Embed Size (px)
Citation preview

124 Biochlmica et Biophvst~'a A eta 845 ~ 19851 124 13()
Elsevier
BBA 11439
Uptake and degradation of insulin and a2-macroglobulin-trypsin complex in rat adipocytes. Evidence for different pathways
Jorgen Gliemann and Ole Sonne Institute of Physiology, University of Aarhus, Universitetsparken, DK-8000 Aarhus C (Denmark)
(Received September 27th, 1984)
Key words: az-Macroglobulin- trypsin; Insulin; Protein degradation; Protein uptake
The cell association and degradation of insulin and a2-macroglobulin-trypsin complex were measured in rat adipocytes with or without various inhibitors in the attempt to clarify whether the two ligands were taken up by the same or by different pathways. Several inhibitors, and particularly those of membrane traffic, lysosomal function and transglutaminase activity, affected the two ligands differently. Thus, chloroquine (I00 /xM) reduced both the uptake of a2-macroglobulin, trypsin and its receptor-mediated degradation by about 70%. In contrast, the uptake of insulin was increased 2-3-times and the receptor-mediated degradation was only slightly reduced. Methylamine (I0 mM) and ammonium chloride (I0 mM) reduced degradation of a2-macroglobulin, trypsin markedly without affecting that of insulin. Leupeptin (I00/~M) increased uptake and reduced degradation of a2-macroglobulin- trypsin without affecting insulin. Dansylcadaverine (500/~M) almost abolished uptake and degradation of a2-macroglobulin.trypsin but had little effect on insulin. Moreover, uptake and degradation of a2-macroglobulin" trypsin was much more sensitive than insulin to the action of metabolic inhibitors such as dinitrophenol and cyanide. The results show that the two ligands are taken up by functionally different systems. In addition, they support the hypothesis that lysosomes play a relatively minor role in the receptor-mediated degradation of insulin.
Introduction
Polypeptide hormones and several glycopro- teins bind to specific receptors on the plasma membrane of the cell. In most cell types, binding is followed by internalization into a vesicular sys- tem followed by degradation of the ligands. The question therefore arises whether different ligands bound to their respective receptors are taken up by the same vesicular system or by different systems. Maxfield et al. [1] followed the uptake of fluo- rescent derivatives of insulin, epidermal growth factor and a2-macroglobulin in Swiss 3T3-4 fibroblasts and suggested a common pathway of
Abbreviation: Hepes, 4-(2-hydroxyethyl)-l-piperazineethane- sulfonic acid.
internalization. However, the fluorescent insulin, a rhodamine lactalbumin-insulin complex, exhibited only about 2% of insulin's potency [2] and this may have influenced the results.
Binding and internalization of insulin in the rat adipocyte is well-characterized and an iodinated insulin derivative with a potency indistinguishable from that of native insulin is available [3]. Several studies have shown that 60-80% of the cell-associ- ated insulin at steady state (37°C) is internalized in the sense that it cannot be removed by trypsin or incubation of the cells at pH 3 [4-6]. We have recently shown that az-macroglobulin, trypsin is specifically taken up by the adipocytes at 37°C and that at least 75% of the cell-associated ligand is internalized [7]. Both insulin [8] and a2-macro- globulin, trypsin [7] are degraded as a conse-
0167-4889/85/$03.30 © 1985 Elsevier Science Publishers B.V. (Biomedical Division)

quence of their binding and internalization. We decided to compare the cell association and de- gradation of the two ligands in the presence of various inhibitors in order to elucidate whether a common or separate pathways were followed.
Materials and Methods
a2-Macroglobulin, prepared from human plasma as described previously [9] using Zn 2+ chelate affinity chromatography, was a gift from Dr. L. Sottrup-Jensen, Institute of Molecular Biol- ogy, University of Aarhus. It was iodinated as described before [7] using 125I- (1.8 Ci//~mol) from Amersham International, except that the de- gree of iodination was 0.3 tool I / tool a2-macro- globulin. 12~I-labelled a2-macroglobulin-trypsin complex was prepared by the addition of a 4-fold molar excess of trypsin (from Boehringer, 60% active) for 5 min at 20°C followed by the addition of soybean trypsin inhibitor at a final molar con- centration twice that of trypsin. Finally, the mix- ture was applied to a 0.9 × 60 cm Sephacryl S-300 column (Pharmacia) and chromatographed in 10 mM Hepes buffer. The protein concentrations in the eluted fractions were determined spectrophoto- metrically using E~8~0 = 9.1, the a2-macroglobulin • trypsin-containing fractions were recovered and used for incubation. Unlabelled a2-macroglobulin was reacted similarly with trypsin followed by soybean trypsin inhibitor and used without further purification.
[125 I]Monoiodoinsulin (essentially 'carrier-free') labelled in residue 14 of the A chain (tyrosine) prepared from pig insulin as described previously [3,10] was a gift from Dr. S. Linde, Hagedorn Research Laboratory, Gentofte, Denmark. Its binding affinity and biological potency on rat adipocytes is closely similar to that of insulin [3]. Unlabelled pig insulin was from Nordisk, Gentofte.
Bovine serum albumin (fraction V) was from Sigma. It was dialyzed against three changes of distilled water and passed through an 0.8 /~m millipore filter before use. Collagense (type 1) was from Worthington. Leupeptin was from the Pro- tein Research Foundation (Osaka). Inorganic salts (analytical grade) were from Merck. All other chemicals were from Sigma.
Rat adipocytes were prepared from epididymal fat pads of ad libitum fed 140-180 g male Wistar
125
rats as described previously [11]. The cells were incubated in buffer containing (in millimolar con- centration) 140 Na+; 4.7 K÷; 2.5 Ca2+; 1.25 Mg2+; 142 C1-; 2.5 H2PO4-/HPO42- ; 1.25 SO42- ; 10.0 Hepes, and 50 mg/ml bovine serum albumin. pH was 7.4 at 37°C. The cell concentration was 5%, v/v, that is about 5 • 10 5 celis/ml. 0.5 rnl cell suspension was incubated in 14 ml round-bottom polyethylene tubes with shaking. The concentra- tion of 125I-labelled a2-macroglobulin, trypsin was 9 nCi /ml (20000 cpm/ml) which is equivalent to 17 pM aa-macroglobulin, trypsin (labelled plus unlabelled). This is very much lower than the apparent K m for the cell association to adipocytes of about 10 nM [7]. The concentration of [125I]monoiodoinsulin was the same in Ci /ml which is equivalent to 5 pM monoiodoinsulin. This is also very much lower than the apparent K m for cell association of insulin of about 2 nM [12].
The incubations were terminated as described previously [8] by the addition of 10 ml 10°C 0.9% NaC1 and 1.2 ml silicone oil (density 0.99) im- mediately followed by centrifugation at about 1000 x g for 40 s. The cell pellet on the surface of the silicone oil was adsorbed to a piece of pipe cleaner and the cell-associated radioactivity de- termined in a gamma counter with an efficiency of 50% and a background of 5 cpm. Nonspecific (not saturable) association of the two labelled ligands were determined from incubations also containing 0.5 ttM unlabelled a2-macroglobulin, trypsin or 1.0 gM unlabelled insulin. The nonspecific cell association of 125I-labelled a2-macroglobulin. trypsin, including trapped extracellular radioactiv- ity, did not exceed 1.5% of the total cell-associated 125I-labelled a2-macroglobulin, trypsin. The equiv- alent figure for [125I]monoiodoinsulin was 4%. All data have been corrected for the small amount of nonspecific cell association. Degradation was mea- sured as the difference in solubility of the tracers in 12% trichloroacetic acid (final concentration) before and after incubation [7]. The solubility of the 125I-labelled a2-macroglobulin • trypsin did not exceed 1% and the solubility of the [125I]mono- iodoinsulin not 2% before incubation.
All experiments were carried out in parallel so that cell association and degradation of the two ligands were always measured in the same cell preparations.
6 -
"~ 5-
~, 4 -
¢1 0
"0
0 u~ co
I 1 -
Results
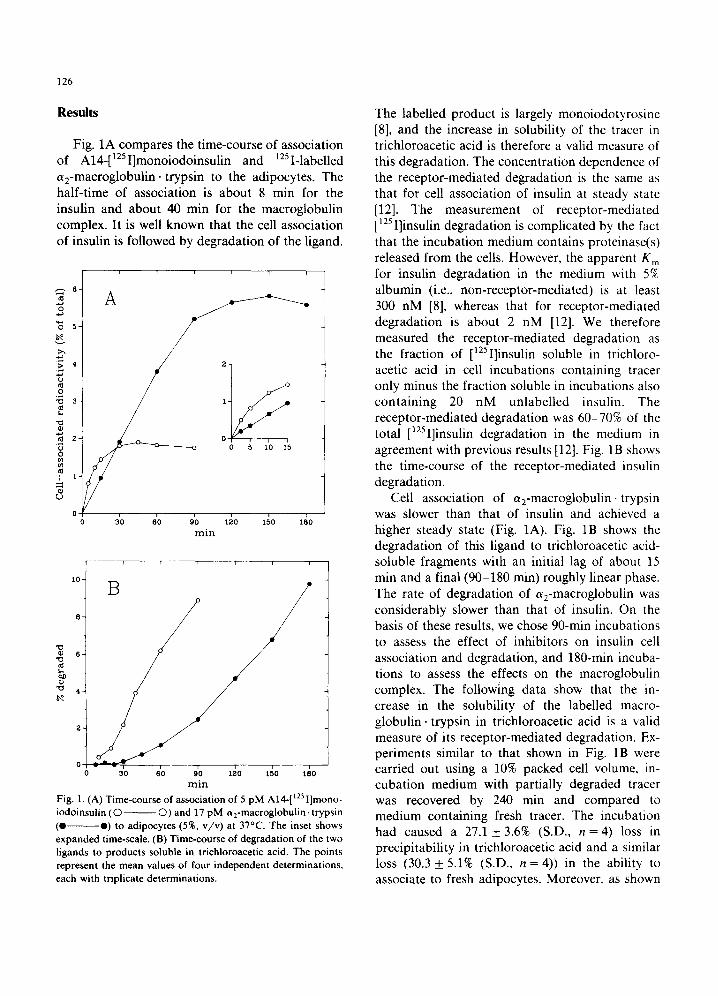
Fig. 1A compares the time-course of association of A14-[lzSI]monoiodoinsulin and azSI-labelled a2-macroglobulin, trypsin to the adipocytes. The half-time of association is about 8 min for the insulin and about 40 min for the macroglobulin complex. It is well known that the cell association of insulin is followed by degradation of the ligand.
"0
A
Z
3'0 6'0 9'0 120 ~ F 150 180
r a i n
w - i
30 do ~o ~o i~o ~o rain
1 0 -
8-
6 -
4 -
2-
O ,
126
Fig. 1, (A) Time-course of association of 5 pM A14-[1251]mono- iodoinsulin (© ©) and 17 pM a2-macroglobulin- trypsin (e O) to adipocytes (5%, v/v) at 37°C. The inset shows expanded time-scale. (B) Time-course of degradation of the two ligands to products soluble in trichloroacetic acid. The points represent the mean values of four independent determinations, each with triplicate determinations.
The labelled product is largely monoiodotyrosine [8], and the increase in solubility of the tracer in trichloroacetic acid is therefore a valid measure of this degradation. The concentration dependence of the receptor-mediated degradation is the same as that for cell association of insulin at steady state [12]. The measurement of receptor-mediated [125 I]insulin degradation is complicated by the fact that the incubation medium contains proteinase(s) released from the cells. However, the apparent K m for insulin degradation in the medium with 5% albumin (i.e., non-receptor-mediated) is at least 300 nM [8], whereas that for receptor-mediated degradation is about 2 nM [12]. We therefore measured the receptor-mediated degradation as the fraction of [tzsI]insulin soluble in trichloro- acetic acid in cell incubations containing tracer only minus the fraction soluble in incubations also containing 20 nM unlabelled insulin. The receptor-mediated degradation was 60-70% of the total [125I]insulin degradation in the medium in agreement with previous results [12]. Fig. 1B shows the time-course of the receptor-mediated insulin degradation.
Cell association of a2-macroglobulin, trypsin was slower than that of insulin and achieved a higher steady state (Fig. 1A). Fig. 1B shows the degradation of this ligand to trichloroacetic acid- soluble fragments with an initial lag of about 15 min and a final (90-180 min) roughly linear phase. The rate of degradation of a2-macroglobulin was considerably slower than that of insulin. On the basis of these results, we chose 90-min incubations to assess the effect of inhibitors on insulin cell association and degradation, and 180-min incuba- tions to assess the effects on the macroglobulin complex. The following data show that the in- crease in the solubility of the labelled macro- globulin, trypsin in trichloroacetic acid is a valid measure of its receptor-mediated degradation. Ex- periments similar to that shown in Fig. 1B were carried out using a 10% packed cell volume, in- cubation medium with partially degraded tracer was recovered by 240 min and compared to medium containing fresh tracer. The incubation had caused a 27.1 + 3.6% (S.D., n =4 ) loss in precipitability in trichloroacetic acid and a similar loss (30.3 _+ 5.1% (S.D., n = 4)) in the ability to associate to fresh adipocytes. Moreover, as shown

in Fig. 2, the concentration dependence of cell association was almost identical to that of de- gradation of a2-macroglobulin, trypsin to trichlo- roacetic acid-soluble products. It is important to note that only a negligible fraction of the degrada- tion is non-saturable.
Table I shows the effect of various inhibitors on the cell association and the receptor-mediated de- gradation of the two ligands. None of these showed any effect on the receptor independent insulin degradation in the medium (data not shown).
Colchicine, an antitubulin, caused a marked increase in the cell association of insulin, but reduced its receptor-mediated degradation. How- ever, colchicine almost blocked both the cell as- sociation and the degradation of a2-macroglobulin • trypsin. Vinblastine (also an antitubulin) had only small effects on insulin but caused some reduction in the cell association of a2-macro- globulin, trypsin and a marked decrease in its degradation. Monensin, an inhibitor of membrane traffic [13] increased insulin association but not its degradation, whereas both parameters were de- creased with the c~2-macroglobulin complex. The effect of chloroquine was similar, although more marked, and it also caused a small reduction in the
127
I -8 6 1
.g
8!
0 o . 6 2 ' 0 . 6 8 ' o .g~ ' 1 .~5 ' g ' 2b ' a b ' a ~ o ' 0
azN.T (nit)
Fig. 2. Concentration dependence of cell association (o O) and degradation (~ A) of a2-macroglobulin -trypsin (a2M.T). Adipocytes were incubated for 180 min as described in the legend to Fig. 1. The points represent the mean values of two independent experiments, each with four repli- cate determinations.
degradation of insulin. Chloroquine is an inhibitor of membrane traffic [13] although its effect in adipocytes has been related to its additional effect on lysosomal function [14,15]. Leupeptin, an effec- tive inhibitor of lysosomal proteolysis [16], de-
TABLE I
THE EFFECT OF INHIBITORS ON CELL ASSOCIATION AND RECEPTOR-MEDIATED DEGRADATION OF INSULIN AND a2-MACROGLOBULIN • TRYPSIN
The numbers indicate the cell association and degradation in percent of that obtained in control cells ± 1 S.D. They represent the mean values of four independent experiments with triplicate determinations for each drug using parallel incubations of the two ligands. The incubation times were 90 min with insulin and 180 min with a2-macroglobulin, trypsin. Receptor-mediated degradation of a2-macroglobulin, trypsin was calculated as the total degradation of tracer minus the degradation in the presence of 500 nM unlabelled macroglobulin complex; that of insulin was calculated as the total degradation of tracer minus the degradation in the presence of 20 nM unlabelled insulin. Zero indicates that cell association or degradation was not measurable.
Insulin a2-Macroglobulin, trypsin
cell degradation cell association association
degradation
Colchicine (10 mM) 298 + 44 20 _+ 8 4 + 2 Vinblastine (100 ~t M) 130 + 12 87 _+ 15 75 + 6 Monensin (100/~M) 180 + 18 96 _+ 12 72 + 6 Chloroquine (100/~ M) 290 + 32 71 ± 14 33 + 4 Leupeptin (100/tM) 98 ± 5 104 ± 11 124 ± 6 NH4CI(10mM) 107+ 8 99± 12 41±3 Methylamine (10 mM) 123 ± 7 82 ± 16 23 ± 4 Dansylcadaverine (500/xM) 126 ± 13 79 ± 21 12 ± 2 Dinitrophenol (500 #M) 92± 9 95 ± 11 49±5 Cyanide (10 raM) 55 ± 4 63 ± 11 0
16+ 7 21+ 4 36+13 18+ 5 33+ 6 26+ 8 19+ 6 0
57+ 6 0

128
creased degradation of the macroglobulin complex markedly and increased its cell association, al- though no effect on insulin was demonstrable. Other experiments (not demonstrated) showed that ieupeptin increased the fraction of cell-associated 125I-labelled macroglobulin not removed by l mg /ml trypsin [7] to more than 80%. NH4C1 and methylamine inhibit lysosomal enzymes by in- creasing intralysosomal pH [17,18]. They had little effect on insulin but decreased the cell association and particularly the degradation of the macro- globulin complex markedly. Monodansylca- daverine almost abolished the cell association and degradation of the macroglobulin complex without affecting insulin much. A similar effect of dansylcadaverine on the uptake of macroglobulin in cultured fibroblasts has been thought of as related to its effect on transglutaminase [19]. Fi- nally, Table I shows that the metabolic inhibitors, dinitrophenol and particularly cyanide, which de- plete the cells of ATP, influence c%-macroglobulin • trypsin much more than insulin• In fact, 10 mM cyanide completely abolished the uptake and de- gradation of the macroglobulin complex.
The experiments presented here were carried out using incubations for 90 min with insulin and 180 min with ct2-macroglobulin-trypsin in order to achieve about 10% degradation of both ligands. Similar results were obtained when 90-min incuba- tions were used for both ligands, although the degradation of the macroglobulin complex was low at that time and the measurements therefore subject to large errors• Cyanide caused some cell lysis in contrast to the other inhibitors and it is therefore particularly important that the results in the presence of this inhibitor were the same after 90 rain as after 180 min, i.e., neither cell associa- tion nor degradation of the a2-macroglobulin com- plex could be demonstrated (data not shown)•
The different effects of primary amines on the cell association and degradation of the two ligands is perhaps one of the most important results, and we therefore followed the time-course of action of 10 mM methylamine. Fig. 3A shows that the in- hibition of cell association of the macroglobulin complex was almost complete by 30 rain. Likewise, addition of methylamine 30 min before the tracer almost abolished the cell association (Fig. 3A) and under these conditions degradation of the complex
2
.o
o
I
r..)
6 -
5 -
4 -
3 -
2 -
1
0 0 3'0 610 9'0 f i 120 150 180
rain
2 2
~n o o
2
O
C_)
2 -
1_/ 0
0
Insu l in
15 3'0 4~5 6'0 7f5
min 9'0
Fig. 3. Effect of 10 mM methylamine on the cell association of (A) a2-macroglobulin, trypsin (a 2 M. T) and (B) insulin. Con- trol (O O), methylamine added at time zero (n D), methylamine added 30 min before the tracer (ll I1). The points represent the mean values of three independent experi- ments, each with triplicate determinations.
was not demonstrable (not shown). In contrast, methylamine added at time zero caused a small increase in the cell association of insulin after about 30 rain and caused only a slight delay when added 30 min before the tracer (Fig. 3B).
Discussion
It is important to consider whether the effect of an inhibitor might interfere with the recognition

step, that is binding to plasma membrane recep- tors. For instance, it has been thought that bacitracin, which inhibits cell association of a2- macroglobulin, trypsin both in fibroblasts [20,21] and adipocytes [7] exerted its action because of its transglutaminase-inhibiting properties [21]. How- ever, this peptide, whose effect on cell association of insulin is absent or trivial in adipocytes [22], turned out to be a competitive inhibitor of binding of the macroglobulin complex [23,7]. It is therefore obvious that the different effects of bacitracin on the two ligands do not allow conclusions with regard to pathways of internalization. The same argument applies to the very marked C a 2÷ depen- dence of a2-macroglobulin, trypsin (in contrast to insulin) cell association in adipocytes [7], and to an observation that a2-macroglobulin • trypsin cell as- sociation is much more sensitive than insulin cell association to the sulfhydryl blocking agent, N- ethylmaleimide (unpublished results). These differ- ent effects may well be exerted at the level of recognition.
The inhibitors listed in Table I almost certainly exert their action on internalization and/or de- gradation. It should be noted that at least 75% of the a2-macroglobulin • trypsin [7] and insulin [4-6] is internalized (not removable by trypsin or acid) in control cells [7]. Thus, major reductions of the cell-associated radioactivity imply inhibition of in- ternalization. It can therefore be concluded from the present study that several inhibitors influence internalization and degradation of erE-macro- globulin, trypsin very differently.
Inhibitors are rarely completely specific for a particular process, and it is therefore more dif- ficult to use them to characterize pathways than for pointing out differences between pathways. However, it has been proposed that a2-macro- globulin, trypsin is internalized into endocytic vesicles in cultured fibroblasts [24] and rabbit al- veolar macrophages [25] followed by fusion with lysosomes and degradation of the ligand. The lag time of degradation (Fig. 1B), the effects of leupeptin, chloroquine and amines suggest that a2-macroglobulin, trypsin is also targeted for lysosomes in adipocytes. On the other hand, lyso- somes appear to be less important for receptor- mediated insulin degradation in agreement with previous studies [22].
129
Primary amines and dansylcadaverine are in- hibitors of transglutaminase and this has been thought to be the reason for their inhibition of ct2-macroglobulin uptake in fibroblasts [19,21]. The drugs are proposed to inhibit clustering of the ligand in coated regions [19]. An alternative, and perhaps more likely, explanation is that such drugs inhibit recycling of the receptors in the endocytic vesicles to the cell surface [26,27]. This may in turn be related to an inhibition of transglutaminase, to acidification of some vesicles, which has been shown to occur before their fusion with lysosomes [24], or to unknown mechanisms. In any case, primary amines, such as methylamine, exhibit strikingly different effects on the uptake of the two ligands (Fig. 3).
In conclusion, the data show very different effects of several inhibitors on the uptake and degradation of insulin on one hand and a2-macro- globulin- trypsin on the other. This indicates that the pathways for internalization of the two ligands are functionally different.
Acknowledgements
This study was supported by the Nordic Insulin Foundation and the NOVO Foundation. Dr. L. Sottrup-Jensen is thanked for providing us with human a2-macroglobulin.
References
1 Maxfield, F.R., Schlessinger, J., Shechter, Y., Pastan, I. and Willingham, M.C. (1978) Cell 14, 805-810
2 Shechter, Y., Schlessinger, J., Jacobs, S., Chang, K.-J. and Cuatrecasas, P. (1978) Proc. Natl. Acad. Sci. USA 75, 2135-2139
3 Gliemann, J., Sonne, O., Linde, S. and Hansen, B. (1979) Biochem. Biophys. Res. Commun. 87, 1183-1190
4 Kahn, C.R. and Baird, K. (1978) J. Biol. Chem. 253, 4900-4906
50lefsky, J.M. and Kao, M. (1982) J. Biol. Chem. 257, 8667-8673
6 Sonne, O., Linde, S., Larsen, T.R. and Gliemann, J. (1983) Hoppe-Seyler's Z. Physiol. Chem. 364, 101-110
7 Gliemann, J., Larsen, T.R. and Sottrup-Jensen, L. (1983) Biochim. Biophys. Acta 756, 230-237
8 Gliemann, J. and Sonne, O. (1978) J. Biol. Chem. 253, 7857-7863
9 Sottrup-Jensen, L., Petersen, T.E. and Magnusson, S. (1980) FEBS Lett. 121,275-279

130
10 Linde, S., Hansen, B., Sonne, O., Hoist, J.J. and Gliemann, J. (1981) Diabetes 30, 1-8
11 Foley, J.E., Foley, R. and Gliemann, J. (1980) Biochim. Biophys. Acta 599, 689-698
12 Foley, J.E., Laursen, A.L., Sonne, O. and Gliemann, J. (1980) Diabetologia 19, 234-241
13 Harford, J., Wolkoff, A.W., Ashwell, G. and Klausner, R.D. (1983) J. Cell Biol. 96, 1824-1828
14 Marshall, S. and Olefsky, J.M. (1979) J. Biol. Chem. 254, 10153-10160
15 Smith, R.M. and Jarett, L. (1982) Proc. Natl. Acad. Sci. USA 79, 7302-7306
16 Umezawa, H. (1976) Methods Enzymol. 45, 678-695 17 DeDuve, C., Barsy, T., Poole, B., Trouet, A., Tulkens, P.
and Van Hoof, F. (1974) Biochem. Pharmacol. 23, 2495-2531
18 Ohkuma, S. and Poole, B. (1978) Proc. Natl. Acad. Sci. USA 75, 3327-3331
19 Davies, P.J.A., Davies, D.R., Levitzki, A., Maxfield, F.R., Milhaud, P., Willlingham, M.C. and Pastan. I.H. (1980) Nature 283, 162-167
20 Maxfield, F.R., Davies, P.J.A., Klempner, L.K., Wil- lingham, M.C. and Pastan, I. (1979) Proc. Natl. Acad. Sci. USA 76, 5731-5735
21 Dickson, R.B., Willingham, M.C. and Pastan, 1. (1981) J. Biol. Chem. 256, 3454-3459
22 Sonne, O. and Gliemann, J. (1983) Mol. Cell. Endocrinol. 31,315-331
23 Van Leuven, F., Maryman, P. Cassiman, J.J. and Van den Berghe, H. (1981) FEBS Lett. 134, 83-87
24 Tycko, B. and Maxfield, F.R. (1982) Cell 28, 643-651 25 Kaplan, J. and Nielsen, M.L. (1979) J. Biol. Chem. 254,
7329-7335 26 Van Leuven, F., Cassiman, J.J. and Van den Berghe, H.
(1980) Cell 20, 37-43 27 Van Leuven, F. (1984) Mol. Cell. Biochem. 58, 121-128





























