Embed Size (px)
Citation preview

UNIVERZITET U NIŠU
PRIRODNO-MATEMATIČKI FAKULTET NIŠ
DEPARTMAN ZA BIOLOGIJU I EKOLOGIJU
Jelena M. Veličković
Zavisnost produkcije biomase mikroalge izolovane iz
Vlasinskog jezera od izvora azota u podlozi
MASTER RAD
Niš, 2016.

UNIVERZITET U NIŠU
PRIRODNO-MATEMATIČKI FAKULTET NIŠ
DEPARTMAN ZA BIOLOGIJU I EKOLOGIJU
MASTER RAD
Zavisnost produkcije biomase mikroalge izolovane iz
Vlasinskog jezera od izvora azota u podlozi
Kandidat: Mentor:
Jelena Veličković 145 Dr Nataša Joković
Niš, 2016.

UNIVERSITY OF NIS
FACULTY OF SCIENCES AND MATHEMATICS
DEPARTMENT OF BIOLOGY AND ECOLOGY
MASTER THESIS
The dependence of biomass production from the
microalga isolated from Vlasina lake on the source of
nitrogen in media
Candidate: Mentor:
Jelena Veličković PhD Nataša Joković
Niš, 2016.

Zahvalnica
Veliku zahvalnost dugujem svom mentoru, prof. dr Nataši Joković, za izdvojeno
vreme, strpljenje i uložen trud. Hvala na savetima, podršci i pomoći koju ste mi
pružili u toku izrade ovog master rada.
Zahvaljujem se i asistentu, Nikoli Stankoviću, za nesebičnu pomoć, dragocene
savete, strpljenje i vreme koje je uložio u toku izvođenja eksperimentalnog dela
rada.
Takođe, zahvaljujem se i dr Ivani Kostić na pomoći i vremenu koje je izdvojila u
toku eksperimentalnog dela rada.
Posebnu zahvalnost dugujem svojoj porodici bez čije podrške i pomoći ne bih
bila ovde. Beskrajno vam hvala na svemu…Volim vas…
Ne mogu ništa drugo reć' do hvala, i hvala i navijek hvala.
(William Shakespeare)

Biografija
Ime i prezime: Jelena Veličković
Datum rođenja: 16. 5. 1992.
Mesto rođenja: Surdulica
Osnovna škola: ,,Vuk Karadžić“ Surdulica (1999 –
2007)
Srednja škola: Gimnazija ,,Svetozar Marković“
Surdulica (2007 – 2011)
Fakultet: Prirodno-matematički fakultet u Nišu,
departman za biologiju i ekologiju
(2011 – 2016)
Osnovne akademske studije: smer biologija (2011 – 2014)
Master akademske studije: smer biologija (2014 – 2016)

Sažetak
Mikroalge predstavljaju značajnu sirovinu u biotehnologiji zbog visokog procenta
ugljenih hidrata, proteina, lipida i drugih metabolita koje sintetišu. Sojevi mikroalgi koji se
koriste u biotehnologiji, izoluju se iz različitih prirodnih sredina, a zatim se vrši njihova
karakterizacija u laboratorijskim uslovima. Kako bi se produkovala dovoljna biomasa
mikroalgi koja se može koristiti za dalja ispitivanja, potrebno je uraditi optimizaciju rasta
mikroalgi. Optimizacija rasta podrazumeva određivanje sastava podloge kao i uslova pod
kojima određeni soj mikroalge daje maksimalnu produkciju biomase.
U ovom radu, iz uzoraka vode sa Vlasinskog jezera, izolovan je jedan izolat
mikroalge koji je identifikovan kao predstavnik roda Closterium. Da bi se ispitalo kako izvori
azota utiču na rast mikroalge, mikroalga je gajena 16 dana na podlogama sa različitim
izvorima azota, amonijum fericitratom, natrijum nitratom i mešavinom ova dva jedinjenja.
Rast mikroalge praćen je spektrofotometrijski dok je usvajanje azota iz podloge određivano
hemijskim spektrofotometrijskim metodama. Nakon mesec dana gajenja mikroalge, određena
je produkovana biomasa gravimetrijskom metodom.
Mikroalga je pokazala dobar rast na podlozi sa amonijum fericitratom kao jedinim
izvorom azota, kao i na podlozi sa mešovitim izvorima azota, amonijum fericitratom i
natrijum nitratom. Rezultati hemijskih analiza uzoraka podloga na kojima je rasla mikroalga
pokazuli su da mikroalga ne usvaja nitrate iz podloga za rast već da koristi amonijumov jon
kao izvor azota. Prinos biomase nakon mesec dana gajenja mikroalge bio je najveći na
podlozi koja je sadržala amonijum fericitrat i natrijum nitrat.
Ključne reči: mikroalga, biomasa, izvori azota

Abstract
Microalgae represent a significant raw material in biotechnology due to their high
percentage of carbon hydrates, proteins, lipids and other metabolites they synthetize.
Microalgal strains which are used in biotechnology are isolated from different natural habitats
and then characterized in laboratory conditions. In order to produce a great enough biomass
of microalgae for use in further study, it is necessary to perform a growth optimization of
microalgae. Growth optimization implies determination of the media composition as well as
conditions under which a certain strain of microalgae gives the maximal production of
biomass.
In this study we have isolated an isolate of the microalga from water samples of
Vlasina lake, which has been identified as a representative of the Closterium genus. In order
to examine how nitrogen sources affect the growth of microalga, the microalga was grown
for 16 days on medias with different nitrogen sources, like ammonium fericitrate, sodium
nitrate and a mixture of these two compounds. The growth of microalga was
spectrophotometrically observed while the nitrogen absorption from the media was
determined by chemical spectrophotometric methods. After a month of microalgal growth,
the produced biomass was determined by a gravimetric method.
The microalga has shown a good growth on the medium with ammonium fericitrate as
the only nitrogen source, as well as on the medium with a mixture of nitrogen sources,
ammonium fericitrate and sodium nitrate. The results of chemical analysis of medium
samples on which the microalga grew have shown that the microalga doesn’t absorb nitrates
from medium for growth, but it instead uses ammonia ion as the source of nitrogen. The
biomass yield after a month of microalgal growth was the biggest on the medium which
contained ammonium fericitrate and sodium nitrate.
Key words: microalgae, biomass, nitrogen source

Sadržaj
1. Uvod ....................................................................................................................................... 1
1.1. Alge ................................................................................................................................. 2
1.2. Klasifikacija algi ............................................................................................................. 3
1.3. Alge u jezerima ............................................................................................................... 9
1.4. Uloga i značaj algi ......................................................................................................... 10
1.5. Upotreba algi u biotehnologiji....................................................................................... 11
1.6. Produkcija biomase algi ................................................................................................ 13
2. Ciljevi rada ........................................................................................................................... 15
3. Materijal i metode ................................................................................................................ 16
3.1. Materijal ........................................................................................................................ 16
3.1.1. Uzorkovanje............................................................................................................ 16
3.1.2. Podloge ................................................................................................................... 16
2.1.3. Reagensi.................................................................................................................. 16
3.2. Metode rada ................................................................................................................... 17
3.2.1. Izolacija algi ........................................................................................................... 17
3.2.2. Identifikacija algi .................................................................................................... 18
3.2.3. Zasejavanje podloga ............................................................................................... 18
3.2.4. Određivanje biomase algi ....................................................................................... 18
3.2.5. Određivanje koncentracije nitrata ........................................................................... 18
3.2.6. Određivanje koncentracije amonijumove soli ........................................................ 19
4. Rezultati i diskusija .............................................................................................................. 20
4.1. Izolacija algi iz Vlasinskog jezera ................................................................................. 20
4.2. Identifikacija mikroalge ................................................................................................ 20
4.3. Rast mikroalge u zavisnosti od izvora azota ................................................................. 21
4.4. Određivanje koncentracije nitrata i amonijum fericitrata u podlogama ........................ 23
4.5. Gravimetrijsko određivanje biomase mikroalge ........................................................... 25
5. Zaključci ........................................................................................................................... 27
6. Literatura .......................................................................................................................... 28

1
1. Uvod
Alge su fotosintetički jednoćelijski i višećelijski organizmi veličine od nekoliko
mikrometara do nekoliko desetina metara. Jednoćelijske mikroskopske alge nazivaju se
mikroalge i predstavljaju heterogenu grupu organizama koji naseljavaju različita staništa.
Mikroalge imaju značajnu ulogu u vodenim ekosistemima gde čine fitoplankton jer
predstavljaju producente organske materije, sintetišu kiseonik i smanjuju količinu ugljen-
dioksida u atmosferi.
Mikroalge se sve više upotrebljavaju u biotehnologiji kao novi izvori hrane za čoveka i
kao industrijska sirovina. S obzirom da veoma brzo rastu u laboratorijskim uslovima pri čemu
produkuju velike količine lipida, ugljenih hidrata i proteine, mogu se koristiti za različite
svrhe u biotehnologiji. Poslednjih godina, mikroalge predstavljaju atraktivnu sirovinu za
proizvodnju novih goriva, naročito biodizela jer neki sojevi produkuju velike količine ulja.
Sojevi mikroalgi koji se koriste u biotehnologiji, izolovani su iz različitih prirodnih
sredina. Danas postoji veliki broj projekata širom sveta koji se bave izolacijom,
identifikacijom i karakterizacijom sojeva mikroalgi sa ciljem upotrebe u biotehnologiji. S
obzirom da je rast mikroalgi u prirodnim uslovima i laboratorijskim uslovima različit,
potrebno je svaki izolovani soj mikroalgi ispitati u laboratoriji. Prvi korak je optimizacija
rasta mikroalgi kako bi se produkovala dovoljna biomasa koja se može koristiti za dalja
ispitivanja. Optimizacija rasta podrazumeva određivanje sastava podloge kao i uslova pod
kojima određeni soj mikroalge daje maksimalnu produkciju biomase.

2
1.1. Alge
Alge su široko rasprostranjena i značajna grupa organizama u prirodi. Naziv alga
potiče od latinske reči algae što znači morska trava. Neki predstavnici su višećelijski i imaju
vegetativno telo koje se naziva talus. Za razliku od biljaka, njihovo telo nikada nije
diferencirano na koren, stablo i list. Kod makroskopskih algi veličina talusa može biti i
nekoliko desetina metara, dok neke vrste morskih algi mogu da dostignu veličinu i do 200
metara. Posebnu grupu algi čine jednoćelijske alge ili mikroalge koje su mikroskopskih
veličina i mogu da formiraju kolonije kada rastu na odgovarajućim podlogama u
laboratorijskim uslovima.
Alge u svojim ćelijama sadrže hlorofil tako da mogu da vrše fotosintezu. Fotosinteza
je najvažniji biohemijski proces u kome se iz ugljen dioksida i vode sintetišu organska
jedninjenja, pri čemu se oslobađa kiseonik. Da bi se fotosinteza odvijala, neophodni su
određeni pigmenti koji su smešteni u organelama koje se zovu plastidi. Osnovni pigmenti su
iz grupe hlorofila, čiji se plastidi nazivaju hloroplasti. Osim njih, za fotosintezu su značajni i
pigmenti iz grupe karotena, ksantofila i fikobilina. Ukoliko oni dominiraju u plastidima, onda
se ti plastidi nazivaju hromoplasti. Plastidi mogu da budu različitog oblika (pločasti,
peharasti, zvezdasti, trakasti i slično). Postoje četiri vrste hlorofila: a, b, c i d. Hlorofil a,
karoteni i ksantofili se nalaze kod svih algi, dok se ostali pigmenti mogu naći kod pojedinih
predstavnika. Karoteni, ksantofili i fikobilini predstavljaju pomoćne pigmente, jer
apsorbovanu svetlost prenose do hlorofila a. Njihov značaj se ogleda u tome što mogu da
apsorbuju svetlost talasne dužine koju hlorofili ne mogu. Takođe, boja algi zavisi od
pigmenata, njihovog sastava i količine, kao i od svetlosnih uslova u kojima se razvijaju. Iako
su alge većinom fotoautotrofni organizmi, postoje i vrste koje se hrane heterotrofno
(Blaženčić, 2000).
Prvobitna sredina u kojoj su alge nastale je voda, ali su se mnoge tokom evolucije
prilagodile životu na kopnu. Možemo ih naći u zemljištu, na kori drveća i svim osvetljenim i
vlažnim mestima na kopnu. Žive samostalno ili u zajednici sa biljnim i životinjskim
organizmima (Blaženčić, 2000). Mogu da naseljavaju površinu biljaka i životinja, ostvaruju
simbiotske odnose kao što su alge i gljive u lišajima i alge i neke životinje primitivne
organizacije – zoohlorele. Takođe, među algama su otkriveni i paraziti (Mihailov-Krstev,
2008).

3
1.2. Klasifikacija algi
Alge su heterogena grupa organizama koja obuhvata prokariotske i eukariotske vrste.
U prokariote spadaju modrozelene alge (Cyanobacteriophyta) dok sve ostale spadaju u
eukariote (Duong et al., 2012). Pretpostavlja se da postoji 200 000 – 800 000 vrsta algi, od
kojih je do sada opisano oko 35 000 (Cardozo et al., 2007).
Klasifikacija algi se vrši na osnovu više kriterijuma. Osnovni kriterijumi su: prisustvo
ili odsustvo i građa pravog jedra, vrste pigmenata, rezervne materije, hemijski sastav
ćelijskog zida, prisustvo, broj i izgled flagela, način razmnožavanja (Mihailov-Krstev, 2008).
Međutim, neka od osnovnih pitanja filogenije nisu u potpunosti razjašnjena i to onemogućava
stvaranje jedinstvenog filogenetskog sistema za klasifikaciju algi. Zbog toga postoji veći broj
klasifikacionih shema gde su ovi organizmi svrstani u 8, 9, 10 ili više razdela (Blaženčić,
2000).
Alge su, na osnovu prethodno pomenutih kriterijuma, podeljene u 10 razdela:
Cyanobacteriophyta, Euglenophyta, Pyrrophyta, Bacillariophyta, Rhodophyta, Xanthophyta,
Chrysophyta, Phaeophyta, Chlorophyta i Charophyta (Mihailov-Krstev, 2008).
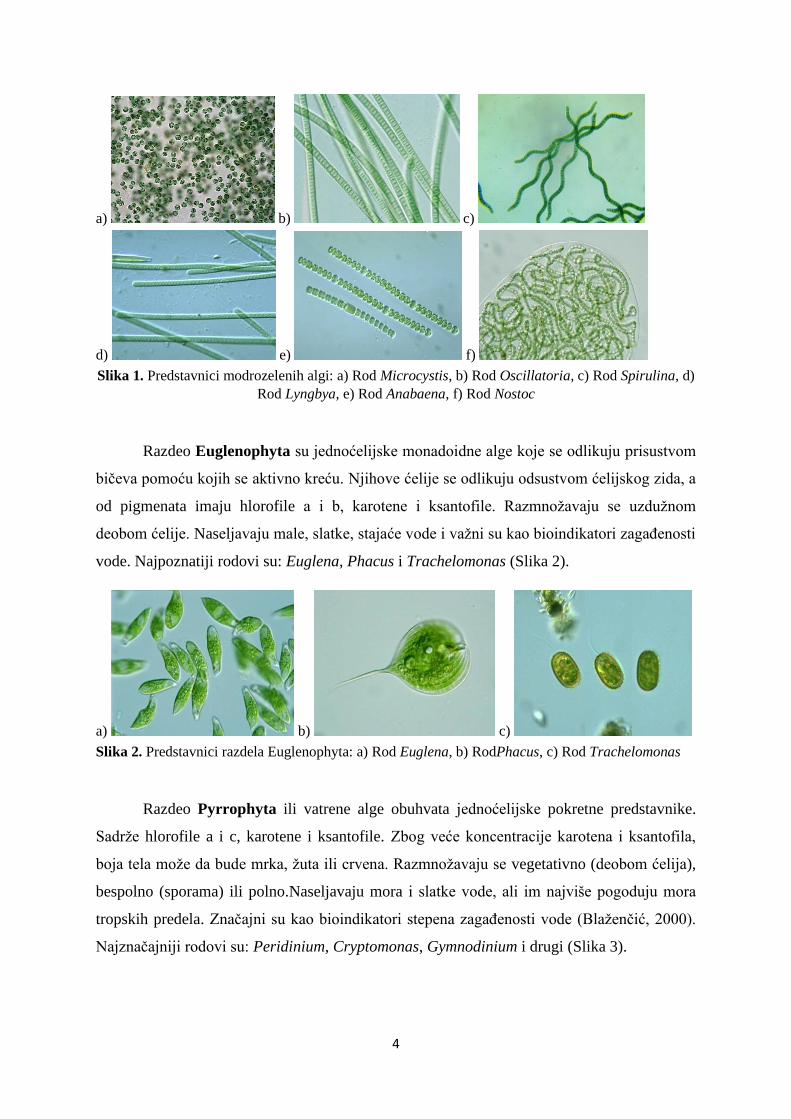
Razdeo Cyanobacteriophyta su modrozelene alge i jedini predstavnici sa
prokariotskim tipom ćelije. Tu spadaju najstariji jednoćelijski fotoautotrofni organizmi.
Sadrže hlorofil a, karotene, ksantofile i fikobilizome (granule sa c-fikocijaninom i c-
fikoeritrinom). Prilikom razmnožavanja deobom ćelija mogu da formiraju kolonije.
Naseljavaju sve tipove ekosistema i otporne su na temperaturne promene i sunčevu radijaciju.
Neki od najpoznatijih predstavnika pripadaju rodovima: Microcystis, Oscillatoria, Spirulina,
Lyngbya, Anabaena, Nostoc i drugi (Slika 1).
4
a) b) c)
d) e) f)
Slika 1. Predstavnici modrozelenih algi: a) Rod Microcystis, b) Rod Oscillatoria, c) Rod Spirulina, d)
Rod Lyngbya, e) Rod Anabaena, f) Rod Nostoc
Razdeo Euglenophyta su jednoćelijske monadoidne alge koje se odlikuju prisustvom
bičeva pomoću kojih se aktivno kreću. Njihove ćelije se odlikuju odsustvom ćelijskog zida, a
od pigmenata imaju hlorofile a i b, karotene i ksantofile. Razmnožavaju se uzdužnom
deobom ćelije. Naseljavaju male, slatke, stajaće vode i važni su kao bioindikatori zagađenosti
vode. Najpoznatiji rodovi su: Euglena, Phacus i Trachelomonas (Slika 2).
a) b) c)
Slika 2. Predstavnici razdela Euglenophyta: a) Rod Euglena, b) RodPhacus, c) Rod Trachelomonas
Razdeo Pyrrophyta ili vatrene alge obuhvata jednoćelijske pokretne predstavnike.
Sadrže hlorofile a i c, karotene i ksantofile. Zbog veće koncentracije karotena i ksantofila,
boja tela može da bude mrka, žuta ili crvena. Razmnožavaju se vegetativno (deobom ćelija),
bespolno (sporama) ili polno.Naseljavaju mora i slatke vode, ali im najviše pogoduju mora
tropskih predela. Značajni su kao bioindikatori stepena zagađenosti vode (Blaženčić, 2000).
Najznačajniji rodovi su: Peridinium, Cryptomonas, Gymnodinium i drugi (Slika 3).

5
a) b) c)
Slika 3. Predstavnici razdela Pyrrophyta: a) Rod Peridinium, b) Rod Cryptomonas, c) Rod
Gymnodinium
Razdelu Bacillariophyta pripadaju jednoćelijske i kolonijalne silikatne alge. Ove alge
se odllikuju prisustvom čvrstog omotača koji se naziva silikatni pancir ili teka. Imaju
hlorofile a i c, karotene i ksantofile. Razmnožavaju se vegetativno (deobom ćelija) i polno.
Naseljavaju slane i slatke vode, tresave, vlažno kamenje, zemljište, sneg i led. Dele se na 2
klase: Centrophyceace (centrične silikatne alge) sa najpoznatijim predstavnicima iz roda
Melosira i Pennatophyceace (penatne silikatne alge) sa poznatim predstavnicima iz rodova
Cocconeis, Navicula, Cymbella i drugi (Slika 4) (Mihailov-Krstev, 2008). Ove alge su
osetljive na koncentraciju hlorida u vodi, što ih čini značajnim bioindikatorima. Takođe se
koriste i u procesima biološkog čišćenja zagađenih voda. Osim što su primarni producenti
organske materije, ove alge su bogate vitaminima, belančevinama i mastima, pa su kvalitetna
hrana, a pritom produkuju i veliku količinu biomase. Ove alge poznate su i po tome što se od
njihovih ljuštura nakon uginuća stvara dijatomejski mulj. Naslage ovog mulja se nazivaju
dijatomit i koriste se u građevinarstvu, prehrambenoj i hemijskoj industriji (Blaženčić,2000).
a) b)
c) d)
Slika 4. Predstavnici razdela Bacillariophyta: a) Rod Melosira, b) Rod Cocconeis, c) Rod Navicula,
d) Rod Cymbella

6
Razdeo Rhodophyta ili crvene alge obuhvata jednoćelijske, kolonijalne alge.
Odlikuju se prisustvom dvoslojnog ćelijskog zida čiji spoljašnji sloj bubri u vodi i sluzav je.
Imaju hlorofile a i d, narandžaste karotene i ksantofile i plave i crvene fikobiline.
Razmnožavaju se vegetativno (deobom, raskidanjem kolonije), bespolno (sporama) ili polno.
Široko su rasprostranjene, naseljavaju bentos u priobalnoj zoni toplih mora, ali i u većim
dubinama, mogu se naći i u planktonu. Predstavnici pripadaju rodovima: Bangia, Porphyra,
Batrachospermum i drugi (Slika 5).
a) b) c)
Slika 5. Predstavnici razdela Rhodophyta:a) Rod Bangia, b) Rod Porphyra, c) Rod Batrachospermum
Razdeo Xanthophyta ili žutozelene alge su jednoćelijski, kolonijalni i višećelijski
organizmi. Imaju hlorofile a i c ili a i e, karotene i ksantofile koji daju boju talusu.
Razmnožavaju se vegetativno, bespolno ili polno. Široko su rasprostranjene i uglavnom
naseljavaju slatke čiste vode, ređe se mogu naći u morima, brakičnim vodama i zemljištu.
Najpoznatiji rodovi su: Tribonema, Vacheria i drugi (Slika 6).
a) b)
Slika 6. Predstavnici razdela Xanthophyta: a) Rod Tribonema, b) Rod Vaucheria
Razdeo Chrysophyta ili zlatne alge obuhvata jednoćelijske, kolonijalne i višećelijske
alge. Imaju hlorofile a i c, karotene i ksantofile. Razmnožavaju se vegetativno, bespolno ili
polno. Žive u slatkim, brakičnim i slanim vodama, uglavnom u planktonu. Predstavljaju
važnu kariku u lancima ishrane, kao i značajne primarne producente organske materije. Neke
vrste mogu da izazovu tzv. cvetanje vode, što je pokazatelj lošeg kvaliteta vode. Najznačajniji
rodovi su: Dinobryon, Chrysamoeba, Hydrurus i drugi (slika 7).

7
a) b) c)
Slika 7. Predstavnici razdela Chrysophyta: a) Rod Dinobryon, b) Rod Chrysamoeba, c) Rod Hydrurus
Razdeo Chlorophyta ili zelene alge obuhvata jednoćelijske, kolonijalne i višećelijske
predstavnike, aktivno pokretne i nepokretne. Zelena boja potiče od hlorofila a i hlorofila b, a
sadrže i karotene i ksantofile. Razmnožavaju se vegetativno, bespolno ili polno. Kosmopoliti
su, a najčešće se mogu naći u planktonu i bentosu slatkovodnih ekosistema. Najpoznatiji su
predstavnici rodova: Chlamidomonas, Scenedesmus, Volvox, Ulotrix, Spirogyra, Chlorella,
Cosmarium, Pediastrum i drugi (Slika 8).
a) b) c)
d) e) f)
g) h)
Slika 8. Predstavnici razdela Chlorophyta: a) Rod Chlamidomonas, b) Rod Scenedesmus, c) Rod
Volvox, d) Rod Ulotrix, e) Rod Spirogyra, f) Rod Chlorella, g) Rod Cosmarium, h) Rod Pediastrum

8
Razdeo Charophyta ili pršljenčice su višećelijske krupne alge koje podsećaju na
rastaviće (Equisetum) i jasno se razlikuju od drugih algi po svom talusu. Sadrže hlorofile a i
b, karotene i ksantofile. Razmnožavaju se vegetativno i polno. Naseljavaju uglavnom slatke
vode i bentosne su. Razdeo sadrži jednu klasu Characeae sa najpoznatijim predstavnicima iz
rodova Chara i Nitella (Slika 9).
a) b)
Slika 9. Predstavnici razdela Charophyta: a) Rod Chara, b) Rod Nitella
Razdeo Phaeophyta ili mrke alge su višećelijske, sesilne alge. Imaju krupan talus,
dvoslojni ćelijski zid koji bubri u vodi. Sadrže hlorofile a i c, narandžaste karotene i
ksantofile i fukoksantin koji je im daje mrku boju. Razmnožavaju se vegetativno, bespolno i
polno (Mihailov-Krstev, 2008). Samo 3 roda žive u slatkim vodama (Pleurocladia, Bodanella
i Lithoderma) (Blaženčić, 2000). Najznačajniji predstavnici pripadaju rodovima: Fucus,
Laminaria, Macrocystis, Ectocarpus i drugim (Slika 10).
a) b)
c) d)
Slika 10. Predstavnici razdela Phaeophyta: a) Rod Fucus, b) Rod Laminaria, c) Rod Macrocystis, d)
Rod Ectocarpus

9
1.3. Alge u jezerima
Alge koje naseljavaju vodenu sredinu dele se na bentosne i planktonske alge. Podela
je izvršena na osnovu dela vodenog basena u kome se razvijaju.
Životni ciklus bentosnih algi vezan je za dno vodenog basena, gde one mogu da budu
pričvršćene za podlogu (sesilne) ili nepričvršćene odnosno slobodne. Najveći broj bentosnih
algi koje mogu da se nađu u slatkim vodama pripadaju razdelu Chlorophyta i većina
predstavnika je sesilna. Osim njih, brojne su i alge iz razdela Bacillariophyta,
Cyanobacteriophyta i Chlorophyta, čiji su predstavnici uglavnom slobodni. Rasprostranjenje
algi zavisi od svetlosnog režima, pa se u čistim i prozračnim jezerima one mogu naći i na
dubinama od 50 m, dok se u manje providnim jezerima ne rasprostiru na većim dubinama od
10 m.
Planktonske alge čine jednoćelijski i kolonijalni organizmi koji slobodno lebde u
vodenoj masi i sa drugim biljnim organizmima čine fitoplankton. Ove alge su razvile brojne
morfološke i fiziološke adaptacije u cilju smanjenja svoje specifične težine kako bi se održale
slobodne u vodi. Neke od tih adaptacija su: gasne vakuole, ulje kao rezervna supstanca, razni
izraštaji, loptasto telo i druge. Rasprostranjenje ovih algi zavisi od svetlosnog režima,
temperature vode, saliniteta, mineralnih soli, ugljen-dioksida i kiseonika. Vegetacioni ciklus
fitoplanktona u jezerima koja se nalaze u umereno-kontinentalnoj klimi počinje u proleće, u
periodu mart-april. Prvi predstavnici jednoćelijskih algi koji se javljaju pripadaju razdelima
Pyrrophyta, Chrysophyta i Bacillariophyta i one su aktivno pokretne. Sa porastom
temperature (10 – 12 ºC) raste broj silikatnih algi, kojima se početkom leta pridružuje sve
veći broj zelenih, modrozelenih i zlatnih algi. Zelene i modrozelene alge su najbrojnije kada
temperatura vode dostigne 15 ºC i više i tada preovladavaju predstavnici rodova: Anabaena,
Microcystis, Oscillatoria, Pediastrum i druge. U jesen, sa opadanjem temperature, raste broj
silikatnih algi, a prisutne su i modrozelene alge. Na razviće fitoplanktona, osim temperature,
utiče i koncentracija nitrata i fosfata. S obzirom da je najveća koncentracija ovih jedinjenja u
proleće i jesen, tada se fitoplakton najviše i razvija. U toku leta dominiraju vrste kojima nije
potreba visoka koncentracija ovih jedinjenja i azot usvajaju iz minimalnih količina
(Blaženčić, 2000).

10
1.4. Uloga i značaj algi
Alge su osnovni producenti organske materije u ekosistemu, a primarna produkcija
predstavlja materijalnu i energetsku osnovu svih produkcionih odnosa u ekosistemu. Njihov
značaj se ogleda u tome što najveći broj ovih organizama pripada fotoautotrofima, odnosno
ima sposobnost fotosinteze. Brojna istraživanja primarne organske produkcije algi pokazala
su da se zahvaljujući njima stvara ogromna količina organskih materija na planeti (Blaženčić,
2000). S obzirom da se u procesu fotosinteze koristi ugljen-dioksid iz atmosfere, alge imaju
važnu ulogu u smanjenju efekta staklene bašte i globalnog zagrevanja (Wang et al., 2008;
Sheehan et al., 1998). Produkcija 100 tona biomase algi uklanja oko 183 tona ugljen-dioksida
iz atmosfere (Chisti, 2007). Osim toga, alge predstavljaju neiscrpan izvor kiseonika. Prema
procenama, mikroalge proizvode polovinu ukupnog atmosferskog kiseonika (Cardozo et al.,
2007).
Alge iz razdela Cyanobacteriophyta (modrozelene alge) imaju sposobnost vezivanja
molekularnog azota. Azot se nalazi u velikim količinama u atmosferi, ali je u tom obliku
nedostupan većini biljaka. Zbog toga je neophodno da se takav azot prevede u stanje u kome
biljke mogu da ga koriste. Ovu sposobnost, pored nekih mikroorganizama i bakterija, imaju i
modrozelene alge. Kako je kruženje azota jedan od najznačajnih bioloških ciklusa u procesu
kruženja materije, to je i značaj ovih algi veći (Blaženčić, 2000).
Alge su značajni indikatori stepena zagađenosti voda, jer su osetljive na kvalitativne i
kvantitativne promene osobina vode. Neke alge su miksotrofi što znači da koriste organske
supstance iz okolne sredine za rast i razvoj. Zahvaljujući ovoj sposobnosti, alge imaju značaj
u procesima biološkog prečišćavanja zagađenih voda. Njihova uloga se ogleda u tome što
koriste ugljen-dioksid koji se oslobađa u procesu truljenja, a za uzvrat oslobađaju i obogaćuju
vodu kiseonikom. Ovo njihovo svojstvo je našlo primenu i pri prečišćavanju otpadnih voda
(Blaženčić, 2000). Pokazalo se da su alge sposobne da uklanjaju policiklične aromatične
ugljovodonike, fenole, organske rastvarače, teške metale i patogene (Muñoz, 2006).
Alge su značajne za ishranu ljudi i životinja. Bogate su ugljenim hidratima,
belančevinama, mastima, vitaminima i mineralnim solima. Sadrže i znatne količine joda,
broma i drugih supstanci koje im daju dodatnu hranljivu vrednost. U ove svrhe više se koriste
morske od slatkovodnih algi (Blaženčić, 2000).

11
Alge su pogodne i za đubrenje zemljišta. Pored kalijumovih, azotnih i fosfornih soli,
preko njih u podlogu dospevaju i jod, arsen, bor i drugi elementi. Uglavnom se za dobijanje
đubriva koriste najkrupnije alge (Tešić i Marinović, 1966).
Alginati, soli alginske kiseline, koje se nalaze kod mrkih algi, koriste se u tekstilnoj,
prehrambenoj, građevinskoj i kozmetičkoj industriji (Tešić i Marinović, 1966). U tekstilnoj
industriji se koriste za slepljivanje vlakana, u građevinarstvu se dodaju cementu i asfaltu, kao
i drugim materijalima kako bi povećale njihove hidroizolacione sposobnosti (Blaženčić,
2000).
Agar-agar je jedna od značajnijih materija koja se dobija iz algi, uglavnom iz crvenih.
Koristi se za pravljenje čvrstih podloga u medicini i biologiji, zatim u tekstilnoj,
prehrambenoj i farmaceutskoj industriji, jer daje čvrstinu proizvodima (Blaženčić, 2000).
1.5. Upotreba algi u biotehnologiji
Glavni pritisak za korišćenje algi u biotehnologiji pojavio se sedamdesetih godina
prošlog veka kako bi se prevazišao manjak hrane i energije (Chen et al., 2009). Veliki
varijetet dostupnih mikroalgi može da se koristi u različite svrhe, kao što su proizvodi koji se
koriste u farmaceutske svrhe, usevi hrane za ljudsku potrošnju i izvori energije (Yin et al.,
2011). Poslednjih godina se mikroalge sve češće upotrebljavaju, jer mogu da obezbede
sirovine za različite tipove goriva kao što su biodizel, etanol, vodonik i metan koji su brzo
biorazgradivi i mogu da budu efikasniji od fosilnih goriva. U Americi je 2011. godine
proizvedeno bilion galona biodizela, naredne godine 1.1 bilion galona, dok je u 2013.
proizvodnja dostigla, čak, 1.8 biliona galona (Anonimus).
Zbog sve veće potražnje biogoriva postoji potreba za otkrićem produktivijih
nehranljivih izvora biomase koja može da se konvertuje u biodizel ili druga goriva (Lee et al.,
2014). Biodizel je netoksično i biorazgradivo alternativno gorivo koje se dobija hemijskom
reakcijom triacilglicerola (biljnih ulja ili životinjske masti) sa alkoholom pri čemu se
formiraju alkil estri masnih kiselina. Neke vrste algi, takođe, mogu da sintetišu
triacilglicerole koji se koriste kao osnovna sirovina za proizvodnju biodizela. Alge su bolja
sirovina za dobijanje biodizela u odnosu na biljke (suncokret, uljana repica i drugo) jer:

12
1. imaju veću produktivnost biomase u odnosu na biljke tako da neke vrste mogu da
udvostruče svoju biomasu za nekoliko sati,
2. neke vrste algi mogu da akumuliraju veliku količinu triacilglicerola u obliku ulja i
3. nije potrebno zemljište za gajenje algi kao što je slučaj sa biljkama (Scott et al., 2010).
Najbolje alge za proizvodnju biodizela su mikroalge, jer produkuju više ulja od
makroalgi i imaju brži rast (Altin et al., 2001), tako da predstavljaju obećavajući izvor ulja za
proizvodnju biodizela (Danilović et al., 2013). Jednoćelijske mikroalge rastu veoma brzo i
mogu da prođu ceo životni ciklus za nekoliko dana ako postoje adekvatni uslovi za njihov
razvoj (svetlost, voda, ugljen-dioksid, nutritijenti) (Li et al., 2008; Liu et al., 2008). Neke
vodene vrste algi imaju visok sadržaj ulja (i do 60% suve težine) i potencijal da postanu
značajno produktivniji izvor za proizvodnju biodizela kada se uporede sa terestričnim
usevima kao što je soja (Chisti, 2007; Hossain et al., 2008; Sakai et al., 1995; Kurano et al.,
1996). Naime, njihova mogućnost proizvodnje je do 100 puta više biodizela po jedinici
površine gajenja u odnosu na suncokret i uljanu repicu (Mata et al., 2010). Takođe, troškovi
transporta i izdvajanja mikroalgi su niži u odnosu na troškove drugih sirovina. Ne zahtevaju
mnogo prostora za gajenje i što je od velikog značaja nemaju negativan uticaj na svetske
zalihe hrane i vode (Hundt i Reddy, 2011). Njihovo gajenje može biti sprovedeno na mestima
koja nisu pogodna za razviće drugih biljaka. Osim toga, mogu se gajiti i na farmama i u
bioreaktorima (Danilović et al., 2013). Alge mogu da produkuju 1000 – 4000 galona ulja po
hektaru godišnje više od soje i drugih useva, mogu da žive u širokim klimatskim i vodenim
uslovima, nisu tradicionalna hrana, pa nisu u kompeticiji sa poljoprivredom (Chen et al.,
2009).
U poređenju sa drugim izvorima (životinjska mast, zrna žitarica i palmino ulje),
postoje izuzetne prednosti kada je u pitanju proizvodnja biodizela iz ulja mikroalgi (Duong et
al., 2012). Biodizel iz ulja algi ne sadrži sumpor, smanjuje emisiju čestica, ugljen-monoksida
i ugljovodonika. Proizvodnja ovakvog biodizela je možda jedini način da se proizvede
dovoljno motornog goriva da bi se zamenila sadašnja potrošnja benzina (Yin et al., 2011).
Veliki problem sa drugim alternativnim gorivima je taj što nisu kompatibilna sa
postojećim vozilima. Biodizel iz algi ima veliku prednost nad ostalim gorivima zato što je
hemijski kompatibilan sa postojećim dizel gorivom (Neltner, 2008).
Osim za proizvodnju biodizela, alge se mogu koristiti i u drugim granama
biotehnologije. Zahvaljujući velikoj sposobnosti adaptacije, mikroalge su postala dominantna

13
grupa organizama na zemlji, bogate su skrobom, uljima, proteinima i mogu da akumuliraju
važne sekundarne metabolite. Neke vrste, kao što je Spirulina sadrže mnogo proteina i gaje se
u različitim zemljama na industrijskom nivou (Chen et al., 2009). Potencijalna industrijska
primena algi se ogleda u tome što je preko 15 000 komponenti (antioksidansi, enzimi, masne
kiseline, peptidi, steroli i toksini) dobijeno iz algi (Danilović et al., 2013). Produkti
ekstrahovani iz algi mogu da se koriste kao izvor za organsko đubrivo ili produkte visoke
vrednosti kao što su omega-3 masne kiseline, steroli, karotenoidi i drugi pigmenti i
antioksidansi (Wang et al., 2008). Istraživanja su pokazala da mikroalge imaju potencijal da
se koriste kao vakcine za ljude i ribe (Rosenberg et al., 2008).
1.6. Produkcija biomase algi
Rast ćelija je jedna od fundamentalnih odlika koja određuje maksimalni potencijal
produkcije biomase algi. Ćelijski rast se sastoji iz dve faze – ranije, ćelijske proliferacije i
kasnije, povećanje obima ćelija. Obe direktno doprinose akumulaciji biomase. Očekuje se da
su brza deoba i rast ćelija najvažniji za produkciju biomase. Za neke manje i okrugle
mikroalge, čiji je dijametar nekoliko mikrometara, produkcija biomase je determinisana
stopom proliferacije (Chen et al., 2009).
Kao što je rečeno, alge su organizmi koji u procesu fotosinteze sintetišu
makromolekule. Konvertuju neorganski ugljenik u organske materije i snabdevaju sve
organizme na planeti energijom i supstancama neophodnim za život i rast. Neke vrste, pak,
koriste gotove organske molekule iz okruženja i ugrađuju ih. Takva metabolička aktivnost je
poznata pod nazivom heterotrofija. Za razliku od autotrofnih algi, heterotrofne rastu veoma
dobro i pri velikoj gustini ćelija, pa je njihova produkcija biomase obično veća. Postoje i
vrste algi koje mogu da koriste i ugljen dioksid i organske molekule, pa je njihova produkcija
biomase, verovatno, negde između prethodna dva načina ishrane (Chen et al., 2009).
Zahvaljujući jednostavnoj jednoćelijskoj strukturi, fotosinteza se odvija veoma brzo,
ugljenik se efikasno vezuje i produkcija biomase je brza (Danilović et al., 2013). Faktori koji
ograničavaju rast mikroalgi su: abiotički (svetlost, temperatura, koncentracija nutritijenata,
kiseonik, ugljen dioksid, pH, salinitet, toksični elementi), biotički (prisustvo patogena i

14
kompeticija sa drugim vrstama algi) i procesni (mešanje, stepen razblaženja i drugi) (Mata et
al., 2010).
Proizvodnja biomase fotoautotrofnih kultura je ograničena, jer se sa povećanjem broja
ćelija smanjuje dostupnost svetlosti. Povećan intenzitet svetlosti i malo iznad intenziteta koji
obezbeđuje maksimalnu brzinu rasta, dovodi do fotoinhibicije, odnosno do smanjenja brzine
rasta algi (Chisti, 2007). Što se tiče uticaja temperature na rast, pokazalo se da su mikroalge
otpornije na niže temperature (mogu preživeti i do 18 ºC niže temperature od optimalnih), a
povećanje temperature za svega 2-4 ºC je letalno (Mata et al., 2010). Mešanje je veoma važan
faktor, dovodi do ravnomerne distribucije ćelija, toplote i metabolita i ubrzava prenos mase
gasova. Rast može da inhibira smanjenje pH vrednosti usled povećanog sadržaja CO2 (Chiu
et al., 2009; De Morais i Costa, 2007).
Metabolizam mikroalgi je sličan metabolizmu viših biljaka. Naravno, nije moguće
predvideti unapred koje podloge alge mogu da koriste (Neilson i Lewin, 1974). Ključni faktor
kod heterotrofne kultivacije mikroalgi je kiseonik. Ograničenje kiseonika u kulturi može da
smanji stopu rasta i produkciju biomase. Takav je slučaj kod Chlorella-espp. (Wu i Shi,
2007).
Manipulacijom metaboličkih puteva može se preusmeriti ćelijska funkcija prema
sintezi željenih proizvoda i na taj način se mogu povećati sposobnosti mikroalgi.
Specijalizovanom kultivacijom mogu da se podstaknu promene u metabolizmu i tako se
dobije biomasa sa ciljanim metabolitom (Rosenberg et al., 2008).

15
2. Ciljevi rada
Imajući u vidu da mikoalge imaju veliki potencijal za primenu u različitim
biotehnološkim procesima, ciljevi ovog rada su:
izolacija mikroalgi iz uzorka vode sa Vlasinkog jezera,
identifikacija dobijenih izolata mikroskopkim metodama i
ispitivanje produkcije biomase mikroalgi u odnosu na različite izvore azota u
podlozi.
16
3. Materijal i metode
3.1. Materijal
3.1.1. Uzorkovanje
U ovom radu za izolaciju mikroalgi korišćeni su uzorci vode iz Vlasinskog jezera.
Uzorci su uzeti sa više lokaliteta i čuvani su u staklenim flašama. Flaše su bile izložene
svetlosti, a zatvarači nisu bili zatvoreni do kraja, dok nisu transportovane u laboratoriju.
3.1.2. Podloge
Za izolaciju, gajenje algi i pripremu inokuluma korišćena je BG11 (Blue-Green)
podloga (Tabela 1).
Tabela 1. Sastav BG11 podloge
Stok 1 (po litru) Stok 2 (po 500 ml) Stok 3 (po litru)
NaNO3 15.0 g K2HPO4 2.0 g H3BO3 2.86 g
MgSO4.7H2O 3.75 g MnCl2.4H2O 1.81 g
CaCl2.2H2O 1.80 g ZnSO4.7H2O 0.22 g
Limunska kiselina 0.30 g Na2MoO4.2H2O 0.39 g
Amonijum fericitrat 0.30 g CuSO4.5H2O 0.08g
EDTANa2 0.05 g Co(NO3)2.6H2O 0.05 g
Na2CO3 1.00 g
Podloga se pravi mešanjem 100 ml stoka 1, 10 ml stoka 2 i 1 ml stoka 3 i dodavanjem
dejonizovane vode do 1 litra. pH podloge je podešavan na 7.1 sa 1M NaOH ili HCl. Za
pravljenje čvrste podloge dodaje se bakteriološki agar 15 g po litru. Pripremljena podloga je
sterilizovana u autoklavu, 15 min 121 ºC.
Za ispitivanje uticaja azota na rast algi pored BG11 podloge korišćene su i
modifikovane BG11 podloge. U modifikovane podloge kao izvori azota dodavani su natrijum
nitrat (15 g/L) ili amonijum fericitrat (0.3 g/L). U podlozi koja je imala natrijum nitrat kao
izvor azota, dodat je feri hlorid (0.3 g/L) kao izvor gvožđa za rast algi.
2.1.3. Reagensi
Za određivanje nitrata u podlozi UV-spektrofotometrijski korišćeni su sledeći
reagensi:
1. demineralizovana voda,

17
2. osnovni rastvor nitrata, dobijen rastvaranjem 7211,9 mg osušenog kalijum-nitrata
(KNO3) i razblaživanjem do 1 litra vodom. Dobijen je rastvor koncentracije 1 ml
= 0,l mg N = 100 μg N,
3. standardni rastvor nitrata, dobijen razblaživanjem 100,0 ml osnovnog rastvora
nitrata do 1 litra vodom; dobijen rastvor koncentracije 1,00 ml = 10,0 μg N = 44,3
μg NO3,
4. hlorovodonična kiselina (HCl) 1 M,
5. suspenzija aluminijum-hidroksida; dobijena odmeravanjem 125 g
AlK(SO4)2·12H2O ili A1NH4(SO4)2. 12H2O i rastvaranjem u litru vode. Zatim je
zagrejana do 60ºC i postepeno je dodato 55 ml koncentrovanog rastvora
amonijaka, uz mešanje. Smeša je ostavljena da stoji jedan sat, zatim prebačena u
široku bocu, a talog ispran sukcesivnim dodavanjem i dekantovanjem vode (bez
sadržaja amonijaka, hlorida, nitrata i nitrite). Nakon taloženja, odekantovano je što
je moguće više bistre tečnosti i ostavljena je samo koncentrovana suspenzija.
Za određivanje koncentracije amonijumovog jona u podlozi sprektrofotometrijski
korišćeni su sledeći reagensi:
1. rastvor amonijum-hlorida dobijen je rastvaranjem 0,0314 g NH4Cl u 100 ml
destilovane vode (0,1 mg NH3/cm3)
2. neslerov reagens
3. rastvor sumporne kiseline, 0,05M dobijen rastvaranjem 2,7 cm3 H2SO4 u 1000 ml
destilovane vode.
3.2. Metode rada
3.2.1. Izolacija algi
Za izolaciju algi korišćene su sledeće metode:
- metoda iscrpljivanja eze (metoda poteza) – Ezom je zahvaćena mala količina početnog
uzorka (10μl) koja je prenešena na jedan deo podloge. Zatim je nanešena kap ezom
prevlačena i na druge delove podloge, tzv. striklovanje.
- metoda razlivanja kapi – Određena količina uzorka (500 μl) je prenešena na podlogu i
razlivena po površini podloge štapićem po Drigalskom.

18
Nakon zasejavanja podloga, Petri ploče su stavljene na inkubaciju u dobro osvetljenoj
prostoriji na sobnoj temperaturi 14 dana. Pojedinačne kolonije algi sa Petri ploča su dalje
prečišćavane prebacivanjem na nove Petri ploče do dobijanja uniformnih kolonija.
3.2.2. Identifikacija algi
Identifikacija algi rađena je na osnovu morfoloških karakteristika. Od kolonija sa Petri
ploča napravljeni su nativni preparati koji su posmatrani svetlosnnim mikroskopom na
uvećanju od 40 x (Leica DM 1000).
3.2.3. Zasejavanje podloga
Priprema inokuluma rađena je zasejavanjem BG11 tečne podloge kolonijama uzetim
sa Petri ploča. Zasejana podloga inkubirana je na sobnoj temperaturi (oko 28 ºC) 15 dana, uz
konstantno prisustvo svetlosti i mešanja na 210 obrtaja u minuti (PSU-20i, Multi-functional
Orbital Shaker).
Određivanje uticaja izvora azota na rast algi rađeno je u 3 podloge:
modifikovana BG11 podloga bez natrijum nitrata, ali sa amonijum fericitratom kao
izvorom azota (1),
modifikovana BG11 podloga bez amonijum fericitrata, ali sa natrijum nitratom kao
izvorom azota (2) i
BG11 podloga koja jej sadržala natrijum nitrat i amonijum fericitrat kako izvore azota
(3).
Podloge su zasejavane 2% inokulumom algi, inkubirane su na sobnoj temperaturi
(oko 28 ºC), uz konstantno prisustvo svetlosti i mešanjem na 210 obrtaja u minuti.
3.2.4. Određivanje biomase algi
Rast algi je praćen u periodu od 16 dana, uzimanjem uzoraka za određivanje biomase
na svakih 2 dana. Nagrađena biomasa algi je određivana spektrofotometrijski merenjem
apsorbance uzoraka na 620 nm.
Na kraju završenog perioda gajenja algi (30 dana), rađeno je gravimetrijsko
određivanje biomase. 10 ml podloge filtrirano je pod vakuumom i sušeno na filter papiru do
konstantne mase. Biomasa algi odrađena je kao razlika mase filter papira sa algama i filter
papira bez algi i izražena je kao masa mikroalgi/L/danu.
3.2.5. Određivanje koncentracije nitrata
Određivanje koncentracije nitrata u uzorcima podloga rađeno je UV-
spektrofotometrijski merenjem apsorbance uzoraka na 220 nm.

19
Najpre su pripremljeni kalibracioni standardi rastvora nitrata u opsegu od 0 do 8 mg
nitrata po litru i to razblaženjem do 50 ml sledećih zapremina standardnog rastvora nitrata: 0;
1,00; 2,00; 4,00; 6,00 i 8,00 ml. Pošto su ispitivani uzorci prilikom određivanja nitrata
razblaživani 1000 puta, obojenje uzorka je tako izbegnuto, a samim tim i postupak uklanjanja
obojenja koji se sastoji u dodavanju suspenzije Al(OH)3 i ceđenju kroz membran-filtar od
0,45 μm, kao i naknadno zakišeljavanje sa HCl.Očitana je apsorbanca svih standardnih
rastvora i uzoraka u odnosu na slepu probu (demineralizovana voda) na 220 i 275 nm.
3.2.6. Određivanje koncentracije amonijumove soli
Određivanje koncentracije amonijumove soli rađeno je Neslerovim reagensom tako
što se uzorak zakiseli pomoću H2SO4. Stvoreni amonijum-sulfat se određuje spektrofoto-
metrijski, jer uzorci koji sadrže amonijumov jon sa Neslerovim reagensom razvijaju žuto
obojenje, čiji intenzitet boje zavisi od količine amonijaka ili amonijumove soli u uzorku.
Najpre je napravljen radni rastvor NH4Cl odmeravanjem 10 ml primarnog rastvora i
rastvaranjem u normalnom sudu od 100 ml, pri čemu je dobijen rastvor koncentracije 0,01
mg NH3/ml. Od tog rastvora napravljena je standardna serija rastvora u normalnim sudovima
od 50 ml odmeravanjem 1, 2, 4, 6, 8 i 10 ml. Dobijeni rastvori su u opsegu koncentracija: 0,2;
0,4; 0,8; 1,2; 1,6 i 2 mg/L. Standardnim rastvorima je zatim dodato 10 ml 0,05 M H2SO4 i 2
ml Neslerovog reagensa, i nakon 15 minuta izmerena je apsorbanca rastvora na 425 nm.
Priprema slepe probe je izvršena tako što je u normalni sud od 50 ml odmereno 10 ml 0,05 M
H2SO4 i dodato 2 ml Neslerovog reagensa, a zatim je normalni sud dopunnjen do crte
destilovanom vodom. Uzorci su najpre razblaženi 100 puta, a onda je kao i kod pripreme
standardnih rastvora dodato 10 ml 0,05 M H2SO4 i 2 ml Neslerovog reagensa, i nakon 15
minuta izmerena je apsorbanca rastvora na 425 nm. Na osnovu konstruisanog kalibracionog
grafika, nanošenjem vrednosti apsorbance na ordinatu i koncentracije amonijumove soli na
apscisu, očitane su vrednosti koncentracije amonijumovog jona u ispitivanim uzorcima.

20
4. Rezultati i diskusija
4.1. Izolacija algi iz Vlasinskog jezera
Alge su veoma heterogena grupa organizama sa različitim morfološkim i fiziološkim
karakteristikama. Posledica ovako velikog diverziteta je heterogenost različitih vrsta algi u
odnosu na uslove njihovog rasta. Zato je lokacija sa koje se alge izoluju ključni faktor za
selekciju sojeva mikroalgi koje se mogu koristiti za produkciju biomase (Lee at al., 2014). U
ovom radu, mikroalge su izolovane iz uzoraka vode uzetih sa površine Vlasinskog jezera.
Vlasinsko jezero izabrano je kao lokalitet za izolaciju algi jer se nalazi na nadmorskoj visini
odod 1 204 m i predstavlja sredinu sa minimalnim uticajem čoveka. Korišćenim metodama
izolacije koje su opisane u delu Metode, dobijen je jedan izolat mikroalgi koji je formirao
kolonije zelene boje (Slika 11). S obzirom da je uzorak vode sa Vlasinskog jezera uzet
početkom aprila, kada su temperature bile niske tako da je tek krenuo vegetacioni ciklus
fitoplanktona, uočena je samo jedna vrsta kolonija pilikom prvog prebacivanja uzorka vode
na Petri ploče. Ovo ukazuje na mali broj mikroalgi u uzorku vode sa Vlasinskog jezera u
ovom periodu godine.
Slika 11. Kolonije mikroalge na Petri ploči
4.2. Identifikacija mikroalge
Slika nativnog preparata izolovane mikroalge dobijena svetlosnim mikroskopom pod
uvećanjem od 40 x prikazana je na Slici 12. Na osnovu morfoloških karakteristika, izolat
mikroalge identifikovan je kao predstavnik roda Closterium. Rod Closterium pripada razdelu
Chlorophyta. Vrste ovog roda su jednoćelijske alge koje imaju kokoidni tip morfološke

21
organizacije. Ćelije su izdužene, polumesečasto savijene i zašiljene na oba kraja. Izgrađene
su od dve simetrične polovine (polućelije) između kojih se nalazi suženje poznato kao istmus.
Dominantna grupa pigmenata su hlorofili a i b koji su smešteni u pločastim hloroplastima sa
pirenoidima.
Slika 12. Nativni preparat izolata mikroalge
Mikroalge iz roda Closterium su slatkovodne i najviše se razvijaju u oligotrofnim
vodama kao što su bare ali su izolovane iz različitih jezera (Descy et al., 1994).
4.3. Rast mikroalge u zavisnosti od izvora azota
Esencijalni elementi za rast algi su N, P, K, Ca, Mg, S, Fe, Cu, Zn i Mn koji se dodaju
u podloge u obliku soli. Koncentracija ovih elemenata varira od podloge do podloge
(Ilavarasi et al., 2011). Azot čini 1-10% suve mase mikroalgi i utiče na sastav ćelija i
metabolizam algi što ga čini jednim od najvažnijih elemenata (Perez-Garcia et al., 2010). Kao
izvor azota se najčešće koriste nitrati, amonijumove soli ili urea (Danilović et al., 2013).
U ovom radu je ispitivan rast mikroalge izolovane iz Vlasinskog jezera u tri podloge
sa različitim izvorima azota. Korišćena je BG11 podloga sa natrijum nitratom i amonijum
fericitratom kao izvorima azota, kao i modifikovane BG11 podloge koje sadrže samo
natrijum nitrat ili amonijum fericitrat. Rast mikroalge u korišćenim podlogama praćen je
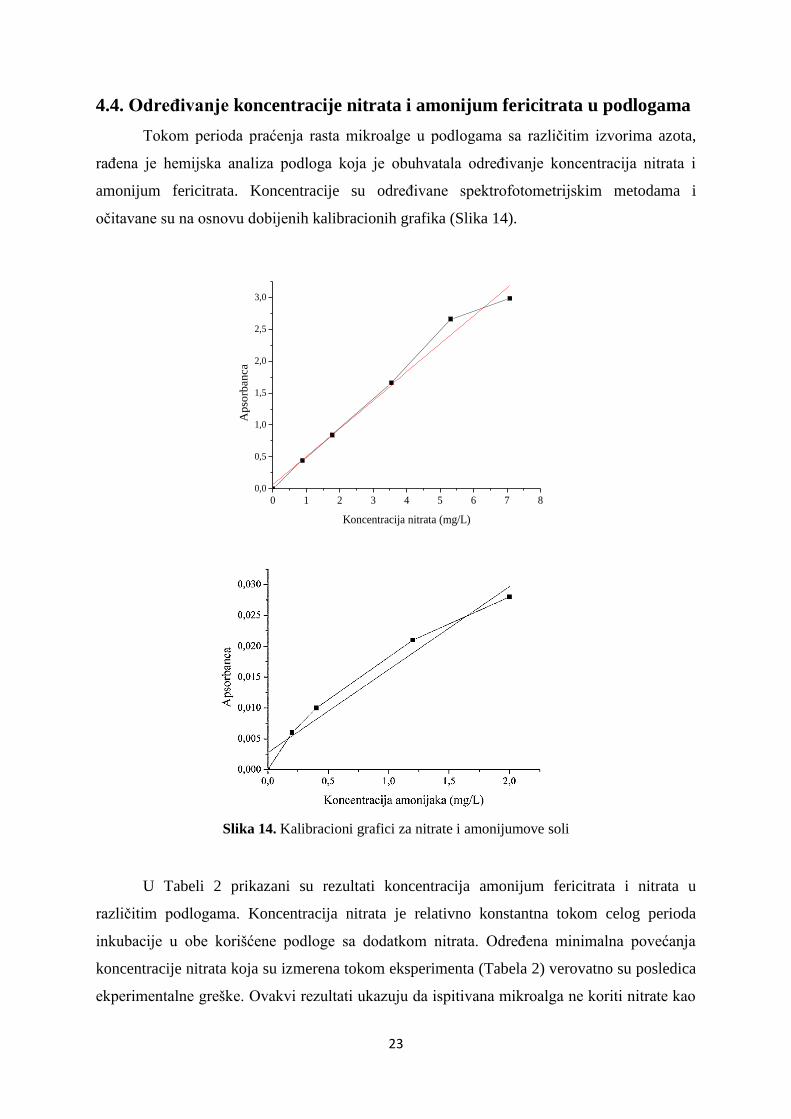
spektrofotometrijski u periodu od 16 dana (Slika 13).

22
Slika 13. Rast mikroalge u podlogama sa različitim izvorima azota
(1 – podloga sa amonijum fericitratom, 2 – podloga sa natrijum nitratom, 3 – podloga sa amonijum
fericitratom i natrijum nitratom)
U svim podlogama sa različitim izvorom azota zapažen je rast mikroalge. U
podlogama sa amonijum fericitratom i mešavinom amonijum fericitrata i natrijum nitrata,
mikroalga ulazi u eksponencijalnu fazu rasta nakon osam dana inkubacije. Na kraju praćenog
perioda produkcije biomase (16 dana), veći rast mikroalge je zabeležen u podlozi sa
amonijum fericitratom (A620 0,25) u odnosu na podlogu sa dva izvora azota (A620 0,14). U
podlozi u kojoj je natrijum nitrat bio jedini izvor azota, rast mikroalge je bio najsporiji.
Početak eksponencijalne faze je tek nakon 14 dana od početka inkubacije, dok je nakon 16
dana vrednost apsorbance jako niska (A620 0,036).
Pretraživanjem literature nisu pronađeni podaci koji bi se odnosili na rast mikroalgi iz
roda Closterium u zavisnosti od izvora azota u podlozi. Rezultati dobijeni za druge zelene
mikroalge su u suprotnosti sa prikazanim rezultatima u ovom radu. Tako je pokazano da
mikroalga Scenedesmus bijugatus najbolju produkciju biomase ima kada su izvori azota
kalijum nitrat i natrijum nitrat u odnosu na ureu, kalcijum nitrat, amonijum nitrat i amonijum
hlorid (Arumugam et al., 2013). Takođe, urea i nitrati su se pokazali kao bolji izvori azota u
odnosu na amonijumove soli za rast mikroalgi iz rodova Chlorella i Neochloris (Converti et
al., 2009; Li et al., 2008). Sa druge strane morska alga iz roda Ellipsoidion je imala bolji rast
na podlozi sa amonijumovim solima u odnosu na podloge sa dodatkom nitrata i uree (Xu et
al., 2001).
0
0.05
0.1
0.15
0.2
0.25
2. 4. 6. 8. 10. 12. 14. 16.
Ap
so
rba
nca
Vreme (dani)
1
2
3
23
4.4. Određivanje koncentracije nitrata i amonijum fericitrata u podlogama
Tokom perioda praćenja rasta mikroalge u podlogama sa različitim izvorima azota,
rađena je hemijska analiza podloga koja je obuhvatala određivanje koncentracija nitrata i
amonijum fericitrata. Koncentracije su određivane spektrofotometrijskim metodama i
očitavane su na osnovu dobijenih kalibracionih grafika (Slika 14).
0 1 2 3 4 5 6 7 8
0,0
0,5
1,0
1,5
2,0
2,5
3,0
Ap
sorb
an
ca
Koncentracija nitrata (mg/L)
Slika 14. Kalibracioni grafici za nitrate i amonijumove soli
U Tabeli 2 prikazani su rezultati koncentracija amonijum fericitrata i nitrata u
različitim podlogama. Koncentracija nitrata je relativno konstantna tokom celog perioda
inkubacije u obe korišćene podloge sa dodatkom nitrata. Određena minimalna povećanja
koncentracije nitrata koja su izmerena tokom eksperimenta (Tabela 2) verovatno su posledica
ekperimentalne greške. Ovakvi rezultati ukazuju da ispitivana mikroalga ne koriti nitrate kao

24
samostalne izvore azota ni u kombinaciji sa amonijumovom soli. Jedan od mogućih razloga
zbog koga mikroalga korišćena u ovom radu ne može da koristi nitrate kao izvore azota za
rast je koncentracija natrijum nitrata u podlozi koja je visoka. Arumugam i saradnici (2013)
pokazali su da koncentracije nitrata u podlozi veće od 10 mM dovode do slabe produkcije
biomase mikroalgi, verovatno zbog štetnog delovanja azota na njihove ćelije što dovodi do
prestanka rasta ćelija.
Tabela 2. Koncentracije amonijum fericitrata i nitrata u različitim podlogama (1 – podloga sa
amonijum fericitratom, 2 – podloga sa natrijum nitratom, 3 – podloga sa amonijum fericitratom i
natrijum nitratom)
Dan Amonijum fericitrat (mg/L) Nitrati (mg/L)
1 3 2 3
2. 4,643 5,159 106,3 116,2
4. 4,006 4,876 115,1 113,3
6. 3,560 3,658 112,1 111,5
8. 3,268 3,071 109,2 121,0
10. 1,944 2,742 119,6 122,1
12. 0 2,136 129,2 135,7
14. 0 2,388 120,5 126,9
16. 0 0 119,0 122,1
Koncentracija amonijum fericitrata u praćenom periodu inkubacije opada kada se
samo amonijumov jon koristi kao izvor azota, kao i kada je u kombinaciji sa nitratima. Na
osnovu ovoga se može zaključiti da mikroalga koristi amonijumov jon kao izvor azota. U
podlozi gde je NH4+ jedini izvor azota, njegova koncentracija opada jako brzo tako da već
nakon 10 dana inkubacije u podlozi nema više amonijum fericitrata. Prisustvo nitrata
smanjuje unos NH4+ pa je brzina smanjenja njegove koncentracije u podlozi sa nitratima i
amonijum fericitratom manja u odnosu na podlogu gde je prisutan samo amonijum fericitrat
(Slika 15). Ukoliko se u podlozi nalaze mešoviti izvori azota, amonijumove soli se obično
prvo koriste za rast mikroalgi, a tek u kasnijim fazama gajenja nitrati ili urea (Eustanace et
al., 2013). Osim toga, u ovom radu je korišćen amonijum fericitrat kao izvor NH4+ koji je u
isto vreme i izvor gvožđa neophodnog za sintezu hlorofila, a samim tim i procesa fotosinteze.

25
Slika 15. Koncentracija amonijum fericitrata tokom gajenja mikroalge (1 – podloga sa amonijum
fericitratom, 3 – podloga sa amonijum fericitratom i natrijum nitratom)
Dobijeni rezultati za koncentraciju amonijum fericitrata i nitrata u podlogama se
poklapaju sa rezultatima produkcije biomase. Na ovaj način je potvrđeno da je rast mikroalge
u korelaciji sa unosom i korišćenjem NH4+ a da mikroalga praktično ne usvaja nitrate u
praćenom periodu gajenja.
4.5. Gravimetrijsko određivanje biomase mikroalge
Nakon mesec dana gajenja mikroalge na rotacionoj mućkalici, dobijen je različit rast u
korišćenim podlogama (Slika 16).
Slika 16. Erlenmajeri u kojima je gajena mikroalga na različitim podlogama
Produkovana biomasa je određivana gravimetrijski i rezultati su prikazani u Tabeli 3.
Najviše biomase produkovano je prilikom rasta mikroalge u podlozi sa mešavinom izvora
azota (natrijum nitrat i amonijum fericitrat). Nakon mesec dana gajenja u ovoj podlozi
proizvedeno je 0,123 g/L/d biomase. U ostalim podlogama produkovna biomasa je bila
značajno manja (Tabela 3). Iako je ispitivana mikroalga imala bolji rast u prvih 16 dana na
0
1
2
3
4
5
6
2. 4. 6. 8. 10. 12. 14. 16.Ko
nce
ntr
acija
(mg
/L)
Vreme (dani)
1
3

26
podlozi sa amonijum fericitratom u odnosu na podlogu sa mešavinom izvora azota, nakon
mesec dana gajenja, prinos biomase je na ovoj podlozi manji. Jedan od problema prilikom
korišćenja NH4+ kao izvora azota jeste smanjenje pH vrednosti podloge što može dovesti do
prestanka rasta ćelija (Eustance et al., 2013). Smanjenje pH vrednosti podloge dešava se zbog
unosa NH4+ katjona u ćeliju tako da ćelija izbacuje protone u spoljašnju sredinu da bi se
održala neutralnost unutar ćelija.
Tabela 3. Produktivnost biomase mikroalge u različitim podlogama (1– podloga sa amonijum
fericitratom, 2– podloga sa natrijum nitratom, 3– podloga sa amonijum fericitratom i natrijum
nitratom)
Produktivnost
biomase (g/L/d)
1 0.011
2 0,020
3 0.123
Da bi se određeni produkti ekstrahovali iz mikroalgi, potrebno je dobiti veliku
količinu biomase mikroalgi što je veoma često problem pri njihovom korišćenju u
biotehnologiji. Prinos biomase mikroalgi je različit i pre svega zavisi od korišćenog soja
mikroalge, kao i od optimizacije samog procesa produkcije odnosno sastava podloge i uslova
gajenja. U literaturi nisu pronađeni podaci koji bi se odnosili na produktivnost biomase vrsta
iz roda Closterium, dok je produktivnost kod ostalih mikroalgi od 0,04 do 4,1 g/L/d
(Danilović et al., 2014).

27
5. Zaključci
Na osnovu prikazanih rezultata mogu se doneti sledeći zaključci:
1. Iz uzoraka vode sa Vlasinskog jezera izolovan je jedan izolat mikroalge koji je
identifikovan kao predstavnik roda Closterium;
2. Mikroalga je pokazala dobar rast na podlozi sa amonijum fericitratom kao jedinim
izvorom azota, kao i na podlozi sa mešovitim izvorima azota, amonijum fericitratom i
natrijum nitratom;
3. Hemijske analize uzoraka podloga na kojima je rasla mikroalga pokazuju da
mikroalga ne usvaja nitrate iz podloga za rast već da koristi amonijumov jon kao izvor
azota;
4. Prinos biomase nakon mesec dana gajenja mikroalge bio je najveći na podlozi koja je
sadržala amonijum fericitrat i natrijum nitrat.

28
6. Literatura
Altin, R., Cetinkaya, S., Yucesu, H. S., 2001: The potential of using vegetable oil fiels
as fuel for diesel engines. – Energy Conversion and Menagement 42, 529-538.
Anonimus: http://biodiesel.org/production/production-statistics
Arumugam, M.,Agarwal, A., Arya M. C., Ahmed, Z., 2013:Influence of nitrogen
sources on biomass productivity of microalgae Scenedesmus bijugatus. -Bioresource
Technology 131, 246–249.
Blaženčić, J., 2000: Sistematika algi. – NNK Internacional, Beograd.
Cardozo, K. H., Guaratini, T., Barros, M. P., Falcao, V. R., Tonon, A. P., Lopes, N.
P., 2007: Metabolites from algae with economical impact. – Com. Biochem. Physiol.,
C 146, 60-78.
Chen, P. et al., 2009: Review of the biological and engineering aspects of algae to
fuels approach. - Int J Agric & Biol Eng 2, 4.
Chisti, Y., 2007: Biodiesel from microalgae. – Biotechnol. Adv. 25, 294-306.
Chiu, S. Y., Kao, C. Y., Tsai M. T., Ong, S. C., Chen, C. H., Lin, C. S., 2009: Lipid
accumulation and CO2 utilization of Nannochloropsis oculata in response to CO2
aeration. - Bioresource Technol. 100, 833–838.
Converti, A., Casazza, A. A., Ortiz, E. Y., Perego, P., Del Borghi, M., 2009: Effect of
temperature and nitrogen concentration on the growth and lipid content of
Nannochloropsis oculata and Chlorellavulgaris for biodiesel production. - Chem.
Eng. Proc. 48, 1146–1151.
Danilović, B. R., Avramović, J. M., Ćirić, J. T., Savić, D. S., Veljković, V. B., 2013:
Proizvodnja biodizela iz ulja mikroalgi. – Savez hemijskih inženjera Srbije 68, 213-
232.
De Morais, M. G., Costa J. A. V., 2007: Carbon dioxide fixation by Chlorella
kessleri, C. vulgaris, Scenedesmus obliquus and Spirulina sp. cultivated in flasks and
vertical tubular photobioreactors. - Biotechnol. Lett. 29, 1349–1352.
Descy, J. P., Reynolds, C. S., Padisák, J., 1994: Phytoplankton in Turbid
Environments: Rivers and Shallow Lakes. - Springer Science & Business Media 100,
2013.
Duong, V. T., Li, Y., Nowak, E., Schenk, P. M., 2012: Microalgae Isolation and
Selection for Prospective Biodiesel Production. – Energies 5, 1835-1849.
Eustance, E., Gardner, R. D., Moll, K. M., Menicucci, J., Gerlach, R., Peyton, B. M.,
2013: Growth, nitrogen utilization and biodiesel potential for two chlorophytes grown
on ammonium, nitrate or urea. - Journal of Applied Phycology 25, 1663–1677.
Hossain, A.B., Salleh, A., Boyce, A. N., Chowdhury, P., Naqiuddin, M., 2008:
Biodiesel Fuel Production from algae as renewable energy. – Am J Biochem.
Biotechnol 4, 250-254.
Hundt, K., Reddy, B. V., 2011: Algal biodiesel production from power plant axhaust
and its potencial to replace petrodiesel and reduce greenhouse gas emissions. – Int. J.
Low-Carbon Technol. 6, 294-298.

29
Kurano, N., Ikemoto, H., Miyashita, H., Hasegawa, T., Hata, H., Miyachi, S., 1996:
Fixation and utilization of carbon dioxide by microalgal photosynthesis. - Energy
Conver Manage 36, 689-92.
Lee, K., Eisterhold, M. L., Rindi, F., Palanisami, S., Nam, P. K., 2014: Isolation and
screening of microalgae from natural habitats in the Midwestern United States of
America for biomass and biodiesel sources. – Journal of Natural Science, Biology and
Medicine 5, 332-339.
Li, Y, Horsman, M., Wang, B., Wu, N., Lan, C. Q., 2008: Effects of nitrogen sources
on cell growth and lipid accumulation of green alga Neochloris oleoabundans. – Appl.
Microbiol. Biotechnol. 81, 629-36.
Liu, Z. Y., Wang, G. C., Zhou, B. C., 2008: Effect of iron on growth and lipid
accumulation in Chlorella vulgaris. - Bioresour. Technol. 99, 4717-4722.
Mata, T., Martins, A., Caetaano, N., 2010: Microalgae for bidiesel production and
other applications. – A review, Renew. Sustain. Energ. Rev. 14, 217-232.
Mihailov-Krstev, T., 2008: Praktikum iz algologije i mikologije. – Prirodno-
matematički fakultet, Niš.
Muñoz, R. B. G., 2006: Water Research 40, 2799-2815.
Neilson, A.H., Lewin, R.A., 1974: The uptake and utilization of organic carbon by
algae: an essay in comparative biochemistry. - Phycologia 13, 227-264.
Neltner, B., 2008: Algae Based Biodiesel. – Algae 2, 5.
Perez-Garcia, O., Escalante, F. M. E., De-Bashan, L.E., Bashan, Y., 2010:
Heterotrophic cultures of microalgae: Metabolism and potencial products. – Elsevier
45, 11-36.
Rosenberg, J. N., Oyler, G. A., Wilkinson, L., Betenbaugh, M. J., 2008: A green light
for engineered algae: redirecting metabolism to fuel a biotechnology revolution. –
Current Opinion in Biotechnology 19, 430-436.
Sakai, N., Sakamoto, Y., Kishimoto, N., Chihara, M., Karube, I., 1995: Chlorella
strains from hot springs tolerant to high temperature and high CO2. – Energy Conver
Manage 36, 693-6.
Scott, S. A. et al., 2010: Biodiesel from algae: challenges and prospects. – Current
Opinion in Biotechnology 21, 277-286.
Sheehan, J., Dunahay, T., Benemann, J., Roessler, P., 1998: A Look Back at the U.S.
Deparment of Energy´s Aquatic Species Program: Biodiesel from microalgae. –
National Renewable Energy Laboratory: Golden, Colorado, USA.
Tešić, Ž., Marinović, R., 1966: Sistematika nižih biljaka, I deo. – Zavod za izdavanje
udžbenika SR Srbije, Beograd.
Wang, B., Li, Y., Wu, N., Lan, C. Q., 2008: CO2 bio-mitigation using microalgae. –
Appl. Microbiol. Biotechnol. 79, 707-718.
Wu, Z., Shi, X., 2007: Optimization for high-density cultivation of heterotrophic
Chlorella based on a hybrid neural network model. - Lett. Appl. Microbiol. 44, 13-18.
Xu, N., Zhang, X., Fan, X., Han, L., Zeng, C., 2001: Effects of nitrogen source and
concentration on growth rate and fatty acid composition of Ellipsoidion sp.
(Eustigmatophyta). - J. Appl. Phycol. 13, 463–469.

30
Yin, N. C., Yaakob, Z., Ali, E., Min, A., Wa, N. S., 2011: Characterization of Various
Microalgae for Biodiesel Fuel Production. – Journal of Materials Science and
Engeineering A 1, 80-86.
Slike algi su preuzete sa sledećih linkova:
http://cyanobacteria.myspecies.info/taxonomy/term/262
https://www.google.rs/search?q=microcystis&espv=2&biw=1366&bih=662&source=lnms&t
bm=isch&sa=X&ved=0ahUKEwisyNvcwefPAhVkLsAKHV0iBTgQ_AUIBigB&dpr=1#tbm
=isch&q=oscillatoria&imgrc=4dapg8x0QYjEqM%3A
http://algae-lab.com/product/spirulina-maxima-live-algae-2/
http://protist.i.hosei.ac.jp/PDB/Images/Prokaryotes/Oscillatoriaceae/Lyngbya/sp_01.jpg
https://www.google.rs/search?q=microcystis&espv=2&biw=1366&bih=662&source=lnms&t
bm=isch&sa=X&ved=0ahUKEwisyNvcwefPAhVkLsAKHV0iBTgQ_AUIBigB&dpr=1#tbm
=isch&q=anabaena&imgrc=Qye_8yU-KxXz_M%3A
https://www.google.rs/search?q=nostoc&espv=2&biw=1366&bih=662&site=webhp&source
=lnms&tbm=isch&sa=X&ved=0ahUKEwjDjsvow-
fPAhXMJ8AKHUeVClsQ_AUIBigB#imgrc=7HD2eGkxzj28BM%3A
https://www.google.rs/search?q=nostoc&espv=2&biw=1366&bih=662&site=webhp&source
=lnms&tbm=isch&sa=X&ved=0ahUKEwjDjsvow-
fPAhXMJ8AKHUeVClsQ_AUIBigB#tbm=isch&q=euglenophyte&imgrc=7jZJ827cnJ1UhM
%3A
https://www.google.rs/search?q=phacus&source=lnms&tbm=isch&sa=X&ved=0ahUKEwiP
_JHKxOfPAhUBuhQKHZd2Da8Q_AUICCgB&biw=1366&bih=662#imgdii=awujxP4Nj9K
4EM%3A%3BawujxP4Nj9K4EM%3A%3BK9yfyc8vAJR6rM%3A&imgrc=awujxP4Nj9K4
EM%3A
https://www.google.rs/search?q=phacus&source=lnms&tbm=isch&sa=X&ved=0ahUKEwiP
_JHKxOfPAhUBuhQKHZd2Da8Q_AUICCgB&biw=1366&bih=662#tbm=isch&q=trachelo
monas&imgrc=D4_0IESY2_FlxM%3A
https://www.google.rs/search?q=phacus&source=lnms&tbm=isch&sa=X&ved=0ahUKEwiP
_JHKxOfPAhUBuhQKHZd2Da8Q_AUICCgB&biw=1366&bih=662#tbm=isch&q=peridini
um&imgrc=YyTZktjd0iixQM%3A
https://www.google.rs/search?q=phacus&source=lnms&tbm=isch&sa=X&ved=0ahUKEwiP
_JHKxOfPAhUBuhQKHZd2Da8Q_AUICCgB&biw=1366&bih=662#tbm=isch&q=cryptom
onas&imgdii=giyD-jpo2JHiLM%3A%3BgiyD-
jpo2JHiLM%3A%3B5Wve2UqS_0OPYM%3A&imgrc=giyD-jpo2JHiLM%3A

31
https://www.google.rs/search?q=phacus&source=lnms&tbm=isch&sa=X&ved=0ahUKEwiP
_JHKxOfPAhUBuhQKHZd2Da8Q_AUICCgB&biw=1366&bih=662#tbm=isch&q=gymnodi
nium&imgrc=BQtvdZzKXUCq9M%3A
https://www.google.rs/search?q=phacus&source=lnms&tbm=isch&sa=X&ved=0ahUKEwiP
_JHKxOfPAhUBuhQKHZd2Da8Q_AUICCgB&biw=1366&bih=662#tbm=isch&q=melosira
&imgrc=IenIudzYNogCAM%3A
https://www.google.rs/search?q=phacus&source=lnms&tbm=isch&sa=X&ved=0ahUKEwiP
_JHKxOfPAhUBuhQKHZd2Da8Q_AUICCgB&biw=1366&bih=662#tbm=isch&q=cocconei
s&imgrc=1qSmqNRJ-6Je5M%3A
https://www.google.rs/search?q=phacus&source=lnms&tbm=isch&sa=X&ved=0ahUKEwiP
_JHKxOfPAhUBuhQKHZd2Da8Q_AUICCgB&biw=1366&bih=662#tbm=isch&q=navicula
+&imgrc=4g3zs4JLbrH8RM%3A
https://www.google.rs/search?q=cymbella&biw=1366&bih=662&tbm=isch&tbo=u&source=
univ&sa=X&ved=0ahUKEwjThI7Py-
fPAhUDLcAKHbbaAIgQsAQIHA#imgrc=LozSRM61c51A4M%3A
https://www.google.rs/search?q=bangia&biw=1366&bih=662&source=lnms&tbm=isch&sa=
X&ved=0ahUKEwiApKSCzOfPAhUFC8AKHTe0ArQQ_AUIBygC#imgrc=7lq8oOK7en_L
PM%3A
https://www.google.rs/search?q=porphyra&biw=1366&bih=662&source=lnms&tbm=isch&s
a=X&ved=0ahUKEwjb5rTQzOfPAhUsIcAKHRTRAxEQ_AUIBigB#imgrc=ot47rb3HdM5
ReM%3A
https://www.google.rs/search?q=batrachospermum&biw=1366&bih=662&tbm=isch&tbo=u
&source=univ&sa=X&ved=0ahUKEwio4seAzefPAhWLI8AKHSbjB5EQsAQIGQ#imgrc=C
YEVBkY9ibHSCM%3A
https://www.google.rs/search?q=tribonema&biw=1366&bih=662&tbm=isch&tbo=u&source
=univ&sa=X&ved=0ahUKEwis16aszefPAhXGBsAKHU5rBigQsAQIGQ#imgrc=WyuhTtU
M7aDDyM%3A
https://www.google.rs/search?q=vaucheria&biw=1366&bih=662&source=lnms&tbm=isch&
sa=X&ved=0ahUKEwi6r9PGzefPAhVBCsAKHaLpBZ0Q_AUIBigB#imgrc=_WCbudsA44
1fVM%3A
http://protist.i.hosei.ac.jp/PDB/images/Heterokontophyta/Dinobryon/sertularia/sp_20.jpg
https://www.google.rs/search?q=chrysamoeba&biw=1366&bih=662&source=lnms&tbm=isc
h&sa=X&ved=0ahUKEwi9x-K8zufPAhVBJcAKHaXSB8YQ_AUIBigB#imgrc=8fbysVR-
wTyXBM%3A
https://www.google.rs/search?q=hydrurus&biw=1366&bih=662&source=lnms&tbm=isch&s
a=X&ved=0ahUKEwi8w_vQz-
fPAhVpIMAKHfVYBMwQ_AUIBigB#imgrc=Uw429BlZUpYvNM%3A

32
https://www.google.rs/search?q=chlamydomonas&biw=1366&bih=662&source=lnms&tbm=
isch&sa=X&ved=0ahUKEwjXntzyz-
fPAhXnLsAKHeWJDo4Q_AUIBigB#imgrc=iEbNZWsIvBOTOM%3A
https://www.google.rs/search?q=chlamydomonas&biw=1366&bih=662&source=lnms&tbm=
isch&sa=X&ved=0ahUKEwjXntzyz-
fPAhXnLsAKHeWJDo4Q_AUIBigB#tbm=isch&q=scenedesmus&imgrc=yYOg4m6-
RgxA0M%3A
https://www.google.rs/search?q=chlamydomonas&biw=1366&bih=662&source=lnms&tbm=
isch&sa=X&ved=0ahUKEwjXntzyz-
fPAhXnLsAKHeWJDo4Q_AUIBigB#tbm=isch&q=volvox&imgrc=TVsiPJj5NVbDyM%3A
https://www.google.rs/search?q=chlamydomonas&biw=1366&bih=662&source=lnms&tbm=
isch&sa=X&ved=0ahUKEwjXntzyz-
fPAhXnLsAKHeWJDo4Q_AUIBigB#tbm=isch&q=ulothrix&imgrc=QzAMjysSoRjEtM%3
A
https://www.google.rs/search?q=chlamydomonas&biw=1366&bih=662&source=lnms&tbm=
isch&sa=X&ved=0ahUKEwjXntzyz-
fPAhXnLsAKHeWJDo4Q_AUIBigB#tbm=isch&q=spirogyra&imgrc=-
PXQB35z7Ty6vM%3A
https://www.google.rs/search?q=chlamydomonas&biw=1366&bih=662&source=lnms&tbm=
isch&sa=X&ved=0ahUKEwjXntzyz-
fPAhXnLsAKHeWJDo4Q_AUIBigB#tbm=isch&q=chlorella&imgrc=z6-
SmqRa45Ab_M%3A
https://www.google.rs/search?q=chlamydomonas&biw=1366&bih=662&source=lnms&tbm=
isch&sa=X&ved=0ahUKEwjXntzyz-
fPAhXnLsAKHeWJDo4Q_AUIBigB#tbm=isch&q=cosmarium&imgrc=TxPb-
Uky50w7lM%3A
https://www.google.rs/search?q=chlamydomonas&biw=1366&bih=662&source=lnms&tbm=
isch&sa=X&ved=0ahUKEwjXntzyz-
fPAhXnLsAKHeWJDo4Q_AUIBigB#tbm=isch&q=pediastrum&imgrc=b47jff34En1idM%3
A
https://www.google.rs/search?q=chara&espv=2&biw=1366&bih=662&source=lnms&tbm=is
ch&sa=X&ved=0ahUKEwjl-O_e0efPAhUHAsAKHdHZD-
IQ_AUIBigB#tbm=isch&q=chara+algae&imgrc=O5YCgAY8RT17FM%3A
https://www.google.rs/search?q=chara&espv=2&biw=1366&bih=662&source=lnms&tbm=is
ch&sa=X&ved=0ahUKEwjl-O_e0efPAhUHAsAKHdHZD-
IQ_AUIBigB#q=nitella&tbm=isch&tbs=rimg%3ACarEFfjCOubxIjj1EDURz9aELVSsegbo
KP3jAbMcccT4YTObUjeTg6RZWGG5px-
HNWl8s89yBD4RIM1NwhveMCfoASoSCfUQNRHP1oQtESxVIZpiiYjMKhIJVKx6Bugo_
1eMRi7jaUPmuO9wqEgkBsxxxxPhhMxE1hZrViWRhqyoSCZtSN5ODpFlYEb6qEXV-
IQZVKhIJYbmnH4c1aXwRf7PMOo7RW34qEgmzz3IEPhEgzREZ-
oT9oFFPwyoSCU3CG94wJ-gBEesZA8-i9Bm_1&imgrc=Tl-KL3JNRrHxjM%3A

33
https://www.google.rs/search?q=fucus&espv=2&biw=1366&bih=662&tbm=isch&tbo=u&so
urce=univ&sa=X&ved=0ahUKEwioitj10ufPAhXFLMAKHRgiC28QsAQIJw#imgrc=GsGIO
XfhqtROzM%3A
https://www.google.rs/search?q=laminaria&espv=2&biw=1366&bih=662&source=lnms&tb
m=isch&sa=X&ved=0ahUKEwiIoJTI0-
fPAhVILcAKHdenDBUQ_AUIBigB#imgrc=ul2ayzEdm9CLWM%3A
https://www.google.rs/search?q=macrocystis&espv=2&biw=1366&bih=662&tbm=isch&tbo
=u&source=univ&sa=X&ved=0ahUKEwiZheDz0-
fPAhWhAsAKHQKwCWwQsAQIJw#imgrc=kY7SaDUSP5pIbM%3A
https://www.google.rs/search?q=ectocarpus&espv=2&biw=1366&bih=662&tbm=isch&tbo=
u&source=univ&sa=X&ved=0ahUKEwje1JWQ1OfPAhWrKsAKHaTeCk4QsAQIGQ#imgr
c=5dJRWLpYF20YvM%3A

34
Прилог 4/1
Образац Q4.09.13 - Издање 1
Редни број, РБР:
Идентификациони број, ИБР:
Тип документације, ТД: Монографска
Тип записа, ТЗ: Текстуални / графички
Врста рада, ВР: Мастер рад
Аутор, АУ: Јелена Величковић
Ментор, МН: Наташа Јоковић
Наслов рада, НР: ЗАВИСНОСТ ПРОДУКЦИЈЕ БИОМАСЕ МИКРОАЛГЕ ИЗОЛОВАНЕ ИЗ ВЛАСИНСКОГ ЈЕЗЕРА ОД ИЗВОРА АЗОТА У ПОДЛОЗИ
Језик публикације, ЈП: Српски
Језик извода, ЈИ: Енглески
Земља публиковања, ЗП: Р. Србија
Уже географско подручје, УГП: Р. Србија
Година, ГО: 2016.
Издавач, ИЗ: ауторски репринт
Место и адреса, МА: Ниш, Вишеградска 33.
Физички опис рада, ФО:
(поглавља/страна/
цитата/табела/слика/графика/прилога)
6 поглавља, 43 стране, 16 слика, 3 табеле
Научна област, НО: Биологија
Научна дисциплина, НД: Алгологија
Предметна одредница/Кључне речи, ПО:
УДК 662.6/.9 : 582.26 + (497.11) : 556.55 + 661.938
Чува се, ЧУ: Библиотека
Важна напомена, ВН: Извод, ИЗ: Микроалге представљају значајну сировину у биотехнологији због високог процента
угљених хидрата, протеина, липида и других метаболита које синтетишу. Сојеви
микроалги који се користе у биотехнологији, изолују се из различитих природних
средина, а затим се врши њихова карактеризација у лабораторијским условима.
Како би се продуковала довољна биомаса микроалги која се може користити за
даља испитивања, потребно је урадити оптимизацију раста микроалги.
Оптимизација раста подразумева одређиванје састава подлоге као и услова под
којима одређени сој микроалге даје максималну продукцију биомасе.
У овом раду, из узорака воде са Власинског језера, изолован је један изолат
микроалге који је идентификован као представник рода Closterium. Да би се
испитало како извори азота утичу на раст микроалге, микроалга је гајена 16 дана на
подлогама са различитим изворима азота, амонијум ферицитратом, натријум
нитратом и мешавином ова два једињења. Раст микроалге праћен је
спектрофотометријски док је усвајање азота из подлоге одређивано хемијским
спектрофотометријским методама. Након месец дана гајења микроалге, одређена је
продукована биомаса гравиметријском методом.
Микроалга је показала добар раст на подлози са амонијум ферицитратом као
јединим извором азота, као и на подлози са мешовитим изворима азота, амонијум
ферицитратом и натријум нитратом. Резултати хемијских анализа узорака подлога
на којима је расла микроалга показали су да микроалга не усваја нитрате из подлога
за раст, већ да користи амонијумов јон као извор азота. Принос биомасе након
месец дана гајења микроалге био је највећи на подлози која је садржала амонијум
ферицитрат и натријум нитрат.
Датум прихватања теме, ДП:
Датум одбране, ДО:
Чланови комисије, КО: Председник:
Члан:
Члан, ментор:
ПРИРОДНО - МАТЕМАТИЧКИ ФАКУЛТЕТ
НИШ
КЉУЧНА ДОКУМЕНТАЦИЈСКА ИНФОРМАЦИЈА

35
Прилог 4/2
ПРИРОДНО - МАТЕМАТИЧКИ ФАКУЛТЕТ
НИШ
KEY WORDS DOCUMENTATION
Accession number, ANO:
Identification number, INO:
Document type, DT: monograph
Type of record, TR: textual / graphic
Contents code, CC: master thesis
Author, AU: Jelena Veličković
Mentor, MN: Nataša Joković
Title, TI: The dependence of biomass production from microalgae isolated from Vlasina
lake on the source of nitrogen in media
Language of text, LT: Serbian
Language of abstract, LA: English
Country of publication, CP: Serbia
Locality of publication, LP: Serbia
Publication year, PY: 2016
Publisher, PB: author’s reprint
Publication place, PP: Niš, Višegradska 33.
Physical description, PD:
(chapters/pages/ref./tables/pictures/graphs/a
ppendixes)
6 chapters, 43 pages, 16 pictures, 3 tables
Scientific field, SF: biology
Scientific discipline, SD: algology
Subject/Key words, S/KW:
UC 662.6/.9 : 582.26 + (497.11) : 556.55 + 661.938
Holding data, HD: library
Note, N:
Abstract, AB: Microalgae represent a significant raw material in biotechnology due to their high percentage of carbon hydrates, proteins, lipids and other metabolites they synthetize. Microalgae breeds which are used in biotechnology are isolated from different natural habitats and then characterized in laboratory conditions. In order to produce a great enough biomass of microalgae for use in further study, it is necessary to perform a growth optimization of microalgae. Growth optimization implies determination of the media composition as well as conditions under which a certain breed of microalgae gives the maximal production of biomass. In this study we have isolated an isolate of a microalgae from water samples of Vlasina lake, which has been identified as a representative of the Closterium genus. In order to examine how nitrogen sources affect the growth of microalgae, the microalgae was grown for 16 days on medias with different nitrogen sources, like ammonium fericitrate, sodium nitrate and a mixture of these two compounds. The growth of microalgae was spectrophotometrically observed while the nitrogen absorption from the media was determined by chemical spectrophotometric methods. After a month of microalgae growth, the produced biomass was determined by a gravimetric method.The microalgae has shown a good growth on the media with ammonium fericitrate as the only nitrogen source, as well as on the media with a mixture of nitrogen sources, ammonium fericitrate and sodium nitrate. The results of chemical analysis of media samples on which the microalgae grew have shown that the microalgae doesn’t absorb nitrates from medias for growth, but it instead uses ammonia ion as the source of nitrogen. The biomass yield after a month of microalgae growth was the biggest on the media which contained ammonium fericitrate and sodium nitrate.
Accepted by the Scientific Board on, ASB:
Defended on, DE:
Defended Board, DB: President:
Member:
Member, Mentor:
Образац Q4.09.13 - Издање 1





























