Embed Size (px)
DESCRIPTION
vet pathol
Citation preview

http://vet.sagepub.com/Veterinary Pathology Online
http://vet.sagepub.com/content/39/5/536The online version of this article can be found at:
DOI: 10.1354/vp.39-5-536
2002 39: 536Vet PatholK. P. Carmichael, D. Bienzle and J. J. Mcdonnell
Feline Leukemia Virus-associated Myelopathy in Cats
Published by:
http://www.sagepublications.com
On behalf of:
Pathologists.American College of Veterinary Pathologists, European College of Veterinary Pathologists, & the Japanese College of Veterinary
can be found at:Veterinary Pathology OnlineAdditional services and information for
http://vet.sagepub.com/cgi/alertsEmail Alerts:
http://vet.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
What is This?
- Sep 1, 2002Version of Record >>
by guest on August 29, 2014vet.sagepub.comDownloaded from by guest on August 29, 2014vet.sagepub.comDownloaded from

536
Vet Pathol 39:536–545 (2002)
Feline Leukemia Virus–associated Myelopathy in CatsK. P. CARMICHAEL, D. BIENZLE, AND J. J. MCDONNELL
Department of Veterinary Pathology, University of Georgia, Athens, GA (KPC); Department of Pathobiology, Universityof Guelph, Guelph, ON (DB); and Department of Clinical Sciences, School of Veterinary Medicine, Tufts University,
North Grafton, MA (JJM)
Abstract. Feline leukemia virus (FeLV) infection is associated with distinct neoplastic, hematologic, andimmunosuppressive diseases. Here we report on a novel neurologic syndrome in 16 cats infected with FeLVfor more than 2 years. Clinical signs consisted of abnormal vocalization, hyperesthesia, and paresis progressingto paralysis. The clinical course of affected cats involved gradually progressive neurologic dysfunction invari-ably resulting in euthanasia. Microscopically, white-matter degeneration with dilation of myelin sheaths andswollen axons was identified in the spinal cord and brain stem of affected animals. Neither neoplastic norhematologic diseases commonly associated with FeLV infection were present. Fungal and protozoal infectionin one animal was suggestive of impaired immune competence. Immunohistochemical staining of affectedtissues revealed consistent expression of FeLV p27 antigens in neurons, endothelial cells, and glial cells. Fur-thermore, proviral DNA was amplified from multiple sections of spinal cord as well as intestine, spleen, andlymph nodes. These findings suggest that in a proportion of chronically FeLV-infected cats, a virus evolvedwith cytopathic potential for cells in the central nervous system.
Key words: Cat; feline leukemia virus; myelopathy; neurotropism; retrovirus.
The feline leukemia virus (FeLV) is a common virusof domestic cats. Vaccines against the virus are widelyused; however, these appear to protect predominantlyagainst tumor formation and not against infection.20FeLV belongs to the oncogenic Retroviridae, a familyof viruses that inserts a double-stranded DNA copy ofits single-stranded RNA genome into the host cells’chromosomes. Once stably inserted, the viral genes be-have like cellular genes and may be transcribed intofunctional protein products or may remain latent forlong periods of time.10 Depending on the infecting vi-ral strain and on the host response, infection withFeLV may result in early viral clearance because of aneffective immune response or in viremia.10,20 In catsthat are viremic, multiple tissues, including bone mar-row, are infected and may retain the latent virus in-definitely. A proportion of viremic cats subsequentlymounts an immune response that results in viral clear-ance. Cats that remain persistently viremic may beclinically healthy or, more commonly, develop im-munosuppressive, hematologic, intestinal, or reproduc-tive disorders, neoplasms such as lymphoma or leu-kemia, or autoimmune diseases.19 Thus, FeLV infec-tion can result in a wide range of potential diseasemanifestations.The specificity of FeLV in infecting cells resides in
the glycoprotein envelope.10 Recombination of exog-enous FeLV subgroup A env genes with complemen-tary endogenous retroviral sequences can result in
FeLV viruses typed as subgroup B or C.6 Opportunitiesfor recombination are expanded during long periods ofactive viral replication and may result in viruses in-fecting endothelial or epithelial cells.20 We have iden-tified a neurologic syndrome in FeLV-positive cats thatappears to be associated with chronic FeLV infection(FeLV-associated myelopathy, FAM). The presentstudy seeks to understand more completely the path-omechanistic spectrum of this important feline patho-gen through characterization of the clinical, morpho-logic, and immunohistochemical features of FAM incats.
Materials and MethodsAnimals
Tissues from 16 cats with a neurologic syndromecharacterized mostly by progressive ataxia were avail-able for study. These cats were FeLV antigenemic (p27antigen detection, IDEXX, Westbrook, ME). Clinicaland history data were obtained from the submittingclinicians or from the necropsy reports. Formalin-fixedtissues were obtained from eight animals (cat Nos. 1–8) submitted for necropsy examination from 1994through 2001, and archived histologic sections fromeight additional cats (cat Nos. 9–16) with a similarneurologic syndrome. The clinical history and pathol-ogy of these cases are summarized in Table 1.Tissues from six control animals were used in the
study. Four of these animals were FeLV-seronegative
by guest on August 29, 2014vet.sagepub.comDownloaded from

Vet Pathol 39:5, 2002 537FeLV Myelopathy
specific pathogen free cats with no clinical signs ofmyelopathy (control cat Nos. 2–5). Two were FeLV-seropositive cats that had no clinical signs of neuro-logic disease (control cat Nos. 1 and 6). Spleen andspinal cord from a FeLV-positive cat with spinal lym-phoma (control cat No. 7) and preparations from apersistently FeLV-infected cell line (3,201 cells stablytransfected with the FeLV-61E plasmid by J. I. Mul-lins, AIDS Research and Reference Reagent Program)were used as positive controls.
Pathology
Samples from 16 cats euthanatized because of pro-gressive neurologic dysfunction were examined. Thetissues were immersion-fixed in 10% neutral bufferedformalin for less than 24 hours and routinely processedfor microscopic examination. Sections of cervical, tho-racic, and lumbar spinal cord, brain stem, cerebellum,and cerebrum from all cases were routinely stainedwith hematoxylin and eosin (HE). Selected replicatesections were stained with Luxol fast blue-cresyl violetand Bodian’s silver stain.
Immunohistochemistry
Five-micrometer sections of spinal cord and spleenfrom all the cases were deparaffinized at �70 C for35 minutes and immersed in Hemo-De (Fisher Scien-tific, Pittsburgh, PA) three times for 3 minutes each.Sections were rehydrated in alcohol, endogenous per-oxidases were quenched with 3% hydrogen peroxide(Sigma, St. Louis, MO), and antigenic sites were un-masked through incubation with citrate buffer (1 : 3 di-lution of Antigen Unmasking Solution, Vector Labo-ratories, Burlingame, CA) for 10 minutes at 96 C.Nonspecific reactivity was blocked with 10% horse se-rum (Atlanta Biologicals, Norcross, GA). Slides wereincubated overnight with goat anti-FeLVp27 antibody(ViroStat Laboratories, Portland, ME) diluted 1 : 100in a mixture of 10% goat serum in PBS/0.005% Tween20 (PBST).11 After extensive washing, the slides wereincubated at 37 C for 1 hour with a 1 : 250 dilution ofa mixture of biotinylated rabbit anti-goat antibody in10% goat serum/PBST (Vector Laboratories). Afterfurther washing and 1 hour exposure to VectastainElite ABC solution (Vectastain Elite ABC Kit, VectorLaboratories), specific staining was visualized withDAB (Vector). The sections were counterstained withhematoxylin and coverslipped. Immunohistochemicalstaining was graded as ���, ��, �, or 0 accordingto the intensity of color development and the numberof cells stained. Negative control tissues (from controlcat Nos. 1–6, FeLV-seronegative cats and FeLV-sero-positive cats lacking neurologic signs), positive controltissues (cytospin preparations of FeLV-infected lym-phocytes and sections from an FeLV-positive cat [con-
trol cat No. 7] with spinal cord lymphoma), and re-agent controls (nonimmune goat serum at the samedilution as the primary antibody and phosphate buff-ered saline as negative controls for the primary anti-body) were included in all assays. The identity of cellsimmunoreactive for FeLV antigens was verified bystaining replicate sections of p27-positive spinal cordfor glial fibrillary acidic protein (DAKO, Carpenteria,CA) and anti-galactosylceramidase (Chemicon, Te-mecula, CA).
Polymerase chain reaction assayGenomic DNA was extracted from 10-�m sections
of paraffin-embedded sections of brain, spinal cord,peripheral nerve, lung, spleen, intestine, bone marrow,lymph node, muscle, kidney, and eye of all cats. Inaddition, DNA was extracted from liver in six cats (catNos. 6, 7, 8, 10, 11, and 12), skeletal muscle in onecat (cat No. 2), and cerebrospinal fluid in another (catNo. 16). Sections were cut using sterile blades to pre-vent sample carryover. The samples were deparaffini-zed through two extractions with xylene and washedwith absolute ethanol. The tissue pellet was incubatedovernight at 37 C in 50 mM Tris/1 mM ethylenedi-amine-tetraacetic acid (EDTA)/0.5% Tween-20 with 1�g/�l proteinase K. DNA was extracted with phenol/chloroform/isoamyl alcohol, precipitated with sodiumacetate, and resuspended in 10 mM Tris/1 mM EDTAand the concentration determined. FeLV-specific pro-viral sequences were amplified from approximately 0.5�g of DNA in a nested polymerase chain reaction(PCR).11 The outer primers were 5� TTA CTC AAGTAT GTT CCC ATG 3� and 5� CTG GGG AGC CTGGAG ACT GCT 3�; they amplified a 166-bp segmentin the viral long terminal repeat in a 50-�l reaction.Cycling parameters consisted of 30 rounds of 94 C for30 seconds, 55 C for 30 seconds, and 72 C for 30seconds. Five microliters of the first-round productwas subjected to a second round of amplification withprimers 5� GGT TAA GCA CCT GGG CCC CGG 3�and 5� GCA GCG GCC TTG AAA CTT CTG 3� withidentical cycling parameters except for an annealingtemperature of 58 C. The final amplicon had a size of85 bp, and sequence determination confirmed comple-mentarity with published FeLV sequences. Positiveand negative controls, as well as all appropriate re-agent controls, consisted of genomic DNA from FeLV-seropositive and -seronegative cats, respectively.
ResultsClinical findingsThe clinical signs of FeLV-seropositive cats con-
sisted predominately of ataxia, hyperesthesia, and pa-resis progressing to paralysis (Table 1). Four cats (catNos. 3, 4, 7, and 11) had a history of weakness, leth-
by guest on August 29, 2014vet.sagepub.comDownloaded from

538 Vet Pathol 39:5, 2002Carmichael, Bienzle, and McDonnellTable1.
Summaryofclinicaldatafrom
FeLV
positivecatswithdegenerativemyelopathy.
Case
No.
Sex
Breed
Age(yr)
FeLV
Diagnosis
ClinicalFindings
ClinicalPathology
PathologicDiagnosis
1MN
DLH
8ELISA,PCR
Progressiveataxia,paralysis,
nodeeppain
Hematology:macrocytosis,
lymphopenia,rubricytosis
Degenerativemyelopathy,
lossofaxonsandmyelin
2FS
Siamese
11ELISA,PCR
Hyperesthesia,flybiting,atax-
ia,progressiveascending
paresisandparalysis
Hematology:lymphopenia;
CSF:NAF*;UA†:protein-
uria
Degenerativemyelopathy,
lossofaxonsandmyelin
3FS
Siamese
18PCR
Poorbalance,difficultywalk-
ing,catatonicbehavior,
weakness,ataxia,urinary
incontinence
Hematology:mildanemia,
lymphopenia;UA:protein-
uria
Degenerativemyelopathy,
lossofaxonsandmyelin
4FS
DLH
Adult
IFA
Abnormalvocalization,hyper-
esthesia,weaknessandleth-
argy
NA‡
Degenerativemyelopathy,
lossofaxonsandmyelin
5FS
DLH
Adult
PCR
Weightloss,abnormalbehav-
ior
NA
Degenerativemyelopathy,
lossofaxonsandmyelin
6FS
DLH
15PCR
Hyperesthesia,progressive
ataxia,anisocoria
Hematology:mildanemia,
lymphopenia;Chemistry:
uremia
Degenerativemyelopathy,
lossofaxonsandmyelin
7FS
DSH
5PCR
Disorientation,progressive
ataxia
NA
Leukoencephalomalacia
8MN
Manx
9ELISA,IFA,PCR
Lowermotorneurondysfunc-
tionthoraciclimbs,upper
motorneurondysfunction
pelviclimbs
Hematology:neutropenia,
lymphopenia;CSF:NAF;
BM§:granulocytichypo-
plasia
Degenerativemyelopathy,
protozoalencephalomyeli-
tis,mesentericmycotic
granulomata
9MN
DSH
7ELISA,PCR
Progressiveparaparesis,dif-
fusepainthroughoutspinal
region
Hematology:mildanemia,
macrocytosis,lymphope-
nia;CSF:NAF
Demyelinatingperipheral
neuropathy,degenerative
myocarditis
10MN
DMH
6ELISA,PCR
Progressiveataxia,paralysis,
nodeeppain,urinaryin-
continence
Hematology:lymphopenia;
Chemistry:uremia,in-
creasedCKactivity
Degenerativemyelopathy,
lossofaxonsandmyelin
11FS
DSH
10ELISA,PCR
Poorbalance,difficultywalk-
ing,catatonicbehavior,
weakness,ataxia,urinary
incontinence
NA
Walleriandegenerationand
axonopathyinspinalcord,
pulmonarystrongylosis,
vulvitis
12MN
DSH
8PCR
Ascendingprogressivelimb
paresis,depression,anisoco-
ria,hyperesthesia,recurrent
constipation
Hematology:lymphopenia;
CSF:NAF;Chemistry:
uremia
Walleriandegenerationand
axonopathyinspinalcord,
AscaridandStrongylein-
fection
13MN
DSH
8ELISA,PCR
Weakness,difficultywalking,
progressiontoparesisand
paralysis
Hematology:NAF;Chemis-
try:increasedCKactivity;
CSF:NAF
Poliomyelomalacia,astrocy-
tosis,skeletalmuscleatro-
phy
by guest on August 29, 2014vet.sagepub.comDownloaded from

Vet Pathol 39:5, 2002 539FeLV MyelopathyTable1.
Continued.
Case
No.
Sex
Breed
Age(yr)
FeLV
Diagnosis
ClinicalFindings
ClinicalPathology
PathologicDiagnosis
14M
DSH
Adult
PCR
Progressiveparaparesis,ataxia
Hematology:anemia,neutro-
penia;BM:granulocytic
hypoplasia;Chemistry:in-
creasedALTandALPac-
tivity
Degenerativemyelopathy
lossofaxonsandmyelin
15MN
DSH
2ELISA,IFA,PCR
Ataxia,propioceptivedeficits
followingenterotomysur-
geryforforeignbodyre-
moval
Hematology:lymphopenia;
Chemistry:mildhypopro-
teinemia;CSF:NAF
Cervicalspinalcordaxono-
pathy,Walleriandegenera-
tion
16MN
DLH
10ELISA
Progressivehindlimbataxia,
proprioceptivedeficitsin
hindlimbs,keratoconjuncti-
vitissicca
Hematology:neutropenia,
lymphopenia;BM:NAF;
CSF:NAF
Degenerativemyelopathy,
lossofaxonsandmyelin
*Noabnormalfindings.
†Urinalysis.
‡Notavailable.
§Bonemarrow.
argy, abnormal behavior, and disorientation. Two cats(cat Nos. 6 and 12) had anisocoria with a diminishedpapillary light reflex, whereas another two (cat Nos. 1and 10) had no deep pain on neurologic examination.Urinary incontinence was seen in three cats (cat Nos.3, 10, and 11), and one cat (cat No. 11) had recurrentconstipation. All cats were FeLV-seropositive for morethan 4 years, except one that was FeLV-seropositivefor 2 years. Four cats had mild anemia (hematocrit0.20–0.30 L/L), one had macrocytosis (mean corpus-cular volume � 55 fL) without anemia, 10 cats hadlymphopenia (�1.5 � 109/L), and three of 12 cats hadmild neutropenia (2.0–2.5 � 109/L). Thrombocytope-nia was not detected, and mean platelet volumes wereinconsistently reported because of frequent plateletclumping. Changes on the biochemistry panel in af-fected cats were nonspecific. Findings from cerebro-spinal fluid examination were unremarkable in all ofseven cases examined.
Gross pathologic findings
Neither typical FeLV-associated hematologic abnor-malities nor tumors were identified grossly or micro-scopically in any of the cats. One cat (No. 8) had mul-tiple nodular masses in the mesentery and lungs thatwere gray-yellow and firm. There was no gross evi-dence of central nervous system (CNS) involvementin this cat.
Histopathology
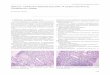
Histologically, all affected cats had widespread le-sions in the spinal cord that most often involved theventromedial funiculus, the superficial lateral funiculi,and the deep ventromedial funiculus (Fig. 1). The le-sions consisted of diffuse white-matter degenerationcharacterized by dilated myelin sheaths (Fig. 2). Someof these dilated myelin sheaths were devoid of axons,whereas others contained either intact or swollen axons(Fig. 3). In some areas, myelin sheaths devoid of cen-tral axons and containing infiltrating gitter cells wereprominent (Fig. 3). There appeared to be a mild de-crease in the numbers of neurons present in the ventralhorns of some affected cats when compared with con-trol animals. Luxol fast blue-cresyl stain showed dif-fuse staining, and there was a decrease in staining es-pecially in the peripheral spinal cord white matter.Bodian’s silver stain revealed axonal material in gittercells. In three cases, the thoracic spinal cord appearedto be more severely affected, whereas in the other cas-es no difference in the severity of lesions in differentsegments of the cord was noted. Sections of peripheralnerves did not contain lesions. Cat No. 8 had multiplegranulomas in the mesentery and the lungs associatedwith the pigmented fungus Cladosporium sp. as iden-
by guest on August 29, 2014vet.sagepub.comDownloaded from

540 Vet Pathol 39:5, 2002Carmichael, Bienzle, and McDonnell
Fig. 1. Spinal cord; FeLV-associated myelopathy–af-fected cat. This low-power photomicrograph shows the dis-tribution of the vacuolar change in the spinal cord. HE. Bar 300 �m.Fig. 2. Spinal cord; FeLV-associated myelopathy–af-
fected cat. The ventral funiculus of the thoracic cord hasdiffuse vacuolar change (arrows). HE. Bar 145 �m.Fig. 3. Spinal cord; FeLV-associated myelopathy–affect-
ed cat. Dilated myelin sheaths contain both gitter cells (largearrows) and intact axons (small arrows). HE. Bar 85 �m.
tified by culture. This cat also had disseminated Toxo-plasma infection involving the spinal cord.
Immunohistochemistry
Positive FeLV p27 immunostaining was present inthe sections of spinal cord examined (Table 2). Thecells stained were glial cells, neurons, and endothelialcells (Table 3). In these cells, diffuse, intense stainingwas present involving the entire cell. In neurons, thecytoplasm of both cell bodies and processes were im-munoreactive for p27 (Fig. 4). Staining of neurons wasnot seen in nonimmune serum tissue controls (Fig. 5)or in p27-stained controls that were FeLV-negative(Fig. 6). Endothelial cell staining was seen predomi-nately in smaller blood vessels in both the gray andwhite matter (Fig. 7). The larger blood vessels of themeninges and ventral median fissure stained less con-sistently. Glial cells stained intensely, and in many cas-es, the immunoreactivity obliterated the nucleus (Fig.7 inset). Control slides of a lymphoma (spinal) andspleen from an FeLV-positive cat showed similar stain-ing. Glial cell neurons and endothelial cells of the neg-ative controls did not express p27 antigen.Neurons and endothelial cells were readily identifi-
able on routine immunohistopathology. The identity ofglial cells immunoreactive for anti-FeLV antigens wasverified by staining replicate sections for glial fibrillaryacidic protein (GFAP; astrocytes) and galactosylcer-amidase (GALC; oligodendroglia). Although both ol-igodendroglial cells and astrocytes expressed p27 pro-tein, in general, sections of spinal cord stained withGFAP and GALC revealed that there were relativelyfewer immunoreactive astrocytes than oligodendroglialcells.
PCR assay
Multiple samples including spinal cord, spleen, andbone marrow sections from affected cats containedamplifiable proviral sequences (Table 2). FeLV pro-viral DNA was extracted from bone marrow of sevencats (cat Nos. 1, 2, 6, 7, 8, 11, and 14). In five cats(cat Nos. 1, 2, 6, 7, 8, and 12), brain, spinal cord,spleen, and intestine were all positive, and in five cats(cat Nos. 3, 4, 5, 13, and 14) proviral DNA was foundin spinal cord only. Lymph node was positive for pro-viral DNA in six cats (cat Nos. 1, 2, 6, 7, 8, and 12),and kidney was positive in only three cats (cat Nos. 1,2, and 12). Cerebrospinal fluid was positive in onlyone cat (cat No. 16). DNA extracted from eyes con-sistently yielded negative results (Fig. 8).
DiscussionIn this report, we have described the clinical and
pathologic features of a novel neurologic syndrome inFeLV-infected cats. FeLV is an oncogenic retrovirus
by guest on August 29, 2014vet.sagepub.comDownloaded from

Vet Pathol 39:5, 2002 541FeLV Myelopathy
Table 2. Results of FeLV detection by PCR and immunohistochemistry.
Case No. Tissues with Proviral DNA Tissues Without Proviral DNA p27 Positive Cells
1 Brain, spinal cord,lung, spleen, intes-tine, bone marrow,lymph node, kidney
Eyes Endothelial cells, neurons
2 Brain, spinal cord,lung, spleen, muscle,intestine, lymphnode, kidney
Eyes Endothelial cells, neurons, glial cells
345
Spinal cordSpinal cordSpinal cord
———
Neurons, glial cellsEndothelial cellsEndothelial cells, neurons, glial cells
6 Brain, spinal cord, in-testine, lymph node,bone marrow, liver,spleen
Peripheral nerve Endothelial cells, neurons, glial cells
7 Brain, spinal cord, in-testine, lymph node,bone marrow, liver,spleen
Peripheral nerve, eyes Neurons
8 Brain, spinal cord,bone marrow, intes-tine, liver, spleen,lymph node
Eyes Endothelial cells, neurons, glial cells
9 Spinal cord — Endothelial cells, neurons, glial cells10 Brain, spinal cord,
skin, kidney, liver,spleen, adrenals,bone marrow, lymphnode
— Endothelial cells
11 Brain, spinal cord,skin, kidney, liver,spleen, adrenals,bone marrow
— Endothelial cells
12 Lung, brain, spinalcord, lymph node,kidney, liver, spleen,intestine
Peripheral nerve, eyes Endothelial cells, neurons, glial cells
1314
Spinal cordSpinal cord
——
Endothelial cellsEndothelial cells
15 Brain, spinal cord, liv-er, intestine, spleen
— Endothelial cells, neurons
16 Brain, spinal cord,spleen, liver, CSF,bone marrow
— Endothelial cells, neurons
Control 1 FeLV� Spinal cord, spleen,kidney
— Endothelial cells, glial cells
Control 2 FeLV�Control 3 FeLV�Control 4 FeLV�Control 5 FeLV�Control 6 FeLV�Control 7 FeLV�
——————
Spinal cordSpinal cordSpinal cordSpinal cordSpinal cord—
—————Neoplastic lymphocytes
by guest on August 29, 2014vet.sagepub.comDownloaded from

542 Vet Pathol 39:5, 2002Carmichael, Bienzle, and McDonnell
Table 3. Relative staining of spinal cord cells with p27 immunohistochemistry.
Case No. Relative Staining p27 Positive Cells
12345678910111213141516Control 1 FeLV�Control 2 FeLV�Control 3 FeLV�Control 4 FeLV�Control 5 FeLV�Control 6 FeLV�Control 7 FeLV�
������������� (few)���������������������������
Endothelial cells, neuronsEndothelial cells, neurons, glial cellsNeurons, glial cellsEndothelial cellsEndothelial cells, neurons, glial cellsEndothelial cells, neurons, glial cellsNeuronsEndothelial cells, neurons, glial cellsEndothelial cells, neurons, glial cellsEndothelial cellsEndothelial cellsEndothelial cells, neurons, glial cellsEndothelial cellsEndothelial cellsEndothelial cells, neuronsEndothelial cells, neuronsEndothelial cells, glial cells—————Neoplastic lymphocytes
* � mild staining; �� moderate staining; ��� intense staining; � no staining.
associated with a wide range of diseases. Strong as-sociations have been reported for T-cell lymphomas inyoung cats, pure red-cell aplasia in cats infected withsubtype C, erythroid myeloproliferative disease, and arapidly progressive immunodeficiency syndrome.20Less well characterized are associations of FeLV in-fection with chronic diarrhea, infertility, and hemato-logic abnormalities such as neutropenia, macrocyticanemia, or both.19 Abnormal neurologic findings havebeen reported as a common feature in a retrospectivesurvey of lesions observed at necropsy of FeLV-se-ropositive cats and in one case of degenerative mye-lopathy in a FeLV-seropositive cat.14,19 In addition, de-scriptions of behavioral and nerve conduction abnor-malities in experimentally infected cats suggest thatneurologic lesions are not an entirely new sequela ofFeLV infection.4,9 However, detailed descriptions of ei-ther the clinical or pathologic characteristics have beenlacking. Hind limb paresis progressing to paralysiswithin approximately 1 year was the most commonclinical abnormality in affected cats. There was evi-dence of neither peripheral nerve deficits nor corticalor brain stem lesions. Lower motor neuron dysfunctionmanifested with urinary incontinence in two cats. He-matologic abnormalities typically associated withFeLV infection (anemia, macrocytosis, rubricytosis,neutropenia, macrothrombocytes, and thrombocytope-nia) were noted in four of 12 cats.5,21 The hematologic
changes were mild but likely a consequence of chronicFeLV infection of hematopoietic precursor cells. Lym-phopenia noted in most of the cats in this study (10of 12 cats examined) was attributed to steroid-inducedleukocyte kinetic changes or immunosuppression re-sulting from cytopathic FeLV infection. Overt immu-nodeficiency, supported by the lymphopenia, may haveresulted in systemic mycotic as well as protozoal in-fections in cat No. 8 and may have contributed to theextensive metazoan infections in cat Nos. 11 and 12.In cases in which serologic testing was done, the
time from the first seropositive test result until the ap-pearance of neurologic signs ranged from 1 to 6 yearswith a mean of 3 years. This time span is probably anunderestimate because the majority of cats were testedfor FeLV infection only after clinical signs becameapparent. Further, the average age of cats with knownbirth dates was 9 years, and FeLV infection is mostcommonly acquired at a young age.10 Tumors were notnoted in any of the cats reported here. In summary,although clinical pathology abnormalities in this groupof cats were either nonspecific or absent, the neuro-logic findings were striking.Although there are etiologic agents that may induce
myelitis and could result in the clinical signs observed,degenerative myelopathy in cats is a rare histopatho-logic diagnosis, typically of unknown etiology. Theassociation of degenerative myelopathy with long-
by guest on August 29, 2014vet.sagepub.comDownloaded from

Vet Pathol 39:5, 2002 543FeLV Myelopathy
Fig. 4. Spinal cord; FeLV-associated myelopathy–affected cat. Positive staining of neurons with anti-p27 antibody(arrows); DAB chromogen. Hematoxylin counterstain. Bar 85 �m.Fig. 5. Spinal cord; FeLV-associated myelopathy–affected cat. No staining of neurons is noted with nonimmune serum.
DAB chromogen. Hematoxylin counterstain. Bar 85 �m.Fig. 6. Spinal cord; FeLV-negative cat. There is no staining of neurons with anti-p27 antibody. DAB chromogen.
Hematoxylin counterstain. Bar 85 �m.Fig. 7. Spinal cord; FeLV-associated myelopathy–affected cat. Positive staining of endothelial cell (center), neuron
(arrow), and glial cells (inset) with anti-p27 antibody. DAB chromogen. Hematoxylin counterstain. Bar 85 �m. Inset:Bar 40�m.
standing FeLV infection in all 16 cats reported heresuggests that the lesions result from the viral infection.In the cats of this study, glial cells (oligodendroglia
and astrocytes) were frequently involved. In vivo in-fection of these cells with FeLV has not been previ-ously reported; however, experimental infection ofbrain cultures with FeLV has been described.9 It is thusnot surprising that these cells were frequently found tocontain immunoreactive viral antigen. Endothelialcells were also commonly affected, and this has pre-viously been described in vitro with a strain of recom-binant FeLV.6 This immunoreactivity is possibly a re-flection of the systemic viremia observed and may in-dicate one possible route of entry into the CNS. In the
CNS, there is close association between capillary en-dothelial cells and astrocytes both physically and func-tionally in the blood-brain barrier, and it is possiblethat viral infection of the endothelial cells may havebeen transferred to astrocytes because of this relation-ship.7,12 It was somewhat surprising to identify viralantigen in neurons because direct in vivo infection ofthese cells has not previously been speculated to occur.But, in vitro experiments have shown FeLV-associatedneuronal infection and damage.8,15 It is possible thatthe supportive role of the astrocytes to neurons andoligodendroglia in the CNS may have facilitated neu-ronal infection.Viral infection of the CNS may have also occurred
by guest on August 29, 2014vet.sagepub.comDownloaded from

544 Vet Pathol 39:5, 2002Carmichael, Bienzle, and McDonnell
Fig. 8. Electrophoretogram of PCR-amplified FeLVDNA. Lane 1, markers (50-bp ladder); 2–5, spinal cord; 6,7, eye; 8, 9, intestine; 10, muscle; 11, positive control; and12, negative control.
subsequent to an earlier episode of lymphocyte infil-tration that receded before the onset of clinical signs.This is a common route of viral dissemination in manyretroviral infections, and a recent histologic observa-tion of perivascular lymphocytic cuffing in a euthan-atized nonneurologic FeLV-positive cat by one of theauthors (K. P. Carmichael) suggests that this is a pos-sibility. The reason for the increased numbers of im-munoreactive oligodendroglia as compared with astro-cytes in this study is not known, but oligodendroglialinfection could be responsible for the degeneration ofaxonal sheaths. Conversely, neuronal infection may di-rectly result in a diffuse axonopathy or in calcium im-balances leading to neuronal dysfunction and death.FeLV subtype A is the prototype of FeLV that is
transmitted among cats through salivary contact. Otherviral subtypes evolve from subtype A and are definedby interference with superinfection in indicator celllines and interaction with different cellular receptors.FeLV subtype A will infect epithelial cells, lympho-cytes, and a range of other cells and is minimally cy-topathic. Infection restricted to the oronasal cavitywith immune clearance of virus localized to craniallymphoid tissues is thought to be the most commonscenario subsequent to salivary exposure with FeLV.Cats infected by this route rapidly clear the virus andare presumed to never become antigenemic and thusto never test FeLV-antigen positive by commonly usedserum enzyme-linked immunosorbent assay tests.14The second most common sequel to oronasal infectionresults in viremia followed by a systemic immune re-sponse and clearance of virus after a more protractedtime period. Cats in which this occurs will test antigen-positive in serum, and viral antigens can be detectedby immunofluorescent labeling of bone marrow cells.It is thought that cell-associated virus is cleared beforethe disappearance of antigenemia.10 Finally, someFeLV-infected cats fail to mount an effective immune
response and become persistently viremic or, on rareoccasions, have persistent latent infection in select cellpopulations. Considering the time span between initialseropositivity, the development of clinical disease, andretesting, it is likely that the cats in this study werepersistently viremic. Persistent replication of FeLV of-fers opportunities for the development of viral diver-sity through the acquisition of nucleotide changes orthrough recombination within the envelope gene ofFeLV. The surface unit of the envelope gene definesreceptor interaction, and therefore cell tropism. Path-ogenic properties of the virus within specific target cellpopulations appear to result from initial infection withFeLV subtype A and subsequent evolution of viruseswith enhanced ability to use an alternate receptor thatis expressed in a cell-restricted manner.16 This conceptwas illustrated by the recent identification of an ac-cessory cellular protein termed FeLIX, which is ex-pressed and secreted by feline T-cells.1 FeLIX supportsreplication of a variant of FeLV subtype A that is T-cell–tropic and results in fatal immunodeficiency.1Similarly, use of a cell-restricted receptor results in thedevelopment of pure red-cell aplasia in cats with aprorubricyte-tropic FeLV variant broadly classified asFeLV subtype C.17 This virus results in impairment ofthe transition of erythroid precursors from the burst-forming unit–erythroid (BFU-E) to the colony-formingunit–erythroid (CFU-E).17 The receptor used by thisvirus encodes an organic anion transporter protein thatis speculated to be involved in the accumulation ofheme compounds necessary for the synthesis of he-moglobin at the BFU-E to CFU-E transition stage.17Another membrane transporter protein, Pit1, involvedin the movement of phosphate across cell membranes,is the receptor used by FeLV subtype B.18 FeLV-B isa variant of FeLV subtype A that arises because ofrecombination with portions of endogenous cat envgenes, and it has the unique ability to infect manydifferent mammalian cells.3 This virus variant contain-ing truncations or mutations in the Env protein is pre-sent in many FeLV-associated malignancies affectinglymphoid cells.13 The syndrome described in this re-port does not bear resemblance to typical diseases as-sociated with FeLV subtypes B, C, or T. Thus, pre-sumptively, the virus may still be of subtype A, oralternatively, a new variant with particular neurotro-pism may be involved. Studies are underway to char-acterize the viral variant involved in this feline degen-erative myelopathy. The histopathologic changes ob-served are similar to those described in human T-cellleukemia virus (HTLV)-1–associated myelopathy ortropical spastic paresis.2 Infection with this human ret-rovirus induces T-cell leukemia/lymphoma in 20–50%and myelopathy in approximately 5% of infected in-dividuals.2 Inflammatory cells in the spinal cord are
by guest on August 29, 2014vet.sagepub.comDownloaded from

Vet Pathol 39:5, 2002 545FeLV Myelopathy
present transiently during the active/chronic stage, andlesions during the chronic stage of the disease consistpredominantly of degeneration of the lateral and dorsalspinal tracts and endothelial fibrosis.2 Whether thepathogenesis or epidemiology of the feline myelopathydescribed in this report is similar to HTLV-associatedmyelopathy in humans is currently unknown.
AcknowledgementsWe would like to thank Roberta Mihut, Amanda Hobe,
and Dr. James Stanton for technical assistance. We also ap-preciate feline spinal cord samples sent to us by Dr. PatriciaBlanchard and Dr. Thomas Van Winkle. This study wasfunded by a grant from the University of Georgia VeterinaryMedical Experiment Station.
References1 Anderson MM, Lauring AS, Burns CC, Overbaugh J:Identification of a cellular cofactor required for infectionby feline leukemia virus. Science 287:1828–1830, 2000
2 Aye MM, Mutsuoka E, Moritoyo, Umehara F, SueharaM, Hokezu Y, Yamanaka H, Isashiki Y, Osame M, IzumoS: Histopathological analysis of four autopsy cases ofHTLV-1 associated myelopathy/tropical spastic parapa-resis: inflammatory changes occur simultaneously in theentire central nervous system. Acta Neuropathol 100:245–252, 2000
3 Boomer S, Eiden M, Burns CC, Overbaugh J: Three dis-tinct envelope domains, variably present in subgroup Bfeline leukemia virus recombinants, mediate Pit1 andPit2 receptor recognition. J Virol 71:8116–23, 1997
4 Boomer S, Gasper P, Whalen LR, Overbaugh J: Isolationof a novel subgroup B feline leukemia virus from a catinfected with FeLV-A. Virology 204:805–810, 1994
5 Boyce JT, Kociba GJ, Jacobs RM, Weiser MG: Felineleukemia virus-induced thrombocytopenia and macroth-rombocytosis in cats. Vet Pathol 23:16–20, 1986
6 Chakrabati R, Hofman FM, Pandey, Mathes LE, Roy-Burman P: Recombination between feline exogenous andendogenous retroviral sequences generates tropism forcerebral endothelial cells. Am J Pathol 144:348–358,1994
7 Chaudhuri JD: Blood brain barrier and infection. MedSci Monit 6:1213–1222, 2000
8 Fails AD, Mitchell TW, Rojko JL, Whalen LR: An oli-gopeptide of the feline leukemia virus envelope glyco-protein is associated with morphological changes andcalcium dysregulation in neuronal growth cones. J Neu-rovirol 3:179–191, 1997
9 Haffer KN, Sharpee RL, Beckenhauer W, Koertje WD,Fanton RW: Is feline leukemia virus responsible for neu-rologic conditions in cats? Vet Med 8:802–805, 1987
10 Hoover EA, Mullins JI: Feline leukemia virus infectionand disease. J Am Vet Med Assoc 199:1392–1401, 1991
11 Jackson ML, Haines DM, Meric SM, Misra V: Felineleukemia virus detection by immunohistochemistry andpolymerase chain reaction in formalin-fixed, paraffin-embedded tumor tissue from cats with lymphosarcoma.Can J Vet Res 57:269–276, 1993
12 Mayhan WG: Regulation of blood-brain barrier perme-ability. Microcirculation 8:89–104, 2001
13 McDougall AS, Terry A, Tzavaras T, Cheney C, RojkoJ, 14. Neil JC: Defective endogenous proviruses are ex-pressed in feline lymphoid cells: evidence for a role innatural resistance to subgroup B feline leukemia viruses.J Virol 68:2151–2160, 1994
14 Mesfin GM, Kusewitt D, Parker A: Degenerative mye-lopathy in a cat. J Am Vet Med Assoc 176:62–64, 1980
15 Mitchell TW, Rojko JL, Hartke JR, Mihajlov AR, Ka-sameyer GA, Gasper PW, Whalen LR: FeLV envelopeprotein (gp70) variable region 5 causes alterations in cal-cium homeostasis and toxicity of neurons. J Acquir Im-mune Defic Syndr Hum Retrovirol 14:307–320, 1997
16 Moser M, Burns CC, Boomer S, Overbaugh J: The hostrange and interference properties of two closely relatedfeline leukemia variants suggest that they use distinctreceptors. Virology 242: 366–377, 1998
17 Quigley JG, Burns CC, Anderson MM, Lynch ED, SaboKM, Overbaugh J, Abkowitz JL: Cloning of the cellularreceptor for feline leukemia virus subgroup C (FeLV-C),a retrovirus that induces red cell aplasia. Blood 95:1093–1099, 2000
18 Rasko JE, Battini JL, Gottschalk RJ, Mazo I, Miller AD:The RD114/simian type D retrovirus receptor is a neutralamino acid transporter. Proc Natl Acad Sci USA 96:2129–2134, 1999
19 Reinacher M: Diseases associated with spontaneous fe-line leukemia virus (FeLV) infection in cats. Vet Im-munol Immunopathol 21:85–95, 1989
20 Rohn JL, Overbaugh J: Pathogenic feline retroviruses:feline leukemia virus and feline immunodeficiency virus.In: Persistent Viral Infections, ed. Chen SY and AhmedR, pp. 379–408. John Wiley & Sons, Inc., New York,NY, 1999
21 Weiser MG, Kociba GJ. Erythrocyte macrocytosis in fe-line leukemia virus associated anemia. Vet Pathol 20:687–697, 1983
Request reprints from Dr. K. Paige Carmichael, Department of Pathology, College of Veterinary Medicine, University ofGeorgia, Athens, GA 30602 (USA). E-mail: [email protected].
by guest on August 29, 2014vet.sagepub.comDownloaded from