Embed Size (px)
Citation preview

1
Wnt/β-catenin pathway represses let-7 microRNAs 1
expression via transactivation of Lin28 to augment breast 2
cancer stem cell expansion 3
4
Running title: Wnt/β-catenin pathway /Lin28/let-7 cascade in 5
breast cancer stem cells 6
7
Wang-Yu Cai1†, Tong-Zhen Wei1†, Qi-Cong Luo2†, Qiu-Wan Wu2, Qing-Feng Liu1, 8
Meng Yang1, Guo-Dong Ye1, Jia-Fa Wu1, Yuan-Yuan Chen1, Guang-Bin Sun1, 9
Yun-Jia Liu1, Wen-Xiu Zhao3, Zhi-Ming Zhang2,*, and Bo-An Li1,* 10
11
12
1State Key Laboratory of Cellular Stress Biology, School of Life Sciences, Xiamen University, 13
Xiamen, Fujian, China 14
2The First Affiliated Hospital of Xiamen University, Xiamen, Fujian, China 15
3 Zhongshan Hospital of Xiamen University, Xiamen, Fujian, China 16
17
* Correspondence 18
Mailing address for Bo-An Li: Department of Biomedical Sciences, Xiamen University School of 19
Life Sciences, 422 South Siming Rd., Xiamen, 361005, Fujian province, China 20
Phone: 86-592-2181987; Fax: 86-592-2181984. E-mail: [email protected]. 21
Mailing address for Zhi-Ming Zhang: Department of Breast Surgery, The First Affiliated Hospital 22
of Xiamen University, 55 Zhenhai Rd., Xiamen, 361003, Fujian province, China 23
Phone: 86-592-2137507; Fax: 86-592-2137509. E-mail: [email protected] 24
25
† These authors contributed equally to this work. 26
27
© 2013. Published by The Company of Biologists Ltd.This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Share Alike License(http://creativecommons.org/licenses/by-nc-sa/3.0), which permits unrestricted non-commercial use, distribution and reproduction inany medium provided that the original work is properly cited and all further distributions of the work or adaptation are subject to the same Creative Commons License terms.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t
JCS Advance Online Article. Posted on 23 April 2013

2
Summary 1
Wnt signaling through β-catenin and the lymphoid-enhancing factor 1/T-cell factor 2
(LEF1/TCF) family of transcription factors maintains stem cell properties in both 3
normal and malignant tissues; however, the underlying molecular pathway involved 4
in this process has not been completely defined. Using a microRNA microarray 5
screening assay, we identified the let-7 miRNAs as downstream targets of 6
Wnt/β-catenin pathway. Expression studies indicated that Wnt/β-catenin pathway 7
suppresses mature let-7 miRNAs but not the primary transcripts, which suggests a 8
posttranscriptional regulation of repression. Furthermore, we identified Lin28, a 9
negative let-7 biogenesis regulator, as a novel direct downstream target of 10
Wnt/β-catenin pathway. Loss of function of Lin28 impairs the Wnt/β-catenin 11
pathway-mediated let-7 inhibition and breast cancer stem cell expansion; enforced 12
expression of let-7 blocks the Wnt/β-catenin pathway-stimulated breast cancer stem 13
cell phenotype. Finally, we demonstrated that Wnt/β-catenin pathway induces Lin28 14
upregulation and let-7 downregulation in both cancer samples and mouse tumour 15
models. Moreover, the delivery of a modified lin28 siRNA or a let-7a agomir into the 16
premalignant mammary tissues of MMTV-wnt-1 mice resulted in a complete rescue 17
of the stem cell phenotype driven by Wnt/β-catenin pathway. These findings highlight 18
a pivotal role for Lin28/let-7 in Wnt/β-catenin pathway mediated cellular phenotypes. 19
Thus, Wnt/β-catenin pathway, Lin28, and let-7 miRNAs, three of the most crucial 20
stem cell regulators, connect in one signal cascade. 21
22
Keywords: breast cancer/ let-7/ Lin28/ stem cell/ Wnt/β-catenin pathway 23
24
25
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

3
Introduction 1
Multiple pathways are implicated in the regulation of stem cell self-renewal and 2
differentiation. These pathways include Hedgehog, Notch, and Wnt (Liu et al., 2010; 3
Merchant and Matsui, 2010; Takebe et al., 2011; Wend et al., 2010), within which 4
Wnt/β-catenin pathway is crucial in both normal mammary development and 5
tumorigenesis (Roarty and Rosen, 2010). β-catenin plays a central role as a 6
transcriptional activator in Wnt/β-catenin pathway (Logan and Nusse, 2004). In the 7
absence of upstream signalling stimulation, cytoplasmic β -catenin is phosphorylated 8
by glycogen synthase kinase-3β (GSK-3β) in a complex that includes Axin and the 9
tumour suppressor Adenomatous polyposis coli (APC) and is targeted for 10
ubiquitin-mediated proteasomal degradation. Stimulation by Wnt ligands leads to the 11
inhibition of phosphorylation and degradation of β-catenin, which subsequently enters 12
the nucleus and binds to the LEF1/TCF family of transcription factors. In turn, the 13
β-catenin–LEF1/TCF complex activates the expression of a cohort of target genes that 14
impact cellular functions (Amit et al., 2002; Liu et al., 2002). As Wnt/β-catenin 15
pathway can regulate mammary stem cell self-renewal and differentiation, the 16
aberrant activation of this pathway in human breast cancer may highlight a key role 17
for it in cancer stem cells. Despite the fact that Wnt/β-catenin pathway is a key 18
regulator of breast cancer stem cells, the downstream transcriptional cascade 19
accounting for this process remains largely unknown. 20
MicroRNAs (miRNAs) play critical roles in many biological processes, including 21
cancer processes, by directly inhibiting the expression of target mRNAs through a 22
variety of molecular mechanisms (Bartel, 2009; Ventura and Jacks, 2009). Also, 23
miRNAs undergo aberrant regulation during carcinogenesis and can act as either 24
oncogenes or tumour suppressors (Lu et al., 2005). Let-7 is an important miRNA 25
family consisting of 12 members with expression that is frequently downregulated in 26
a number of human cancers (Calin et al., 2004; Johnson et al., 2005; Kumar et al., 27
2008). Moreover, let-7 is also the key regulator of breast cancer stem cell self-renewal 28
and differentiation (Yu et al., 2007). A large body of evidence indicates that many 29
miRNAs can regulate Wnt/β-catenin pathway through the targeting of Wnt 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

4
components (Huang et al., 2010); however, little is known regarding whether 1
Wnt/β-catenin pathway regulates the expression of miRNAs, especially in cancer 2
stem cells. 3
Recent studies on iPS cells support the concept that cancer stem cells arise through 4
a reprogramming-like mechanism (Krizhanovsky and Lowe, 2009). By enforcing the 5
expression of so-called reprogramming factors (OCT4, KLF4, SOX2, and Lin28 or 6
c-MYC), differentiated somatic cells can be converted to iPS cells, which can 7
differentiate into any tissue type. Several of these reprogramming factors are 8
upregulated in human tumours and are considered to be putative oncogenes 9
(Krizhanovsky and Lowe, 2009). Thus, iPS cells and cancer stem cells may share 10
similar mechanisms that control the stemness of the cells. Lin28, one of these 11
reprogramming-like factors, is highly abundant in embryonic stem cells and 12
developing cells, but in later stages, its expression declines (Moss and Tang, 2003). 13
Lin28 may be overexpressed in some tumours (Viswanathan and Daley, 2010). 14
Moreover, Lin28 has been found to be positively correlated with the percentage of 15
ALDH1+ tumour cells in breast cancer, suggesting that Lin28 plays an important role 16
in the regulation of breast cancer stem cells (Yang et al., 2010). Lin28 and Lin28B 17
function as negative regulators of let-7 biogenesis through their ability to directly 18
interact with the loop region of let-7 hairpins. Lin28/Lin28B accelerates the turnover 19
of let-7 precursors and prevents both Drosha- and Dicer-mediated let-7 processing 20
(Heo et al., 2008; Newman et al., 2008; Piskounova et al., 2008; Rybak et al., 2008). 21
Conversely, let-7 targets Lin28 and downregulates its expression (Yang et al., 2010). 22
These results suggest that a regulatory feedback loop exists between let-7 and Lin28 23
(Yang et al., 2010) that is critical in modulating the self-renewal and differentiation of 24
breast cancer stem cells. 25
In this study, we have demonstrated that the activation of Wnt/β-catenin pathway 26
suppresses mature let-7 miRNAs but not the primary transcripts, which suggests a 27
posttranscriptional regulation of let-7 expression. We further identified Lin28 as a 28
novel direct downstream target of Wnt/β-catenin pathway. Accordingly, loss of 29
function of Lin28 impairs Wnt/β-catenin pathway-mediated let-7 inhibition and breast 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

5
cancer stem cell expansion. Moreover, enforced expression of let-7 blocks 1
Wnt/β-catenin pathway-stimulated breast cancer stem cell expansion. Thus, 2
Wnt/β-catenin pathway, Lin28, and let-7 miRNAs, three of the most crucial stem cell 3
regulators, connect in one signal cascade. 4
5
Results 6
Posttranscriptional repression of let-7 family miRNAs by Wnt/β-catenin 7
pathway 8
Studies have shown that miRNAs play pivotal roles in controlling the maintenance 9
and function of stem cells. We sought to identify the downstream target miRNAs of 10
Wnt/β-catenin pathway by screening the ZR-75-30 breast cancer cell line using a 11
miRNA microarray system. Specifically, miRNA expression profiles were examined 12
in high-Wnt and low-Wnt states with stable overexpression of β-catenin and GFP. All 13
miRNAs showing a 1.5-fold or greater upregulation or downregulation in the 14
high-Wnt state were chosen for further analysis (Table S1). Of these miRNAs, we 15
found that 7 members of the let-7 miRNA family are downregulated by approximately 16
2.5-fold in β-catenin stable overexpression cells and quantitative real-time PCR 17
(qPCR) results confirmed the changes (Figure 1A). To verify the expression changes 18
in more breast cancer cells and more Wnt stimulations, MDA-MB-231 and T-47D 19
cells were also used. Consistent with the results in β-catenin activated ZR-75-30 cells, 20
the expression levels of let-7a, let-7f and let-7g in these cells were dramatically 21
decreased when treated with LiCl, a GSK3β inhibitor, as well as stably 22
overexpressing of HA-Wnt1 (Figure 1B and Figure S1A). Conversely, knockdown of 23
β-catenin expression resulted in the induction of let-7a and let-7f in both ZR-75-30 24
and MDA-MB-231 cells (Figure 1C and Figure S1B). These results suggest that let-7 25
miRNAs are downstream target miRNAs of Wnt/β-catenin pathway in breast cancer 26
cells. 27
To investigate the mechanism through which Wnt/β-catenin pathway regulates let-7 28
miRNAs, we used qPCR analysis to examine further the abundance of their primary 29
transcripts in ZR-75-30 cells. Unexpectedly, we found that the expression of the 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

6
let-7a/let-7f/let-7d and let-7g primary transcripts was not repressed in β-catenin stable 1
overexpression cells (Figure 1D). Consistently, these primary transcripts were also not 2
induced in β-catenin knockdown cells (Figure 1E). These findings imply that 3
Wnt/β-catenin pathway utilises alternative mechanisms to downregulate multiple let-7 4
family members through a posttranscriptional pathway. 5
6
Wnt/β-catenin pathway activates Lin28 expression in breast cancer cells 7
Lin28 and Lin28B RNA binding proteins have been demonstrated to bind to the 8
stem loops of let-7 miRNAs and to inhibit their biogenesis by blocking Drosha- and 9
Dicer-mediated cleavage and accelerating the turnover of let-7 precursors (Heo et al., 10
2008; Newman et al., 2008; Piskounova et al., 2008; Rybak et al., 2008). Therefore, 11
we speculated that Wnt/β-catenin pathway may inhibit let-7 family members through 12
the transactivation of Lin28 or Lin28B. To verify this hypothesis, we used 13
semi-quantitative RT-PCR to examine Lin28 and Lin28B expression in different 14
breast cancer cell lines upon LiCl stimulation. While Lin28B expression was 15
undetected in 3 of 4 cell lines tested, and no changes were found in the ZR-75-30 cell 16
line, Lin28 expression was dramatically induced in all cell lines treated with LiCl 17
(Figure 2A, left). Similar results were achieved by qPCR analysis (Figure 2A, right). 18
Conversely, knockdown of β -catenin expression resulted in greatly reduced Lin28 19
expression in ZR-75-30 cells (Figure 2B). We previously demonstrated that Pygo2, a 20
newly found β-catenin interaction protein, is overexpressed in breast cancer stem cells 21
and augments Wnt/β-catenin pathway activity (Chen et al., 2010). We examined 22
whether Pygo2 knockdown also affects Lin28 expression. As shown in Figure 2C, 23
depletion of Pygo2 in ZR-75-30 cells caused a remarkable decrease in Lin28 24
expression. Next, we investigated whether the let-7 family members responded to 25
Lin28 in the breast cancer cells in our experiments. As expected, when Lin28 mRNA 26
and protein expression were effectively knocked down (Figure S2A), the levels of 27
mature let-7a and let-7f transcripts were greatly increased (Figure S2B), whereas 28
overexpression of Lin28 resulted in reduced mature let-7a and let-7f levels (Figure 29
S2C). In contrast, the primary transcript levels of let-7a/let-7f/let-7d and let-7g were 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

7
not changed in either experiment (Figure S2D and S2E). To further verify that both 1
β-catenin and Lin28 could inhibit let-7’s biological function as microRNAs, we 2
performed a luciferase assay using a let-7 sensor containing a constitutively expressed 3
luciferase reporter bearing two copies of sequences complementary to let-7a in the 4
downstream 3’ UTR. As shown in Figure 2D, endogenous let-7 miRNAs in the 5
MDA-MB-231 cells greatly inhibited let-7 sensor activity indicating that these 6
miRNAs indeed target the 3’ UTR of luciferase and inhibit its expression. Enforced 7
expression of β-catenin and Lin28 both remarkably rescued the sensor activity almost 8
to the control vector levels, which suggests that let-7’s biological function is inhibited. 9
Finally, we treated the MDA-MB-231, ZR-75-30 and T-47D cells with recombinant 10
Wnt1 to see weather the Wnt ligand also triggers the same activity. As expected, 11
β-catenin expression was up-regulated in all cell lines treated with recombinant Wnt1. 12
Moreover, Lin28 was up-regulated whereas let-7a/let-7f was down-regulated in these 13
cells (Figure 2E). The regulatory consistency of let-7 suggests that Wnt/β-catenin 14
pathway and Lin28 may share the same signalling cascade and supports the 15
hypothesis that Wnt/β-catenin pathway inhibits let-7 miRNAs by activating Lin28 16
expression in breast cancer cells. 17
18
Lin28 is a novel direct downstream target gene of Wnt/β-catenin pathway 19
To determine whether Lin28 is a direct Wnt/β-catenin pathway target gene, we 20
examined the genomic sequence within a 10-kb window centred on the transcriptional 21
start site (TSS) of Lin28. The genomic sequence extending from 1.0 kb upstream to 22
114 bp (ATG site) downstream of the TSS contains 7 consensus LEF/TCF binding 23
sites of CTTTG or GAAAC, with 4 sites showing conservation between human and 24
mouse (Figure 3A, conserved sites shown in black dots). This fragment was cloned 25
into a PGL3-basic promoter-less luciferase reporter cassette. The fragment was 26
sufficient to drive the luciferase reporter activity in a dose-dependent manner when 27
stimulated with LiCl, β-catenin or Wnt-1 in 293T cells (Figure 3B). Equivalent 28
Wnt-dependent reporter activity was conferred by a truncated fragment without the 29
last 3 sites, which indicates that the conserved sites are responsible for the luciferase 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

8
activity. While mutation of the site 2 or 3 binding sequence had no effect, deletion or 1
mutation of site 1 or 4 resulted in significantly compromised luciferase activity 2
induced with LiCl, albeit the decrease for the site 1 mutation was minor (Figure 3C). 3
Similar results were achieved in MDA-MB-231 breast cancer cells when stimulated 4
Wnt/β-catenin activity by recombinant Wnt1 (Figure 3D). In a chromatin 5
immunoprecipitation (ChIP) assay, both LEF1 and β-catenin were found to occupy 6
the endogenous Lin28 promoter at site 4 but not at site 1 in both MDA-MB-231 and 7
T-47D cells (Figure 3E), suggesting that only site 4 functions as a true endogenous 8
LEF1/β-catenin binding site on the Lin28 promoter. Taken together, our findings 9
establish that Wnt/β-catenin pathway activates Lin28 expression via direct binding to 10
the Lin28 promoter, which provides compelling evidence that Lin28 is a direct target 11
gene of Wnt/β-catenin pathway. 12
13
Activation of Lin28 is necessary for Wnt/β-catenin pathway-induced let-7 14
repression and cell proliferation 15
To examine whether Lin28 is necessary for Wnt/β-catenin pathway-mediated 16
repression of let-7 family members, we used 2 different shRNAs to inhibit Lin28 17
expression in MDA-MB-231 cells in the high Wnt state. Both of the shRNAs were 18
observed to knock down the Lin28 expression significantly to levels comparable to 19
those of negative control cells (Figure 4A, left). Knockdown of Lin28 completely 20
reversed the β-catenin-mediated repression of mature let-7a and let-7f, suggesting the 21
essential role of Lin28 in this process (Figure 4A, right). Conversely, overexpression of 22
Lin28 almost completely compromised the β -catenin knockdown-induced let-7a and 23
let-7f expression (Figure 4B). As HRAS and HMGA2 are known let-7 targets, we 24
examined the relationship between these proteins and Wnt/β-catenin pathway. 25
Multiple panel results indicated that Wnt/β-catenin pathway enhances HRAS and 26
HMGA2 protein expression and that this activity depends on Lin28 activation and 27
let-7 repression (Figure 4A-C). 28
Wnt/β-catenin pathway is known to enhance the proliferation of many cancer types 29
including in breast cancer (MacDonald et al., 2009; Matsuda et al., 2009). As 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

9
expected, enforced expression of β -catenin resulted in a significantly increased 1
growth rate in MDA-MB-231 cells. The simultaneous knockdown of Lin28 was found 2
to almost completely compromise the β-catenin activity (Figure 4D). When plated at a 3
clonal density, a significant decrease in the number and size of colonies was observed 4
for Lin28-depleted cells (Figure 4E). These results suggest that the transactivation of 5
Lin28 is essential for Wnt/β-catenin pathway-mediated let-7 inhibition and cell 6
proliferation. Moreover, overexpression of let-7a in Wnt-activated MDA-MB-231 7
cells was also observed to completely inhibit β -catenin-activated cell growth and 8
colony formation (Figure 4C-E), which underscores the importance of let-7 miRNAs 9
as Wnt/β-catenin pathway downstream targets in regulating cell proliferation. 10
Collectively, the above results verify the hypothesis that the repression of let-7 11
through the activation of Lin28 is necessary for Wnt/β-catenin pathway-induced cell 12
proliferation. 13
14
Let-7 repression through activation of Lin28 is necessary for Wnt/β-catenin 15
pathway-induced expansion of breast cancer stem cell populations 16
Although previous studies have demonstrated that Wnt/β-catenin pathway, Lin28 17
and let-7 miRNAs all play central roles in stem cell self-renewal and differentiation, 18
their possible involvement in the same signalling cascade that regulates cancer stem 19
or cancer-initiating cells has not been addressed. Such cells in breast cancer can be 20
enriched from established cancer cell lines using either mammosphere cultures or 21
FACS sorting for a CD44+ CD24- population (Al-Hajj et al., 2003; Ponti et al., 2005). 22
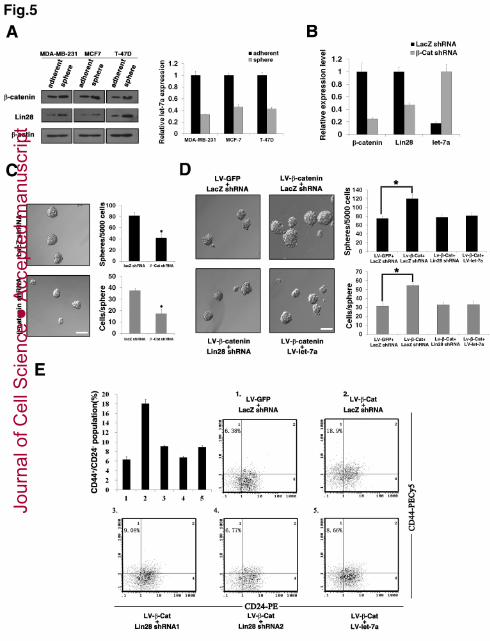
Using Western blotting and qPCR, we detected higher levels of β-catenin and Lin28 23
proteins but lower levels of let-7 members in mammospheres of the MDA-MB-231, 24
MCF7, and T-47D cell lines in comparison to their corresponding adherent cultures 25
(Figure 5A). Knockdown of β -catenin expression in MDA-MB-231 cells resulted in 26
decreased Lin28 expression and increased let-7a transcript levels in the 27
mammospheres (Figure 5B). Consequently, β -catenin-depleted cells showed a 28
reduced mammosphere number and size (Figure 5C). The expression trend of let-7 29
and Lin28 regulated by β-catenin in mammospheres implies a functional relevance of 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

10
these factors in breast cancer stem cells. Given our result that the Wnt/β-catenin 1
pathway/Lin28/let-7 cascade promotes breast cancer cell proliferation, we reasoned 2
that this phenomenon is caused by stem cell expansion. To verify this hypothesis, we 3
infected MDA-MB-231 cells with β-catenin-expressing lentiviruses and examined the 4
mammosphere formation. The enforced expression of β -catenin resulted in a 5
significantly increased mammosphere number and size in MDA-MB-231 cells. 6
Simultaneous knockdown of Lin28 or overexpression of let-7a was found to 7
compromise the β -catenin-enhanced mammosphere formation almost completely 8
(Figure 5D). We also examined whether the Wnt/β-catenin pathway/Lin28/let-7 9
cascade affects the size of the CD44+ CD24- population in breast cancer cells. T-47D 10
cells were used for this analysis as MDA-MB-231 cells contain too high a percentage 11
(�90%) of such cells, making it difficult to score any potential increase. As shown in 12
Figure 5E, the enforced overexpression of β -catenin in T-47D cells via lentiviral 13
infection yielded a greatly increased CD44+ CD24- population, but the overexpressed 14
β-catenin was no longer able to induce an increase in the CD44+ CD24- population in 15
the Lin28-depleted or let-7a-overexpressed cells. Collectively, these results 16
demonstrate that let-7 repression through the activation of Lin28 is necessary for 17
Wnt/β-catenin pathway-induced expansion of the breast cancer stem cell population. 18
19
In vivo analyses of the Wnt/β-catenin pathway/Lin28/let-7 cascade 20
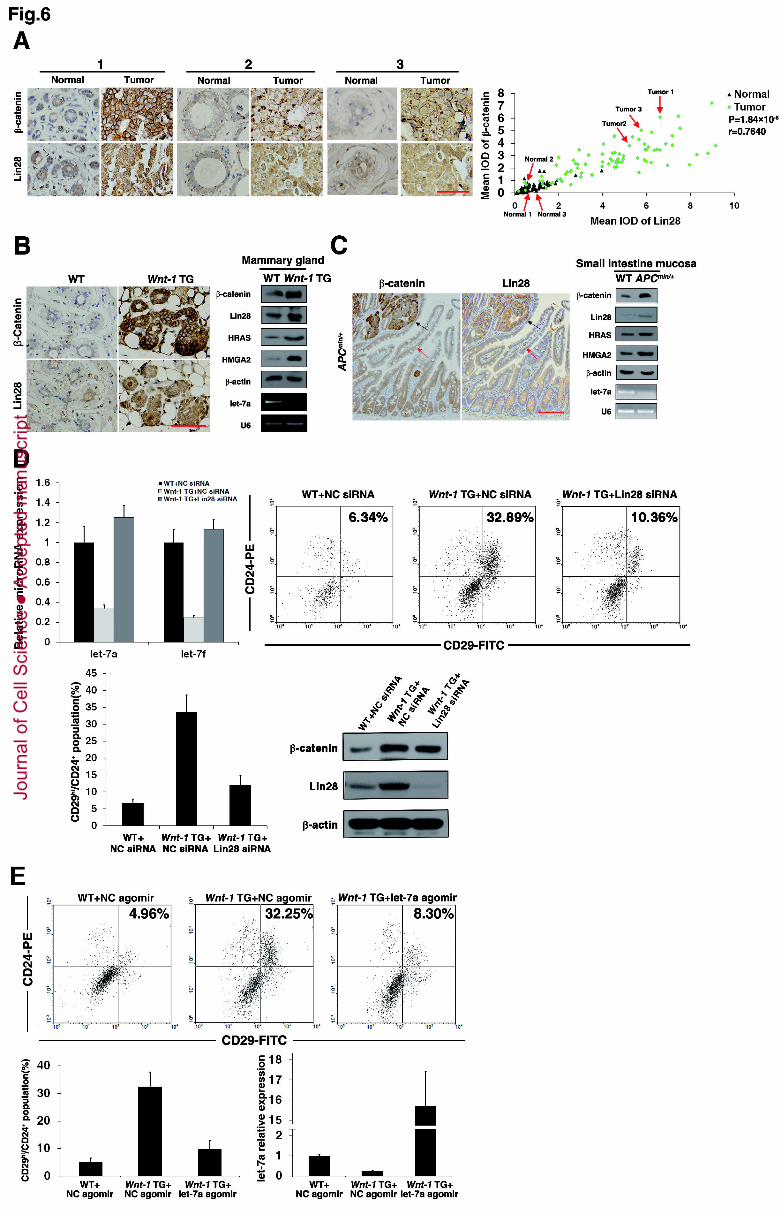
To investigate whether Wnt/β-catenin pathway is associated with Lin28 expression 21
in breast cancer patients, tissue microarrays from 82 patients with breast cancer who 22
had undergone mammary gland resection were examined via immunostaining with 23
β-catenin and Lin28 antibodies. The overall expression levels of both the β -catenin 24
and Lin28 proteins were significantly higher in breast cancers than in adjacent tissues 25
as indicated by the representative samples in the tissue microarrays (Figure 6A, left) 26
and the summary of all the samples (Table S2). Moreover, correlation analyses 27
revealed strong correlations between β-catenin expression and Lin28 levels (Table S2 28
and Figure 6A, right). To analyse further the relationship among Wnt/β-catenin 29
pathway, Lin28 and let-7, 16 pairs of fresh tumour samples along with their adjacent 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

11
tissues were used to detect the expression of β-catenin, Lin28 and let-7a by Western 1
blotting and qPCR. As shown in Figure S3, the β-catenin and Lin28 expression levels 2
in the cancers are higher than in the adjacent tissues, which is consistent with the 3
tissue microarray results. Conversely, the let-7a expression levels in the cancers are 4
dramatically lower than those of the adjacent tissues. Moreover, Lin28 expression 5
showed a positive relationship, whereas let-7a expression was found to be reversely 6
related to β-catenin. It has been well documented for the MMTV-wnt-1 mouse that 7
the stem cell-enriched Lin- CD29hi CD24+ population in premalignant mammary 8
tissue is greatly expanded (Shackleton et al., 2006), indicating the critical role for 9
Wnt/β-catenin pathway in the expansion of breast cancer stem cells. We therefore 10
measured the expression levels of β-catenin, Lin28 and let-7a in this model via 11
immunostaining, Western blotting and qPCR. As expected, β-catenin expression was 12
greatly increased in premalignant mammary tissues compared with tissues from 13
wild-type littermates. Lin28 and let-7a showed coordinating increased and decreased 14
expression levels. Moreover, the let-7 targets HRAS and HMGA2 also showed 15
increased protein expression levels in premalignant mammary tissues of the 16
MMTV-wnt-1 mouse (Figure 6B). To confirm the regulating cascade in vivo in a 17
second setting, we examined an additional model with a high Wnt state. The model 18
used, the Apc min/+ mouse, is a mouse intestine adenoma model bearing a truncated 19
Apc allele. Results similar to those of the MMTV-wnt-1 mice were achieved and 20
further confirm the relationship among Wnt/β-catenin pathway, Lin28 and let-7a 21
(Figure 6C). Finally, the in vivo functional relevance of Wnt/β-catenin 22
pathway/Lin28/let-7 cascade was examined. We delivered 23
cholesterol-,OMe-conjugated Lin28 siRNA into the premalignant mammary tissue of 24
MMTV-wnt-1 mice through an intraductal injection and examined whether Lin28 25
knockdown could rescue the let-7 repression and the stem cell phenotype driven by 26
Wnt/β-catenin pathway. As illustrated in Figure 6D, premalignant mammary 27
tissues exhibited decreased let-7a and let-7f levels and an expanded Lin- CD29hi 28
CD24+ population when compared with tissues from wild-type littermates; however, 29
knockdown of Lin28 dramatically recovered the let-7 expression and the size of the 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

12
stem cell population when compared with the wild-type littermates. Similarly, 1
delivery of a cholesterol-conjugated let-7a agomir into the premalignant mammary 2
tissue of MMTV-wnt-1 mice was also observed to dramatically rescue the stem cell 3
phenotype (Figure 6E). Taken together, these results provide solid evidence implying 4
the existence of a Wnt/β-catenin pathway/Lin28/let-7 cascade in vivo. 5
6
Discussion 7
Aberrant activation of the Wnt/β-catenin pathway has been found in a wide range 8
of cancers, especially in cancers derived from intestine, skin, mammary gland and 9
haematopoietic cells. Moreover, Wnt/β-catenin pathway may preferentially influence 10
stem/progenitor cell expansion in these cancers (Wend et al., 2010). Although many 11
downstream target genes that account for either normal development or tumorigenesis 12
have been identified in different cellular contexts, the genes that mediate 13
Wnt/β-catenin pathway activity in maintaining the stemness process are still not very 14
clear. c-Myc, a reprogramming factor and a well-known Wnt target, was recently 15
demonstrated to be an important stem cell regulator in normal and cancerous cells 16
(Kim et al., 2010; Smith et al., 2011). However, no study has established a convincing 17
relationship between Wnt/β-catenin pathway and c-Myc in stem cells. In contrast, 18
recent studies on both iPS and ESCs suggest that the role of Wnt/β-catenin pathway in 19
reprogramming and maintaining stem cells could be independent of c-Myc induction 20
(Marson et al., 2008; Ying et al., 2008). In breast cancer, current studies indicate that 21
Hedgehog, Notch, and Wnt/β-catenin pathway, as well as Bmi1 (Liu et al., 2006) and 22
Lin28 (Yang et al., 2010) transcription factors are major regulators that affect cancer 23
stem cell properties. However, the downstream molecular events of Wnt/β-catenin 24
pathway responsible for this stemness-related property remain largely unknown. In 25
this study, we identified Lin28 as a novel downstream target of Wnt/β-catenin 26
pathway and found that Lin28 is necessary for Wnt/β-catenin pathway-mediated 27
breast cancer stem cell expansion, which highlights a molecular mechanism for 28
Wnt/β-catenin pathway in regulating the stemness of cancer cells. More recently, it 29
was demonstrated that Wnt/β-catenin pathway acts as a new reprogramming factor to 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

13
promote somatic cells to pluripotency, although the underlying molecular rationale is 1
unclear (Ying et al., 2008). Thus, our studies have shed light on the connection 2
between Wnt/β-catenin pathway and the reprogramming process. 3
In regulating stem cell functions, miRNAs are proposed to be important factors 4
because expression levels of certain miRNAs in embryonic stem cells are different 5
from those of differentiated embryoid bodies (Suh et al., 2004). Consistently, tumours 6
analysed via miRNA profiling have shown significantly different miRNA profiles for 7
stem cells compared with differentiated cells from the same sample 8
(Papagiannakopoulos and Kosik, 2008; Sun et al., 2010). In breast cancer, the 9
inhibition of let-7, miR-30, and miR-200 and the induction of miR-181 and miR-495 10
are required for the maintenance of stem cell properties (Hwang-Verslues et al., 2011; 11
Iliopoulos et al., 2010; Shimono et al., 2009; Wang et al., 2011; Yu et al., 2010; Yu et 12
al., 2007), implying the importance of miRNAs in regulating breast cancer stem cells. 13
As Wnt/β-catenin pathway is a dominant regulator of breast cancer stem cells, we 14
speculate that this pathway may affect the stemness via modulations in miRNA 15
expression. The relationship between Wnt/β-catenin pathway and miRNAs has been 16
verified in different cellular contexts, although it remains obscure in stem cells, 17
especially cancer stem cells. Compared with rare downstream target miRNAs, many 18
upstream modulated miRNAs of Wnt/β-catenin pathway have been identified, of 19
which miR-135a/b (Nagel et al., 2008) and miR-315 (Silver et al., 2007) upregulate, 20
whereas miR-200a (Korpal et al., 2008; Saydam et al., 2009), miR-21 (Hashimi et al., 21
2009), miR-203 (Thatcher et al., 2008) and miR-8 (Kennell et al., 2008) downregulate 22
β-catenin transcriptional activity through targeting Wnt/β-catenin pathway 23
components. Among the few Wnt-regulated miRNAs, Wnt/β-catenin pathway 24
regulates miR-15/16 maturation, rather than its transcription. However, the underlying 25
mechanism is unknown (Martello et al., 2007). Another study showed that miR-122a 26
expression is downregulated in APC-driven gastrointestinal cancers. The mechanisms 27
by which Wnt/β-catenin pathway regulates miR-122a and 122a inhibition are 28
unknown (Wang et al., 2009). Additionally, miR-375 is downregulated by β-catenin. 29
However, the function of miR-375 and the transcriptional mechanism through which 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

14
miR-375 is regulated by Wnt/β-catenin pathway are not clear and are left for further 1
investigation (Ladeiro et al., 2008). In an attempt to identify novel downstream target 2
miRNAs of Wnt/β-catenin pathway, we performed screening experiments in breast 3
cancer cells. In our screening, 15 miRNAs were upregulated, while 48 miRNAs were 4
downregulated upon Wnt/β-catenin pathway stimulation, which is consistent with the 5
notion that an overall downregulation of miRNAs is present in many cancers 6
compared with their normal tissue counterparts (Hammond, 2006). It is noted that 2 7
known downregulated miRNAs, miR-15 and miR-375, are present in our list, 8
suggestive of a successful screening strategy for our experiment. The obvious 9
downregulation for let-7 members is remarkable, as let-7 miRNAs have been 10
demonstrated to play an important role in breast cancer stem cells. Both qPCR and 11
reporter assays in several cell lines verified that let-7 miRNAs are novel downstream 12
target miRNAs of Wnt/β-catenin pathway. Furthermore, we found that Wnt/β-catenin 13
pathway regulates let-7 miRNAs through a posttranscriptional pathway by directly 14
transactivating Lin28 expression. To the best of our knowledge, this report is the first 15
to find that Wnt/β-catenin pathway inhibits let-7 miRNAs via Lin28 transactivation. 16
Recent advances in the characterisation of stem cells in mammary epithelium and 17
breast cancer cells have opened the door for understanding the signalling and 18
transcriptional programming underlying both the development of normal mammary 19
stem cells and the proliferation/differentiation of their malignant counterparts 20
(Al-Hajj et al., 2003; Ponti et al., 2005; Shackleton et al., 2006; Stingl et al., 2006). 21
Wnt/β-catenin pathway, Lin28 and let-7 are defined as critical mammary/breast 22
cancer stem cell regulators. However, the relationship among them is unknown. In 23
this study, we have functionally established a connection between Wnt/β-catenin 24
pathway and Lin28/ let-7 that is essential for Wnt/β-catenin pathway-mediated breast 25
cancer stem cell expansion. This model is strongly supported by (i) the converging 26
effects of Wnt/β-catenin pathway, Lin28 and let-7 on breast cancer stem cell 27
properties, (ii) the dependence of Wnt/β-catenin pathway in let-7 inhibition and breast 28
cancer stem cell expansion on Lin28 activation, (iii) the dependence of Wnt/β-catenin 29
pathway in breast cancer stem cell expansion on let-7 inhibition, and (iv) the 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

15
expression pattern and functional relevance of β-catenin, Lin28 and let-7a in cancer 1
samples and tumour models. As such, our study presents the first mechanistic 2
characterisation of the Wnt/β-catenin pathway function in breast cancer stem cells and 3
points to Lin28/let-7 as important downstream targets in these cells in vitro and in 4
vivo. As Wnt/β-catenin pathway is aberrantly activated in many cancer types, future 5
work should address whether the Wnt/β-catenin pathway/Lin28/let-7 cascade is also 6
involved in the regulation of other cancers. Moreover, reexpression of let-7 miRNAs 7
may represent a new therapeutic option in treating cancers with aberrantly activated 8
Wnt/β-catenin pathway. 9
10
Materials and Methods 11
Cell culture. HEK 293T (human embryonic kidney 293T) and human breast 12
cancer cell lines ZR-75-30 (human breast ductal carcinoma), MDA-MB-231 (human 13
breast adenocarcinoma), MCF7 (human breast adenocarcinoma) and T-47D (human 14
breast ductal carcinoma) were obtained from the American Type Culture Collection 15
and grown in Dulbecco’s Modified Eagle’s medium (DMEM) supplemented with 16
10% foetal calf serum (Gibco), penicillin, and streptomycin. To induce Wnt/β-catenin 17
pathway, cells were grown in the presence of 20 mM LiCl (Sigma) for 24-36 h or 100 18
ng/ml Wnt1 protein(Abcam,Cat#ab84080)for 48h. All cell lines were grown at 37°C 19
with 5% carbon dioxide. 20
Reagents. The following antibodies were used:anti-Lin28(Abcam,Cat#ab46020), 21
anti-β-catenin(Cell Signaling Technology,Cat#9562), anti-β-actin(Sigma-Aldrich,Cat#A1978), 22
anti-HRAS(Proteintech Group,Cat#18295-1-AP), anti-HMGA2(R&D Systems,Cat#AF3184), 23
anti-LEF-1(Santa Cruz Biotechnology,Cat#sc-8591), anti-HA (Sigma-Aldrich, Cat#H9658). 24
The human cell surface markers utilised were CD24-PE(BD Biosciences) and 25
CD44-PE-Cy5(eBioscience). Cholesterol-conjugated mmu-let-7a agomir and negative 26
control agomir as well as cholesterol-,OMe-conjugated mouse lin28 siRNA and 27
negative control siRNA for in vivo delivery were obtained from Ribobio Co. 28
(Guangzhou, China). 29
Quantitative real-time PCR. Total RNAs were isolated using Trizol reagent 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

16
(Invitrogen). cDNAs were prepared from these RNAs using a ReverTra Ace qPCR 1
RT kit from Toyobo. Quantitative PCR was performed using a Rotor gene 6000 2
Sequence Detection System with the Thunderbird SYBR qPCR Mix (Toyobo). 3
Mature let-7a/let-7f/let-7g miRNAs were quantified using a predesigned miRCURY 4
LNATM Universal RT microRNA PCR Assay (Exiqon). A eukaryotic 18S rRNA or 5
U6 snRNA endogenous control was used as an internal standard, and the results were 6
calculated using the ΔΔCT (where CT is threshold cycle) method. Primer sequences 7
are provided in Table S3. 8
Cell proliferation. Stable transfected cells were plated in 96-well dishes at 1000 9
cells/well. Cell growth rates were monitored using a Cell Counting Kit-8 (CCK-8, 10
Dojindo Molecular Technologies) and an MTT Assay Kit (Promega). 11
Colony formation assays. Stable transfected cells were plated in 6-well dishes at 12
500 cells/well. After 10 days, the cultures were washed twice with PBS, incubated 13
with methanol for 20 min, stained with crystal violet for 30 min, and washed with tap 14
water. The colonies were counted under low magnification microscope and the cell 15
group that contains more than 10 cells is defined as a colony. 16
FACS analysis. Confluent cells were trypsinised into single-cell suspensions. 17
These culture cell suspensions and the tissue digested cell suspensions were washed 18
with fluorescence-activated cell sorting (FACS) buffer (2% foetal bovine serum in 19
PBS), counted, and stained with fluorophore-conjugated antibodies. A total of 106 20
cells in 100 µ l of FACS buffer were incubated with antibodies for 30 min at 4°C. 21
Unbound antibodies were washed off, and the cells were sorted using a Beckman 22
EPICS XL instrument. 23
Mammosphere culture. Cells (104 cells/ml) were cultured in ultra-low attachment 24
plates in serum-free DMEM/F12 (Invitrogen) supplemented with B-27 (1:50; 25
Invitrogen), 20 ng/ml epidermal growth factor (EGF; BD Biosciences), 20 ng/ml 26
basic fibroblast growth factor (bFGF; BD Biosciences), and 4 µg/ml insulin (Sigma) 27
and fed every 3 days. 28
miRNA microarrays. Total RNAs were harvested using TRIzol (Invitrogen) and 29
an RNeasy mini kit (QIAGEN) according to the manufacturer’s instructions. The 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

17
samples were labelled using a miRCURY™ Hy3™/Hy5™ Power labelling kit and 1
hybridised on a miRCURY™ LNA Array (v.14.0). Scanning was performed on an 2
Axon GenePix 4000B microarray scanner. GenePix pro V6.0 was used to read the 3
raw intensity of the images. Signals that were <2 times the background were 4
removed, and datasets were median-centred before calculating fold change values. 5
ChIP. A chromatin immunoprecipitation (ChIP) assay was performed following 6
the Upstate Biotechnology protocol. Briefly, T-47D cells were fixed with 1% 7
paraformaldehyde at room temperature for 10 min, washed, and lysed with SDS lysis 8
buffer (50 mM Tris-HCl, 1% SDS, 10 mM EDTA, and protease inhibitors). The 9
lysates were sonicated to reduce DNA lengths to between 300 and 600 bp. The 10
soluble fraction was diluted, precleared with salmon sperm DNA-protein A-agarose, 11
and divided into two tubes and incubated with specific antibodies or control IgG. The 12
immune complexes were then precipitated with protein A/G beads and eluted with 13
elution buffer (0.1 M NaHCO3 and 1% SDS). The eluted samples were reverse 14
cross-linked and treated with proteinase K. DNA was purified by phenol-chloroform 15
extraction and dissolved in distilled water. Real-time PCR quantification of the ChIP 16
samples was performed in triplicate using the Thunderbird SYBR qPCR Mix 17
(Toyobo), and primer sequences for the Lin28 promoter are provided in Table S3. 18
Generation of miRNA- and shRNA- or cDNA-expressing lentiviruses. 19
Oligonucleotides encoding let-7a1 pre-miRNA or shRNA targeting Lin28, β-catenin, 20
Pygo2 or LacZ (control) were synthesised by Invitrogen and cloned under the control 21
of the U6 promoter in the lentiviral vector lentilox pLL3.7. For overexpression of 22
Lin28, β -catenin S33Y, HA-Wnt1 or GFP (control), the cDNAs were cloned under 23
the control of the EF1α promoter in the lentiviral vector pLV-CS2.0. The generation 24
of lentivirus vectors was performed by co-transfecting pLL3.7 or pLV-CS2.0 carrying 25
the expression cassette with helper plasmids pVSV-G and pHR into 293T cells using 26
Lipofectamine 2000 (Invitrogen). The viral supernatant was collected 48 h after 27
transfection, and viral titers were determined by transducing HeLa cells at serial 28
dilutions and analysing the GFP expression using flow cytometry. Cells at 50 to 70% 29
confluence were infected with viral supernatants containing 10 µg/ml Polybrene for 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

18
24 h after which fresh medium was added to the infected cells, which were later 1
selected with puromycin or G418. The oligonucleotide sequences are provided in 2
Table S4. 3
Let-7 luciferase assay. To evaluate the miRNA function of let-7, a 4
pMIR-REPORTTM luciferase reporter vector (Ambion) with two copies of sequences 5
complementary to let-7a cloned into its 3’UTR (let-7 sensor) was used. The reporter 6
vector plasmid was transfected into MDA-MB-231 using Lipofectamine 2000 7
according to the manufacturer’s instructions. To correct for the transfection efficiency, 8
a luciferase reporter vector without let-7 target was transfected in parallel. The 9
luciferase activity was assayed using a luciferase assay kit (Promega). let-7 miRNA 10
function was expressed as a percent reduction in the luciferase activity of cells 11
transfected with the reporter vector containing the let-7 target sequences compared 12
with cells transfected with the vector without the let-7 target. 13
Lin28 promoter luciferase assay. To generate the luciferase reporter vectors, the 14
Lin28 promoter fragment was amplified from human genomic DNA and cloned into 15
the KpnI-Hind� site of the firefly luciferase plasmid pGL3-basic-IRES. For reporter 16
assays, HEK 293T cells in 24-well plates were transfected at 50 to 60% confluence 17
using a calcium-phosphate method. Both Lin28 promoter constructs (50 ng) and 18
cytomegalovirus (CMV)–β-galactosidase (25 ng) reporter plasmids were 19
co-transfected with the β -catenin S33Y or Wnt-1 expression plasmids or in the 20
presence of LiCl (10 to 20 mM) for 24 h. The total amount of plasmid DNA 21
transfected was made equivalent by adding empty vectors. Cells were harvested after 22
24 h and processed for luciferase and β -galactosidase assays, and the data were 23
normalised to the β-galactosidase levels. 24
In vivo stem cell assay. Fourteen-week-old virgin female MMTV-wnt-1 (FVB) 25
and wild-type (FVB) mice were anesthetized. The keratin plugs were removed from 26
the surface of the nipple, revealing the duct orifice. The mammary ducts were 27
cannulated using a 1.0-cm, 34-gauge, blunt-ended needle attached to a 1-mL 28
tuberculin syringe. Next, 5 nmol of cholesterol-conjugated microRNA agomir or 29
cholesterol-, OMe-conjugated siRNA in 25 µ l PBS was infused into the mammary 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

19
gland once every 3 days for a total of 4 times. Twenty days after the first injection, the 1
mammary glands of the injected site were dissected from the female mice. After 2
mechanical dissociation with surgical scissors, the tissue was placed in culture 3
medium 1640 supplemented with 5% bovine calf serum and containing 300 U/ml 4
collagenase (Sigma) and 100 U/ml hyaluronidase (Sigma) and digested for 3 h at 5
37°C. The resultant organoid suspension was vortexed and lysis of the red blood cells 6
in NH4Cl. A single-cell suspension was obtained by sequential dissociation of the 7
fragments by gentle pipetting for 1–2 min in 0.25% trypsin and then for 2 min in 5 8
mg/ml dispase II plus 0.1 mg/ml DNase I (Sigma-Aldrich) followed by filtration 9
through a 40-µm mesh. Cells were resuspended to 106 cells in 100 µl of cold FACS 10
buffer. Staining was performed using the following antibodies and reagents on ice for 11
30 min as follows: Biotin-conjugated Lin (CD45 , CD31 , TER119 , 50 12
µl/ml)(EasySep,Cat#19757), PE-conjugated CD24 (10 µ l/100 µl) (StemCell 13
Technologies,Cat#10814), FITC-conjugated CD29 (10 µl/100 14
µl)(BioLegend,Cat#102205), PE-conjugated IgG isotype control (10 µl/ 100 µl), 15
FITC-conjugated IgG isotype control (10 µl/ 100 µl). Unbound antibodies were 16
washed off, Propidium iodide (2 µg/ml final concentration) was added and incubated 17
on ice for 20 min. The cells were sorted using a Beckman EPICS XL instrument. 18
Forward scatter (FS) and side scatter (SS) were used to select most of the single cells 19
population. The PI-positive, non-viable cells were excluded from the final analysis. 20
Lin+ cells were gated out of the final analysis. Single stained samples were used as 21
compensation controls. Cells incubated in non-specific IgG isotype control 22
(conjugated to PE or FITC) were used to set the gates that define Lin-, CD24+ or 23
CD29hi cells respectively. 24
Histological analysis. Tissue microarrays from 82 patients with breast cancer were 25
purchased from Shanghai Outdo Biotech Co., Ltd. The tissues were fixed in 10% 26
formalin, embedded in paraffin, and sectioned. After dewaxing and rehydration, the 27
sections were pretreated with peroxidase blocking buffer (Maxim, Fuzhou, China) for 28
20 min at room temperature. Antigen retrieval was performed by boiling in 29
Tris-EDTA (pH 9.0) for 20 min. After treatment with a blocking buffer (5% normal 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

20
goat serum in PBS) for 1 h at room temperature, incubation with anti-Lin28 antibody 1
(1:300, overnight at 4°C, Abcam) and anti-β-catenin antibody (1:200, overnight at 2
4°C, Cell Signaling Technology) was performed in the blocking buffer. Secondary 3
antibody reagents were obtained from the SABC kit (Boster Biological Technology, 4
Wuhan, China) or DAB kit (Maxim Biological Technology, Fuzhou, China). 5
The pictures were taken by the use of a computerized image system (Leica 6
Microsystems, Imaging Solutions Ltd., Cambridge, United Kingdom). Under 7
high-power magnification (×200), photographs of three representative fields were 8
captured using Leica QWin Plus v3 software; identical settings were used for each 9
photograph. For the reading of each antibody staining, a uniform setting for all of the 10
slides was applied. The mean integrated optical density (IOD) was used to represent 11
the relative expression of indicated protein. IOD is a computer-assisted method which 12
is commonly used to quantify both the area and the intensity of the positive staining 13
of immunohistochemistry. We calculated the IOD of each photograph acquired from 14
the tissue microarray sections by using Image-Pro Plus v6.2 software (Media 15
Cybernetics Inc, Bethesda, MD). The typical positive staining area was located with 16
the help of a pathologist in the Segmentation panel; Standard Optical Density was 17
chosen in the Intense Calibration panel, and background was subtracted in the panel 18
of Optical Density Calibration, which is chosen to exclude all the areas without 19
epithelial or tumor tissues. We also subtract the background in a serial section by 20
using a non-specific IgG to instead of primary antibody. The “mean IOD” (IOD/tissue 21
area) represents the expression level of indicated protein. 22
Statistical analysis. For the tissue microarrays, Fisher’s exact test was used to 23
compare qualitative variables; quantitative variables were analysed using the t-test 24
and Spearman’s rank correlation test. The data were presented as the mean ± SD. All 25
statistical tests were two-sided Student's t-tests, and a P<0.05 was considered to be 26
statistically significant. The animal experiments were reviewed and approved by the 27
Institutional Animal Care and Use Committee of Xiamen University College of 28
Medicine. The human tissue preparation and analysis were approved by the 29
Institutional Review Board of The First Affiliated Hospital of Xiamen University. 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

21
1
Conflict of interest 2
The authors declare no conflict of interest. 3
4
Acknowledgements 5
This work was supported by grants from the "973" Project of the Ministry of 6
Science and Technology (2009CB52220, 2013CB530600), the National Natural 7
Science Foundation of China (U1205023,81272384,90919037,81201617,81201616),8
The 2013 major project of science and technology from Department of Education 9
(313051), the Natural Science Foundation of Fujian Province (13111125), the key 10
Projects of Fujian Province (2011Y01010460), the project of the Fujian Health 11
Department (2011-CXB-34), the project of the Xiamen Technique Bureau 12
(3502Z20114003), and "Project 111" sponsored by the State Bureau of Foreign 13
Experts and Ministry of Education (B06016). 14
15
16
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

22
References 1
Al-Hajj, M., Wicha, M. S., Benito-Hernandez, A., Morrison, S. J. and Clarke, M. F. (2003). 2 Prospective identification of tumorigenic breast cancer cells. Proc Natl Acad Sci U S A 100, 3983-8. 3 Amit, S., Hatzubai, A., Birman, Y., Andersen, J. S., Ben-Shushan, E., Mann, M., Ben-Neriah, 4 Y. and Alkalay, I. (2002). Axin-mediated CKI phosphorylation of beta-catenin at Ser 45: a molecular 5 switch for the Wnt pathway. Genes Dev 16, 1066-76. 6 Bartel, D. P. (2009). MicroRNAs: target recognition and regulatory functions. Cell 136, 215-33. 7 Calin, G. A., Sevignani, C., Dumitru, C. D., Hyslop, T., Noch, E., Yendamuri, S., Shimizu, 8 M., Rattan, S., Bullrich, F., Negrini, M. et al. (2004). Human microRNA genes are frequently 9 located at fragile sites and genomic regions involved in cancers. Proc Natl Acad Sci U S A 101, 10 2999-3004. 11 Chen, J., Luo, Q., Yuan, Y., Huang, X., Cai, W., Li, C., Wei, T., Zhang, L., Yang, M., Liu, Q. 12 et al. (2010). Pygo2 associates with MLL2 histone methyltransferase and GCN5 histone 13 acetyltransferase complexes to augment Wnt target gene expression and breast cancer stem-like cell 14 expansion. Mol Cell Biol 30, 5621-35. 15 Hammond, S. M. (2006). MicroRNAs as oncogenes. Curr Opin Genet Dev 16, 4-9. 16 Hashimi, S. T., Fulcher, J. A., Chang, M. H., Gov, L., Wang, S. and Lee, B. (2009). 17 MicroRNA profiling identifies miR-34a and miR-21 and their target genes JAG1 and WNT1 in the 18 coordinate regulation of dendritic cell differentiation. Blood 114, 404-14. 19 Heo, I., Joo, C., Cho, J., Ha, M., Han, J. and Kim, V. N. (2008). Lin28 mediates the terminal 20 uridylation of let-7 precursor MicroRNA. Mol Cell 32, 276-84. 21 Huang, K., Zhang, J. X., Han, L., You, Y. P., Jiang, T., Pu, P. Y. and Kang, C. S. (2010). 22 MicroRNA roles in beta-catenin pathway. Mol Cancer 9, 252. 23 Hwang-Verslues, W. W., Chang, P. H., Wei, P. C., Yang, C. Y., Huang, C. K., Kuo, W. H., 24 Shew, J. Y., Chang, K. J., Lee, E. Y. and Lee, W. H. (2011). miR-495 is upregulated by E12/E47 in 25 breast cancer stem cells, and promotes oncogenesis and hypoxia resistance via downregulation of 26 E-cadherin and REDD1. Oncogene 30, 2463-74. 27 Iliopoulos, D., Lindahl-Allen, M., Polytarchou, C., Hirsch, H. A., Tsichlis, P. N. and Struhl, 28 K. (2010). Loss of miR-200 inhibition of Suz12 leads to polycomb-mediated repression required for 29 the formation and maintenance of cancer stem cells. Mol Cell 39, 761-72. 30 Johnson, S. M., Grosshans, H., Shingara, J., Byrom, M., Jarvis, R., Cheng, A., Labourier, E., 31 Reinert, K. L., Brown, D. and Slack, F. J. (2005). RAS is regulated by the let-7 microRNA family. 32 Cell 120, 635-47. 33 Kennell, J. A., Gerin, I., MacDougald, O. A. and Cadigan, K. M. (2008). The microRNA 34 miR-8 is a conserved negative regulator of Wnt signaling. Proc Natl Acad Sci U S A 105, 15417-22. 35 Kim, J., Woo, A. J., Chu, J., Snow, J. W., Fujiwara, Y., Kim, C. G., Cantor, A. B. and Orkin, 36 S. H. (2010). A Myc network accounts for similarities between embryonic stem and cancer cell 37 transcription programs. Cell 143, 313-24. 38 Korpal, M., Lee, E. S., Hu, G. and Kang, Y. (2008). The miR-200 family inhibits 39 epithelial-mesenchymal transition and cancer cell migration by direct targeting of E-cadherin 40 transcriptional repressors ZEB1 and ZEB2. J Biol Chem 283, 14910-4. 41 Krizhanovsky, V. and Lowe, S. W. (2009). Stem cells: The promises and perils of p53. Nature 42 460, 1085-6. 43 Kumar, M. S., Erkeland, S. J., Pester, R. E., Chen, C. Y., Ebert, M. S., Sharp, P. A. and 44
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

23
Jacks, T. (2008). Suppression of non-small cell lung tumor development by the let-7 microRNA 1 family. Proc Natl Acad Sci U S A 105, 3903-8. 2 Ladeiro, Y., Couchy, G., Balabaud, C., Bioulac-Sage, P., Pelletier, L., Rebouissou, S. and 3 Zucman-Rossi, J. (2008). MicroRNA profiling in hepatocellular tumors is associated with clinical 4 features and oncogene/tumor suppressor gene mutations. Hepatology 47, 1955-63. 5 Liu, C., Li, Y., Semenov, M., Han, C., Baeg, G. H., Tan, Y., Zhang, Z., Lin, X. and He, X. 6 (2002). Control of beta-catenin phosphorylation/degradation by a dual-kinase mechanism. Cell 108, 7 837-47. 8 Liu, J., Sato, C., Cerletti, M. and Wagers, A. (2010). Notch signaling in the regulation of stem 9 cell self-renewal and differentiation. Curr Top Dev Biol 92, 367-409. 10 Liu, S., Dontu, G., Mantle, I. D., Patel, S., Ahn, N. S., Jackson, K. W., Suri, P. and Wicha, M. 11 S. (2006). Hedgehog signaling and Bmi-1 regulate self-renewal of normal and malignant human 12 mammary stem cells. Cancer Res 66, 6063-71. 13 Logan, C. Y. and Nusse, R. (2004). The Wnt signaling pathway in development and disease. 14 Annu Rev Cell Dev Biol 20, 781-810. 15 Lu, J., Getz, G., Miska, E. A., Alvarez-Saavedra, E., Lamb, J., Peck, D., Sweet-Cordero, A., 16 Ebert, B. L., Mak, R. H., Ferrando, A. A. et al. (2005). MicroRNA expression profiles classify 17 human cancers. Nature 435, 834-8. 18 MacDonald, B. T., Tamai, K. and He, X. (2009). Wnt/beta-catenin signaling: components, 19 mechanisms, and diseases. Dev Cell 17, 9-26. 20 Marson, A., Foreman, R., Chevalier, B., Bilodeau, S., Kahn, M., Young, R. A. and Jaenisch, 21 R. (2008). Wnt signaling promotes reprogramming of somatic cells to pluripotency. Cell Stem Cell 3, 22 132-5. 23 Martello, G., Zacchigna, L., Inui, M., Montagner, M., Adorno, M., Mamidi, A., Morsut, L., 24 Soligo, S., Tran, U., Dupont, S. et al. (2007). MicroRNA control of Nodal signalling. Nature 449, 25 183-8. 26 Matsuda, Y., Schlange, T., Oakeley, E. J., Boulay, A. and Hynes, N. E. (2009). WNT signaling 27 enhances breast cancer cell motility and blockade of the WNT pathway by sFRP1 suppresses 28 MDA-MB-231 xenograft growth. Breast Cancer Res 11, R32. 29 Merchant, A. A. and Matsui, W. (2010). Targeting Hedgehog--a cancer stem cell pathway. Clin 30 Cancer Res 16, 3130-40. 31 Moss, E. G. and Tang, L. (2003). Conservation of the heterochronic regulator Lin-28, its 32 developmental expression and microRNA complementary sites. Dev Biol 258, 432-42. 33 Nagel, R., le Sage, C., Diosdado, B., van der Waal, M., Oude Vrielink, J. A., Bolijn, A., 34 Meijer, G. A. and Agami, R. (2008). Regulation of the adenomatous polyposis coli gene by the 35 miR-135 family in colorectal cancer. Cancer Res 68, 5795-802. 36 Newman, M. A., Thomson, J. M. and Hammond, S. M. (2008). Lin-28 interaction with the 37 Let-7 precursor loop mediates regulated microRNA processing. Rna 14, 1539-49. 38 Papagiannakopoulos, T. and Kosik, K. S. (2008). MicroRNAs: regulators of oncogenesis and 39 stemness. BMC Med 6, 15. 40 Piskounova, E., Viswanathan, S. R., Janas, M., LaPierre, R. J., Daley, G. Q., Sliz, P. and 41 Gregory, R. I. (2008). Determinants of microRNA processing inhibition by the developmentally 42 regulated RNA-binding protein Lin28. J Biol Chem 283, 21310-4. 43 Ponti, D., Costa, A., Zaffaroni, N., Pratesi, G., Petrangolini, G., Coradini, D., Pilotti, S., 44
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

24
Pierotti, M. A. and Daidone, M. G. (2005). Isolation and in vitro propagation of tumorigenic breast 1 cancer cells with stem/progenitor cell properties. Cancer Res 65, 5506-11. 2 Roarty, K. and Rosen, J. M. (2010). Wnt and mammary stem cells: hormones cannot fly 3 wingless. Curr Opin Pharmacol 10, 643-9. 4 Rybak, A., Fuchs, H., Smirnova, L., Brandt, C., Pohl, E. E., Nitsch, R. and Wulczyn, F. G. 5 (2008). A feedback loop comprising lin-28 and let-7 controls pre-let-7 maturation during neural 6 stem-cell commitment. Nat Cell Biol 10, 987-93. 7 Saydam, O., Shen, Y., Wurdinger, T., Senol, O., Boke, E., James, M. F., Tannous, B. A., 8 Stemmer-Rachamimov, A. O., Yi, M., Stephens, R. M. et al. (2009). Downregulated 9 microRNA-200a in meningiomas promotes tumor growth by reducing E-cadherin and activating the 10 Wnt/beta-catenin signaling pathway. Mol Cell Biol 29, 5923-40. 11 Shackleton, M., Vaillant, F., Simpson, K. J., Stingl, J., Smyth, G. K., Asselin-Labat, M. L., 12 Wu, L., Lindeman, G. J. and Visvader, J. E. (2006). Generation of a functional mammary gland 13 from a single stem cell. Nature 439, 84-8. 14 Shimono, Y., Zabala, M., Cho, R. W., Lobo, N., Dalerba, P., Qian, D., Diehn, M., Liu, H., 15 Panula, S. P., Chiao, E. et al. (2009). Downregulation of miRNA-200c links breast cancer stem cells 16 with normal stem cells. Cell 138, 592-603. 17 Silver, S. J., Hagen, J. W., Okamura, K., Perrimon, N. and Lai, E. C. (2007). Functional 18 screening identifies miR-315 as a potent activator of Wingless signaling. Proc Natl Acad Sci U S A 104, 19 18151-6. 20 Smith, K. N., Lim, J. M., Wells, L. and Dalton, S. (2011). Myc orchestrates a regulatory 21 network required for the establishment and maintenance of pluripotency. Cell Cycle 10, 592-7. 22 Stingl, J., Eirew, P., Ricketson, I., Shackleton, M., Vaillant, F., Choi, D., Li, H. I. and Eaves, 23 C. J. (2006). Purification and unique properties of mammary epithelial stem cells. Nature 439, 993-7. 24 Suh, M. R., Lee, Y., Kim, J. Y., Kim, S. K., Moon, S. H., Lee, J. Y., Cha, K. Y., Chung, H. 25 M., Yoon, H. S., Moon, S. Y. et al. (2004). Human embryonic stem cells express a unique set of 26 microRNAs. Dev Biol 270, 488-98. 27 Sun, J. G., Liao, R. X., Qiu, J., Jin, J. Y., Wang, X. X., Duan, Y. Z., Chen, F. L., Hao, P., Xie, 28 Q. C., Wang, Z. X. et al. (2010). Microarray-based analysis of microRNA expression in breast cancer 29 stem cells. J Exp Clin Cancer Res 29, 174. 30 Takebe, N., Harris, P. J., Warren, R. Q. and Ivy, S. P. (2011). Targeting cancer stem cells by 31 inhibiting Wnt, Notch, and Hedgehog pathways. Nat Rev Clin Oncol 8, 97-106. 32 Thatcher, E. J., Paydar, I., Anderson, K. K. and Patton, J. G. (2008). Regulation of zebrafish 33 fin regeneration by microRNAs. Proc Natl Acad Sci U S A 105, 18384-9. 34 Ventura, A. and Jacks, T. (2009). MicroRNAs and cancer: short RNAs go a long way. Cell 136, 35 586-91. 36 Viswanathan, S. R. and Daley, G. Q. (2010). Lin28: A microRNA regulator with a macro role. 37 Cell 140, 445-9. 38 Wang, X., Lam, E. K., Zhang, J., Jin, H. and Sung, J. J. (2009). MicroRNA-122a functions as 39 a novel tumor suppressor downstream of adenomatous polyposis coli in gastrointestinal cancers. 40 Biochem Biophys Res Commun 387, 376-80. 41 Wang, Y., Yu, Y., Tsuyada, A., Ren, X., Wu, X., Stubblefield, K., Rankin-Gee, E. K. and 42 Wang, S. E. (2011). Transforming growth factor-beta regulates the sphere-initiating stem cell-like 43 feature in breast cancer through miRNA-181 and ATM. Oncogene 30, 1470-80. 44
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

25
Wend, P., Holland, J. D., Ziebold, U. and Birchmeier, W. (2010). Wnt signaling in stem and 1 cancer stem cells. Semin Cell Dev Biol 21, 855-63. 2 Yang, X., Lin, X., Zhong, X., Kaur, S., Li, N., Liang, S., Lassus, H., Wang, L., Katsaros, D., 3 Montone, K. et al. (2010). Double-negative feedback loop between reprogramming factor LIN28 and 4 microRNA let-7 regulates aldehyde dehydrogenase 1-positive cancer stem cells. Cancer Res 70, 5 9463-72. 6 Ying, Q. L., Wray, J., Nichols, J., Batlle-Morera, L., Doble, B., Woodgett, J., Cohen, P. and 7 Smith, A. (2008). The ground state of embryonic stem cell self-renewal. Nature 453, 519-23. 8 Yu, F., Deng, H., Yao, H., Liu, Q., Su, F. and Song, E. (2010). Mir-30 reduction maintains 9 self-renewal and inhibits apoptosis in breast tumor-initiating cells. Oncogene 29, 4194-204. 10 Yu, F., Yao, H., Zhu, P., Zhang, X., Pan, Q., Gong, C., Huang, Y., Hu, X., Su, F., Lieberman, 11 J. et al. (2007). let-7 regulates self renewal and tumorigenicity of breast cancer cells. Cell 131, 12 1109-23. 13 14
15
Figure legends 16
Figure 1. Posttranscriptional repression of let-7 family miRNAs by 17
Wnt/β-catenin pathway. 18
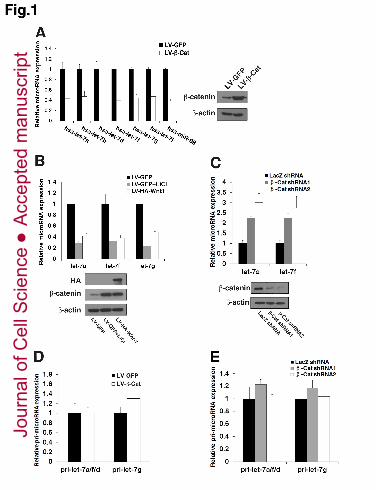
(A) Relative mature microRNA (miRNA) levels of individual let-7 family members 19
in ZR-75-30 cells stably expressing LV-GFP (control) and LV-β-Cat (left) using 20
qPCR analysis. Western blotting results showing β-catenin overexpression levels 21
(right). (B) Relative levels of mature let-7a, let-7f and let-7g miRNAs in control and 22
LiCl- (36 h) or LV-HA-Wnt1-treated ZR-75-30 cells using qPCR analysis (top). 23
Western blotting showing the indicated proteins expression (bottom). (C) qPCR 24
analysis of the mature let-7a, let-7f miRNA levels in ZR-75-30 cells stably expressing 25
LacZ shRNA (control) and 2 different β -catenin (β-Cat) shRNAs (top). Western 26
blotting indicating the β-catenin knockdown efficiency (bottom). (D) qPCR analysis 27
of the let-7a/let-7f/let-7d and let-7g pri-miRNA levels in ZR-75-30 cells stably 28
expressing LV-GFP and LV-β-Cat. (E) qPCR analysis of the let-7a/let-7f/let-7d and 29
let-7g pri-miRNA levels in ZR-75-30 cells stably expressing LacZ shRNA and 2 30
different β -Cat shRNAs. (A-E) Data are the mean ± SD of three independent 31
experiments. 32
33
Figure 2. Wnt/β-catenin pathway activates Lin28 expression in breast cancer 34
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

26
cells. 1
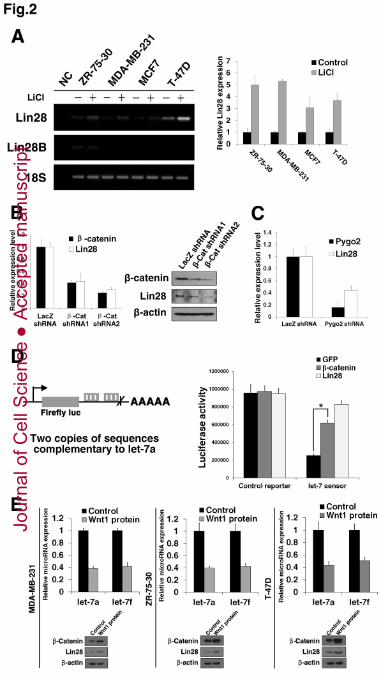
(A) Expression levels of Lin28 and Lin28B mRNA upon Wnt/β-catenin pathway 2
stimulation in breast cancer cell lines. ZR-75-30, MDA-MB-231, MCF7 and T-47D 3
cell lines were treated with or without LiCl for 36 h. A no-cDNA template group was 4
used as a negative control (NC), whereas 18S rRNA was used as an internal control. 5
Semi-quantitative RT-PCR analysis was used to measure the Lin28 and Lin28B levels 6
(left). The expression levels of Lin28 mRNA were confirmed using qPCR (right). (B) 7
Relative expression of Lin28 mRNA (left) and protein (right) measured using qPCR 8
and Western blotting in ZR-75-30 cells stably expressing LacZ shRNA (control) and 9
2 different β -Cat shRNAs. (C) Relative expression of Lin28 mRNA measured by 10
qPCR in ZR-75-30 cells stably expressing LacZ shRNA and Pygo2 shRNA. (D) 11
Luciferase sensor assay. Schematic representation of the let-7 sensor, which contained 12
a constitutively expressed firefly luciferase reporter bearing two copies of sequences 13
complementary to let-7a in the downstream 3’UTR (left). Luciferase activity of the 14
let-7 sensor in β-catenin , Lin28, or GFP-transfected MDA-MB-231 cells (right). The 15
vehicle vector pMIR reporter was used as a control reporter. (E) MDA-MB-231, 16
ZR-75-30 and T-47D cell lines were treated with or without Wnt1 protein (100ng/ml) 17
for 48 h. qPCR analysis was used to measure the mature let-7a and let-7f levels (top); 18
Western blotting was used to measure β -catenin and Lin28 levels (bottom). (A-E) 19
Data are the mean ± SD of three independent experiments (*, P<0.001). 20
21
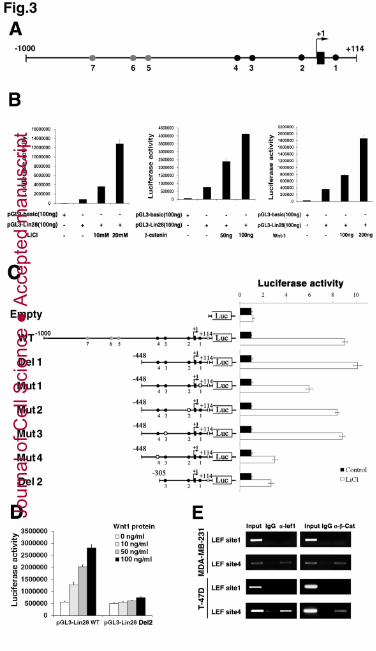
Figure 3. Lin28 is a novel direct downstream target gene of Wnt/β-catenin 22
pathway. 23
(A) Schematic representation of the genomic region near the transcription start site of 24
Lin28. The dots represent the putative LEF/TCF binding sites CTTTG or GAAAC; 25
those in black are conserved between human and mouse. (B) The luciferase activity of 26
the pGL3-Lin28 vector containing the putative LEF/TCF binding sites was activated 27
in a dose-dependent manner upon LiCl (left), β-catenin (middle) or Wnt-1(right) 28
stimulation in 293T cells. The empty pGL3-basic vector was used as a negative 29
control. (C) Luciferase activity of various pGL3-Lin28 vectors (wild type, deleted or 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

27
mutated in LEF/TCF sites) in untreated and LiCl-treated (24 h) 293T cells. (D) 1
Luciferase activity of pGL3-Lin28 vectors (wild type or Del2) in Wnt1 protein-treated 2
(0, 10, 50, 100ng/ml) MDA-MB-231 cells for 48h. (E) Chromatin 3
immunoprecipitation (ChIP) assay of LEF1 or β -catenin for the Lin28 promoter in 4
MDA-MB-231 (top) and T-47D (bottom) cells using semi-quantitative RT-PCR 5
analyses. (B-D) Data are the mean ± SD of three independent experiments. 6
7
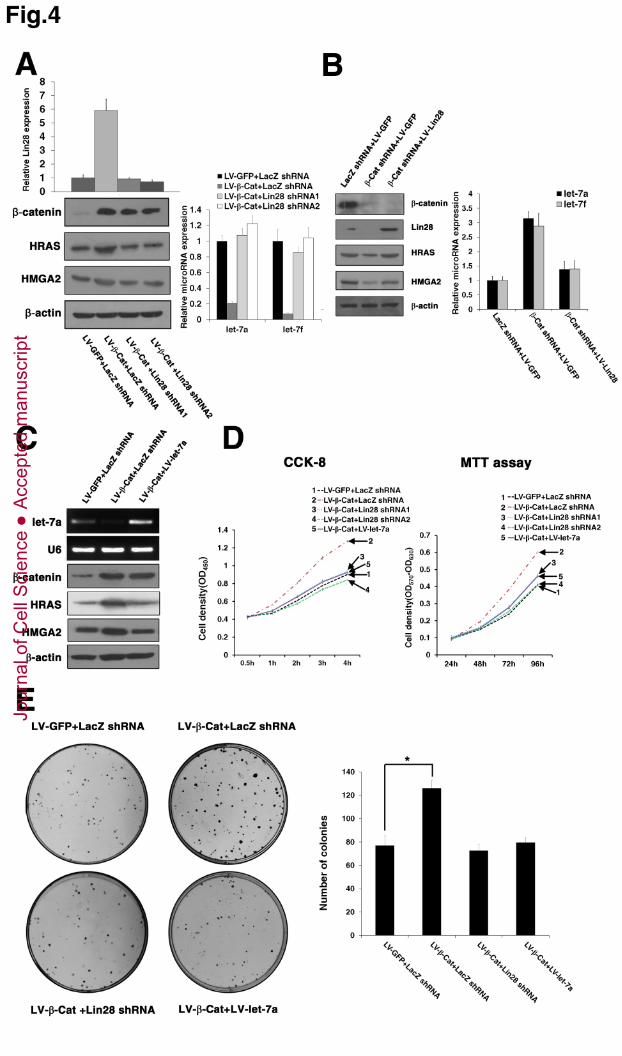
Figure 4. Activation of Lin28 is necessary for Wnt/β-catenin pathway-induced 8
Let-7 repression and cell proliferation 9
(A) Knockdown of Lin28 completely reversed the β -catenin-mediated repression of 10
mature let-7a and let-7f. MDA-MB-231 cells were infected with β-catenin lentiviruses 11
(LV-β-Cat) and treated with 2 different Lin28 shRNAs simultaneously. qPCR was 12
used to measure Lin28 levels (left, top); Western blotting was used to measure 13
β-catenin, HRAS and HMGA2 levels (left, bottom); semi-quantitative RT-PCR was 14
used to measure let-7a/let-7f abundance (right). (B) Overexpression of Lin28 15
completely compromised β-catenin knockdown-induced let-7a and let-7f expression. 16
MDA-MB-231 cells were infected with β -Cat shRNA and treated by Lin28 17
lentiviruses (LV-Lin28) simultaneously. Western blotting was used to measure 18
β-catenin, Lin28, HRAS and HMGA2 protein levels (left); qPCR was used to 19
measure let-7a/let-7f abundance (right). (C) Upregulation of HRAS and HMGA2 by 20
β-catenin is dependent on let-7 suppression. MDA-MB-231 cells were infected with 21
β-catenin lentiviruses (LV-β-Cat) and treated by let-7a lentiviruses (LV-let-7a) 22
simultaneously. Semi-quantitative RT-PCR was used to measure let-7a abundance 23
(top); Western blotting was used to measure β -catenin, HRAS and HMGA2 levels 24
(bottom). (D and E) Knockdown of Lin28 or enforced expression of let-7a 25
compromised β-catenin-induced cell proliferation. MDA-MB-231 cells from A and C 26
were used. Cell growth rates (D) and colony formations (E) were measured. (E) A 27
representative experiment of three independent experiments is shown. Photographs of 28
colonies were taken at day 10 (left), and the number of colonies was quantified (right). 29
(A-E) Data are the mean ± SD of three independent experiments (*, P<0.001). 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

28
1
Figure 5. Let-7 repression through activation of Lin28 is necessary for 2
Wnt/β-catenin pathway-induced expansion of breast cancer stem cell 3
populations. 4
(A) β-catenin and Lin28 expressions are enriched, whereas let-7 expression is reduced 5
in breast cancer stem cells. Shown are the Western blotting results for β-catenin and 6
Lin28 protein levels (left) and the qPCR analysis of let-7a transcripts (right) in 7
mammospheres or corresponding adherent cells of different breast cancer cell lines. 8
(B) Knockdown of β -catenin expression resulted in decreased Lin28 expression and 9
increased let-7a transcript levels in mammospheres. MDA-MB-231 cells stably 10
expressing β-Cat shRNA were cultured in mammospheres for 14 days, and qRT-PCR 11
analysis measured the expression levels of β -catenin, Lin28 and let-7a. (C) 12
β-catenin-depleted cells showed a reduced mammosphere number and size. 13
Representative photographs of mammospheres were taken at day 14 (left); the sphere 14
number and size were quantified (right). (D) Knockdown of Lin28 or overexpression 15
of let-7a compromised the β -catenin-enhanced mammosphere formation. 16
MDA-MB-231 cells were infected with LV-β-Cat and treated by Lin28 shRNA or 17
LV-let-7a simultaneously, and the cells were cultured in mammospheres for 12 days. 18
Representative photographs of mammospheres were taken at day 12 (left); the sphere 19
number and size were quantified (right). (E) Knockdown of Lin28 or overexpression 20
of let-7a compromised β-catenin-enhanced CD44+ CD24- population. MDA-MB-231 21
cells were infected with corresponding lentiviruses, and the percentage of 22
CD44+CD24- cells was determined using FACS analysis. Representative FACS 23
profiles from a single pair are shown, and the diagram indicates the mean values for 24
three different pairs. (C and D) A representative experiment of four independent 25
experiments. Data are the mean ± SD of four independent experiments (*, P<0.001). 26
(A-B and E) Data are the mean ± SD of three independent experiments. Bars, 100 µm. 27
28
Figure 6. In vivo analyses of the Wnt/β-catenin pathway/Lin28/let-7 cascade. 29
(A) Breast cancer samples derived from 82 patients in a tissue microarray along with 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

29
their adjacent normal tissues were immunostained with β -catenin and Lin28 1
antibodies, respectively. Left, Representative immunostaining for β-catenin and Lin28 2
is shown for three patient samples. Right, Correlations between β-catenin and Lin28 3
protein levels in tumour and normal tissues were measured using the Spearman rank 4
correlation test (r=0.7640, P=1.84×10-8). The three Representative samples were 5
marked in the correlation map. Bars, 50 µm. (B) Expression levels of β-catenin, Lin28 6
and let-7a in MMTV-Wnt-1 transgenic mouse tissue (Wnt-1 TG). 7
Immunohistochemical analysis of β -catenin and Lin28 expression in premalignant 8
mammary tissues derived from a Wnt-1 TG mouse and mammary tissues from its 9
wild-type (WT) littermate, at an age of 14 weeks (left). Bars, 50 µm. Western blotting 10
and semi-quantitative RT-PCR analysis of β -catenin/Lin28/HRAS/HMGA2/let-7a 11
levels in the same mammary tissues (right). (C) Immunohistochemical analysis of 12
β-catenin and Lin28 expression in small intestine tissues derived from an Apcmin/+ 13
mouse at the age of 16 weeks. Black arrows indicate adenoma tissues; red arrows 14
indicate villus tissues (left). Bars, 200 µm. Western blotting and RT-PCR analysis of 15
β-catenin/Lin28/HRAS/HMGA2/let-7a levels in small intestine mucosa (free of 16
muscle) derived from an Apcmin/+ mouse and its wild-type (WT) littermate, at an age 17
of 16 weeks (right). (D) Knockdown of Lin28 compromised the let-7 repression and 18
the stem cell expansion of premalignant mammary tissues from the Wnt-1 transgenic 19
(TG) mice. WT and Wnt-1 TG mice (n=3) at 14 weeks of age were administered 20
cholesterol-,OMe-conjugated negative control (NC) or Lin28 siRNA through 21
intraductal injection. The indicated let-7 expression was detected by qPCR(top, left) 22
and the percentage of Lin- CD29hi CD24+ cells in cell suspensions of mammary 23
tissues was determined by FACS analysis(top, right). Shown are representative FACS 24
profiles from a single pair, and the diagram indicates the mean values of three 25
different pairs (bottom, left). The Western blotting indicates a successive knockdown 26
of Lin28 in Wnt-1 TG mammary cells (bottom, right). (E) Enforced expression of 27
let-7a rescued the stem cell population of premalignant mammary tissues from the 28
Wnt-1 TG mice. WT and Wnt-1 TG mice (n=3) at 14 weeks of age were administered 29
cholesterol-conjugated negative control (NC) or let-7a agomir through intraductal 30
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

30
injection, and the percentage of Lin- CD29hi CD24+ cells in cell suspensions of 1
mammary tissues was determined by FACS analysis. Shown are representative FACS 2
profiles from a single pair (top), and the diagram indicates the mean values of three 3
different pairs (bottom, left). The qPCR results indicate a successive intake of let-7a 4
in Wnt-1 TG mammary cells (bottom, right). (B-E) A representative experiment of 5
three independent experiments. (D and E) Data are the mean ± SD of three 6
independent experiments. 7
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t





























