, was gut zu den ∆H°-Werten aus ITC-Messungen passt (bei Vergleich mit der Raf-RBDN56K,A85K-Mutante (Tabelle 3.20): Bei 15 °C ist ∆H° 1.9 kcal/mol und da ∆H° bei 30 °C schwach negativ ist, ist anzunehmen, dass das ∆H° aus ITC-Messungen bei tieferen Temperaturen als 20 °C noch größer positiv wird und daher mit den Ergebnissen ermittelt aus der Temperaturabhängigkeit von Ka aus NMR-Titrationen gut übereinstimmt.

ERGEBNISSE
90
ABBILDUNG 3.51 Bestimmung der Enthalpieänderung ∆H° der Bindung von Ras⋅GppNHpT35A an Raf-RBDA85K. (A) Das molare Verhältnis von Raf-RBDA85K zu Ras⋅GppNHpT35A betrug 3:1. Alle Messungen erfolgten in 40 mM Hepes (pH 7.4), 10 mM MgCl2 und 150 mM NaCl entsprechend Tabelle 3.27. Nach Gleichung 3.16 berechnet sich ∆H° zu 6.0 kcal/mol.
3.8.1.2 Bindung von Raf-RBDWT und Raf-RBDA85K an Ras⋅GDP
Die 31P-NMR-Titration von Ras⋅GDP mit Raf-RBDA85K ist in Abbildung 3.52-A dargestellt. Ras⋅GDP ohne Raf-RBD führt zu zwei Resonanzlinien, die den Resonanzen des α-Phosphats ( = α1, bei -10.6 ppm, Zustand 1) und ß-Phosphats (bei -1.98 ppm) zuzuordnen sind. Bei Titration von Raf-RBDA85K zu der Ras⋅GDP Lösung kommt es bereits bei einem molaren Verhältnis von 0.5 zu einer deutlichen Aufspaltung der Resonanzlinien am α-Phosphat (Zustand 1 und Zustand 2) mit einer chemischen Verschiebung am α2 von -11.6 ppm. Bei einem 4-fachen molaren Verhältnis verschwindet der zu Anfang beobachtete Zustand vollständig.
A B
ABBILDUNG 3.52 31P-NMR Spektren der Titration von Ras ⋅GDP mit Raf-RBDA85K und Raf-RBDWT. (A) Raf-RBDA85K wurde zu einer 1.7 mM Lösung von Ras⋅GDP titriert, wobei das molare Verhältnis von Raf zu Ras 0, 0.5, 1, 2 bzw. 4 betrug. (B) Raf-RBDWT wurde zu einer 1.7 mM Lösung von Ras ⋅GDP titriert an, wobei das molare Verhältnis von Raf zu Ras 0, 1, 2, 4 bzw. 7 betrug. Alle Messungen erfolgten bei 20 °C (293 K) in 40 mM Hepes (pH 7.4), 10 mM MgCl2 und 150 mM NaCl.

ERGEBNISSE
91
Im Gegensatz dazu gibt es bei der Titration von Ras⋅GDP mit Raf-RBDWT keine eindeutige Aufspaltung der Resonanzlinien am α-Phosphat und sogar bei einem 7-fachen molaren Überschuss von Raf zu Ras ist der Ausgangszustand immer noch stärker populiert als der zweite Zustand (Abbildung 3.52-B). Anhand der Veränderung des Flächenverhältnisses der chemischen Verschiebungen am α-Phosphat für Zustand 1 und 2 und den Konzentrationen, können die Affinitäten der Bindungen von Raf-RBDWT und Raf-RBDA85K an Ras⋅GDP berechnet werden (Tabelle 3.22).
TABELLE 3.22: Flächen der Verschiebungen am α-Phosphat von Phosphor -NMR Titrationen der Bindung von Raf-RBDWT und Raf-RBDA85K an Ras⋅GDP. Die Messungen erfolgten bei 20 °C in 40 mM Hepes (pH 7.4), 10 mM MgCl2 und 150 mM NaCl. Die chemische Verschiebung des ß-Phosphats ist -1.98 ppm.
Die auf diese Weise bestimmten Affinitäten für die Bindung von Raf-RBDA85K an Ras⋅GDP liegen zwischen 0.88 und 0.15 mM (Tabelle 3.22). Es bestehen sehr große Abweichungen zwischen den einzelnen Werten, was insbesondere durch die großen Fehler bei der Berechnung der Flächen begründet ist. Auf diese Weise berechnet sich die Affinität der Bindung von Raf-RBDA85K an Ras⋅GDP zu 0.88 mM. Die Flächenbestimmung sowie die Affinitäten der Bindung von Raf-RBDWT an Ras⋅GDP sind in Tabelle 3.25 zusammengefasst. Die Kd-Werte liegen zwischen 2.1 und 4.5 mM. Die Affinität der Bindung von Raf-RBDWT an Ras⋅GDP bestimmt aus einer Bindungskurve beträgt 3.5 mM. Die Affinitäten bestimmt aus ITC-Messungen sind wesentlich höher als durch NMR-Titration bestimmt (0.76 bzw. 78 µM für die Bindung von Raf-RBDA85K bzw. Raf-RBDWT an Ras⋅GDP in Puffer ohne NaCl). Hier ist zu beachten, dass die Lösung bei der NMR-Titration teilweise mit jedem Titrationsschritt etwas trüber wurde, da Protein ausgefallen ist. Dadurch berechnet sich die Affinität natürlich viel zu niedrig, da die zur Berechnung der Kd-Werte verwendeten Konzentrationen nicht stimmen.
Im Folgenden sollte untersucht werden, ob bei einem Überschuss an fr eiem Phosphat dieses die Stelle des γ-Phosphats einnehmen kann. Dieses wäre dann die Beobachtung von
Raf-RBD Molares Verhältnis Raf/Ras
Fläche α1 (%) (-10.6 ppm)
Fläche α2 (%) (-11.2 ppm)
Kd (mM)
WT 0 100 0 WT 1 61 39 2.1 WT 2 59 41 2.3 WT 4 56 44 4.5 WT 7 n.a. n.a. n.a.
A85K 0 100 0 A85K 0.5 73 27 0.88 A85K 1 41 59 0.35 A85K 2 12 88 0.15 A85K 4 >90
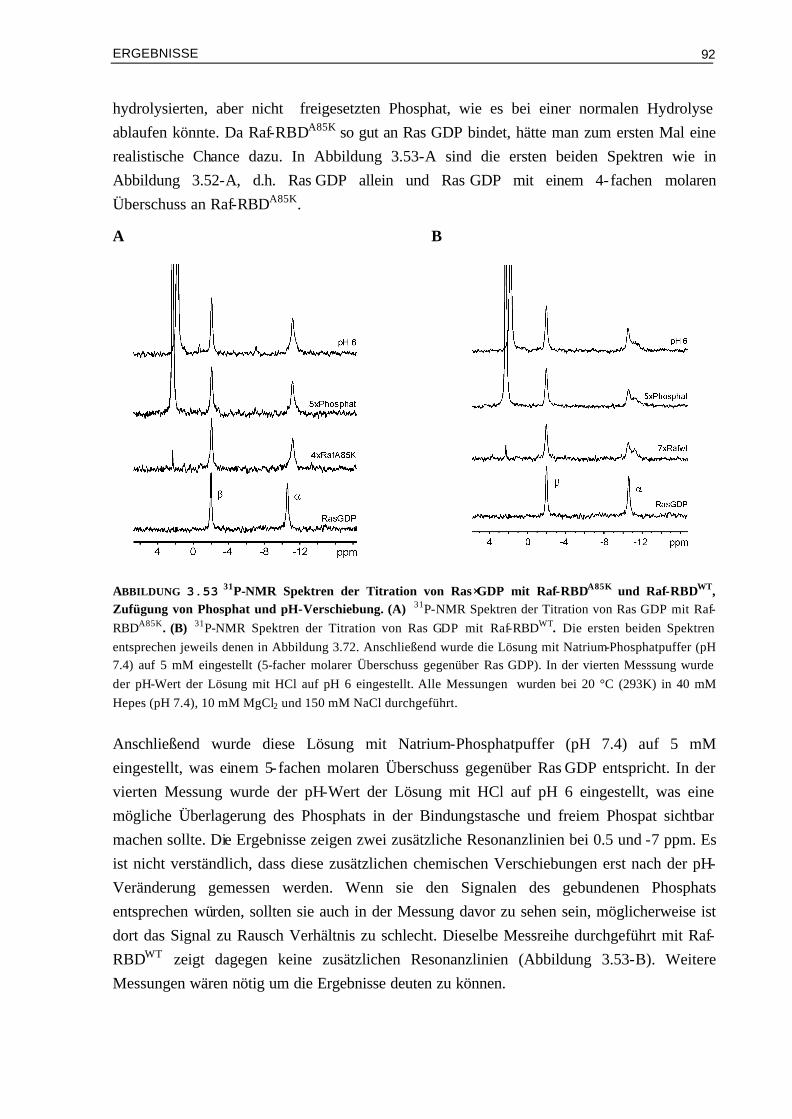
ERGEBNISSE
92
hydrolysierten, aber nicht freigesetzten Phosphat, wie es bei einer normalen Hydrolyse ablaufen könnte. Da Raf-RBDA85K so gut an Ras⋅GDP bindet, hätte man zum ersten Mal eine realistische Chance dazu. In Abbildung 3.53-A sind die ersten beiden Spektren wie in Abbildung 3.52-A, d.h. Ras⋅GDP allein und Ras⋅GDP mit einem 4-fachen molaren Überschuss an Raf-RBDA85K.
A B
ABBILDUNG 3.53 31P-NMR Spektren der Titration von Ras ⋅GDP mit Raf-RBDA85K und Raf-RBDWT, Zufügung von Phosphat und pH-Verschiebung. (A) 31P-NMR Spektren der Titration von Ras ⋅GDP mit Raf-RBDA85K. (B) 31P-NMR Spektren der Titration von Ras ⋅GDP mit Raf-RBDWT. Die ersten beiden Spektren entsprechen jeweils denen in Abbildung 3.72. Anschließend wurde die Lösung mit Natrium-Phosphatpuffer (pH 7.4) auf 5 mM eingestellt (5-facher molarer Überschuss gegenüber Ras ⋅GDP). In der vierten Messsung wurde der pH-Wert der Lösung mit HCl auf pH 6 eingestellt. Alle Messungen wurden bei 20 °C (293K) in 40 mM Hepes (pH 7.4), 10 mM MgCl2 und 150 mM NaCl durchgeführt.
Anschließend wurde diese Lösung mit Natrium-Phosphatpuffer (pH 7.4) auf 5 mM eingestellt, was einem 5-fachen molaren Überschuss gegenüber Ras⋅GDP entspricht. In der vierten Messung wurde der pH-Wert der Lösung mit HCl auf pH 6 eingestellt, was eine mögliche Überlagerung des Phosphats in der Bindungstasche und freiem Phospat sichtbar machen sollte. Die Ergebnisse zeigen zwei zusätzliche Resonanzlinien bei 0.5 und -7 ppm. Es ist nicht verständlich, dass diese zusätzlichen chemischen Verschiebungen erst nach der pH-Veränderung gemessen werden. Wenn sie den Signalen des gebundenen Phosphats entsprechen würden, sollten sie auch in der Messung davor zu sehen sein, möglicherweise ist dort das Signal zu Rausch Verhältnis zu schlecht. Dieselbe Messreihe durchgeführt mit Raf-RBDWT zeigt dagegen keine zusätzlichen Resonanzlinien (Abbildung 3.53-B). Weitere Messungen wären nötig um die Ergebnisse deuten zu können.
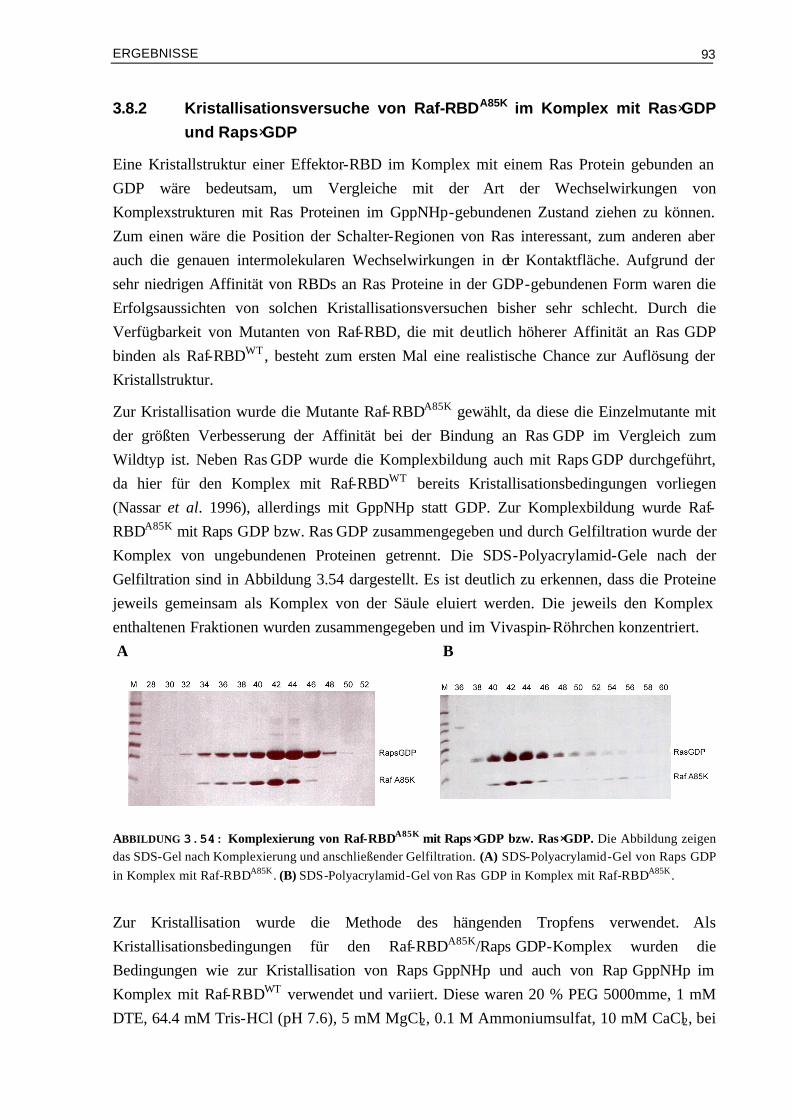
ERGEBNISSE
93
3.8.2 Kristallisationsversuche von Raf-RBDA85K im Komplex mit Ras⋅GDP und Raps⋅GDP
Eine Kristallstruktur einer Effektor-RBD im Komplex mit einem Ras Protein gebunden an GDP wäre bedeutsam, um Vergleiche mit der Art der Wechselwirkungen von Komplexstrukturen mit Ras Proteinen im GppNHp-gebundenen Zustand ziehen zu können. Zum einen wäre die Position der Schalter-Regionen von Ras interessant, zum anderen aber auch die genauen intermolekularen Wechselwirkungen in der Kontaktfläche. Aufgrund der sehr niedrigen Affinität von RBDs an Ras Proteine in der GDP-gebundenen Form waren die Erfolgsaussichten von solchen Kristallisationsversuchen bisher sehr schlecht. Durch die Verfügbarkeit von Mutanten von Raf-RBD, die mit deutlich höherer Affinität an Ras⋅GDP binden als Raf-RBDWT , besteht zum ersten Mal eine realistische Chance zur Auflösung der Kristallstruktur.
Zur Kristallisation wurde die Mutante Raf-RBDA85K gewählt, da diese die Einzelmutante mit der größten Verbesserung der Affinität bei der Bindung an Ras⋅GDP im Vergleich zum Wildtyp ist. Neben Ras⋅GDP wurde die Komplexbildung auch mit Raps⋅GDP durchgeführt, da hier für den Komplex mit Raf-RBDWT bereits Kristallisationsbedingungen vorliegen (Nassar et al. 1996), allerdings mit GppNHp statt GDP. Zur Komplexbildung wurde Raf-RBDA85K mit Raps⋅GDP bzw. Ras⋅GDP zusammengegeben und durch Gelfiltration wurde der Komplex von ungebundenen Proteinen getrennt. Die SDS-Polyacrylamid-Gele nach der Gelfiltration sind in Abbildung 3.54 dargestellt. Es ist deutlich zu erkennen, dass die Proteine jeweils gemeinsam als Komplex von der Säule eluiert werden. Die jeweils den Komplex enthaltenen Fraktionen wurden zusammengegeben und im Vivaspin-Röhrchen konzentriert. A B
ABBILDUNG 3.54: Komplexierung von Raf-RBDA85K mit Raps ⋅GDP bzw. Ras⋅GDP. Die Abbildung zeigen das SDS-Gel nach Komplexierung und anschließender Gelfiltration. (A) SDS-Polyacrylamid-Gel von Raps⋅GDP in Komplex mit Raf-RBDA85K . (B) SDS-Polyacrylamid-Gel von Ras⋅GDP in Komplex mit Raf-RBDA85K .
Zur Kristallisation wurde die Methode des hängenden Tropfens verwendet. Als Kristallisationsbedingungen für den Raf-RBDA85K/Raps⋅GDP-Komplex wurden die Bedingungen wie zur Kristallisation von Raps⋅GppNHp und auch von Rap⋅GppNHp im Komplex mit Raf-RBDWT verwendet und variiert. Diese waren 20 % PEG 5000mme, 1 mM DTE, 64.4 mM Tris-HCl (pH 7.6), 5 mM MgCl2, 0.1 M Ammoniumsulfat, 10 mM CaCl2, bei

ERGEBNISSE
94
18 °C mit 15 bis 20 mg/ml Protein (Nassar et al., 1995) und 11% PEG 4000, 2 mM DTE, 80 mM Tris-HCl (pH 7.6), 5 mM MgCl2, 20 bis 50 mM Ca-Acetat bei 18 °C mit 15 mg/ml Protein (Nassar et al., 1996). Es wurden unter verschiedenen Variationen dieser Bedingungen kleine Kristalle, zumeist nach ca. 4-7 Tagen sichtbar. Größere Kristalle (mit Ausmaßen von ca. 300 mal 50 µm) wurden mit kleineren Prozenten an PEG, aber bei Verwendung von PEG 6000 statt 4000 für den Ras⋅GDP/Raf-RBDA85K Komplex erhalten. Beispiele hierfür sind in Abbildung 3.55-A und -B dargestellt. Zur Kontrolle, das es sich wirklich um Protein-Kristalle handelt, wurde ein Kristall mit Isit angefärbt was zur Blaufärbung führte (Abbildung 3.55-C).
A B
C ABBILDUNG 3.55: Kristalle von Raf-RBDA85K im Komplex mit Ras ⋅GDP. (A) und (B) Die Kristalle wurden nach 6 Tagen Wachstum bei 18 °C in 16 % PEG 6000, 80 mM Tris (pH 6), 2 mM DTE, 5 mM MgCl2 und 50 mM Ca-Acetat erhalten. Die Konzentration des Protein-komplexes betrug 16.2 mg ml-1. Die Größe war ca. 300 mal 50 µm. (C) Die Anfärbung des Kristalls mit Isit zeigt, dass es sich um einen Proteinkristall handelt.
Leider streuten diese Kristalle nicht. Es wurden weitere Variationen der Kristallisationsbedingungen durchgeführt und es wurden die Tropfen der Kristallisationsansätze mit Impfkristallen versehen („seeding“), was zu schnellerem Kristallwachstum führt. Allerdings wurden die Kristalle bei diesem Verfahren eher kleiner. Parallel dazu wurde der Hampton Screen 1 und 2 mit 9 mg/ml Protein bei 4 °C und bei 18 °C für den Raf-RBDA85K/Raps⋅GDP-Komplex durchgeführt. Bis zum Ende dieser Arbeit konnten keine geeigneten Kristalle erhalten werden, die eine Strukturauflösung möglich machen würden.

ERGEBNISSE
95
3.8.3 Doppel-Mutanten-Analyse zur Ermittlung der energetischen Beiträge einzelner Aminosäure-Kontakte
Da die Komplexstruktur zwischen Raf-RBDA85K im Komplex mit Ras⋅GDP nicht gelöst werden konnte, wurden Doppel-Mutanten-Analysen durchgeführt, was die Ermittlung der energetischen Beiträge einzelner Aminosäure-Kontakte ermöglicht (Schreiber & Fersht, 1995). In dieser Methode wird jeweils die Affinität zwischen den Wildtyp-Proteinen, jeweils einer Alanin-Mutante mit dem anderen Wildtyp-Protein und den beiden Alanin-Mutanten verglichen, was anhand des Schemas in Abbildung 3.56 gezeigt ist. ABBILDUNG 3.56: Prinzipielles Schema eines Doppel-Mutanten-Zyklus. Der Vergleich der Affinitäten zwischen den beiden Wildtyp-Proteinen, ∆G° (RasWT/RafWT), jeweils einer Alanin-Mutante mit dem anderen Wildtyp-Protein, ∆G° (RasAlaT/RafWT), ∆G° (RasWT/RafAla) und den beiden Alanin-Mutanten liefert die Kopplungsenergie der Aminosäuren X und Y (∆∆G°int). Durch Addition und Subtraktion der Änderungen der freien Enthalpie entsprechend dem Schema in Abbildung 3.56 entspricht die Kopplungsenergie (∆∆G°int) dem energetischen Beitrag zwischen der Aminosäure X und Y; alle anderen Kontakte heben sich auf. Negative Werte für ∆∆G°int entsprechen dabei anziehenden Wechselwirkungen, positive Werte zeigen, dass die Aminosäuren sich gegenseitig abstoßen. Ist die Kopplungsenergie Null, wechselwirken die Aminosäuren X und Y nicht. Die Voraussetzung ist allerdings in allen Fällen, dass die Mutation zu Alanin zu keinen größeren strukturellen Änderungen führt.
3.8.3.1 Kopplungsenergien zwischen Lys85 von Raf-RBD und Glu31 von Ras⋅GDP
Für Lys84 von Raf-RBD konnte anhand der Struktur gezeigt werden, dass sie ionische Kontakte zu Glu31 und Asp33 von Ras (bzw. Raps) macht (Nassar et al., 1996). Außerdem konnte durch Doppel-Mutanten-Analyse ermittelt werden, dass diese Aminosäuren mit großen negativen Kopplungenergien miteinander wechselwirken (Kiel et al., eingereicht). Die Aminosäure Ala85 von Raf-RBD befindet sich neben Lys84 und daher sollte getestet werden,

ERGEBNISSE
96
ob die Mutante von Ala85 zu Lys ebenfalls mit Glu31 von Ras⋅GDP wechselwirken kann. Das Schema für diesen Doppel-Mutanten-Zyklus ist in Abbildung 3.57 dargestellt: ABBILDUNG 3.57: Doppel-Mutanten-Analyse zur Bestimmung der Kopplungsenergie zwischen den Aminosäuren Glu31 von Ras⋅GDP und Lys85 von Raf-RBD. Die entsprechenden Werte für ∆G° (∆H°, T∆S° siehe unten) wurden der Tabellen 3.14 entnommen. Die Kopplungsenergien (∆∆G°int, ∆∆H°int und T∆∆S°int) berechnen sich folgendermaßen:
∆∆G°int = ∆G° (Ras⋅GDPWT /Raf-RBDA85K) - ∆G° (Ras⋅GDPWT /Raf-RBDWT) - ∆G° (Ras⋅GDPE31A/Raf-RBDA85K) + ∆G° (Ras⋅GDPE31A/Raf-RBDWT) (-7.8 + 5.6 + 6.9 – 6.9) kcal/mol = -2.2 kcal/mol
∆∆H°int = ∆H° (Ras⋅GDPWT /Raf-RBDA85K) - ∆H° (Ras⋅GDPWT /Raf-RBDWT) - ∆H° (Ras⋅GDPE31A/Raf-RBDA85K) + ∆H° (Ras⋅GDPE31A/Raf-RBDWT) = (- 6.3 + 5.1 + 2.4 - 0.8) kcal/mol = 0.4 kcal/mol
Τ∆∆S°int = T∆S° (Ras⋅GDPWT /Raf-RBDA85K) - T∆S° (Ras⋅GDPWT /Raf-RBDWT) - T∆S° (Ras⋅GDPE31A/Raf-RBDA85K) + T∆S° (Ras⋅GDPE31A/Raf-RBDWT) = (1.5 - 0.5 - 4.5 + 6.1) kcal/mol = 2.6 kcal/mol
Es bestehen somit anziehende Wechselwirkungen zwischen Lys85 von Raf-RBD und Glu31 von Ras⋅GDP mit einer Kopplungsenergie von ∆∆G°int von -2.2 kcal/mol. Der Vergleich der Werte für ∆∆H°int und T∆∆S°int zeigt, dass die günstigen Wechselwirkungen hauptsächlich die Folge von günstigen Entropieänderungen ist.
3.8.3.2 Kopplungsenergien zwischen Lys85 von Raf-RBD und Glu31 von
Ras⋅GppNHp
Das Schema des Doppel-Mutanten-Zyklus zur Bestimmung der Kopplungsenergie zwischen Lys85 von Raf-RBD und Glu31 von Ras⋅GppNHp ist in Abbildung 3.58 dargestellt. Die Kopplungsenergien (∆∆G°int, ∆∆H°int und T∆∆S°int) berechnen sich wie folgt:

ERGEBNISSE
97
∆∆G°int = ∆G° (Ras⋅GppNHpWT /Raf-RBDA85K) - ∆G° (Ras⋅GppNHpWT /Raf-RBDWT) - ∆G° (Ras⋅GppNHpE31A/Raf-RBDA85K) + ∆G° (Ras⋅GppNHpE31A/Raf-RBDWT) (-10.8 + 10.9 + 9.1 – 10.6) kcal/mol = -1.4 kcal/mol
∆∆H°int = ∆H° (Ras⋅GppNHpWT /Raf-RBDA85K) - ∆H° (Ras⋅GppNHpWT /Raf-RBDWT) - ∆H° (Ras⋅GppNHpE31A/Raf-RBDA85K) + ∆H° (Ras⋅GppNHpE31A/Raf-RBDWT) = (-6.4 + 5.5 + 2.6 – 2.4) kcal/mol = -0.7 kcal/mol
Τ∆∆S°int = T∆S° (Ras⋅GppNHpWT /Raf-RBDA85K) - T∆S° (Ras⋅GppNHpWT /Raf-RBDWT) - T∆S° (Ras⋅GppNHpE31A/Raf-RBDA85K) + T∆S° (Ras⋅GppNHpE31A/Raf-RBDWT) = (4.4 – 5.4 – 6.5 + 8.2) kcal/mol = 0.7 kcal/mol ABBILDUNG 3.58: Doppel-Mutanten-Analyse zur Bestimmung der Kopplungsenergie zwischen den Aminosäuren Glu31 von Ras⋅GppNHp und Lys85 von Raf-RBD. Die entsprechenden Werte für ∆G° (∆H°, T∆S° siehe unten) bei 25 °C wurden der Tabelle 3.12 entnommen. Es bestehen somit ebenfalls negative, d.h. günstige Kopplungsenergien für ∆∆G°int zwischen Lys85 von Raf-RBD und Glu31 von Ras im Komplex mit GppNHp (∆∆G°int, = -1.4 kcal/mol), allerdings sind diese geringer als im Komplex mit Ras gebunden an GDP (∆∆G°int, = -2.2 kcal/mol). Die Beiträge von ∆∆H°int und Τ∆∆S°int tragen dabei mit dem gleichen Betrag zu ∆∆G°int bei.
3.8.3.3 Kopplungsenergien zwischen Lys85 von Raf-RBD und dem ?-Phosphat von
Ras⋅GppNHp
Mit einem weiteren Doppel-Mutanten-Zyklus sollte analysiert werden, ob günstige Wechselwirkungen zwischen Lys85 von Raf-RBD und dem γ-Phosphat von Ras⋅GppNHp bestehen. Das Schema des entsprechenden Zyklus ist in Abbildung 3.59 dargestellt. Hier werden somit Messungen von RafRBDA85K und Raf-RBDWT mit Ras im GDP- und GppNHp-gebundenen Zustand miteinander verglichen.

ERGEBNISSE
98
ABBILDUNG 3.59: Doppel-Mutanten Analyse zur Bestimmung der Kopplungsenergie zwischen dem γ-Phosphat und Lys85 von Raf-RBD. Die entsprechenden Werte für ∆G° (∆H°, T∆S° siehe unten) bei 25 °C wurden den Tabellen 3.12 und 3.14 entnommen.
Die Kopplungsenergien (∆∆G°int, ∆∆H°int und T∆∆S°int) berechnen sich zu folgenden Werten:
∆∆G°int = ∆G°(Ras⋅GppNHp/Raf-RBDA85K) - ∆G°(Ras⋅GDP/Raf-RBDA85K) - ∆G°(Ras⋅GppNHp/Raf-RBDWT) + ∆G°(Ras⋅GDP/Raf-RBDWT) = (-10.8 + 7.7 + 10.9 – 5.6) kcal/mol = 2.3 kcal/mol
∆∆H°int = ∆H°(Ras⋅GppNHp/Raf-RBDA85K) - ∆H°(Ras⋅GDP/Raf-RBDA85K) - ∆H°(Ras⋅GppNHp/Raf-RBDWT) + ∆H°(Ras⋅GDP/Raf-RBDWT) = (-6.4 + 6.3 + 5.5 –5.1) kcal/mol = 0.3 kcal/mol
Τ∆∆S°int = T∆S°(Ras⋅GppNHp/Raf-RBDA85K) - T∆S°(Ras⋅GDP/Raf-RBDA85K) - T∆S°(Ras⋅GppNHp/Raf-RBDWT) + T∆S°(Ras⋅GDP/Raf-RBDWT) = (4.4 – 1.5 –5.4 + 0.5) kcal/mol = -2.0 kcal/mol Es bestehen somit abstoßende Wechselwirkungen zwischen Lys85 von Raf-RBD und dem γ-Phosphat von Ras⋅GppNHp. Dies Ergebnis ist erstaunlich, da man entweder keine Wechselwirkungen (D.h ∆∆G°int wäre ungefähr Null) oder eine negative Kopplungsenergie für ∆∆G°int (aufgrund der gegensätzlichen Ladungen) erwarten würde. Eine mögliche Erklärung ist, dass es zu sterischen Hinderungen kommt, worauf die ungünstige Kopplungsenergie der Entropieänderung ein Hinweis sein könnte. Wahrscheinlicher ist jedoch, dass die strukturellen Unterschiede zwischen Ras in der GDP- und der GppNHp-gebundenen Form die Ursache ist, so dass die Voraussetzungen für die Doppel-Mutanten-Analyse (keine größeren strukturellen Änderungen) nicht gegeben sind.

ERGEBNISSE
99
3.9 Zelluläre Signalweiterleitung von Ras/Effektor-Mutanten-Komplexen mit veränderter Elektrostatik
Nach umfangreicher kinetischer und thermodynamischer Charakterisierung der Effektor-RBD-Mutanten im Komplex mit Ras, sollten die RalGDS- und Raf-Mutanten mit veränderten Eigenschaften auf ihre Fähigkeit zur Signalweiterleitung in der Zelle analysiert werden.
3.9.1 Der Ral-Beladungs-Assay zur Quantifizierung der Signalweiterleitung
über RalGDS
In diesem Assay erfolgte die Messung der Aktivität von RalGDS über die GTP-Beladung der GTPase RalA, da RalGDS als Austauschfaktor für RalA wirkt. Es konnte gezeigt werden, dass bei Koexpression von RalGDS und RalA in COS-7 Zellen der GTP-Gehalt von RalA signifikant über den basalen GTP-Gehalt auf ca. 23 % erhöht wird (Urano et al., 1996; Linnemann et al., 2002). Dieser Gehalt kann auf 37 % gesteigert werden, wenn zusätzlich RasG12V als konstitutiv aktive Ras Mutante kotransfiziert wird. Wird dagegen RalGDS mit einer Mutation innerhalb der RBD (Lys48 zu Ala), die in vitro zu einer deutlich verringerten Affinität im Komplex mit Ras⋅GppNHp führt, exprimiert, kommt es zu keiner erhöhten GTP-Beladung von RalA bei Kotransfektion mit RasG12V (Abbildung 3.60). Der Assay funktioniert qualitativ also sehr überzeugend, aber das experimentelle Fenster ist sehr klein (die Signaländerung erfolgt von 23 auf 37 %, also nicht einmal 2- fach).
ABBILDUNG 3.60: Der Ral-Beladungs-Assay zur Quantifizierung der Ras-vermittelten Stimulierung von RalGDS. In jedem Experiment wurden 1 µg pMT2:RalA transfiziert. Zusätzlich wurden jeweils 1 µg pcDNA3:RalGDSWT bzw. pcDNA3:RalGDSK835A (K835 entspricht der Numerierung K48 in dieser Arbeit) und 3 µg pSVK3:RasG12V wie beschriftet kotransfiziert. Nach Immunpräzipitation von RalA wurden die gebundenen Nukleotide durch Dünnschicht-chromatographie getrennt und im Phosphor-Imager sichtbar gemacht und quantifiziert (oberer Teil der Abbildung). Die daraus bestimmte GTP-Beladung ist im unteren Teil der Abbildung dargestellt. Außerdem ist ein Western-Blot (mit dem monoklonalen Antikörper α-RG139) von RalGDSWT und RalGDSK835A dargestellt, was zeigt, dass die beiden Konstrukte vergleichbar exprimiert werden.
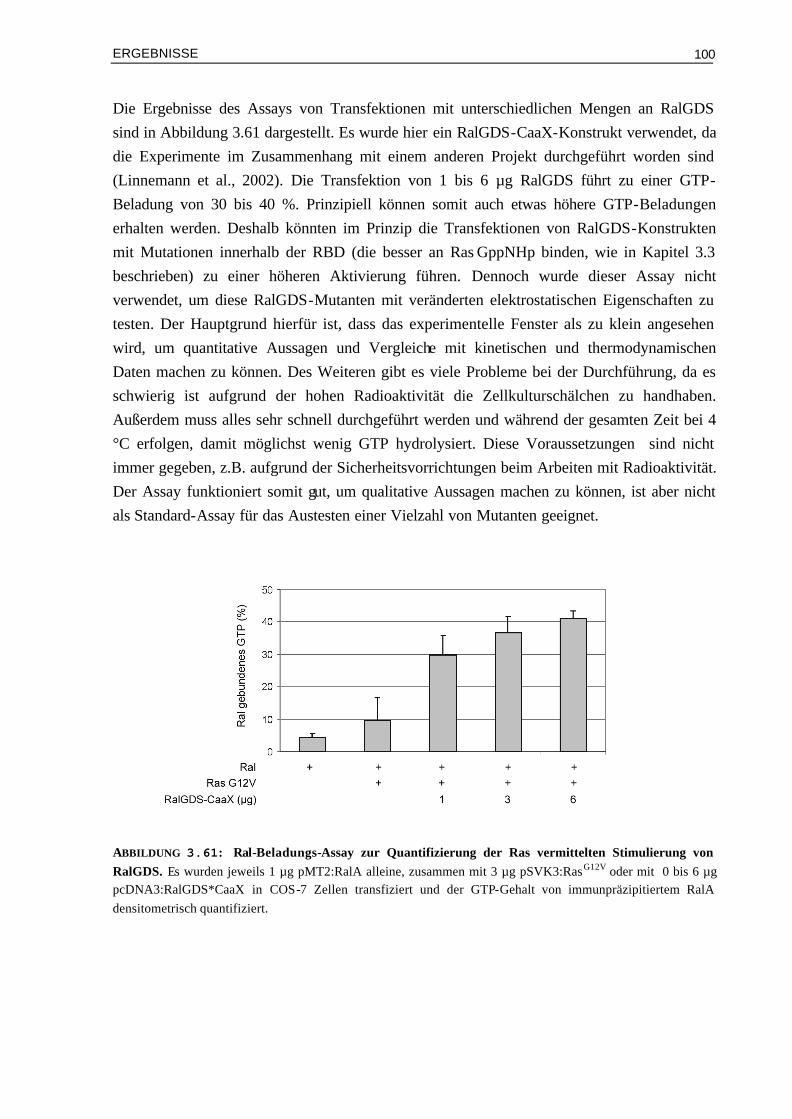
ERGEBNISSE
100
Die Ergebnisse des Assays von Transfektionen mit unterschiedlichen Mengen an RalGDS sind in Abbildung 3.61 dargestellt. Es wurde hier ein RalGDS-CaaX-Konstrukt verwendet, da die Experimente im Zusammenhang mit einem anderen Projekt durchgeführt worden sind (Linnemann et al., 2002). Die Transfektion von 1 bis 6 µg RalGDS führt zu einer GTP-Beladung von 30 bis 40 %. Prinzipiell können somit auch etwas höhere GTP-Beladungen erhalten werden. Deshalb könnten im Prinzip die Transfektionen von RalGDS-Konstrukten mit Mutationen innerhalb der RBD (die besser an Ras⋅GppNHp binden, wie in Kapitel 3.3 beschrieben) zu einer höheren Aktivierung führen. Dennoch wurde dieser Assay nicht verwendet, um diese RalGDS-Mutanten mit veränderten elektrostatischen Eigenschaften zu testen. Der Hauptgrund hierfür ist, dass das experimentelle Fenster als zu klein angesehen wird, um quantitative Aussagen und Vergleiche mit kinetischen und thermodynamischen Daten machen zu können. Des Weiteren gibt es viele Probleme bei der Durchführung, da es schwierig ist aufgrund der hohen Radioaktivität die Zellkulturschälchen zu handhaben. Außerdem muss alles sehr schnell durchgeführt werden und während der gesamten Zeit bei 4 °C erfolgen, damit möglichst wenig GTP hydrolysiert. Diese Voraussetzungen sind nicht immer gegeben, z.B. aufgrund der Sicherheitsvorrichtungen beim Arbeiten mit Radioaktivität. Der Assay funktioniert somit gut, um qualitative Aussagen machen zu können, ist aber nicht als Standard-Assay für das Austesten einer Vielzahl von Mutanten geeignet.
ABBILDUNG 3.61: Ral-Beladungs-Assay zur Quantifizierung der Ras vermittelten Stimulierung von RalGDS. Es wurden jeweils 1 µg pMT2:RalA alleine, zusammen mit 3 µg pSVK3:RasG12V oder mit 0 bis 6 µg pcDNA3:RalGDS*CaaX in COS-7 Zellen transfiziert und der GTP-Gehalt von immunpräzipitiertem RalA densitometrisch quantifiziert.
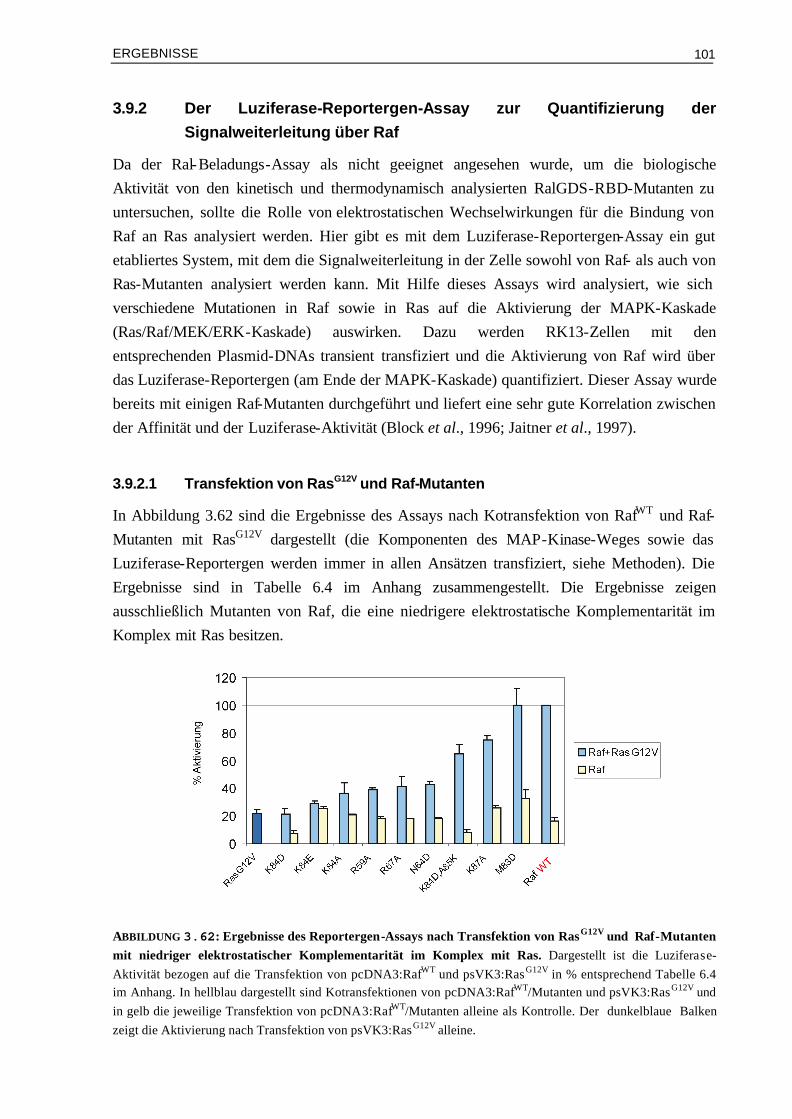
ERGEBNISSE
101
3.9.2 Der Luziferase-Reportergen-Assay zur Quantifizierung der Signalweiterleitung über Raf
Da der Ral-Beladungs-Assay als nicht geeignet angesehen wurde, um die biologische Aktivität von den kinetisch und thermodynamisch analysierten RalGDS-RBD-Mutanten zu untersuchen, sollte die Rolle von elektrostatischen Wechselwirkungen für die Bindung von Raf an Ras analysiert werden. Hier gibt es mit dem Luziferase-Reportergen-Assay ein gut etabliertes System, mit dem die Signalweiterleitung in der Zelle sowohl von Raf- als auch von Ras-Mutanten analysiert werden kann. Mit Hilfe dieses Assays wird analysiert, wie sich verschiedene Mutationen in Raf sowie in Ras auf die Aktivierung der MAPK-Kaskade (Ras/Raf/MEK/ERK-Kaskade) auswirken. Dazu werden RK13-Zellen mit den entsprechenden Plasmid-DNAs transient transfiziert und die Aktivierung von Raf wird über das Luziferase-Reportergen (am Ende der MAPK-Kaskade) quantifiziert. Dieser Assay wurde bereits mit einigen Raf-Mutanten durchgeführt und liefert eine sehr gute Korrelation zwischen der Affinität und der Luziferase-Aktivität (Block et al., 1996; Jaitner et al., 1997).
3.9.2.1 Transfektion von RasG12V und Raf-Mutanten
In Abbildung 3.62 sind die Ergebnisse des Assays nach Kotransfektion von RafWT und Raf-Mutanten mit RasG12V dargestellt (die Komponenten des MAP-Kinase-Weges sowie das Luziferase-Reportergen werden immer in allen Ansätzen transfiziert, siehe Methoden). Die Ergebnisse sind in Tabelle 6.4 im Anhang zusammengestellt. Die Ergebnisse zeigen ausschließlich Mutanten von Raf, die eine niedrigere elektrostatische Komplementarität im Komplex mit Ras besitzen.
ABBILDUNG 3.62: Ergebnisse des Reportergen-Assays nach Transfektion von RasG12V und Raf-Mutanten mit niedriger elektrostatischer Komplementarität im Komplex mit Ras. Dargestellt ist die Luziferase-Aktivität bezogen auf die Transfektion von pcDNA3:RafWT und psVK3:RasG12V in % entsprechend Tabelle 6.4 im Anhang. In hellblau dargestellt sind Kotransfektionen von pcDNA3:RafWT/Mutanten und psVK3:RasG12V und in gelb die jeweilige Transfektion von pcDNA3:RafWT/Mutanten alleine als Kontrolle. Der dunkelblaue Balken zeigt die Aktivierung nach Transfektion von psVK3:RasG12V alleine.
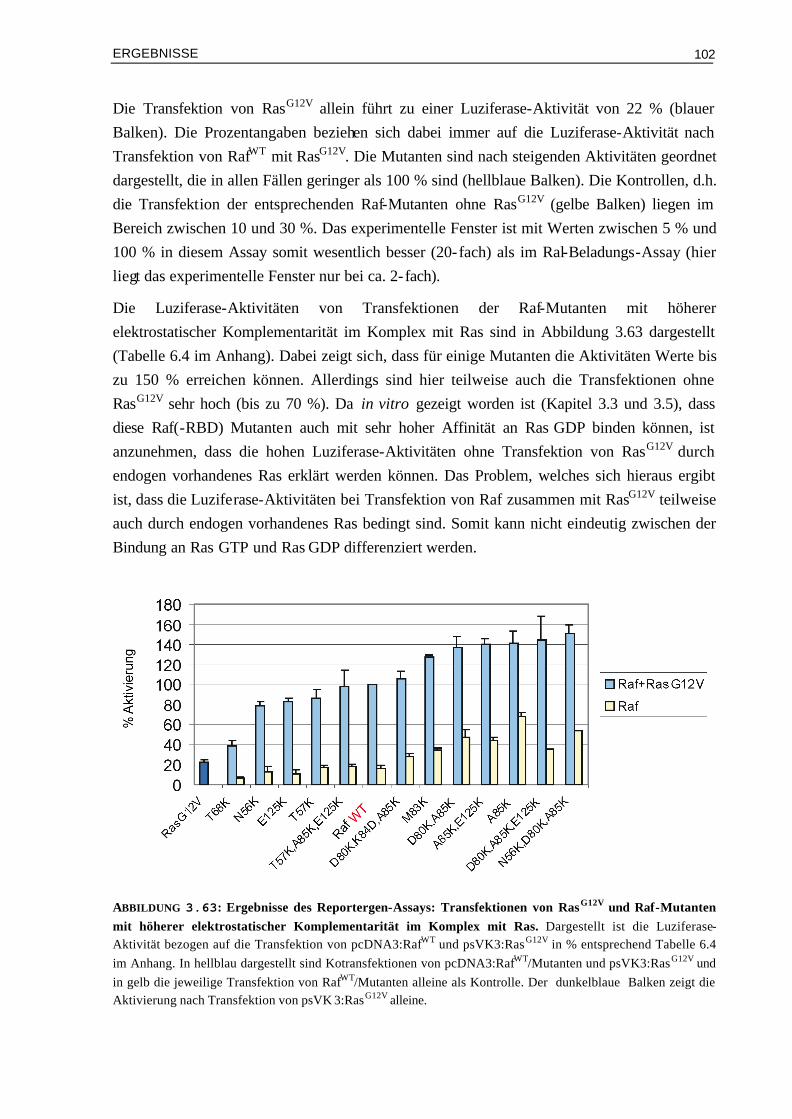
ERGEBNISSE
102
Die Transfektion von RasG12V allein führt zu einer Luziferase-Aktivität von 22 % (blauer Balken). Die Prozentangaben beziehen sich dabei immer auf die Luziferase-Aktivität nach Transfektion von RafWT mit RasG12V. Die Mutanten sind nach steigenden Aktivitäten geordnet dargestellt, die in allen Fällen geringer als 100 % sind (hellblaue Balken). Die Kontrollen, d.h. die Transfektion der entsprechenden Raf-Mutanten ohne RasG12V (gelbe Balken) liegen im Bereich zwischen 10 und 30 %. Das experimentelle Fenster ist mit Werten zwischen 5 % und 100 % in diesem Assay somit wesentlich besser (20-fach) als im Ral-Beladungs-Assay (hier liegt das experimentelle Fenster nur bei ca. 2-fach).
Die Luziferase-Aktivitäten von Transfektionen der Raf-Mutanten mit höherer elektrostatischer Komplementarität im Komplex mit Ras sind in Abbildung 3.63 dargestellt (Tabelle 6.4 im Anhang). Dabei zeigt sich, dass für einige Mutanten die Aktivitäten Werte bis zu 150 % erreichen können. Allerdings sind hier teilweise auch die Transfektionen ohne RasG12V sehr hoch (bis zu 70 %). Da in vitro gezeigt worden ist (Kapitel 3.3 und 3.5), dass diese Raf(-RBD) Mutanten auch mit sehr hoher Affinität an Ras⋅GDP binden können, ist anzunehmen, dass die hohen Luziferase-Aktivitäten ohne Transfektion von RasG12V durch endogen vorhandenes Ras erklärt werden können. Das Problem, welches sich hieraus ergibt ist, dass die Luziferase-Aktivitäten bei Transfektion von Raf zusammen mit RasG12V teilweise auch durch endogen vorhandenes Ras bedingt sind. Somit kann nicht eindeutig zwischen der Bindung an Ras⋅GTP und Ras⋅GDP differenziert werden.
ABBILDUNG 3.63: Ergebnisse des Reportergen-Assays: Transfektionen von RasG12V und Raf-Mutanten mit höherer elektrostatischer Komplementarität im Komplex mit Ras. Dargestellt ist die Luziferase-Aktivität bezogen auf die Transfektion von pcDNA3:RafWT und psVK3:RasG12V in % entsprechend Tabelle 6.4 im Anhang. In hellblau dargestellt sind Kotransfektionen von pcDNA3:RafWT/Mutanten und psVK3:RasG12V und in gelb die jeweilige Transfektion von RafWT/Mutanten alleine als Kontrolle. Der dunkelblaue Balken zeigt die Aktivierung nach Transfektion von psVK 3:RasG12V alleine.
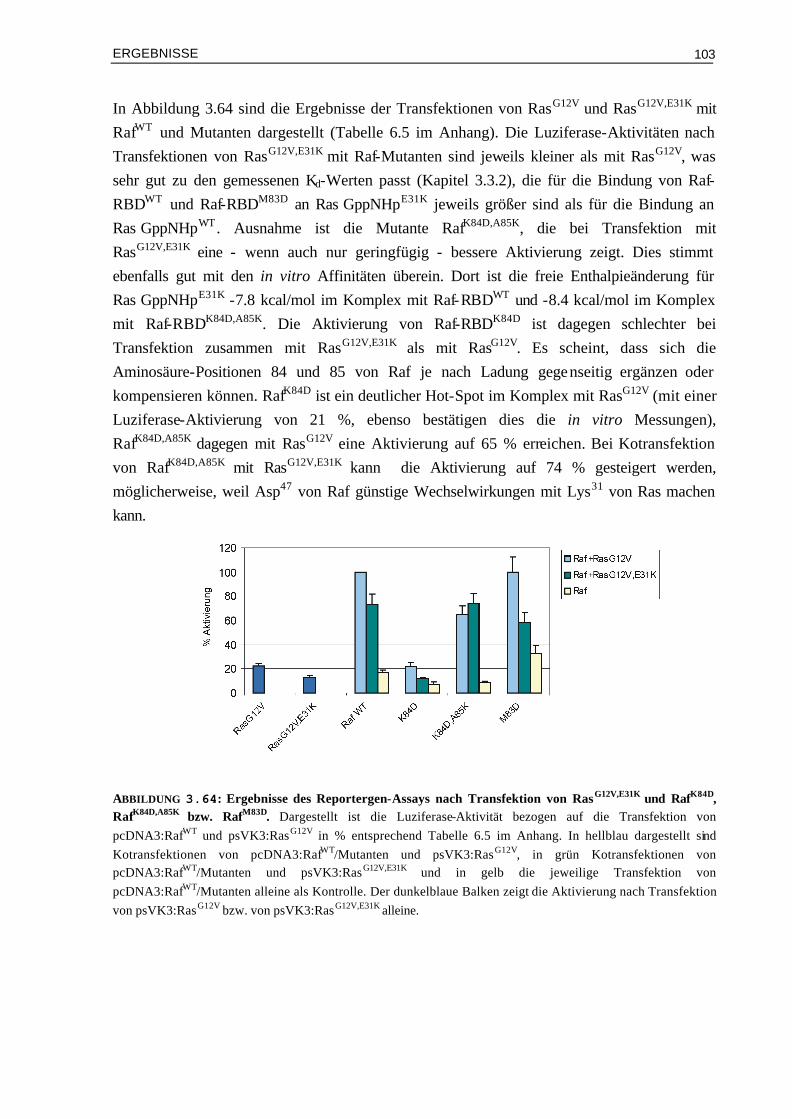
ERGEBNISSE
103
In Abbildung 3.64 sind die Ergebnisse der Transfektionen von RasG12V und RasG12V,E31K mit RafWT und Mutanten dargestellt (Tabelle 6.5 im Anhang). Die Luziferase-Aktivitäten nach Transfektionen von RasG12V,E31K mit Raf-Mutanten sind jeweils kleiner als mit RasG12V, was sehr gut zu den gemessenen Kd-Werten passt (Kapitel 3.3.2), die für die Bindung von Raf-RBDWT und Raf-RBDM83D an Ras⋅GppNHpE31K jeweils größer sind als für die Bindung an Ras⋅GppNHpWT . Ausnahme ist die Mutante RafK84D,A85K, die bei Transfektion mit RasG12V,E31K eine - wenn auch nur geringfügig - bessere Aktivierung zeigt. Dies stimmt ebenfalls gut mit den in vitro Affinitäten überein. Dort ist die freie Enthalpieänderung für Ras⋅GppNHpE31K -7.8 kcal/mol im Komplex mit Raf-RBDWT und -8.4 kcal/mol im Komplex mit Raf-RBDK84D,A85K. Die Aktivierung von Raf-RBDK84D ist dagegen schlechter bei Transfektion zusammen mit RasG12V,E31K als mit RasG12V. Es scheint, dass sich die Aminosäure-Positionen 84 und 85 von Raf je nach Ladung gegenseitig ergänzen oder kompensieren können. RafK84D ist ein deutlicher Hot-Spot im Komplex mit RasG12V (mit einer Luziferase-Aktivierung von 21 %, ebenso bestätigen dies die in vitro Messungen), RafK84D,A85K dagegen mit RasG12V eine Aktivierung auf 65 % erreichen. Bei Kotransfektion von RafK84D,A85K mit RasG12V,E31K kann die Aktivierung auf 74 % gesteigert werden, möglicherweise, weil Asp47 von Raf günstige Wechselwirkungen mit Lys31 von Ras machen kann. ABBILDUNG 3.64: Ergebnisse des Reportergen-Assays nach Transfektion von RasG12V,E31K und RafK84D, RafK84D,A85K bzw. RafM83D. Dargestellt ist die Luziferase-Aktivität bezogen auf die Transfektion von pcDNA3:RafWT und psVK3:RasG12V in % entsprechend Tabelle 6.5 im Anhang. In hellblau dargestellt sind Kotransfektionen von pcDNA3:RafWT/Mutanten und psVK3:RasG12V, in grün Kotransfektionen von pcDNA3:RafWT/Mutanten und psVK3:RasG12V,E31K und in gelb die jeweilige Transfektion von pcDNA3:RafWT/Mutanten alleine als Kontrolle. Der dunkelblaue Balken zeigt die Aktivierung nach Transfektion von psVK3:RasG12V bzw. von psVK3:RasG12V,E31K alleine.
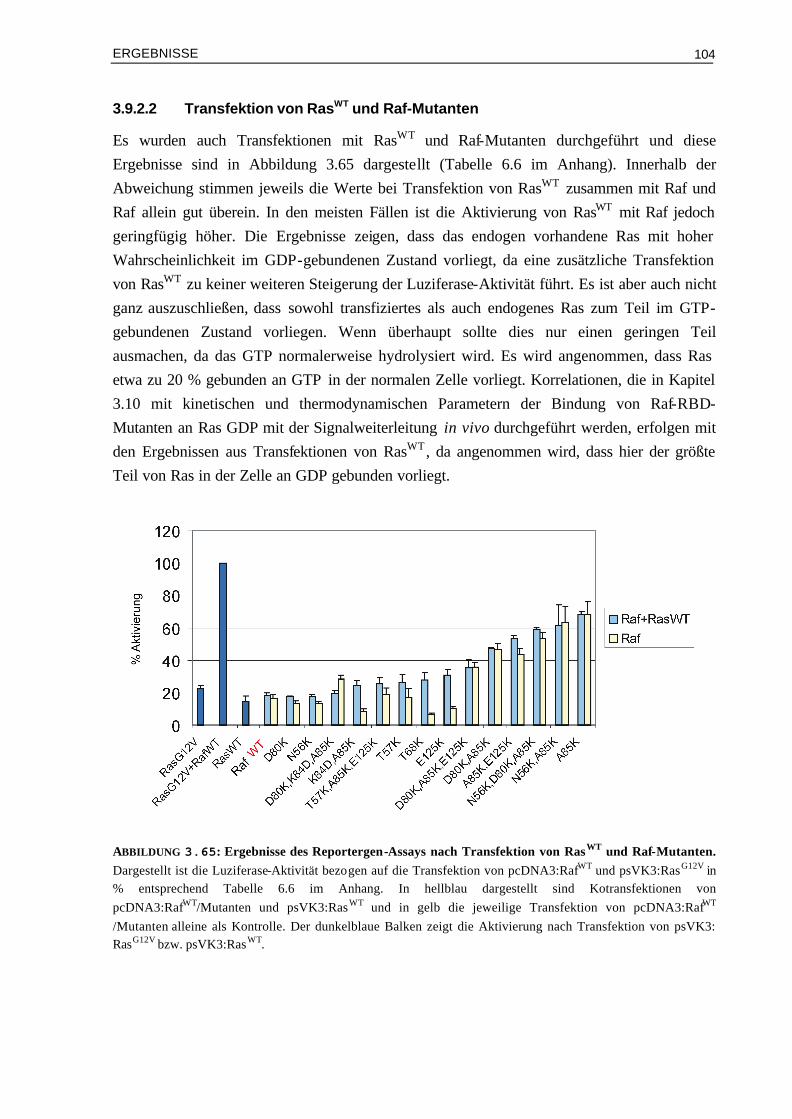
ERGEBNISSE
104
3.9.2.2 Transfektion von RasWT und Raf-Mutanten
Es wurden auch Transfektionen mit RasWT und Raf-Mutanten durchgeführt und diese Ergebnisse sind in Abbildung 3.65 dargestellt (Tabelle 6.6 im Anhang). Innerhalb der Abweichung stimmen jeweils die Werte bei Transfektion von RasWT zusammen mit Raf und Raf allein gut überein. In den meisten Fällen ist die Aktivierung von RasWT mit Raf jedoch geringfügig höher. Die Ergebnisse zeigen, dass das endogen vorhandene Ras mit hoher Wahrscheinlichkeit im GDP-gebundenen Zustand vorliegt, da eine zusätzliche Transfektion von RasWT zu keiner weiteren Steigerung der Luziferase-Aktivität führt. Es ist aber auch nicht ganz auszuschließen, dass sowohl transfiziertes als auch endogenes Ras zum Teil im GTP-gebundenen Zustand vorliegen. Wenn überhaupt sollte dies nur einen geringen Teil ausmachen, da das GTP normalerweise hydrolysiert wird. Es wird angenommen, dass Ras etwa zu 20 % gebunden an GTP in der normalen Zelle vorliegt. Korrelationen, die in Kapitel 3.10 mit kinetischen und thermodynamischen Parametern der Bindung von Raf-RBD-Mutanten an Ras⋅GDP mit der Signalweiterleitung in vivo durchgeführt werden, erfolgen mit den Ergebnissen aus Transfektionen von RasWT , da angenommen wird, dass hier der größte Teil von Ras in der Zelle an GDP gebunden vorliegt.
ABBILDUNG 3.65: Ergebnisse des Reportergen-Assays nach Transfektion von RasWT und Raf-Mutanten. Dargestellt ist die Luziferase-Aktivität bezogen auf die Transfektion von pcDNA3:RafWT und psVK3:RasG12V in % entsprechend Tabelle 6.6 im Anhang. In hellblau dargestellt sind Kotransfektionen von pcDNA3:RafWT/Mutanten und psVK3:RasWT und in gelb die jeweilige Transfektion von pcDNA3:RafWT
/Mutanten alleine als Kontrolle. Der dunkelblaue Balken zeigt die Aktivierung nach Transfektion von psVK3: RasG12V bzw. psVK3:RasWT.
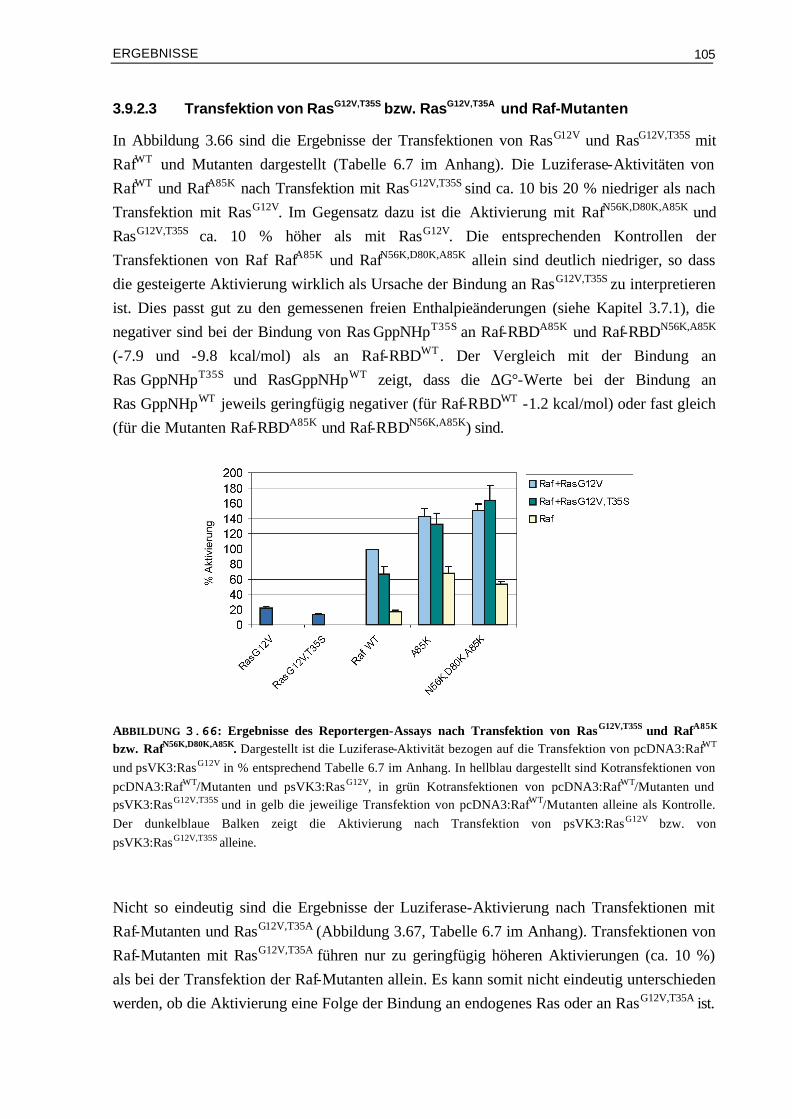
ERGEBNISSE
105
3.9.2.3 Transfektion von RasG12V,T35S bzw. RasG12V,T35A und Raf-Mutanten
In Abbildung 3.66 sind die Ergebnisse der Transfektionen von RasG12V und RasG12V,T35S mit RafWT und Mutanten dargestellt (Tabelle 6.7 im Anhang). Die Luziferase-Aktivitäten von RafWT und RafA85K nach Transfektion mit RasG12V,T35S sind ca. 10 bis 20 % niedriger als nach Transfektion mit RasG12V. Im Gegensatz dazu ist die Aktivierung mit RafN56K,D80K,A85K und RasG12V,T35S ca. 10 % höher als mit RasG12V. Die entsprechenden Kontrollen der Transfektionen von Raf RafA85K und RafN56K,D80K,A85K allein sind deutlich niedriger, so dass die gesteigerte Aktivierung wirklich als Ursache der Bindung an RasG12V,T35S zu interpretieren ist. Dies passt gut zu den gemessenen freien Enthalpieänderungen (siehe Kapitel 3.7.1), die negativer sind bei der Bindung von Ras⋅GppNHpT35S an Raf-RBDA85K und Raf-RBDN56K,A85K
(-7.9 und -9.8 kcal/mol) als an Raf-RBDWT . Der Vergleich mit der Bindung an Ras⋅GppNHpT35S und RasGppNHpWT zeigt, dass die ∆G°-Werte bei der Bindung an Ras⋅GppNHpWT jeweils geringfügig negativer (für Raf-RBDWT -1.2 kcal/mol) oder fast gleich (für die Mutanten Raf-RBDA85K und Raf-RBDN56K,A85K) sind. ABBILDUNG 3.66: Ergebnisse des Reportergen-Assays nach Transfektion von RasG12V,T35S und RafA85K bzw. RafN56K,D80K,A85K. Dargestellt ist die Luziferase-Aktivität bezogen auf die Transfektion von pcDNA3:RafWT und psVK3:RasG12V in % entsprechend Tabelle 6.7 im Anhang. In hellblau dargestellt sind Kotransfektionen von pcDNA3:RafWT/Mutanten und psVK3:RasG12V, in grün Kotransfektionen von pcDNA3:RafWT/Mutanten und psVK3:RasG12V,T35S und in gelb die jeweilige Transfektion von pcDNA3:RafWT/Mutanten alleine als Kontrolle. Der dunkelblaue Balken zeigt die Aktivierung nach Transfektion von psVK3:RasG12V bzw. von psVK3:RasG12V,T35S alleine. Nicht so eindeutig sind die Ergebnisse der Luziferase-Aktivierung nach Transfektionen mit Raf-Mutanten und RasG12V,T35A (Abbildung 3.67, Tabelle 6.7 im Anhang). Transfektionen von Raf-Mutanten mit RasG12V,T35A führen nur zu geringfügig höheren Aktivierungen (ca. 10 %) als bei der Transfektion der Raf-Mutanten allein. Es kann somit nicht eindeutig unterschieden werden, ob die Aktivierung eine Folge der Bindung an endogenes Ras oder an RasG12V,T35A ist.
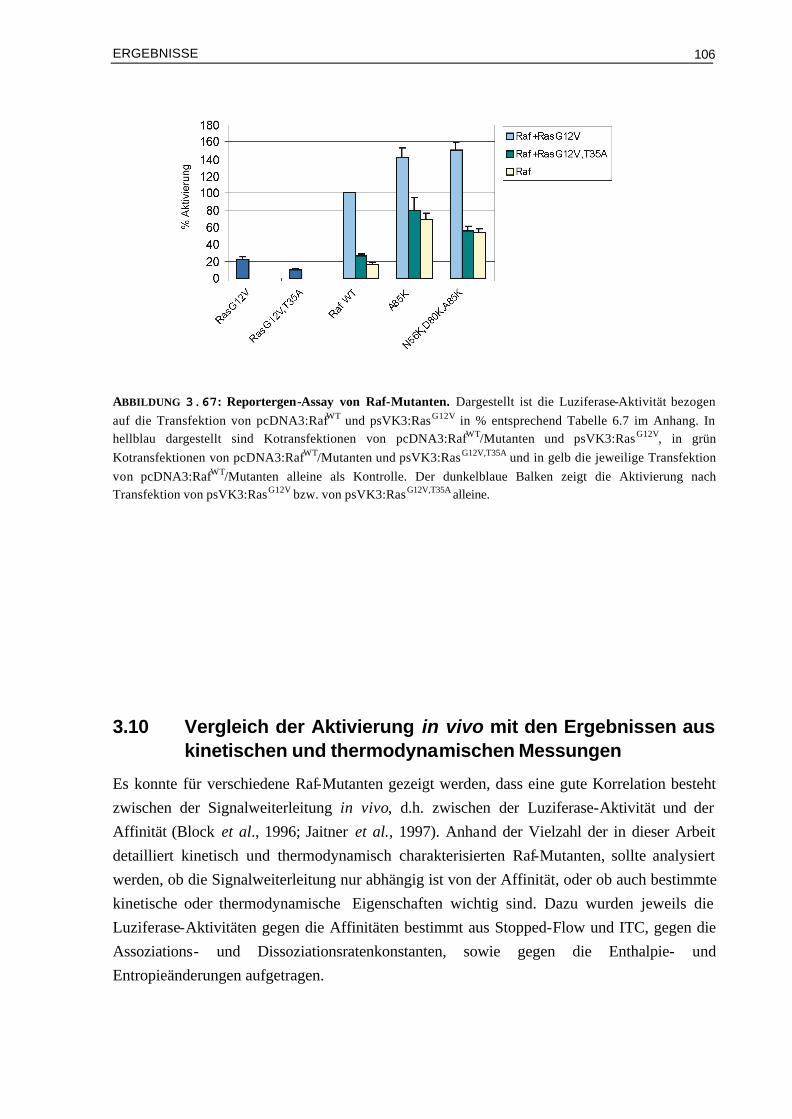
ERGEBNISSE
106
ABBILDUNG 3.67: Reportergen-Assay von Raf-Mutanten. Dargestellt ist die Luziferase-Aktivität bezogen auf die Transfektion von pcDNA3:RafWT und psVK3:RasG12V in % entsprechend Tabelle 6.7 im Anhang. In hellblau dargestellt sind Kotransfektionen von pcDNA3:RafWT/Mutanten und psVK3:RasG12V, in grün Kotransfektionen von pcDNA3:RafWT/Mutanten und psVK3:RasG12V,T35A und in gelb die jeweilige Transfektion von pcDNA3:RafWT/Mutanten alleine als Kontrolle. Der dunkelblaue Balken zeigt die Aktivierung nach Transfektion von psVK3:RasG12V bzw. von psVK3:RasG12V,T35A alleine.
3.10 Vergleich der Aktivierung in vivo mit den Ergebnissen aus
kinetischen und thermodynamischen Messungen
Es konnte für verschiedene Raf-Mutanten gezeigt werden, dass eine gute Korrelation besteht zwischen der Signalweiterleitung in vivo, d.h. zwischen der Luziferase-Aktivität und der Affinität (Block et al., 1996; Jaitner et al., 1997). Anhand der Vielzahl der in dieser Arbeit detailliert kinetisch und thermodynamisch charakterisierten Raf-Mutanten, sollte analysiert werden, ob die Signalweiterleitung nur abhängig ist von der Affinität, oder ob auch bestimmte kinetische oder thermodynamische Eigenschaften wichtig sind. Dazu wurden jeweils die Luziferase-Aktivitäten gegen die Affinitäten bestimmt aus Stopped-Flow und ITC, gegen die Assoziations- und Dissoziationsratenkonstanten, sowie gegen die Enthalpie- und Entropieänderungen aufgetragen.
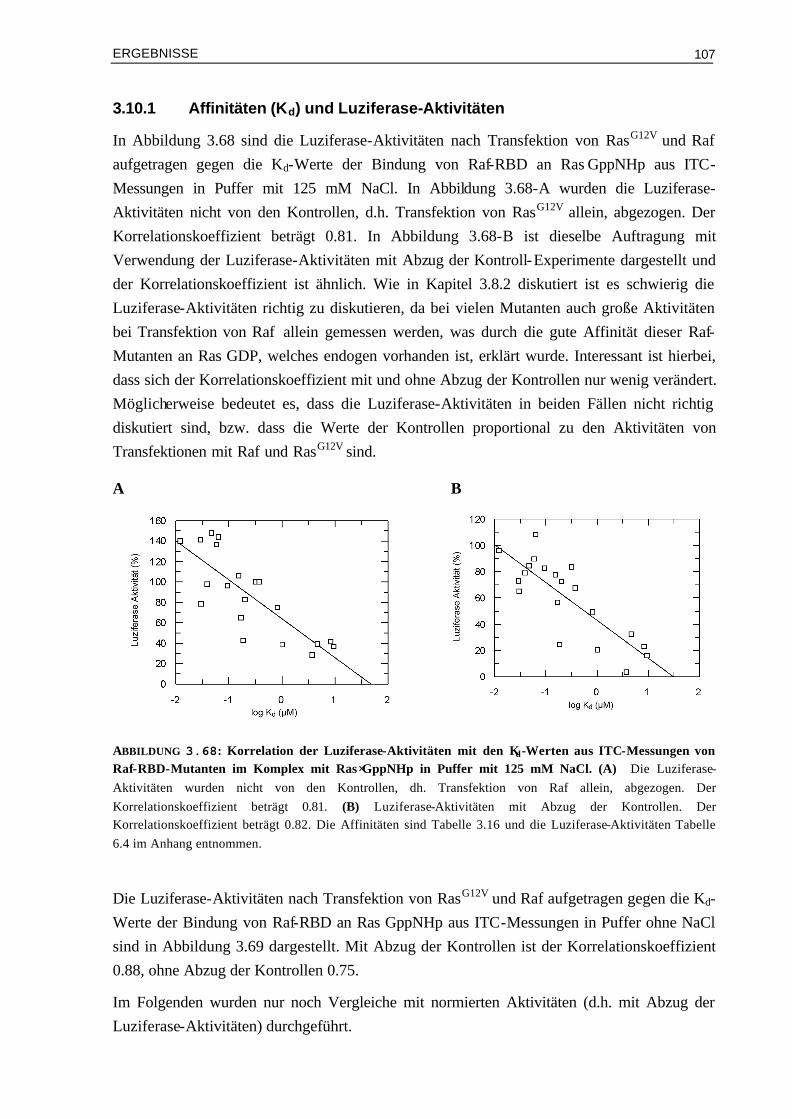
ERGEBNISSE
107
3.10.1 Affinitäten (Kd) und Luziferase-Aktivitäten
In Abbildung 3.68 sind die Luziferase-Aktivitäten nach Transfektion von RasG12V und Raf aufgetragen gegen die Kd-Werte der Bindung von Raf-RBD an Ras⋅GppNHp aus ITC-Messungen in Puffer mit 125 mM NaCl. In Abbildung 3.68-A wurden die Luziferase-Aktivitäten nicht von den Kontrollen, d.h. Transfektion von RasG12V allein, abgezogen. Der Korrelationskoeffizient beträgt 0.81. In Abbildung 3.68-B ist dieselbe Auftragung mit Verwendung der Luziferase-Aktivitäten mit Abzug der Kontroll-Experimente dargestellt und der Korrelationskoeffizient ist ähnlich. Wie in Kapitel 3.8.2 diskutiert ist es schwierig die Luziferase-Aktivitäten richtig zu diskutieren, da bei vielen Mutanten auch große Aktivitäten bei Transfektion von Raf allein gemessen werden, was durch die gute Affinität dieser Raf-Mutanten an Ras⋅GDP, welches endogen vorhanden ist, erklärt wurde. Interessant ist hierbei, dass sich der Korrelationskoeffizient mit und ohne Abzug der Kontrollen nur wenig verändert. Möglicherweise bedeutet es, dass die Luziferase-Aktivitäten in beiden Fällen nicht richtig diskutiert sind, bzw. dass die Werte der Kontrollen proportional zu den Aktivitäten von Transfektionen mit Raf und RasG12V sind. A B
ABBILDUNG 3.68: Korrelation der Luziferase-Aktivitäten mit den Kd-Werten aus ITC-Messungen von Raf-RBD-Mutanten im Komplex mit Ras ⋅GppNHp in Puffer mit 125 mM NaCl. (A) Die Luziferase-Aktivitäten wurden nicht von den Kontrollen, dh. Transfektion von Raf allein, abgezogen. Der Korrelationskoeffizient beträgt 0.81. (B) Luziferase-Aktivitäten mit Abzug der Kontrollen. Der Korrelationskoeffizient beträgt 0.82. Die Affinitäten sind Tabelle 3.16 und die Luziferase-Aktivitäten Tabelle 6.4 im Anhang entnommen.
Die Luziferase-Aktivitäten nach Transfektion von RasG12V und Raf aufgetragen gegen die Kd-Werte der Bindung von Raf-RBD an Ras⋅GppNHp aus ITC-Messungen in Puffer ohne NaCl sind in Abbildung 3.69 dargestellt. Mit Abzug der Kontrollen ist der Korrelationskoeffizient 0.88, ohne Abzug der Kontrollen 0.75.
Im Folgenden wurden nur noch Vergleiche mit normierten Aktivitäten (d.h. mit Abzug der Luziferase-Aktivitäten) durchgeführt.
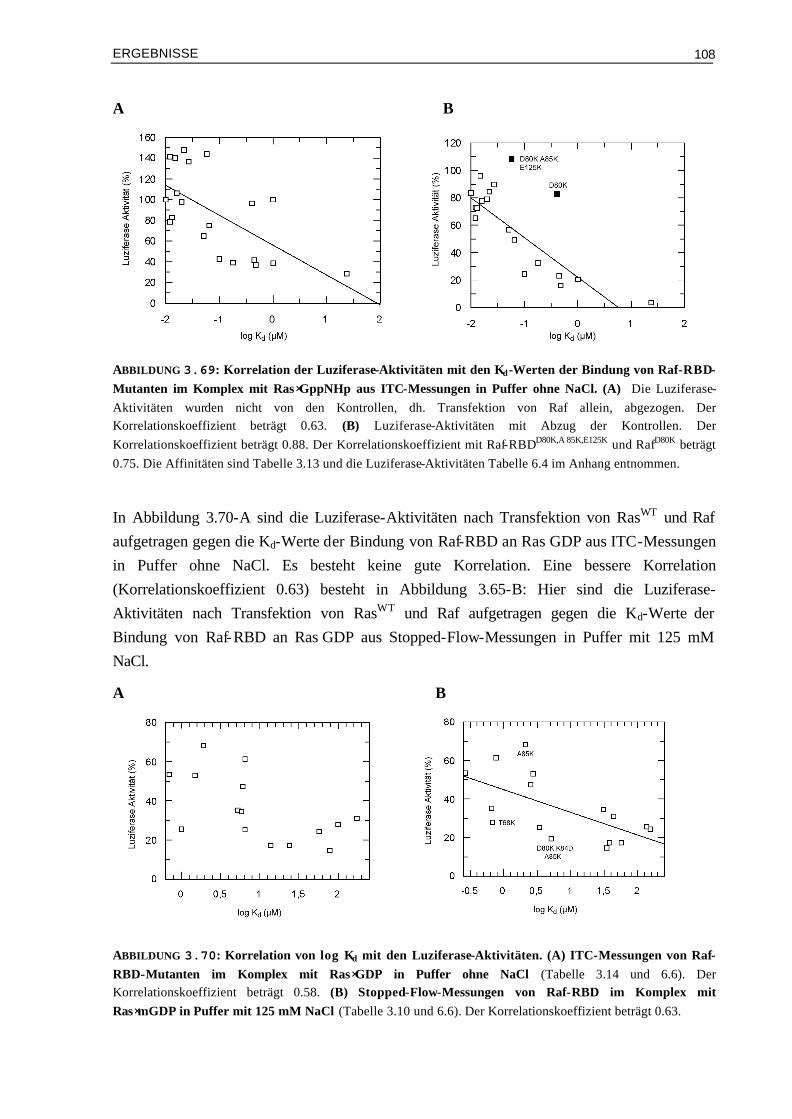
ERGEBNISSE
108
A B
ABBILDUNG 3.69: Korrelation der Luziferase-Aktivitäten mit den Kd-Werten der Bindung von Raf-RBD-Mutanten im Komplex mit Ras ⋅GppNHp aus ITC-Messungen in Puffer ohne NaCl. (A) Die Luziferase-Aktivitäten wurden nicht von den Kontrollen, dh. Transfektion von Raf allein, abgezogen. Der Korrelationskoeffizient beträgt 0.63. (B) Luziferase-Aktivitäten mit Abzug der Kontrollen. Der Korrelationskoeffizient beträgt 0.88. Der Korrelationskoeffizient mit Raf-RBDD80K,A 85K,E125K und RafD80K beträgt 0.75. Die Affinitäten sind Tabelle 3.13 und die Luziferase-Aktivitäten Tabelle 6.4 im Anhang entnommen.
In Abbildung 3.70-A sind die Luziferase-Aktivitäten nach Transfektion von RasWT und Raf aufgetragen gegen die Kd-Werte der Bindung von Raf-RBD an Ras⋅GDP aus ITC-Messungen in Puffer ohne NaCl. Es besteht keine gute Korrelation. Eine bessere Korrelation (Korrelationskoeffizient 0.63) besteht in Abbildung 3.65-B: Hier sind die Luziferase-Aktivitäten nach Transfektion von RasWT und Raf aufgetragen gegen die Kd-Werte der Bindung von Raf-RBD an Ras⋅GDP aus Stopped-Flow-Messungen in Puffer mit 125 mM NaCl.
A B
ABBILDUNG 3.70: Korrelation von log Kd mit den Luziferase-Aktivitäten. (A) ITC-Messungen von Raf-RBD-Mutanten im Komplex mit Ras⋅GDP in Puffer ohne NaCl (Tabelle 3.14 und 6.6). Der Korrelationskoeffizient beträgt 0.58. (B) Stopped-Flow-Messungen von Raf-RBD im Komplex mit Ras⋅mGDP in Puffer mit 125 mM NaCl (Tabelle 3.10 und 6.6). Der Korrelationskoeffizient beträgt 0.63.
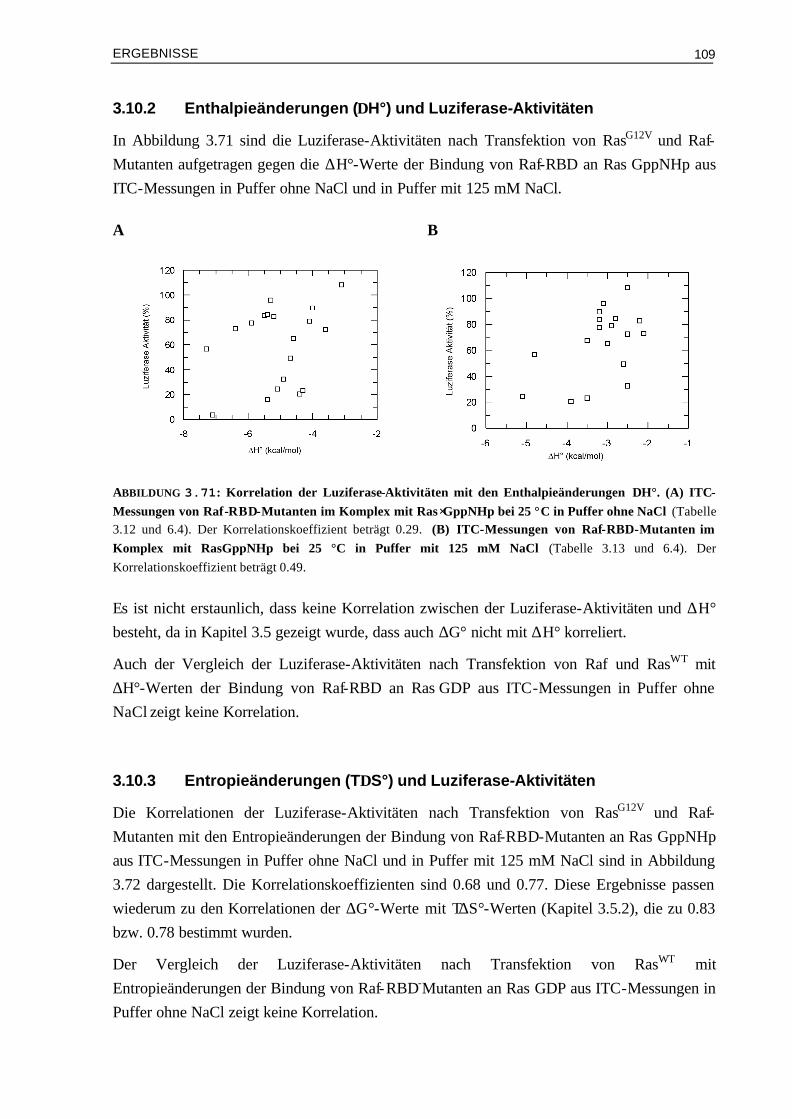
ERGEBNISSE
109
3.10.2 Enthalpieänderungen (∆H°) und Luziferase-Aktivitäten
In Abbildung 3.71 sind die Luziferase-Aktivitäten nach Transfektion von RasG12V und Raf-Mutanten aufgetragen gegen die ∆H°-Werte der Bindung von Raf-RBD an Ras⋅GppNHp aus ITC-Messungen in Puffer ohne NaCl und in Puffer mit 125 mM NaCl.
A B
ABBILDUNG 3.71: Korrelation der Luziferase-Aktivitäten mit den Enthalpieänderungen ∆H°. (A) ITC-Messungen von Raf-RBD-Mutanten im Komplex mit Ras ⋅GppNHp bei 25 °C in Puffer ohne NaCl (Tabelle 3.12 und 6.4). Der Korrelationskoeffizient beträgt 0.29. (B) ITC-Messungen von Raf-RBD-Mutanten im Komplex mit RasGppNHp bei 25 °C in Puffer mit 125 mM NaCl (Tabelle 3.13 und 6.4). Der Korrelationskoeffizient beträgt 0.49.
Es ist nicht erstaunlich, dass keine Korrelation zwischen der Luziferase-Aktivitäten und ∆H° besteht, da in Kapitel 3.5 gezeigt wurde, dass auch ∆G° nicht mit ∆H° korreliert.
Auch der Vergleich der Luziferase-Aktivitäten nach Transfektion von Raf und RasWT mit ∆H°-Werten der Bindung von Raf-RBD an Ras⋅GDP aus ITC-Messungen in Puffer ohne NaCl zeigt keine Korrelation.
3.10.3 Entropieänderungen (T∆S°) und Luziferase-Aktivitäten
Die Korrelationen der Luziferase-Aktivitäten nach Transfektion von RasG12V und Raf-Mutanten mit den Entropieänderungen der Bindung von Raf-RBD-Mutanten an Ras⋅GppNHp aus ITC-Messungen in Puffer ohne NaCl und in Puffer mit 125 mM NaCl sind in Abbildung 3.72 dargestellt. Die Korrelationskoeffizienten sind 0.68 und 0.77. Diese Ergebnisse passen wiederum zu den Korrelationen der ∆G°-Werte mit T∆S°-Werten (Kapitel 3.5.2), die zu 0.83 bzw. 0.78 bestimmt wurden.
Der Vergleich der Luziferase-Aktivitäten nach Transfektion von RasWT mit Entropieänderungen der Bindung von Raf-RBD-Mutanten an Ras⋅GDP aus ITC-Messungen in Puffer ohne NaCl zeigt keine Korrelation.
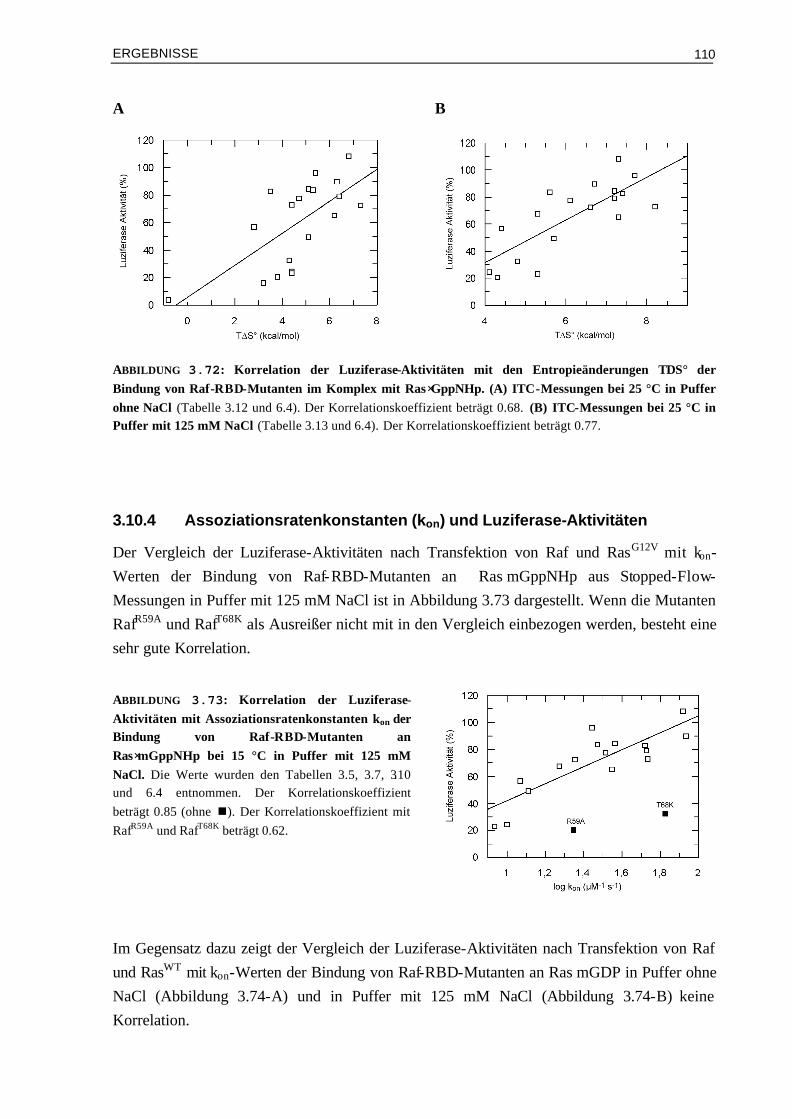
ERGEBNISSE
110
A B
ABBILDUNG 3.72: Korrelation der Luziferase-Aktivitäten mit den Entropieänderungen T∆S° der Bindung von Raf-RBD-Mutanten im Komplex mit Ras ⋅GppNHp. (A) ITC-Messungen bei 25 °C in Puffer ohne NaCl (Tabelle 3.12 und 6.4). Der Korrelationskoeffizient beträgt 0.68. (B) ITC-Messungen bei 25 °C in Puffer mit 125 mM NaCl (Tabelle 3.13 und 6.4). Der Korrelationskoeffizient beträgt 0.77.
3.10.4 Assoziationsratenkonstanten (kon) und Luziferase-Aktivitäten
Der Vergleich der Luziferase-Aktivitäten nach Transfektion von Raf und RasG12V mit kon-Werten der Bindung von Raf-RBD-Mutanten an Ras⋅mGppNHp aus Stopped-Flow-Messungen in Puffer mit 125 mM NaCl ist in Abbildung 3.73 dargestellt. Wenn die Mutanten RafR59A und RafT68K als Ausreißer nicht mit in den Vergleich einbezogen werden, besteht eine sehr gute Korrelation.
ABBILDUNG 3.73: Korrelation der Luziferase-Aktivitäten mit Assoziationsratenkonstanten kon der Bindung von Raf-RBD-Mutanten an Ras⋅mGppNHp bei 15 °C in Puffer mit 125 mM NaCl. Die Werte wurden den Tabellen 3.5, 3.7, 3.10 und 6.4 entnommen. Der Korrelationskoeffizient beträgt 0.85 (ohne n). Der Korrelationskoeffizient mit RafR59A und RafT68K beträgt 0.62.
Im Gegensatz dazu zeigt der Vergleich der Luziferase-Aktivitäten nach Transfektion von Raf und RasWT mit kon-Werten der Bindung von Raf-RBD-Mutanten an Ras⋅mGDP in Puffer ohne NaCl (Abbildung 3.74-A) und in Puffer mit 125 mM NaCl (Abbildung 3.74-B) keine Korrelation.

ERGEBNISSE
111
A B
ABBILDUNG 3.74: Korrelation der Luziferase-Aktivitäten mit log kon der Bindungen von Raf-RBD-Mutanten an Ras⋅mGDP. (A) Stopped-Flow-Messungen ohne NaCl. Der Korrelationskoeffizient beträgt 0.26. Puffer ohne NaCl. Die Werte wurden den Tabellen 3.9 und 6.6 entnommen. (B) Stopped-Flow-Messungen in Puffer mit 125 mM NaCl . Der Korrelationskoeffizient beträgt 0.41. Die Werte wurden den Tabellen 3.10 und 6.6 entnommen.
3.10.5 Dissoziationsratenkonstanten (koff) und Luziferase-Aktivitäten
In Abbildung 3.75 ist eine Korrelation der Luziferase-Aktivitäten nach Transfektion von Raf-Mutanten und RasG12V mit koff-Werten der Bindung von Raf-RBD an Ras⋅mGppNHp in Puffer mit 125 mM NaCl dargestellt. Der Korrelationskoeffizient ist mit 0.74 relativ gut.
ABBILDUNG 3.75: Korrelation der Luziferase-Aktivitäten mit den log koff-Werten aus Stopped-Flow-Messungen der Bindung von Raf-RBD an Ras⋅mGppNHp bei 15 °C in Puffer mit 125 mM NaCl. Die Werte wurden den Tabellen 3.5, 3.7 und 6.4 entnommen. Der Korrelationskoeffizient beträgt 0.74.
Ebenso ist die Korrelation der Luziferase-Aktivitäten nach Transfektion von Raf-Mutanten und RasWT mit koff-Werten der Bindung von Raf-RBD an Ras⋅mGDP sowohl in Puffer ohne NaCl als auch in Puffer mit 125 mM NaCl sehr gut (Abbildung 3.76). Hier ist somit die langsame Dissoziation, d.h. die Lebensdauer der Komplexe entscheidend für die Aktivierung.
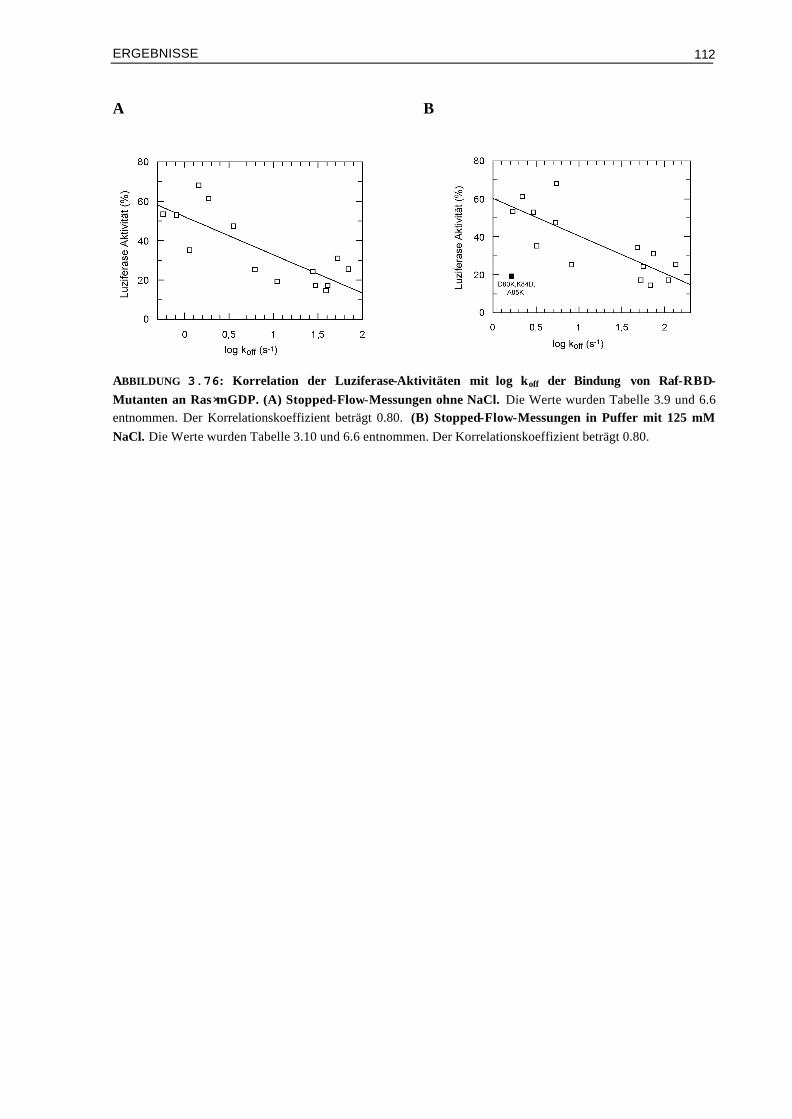
ERGEBNISSE
112
A B
ABBILDUNG 3.76: Korrelation der Luziferase-Aktivitäten mit log koff der Bindung von Raf-RBD-Mutanten an Ras ⋅mGDP. (A) Stopped-Flow-Messungen ohne NaCl. Die Werte wurden Tabelle 3.9 und 6.6 entnommen. Der Korrelationskoeffizient beträgt 0.80. (B) Stopped-Flow-Messungen in Puffer mit 125 mM NaCl. Die Werte wurden Tabelle 3.10 und 6.6 entnommen. Der Korrelationskoeffizient beträgt 0.80.

DISKUSSION
113
IV DISKUSSION 4.1 Die Oberflächeneigenschaften von Ras und Effektor-RBDs
und thermodynamische und kinetische Parameter bei der Bindung
Wie durch Röntgen- und NMR-Strukturen gezeigt worden ist, sind sowohl die Oberflächen der Ras Proteine als auch die der Effektor-RBDs sehr polar, d.h. die Seitenketten von polaren und geladenen Aminosäuren befinden sich an der Oberfläche und sind von Wasser solvatisiert. Auch die Kontaktflächen, über die die Bindung in Komplexen aus Ras Proteinen und RBDs erfolgt, bestehen zum größten Teil aus Aminosäuren mit polaren und geladenen Seitenketten. Dies scheint eine allgemeine Eigenschaft von Proteinkontaktflächen zu sein, wie durch die Analyse einer großen Anzahl von Strukturen von Proteinen im gebundenen und ungebundenen Zustand gezeigt wurde (Übersicht: Jones & Thornton, 1996; Stites, 1997; Conte et al., 1999): Es wird eine signifikant größere Anzahl von geladenen und polaren Resten in Protein-Kontaktflächen als im Innern der Proteine gefunden; diese sind für die thermodynamischen und kinetischen Eigenschaften von Protein-Protein-Wechselwirkungen verantwortlich und tragen zur Spezifität und Affinität von Protein Komplexen bei.
4.1.1 Die thermodynamischen Parameter der Bindung von Effektor-RBDs
an Ras Proteine geben Hinweis auf die Art der Bindung.
Nach der Gibbs-Helmholtz Gleichung (∆G° = ∆H° - T∆S°) ist die Entscheidung, ob zwei Proteine aneinander binden, eine Folge der energetischen Beiträge von Änderungen der Enthalpie (∆H°) und der Entropie (T∆S°) der Komplexbildung. Entropisch gesehen ist der Verlust der Bewegungsfreiheit der Proteine und die Einschränkung der Beweglichkeit der Seitenketten der Aminosäuren bei der Assoziation ungünstig. Günstig auf die Entropie dagegen wirken sich die Freisetzung von Wassermolekülen der Solvatation von Aminosäureresten aus. Je nach dem, welcher Effekt überwiegt, kann die Entropie positiv oder negativ sein (Dunitz, 1994). So ist bei der Bindung von Raf-RBD an Ras Proteine (Rudolph et al., 2001; diese Arbeit; Abbildung 4.1) die Entropieänderung positiv, d.h. der Verlust der Bewegungsfreiheit der Proteine wird durch einen Gewinn durch die Freisetzung von Wassermolekülen überkompensiert. Dieses ist nicht der Fall bei der Bindung von RalGDS-RBD an verschiedene Ras Proteine (Rudolph et al., 2001; diese Arbeit); hier ist die Entropieänderung negativ, was möglicherweise dadurch erklärt werden kann, dass nicht so viele Wassermoleküle freigesetzt werden, sondern z. T. in intermolekulare H-Brücken mit einbezogen oder weniger Seitenketten desolvatisiert werden. Die Rolle von Wassermolekülen und deren Analyse hat daher eine große Bedeutung – wenn nicht sogar die entscheidende
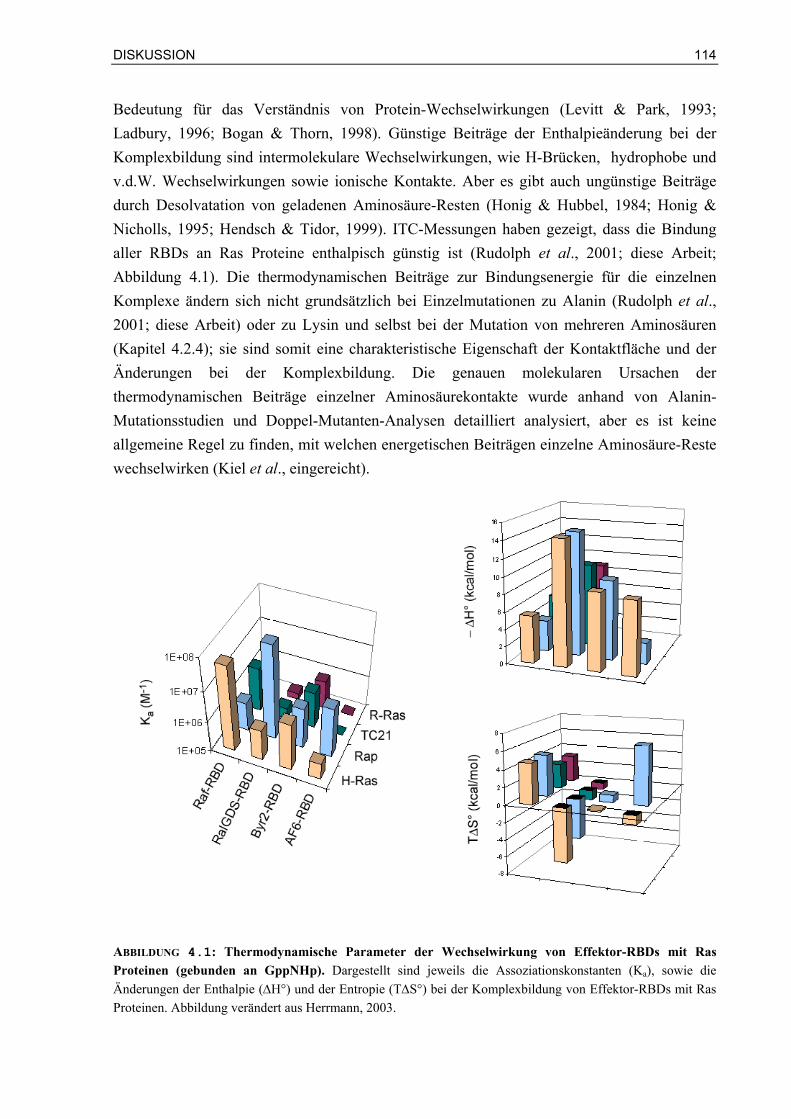
DISKUSSION
114
Bedeutung für das Verständnis von Protein-Wechselwirkungen (Levitt & Park, 1993; Ladbury, 1996; Bogan & Thorn, 1998). Günstige Beiträge der Enthalpieänderung bei der Komplexbildung sind intermolekulare Wechselwirkungen, wie H-Brücken, hydrophobe und v.d.W. Wechselwirkungen sowie ionische Kontakte. Aber es gibt auch ungünstige Beiträge durch Desolvatation von geladenen Aminosäure-Resten (Honig & Hubbel, 1984; Honig & Nicholls, 1995; Hendsch & Tidor, 1999). ITC-Messungen haben gezeigt, dass die Bindung aller RBDs an Ras Proteine enthalpisch günstig ist (Rudolph et al., 2001; diese Arbeit; Abbildung 4.1). Die thermodynamischen Beiträge zur Bindungsenergie für die einzelnen Komplexe ändern sich nicht grundsätzlich bei Einzelmutationen zu Alanin (Rudolph et al., 2001; diese Arbeit) oder zu Lysin und selbst bei der Mutation von mehreren Aminosäuren (Kapitel 4.2.4); sie sind somit eine charakteristische Eigenschaft der Kontaktfläche und der Änderungen bei der Komplexbildung. Die genauen molekularen Ursachen der thermodynamischen Beiträge einzelner Aminosäurekontakte wurde anhand von Alanin-Mutationsstudien und Doppel-Mutanten-Analysen detailliert analysiert, aber es ist keine allgemeine Regel zu finden, mit welchen energetischen Beiträgen einzelne Aminosäure-Reste wechselwirken (Kiel et al., eingereicht).
ABBILDUNG 4.1: Thermodynamische Parameter der Wechselwirkung von Effektor-RBDs mit Ras Proteinen (gebunden an GppNHp). Dargestellt sind jeweils die Assoziationskonstanten (Ka), sowie die Änderungen der Enthalpie (∆H°) und der Entropie (T∆S°) bei der Komplexbildung von Effektor-RBDs mit Ras Proteinen. Abbildung verändert aus Herrmann, 2003.

DISKUSSION
115
4.1.2 Komplementäre elektrostatische Ladungen in der Kontaktfläche von
Ras/Effektor-RBD-Komplexen beschleunigen die Assoziation
Neben den polaren und ionischen Eigenschaften der Oberfläche von Ras und Effektor Proteinen sind die Kontaktflächen entgegengesetzt, also elektrostatisch komplementär geladen (Abbildung 4.2). Aufgrund der hohen elektrostatischen Komplementarität sind die Werte für kon in allen Ras-Effektor-Komplexen sehr hoch (Abbildung 4.3). Die Änderungen der Affinität verschiedener RBD/Ras Protein-Komplexe sind dabei im Wesentlichen die Folge von veränderten kon-Werten und die Dissoziation ist in allen Komplexen ähnlich. Insbesondere die Assoziationsratenkonstanten von RalGDS-RBD und Raf-RBD unterscheiden sich deutlich, was anhand der Elektrostatik der Kontaktflächen erklärt werden kann (Abbildung 4.2). Während die Oberflächen von Ras und Rap im Effektor Bindungsbereich überwiegend negative Ladungen enthalten, sind die Oberflächen der RBDs von RalGDS und Raf hauptsächlich positiv geladen. Insbesondere zwischen der Kontaktfläche von Raf-RBD und Ras besteht eine sehr starke elektrostatische Komplementarität, was den sehr hohen Wert für kon als Folge von elektrostatischer Lenkung erklären kann. Im Gegensatz dazu ist die Kontaktfläche zwischen RalGDS-RBD und Ras nicht so stark elektrostatisch komplementär geladen, was eine Erklärung für den niedrigeren kon-Wert ist.
Die Dissoziationsratenkonstanten liegen alle in einer ähnlichen Größenordnung und sind relativ groß. Die Bindungen von Effektor-RBDs an Ras Proteine gebunden an mGppNHp sind somit gekennzeichnet durch sowohl schnelle Assoziations- als auch schnelle Dissoziationsreaktionen; die Komplexe sind somit sehr dynamisch, was eine Bedeutung für die Bindung von regulativen Proteinen wie z. B. GAPs haben könnte: Effektoren und GAPs binden an teilweise überlappende Regionen von Ras (Vetter & Wittinghofer, 2001).
A B
ABBILDUNG 4.2: Oberflächenpotentiale zweier Ras/Effektor-RBD-Komplexe. Dargestellt sind jeweils die Oberflächenpotentiale der aufgeklappten Komplexe. In blau dargestellt sind positive und in rot negative Ladungen. (A) Komplex zwischen Raf-RBD und Rap⋅GppNHp. (B) Komplex zwischen RalGDS-RBD und Ras⋅GppNHp.

DISKUSSION
116
ABBILDUNG 4.3: Kinetische Parameter der Wechselwirkung von Effektor-RBDs mit Ras Proteinen. Dargestellt sind jeweils die Assoziationsratenkonstanten (kon), die Dissoziationsratenkonstanten (koff) und die Dissoziationskonstanten (Kd) bei der Komplexbildung von Effektor-RBDs mit Ras und Rap gebunden an mGppNHp. Es sind die absoluten Werte auf einer logarithmischen Skala mit den Einheiten µM-1s-1 (kon), s-1 (koff) und µM (Kd) angegeben.
Auffallend anders ist dagegen der Wert für koff von RalGDS-RBD im Komplex mit Rap⋅mGppNHp: Hier ist die sehr langsame Dissoziation die Ursache für die sehr hohe Affinität, während die Assoziationsratenkonstante ähnlich wie bei der Bindung an Ras⋅mGppNHp ist. Möglicherweise gibt diese extrem langsame Dissoziation einen weiteren Hinweis auf die vielfach diskutierte inhibitorische Funktion von Rap. So konnte in vivo gezeigt werden, dass Ras-Effektoren an Rap Proteine binden können, aber nicht zu deren Aktivierung führen (Boussiotis et al., 1997). Diese inhibitorische Wirkung wird allerdings kontrovers diskutiert, denn in anderen in vivo Studien wurde eine gleichzeitige Aktivierung von Ras und Rap gefunden, ohne die Effekte von Ras zu beeinflussen (Zwartkruis et al., 1998).
4.1.3 Es werden unterschiedliche thermodynamische und kinetische Hot-Spots in Ras/Effektor-RBD-Komplexen gefunden
Die Wechselwirkung der Bindung verschiedener Effektor-RBDs an Ras Proteine ist sehr spezifisch: Obwohl die RBDs alle denselben Faltungstyp der Ubiquitinfaltung aufweisen (Abbildung 4.4) und die Ras Proteine – insbesondere in der Effektor-Bindungs Region – sehr homolog sind, binden Ras und Effektor-RBD Proteine mit teilweise sehr unterschiedlichen Affinitäten aneinander (Kapitel 4.1.1). Diese spezifische Erkennung eines Partnerproteins von nur einem Mitglied aus einer Protein-Familie, obwohl alle Mitglieder denselben Faltungstyp

DISKUSSION
117
aufweisen ist eine zentrale Eigenschaft in vielen biologischen Prozessen (Übersicht: Ponting & Russel, 2002). Die spezifischen Unterschiede müssen daher auf der Protein Oberfläche als verschiedene Determinanten der Bindungsenergie kodiert sein. Die Kenntnis einer bestimmten Faltung bedeutet somit nicht, dass man die Funktion eines Proteins kennt (Orengo et al., 1994). Nach der vollständigen Sequenzierung des menschlichen Genoms, ist ein neues großes Ziel zu verstehen, in welcher Beziehung Aminosäuresequenz, Proteinstruktur und Funktion zueinander stehen. Bisher wurden einige theoretische Ansätze entwickelt, um funktionell wichtige Regionen und Bindungsstellen auf Protein Oberflächen vorherzusagen (Casari et al., 1995; Aloy et al., 2001; Landgraf et al., 2001); auch hier wird noch einmal deutlich, wie wichtig es wäre, mehr über die energetischen Beiträge in Protein Komplexen zu verstehen.
ABBILDUNG 4.4: Ubiquitinfaltung der RBDs dreier Effektoren. Überlagerung der RBDs von Raf (grün), RalGDS (rot) und PI3K (blau). Abbildung aus Herrmann 2003.
Eine vielfach verwendete experimentelle Methode zur Ermittlung der Beiträge einzelner Aminosäure-Reste in der Kontaktfläche ist die Alanin-Mutanten Analyse (Übersicht: Bogan & Thorn, 1998). Des Weiteren werden Daten aus diesen Messungen häufig verwendet, um theoretische Berechnungen mit experimentellen Ergebnissen vergleichen zu können, wobei die Interpretation von Daten aus Mutanten-Analysen hinsichtlich struktureller Veränderungen problematisch sein können (DeLano, 2002).
In Abbildung 4.5 sind die Änderungen der freien Enthalpieänderung der Bindung zwischen Wildtyp und Mutanten (∆∆G° = ∆G° WT- ∆G° Mut) als farbliche Abstufungen in den RalGDS-/ und Raf-RBD/Ras⋅GppNHp-Komplex-Strukturen dargestellt (Ergebnisse aus Kiel et al., eingereicht und dieser Arbeit). Negative Werte für ∆∆G° (rot in der Struktur) zeigen, dass die Affinität bei Mutation zu Alanin schlechter wird, postive Werte (blau) bedeuten, dass die Affinität der Alanin Mutante besser ist als beim Wildtyp-Protein Komplex. Diese Energieverteilungen basierend auf Alanin-Mutanten-Analysen unterscheiden sich somit deutlich für Raf- und RalGDS-RBD im Komplex mit Ras⋅GppNHp (Abbildung 4.5). Im Raf-

DISKUSSION
118
RBD/Ras⋅GppNHp-Komplex ist Arg89 von Raf ein starker Hot-Spot der Bindungsenergie; die Affinität der Mutante von Arg89 zu Alanin konnte nicht im Komplex mit Ras⋅GppNHp gemessen werden, da diese zu klein ist und daher ist der Wert für ∆∆G° ein Mindestwert. Arginin wurde häufig als sehr wichtige Aminosäure in Protein-Komplexen gefunden (Bogan & Thorn, 1998). In der Raf-RBD/RapsGppNHp Struktur sieht man, dass Arg89 direkt in die Mitte der Kontaktfläche zeigt. Möglicherweise handelt es sich hier um eine spezielle Form der Komplexbildung, als O-Ring Modell bezeichnet (Bogan & Thorn, 1998), wobei ein Aminosäurerest, der einen entscheidenden Beitrag zur Bindungsenergie liefert, sich in einer von Wasser abgeschirmten Umgebung befinden muss, wozu andere Aminosäuren in der Kontaktfläche, die keinen so großen Einfluss auf die Bindungsenergie haben, eine Art Ring um diesen Rest bilden.
ABBILDUNG 4.5: Energieverteilung in den Komplexen von Ras⋅GppNHp mit Raf-RBD (links) und RalGDS-RBD (rechts) . Die Änderungen der freien Enthalpie bei Mutation zu Alanin im Vergleich zu Wildtyp (∆∆G°) wurden in den Komplex-Strukturen (als farbliche Änderungen sichtbar gemacht. Abbildung aus Kiel et al., eingereicht.
Hot-Spots in Protein Wechselwirkungen sind somit definiert als Aminosäure-Reste, die nach Mutation zu einer großen Verringerung in der Bindungsenergie (Affinität) führen. Kinetische Messungen zeigen, dass diese Hot-Spots meistens eine Folge von größeren Dissoziationsratenkonstanten sind, aber es gibt auch Fälle bei denen kleinere Assoziationsratenkonstanten beobachtet werden (Schreiber, 2002).
Die kinetischen Beiträge die zu den in Abbildung 4.5 dargestellten „thermodynamischen“ Hot-Spots in den Raf-/ und RalGDS-RBD/Ras⋅mGppNHp Komplexen führen, sind in Tabelle 4.1 dargestellt. Im Komplex von RalGDS-RBD mit Ras⋅mGppNHp zeigen die Ergebnisse aus Stopped-Flow-Messungen, dass bei der Mutation von positiv geladenen Aminosäuren zu Alanin (R16A, K28A, K44A und K48A) in allen Fällen die Assoziationsratenkonstante
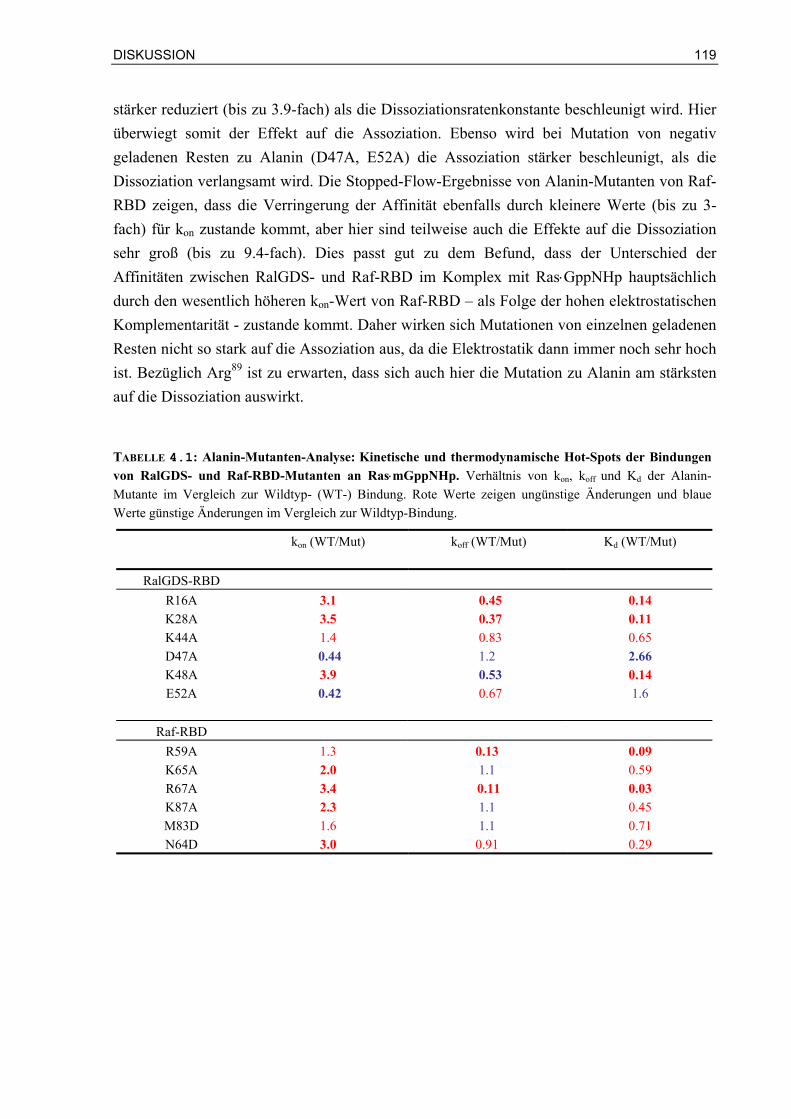
DISKUSSION
119
stärker reduziert (bis zu 3.9-fach) als die Dissoziationsratenkonstante beschleunigt wird. Hier überwiegt somit der Effekt auf die Assoziation. Ebenso wird bei Mutation von negativ geladenen Resten zu Alanin (D47A, E52A) die Assoziation stärker beschleunigt, als die Dissoziation verlangsamt wird. Die Stopped-Flow-Ergebnisse von Alanin-Mutanten von Raf-RBD zeigen, dass die Verringerung der Affinität ebenfalls durch kleinere Werte (bis zu 3-fach) für kon zustande kommt, aber hier sind teilweise auch die Effekte auf die Dissoziation sehr groß (bis zu 9.4-fach). Dies passt gut zu dem Befund, dass der Unterschied der Affinitäten zwischen RalGDS- und Raf-RBD im Komplex mit Ras⋅GppNHp hauptsächlich durch den wesentlich höheren kon-Wert von Raf-RBD – als Folge der hohen elektrostatischen Komplementarität - zustande kommt. Daher wirken sich Mutationen von einzelnen geladenen Resten nicht so stark auf die Assoziation aus, da die Elektrostatik dann immer noch sehr hoch ist. Bezüglich Arg89 ist zu erwarten, dass sich auch hier die Mutation zu Alanin am stärksten auf die Dissoziation auswirkt.
TABELLE 4.1: Alanin-Mutanten-Analyse: Kinetische und thermodynamische Hot-Spots der Bindungen von RalGDS- und Raf-RBD-Mutanten an Ras⋅mGppNHp. Verhältnis von kon, koff und Kd der Alanin-Mutante im Vergleich zur Wildtyp- (WT-) Bindung. Rote Werte zeigen ungünstige Änderungen und blaue Werte günstige Änderungen im Vergleich zur Wildtyp-Bindung.
kon (WT/Mut)
koff (WT/Mut)
Kd (WT/Mut)
RalGDS-RBD R16A 3.1 0.45 0.14 K28A 3.5 0.37 0.11 K44A 1.4 0.83 0.65 D47A 0.44 1.2 2.66 K48A 3.9 0.53 0.14 E52A 0.42 0.67 1.6
Raf-RBD
R59A 1.3 0.13 0.09 K65A 2.0 1.1 0.59 R67A 3.4 0.11 0.03 K87A 2.3 1.1 0.45 M83D 1.6 1.1 0.71 N64D 3.0 0.91 0.29

DISKUSSION
120
4.1.4 Ergebnisse aus Doppel-Mutanten-Analysen können die
Bindungsspezifitäten der Bindung von RalGDS- und Raf-RBD an Ras und Rap quantifizieren
Die Abbildung 4.6 zeigt die zwei Komplexe mit hoher Affinität, RalGDS-RBD/Rap⋅mGppNHp (-11.4 kcal/mol) und Raf-RBD/Ras⋅mGppNHp (-11.1 kcal/mol) sowie die beiden Komplexe mit niedrigeren Affinität, RalGDS-RBD/Ras⋅mGppNHp (-8.6 kcal/mol) und Raf-RBD/Rap⋅mGppNHp (-8.5 kcal/mol) (∆G°-Werte aus Kiel & Herrmann, 2001). Es besteht somit eine Kreuz-Spezifität der Bindung von RalGDS- und Raf-RBD an Ras und Rap, d.h. RalGDS-RBD bindet mit etwa 100-fach höherer Affinität an Rap als an Ras während Raf-RBD ca. 60-fach besser an Ras als an Rap bindet. Die Ergebnisse von Doppel-Mutanten-Analysen für die Raf-/und Ral-GDS–RBD/RasGppNHp Komplexe können verwendet werden, um die gemessenen Kreuz-Bindungsspezifitäten von Raf- und RalGDS-RBD an Ras bzw. Rap auf molekularer Ebene zu erklären.
Die Doppel-Mutanten-Analyse ist eine Methode zur Bestimmung der energetischen Beiträge einzelner Aminosäure-Wechelwirkungen in der Kontaktfläche (Schreiber & Fersht, 1995). Dabei wird jeweils die Affinität zwischen zwei Wildtyp-Proteinen, jeweils einer Alanin-Mutante mit dem anderen Wildtyp-Protein und zwischen beiden Alanin-Mutanten verglichen. Dies liefert die interne Kopplungsenergie zwischen einzelnen Aminosäure-Paaren nach folgendem Schema:
∆∆G°int = ∆G°X,Y - ∆G°X→A,Y - ∆G°X,Y→A + ∆G°X→A,Y→A
Energetisch günstige Kontakte zeigen dabei negative Werte für ∆∆G°int, abstoßende Kontakte zeigen positive Werte und für Aminosäuren, die nicht wechselwirken ist die Kopplungsenergie Null.
Die Ergebnisse von Doppel-Mutanten-Analysen zeigen, dass Lys84 von Raf-RBD energetisch günstige Wechselwirkungen mit den Aminosäuren Glu31 (-1.0 kcal/mol) und Asp33 (-0.8 kcal/mol) von Ras macht (Abbildung 4.6; Kiel & Herrmann, 2001). Die Summe dieser beiden Wechselwirkungen trägt mehr als die Hälfte der Differenz der freien Enthalpie der Bindung zwischen Raf-RBD im Komplex mit Ras und Rap bei (-2.6 kcal/mol). Der Rest der Differenz könnte durch abstoßende Wechselwirkungen zwischen Lys84 von Raf-RBD und Lys31 in dem Raf-RBD/Rap-Komplex erklärt werden. Wie in der Raf-RBD/Raps-Struktur sichtbar ist, kann Lys84 Salzbrücken zu Glu31 und Asp33 ausbilden (Nassar et al., 1996), die zu ähnlichen energetischen Kontakten führen wie hier durch Doppel-Mutanten-Zyklen im Raf-RBD/Ras-Komplex gezeigt wurde. Die Komplexe von RalGDS-RBD mit Ras und Rap zeigen eine Differenz der Änderungen der freien Enthalpie von 2.8 kcal/mol. In diesem System ergeben die Ergebnisse aus Doppel-Mutanten-Analysen abstoßende Wechselwirkungen zwischen

DISKUSSION
121
Asp47 von RalGDS-RBD und Glu31 von Ras (1.3 kcal/mol), was ungefähr der Hälfte dieser Differenz entspricht. Hier könnte eine günstige Wechselwirkung zwischen Lys31 von Rap und Asp47 von RalGDS-RBD den Rest der Differenz erklären.
Anhand der Ergebnisse lassen sich somit die energetischen Beiträge einzelner Aminosäure-Kontakte quantifizieren, aber es ist nicht möglich anhand der Komplexstrukturen diese Beiträge vorherzusagen.
D47 E52E53
D47 E52 E53
RalGDS-RBD
RalGDS-RBD
Raf-RBD
Raf-RBD
H-Ras H-Ras
Rap Rap
E31
K84
E31 D33D33
K31
K84
D33D33K31
G° = - 2.6 kcal/molG° = 2.8 kcal/mol
G° = - 8.5 kcal/molG° = - 11.4 kcal/mol
G° = - 8.6 kcal/mol G° = - 11.1 kcal/mol∆ ∆
∆
∆
∆
∆
∆ ∆
ABBILDUNG 4.6: Schematische Darstellung der molekularen Ursachen der Bindungsspezifitäten von RalGDS- und Raf-RBD an Ras und Rap basierend auf Doppel-Mutanten-Analysen. Abbildung aus Kiel & Herrmann, 2001.
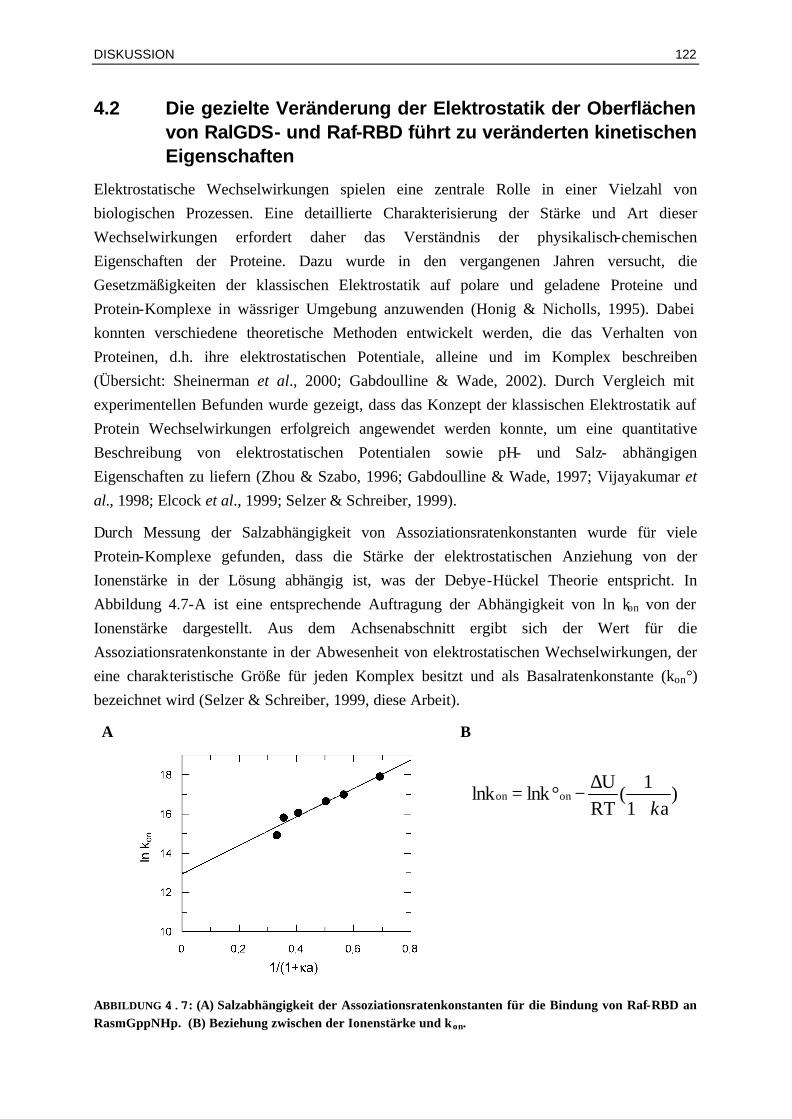
DISKUSSION
122
)a1
1(
RTU
lnklnk ononκ+
∆−°=
4.2 Die gezielte Veränderung der Elektrostatik der Oberflächen von RalGDS- und Raf-RBD führt zu veränderten kinetischen Eigenschaften
Elektrostatische Wechselwirkungen spielen eine zentrale Rolle in einer Vielzahl von biologischen Prozessen. Eine detaillierte Charakterisierung der Stärke und Art dieser Wechselwirkungen erfordert daher das Verständnis der physikalisch-chemischen Eigenschaften der Proteine. Dazu wurde in den vergangenen Jahren versucht, die Gesetzmäßigkeiten der klassischen Elektrostatik auf polare und geladene Proteine und Protein-Komplexe in wässriger Umgebung anzuwenden (Honig & Nicholls, 1995). Dabei konnten verschiedene theoretische Methoden entwickelt werden, die das Verhalten von Proteinen, d.h. ihre elektrostatischen Potentiale, alleine und im Komplex beschreiben (Übersicht: Sheinerman et al., 2000; Gabdoulline & Wade, 2002). Durch Vergleich mit experimentellen Befunden wurde gezeigt, dass das Konzept der klassischen Elektrostatik auf Protein Wechselwirkungen erfolgreich angewendet werden konnte, um eine quantitative Beschreibung von elektrostatischen Potentialen sowie pH- und Salz- abhängigen Eigenschaften zu liefern (Zhou & Szabo, 1996; Gabdoulline & Wade, 1997; Vijayakumar et al., 1998; Elcock et al., 1999; Selzer & Schreiber, 1999).
Durch Messung der Salzabhängigkeit von Assoziationsratenkonstanten wurde für viele Protein-Komplexe gefunden, dass die Stärke der elektrostatischen Anziehung von der Ionenstärke in der Lösung abhängig ist, was der Debye-Hückel Theorie entspricht. In Abbildung 4.7-A ist eine entsprechende Auftragung der Abhängigkeit von ln kon von der Ionenstärke dargestellt. Aus dem Achsenabschnitt ergibt sich der Wert für die Assoziationsratenkonstante in der Abwesenheit von elektrostatischen Wechselwirkungen, der eine charakteristische Größe für jeden Komplex besitzt und als Basalratenkonstante (kon°) bezeichnet wird (Selzer & Schreiber, 1999, diese Arbeit).
A B
ABBILDUNG 4.7: (A) Salzabhängigkeit der Assoziationsratenkonstanten für die Bindung von Raf-RBD an RasmGppNHp. (B) Beziehung zwischen der Ionenstärke und kon.

DISKUSSION
123
Somit ist kon die Summe aus zwei Beiträgen: der Basalratenkonstante, in Abwesenheit von elektrostatischen Kräften, und den elektrostatischen Wechselwirkungen zwischen den Proteinen. Basierend auf den elektrostatischen Beiträgen wurde die in dieser Arbeit angewendete Methode zur gezielten Veränderung von Assoziationsratenkonstanten entwickelt (Selzer et al., 2000).
4.2.1 Die kinetischen Parameter werden in Komplexen von RalGDS- und Raf-RBD unterschiedlich beeinflusst
Durch gezielte Veränderung der elektrostatischen Komplementarität in der Kontaktfläche von RalGDS- und Raf-RBD wurden Komplexe mit veränderten elektrostatischen Eigenschaften erhalten. So führte die Einführung von positiv geladenen Aminosäure-Resten am Rand der Kontaktfläche von RalGDS-RBD zu einer schnelleren Assoziationsratenkonstante im Komplex mit Ras⋅mGppNHp. Da die Dissoziationsratenkonstanten in den meisten Fällen nicht verändert sind, korrelieren höhere Werte für kon mit einer besseren Affinität.
Der größte Effekt wird für die Mutante RalGDS-RBDM26K,D47K,E54K erhalten: die Affinität wird von Kd = 1.9 µM (Wildtyp) auf Kd = 0.071 µM (27-fach) gesteigert. Dabei wird kon 14.3-fach schneller, aber auch koff wird 2-fach langsamer und trägt somit einen kleinen Teil zur besseren Affinität bei. Der Grund hierfür liegt darin, dass die Aminosäure Asp47 von RalGDS-RBD in der Kontaktfläche liegt und abstoßende Wechselwirkungen zu Glu31 von Ras macht (Kiel et al., eingereicht). Bei Mutation von Asp47 zu Lysin kann sich somit wahrscheinlich eine zusätzliche Salzbrücke ausbilden, die die verlangsamte Dissoziation erklären könnte. Den größten Effekt auf den Kd-Wert für eine Einzel-Mutante hat RalGDS-RBDD90K (7-fach besser), wobei die Assoziationsratenkonstante 5.5-fach größer wird und die Dissoziation sich 1.9-fach verlangsamt. Im Gegensatz dazu ergab die Mutation von Aminosäuren am Rand und in der Kontaktfläche von Raf-RBD zu positiv geladenen Resten keinen so großen Effekt auf die Assoziationsratenkonstante im Komplex mit Ras⋅mGppNHp. Der größte Effekt wird für die dreifach Mutante Raf-RBDD80K,A85K,E125K erhalten. Hier ist der Wert für kon 2.8-fach höher im Vergleich zu Wildtyp. Die möglichen Gründe, dass nicht so hohe Beschleunigungen wie im RalGDS-RBD/Ras⋅mGppNHp-System erhalten wurden, sind zum einen, dass die elektrostatische Komplementarität in der Kontaktfläche von Raf-RBDWT und Ras⋅GppNHp bereits sehr hoch ist und, dass die Assoziation sehr schnell und schon nahe an der Grenze einer diffusions-kontrollierten Assoziation ist, wodurch eine weitere Beschleunigung durch zusätzliche günstige elektrostatische Wechselwirkungen fast nicht mehr möglich ist. Zum anderen erfolgten die Messungen der Raf-RBD-Mutanten/Ras⋅GppNHp-Komplexe in Puffer mit 125 mM NaCl. Diese hohe Ionenstärke führt aufgrund der Abschwächung des elektrostatischen Effekts, zu einer deutlichen Reduzierung der Assoziationsratenkonstanten und demzufolge können auch nicht so hohe Beschleunigungen erreicht werden. Dennoch ist die Affinität dieser Mutante und aller

DISKUSSION
124
weiteren Mutanten mit A85K sehr hoch. Die Einzelmutante von Raf-RBD, die im Komplex mit Ras⋅mGppNHp den größten Effekt zeigt ist A85K. Die Affinität wird hier 7.4-fach besser im Vergleich zu Raf-RBDWT , aber hier ist dies hauptsächlich die Folge einer (4.1-fach) verlangsamten Dissoziation, während der Wert für kon nur 1.8-fach größer wird. Noch drastischere Effekte auf die Dissoziationsratenkonstanten werden bei Raf-RBD-Mutanten im Komplex mit Ras⋅mGDP gefunden: hier führt die Einzelmutante Raf-RBDA85K im Komplex mit Ras⋅mGDP zu einem 16.5-fach besseren Kd-Wert, wobei die Dissoziation hier sehr stark verlangsamt ist (12.4-fach) aber der Wert für kon nur 1.3-fach verbessert wird. Es werden somit in allen drei untersuchten Systemen höhere Affinitäten gefunden, aber nur für die Bindungen von RalGDS-RBD-Mutanten an Ras⋅mGppNHp ist dies hauptsächlich die Folge von schnelleren Assoziationsratenkonstanten. Für die Bindung von Raf-RBD-Mutanten an Ras⋅mGppNHp und Ras⋅mGDP ist die höhere Affinität eine Folge einer wesentlich verlangsamten Dissoziation.
Die verbesserten Affinitäten der RalGDS-Mutanten wurden durch ITC-Messungen im Komplex mit Ras⋅GppNHp als unabhängige Methode bestätigt, wobei insgesamt eine sehr gute Übereinstimmung zwischen den Affinitäten ermittelt aus Stopped-Flow und ITC besteht. Im Gegensatz dazu bestätigen ITC-Messungen von Raf-RBD Mutanten im Komplex mit Ras⋅GppNHp und Ras⋅GDP nicht in allen Fällen die Ergebnisse aus Stopped-Flow-Messungen. Sehr deutliche Abweichungen bestehen bei den Messungen von Raf-RBD-Mutanten an Ras⋅GDP, insbesondere bei allen Mutanten, die die Mutation Ala85 zu Lys enthalten. Hier sind die Ratenkonstanten der Dissoziation extrem klein, sogar kleiner als in dem hoch affinen Raf-RBD/Ras⋅mGppNHp-Komplex und führen zusammen mit den Assoziationsratenkonstanten zu sehr hohen Affinitäten. Möglicherweise werden hier Teilschritte der Reaktion mit einer der beiden Methoden nicht erfasst.
4.2.2 Die Korrelation von experimentellen und berechneten Werten für kon ist für Komplexe mit RalGDS-RBD-Mutanten am besten
Von den untersuchten Systemen ist die Übereinstimmung zwischen experimentellen und berechneten Assoziationsratenkonstanten für die Bindungen von RalGDS-RBD-Mutanten an Ras⋅mGppNHp und Rap⋅mGppNHp am besten (Abbildung 4.8). Der Korrelationskoeffizient ist mit 0.87 sehr gut, wichtiger aber ist die Steigung (0.69), die anzeigt, wie gut die absoluten experimentellen und berechneten Werte von kon übereinstimmen. Entscheidend anders sieht der Vergleich von kon (exp.) und kon (ber.) für die Bindungen von Raf-RBD-Mutanten an Ras⋅mGppNHp (Abbildung 4.8) und Ras⋅mGDP aus: Während auch hier die Korrelations-koeffizienten im Bereich von 0.7 (Ras⋅mGppNHp) und 0.9 (Ras⋅mGDP) liegen, weichen die Steigungen deutlich von 1 ab (0.2 bis 0.4). Die Ursachen liegen darin, dass im Gegensatz zum
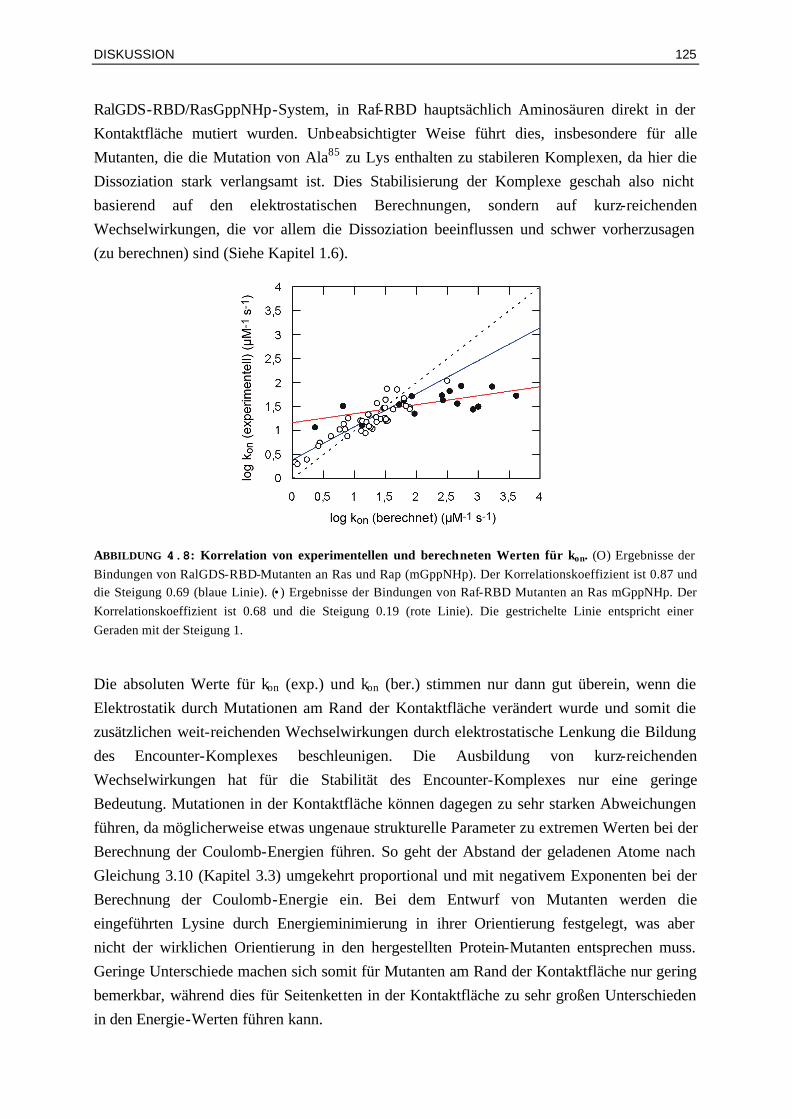
DISKUSSION
125
RalGDS-RBD/RasGppNHp-System, in Raf-RBD hauptsächlich Aminosäuren direkt in der Kontaktfläche mutiert wurden. Unbeabsichtigter Weise führt dies, insbesondere für alle Mutanten, die die Mutation von Ala85 zu Lys enthalten zu stabileren Komplexen, da hier die Dissoziation stark verlangsamt ist. Dies Stabilisierung der Komplexe geschah also nicht basierend auf den elektrostatischen Berechnungen, sondern auf kurz-reichenden Wechselwirkungen, die vor allem die Dissoziation beeinflussen und schwer vorherzusagen (zu berechnen) sind (Siehe Kapitel 1.6).
ABBILDUNG 4.8: Korrelation von experimentellen und berechneten Werten für kon. (Ο) Ergebnisse der Bindungen von RalGDS-RBD-Mutanten an Ras und Rap (mGppNHp). Der Korrelationskoeffizient ist 0.87 und die Steigung 0.69 (blaue Linie). (•) Ergebnisse der Bindungen von Raf-RBD Mutanten an Ras ⋅mGppNHp. Der Korrelationskoeffizient ist 0.68 und die Steigung 0.19 (rote Linie). Die gestrichelte Linie entspricht einer Geraden mit der Steigung 1.
Die absoluten Werte für kon (exp.) und kon (ber.) stimmen nur dann gut überein, wenn die Elektrostatik durch Mutationen am Rand der Kontaktfläche verändert wurde und somit die zusätzlichen weit-reichenden Wechselwirkungen durch elektrostatische Lenkung die Bildung des Encounter-Komplexes beschleunigen. Die Ausbildung von kurz-reichenden Wechselwirkungen hat für die Stabilität des Encounter-Komplexes nur eine geringe Bedeutung. Mutationen in der Kontaktfläche können dagegen zu sehr starken Abweichungen führen, da möglicherweise etwas ungenaue strukturelle Parameter zu extremen Werten bei der Berechnung der Coulomb-Energien führen. So geht der Abstand der geladenen Atome nach Gleichung 3.10 (Kapitel 3.3) umgekehrt proportional und mit negativem Exponenten bei der Berechnung der Coulomb-Energie ein. Bei dem Entwurf von Mutanten werden die eingeführten Lysine durch Energieminimierung in ihrer Orientierung festgelegt, was aber nicht der wirklichen Orientierung in den hergestellten Protein-Mutanten entsprechen muss. Geringe Unterschiede machen sich somit für Mutanten am Rand der Kontaktfläche nur gering bemerkbar, während dies für Seitenketten in der Kontaktfläche zu sehr großen Unterschieden in den Energie-Werten führen kann.
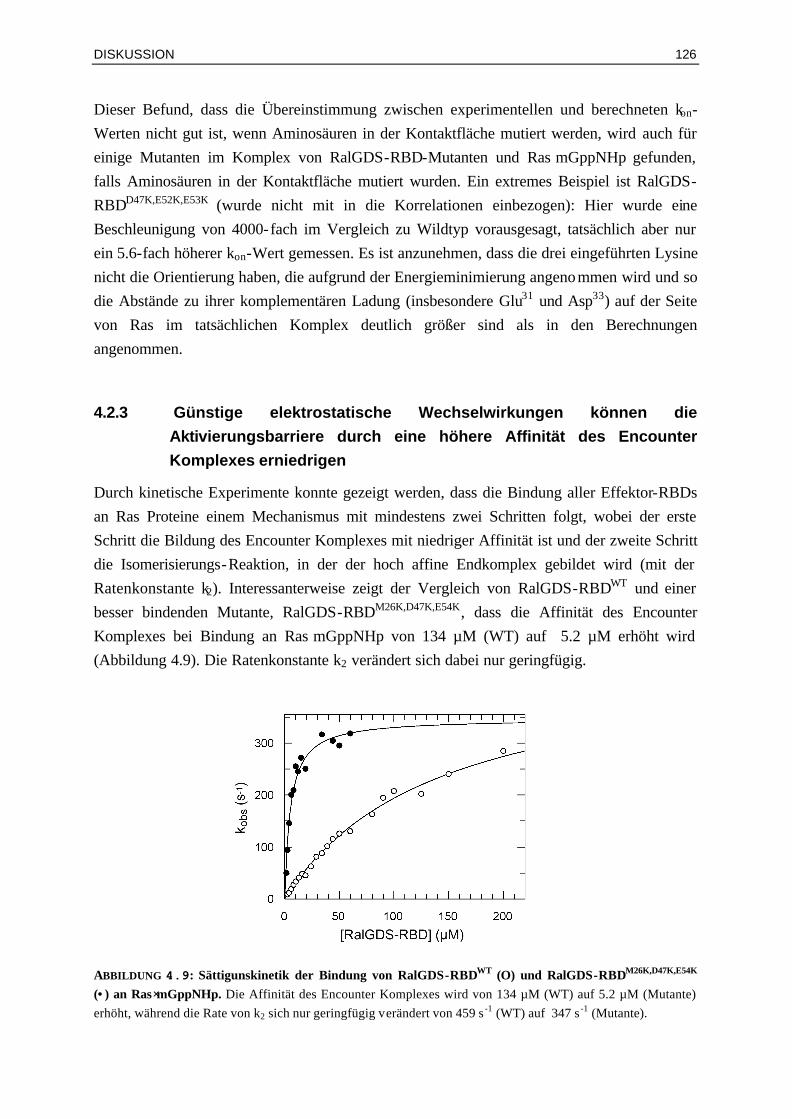
DISKUSSION
126
Dieser Befund, dass die Übereinstimmung zwischen experimentellen und berechneten kon-Werten nicht gut ist, wenn Aminosäuren in der Kontaktfläche mutiert werden, wird auch für einige Mutanten im Komplex von RalGDS-RBD-Mutanten und Ras⋅mGppNHp gefunden, falls Aminosäuren in der Kontaktfläche mutiert wurden. Ein extremes Beispiel ist RalGDS-RBDD47K,E52K,E53K (wurde nicht mit in die Korrelationen einbezogen): Hier wurde eine Beschleunigung von 4000-fach im Vergleich zu Wildtyp vorausgesagt, tatsächlich aber nur ein 5.6-fach höherer kon-Wert gemessen. Es ist anzunehmen, dass die drei eingeführten Lysine nicht die Orientierung haben, die aufgrund der Energieminimierung angenommen wird und so die Abstände zu ihrer komplementären Ladung (insbesondere Glu31 und Asp33) auf der Seite von Ras im tatsächlichen Komplex deutlich größer sind als in den Berechnungen angenommen.
4.2.3 Günstige elektrostatische Wechselwirkungen können die Aktivierungsbarriere durch eine höhere Affinität des Encounter Komplexes erniedrigen
Durch kinetische Experimente konnte gezeigt werden, dass die Bindung aller Effektor-RBDs an Ras Proteine einem Mechanismus mit mindestens zwei Schritten folgt, wobei der erste Schritt die Bildung des Encounter Komplexes mit niedriger Affinität ist und der zweite Schritt die Isomerisierungs-Reaktion, in der der hoch affine Endkomplex gebildet wird (mit der Ratenkonstante k2). Interessanterweise zeigt der Vergleich von RalGDS-RBDWT und einer besser bindenden Mutante, RalGDS-RBDM26K,D47K,E54K, dass die Affinität des Encounter Komplexes bei Bindung an Ras⋅mGppNHp von 134 µM (WT) auf 5.2 µM erhöht wird (Abbildung 4.9). Die Ratenkonstante k2 verändert sich dabei nur geringfügig.
ABBILDUNG 4.9: Sättigunskinetik der Bindung von RalGDS-RBDWT (Ο) und RalGDS-RBDM26K,D47K,E54K
(•) an Ras⋅mGppNHp. Die Affinität des Encounter Komplexes wird von 134 µM (WT) auf 5.2 µM (Mutante) erhöht, während die Rate von k2 sich nur geringfügig verändert von 459 s -1 (WT) auf 347 s -1 (Mutante).

DISKUSSION
127
Der Encounter Komplex kann als ein Ensemble von Konformationen betrachtet werden, in denen die Moleküle aneinander vorbei diffundieren und an vielen kleinen Mikrokollisionen beteiligt sind (Camacho et al., 1999). Die Art der Wechselwirkungen, die zur Stabilität des Encounter Komplexes führen wurde vielfach diskutiert, und beinhalten van-der-Waals-Kontakte und Wasserstoffbrücken-Bindungen sowie elektrostatische und auch hydrophobe Effekte (Sommer et al., 1982; Berg & von Hippel, 1985; Schreiber & Fersht, 1996) Allerdings ist der Encounter Komplex experimentell sehr schwer zugänglich. Für die meisten Protein-Protein Wechselwirkungen wurde kein Encounter Komplex gefunden, was aber seine Existenz nicht ausschließt, da dieser häufig nur bei sehr hohen Protein-Konzentrationen nachgewiesen werden kann, was zu sehr großen Werten für kobs führt (Schreiber, 2002). Diese kobs-Werte sind dann an der Grenze dessen, was mit Stopped-Flow-Messungen detektiert werden kann.
Die Korrelation von Änderungen der Aktivierungsenergie zwischen RalGDS-RBDWT und Mutanten (∆∆G#) im Komplex mit Ras⋅mGppNHp zeigt, dass diese Werte sehr gut mit den entsprechenden Änderungen der freien Enthalpie (∆∆G°) bei der Komplexbildung übereinstimmen (die Korrelationskoeffizienten liegen bei 0.9 und die Steigungen nahe bei 1). Die besseren Affinitäten von RalGDS-RBD Mutanten mit höherer elektrostatischer Komplementarität im Komplex mit Ras⋅mGppNHp sind somit eine Folge der erniedrigten Aktivierungsbarriere zwischen den Ausgangsproteinen und dem Endkomplex (Abbildung 4.10).
Wie bereits diskutiert ist die Verwendung von kon-Werten zur Berechnung von Aktivierungsenergien eigentlich nicht möglich, da die Gleichung nur für mikroskopische Ratenkonstanten, d.h. für die Betrachtung von einzelnen Schritten verwendet werden kann. Die absoluten Werte sind somit nicht zu erhalten, aber die Betrachtung von Änderungen ist möglich (siehe Kapitel 3.6).
Zusammen mit dem Befund, dass bei diesen Mutanten die Affinität des Encounter Komplexes erhöht ist, lässt den Schluss zu, dass günstige elektrostatische Wechselwirkungen in Protein-Komplexen die Aktivierungsbarriere erniedrigen und um denselben Betrag die Affinität des Encounter Komplexes erniedrigen. Im Gegensatz dazu wurden für die Wechselwirkungen von Raf-RBD-Mutanten im Komplex mit Ras⋅mGppNHp und Ras⋅mGDP keine guten Korrelationen von ∆∆G# und ∆∆G° gefunden werden. Die Korrelationskoeffizienten liegen im Bereich von 0.8 oder schlechter, was abhängig davon ist, welche Komplexe als Ausreißer nicht mit in den Vergleich einbezogen wurden. Wichtiger ist aber, dass hier die Steigungen mit 1.7 (Ras⋅GppNHp) und 2.6 (Ras⋅GDP) deutlich von 1 abweichen. Dies zeigt, dass nicht nur die geringeren Aktivierungsbarrieren für die besseren Affinitäten verantwortlich sind, sondern auch Änderungen im zweiten Schritt des Bindungsmechanismus. Dies ist gleichzeitig eine weitere Erklärung für die Abweichungen von experimentellen und berechneten Werten für kon, da die Berechnungen darauf beruhen, dass dieser zweite Schritt nicht verändert wird
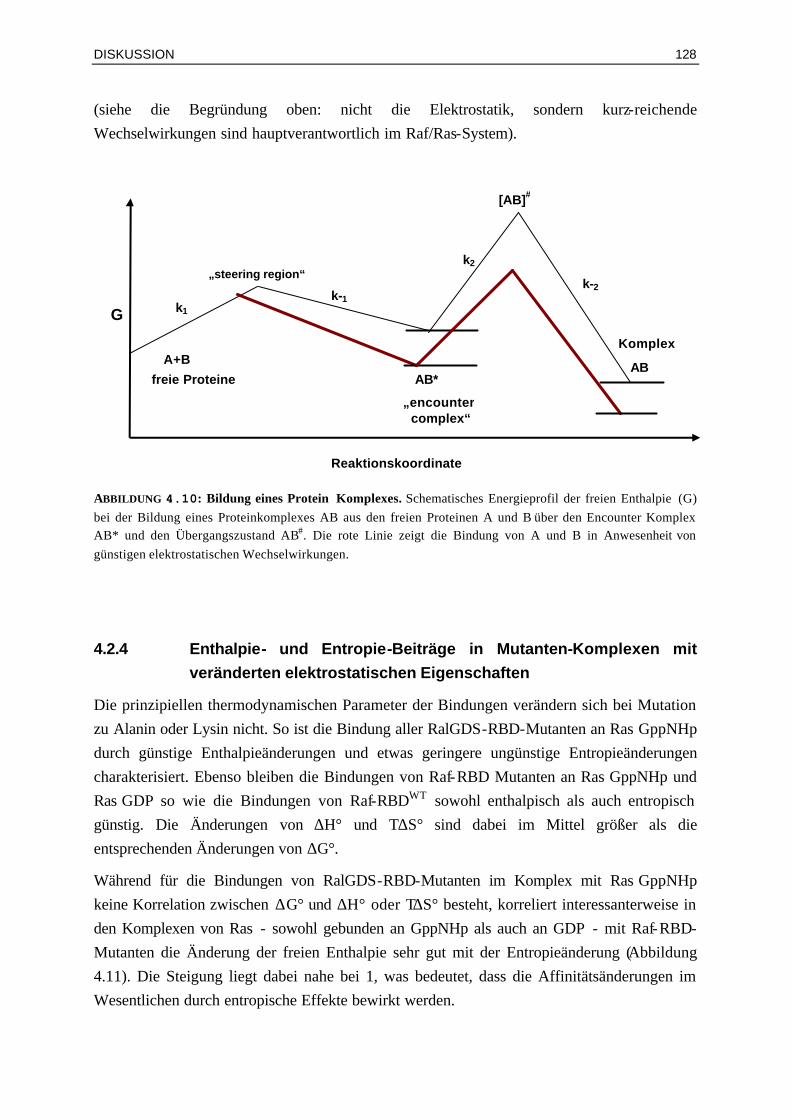
DISKUSSION
128
(siehe die Begründung oben: nicht die Elektrostatik, sondern kurz-reichende Wechselwirkungen sind hauptverantwortlich im Raf/Ras-System).
ABBILDUNG 4.10: Bildung eines Protein Komplexes. Schematisches Energieprofil der freien Enthalpie (G) bei der Bildung eines Proteinkomplexes AB aus den freien Proteinen A und B über den Encounter Komplex AB* und den Übergangszustand AB#. Die rote Linie zeigt die Bindung von A und B in Anwesenheit von günstigen elektrostatischen Wechselwirkungen.
4.2.4 Enthalpie- und Entropie-Beiträge in Mutanten-Komplexen mit veränderten elektrostatischen Eigenschaften
Die prinzipiellen thermodynamischen Parameter der Bindungen verändern sich bei Mutation zu Alanin oder Lysin nicht. So ist die Bindung aller RalGDS-RBD-Mutanten an Ras⋅GppNHp durch günstige Enthalpieänderungen und etwas geringere ungünstige Entropieänderungen charakterisiert. Ebenso bleiben die Bindungen von Raf-RBD Mutanten an Ras⋅GppNHp und Ras⋅GDP so wie die Bindungen von Raf-RBDWT sowohl enthalpisch als auch entropisch günstig. Die Änderungen von ∆H° und T∆S° sind dabei im Mittel größer als die entsprechenden Änderungen von ∆G°.
Während für die Bindungen von RalGDS-RBD-Mutanten im Komplex mit Ras⋅GppNHp keine Korrelation zwischen ∆G° und ∆H° oder T∆S° besteht, korreliert interessanterweise in den Komplexen von Ras - sowohl gebunden an GppNHp als auch an GDP - mit Raf-RBD-Mutanten die Änderung der freien Enthalpie sehr gut mit der Entropieänderung (Abbildung 4.11). Die Steigung liegt dabei nahe bei 1, was bedeutet, dass die Affinitätsänderungen im Wesentlichen durch entropische Effekte bewirkt werden.
A+B
AB*
[AB]#
AB
G
Reaktionskoordinate
„steering region“
freie Proteine
„encounter complex“
Komplex
k-1 k1
k2
k-2
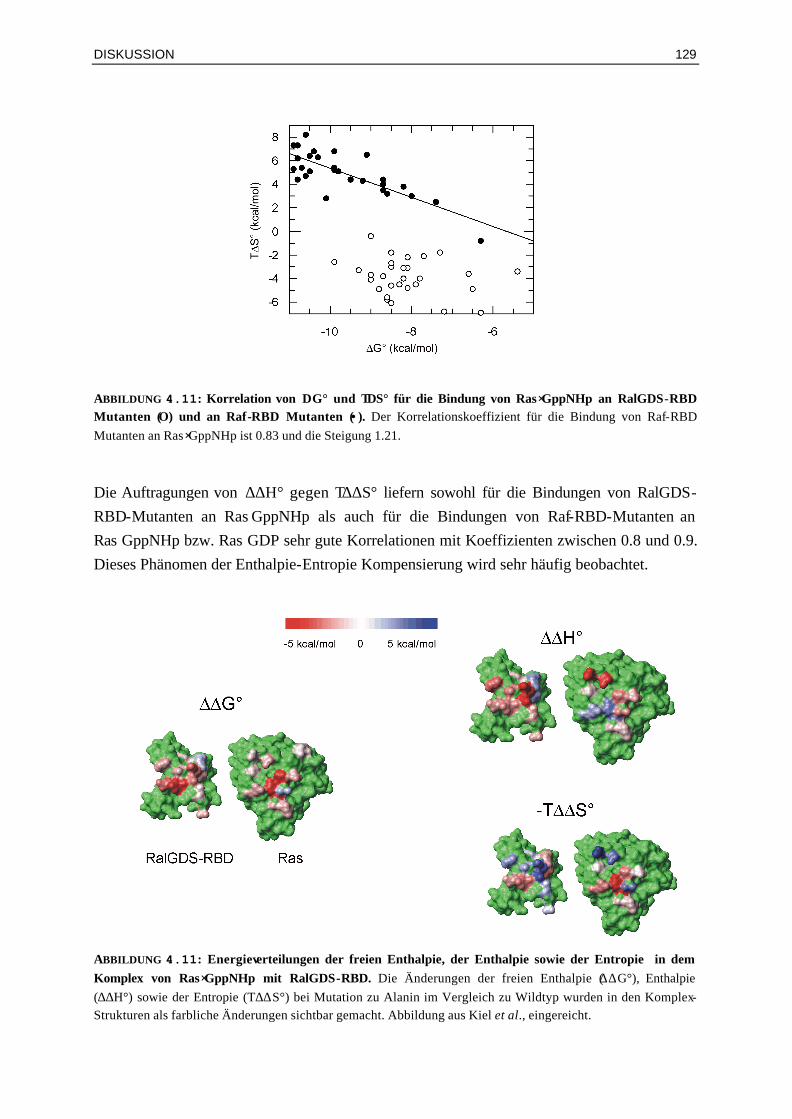
DISKUSSION
129
ABBILDUNG 4.11: Korrelation von ∆ G° und T∆S° für die Bindung von Ras⋅GppNHp an RalGDS-RBD Mutanten (Ο) und an Raf-RBD Mutanten (•). Der Korrelationskoeffizient für die Bindung von Raf-RBD Mutanten an Ras ⋅GppNHp ist 0.83 und die Steigung 1.21.
Die Auftragungen von ∆∆H° gegen T∆∆S° liefern sowohl für die Bindungen von RalGDS-RBD-Mutanten an Ras⋅GppNHp als auch für die Bindungen von Raf-RBD-Mutanten an Ras⋅GppNHp bzw. Ras⋅GDP sehr gute Korrelationen mit Koeffizienten zwischen 0.8 und 0.9. Dieses Phänomen der Enthalpie-Entropie Kompensierung wird sehr häufig beobachtet.
ABBILDUNG 4.11: Energieverteilungen der freien Enthalpie, der Enthalpie sowie der Entropie in dem Komplex von Ras⋅GppNHp mit RalGDS-RBD. Die Änderungen der freien Enthalpie (∆∆G°), Enthalpie (∆∆H°) sowie der Entropie (T∆∆S°) bei Mutation zu Alanin im Vergleich zu Wildtyp wurden in den Komplex-Strukturen als farbliche Änderungen sichtbar gemacht. Abbildung aus Kiel et al., eingereicht.
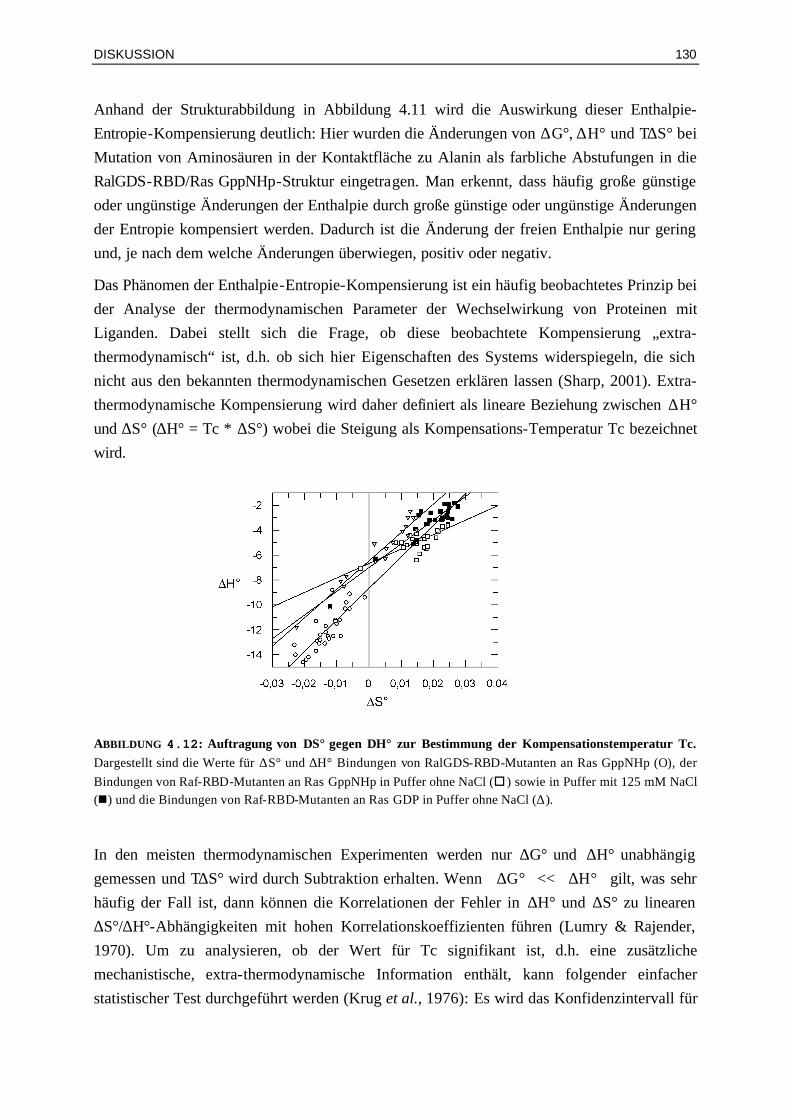
DISKUSSION
130
Anhand der Strukturabbildung in Abbildung 4.11 wird die Auswirkung dieser Enthalpie-Entropie-Kompensierung deutlich: Hier wurden die Änderungen von ∆G°, ∆H° und T∆S° bei Mutation von Aminosäuren in der Kontaktfläche zu Alanin als farbliche Abstufungen in die RalGDS-RBD/Ras⋅GppNHp-Struktur eingetragen. Man erkennt, dass häufig große günstige oder ungünstige Änderungen der Enthalpie durch große günstige oder ungünstige Änderungen der Entropie kompensiert werden. Dadurch ist die Änderung der freien Enthalpie nur gering und, je nach dem welche Änderungen überwiegen, positiv oder negativ.
Das Phänomen der Enthalpie-Entropie-Kompensierung ist ein häufig beobachtetes Prinzip bei der Analyse der thermodynamischen Parameter der Wechselwirkung von Proteinen mit Liganden. Dabei stellt sich die Frage, ob diese beobachtete Kompensierung „extra-thermodynamisch“ ist, d.h. ob sich hier Eigenschaften des Systems widerspiegeln, die sich nicht aus den bekannten thermodynamischen Gesetzen erklären lassen (Sharp, 2001). Extra-thermodynamische Kompensierung wird daher definiert als lineare Beziehung zwischen ∆H° und ∆S° (∆H° = Tc * ∆S°) wobei die Steigung als Kompensations-Temperatur Tc bezeichnet wird.
ABBILDUNG 4.12: Auftragung von ∆S° gegen ∆H° zur Bestimmung der Kompensationstemperatur Tc. Dargestellt sind die Werte für ∆S° und ∆H° Bindungen von RalGDS-RBD-Mutanten an Ras ⋅GppNHp (Ο), der Bindungen von Raf-RBD-Mutanten an Ras ⋅GppNHp in Puffer ohne NaCl (o ) sowie in Puffer mit 125 mM NaCl (n) und die Bindungen von Raf-RBD-Mutanten an Ras ⋅GDP in Puffer ohne NaCl (∆).
In den meisten thermodynamischen Experimenten werden nur ∆G° und ∆H° unabhängig gemessen und T∆S° wird durch Subtraktion erhalten. Wenn ∆G° << ∆H° gilt, was sehr häufig der Fall ist, dann können die Korrelationen der Fehler in ∆H° und ∆S° zu linearen ∆S°/∆H°-Abhängigkeiten mit hohen Korrelationskoeffizienten führen (Lumry & Rajender, 1970). Um zu analysieren, ob der Wert für Tc signifikant ist, d.h. eine zusätzliche mechanistische, extra-thermodynamische Information enthält, kann folgender einfacher statistischer Test durchgeführt werden (Krug et al., 1976): Es wird das Konfidenzintervall für
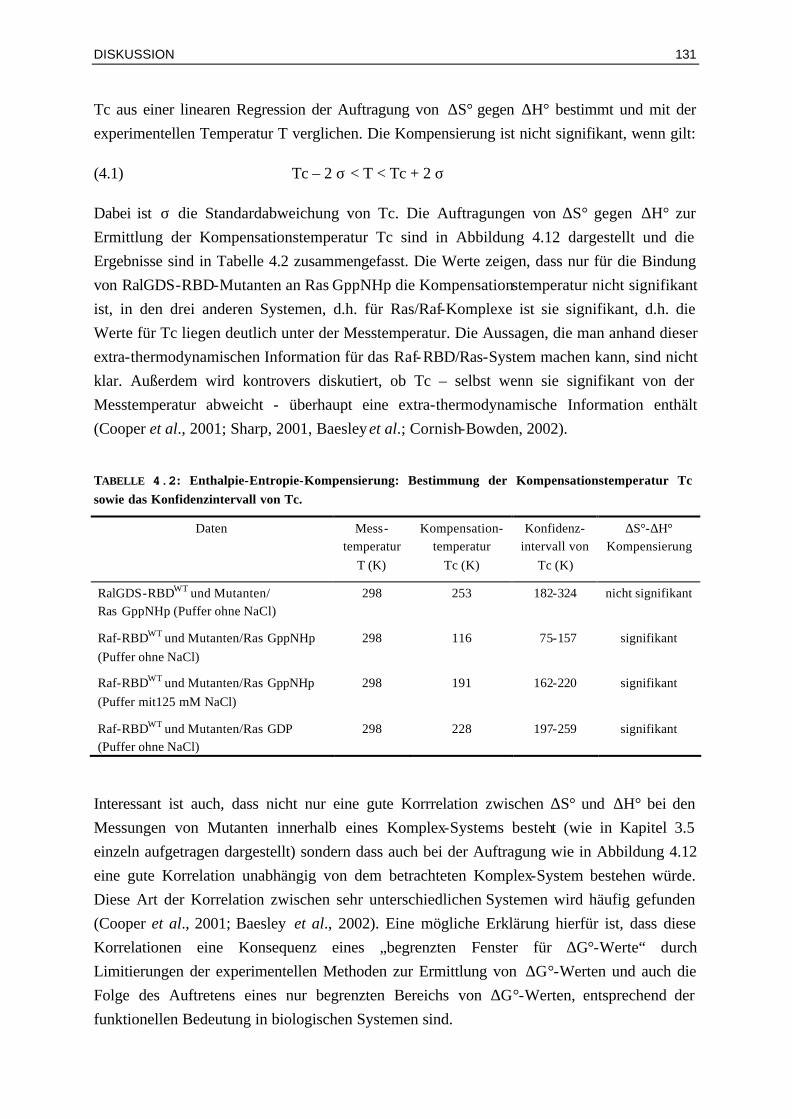
DISKUSSION
131
Tc aus einer linearen Regression der Auftragung von ∆S° gegen ∆H° bestimmt und mit der experimentellen Temperatur T verglichen. Die Kompensierung ist nicht signifikant, wenn gilt:
(4.1) Tc – 2 σ < T < Tc + 2 σ
Dabei ist σ die Standardabweichung von Tc. Die Auftragungen von ∆S° gegen ∆H° zur Ermittlung der Kompensationstemperatur Tc sind in Abbildung 4.12 dargestellt und die Ergebnisse sind in Tabelle 4.2 zusammengefasst. Die Werte zeigen, dass nur für die Bindung von RalGDS-RBD-Mutanten an Ras⋅GppNHp die Kompensationstemperatur nicht signifikant ist, in den drei anderen Systemen, d.h. für Ras/Raf-Komplexe ist sie signifikant, d.h. die Werte für Tc liegen deutlich unter der Messtemperatur. Die Aussagen, die man anhand dieser extra-thermodynamischen Information für das Raf-RBD/Ras-System machen kann, sind nicht klar. Außerdem wird kontrovers diskutiert, ob Tc – selbst wenn sie signifikant von der Messtemperatur abweicht - überhaupt eine extra-thermodynamische Information enthält (Cooper et al., 2001; Sharp, 2001, Baesley et al.; Cornish-Bowden, 2002).
TABELLE 4.2: Enthalpie-Entropie-Kompensierung: Bestimmung der Kompensationstemperatur Tc sowie das Konfidenzintervall von Tc.
Interessant ist auch, dass nicht nur eine gute Korrrelation zwischen ∆S° und ∆H° bei den Messungen von Mutanten innerhalb eines Komplex-Systems besteht (wie in Kapitel 3.5 einzeln aufgetragen dargestellt) sondern dass auch bei der Auftragung wie in Abbildung 4.12 eine gute Korrelation unabhängig von dem betrachteten Komplex-System bestehen würde. Diese Art der Korrelation zwischen sehr unterschiedlichen Systemen wird häufig gefunden (Cooper et al., 2001; Baesley et al., 2002). Eine mögliche Erklärung hierfür ist, dass diese Korrelationen eine Konsequenz eines „begrenzten Fenster für ∆G°-Werte“ durch Limitierungen der experimentellen Methoden zur Ermittlung von ∆G°-Werten und auch die Folge des Auftretens eines nur begrenzten Bereichs von ∆G°-Werten, entsprechend der funktionellen Bedeutung in biologischen Systemen sind.
Daten Mess- temperatur
T (K)
Kompensation- temperatur
Tc (K)
Konfidenz-intervall von
Tc (K)
∆S°-∆H° Kompensierung
RalGDS-RBDWT und Mutanten/ Ras⋅GppNHp (Puffer ohne NaCl)
298 253 182-324 nicht signifikant
Raf-RBDWT und Mutanten/Ras ⋅GppNHp (Puffer ohne NaCl)
298 116 75-157 signifikant
Raf-RBDWT und Mutanten/Ras ⋅GppNHp (Puffer mit125 mM NaCl)
298 191 162-220 signifikant
Raf-RBDWT und Mutanten/Ras ⋅GDP (Puffer ohne NaCl)
298 228 197-259 signifikant

DISKUSSION
132
4.3 Bindung von Raf-RBDWT und Mutanten an Ras⋅GDP und
Ras⋅GppNHpT35A Durch Phosphor-NMR-Messungen konnte gezeigt werden, dass Ras⋅GppNHp in zwei Zuständen vorliegt, die durch unterschiedliche Konformationen (Zustand 1 und 2) gekennzeichnet sind, was durch eine Aufspaltung der Resonanzfrequenzen am ß- und geringfügig auch am α-Phosphat sichtbar wird (Geyer et al., 1996). Diese Aufspaltung wurde durch die Mobilität der Schalter-Regionen von Ras gedeutet, wobei in einem Zustand Tyr32 von Ras in die Nähe des γ-Phosphates gelangt und dadurch der π-Elektronen-Ringstrom-Effekt des aromatischen Rings die beobachtete Hochfeldverschiebung der Resonanzlinien (Zustand 2) verursacht. Bei Titration mit einer Effektor-RBD wird Zustand 2 schon bei geringen RBD-Konzentrationen stabilisiert, bis Zustand 1 bei höheren Konzentrationen in Abhängigkeit von der Affinität des Ras/RBD-Komplexes nicht mehr detektiert wird. (Geyer et al., 1996, Linnemann et al., 1999, Spoerner et al., 2001). Somit wurde Zustand 1 als ungebundene und Zustand 2 als gebundene Konformation gedeutet. Des Weiteren konnte durch kinetische Messungen gezeigt werden, dass die Assoziation von Ras⋅GppNHp an alle RBDs zu einer Sättigung führt, was einen Bindungsmechanismus mit mindestens 2 Schritten anzeigt. Dabei entspricht die Ratenkonstante des zweiten Schrittes (k2) ungefähr der Umwandlungsrate zwischen Zustand 1 und Zustand 2 aus NMR-Experimenten. Dies wird so gedeutet, dass dieser Konformationswechsel der geschwindigkeitsbestimmende Schritt der Bindung ist.
Für die Ras-Mutanten Ras⋅GppNHpT35A und Ras⋅GppNHpT35S konnte keine Aufspaltung der Resonanzlinien am ß-Phosphat gezeigt werden (Geyer et al., 1996, Spoerner et al., 2001) und die chemische Verschiebung entspricht der von Zustand 1, d.h. der nicht bindenden Konformation. Bei Titration von Raf- oder RalGDS-RBDWT kann im Fall von Ras⋅GppNHpT35S der bindende Zustand 2 induziert werden. Außerdem kann bei der Assoziationskinetik eine Sättigung gemessen werden, was zeigt, dass erneut die Umwandlung in Zustand 2 der geschwindigkeitsbestimmende Schritt ist. Im Fall von Ras⋅GppNHpT35A kann dagegen Konformation 1 nicht in den bindenden Zustand überführt werden (Spoerner et al., 2001) und es wird auch keine Sättigungskinetik beobachtet. Die Assoziation von Ras⋅GppNHpT35A an RBDs erfolgt somit ohne geschwindigkeitsbestimmende Konformationsänderung der Schalter Bereiche.
Beim Vergleich von Ras-Strukturen in den GDP- und GTP- bzw. GppNHp- gebundenen Zuständen zeigt sich, dass die hoch konservierte Aminosäure Thr 35 von Ras eine zentrale Rolle besitzt. Im GppNHp-gebundenen Zustand koordiniert Thr35 das Mg2+ und ist zusammen mit Gly60 an der Bindung des γ-Phosphats beteiligt (Vetter & Wittinghofer, 2001). Bei GTP-Hydrolyse und damit bei Verlust der Kontakte zum γ-Phosphat, relaxieren Schalter I und II, was mit dem Mechanismus einer gespannten Feder („loaded-spring mechanism“) verglichen werden kann (Abbildung 4.13). Dabei kommt es zu den beschriebenen Positionsänderungen

DISKUSSION
133
der Schalterregionen beim Wechsel zwischen dem GDP- und GTP-gebundenen Zustand. Möglicherweise besitzen somit Ras⋅GDP und Ras⋅GppNHpT35A ähnliche Eigenschaften, da in beiden Fällen Ras nicht in dem „gespannten Zustand“ vorliegt, welche günstig für die Effektorbindung ist. Daher wird im Folgen die Bindung von Raf-RBDWT und Mutanten an Ras⋅GppNHpT35A und Ras⋅GDP vergleichend diskutiert.
ABBILDUNG 4.13: Schematische Darstellung des Schalter-Mechanismus von Ras. Schalter I und II (switch I und II) sind an das γ-Phosphat über die Hauptketten-NH-Gruppen der Aminosäuren Thr35 und Gly60 gebunden. Die Hydrolyse von GTP, und damit der Verlust der Kontakte zum γ-Phosphat, führt zu Positionsänderungen von Schalter I und II. Dies kann mit dem Mechanismus einer gespannten Feder verglichen werden („loaded-spring-mechanism“). Abbildung aus Vetter & Wittinghofer, 2001.
.
4.3.1 Zusammenhang von kinetischen Parametern, Sättigungskinetik und Aufspaltung der Phosphor-Resonanzen
Alle Effektor-RBDs zeigen bei der Assoziation an Ras Proteine eine Abnahme der mant-Fluoreszenz bei der Assoziation und eine Zunahme bei der Dissoziation. Im Gegensatz dazu sind diese Fluoreszenzänderungen genau umgekehrt bei der Bindung von Raf-RBDWT und Mutanten an Ras⋅mGDP und an Ras⋅mGppNHpT35A, d.h. Zunahme der Fluoreszenz bei Assoziation und Abnahme bei Dissoziation. Dieses gibt einen ersten Hinweis darauf, dass der Mechanismus der Bindung möglicherweise anders ist, bzw. dass die Kontaktfläche in den Komplexen etwas anders gebildet wird. Während für Ras⋅mGppNHpT35A im Komplex mit Raf-RBDWT und Mutanten die Amplituden sehr groß sind (bei 15 und 25 °C), sind diese bei der Bindung von Ras⋅mGDP an Raf-RBDWT und Mutanten bei 25 °C etwas kleiner, nehmen bei Erniedrigung der Temperatur ab und bei 15 °C und niedrigeren Temperaturen kann keine Fluoreszenzänderung beobachtet werden.
Die Raf-RBD-Mutanten, die zu einer höheren elektrostatischen Komplementarität im Komplex mit Ras führen, zeigen bei der Bindung an Ras⋅mGppNHpT35A - ebenso wie bei der Bindung an Ras⋅mGppNHpWT und Ras⋅mGDP – einen höheren Wert für kon. In Tabelle 4.2 sind die kinetischen Parameter zum Vergleich für Raf-RBDA85K und Raf-RBDWT bei der Bindung an Ras⋅GDP, Ras⋅mGppNHpT35A und Ras⋅mGppNHpWT gegenübergestellt.
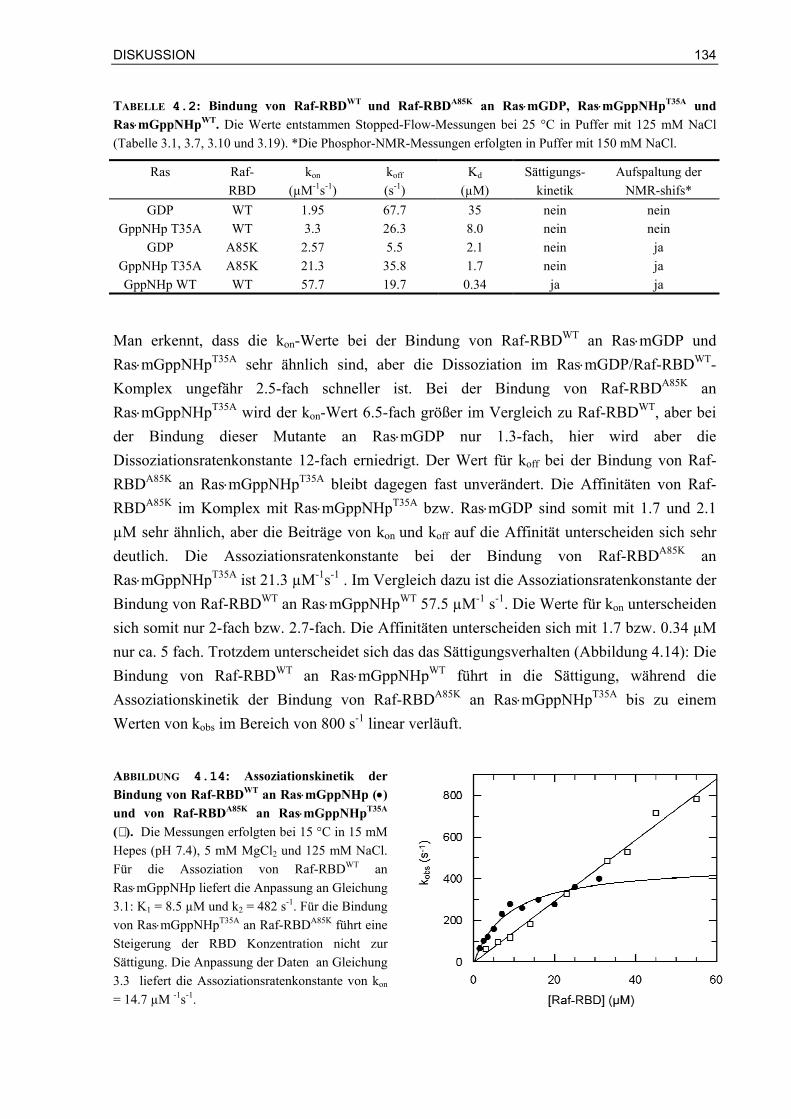
DISKUSSION
134
TABELLE 4.2: Bindung von Raf-RBDWT und Raf-RBDA85K an Ras⋅mGDP, Ras⋅mGppNHpT35A und Ras⋅mGppNHpWT. Die Werte entstammen Stopped-Flow-Messungen bei 25 °C in Puffer mit 125 mM NaCl (Tabelle 3.1, 3.7, 3.10 und 3.19). *Die Phosphor-NMR-Messungen erfolgten in Puffer mit 150 mM NaCl.
Ras Raf-RBD
kon (µM-1s-1)
koff (s-1)
Kd (µM)
Sättigungs- kinetik
Aufspaltung der NMR-shifs*
GDP WT 1.95 67.7 35 nein nein GppNHp T35A WT 3.3 26.3 8.0 nein nein
GDP A85K 2.57 5.5 2.1 nein ja GppNHp T35A A85K 21.3 35.8 1.7 nein ja GppNHp WT WT 57.7 19.7 0.34 ja ja
Man erkennt, dass die kon-Werte bei der Bindung von Raf-RBDWT an Ras⋅mGDP und Ras⋅mGppNHpT35A sehr ähnlich sind, aber die Dissoziation im Ras⋅mGDP/Raf-RBDWT-Komplex ungefähr 2.5-fach schneller ist. Bei der Bindung von Raf-RBDA85K an Ras⋅mGppNHpT35A wird der kon-Wert 6.5-fach größer im Vergleich zu Raf-RBDWT, aber bei der Bindung dieser Mutante an Ras⋅mGDP nur 1.3-fach, hier wird aber die Dissoziationsratenkonstante 12-fach erniedrigt. Der Wert für koff bei der Bindung von Raf-RBDA85K an Ras⋅mGppNHpT35A bleibt dagegen fast unverändert. Die Affinitäten von Raf-RBDA85K im Komplex mit Ras⋅mGppNHpT35A bzw. Ras⋅mGDP sind somit mit 1.7 und 2.1 µM sehr ähnlich, aber die Beiträge von kon und koff auf die Affinität unterscheiden sich sehr deutlich. Die Assoziationsratenkonstante bei der Bindung von Raf-RBDA85K an Ras⋅mGppNHpT35A ist 21.3 µM-1s-1 . Im Vergleich dazu ist die Assoziationsratenkonstante der Bindung von Raf-RBDWT an Ras⋅mGppNHpWT 57.5 µM-1 s-1. Die Werte für kon unterscheiden sich somit nur 2-fach bzw. 2.7-fach. Die Affinitäten unterscheiden sich mit 1.7 bzw. 0.34 µM nur ca. 5 fach. Trotzdem unterscheidet sich das das Sättigungsverhalten (Abbildung 4.14): Die Bindung von Raf-RBDWT an Ras⋅mGppNHpWT führt in die Sättigung, während die Assoziationskinetik der Bindung von Raf-RBDA85K an Ras⋅mGppNHpT35A bis zu einem Werten von kobs im Bereich von 800 s-1 linear verläuft.
ABBILDUNG 4.14: Assoziationskinetik der Bindung von Raf-RBDWT an Ras⋅mGppNHp (•) und von Raf-RBDA85K an Ras⋅mGppNHpT35A ( ). Die Messungen erfolgten bei 15 °C in 15 mM Hepes (pH 7.4), 5 mM MgCl2 und 125 mM NaCl. Für die Assoziation von Raf-RBDWT an Ras⋅mGppNHp liefert die Anpassung an Gleichung 3.1: K1 = 8.5 µM und k2 = 482 s-1. Für die Bindung von Ras⋅mGppNHpT35A an Raf-RBDA85K führt eine Steigerung der RBD Konzentration nicht zur Sättigung. Die Anpassung der Daten an Gleichung 3.3 liefert die Assoziationsratenkonstante von kon = 14.7 µM -1s-1.

DISKUSSION
135
Ebenso wird keine Sättigungskinetik gemessen für Raf-RBDWT und Raf-RBDA85K im Komplex mit Ras⋅mGDP (Sydor et al., 1998; diese Arbeit). Umso bemerkenswerter sind die Ergebnisse aus Phosphor-NMR-Messungen: In 31P-NMR-Titrationen von Raf-RBDA85K an Ras⋅GppNHpT35A kommt es wieder zu einer Aufspaltung am ß-Phosphat, wobei ein zweiter, Hochfeld verschobener Zustand 2 entsteht wie bei Ras⋅GppNHpWT. Ebenso zeigen Messungen von Raf-RBDA85K an Ras⋅GDP ebenfalls eine Hochfeld verschobene Aufspaltung der Resonanzfrequenzen, allerdings am α-Phosphat. Diese Aufspaltung wird bereits bei einem molaren Verhältnis 0.5 (Raf zu Ras) sichtbar und ist bei einem molaren Verhältnis von 4 vollständig in Zustand 2 überführt. Dieselbe Titration mit Raf-RBDWT zeigt keine eindeutige Aufspaltung der Signale und selbst bei einem molaren Verhältnis von 7 ist immer noch die Fläche von Zustand 1 größer.
Die Bindung von Effektor-RBDs an Ras Proteine kann unabhängig von dem Nukleotid-Zustand und der Mutation anhand des folgenden Schemas verdeutlicht werden, wobei die Ursachen der verschiedenen Eigenschaften nur in der unterschiedlichen Gleichgewichtslage liegen (Abbildung 4.15):
ABBILDUNG 4.15: Gleichgewichte bei der Bindung von Ras an Effektor-RBDs. Ras (1) und Ras (2) kennzeichnen die verschiedenen möglichen Konformationen von Ras, wobei Ras gebunden an GppNHp, GTP oder GDP sein kann und auch jeder Mutante entsprechen kann. K1, K2, K1’ und K2’ sind die jeweiligen Gleichgewichtskonstanten (Affinitäten) der entsprechenden Teilschritte.
Für die Bindung von RBDs an RasGppNHp kann die Bindung fast vollständig in den Zustand Ras (2)*RBD überführt werden, da Ras (1) und Ras (2) gebunden an GppNHp energetisch sehr ähnlich sind. Dies wurde durch Phosphor-NMR-Messungen gezeigt, da auch ohne Bindung von RBD zwei Zustände vorliegen, die ungefähr gleich populiert, und somit auf einem ähnlichen energetischen Niveau sind. Der geschwindigkeitsbestimmende Schritt der Reaktion ist die Bildung von Ras (2)*RBD und K1 entspricht der Afffinität des Encounter Komplexes.
Ras (1) und Ras (2) gebunden an GDP, bzw. die Mutante T35A gebunden an GppNHp besitzen dagegen große Energiedifferenzen, die ebenfalls aus Phosphor-NMR-Messungen

DISKUSSION
136
abgeschätzt werden können: Ohne Bindung von RBD liegt das Gleichgewicht weit auf der Seite von Ras (1), wobei ein Wert von ca. 3 kcal/mol für diese Energiedifferenz von Ras (1) und Ras (2) abgeschätzt werden kann (für K2’ ca. 100 M-1). Bei der Bindung von RBD an Ras⋅GDP, bzw. an Ras⋅GppNHpT35A kann der Zustand Ras (2)*RBD nicht erreicht werden, da die Energiedifferenz zwischen Ras (1)*RBD und Ras (2)*RBD zu groß und daher ungünstig ist. Das Gleichgewicht der Bindung ist somit stark zu Ras (1)*RBD verschoben, was erklärt, dass keine Sättigungskinetik und keine Aufspaltung der Phosphor-Resonanzen gemessen wird.
Die Ergebnisse der Bindung von Raf-RBDA85K an Ras⋅GDP und Ras⋅GppNHpT35A können dann folgendermaßen erklärt werden: Das Gleichgewicht K2’ ist unverändert und Ras (1) und Ras (2) besitzen wieder eine hohe Energiedifferenz, aber durch die günstigen elektrostatischen Wechselwirkungen wird die Affinität des Encounter Komplexes besser, d.h. K1 wird größer. Das Gleichgewicht ist somit deutlich weiter auf die Seite von Ras (1)*RBD verschoben als bei der Bindung von Raf-RBDWT. Durch die deutliche Verschiebung dieser Gleichgewichtslage ist anzunehmen, dass auch teilweise Ras (2)*RBD vorliegt, was die beobachtete Aufspaltung der Resonanzfrequenzen bei der Bindung von Raf-RBDA85K an Ras⋅GDP und Ras⋅GppNHpT35A erklären kann. Des Weiteren liefert dieses Schema eine Erklärung für die Abweichungen der Kd-Werte bestimmt aus NMR- und Stopped-Flow-Messungen, da bei NMR-Titrationen nur Ras (2)*RBD detektiert wird, aber nicht die Bildung von Ras (1)*RBD.
Der Grund weshalb keine Sättigungskinetik für die Bindung von Raf-RBD an Ras⋅GDP und Ras⋅GppNHpT35A gemessen werden kann ist, dass die Affinität des Encounter Komplexes so niedrig ist, dass diese im Stopped-Flow nicht detektiert werden kann, da die Ratenkonstanten von kobs am Limit sind, so dass keine weitere Erhöhung der Konzentration möglich ist.
4.3.2 Affinitäten und Wärmekapazitäten
Interessant ist, dass bei der Bindung von Raf-RBDA85K an Ras·GppNHpT35A bei steigender Temperatur die Fläche von Zustand 2 bei gleichem Konzentrationsverhältnis größer wird. Somit nimmt die Affinität mit steigender Temperatur zu. Daraus folgt, dass die Enthalpiänderung positiv ist, was auch durch ITC-Experimente bestätigt wurde. Möglicherweise sind die Schalter-Bereiche bei höherer Temperatur beweglicher und dadurch kann der Effektor das Ras Protein leichter in die für die Bindung günstigere Konformation drängen.
Die Wärmekapazitäten der Bindung von Effektor-RBDs an Ras Proteine im GppNHp-Zustand liegen zwischen -100 und -630 cal mol-1 K-1 (Rudolph et al., 2001). Die Werte für ∆Cp° von Raf-RBDWT im Komplex mit Ras⋅GppNHpT35A und Ras⋅GDP sind -98 bzw. -55 cal mol-1 k-1, also weniger negativ als für die Bindung der meisten RBDs an Ras⋅GppNHp. Bei

DISKUSSION
137
Assoziation von besser bindenden Raf-RBD-Mutanten an Ras⋅GppNHpT35A und Ras⋅GDP werden die ∆Cp°-Werte mit -250 (für Raf-RBDN56K,A85K im Komplex mit Ras⋅GppNHpT35A) und -306 bzw. -684 (für Raf-RBDA85K und Raf-RBDD80K,A85K,E125K im Komplex mit Ras⋅GDP) wieder deutlich negativer. Aus strukturellen Untersuchungen und Studien zur Protein-Faltung wurde folgende empirische Beziehung zwischen der Wärmekapazitätsänderung ∆Cp° und den Änderungen der Zugänglichkeit der Oberfläche für Wasser (∆ASA) gefunden (Murphy & Freire, 1992):
∆Cp° = 0.45 cal/(mol K Å2) ∆ASAapolar – 0.26 cal/(mol K Å2) ∆ASApolar
Nach dieser Beziehung trägt die apolare Oberfläche, die bei der Bindung abgedeckt wird negativ und die polare Oberfläche positiv zu ∆Cp° bei. Die berechneten Werten für ∆Cp° für die Bindung von Effektor-RBDs an Ras⋅GppNHp liegen alle bei Null, d.h. sind schwach positiv oder negativ (Rudolph et al., 2001). Möglicherweise wird bei den Berechnungen der Effekt der Schalter-Änderungen nicht berücksichtigt, und daher stimmen die Werte für die Bindungen von Raf-RBDWT an Ras⋅GppNHpT35A und Ras⋅GDP mit berechneten Werten besser überein, als die von Effektor-RBDs in Komplex mit Ras·GppNHp.
4.3.3 Mögliche Rolle der Mutation Lys85 in Raf-RBD
Durch die höhere Affinität von Raf-RBDA85K im Gegensatz zu Raf-RBDWT bewirkt die Bindung möglicherweise wieder Änderungen der Positionen der Schalter Regionen – auch ohne Thr35 von Ras. Eventuell kann diese Überführung in den zweiten Zustand – auch ohne Stabilisierung durch Thr35 von Ras durch starke energetisch günstige Wechselwirkungen zwischen Lys85 von Raf-RBD und Glu31 von Ras erklärt werden. Durch Doppel-Mutanten-Analysen wurden sowohl für Ras gebunden an GppNHp als auch an GDP, energetisch günstige Kopplungsenergien von -1.4 und -2.2 kcal/mol zwischen diesen Aminosäure-Resten gemessen. In Abbildung 4.16 ist ein Ausschnitt aus der Raf-RBD/Raps⋅GppNHp Komplexstruktur dargestellt, in der die Mutation von Ala85 zu Lys hineinmodelliert worden ist. Lys85 befindet sich neben Lys84 von Raf-RBD, das günstige Wechselwirkungen mit Glu31 und Asp33 von Ras machen kann, was durch Doppel-Mutanten-Analyse und durch strukturelle Daten gezeigt wurde (Nassar et al., 1996; Kiel et al., eingereicht). Möglicherweise kann sich Lys85 an der Ausbildung der Salzbrücke zu Glu31 und Asp33 beteiligen und diese so stark verstärken, dass Tyr32 wegklappen kann, in räumliche Nähe zum γ-Phosphat gelangen kann und die Aufspaltung der Resonanzlinien am ß-Phosphat erklären – ohne dass Thr35 die Schalter Regionen stabilisiert. Ebenso wäre dies eine mögliche Erklärung für die Bindung von Raf-RBDA85K an Ras⋅GDP: Lys85 macht energetisch günstige Wechselwirkungen mit Glu31 und Asp33 von Ras⋅GDP wodurch Tyr32 in die Nähe des α-Phospats gelangt und so die Aufspaltung der Resonanzlinien am α-Phosphat verursacht.

DISKUSSION
138
ABBILDUNG 4.16: Ausschnitt aus der Komplexstruktur von Raf-RBD/Raps·GppNHp mit der modellierten Mutation Ala85 zu Lys von Raf-RBD. Möglicherweise kann sich Lys85 zusammen mit Lys84 von Raf-RBD an der Ausbildung von ionischen Kontakten zu Glu31 und Asp33 von Ras beteiligen. Der Abstand von Lys85 zu Glu31 ist ca. 6.6 Å.
4.4 Biologische Bedeutung der Bindungskinetik Die zentrale Frage für das Verständnis der Bedeutung von Bindungsprozessen in biologischen Systemen ist, ob die Bindungsaffinität entscheidend für die Signalweiterleitung in vivo ist, oder ob die unabhängigen Beiträge von Ratenkonstanten der Assoziation und Dissoziation ebenfalls wichtig sind, d.h.: Ist es ausreichend, dass Bindungspartner fest aneinander binden oder müssen sie auch schnell aneinander binden?
Mehrere Studien der letzen Jahre geben Hinweise darauf, dass die individuellen kinetischen Ratenkonstanten wichtig sind, wie z.B. im System von Cytokin-Rezeptor Wechselwirkungen. So konnte für die Wechselwirkungen von Interleukin-4, dem humanen Wachstumshormon hGH sowie von Interferon mit ihren entsprechenden zellulären Rezeptoren gezeigt werden, dass eine lineare Abhängigkeit zwischen der Dissoziationsratenkonstanten und der biologischen Aktivität besteht (Wang et al., 1997; Pearce et al., 1999; Piehler et al., 2000). Diese lineare Beziehung gilt so lange wie koff größer als 0.02 s-1 ist, langsamere Dissoziationsratenkonstanten führen dabei zu keiner gesteigerten biologischen Aktivität.
Die Bedeutung der Optimierung von elektrostatischen Wechselwirkungen zur Beschleunigung der Assoziation zeigt ein weit verbreitetes Prinzip in biologischen Systemen, wie z.B. bei den Elektrostatinen als einer Klasse von Adhäsionsmolekülen mit konservierten elektrostatischen und konservierten Motiven (Botti et al., 1998) und bei der kleinen Ribonuklease Barnase mit seinem zellulären Inhibitor Barstar (Schreiber et al., 1994; Lee &

DISKUSSION
139
Tidor, 2001). Kinetische Untersuchungen von Antikörper-Antigen Wechselwirkungen zeigen eine Korrelation der Affinität und eine stärkere Korrelation der Assoziationsratenkonstante mit der biologischen Aktivität (Roost et al., 1995). Für die Bindung verschiedener Raf-RBD-Mutanten wurde eine Korrelation der Affinität mit der Signalweiterleitung in vivo festgestellt (Block et al., 1996; Jaitner et al., 1997). Da die Bindung ebenfalls durch eine elektrostatisch optimierte Kontaktfläche erfolgt und die Assoziation sehr schnell ist, könnte die Signalübertragung in diesem System ebenfalls assoziations-kontrolliert sein.
4.4.1 Ist die Signalweiterleitung in vivo nur abhängig von der Affinität oder
auch von der Assoziationsratenkonstante?
Die Ergebnisse des Reportergen-Assays zeigen, dass nach Transfektion von RasG12V und Raf-Mutanten eine gute Korrelation der Affinität (der entsprechenden Raf-RBD Mutanten im Komplex mit Ras⋅GppNHp) mit der Signalweiterleitung in vivo (gemessen an der Luziferase Aktivität) besteht. Die Korrelationskoeffizienten liegen bei 0.83, wenn die korrigierten (das bedeutet mit Abzug der Aktivitäten bei Transfektion von Raf-RBD Mutanten ohne RasG12V) Luziferase-Aktivitäten verwendet werden. Problematisch für die Auswertung der Ergebnisse ist, dass diese Luziferase-Aktivitäten der Kontrollen teilweise sehr hoch sind, was durch endogen vorhandenes Ras erklärt wird. Dieses sollte zwar hauptsächlich in der GDP-gebundenen Form vorliegen, aber wie gezeigt wurde, bestehen teilweise sehr hohe Affinitäten der entsprechenden Raf-RBD-Mutanten an Ras⋅GDP. Zur Auswertung wurden die Kontrollen abgezogen, was nicht richtig sein muss, da man nicht sagen kann, wie viel der Luziferase-Aktivität durch die Aktivierung über Ras⋅GDP oder Ras⋅GTP erfolgt. Dies erklärt auch, weshalb die Korrelationkoeffizienten mit und ohne Abzug der Kontrollen sehr ähnlich sind. Die Unterschiede in den Korrelationen bei Verwendung von Ergebnissen aus Messungen in Puffer mit 125 mM NaCl und in Puffer ohne Salz sind dagegen sehr eindeutig: Die Korrelationskoeffizienten sind in Puffer mit 125 mM NaCl wesentlich besser, was zu erwarten war, da dies der physiologischen Ionenstärke entspricht.
Die Korrelation der biologischen Aktivität mit den Assoziationsratenkonstanten (0.75) ist besser als mit den Affinitäten (0.85), wenn die Werte von RafR59A und RafT68K nicht mit einbezogen werden. Dies zeigt aber auch die Problematik beim Vergleich der absoluten Werte verschiedener Korrelationskoeffizienten, da teilweise einzelne Punkte eindeutig von der Korrelationsgeraden abweichen, während in anderen Fällen die Punkte sehr stark um eine Ausgleichsgerade streuen, was aber auch Berücksichtigung finden sollte. So zeigen die Korrelationen der Affinität mit der Luziferase-Aktivität viele um die Ausgleichsgerade verteilte Punkte, während bei der Auftragung von kon-Werten die Punkte recht gut auf der Geraden liegen, wenn einzelne Punkte herausgelassen werden. Zu beachten ist auch, dass die Fehlergrenzen in diesem biologischen Assay relativ hoch sind (in Einzelfällen bis zu 20 %).

DISKUSSION
140
Die Dissoziationsratenkonstanten korrelieren schlechter mit der in vivo Aktivität (Korrelationskoeffizient: 0.74). Auch hier streuen die Werte stark um die Ausgleichsgerade und es gibt keine auffälligen Ausreißer. Man kann somit sagen, dass eine Korrelation mit der Affinität, und in geringerem Maße mit der Dissoziationsratenkonstanten besteht, aber eine bessere Korrelation mit den Assoziationsratenkonstanten.
4.4.2 Für die Signalweiterleitung über Ras⋅GDP sind nicht die Assoziationsratenkonstanten, sondern die Lebenszeiten der Komplexe entscheidend
Nach Transfektion von RasWT und Raf-Mutanten zeigen die Ergebnisse des Reportergen-Assays, dass nur eine geringe Korrelation der Affinität (der entsprechenden Raf-RBD Mutanten im Komplex mit Ras⋅GDP gemessen in Puffer mit 125 mM NaCl) mit der Signalweiterleitung in vivo besteht. Der Korrelationskoeffizient liegt bei 0.63. Dabei ist die Übereinstimmung der Luziferase-Aktivitäten nach Transfektion von Raf-Mutanten und RasWT identisch (innerhalb des Fehlerbereichs) mit denen nach Transfektion von Raf alleine, was aufgrund des endogen vorhandenen Ras·GDP zu erwarten war. Die Assoziationsratenkonstanten korrelieren in diesem System gar nicht mit der Signalweiterleitung in vivo, aber es besteht eine sehr gute Korrelation mit der Dissoziationsratenkonstante. Dies entspricht den Erwartungen, da eine Korrelation von Kd mit koff besteht und nicht mit kon. Dabei liefern Korrelationen von Luziferase-Aktivitäten mit den koff-Werten gleiche Korrelationskoeffizienten (0.8), wenn die Ergebnisse aus Stopped-Flow Messungen in Puffer ohne NaCl und in Puffer mit 125 mM NaCl verglichen werden. Dies ist sehr interessant, da es noch einmal verdeutlicht, dass die Dissoziationsratenkonstanten bei niedrigen Salzkonzentrationen unabhängig von der Ionenstärke sind.
4.4.3 Vergleich der Effizienz der Signalweiterleitung über Ras⋅GDP und Ras⋅GTP
In Abbildung 4.17 ist ein Vergleich von log Kd und den Luziferase-Aktivitäten für die Bindung von RafWT und Mutanten an Ras⋅GDP und an Ras⋅GppNHp in einem Diagramm dargestellt. Es wurden jeweils die ITC- oder Stopped-Flow-Ergebnisse in Puffer mit 125 mM NaCl verwendet. Der Korrelationskoeffizient beträgt 0.79. Die Signalweiterleitung kann somit sowohl über Ras im GTP-Zustand als auch über Ras⋅GDP verlaufen und zeigt, dass nur die Affinität entscheidend ist und nicht, im welchem Nukleotid-Zustand Ras vorliegt. Die unterschiedlichen Steigungen (Abbildung 4.17) bei der Bindung von RafWT und Mutanten für die Signalweiterleitung über GTP und GTP könnten andeuten, dass die Signalübertragung über Ras⋅GTP effizienter ist. Möglicherweise ist dies aber die Folge davon, dass endogenes Ras auch teilweise im GTP-gebundenen Zustand vorliegt.

DISKUSSION
141
ABBILDUNG 4.16: Korrelation von log Kd und der Luziferase Aktivität für die Bindung von RafWT und Mutanten an Ras⋅GDP und Ras⋅GTP/GppNHp. Der Korrelationskoeffizient für alle Werte beträgt 0.79. Die Korrelation für die Bindung von RafWT und Mutanten an Ras⋅GTP ist 0.82 (blaue Linie). Die Korrelation für die Bindung von RafWT und Mutanten an Ras⋅GDP ist 0.63 (rote Linie).
4.4.4 Besteht eine Korrelation mit thermodynamischen Parametern und den Luziferase-Aktivitäten ?
Der Vergleich der thermodynamischen Parameter zeigt, dass keine zusätzliche Korrelation darüber hinaus besteht, was nicht auch schon bei der Korrelation mit den thermodynamischen Parametern und Affinitäten in vitro beobachtet wurde. So besteht keine Korrelation mit den Luziferase-Aktivitäten und den Enthalpieänderungen, aber es besteht eine Korrelation der Luziferase-Aktivitäten mit T∆S° für Raf-Mutanten bei der Signalweiterleitung über Ras⋅GTP und Ras⋅GDP (als Folge der Korrelation von Kd mit T∆S°).
4.4.5 Signalweiterleitung über RasT35A und RasT35S
Die Signalweiterleitung in vivo über RasG12V,T35S ist möglich und dabei korreliert die Affinität zwischen RasGppNHpT35S bei der Bindung an Raf-RBDWT und Mutanten mit der Luziferase-Aktivität, wobei diese Werte deutlich höher sind als die Kontrollen (Transfektion der entsprechenden Raf-Konstrukte allein). Die Signalweiterleitung von RafWT und Mutanten an RasG12V,T35A kann dagegen nicht wesentlich über die Luziferase-Werte von Transfektionen der entsprechenden Raf-Konstrukte allein gesteigert werden. Dies zeigt noch einmal, die ähnlichen Eigenschaften von RasGppNHpT35A und Ras·GDP: Auch bei Ras⋅GDP entspricht die Luziferase-Aktivität nach Transfektion von Raf-Mutanten mit RasWT ungefähr den Aktivitäten nach Transfektion von Raf-Mutanten allein.

ZUSAMMENFASSUNG
142
VI ZUSAMMENFASSUNG
Ziel der Arbeit war die Untersuchung der Bedeutung von elektrostatischen Wechselwirkungen in Ras/Effektor-Komplexen für die kinetischen und thermodynamischen Eigenschaften in vitro und für die Signalweiterleitung in vivo.
Die Mitglieder der Familie der Ras Proteine sind wichtige Schaltermoleküle in der Zelle, die zwischen einem GDP-gebundenen inaktiven und einem GTP-gebundene aktiven Zustand wechseln können. Ras⋅GTP kann mit nach geschalteten Proteinen (Effektoren) wechselwirken und dadurch wird eine Vielzahl von zellulären Prozessen, wie z. B. die Proliferation und Differenzierung reguliert. Alle Effektoren, die bisher analysiert wurden, besitzen einen gemeinsamen strukturellen Bereich (Ras-Bindungs-Domäne, RBD), über den die Bindung an den Effektor-Bereich von Ras⋅GTP erfolgt. Obwohl keine oder nur geringe Sequenzhomologie besteht, besitzen alle strukturell charakterisierten RBDs dieselbe Ubiquitin-ähnliche Faltung.
Es konnte gezeigt werden, dass die Bindung von Effektor-RBDs an Ras Proteine sehr spezifisch ist, obwohl die Primär-Sequenzen der Ras Proteine im Effektor-Bindungs-Bereich fast identisch sind und die RBDs alle denselben Faltungstyp zeigen. Außerdem sind die individuellen thermodynamischen Beiträge von Enthalpie und Entropie auf die freie Enthalpie der Bindung sehr unterschiedlich. Durch Alanin-Mutanten-Analyse konnte für die Komplexe zwischen Raf- bzw. RalGDS-RBD mit Ras·GppNHp gezeigt werden, dass die Energieverteilungen in den Komplexen sehr unterschiedlich sind. Dies zeigt, dass die spezifischen Unterschiede der Affinitäten als verschiedene Determinanten der Bindungsenergie auf der Protein Oberfläche kodiert sind.
Durch Doppel-Mutanten-Analyse konnten die Beiträge einzelner Aminosäurekontakte zur Bindungsenergie bestimmt werden und die Bindungsspezifitäten quantitativ analysiert werden, aber es ist nicht möglich, basierend auf den Röntgenstrukturen Bindungsaffinitäten zu berechnen. Der Grund hierfür ist, dass die energetischen Beiträge von kurz-reichenden Wechselwirkungen auf die Affinität bisher nur wenig verstanden sind und bisher kaum allgemeine Gesetzmäßigkeiten benannt werden können.
Im Gegensatz dazu konnte das Verständnis von weit-reichenden günstigen elektrostatischen Wechselwirkungen dazu benutzt werden, um gezielt Ras/Effektor-Komplexe mit veränderten Affinitäten herzustellen. Es konnte gezeigt werden, dass komplementäre elektrostatische Ladungen in der Kontaktfläche von Ras/Effektor-RBD-Komplexen die Assoziation beschleunigen, da die Bildung des Encounter Komplexes durch elektrostatische Lenkung beschleunigt wird. Die Dissoziation ist dagegen mit elektrostatischen Wechselwirkungen allein nicht zu beschreiben und zu berechnen.

ZUSAMMENFASSUNG
143
Durch Mutationen wurden gezielt zusätzliche positiv geladene Aminosäure-Reste in RalGDS- und Raf-RBD eingeführt, um die elektrostatische Komplementarität im Komplex mit Ras zu erhöhen. Stoppd-Flow-Messungen der RalGDS-RBD-Mutanten, die am Rand der Kontaktfläche eingeführt wurden, zeigen bei der Komplexbildung mit Ras⋅mGppNHp, dass die Assoziation beschleunigt ist, was zu einer verbesserten Affinität führt, da die Dissoziation nur wenig beeinflusst wird. Die Korrelation von experimentellen und berechneten Werten für kon zeigt eine gute Übereinstimmung. Des Weiteren konnte gezeigt werden, dass diese zusätzlichen günstigen elektrostatischen Wechselwirkungen die Aktivierungsbarriere durch eine höhere Affinität des Encounter Komplexes erniedrigen können. Ebenso konnten die Affinitäten in Komplexen von Raf-RBD-Mutanten mit Ras⋅mGppNHp und Ras⋅mGDP gesteigert werden. Allerdings ist die Übereinstimmung der Werte für kon nicht so gut, da hier hauptsächlich Mutanten in der Kontaktfläche analysiert wurden, was zu einer zusätzlichen Beeinflussung der Dissoziation führt, da kurz-reichende Wechselwirkungen beteiligt sind.
Die Affinitäten aus Stopped-Flow-Experimenten konnten für die RalGDS-RBD-Mutanten/Ras·GppNHp-Komplexe durch ITC-Messungen bestätigt werden. Im Fall der Bindungen von Raf-RBD-Mutanten an Ras⋅GppNHp und Ras⋅GDP ergeben sich jedoch teilweise deutliche Unterschiede. Während für die RalGDS-RBD-Mutanten im Komplex mit Ras·GppNHp keine Korrelation zwischen ∆G° und ∆H° bzw. T∆S° besteht, wird für die Bindungen von Raf-RBD-Mutanten sowohl im Komplex mit Ras⋅GppNHp als auch mit Ras⋅GDP eine gute Korrelation von ∆G° mit T∆S° gefunden.
Die Unterschiede der Bindungen von RBDs an Ras·GppNHp im Vergleich zu Ras·GppNHpT35A und Ras·GDP sind die Folge der unterschiedlichen Gleichgewichtslagen der Konformationen von Ras (Zustand 1 und 2). Im Fall der Bindung von RBDs an Ras·GppNHp kann die für die Bindung günstigere Konformation (Zustand 2 von Ras) einfacher eingenommen werden, da bereits ohne Anwesenheit von RBDs beide Zustände von Ras in etwa gleichen Verhältnissen vorliegen. Im Fall der Bindung an Ras·GDP und Ras·GppNHpT35A muss zu dieser Überführung eine Energie von ca. 3 kcal/mol aufgewendet werden und daher ist hier das Gleichgewicht auch im Komplex mit RBDs stark auf die Seite von Zustand 1 von Ras verschoben.
Die Untersuchung der Signalweiterleitung in vivo von Ras/Effektor Wechselwirkungen zeigt für die Weiterleitung über Ras⋅GTP, dass die Luziferase-Aktivität nicht nur abhängig von der Affinität sondern auch von der Assoziationsratenkonstante ist. Für die Signalweiterleitung über Ras⋅GDP sind dagegen nicht die Assoziationsratenkonstanten, sondern die Lebenszeiten der Komplexe entscheidend.

ANHANG
144
VI ANHANG 6.1 Abkürzungen Aminosäuren A (Ala, Alanin), C (Cys, Cystein), D (Asp, Aspartat) E Glutamat, F (Phe, Phenylalanin), G (Gly, Glycin), H (His, Histidin), I (Ile, Isoleucin), K (Lys, Lysin), L, (Leu, Leucin), M (Met, Methionin), N (Asp, Asparagin), P (Pro, Prolin), Q (Gln, Glutamin), R (Arg, Arginin), S (Ser, Serin), T (Thr, Threonin), V (Val, Valin), W (Trp, Tryptophan), Y (Tyr, Tyrosin) Amp Ampicillin APS Ammoniomperoxodisulphat DNA Desoxyribonukleinsäure DTE 1,4-Dithioerytrit EDTA Ethylene-diamine-tetraacetic acid ERK extrazellulär regulierte Kinase FPLC fast performance-liquid-chromatography g Gramm GAP GTPase-aktivierendes Protein GDP Guanin-5`-diphosphat GEF Guaninnukleotid Austauschfaktor (exchange factor) GppNHp 5`-Guanosyl-β, γ-Imido-Triphosphat GSH reduziertes Glutathion GST Glutathion-S-Transferase GTP Guanin-5`-triphosphat h Stunde IPTG Isopropyl-β-D-1-thiogalactopyranosid ITC isothermes Titrationskalorimeter Kan Kanamycin KDa Kilodalton kcal Kilokalorie mant 2`,3` - (N-methylanthraniloyl) MAPK mitogen aktivierte Protein-Kinase MAPKK MAPK-Kinase MAPKKK MAPKK-Kinase MEK MAP/ERK-Kinase min Minute ml Milliliter µM mikromolar OD optische Dichte PAGE Polyacrylamid-Gelelektrophorese PIA Protein Interface Analyse (protein interface analysis) PMSF Phenylmethansulfonyl-Fluorid Raf-RBD Ras-Bindungsdomäne der Proteinkinase c-Raf-1, kurz: Raf RalGDS-RBD Ras-Bindungsdomäne des RalGDS RalGEF Ral Guaninnukleotid-Austauschfaktor = RalGDS RapC` Rap1A am C-Terminus verkürzt Raps Rap1A (E30D, K31E), am C-Terminus verkürzt RBD Ras-Bindungsdomäne rpm Umdrehungen pro Minute SDS Natriumdodecylsulfat TEMED N,N,N`,N`-Tetramethylethyldiamin Tris Tris-(hydroxymethyl)-aminomethan UV Ultaviolett µl Mikroliter

ANHANG
145
6.2 Oligonukleotide Oligonukleotide für Sequenzierungen: pGEXs GGT GGT GGC GAC CAT CCT CC pGEXas GTC AGA GGT TTT CAC CGT CAT CAC CG pCDNA3s CCC ACT GCT TAC TGG CTT ATC GAA pCDNA3as GCC GCC AGT GTG ATG GAT ATC TGC Raf-RBDs C AAG GTG AGG GGC CTG CAA CAA GAG Raf as CT TGG TCT CCT GTA CAT GCA G
Oligonukleotide für Mutationen in RalGDS: RGS(16A)s TGC TGT ATC ATC GCC GTC AGC CTG GAC RGS(16A)as GTC CAG GCT GAC GGC GAT GAT ACA GCA RGS(18K)s TGT ATC ATC CGC GTC AAA CTG GAC GTG GAC RGS(18K)as GTC CAC GTC CAG TTT GAC GCG GAT GAT ACA RGS(19K)s ATC ATC CGC GTC AGC AAG GAC GTG GAC AAT RGS(19K)as ATT GTC CAC GTC CTT GCT GAC GCG GAT GAT RGS(20K)s ATC CGC GTC AGC CTG AAA GTG GAC AAT GGC RGS(20K)as GCC ATT GTC CAC TTT CAG GCT GAC GCG GAT RGS(23K)s CTG GAC GTG GAC AAA GGC AAC ATG TAC AAG RGS(23K)as CTT GTA CAT GTT GCC TTT GTC CAC GTC CAG RGS(26K)s GAC AAT GGC AAC AAG TAC AAG AGC ATC CTG RGS(26K)as CAG GAT GCT CTT GTA CTT GTT GCC ATT GTC RGS(47K)s CGC AAG GCC ATG AAG AAA CAC AAC CTG RGS(47K)as CAG GTT GTG TTT CTT CAT GGC CTT GCG RGS(47K/50K)s GCC ATG AAG AAA CAC AAA CTG GAG GAG RGS(47K/50K)as CTC CTC CAG TTT GTG TTT CTT CAT GGC RGS(48A)s CGC AAG GCC ATG GAC GCA CAC AAC CTG GAG GAG RGS(48A)as CTC CTC CAG GTT GTG TGC GTC CAT GGC CTT GCG RGS(51K)s GAC AAA CAC AAC AAG GAG GAG GAG GAG CCG RGS(51K)as CGG CTC CTC CTC CTC CTT GTT GTG TTT GTC RGS(54K)s CAC AAC CTG GAG GAG AAG GAG CCG GAG GAC RGS(54K)as GTC CTC CGG CTC CTT CTC CTC CAG GTT GTG RGS(57K)s GAG GAG GAG CCG AAG GAC TAT GAG CTG RGS(57K)as CAG CTC ATA GTC CTT CGG CTC CTC CTC RGS(88K)s AAC TCT ACC GCC AAG TAT GAC TTT GTC CTC RGS(88K)as GAG GAC AAA GTC ATA CTT GGC GGT AGA GTT RGS(89K)s TCT ACC GCC AAC AAA GAC TTT GTC CTC AAG RGS(89K)as CTT GAG GAC AAA GTC TTT GTT GGC GGT AGA RGS(90K)s TCT ACC GCC AAC TAT AAA TTT GTC CTC AAG AAG RGS(90K)as CTT CTT GTG GAC AAA TTT ATA GTT GGC GGT AGA
Oligonukleotide für Mutationen in Raf: Raf(56K)s TCT AAG ACA AGC AAG ACT ATC ATC CGT GTT TTC Raf(56K)as GAA AAC ACG GAT AGT GTT GCT TGT CTT AGA Raf(57K)s ACA AGC AAC AAG ATC CGT GTT TTC TTG CCG Raf(57K)as CGG CAA GAA AAC ACG GAT CTT GTT GCT TGT Raf(68K)s AAC AAG CAA AGA AAA GTG GTC AAT GTG CGA Raf(68K)as TCG CAC ATT GAC CAC TTT TCT TTG CTT GTT Raf(69K)s AAC AAG CAA AGA ACA AAG GTC AAT GTG CGA Raf(69K)as TCG CAC ATT GAC CTT TGT TCT TTG CTT GAA Raf(80K)s AAT GGA ATG AGC TTG CAT AAG TGC CTT ATG Raf(80K)as CAT AAG GCA CTT ATG CAA GCT CAT TCC ATT Raf(83K)s G CA T GAC TGC CT T AAG AAA GCA CTC AAG GTG Raf(83K)as CAC CTT GAG TGC TTT CTT AAG GCA GTC ATG C Raf(83D)s G CAT GAC TGC CTT GAC AAA GCA CTC AAG GTG Raf(83D)as CAC CTT GAG TGC TTT GTC AAG GCA GTC ATG C Raf(84D)s TGC CTT ATG GAC GCA CTC AAG GTG AGG Raf(84D)as CCT CAC CTT GAG TGC GTC CAT AAG GCA

ANHANG
146
Raf(85K)s TGC CTT ATG AAA AAA CTC AAG GTG AGG GGC Raf(85K)as GCC CCT CAC CTT GAG TTT TTT CAT AAG GCA Raf(84D/85K)s TGC CTT ATG GAC AAA CTC AAG GTG AGG GGC Raf(84D/85K)as GCC CCT CAC CTT GAG TTT GTC CAT AAG GCA Raf(87A)s ATG AAA GCA CTC GCG GTG AGG GGC CTG Raf(87A)as CAG GCC CCT CAC CGC GAG TGC TTT CAT Raf(125K)s TCT TTG ATT GGA GAA AAA CTT CAA GTA GAT Raf(125K)as ATC TAC TTG AAG TTT TTC TCC AAT CAA AGA
Oligonukleotide für Mutationen in Ras: Ras(31K)s C CAT TTT GTC GAC AAA TAC GAC CCC Ras(31K)as GGG GTC GTA TTT GTC GAC AAA ATG G Ras(35A)s GAA TAC GAC CCC GCC ATA GAG GAT TC Ras(35A)as GA ATC CTC TAT GGC GGG GTC GTA TTC Ras(35S)s GAC GAA TAC GAC CCC TCT ATA GAG GAT TCC Ras(35S)as GGA ATC CTC TAT AGA GGG GTC GTA TTC GTC
6.3 Klonierungen Im folgenden sind für alle Mutagenesen die jeweiligen verwendeten Template-Vektoren und Oligonukleotid-Paare gelistet (T = Template-DNA, O = Oligonukleotide)
Mutationen in pGEX2T:RalGDS-RBD: 1) pGEX2T RalGDS-RBD S18K (T: pGEX2T RalGDS-RBD WT, O: RGS(18K)s und RGS(18K)as) 2) pGEX2T RalGDS-RBD S18K,Y89K (T: pGEX2T RalGDS-RBD S18K, O: RGS(89K)s und RGS(89K)as) 3) pGEX2T RalGDS-RBD L19K (T: pGEX2T RalGDS-RBD WT, O: RGS(19K)s und RGS(19K)as) 4) pGEX2T RalGDS-RBD D20K (T: pGEX2T RalGDS-RBD WT, O: RGS(20K)s und RGS(20K)as) 5) pGEX2T RalGDS-RBD N23K (T: pGEX2T RalGDS-RBD WT, O: RGS(23K)s und RGS(23K)as) 6) pGEX2T RalGDS-RBD N23K,N88K (T: pGEX2T RalGDS-RBD N23K, O: RGS(88K)s und
RGS(88K)as) 7) pGEX2T RalGDS-RBD M26K (T: pGEX2T RalGDS-RBD WT, O:RGS(26K)s und RGS(26K)as) 8) pGEX2T RalGDS-RBD M26K,E54K (T: pGEX2T RalGDS-RBD M26K, O: RGS(54K)s und RGS(54K)as) 9) pGEX2T RalGDS-RBD M26K,D47K,E54K (T: pGEX2T RalGDS-RBD M26K,E54K O: RGS(264754)s
und RGS(264754)as) 10) pGEX2T RalGDS-RBD M26K,D90K (T: pGEX2T RalGDS-RBD D90K, O:RGS(26K)s und RGS(26K)as) 11) pGEX2T RalGDS-RBD M26K,K48A,D90K (T: pGEX2T RalGDS-RBD M26K,D90K, O:RGS(48A)s und RGS(48a)as) 12) pGEX2T RalGDS-RBD M26K,D47K,E54K,D90K (T: pGEX2T RalGDS-RBD M26K,E54K,D90K O: RGS(264754)s und RGS(264754)as) 13) pGEX2T RalGDS-RBD D47K,E52K,E53K (T: pGEX2T RalGDS-RBD D47K, O: RGS(52K/53K)s und RGS(52K/53K)as) 14) pGEX2T RalGDS-RBD L51K (T: pGEX2T RalGDS-RBD WT, O: RGS(51K)s und RGS(51K)as) 15) pGEX2T RalGDS-RBD E54K (T: pGEX2T RalGDS-RBD WT, O: RGS(54K)s und RGS(54K)as) 16) pGEX2T RalGDS-RBD E54K,D90K (T: pGEX2T RalGDS-RBD E54K, O: RGS(90K)s und RGS(90K)as) 17) pGEX2T RalGDS-RBD N88K (T: pGEX2T RalGDS-RBD WT, O: RGS(88K)s und RGS(88K)as) Mutationen in pGEX2T:Raf-RBD: 18) pGEX2T Raf-RBD N56K (T: pGEX2T Raf-RBD WT, O: Raf(56K)s und Raf(56K)as) 19) pGEX2T Raf-RBD N56K,A85K (T: pGEX2T Raf-RBD A85K, O: Raf(56K)s und Raf(56K)as) 20) pGEX2T Raf-RBD N56K,D80K,A85K (T: pGEX2T Raf-RBD D80K,A85K, O: Raf(56K)s und Raf(56K)as) 21) pGEX2T Raf-RBD N56K,D80K,A85K,E125K (T: pGEX2T Raf-RBD N56K,D80K,A85K, O: Raf(125K)s und Raf(125K)as) 22) pGEX2T Raf-RBD T57K (T: pGEX2T Raf-RBD WT, O: Raf(57K)s und Raf(57K)as)

ANHANG
147
23) pGEX2T Raf-RBD T57K,A85K,E125K (T: pGEX2T Raf-RBD A85K,E125K, O: Raf(57K)s und Raf(57K)as) 24) pGEX2T Raf-RBD T68K (T: pGEX2T Raf-RBD WT, O: Raf(68K)s und Raf(68K)as) 25) pGEX2T Raf-RBD V69K (T: pGEX2T Raf-RBD WT, O: Raf(69K)s und Raf(69K)as) 26) pGEX2T Raf-RBD D80K (T: pGEX2T Raf-RBD WT, O: Raf(80K)s und Raf(80K)as) 27) pGEX2T Raf-RBD D80K,K84D (T: pGEX2T Raf-RBD D80K, O: Raf(84D)s und Raf(84D)as) 28) pGEX2T Raf-RBD D80K,A85K (T: pGEX2T Raf-RBD D80K, O: Raf(85K)s und Raf(85K)as) 29) pGEX2T Raf-RBD D80K,K84D,A85K (T: pGEX2T Raf-RBD K84D,A85K, O: Raf(80K)s und Raf(80K)as) 30) pGEX2T Raf-RBD K84D,A85K (T: pGEX2T Raf-RBD K84D, O: Raf(84D/85K)s und Raf(84D/85K)as) 31) pGEX2T Raf-RBD A85K (T: pGEX2T Raf-RBD WT, O: Raf(85K)s und Raf(85K)as) 31) pGEX2T Raf-RBD A85K,E125K (T: pGEX2T Raf-RBD A85K, O: Raf(125K)s und Raf(125K)as) 32) pGEX2T Raf-RBD K87A (T: pGEX2T Raf-RBD WT, O: Raf(87A)s und Raf(87A)as) 33) pGEX2T Raf-RBD E125K (T: pGEX2T Raf-RBD WT, O: Raf(125K)s und Raf(125K)as)
Mutationen in pCDNA3:Raf f.l. 34) pCDNA3 Raf fl. N56K (T: pCDNA3 Raf fl. WT, O: Raf(56K)s und Raf(56K)as) 35) pCDNA3 Raf fl. N56K, A85K (T: pCDNA3 Raf fl. A85K, O: Raf(56K)s und Raf(56K)as) 36) pCDNA3 Raf fl. N56K,D80K,A85K (T: pCDNA3 Raf fl. N56K,A85K, O: Raf(80K)s und Raf(80K)as) 37) pCDNA3 Raf fl. N56K,D80K,A85K,E125K (T: pCDNA3 Raf fl. D80K, A85K, E125K, O: Raf(56K)s und Raf(56K)as) 38) pCDNA3 Raf fl. T57K (T: pCDNA3 Raf fl. WT, O: Raf(57K)s und Raf(57K)as) 39) pCDNA3 Raf fl. T57K,A85K,E125K (T: pCDNA3 Raf fl. A85K, E125K, O: Raf(57K)s und Raf(57K)as) 40) pCDNA3 Raf fl. T68K (T: pCDNA3 Raf fl. WT, O: Raf(68K)s und Raf(68K)as) 41) pCDNA3 Raf fl. V69K (T: pCDNA3 Raf fl. WT, O: Raf(69K)s und Raf(69K)as) 42) pCDNA3 Raf fl. D80K (T: pCDNA3 Raf fl. WT, O: Raf(80K)s und Raf(80K)as) 43) pCDNA3 Raf fl. D80K,A85K (T: pCDNA3 Raf fl. A85K, O: Raf(80K)s und Raf(80K)as) 44) pCDNA3 Raf fl. D80K,A85K,E125K (T: pCDNA3 Raf fl. A85K, E125K O: Raf(80K)s und Raf(80K)as) 45) pCDNA3 Raf fl. D80K,K84D,A85K (T: pCDNA3 Raf fl. K84D, A85K, O: Raf(80K)s und Raf(80K)as) 46) pCDNA3 Raf fl. D80K,K84D,A85K,E125K (T: pCDNA3 Raf fl. D80K,K84D,A85K, O: Raf(125K)s und Raf(125K)as) 47) pCDNA3 Raf fl. M83K (T: pCDNA3 Raf fl. WT, O: Raf(83K)s und Raf(83K)as) 48) pCDNA3 Raf fl. M83D (T: pCDNA3 Raf fl. WT, O: Raf(83D)s und Raf(83D)as) 49) pCDNA3 Raf fl. K84D,A85K (T: pCDNA3 Raf fl. K84D, O: Raf(84D/85K)s und Raf(84D/85K)as) 50) pCDNA3 Raf fl. K84D (T: pCDNA3 Raf fl. WT, O: Raf(84D/)s und Raf(84D)as) 51) pCDNA3 Raf fl. A85K (T: pCDNA3 Raf fl. WT, O: Raf(85K)s und Raf(85K)as) 52) pCDNA3 Raf fl. A85K,E125K (T: pCDNA3 Raf fl. A85K, O: Raf(125K)s und Raf(125K)as) 53) pCDNA3 Raf fl. K87A (T: pCDNA3 Raf fl. WT, O: Raf(87A)s und Raf(87A)as) 54) pCDNA3 Raf fl. E125K (T: pCDNA3 Raf fl. WT, O: Raf(125K)s und Raf(125K)as)
Mutationen in pSVK3 Ras f.l. 55) pSVK3 Ras fl. G12V, E31K (T: pSVK3 Ras fl. G12V, O: Ras(31K)s und Raf(31K)as) 56) pSVK3 Ras fl. G12V, T35A (T: pSVK3 Ras fl. G12V, O: Ras(35A)s und Raf(35A)as) 57) pSVK3 Ras fl. G12V, T35S (T: pSVK3 Ras fl. G12V, O: Ras(35S)s und Raf(35S)as)
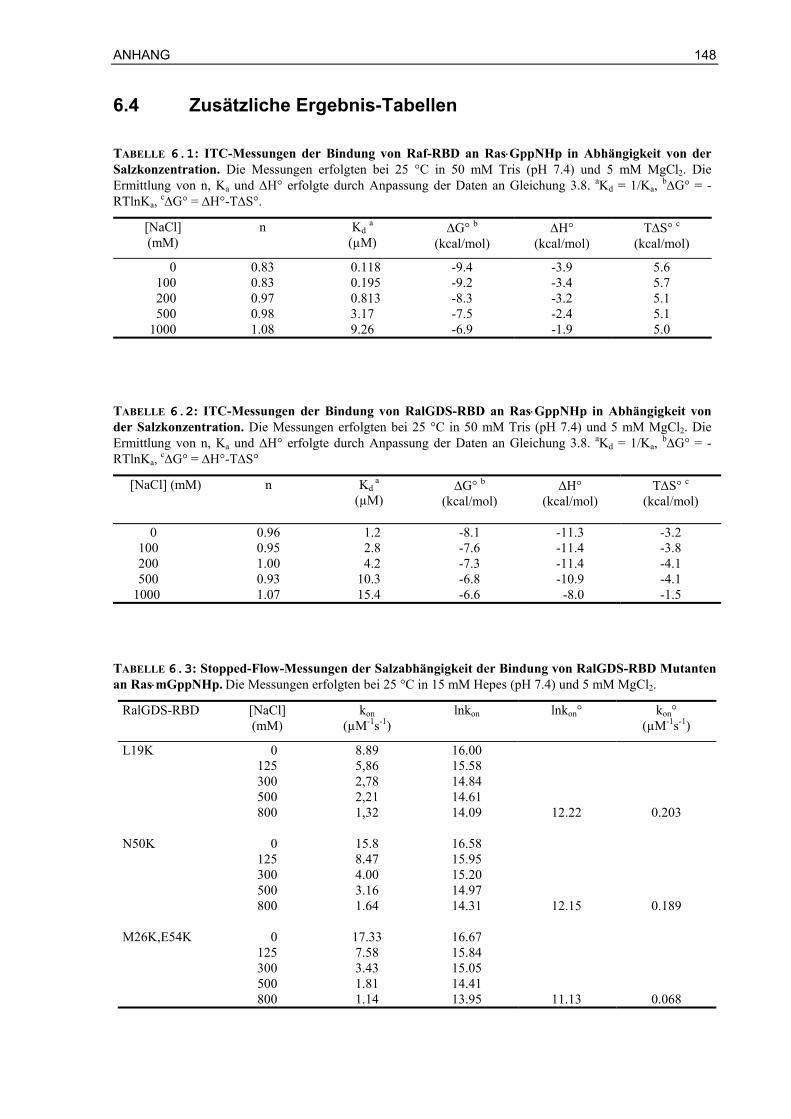
ANHANG
148
6.4 Zusätzliche Ergebnis-Tabellen TABELLE 6.1: ITC-Messungen der Bindung von Raf-RBD an Ras⋅GppNHp in Abhängigkeit von der Salzkonzentration. Die Messungen erfolgten bei 25 °C in 50 mM Tris (pH 7.4) und 5 mM MgCl2. Die Ermittlung von n, Ka und ∆H° erfolgte durch Anpassung der Daten an Gleichung 3.8. aKd = 1/Ka, b∆G° = -RTlnKa, c∆G° = ∆H°-T∆S°.
[NaCl] (mM)
n Kd a (µM)
∆G° b (kcal/mol)
∆H° (kcal/mol)
T∆S° c (kcal/mol)
0 0.83 0.118 -9.4 -3.9 5.6 100 0.83 0.195 -9.2 -3.4 5.7 200 0.97 0.813 -8.3 -3.2 5.1 500 0.98 3.17 -7.5 -2.4 5.1 1000 1.08 9.26 -6.9 -1.9 5.0
TABELLE 6.2: ITC-Messungen der Bindung von RalGDS-RBD an Ras⋅GppNHp in Abhängigkeit von der Salzkonzentration. Die Messungen erfolgten bei 25 °C in 50 mM Tris (pH 7.4) und 5 mM MgCl2. Die Ermittlung von n, Ka und ∆H° erfolgte durch Anpassung der Daten an Gleichung 3.8. aKd = 1/Ka, b∆G° = -RTlnKa, c∆G° = ∆H°-T∆S°
[NaCl] (mM) n Kd a
(µM) ∆G° b
(kcal/mol) ∆H°
(kcal/mol) T∆S° c
(kcal/mol)
0 0.96 1.2 -8.1 -11.3 -3.2 100 0.95 2.8 -7.6 -11.4 -3.8 200 1.00 4.2 -7.3 -11.4 -4.1 500 0.93 10.3 -6.8 -10.9 -4.1 1000 1.07 15.4 -6.6 -8.0 -1.5
TABELLE 6.3: Stopped-Flow-Messungen der Salzabhängigkeit der Bindung von RalGDS-RBD Mutanten an Ras⋅mGppNHp. Die Messungen erfolgten bei 25 °C in 15 mM Hepes (pH 7.4) und 5 mM MgCl2.
RalGDS-RBD [NaCl] (mM)
kon (µM-1s-1)
lnkon lnkon° kon° (µM-1s-1)
L19K 0 8.89 16.00 125 5,86 15.58 300 2,78 14.84 500 2,21 14.61 800 1,32 14.09 12.22 0.203 N50K 0 15.8 16.58 125 8.47 15.95 300 4.00 15.20 500 3.16 14.97 800 1.64 14.31 12.15 0.189 M26K,E54K 0 17.33 16.67 125 7.58 15.84 300 3.43 15.05 500 1.81 14.41 800 1.14 13.95 11.13 0.068
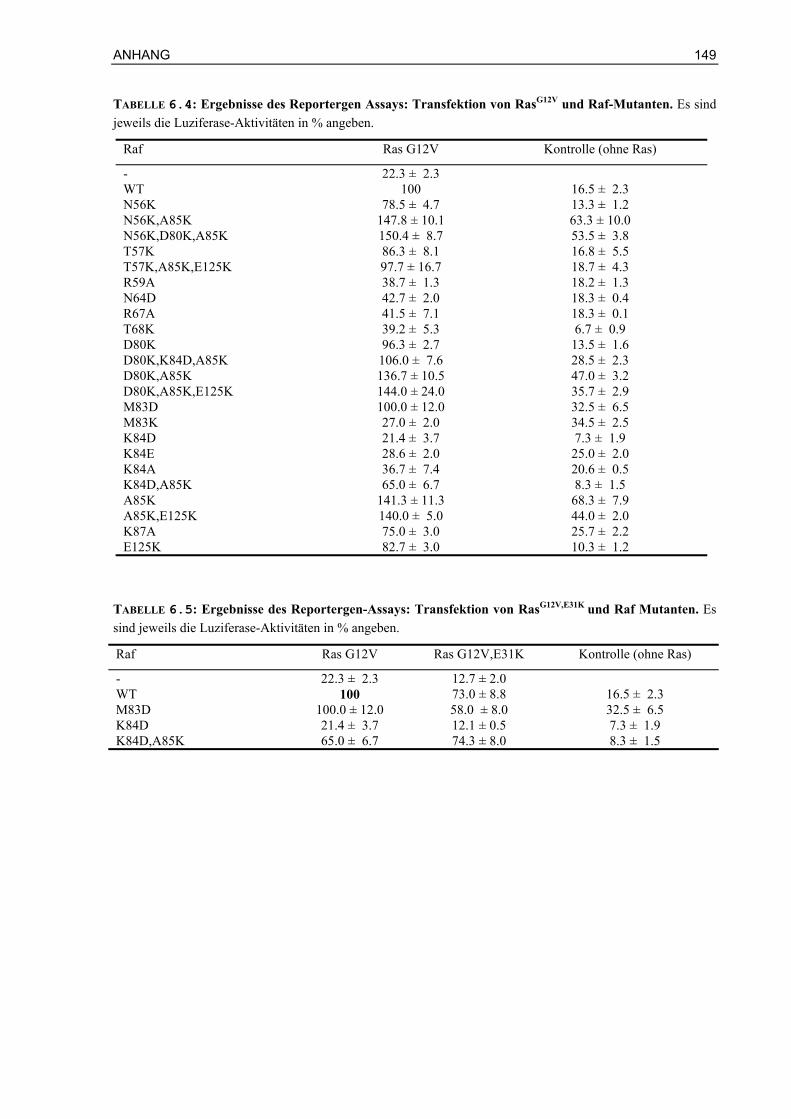
ANHANG
149
TABELLE 6.4: Ergebnisse des Reportergen Assays: Transfektion von RasG12V und Raf-Mutanten. Es sind jeweils die Luziferase-Aktivitäten in % angeben.
Raf Ras G12V Kontrolle (ohne Ras)
- 22.3 ± 2.3 WT 100 16.5 ± 2.3 N56K 78.5 ± 4.7 13.3 ± 1.2 N56K,A85K 147.8 ± 10.1 63.3 ± 10.0 N56K,D80K,A85K 150.4 ± 8.7 53.5 ± 3.8 T57K 86.3 ± 8.1 16.8 ± 5.5 T57K,A85K,E125K 97.7 ± 16.7 18.7 ± 4.3 R59A 38.7 ± 1.3 18.2 ± 1.3 N64D 42.7 ± 2.0 18.3 ± 0.4 R67A 41.5 ± 7.1 18.3 ± 0.1 T68K 39.2 ± 5.3 6.7 ± 0.9 D80K 96.3 ± 2.7 13.5 ± 1.6 D80K,K84D,A85K 106.0 ± 7.6 28.5 ± 2.3 D80K,A85K 136.7 ± 10.5 47.0 ± 3.2 D80K,A85K,E125K 144.0 ± 24.0 35.7 ± 2.9 M83D 100.0 ± 12.0 32.5 ± 6.5 M83K 27.0 ± 2.0 34.5 ± 2.5 K84D 21.4 ± 3.7 7.3 ± 1.9 K84E 28.6 ± 2.0 25.0 ± 2.0 K84A 36.7 ± 7.4 20.6 ± 0.5 K84D,A85K 65.0 ± 6.7 8.3 ± 1.5 A85K 141.3 ± 11.3 68.3 ± 7.9 A85K,E125K 140.0 ± 5.0 44.0 ± 2.0 K87A 75.0 ± 3.0 25.7 ± 2.2 E125K 82.7 ± 3.0 10.3 ± 1.2
TABELLE 6.5: Ergebnisse des Reportergen-Assays: Transfektion von RasG12V,E31K und Raf Mutanten. Es sind jeweils die Luziferase-Aktivitäten in % angeben.
Raf Ras G12V Ras G12V,E31K Kontrolle (ohne Ras)
- 22.3 ± 2.3 12.7 ± 2.0 WT 100 73.0 ± 8.8 16.5 ± 2.3 M83D 100.0 ± 12.0 58.0 ± 8.0 32.5 ± 6.5 K84D 21.4 ± 3.7 12.1 ± 0.5 7.3 ± 1.9 K84D,A85K 65.0 ± 6.7 74.3 ± 8.0 8.3 ± 1.5
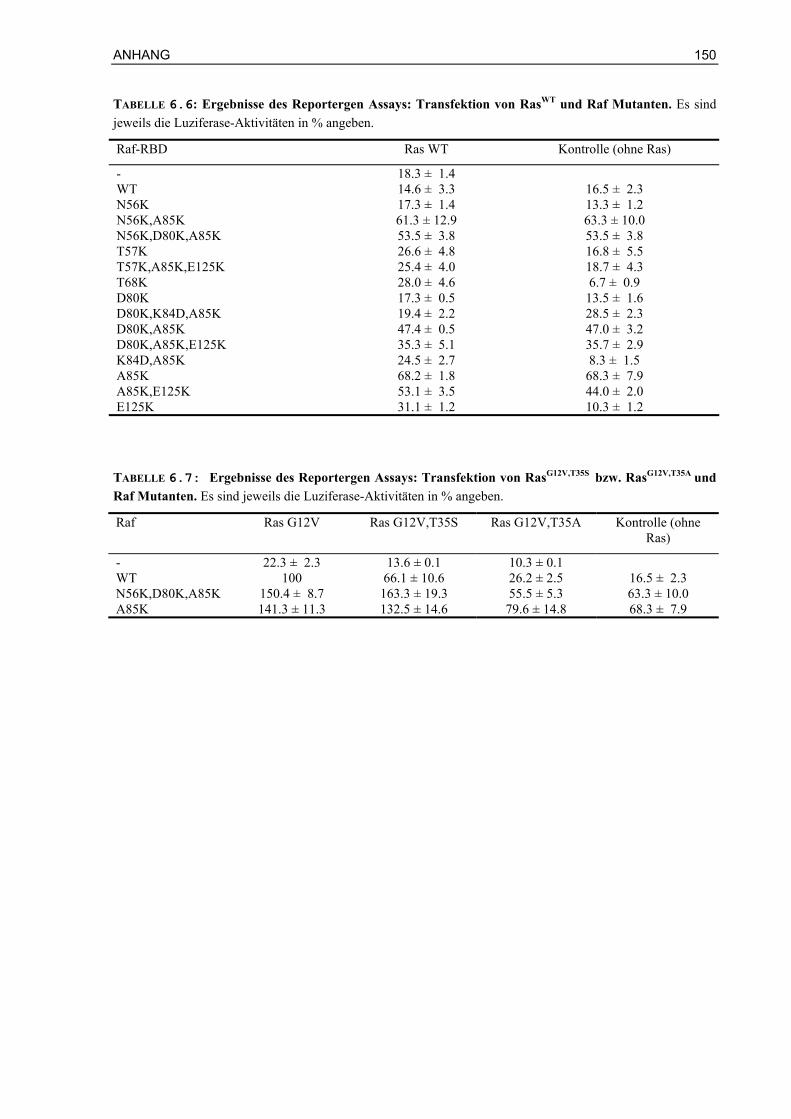
ANHANG
150
TABELLE 6.6: Ergebnisse des Reportergen Assays: Transfektion von RasWT und Raf Mutanten. Es sind jeweils die Luziferase-Aktivitäten in % angeben.
Raf-RBD Ras WT Kontrolle (ohne Ras)
- 18.3 ± 1.4 WT 14.6 ± 3.3 16.5 ± 2.3 N56K 17.3 ± 1.4 13.3 ± 1.2 N56K,A85K 61.3 ± 12.9 63.3 ± 10.0 N56K,D80K,A85K 53.5 ± 3.8 53.5 ± 3.8 T57K 26.6 ± 4.8 16.8 ± 5.5 T57K,A85K,E125K 25.4 ± 4.0 18.7 ± 4.3 T68K 28.0 ± 4.6 6.7 ± 0.9 D80K 17.3 ± 0.5 13.5 ± 1.6 D80K,K84D,A85K 19.4 ± 2.2 28.5 ± 2.3 D80K,A85K 47.4 ± 0.5 47.0 ± 3.2 D80K,A85K,E125K 35.3 ± 5.1 35.7 ± 2.9 K84D,A85K 24.5 ± 2.7 8.3 ± 1.5 A85K 68.2 ± 1.8 68.3 ± 7.9 A85K,E125K 53.1 ± 3.5 44.0 ± 2.0 E125K 31.1 ± 1.2 10.3 ± 1.2
TABELLE 6.7: Ergebnisse des Reportergen Assays: Transfektion von RasG12V,T35S bzw. RasG12V,T35A und Raf Mutanten. Es sind jeweils die Luziferase-Aktivitäten in % angeben.
Raf Ras G12V Ras G12V,T35S Ras G12V,T35A Kontrolle (ohne Ras)
- 22.3 ± 2.3 13.6 ± 0.1 10.3 ± 0.1 WT 100 66.1 ± 10.6 26.2 ± 2.5 16.5 ± 2.3 N56K,D80K,A85K 150.4 ± 8.7 163.3 ± 19.3 55.5 ± 5.3 63.3 ± 10.0 A85K 141.3 ± 11.3 132.5 ± 14.6 79.6 ± 14.8 68.3 ± 7.9

ANHANG
151
6.5 Lebenslauf
Christina Kiel Bahnhofstraße 92 32219 Rödinghausen Geburtsdatum 25.11.1973 Geburtsort Bünde Familienstand ledig Staatsangehörigkeit deutsch 1980-1984 Grundschule Bieren 1984-1993 Freiherr-vom-Stein-Gymnasium in Bünde 16.06.1993 Allgemeine Hochschulreife 1993-1999 Studium der Biochemie an der Ruhr-Universität Bochum 30.9.1996 Vordiplom im Fach Biochemie 06.1999-12.1999 Diplomarbeit am Max-Planck-Institut für molekulare
Physiologie, Abteilung Strukturelle Biologie, Titel: Thermodynamische Analyse der Interaktion von H-Ras
und Rap1A mit RalGDS 22.12.1999 Abschluss: Diplom Biochemikerin seit 01.01.2000 Studium der Chemie an der Ruhr-Universität Bochum
01.2000 – 03.2002 Doktorarbeit am Max-Planck-Institut für molekulare Physiologie, Abteilung Strukturelle Biologie

ANHANG
152
6.6 Vorträge und Konferenzen
Vorträge:
18. Rabenstein Kolleg (22.6.-24.6.2000) in Pottenstein. „Detaillierte Analyse der Interaktion von H-Ras und Rap1A mit RalGDS“.
NATO/FEBS Advanced Study Institute International Summerschool: Protein Modules in cellular Signalling (13.9.-22.9.2000) in St. Martin de Londres France. “Analysis of the H-Ras-RalGDS/Raf Interface by Double Mutant Cycles”.
19. Rabenstein Kolleg (14.6.-16.6.2001) in Weißenstadt. “Kinetische Kontrolle der Signalweiterleitung über Ras-Proteine”.
Einladung von Prof. Rob. C. Aalberse, Sanquin Research at CLB, Department of Immunopathology, Amsterdam (6.5.2002). “Quantification of Ras/effector interactions: Thermodynamic and kinetic investigations”.
20. Rabenstein Kolleg (30.5.-1.6.2002) in Pottenstein. “Thermodynamische, strukturelle und zelluläre Untersuchung von Ras/Effektor-Komplexen mit veränderten kinetischen Eigenschaften“.
Targets for Cancer Therapy consortium meeting (27.6.-28.6.2002) am CNIO, in Madrid. “Quantification of Ras/Effector interactions and Assays for Drug-Screening”.
Graduate School of Chemistry and Biochemistry (4.7.-5.7. 2002) in Bielefeld. “Energy landscapes of proteins: Thermodynamic and kinetic properties of Ras/Effector interfaces”.
17th IUPAC Conference on Chemical Thermodynamics (28.7.-2.8.2002) in Rostock “Energy landscapes of proteins: Thermodynamic and kinetic investigations of Ras/Effector interfaces”
Teilnahme an Konferenzen:
2nd International Conference on Applications of Biocalorimetry (28.3.-31.3.1999) in Halle/S., Germany.
17. Rabenstein Kolleg (3.6.-5.6.1999) in Pottenstein.
SFB394-Symposium (10.-11.12.1999) in Hamminkeln-Dingden.
51. Mosbacher Kolloquium (3.4.-5.4.2000) in Mosbach, Baden. GTP-Binding Proteins: Central Regulators in Cell Biology. Poster: „Analysis of the H-Ras-RalGDS/Raf Interface by

ANHANG
153
Double Mutant Cycles” (Christina Kiel, Daniel Schwarz, Alfred Wittinghofer & Christian Herrmann).
Workshop: Fundamentals of BioCalorimetry (27.10.2000) in Frankfurt.
Second International Symposium on Structures and Mechanism in Biological Signal and Energy Transduction (23.11.-25.11.2000), Dabringhausen, Germany. Poster: “Thermodynamic, Kinetik and Structural Analysis of Ras Proteins and their Effektors” (Christina Kiel., Daniel Schwarz, Sabine Wolgemuth, Mathias Geyer, Hans Robert Kalbitzer, Alfred Wittinghofer & Christian Herrmann).
Biocalorimetry 2001 Conference: Current Trends in Microcalorimetry (26.7.-28.7.2001) in Philadelphia, USA. Poster: “Thermodynamic, Kinetic and Structural Analysis of Ras Proteins and their Effectors” (Christina Kiel, Gideon Schreiber, Sabine Wohlgemuth & Christian Herrmann).
Herbsttagung der Gesellschaft für Biochemie und Molekularbiologie (9.9.-12.9.2001) in Bochum. Poster: “Thermodynamic, Kinetic and Structural Analysis of Ras Proteins and their Effectors” (Christina Kiel, Gideon Schreiber, Sabine Wohlgemuth & Christian Herrmann).

LITERATUR
154
VII LITERATUR Ahmadian, M. R., Zor, T., Vogt, D., Kabsch, W., Selinger, Z., Wittinghofer, A. & Scheffzek, K. (1999)
Guanosine triphosphatase stimulation of oncogenic Ras mutants. Proc. Natl. Acad. Sci. USA 96, 7065-7070.
Aloy, P., Querol, E., Aviles, F. X. & Sternberg, M. J. E. (2001) Automated structure-based prediction of
functional sites in proteins: applications to assessing the validity of inheriting protein function from homology in genome annotation and to protein docking. J. Mol. Biol. 311, 395-408.
Albright, C. F., Giddings, B. W., Liu, J., Vito, M. & Weinberg, R. A. (1993) Characterization of a guanine
nucleotide dissociation stimulator for a ras-related GTPase. EMBO J. 12, 339-347. Avruch, J., Zhang, X. F. & Kyriakis, J. M. (1994) Raf meets Ras: completing the framework of a signal
transduction pathway. Trends Biochem. Sci. 19, 279-283. Baerga-Ortiz, A., Rezaie, A. R. & Komives, E. A. (2000) Electrostatic dependence of the thrombin-
thrombomudulin interaction. J. Mol. Biol. 296, 651-658. Baesley, J. R., Doyle, D. F., Chen, L., Cohen, D. S., Fine, B. R. & Pielak, G. J. (2002) Searching for
quantitative entropy-enthalpy compensation among protein variants. Proteins 49, 398-402. Barbacid, M. (1987) Ras genes. Annu. Rev. Biochem. 56, 779-827. Barnard, D., Diaz, B., Hettich, L., Chuang, E., Zhang, X., Avruch, J. & Marshall, M. (1995) Identification
of the sites of interaction between c-Raf-1 and Ras-GTP. Oncogene 10, 1283-1290. Berg, O. G. & von Hippel, P. H. (1985) Diffusion-controlled macromolecular interactions. Ann. Rev. Biophys.
Biophys. Chem. 14, 131-160. Bilan, P. J., Mayers, J. S. & Kahn, C. R. (1998) The ras-related protein rad associates with the cytoskeleton in
a non-lipid-dependent manner. Exp. Cell Res. 242, 391-400. Block, C., Janknecht, R., Herrmann, C., Nassar, N. & Wittinghofer, A. (1996) Quantitative structure-activity
analysis correlating Ras/Raf interaction in vitro to Raf activation in vivo. Nature Struct. Biol. 3, 255-251.
Boettner, B., Govek, E. E., Cross, J. & Van Aelst, L. (2000) The junctional multidomain protein AF-6 is a
binding partner of the Rap1A GTPase and associates with the actin cytoskeletal regulator profilin. Proc. Natl. Acad. Sci. USA 97, 9064-9069.
Bogan, A. A. & Thorn, K. S. (1998) Anatomy of Hot Spots in Protein Interfaces. J. Mol. Biol., 280, 1-9. Boguski, M. S. & McCormick, F. (1993) Proteins regulating ras and its relatives. Nature 366, 643-654. Bourne, H. R., Sanders, D. A. & McCormick, F. (1990) The GTPase superfamily: a conserved switch for
diverse cell functions. Nature 348, 125-132.

LITERATUR
155
Bourne, H. R., Sanders, D. A. & McCormick, F. (1991) The GTPase superfamily: conserved structure and molecular mechanism. Nature 348, 117-126.
Bos, J. L. (1989) Ras oncogenes in human cancer: a review. Cancer Res. 49, 4682-4689. Bos, J. L. (1998) All in the family? New insights and questions regarding interconnectivity of Ras, Rap and Ral.
EMBO J. 17, 6776-6782. Botti, S., A., Felder, C. E., Sussman, J. L. & Silman, I. (1998) Electrotactins: a class of adhesion proteins with
conserved electrostatic and structural motifs. Protein Eng. 11, 415-420. Boussiotis, V. A., Freeman, G. J., Berezovskaya, A., Barber, D. L. & Nadler, L. M. (1997) Maintenance of
human T cell anergy: blocking of IL-2 gene transcription by activated Rap1. Science 278, 124-128. Bradford, M. M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein
utilizing the principle of protein dye binding. Analyt. Biochem. 72, 248-253. Camacho, C. J., Weng, Z., Vajda, S. & DeLisi, C. (1999) Free energy landscapes of encounter complexes in
protein-protein association. Biophys. J. 76, 1166-1178. Camacho, C. J., Kimura, S. R., DeLisi, C. & Vajda, S. (2000) Kinetics of desolvation mediated protein-
protein binding. Biophys. J. 78, 1094-1105. Carter, P. J., Winter, G., Wilkinson, A. J. & Fersht, A. R. (1984) The use of double mutants to detect
structural changes in the active site of the tyrosyl-tRNA synthetase (Bacillus Stearothermophilus). Cell 38, 835-840.
Casari, G., Sander, C. & Valencia, A. (1995) A method to predict functional residues in proteins. Nat. Struct.
Biol. 2, 171-178. Castro, M. J. & Anderson, S. (1996) Alanine point-mutations in the reactive region of bovine pancreatic
trypsin inhibitor: effects on the kinetics and thermodynamics of binding to beta-trypsin and alpha-chymotrypsin. Biochemistry 35, 11435-11446.
Chuang, C. T., Niemela, S. L. & Miller, R. H. (1989) One-Step preparation of competent Escherichia coli:
transformation and storage of bacterial cells in the same solution. Proc. Natl. Acad. Sci. USA 89, 5789-5793.
Clackson, T., Ultsch, M. H., Wells, J. A. & de Vos, A. M. (1998) Structural and functional analysis of the 1:1
growth hormone:receptor complex reveals the molecular basis for receptor affinity. J. Mol. Biol. 277, 1111-1128.
Cooper, A. (1999) Thermodynamic analysis of biomolecular interactions. Curr. Opin. Chem. Biol. 3, 557-563. Cooper, A., Johnson, C. M., Lakey, J. H. & Nöllmann, M. (2001) Heat does not come in different colours:
entropy-enthalpy compensation, free energy windows, quantum confinement, pressure perturbation calorimetry, salvation and the multiple causes of heat capacity effects in biomolecular interactions. Biophys. Chem. 93, 215-230.
Cornish-Bowden, A. (2002) Enthalpy-entropy compensation: a phantom phenomenon. J. Biosci. 27, 121-126.

LITERATUR
156
Cox, A. D. & Der, C. J. (2002) Farnesyltransferase inhibitors: promise and realities. Curr. Opin. Pharmacol. 2, 388-393.
Datta, S. R., Brunet, A. & Greenberg, M. E. (1999) Cellular survival: a play in three Akts. Genes Dev. 13,
2905-2927. Daum, G., Eisenmann-Tappe, I., Fries, H.-W., Troppmair, J. & Rapp, U. R. (1994) The ins and outs of Raf
kinases. Trends biochem. Sci. 19, 474-480. DeLano, W. L. (2002) Unraveling hot spots in binding interfaces: progress and challenges. Curr. Opin. Struct.
Biol. 12, 14-20. Downward, J. (2003) Targeting Ras signalling pathways in cancer therapy. Nature Rev. Cancer 3, 11-22. Dunitz, J. P. (1994) The entropic cost of bound water in crystals and biomolecules. Science 264, 670. Dunitz, J. P. (1995) Win some, lose some – enthalpy-entropy compensation in weak intermolecular interactions.
Chem. Biol. 2, 709-712. Elcock, A. H., Gabdoulline, R. R., Wade, R. C. & McCammon, J. A. (1999) Computer simulation of protein-
protein association kinetics: acetylcholinesterase-fasiculin. J. Mol. Biol. 291, 149-162. Emerson, S. D., Madison, V. S., Palermo, R. E., Waugh, D. S., Scheffler, J. E., Tsao, K. L., Kiefer, S. E.,
Liu, S. P. & Fry, D. C. (1995) Solution structure of the Ras-binding domain of c-Raf-1 and identification of its Ras interaction surface. Biochemistry 34, 6911-6918.
Esser, D., Bauer, B., Wolthuis, R. M. F., Wittinghofer, A., Cool, R. H. & Bayer, P. (1998) Structure
determination of the Ras-binding domain of the Ral-specific guanine nucleotide exchange factor Rlf. Biochemistry 37, 13453-13462.
Frisch, C., Schreiber, G., Johnson, C. M. & Fersht, A. R. (1997) Thermodynamics of the interaction of
barnase and barstar: changes in free energy versus changes in enthalpy on mutation. J. Mol. Biol. 267, 696-706.
Frisch, C., Fersht, A. R. & Schreiber, G. (2001) Experimental assignment of the structure of the transition
state of Barnase and Barstar. J. Mol. Biol. 308, 69-77. Fry, M. J. (1994) Structure, regulation and function of phosphoinositide 3-kinase. Biochem. biophys. Acta 1226,
237-268. Gabdoulline, R. R. & Wade, R. C. (1997) Simulation of the diffusional association of barnase and barstar.
Biophys. J. 72, 1917-1927. Gabdoulline, R. R. & Wade, R. C. (2002) Biomolecular diffusional association. Curr. Opin. Struct. Biol. 12,
204-213. Garvey, W. T., Maianu, L., Kennedy, A., Wallace, P., Ganaway, E., Hamacher, L. L., Lenhard, J. M. &
Burns, D. K. (1997) Muscle ras expression and human metabolism: potential role of the novel Ras-related GTPase in expenditure and body composition. Diabetes 46, 444-450.

LITERATUR
157
Geyer, M., Schweins, T., Herrmann, C., Prisner, T., Wittinghofer, A. & Kalbitzer, H. R. (1996) Conformational transitions in p21ras and its complexes with the effector protein Raf-RBD and the GTPase activating protein GAP. Biochemistry 35, 10308-10320.
Geyer, M., Herrmann, C., Wohlgemuth, S., Wittinghofer, A. & Kalbitzer, H. R. (1997) Structure of the Ras
binding domain of RalGEF and implications for Ras binding and signalling. Nature Struct. Biol. 3, 694-699.
Gibson, T. J. (1984) Studies on the Eppstein-Barr virus genome. Ph. D. thesis, University of Cambridge, U.K. Gideon, P., John, J., Frech, M., Lautwein, A., Clark, R., Scheffler, J. E. & Wittinghofer, A. (1992)
Mutational and kinetic analysis of the GTPase-activating protein (GAP)-p21 interaction: The C-terminal domain of GAP is not efficient for full activity. Mol. Cell Biol. 12, 2050-2056.
Glasstone, S., Laidler, K. J. & Eyring, H. (1941) In the theory of rate processes. McGraw-Hill Book Co., New
York and London. Gohlke, H. & Klebe, G. (2002) Approaches to the description and prediction of the binding affinity of small-
molecule ligands to macromolecular receptors. Angew. Chem. Int. Ed. 41, 2644-2676. Gomez, J. & Freire, E. (1995) Thermodynamic mapping of the inhibitor site of the aspartic protease
endothiapepsin. J. Mol. Biol. 252, 337-350. Gomez, J., Hilser, V. J., Xie, D. & Freire, E. (1995) The heat capacity of proteins. Proteins 22, 404-412. Gronwald, W., Huber, F., Grünewald, P., Spörner, M., Wohlgemuth, S., Herrmann, C. & Kalbitzer, H. R.
(2001) Solution structure of the Ras-binding domain of the protein kinase Byr2 from Schizosaccharomyces pombe. Structure 9, 1029-1041.
Guerois, R., Nielsen, J. E. & Serrano, L. (2002) Predicting changes in the stability of proteins and protein
complexes: a study of more than 1000 mutations. J. Mol. Biol. 320, 369-387. Han, L. M. & Colicelli, J. (1995) A human protein selected for interference with Ras functions interacts directly
with Ras and competes with Raf. Mol. Cell. Biol. 15, 1318-1323. Hancock, J. F., Paterson, H., Marshall, C. J. (1990) A polybasic domain or palmitoylation is required in
addition to the CAAX motif to localize p21ras to the plasma membrane. Cell 63, 133-139. Harel, M., Kleywegt, G. J., Ravelli, R. B., Silman, I. & Sussman, J. L. (1995) Crystal structure of an
acetylcholinesterase-fasciculin complex: interaction of a three-fingered toxin from snake venom with its target. Structure 3, 1355-1366.
Hendsch, Z. S. & Tidor, B. (1994) Do salt bridges stabilize proteins? A continuum electrostatic analysis.
Protein Sci. 3, 211-226. Hendsch, Z. & Tidor, B. (1999) Electrostatic interactions in the GCN4 leucine zipper: substantial contributions
arise from intramolecular interactions enhanced on binding. Protein Sci. 8, 1381-1392. Herrmann, C., Martin, G. A. & Wittinghofer, A. (1995) Quantitative analysis of the complex between p21 ras
and the Ras-binding domain of the human Raf-1 protein kinase. J. Biol. Chem. 270, 2901-2905.

LITERATUR
158
Herrmann, C., Horn, G., Spaargaren, M. & Wittinghofer, A. (1996) Differential interaction of the ras family GTP-binding proteins H-Ras, Rap1A and R-Ras with the putative effector molecules raf kinase and ral-guanine nucleotide exchange factor. J. Biol. Chem. 271, 6794-6800.
Herrmann, C. & Nassar, N. (1996) Ras and its effectors. Prog. Biophys. molec. Biol. 66, 1-41. Herrmann, C., Block, C., Geisen, C., Haas, K., Weber, C., Winde, G., Moroy, T. & Müller, O. (1998)
Sulindac sulfide inhibits Ras signaling. Oncogene 17, 1769-1776. Herrmann, C. (2003) Ras-effector interactions: after one decade. Curr. Opin. Struct. Biol. 13, 122-129. Hilser, V. J., Gomez, J. & Freire, E. (1996) The enthalpy change in protein folding and binding: refinement of
parameters for structure-based calculations. Proteins 26, 123-133. Hofer, F., Fields, S., Schneider, C. & Martin, G. S. (1994) Activated Ras interacts with the Ral guanine
nucleotide dissociation stimulator. Proc. Natl. Acad. Sci. USA 91, 11089-11093. Honig, B. H. & Hubbel, W. L. (1984) Stability of `salt bridges´ in membrane proteins. Proc. Natl. Acad. Sci.
USA 81, 54-12-5416. Honig, B. & Nicolls, A. (1995) Classical electrostatics in biology and chemistry. Science 268, 1144-1149. Horovitz, A. (1987) Non-additivity in Protein-Protein Interactions. J. Mol. Biol. 196, 733-735. Huang, L., Weng, X., Hofer, F., Martin, G. S. & Kim, S. H. (1997) Three-dimensional structure of the Ras-
interacting domain of RalGDS. Nature Struct. Biol. 4, 609-615. Huang, L., Weng, X., Hofer, F., Martin, G. S. & Kim, S. H. (1998) Structural basis for the interaction of Ras
with RalGDS. Nature Struct. Biol. 5, 422-426. Huber, F., Gronwald, W., Wohlgemuth, S., Herrmann, C., Geyer, M., Wittinghofer, A. & Kalbitzer, H. R.
(2000) Sequential NMR assignment of the RAS-binding domain of Byr2. J. Biomol. NMR 18, 355-356.
Ito, W., Yasui, H. & Kurosawa, Y. (1995) Mutations in the complementarity-determining regions do not cause
differences in free energy during the process of formation of the activated complex between an antibody and the corresponding protein antigen. J. Mol. Biol. 248, 729-732.
Jaitner, B. K., Becker, J., Linnemann, T., Herrmann, C., Wittinghofer, A. & Block, C. (1997)
Discrimination of amino acids mediating Ras binding from noninteracting residues affecting Raf activation by double mutant analysis. J. Biol. Chem. 272, 29927-29933.
Janin, J. (1997) The Kinetics of Protein-Protein Recognition. Proteins 28, 153-161. John, J., Sohmen, R., Feuerstein, J., Linke, R., Wittinghofer, A. & Goody, R. S. (1990) Kinetics of
nucleotides with nucleotide-free H-ras p21. Biochemistry 29, 6058-6065. Jones, S. & Thornton, J. M. (1996) Principles of protein-protein interactions. Proc. Natl. Acad. Sci. USA 93,
13-20.

LITERATUR
159
Karaguni, I. M., Herter, P., Debruyne, P., Chtarbova, S., Kasprzynski, A., Herbrand, U., Ahmadian, M. R., Glüsenkamp, K. H., Winde, G., Mareel, M., Möröy, T. & Müller, O. (2002) The new Sulindac derivative IND 12 reverses Ras-induced cell transformation. Cancer Res. 62, 1718-1723.
Kelley, G. S., Reks, S. E., Ondrako, J. M. & Smrcka, A. V. (2001) Phospholipase Ce: a novel Ras effector.
EMBO J. 20, 743-754. Khokhlatchev, A., Rabizadeh, S., Xavier, R., Nedwidek, M., Chen, T., Zhong, X. F., Seed, B. & Avruch, J.
(2002) Identification of a novel Ras-regulated proapoptotic pathway. Curr. Biol. 12, 253-265. Kiel, C. (1999) Thermodynamische Analyse der Interaktion von H-Ras und Rap1A mit RalGDS. Diplomarbeit,
Ruhr-Universität Bochum.
Kiel, C. & Herrmann, C. (2001) Double mutant cycle analysis of Ras proteins and their effektors: Quantification of binding specificity. Published in “Protein Modules in Cellular Signaling”, edited by Heilmeyer, L. & Friedrich, P. IOS Press.
Kigawa, T., Endo, M., Ito, Y., Shirouzu, M., Kikuchi, A. & Yokoyama, S. (1998) Solution structure of the
Ras-binding domain of Rgl. FEBS Lett. 441, 413-418. Kikuchi, A., Demo, S. D., Ye, Z. H., Chen, Y. W. & Williams, L. T. (1994) RalGDS family members interact
with the effector loop of ras p21. Mol. Cell. Biol. 14, 7483-7491. Kodaki, T., Woscholski, R., Hallberg, B., Rodriguez-Viciana, P., Downward, J. & Parker, P. J. (1994) The
activation of phosphatidylinositol 3-kinase by Ras. Curr. Biol. 4, 798-806. Kohl, N. E., Omer, C. A., Conner, M. W., Anthony, N. J., Davide, J. P., de Solms, S. J., Guiliani, E. A.,
Gomez, P. R. & Graham, S. L. (1995) Inhibition of farnesyltransferase induces regression of mammary and salivary carcinomas in ras transgenic mice. Nature Med. 1, 792-297.
Kraulis, P. J. (1991) MOLSCRIPT - a program to produce both detailed and schematic plots of protein
structures. J. Appl. Cryst. 24, 946-950. Krug, R., Hunter, W. & Grieger, R. (1976) Statistical interpretation of enthalpy-entropy compensation. Nature
261, 566-567. Kumar, S., Ma, B., Tsai, C. J. & Nussinov, R. (2000) Electrostatic strengths of salt bridges in thermophilic and
mesophilic glutamate dehydrogenase monomers. Proteins: Struct. Funct. Genet. 38, 368-383. Kuriyama, M., Harada, N., Kuroda, S., Yamamoto, T., Nakafuku, M., Iwamatsu, A., Yamamoto, D.,
Prasad, R., Croce, C. & Canaani, E. (1996) Identification of AF-6 and Canoe as putative targets for Ras. J. Biol. Chem. 271, 607-610.
Ladbury, J. E. (1996) Just add water! The effect of water on the specificity of protein-ligand binding sites and
its potential application to drug design. Chemistry & Biology 3, 973-980. Ladbury, J. E. & Chowdhry, B. Z. (1996) Sensing the heat: the application of isothermal titration calorimetry
to thermodynamic studies of biomolecular interactions. Chemistry & Biology 3, 791-801.

LITERATUR
160
Laevitt, S. & Freire, E. (2001) Direct measurement of protein binding energetics by isothermal titration calorimetry. Curr. Opin. Struct. Biol. 11, 560-566.
Landgraf, R., Xenarios, I. & Eisenberg, D. (2001) Three-dimensional cluster analysis identifies interfaces and
functional residue clusters in proteins. J. Mol. Biol. 307, 1487-1502. Lange-Carter, C. A., Pleiman, C. M., Gardner, A. M., Blumer, K. J. & Johnson, G. L. (1993) A divergence
in the MAP kinase network defined by MEK kinase and raf. Science 260, 315-319. Lazaridis, T. & Karplus, M. (2000) Effective energy functions for protein structure prediction. Curr. Opin.
Struct. Biol. 10, 139-145. Lee, F. S., Auld, D. S. & Vallee, B. L. (1989) Tryptophan fluorescence as a probe of placental ribonuclease
inhibitor binding to angiogenin. Biochemistry 28, 219-224. Lee, C. H. J., Della, N. G., Chew, C. E. & Zack, D. J. (1996) Rin, a neuron-specific and calmodulin-binding
small G-protein. J. Neuroscience 16, 6784-6794. Lee, L. P. & Tidor, B. (2001) Barstar is electrostatically optimized for tight binding of barnase. Nature Struct.
Biol. 8, 73-76. Levers, S. J., Paterson, H. F. & Marshall, C. J. (1994) Requirement for Ras in Raf activation is overcome by
targeting Raf to the plasma membrane. Nature 369, 411-414. Levitt, M. & Park, B. H. (1993) Water: now you see it, now you don`t. Structure 15, 223-226. Li, Y., Lipschultz, C. A., Mohan, S. & Smith-Gill, S. J. (2001) Mutations of an epitope hot-spot residue alter
rate limiting steps of antigen-antibody protein-protein associations. Biochemistry 40, 2011-2022. Linnemann, T., Geyer, M., Jaitner, B. K., Block, C., Kalbitzer, H. R., Wittinghofer, A. & Herrmann, C.
(1999) Thermodynamic and kinetic characterization of the interaction between the Ras binding domain of AF6 and members of the Ras subfamily. J. Biol. Chem. 274, 13556-13562.
Linnemann, T., Kiel, C., Herter, P. & Herrmann, C. (2002) The activation of RalGDS can be achieved
independently of its Ras binding domain: Implications for an activation mechanism in Ras effector specificity and signal distribution. J. Biol. Chem. 10, 7831-7837.
Lo Conte, L., Chothia, C. & Janin, J. (1999) The atomic structure of protein-protein recognition sites. J. Mol.
Biol. 285, 2177-2198. Lumry, R. & Rajender, S. (1970) Enthalpy-entropy compensation phenomena in water solutions of proteins
and small molecules: A ubiquitous property of water. Biopolymers 9, 1125-1227. Mackay, D. J. (1998) Rho GTPases. J. Biol. Chem. 273, 20685-20688. Marshal, C. J. (1996) Ras effectors. Curr. Opin. Cell. Biol. 8, 197-204. Maurer, T. & Kalbitzer, H. R. (1996) Indirect Referencing of 31P and 19F NMR Spectra J. Magn. Reson. 113,
177-178.

LITERATUR
161
Mendelsohn, J. & Baselga, J. (2000) The EGF receptor family as targets for cancer therapy. Oncogene 19, 6550-6565.
Milburn, M. V., Tong, L., DeVos, A. M., Brünger, A., Yamaizumi, Z., Nishimura, S. & Kim, S. H. (1990)
Molecular switch for signal transduction: structural differences between active and inactive forms of protooncogenic ras proteins. Science 247, 939-945.
Moore, M. S. (1998) Ran and nuclear transport. J. Biol. Chem. 273, 22857-22860. Moss, J. & Vougham, M. (1998) Molecules in the ARF orbit. J. Biol. Chem. 273, 2143-2144. Murphy, K. P. & Freire, E. (1992) Thermodynamics of structural stability and cooperative folding behavior in
proteins. Adv. Protein Chem. 43, 313-361. Myles, T., Le Bonniec, B. F., Netz, A. & Stone, S. R. (2001) Electrostatic steering and ionic tethering in the
formation of thrombin-hirudin complexes: the role of the thrombin anion-binding exosite-I. Biochemistry 40, 4972-4979.
Nassar, N., Horn, G., Herrmann, C., Scherer, A., McCormick, F. & Wittinghofer, A. (1995) The 2.2 Å
crystal structure of the Ras-binding domain of the serine/threonine kinase c-Raf1 in complex with Rap1A and a GTP analogue. Nature 375, 554-560.
Nassar, N., Horn, G., Herrmann, C., Block, C., Janknecht, R. & Wittinghofer, A. (1996) Ras/Rap effector
specifity determined by charge reversal. Nature Struct. Biol. 3, 723-729. Nielsen, J. E., Andersen, K. V., Honig, B., Hooft, R. W. W., Klebe, G., Vriend, G. & Wade, R. C. (1999)
Improving macromolecular electrostatics calculations. Protein Eng. 12, 657-662. Noonan, T., Brown, N., Dudycz, L. & Wright, G. (1991) Interaction of GTP derivatives with cellular and
oncogenic ras-p21 proteins. J. Med. Chem. 34, 1303-1307. Norel, R., Sheinerman, F., Petrey, D. & Honig, B. (2001) Electrostatic contributions to protein-protein
interactions: fast energetic filters for docking and their physical basis. Protein Sci. 10, 2147-2161. Northrup, S. H. & Erickson, H. P. (1992) Kinetics of protein-protein-association explained by Brownian
dynamics computer simulation. Proc. Natl. Acad. Sci. USA 89, 3338-3342. Oliveberg, M., Tan, Y. J. & Fersht, A. R. (1995) Negative activation enthalpies in the kinetics of protein
folding. Proc. Natl. Acad. Sci. USA 92, 8926-8929. Orengo, C. A., Jones, D. T. & Thornton, J. M. (1994) Protein superfamilies and domain superfold. Nature
372, 631-634. Pacold, M. E., Suire, S., Perisic, O., Lara-Gonzalez, S., Davis, C. T., Walker, E. H, Hawkins, P. T.,
Stephens, L., Eccleston, J. F. & Williams, R. L. (2000) Crystal structure and functional analysis of Ras binding to its effector phosphoinoside 3-kinase gamma. Cell 103, 931-943.
Pai, E. F., Kabsch, W., Krengel, U., Holmes, K. C., John, J. & Wittinghofer, A. (1989) Structure of the
guanine-nucleotide-binding domain of the Ha-ras oncogene product p21 in the triphosphate conformation. Nature 341, 209-214.

LITERATUR
162
Pai, E. F., Krengel, U., Petsko, G. A., Goody, G. S., Kabsch, W. & Wittinghofer, A. (1990) Refined crystal structure of the triphosphate conformation of Ha-Ras p21 at 1.35 Å resolution. EMBO Journal 9, 2351-2359.
Piehler, J. & Schreiber, G. (1999) Mutational and structural analysis of the binding interface between type I
interferons and their receptor Ifnar2. J. Mol. Biol. 294, 223-237. Pearce, K. H. J., Cunningham, B. C., Fuh, G., Teeri, T. & Wells, J. A. (1999) Growth hormone binding
affinity for its receptor surpasses the requirements for cellular activity. Biochemistry 38, 81-89. Piehler, J., Roisman, L. C. & Schreiber, G. (2000) New structural and functional aspects of the IFN-receptor
interaction revealed by comprehensive mutational analysis of the binding interface. J. Biol. Chem. 275, 40425-40433.
Ponting, C. P. & Benjamin, D. R. (1996) A novel family of ras-binding domains. Trends Biochem. Sci. 21,
422-425. Ponting, C. P. & Russell, R. R. (2002) The natural history of protein domains. Annu. Rev. Biophys. Biomol.
Struct. 31, 45-71. Prasad, R., Gu Y., Alder, H., Nakamura, T., Canaani, O., Saito, H., Huebner, K., Gale, R. P., Nowell, P. C.
& Kuriyama, K. (1993) Cloning the ALL-1 fusion partner; the AF-6 gene, involved in acute myeloid leucemias with the t(6;11) chromosome translocation. Cancer Res. 53, 5624-5628.
Prior, I. A., Harding, A., Yan, J., Sluimer, J., Parton, R. G. & Hancock, J. F. (2001) GTP-dependent
segregation of H-ras from lipid rafts is required for biological activity. Nature Cell Biol. 3, 368-375. Prive, G. G., Milburn, M. V., Tong, L., de Vos, A. M., Yamaizumi, Z., Nishimura, S. & Kim, S. H. (1992)
X-ray crystal structures of transforming p21 ras mutants suggest a transition-state stabilization mechanism for GTP hydrolysis. Proc. Natl. Acad. Sci. U S A 89, 3649-3653.
Qian, H. & Hopfield, J. J. (1996) Entropy-enthalpy compensation: Perturbation and relaxation in
thermodynamic systems. J. Chem. Phys. 105, 9292-9298. Radic, Z., Kirchhoff, P. D., Quinn, D. M., McCammon, J. A. & Taylor, P. (1997) Electrostatic influence of
the kinetic of ligand binding to acetylcholinesterase. J. Biol. Chem. 37, 23265-23277. Rajpal, A., Taylor, M. G. & Kirsch, J. F. (1998) Quantitative evaluation of the chicken lysozyme epitope in
the HyHEL-10 Fab complex: Free energies and kinetics. Protein Sci. 7, 1868-1874. Rapp, U. R., Goldsborough, M. D., Mark, G. E., Bonner, T. I., Groffen, J., Reynolds, F. H. Jr. &
Stephenson, J. (1983) Structure and biological activity of v-raf, a unique oncogene transduced by a retrovirus. Proc. Natl. Acad. Sci. U.S.A. 80, 4218-4222.
Rashin, A. A. & Honig, B. (1984) On the environment of ionizable groups in globular proteins. J. Mol. Biol.
173, 515-521. Rapp, U. R., Cleveland, J. L., Bonner, T. I. & Storm, S. M. (1988) The raf oncogene. In The Oncogene
Handbook, pp. 213-253 (eds. E.P. Reddy, A.M. Skalka and T. Curran), Elsevier/North Holland Publishing Co., Amsterdam.

LITERATUR
163
Rodriguez-Viciana, P., Warne, P. H., Dhand, R., Van Haesebroeck, B., Gout, I., Fry, M. J., Waterfield, M. D. & Downward, J. (1994) Phosphatidylinositol-3-OH kinase as a direct target of Ras. Nature 370, 527-532.
Roost, H. P., Bachmann, M. F., Haag, A., Kalinke, U., Pliska, V., Hengartner, H. & Zinkernagel, R. M.
(1995) Early high-affinity neutralizing anti-viral IgG responses without further overall improvements of affinity. Proc. Natl. Acad. Sci. USA 28, 1257-1261.
Rudolph, M. G., Linnemann, T., Grünewald, P., Wittinghofer, A., Vetter, I. R. & Herrmann, C. (2001)
Thermodynamics of Ras/effector interactions probed by isothermal titration calorimetry. J. Biol. Chem. 276, 23914-23921.
Scheffzek, K., Grünewald, P., Wohlgemuth, S., Kabsch, W., Tu, H., Wigler, M., Wittinghofer, A. &
Herrmann, C. (2001) The Ras-Byr2RBD complex: structural basis for Ras effector recognition in yeast. Structure 9, 1043-1050.
Sanger, F., Nicklen, S. & Coulson, A. R. (1977) DNA sequencing with chain-terminating inhibitors. Proc.
Natl. Acad. Sci. USA 74, 5463-5467. Saraste, M., Sibbald, P. R. & Wittinghofer, A. (1990) The P-loop – a common motif in ATP-and GTP-binding
proteins. Trends Biochem. Sci. 15, 430-434. Scheffzeck, K., Grünewald, P., Wohlgemuth, S., Kabsch, W., Tu, H., Wigler, M., Wittinghofer, A. &
Herrmann, C. (2001) The Ras-Byr2RBD complex: Structural basis for Ras effector recognition in yeast. Structure 9, 1043-1050.
Schimmoller, F., Itin, C. & Pfeffer, S. (1997) Vesicle transport, get your coat. Curr. Biol. 7, 235-237 Schreiber, G. & Fersht, A. (1993) Interaction of Barnase with its polypeptide inhibitor Barstar studied by
protein engineering. Biochemistry. 32, 5145-5150. Schreiber, G., Buckle, A. M. & Fersht, A. R. (1994) Stability versus function: two competing forces in the
evolution of barstar. Structure 2, 945-951. Schreiber, G. & Fersht, A. R. (1995) Energetics of protein-protein interactions: analysis of the barnase-barstar
interface by single mutations and double mutant cycles. J. Mol. Biol. 248, 478-486. Schreiber, G. & Fersht, A. (1996) Rapid, electrostatically assisted association of proteins. Nature Struct. Biol.
3, 427-431. Schreiber, G. (2002) Kinetic studies on protein-protein interactions. Curr. Opin. Struct. Biol. 12, 41-47. Schürmann, A. (1996) Cloning a new family of mammalian GTP-binding proteins (RagA, RagB5, RagB1) with
remote similarity to the Ras-related GTPases. J. Biol. Chem. 270, 28, 982-988. Schweins, T. & Wittinghofer, A. (1994) GTP binding proteins, structures, interactions and relationships. Curr.
Biol. 4, 547-550. Schweins, T., Scheffzek, K., Aßheuer, R. & Wittinghofer, A. (1997) The role of the metal ion in the p21ras
catalysed GTP-hydrolysis: Mn2+ versus Mg2+. J. Mol. Biol. 266, 847-856.

LITERATUR
164
Scolnick, E. M., Papageorge, A. G. & Shit, T. Y. (1979) Guanine nucleotide binding activity as an assay for src protein of rat-derived murine sarcoma viruses. Proc. Natl. Acad. Sci. USA 76, 5355-5359.
Selzer, T. & Schreiber, G. (1999) Predicting the rate enhancement of protein complex formation from the
electrostatic energy of interaction. J. Mol. Biol. 287, 409-419. Selzer, T., Albeck, S. & Schreiber, G. (2000) Rational design of faster associating and tighter binding protein
complexes. Nature Struct. Biol. 7, 537-541. Selzer, T. & Schreiber, G. (2001) New insights into the mechanism of protein-protein association. Proteins 45,
190-198. Shaka, A. J., Baker, P. B. & Freeman, R. (1985) Computer-optimized decoupling scheme for wideband
applications and low-level operation. J. Magn. Res. 96, 547-552. Sharp, K. (2001) Entropy-enthalpy compensation: Fact or artefact? Protein Science 10, 661-667. Sheinerman, F. B., Norel, R. & Honig, B. (2000) Electrostatic aspects of protein-protein interactions. Curr.
Opin. Struct. Biol. 10, 153-159. Sheinerman, F. B. & Honig, B. (2002) On the role of electrostatic interactions in the design of protein-protein
interfaces. J. Mol. Biol. 318, 161-177. Shi, Y. Y., Mark, A. E., Wang, C. X., Huang, F., Berendsen, H. J. & van Gunsteren, W. F. (1993) Can the
stability of protein mutants be predicted by free energy calculations? Protein Eng. 6, 289-295. Shoup, D. & Szabo, A. (1982) Role of diffusion in ligand binding to macromolecules and cell-bound receptors.
Biophys. J. 40, 33-39. Smoluchowski, M. V. (1917) Versuch einer mathematischen Theorie der Koagulationskinetik kolloider
Loesungen. Z. Phys. Chem. 92, 129-168. Sommer, J., Jonah, C., Fukuda, R. & Bergsohn, R. (1982) Production and subsequent second-order
decomposition of protein disulfide anions. J. Mol. Biol. 159, 721-744. Spaargaren, M. & Bischoff, J. R. (1994) Identification of the guanine nucleotide dissociation stimulator for Ral
as a putative effector molecule of R-ras, H-ras, K-ras and Rap. Proc. Natl. Acad. Sci. USA 91, 12609-12613.
Spoerner, M., Herrmann, C., Vetter, I. R., Kalbitzer, H. R. & Wittinghofer, A. (2001) Dynamic properties
of the Ras switch I region and its importance for binding to effectors. Proc. Nat. Acad. Sci. USA 98, 4944-4949.
Stephens, L. R., Jackson, T. R. & Hawkins, P. T. (1993) Agonist-stimulated synthesis of phosphatidylinositol
(3,4,5,) triphosphate: A new intracellular signalling system? Biochem. Biophys. Acta 1179, 27-75. Stites, W. E. (1997) Protein-Protein Interactions: Interface structure, binding thermodynamics, and mutational
analysis. Chem. Rev. 97, 1233-1250. Stokoe, D., MacDonald, S. G., Cadwallader, K., Symons, M. & Hancock, J. F. (1994) Activation of Raf as a
result of recruitment to the plasma membrane. Science 264, 1463-1467.

LITERATUR
165
Stokoe, D. & McCormick, F. (1997) Activation of c-Raf-1 by Ras and Src through different mechanism:
activation in vivo and in vitro. EMBO J. 16, 2384-2396. Stone, S. R., Dennis, S. & Hofsteenge, J. (1989) Quantitative evaluation of the contribution of ionic
interactions to the formation of the thrombin-hirudin complex. Biochemistry 28, 6857-6863. Studier, F. W. & Moffat, B. A. (1986) Use of bacteriophage T7 RNA polymerase to direct selective high-level
expression of cloned genes. J. Mol. Biol. 189, 113-130. Sydor, J. R., Engelhard, M., Wittinghofer, A., Goody, R. & Herrmann, C. (1998) Transient kinetic studies
on the interaction of Ras and the Ras-binding domain of c-Raf-1 reveal rapid equilibrium of the complex. Biochemistry 37, 14292-14299.
Tamada, M., Hu, C. D., Kariya, K., Okada, T. & Kataoka, T. (1997) Membrane recruitment of Raf-1 is not
the only funtion of ras in Raf-1 activation. Oncogene 15, 2959-2964. Taveras, G., Remiszewski, S. W., Doll, R. J., Cesarz, D., Huang, E. C., Kirschmeier, P., Pramanik, B. N.,
Snow, M. E., Wang, Y. S., del Rasio, J. D., Bauer, B. B., Brown, J. E., Carr, D., Catino, J., Evans, C. A., Girijavallabhan, V., Heimark, L., Libverles, J. L., Nash., C., Perkins, L., Senior, M. M., Tsarbapoulos, A., Ganguly, A. K., Aust, R., Brown, E., Delisle, D., Fuhrmann, S., Hendrickson, T., Kissinger, C., Love, R., Sisson, W., Villafranca, E. & Webber, S. E. (1997) Ras oncoprotein inhibitors: the discovery of potent, ras nucleotide exchange inhibitors and the structural determination of a drug-protein complex. Bioorg. Med. Chem. 5, 125-133.
Thompson, H. J., Jiang, C., Lu, J., Metha, R. G., Piazza, G. A., Paranka, N. S., Paucku, R. & Ahnen, D. J.
(1997) Sulfone metabolite of sulindac inhibits mammary carcinogenesis. Cancer Res. 57, 267-271. Trahey, M. & McCormick, F. (1987) A cytoplasmic protein stimulates normal N-ras p21 GTPase, but does not
affect oncogenic mutants. Science 238, 542-545. Tsai, C. J., Lin, S. L., Wolfson, H. J. & Nussiniv, R. (1997) Studies of protein-protein interfaces: a statistical
analysis of the hydrophobic effect. Protein Sci. 6, 53-64. Urano, T., Emkey, R. & Feig, L. A. (1996) Ral-GTPases mediate a distinct downstream signalling pathway
from Ras that facilitates cellular transformation. EMBO J. 15, 810-816. Valencia, A., Chardin, P., Wittinghofer, A. & Sander, C. (1991) The ras protein family: evolutionary tree and
role of conserved amino acids. Biochemistry 30, 4637-4648. Vavvas, D., Li, X., Avruch, J. & Zhang, X. F. (1998) Identification of NORE1 as a potential Ras effector. J.
Biol. Chem. 273, 5439-5442. Vetter, I. R., Linnemann, T., Wohlgemuth, S., Geyer, M., Kalbitzer, H. R., Herrmann, C. & Wittinghofer,
A. (1999) Structural and biochemical analysis of Ras-effector signaling via RalGDS. FEBS Letters 451, 175-180.
Vetter, I. R. & Wittinghofer, A. (2001) Signal transduction – the guanine nucleotide-binding switch in three
dimensions. Science 294, 1299-1304.

LITERATUR
166
Vijayakumar, M., Wong, K.-Y., Schreiber, G., Fersht, A. R., Szabo, A. & Zhou, H.-X. (1998) Electrostatic enhancement of diffusion-controlled protein-protein association: Comparison of theory and experiment on barnase and barstar. J. Mol. Biol. 278, 1015-1024.
Vojtek, A. B. & Der, C. J. (1998) Increasing complexity of the Ras signaling pathway. J. Biol. Chem. 273,
19925-19928. Walker, J. E., Saraste, M., Runswick, M. J. & Gay, N. J. (1982) Distantly related seuences in the alpha- and
beta-subunits of ATP synthase, myosin, kinases and other ATP-requiring enzymes and a common nucleotide binding fold. EMBO Journal 1, 945-951.
Walker, E. H., Perisic, O., Ried, C., Stephens, L. & Williams, R. L. (1999) Structural insights into
phosphoinositide 3-kinase catalysis and signalling. Nature 402, 313-320. Wallis, R., Moore, G. R., James, R. & Kleanthous, C. (1995) Protein-protein interactions in colicin E9
DNase-immunity protein complexes. 1. Diffusion-controlled association and femtomolar binding for the cognate complex. Biochemistry 34, 13743-13750.
Wang, Y., Shen, B. J. & Sebald, W. (1997) A mixed-charge pair in human interleukin 4 dominates high-
affinity interaction with the receptor alpha chain. Proc. Natl. Acad. Sci. USA 94, 1657-1662. Wang, Y., Waldron, R. T., Dhaka, A., Patel, A., Riley, M. M., Rozengurt, E. & Colicelli, J. (2002) The Ras
effektor RIN1 directly competes with RAF and is regulated by 14-3-3 proteins. Mol. Cell. Biol. 22, 916-926.
Weiss, B., Bollag, G. & Shannon, K. (1999) Hyperactive Ras as a therapeutic target in neurofibromatosis type
1. Am. J. Med. Genet. 89, 14-22. Wells, J. A. (1996) Binding in the growth hormone receptor complex. Proc. Natl. Acad. Sci. USA 93, 1-6. Wendt, H., Leder, L., Haermae, H., Jelesarov, I., Baici, A. & Bosshard, H. R. (1997) Very rapid, ionic
strength-dependent association and folding of a heterodimeric leucine zipper. Biochemistry 36, 204-213.
Wiesmüller, L. & Wittinghofer, F. (1994) Signal transduction pathways involving ras. Cell. Signalling 6, 247-
267. Wiseman, T., Williston, S., Brandts, J. F. & Lin, L.-N. (1989) Rapid measurement of binding constants and
heats of binding using a new titration calorimeter. Analyt. Biochem. 179, 131-137. Wittinghofer, A., Pai, E. F. & Goody, R. S. (1993) Structural and mechanistic aspects of the GTPase reaction
of H-Ras p21. Handbook of Exp. Pharmacology 108, 196-211. Wittinghofer, A. & Herrmann, C. (1995) Ras-effector interactions, the problem of specificity. FEBS Lett. 369,
52-56. Wittinghofer, A. & Nassar, N. (1996) How Ras-related proteins talk to their effectors. TIBS 21, 488-491. Wittinghofer, A & Waldmann, H. (2000) Ras – A molecular switch involved in tumor formation. Angew.
Chem. Int. Ed. 39, 4192-4214.

LITERATUR
167
Wolin, R., Wang, D., Kelley, J., Afonso, A., James, L., Kirschmeier, P. & Mc Phail, A. (1996) Synthesis of isomeric 3-piperidinyl and 3-pyrrolidinyl benzo[5,6]cyclohepta[1,2-b]pyridines: sulfonamido derivatives as inhibitors of Ras prenylation. Bioorg. Med. Chem. Lett. 6, 195-200.
Wolthuis, R. M. F., Bauer, B., van`t Veer, L. J., de Vries-Smits, A. M. M., Cool, R. H., Spaargaren, M.,
Wittinghofer, A., Burgering, B. M. T. & Bos., J. L. (1996) RalGDS-like factor (Rlf) is a novel Ras and Rap1A-associating protein. Oncogene 13, 353-362.
Xiao, L. & Honig, B. (1999) Electrostatic contributions to the stability of hyperthermophilic proteins. J. Mol.
Biol. 289, 1435-1444. Zhou, H. X. & Szabo, A. (1996) Theory and simulation of the time-dependent rate coefficients of diffusion-
influenced reactions. Biophys. J. 71, 2440-2457. Zor, T., Bar-Yaacov, M., Elgavish, S., Shaanan, B. & Selinger, Z. (1997) Rescue of a mutant G-protein by
substrate-assisted catalysis. Eur J Biochem. 249, 330-336. Zor, T., Andorn, R., Sofer, I., Chorev, M. & Selinger Z. (1998) GTP analogue hydrolysis by the Gs protein:
implication for the role of catalytic glutamine in the GTPase reaction. FEBS Lett. 433, 326-330. Zwartkruis, F. J. T., Wolthuis, R. M. F., Nabben, N. M. J. M., Franke, B. & Bos, J. L. (1998) Extracellular
signal-regulated activation of Rap1 fails to interfere in Ras effector signalling. EMBO J. 17, 5905-5912.
Bisherige eigene Veröffentlichungen: Kiel, C. & Herrmann, C. (2001) Double mutant cycle analysis of Ras proteins and their effektors:
Quantification of binding specificity. Published in “Protein Modules in Cellular Signaling”, edited by Heilmeyer, L. & Friedrich, P. IOS Press
Linnemann, T., Kiel, C., Herter, P. & Herrmann, C. (2002) The activation of RalGDS can be achieved
independently of its Ras binding domain: Implications for an activation mechanism in Ras effector specificity and signal distribution. J. Biol. Chem. 10, 7831-7837.
Ahmadian, M. R., Kiel, C., Stege, P. & Scheffzek, K. Fingerprints of the Ras-GTPase activating proteins
neurofibromin and p120GAP. J. Mol. Biol., in Revision. Gohlke, H., Kiel, C. & Case, D. A. Insights into protein-protein binding by binding free-energy calculation and
free energy decomposition for the Ras-Raf and Ras-RalGDS complexes, eingereicht. Kiel, C., Serrano, L. & Herrmann, C. Energy landscapes of proteins: A detailed thermodynamic analysis of
Ras/effector interfaces. J. Mol. Biol., eingereicht.

DANKSAGUNG
An erster Stelle bedanke ich mich bei Prof. Alfred Wittinghofer für die Möglichkeit diese Doktorarbeit in seiner Abteilung durchzuführen. Des Weitern bedanke ich mich für sein fortlaufendes Interesse an meiner Arbeit und für die Übernahme des Erstgutachtens.
* Bei Prof. Rolf Heumann bedanke ich mich für die Übernahme des Zweitgutachtens und bei Prof. Martin Feigel als dritten Prüfer.
* Ein besonderer Dank geht an Prof. Christian Herrmann für die gute Unterstützung. Insbesondere danke ich ihm, sowie Prof. Gideon Schreiber vom Weizmann-Institut in Israel, für die Initiierung des GIF-Projektes und für ihren Einsatz hinsichtlich der guten Zusammenarbeit und der gegenseitigen Besuche im Rahmen dieses Projektes.
* Die finanzielle Unterstützung erfolgte durch die Deutsch-Israelische Stiftung für Wissenschaftliche Forschung und Entwicklung (GIF).
* Ich danke Prof. Hans Robert Kalbitzer und Dr. Michael Spoerner von der Universität Regensburg für die Hilfe bei den NMR-Messungen.
* Ich bedanke mich bei Beate Voss und Ulrike Herbrand für ihre Hilfe und Anregungen in der Zellkultur.
* Des Weiteren danke ich Sabine Wohlgemuth für viele Ideen im Labor und die schönen Milchkaffee-Pausen auf der Terrasse. Meinen Laborkollegen und allen weiteren Mitarbeitern danke ich für das gute Arbeitsklima.
* Ganz besonders möchte ich mich bei Christine Welz-Kiel und Achim Kiel für ihre Hilfe und Freundschaft bedanken.
* Meinen lieben Eltern danke ich für ihre ständige Unterstützung und Förderung.
![) [Uyumluluk Modu]) · 2016. 11. 22. · Diyaliz Travma Kesici , delici alet yaralanmalar ... 2.2.3.1: Radyasyon güvenli ği el kitabı 2.2.3.3: Dozimetre takipleri 2.2.3.4: Çekim](https://img.pdfslide.tips/doc/110x75/612681d9baefd332493a1fdf/-uyumluluk-modu-2016-11-22-diyaliz-travma-kesici-delici-alet-yaralanmalar.jpg)




























