Embed Size (px)
Citation preview

Chiang Mai J. Sci. 2014; 41(3) 491
Chiang Mai J. Sci. 2014; 41(3) : 491-502http://epg.science.cmu.ac.th/ejournal/Contributed Paper
Xylitol Production by Thermotolerant MethylotrophicYeast Ogataea siamensis and Its Xylose ReductaseGene (XYL1) CloningWanatchabhorn Boontham [a], Nantana Srisuk*[a,b], Kanya Kokaew [a],Pantida Treeyoung [a], Savitree Limtong [a,b], Arinthip Thamchaipenet [b,c] andHiroya Yurimoto [d][a] Department of Microbiology, Faculty of Science, Kasetsart University, Bangkok 10900, Thailand.[b] Center for Advanced Studies in Tropical Natural Resources, NRU-KU, Kasetsart University,
Chatuchak, Bangkok 10900, Thailand.[c] Department of Genetics, Faculty of Science, Kasetsart University, Bangkok 10900, Thailand.[d] Division of Applied Life Sciences, Graduate School of Agriculture, Kyoto University,
Kitashirakawa-Oiwake, Sakyo-ku, Kyoto 606-8502, Japan.*Author for correspondence; e-mail: [email protected]
Received: 9 May 2013Accepted: 8 January 2014
ABSTRACTThermotolerant methylotrophic yeast, Ogataea siamensis N22, possesses high
yield of xylitol from D-xylose at high temperature. Xylitol production was comparedat 37oC and 40oC in a 2.5L fermenter with 1.5% (v/v) methanol and 100 g/L D-xylosesupplementation. The maximum yield of xylitol (Yp/s) and volumetric productivity (Qp)were 0.83 g/g and 0.36 g/L/h at 37oC, respectively; while at 40oC the values appeared tobe 0.87 g/g and 0.34 g/L/h, respectively. The yeast strain was subjected to characterizationfor XYL1 gene coding for xylose reductase which catalyzes an initial step of xylitolproduction from D-xylose. XYL1 gene was cloned from O. siamensis N22 genomicDNA and subsequently sequenced. As a result, an open reading frame of 960 bp encoding319 amino acids was obtained with a predicted protein molecular mass of 36.3 kD.Alignment of deduced amino acids sequence indicated that xylose reductase of O. siamensisN22 was closely related to aldose reductase of Candida boidinii with 72% similarity. O.siamensis N22 recombinant carrying an extra copy of XYL1 integrated into thechromosome showed slightly higher xylose reductase activity and hence xylitiolproduction than that of the original host strain.
Keywords: Ogataea siamensis, xylitol production, xylose reductase, gene cloning
1. INTRODUCTIONXylitol is a five-carbon sugar alcohol
naturally found in many fruits and vegetableswith minor quantity. Despite of the samesweetness level as sucrose, xylitol possessesa reduced calorie sweetening power as well
as non- and anti-cariogenic properties [1].Another beneficial health property of xylitolhas been found fascinating to all diabetics asinsulin is not required for its metabolism [2].Broad applications of xylitol have long been

492 Chiang Mai J. Sci. 2014; 41(3)
reported in pharmaceuticals and foodindustries. These include the production ofchewing gum as a result of its coolingeffect and negative heat of solution [3]. It isalso commonly used in production ofsugar free oral tablets and oral hygieneproducts. Alternatively, xylitol has beenapplied as an alternative preservative forcosmetic products [4]. It was claimed thatxylitol shared an increasing market value upto $340 million with 43,000 tons of globalxylitol consumption in 2005 [5].
Industrial production of xylitol isachieved by a nickel-catalyzed hydrogenationof D-xylose which is costly and energyconsumed [1]. These disadvantages areovercome by a more environmental friendlybiological process using xylose-assimilatingmicroorganisms. Microbial strains able topotentially assimilate xylose to xylitol havebeen screened and used as a tool to fulfillthis approach. Among microbes able toassimilate xylose to xylitol, yeast appears tobe an organism of choice that xylitolproduction of various genus and species hasbeen investigated such as Debaryomyces hansenii[5], Candida tropicalis [6-7], C. guilliermondii [8],Pichia guilliermondii [9] and methylotrophicyeasts, extensively focused on C. boidinii [10].Yeast strains those keen on xylitol productionsucceeded to employ xylose reductase, thefirst enzyme in xylose metabolic pathway,to utilize xylose as their substrate for growth.
Xylose reductase, XR, (EC. 1.1.1.21) isresponsible for reduction of D-xyloseto xylitol with concomitant NAD(P)Hoxidation. XR is important in fermentingxylose to xylitol. Yeast XRs were categorizedinto the aldose reductase (ALR) family,a member of the aldo-ketoreductase(AKR) superfamily, based on sequence andstructural similarities [11]. The enzymes werealso classified into two groups accordingto their coenzyme specificity [12]. Generally,
XR is specific for NADPH, as cofactor such asthat from Candida intermedia, C. guilliermondiiand Debaryomyces hansenii UFV-17 [13-15]. However, both NADH and NADPHare usable for yeasts Pichia stipitis, C. tenuisand C. shehatae [16-18].
Several yeast XR genes have been cloned,characterized and expressed includingKluyveromyces lactis [21], C. tropicalis [22],C. guiliermondii [23], C. parapsilosis [20],C. boidinii [19], Pachysolen tannophilus [24]and Pichia stipitis [25] using different methods.However, up to date, neither xylitolproduction nor xylose reductase enzymefrom Ogataea siamensis has not yet beenreported.
It has been well agreed that usingthermotolerant microorganisms wouldreduce the cooling cost during industrialfermentation process especially in tropicalcountries. We therefore report here xylitolproduction of O. siamensis N22 at hightemperatures i.e. 37°C and 40°C. Inaddition, we cloned and characterized O.siamensis N22 XYL1 gene encoding xylosereductase, the first enzyme catalyzing thereduction of D-xylose to xylitol in xyloseassimilating pathway.
2. MATERIALS AND METHODS2.1 Yeast Strain and Xylitol Production
Thermotolerant yeast Ogataea siamensisN22 was maintained in YPD medium(1% yeast extract, 2% peptone and 2%dextrose) with or without 1.5% agardepending on purpose of cultivation.For xylitol production, yeast culture wasgrown in basal medium [10] supplementedwith 1.5% (v/v) methanol, 10g/L xylose,5 g/L casamino acid and 0.5 g/L MgSO4.7H2O at pH 7.0 in a 2.5 L bioreactor(Biostat B, Braun Biotech International,Germany) with aeration rate 1.75 vvmand agitation speed 150 rpm.

Chiang Mai J. Sci. 2014; 41(3) 493
2.2 Growth, Xylose and XylitolDetermination
Yeast growth was assessed as cell dryweight. Xylose and xylitol were analyzedby HPLC (Hewlett Packard1100, Germany)with RI detector and a sugar column(ULTRON PS-80, Shinwa ChemicalIndustries, Kyoto, Japan).
2.3 XYL1 Gene Cloning and SequencingThe genomic DNA of O. siamensis N22
was prepared [26] and partially digestedby Bsp143I (Fermentas, USA). The DNAlibrary was constructed using pJET1.2/blunting vector (Fermentas, USA) andintroduced into E. coli XL-1 Blue. Recom-binants were grown on LB agar containing100 μg/mL ampicillin and screened by PCRamplification using primers OSXRf55(5’- TTYGGCTGYTGGAAAGTCG -3’)and OSXRr57(5’- YTTKSYTGGGTCAAGTATGGRTGRTG -3’) specifically designedfor XR. Polymerase chain reaction (PCR)was performed in a 25 μL reaction mixturecontaining 5ng genomic DNA template(2 μL), 2.5 μL 10x PCR Buffer (25 mMMgCl2), 0.6 μL dNTP mixture (2.5 mmol/L), 2 μL primer mixture (1pmol/L), 0.13 μLTaq DNA polymerase (5 U/L; Fermentas,USA) and 15.8 μL distilled water. The PCRconditions were as follow: 3cycles of 1 minat 94.0°C, 1 min at 64.0°C, and 1 min at72.0°C, preceeded by 5 min at 94.0°C andfollowed by 1min at 72.0°C. The positiveclones were sequenced (First Base Inc.,Malaysia) and analyzed.
2.4 Multiple Alignment and PhylogeneticAnalysis of Protein Sequence
Nucleotide and amino acid sequencessimilarity as well as sequences identitywere determined using BLAST programs[27]. Amino acid sequence alignment wasperformed using ClustalW2 program [28].
A phylogenetic tree was generated from thealignments using the neighbor-joiningmethod [29] and bootstrap analysis wassampled 1,000 times.
2.5 Characterization of XYL1 PutativeProtein
The putative protein of XYL1 wasdetermined using ORF Finder programhttp://www.ncbi.nlm.nih.gov/projects /gorf). The predicted protein molecularmass was achieved using Compute pI/Mw(http://web.expasy.org/compute_pi).Various yeast XR amino acid sequencesand the deduced amino acid sequence ofOgataea siamensis N22 XR were alignedand the putative active site was determinedusing ScanProsite program (http://au.expasy.org/tools/scanprosite/) .Nucleotide sequence of O. siamensis N22XYL1 coding region was deposited inGenBank, accession no. FJ763639.
2.6 Construction and Analysis of Ogataeasiamensis N22 Overexpressing XYL1
The O. siamensis N22 XYL1 codingregion was amplified from genomic DNAby PCR using specific primers OSXRf-78 (5’-GCTTCGAATATTGTGTGGGGAATTTCTTG-3’ with the underlined SfuI site for otherpurpose) and OSXRr95 (5- CGGAATTCGGCCTTTATTTACTGTTTCTAG -3 withthe underlined EcoRI site) under thesame PCR condition described above.PCR product was purified and inserted atEcoRI site of pGAPZ B vector (Invitrogen,USA), an integrative vector containingZeocin resistant marker and a constitutivepromoter, PGAP. The resulting plasmid,pGAPXR, was then transformed intosorbitol treated competent yeast cellby Bio-Rad gene pulser (Bio-Rad, USA) at25 μF and 2.5 KV. Using similar technique,the PCR product was purified and inserted

494 Chiang Mai J. Sci. 2014; 41(3)
into pPICZ B vector (Invitrogen, USA),an integrative vector containing Zeocinresistant marker and a regulated promoter,PAOX. The resulting plasmid pPICXR wastransformed into competent O. siamensisN22 cells using the same procedure thatyielded pGAPXR. Transformants wereselected on YPD agar supplementedwith 5μg/mL Zeocin™ (Invitrogen, USA).O. siamensis N22 recombinants containing theconstruct integrated into the chromosomewere investigated for their XR activity incomparison to the original host strain bycultivation in 50 mL of YPX medium (1%yeast extract, 2% peptone and 2% xylose) on170 rpm rotary shaker at 37°C for one day.Cells were harvested and resuspendedin extraction buffer prior to cell breakage.
2.7 Xylose Reductase Activity AssayCell crude extracts were prepared using
glass beads and mini-bead beater (Biospec,UK). XR activity in cell crude extracts wasdetermined spectrophotometrically [20].NADH was used, instead of NADPH, todetermine cofactor preference of O. siamensisN22 XR activity. Protein concentration incell extracts was determined by Bradfordsmini assay [20]. All assay experiments wereperformed in triplicates.
2.8 Xylitol Production by YeastTransformants
Ogataea siamensis N22 and trans-formants were cultivated in 50 mL of basalmedium [10] supplemented with 1.5%(v/v) methanol, 100 g/L xylose, 5 g/Lcasamino acid and 0.5 g/L MgSO4. 7H2Oat pH 7.0 in a 250 mL flask with shakingspeed 150 rpm for 14 days. Samples weretaken for determination of growth everyday whereas xylose and xylitol concentra-tion were assessed every two days.
3. RESULTS AND DISCUSSION3.1 Thermotolerant Ogataea siamensisN22
Ogataea siamensis (formerly Pichia siamensis)is a methylotrophic yeast isolated from treeexudates in Thailand and was taxonomicallyvalidated earlier [31-32]. O. siamensis N22 isclaimed as a thermotolerant yeast strainaccording to higher specific growth rateappeared at 30°C when compared to thevalue observed at 40°C. Moreover, growthon YPD agar at 8°C has been observed(data not shown). This is in consistent toprevious report on characteristics ofthermotolerant yeast [33].
3.2 Xylitol Production in 2.5L FermenterThermotolerant methylotrophic yeast
Ogataea siamensis N22 was grown in 2.5Lbioreactor under aerobic batch cultivationand xylitol production at 37°C and 40°C(Figure 1). Xylose was completely used upwithin 10-11 days of cultivation in accordancewith observed stationary phase growth.The maximum yeast growth at 40°C wasslightly lower (14.4 g/L) than that observedat 37°C (15 g/L). However, comparativemaximum xylitol concentration of 82.8 g/Land 80.8 g/L were observed after 11 daysof cultivation at 37°C and 40°C respectively.The theoretical xylitol production yieldfrom xylose (Yp/s) as well as volumetricproductivity (Qp) were calculated. Resultsexhibited the maximum yield of xylitolproduction and volumetric productivityof 0.83 g/g and 0.36 g/L/h at 37°Cwhile the values appeared to be 0.87 g/gand 0.34 g/L/h respectively at 40°C(Table 1). Interestingly, the higher maximumyield was observed within 8 days ofcultivation at 40°C, whereas the maximumyield at 37°C was not observed untilthe cultivation reached 11 days.
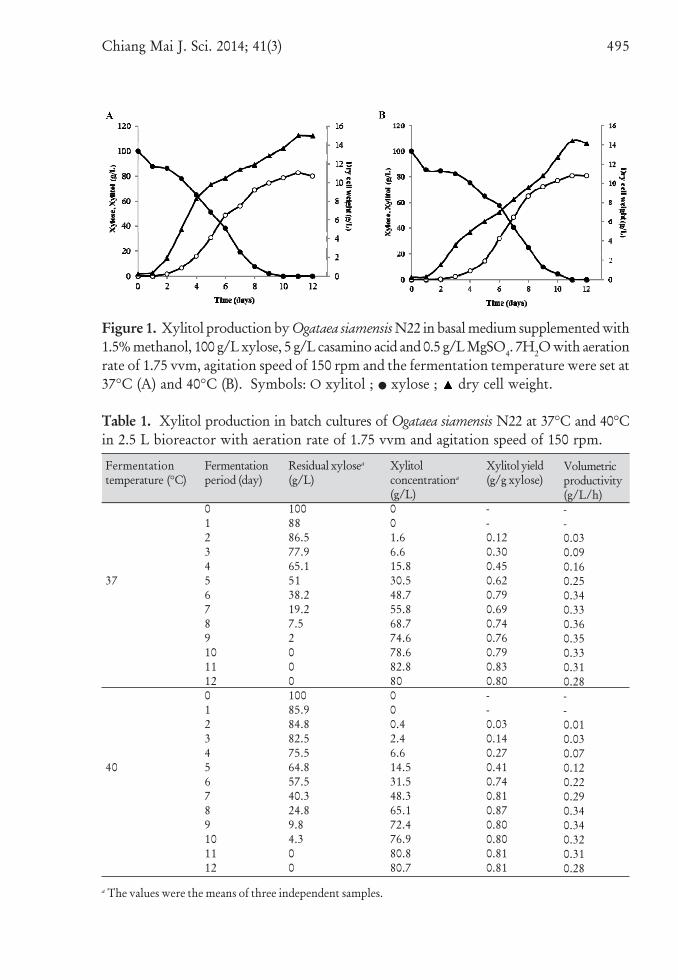
Chiang Mai J. Sci. 2014; 41(3) 495
Figure 1. Xylitol production by Ogataea siamensis N22 in basal medium supplemented with1.5% methanol, 100 g/L xylose, 5 g/L casamino acid and 0.5 g/L MgSO4. 7H2O with aerationrate of 1.75 vvm, agitation speed of 150 rpm and the fermentation temperature were set at37°C (A) and 40°C (B). Symbols: O xylitol ; • xylose ; dry cell weight.
Table 1. Xylitol production in batch cultures of Ogataea siamensis N22 at 37°C and 40°Cin 2.5 L bioreactor with aeration rate of 1.75 vvm and agitation speed of 150 rpm.
a The values were the means of three independent samples.
Fermentationtemperature (°C)
37
40
Fermentationperiod (day)
01234567891011120123456789101112
Residual xylosea
(g/L)
1008886.577.965.15138.219.27.5200010085.984.882.575.564.857.540.324.89.84.300
Xylitolconcentrationa
(g/L)001.66.615.830.548.755.868.774.678.682.880000.42.46.614.531.548.365.172.476.980.880.7
Xylitol yield(g/g xylose)
--0.120.300.450.620.790.690.740.760.790.830.80--0.030.140.270.410.740.810.870.800.800.810.81
Volumetricproductivity(g/L/h)--0.030.090.160.250.340.330.360.350.330.310.28--0.010.030.070.120.220.290.340.340.320.310.28

496 Chiang Mai J. Sci. 2014; 41(3)
Results appeared that, even at hightemperature as 40°C, O. siamensis N22could produce higher xylitol concentrationthan that has been reported in othermethylotrophic yeasts. After 5 days ofcultivation, C. boidinii No. 2201 grown inxylose (10 g/L) medium supplemented with2% (v/v) methanol showed only 48.5 g/Lxylitol at 28°C [34] whereas, under the samexylose content and 1.5% (v/v) methanolsupplementation, O. siamensis N22produced 55.8 g/L and 65.1 g/L xylitolafter 7 and 8 days of cultivation at 37°Cand 40°C, respectively. Hansenulapolymorpha DL1 was shown to give highyield of xylitol as 57 g/L xylitol frommedium plus 110 g/L D-xylose and 1%(v/v) methanol after 3 days of 30°Ccultivation [10]. Although these twomethylotrophic yeasts seem to producexylitol at earlier fermentation time thanO. siamensis N22, it should be, however,noted that we presented here the xylitolproduction at higher temperature i.e. 37oCand 40oC. The other thermotolerant yeast,Debaryomyces hansenii, showed the maximumxylitol produced by free cells as 68.6 g/Lxylitol at 40°C from 100 g/L of xylose, witha yield of 0.76 g/g after 7 days of cultivation[9]. O. siamensis N22, although at longerperiod of incubation, obviously showedhigher xylitol yield than that reported fromD. hansenii. This therefore confirms thatO. siamensis N22 has the ability to producexylitol from D-xylose at high temperature.
Xylitol production in methylotrophicyeast was significantly increased whensupplemented with methanol. This had beenproposed to be due to the oxidation ofmethanol that supplies NADH to enhancethe reduction of D-xylose [34]. However, thisexplanation may not be applied to ourO. siamensis N22 and its transformant sincewe eventually found that NADPH was
more preferable than NADH. The highestxylitol production yield of O. siamensisN22 observed at 37°C (0.83 g xylitol/gxylose consumed) appeared to be comparableto the highest value reported in Candidasp. [10] and higher than those previouslyreported [35].
3.3 Cloning and Sequencing of XYL1Gene from Ogataea siamensis N22
Partially Bsp143I digested genomicDNA of O. siamensis N22 was purified andselected for the DNA fragment size about1-3 kb for cloning into pJET1.2/bluntvector. The recombinant plasmids harboringXR gene (XYL1) were then screened byPCR using specific primers, OSXRf55 andOSXRr570. Nucleotide sequence analysis ofthe positive clone showed a full-length960 bp of XYL1 gene encoding a putative319 amino acids as well as the potentialpromoter element containing a TATA-likesequence (-112TATAA-108) upstream fromstart codon and a polyadenylation signal(974AATAAA979) downstream from stopcodon (Figure 2). This consistently appearedin the reports on the existence of TATA-likesequence and polyadenylation signalappeared in yeast genes responsible forxylose metabolism [21-22].
Amino acids sequence of O. siamensisN22 XR was examined and the active siteconsists 269IPKSNQKERLLQNLSV284
indicating the signature of yeast XR active site,269IPKS272 , was identified (Figurre 2) [22, 24].Multiple alignment of the deduced amino acidsequences of O. siamensis N22 XR and otheryeasts revealed the conserved amino sequenceof aldo/ketoreductase family putative activesite signature i.e. IPKS (Figure 3). Molecularmass of O. siamensis N22 XR was predictedfrom deduced amino acids sequence as36.3 kD. This prediction on XR molecularmass well agrees with that reported for

Chiang Mai J. Sci. 2014; 41(3) 497
Figure 2. Nucleotide and deduced amino acid sequences of XR gene (XYL1) of Ogataeasiamensis N22. An asterisk (*) indicates stop codon. The deduced TATAA promotersite is underlined (thick line). The AATAA putative polyadenylation signal is alsounderlined (thin line). Amino acid sequence in the gray box show aldo/ketoreductasefamily putative active site signature.
Candida tropicalis XR [22]. Phylogeneticanalysis of XR homologs suggested that O.siamensis N22 XR is closely related toaldose reductase of Candida boidinii (Figure4). Both results show close relationshipbetween XR of O. siamensis N22 and twomembers of the genus Candida. However,
it should be noted that, O. siamensis N22was formerly classified as a member of thegenus Pichia [31-32]. More details onenzyme properties may be needed to ruleout the concrete relation of O. siamensisN22 XR and other yeast XRs.

498 Chiang Mai J. Sci. 2014; 41(3)
Figure 3. Amino acid sequences alignment of 12 yeast xylose reductases and the deducedamino acid sequence of Ogataea siamensis N22 xylose reductase. A putative active site signaturewas shown as underlined.
Figure 4. Phylogenetic tree of Ogataea siamensis N22 xylose reductase and otherhomologous xylose reductase reported in yeasts. A phylogenetic tree was constructedby Neighbor joining algorithm and the aligned regions after excluding sequences creatinggaps. Boot strap values are shown on the branches and expressed in percentages.
3.4 Xylose Reductase Activity andXylitol Production of Ogataea siamensisN22 Recombinants
A copy of O. siamensis N22 XYL1 genewas expected to integrate into its chromosomeusing pGAPZ B vector. Three recombinantsshowed 0.5 fold higher XR specific activity(90-93 mU/mg protein) than that of theoriginal host (60 mU/mg protein) while thestrain habouring naked pGAPZ B showedequal level (61 mU/mg protein). We alsoconstructed O. siamensis N22 transformants
which was expected to possess extra copiesof XYL1 gene into yeast genome via anotherintegrative vector carries regulated promoterPAOX. However, two recombinants carryingextra copies of XYL1 gene into their genomevia pPICZ B vector also showed only0.5 fold increased in XR specific activity.This unpleasant outcome may be due toinappropriate promoters used in this studyi.e. PGAP and PAOX in pGAPZ B and pPICZ B,respectively. Expression of the XYL1 geneon its own promoter may be a successive
Chiang Mai J. Sci. 2014; 41(3) 499
task to solve this obstruct. Noticeably thatXR activity of the original host appeared tobe closed to that of C. boidinii (78 mU/mgprotein) but only half of that of C. tenuis(150 mU/mg protein) [36]. Review on yeastXR activity with NADPH as cofactor hasbeen shown to be various between 110 to6430 mU/mg protein [37]. Investigation ofcofactor preference revealed that NADPHwas more favorable to O. siamensis N22xylose reductase reaction than NADH.
We studied on optimal temperaturefor XR activity of O. siamensis N22 andits transformants at 30°C, 37°C and 40°C(data not shown). Results revealed thatO. siamensis N22 and the two transformantscarrying pPICZ B:: XYL1 or pGAPZ
B:: XYL1 in their genome showed similarresults i.e increased XR activity as thetemperature increased up to 40°C.
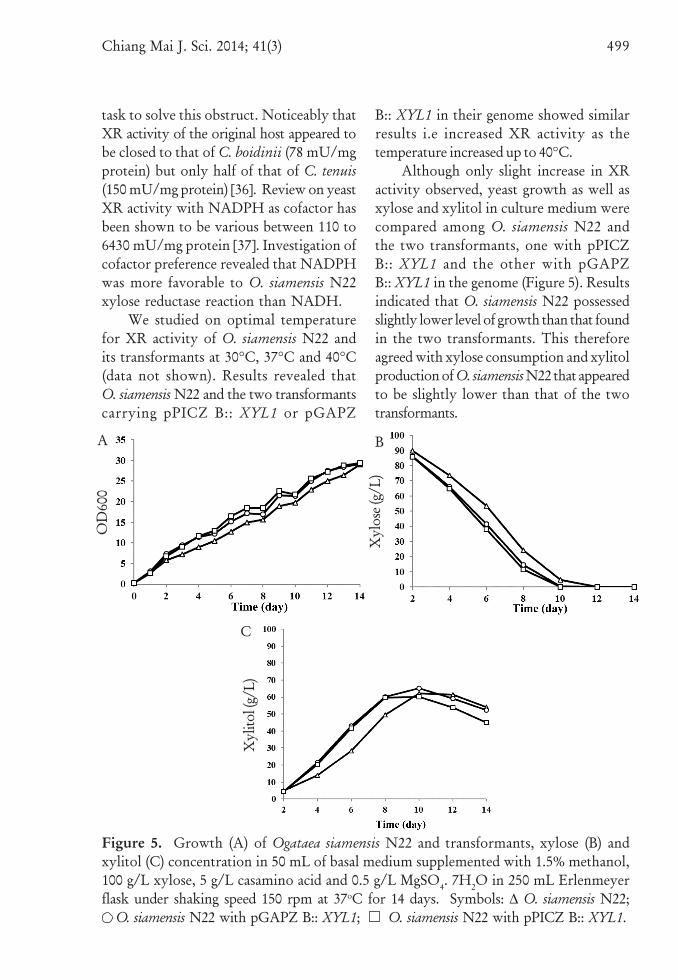
Although only slight increase in XRactivity observed, yeast growth as well asxylose and xylitol in culture medium werecompared among O. siamensis N22 andthe two transformants, one with pPICZB:: XYL1 and the other with pGAPZB:: XYL1 in the genome (Figure 5). Resultsindicated that O. siamensis N22 possessedslightly lower level of growth than that foundin the two transformants. This thereforeagreed with xylose consumption and xylitolproduction of O. siamensis N22 that appearedto be slightly lower than that of the twotransformants.
Figure 5. Growth (A) of Ogataea siamensis N22 and transformants, xylose (B) andxylitol (C) concentration in 50 mL of basal medium supplemented with 1.5% methanol,100 g/L xylose, 5 g/L casamino acid and 0.5 g/L MgSO4. 7H2O in 250 mL Erlenmeyerflask under shaking speed 150 rpm at 37oC for 14 days. Symbols: ∆ O. siamensis N22;
O. siamensis N22 with pGAPZ B:: XYL1; O. siamensis N22 with pPICZ B:: XYL1.
A B
C
OD
600
Xyl
ose (
g/L)
Xyl
itol (
g/L)

500 Chiang Mai J. Sci. 2014; 41(3)
4. CONCLUSIONSWe describe here a report on xylitol
production at high temperature ofOgataea siamensis N22, thermotolerantmethylotrophic yeast isolated in Thailand.Xylitol production by O. siamensis N22at 40oC appears to be promising since highxylitol production yield was obtained. Theprocess optimization is considered tofurther obtain the highest yield as well asthe shortest time of fermentation. We alsopublish here the nucleotide sequence of O.siamensis XYL1 and attempt to overproducexylose reductase enzyme in O. siamensis N22.Although high level of XR overproductionwas not successively obtained, experimentalscheme could be drawn for further fruitfulresearch goal.
ACKNOWLEDGMENTSThis work was partially supported by
Kasetsart University Research andDevelopment Institute to Nantana Srisuk.The authors would also like to thankDr. Savitr Trakulnaleumsai for her valuablecomments and suggestions to this work.
REFERENCES
[1] Prakasham R.S., Sreenivas R.R. andHobbs P.J., Current trends inbiotechnology production of xylitol andfuture prospects, Curr. Trends Biotechnol.Pharm., 2009; 3: 8-36.
[2] Emodi A., Xylitol: Its properties andfood applications, Food Technol., 1978;32: 20-32.
[3] Pepper T. and Olinger P.M., Xylitolin sugar-free confections, FoodTechnol., 1988; 10: 98-106.
[4] Amaral L.F.B., Camilo N.S., PeredaM.D.C.V., Levy C.E., Moriel P.and Mazzola P.G., Evaluation ofantimicrobial effectiveness of C-8
xylitol monoester as an alternativepreservative for cosmetic products,Int. J. Cosmt. Sci., 2011; 33: 391-397.
[5] Prakash G., Varma A.J., Prabhune A.,Shouche Y. and Rao M., Microbialproduction of xylitol from D-xylose andsugarcane bagasse hemicelluloses usingnewly isolated thermotolerant yeastDebaryomyces hansenii, Bioresour.Technol., 2011; 102: 3304-3308.
[6] Wang L., Yang M., Fan X., Zhu X., XuT. and Yuan Q., An environmentallyfriendly and efficient method for xylitolbioconversion with high-temperature-steaming corncob hydrolysate byadapted Candida tropicalis, ProcessBiochem., 2011; 46: 1619-1626.
[7] Misra S., Gupta P., Raghuwanshi S.,Dutt K. and Saxena R.K., Comparativestudy on different strategies involved forxylitol purification from culture mediafermented by Candida tropicalis, Sep.Purif. Technol., 2011; 78: 266-273.
[8] Sarrouh B.F. and Silva S.S., Applicationof response surface methodology foroptimization of xylitol productionfrom lignocellulosic hydrolysate in afluidized bed reactor, Chem. Eng.Technol., 2010; 33: 1481-1487.
[9] Zou Y.Z., Qi K., Chen X., Miao X.L.and Zhong J.J., Favorable effect ofvery low initial KLa value on xylitolproduction from xylose by a self-isolated strain of Pichia guilliermondii,J. Biosci. Bioeng., 2010; 109: 149-152.
[10] Suryadi H., Katsuragi T., Yoshida N.,Suzuki S., and Tani Y., Polyol productionby culture of methanol-utilizing yeast,J. Biosci. Bioeng., 2000; 89: 236-240.
[11] Jez J.M. and Penning T.M., The aldo-ketoreductase (AKR) superfamily anupdate, Chem. Biol. Interact., 2001;130-132: 499-525.

Chiang Mai J. Sci. 2014; 41(3) 501
[12] Mishra P. and Singh A., Microbialpentose utilization, Adv. Appl.Microbiol., 1993; 39: 91-152.
[13] Sugai J.K. and Delgenes J.P., Cataboliterepression of induction of aldosereductase activity and utilization ofmixed hemicellulosic sugars in Candidaguilliermondii, Curr. Microbiol., 1995;31: 239-244.
[14] Mayr P., Bruggler K., Kulbe K.D. andNidetzky B., D-xylose metabolism byCandida intermedia isolation andcharacterisation of two forms of aldosereductase with different coenzymespecificities, J. Chromatogr. B, 2000;737: 195-202.
[15] Sampaio F.C., Faria J.T., PassosF.M.L., Converti A. and Minin L.A.,Optimal activity and thermostabilityof xylose reductase from Debaryomyceshansenii UFV-170, J. Ind. Microbiol.Biotechnol., 2009; 36: 293-300.
[16] Verduyn C., Kleef R.V., Frank J.,Schreuder H. and Dijken J.P.V.,Properties of the NAD(P)H-dependentxylose reductase from the xylosefermenting yeast Pichia stipitis,Biochem. J., 1985; 226: 669-667.
[17] Neuhauser W., Haltrich D., KulbeK.D. and Nidetzky B., NAD(P)H-dependent aldose reductase from thexylose-assimilating yeast Candidatenuis, J. Biochem., 1997; 326: 683-692.
[18] Wang X., Fang B., Luo J., Li W. andZhang L., Heterologous expression,purification, and characterization ofxylose reductase from Candidashehatae, Biotechnol. Lett., 2007; 29:1409-1412.
[19] Kang M.H., Ni H. and Jeffries T.W.,Molecular characterization of a gene foraldose reductase (CbXYL1) fromCandida boidinii and its expressionin Saccharomyces cerevisiae, Appl.Biochem. Biotechnol., 2003; 105: 265-276.
[20] Lee J.K., Koo B.S. and Kim S.Y.,Cloning and characterization of the xyl1gene encoding an NADH-preferingxylose reductase from Candidaparapsilosis and its functional expressionin Candida tropicalis, Appl. Environ.Microbiol., 2003; 69: 6179-6188.
[21] Billard P., Menart S., Fleer R. andBolotin-Fukuhara M., Isolation andcharacterization of the gene encodingxylose reductase from Kluyveromyceslactis, Gene, 1995; 162: 93-97.
[22] Yokoyama S.I., Kinoshita Y., SuzukiT., Kawai K., Horitsu H. andTakamizawa K., Cloning andsequencing of two D-xylose reductasegenes (xyrA and xyrB) from Candidatropicalis, Ferment. Bioeng., 1995; 80:603-605.
[23] Handumrongkul C., Ma D.P. and SilvaJ.L., Cloning and expression of Candidaguilliermondii xylose reductase gene(xyl1) in Pichia pastoris, Appl. Microbiol.Biotechnol., 1998; 49: 399-404.
[24] Bolen P., Hayman G.T. and ShepherdH.S., Sequence and analysis of analdose (xylose) reductase gene from thexylose-fermenting yeast Pachysolentannophilus, Yeast, 1996; 12: 1367-1375.
[25] Amore R., K tter P., K ster C.,Ciriacy M. and Hollenberg C.P.,Cloning and expression inSaccharomyces cerevisiae of theNAD(P)H-dependent xylosereductase-encoding gene (XYL1) fromthe xylose-assimilating yeast Pichiastipitis, Gene, 1991; 109: 89-97.
[26] Higgins D.R. and Cregg J.M., Methodsin Molecular Biology Pichia Protocols,Humana Press Inc, New Jersey, 1998.
[27] Altschul S.F., Gish W., Miller W., MyersE.W. and Lipman D.J., Basic localalignment search tool, J. Mol. Biol.,1990; 215: 403-410.

502 Chiang Mai J. Sci. 2014; 41(3)
[28] Larkin M.A, Blackshields G., BrownN.P., Chenna R., Mc Gettigan P.A.,McWilliam H., Valentin F., WallaceI.M.,Wilm A., Lopez R., ThompsonJ.D., GibsonT.J. and Higgin D.G.,ClustalW and ClustalX version 2,Bioinform., 2007; 23: 2947-2948.
[29] Saitou N. and Nei M., The neighbor-joining method a new method forreconstructing phylogenetic trees, Mol.Biol. Evol., 1987; 4: 406-425.
[30] Bradford M.M., A rapid and sensitivemethod for the quantitation ofmicrogram quantities of protein utilizingthe principle of protein-dye binding,Anal. Biochem., 1976; 72: 248-254.
[31] Limtong S., Srisuk N., YongmanitchaiW., Kawasaki H., Yurimoto H.,Nakase T. and Kato N., Three newthermotolerant methylotrophic yeasts,Candida krabiensis sp. nov., Candidasithepensis sp. nov., and Pichia siamensissp. nov., isolated in Thailand, J. Gen.Appl. Microbiol., 2004; 50: 119-127.
[32] Limtong S., Srisuk N., YongmanitchaiW., Yurimoto H. and Nakase T.,Ogataea chonburiensis sp. nov. andOgataea nakhonphanomensis sp. nov.thermotolerant, methylotriphic yeastspecies isolated in Thailand, and transferof Pichia siamensis and Pichiathermomethanolica to the genus
Ogataea, Int. J. Syst. Evol. Microbiol.,2008; 58: 302-307.
[33] Arthur H. and Watson K., Thermaladaptation in yeast growth temperatures,membrane lipid, and cytochromecomposition of psychrophilic, mesophilicand thermophilic yeasts, J. Bacteriol.,1976; 128: 56-68.
[34] Vongsuvanlert V. and Tani Y., L-iditolproduction from L-sorbose by amethanol yeast, Candida boidinii(Kloeckera sp.) No. 2201, J. Ferment.Technol., 1988; 66: 517-523.
[35] Branco R.F., Santos J.C., MurakamiL.Y., Mussatto S.I., Dragone G. andSilva S.S., Xylitol production in abubble column bioreactor influence ofthe aeration and immobilized systemconcentration, Proc., Biochem., 2007; 42:258-262.
[36] Khoury G.A., Fazelinia H., Chin J.W.,Pantazes R.J., Cirino P.C. and MaranasC.D., Computational design of Candidaboidinii xylose reductase for alteredcofactor specificity, Protein Sci., 2009;18: 2125-2138.
[37] Yablochkova E.N., Bolotnikova O.I.,Mikhailova N.P., Nemova N.N., andGinak A.I., The activity of xylosereductase and xylitol dehydrogenase inyeasts, Microbiol., 2003; 72: 414-417.





























