Embed Size (px)
Citation preview

Variabilidad ambiental y recursos marinos
Lat. Am. J. Aquat. Res., 41(1): 1-28, 2013 DOI: 103856/vol41-issue1-fulltext-1
Review
Variabilidad ambiental y recursos pesqueros en el Pacífico suroriental: estado
de la investigación y desafíos para el manejo pesquero
Carolina Parada1,2, Beatriz Yannicelli3, Samuel Hormazábal4, Sebastián Vásquez1,5 Javier Porobić5, Billy Ernst6, Claudio Gatica1, Marcos Arteaga1, Aldo Montecinos2
Sergio Núñez1 & Alexandre Gretchina1
1Instituto de Investigación Pesquera, Avda. Cristóbal Colón 2780, Talcahuano, Chile 2Departamento de Geofísica, Universidad de Concepción, P.O. Box 160-C, Concepción, Chile
3Centro de Estudios Avanzados en Zonas Áridas (CEAZA), Facultad de Ciencias del Mar Universidad Católica del Norte, P.O. Box 117, Coquimbo, Chile
4Escuela de Ciencias del Mar, Pontificia Universidad Católica de Valparaíso P.O. Box 1020, Valparaíso, Chile
5Programa de Magíster en Ciencias mención Pesquerías, Departamento de Oceanografía Universidad de Concepción, P.O. Box 160-C, Concepción, Chile
6Sección Pesquerías, Departamento de Oceanografía, Universidad de Concepción P.O. Box 160-C, Concepción, Chile
RESUMEN. Las fluctuaciones en abundancia, biomasa, estructura de edad y patrones de distribución de los recursos pesqueros responden, entre otros, a la variabilidad ambiental. Estas respuestas son consecuencia tanto de efectos climáticos directos sobre los recursos como indirectos actuando sobre niveles tróficos relacionados. En este estudio se revisa: i) el estado del conocimiento de los mecanismos físicos asociados a la variabilidad océano-atmósfera y las escalas de variabilidad espaciales y temporales del ambiente y la relación con recursos marinos, basados en datos observacionales y modelos hidrodinámicos, ii) la relación ambiente-recurso para pesquerías pelágicas, demersales y bentónicas, y el uso de modelos biofísicos para entender estas relaciones, iii) manejo actual de recursos pelágicos, demersales y bentónicos y los alcances en relación al uso de variables ambientales, y iv) se discuten los desafíos hacia la asesoría en relación a las estrategias para mejorar la comprensión de la relación ambiente-recurso, así como, las estrategias para incorporar la modelación biofísica y variables ambientales en modelos operacionales para la asesoría hacia el manejo. Palabras clave: variabilidad ambiental, sistemas de surgencia, recursos pesqueros, modelación biofísica, dinámica poblacional, modelos operacionales para el manejo pesquero, Pacífico suroriental, Chile.
Environmental variability and fisheries in the southeastern Pacific:
research status and challenges for fisheries management
ABSTRACT. Fluctuations in abundance, biomass, age structure and distribution patterns of fisheries often respond to environmental variability. These responses are a result of both direct climatic effects on resources and indirect action on related trophic levels. In this study we review: i) the state of knowledge about the physical mechanisms associated with ocean-atmosphere variability and scales of spatial and temporal variability of the environment and related marine resources, based on observational data and hydrodynamic models, ii) environment relationship with pelagic, demersal and benthic fisheries, and the use of biophysical models to understand these relationships, iii) the current management of pelagic, demersal and benthic resources and the scope in relation to the use of environmental variables, and iv) strategies to improve understanding of the resource-environment relationship, as well as strategies for modeling biophysical and environmental variables in operational models for advice towards management are discussed. Keywords: environmental variability, fishing resources, biophysical modeling, population dynamics, operational models for management, southeastern Pacific, Chile.
___________________ Corresponding author: Carolina Parada ([email protected])
1

Latin American Journal of Aquatic Research
INTRODUCCIÓN
Las fluctuaciones en abundancia, biomasa, estructura de edad y patrones de distribución de los recursos pesqueros responden, entre otros, a la variabilidad ambiental. Esas respuestas son consecuencia tanto de efectos climáticos y oceanográficos (viento, tur-bulencia, concentración de oxígeno, temperatura) directos sobre los recursos (i.e., sincronización de los períodos reproductivos, retención/advección larval) como indirectos, mediados por cambios inducidos en otros niveles tróficos relacionados (i.e., alteración en la cantidad y calidad de la oferta de alimento de adultos y larvas, mecanismos de control del ecosistema). Sin embargo, aun se desconoce cómo estos procesos interactúan entre sí (Werner et al., 2004; Ottersen et al., 2010), ni de qué manera afectan las cadenas tróficas (Yoo et al., 2008) y en último término el ecosistema (Cury & Shannon, 2004; Kenny et al., 2009). Se entrega una revisión general de las relaciones ambiente-recurso a diferentes escalas espaciales y de los aspectos específicos que han sido abordados para los recursos objetivos de las pesque-rías chilenas en el Sistema de la Corriente de Humboldt (Fig. 1, Tabla 1). A pesar que los factores ambientales modulan los procesos biológicos durante todo el ciclo de vida de los recursos, en un importante número de estudios destaca el impacto poblacional que ejercen sus fluctuaciones actuando directamente sobre los estadios de vida temprana y el reclutamiento (Leggett & DeBlois, 1994). No obstante, los mecanismos involucrados en la fluctuación interanual del recluta-miento, en la mayor parte de los recursos, sigue siendo incierto. La incorporación explícita de factores ambientales en el proceso de asesoría para el manejo considera dos etapas: i) sintetizar el conocimiento multidisciplinario empírico para vincular los procesos físicos con las respuestas biológicas, y ii) desarrollar modelos numéricos o estadísticos e implementar modelos operativos capaces de evaluar alternativas de manejo en un ambiente variable. En Chile, destacan tres investigaciones en esta línea, en las cuales se han aplicado modelos de producción global para entender las capturas de anchoveta (Freón & Yáñez, 1995), modelos de redes neuronales para explicar las capturas de sardina en el norte de Chile asociadas a El Niño (Gutiérrez-Estrada et al., 2009), y el uso de escenarios de cambio climático (E. Yañez, com. pers.).
En las últimas décadas, se reconocen principal-mente dos aproximaciones al estudio de las relaciones ambiente-recurso. La primera se basa en la búsqueda de correlaciones entre variables ambientales y temporales de abundancia poblacional (Bakun, 2010),
construidas a partir de modelos poblacionales denso-dependientes en equilibrio, y/o al establecimiento de relaciones multivariadas con variables demográficas asociadas a los recursos marinos. Sin embargo, los proxies ambientales genéricos (e.g., TSM) muchas veces están desacoplados de las condiciones reales asociadas a los hábitats característicos de los recursos y sus estadios de vida específicos, generando rela-ciones ambiente-recurso difusas y de escaso valor predictivo. La segunda aproximación es la modelación biofísica, la cual se compone en sus versiones más simples de dos partes principales (Fig. 2): un modelo hidrodinámico que describe los procesos océano-gráficos a una escala espacio-temporal adecuada al recurso y a la etapa del ciclo de vida a modelar; y un modelo biológico que describe los procesos y cambios que afectan al individuo durante su ontogenia (sobrevivencia, movimiento, crecimiento, procesos bioenergéticos). En estos modelos la dinámica poblacional es el resultado del destino y evolución de individuos en su ambiente al cual están ligados por relaciones funcionales. Esta aproximación ha permi-tido mejorar la comprensión de la variabilidad temporal y espacial de los recursos marinos y sus relaciones con el ambiente aportando significa-tivamente a las ciencias pesqueras (Brochier et al., 2008; Xue et al., 2008; Parada et al., 2010a), así como también aparece como la aproximación más prome-tedora hacia el asesoramiento pesquero.
El mejoramiento sistemático en los modelos hidrodinámicos ha permitido que los modelos biofísicos se transformen en herramientas poderosas para el estudio de la dinámica de recursos marinos pelágicos y bentónicos, donde sus resultados pasan a ser insumos para la evaluación de estrategias de manejo que incluyen fuentes de información de diferentes áreas (Fulton et al., 2007; Melbourne-Thomas et al., 2011). Los modelos biofísicos han estado mayormente enfocados en la puesta a prueba de hipótesis de trabajo acerca del entendimiento de procesos biofísicos espacio-temporales, transporte y conectividad larval, análisis de retención, asenta-miento y éxito de pre-reclutas, exploración de zonas de crianza, entendimiento de la historia de vida (Hermann et al., 1996; Hinckley et al., 1996; Parada et al., 2010a, 2010b) y, últimamente, la puesta a prueba de hipótesis relacionadas con el cambio climático (Lett et al., 2009; Munday et al., 2009). Esta aproximación proporciona un potente enfoque para el estudio ecológico de las poblaciones, la evaluación estadística de hipótesis relacionadas con la conectividad poblacional (Cowen et al., 2000) y la evaluación de los efectos que generaría el cambio climático sobre la dispersión, conectividad y variabilidad espacio-temporal de las poblaciones
2

Variabilidad ambiental y recursos marinos
Figura 1. Escalas temporales, procesos físicos y biológicos asociados. Modificado de tabla generada en el marco del Grupo de Trabajo (E. Acuña, S. Hormazábal, J.L. Blanco, C.Parada) de Variabilidad Ambiental y Recursos, entregado oficialmente a la Subsecretaría de Pesca. marinas (Lett et al., 2010). Los modelos iniciales puramente lagrangeanos (Paris et al., 2002; Di Franco & Guidetti, 2011), evolucionaron para incorporar mayor complejidad biológica, de forma de poder incluir procesos evolutivos y bioenergéticos (Megrey & Hinckley, 2001). Estos últimos, requieren además de la implementación de modelos de bajo nivel trófico o biogeoquímicos acoplados (i.e., Modelos NPZ: nitrógeno, fitoplancton, zooplancton). El acoplamiento de los modelos biofísicos con los de bajo nivel trófico es esencial para evaluar la modulación indirecta que ejerce el ambiente (disponibilidad de alimento) sobre
los estadios tempranos, implementación que dista de ser trivial. En un nivel de complejidad mayor, modelos multiespecíficos, que consideran el ciclo de vida completo de peces, han sido desarrollados para utilizar salidas de modelos de bajo nivel trófico acoplados, permitiendo evaluar la modulación am-biental de organismos de niveles tróficos superiores, y sus interacciones (Travers & Shin, 2010). En términos operacionales, los modelos biofísicos acoplados a modelos de nivel trófico inferior pueden ser utilizados en el asesoramiento como indicadores independientes del reclutamiento, implementaciones explícitas de
3
Latin American Journal of Aquatic Research
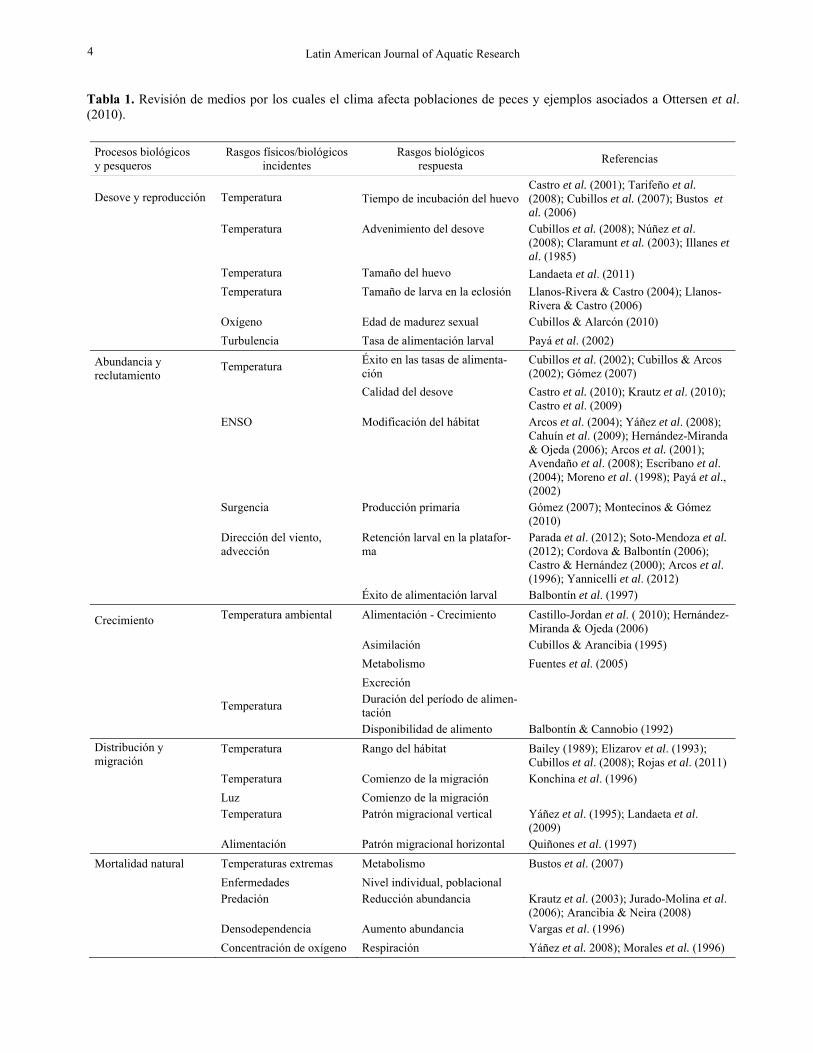
Tabla 1. Revisión de medios por los cuales el clima afecta poblaciones de peces y ejemplos asociados a Ottersen et al. (2010).
Procesos biológicos y pesqueros
Rasgos físicos/biológicos incidentes
Rasgos biológicos respuesta Referencias
Desove y reproducción Temperatura Tiempo de incubación del huevoCastro et al. (2001); Tarifeño et al. (2008); Cubillos et al. (2007); Bustos et al. (2006)
Temperatura Advenimiento del desove Cubillos et al. (2008); Núñez et al.
(2008); Claramunt et al. (2003); Illanes et al. (1985)
Temperatura Tamaño del huevo Landaeta et al. (2011)
Temperatura Tamaño de larva en la eclosión Llanos-Rivera & Castro (2004); Llanos-Rivera & Castro (2006)
Oxígeno Edad de madurez sexual Cubillos & Alarcón (2010) Turbulencia Tasa de alimentación larval Payá et al. (2002)
Abundancia y reclutamiento
Temperatura Éxito en las tasas de alimenta-ción
Cubillos et al. (2002); Cubillos & Arcos (2002); Gómez (2007)
Calidad del desove Castro et al. (2010); Krautz et al. (2010); Castro et al. (2009)
ENSO Modificación del hábitat Arcos et al. (2004); Yáñez et al. (2008); Cahuín et al. (2009); Hernández-Miranda & Ojeda (2006); Arcos et al. (2001); Avendaño et al. (2008); Escribano et al. (2004); Moreno et al. (1998); Payá et al., (2002)
Surgencia Producción primaria Gómez (2007); Montecinos & Gómez (2010)
Dirección del viento, advección
Retención larval en la platafor-ma
Parada et al. (2012); Soto-Mendoza et al. (2012); Cordova & Balbontín (2006); Castro & Hernández (2000); Arcos et al. (1996); Yannicelli et al. (2012)
Éxito de alimentación larval Balbontín et al. (1997)
Crecimiento Temperatura ambiental Alimentación - Crecimiento Castillo-Jordan et al. ( 2010); Hernández-Miranda & Ojeda (2006)
Asimilación Cubillos & Arancibia (1995) Metabolismo Fuentes et al. (2005) Excreción
Temperatura Duración del período de alimen-tación
Disponibilidad de alimento Balbontín & Cannobio (1992) Distribución y migración
Temperatura Rango del hábitat Bailey (1989); Elizarov et al. (1993); Cubillos et al. (2008); Rojas et al. (2011)
Temperatura Comienzo de la migración Konchina et al. (1996) Luz Comienzo de la migración
Temperatura Patrón migracional vertical Yáñez et al. (1995); Landaeta et al. (2009)
Alimentación Patrón migracional horizontal Quiñones et al. (1997) Mortalidad natural Temperaturas extremas Metabolismo Bustos et al. (2007) Enfermedades Nivel individual, poblacional
Predación Reducción abundancia Krautz et al. (2003); Jurado-Molina et al. (2006); Arancibia & Neira (2008)
Densodependencia Aumento abundancia Vargas et al. (1996) Concentración de oxígeno Respiración Yáñez et al. 2008); Morales et al. (1996)
4

Variabilidad ambiental y recursos marinos
Data de cruceros e
información satelital
Piezas de información
de IR
Indices de reclutamiento (IR)
biofísico independiente
Variables dinámicas de salida
(u,v,w,sal,temp,zeta, chla, phyto, zoo)
Modelación hidrodinámica
Condiciones de borde Condiciones Iniciales
ROMS
Grillas Forzantes
Validación
Modelo Biofísico
Submodelos Desove
Crecimiento Movimeinto
Alimentación
Forzantes
Estadísticos de contrastación Autocorrelación, correlación,
correlación cruzada, Funciones ortogonales empíricas, wavelets
Modelo de dinámica
Evaluación hidroacústica
Pesquería
Lineamientos hacia la asesoría
Transporte Conectividad Zonas
de crianza
Constrastación IRsA
copl
amie
nto
Figura 2. Sistema de modelación biofísica constituido por un modelo hidrodinámico y un modelo biológico y generación de índices independientes de la pesquería que podrían servir para generar lineamientos hacia la asesoría. variables biofísicas simuladas en modelos de dinámica, o como una componente en modelos ecosis-témicos operativos.
El objetivo de este trabajo es realizar una revisión del estado actual del conocimiento de las escalas de variabilidad del clima, del sistema oceánico, de la relación ambiente-recurso para pesquerías pelágicas, demersales y bentónicas del Pacífico suroriental, y el uso de modelos biofísicos para entender esta relación. Por otro lado, se realiza una revisión del manejo actual de algunos recursos pelágicos, demersales y bentónicos. Se identifican los desafíos que enfrenta la investigación pesquera en Chile y las posibles estrategias para la incorporación de variables ambientales en la estructura de modelos operacionales (biofísicos), que permitan apoyar el manejo de recursos pesqueros.
Revisión de la relación ambiente-recurso para pesquerías pelágicas, demersales y bentónicas en el Pacífico suroriental Escalas y forzantes de variabilidad climática y océa-nográfica en el Sistema de la Corriente de Humboldt El acoplamiento océano-atmósfera modula la inten-sidad de los procesos dinámicos y termodinámicos que operan en diversas escalas, determinando las principales características del océano en un ecosistema particular (Fig. 1). Por tanto, los modos climáticos de variabilidad (patrones de anomalías de sistemas de circulación atmosféricos), modulan las condiciones
oceanográficas que afectan directa y/o indirectamente desde los niveles de organización biológica pobla-cionales a los ecosistémicos. Los ciclos biológicos de las especies explotadas se encuentran mayorita-riamente acoplados al ciclo anual del ambiente físico y es a esas escalas que se han explorado las relaciones ambiente-recurso. El modo climático de variabilidad interanual más prominente es El Niño-Oscilación del Sur (ENOS) que corresponde a un proceso acoplado océano-atmósfera que se desarrolla en el Pacífico tropical, pero que tiene impactos en amplias regiones del planeta, a través de patrones de teleconexión tanto atmosféricas como oceánicas (Aceituno, 1988; Sarachik & Cane, 2010). Abruptos cambios climáticos han sido descritos como parte de oscilaciones en la escala interdecadal, llevando a definir oscilaciones climáticas conocidas como Pacific Decadal Oscillation (PDO) y North Pacific Oscillation (NPO) (Mantua et al., 1997; Salinger et al., 2001).
Estas oscilaciones muestran al Pacífico norte como la región donde esta señal se desarrolla con mayor intensidad. Sin embargo, existen evidencias que sugieren que la oscilación interdecadal posee un patrón asimétrico en la variabilidad del clima del hemisferio norte y sur, brindando importancia al estudio de las características de estas fluctuaciones en el hemisferio sur (Garreaud & Battisti, 1999). En este último, en latitudes altas, el modo de variabilidad climática más prominente corresponde a la Oscilación Antártica (AAO), la cual presenta fluctuaciones en
5
funciones
Movimiento
Conectividad, zonas

Latin American Journal of Aquatic Research
escalas interanual e interdecadal que, a su vez generan fluctuaciones en la presión atmosférica, campo de viento, temperatura del mar, además de generar perturbaciones sobre los vientos de superficie en latitudes medias (Thompson & Solomon, 2002; Fyfe, 2003). El efecto de la AAO sobre la variabilidad climática en latitudes medias, particularmente en el Pacífico suroriental, ha sido menos estudiada que para ENSO, aunque han habido avances en su relación con la precipitación (Quintana, 2004; Gillet et al., 2006; Garreaud et al., 2008). Todos estos modos de variabilidad climática son reconocidos por sus impactos en el sistema climático del Pacífico suroriental. Sin embargo, a pesar de que se ha mejorado el conocimiento sobre este tema, aun existen muchas interrogantes sobre los mecanismos respon-sables de la variabilidad climática y el impacto de estos procesos sobre los recursos marinos en esta región.
La variabilidad del ambiente en el Pacífico suroriental está determinada principalmente por la acción combinada de: a) fluctuaciones, desde sinópticas a intraestacionales, ligadas al proceso de surgencia costera (Strub et al., 1998; Leth & Shaffer, 2001; Rutllant et al., 2004), b) variabilidad interanual asociada a la alternancia de eventos El Niño y La Niña (Shaffer et al., 1999; Hormazábal et al., 2001; Escribano et al., 2004), incluyendo ondas de Kelvin ecuatoriales, ondas atrapadas a la costa y ondas de Rossby (Pizarro, 1991, 1999; Strub et al., 1998; Fuenzalida et al., 2008), y c) fluctuaciones interdecadales (Klyashtorin, 1998; McFarlane et al., 2000; Yáñez et al., 2001). Se ha sugerido que este tipo de variabilidad, en conjunto con la dinámica de estructuras de mesoescala (remolinos y meandros, Nakata et al., 2000; Hormazábal et al., 2004; Correa-Ramírez et al., 2007), pueden causar cambios significativos de producción y abundancia, tanto en el océano costero como en alta mar que pudiesen afectar la distribución y abundancia de recursos pesqueros. Dentro de la variabilidad climática asociada a ENOS en el Pacífico ecuatorial, las oscilaciones de Madden-Julian son la fuente principal de la generación de ondas de Kelvin intraestacionales (30-90 días) (Zhang, 2005; Xu et al., 2005). Durante El Niño, las fluctuaciones de los vientos alisios, asociadas a las Madden-Julian, fuerzan ondas Kelvin oceánicas que viajan por el ecuador hacia la costa de Sudamérica.
Una vez que estas ondas llegan a la costa, parte de su energía se propaga hacia el polo como ondas atrapadas a la costa y otra parte se refleja hacia el oeste como ondas de Rossby. Mediante observaciones directas y resultados de modelos numéricos se ha
mostrado que las corrientes, nivel del mar e isotermas bajo la termoclina muestran oscilaciones en la escala intraestacional hasta los 33°S (Shaffer et al., 1997; Hormazábal et al., 2002). Al sur de los 33ºS, el viento a lo largo de la costa aparece contribuyendo en forma significativa al forzamiento de ondas intraestacionales atrapadas a la costa (Hormazábal et al., 2002; Leth & Middleton, 2006). Por otra parte, se han observado oscilaciones semianuales e interanuales asociadas a ondas de Rossby forzadas por ondas de Kelvin ecuatoriales (Pizarro et al., 2001; Vega et al., 2003; Hormázabal et al., 2004).
El ENOS afecta en forma diferente al esfuerzo del viento a lo largo de la costa de Sudamérica. Asociados al ENOS, en la zona centro-sur de Chile, se han observado patrones de teleconexión atmosféricos que afectan el período de inicio y final de la estación de surgencia (Montecinos & Gómez, 2010) y, a través de éste, la concentración de clorofila y el reclutamiento de la sardina común (Gómez et al., 2012). Sin embargo, en la zona norte de Chile, no se observa un efecto significativo del ENOS sobre el viento costero (Montecinos, 1991), a diferencia de lo observado frente a Perú (Halpern, 2002).
En la zona norte de Chile, se ha observado que la variabilidad del nivel del mar, temperatura superficial del mar, salinidad, oxígeno disuelto y profundidad de las isotermas, se encuentra más relacionada con ondas oceánicas forzadas por los vientos ecuatoriales que con el esfuerzo del viento costero (Morales et al., 1996; Ulloa et al., 2001; Gómez, 2007). En esta zona las ondas atrapadas a la costa y las ondas de Rossby, forzadas ecuatorialmente, profundizan (levantan) la interfase termoclina-oxiclina-nutriclina, expandiendo (comprimiendo) el hábitat vertical de la anchoveta (Engraulis ringens), modulando su distribución vertical.
Recursos pesqueros y variabilidad ambiental Pequeños pelágicos y variabilidad ambiental Los desembarques artesanales e industriales repor-tados para anchoveta (Engraulis ringens) ascienden a 41.000 ton, para sardina común (Strangomera bentin-cki) 470.000 ton en la zona centro-sur de Chile durante el primer semestre del año 2012 y, para sardina española (Sardinops sagax), 159 ton en el mismo período en la zona norte de Chile (SUBPESCA, 2012). Debido a los rangos distribucionales de las especies de sardina y de los stocks de anchoveta (Ferrada et al., 2002), se desarrollan pesquerías independientes en la zona norte del país (Arica-Antofagasta) constituida por anchoveta y sardina española, y en la zona centro-sur
6

Variabilidad ambiental y recursos marinos
(Valparaíso-Valdivia) por sardina común y anchoveta (Serra, 1978; Arrizaga, 1981). Estas especies habitan a lo largo de la costa hasta las 30 mn, presentando una componente estacional en el crecimiento (Feltrim & Ernst, 2010), reclutamiento (Cubillos & Arcos, 2002) y pesca (Feltrim, 2009), asociada a la zona de influencia de surgencia costera y mayor productividad primaria durante la primavera-verano (Arcos et al., 1987; Yuras et al., 2005).
Zona norte Los pequeños pelágicos del norte de Chile destacan por ser los recursos más estudiados a nivel nacional en función de las relaciones ambiente-recurso. Grandes fluctuaciones de sardina española y anchoveta alternan en desfase a escala interdecadal, manifestándose directamente sobre los niveles de captura. No obstante, se ha verificado que no existen fases consecutivas en la dominancia de su abundancia, sino más bien como resultado de un proceso estocástico ligado a la variabilidad ambiental (Gutiérrez-Estrada et al., 2009). Dos décadas de estudio fueron sintetizadas en un modelo conceptual de variabilidad de pelágicos-ambiente a diferentes escalas por Yáñez et al. (2008). El modelo describe las fluctuaciones interdecadales asociadas a cambios de régimen, e interanuales asociadas a los eventos El Niño del Pacífico suroriental. En la zona norte de Chile, se observan efectos en el ciclo anual, la dinámica de las ondas atrapadas a la costa y la surgencia costera (Yañez et al., 2008). En este marco, las fluctuaciones interdecadales juegan un rol importante en la secuencia del remplazo anchoveta-sardina española-anchoveta. Las fluctuaciones del stock de anchoveta del sur de Perú y norte de Chile, están en sincronía con las del stock del norte-centro de Perú (Cubillos et al., 2007). Los períodos de altos desembarques de anchoveta (ambas escalas), obedecen tanto a incrementos poblacionales (cambios en termoclina-nutriclina modulan poblaciones plantónicas), como a cambios de capturabilidad (aumenta con reducción de profundidad de la oxiclina, disminuyendo el hábitat de fracción adulta). Ambos factores determinantes de la sobrevivencia operan en forma inversa para anchoveta y sardina española. Existe evidencia de que el ingreso de aguas subtropicales afecta la distribución horizontal de anchoveta frente a Perú y norte de Chile, haciéndose más costeras (Yáñez et al., 2001; Bertrand et al., 2004) y más australes (Yáñez et al., 1995; Ñiquén & Bouchon, 2004). Por otro lado, un descenso de la actividad reproductiva observado mediante el índice gona-dosomático, y una disminución de la abundancia de la fracción juvenil, se relacionan con el efecto de El Niño (Ñiquén & Bouchon, 2004). Por otra
parte, durante períodos cálidos, se ha observado un cambio en la estructura de la comunidad planctónica (Alheit & Ñiquén, 2004), con una reducción de la abundancia de copépodos y eufáusidos (González et al., 1998). Estas obser-vaciones pesqueras (registro de los últimos 40 años) contrastan con lo señalado por Valdés et al., 2008, quienes encontraron respuestas no alternantes en algunos períodos de los 250 años reconstruidos mediante el análisis de escamas en depositación. Es decir, ambas especies se hallaron en momentos positivos de abundancia, a veces una dominando sobre la otra. Esto sugiere que la modulación en las abundancias entre sardina y anchoveta es gobernada por otros procesos oceano-climáticos especie-específicos no claramente identificados. Todos estos antecedentes sugieren que estudios ecosistémicos que involucren variabilidad del ambiente más la interacción de múltiples niveles tróficos son necesarios para el entendimiento de la variabilidad de los stocks de pequeños pelágicos.
Zona centro-sur La sardina común y la anchoveta sustentan una importante pesquería mixta en la zona centro-sur de Chile y juegan un rol clave en la dinámica trófica del sistema, transfiriendo energía desde los productores primarios hacia los eslabones superiores de la trama trófica. Ambas especies presentan un ciclo de vida corto y rápido crecimiento, con desoves centrados a fines del invierno austral y una longevidad no superior a 4 y 5 años (sardina y anchoveta respectivamente; Cubillos et al., 1999). Otro rasgo característico de estas poblaciones es la marcada fluctuación en el tamaño de los stocks, altamente dependiente de la fuerza del pulso de reclutamiento anual (Gatica et al., 2007). Especies de pequeños pelágicos, que habitan sistemas de surgencia costera, son especialmente sensibles a la variabilidad del ambiente en todas las etapas de su ciclo vital (Cury et al., 2000).
A escala interdecadal, la pesquería de anchoveta centro-sur ha fluctuado en sincronía y fase con las pesquerías Chile N-Perú S y Perú N-centro (1960-2000). Sin embargo, a escala interanual, el patrón difiere y las anomalías de abundancia no se correlacionan con índices El Niño (Cubillos et al., 2007). La sardina común fluctúa en sincronía y fase con la anchoveta a escala interdecadal, pero a escala interanual, esta sí se correlaciona con índices de El Niño ecuatoriales y anomalías de temperatura locales. Estos patrones no han sido esclarecidos al momento, y las relaciones propuestas (Arcos et al., 2004) deben ser revisadas críticamente a la luz de nuevos resultados que asocian la condición materna y la calidad de huevos con la variabilidad ambiental en la
7

Latin American Journal of Aquatic Research
plataforma continental de Chile centro-sur (Castro et al., 2009). Así mismo, evaluar las diferencias latitudinales en cuanto a la amplitud de las fluctua-ciones ambientales interanuales, y variaciones latitu-dinales en rasgos reproductivos (i.e., tamaño de huevo y consecuente disponibilidad de reservas de la larva recién eclosionada, Llanos-Rivera & Castro, 2004), pre-reclutas ayudar a interpretar las diferencias descritas entre patrones interanuales.
El reclutamiento de sardina común en la zona centro-sur de Chile, pareciera estar afectado por teleconexiones asociadas a El Niño y La Niña, que modifican considerablemente el ambiente nerítico. Durante eventos cálidos, los niveles de reclutamiento disminuyen en consistencia con una disminución en la disponibilidad de alimento para los pre-reclutas y a cambios en la estructura vertical de la concentración de oxígeno, generando una disminución en la sobrevivencia (Gómez, 2007). Este escenario propi-ciaría un aumento en la abundancia de anchoveta producto de una interacción biológica con las cohortes menos abundantes de sardina (Cubillos & Arcos, 2002). Por otra parte, años fríos (La Niña) están altamente relacionados con el ingreso de clases anuales de sardina, que podrían asociarse con el incremento de la producción biológica. A la fecha, la información sobre la relación entre la variabilidad ambiental y su impacto sobre pequeños pelágicos en la zona centro-sur es dispersa, lo que dificulta esbozar un modelo conceptual de manejo ambientalmente explí-cito. Sin embargo, la integración y síntesis de información ambiental, pesquera y biológica existente podría permitir plantear hipótesis más compre-hensivas, y evaluables, a través de esfuerzos de modelación.
Variabilidad ambiental, la estructura poblacional y el hábitat del jurel El jurel (Trachurus murphyi) cuya distribución geográfica abarca la región del Pacifico suroriental ha sostenido una de las pesquerías pelágicas más importantes en el mundo. En las últimas dos décadas los desembarques se han reducido desde un máximo de 4,4 millones de ton en 1994 a 1,2 millones en 1999, alcanzando sólo 190.000 ton en 2010. Existen dos hipótesis, probablemente no excluyentes, acerca del descenso de esta población, asociadas a la sobrepesca y a la variabilidad ambiental. Las regulaciones de manejo pesquero en Chile establecieron un mínimo legal en 26 cm en 1982, cerrando la entrada de nuevas embarcaciones en 2003. Actualmente, la Organización Regional de Pesca (SPRFMO, South Pacific Regional Fisheries Management Organization) en 2011, aplicó
una medida interina adoptada como una aproximación provisional para manejar el recurso jurel (http://www. southpacificrfmo.org/interim-measures/). Esta especie es altamente migratoria y ampliamente distribuida en las costas chilenas y peruanas, alcanzando Nueva Zelandia y Tasmania en el Pacífico sur (Bailey, 1989; Serra, 1991; Elizarov et al., 1993; Arcos & Gretchina, 1994; Gretchina et al., 1998, Arcos et al., 2001). Dos subpoblaciones han sido propuestas en base a estudios merísticos, en contraposición a hipótesis panmícticas (Serra, 1991). Estudios genéticos entregan nueva evidencia que apoya la hipótesis de una única población genética a través del rango distribucional de T. murphyi (Cárdenas et al., 2009). El período principal de desove está centrado en noviembre, extendiéndose desde octubre a diciembre (Gretchina et al., 1998; Oyarzún et al., 1998). Durante este período, se dispersan las agregaciones de jurel (sin formación de cardúmenes) (Konchina et al., 1996; Barbieri et al., 1999), lo cual impide su detección acústica (Gatica et al., 2003). El hábitat de desove coincide con la región de alta variabilidad de mesoescala del frente subtropical (alrededor de los 35ºS), donde convergen aguas subtropicales cálidas y de alta salinidad con aguas más frías y menos salinas de origen subantártico (Chaigneau & Pizarro, 2005). Posterior a la repro-ducción, dos procesos co-ocurren: una migración estacional hacia la costa a hábitats de alimentación, donde se sugiere que la disponibilidad de alimento es el factor determinante de la migración (Cornejo-Rodríguez, 1991; Quiñones et al., 1997; Miranda et al., 1998), como también el transporte/ advección de huevos y larvas eclosionadas hacia la zona disponible de crianza (Norte de 30ºS, Elizarov et al., 1993; Gretchina et al., 1998; Arcos et al., 2001). Estudios de química de otolitos (Ashford et al., 2010), de estructura de edad de larvas de jurel océano-costa, y su asociación a procesos oceanográficos (Vásquez, 2012), sustentan la hipótesis de conectividad entre las zonas de desove oceánico y de crianza. A lo largo de la costa de Perú y Chile, las inestabilidades baroclínicas gatillan una alta actividad de mesoescala, con remolinos que se propagan hacia el oeste (Hormazábal et al., 2004), con contribución de clorofila hacia fuera de la costa (Correa-Ramírez et al., 2007), en sentido inverso a la dirección de conectividad con zonas de crianza costera.
A la fecha, la información ha permitido esbozar modelos conceptuales que han servido de base para poner a prueba hipótesis en relación a la historia de vida de este recurso (ver sección jurel y modelación biofísica). Sin embargo, aun quedan numerosas interrogantes, acerca de relaciones ambiente-recurso con la demografía y la variabilidad interanual del reclutamiento.
8

Variabilidad ambiental y recursos marinos
Recursos demersales y variabilidad ambiental El recurso merluza común (Merluccius gayi) habita en el sistema de corriente de Humboldt (Aguayo, 1995; Ojeda et al., 2001), que se caracteriza por su alta productividad e intensa variabilidad ambiental semi- e interanual, de la misma manera que especies congénericas en otros sistemas de borde oriental (Alheit & Pitcher, 1995). Este recurso constituye la pesquería demersal más importante de Chile (Aguayo, 1995; Payá et al., 2003), distribuyéndose desde Antofagasta (23º40’S) hasta el canal Cheap (47º00’S), con las capturas más importantes en la zona central de Chile (31º-41ºS) (Aguayo, 1995; Lillo et al., 2001), con una distribución batimétrica que varía principalmente entre 100-350 m de profundidad. Con posterioridad a la declinación de los desembarques (30.000 ton) ocurrida en 1976-1987, el recurso mostró una importante recuperación de los índices de abundancia y estructura poblacional durante 1987-2001, dando lugar a desembarques sobre 110.000 ton (Payá & Zuleta, 1999; Lillo et al., 2001). Sin embargo, desde 2004-2006 el recurso ha declinado, reportándose desembarques inferiores a 50.000 ton y una estructura poblacional desmejorada, lo que se ha atribuido principalmente a altos niveles de depredación por jibia (Dosidicus gigas, Payá, 2006), que han afectado tanto la abundancia como la disponibilidad del recurso (Gálvez, 2006), altas tasas de canibalismo (Jurado-Molina et al., 2006), una sobre-estimación del tamaño del stock en los años previos y consecuente sobre-explotación (Arancibia & Neira, 2008), a fluctuaciones ambientales en la escala interanual (Payá, 2006), y a los niveles de captura. Por otra parte, se ha indicado que su reclutamiento está estrechamente vinculado a las condiciones ambien-tales imperantes en la época de desove (fines de invierno, Balbontín & Fischer, 1981; Alarcón & Arancibia, 1993; Payá, 2006), observándose mayores reclutamientos cuando el desove se produce en años cálidos (Payá, 2006; Payá & Zuleta, 1999). Esta asociación se ha postulado como una explicación para los reclutamientos exitosos de los años 1982-1983, 1990-1993 y 1996-1997 (Payá, 2006), y ha sido informada también para Merluccius gayi peruanus frente a la costa de Perú (Espino & Wosnitza-Mendo, 1989), señalando una dispersión de las agregaciones que incrementan el área de distribución y disminuyen la competencia y el canibalismo, favoreciendo el proceso de reclutamiento. Aunque la merluza común desova en forma continúa todo el año, entre Antofagasta y Puerto Montt (Balbontín & Fischer, 1981), presenta dos máximos principales de desove, durante finales de invierno y principios de primavera (agosto-noviembre), asociados con frentes de
surgencia (Vargas & Castro, 2001), y un desove menor hacia finales de verano (marzo) (Cerna & Oyarzún, 1998; Alarcón et al., 2004). El desove de invierno-primavera ocurre entre 50-60 km fuera de la costa desde donde huevos y larvas tempranas son transportados por corrientes subsuperficiales hacia la costa, donde ocurre el asentamiento (Avilés et al., 1979; Vargas et al., 1996; Bernal et al., 1997). En el desove secundario, los adultos migran desde la plataforma hacia zonas someras donde desovan (Avilés et al., 1979; Arancibia, 1992). A pesar que esta pesquería demersal es de gran importancia económica, existe escasa información acerca de su historia de vida temprana (Landaeta & Castro, 2006), así como de las asociaciones ambientales relacionadas.
Recursos bentónicos y variabilidad ambiental A lo largo de la costa, plataforma continental, islas oceánicas y mar interior de Chile se extraen más de 60 especies de invertebrados (Anuario SERNAPESCA, 1992-2002), de hábitat bentónico en su mayoría (excepto algunos cefalópodos). Las relaciones ambiente-recurso en el caso de las pesquerías bentónicas de invertebrados se han abordado explícitamente en la zona norte de Chile (y Perú), donde se manifiestan respuestas positivas y negativas asociadas a El Niño (Arntz, 1986). A continuación se revisa el caso de algunas pesquerías bentónicas de la zona norte y central, y otras de amplio rango de distribución latitudinal, cuyas especies objetivo muestran variabilidad interanual conspicua.
Mesodesma donacium y variabilidad ambiental La macha (Mesodesma donacium) es un bivalvo filtrador que habita playas de arena expuestas intermedias-disipativas en el inter- y submareal, con desarrollo larval pelágico prolongado y caracterizado por altas fluctuaciones interanuales (Defeo & McLachland, 1995). En la zona norte de Chile y sur de Perú en particular, la desaparición de grandes poblaciones de macha ha coincidido con la ocurrencia de eventos El Niño intensos (Riascos et al., 2008; Carstensen et al., 2010), con aumentos en la temperatura superficial del mar y disminuciones en salinidad al norte de los 30°S. El rango de distribución de la macha entre los 5°N y la isla de Chiloé (Tarifeño, 1980), se redujo en su extremo norte en cerca de 20° (Arntz et al., 1987) luego de El Niño 1982-1983, mientras que durante El Niño 1997-1998 importantes poblaciones de Arica (~18°S) y Coquimbo (~29°55’S) fueron diezmadas, para posteriormente recuperarse (Aburto & Stotz, 2003; Stotz et al., 2005). Estudios de laboratorio y de campo sugieren que el aumento de temperatura (Riascos et
9

Latin American Journal of Aquatic Research
al., 2009), disminuciones en la salinidad en conjunto con la expansión del parásito Polydora occipitalis (afecta negativamente crecimiento y fecundidad de la macha; Riascos et al., 2011), generan altas tasas de mortalidad, responsables de la masiva desaparición de machas asociadas a El Niño en playas del sector norte de su rango de distribución. Sin embargo, la reducción poblacional durante El Niño parece responder al aumento de la carga de sedimentos y caudal aportado por el río aledaño, a consecuencia del régimen de lluvias durante este período (Aburto & Stotz, 2003). Así, un evento de gran escala espacial puede afectar negativamente las poblaciones locales a través de diferentes mecanismos, en un amplio rango latitudinal. Al momento de evaluar la variabilidad interanual de los recursos costeros, debe considerarse cómo los fenómenos de gran escala se manifiestan regional/ localmente, tanto en las condiciones marinas (i.e., temperatura, productividad marina, condiciones de retención) como en el ambiente terrestre aledaño (i.e., caudales y aportes terrígenos asociados a los usos del suelo).
Argopecten purpuratus y variabilidad ambiental El ostión del norte (Argopecten purpuratus) es un pectínido filtrador; hermafrodita facultativo, de fecundación externa y desarrollo larval planctotrófico pelágico; habita zonas submareales de sustrato blando (arenoso), en bahías someras a lo largo de la costa Pacífica, principalmente entre Paita, Perú (5ºS, 81ºW) hasta Valparaíso Chile (33ºS, 71º37’W) (Uribe et al., 1995). Contrariamente al caso de la macha, las poblaciones y capturas de ostión del norte se incrementan en más de un orden de magnitud tras eventos El Niño (Wolff, 1987). Durante los períodos de El Niño (82-83, 97-98), efectos tales como aumentos en la actividad reproductiva, mayores asentamientos, mayor crecimiento en juveniles y/o producción somática (dentro del rango 12-22°C, B. Yannicelli, pers. comm.), acortamiento del período larval y mayor sobrevivencia, se han descrito para las bahías Arenas, Independencia, Mejillones y Tongoy (Illanes et al., 1985; Tarazona et al., 2007; Avendaño et al., 2008). Otros factores, como la disminución de predadores y aumento de oxigenación de fondo, también potenciarían el éxito poblacional favore-ciendo la sobrevivencia de juveniles y adultos. Los factores más importantes en el crecimiento de los pectínidos son la temperatura y disponibilidad de alimento (Farías et al., 2003), tanto en su cantidad como calidad (Shumway et al., 1987; Orensanz et al., 1991), la cual juega un rol importante en el crecimiento y desarrollo de las etapas larvales y en etapas adultas de acondicionamiento reproductivo (Martínez et al., 2000; Navarro et al., 2000). El
alimento, más que la temperatura, afecta el potencial de crecimiento de adultos de A. purpuratus (Navarro et al., 2000) en el rango 16-20°C. El efecto ‘positivo’ de El Niño sobre A. purpuratus es claro, aunque los mecanismos que ligan dicha relación debe ser evaluada en más detalle. Las respuestas ante un evento de El Niño no sólo depende de la intensidad del mismo sino en la fase del ciclo anual en la cual se produce (se esperan mayores efectos cuando se sobrepone a los períodos reproductivos ‘normales’, Thatje et al., 2008) y variarán latitudinalmente. Por eso, es necesario cuantificar la importancia relativa de la sobrevivencia larval versus el potencial reproductivo en el aumento poblacional. El Niño se asocia a menores tasas y biomasas de productores primarios, pero también afecta su estructura de tamaños en la costa norte de Chile. Las larvas de A. purpuratus se encuentran en el rango de tallas de 20 a 200 µ, por lo que el incremento de fracciones de menor tamaño (Iriarte et al., 2000; Iriarte & González et al., 2004) podrían ser las relevantes para su éxito, aunque estas hipótesis no han sido analizadas.
Octopus mimus y variabilidad ambiental El pulpo (Octopus mimus) se distribuye desde el norte de Perú hasta Chile central. Se trata de una especie dioica con dimorfismo sexual marcado, semélpara y de ciclo de vida corto (Cortéz, 1995), que presenta un estadio larval de paralarva. La especie fue identificada en forma reciente (Guerra et al., 2009). Su pesquería se desarrolla a nivel artesanal en Perú y norte de Chile durante prácticamente todo el año (Rocha & Vega, 2003). Si bien la historia de capturas a nivel interanual refleja el incremento en la demanda del recurso (Defeo & Castilla 1998; Cardoso et al., 2004), aumentos marcados en las capturas totales de Perú se asocian a los eventos de El Niño (Cardoso et al. 2004), al igual que la expansión hacia el sur de las zonas donde se registran desembarques en Chile. Frente a las regiones de Atacama y Coquimbo, las capturas de pulpo se registran casi exclusivamente después de eventos El Niño. Se han realizado estudios de biología básica en terreno para sustentar decisiones de manejo tradicionales (Olivares et al., 1996) y en cultivo (Cortéz et al., 1999). Sin embargo, no existen estudios que aborden la causalidad de los patrones de captura observados y la ocurrencia de El Niño. Dada su condición de adultos carnívoros y vida larval pelágica, serían esperables efectos ambientales directos e indirectos (sobrevivencia larval, disponibilidad de alimentación bentónica entre otros). Se ha enfatizado la necesidad de evaluar los efectos de El Niño sobre los estadios larvales de invertebrados para comprender las respuestas poblacionales (Gaymer et al., 2010). Sin
10

Variabilidad ambiental y recursos marinos
embargo, en esta especie, no existe información disponible para dicho estadio.
Langostino colorado y variables ambientales Las pesquerías de crustáceos industriales más impor-tantes corresponden al camarón nylon (Heterocarpus reedi), langostino amarillo (Cervimunida johni) y langostino colorado (Pleuroncodes monodon). En ésta sección sólo se considerará el langostino colorado. El rango de distribución comúnmente citado para la especie va desde Lobos de Afuera (Perú, ~6°S) hasta Ancud (Chile, ~42°S) (Haig, 1955; Palma & Arana, 1997), donde estudios genéticos han confirmado que especímenes en ese rango pertenecen a la misma especie. Sin embargo, también se la ha citado para Centro América y Méjico. En la plataforma chilena (~22°S a 37°S), los adultos tienen hábitos bentónicos asociados a zonas hipóxicas, sobre plataforma y hasta ~500 m de profundidad, donde se agregan en sustratos de tipo arenoso, dominando la comunidad. En el norte de Chile y Perú, P. monodon es una especie clave en el hábitat pelágico, donde se distribuye por sobre la oxiclina (Gutiérrez et al., 2008). Este fenómeno es un caso de plasticidad fenotípica heterocrónica que sería gatillada por algún factor de estrés ambiental (Haye et al., 2010). A partir de El Niño de 1997, la biomasa de P. monodon pelágico frente a Perú ha aumentado y últimamente, su presencia en aguas costeras ha dejado de estar restringida a condiciones frías (Gutiérrez et al., 2011). Tanto en la plataforma frente a Chile (Palma, 1994; Canales et al., 2002; Yannicelli et al., 2012), como frente a Perú (Franco-Meléndez, 2012) P. monodon presenta períodos reproductivos exten-didos, una característica asociada a ambientes variables que tendería a favorecer la ocurrencia de reclutamientos más estables.
En Chile, la pesquería de arrastre de P. monodon está dividida en las zonas norte y sur. En el sur, se ha desarrollado desde los años sesenta entre 33º y 37ºS, ha sufrido fluctuaciones interanuales de biomasa total y rango de distribución latitudinal que han instando al cierre de la pesquería en varias oportunidades (Roa & Bahamonde, 1993). La historia de la pesquería en la zona norte (Coquimbo) es más reciente (1998). Los cambios de biomasa en el período 1990-2007 en la zona sur se explican por la variabilidad de recluta-miento anual a la pesquería, detectándose fallas de reclutamiento no asociadas a la abundancia del stock parental (Acuña et al., 2005) que, sumadas a altas tasas de explotación, han determinado caídas de la biomasa explotable después de 1998 (Acuña et al., 2009). La variabilidad del éxito de reclutamiento se debería a alteraciones de la fecundidad, sobrevivencia larval o sobrevivencia de juveniles post-asentamiento.
Recientemente se ha constatado que, contrariamente a lo asumido, la especie presenta múltiples puestas por hembra a lo largo del período reproductivo (M. Thiel, com. pers.) y, por tanto, que la fecundidad habría sido subestimada a lo largo de la historia de la pesquería. Subrayando la susceptibilidad de esta fase del ciclo de vida, Gallardo et al. (2004), sugirieron que la ocurrencia de colonias de la bacteria gigante Thioploca en la plataforma frente a Concepción, sería un refugio para juveniles de P. monodon. La disminución de la bacteria durante El Niño, producida por oxigenación del fondo, podría relacionarse a una mayor mortalidad juvenil y falla de reclutamiento post-El Niño. Esta hipótesis, sin embargo, no ha sido evaluada a pesar de existir series temporales que podrían permitir una primera evaluación. Por otro lado, estudios ecofisiológicos y metabólicos indicarían que las condiciones ambientales (temperatura y oxigeno), bajo las cuales eclosionan las larvas de P. monodon en Chile centro-sur, no se encuentran dentro del rango óptimo (B. Yannicelli, com. pers.)
La pesquería de la langosta de Juan Fernández y su forzamiento ambiental
La pesquería de la langosta de Juan Fernández (Jasus frontalis) es una de las pesquerías de crustáceos más antiguas de nuestro país y cuenta con registros de captura desde los años 1930's (Yáñez et al., 1985). Existen tres flotas que operan en este sistema: una en las islas Robinson Crusoe y Santa Clara (RC-SC), otra en la isla Alejandro Selkirk (AS), y una tercera dependiente de RC-SC, que opera esporádicamente en las islas Desventuradas (ID) (Arana, 1987). El proceso de pesca ocurre asociado a unidades geográficas discretas, conocidas como “Marcas”, las que funcionan como un sistema no escrito de tenencia de lugares discretos de pesca (Ernst et al., 2010a). Los registros pesqueros de J. frontalis muestran cambios significativos a través del tiempo, donde destaca un período de elevados desembarques, entre la década de los 1930’s y fines de los años 1960’s (con un valor medio cercano a las 90 ton). Posteriormente, una caída sistemática en los desembarques entre comienzos de los años 1970’s y fines de los años 1990’s. Finalmente, un repunte en los desembarques a partir del año 2002 hasta alcanzar en el año 2011 las 81 ton (P. Alarcón, pers. comm.). Estos aumentos se han visto asociados a alzas importantes en las tasas de captura reflejando incrementos en las abundancias poblacionales de este recurso. La disminución de las capturas y de los rendimientos de pesca hacia la década del año 2000 fue atribuida principalmente a una sobre pesca (Arana et al., 2006; Eddy et al., 2010). En conjunto con las variaciones de baja
11

Latin American Journal of Aquatic Research
frecuencia presentes en esta pesquería, también existen variaciones de alta frecuencia con características interanuales, como se manifiesta en la CPUE. (Larraín & Yáñez 1985; Ernst et al., 2010b). Estas variaciones en la CPUE durante la temporada de pesca han sido atribuidas a factores físicos (temperatura), en especies congenéricas, los cuales repercutirían sobre procesos biológicos tales como la muda o sobre aspectos conductuales (Ziegler et al., 2004). En el caso de J. frontalis, estas variaciones han sido atribuidas al desfase generado por la muda de los machos, la cual sería regulada, entre otros factores, por la temperatura (Ernst et al., 2010b). La temperatura afectaría también el nivel de actividad, influyendo de esta forma sobre la captura-bilidad del recurso (Ziegler et al., 2002, 2003, 2004). La época y duración del período de portación de huevos sería atribuido a factores ambientales (Larraín & Yáñez, 1985), donde la temperatura, que experimenta cambios intra-anuales de al menos 4ºC a una profundidad media de 80 m, podría modular estos procesos (Ernst et al., 2010b). Estas variaciones inducirían cambios importantes en la CPUE debido a que las hembras de talla comercial quedan disponibles para la pesquería una vez que eclosionan los huevos. Bajo los recientes escenarios de explotación, surgen interrogantes respecto de la importancia del ambiente (señales de alta y baja frecuencia) en la regulación de la dinámica poblacional de este recurso y su pesquería (Ernst et al., 2011). Sin embargo, pocos son los estudios dedicados a comprender la dinámica océano-gráfica del sistema de islas oceánicas y la influencia del ambiente en la dinámica de la población de la langosta.
Concholepas concholepas y variabilidad ambiental El loco (Concholepas concholepas) se distribuye desde la costa central de Perú a Magallanes (Moreno et al., 1998) siendo extraído por las comunidades litorales desde el Holoceno temprano (Báez et al., 2004; Rivadeneira et al., 2010). A fines de la década del 70’s y comienzos de los 80’s, los desembarques aumentaron (incentivos por demanda y precio) disminuyendo drásticamente poco después de El Niño 82-83 en la zona norte (Regiones I-III), segunda mitad de la década (Regiones la IV y X), y a mediados de los 90’s al sur (B. Yannicelli, com. pers.). Los niveles previos no se han recuperado a pesar de que la pesquería está sujeta a un régimen de manejo de uso territorial (AMERBs). En consecuencia, los altos niveles de captura a nivel nacional sostenidos durante la década de los 80’s, son el resultado de cambios en la distribución espacial del esfuerzo de pesca y, en ocasiones, al agotamiento secuencial, dos procesos
típicos en pesquerías bentónicas. Estos, entre otros motivos (escalas metapoblacionales, captura ilegal), dificultan la utilización de series históricas de desem-barque en el análisis recurso-ambiente, requiriéndose estudios con mayor detalle espacial e indicadores adicionales (esfuerzo, estructura de talla).
Moreno et al. (1998) y Moreno (2004) a través del seguimiento interanual de reclutamiento, han correlacionado fallas del reclutamiento en la zona de Valdivia durante períodos El Niño-La Niña que podrían obedecer a cambios en los patrones de vientos. Sin embargo, las consecuencias poblacionales no han sido evaluadas. González et al., 2006, detectaron fuerte variabilidad interanual en el reclutamiento en la zona de Coquimbo (30ºS). En la costa del Perú se mencionan efectos negativos de El Niño a consecuencia de una menor disponibilidad de alimento y mayor capturabilidad.
La oceanografía de la zona austral de Chile no ha sido estudiada con intensidad, por lo que se desconoce la variabilidad interanual física del sistema. Se trata de una zona de topografía compleja y sitios remotos, lo cual dificulta el seguimiento y análisis de la historia de los recursos que en ella se explotan siendo, por tanto, un área prácticamente desconocida desde este enfoque.
Modelación biofísica de recursos pesqueros en el Pacífico sudoriental A pesar que el diseño y desarrollo de modelación biofísica es una herramienta clave para abordar el estudio de la relación ambiente-recursos desde una perspectiva mecanística, en Chile son escasos los estudios de modelación biofísica aplicados a recursos marinos. Los estudios disponibles, sin embargo, contribuyen significativamente al entendimiento de la relación ambiente-recurso y al planteamiento de hipótesis acerca de los mecanismos que los vinculan. A continuación, se describen los estudios que incorporan esta aproximación.
Recursos pelágicos y modelación biofísica Dos estudios basados en modelación biofísica han contribuido a entender factores que afectan los patrones de transporte, distribución y éxito de estadios tempranos de anchoveta en la zona centro-sur de Chile (Parada et al., 2012; Soto-Mendoza et al., 2012). Durante el verano, se observaron patrones típicos de surgencia con una fuerte componente de la corriente hacia fuera de la costa y hacia el norte. En invierno, la advección media fue más débil, con corrientes más estables cerca de la costa que en verano. El modelo biofísico acoplado reprodujo la región histórica de reclutamiento (RHR) y permitió identificar que
12

Variabilidad ambiental y recursos marinos
Dichato y golfo de Arauco operaron como zonas de suplemento de larvas a la RHR (Parada et al., 2012). Un resultado relevante que emergió de las simulaciones fue que la zona de Lebu-Corral (la más al sur de la región modelada) fue identificada como una zona de pre-reclutamiento/desove con un alto nivel de retención (Parada et al., 2012; Soto-Mendoza et al., 2012). Soto-Mendoza et al. (2012), mostraron que la variabilidad espacial y temporal de las zonas de desove tiene un efecto en el transporte, retención y sobrevivencia de huevos y larvas con saco vitelino en la zona centro-sur de Chile. La información generada a través del modelo podría ser utilizada para explorar medidas de manejo, tales como redefinición de zonas de monitoreo para evaluar la dinámica de transporte, conectividad y retención de huevos y larvas desovadas en la zona de Lebu-Corral (Soto-Mendoza et al., 2012). Este punto es de gran importancia dado que el asesoramiento científico para la asignación de cuotas de captura utiliza, entre otros, el método de producción de huevos. Estos resultados plantean nuevas hipótesis para explicar la relevancia de zonas de crianza asociadas con la región sur de Chile y sus implicancias para el reclutamiento, así como para evaluar consecuencias de los cambios en la distribución adulta. Una de las limitaciones de estos dos estudios radica en las limitaciones del modelo hidrodinámico (climatológico) utilizado en este estudio, debido a su falta de capacidad para reproducir las señales ecuatoriales en el modelo, y la falta del efecto de forzamiento con alta resolución espacial y temporal usando modelos atmosféricos regionales (Parada et al., 2012). Para poder abordar desde la perspectiva biofísica la temática de la relación ambiente-recurso de pequeños pelágicos, se requiere el diseño y desarrollo de modelamiento hidrodinámico orientado a resolver los procesos, escalas físicas adecuadas a los procesos biológicos y el fortale-cimiento de planes monitoreo para validar el trabajo de modelación. Estudios en relación a la sardina y sardina común no han sido abordados desde esta perspectiva.
Jurel y modelación biofísica Recientemente, se han realizado estudios biofísicos (Parada et al., 2010a; Vásquez, 2012) para evaluar la hipótesis de conectividad de las zonas de desove-crianza derivadas del modelo conceptual del ciclo de vida del jurel (Arcos et al., 2001). Las aplicaciones biofísicas que se implementaron, se realizaron en el marco del proyecto CONICYT 78090007, las cuáles permitieron abordar la conectividad entre las zonas de desove y crianza, y la variabilidad del reclutamiento. El modelo hidrodinámico utilizado permitió resolver adecuadamente diferentes escalas de variabilidad
física, incluyendo señales ecuatoriales en el modelo, lo que posibilitó evaluar dinámicamente la conectividad y el transporte de huevos y larvas de jurel, desde zonas de desove oceánicas a la zona de crianza histórica, así como también realizar estimaciones de reclutamiento comparables con los modelos de evaluación de stock (Parada et al., 2010a; Vásquez, 2012). Por otro lado, se evaluó biofísicamente el rol ecológico de bajos submarinos asociados al área entre 34º-36ºS y109º-112ºW, como región de desove-crianza para el jurel, dada la evidencia empírica de juveniles (durante el crucero Atlántida durante octubre de 2009), (Anikev, 2010) asociados a esta zona de montes submarinos (Parada et al., en revisión). Se desarrolló un estudio de la dinámica de la circulación junto con la implementación biofísica, la cual generó índices con alta retención de partículas del orden del 26-32% y tiempos de residencia de hasta cuatro meses, en asociación a giros estacionarios que promueven la retención. Evidencia de jurel adulto, entre 2009-2011, de peces de potencialmente pertenecientes a la misma cohorte asociados a estas zonas en conjunto con los resultados de la modelación, sugieren que estos montes submarinos podrían cumplir un rol de zonas temporales de “crianza-reclutamiento” y potencial-mente de desove, lo que cuestiona el modelo conceptual (Arcos et al., 2001) del ciclo de vida del jurel en el Pacífico sur. La relevancia, de este tipo de estudios, es que permiten poner a prueba hipótesis acerca de la historia de vida de los recursos, generando un refinamiento o cambio de los modelos conceptuales imperantes. Por otro lado, la vinculación de procesos físicos con parámetros poblacionales (i.e., recluta-miento), a través de mecanismos biofísicos explícitos, ofrece oportunidades predictivas que podrían permitir un mejor manejo del recurso.
Langostino colorado y modelación biofísica Siendo la fluctuación interanual del reclutamiento un factor clave de la dinámica poblacional de este recurso en la zona de Chile centro-sur, se planteó un estudio de modelación biofísica para evaluar la influencia de la variabilidad temporal y espacial del desove en el éxito de reclutamiento (Yannicelli et al., 2012). Se incorporaron en el modelo biofísico, ecuaciones de desarrollo y sobrevivencia dependientes de la tempe-ratura y movimiento vertical, cuyas formas y parametrizaciones se realizaron a partir de experi-mentos de laboratorio. La fracción de propágulos originados en las proximidades del área de crianza que reclutan exitosamente, es mayor y más constante a lo largo del tiempo que aquella de propágulos generados en agregaciones adultas ubicadas hacia el norte. El éxito de larvas originadas en estos últimos, varió en el tiempo, incrementándose para aquellas liberadas hacia
13

Latin American Journal of Aquatic Research
el fin de la temporada de surgencia. El éxito larval fue dependiente de la distribución espacio-temporal de su origen y su acoplamiento a la hidrodinámica costera, que respondió a la geomorfología y distribución temporal de vientos. Por tanto, esas dependencias podrían explicar parte de la incertidumbre asociada a la relación del potencial reproductivo con el reclutamiento, lo cual deberá ser evaluado a través de simulaciones interanuales, y podría ser considerado al establecer zonas y niveles de captura. Estos resultados podrían ser explorados más profundamente e incorporados operacionalmente en los modelos de toma de decisión para el manejo de este recurso.
Langosta de Juan Fernández Estudios recientes, han abordado la variabilidad interanual de la conectividad/retención de los estadios tempranos larvales de J. frontalis y su acoplamiento con los procesos oceanográficos imperantes en la zona, a través de herramientas de modelación biofísica (Porobić et al., 2012). Los resultados indican una fuerte variabilidad interanual de los índices de conectividad-retención para las poblaciones de J. frontalis, determinadas principalmente por el acopla-miento entre los factores biológicos reproductivos y por factores oceanográficos tales como las variaciones en la intensidad y presencia de estructuras de mesoescala. En conjunto con lo anterior, también se propone que J. frontalis presenta una estructura metapoblacional, con un alto nivel de conectividad en el archipiélago de Juan Fernández (AJF) y un fuerte aporte de larvas hacia las islas Desventuradas (ID). Se establece de esta forma la presencia de flujos migratorios diferenciales en el sistema de islas, encontrando que AJF actuaría como una zona fuente y que las ID actuarían como sumidero con altos niveles de advección. Estos estudios son de alto interés desde la perspectiva de la ecología de este recurso, así como la potencialidad que estos resultados tendrían sobre la evaluación de algunas propuestas de manejo sugeridas actualmente para este sistema (Eddy et al., 2010). Estas estrategias de manejo presentan la posibilidad de instaurar un área marina protegida (AMP) en la franja costera alrededor de la isla Robinson Crusoe, considerando principalmente la fase bentónica de la especie. Los resultados de la modelación biofísica (conectividad-retención), de los estadios planctónicos dentro del sistema de islas, plantean varias interrogantes relacionadas con la validez de desacoplar las fases pelágicas y bentónicas al modelar la instauración de un AMP.
Modelación biofísica y conectividad de metapo-blaciones costeras En relación a la conectividad de metapoblaciones costeras destacan dos estudios de modelación
biofísica. Aiken et al. (2007), mostraron que ejes de dispersión genéricos son variables y asimétricos a lo largo de la costa (33º-34°S), y que la dependencia temporal de los mismos es importante. Sus resultados apoyan el uso de modelos numéricos hidrodinámicos y biológicos acoplados más que estadísticas de flujo anual y modelos simples de difusión-advección para abordar el tema de conectividad. Por otro lado, Aiken & Navarrete (2011) mostraron que la circulación costera forzada con campos de viento más fuertes que los actuales, derivados de escenarios climáticos del IPCC (Intergovernmental Panel on Climate Change), inducirían mayores pérdidas costa afuera de larvas distribuidas en superficie, pero mayor retención de aquellas con comportamiento de migración vertical. Estos estudios, con resolución de mesoescala, indican que el comportamiento larval específico es un factor determinante en las tendencias poblacionales y comunitarias futuras. Estos significativos aportes fueron derivados de la utilización de configuraciones hidrodinámicas validadas a mesoescala. Sin embargo, la variabilidad hidrodinámica costera tiene una componente forzada topográficamente que es de mayor importancia para los procesos de retención y exportación de material orgánico. Si bien se sabe que los resultados biofísicos dependen de los campos hidrodinámicos utilizados, rara vez se evalúa cómo son afectados por la utilización de configuraciones físicas diferentes en cuanto a la resolución espacio-temporal propia y de sus forzantes. En este sentido, la circulación costera (al nivel de bahías, sombras de surgencia) obtenida con el modelo no captura la variabilidad correctamente a menos que la resolución del modelo oceánico se aumente en función de la topografía costera paralelamente al aumento de la resolución del forzante atmosférico y se incorporen detalles batimé-tricos de alta resolución en la costa (B. Yannicelli, com. pers.). Además, como el estable-cimiento empírico de patrones de conectividad está restringido por aspectos metodológicos, el trabajo propuso la factibilidad de utilizar información auxiliar en la evaluación de los resultados biofísicos. Por lo tanto, el refinamiento de los modelos hidrodinámicos y la evaluación de sus implicancias biológicas constituyen un paso esencial previo a la incorporación de las herramientas biofísicas en modelos operativos de manejo. El esclarecimiento de la estructura de conectividad metapoblacional media, su variabilidad y factores determinantes, son elementos considerados clave para el establecimiento de medidas de conservación, escalas de manejo pesquero y selección de sitios de repoblamiento.
Manejo de recursos en Chile y variabilidad am-biental en el asesoramiento A continuación se ejemplifica, con algunos recursos pelágicos, demersales y bentónicos, las regulaciones
14

Variabilidad ambiental y recursos marinos
de manejo y la potencialidad de la inclusión de aspectos de variabilidad ambiental que permitan apoyar el manejo de los recursos pesqueros.
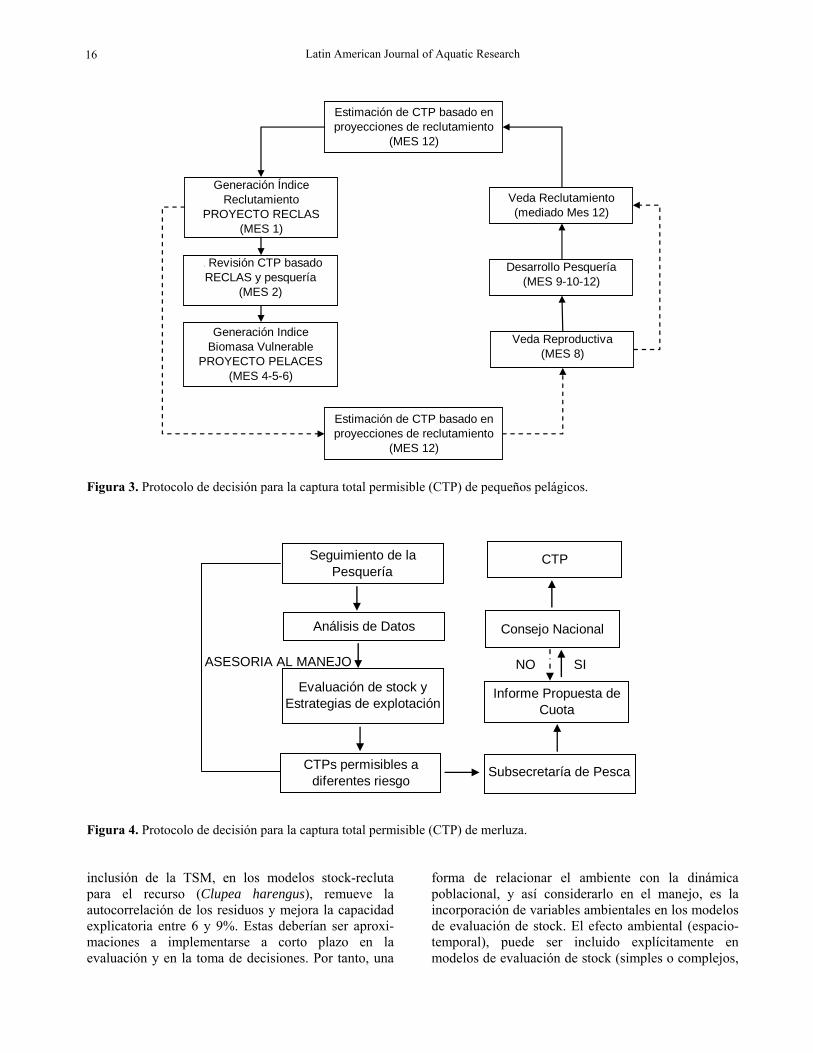
Manejo de recursos pelágicos y demersales y varia-bilidad ambiental El objetivo de manejo de pequeños pelágicos establece un nivel de biomasa reproductiva o desovante equivalente al 66% del stock desovante en estado virginal (sin explotación) considerando, además, el establecimiento de un nivel de riesgo de no alcanzar el objetivo de conservación de un 10%. Así, el estado de cada recurso es determinado mediante la utilización de modelos de evaluación de stock que incorporan información biológica y pesquera, índices de abundancia relativa, como la captura por unidad de esfuerzo (CPUE), desembarques oficiales informados por el programa de fiscalización del Servicio Nacional de Pesca y estimaciones directas de biomasa desde el año 2000 mediante hidroacústica (Castillo et al., 2009). Esta información es utilizada en la calibración de los modelos de evaluación de stock, y se obtienen las variables de estado representadas por abundancias, biomasas, y niveles de mortalidad por pesca relacionadas a la explotación pesquera. Estas variables dan cuenta del estado de los recursos que, sumado a la estrategia de explotación utilizada y al riesgo de no sobrepasar el criterio de conservación de la biomasa desovante (basado en indicadores de desempeño), dan pie a la conformación de la Captura Total Permisible (CTP) en términos biológicos aceptables (Feltrim, 2009). En la pesquería de pequeños pelágicos, sardina común y anchoveta, los problemas de manejo se refieren a que es una pesquería de carácter mixto y con alta presencia de ejemplares reclutas en las capturas. En el caso de los pelágicos se actualiza el modelo de evaluación y se analizan los escenarios de explotación para el año en curso, basado en la incorporación de la información acústica del crucero Reclas (Fig. 3). El resultado es una actualización de la captura biológicamente aceptable (CBA).
En la pesquería de la merluza común, el problema de manejo se refiere a la menor abundancia de ejemplares adultos y a la baja en la abundancia ocurrida a partir del año 2003, la cual se atribuye como responsabe a la jibia. En el aspecto administrativo, anualmente la Subsecretaría de Pesca, sobre la base de los estudios señalados, con asesoramiento por parte de los Comités Científicos, propone una CBA, la cual debe ser aprobada o rechazada por el Consejo Nacional de Pesca (CNP conformado por representantes de los grupos laborales, pescadores, y representantes del sector productivo), donde participa como presidente del
Consejo, el Subsecretario de Pesca (Fig. 4). La Subsecretaría de Pesca, sobre la base de la información disponible, realiza una propuesta de cuota que los sectorialistas evalúan y someten al CNP, quien tiene la instancia de decisión del CBA. Es claro que en el actual régimen de manejo no existe la inclusión de variabilidad ambiental y que existe suficiente evidencia que sustenta su potencial uso.
Los esquemas de administración para el manejo de los recursos, señalan que estos deben propender a la sustentabilidad de las poblaciones marinas y conservación de los ecosistemas. Sin embargo, el manejo se encuentra basado en información biológica y pesquera emanada de programas de monitoreo de la pesquería, y de estudios del estado de las poblaciones mediante modelos indirectos edad y talla estruc-turados. Aspectos relevantes como la variabilidad ambiental y elementos del ecosistema no son parte del modelo de asesoría y, por lo tanto, no son considerados de manera formal en la decisión. Estas características, consolidan al manejo en un esquema de corto plazo y sin elementos que permitan verificar las consecuencias de las decisiones, generando una administración de carácter monoespecífica sin consi-deraciones ecosistémicas. Es necesario, incluir ele-mentos de relaciones ambiente-recurso y ecosis-témicos para la administración con programas paralelos al monitoreo de las pesquerías, pero enfocados al estudio del ambiente y sus efectos en las poblaciones, incluyendo el análisis de las interac-ciones entre especies.
A nivel internacional se han realizado esfuerzos para vincular la variabilidad ambiental con el manejo. De Oliveira & Butterworth (2005) incorporaron factores ambientales en relación a las predicciones de reclutamientos futuros en los procedimientos de manejo de las pesquerías de sardina (Sardinops sagax) y anchoveta (Engraulis capensis), en Sudáfrica. Por otro lado, la regla de cosecha en el manejo de la sardina japonesa (Hurtado-Ferro et al., 2010) considera un índice ambiental en función de la TSM. En Perú, no se han establecido Puntos Biológicos de Referencia (PBR) formales, sólo existe una regla de decisión que indica que las cuotas de pesca de anchoveta serán la diferencia entre la biomasa actual menos 5 millones de ton de adultos (R. Guevara, com. pers.). Walters & Korman (1999), reconocen que existen otras fuentes que encubren la verdadera señal de reclutamiento desde el tamaño del stock desovante, tal como la depredación, competencia y el ambiente físico. Esto determina que el reclutamiento de las especies debe ser determinado por medio de una función stock-recluta que incorpore variabilidad ambiental. Fiksen & Slotte (2002) encuentran que la
15
Latin American Journal of Aquatic Research
Generación Índice Reclutamiento
PROYECTO RECLAS(MES 1)
1 Revisión CTP basado RECLAS y pesquería
(MES 2)
Generación Indice Biomasa Vulnerable
PROYECTO PELACES(MES 4-5-6)
Veda Reclutamiento(mediado Mes 12)
Desarrollo Pesquería(MES 9-10-12)
Veda Reproductiva(MES 8)
Estimación de CTP basado en proyecciones de reclutamiento
(MES 12)
Estimación de CTP basado en proyecciones de reclutamiento
(MES 12)
Figura 3. Protocolo de decisión para la captura total permisible (CTP) de pequeños pelágicos.
Seguimiento de la Pesqueria
ASESORIA AL MANEJO
Análisis de Datos
Evaluación de stock y Estrategias de explotación
CTPs permisibles a diferentes riesgo Subsecretaria de Pesca
Informe Propuesta de Cuota
Consejo Nacional
CTP
SINO
Figura 4. Protocolo de decisión para la captura total permisible (CTP) de merluza.
inclusión de la TSM, en los modelos stock-recluta para el recurso (Clupea harengus), remueve la autocorrelación de los residuos y mejora la capacidad explicatoria entre 6 y 9%. Estas deberían ser aproxi-maciones a implementarse a corto plazo en la evaluación y en la toma de decisiones. Por tanto, una
forma de relacionar el ambiente con la dinámica poblacional, y así considerarlo en el manejo, es la incorporación de variables ambientales en los modelos de evaluación de stock. El efecto ambiental (espacio-temporal), puede ser incluido explícitamente en modelos de evaluación de stock (simples o complejos,
16
Subsecretaría de Pesca
Pesquería

Variabilidad ambiental y recursos marinos
mono o multiespecíficos) modulando el crecimiento individual, la mortalidad y/o el reclutamiento (entre otros), en forma determinística o estocástica, así como pueden ser utilizados en la estandarización de la captura por unidad de esfuerzo como índice de biomasa (Wiff & Quiñones, 2004). Además, la formulación (y parametrización) de la modulación de los procesos biológicos por el ambiente físico puede tener base mecanística o correlacional y muy diverso grado de complejidad (Keyl & Wolff, 2008).
Manejo de recursos bentónicos y variabilidad ambiental La pesquería de un buen número de crustáceos, moluscos y equinodermos, se encuentra sujeta a medidas de regulación como período de veda, talla mínima de extracción o asignación de cuotas de extracción. Estas especies son objeto de estudios biológico-pesqueros de diversa cobertura espacio-temporal, y además, del registro de desembarques que realiza la autoridad fiscalizadora (SERNAPESCA). Estos recursos son monitoreados a través de programas de seguimientos de pesquerías, evalua-ciones directas, pescas de investigación y estudios biológico-pesqueros conducidos a requerimiento de la Subsecretaría de Pesca. En el año 2011, la Subse-cretaría de Pesca convocó a expertos nacionales a un Comité Científico Bentónico (CCB) para avanzar hacia un manejo ecosistémico y sustentable, propuesto en el proyecto actual de ley de pesca. Sin embargo, no existe ningún documento de referencia en cuanto al proceso de toma de decisiones referidas a la pesca bentónica conducida en áreas no asignadas como AMERBs (J. González presidente CCB, com. pers.). La consideración del efecto de la variabilidad ambiental espacio-temporal en los recursos bentónicos explotados no es explícita en las regulaciones. Sin embargo, en recursos de amplia distribución latitudinal, la variabilidad ambiental asociada es considerada a través del ajuste de valores de parámetros como la tasa de crecimiento por macrozonas. Actualmente, se evalúa un plan de manejo de 'pesquerías bentónicas' multiespecíficas en las regiones de Los Lagos y Aysén, basado en una zonificación para ser aplicada en la evaluación y manejo de más de seis recursos, y que considera la variabilidad ambiental espacial junto con otros factores (Molinet et al., 2011). Sin embargo, las regulaciones extractivas de recursos bentónicos no consideran lineamientos que guíen respuestas administrativas ante la ocurrencia de eventos climáticos interanuales, a pesar de la recurrencia de eventos climáticos conocidos con consecuencias biológicas típicas (i.e., expansión del pulpo a la región de Coquimbo).
Bajo el régimen de áreas de manejo y explotación de recursos bentónicos, la abundancia, estructura de tallas y biomasa de las especies animales explotadas, en particular el loco, son evaluadas anualmente, en base a lo cual se establece una cuota de extracción año a año para cada AMERB y recurso, que no puede exceder el 30% del stock explotable. En principio la mantención de una tasa de explotación constante serviría como enfoque precautorio para especies de longevidad mayor a 1-2 años, en escenarios de variabilidad ambiental interanual de baja frecuencia (Walters & Parma, 1996). Dada la complejidad de implementación que requieren dichos sistemas, y la incertidumbre en los procesos, una sencilla regla como la que efectivamente es aplicada en las AMERBS, sería en principio adecuada para lograr mantener las poblaciones en niveles explotables. Sin embargo, la estructura espacial de una misma meta población sujeta a explotación en numerosas concesiones independientes entre sí, podría requerir alteraciones espacialmente explícitas a la regla general, debido a la heterogeneidad espacial de la respuesta biológica ante eventos climáticos, cuya manifestación difiere entre localidades, afectando además la estructura de conectividad. La evaluación de la relación espacio-temporal ambiente/recurso, en forma espacialmente explícita, en configuraciones costeras realistas y escenarios ambientales contrastantes, es parte de investigaciones colaborativas en desarrollo que permitirían adecuar regulaciones como parte de la revisión estratégica de la medida administrativa, para su aplicación a nivel táctico anual.
SÍNTESIS
El estado del conocimiento de la relación ambiente-recurso Actualmente, la información en relación a las variabilidad ambiental y su impacto en jurel, merluza y pequeños pelágicos en la región centro sur es dispersa, careciendo a nivel nacional de un estudio que logre integrar y sintetizar la información ambiental disponible y su variabilidad en un marco que haga posible incorporarlo operativamente en un modelo de manejo. En el caso de los pequeños pelágicos en la zona norte existen estudios más orientados, aunque no se han realizados esfuerzos para implementar las relaciones encontradas en función de modelos operativos. La revisión sobre la relación recursos bentónicos-ambiente en el Sistema de la Corriente de Humboldt sugiere que las respuestas biológicas locales, no deberían relacionarse directamente a indicadores globales de variabilidad interanual. En el marco del nuevo objetivo de la política pesquera
17

Latin American Journal of Aquatic Research
nacional de manejo ecosistémico, el estudio de la variabilidad ambiental-recursos se integra como una prioridad de investigación.
Contribuciones a partir de modelación biofísica y pasos hacia su implementación El estado del arte de modelación biofísica de recursos pesqueros dentro de la jurisdicción chilena se circunscribe a un grupo reducido de contribuciones, orientadas a dilucidar aspectos básicos de la dinámica larval de anchoveta, langostino, jurel, langosta de Juan Fernández y otros en desarrollo, como el estudio del loco. Los resultados de la aplicación de modelos biofísicos de pequeños pelágicos han dado lugar al planteamiento de nuevas hipótesis en relación a la relevancia de zonas de crianza, asociadas con la región sur de Chile, y sus implicaciones para el reclu-tamiento. Estos resultados podrían ser utilizados para explorar medidas de manejo, tales como redefinición de zonas de monitoreo para evaluar la dinámica de transporte, conectividad y retención de huevos y larvas desovadas en áreas específicas. Así mismo, los resultados de la modelación biofísica del jurel han mejorado el conocimiento de su historia de vida y permitido expandir el modelo conceptual de este recurso. La relevancia topográfica en la modulación hidrodinámica costera y sus implicancias en la variabilidad de la sobrevivencia de larvas bajo consideraciones de origen espacial y temporal explícito fue evidenciada en simulaciones biofísicas de invertebrados demersales. Estos resultados sugieren la re-evaluación de procedimientos de evaluación tradicionales. Los modelos biofísicos pueden ser acoplados a los modelos de dinámica poblacional para reducir incertezas en las estimaciones (i.e., recluta-miento, biomasa vulnerable y biomasa desovante entre otras). Los modelos biofísicos pueden ser utilizados como índices ambientales los que pueden ser utilizados como indicadores independientes de recluta-miento y asentamiento, determinación de índices de sobrevivencia, así como estimaciones regionales de estos índices. Estos indicadores ambientales pueden ser incorporados en las estimaciones del modelo de evaluación de stock, asociado a la relación stock-recluta y, finalmente, como una herramienta de simulación que permita evaluar hipótesis en función de la historia de vida de los recursos y escenarios ambientales potenciales. La implementación de estos enfoques de modelación es un desafío, ya que está determinado por la estructura institucional de la investigación pesquera en Chile y la limitación de instrumentos disponibles de financiamiento. Por una parte, se requiere un énfasis en los desarrollos multidisciplinarios, un cambio en los paradigmas que guían el planteamiento y evaluación de hipótesis a
nivel observacional y numérico, para realizar avances más certeros y conceptualmente significativos. Se requiere el desarrollo de un programa orientado al establecimiento de iniciativas de implementación de sistemas de modelación acoplados entre modelos hidrodinámicos y biológicos con la incorporación de niveles tróficos inferiores y superiores, la integración y síntesis de la información disponible de los recursos pelágicos, demersales y bentónicos (análisis retros-pectivo), y el reforzamiento de programas nacionales de investigación y de políticas de cooperación inter-institucional y colaboración multidisciplinaria. Alta-mente recomendable es la formalización de planes de monitoreo y desarrollo de la oceanografía opera-cional de largo plazo, como programas establecidos, y la instauración de programas de mejoramiento de capacidades. La modelación biofísica es un elemento nuevo a nivel nacional y relevante hacia estrategias de manejo y el manejo de pesquerías basado en el ecosistema. Sin embargo, futuros planes de manejo pesquero deberían considerar la integración de modelos con todos los componentes del ecosistema marino (compartimentos biofísicos, económicos y sociales, Fulton et al., 2004). De esta manera, se considera que el acoplamiento con la componente biofísica es un elemento fundamental en la transición de modelos operacionales tradicionales hacia un enfoque más ecosistémico.
AGRADECIMIENTOS
C.P. agradece a la Comisión Nacional de Ciencia y Tecnología (CONICYT) por el financiamiento a través del proyecto 78090007; B.E. agradece a la Fundación CREO por el financiamiento del proyecto “Meso-scale and coastal oceanographic analysis around Juan Fernández and Desventuradas Archi-pelagos”, S.V. fue parcialmente financiado por la beca de magister de CONICYT; B.Y. agradece al proyecto INNOVA-CORFO 07CN13IXM-150 (Sistema de Monitoreo y Pronósticos de las Condiciones Oceano-gráficas para la Gestión Productiva y Pública del Océano Costero: Fase I).
REFERENCIAS
Aburto, J. & W. Stotz. 2003. Una experiencia de co-manejo de bivalvos en el marco de una nueva herramienta de administración pesquera en Chile: las áreas de manejo. Pol. Matt., 12: 200-204.
Aceituno, P. 1988. On the functioning of the Southern Oscillation in the South American sector, I, Surface climate. Mon. Weather Rev., 116: 505-525.
18

Variabilidad ambiental y recursos marinos
Acuña, E., R. Alarcón, L. Cid, H. Arancibia, L. Cubillos & A. Cortés. 2005. Evaluación directa de langostino colorado y langostino amarillo entre la II y VIII Regiones, año 2005. Proyecto FIP Nº 2005-09: 354 pp.
Acuña, E., R. Alarcón, H. Arancibia, A. Cortés, L. Cid, L. Cubillos & R. León. 2009. Evaluación directa de langostino colorado y langostino amarillo entre la II y VIII Regiones, año 2008. Informes técnicos FIP. FIP/IT Nº 2008-16: 420 pp.
Aguayo, M. 1995. Biology and fisheries of Chilean hakes (M. gayi and M. australis). In: J. Alheit & T.J. Pitcher (eds.). Biology, fisheries and markets. Chapman & Hall, London, pp. 305-338.
Aiken, C., S. Navarrete, M. Castillo & J.C. Castilla. 2007. Alongshore larval dispersal kernels in a numerical ocean model of the central Chilean coast. Mar. Ecol. Prog. Ser., 339: 13-24.
Aiken, C. & S. Navarrete. 2011. Environmental fluctuations and asymmetrical dispersal: generalized stability theory for studying metapopulation persistence and marine protected areas. Mar. Ecol. Prog. Ser., 428: 77-88.
Alarcón, R. & H. Arancibia. 1993. Talla de primera madurez sexual y fecundidad parcial en la merluza común, Merluccius gayi gayi (Guichenot, 1848). Cienc. Tec. Mar., 16: 33-45.
Alarcón, C., L. Cubillos & C. Oyarzún. 2004. Influence of female size on the duration and intensity of the reproductive activity of Merluccius gayi gayi off central-south Chile. Invest. Mar., Valparaíso, 32: 59-69.
Alheit, J. & M. Ñiquen. 2004. Regime shift in the Humboldt Current ecosystem. Progr. Oceanogr., 60: 201-222.
Alheit, J. & T. Pitcher. 1995. Hake: fisheries, ecology and markets, fish and fisheries series vol. 15, Chapman and Hall, London, 487 pp.
Anikev, V. 2010. Reporte de expedición de investigación científica de STM K-1704, Atlántida (crucero 53) a las regiones del Atlántico norte y del Pacífico sur fuera de las zonas económicas exclusivas de los países ribereños, 15 Julio al 28 diciembre 2009. AtlanNiro, 286 pp.
Arana, P. 1987. Perspectivas históricas y proyecciones de la actividad pesquera realizada en el archipiélago de Juan Fernández, Chile. In: J.C. Castilla (ed.). Islas oceánicas chilenas: conocimiento científico y nece-sidades de investigación. Edit. Católica de Chile, Santiago, 319-353.
Arana, P., S. Palma, A. Guerrero, M. Ahumada & A. Jofré. 2006. Monitoreo biológico-pesquero de la langosta y cangrejo dorado en el archipiélago de Juan Fernández. (Proyecto FIP Nº 2004-48). Informe Final. Estud. Doc, Pont. Univ. Católica Valparaíso, 32: 288.
Arancibia, H. 1992. Patrones de distribución de la asociación de peces demersales frente a Chile central. Rev. Biol. Pesq., 21: 43-53.
Arancibia, H. & S. Neira. 2008. Overview of the Chilean hake (Merluccius gayi) stock, a biomass forecast, and the jumbo squid (Dosidicus gigas) predator-prey relationship off central Chile (33ºS-39ºS). CalCOFI Rep., 49: 104-115.
Arcos, D., & A. Grechina. 1994. Biología y pesca comercial del jurel en el Pacífico sur. Editorial Aníbal Pinto, Concepción, 203 pp.
Arcos, D., L. Cubillos & S. Núñez. 2004. Efectos de El Niño 1997-1998 sobre las principales pesquerías pelágicas de la zona centro-sur de Chile. In: S. Avaria, J. Carrasco, J. Rutllant & E. Yáñez (eds.). 2004. El Niño-La Niña 1997-2000. Sus efectos en Chile. CONA, Chile, Valparaíso, pp. 153-177.
Arcos, D., S. Núñez & A. Acuña. 1996. Variabilidad de pequeña escala en la zona nerítica del sistema de surgencia de Talcahuano (Chile central): identi-ficación y dinámica de áreas de retención larval. Gayana Oceanol. (Chile), 4: 21-58.
Arcos, D., L. Cubillos & S. Núñez. 2001. The jack mackerel fishery and El Niño 1997-1998 effects off Chile. Progr. Oceanogr., 49: 597-617.
Arcos, D., S. Núñez, L. Castro & N. Navarro. 1987. Variabilidad vertical de clorofila-a en un área de surgencia frente a Chile central. Invest. Pesq., 34: 47-55.
Arntz, W.E. 1986. The two faces of El Niño 1982-83. Meeresforsch., 31: 1-46.
Arntz, W.E., T. Brey, J. Tarazona & A. Robles. 1987. Changes in the structure of a shallow sandy-beach community in Peru during an El Niño event. S. Afr. J. Mar. Sci., 5: 645-658.
Arrizaga, A. 1981. Nuevos antecedentes biológicos para la sardina común, Clupea (Strangomera) bentincki Norman 1936. Bol. Soc. Biol. Concepción, 52: 5-66.
Ashford, J., R. Serra, J. Saavedra & J. Letelier. 2010. Otolith chemistry indicates large-scale connectivity in Chilean jack mackerel (Trachurus murphyi), a highly mobile species in the southern Pacific Ocean. Fish. Res., 107(1-3): 291-299.
Avendaño, M., M. Cantillánez, M. Le Pennec & G. Thouzeau. 2008. Reproductive and larval cycle of the scallop Argopecten purpuratus (Ostreida: Pectinidae), during El Niño, La Niña events and normal condition in Antofasta, northen Chile. Rev. Biol. Trop., 56(1): 121-132.
Avilés, S., M. Aguayo, F. Inostroza & J. Cañón. 1979. Merluza común. En: Estado actual de las principales pesquerías nacionales. Bases para un desarrollo pesquero, IFOP-CORFO/AP, pp. 79-18.
19

Latin American Journal of Aquatic Research
Bailey, K. 1989. Description and surface distribution of juvenile Peruvian jack mackerel, Trachurus murphyi, Nichols from the subtropical convergence zone of central south Pacific. Fish. Bull., 87: 273-278.
Báez, P., J. Arata & D. Jackson. 2004. El loco Con-cholepas concholepas (Bruguière, 1789) (Mollusca: Gastropoda: Muricidae) como recurso durante el Holoceno Temprano-Medio en Los Vilos, Chile central. Invest. Mar., Valparaíso, 32(1): 107-113.
Bakun, A. 2010. Linking climate to population varia-bility in marine ecosystems characterized by nonsim-ple dynamics: Conceptual templates and schematic constructs. J. Mar. Syst., 79: 361-373.
Balbontín, F. & A. Cannobio. 1992. Growth and survival of Chilean sardine Sardinops sagax larvae reared at different densities of food. Rev. Biol. Mar., 27(1): 101-112.
Balbontín, F., A. Llanos & V. Valenzuela. 1997. Sobreposicion trófica e incidencia alimentaria en larvas de peces de Chile central. Rev. Chil. Hist. Nat., 70: 381-390.
Balbontín, F. & W. Fisher. 1981. Ciclo sexual y fecun-didad de la merluza, Merluccius gayi gayi, en la costa de Chile. Rev. Biol. Mar., 17(3): 285-334.
Barbieri, M., J. Córdova, S. Lillo, H. Peña, A. Grechina, S. Núñez, A. Sepúlveda, L. Miranda & H. Rebolledo. 1999. Análisis de la estructura del stock de jurel fuera de las aguas jurisdiccionales. Inf. Tec. FIP-IT/97-05B: 121 pp.
Bernal, R., F. Balbontín & O. Rojas. 1997. Patrones de distribución de huevos y larvas de MerluccÍus gayi gayi en la costa de Chile y factores ambientales asociados. Rev. Biol. Mar. Oceanogr., 32: 45-66.
Bertrand, A., M. Segura, M. Gutiérrez & L. Vásquez. 2004. From small-scale habitat loopholes to decadal cycles: a habitat-based hypothesis explaining fluctuation in pelagic fish populations off Peru. Fish. Fish., 5: 296-316.
Brochier, T., C. Lett, J. Tam, P. Fréon, F. Colas & P. Ayón. 2008. An individual-based model study of anchovy early life history in the northern Humboldt Current System. Progr. Oceanogr., 79: 313-325.
Bustos, C. & M. Landaeta. 2006. Desarrollo de huevos y larvas tempranas de la merluza del sur, Merluccius australis, cultivados bajo condiciones de laboratorio. Gayana, 69(2): 402-408.
Bustos, C., M. Landaeta, E. Bay-Schmith, R. Lewis & X. Moraga. 2007. Effects of temperature and lipid droplet adherence on mortality of hatchery-reared southern hake Merluccius australis larvae. Aqua-culture, 270: 535-540.
Cahuín, S., L. Cubillos, M. Ñiquen & R. Escribano. 2009. Climatic regimes and the recruitment rate of
anchoveta, Engraulis ringens, off Peru. Estuar. Coast. Shelf Sci., 84: 591-597.
Canales, C., M.A. Barbieri, R. Bahamonde, B. Leiva, P. Arana, S. Palma & T. Melo. 2002. Evaluación directa de langostino colorado y langostino amarillo entre la II and VIII Regiones, año 2001. Informe final FIP 2001-06: 269 pp.
Cárdenas, L., A. Silva, A. Magoulas, J. Cabezas, E. Poulin & F. Ojeda. 2009. Genetic population structure in the Chilean jack mackerel, Trachurus murphyi (Nichols) across the south-eastern Pacific ocean. Fish. Res., 100: 109-115.
Cardoso, F., P. Villegas & C. Estrella. 2004. Obser-vaciones sobre la biología de Octopus mimus (Cephalopoda: Octopoda) en la costa peruana. Rev. Peru. Biol., 11: 45-50.
Carstensen, D., J. Riascos, O. Heilmayer, W.E. Arntz & J. Laudien. 2010. Recurrent, thermally-induced shifts in species distribution range in the Humboldt current upwelling system. Mar. Environ. Res., 70(3-4): 293-299.
Castillo, J., E. Molina, F. Leiva, S. Núñez, S. Vásquez & E. Navarro. 2009. Evaluación hidroacústica del reclutamiento de anchoveta y sardina común entre la V y X Regiones, año 2009. Informe de Avance proyecto FIP 2008-03, Inst. Fom. Pesq. Valparaíso, 62 pp.
Castillo-Jordan, C., L. Cubillos & E. Navarro. 2010. Inter-cohort growth rate changes of common sardine (Strangomera bentincki) and their relationship with environmental conditions off central southern Chile. Fish. Res., 105: 228-236.
Castro, L. & E. Hernández. 2000. Early life survival of the Anchoveta Engraulis ringens off central Chile during the 1995 and 1996 winter spawning seasons. Trans. Am. Fish. Soc., 129: 1107-1117.
Castro, L., G. Claramunt, M. Krautz, A. Llanos-Rivera & P. Moreno. 2009. Egg trait variation in anchoveta Engraulis ringens: a maternal response to changing environmental conditions in contrasting spawning habitats. Mar. Ecol. Prog. Ser., 381: 237-248.
Castro, L., A. Llanos, J.L. Blanco, E. Tarifeño, R. Escribano & M. Landaeta. 2001. Latitudinal varia-tions in spawning habitat characteristics: influence on the early life history traits of the anchoveta, Engraulis ringens, off northern and central Chile. GLOBEC Report, 16: 42-45.
Castro, L., G. Claramunt, H. González, M. Krautz, A. Llanos-Rivera, J. Méndez, W. Schneider & S. Soto. 2010. Fatty acids in eggs of anchoveta Engraulis ringens during two contrasting winter spawning seasons. Mar. Ecol. Prog. Ser., 420: 193-205.
20

Variabilidad ambiental y recursos marinos
Cerna, J.F. & C. Oyarzún. 1998. Talla de primera madurez sexual y fecundidad parcial en la merluza común Merluccius gayi (Guichenot, 1848) del área de la pesquería industrial de la zona de Talcahuano, Chile. Invest. Mar., Valparaíso, 26: 31-40.
Chaigneau, A. & O. Pizarro. 2005. Surface circulation and fronts of the south Pacific Ocean, east of 120ºW. Geophys. Res. Lett., 32(8), LO8605.
Claramunt, G., R. Roa & L. Cubillos. 2003. Estimating daily spawning fraction using the gonadosomatic index: application to three stocks of small pelagic fish from Chile. In: O.S. Kjesbu, J.R. Hunter & P.R. Witthames (eds). Modern approaches to assess matu-rity and fecundity of warm-and cold-water fish and squids. IMR series Fisken og Havet, 12: 43-50.
Cordova, G. & F. Balbontín. 2006. Spatial distribution of abundance and size of eight kinds of fish larvae from Guafo outlet to anna pink bay, southern Chile. Cienc. Tecnol. Mar, 29(1): 153-161.
Cornejo-Rodríguez, M.H. 1991. Causas posibles de las agregaciones de jurel (Trachurus murphyi, Nichols): disponibilidad de alimento. Tesis de Magister en Ciencias con Mención en Oceanografía. Universidad de Concepción, Concepción, 108 pp.
Correa-Ramírez, M., S. Hormazábal & G. Yuras. 2007. Mesoscale eddies and high chlorophyll concen-trations off central Chile (29º-39ºS). Geophys. Res. Lett. 34, L12604, doi: 10.1029/2007gl029541.
Cortéz, T. 1995. Biología y ecología del pulpo común Octopus mimus Gould, 1852 (Mollusca: Cephalopo-da) en aguas litorales del norte de Chile. Ph.D. Thesis, Universidad de Vigo, Vigo, 293 pp.
Cortéz, T., A. González & A. Guerra. 1999. Growth of Octopus mimus (Cephalopoda, Octopodidae) in wild populations. Fish. Res., 42: 31-39.
Cowen, R., K. Lwiza, S. Sponaugle, C. Paris & D. Olson. 2000. Connectivity of marine populations: open or closed? Science, 287: 857-859.
Cubillos, L. & C. Alarcón. 2010. Estimación de la talla media de madurez sexual en Trachurus murphyi mediante parámetros del consumo relativo de oxígeno. Lat. Am. J. Aquat. Res., 38(2): 178-187.
Cubillos, L. & H. Arancibia. 1995. Comparative growth performance of horse mackerel of the genus Trachurus, with emphasis on T. symmetricus mur-phyi. In: C. Bas, J.J. Castro & J.Ma. Lorenzo (eds). International symposium on middle-sized pelagic fish. Sci. Mar., 59(3-4): 647-652.
Cubillos, L. & D. Arcos. 2002. Recruitment of common sardine (Strangomera bentincki) and anchovy (En-graulis ringens) off central-south Chile in the 1990s and the impact of the 1997-1998 El Niño. Aquat. Living Resour., 15: 87-94.
Cubillos, L., M. Canales, D. Bucarey, A. Rojas & R. Alarcón. 1999. Época reproductiva y talla media de primera madurez sexual de Strangomera bentincki y Engraulis ringens en el período 1993-1997 en la zona centro-sur de Chile. Invest. Mar., Valparaíso, 27: 73-85.
Cubillos, L.A., D.A. Bucarey & M. Canales. 2002. Monthly abundance estimation for common sardine Strangomera bentincki and anchovy Engraulis ringens in the central-south Chile (34-40°S). Fish. Res., 57: 117-130.
Cubillos, L.A, J. Paramo, P. Ruiz, S. Núñez & A. Sepúlveda. 2008. The spatial structure of the oceanic spawning of jack mackerel (Trachurus murphyi) off central Chile (1998-2001). Fish. Res., 90: 261-270.
Cubillos, L., P. Ruiz, G. Claramunt, S. Gacitúa, S. Núñez, L. Castro, K. Riquelme, C. Alarcón, C. Oyarzún & A. Sepúlveda. 2007. Spawning, daily egg production, and spawning stock biomass estimation for common sardine (Strangomera bentincki) and anchovy (Engraulis ringens) off central southern Chile in 2002. Fish. Res., 86: 228-240.
Cury, P. & L.J. Shannon. 2004. Regime shifts in upwelling ecosystems: observed changes and possible mechanisms in the northern and southern Benguela. Progr. Oceanogr., 60: 223-243.
Cury, P., A. Bakun, R. Crawford, A. Jarre, R. Quiñones, L. Shannon & H. Verheye. 2000. Small pelagics in upwelling systems: patterns of interaction and structural changes in ‘‘wasp-waist’’ ecosystems. ICES J. Mar. Sci., 57: 603-618.
De Oliveira, J. & D. Butterworth. 2005. Limits to the use of environmental indices to reduce risk and/or increase yield in the south African anchovy fishery. Afr. J. Mar. Sci., 27: 191-203.
Defeo, M. & J.C. Castilla. 1998. Harvesting and economic patterns in the artisanal Octopus mimus (Cephalopoda) fishery in a northern Chile cove. Fish. Res., 38: 121-130.
Defeo, O. & A. McLachlan. 1995. Patterns, processes and regulatory mechanisms in sandy beach macro-fauna: a multi-scale analysis. Mar. Ecol. Prog. Ser., 295: 1-20.
Di Franco, A. & P. Guidetti. 2011. Patterns of variability in early-life traits of fishes depend on spatial scale of analysis. Biol. Lett., 7(3): 454-456.
Eddy, T., J. Gardner & A. Pérez-Matus. 2010. Applying fishers ecological knowledge to construct past and future lobster stocks in the Juan Fernández archipelago, Chile. PloS One, 5(11): e13670. doi: 10.1371/journal.pone.0013670.
Elizarov, A.A., A.S. Grechina, B.N. Kotenev & A.N. Kuzetsov. 1993. Peruvian jack mackerel, Trachurus
21

Latin American Journal of Aquatic Research
symmetricus murphyi, in the open waters of the South Pacific. J. Ichthyol., 33: 86-104.
Ernst, B., C. Parada, P. Manríquez, J. Chamorro & P. Retamal. 2010a. Dinámica poblacional y pesquera de la langosta en la isla Alejandro Selkirk. Proyecto FIP N°2008-24: 166 pp.
Ernst, B., J.M. (Lobo) Orensanz, P. Manríquez, J. Chamorro, C. Roman & J. Porobić. 2011. Monitoreo a la pesquería de crustáceos en el archipiélago Juan Fernández, año 2011. Proyecto FIP N°4728-49: 142 pp.
Ernst, B., P. Manríquez, J.M. (Lobo) Orensanz, R. Roa, J. Chamorro & C. Parada. 2010b. Strengthening of a traditional territorial tenure system through prota-gonism in monitoring activities by lobster fishermen from the Juan Fernandez Islands, Chile. Bull. Mar. Sci., 86(2): 1-44.
Escribano, R., G. Daneri, L. Farías, V.A. Gallardo, H. González, D. Gutiérrez, C.B. Lange, C. Morales, O. Pizarro, O. Ulloa & M. Braun. 2004. Biological and chemical consequences of the 1997-1998 El Niño in the Chilean coastal upwelling system: a synthesis. Deep-Sea Res. II, 51: 2389-2411.
Espino, M. & C. Wosnitza-Mendo. 1989. Biomass of hake (Merluccius gayi) of Peru, 1953-1987. In: D. Pauly, P. Muck, J. Mendo & I. Tsukayama (eds.). The Peruvian upwelling ecosystem: dynamics and inte-ractions. ICLARM Conf. Proc., 18: 297-305.
Farías, A., J.G. Bell, I. Uriarte & J. Sargent. 2003. Polyunsaturated fatty acids in total lipid and phospholipids of Chilean scallop Argopecten purpu-ratus (L.) larvae: effects of diet and temperature. Aquaculture, 228: 289-305.
Feltrim, M. & B. Ernst. 2010. Inter-cohort growth variability and its implication for fishery management of the common sardine (Strangomera bentincki) stock off the coast of south-central Chile. Fish. Res., 106(3): 368-377.
Feltrim, M. 2009. Investigación, evaluación de stock y CTP de sardina común y sardina austral centro sur. Informe Final. Instituto de Fomento Pesquero-SUB PESCA, 60 pp.
Ferrada, S., K. Hernández, R. Montoya & R. Galleguillos. 2002. Estudio poblacional del recurso anchoveta (Engraulis ringens Jenyns, 1842) (Clupeiformes, Engraulidae), mediante análisis de ADN. Gayana, 66(2): 243-248.
Fiksen, O. & A. Slotte. 2002. Stock-recruitment models for Norwegian spring spawning herring (Clupea harengus). Can. J. Fish. Aquat. Sci., 59: 211-217.
Franco-Meléndez, M.T. 2012. Breeding behavior and sex ratio variation of Pleuroncodes monodon (Crustacea: Galatheidae) off the Peruvian coast. Cienc. Mar., 38: 441-457.
Freón, P. & E. Yáñez. 1995. Influencia del medio ambiente en evaluación de stock: una aproximación con modelos globales de producción. Invest. Mar., Valparaíso, 23: 25-47.
Fuentes, L., I. Vila & M. Contreras. 2005. Temperature, energy acquisition and energy use in the Chilean silverside Basilichthys australis Eigenmann (Atheri-nopsidae). Rev. Chil. Hist. Nat., 78(2): 303-311.
Fuenzalida, R., W. Schneider, J. Garcés-Vargas & L. Bravo. 2008. Satellite altimetry data reveal jet-like dynamics of the Humboldt Current. J. Geophys. Res., 113: c07043, doi: 10.1029/2007jc004684.
Fulton, E.A., J.S. Parslow, A.D.M. Smith & C.R. Johnson. 2004. Biogeochemical marine ecosystem models II: the effect of physiological detail on model performance. Ecol. Model., 173: 371-406.
Fulton, E., A. Smith & D. Smith. 2007. Alternative management strategies for southeast australian commonwealth Fisheries: Stage 2: Quantitative Ma-nagement Strategy Evaluation. Australian Fisheries Management Authority, Fisheries Research and Development Corporation Report, 378 pp.
Fyfe, J. 2003. Extratropical southern hemisphere cyclones: harbingers of climate change. J. Climate, 16: 2802-2805.
Gallardo, V.A., M. Palma, F.D. Carrasco, D. Gutiérrez, A. Levin & J.I. Cañete. 2004. Macrobenthic zonation caused by the oxygen minimum zone on the shelf and slope off central Chile. Deep-Sea Res. II, 51(20-21): 2475-2490.
Gálvez, P., J. Sateler, R. Tascheri, R. Escobar, Z. Young, J. Olivares, V. Ojeda, J. González & G. Toledo. 2006. Investigación situación pesquería demersal centro sur, 2005. Informe final, IFOP, 87 pp.
Garreaud, R. & D. Battisti. 1999. Interannual (ENSO) and interdecadal (ENSO-like) variability in the southern hemisphere tropospheric circulation. J. Climate, 12: 2113-2123.
Garreaud, R., M. Vuille, R. Compagnucci & J. Marengo. 2008. Present-day South American climate. PALAEO3 Special Issue (LOTRED South America), 281: 180-195, doi:10.1016/j.paleo.2007.10.032.
Gatica, C., C. Alarcón & L. Cubillos. 2003. Precisión en los estimados de distribución de frecuencias de tallas de jurel, Trachurus symmetricus. Invest. Mar., Valparaíso, 31: 101-106.
Gatica, C., M. Arteaga, J. Giacaman & P. Ruiz. 2007. Tendencias en la biomasa de sardina común (Strangomera bentincki) y anchoveta (Engraulis ringens) en la zona centro-sur de Chile, entre 1991 y 2005. Invest. Mar., Valparaíso, 35(1): 13-24.
Gaymer, C.F., W. Stotz, R. Garay-Fluhmann, G. Luna-Jorquera & M. Ramos. 2010. Evaluación de línea base
22

Variabilidad ambiental y recursos marinos
de Las Reservas Marinas Isla Chañaral e Isla Choros-Damas. Informe Final Proyecto FIP 2006-56: 532 pp.
Gillet, N.P., T.D. Kell & P.D. Jones. 2006. Regional climate impacts of the southern Annular Mode, Geophys. Res. Lett., 33, L23704, doi:10.1029/2006 GL027721.
Gómez, F. 2007. Variabilidad ambiental y pequeños pelágicos de la zona norte y centro-sur de Chile. Tesis Magíster en Ciencias, mención Pesquerías. Univer-sidad de Concepción, Concepción, 90 pp.
Gómez, F., A. Montecinos, S. Hormazábal, L.A. Cubillos, M. Correa-Ramírez & F.P. Chávez. 2012. Impact of spring upwelling variability off southern-central Chile on common sardine (Strangomera bentincki) recruitment. Fish. Oceanogr., 21(6): 405-414.
Gómez, F. 2007. Variabilidad ambiental y pequeños pelágicos de la zona Norte y centro-sur de Chiloé. Tesis de Magíster en Ciencias, Sección Pesquerías. Universidad de Concepción, Concepción, 90 pp.
González, H, G. Daneri, D. Figueroa, J.L. Iriarte, N. Lefevre, G. Pizarro, R. Quiñones, M. Sobarzo & A. Troncoso. 1998. Producción primaria y su destino en la trama trófica pelágica y océano profundo e intercambio océano-atmósfera de CO2 en la zona norte de la Corriente de Humboldt (23ºS): posibles efectos del evento El Niño, 1997-1998 en Chile. Rev. Chil. Hist. Nat., 71(4): 429-458.
González, J., C. Tapia, A. Wilson, W. Stotz, J.M. Orensanz, A.M. Parma, J. Valero, M. Catrilao & J. Garrido. 2004. Bases biológicas para la evaluación y manejo de metapoblaciones de loco en las III y IV Regiones. IFOP (Chile). IFOP, Project FIP No 2002-16, Final Report, Valparaíso, 101 pp.
González, J., W. Stotz, J. Garrido, J.M. (Lobo), Orensanz, A.M. Parma, C. Tapia & A. Zuleta. 2006. The Chilean TURF system: how is it performing in the case of the loco fishery? Bull. Mar. Sci., 78: 499-527.
Gretchina, A., S. Núñez & D. Arcos. 1998. Biología reproductiva del jurel (Trachurus symmetricus murphyi) en el Pacífico sur. In: D. Arcos (ed.). Biología y ecología del jurel en aguas chilenas. Instituto de Investigación Pesquera, Talcahuano, pp. 77-99.
Guerra, A., T. Cortéz & F. Rocha. 2009. Redescripción del pulpo de los changos, Octopus mimus Gould 1852, del litoral Chileno-Peruano (Mollusca, Cephalopoda). Iberus, 17(2): 37-57.
Gutiérrez, M., A. Ramírez, S. Bertrand, O. Moron & A. Bertrand. 2008. Ecological niches, patterns of distribution and areas of overlap of the squat lobster
munida (Pleuroncodes monodon) and anchoveta (Engraulis ringens) off Peru. Progr. Oceanogr., 79: 256-263.
Gutiérrez, D., A. Bertrand, C. Wosnitza-Mendo, B. Dewitte, S. Purca, C. Peña, A. Chaigneau, J. Tam, M. Graco, V. Echevin, C. Grados, P. Fréon & R. Guevara-Carrasco. 2011. Sensibilidad del sistema de afloramiento costero del Perú al cambio climático e implicancias ecológicas. RPGA, 3: 1-26.
Gutiérrez-Estrada, J.C., E. Yáñez, I. Pulido-Calvo, C. Silva, F. Plaza & C. Bórquez. 2009. Pacific sardine (Sardinops sagax Jenyns, 1842) landings prediction. A neural network ecosystemic approach. Fish. Res., 100: 116-125.
Haig, J. 1955. The Crustacean Anomura of Chile. Report of the Lunds University Chile Expedition 1848-49. Lund Univ. Arsskr. (N.F. Avd. 2), 51(12): 1-60.
Halpern, D. 2002. Offshore Ekman transport and Ekman pumping off Peru during the 1997-1998 El Niño. Geophys. Res. Lett., 29(5): 1075, doi: 10.1029/2001 GL014097.
Haye, P., P. Salinas, E. Acuña & E Poulin. 2010. Heterochronic phenotypic plasticity with lack of genetic differentiation in the southeastern Pacific squat lobster Pleuroncodes monodon. Evol. Dev., 12(6): 628-634.
Hermann, A., S. Hinckley, B. Megrey & P. Stabeno. 1996. Interannual variability of the early life history of walleye pollock near Shelikof Strait, as inferred from a spatially explicit, individual-based model. Fish. Oceanogr., 5(1): 39-57.
Hernández-Miranda, E. & P. Ojeda. 2006. Inter-annual variability in somatic growth rates and mortality of coastal fishes off central Chile: an ENSO driven process? Mar. Biol., 149: 925-936.
Hinckley, S., A. Hermann & B. Megrey. 1996. Develop-ment of a spatially explicit, individual-based model of marine fish early life history. Mar. Ecol. Prog. Ser., 139: 47-68.
Hormazábal, S., G. Shaffer & O. Leth. 2004. The coastal transition zone off Chile. J. Geophys. Res., 109(C01021), doi:10.1029 2003JC001956.
Hormazábal, S., G. Shaffer & O. Pizarro. 2002. Tropical Pacific control of intraseasonal oscillations off Chile by way of oceanic and atmospheric pathways. Geophys. Res. Lett., 29(6): doi: 10.1029/2001GL0 13481.
Hormazábal, S., G. Shaffer, J. Letelier & O. Ulloa, 2001. Local and remote forcing of sea surface temperature in the coastal upwelling system off Chile. J. Geophys. Res., 106: 16657-16672.
23

Latin American Journal of Aquatic Research
Hurtado-Ferro, F., K. Hiramatsu & K. Shirakihara. 2010. Allowing for environmental effects in a management strategy evaluation for Japanese sardine. ICES J. Mar. Sci., 67: 2012-2017.
Illanes, J., S. Akaboshi & E. Uribe. 1985. Efectos de la temperatura en la reproducción del ostión del norte Chlamys (Argopecten) purpuratus en la Bahía Tongoy durante el fenómeno El Niño 1982-83. Invest. Pesq., 32: 167-173.
Iriarte, J.L. & H.E. González. 2004. Phytoplankton size structure during and after the El Niño of 1997-1998 in a coastal upwelling area of the northern Humboldt Current System. Mar. Ecol. Prog. Ser., 269: 83-90.
Iriarte, J.L., G. Pizarro, V.A. Troncoso & M. Sobarzo. 2000. Primary production and biomass of size-fractionated phytoplankton off Antofagasta, Chile (23-24°S) during pre-El Niño and El Niño 1997. J. Mar. Syst., 26: 37-51.
Jurado-Molina, J., C. Gatica & L. Cubillos. 2006. Incorporating cannibalism into age-structured model for the Chilean hake. Fish. Res., 82: 30-40.
Kenny, A., H. Skjoldal, G. Engelhard, P. Kershaw & J. Reid. 2009. An integrated approach for assessing the relative significance of human pressures and environmental forcing of the status of Large Marine Ecosystems. Progr. Oceanogr., 81: 132-148.
Keyl F. & M. Wolff. 2008. Environmental variability and fisheries: what can models do? Rev. Fish. Biol. Fish., 18: 273-299.
Klyashtorin, L. 1998. Long-term climate change and main commercial fish production in the Atlantic and Pacific. Fish Res., 37: 115-25.
Konchina, Y., A. Nesin, N. Onishchik & Y. Pavlov. 1996. On the migration and feeding of the jack mackerel Trachurus symmetricus murphyi in the Eastern Pacific. J. Ichthyol., 36: 753-766.
Krautz, M.C., M. González & L. Castro. 2003. Detection of anchoveta (Engraulis ringens Jenyns,1842) eggs in euphausiid diets using immunoassays (ELISA). J. Exp. Mar. Biol. Ecol., 294(1): 27-39.
Krautz, M.C., S. Vásquez, L.R. Castro, M. González, A. Llanos-Rivera & S. Pantoja. 2010. Changes in metabolic substrates during early development in anchoveta Engraulis ringens (Jenyns, 1842) in the Humboldt Current. Mar. Biol., 157(5): 1137-1149.
Landaeta, M.F. & L.R. Castro. 2006. Larval distribution and growth of the rockfish, Sebastes capensis (Sebastidae, Pisces), in the fjords of southern Chile. ICES J. Mar. Sci., 63: 714-724.
Landaeta, M., M.I. Muñoz & L. Castro. 2009. Variabilidad estacional y a pequeña escala en la distribución vertical del ictioplancton en un fiordo
estratificado del sur de Chile. Cienc. Tecnol. Mar, 32(2): 27-42.
Landaeta, M., C. Bustos, P. Palacios-Fuentes, P. Rojas & F. Balbontín. 2011. Distribución del ictioplancton en la Patagonia austral de Chile: potenciales efectos del deshielo de Campos de Hielo Sur. Lat. Am. J. Aquat. Res., 39(2): 236-249.
Larraín, F. & E. Yáñez. 1985. Diseño de un sistema de recolección y procesamiento de datos de captura y esfuerzo para la pesquería de langosta de Juan Fernández (Jasus frontalis, H. Milne Edwards, 1837) In: P. Arana (ed.). Investigaciones marinas en el archipiélago de Juan Fernández. Universidad Católica de Valparaíso, Valparaíso, pp. 373-278.
Leggett, W.C. & E. DeBlois. 1994. Recruitment in marine fishes: is it regulated by starvation and predation in the egg and larval stages. Neth. J. Sea. Res., 32: 119-134.
Leth, O. & G. Shaffer. 2001. A numerical study of seasonal variability in the circulation off central Chile. J. Geophys. Res., 106(C10): 22229-22248.
Leth, O. & J.F. Middleton. 2006. A numerical study of the upwelling circulation off central Chile: effects of remote oceanic forcing. J. Geophys. Res., 111, C12003, doi: 10.1029/2005JC003070.
Lett, C., K. Rose & B. Megrey. 2009. Biophysical models. In: D.M. Checkley, J. Alheit, Y. Oozeki & C. Roy (eds.). Climate change and small pelagic fish. Cambridge University Press, Cambridge, pp. 88-111.
Lett, C., S.D. Ayata, M. Huret & J.O. Irisson. 2010. Biophysical modelling to investigate the effects of climate change on marine population dispersal and connectivity. Progr. Oceanogr., 87(1-4): 106-113.
Lillo, S., J. Córdova, M. Rojas, V. Ojeda, J. Olivares, R. Tascheri, M. Braun, S. Núñez & J. Ortiz. 2001. Evaluación hidroacústica del stock de merluza común explotado en la zona centro-sur. Informe Final Proyecto FIP 2000-04: 276 pp.
Llanos-Rivera, A. & L. Castro. 2004. Latitudinal and seasonal egg size variations of the anchoveta Engraulis ringens off the Chilean coast. Fish. Bull., 102: 207-212.
Llanos-Rivera, L. & R. Castro. 2006. Inter-population differences in temperature effects on Engraulis ringens yolk-sac larvae. Mar. Ecol. Prog. Ser., 312: 245-253.
Mantua, N.J., S.R. Hare, Y. Zhang, J.M. Wallace & R.C. Francis. 1997. A Pacific decadal climate oscillation with impacts on salmon. B.A.M. Meteorol. Soc., 78: 1069-1079.
Martínez, G., C. Aguilera & L. Mettifogo. 2000. Interactive effects of diet and temperature on
24

Variabilidad ambiental y recursos marinos
reproductive conditioning of Argopecten purpuratus broodstock. Aquaculture, 183: 149-159.
McFarlane, G.A., J.R. King & R.J. Beamish. 2000. Have there been recent changes in climate? Ask the fish. Progr. Oceanogr., 47: 147-169.
Megrey, B.A. & S. Hinckley. 2001. The effect of turbulence on feeding of larval fishes: a sensitivity analysis using an individual based model. ICES J. Mar. Sci., 58(5): 1015-1029.
Melbourne-Thomas, J., C.R. Johnson, P.M. Aliño, R.C. Geronimo, C.L. Villanoy & G.G. Gurney. 2011. A multi-scale biophysical model to inform regional management of coral reefs in the western Philippines and south China sea. Environ. Modell. Softw., 26: 66-82.
Miranda, L., A. Hernández, A. Sepúlveda & M. Landaeta. 1998. Alimentación de jurel y análisis de la selectividad en la zona centro-sur de Chile. In: D. Arcos (ed.). Biología y ecología del jurel en aguas chilenas. Instituto de Investigación Pesquera, Talcahuano, 216 pp.
Molinet, C., M. Cáceres, M. González, J. Carvajal, G. Asencio, M. Díaz, P. Díaz, M. Castro & H. Codjambassis. 2011. Population dynamic of early stages of Caligus rogercresseyi in an embayment used for intensive salmon farms in Chilean inland seas. Aquaculture, 312: 62-71.
Montecinos, A. 1991. Efecto del fenómeno El Niño en los vientos favorables a la surgencia costera en la costa norte de Chile. Tesis de Oceanografía. Univer-sidad Católica de Valparaíso, Valparaíso, 143 pp.
Montecinos, A. & F. Gómez. 2010. Interdecadal variability of the upwelling season off southern-central Chile (37ºS): impact on common sardine (Strangomera bentincki). http://www.dgf.uchile.cl/ ACT19/COMUNICACIONES/Revistas/MG_PO07.pdf.
Morales, C., M. Braun, H. Reyes, J. Blanco & A. Davies. 1996. Anchovy larval distribution in the coastal zone off northern Chile: the effect of low dissolved oxygen concentrations and of a cold-warm sequence (1990-95). Invest. Mar., Valparaíso, 24: 77-96.
Moreno, C., G. Asencio, W.E. Duarte & V. Marín. 1998. Settlement of tye muricid Concholepas concholepas (Brugière) ans its relatonship with El Niño and coastal upwellings in southern Chile. Mar. Ecol. Prog. Ser., 167: 171-175.
Moreno, C. 2004. Efectos de El Niño en el reclutamiento de Concholepas concholepas y Tegula atra (Mollusca, Gastropoda) en la costa de Valdivia, Chile. In: S. Avaria, J. Carrasco, J. Rutllant & E. Yaez. (eds.). El Niño-La Niña 1997-2000. Sus efectos
en Chile. CONA, Chile, Valparaíso. PDS Code: 11932, 1(1): 179-189.
Munday, P., J. Leis, J. Lough, C. Paris, M. Kingsford, M. Berumen & J. Lambrechts. 2009. Climate change and coral reef connectivity. Coral Reefs, 28(2): 379-395.
Nakata, H., S. Kimura, Y. Okazaki & A. Kasai. 2000. Implications of meso-scale eddies caused by frontal disturbances of the Kuroshio Current for anchovy recruitment. ICES J. Mar. Sci., 57: 143-152.
Navarro, J.M., G.E. Leiva, G. Martínez & C. Aguilera. 2000. Interactive effects of diet and temperature on the scope for growth of the scallop Argopecten purpuratus during reproductive conditioning. J. Exp. Mar. Biol. Ecol., 247: 67-83.
Ñiquen, M. & M. Bouchon. 2004. Impact of El Niño event on pelagic fisheries in Peruvian waters. Deep–Sea Res. II, 51: 563–574.
Ojeda, V., F. Cerna, H. Hidalgo, R. Bravo, L. Cid, L. Muñoz & L. Chong. 2001. Determinación de edad Informe Final II Parte. En: Programa de seguimiento del estado de situación de las principales pesquerías nacionales. Proyecto investigación situación pes-quería demersal sur-austral, 2000.
Olivares, A., O. Zúñiga, G. Castro, C. Segura & J. Sánchez. 1996. Bases biológicas para el manejo de Octopus mimus: reproducción y crecimiento. Estud. Oceanol., 15: 61-74.
Orensanz, J., A. Parma & O. Iribarne. 1991. Population dynamics and management of natural stocks. In: S. Shumway (ed.). Scallops: biology, ecology and aquaculture. Developments in aquaculture and fisheries science. Elsevier, Amsterdam, 21: 625-713.
Ottersen, G., S. Kim, G. Hused, J. Polovina & N. Stenseth. 2010. Major pathways by which climate may force marine fish populations. J. Mar. Syst., 79(3-4): 343-360.
Oyarzún, C., J. Chong & M. Malagueño. 1998. Fenología reproductiva en el jurel Trachurus symmetricus (Ayres, 1855) (Perciformes, Carangidae) en el área de Talcahuano, Chile: 1982-1984. In: D. Arcos (ed.). Biología y ecología del jurel en aguas chilenas, Instituto de Investigación Pesquera, Talcahuano, pp. 67-75.
Palma, S. 1994. Distribución y abundancia de larvas de langostino colorado Pleuroncodes monodon frente a la costa de Concepción, Chile. Invest. Mar., Valparaíso, 22: 13-29.
Palma, S. & P. Arana. 1997. Aspectos reproductivos de langostino colorado (Pleuroncondes monodon H. Milne-Edwards, 1837), frente a la costa de Concepción, Chile. Invest. Mar., Valparaíso, 25: 203-221.
25

Latin American Journal of Aquatic Research
Parada, C., F. Colas, S. Soto-Mendoza & L. Castro. 2012. Effects of seasonal variability in across- and alongshore transport of anchoveta (Engraulis ringens) larvae on model-based pre-recruitment indices off central Chile. Progr. Oceanogr., 92(1): 192-205.
Parada, C., S. Nuñez, M. Correa-Ramírez, S. Vásquez, A. Sepúlveda, S. Hormazábal, V. Combes & E. Di Lorenzo. 2010a. Advances in biophysical modeling of chilean jack mackerel in the south Pacific. ICES CM 2010/L 20: 32 pp.
Parada, C., D. Armstrong, B. Ernst, S. Hinckley & J.M. (Lobo) Orensanz. 2010b. Spatial dynamics of snow crab (Chionoecetes opilio) in the Eastern Bering Sea-Putting together the pieces of the puzzle. Bull. Mar. Sci., 86(2): 413-437.
Parada, C., A. Gretchina, S. Vásquez, V. Combes, B. Ernst, S. Núñez, A. Sepúlveda & E. DiLorenzo. Expanding the conceptual framework of the spatial population structure and life history of jack mackerel in the southeastern Pacific off Central-Chile: Oceanic seamount region as potential spawning/ nursery habitat, 36 pp. (Submitted to ICES).
Paris, C.B., R.K. Cowen, K.M. Lwiza, D.P Wang & D.B. Olson. 2002. Objective analysis of three-dimensional circulation in the vicinity of Barbados, West Indies: Implication for larval transport. Deep-Sea Res., 49: 1363-1386.
Payá, I. 2006. Investigación evaluación de stock y CTP merluza común, 2006. Informe Prefinal BIP N˚ 30043787-0, Instituto de Fomento Pesquero (IFOP), Valparaíso, 39 pp.
Payá, I. & A. Zuleta. 1999. Investigación de CTP de merluza común, año 2000. Instituto de Fomento Pesquero, Valparaíso, 57 pp.
Payá, I., P. Rubilar, H. Pool, R. Céspedes, H. Reyes, N. Ehrhardt, L. Adarme & H. Hidalgo, 2002. Evaluación de la merluza de cola y merluza tres aletas. Informe Final Proyecto FIP 2000-15: 163 pp.
Payá, I., L. Caballero, H. Hidalgo & M. Montecinos. 2003. Captura total permisible regionalizada de merluza de cola 2003. Informe Final. Instituto de Fomento Pesquero, Valparaíso, 47 pp.
Pizarro, O. 1991. Propagación y forzamiento de perturba-ciones de baja frecuencia del nivel del mar en la costa norte de Chile. Tesis de Oceanografía. Universidad Católica de Valparaiso, Valparaiso, 105 pp.
Pizarro, O. 1999. Low frequency fluctuations in the eastern boundary current circulation off south America: remote and local forcing. Ph.D. Thesis, University of Gothenburg, Sweden, 102 pp.
Pizarro, O., A. Clarke & S. Van Gorder. 2001. El Niño sea level and currents along the South America coast: comparison of observations with theory. J. Phys. Oceanogr., 31: 1891-1903.
Porobić, J., C. Parada, B. Ernst, S. Hormazabal & V. Combes. 2012. Modelación de la conectividad de las subpoblaciones de la langosta de Juan Fernández a través de un modelo biofísico. Lat. Am. J. Aquat. Res., 40(3): 613-632.
Quintana, J. 2004. Estudio de los factores que explican la variabilidad de la precipitación en Chile en escalas de tiempo interdecadal. Tesis de Magíster en Geofísica, Universidad de Chile, Santiago, 63 pp.
Quiñones, R., R. Serra, S. Nuñez, H. Arancibia, J. Córdova & F. Bustos. 1997. Relación espacial entre el jurel y sus presas en la zona centro-sur de Chile. In: E. Tarifeño (ed.). Gestión de sistemas oceanográficos del Pacífico oriental. UNESCO COI/INF-1046, pp. 187-202.
Riascos, J., O. Heilmayer, M.E. Oliva & J. Laudien. 2011. Environmental stress and parasitism as drivers of population dynamics of Mesodesma donacium at its northern biogeographic range. ICES J. Mar. Sci., 68: 823-833.
Riascos, J., O. Heilmayer, M. Oliva, J. Laudien & W.E. Arntz. 2008. Infestation of the surf clam Mesodesma donacium by the spionid polychaete Polydora bioccipitalis. J. Sea Res., 59: 217-227.
Riascos, J., D. Carstensen, J. Laudien, W.E. Arntz, M. Oliva, A. Güntner & O. Heilmayer. 2009. Thriving and declining: climate variability shaping life-history and population persistence of Mesodesma donacium in the Humboldt upwelling system. Mar. Ecol. Prog. Ser, 385: 151-163.
Rivadeneira, M., C.M. Santoro & P.A. Marquet. 2010. Reconstructing the history of human impacts on coastal biodiversity in Chile: constraints and opportunities. Aquat. Conserv. Mar. Freshwat. Ecosyst., 20: 74-82.
Roa, R. & R. Bahamonde. 1993. Growth and expansion of an exploited population of the squat lobster (Pleuroncodes monodon) after 3 years without harvesting. Fish. Res., 18: 305-319.
Rocha, F. & M. Vega. 2003. Overview of cephalopod fisheries in Chilean waters. Fish. Res., 60: 151-159.
Rojas, P., M. Landaeta & R. Ulloa. 2011. Eggs and larvae of anchoveta Engraulis ringens off northern Chile during the 1997-1998 El Niño event. Rev. Biol. Mar. Oceanogr., 46(3): 405-419.
Rutllant, J., B. Rosenbluth & S. Hormazábal. 2004. Intraseasonal variability in the local wind forcing of coastal upwelling off central Chile (30°S). Cont. Shelf. Res., 24: 789-804.
26

Variabilidad ambiental y recursos marinos
Salinger, M., J. Renwick & A. Mullan. 2001. The interdecadal Pacific oscillation and South Pacific climate, Int. J. Climatol., 21: 1705-1721.
Sarachik, E.S. & M.A. Cane. 2010. The El Nino-southern oscillation phenomenon. Cambridge University Press, Cambridge, 369 pp.
Serra, J.R. 1978. La pesquería de sardina común (Clupea (Strangomera) bentincki) y anchoveta (Engraulis ringens) de Talcahuano: análisis de su desarrollo y situación actual. Invest. Pesq., 29: 1-21.
Serra, J.R., 1991. Important life history aspects of the Chilean jack mackerel, Trachurus symmetricus murphyi. Invest. Pesq., 36: 67-83.
Shaffer, G., O. Pizarro, L. Djurfeldt, S. Salinas & J. Rutlant. 1997. Circulation and low-frequency variability near the Chilean coast: remotely forced fluctuations during the 1991-92 El Niño. J. Phys. Oceanogr., 27: 217-235.
Shaffer, G., S. Hormázabal, O. Pizarro & S. Salinas. 1999. Seasonal and interannual variability of currents and temperature over the slope of central Chile. J. Geophys. Res., 104(C12): 29951-29961.
Shumway, S.E., R. Selvin & D.F. Schick. 1987. Food resources related to habitat in the scallop Placopecten magellanicus (Gmelin, 1791). J. Shellfish Res., 7: 77-82.
Soto-Mendoza, S., C. Parada, L. Castro, F. Colas & W. Schneider. 2012. Modeling transport and survival of anchoveta eggs and yolk–sac larvae in the coastal zone off central-southern Chile: Assessing spatial and temporal spawning parameters. Prog. Oceanogr., 92(1): 178-191.
Stotz, W.B., L.M. Caillaux & D. Lancelloty. 2005. Formulación de metodologías para la evaluación de Áreas de Manejo. Proyecto FIP 2003-18: 302 pp.
Strub, P.T., J.M. Mesías, V. Montecino, J. Rutllant & S. Salinas. 1998. Coastal ocean circulation off western South America. Coastal segment (6, E). In: A.R. Robinson & K.H. Brink (eds.). The Sea, 11: 273-308.
Servicio Nacional de Pesca (SERNAPESCA). 1992-2002. Anuarios Estadísticos de pesca http://www. sernapesca.cl/index.php?option=com_remository Ite mid=246&func=select&id=2.
Subsecretaría de Pesca (SUBPESCA). 2012. Informe sectorial de pesca y acuicultura. Departamento de análisis sectorial. Subsecretaría de Pesca, Valparaíso, 26 pp.
Tarazona, J., R. Espinoza, M. Solís & W. Arntz. 2007. Crecimiento y producción somática de la concha de abanico (Argopecten purpuratus) en Bahía Indepen-dencia, Pisco (Perú) comparados entre eventos El
Niño y La Niña. Rev. Biol. Mar. Oceanogr., 42(3): 275-285.
Tarifeño, E. 1980. Studies on the biology of surf clam Mesodesma donacium (Lamarck, 1818) (Bivalvia: Mesodesmatidae) from Chilean sandy beaches. University of California, California, 229 pp.
Tarifeño, E., M. Carmona, A. Llanos-Rivera & L.R. Castro. 2008. Temperature effects on the anchoveta Engraulis ringens egg development: do latitudinal differences occur? Environ. Biol. Fish., 81(4): 387-395.
Thatje, S., O. Heilmeyer & J. Laudien. 2008. Climate variability and El Niño Southern thermally-induced shifts in species distribution range in the Humboldt Current upwelling system. Mar. Environ. Res., 70: 293-299.
Thompson, D. & S. Solomon. 2002. Interpretation of recent Southern Hemisphere climate change. Science, 296: 895-899.
Travers, M. & Y.J. Shin. 2010. Spatio-temporal variability in fish-induced predation mortality on plankton: a simulation approach using a coupled trophic model of the Benguela ecosystem. Prog. Oceanogr., 84: 118-120.
Ulloa, O., R. Escribano, S. Hormazábal, R. Quiñones, R. González & M. Ramos. 2001. Evolution and biological effects of the 1997-98 El Niño in the upwelling ecosystem off northern Chile. Geophys. Res. Lett., 28: 1591-1594.
Uribe, T., J. Olivares, J. Moraga, J. Illanes, J. Rutllant, D. Boré, J. Blanco, C. Jélvez & G. Ledermann. 1995. Determinación de la capacidad de carga de la Bahía Inglesa (III Región) y Tongoy (IV Región). Informe Final FIP-IT/ 93-28: 130 pp.
Valdés, J, L. Ortlieb, D. Gutiérrez, L. Marinovic, G. Vargas & A. Sifeddine. 2008. A 250 years – sedimentary record of sardine and anchovy scale deposition in Mejillones Bay, 23ºS, Northern Chile. Prog. Oceanogr., 79: 198-207.
Vargas, C., G. Valenzuela, S. Núñez & D. Arcos. 1996. Mortalidad natural de larvas de merluza común (Merluccius gayi gayi) frente a Chile centro-sur en relación a su distribución espacial y grado de agregación. Invest. Mar., Valparaíso, 24: 19-30.
Vargas, C.A. & L. Castro. 2001. Spawning of the Chilean hake (Merluccius gayi gayi) in the upwelling system off central-southern Chile in relation to oceanographic and biological features. Sci. Mar., 65(2): 101-110.
Vásquez, S. 2012. Modelación de la historia de vida temprana del jurel (Trachurus murphyi) en el Pacífico suroriental: influencia de factores biofísicos en la
27

Latin American Journal of Aquatic Research
conectividad de zonas de desove-crianza. Tesis de Magister en Pesquerías. Universidad de Concepción, Concepción, 144 pp.
Vega, A., Y. Du-Penhoat, B. Dewitte & O. Pizarro. 2003. Equatorial forcing of interannual Rossby waves in the eastern South Pacific, Geophys. Res. Lett., (30)5: 1197.
Walters, C. & J. Korman. 1999. Linking recruitment to trophic factors: revisiting the Beverton-Holt recruit-ment model from a life history and multispecies perspective. Rev. Fish Biol. Fish., 9: 187-202.
Walters, C.J. & A. Parma. 1996. Fixed exploitation rate strategies for coping with the effects of climate change. Can. J. Fish. Aquat. Sci., 53: 110-23.
Werner, F.E., A. Aretxabaleta & K.P. Edwards. 2004. Modelling marine ecosystems and their environ-mental forcing. In: N.C. Stenseth, G. Ottersen, J.W. Hurrell & A. Belgrano (eds.). Marine ecosystems and climate variation: the North Atlantic. Oxford University Press, Oxford, pp. 33-46.
Wiff, R. & R. Quiñones. 2004. Parametrización ambiental en modelos biológico-pesqueros. Una revisión. Gayana, 68(1): 76-92.
Wolff, M. 1987. Population dynamics of the Peruvian scallop Argopecten purpuratus during the El Niño phenomena of 1983. Can. J. Fish. Aquat. Sci., 44: 1684-1691.
Xu, H., S.P. Xie, Y. Wang & R.J. Small. 2005. Effects of Central American mountains on the eastern Pacific winter ITCZ and moisture transport. J. Climate, 18: 3856-3873.
Xue, H., L. Incze, D. Xu, N. Wolff & N. Pettigrew. 2008. Connectivity of lobster populations in the coastal gulf of Maine-Part I: circulation and larval transport potential. Ecol. Model., 210(1-2): 193-211.
Yannicelli, B., L. Castro, C. Parada, W. Schneider, F. Colas & D. Donoso. 2012. Distribution of Pleuron-codes monodon larvae over the continental shelf of south central Chile: field and modelling evidence for partial local retention and transport. Prog. Oceanogr., 92-95: 206-227.
Yáñez, E., A. González & M.A Barbieri. 1995. Estruc-tura térmica superficial del mar asociada a la distribución espacio-temporal de sardina y anchoveta en la zona norte de Chile entre 1987 y 1992. Invest. Mar., Valparaíso, 23: 123-147.
Received: 6 June 2012; Accepted: 26 February 2013
Yáñez, E., L. Pizarro, M.A. Barbieri & O. Barra. 1985. Dinámica del stock de langosta (Jasus frontalis, H. Milne Edwards 1837) explotado en el archipiélago de Juan Fernández (30º40'S-80ºW). En: P. Arana (ed.). Investigaciones marinas en el archipiélago de Juan Fernández. Universidad Católica de Valparaíso, Valparaíso, pp. 251-271.
Yáñez, E., M.A. Barbieri, C. Silva, K. Nieto & F. Espíndola. 2001. Climate variability and pelagic fisheries in northern Chile. Prog. Oceanogr., 49: 581-596.
Yáñez, E., S. Hormazábal, C. Silva, A. Montecinos, M.A. Barbieri, A. Valdenegro, A. Órdenes & F. Gómez. 2008. Coupling between the environment and the pelagic resources exploited off northern Chile: ecosystem indicators and a conceptual model. Lat. Am. J. Aquat. Res., 36(2): 159-181.
Yoo, S., H. Batchelder, W. Peterson & W. Sydeman. 2008. Seasonal, interannual and event scale variation in North Pacific ecosystems. Prog. Oceanogr., 77(2-3): 155-181.
Yuras, G., O. Ulloa & S. Hormazábal. 2005. On the annual cycle of coastal and open ocean satellite chlorophyll of Chile (18º-40ºS). Geophys. Res. Lett. 32, L23604. doi:10.1029/2005GL023946.
Zhang, C. 2005. The Madden Julian Oscillation, Rev. Geophys., RG2003, 43 pp.
Ziegler, P.E., M. Haddon, S.D. Frusher & C. Johnson. 2004. Modelling seasonal catchability of the southern rock lobster Jasus edwardsii by water temperature, moulting and mating. Mar. Biol., 145: 179-190.
Ziegler, P., S. Frusher & C. Johnson. 2003. Space-time variation in catchability of southern rock lobster Jasus edwardsii in Tasmania explained by environ-mental, physiological and density-dependent proce-sses. Fish. Res., 61(1-3): 107-123.
Ziegler, P., C. Johnson, S. Frusher & C. Gardner 2002. Catchability of southern rock lobster Jasus edwardsii. II Efects of size. Mar. Freshw. Res., 53: 1149-1159.
28





























