ΘΕΜΑ 1
Περίληψη
Κάθε νουκλεοτίδιο αποτελείται από μια βάση πουρίνης ή πυριμιδίνης, από μια
πεντόζη (2-δεοξυ-D-ριβόζη ή D-ριβόζη για DNA ή RNA αντίστοιχα) και από μια
φωσφορική ομάδα. Τα φωσφορικά οι ζωντανοί οργανισμοί τα προσλαμβάνουν με τις
τροφές τους. Η ριβόζη δημιουργείται στον κύκλο των φωσφορικών πεντοζών και
είναι δυνατό να αναχθεί σε δεοξυριβόζη. Όσο για τη σύνθεση των πουρινών και
πυριμιδινών θα πρέπει να παρατηρήσουμε ότι οι πρόδρομες ενώσεις για τη σύνθεση
του DNA και του RNA είναι οι τριφωσφορικοί εστέρες των νουκλεοζιτών, επομένως
δε θα εστιάσουμε την προσοχή μας μόνο στις βάσεις αλλά και στη σύνθεση ATP,
GTP, CTP και UTP.
Οι πορείες για τη βιοσύνθεση των νουκλεοτιδίων κατατάσσονται σε δύο
ομάδες, στις πορείες de novo και στις πορείες περίσωσης. Στις πορείες de novo οι
νουκλεοτιδικές βάσεις συναρμολογούνται από απλούστερες ενώσεις, ενώ στις
πορείες περίσωσης, προσχηματισμένες βάσεις ανακτώνται και επανασυνδέονται με
μια μονάδα ριβόζης.
To φωσφορικό καρβαμίδιο και το ασπαραγινικό οξύ απαιτούνται αρχικά για
το σχηματισμό του δακτυλίου της πυριμιδίνης. Στη συνέχεια ο δακτύλιος κλείνει,
σχηματίζεται με οξείδωση το οροτικό οξύ, έπειτα το οροτιδυλικό οξύ και ακολουθεί
το ουριδυλικό οξύ (UMP) το οποίο με τροποποιήσεις είναι δυνατό να καταλήξει στο
κυτιδυλικό οξύ (CMP). Η φωσφορική ριβόζη αποτελεί το δομικό λίθο για το
σχηματισμό του πουρινικού δακτυλίου, επάνω στην οποία ενσωματώνονται αρχικά
ένα άτομο Ν και η ριβόζη, καθώς και άτομα C και Ν από διάφορες ενώσεις και
συνένζυμα. Ακολουθούν οι αμινομάδες από το ασπαραγινικό οξύ και την αμιδική
ομάδα της γλουταμίνης. Οι πουρίνες στον άνθρωπο και σε ορισμένα άλλα είδη
αποικοδομούνται προς ουρικό οξύ, ενώ στα περισσότερα ζώα συνεχίζουν να
διασπώνται σε αλλαντοίνη, η οποία και αποβάλλεται. Οι πυριμιδίνες τέλος
μετατρέπονται σε β-αμινοξέα, NH3 και CO2.
Οι διαταραχές του μεταβολισμού των πουρινών και πυριμιδινών (PP) οδηγούν
σε μια ευρεία ομάδα ασθενειών, με ετερογένεια στα συμπτώματα και
υποεκπροσώπηση στη διεθνή βιβλιογραφία σε σχέση με τη σημασία τους. Η έγκαιρη
διάγνωση και η πρόληψη των ασθενειών αυτών αποτελεί ζητούμενο που είναι δυνατό
να ικανοποιηθεί με την ανάπτυξη πολύ ευαίσθητων εργαστηριακών τεχνικών, όπως
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

HPLC και Mass Spectrometry. Συγκεκριμένα συμπτώματα που δεν ερμηνεύονται στη
βάση κοινών ασθενειών πρέπει να μας οδηγούν στην αναζήτηση τέτοιων διαταραχών.
Ο πιο κοινός εκπρόσωπος των διαταραχών PP είναι η ουρική αρθρίτιδα, ενώ το
σύνδρομο Lesch-Nyhan αποκαλύπτει ότι η ανώμαλη συμπεριφορά όπως είναι ο
αυτοακρωτηριασμός και η εξαιρετική εχθρικότητα, μπορεί να προκληθούν από την
απουσία ενός μόνο ενζύμου.
Νουκλεϊνικά οξέα και νουκλεοτίδια
Στα δεοξυριβονουκλεοτίδια απαντούν τέσσερις διαφορετικές ετεροκυκλικές
βάσεις. Οι δύο είναι υποκατεστημένες πουρίνες (αδενίνη και γουανίνη), ενώ οι άλλες
δύο είναι υποκατεστημένες πυριμιδίνες (κυτοσίνη και θυμίνη). Η αδενίνη, η γουανίνη
και η κυτοσίνη αποτελούν επίσης συστατικά των RNA, όμως η θυμίνη δεν απαντά σε
αυτά και αντικαθίσταται από μια άλλη πυριμιδινική βάση που λέγεται ουρακίλη. [1]
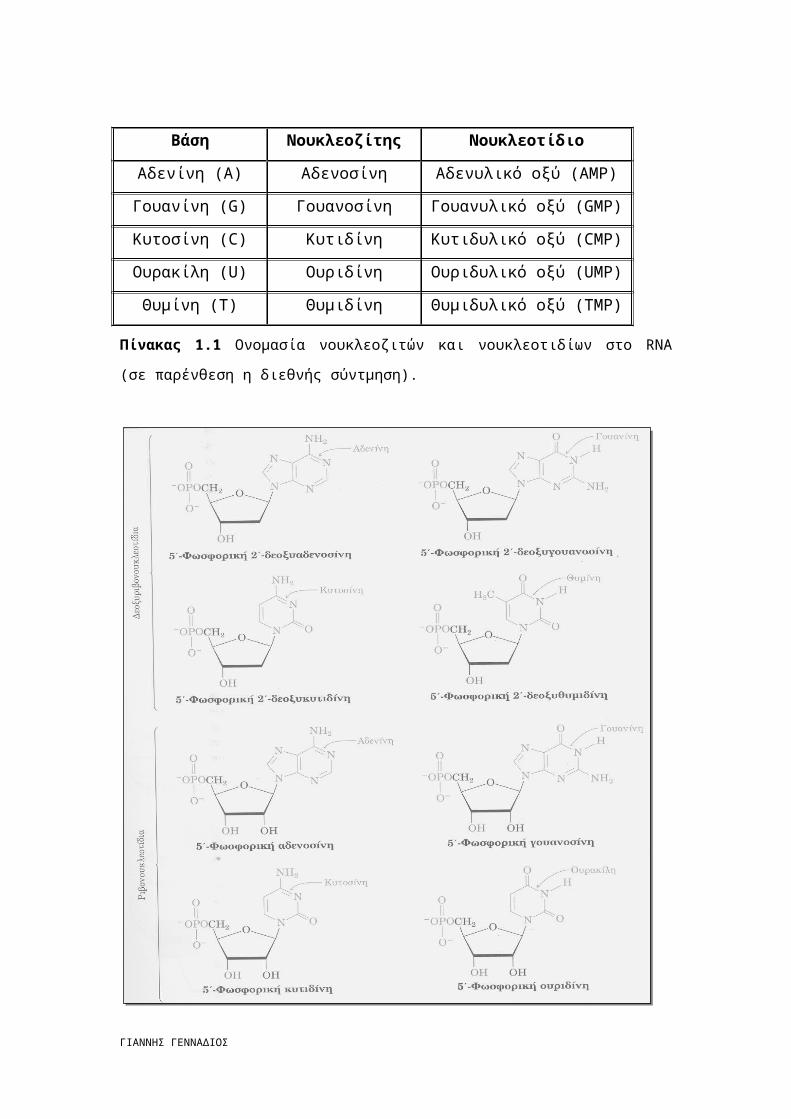
Σχήμα 1.1 Οι πουρίνες και πυριμιδίνες των νουκλεϊκών οξέων.
Οι πλήρεις δομές των τεσσάρων δεοξυριβονουκλεοτιδίων και των τεσσάρων
ριβονουκλεοτιδίων παρατίθενται στο Σχήμα 1.2.
Για τις πορείες της βιοσύνθεσης και αποδόμησης πουρινών και πυριμιδινών
θα χρησιμοποιήσουμε τη σύντμηση των ονομάτων τους που παρουσιάζεται στον
Πίνακα 1.1. Οι νουκλεοζίτες και τα νουκλεοτίδια που περιέχουν δεοξυριβόζη, όπως
στην περίπτωση του DNA, παίρνουν το πρόθεμα δεοξυ-. [2]
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Βάση Νουκλεοζίτης Νουκλεοτίδιο
Αδενίνη (A) Αδενοσίνη Αδενυλικό οξύ (AMP)
Γουανίνη (G) Γουανοσίνη Γουανυλικό οξύ (GMP)
Κυτοσίνη (C) Κυτιδίνη Κυτιδυλικό οξύ (CMP)
Ουρακίλη (U) Ουριδίνη Ουριδυλικό οξύ (UMP)
Θυμίνη (T) Θυμιδίνη Θυμιδυλικό οξύ (TMP)
Πίνακας 1.1 Ονομασία νουκλεοζιτών και νουκλεοτιδίων στο RNA (σε παρένθεση η διεθνής
σύντμηση).
Σχήμα 1.2 Ονομασίες και δομές των τεσσάρων δεοξυριβονουκλεοτιδίων και των τεσσάρων
ριβονουκλεοτιδίων.
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ
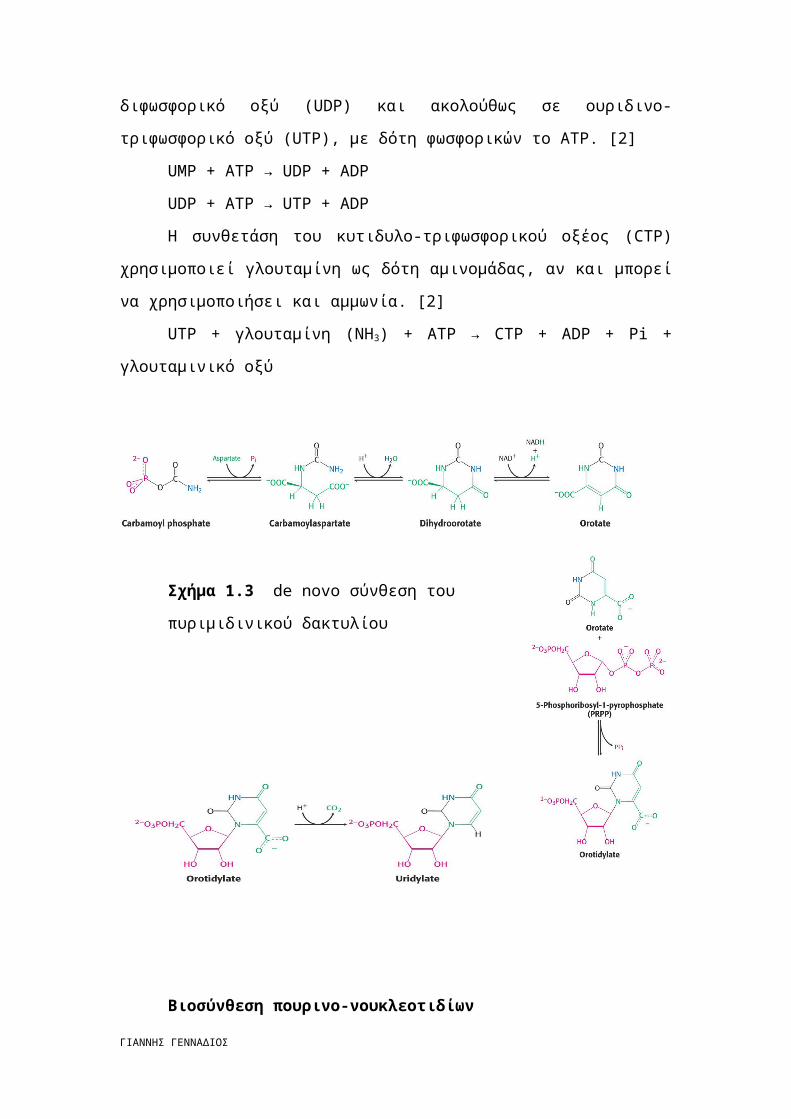
Βιοσύνθεση πυριμιδινο-νουκλεοτιδίων
Ως πρόδρομες ενώσεις για τη σύνθεση του δακτυλίου της πυριμιδίνης
χρησιμεύουν το φωσφορικό καρβαμίδιο και το ασπαραγινικό οξύ. [2]
Ο δακτύλιος κλείνει, και η ένωση που προκύπτει οξειδώνεται σε οροτικό οξύ.
Στο σημείο αυτό ενσωματώνεται η φωσφορική ριβόζη και προκύπτει το οροτιδυλικό
οξύ, που με τη σειρά του αποκαρβοξυλιώνεται σε ουριδυλικό οξύ (UMP), ένα από τα
τέσσερα κύρια νουκλεοτίδια του RNA. [2]
Το κυτιδυλικό οξύ (CMP) προέρχεται από το ουριδυλικό οξύ, αφού όμως το
τελευταίο μετατραπεί πρώτα σε ουριδινο-διφωσφορικό οξύ (UDP) και ακολούθως σε
ουριδινο-τριφωσφορικό οξύ (UTP), με δότη φωσφορικών το ATP. [2]
UMP + ATP → UDP + ADP
UDP + ATP → UTP + ADP
Η συνθετάση του κυτιδυλο-τριφωσφορικού οξέος (CTP) χρησιμοποιεί
γλουταμίνη ως δότη αμινομάδας, αν και μπορεί να χρησιμοποιήσει και αμμωνία. [2]
UTP + γλουταμίνη (ΝΗ3) + ATP → CTP + ADP + Pi + γλουταμινικό οξύ
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Σχήμα 1.3 de novo σύνθεση του
πυριμιδινικού δακτυλίου
Βιοσύνθεση πουρινο-νουκλεοτιδίων
Σε αντίθεση με τα πυριμιδινο-νουκλεοτίδια, όπου συντίθεται πρώτα ο
πυριμιδινικός δακτύλιος και ακολούθως ενσωματώνεται η φωσφορική ριβόζη, στην
περίπτωση των πουρινο-νουκλεοτιδίων οι αντιδράσεις ξεκινούν από τη φωσφορική
ριβόζη επάνω στην οποία κτίζεται κατά κάποιον τρόπο ο πουρινικός δακτύλιος. [2]
Το πρώτο άτομο που ενσωματώνεται είναι το Ν, μέσω του οποίου ο δακτύλιος
δεσμευεται με τη ριβόζη. Δότης του αζώτου είναι η γλουταμίνη. [2]
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ
Σχήμα 1.4 Συναρμολόγηση πουρινικού δακτυλίου σε εννιά βήματα.
Δότες των υπολοίπων ατόμων C και Ν του δακτυλίου είναι διάφορες ενώσεις
και συνένζυμα, όπως φαίνεται στο σχήμα 1.5. [2]
Η αμινομάδα της αδενίνης προέρχεται από το ασπαραγινικό οξύ, εκείνη δε της
γουανίνης από την αμιδική ομάδα της γλουταμίνης. [2]
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Σχήμα 1.5 Προέλευση των ατόμων C και Ν του πουρινικού δακτυλίου.
Οι πουρίνες οξειδώνονται και αποβάλλονται με διάφορες μορφές ανάλογα με
τον οργανισμό.
Τόσο η αδενίνη όσο και η γουανίνη απαμινώνονται και μετατρέπονται σε
υποξανθίνη και ξανθίνη αντίστοιχα. Η υποξανθίνη οξειδώνεται επίσης προς ξανθίνη
και αυτή σε ουρικό οξύ. Στο σημείο αυτό σταματά ο περαιτέρω μεταβολισμός των
πουρινών στον άνθρωπο και σε ορισμένα άλλα είδη (το ουρικό οξύ αποβάλλεται),
ενώ προχωρεί με διάσπαση του δακτυλίου στα περισσότερα ζώα δίνοντας
αλλαντοΐνη, η οποία αποβάλλεται στα περισσότερα θηλαστικά. Στα περισσότερα
ψάρια και αμφίβια η αλλαντοΐνη μετατρέπεται σε ουρία και αποβάλλεται, ενώ σε
ορισμένα θαλάσσια ασπόνδυλα μετατρέπουν και την ουρία σε ΝΗ3 και CO2. [2]
Σχήμα 1.6 Οι χημικοί τύποι των ενώσεων που σχετίζονται με την αποδόμηση των
πουρινών.
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ
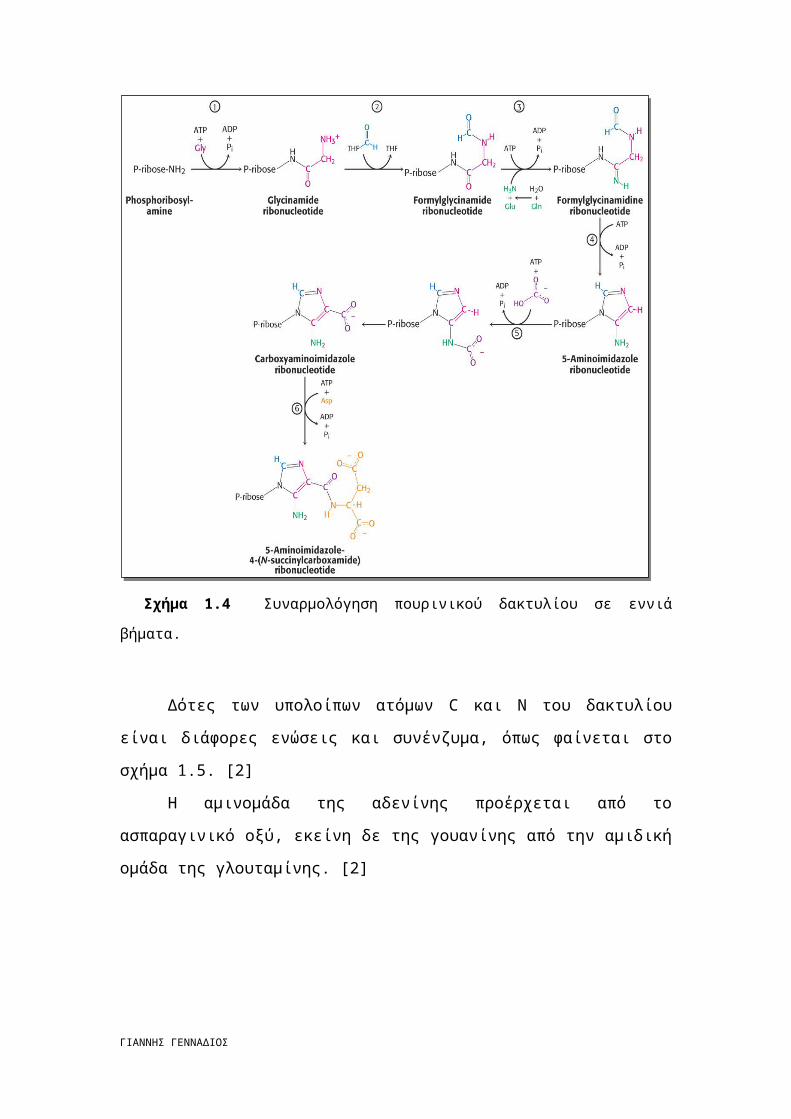
Σχήμα 1.7 Η AMP αποικοδομείται προς την ελεύθερη βάση υποξανθίνη μέσω
απαμίνωσης και υδρολυτικής διάσπασης του γλυκοζιτικού δεσμού. Η οξειδάση της ξανθίνης
οξειδώνει την υποξανθίνη σε ξανθίνη και στη συνέχεια σε ουρικό οξύ. Το ουρικό οξύ χάνει
ένα πρωτόνιο σε συνθήκες φυσιολογικού pH για να σχηματίσει ουρικό ιόν. Στον άνθρωπο, το
ουρικό ιόν είναι το τελικό προϊόν αποδόμησης και απεκκρίνεται από τα ούρα.
Οι πυριμιδίνες μετατρέπονται σε β-αμινοξέα, ΝΗ3 και CO2.
Η κυτοσίνη και η ουρακίλη έχουν κοινή μεταβολική τύχη, μια και η πρώτη
απαμινώνεται και μετατρέπεται στη δεύτερη. Η ουρακίλη ανάγεται σε
διυδροουρακίλη και ακολούθως ο δακτύλιος διασπάται και δίνει β-ουρεϊδοπροπιονικό
οξύ, το οποίο με τη σειρά του υδρολύεται προς β-αλανίνη, ΝΗ3 και CO2. Η θυμίνη
έχει παρόμοια τύχη με την ουρακίλη, μόνο που το τελικό προϊόν είναι β-
αμινοϊσοβουτυρικό οξύ αντί για β-αλανίνη. [2]
Σχήμα 1.8 Οι χημικοί τύποι της αλανίνης και του αμινοϊσοβουτυρικού οξέος.
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Γενικά για τις διαταραχές του μεταβολισμού πουρινών και πυριμιδινών
Οι γενετικές διαταραχές του μεταβολισμού των πουρινών και πυριμιδινών
(PP) δεν απαντώνται συχνά στη βιβλιογραφία και αναφέρονται συνήθως σε reviews
σχετικά με άλλες εκ γενετής διαταραχές του μεταβολισμού. Εξαιτίας του
περιορισμένου - σε σχέση με τη σημασία τους - ενδιαφέροντος και της σχετικά
πρόσφατης αναγνώρισή τους, οι ανωμαλίες αυτές είναι δυνατό συχνά να μην
τυγχάνουν διάγνωσης ή να ακολουθούνται από μια λάθος διάγνωση. Οι ανωμαλίες
που παρουσιάζονται ως αποτέλεσμα κακής λειτουργίας του μεταβολισμού PP
αντιπροσωπεύουν ένα από τα πλέον ενδιαφέροντα διαγνωστικά προβλήματα στην
ιατρική, ιδιαίτερα από τη στιγμή που εμφανίζονται με ένα εύρος συμπτωμάτων.
Μπορούν να προσβάλλουν οποιοδήποτε σύστημα του ανθρώπου με διάφορους
τρόπους και συχνά προσομοιάζουν σε άλλες, πιο γνωστές διαταραχές. Τα
διαγνωστικά προβλήματα σχετίζονται πολλές φορές με το ότι μερικοί ασθενείς είναι
ασυμπτωματικοί. Εργαστηριακές τεχνικές που αναπτύσσονται ταχύτατα όπως η υγρή
χρωματογραφία υψηλής πίεσης (high-performance liquid chromatography, HPLC), σε
συνδυασμό με τη φασματοσκοπία μαζών (mass spectrometry), έχουν πλέον
αποδειχτεί πολύ χρήσιμες στην ανίχνευση των ασθενειών αυτών, αλλά το πιο κρίσιμο
βήμα θα αποτελέσει η διάγνωση που μπορεί να προβλέπει τις ανωμαλίες. Η έγκαιρη
διάγνωση είναι σήμερα θέμα ζωτικής σημασίας επειδή η γενετική καθοδήγηση
μπορεί να επιτρέπει για κάθε περίπτωση ειδική θεραπεία που θα επιβραδύνει ή και θα
αποτρέπει τα κλινικά συμπτώματα. Εάν οι ανωμαλίες αυτές δε διαγνωστούν, μπορεί
να αποδειχθούν καταστροφικές για τους ασθενείς και τις οικογένειές τους, οδηγώντας
σε θάνατο ή σε μόνιμη κατάσταση των συμπτωμάτων για όλη τη ζωή του ασθενούς.
[4]
Το να υποπτευθεί κανείς και έτσι να ερευνήσει τις εκ γενετής διαταραχές στο
μεταβολισμό των πουρινών και πυριμιδινών είναι ένα δίλλημα, ακόμη και για τον πιο
παρατηρητικό ερευνητή. Συχνά οι γονείς των παιδιών που πάσχουν ή έχουν ιστορικό
με αμφιθαλή αδέρφια, μπορούν να παρέχουν πολύτιμες ενδείξεις. Η αναγνώριση μιας
νέας ασθένειας που οφείλεται σε μεταβολισμό PP απαιτεί ικανότητες και εύνοια σε
τυχαίες ανακαλύψεις. Αλλά ακόμη και το να αναγνωρίσει κανείς τις ήδη γνωστές
διαταραχές μπορεί να αποδειχτεί δύσκολο από τη στιγμή που αυτές καλύπτουν ένα
ευρύ φάσμα ασθενειών με περισσότερα από ένα συμπτώματα ή και πρόωρο θάνατο.
Αυτό το πρόβλημα συνδέεται με το γεγονός ότι οι παθολογικές καταστάσεις του
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ
μεταβολισμού PP έχουν περιγραφεί σχετικά πρόσφατα και για αυτό λίγα είναι όσα
γνωρίζουμε, είτε σε επίπεδο κλινικών συμπτωμάτων είτε σε εργαστηριακό επίπεδο. Η
ετερογένεια στην κλινική έκφραση μέσα στις οικογένειες σημαίνει ότι οι
ασυμπτωματικοί ομoζυγώτες δε μπορούν εύκολα να αναγνωριστούν. Τα συμπτώματά
τους είναι δυνατό να παρουσιαστούν από την παιδική ή την εφηβική τους ηλικία έως
και την όγδοη δεκαετία της ζωής τους. [5]
Οι ανωμαλίες αυτές θα πρέπει να αναζητούνται σε κάθε περίπτωση
ανεξήγητης αναιμίας, αρρυθμίας, επιδεκτικότητας σε επαναλαμβανόμενες μολύνσεις,
ή νευρολογικές διαταραχές χωρίς διάγνωση, συμπεριλαμβανομένων των αυτισμού,
εγκεφαλικής παράλυσης, καθυστέρησης στην ανάπτυξη, κώφωσης, επιληψίας, μυικής
ατονίας, αδυναμίας σε βάδιση και ομιλία και ασυνήθιστες για την παιδική και
εφηβική ηλικία ασθένειες τύπου gout συνοδευόμενες μερικές φορές από νεφρική
ανεπάρκεια. Μερικές ανωμαλίες οδηγούν σε πέτρα στα νεφρά, σε οξεία ή χρόνια
νεφρική ανεπάρκεια μόνη της ή σε συνδυασμό με τα παραπάνω συμπτώματα ή
οδηγούν επίσης σε μεγάλη ευαισθησία και μη ανοχή σε θεραπείες (π.χ. 5-fluorouracil
σε κακοήθεις νεοπλασίες ή azathioprine για την ανοσοκαταστολή σε περίπτωση
μεταμόσχευσης). Συμπερασματικά, διάφοροι είναι οι παράμετροι που πρέπει να
συνεκτιμηθούν για να διασφαλιστεί η σωστή διάγνωση. [5]
Ουρική αρθρίτιδα
Η ουρική αρθρίτιδα (gouty arthritis) είναι πάθηση των αρθρώσεων που είναι
αποτέλεσμα εναπόθεσης σε αυτές κρυστάλλων ουρικού μονονατρίου, λόγω
υπερκορεσμού του αρθρικού υγρού σε ουρικό οξύ. Αν και στην καθομιλουμένη
λέγεται «ποδάγρα» (αρχαία λέξη από το πους + άγρα=παγίδα για τα πόδια), η
επιστημονική της ονομασία είναι «υποτροπιάζουσα οξεία ή χρόνια
κρυσταλλογενής αρθρίτιδα περιφερικών αρθρώσεων». [3]
Ουρικό οξύ και ουρική αρθρίτιδα
Το ουρικό οξύ είναι το τελικό προϊόν του μεταβολισμού των νουκλεϊνικών
οξέων και συγκεκριμένα των πουρινών, που αποτελούν τα δομικά συστατικά του
γενετικού υλικού όλων των κυττάρων μας. Σχεδόν το 1/3 των πουρινών πού
υπάρχουν στο σώμα μας προέρχονται από εξωγενή χορήγηση από τη διατροφή, ενώ
τα 2/3 από την ενδογενή παραγωγή. Οι φυσιολογικές τιμές του ουρικού οξέος του
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

αίματος είναι 3,5-7,2 mg/dL στους άνδρες, 2,6-6,0 mg/dL για τις γυναίκες και 2,0-5,5
mg/dL για τα παιδιά. Θεωρείται ότι όταν το ουρικό οξύ ξεπεράσει τα 7 mg/dL
(υπερουριχαιμία, hyperuricaemia), επέρχεται κορεσμός, με αποτέλεσμα να
δημιουργούνται άλατα, τα οποία με τη μορφή κρυστάλλων (κρύσταλλοι ουρικού
μονονατρίου), να καθιζάνουν και να εναποτίθενται στο αρθρικό υγρό, στα άλλα
ανατομικά μόρια της άρθρωσης και σε άλλους ιστούς. Αποτέλεσμα της καθίζησης
αυτής, είναι η πρόκληση φλεγμονής (αρθρίτιδας). Αθροίσεις τέτοιων κρυστάλλων,
ονομάζονται ουρικοί τόφοι και είναι αρκετά μεγάλοι ώστε να είναι ορατοί σε
ακτινογραφίες ή να είναι αισθητοί σαν υποδόρια μορφώματα. Η οξεία ουρική
αρθρίτιδα συνήθως εκδηλώνεται μετά από ένα πλούσιο και «βαρύ» δείπνο και μετά
από μεγάλη κατανάλωση αλκοόλ και εμφανίζεται ξαφνικά. Η νόσος συχνά προσβάλει
υπέρβαρους άνδρες άνω των 40 ετών, χωρίς να αποκλείονται και άλλες ηλικίες. Σε
μερικές περιπτώσεις oξείας ουρικής αρθρίτιδας, η τιμή του ουρικού οξέος μπορεί να
είναι φυσιολογική. Αυτό εξηγείται ως εξής: για κάποιο λόγο δημιουργείται υψηλή
τιμή ουρικού οξέος αίματος (peak), η οποία σε συνδυασμό με εκλυτικούς
παράγοντες, προκαλεί οξεία ουρική αρθρίτιδα. Μετά από επαρκή όμως ενυδάτωση,
επέρχεται μείωση τιμών του ουρικού οξέος, οπότε την επόμενη ή μεθεπόμενη ημέρα
οι τιμές είναι φυσιολογικές. Στο μεταξύ η ουρική αρθρίτιδα συνεχίζει να υπάρχει.
Στατιστικά, το 30% των ασθενών με οξεία ουρική αρθρίτιδα έχει φυσιολογικό ουρικό
οξύ αίματος. [3]
Ουρική Νόσος
Ουρική νόσος (Αγγλικά Gout = γκάουτ, Γερμανικά Ζipperlein), ονομάζεται το
σύνολο των κλινικών, εργαστηριακών και ιστολογικών διαταραχών, πού οφείλονται
σε διαταραχή του μεταβολισμού η οποία έχει σαν αποτέλεσμα την υπερουριχαιμία
και την εναπόθεση κρυστάλλων ουρικού νατρίου στους ανθρώπινους ιστούς. Ακριβής
μονολεκτική απόδοση του Αγγλικού όρου Gout στα Ελληνικά δεν υπάρχει, πλην του
παλαιού αρχαίου όρου «ποδάγρα» ο οποίος όμως έχει σχεδόν καταργηθεί ως
αδόκιμος. Συνεπώς πρέπει να χρησιμοποιείται ο σωστός όρος Ουρική Νόσος. Ο όρος
Ουρική Αρθρίτιδα δεν είναι πάντα επιτυχής διότι μπορεί να έχουμε Ουρική
Ελυτρίτιδα, οπότε δεν υπάρχει αρθρίτις. H ουρική αρθρίτιδα (αγγλικά Gouty
Αrthritis), είναι φλεγμονώδης αρθρίτιδα, οφειλόμενη σε εναπόθεση κρυστάλλων
Ουρικού Νατρίου, σε μία ή περισσότερες αρθρώσεις, ενίοτε όμως και σε
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

παρααρθρικά μαλακά μόρια (θύλακοι, τένοντες, έλυτρα, κλπ). Η συνηθέστερη
άρθρωση που προσβάλλεται είναι η Πρώτη Μεταταρσιοφαλαγγική στο πόδι (παλαιά
ονομασία «Ποδάγρα»), και ακολουθούν κατά σειρά συχνότητας η Ποδοκνημική
Άρθρωση, οι Μεταταρσιοφαλαγγικές αρθρώσεις των ποδιών, οι Μεσοφαλαγγικές
Αρθρώσεις των χεριών, τα γ όνατα , οι αγκώνες (ορογονοθυλακίτις του ωλεκράνου,
κλπ). Η αρθρίτιδα της Μεσοφαλαγγικής αρθρώσεως (ΦΦ) στο μεγάλο δάκτυλο του
ποδιού δεν είναι ουρική αρθρίτιδας, αλλά οστεοαρθρίτιδα ή θυλακίτις της κεφαλής
μεταταρσίου. [3]
Αίτια
Το αίτιο που προκαλεί την ουρική αρθρίτιδα είναι η αύξηση του ουρικού
οξέος αίματος (υπερουριχαιμία). Αυτή, μπορεί να είναι πρωτοπαθής (ιδιοπαθής) και
δευτεροπαθής. Οι παράγοντες που ευνοούν την καθίζηση των κρυστάλλων του
ουρικού μονονατρίου στις αρθρώσεις και την πρόκληση της ουρικής αρθρίτιδας είναι:
1. Αυξημένη πρόσληψη πουρινών με την τροφή, δηλαδή αυξημένη
κατανάλωση εντοσθίων, μικρών ζώων, θαλασσινών, προϊόντων
κυνηγίου κ.α.
2. Αυξημένη κατανάλωση αλκοόλ.
3. Αυξημένη ανακύκλωση πυρηνοπρωτεϊνών που συμβαίνει σε ορισμένα
αιματολογικά νοσήματα όπως λεμφώματα, λευχαιμίες, αιμολυτικές
αναιμίες καθώς και σε άλλα νοσήματα όπως ψωρίαση.
4. Ύπαρξη γενετικών ανωμαλιών ενζύμων, που συμμετέχουν στο
μεταβολισμό του ουρικού οξέος (π.χ. σύνδρομο Lesch-Nyhan).
5. Χωρίς να διαπιστώνεται το αίτιο (ιδιοπαθής υπερουριχαιμία).
6. Ελαττωμένη αποβολή του ουρικού οξέος από τους νεφρούς: Νεφρική
βλάβη.
7. Διάφορα φάρμακα.
8. Καταστάσεις που προκαλούν μείωση του pH των ούρων.
9. Καταστάσεις οξέωσης (διαβητική, γαλακτική, νηστεία).
Όλες οι καταστάσεις που αναφέρθηκαν, προκαλούν υπερουριχαιμία, (αύξηση
της τιμής του ουρικού οξέος στο αίμα). Παραδόξως, υπάρχουν ασθενείς με αυξημένες
τιμές ουρικού οξέος αίματος, που δεν αναπτύσσουν ποτέ ουρική αρθρίτιδα. [3]
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Αίτια της πρωτοπαθούς υπερουριχαιμίας
Το αίτιο της πρωτοπαθούς ουρικής αρθρίτιδας φαίνεται να είναι η μερική ή
πλήρης (σύνδρομο Lesh-Nyhan) έλλειψη του ενζύμου ΗGPRT (υποξανθίνη-
γουανίνη-φωσφο-ριβόζυλο-τρανσφεράση) το οποίο μετατρέπει την υποξανθίνη σε
ινοσινικό οξύ. H πρωτοπαθής υπερουριχαιμία μπορεί να είναι:
(α) Ιδιοπαθής.
(β) Επί Συνδρόμου Lesch-Nyhan (το οποίο οφείλεται σε πλήρη έλλειψη του
αναφερθέντος ενζύμου).
(γ) Νόσος Εναποθέσεως Γλυκογόνου (Glycogen Storage Disease).
(δ) Ιδιοπαθής Ανεπάρκεια Νεφρικής Κάθαρσης Ουρικού οξέος. Εκδηλώνεται
είτε μετά από διαιτητικές παρεκτροπές, είτε και σε επιτρεπτό διαιτολόγιο (ενδογενής).
Στην περίπτωση αυτή, δεν υποχωρεί μόνο με την ειδική δίαιτα. [3]
Αίτια της δευτεροπαθούς υπερουριχαιμίας
Δευτεροπαθής υπερουριχαιμία παρατηρείται σε δύο μεγάλες κατηγορίες
νοσημάτων:
Νοσήματα με αυξημένο καταβολισμό και εναλλαγή πουρινών (Purine
turnover):
Μυελοϋπερπλαστικά νοσήματα, λευχαιμία, υπερερυθραιμία.
Λεμφοϋπερπλαστικά νοσήματα, λέμφωμα, λεμφοκοκκίωμα.
Καρκινώματα και σαρκώματα.
Xρόνιες αιμολυτικές αναιμίες.
Λήψη κυτταροστατικών φαρμάκων (Cytotoxic drugs).
Ψωρίαση , ψωριασική αρθροπάθεια, σαρκοειδική αρθροπάθεια, κλπ.
Νοσήματα με μείωση νεφρικής κάθαρσης ουρικού οξέος:
Εσωτερικά οργανικά νοσήματα των νεφρών (χρόνια νεφρική
ανεπάρκεια, κλπ).
Λειτουργική μείωση της σπειραματικής διήθησης ουρικού οξέος. [3]
Χρόνια τοφώδης ουρική αρθρίτιδα
Είναι το αποτέλεσμα μακροχρόνιας συνήθως υπερουριχαιμίας, με συχνές
προσβολές και εναπόθεση ουρικών τόφων που προκαλούν παραμόρφωση της
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

άρθρωσης και δυσλειτουργία της. Συναντιέται ενίοτε στα χέρια, τα δάχτυλα και τα
γόνατα. [3]
Σύνδρομο Lesch-Nyhan
Σύμφωνα με τη βάση δεδομένων ΟΜΙΜ του NCBI, τα χαρακτηριστικά του
συνδρόμου Lesch-Nyhan (LNS) είναι τα εξής: υπερουριχαιμία, χαρακτηριστικές
νευρολογικές ανωμαλίες, χορειοαθέτωση, αυτοκαταστροφικό δάγκωμα στα δάχτυλα
ή στα χείλη, σπαστικότητα, πνευματική καθυστέρηση και νεφρική ανεπάρκεια. Η
μεγαλοβλαστική αναιμία έχει επίσης προταθεί από κάποιους (van der Zee et al.,
1968). [6]
Η ολική ανεπάρκεια του ενζύμου Υποξανθινική Φωσφοριβοσυλοτρανφεράση
(HPRT) σχετίζεται με το σύνδρομο Lesch-Nyhan ενώ η μερική ανεπάρκεια
σχετίζεται με το σύνδρομο Kelley-Seegmiller. To LNS χαρακτηρίζεται επίσης από μη
φυσιολογικό μεταβολισμό και νευρολογικές εκδηλώσεις. Σε αντίθεση, το Kelley-
Seegmiller σύνδρομο συνήθως σχετίζεται μόνο με κλινικά συμπτώματα υπερβολικής
παραγωγής πουρινών. Πέτρα στα νεφρά, νεφροπάθεια εξαιτίας του ουρικού οξέος και
νεφρική απόφραξη είναι τα πλέον συνήθη συμπτώματα του συνδρόμου Kelley-
Seegmiller, αλλά σπάνια του LNS. Μετά την εφηβεία, η υπερουριχαιμία του
συνδρόμου Kelley-Seegmiller είναι δυνατό να προκαλέσει την ουρική νόσο (gout).
Μια τρίτη ομάδα ασθενών με ενδιάμεση ενεργότητα ενζύμου (1,5 έως 8%),
σχετίζεται αυξημένο ουρικό οξύ και νευρολογική ανικανότητα που διαφέρει από
απλή αδεξιότητα και είναι δυνατό να προκαλέσει κινητικά προβλήματα (Jinnah and
Friedman, 2001). [7]
Το 1979, ο Bakay επανεξέτασε έναν ασθενή με ανεπάρκεια της HPRT, με
χορειοαθέτωση, σπαστικότητα, δυσαρθρία και υπερουριχαιμία αλλά φυσιολογική
νοημοσύνη και καμία τάση αυτοακρωτηριασμού. (Ένας θείος της μητέρας του
εμφάνιζε την ίδια διαταραχή). Παρόλο που η ανεπάρκεια της HPRT έδειχνε να είναι
πλήρης, κύτταρα ινοβλαστών του σε καλλιέργεια είχαν κάποια ικανότητα να
μεταβολίζουν τις ξανθίνη και υποξανθίνη. [8]
Ο Page περιέγραψε δύο αδέρφια και δύο θείους τους που προέρχονταν από
τους συγγενείς της μητέρας τους, που εμφάνιζαν ανεπάρκεια HPRT, διανοητική
καθυστέρηση και βάδισμα με σπασμούς. Στα κύτταρα αυτών των ασθενών
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

παρατηρήθηκαν διαφορές καθώς σε ορισμένους τύπους κυττάρων υπήρχε πλήρης
ανεπάρκεια ενώ σε κάποιους άλλους η ενεργότητα του ενζύμου έφτανε το 7,5% της
φυσιολογικής. Οι κινητικές μελέτες έδειξαν επίσης διαφορές. Η αδερφή τους ήταν, σε
σχέση με το ένζυμο, ετερόζυγη. [9]
Το 2008, ο Hladnik και οι συνεργάτες του, ανέφεραν στην εργασία τους μια
οικογένεια στην οποία πέντε άτομα είχανε μετάλλαξη στο ίδιο σημείο του γονιδίου
της HPRT αλλά εμφάνιζαν φαινοτυπικές διαφορές. Ο ένας ασθενής εμφάνιζε την
κλασσική συμπτωματολογία του συνδρόμου Lesch-Nyhan, με καθυστέρηση στην
ανάπτυξη, σπαστικότητα, δυστονία και τάσεις αυτοτραυματισμού. Δύο ασθενείς
είχαν ενδιάμεσους φαινοτύπους, με κάποιες γνωστικές και μαθησιακές δυσκολίες,
δυστονία και αυξημένα επίπεδα ουρικού οξέος, αλλά χωρίς τάσεις
αυτοτραυματισμών. Οι υπόλοιποι δύο είχαν ήπια σπαστικότητα, gout και κανονικό
IQ. Έτσι, διαπίστωσαν ότι κάθε ένας ασθενής είχε διαφορετική έκφραση του
μεταλλαγμένου μετάγραφου και τόνισαν ότι ασθενείς με τον ίδιο γονότυπο δε θα
έχουν απαραίτητα ταυτόσημο φαινότυπο. [10]
Το 1977, ο Vogel μελετώντας γεγονότα που αφορούσαν στην αιμοφιλία και
το σύνδρομο Lesch-Nyhan, οδηγήθηκε στο συμπέρασμα ότι ο ρυθμός των
μεταλλάξεων είναι υψηλότερος στους άνδρες σε σχέση με τις γυναίκες. [11] Από το
1965 είχε προταθεί η φυλοσύνδετη κληρονομικότητα (Hoefnagel et al.) και
υποστηρίχθηκε από μια σειρά παρατηρήσεων σε οικογένειες με ανεπάρκεια σε
HPRT. [12]
Βιβλιογραφία
1. McMurry J. ‘‘Οργανική Χημεία Τόμος ΙΙ’’, Πανεπιστημιακές Εκδόσεις
Κρήτης, Κεφ. 29, σελ. 1395-1398, 2004.
2. Γεωργάτσος Ι. ‘‘Βιοχημεία,’’, Εκδόσεις ΕΑΠ.; Τόμος Β΄ Κεφ. 6, σελ. 134-
158, 2000.
3. Βικιπαίδεια – ουρική αρθρίτιδα.
4. Jurecka A. ‘‘Inborn errors of purine and pyrimidine metabolism’’, J Inherit
Metab Dis. 2009 Apr; 32(2):247-63, Epub 2009 Mar 15.
5. Simmons HA, Duley JA, FairbanksLD, McBride MB. ‘‘When to investigate
for pyurine and pyrimidine disorders. Introduction and review of clinical and
laboratory indications.’’, J Inherit Metab Dis. 1997 Jun; 20(2):214-26.
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

6. Van der Zee, S. P. M.; Schretlen, E. D. A. M.; Monnens, L. A. H. :
Megaloblastic anaemia in the Lesch-Nyhan syndrome. (Letter) Lancet 291:
1427 only, 1968. Note: Originally Volume I.
7. Jinnah, H. A.; Friedmann, T. : Lesch-Nyhan disease and its variants.In:
Scriver, C. R.; Beaudet, A. L.; Sly, W. S.; Valle, D. (eds.) : The Metabolic &
Molecular Bases of Inherited Disease. Vol. II. 8th ed. New York: McGraw-
Hill 2001. P. 2537.
8. Bakay, B.; Nissinen, E.; Sweetman, L.; Francke, U.; Nyhan, W. L. :
Utilization of purines by an HPRT variant in an intelligent, nonmutilative
patient with features of the Lesch-Nyhan syndrome. Pediat. Res. 13: 1365-
1370, 1979. PubMed ID : 523196
9. Page, T.; Nyhan, W. L.; Morena de Vega, V. : Syndrome of mild mental
retardation, spastic gait, and skeletal malformations in a family with
partial deficiency of hypoxanthine-guanine phosphoribosyltransferase.
Pediatrics 79: 713-717, 1987. PubMed ID : 3575027
10. Hladnik, U.; Nyhan, W. L.; Bertelli, M. : Variable expression of HPRT
deficiency in 5 members of a family with the same mutation. Arch. Neurol.
65: 1240-1243, 2008. PubMed ID : 18779430
11. Vogel, F. : A probable sex difference in some mutation rates. (Editorial)
Am. J. Hum. Genet. 29: 312-319, 1977. PubMed ID : 868880
12. Hoefnagel, D.; Andrew, E. D.; Mireault, N. G.; Berndt, W. O. : Hereditary
choreoathetosis, self-mutilation and hyperuricemia in young males. New
Eng. J. Med. 273: 130-135, 1965. PubMed ID : 14303658
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

ΘΕΜΑ 2
Περίληψη
Η θέση ότι μόνο οι πρωτεΐνες έχουν αρκετή ποικιλομορφία ώστε να φέρουν
εις πέρας ενζυμικές αντιδράσεις, έχει αλλάξει τα τελευταία χρόνια με την ανακάλυψη
ότι RNA μόρια έχουν τη δυνατότητα να πραγματοποιούν μια σειρά από διαφορετικές
καταλυτικές διαδικασίες. Την ενζυμική ενεργότητα τέτοιων RNA μορίων, των
ριβοενζύμων, μπορούμε να μελετήσουμε με τον ίδιο τρόπο όπως και των πιο
συμβατικών πρωτεϊνικών ενζύμων, καταλήγοντας σε συμπέρασμα για μεγάλη
συγγένεια υποστρώματος και μικρό καταλυτικό ρυθμό. Τα ριβοένζυμα μπορούν να
χρησιμοποιήσουν διαφορετικά μόρια RNA ως υπόστρωμα και να ανακυκλωθούν ή να
χρησιμοποιήσουν τον ίδιο τον εαυτό τους σαν υπόστρωμα οπότε χρησιμοποιούνται
μόνο μια φορά. Τα παραδείγματα RNA καταλυτών που θα μελετηθούνε είναι το
ένζυμο ριβονουκλεάση-P, τα εσώνια των ομάδων Ι και ΙΙ, το σφυροκέφαλο
ριβοένζυμο, το ριβοένζυμο HDV και το glmS. Η χρήση των ριβοενζύμων με την
αξιοποίηση των ρυθμιστικών τους ρόλων για τους σκοπούς της Ιατρικής είναι ένα
πεδίο με πολλές προκλήσεις που απαιτεί όμως περαιτέρω έρευνα.
Τα ριβοένζυμα καταλύουν διάφορες ενζυμικές αντιδράσεις, όχι πάντα με τον
ίδιο τρόπο, αλλά παρουσιάζουν μια ποικιλία από κατάλληλες θέσεις ως αντιδρώντα.
Για να επιτευχθεί μια χημική RNA κατάλυση, μερικά τμήματα από το μόριο του
ριβοενζύμου πρέπει να υιοθετήσουν πολύ ακριβείς σχετικές θέσεις και
προσανατολισμούς. Σχεδόν πάντα όμως δρουν επάνω στον φωσφοδιεστερικό
σκελετό, συνδέοντας δυο διαδοχικά νουκλεοτίδια. Όσον αφορά στη δομή τους, ο
αριθμός των συντηρημένων νουκλεοτιδίων και η διασπορά τους από μόριο σε μόριο
διαφέρει σημαντικά.
H ανακάλυψη των ριβοενζύμων και η προσεκτική μελέτη της δράσης και της
δομής τους μας οδηγεί σε σημαντικά συμπεράσματα που αφορούν στην εξέλιξη.
Μπορούμε να υποθέσουμε ότι τα πρώτα συστήματα με δυνατότητα αναπαραγωγής
του εαυτού τους αποτελούνται μόνο από νουκλεϊνικά οξέα με απλές καταλυτικές
λειτουργίες. Οι πρώτες πρωτεΐνες ανέλαβαν αρχικά τη σταθεροποίηση της δομής του
RNA και σταδιακά λόγω της πολυμορφίας τους ανέλαβαν και την πληθώρα των
ενζυμικών αντιδράσεων.
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Σήμερα, διάφοροι μηχανισμοί έχουν προταθεί για να περιγράψουν την
κατάλυση στις περιπτώσεις των ριβοενζύμων, αν και αρχικά ο ρόλος των δισθενών
κατιόντων θεωρήθηκε σχεδόν ως ο μοναδικός. Η γνώση όμως λεπτομερειών για τις
περισσότερες κατηγορίες ριβοενζύμων, επιτρέπουν πλέον την ανάπτυξη και άλλων
μηχανιστικών προσεγγίσεων. Στα ιντρόνια της ομάδας Ι επικρατεί πράγματι ο
μηχανισμός των δύο μετάλλων. Ο ρόλος όμως του καταλύτη αποδίδεται στην ιστιδίνη
στην περίπτωση του ριβοενζύμου HDV. Για το σχηματισμό του πεπτιδικού δεσμού
στα ριβοσώματα καθοριστικός είναι ο ρόλος του υποστρώματος ενώ στην περίπτωση
του ριβοενζύμου glmS η χρήση καταλυτικού συμπαράγοντα.
Γενικά για τον καταλυτικό ρόλο των μορίων RNA
Τα ριβoένζυμα είναι μόρια RNA που καταλύουν μια χημική αντίδραση, π.χ.
τη διάσπαση ενός ομοιοπολικού δεσμού. Ο Cech και οι συνεργάτες του πρώτοι
περιέγραψαν ένα αυτοδιασπώμενο μόριο RNA στις μελέτες τους. Αυτό ήταν το
ιντρόνιο της ομάδας Ι (στο 413 νουκλεοτίδιο) από τον μικροοργανισμό Tetrahymena
thermophila. Μερικά χρόνια αργότερα, ο Zaug και οι συνεργάτες του περιέγραψαν
μια παραλλαγή του Τ. Thermophila που μπορούσε να δράσει in trans, (π.χ. ως μια
ειδική αλληλουχία ενδοριβονουκλεάσης που επιδρά επάνω σε άλλα υποστρώματα
RNA). Η εργασία των Foster και Symons σε ιούς φυτών RNA (virusoid RNAs)
πρότεινε μικρότερες RNA υπομονάδες που ήταν ικανές για καταλυτική
δραστηριότητα. Ο Uhlenbeck ήταν ο πρώτος που ανακάλυψε ένα μικρό δραστικό
ριβοένζυμο, το σφυροκέφαλο ριβοένζυμο (hammerhead ribozyme), στο οποίο η
καταλυτική υπομονάδα και η υπομονάδα που δρα ως υπόστρωμα ήταν
διαχωρισμένες. Τέτοια ριβοένζυμα δένονται με το υπόστρωμα RNA διαμέσου
αλληλεπιδράσεων ζευγαρώματος βάσεων, διασπούν το μόριο-στόχο RNA,
απελευθερώνουν το προϊόν διάσπασης και επανενώνονται έτσι ώστε να είναι ικανά να
επαναλάβουν τη διαδικασία για πολλές φορές. Οι Haseloff και Gerlach απαρίθμησαν
γενικούς κανόνες σχεδιασμού για απλά σφυροκέφαλα ριβοένζυμα, ικανά να δράσουν
in trans. Εξοπλισμένοι με αυτούς τους απλούς κανόνες, οι ειδικοί της Γενετικής
Μηχανικής ήταν έτοιμοι να αναπτύξουν τεχνικές γονιδιακής λειτουργίας in vivo. Μια
από τις μελέτες για την in vivo αποτελεσματικότητα των σφυροκέφαλων ριβοενζύμων
έδειξε 60% εξειδικευμένη μείωση της γενετικής έκφρασης σε κύτταρα πιθήκων. Για
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

να καταστεί επομένως η θεραπεία με ριβοένζυμα βιώσιμη, απαιτείται περαιτέρω
έρευνα. [1]
Η δραστικότητα των ριβοενζύμων
Για να έχουμε ένα ριβοένζυμο αποτελεσματικό σε in vivo συνθήκες,
χρειάζεται εκτεταμένη in vitro έρευνα με καλλιέργεια κυττάρων. Η κατανόηση που
προκύπτει από αυτές τις μελέτες χρησιμοποιείται έπειτα για να λάβουμε ενεργά
ριβοένζυμα σε φυσιολογικές συνθήκες με καλές φαρμακοκινητικές ιδιότητες. [1]
Τα ριβοένζυμα φαίνονται να υπακούουν σε κινητική του τύπου Michaelis-
Menton. Σε συνθήκες κορεσμού, ο ρυθμός διάσπασης καθορίζεται από το λόγο
Kcat/Km. Τα πιο αποτελεσματικά ριβοένζυμα εμφανίζουν λόγους με τιμές από 108 Μ-1
min-1. Ο λόγος Κcat/Km είναι συνήθως χαμηλός, όπου το Km και το Kcat έχουν χαμηλή
τιμή, υποδηλώνοντας υψηλή συγγένεια για τα υποστρώματά τους και αργή
απελευθέρωση των προϊόντων αντίστοιχα, με αποτέλεσμα τον γρήγορο κορεσμό του
καταλυτικού τους κέντρου [1]. Συμπερασματικά τα ριβοένζυμα είναι, συγκριτικά με
τα πρωτεϊνικά ένζυμα, πολύ πιο αργοί καταλύτες. Αυτά όμως τα κινητικά
χαρακτηριστικά ταιριάζουν με τη λειτουργία των ριβοενζύμων, η οποία για τις
περισσότερες περιπτώσεις είναι η κατάλυση μιας αντίδρασης ενός κύκλου (single
turnover reaction), όπως είναι η αποκοπή τους από ένα μεγαλύτερο μόριο. Τα
ριβοένζυμα έχουν αποτελέσει ελκυστικό ερευνητικό εργαλείο στην ιατρική βιοχημεία
για θεραπεία γενετικών ασθενειών με τη μέθοδο της γονιδιακής θεραπείας
(Kalavrizioti et al., 2006). [6]
Η παρατήρηση και η μελέτη της δραστικότητας των ριβοενζύμων in vitro,
είναι σχετικά απλή: η επώαση για ένα ριβοένζυμο και το υπόστρωμά του
ακολουθείται από ανάλυση με ηλεκτροφόρηση σε gel πολυακρυλαμιδίου, η οποία
μας επιτρέπει την ανίχνευση των προϊόντων διάσπασης. Η μελέτη της δραστικότητας
των ριβοενζύμων στα κύτταρα είναι πιο πολύπλοκη. Πρώτον, απαιτείται ένα μέσο
που θα εισάγει τα ριβοένζυμα μέσα στα κύτταρα. Αυτό συνήθως επιτυγχάνεται με
έναν από τους παρακάτω τρεις τρόπους: εισαγωγή των προσχηματισμένων
ριβοενζύμων με μικροέγχυση ή με κατιονικά λιποσώματα, εισαγωγή με μετάδοση
μέσω πλασμιδίων ή εισαγωγή διαμέσου επιμόλυνσης με ρετροϊό που φέρει το
ριβοένζυμο. Δεύτερον, τα προϊόντα διάσπασης που παράγονται από τα ριβοένζυμα
συνήθως δεν ανιχνεύονται μέσα στα κύτταρα. Αυτό πιθανώς οφείλεται στη
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

δραστικότητα των διαμοριακών ενδονουκλεασών που γρήγορα αποσυνθέτουν το
διασπασμένο υπόστρωμα RNA. Για αυτό, άλλα παράπλευρα γεγονότα
παρουσιάζονται συνήθως και δίνουν πληροφορίες για τη δραστικότητα των
ριβοενζύμων. [1]
Σε κλινικές δοκιμές χρησιμοποιήθηκαν ριβοένζυμα σε ασθενείς με λοιμώδη
νοσήματα ή καρκίνο. Σε αυτές τις μελέτες, τα ριβοένζυμα χορηγήθηκαν στον ασθενή
είτε διαμέσου μεθόδων γονιδιακής θεραπείας είτε με απευθείας έγχυση με συνθετικά
ριβοένζυμα. [1]
Η κατάλυση από μόρια RΝA έχει ανακαλυφθεί εδώ και είκοσι πέντε περίπου
χρόνια. Οι ανακαλύψεις ότι συγκεκριμένα ιντρόνια ήταν ικανά να αυτοδιασπώνται
(self-splicing ή εκτομή RNA) και ότι τμήμα του RNA της βακτηριακής
ριβονουκλεάσης P (RNase P) από μόνο του μπορούσε να δώσει πρόδρομα tRNAs,
ήταν οι πρώτες ενδείξεις ότι υπολείμματα μορίων RNA με καταλυτική δράση
υπάρχουν μέχρι σήμερα. Από το 1980, οι καταλυτικές δυνατότητες του RNA είχαν
αποκαλυφθεί με την ανακάλυψη των αποκαλούμενων μικρών νουκλεολυτικών
ριβοενζύμων (small nucleolytic ribozymes or RNA-based enzymes). Η οικογένεια
αυτή αποτελείται από τέσσερα μέλη: το σφυροκέφαλο (hammerhead) ριβοένζυμο, το
ριβοένζυμο φουρκέτα (hairpin ribozyme), το ριβοένζυμο του ιού της ηπατίτιδας δ
(hepatitis delta virus, HDV) και το ριβοένζυμο Neurospora crassa Varkud satellite
(VS). Όλα τα μικρά νουκλεολυτικά ριβοένζυμα εμπλέκονται στη διαδικασία της
RNA αντιγραφής και καταλύουν ως ενδιάμεσα μια απλή αντίδραση RNA
διαχωρισμού ή συνένωσης νουκλεοτιδίων (ligation reaction). [2]
Τα περισσότερα γνωστά ριβοένζυμα έχουν, όπως και το υπόστρωμά τους,
τους συμβατικούς 3΄,5΄-φωσφοδιεστερικούς δεσμούς στο RNA. Σε μια από τις
απλούστερες αντιδράσεις, μια περιοχή του RNA της RNase P καταλύει την υδρόλυση
του πρόδρομου tRNAs (Σχήμα 1a). Περισσότερο συχνά ωστόσο, τα ριβoένζυμα
καταλύουν αντιδράσεις τρανσεστεροποίησης (transesterification), όπως κάνουν και
τα μικρά νουκλεολυτικά ριβοένζυμα (Σχήμα 1b) και τα αυτοδιασπώμενα ιντρόνια
(Σχήμα 1c,d). Τα μικρά νουκλεολυτικά ριβοένζυμα καταλύουν την ενός βήματος
διάσπαση του 3΄,5΄-φωσφοδιεστερικού δεσμού, με το σχηματισμό δύο τμημάτων
RNA που το ένα έχει το 2΄,3΄- κυκλικό φωσφοδιεστερικό άκρο και το άλλο ένα 5΄-
υδροξύλιο στα προϊόντα διάσπασης (Σχήμα 1b). Παρά το ότι έχουν τον ίδιο
μηχανισμό, τα μικρά νουκλεολυτικά ριβoένζυμα διαφέρουν έντονα μεταξύ τους στη
δομή, στην καταλυτική τους δραστικότητα σε διάφορες τιμές pH και στα
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

απαιτούμενα μεταλλικά ιόντα για την κατάλυση. Φαίνεται ο μηχανισμός αυτός να
είναι ο πιο κατάλληλος για τη διάσπαση του RNA, όπως συμβαίνει κατά τη διάσπαση
των πολυμερών σε μονομερή. Άλλα μόρια που χρησιμοποιούν τον μηχανισμό αυτό
είναι τα in vitro επιλεγμένα leadzyme, η πρωτεϊνική RNase A και το προσφάτως
ανακαλυφθέν riboswitch glmS, ένα στοιχείο RNA που ελέγχει τη γενετική έκφραση
δια μέσου της καταλυτικής του δραστικότητας. [2]
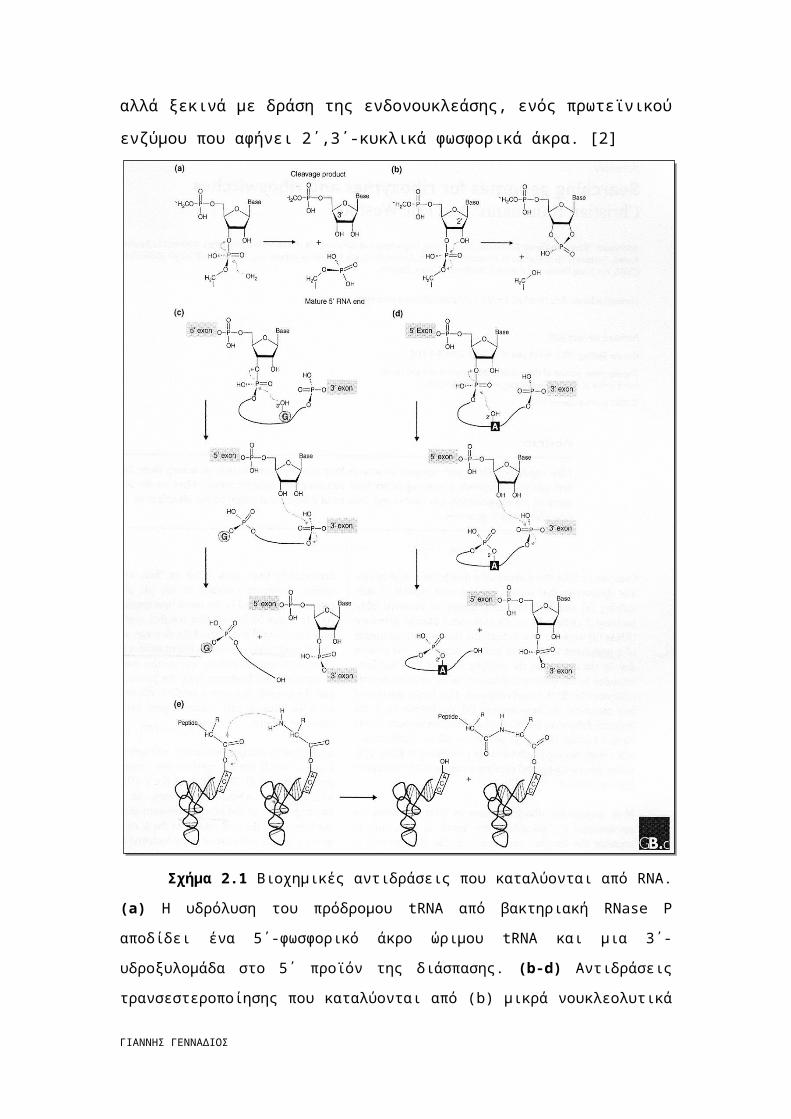
Σε αντίθεση με αυτή την απλή αντίδραση, η αυτοδιάσπαση των ιντρονίων της
ομάδας Ι και της ομάδας ΙΙ περιλαμβάνει δύο διαδοχικά στάδια (σχήμα 1c,d). Το
πρώτο ελευθερώνει το 3΄-OH από το 5΄ εξώνιο, το οποίο επιτρέπει σε ένα δεύτερο
στάδιο, την προσβολή στη συμβολή του φωσφοδιεστερικού δεσμού μεταξύ του
τελευταίου υπολείμματος του ιντρονίου και του πρώτου υπολείμματος του 3΄
εξωνίου. Τα αυτοδιασπώμενα ιντρόνια της ομάδας Ι χρησιμοποιούν το 3΄-υδροξύλιο
μιας εξωγενούς γουανοσίνης ως το αρχικά αντιδρών πυρηνόφιλο. Η γουανοσίνη
φωσφορυλιώνεται κατά την αντίδραση και ελευθερώνεται (Σχήμα 1c). Στα
αυτοδιασπώμενα ιντρόνια της ομάδας Ι, ο σχηματισμός ενός ενδιάμεσου με 2΄,3΄-
κυκλικό φωσφοδιεστερικό δεσμό δεν έχει παρατηρηθεί, πιθανώς επειδή το ενδιάμεσο
αυτό συνεπάγεται απώλεια της δομικής ακεραιότητας στα τετμημένα εξώνια με το
σχηματισμό 2΄,5΄-φωσφοδιεστερικής σύνδεσης στο δεύτερο στάδιο της αντίδρασης.
Μια παρόμοια στρατηγική δύο σταδίων υιοθετείται από τα αυτοδιασπώμενα ιντρόνια
της ομάδας ΙΙ, αλλά σε αυτή την περίπτωση το προσβάλλον πυρηνόφιλο είναι το 2΄-
υδροξύλιο της συντηρημένης αδενοσίνης της κλαδικής περιοχής (σχήμα 1d). Ενώ
σχηματίζεται μια RNA θηλιά στο ιντρόνιο, η δομική ακεραιότητα των συνδεδεμένων
εξωνίων εξασφαλίζεται. Θα πρέπει να σημειωθεί ότι η διάσπαση των tRNA ιντρονίων
στους μικροοργανισμούς Eukarya και Archaea δεν προέρχεται από την αυτοδιάσπαση
όπως στα βακτήρια, αλλά ξεκινά με δράση της ενδονουκλεάσης, ενός πρωτεϊνικού
ενζύμου που αφήνει 2΄,3΄-κυκλικά φωσφορικά άκρα. [2]
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Σχήμα 2.1 Βιοχημικές αντιδράσεις που καταλύονται από RNA. (a) Η υδρόλυση του
πρόδρομου tRNA από βακτηριακή RNase P αποδίδει ένα 5΄-φωσφορικό άκρο ώριμου tRNA
και μια 3΄-υδροξυλομάδα στο 5΄ προϊόν της διάσπασης. (b-d) Αντιδράσεις
τρανσεστεροποίησης που καταλύονται από (b) μικρά νουκλεολυτικά ριβοένζυμα, (c) ιντρόνια
της ομάδας Ι, (d) ιντρόνια της ομάδας ΙΙ στα οποία διαφορετικές χαρακτηριστικές ομάδες
παίζουν το ρόλο του πυρηνόφιλου. Στα μικρά νουκλεολυτικά ριβοένζυμα (b), το
καθορισμένο 2΄-υδροξύλιο προσβάλλει τον γειτονικό 3΄,5΄-φωσφοδιεστερικό δεσμό,
οδηγώντας σε μια 2΄,3΄-κυκλική φωσφορική ομάδα και σε ένα 5΄-υδροξύλιο. Στο πρώτο
βήμα της διάσπασης των ιντρονίων της ομάδας Ι (c), το 3΄-υδροξύλιο του συμπαράγοντα της
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

εξωγενούς γουανοσίνης (G) προσβάλει τη συμβολή του 5΄-εξωνίου με το ιντρόνιο και
ελευθερώνει το 5΄ εξώνιο, το οποίο οδηγεί στην πρόσδεση του συμπαράγοντα στο 5΄ άκρο
του ιντρονίου. Σε μια δεύτερη αντίδραση τρανσεστεροποίησης, το 5΄ εξώνιο σχηματίζει έναν
συμβατικό 3΄,5΄ δεσμό με το 3΄ εξώνιο, απελευθερώνοντας το γραμμικό ιντρόνιο με την
επιπρόσθετη γουανοσίνη. Στα ιντρόνια της ομάδας ΙΙ (d), η αδενοσίνη συμπεριφέρεται ως
πυρηνόφιλο, οδηγώντας στο σχηματισμό ιντρονίου με μορφή θηλειάς. (e) Σχηματισμός
πεπτιδικού δεσμού που καταλύεται από ριβόσωμα.
Η συντήρηση στον κόσμο των μορίων RNA έχει επιβεβαιωθεί από την
αποκάλυψη ότι το ριβόσωμα είναι ένα ριβοένζυμο. Στοιχεία δηλαδή του ριβοσωμικού
RNA έχουν καταλυτική δραστηριότητα στην πολυπεπτιδική σύνθεση, τοποθετώντας
την ενεργότητα των ριβοενζύμων στην καρδιά του σημερινού κυττάρου και
αποδεικνύοντας ότι τα ριβοένζυμα μπορούν να καταλύσουν αντιδράσεις πέρα από τη
διάσπαση και το σχηματισμό φωσφοδιεστερικών δεσμών στο RNA (Σχήμα 1e). Η
πρώτη ένδειξη καταλυτικού RNA στο ριβόσωμα προήλθε από βιοχημικά δεδομένα
που έδειξαν διατήρηση της καταλυτικής ενεργότητας μετά την αποδιάταξη των
πρωτεϊνών τους. Η τελική απόδειξη ότι το RNA είναι καταλύτης σε πρωτεϊνική
βιοσύνθεση προήλθε από κρυσταλλογραφική έρευνα που έδειξε ότι το ενεργό κέντρο
της πεπτυδιλοτρανσφεράσης στο ριβόσωμα στερείται κάθε πρωτεϊνικού στοιχείου και
είναι δομημένο αποκλειστικά από υπολείμματα rRNA. [2]
Στα προηγούμενα χρόνια, ένας αριθμός νέων καταλυτικών RNA μορίων είχαν
ανακαλυφθεί, συμπεριλαμβανομένων των καταλυτικών μορίων riboswitch και
γνωστών στοιχείων που έχουν ανιχνευθεί σε περιοχές του γονιδιώματος. Στον πίνακα
1 απαριθμούνται όλα τα γνωστά καταλυτικά RNAs. Σήμερα όμως γνωρίζουμε όλο το
φάσμα των καταλυτικών μορίων RNA ή μένουν πολλά ακόμα να ανακαλύψουμε; Για
να λυθεί το πρόβλημα αυτό, νέα εργαλεία της βιοπληροφορικής που θα
ολοκληρώσουν τις τρέχουσες γνώσεις μας για τη δομή του RNA, απαιτούνται να
αναπτυχθούν. [2]
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Πίνακας 2.1 Τα σημερινά ριβοένζυμα και τα riboswitches.
Για να επιτευχθεί μια χημική RNA κατάλυση, μερικές περιοχές από το μόριο
του ριβοενζύμου πρέπει να υιοθετήσουν πολύ ακριβείς σχετικές θέσεις και
προσανατολισμούς. Εκτός από την ειδική αναγνώριση, πρέπει να υπάρχουν δυναμικοί
μηχανισμοί για την πρόσδεση με το υπόστρωμα και την απελευθέρωση του
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

προϊόντος. Με την αξιοσημείωτη εξαίρεση του ριβοσώματος, τα ριβοένζυμα δρουν
επάνω στον φωσφοδιεστερικό σκελετό, συνδέοντας δυο διαδοχικά νουκλεοτίδια.
Παρότι οι καταλυτικές διαδικασίες τέτοιων αντιδράσεων είναι βασικά ίδιες, μπορούν
να επιτευχθούν με πολλούς και διαφορετικούς τρόπους και επιπλέον εάν η σύγκλιση
είναι εύκολη, τα ριβοένζυμα παρουσιάζουν μια ποικιλία από κατάλληλες θέσεις ως
αντιδρώντα. [2]
Επιπλέον, ο αριθμός των συντηρημένων νουκλεοτιδίων και η διασπορά τους
από μόριο σε μόριο διαφέρει σημαντικά. Για παράδειγμα, το σφυροκέφαλο
ριβοένζυμο και τα ιντρόνια της ομάδας Ι έχουν περίπου τον ίδιο αριθμό
συντηρημένων υπολειμματικών περιοχών - περίπου 7 - παρόλο που το δεύτερο είναι
πάνω από τέσσερις φορές μεγαλύτερο. Η θέση και η σχετική διάταξη των
συντηρημένων δομικών στοιχείων σε σχέση με την αρχή και το τέλος του
ριβοενζύμου, επίσης διαφέρουν (Σχήμα 2.2). Οι περισσότερες οικογένειες
ριβοενζύμων μπορούν να υποδιαιρεθούν σε κατηγορίες με βάση τα μη ομόλογα
περιφερειακά τους χαρακτηριστικά στοιχεία. Ωστόσο, ο πυρήνας της τρισδιάστατης
μορφής ενός ριβοενζύμου που ανήκει στην ίδια οικογένεια αναμένεται να είναι ίδιος
επειδή αυτά διατηρούνται εξαιτίας περιορισμών στην τριτοταγή δομή, οι οποίες παρά
τη συντήρηση μικρών αλληλουχιών, μπορούν να δώσουν σχήματα με διάφορους
τρόπους. [2]
Σχήμα 2.2 Τα σφυροκέφαλα ριβοένζυμα βασίζουν τη δομή τους σε τρεις τρόπους
συνδέσεων και διακρίνονται σε δύο κύριους τύπους. (a) Ο τύπος Ι έχει τα άκρα της απλής
αλυσίδας RNA στη θηλιά Ι. (b) Ο τύπος ΙΙΙ έχει τα άκρα του μονόκλωνου RNA στη θηλιά ΙΙΙ.
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Για λόγους που δεν είναι μέχρι σήμερα γνωστοί, ριβοένζυμα του τύπου ΙΙ (με τα μονόκλωνα
άκρα σε θηλιά ΙΙ) δεν έχουν παρατηρηθεί.
Το σφυροκέφαλο ριβοένζυμο απεικονίζει ανάγλυφα της δυσκολίες στην
αναγνώριση νέων ριβοενζύμων είτε πειραματικά, είτε μέσω in silico προσεγγίσεων.
Πράγματι, μια ατελής καταλυτική RNA αναδίπλωση, η οποία δεν περιλαμβάνει
επαφές μεταξύ στοιχείων μακριά από καταλυτικές περιοχές, ήταν αποδεχτή για πολύ
καιρό, μέχρι να αποκαλυφθεί ολόκληρη η δομή του σφυροκέφαλου ριβοενζύμου. Μια
πρόσφατη κρυσταλλική δομή, δείχνει πως η παρουσία τριτοταγών επαφών μεταξύ
των θηλειών, πολύ απομακρυσμένων από τις συντηρημένες καταλυτικές περιοχές,
επάγει δομικές αλλαγές στον πυρήνα που προωθούν την ενεργή κατάσταση του
ριβοενζύμου. Κατά ένα πολύ σημαντικό τρόπο, όλες αυτές οι επαφές περιλαμβάνουν
ένα δίκτυο ζευγαρωμένων βάσεων που δεν ακολουθεί το κλασσικό πρότυπο της
διπλής έλικας (non-Watson-Crick base pairing). Για να είναι πλήρως βιολογικά
ενεργά τα σφυροκέφαλα ριβοένζυμα, απαιτούν δομική ακρίβεια και αυστηρές
προδιαγραφές στην αλληλουχία τους (σχήμα 2.3b). Εξαιτίας όμως του τρόπου
ζευγαρώματος κατά non-Watson-Crick αυτό δεν καθίσταται άμεσα εμφανές μόνο από
την αλληλουχία τους. Σε αντίθεση, εξαιτίας της κουλουριασμένης δομής τους που
βασίζεται σε κατά Watson-Crick ζευγάρωμα, τα ριβοένζυμα HDV αποκαλύπτουν
άμεσα την πολυπλοκότητα της δομής τους (σχήμα 2.3a). Ατελή σφυροκέφαλα
ριβοένζυμα χωρίς περιφερειακά στοιχεία και με μικρή πολυπλοκότητα στην
αλληλουχία και στη δομή τους παρουσιάζουν μειωμένη καταλυτική δραστηριότητα.
[2]
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Σχήμα 2.3 Διάγραμμα του δικτύου αλληλεπιδράσεων που συμβάλλει στη διατήρηση
της τρισδιάστατης δομής δύο διαφορετικών ριβοενζύμων. (a) Το HDV ριβοένζυμο και (b) το
σφυροκέφαλο ριβοένζυμο. To HDV ριβοένζυμο έχει μια δομή κουλουριασμένη (convoluted
pseudoknotted topology). Κάθε νουκλεοτίδιο έχει τρεις θέσεις υποψήφιες για δεσμό
υδρογόνου, τη θέση Watson-Crick (υποδεικνύεται με κύκλο), τη θέση Hoogsteen
(υποδεικνύεται με τετράγωνο) και την υποδοχή στη θέση του σακχάρου (υποδεικνύεται με
τρίγωνο). Η αλληλεπίδραση μεταξύ βάσεων μπορεί να λάβει χώρα είτε μέσω σακχάρου από
την πλευρά της γραμμής προσέγγισης (cis διαμόρφωση, κλειστό σύμβολο) ή μέσω σακχάρων
στις εκατέρωθεν πλευρές της γραμμής προσέγγισης (trans-διαμόρφωση, ανοιχτό σύμβολο).
Όταν μνημονεύουμε τριτοταγείς επαφές, τα νουκλεοτίδια που εμπλέκονται βρίσκονται μέσα
σε τετράγωνα πλαίσια. Όταν η βάση του νουκλεοτιδίου έχει syn-διαμόρφωση σε σχέση με το
σάκχαρο σημειώνεται με Bold. Τα ορθογώνια παρουσιάζουν την ακριβή θέση που κατέχει
στο χώρο κάθε νουκλεοτίδιο.
Πράγματι, σε πειραματικές διαδικασίες που εξελίσσονται in vitro, ξεκινώντας
από τυχαίες βιβλιοθήκες, έχουν παραχθεί ποικίλα δομικά ριβοένζυμα με χαμηλή
δραστηριότητα, τα οποία περιέχουν μερικές παραλλαγές του τύπου των
σφυροκέφαλων. Άλλα in vitro πειράματα απέδειξαν ένα πρότυπο αυτοδιάσπασης με
βιοχημική δραστηριότητα τύπου σφυροκέφαλου ριβοενζύμου και οδήγησαν στο
συμπέρασμα ότι αυτό το πρότυπο κάνει το πιο κοινό ριβοένζυμο να αναδιπλώνεται.
Προτείνει επίσης ότι αυτό το πρότυπο είχε πολλαπλές και ανεξάρτητες μεταξύ τους
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

καταγωγές. Η μεγάλη εύρους διαδραστικότητα, δε θεωρήθηκε στην εποχή των δυο in
vitro πειραμάτων ως ένα σχήμα επιλογής, καθώς η σημασία τους δεν είχε
αναγνωριστεί ως σημαντική μέχρι εκείνη τη στιγμή. [2]
Η σημασία των ριβοενζύμων στην επιστήμη της εξέλιξης
Η ανακάλυψη των ριβοενζύμων έφερε ξανά στην επικαιρότητα ένα πρόβλημα
που για πολλά χρόνια ταλανίζει την επιστημονική κοινότητα. Ποιο ήταν το πρώτο
πληροφοριακό μόριο κατά τα αρχικά στάδια της εξέλιξης; Το DNA ή τα ένζυμα; Το
ερώτημα δημιουργείται επειδή δε φαίνεται να υπάρχει επικάλυψη στις λειτουργίες
που επιτελούν αυτά τα δύο μόρια, αφού το ένα αποθηκεύει τη γενετική πληροφορία,
ενώ το άλλο μπορεί να καταλύει αντιδράσεις. Πως όμως έγινε η σύνθεση DNA χωρίς
ένζυμα ή αντίστροφα πως έγινε η σύνθεση ενζύμων χωρίς DNA; [3]
Μια απάντηση θα μπορούσε να είναι ότι το RNA επιτελεί και τις δυο
λειτουργίες, αφού μπορεί να είναι και γενετικό υλικό κα να καταλύει αντιδράσεις,
άρα μπορεί να είναι το πληροφοριακό μόριο που εμφανίστηκε πρώτο εξελικτικά.
Αργότερα οι λειτουργίες αυτές μοιράστηκαν στο DNA και στις πρωτεΐνες και
απέμεινε στο RNA η λειτουργία του συνδέσμου μεταξύ των δύο μορίων κατά τη ροή
της γενετικής πληροφορίας. [3]
Αυτές οι σκέψεις μας επιτρέπουν να φανταζόμαστε ένα κόσμο RNA στην
αρχή της ζωής, πριν την εμφάνιση του DNA και των πρωτεϊνών. Ο Walter Gilbert
πρότεινε ότι τα μόρια RNA πρώτα κατέλυσαν τη δική τους αντιγραφή και ανέπτυξαν
ένα ρεπερτόριο ενζυμικής δραστηριότητας. Σε δεύτερο στάδιο το RNA άρχισε να
συνθέτει πρωτεΐνες που εμφανίστηκαν ως καλύτερης ποιότητας ένζυμα. Τελικά το
DNA σχηματίστηκε από την αντίστροφη μεταγραφή του RNA και αντικατέστησε το
RNA ως γενετικό υλικό διότι η δομή της διπλής έλικας αποτελεί καλύτερο, δηλαδή
πιο σταθερό μέσο αποθήκευσης της γενετικής πληροφορίας από ότι τα μονόκλωνα
μόρια του RNA. Έτσι, το RNA έμεινε με τους ρόλους που διατηρεί μέχρι σήμερα. [4]
Ο καταλυτικός ρόλος των ριβοενζύμων και των ριβοσωμάτων
Ιντρόνια της ομάδας Ι
Η διάσπαση του RNA περιλαμβάνει δύο αντιδράσεις μεταφοράς
φωσφορυλίου που καταλύονται και οι δύο από το ίδιο το ιντρόνιο (σχήμα 2.4). Στην
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

πρώτη αντίδραση, μια εξωγενής γουανίνη προσβάλλει το 5΄ άκρο για να ελευθερώσει
το 5΄ εξώνιο από το ιντρόνιο. Σε μια δεύτερη αντίδραση, το 5΄ εξώνιο προσβάλλει το
3΄ άκρο στη φωσφορική ομάδα στην τελευταία γουανίνη του ιντρονίου για να
δημιουργηθεί το συνδεδεμένο πλέον εξώνιο και να ελευθερωθεί το ιντρόνιο. Η
δεύτερη αντίδραση είναι ουσιαστικά η αντίστροφη της πρώτης. Διαφέρει μόνο στο
ότι η εξωγενής γουανίνη είναι συνδεδεμένη στο ενεργό άκρο. Ένα νεότερο μοντέλο
για την κατάλυση των ιντρονίων της ομάδας Ι προτάθηκε από τους Steitz και Steitz
βασισμένο στο ανάλογο των πρωτεϊνικών ενζύμων που καταλύουν τη μεταφορά
φωσφορυλίου. Η πρότασή τους περιλαμβάνει μηχανισμό βασισμένο σε δισθενή
κατιόντα. [5]
Η δομή του ιντρονίων της ομάδας Ι έγινε γνωστή με κρυσταλλογραφία
ακτίνων X. Οι δομές από τρία διαφορετικά ιντρόνια σε τρεις διαφορετικές
αντιδράσεις αναφέρθηκαν το 2004. Αυτές μας έδωσαν τρισδιάστατες διαστάσεις των
ιντρονίων της ομάδας Ι, αλλά από το RNΑ, σε κάθε μια από αυτές τις δομές, έλειπε
τουλάχιστον μια ενεργή πλευρική λειτουργική ομάδα που είναι γνωστό ότι συνεργεί
με τα μεταλλικά ιόντα. Η δομή του καταλυτικού συμπλόκου, συμπεριλαμβανομένου
του ιντρονίου, των δύο εξωνίων και των μεταλλικών συναρμοστών, αναφέρθηκε ένα
χρόνο αργότερα. Η συνολική δομή ήταν η ίδια με αυτή των προηγούμενων αναφορών
αλλά παρατηρήθηκαν διαφορές στο ενεργό κέντρο. [5]
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

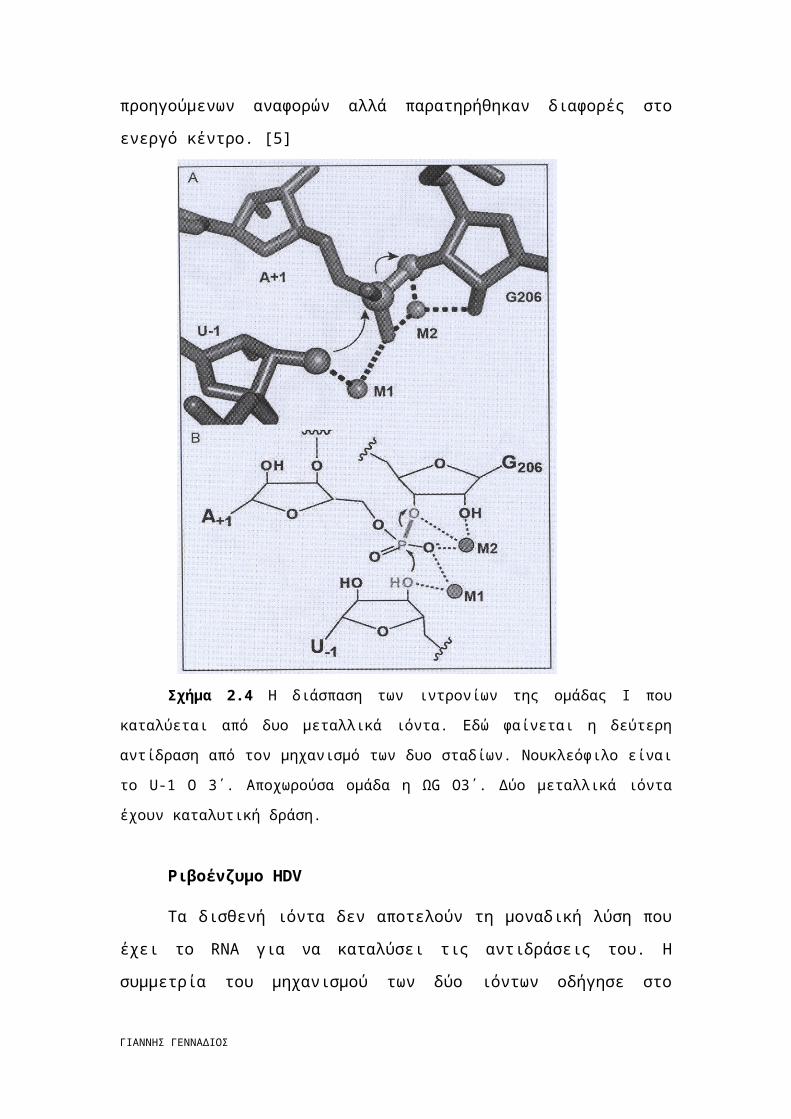
Σχήμα 2.4 Η διάσπαση των ιντρονίων της ομάδας Ι που καταλύεται από δυο
μεταλλικά ιόντα. Εδώ φαίνεται η δεύτερη αντίδραση από τον μηχανισμό των δυο σταδίων.
Νουκλεόφιλο είναι το U-1 O 3΄. Αποχωρούσα ομάδα η ΩG O3΄. Δύο μεταλλικά ιόντα έχουν
καταλυτική δράση.
Ριβοένζυμο HDV
Τα δισθενή ιόντα δεν αποτελούν τη μοναδική λύση που έχει το RNA για να
καταλύσει τις αντιδράσεις του. Η συμμετρία του μηχανισμού των δύο ιόντων
οδήγησε στο πρόωρο συμπέρασμα ότι όλα τα ριβοένζυμα είναι μεταλλοένζυμα.
Σήμερα βέβαια γνωρίζουμε ότι αυτό δεν είναι αλήθεια. Για παράδειγμα, η
ριβονουκλεάση Α συναρμόζει δύο μόρια ιστιδίνης ως καταλύτες για να
ενεργοποιήσει το πυρηνόφιλο και να σταθεροποιήσει την αποχωρούσα ομάδα κατά
της διάρκεια της εκτομής του RNA. Η ιστιδίνη έχει pKa 6,8. Αποτελεί επομένως
ιδανικό μόριο για αυτούς τους ρόλους σε φυσιολογικές τιμές pH. [5]
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

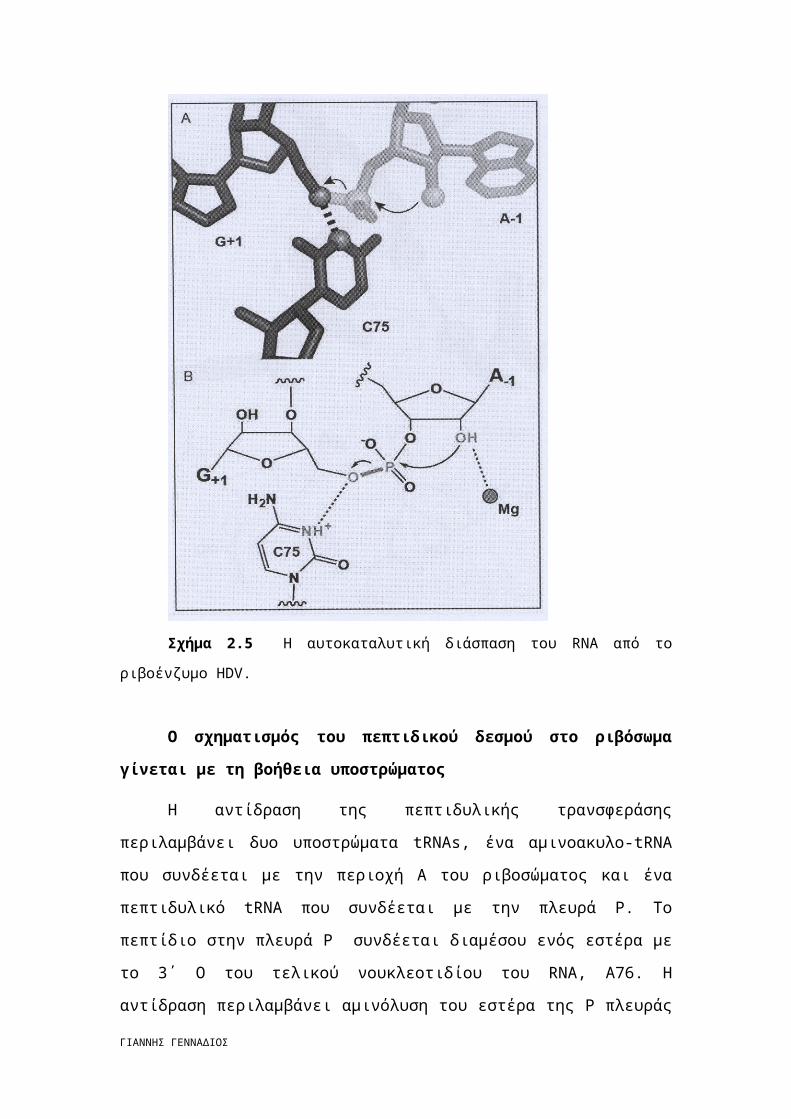
Το HDV ριβοένζυμο είναι ένα μικρό αυτο-νουκλεολυτικό RNA το οποίο έχει
την ίδια χημική συμπεριφορά με τα άλλα νουκλεολυτικά ριβοένζυμα. Το 2΄-
υδροξύλιο δρα ως πυρηνόφιλο για να αντικαταστήσει το Ο 5΄ οδηγώντας σε δύο
προϊόντα RNA, ένα με 2΄,3΄-κυκλικό φωσφορικό άκρο και ένα με 5΄-τελικό
υδροξύλιο (σχήμα 2.5). Αυτή είναι η ίδια χημική αντίδραση που καταλύεται από τη
ριβονουκλεάση Α. Η πρώτη κρυσταλλική δομή του HDV από ένα προϊόν που
σχηματίστηκε από ριβοένζυμο, έδειξε ότι το 5΄-υδροξύλιο ήταν σε τέτοια απόσταση
ώστε να σχηματίζει δεσμό υδρογόνου με το Ν3 του C75, ένα σταθερό υπόλειμμα που
δίνει αποτελεσματικές διασπάσεις. Ο C75 μπορεί να λειτουργήσει ως οξύ που μπορεί
να πρωτονιώσει την O5΄ αποχωρούσα ομάδα. [5]
Σχήμα 2.5 Η αυτοκαταλυτική διάσπαση του RNA από το ριβοένζυμο HDV.
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ
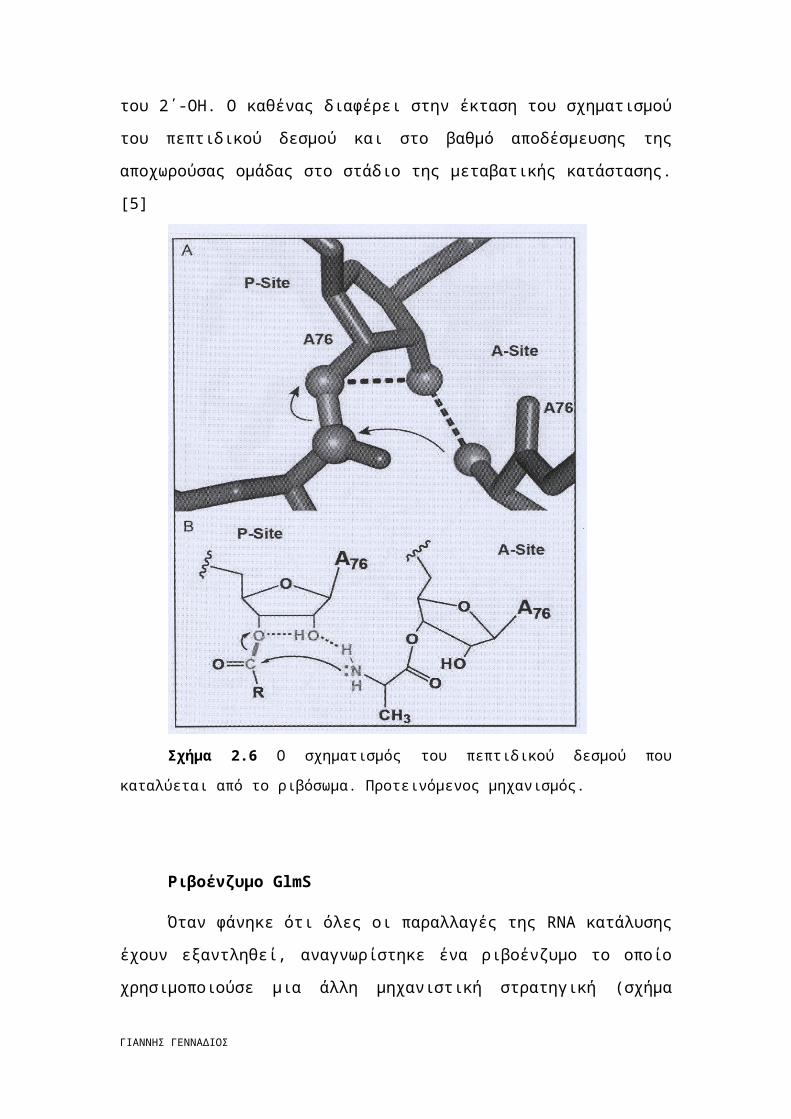
Ο σχηματισμός του πεπτιδικού δεσμού στο ριβόσωμα γίνεται με τη
βοήθεια υποστρώματος
Η αντίδραση της πεπτιδυλικής τρανσφεράσης περιλαμβάνει δυο υποστρώματα
tRNAs, ένα αμινοακυλο-tRNA που συνδέεται με την περιοχή Α του ριβοσώματος και
ένα πεπτιδυλικό tRNA που συνδέεται με την πλευρά P. To πεπτίδιο στην πλευρά P
συνδέεται διαμέσου ενός εστέρα με το 3΄ O του τελικού νουκλεοτιδίου του RNA,
A76. H αντίδραση περιλαμβάνει αμινόλυση του εστέρα της P πλευράς από μια
αμινοομάδα της Α-πλευράς. Το ριβόσωμα ευθυγραμμίζει τα υποστρώματα με τέτοιον
τρόπο ώστε η αμίνη να προσεγγίζει τον εστέρα οδηγώντας σε μια μεταβατική
κατάσταση χειλικής μορφής με στερεοχημεία S. [5]
Το Α76 2΄-ΟΗ δίπλα στην πλευρά P παίζει ουσιαστικό ρόλο στην αντίδραση.
Σημειακή μετάλλαξη σε αυτή τη λειτουργική ομάδα οδηγεί σε πλήρη απώλεια της
δραστικότητας της πεπτιδυλικής τρανσφεράσης. Χωρίς τη συγκεκριμένη
υδροξυλομάδα, ο ρυθμός υδρόλυσης του εστέρα είναι ταχύτερος από το ρυθμό
σχηματισμού του πεπτιδικού δεσμού. Η συνεισφορά αυτή είναι μεγαλύτερη από την
αντίστοιχη κάθε rRNA αν και το 2΄-ΟΗ του Α2451 έχει επίσης αποδειχτεί ότι
συμβάλει στην πραγματοποίηση της αντίδρασης. [5]
Όσον αφορά στο μηχανισμό μεταφοράς πρωτονίου, οι ομάδες υδροξυλίου
μάλλον δεν είναι ικανές να εμπλακούν σε ένα τέτοιο ρόλο. Με τιμή pΚa περίπου 12,
το 2΄-ΟΗ είναι λιγότερο πιθανό από ότι οι βάσεις των νουκλεοτιδίων να εμπλέκεται
άμεσα στη μεταφορά πρωτονίου, ακόμη και εάν προσπαθήσουμε να ρυθμίσουμε την
τιμή του pH. [5]
Η θέση του ριβοσώματος έχει μεγάλη σημασία για το Α76 2΄-ΟΗ μεταξύ του
α-αμινο πυρηνόφιλου και της 3΄-ΟΗ αποχωρούσας ομάδας (σχήμα 2.6). Καθώς η
αμίνη πρέπει να χάσει ένα πρωτόνιο και το Ο3΄ πρέπει να κερδίσει ένα, έχουν
προταθεί μηχανισμοί που περιλαμβάνουν μεταφορά πρωτονίου από την α-αμίνη του
Α76 Ο3΄ διαμέσου του Α76 2΄-ΟΗ. Οι μηχανισμοί αυτοί εξηγούν την ανάγκη για το
ρόλο του 2΄-ΟΗ. Ο καθένας διαφέρει στην έκταση του σχηματισμού του πεπτιδικού
δεσμού και στο βαθμό αποδέσμευσης της αποχωρούσας ομάδας στο στάδιο της
μεταβατικής κατάστασης. [5]
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Σχήμα 2.6 Ο σχηματισμός του πεπτιδικού δεσμού που καταλύεται από το ριβόσωμα.
Προτεινόμενος μηχανισμός.
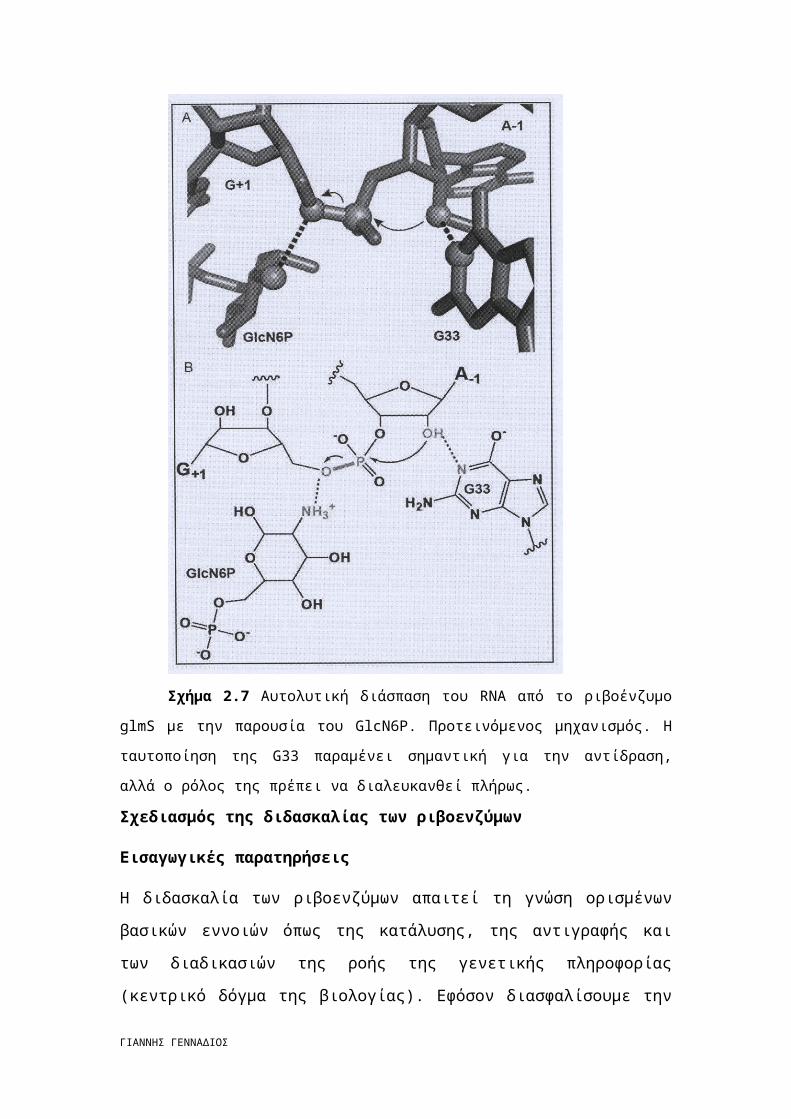
Ριβοένζυμο GlmS
Όταν φάνηκε ότι όλες οι παραλλαγές της RNA κατάλυσης έχουν εξαντληθεί,
αναγνωρίστηκε ένα ριβοένζυμο το οποίο χρησιμοποιούσε μια άλλη μηχανιστική
στρατηγική (σχήμα 2.7). Το glmS riboswitch είναι τοποθετημένο στην 5΄-
αμετάφραστη περιοχή του γονιδίου που κωδικοποιεί την ένωση glycosamine-6-
phosphate (GlcN6P) synthetase. Η παρουσία της GlcN6P αρκεί για να διασπάσει το
δικό της mRNA, ρυθμίζοντας έτσι τη μειωμένη σύνθεση της συνθετάσης. Η GlcN6P
είναι απαραίτητη για μια αποτελεσματική αντίδραση διάσπασης, αν και άλλες
πρωτοταγείς αμίνες με γειτονικά υδροξύλια μπορούν να ενεργοποιήσουν το
ριβοένζυμο in vitro. H πρωτοταγής αμίνη είναι απαραίτητη για την αντίδραση επειδή
η υποκατάσταση της αμίνης με ένα υδροξύλιο οδηγεί στην οριστική απώλεια της
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

δραστικότητας. Η GlcN6P είναι ακόμη ικανή να προσδεθεί με το ριβοένζυμο αλλά
λειτουργεί ως συναγωνιστικός παρεμποδιστής. [5]
Πώς η GlcN6P δρα ως καταλυτικός συμπαράγοντας; Έχει προταθεί ότι η
αμίνη κατέχει το ρόλο της βάσης που αποπρωτονιώνει το Ο2΄ πυρηνόφιλο. Αν και το
υδροξύλιο του C2 δεν είναι κατάλληλα τοποθετημένο για αυτό το ρόλο, έχει προταθεί
ότι μπορεί να δρα διαμέσου αλυσίδας δυο μορίων νερού που υπάρχουν στο ενεργό
κέντρο. Η αμίνη είναι σε τέτοια απόσταση ώστε να μπορεί να σχηματίσει δεσμό
υδρογόνου με την Ο5΄ αποχωρούσα ομάδα. [5]
Σχήμα 2.7 Αυτολυτική διάσπαση του RNA από το ριβοένζυμο glmS με την
παρουσία του GlcN6P. Προτεινόμενος μηχανισμός. Η ταυτοποίηση της G33 παραμένει
σημαντική για την αντίδραση, αλλά ο ρόλος της πρέπει να διαλευκανθεί πλήρως.
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Σχεδιασμός της διδασκαλίας των ριβοενζύμων
Εισαγωγικές παρατηρήσεις
Η διδασκαλία των ριβοενζύμων απαιτεί τη γνώση ορισμένων βασικών εννοιών όπως
της κατάλυσης, της αντιγραφής και των διαδικασιών της ροής της γενετικής
πληροφορίας (κεντρικό δόγμα της βιολογίας). Εφόσον διασφαλίσουμε την
κατανόηση των εννοιών αυτών από τους μαθητές, έτσι ώστε να είμαστε σε θέση να
χτίσουμε έννοιες και να μην παραθέσουμε απλά ορισμένες πληροφορίες, είναι δυνατό
να θέσουμε τον εξής προβληματισμό: Ποιο ήταν κατά τη γνώμη σας το αρχικό μόριο-
φορέας γενετικής πληροφορίας στα αρχικά στάδια της εξέλιξης; Αν ήταν το DNA,
πώς έγινε η σύνθεσή του χωρίς ένζυμα; Αν ήταν οι πρωτεΐνες, πώς έγινε η σύνθεσή
τους χωρίς DNA; Αυτός θα είναι ο πυρήνας του προβληματισμού μας, ώστε οι
μαθητές να προετοιμαστούν για τη σημασία των ρυθμιστικών ρόλων που γνωρίζουμε
σήμερα ότι έχουν αποκτήσει τα ριβοένζυμα.
Ανάκληση γνωστικών προαπαιτήσεων
Ακόμη και αν δε γνωρίζουν όλοι οι μαθητές τις διαδικαστικές λεπτομέρειες της
πορεία της αντιγραφής, μεταγραφής, αντίστροφης μεταγραφής, ματίσματος και
μετάφρασης, σχεδιάζουμε στον πίνακα το κεντρικό δόγμα της βιολογίας και
εστιάζουμε στη σημασία που έχουν για τη βιολογία και όχι στις λεπτομέρειες της
πορείας των παραπάνω διαδικασιών. Οι διαδικασίες αυτές απαιτούν «υποβοήθηση»;
Απαιτούν ένζυμα; Εδώ σίγουρα όλοι οι μαθητές μπορούν να απαριθμήσουν κάποια
ένζυμα, π.χ. της αντιγραφής ή μεταγραφής. Μήπως όμως υπάρχουν διαδικασίες οι
οποίες δεν απαιτούν εξωγενή ένζυμα; Αποδίδουμε έμφαση στο μάτισμα και στις
αλληλουχίες εσωνίων και εξωνίων.
Ενημέρωση των μαθητών για τους στόχους του μαθήματος. Διέγερση της
προσοχής των μαθητών.
Με σκοπό να προετοιμαστούν γνωστικά και ψυχολογικά οι μαθητές αλλά και
να οργανώσουν τη σκέψη τους ώστε να μην αποδεχτούν παθητικά τις πληροφορίες
του μαθήματος, μπορεί να γίνει αναφορά στον κεντρικό προβληματισμό: Ποιο είναι
το πληροφοριακό μόριο που εμφανίστηκε πρώτο; Από τις απαντήσεις που θα
λάβουμε μπορούμε να αναδείξουμε τα αδιέξοδα στα οποία μας οδηγούν οι αναφορές
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

στα ένζυμα και στο DNA. Η χρήση των ριβοενζύμων με την αξιοποίηση των
ρυθμιστικών τους ρόλων για τους σκοπούς της Ιατρικής είναι ένα πεδίο με πολλές
προκλήσεις που αποδεικνύει τις πρακτικές εφαρμογές του μαθήματος που θα
διδαχτούν οι μαθητές.
Παρουσίαση – επεξεργασία του μαθήματος
1. Τα μόρια RNA που αποτελούν την πλειοψηφία στα σύγχρονα βιολογικά
συστήματα είναι τα εξής: rRΝA, mRNA και tRNA. Άραγε είναι και τα
μοναδικά;
2. Μια πολύ σύντομη αναφορά στο μη κωδικοποιόν RNA (ncRNA) και στο
γεγονός ότι υπάρχουν διάφορα είδη RNA τα οποία δεν έχουν γίνει ακόμη
γνωστά σε επίπεδο λυκείου. Η ραγδαία όμως ανάπτυξη της επιστήμης της
βιολογίας θα οδηγήσει στην εισαγωγή των κυριότερων από αυτών στα
σχολικά βιβλία της βιολογίας. Οι διαφάνειες 4 και 5 θα βοηθήσουν τους
μαθητές να εξοικειωθούν με τη δομή των μορίων RNA.
3. Καταλυτικό RNA. Πώς ανακαλύφθηκε; Παρουσίαση των αποτελεσμάτων των
αρχικών εργασιών επάνω στα ριβοένζυμα.
4. Ιντρόνια σε ορισμένα RNAs υπόκεινται σε συρραφή εν απουσία ενζύμων.
Διαφάνεια 2.
5. Κατάταξη των ριβοενζύμων σε μικρά και μεγάλα ριβοένζυμα. Στα μικρά
ριβοένζυμα ανήκουν το ριβοένζυμο φουρκέτα, το οποίο εντοπίστηκε στον
δακτυλιοειδή ιό του καπνού, το ριβοένζυμο του ιού της ηπατίτιδας δ, το
ριβοένζυμο από τα μιτοχόνδρια της Neurospora crassa και το σφυροκέφαλο
ριβοένζυμο. Το μέγεθός τους κυμαίνεται από ~35 έως ~164 νουκλεοτίδια. Στα
μεγάλα ριβοένζυμα ανήκουν τα ιντρόνια της ομάδας Ι, των οποίων το μέγεθος
ποικίλει από 200 νουκλεοτίδια μέχρι 1500 νουκλεοτίδια και έχουν ανιχνευθεί
σε σχεδόν όλους τους οργανισμούς, τόσο σε προκαρυωτικούς και
ευκαρυωτικούς όσο και σε υποκυτταρικά οργανίδια και στον Τ4
βακτηριοφάγο. Στα μεγάλα ριβοένζυμα ανήκουν επίσης τα ιντρόνια της
ομάδας ΙΙ που έχουν βρεθεί σε προκαρυωτικούς οργανισμούς αλλά και σε
αρκετά υποκυτταρικά οργανίδια. Το μέγεθός τους κυμαίνεται από 300 μέχρι
3000 νουκλεοτίδια. Τέλος, στα μεγάλα ριβοένζυμα ανήκει και η RNA
υπομονάδα της ριβονουκλεάσης P (RNase P) από τα βακτήρια και κάποια
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

αρχαία. Τα μόρια αυτά έχουν μέγεθος που φτάνει μέχρι ~ 400-500
νουκλεοτίδια. Διαφάνεια 1.
6. Καταλυτική δράση των ριβοενζύμων. Για τα μεγάλα ριβοένζυμα τα δισθενή
κατιόντα - κυρίως το Mg2+ - είναι απαραίτητα. Το γενικό μοντέλο του
μηχανισμού αντίδρασης είναι βασισμένο στο μηχανισμό αντίδρασης των
πρωτεϊνικών μεταλλοενζύμων και απαιτεί δύο κατιόντα μετάλλου. Δεν
αποτελεί όμως τον μοναδικό καταλυτικό μηχανισμό.
7. Ένας πιθανός μηχανισμός δράσης για τα ριβοένζυμα. Στη διαφάνεια 3
παρατηρούμε τη διάσπαση του φωσφοδιεστερικού δεσμού μετά από τις
βάσεις ουρακίλη και κυτοσίνη.
8. Καταλυτικά χαρακτηριστικά των ριβοενζύμων. Παρουσιάζουν υψηλή
συγγένεια για τα υποστρώματά τους και αργή απελευθέρωση των προϊόντων
αντίστοιχα, με αποτέλεσμα τον γρήγορο κορεσμό του καταλυτικού τους
κέντρου. Τα ριβοένζυμα, συγκριτικά με τα πρωτεϊνικά ένζυμα, είναι πολύ πιο
αργοί καταλύτες.
9. Από τις αρχές του 1990, ο Noller και οι συνεργάτες του είχαν υποβάλλει το
ριβόσωμα σε εξαντλητική αποπρωτεΐνωση και είχαν διαπιστώσει ότι η
δραστικότητα της πεπτιδυλο-τρανσφεράσης παρέμενε. Υπήρχε λοιπόν ισχυρή
ένδειξη για το ότι και το ριβόσωμα αποτελεί ένα ριβοένζυμο. Σήμερα με την
ανάπτυξη κρυσταλλογραφικών μεθόδων επιβεβαιώθηκε ότι στο ενεργό
κέντρο της πεπτιδυλο-τρανσφεράσης του ριβοσώματος (κατά τη δράση της
σχηματίζεται ο πεπτιδικός δεσμός στις νεοσυντιθέμενες πρωτεΐνες) υπάρχει
μόνο RNA. Με αυτόν τον τρόπο αποδεικνύεται ότι το ριβόσωμα αποτελεί ένα
ριβοένζυμο.
Ενίσχυση της συγκράτησης και μεταφορά της νέας μάθησης
Στο σημείο αυτό τονίζουμε τα κύρια σημεία του μαθήματος και στη συνέχεια
δείχνουμε τη σημασία των ριβοενζύμων στην επιστήμη της εξέλιξης. Αυτή όμως δεν
είναι και η μοναδική συμβολή των ριβοενζύμων στην επιστήμη. Ο ρυθμιστικός τους
ρόλος είναι δυνατό να δώσει, για παράδειγμα, νέα ώθηση στην αντιμετώπισης
γενετικών ασθενειών με τη μέθοδο της γονιδιακής θεραπείας.
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

ΟΜΑΔΟΠΟΙΗΣΗ ΤΩΝ ΦΥΣΙΚΩΝ ΡΙΒΟΕΝΖΥΜΩΝ ΜΕ
ΒΑΣΗ
ΤΟ ΜΕΓΕΘΟΣ ΤΟΥΣ
Μεγάλα ριβοένζυμα Μικρά ριβοένζυμα
Ιντρόνια της ομάδας ΙΡιβοένζυμο του ιού της
ηπατίτιδας δ
Ιντρόνια της ομάδας ΙΙ Ριβοένζυμο φουρκέτα
RNA υπομονάδα της RNase P Σφυροκέφαλο ριβοένζυμο
Ριβοένζυμο της Neurospora crassa
Διαφάνεια 1
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Διαφάνεια 2
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Διαφάνεια 3
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

ΔΟΜΕΣ RNA: ΤΟ ΕΞΕΛΙΣΣΟΜΕΝΟ «ΡΙΒΟΕΝΖΥΜΟ»
Διαφάνεια 4
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Διαφάνεια 5
Βιβλιογραφία
1. Antony M. Jose ‘‘Ribozyme therapy: RNA Enzymes to the Resque’’, Yale
Journal of Biology and Medicine 75 (2002), pp 215-219.
2. Cristian Hamman and Eric Westhof ‘‘Searcing genomes for ribozymes and
riboswitches’’, Genome Biology 2007, 8:210.
3. Αλέπορου, Αργυροκαστρίτης, Κομητοπουλου, Σγουρίτσα ‘‘Βιολογία Θετικής
Κατεύθυνσης Γ΄ Λυκείου’’, σελ 33.
4. Lubert Streyer ‘‘ΒΙΟΧΗΜΕΙΑ’’, Πανεπιστημιακές Εκδόσεις Κρήτης, σελ.
222-223.
5. Scott Strobel and Jesse C. Cochrane ‘‘RNA Catalysis: Ribozymes, Ribosomes
and Riboswitches’’ Curr Opin Chem Biol. 2007 December; 11(6): 636-643.
6. Kalavrizioti D., Vourekas A., Stamatopoulou V., Toumpeki C., Giannouli S.,
Stathopoulos K., Drainas D., ‘‘RNA Mediated Therapeutics: From Gene
Inactivation to Clinical Application’’ Current Topics in Medicinal Chemistry,
Volume 6, Number 16, p. 1737-1758, 2006.
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ
ΘΕΜΑ 3
Περίληψη
Ως αποτέλεσμα σημειακών μεταλλάξεων, παρά την εξαιρετικά περιορισμένη
τους έκταση, είναι δυνατό να προκύψουν φαινότυποι με μεγάλες διαφορές σε σχέση
με τους θεωρούμενους ως φυσιολογικούς. Έτσι οδηγούμαστε σε μια σειρά
συνδρόμων, τα οποία για τον προσδιορισμό και την κατηγοριοποίησή τους απαιτούν
προσεκτική παρατήρηση και καταγραφή. Αρχικά μελετούμε το σύνδρομο
επεισοδιακής αταξίας για το οποίο έχουν αναφερθεί οι τύποι ΕΑ1, ΕΑ2, ΕΑ3, ΕΑ4,
ΕΑ5, ΕΑ6 και ΕΑ7. Η επεισοδιακή αταξία είναι μια γενετικά κληρονομική
νευρολογική κατάσταση που χαρακτηρίζεται από ετερογένεια στα συμπτώματα. Η
μελέτη της ΕΑ1 ξεκίνησε από το 1975 και σχετίζεται με τη μυοκυμία. Το σύνδρομο
Dejerine-Sottas χαρακτηρίζεται κυρίως από υπερτροφική νευροπάθεια και
απομυελίνωση. Εμφανίζει συμπτώματα παρόμοια, αλλά πιο έντονα, από αυτά του
συνδρόμου Charot-Marie-Tooth τύπου 1Α. Οφείλεται σε μετάλλαξη που συμβαίνει
στο γονίδιο της πρωτεΐνης PMP22 που αποτελεί το 2 έως 5% της μυελίνης του
περιφερειακού νευρικού συστήματος. Για το Li-Fraumeni σύνδρομο έχουν αναφερθεί
τρεις τύποι (LFS1, LFS2 και LFS3) καθώς και ο τύπος Li-Fraumeni-like.
Διαγιγνώσκεται εφόσον πληρούνται τα κριτήρια που έχουν τεθεί το 1994 (Birch et
al.) και σχετίζεται με σαρκώματα και εμφάνιση καρκίνου σε νεαρή ηλικία. Τα άτομα
με LFS βρίσκονται σε κίνδυνο για ένα ευρύ φάσμα κακοηθειών. Ακολουθεί η μελέτη
μιας οικογένειας υποδοχέων (FGFR2), οι μεταλλάξεις των οποίων οδηγούν σε
σύνδρομο Pfeiffer ή σύνδρομο Crouzon. Για το σύνδρομο Pfeiffer ορίζονται τρεις
υποπεριπτώσεις με τον τύπο 1 να αποτελεί τον πιο κοινό και να οδηγεί κυρίως σε
πρόωρη συγχώνευση ορισμένων οστών του κρανίου επηρεάζοντας το σχήμα του
κεφαλιού. Οι μεταπτώσεις, κυρίως στη θέση του 342 αμινοξέος, οδηγούν στο
σύνδρομο Crouzon. Τέλος, το Denys-Drash σύνδρομο προκαλείται από μετάλλαξη
στο γονίδιο WT1 και έχει τις νεφροπάθειες ως κοινό παρανομαστή στα συμπτώματά
του, τα οποία μπορεί να περιλαμβάνουν επίσης τον ψευδοερμαφροδιτισμό και την
εμφάνιση όγκων Wilms.
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Γενικά για τις μεταλλάξεις
Τα γονιδιώματα όλων των οργανισμών είναι δυναμικές οντότητες που
μπορούν να αλλάζουν στη διάρκεια του χρόνου ως αποτέλεσμα συσσώρευσης
αλλαγών. Οι αλλαγές αυτές, που καλούνται μεταλλάξεις, μπορεί να αφορούν αλλαγές
στην αλληλουχία των νουκλεοτιδίων ή αλλαγές που αφορούν αναδιατάξεις
(rearrengments) μεγαλύτερων περιοχών ή και χρωμοσωμάτων και καλούνται
χρωμοσωματικές μεταλλάξεις.
Σημειακή μετάλλαξη είναι μια αλλαγή που αφορά σε μια βάση ενός
γονιδίου. Ενδεχομένως να δημιουργεί ή και να μη δημιουργεί αλλαγή στην πρωτεΐνη
που παράγεται από το συγκεκριμένο γονίδιο. Οι μεταλλάξεις είναι αποτέλεσμα λαθών
κατά τη διάρκεια αντιγραφής του DNA, ή μπορεί να προέλθουν εξαιτίας
μεταλλαξιγόνων παραγόντων.
Η υποκατάσταση ενός ζευγαριού από ένα άλλο αποτελεί ένα τύπο μετάλλαξης
που οδηγεί σε ορισμένα από τα σύνδρομα που παρουσιάζονται στην εργασία αυτή.
Συμβαίνει όταν σχηματίζονται παράταιρα ζεύγη βάσεων (mismatched base pairs)
κατά τη διάρκεια της αντιγραφής του DNA και βασίζεται στην ιδιότητα των βάσεων
να απαντούν σε διάφορες μορφές, που είναι γνωστές ως ταυτομερείς μορφές και που
αποτελούν ισομερή του κάθε μορίου.
Κατατάσσονται σε δύο είδη:
Τις μεταπτώσεις, όπου μια πουρίνη αντικαθίσταται από πουρίνη ή μια
πυριμιδίνη από μια πυριμιδίνη.
Τις μεταστροφές, όπου μια πουρίνη αντικαθίσταται από μια
πυριμιδίνη ή μια πυριμιδίνη από μια πουρίνη.
Υπάρχουν μεταλλάξεις (σιωπηλές και ουδέτερες) που δεν αλλάζουν τη
λειτουργία τις πρωτεΐνης και δε θα μας απασχολήσουν στη μελέτη των συνδρόμων.
Οι μεταλλάξεις που οδηγούν σε διαφορετικό φαινότυπο είναι οι εξής:
μεταλλάξεις με λάθος νόημα (missense mutations), μεταλλάξεις χωρίς νόημα
(nonsense mutations) και μεταλλάξεις μετατόπισης αναγνωστικού πλαισίου. Τα
σύνδρομα που αναφέρονται στη συνέχεια οφείλονται αποκλειστικά σε σημειακές
μεταλλάξεις και κυρίως σε μεταλλάξεις με λάθος νόημα (missense).
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ
Σύνδρομο Επεισοδιακής Αταξίας, Τύπου 1
Το σύνδρομο της Επεισοδιακής Αταξίας, Episodic ataxia syndrome, σύμφωνα
με το διεθνή οργανισμό National Ataxia Foundation, είναι μια σπάνια οικογενής
κληρονομική νόσος του κεντρικού νευρικού συστήματος. Η επεισοδιακή αταξία
τύπου 1 (ΕΑ1) προκαλείται από μετάλλαξη στο γονίδιο της αντλίας του καλίου,
KCNA 1, στο χρωμόσωμα 12p13.
Επίσης, σύμφωνα με τη βάση δεδομένων ΟΜΙΜ του NCBI, αναφέρονται οι
τύποι: ΕΑ2 που προκαλείται από μετάλλαξη στο γονίδιο CACNA1A (CALCIUM
CHANNEL, VOLTAGE-DEPENDENT, P/Q TYPE, ALPHA-1A SUBUNIT) του
χρωμοσώματος 19q13, EA3 που έχει χαρτογραφηθεί στο χρωμόσωμα 1q42, EA4,
EA5 που οφείλεται σε μετάλλαξη στο γονίδιο CACNB4 (CALCIUM CHANNEL,
VOLTAGE-DEPENDENT, BETA-4 SUBUNIT) του χρωμοσώματος 2q22-q23,
EA6 από μετάλλαξη στο γονίδιο 5q13 και ΕΑ7 που έχει χαρτογραφηθεί στο
χρωμόσωμα 19q13.
H μυοκυμία (myokymia) σχετίζεται με μετάλλαξη στο γονίδιο KCNQ2
(POTASSIUM CHANNEL, VOLTAGE-GATED, KQT-LIKE SUBFAMILY,
MEMBER 2).
H επεισοδιακή αταξία είναι μια γενετικά κληρονομική νευρολογική
κατάσταση που χαρακτηρίζεται από ετερογένεια στα συμπτώματα τα οποία
περιλαμβάνουν δυσαρμονία, συχνά συσχετισμένη με προοδευτική αταξία. [1]
Σχήμα 3.1 Απλοποιημένο κληρονομικό πρότυπο του συνδρόμου από τον Οργανισμό
National Ataxia Foundation.
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Οι Van Dyke, Griggs, Merphy και Goldstein το 1975 μελέτησαν μια
οικογένεια έντεκα ατόμων για τρεις συνεχόμενες γενιές που εμφάνιζαν μυοκυμία και
περιοδική αταξία. Μόνο δύο από τα έντεκα συνολικά άτομα ήταν αρσενικά και καμία
μεταβίβαση της ασθένειας από αρσενικό σε αρσενικό δεν παρατηρήθηκε. Η
διαταραχή εμφανίζεται στην πρώτη βρεφική ηλικία με επεισόδια διάρκειας 1 με 2
λεπτών. Τα επεισόδια σχετίζονταν με κινήσεις απότομες στο κεφάλι, στα χέρια και
στα πόδια. Τα επεισόδια προκαλούνταν από απότομές αλλαγές στη θέση, από έντονα
συναισθηματικά ερεθίσματα ή σε μερικές περιπτώσεις από γεύματα με υψηλή
θερμιδική αξία, κυρίως υψηλής περιεκτικότητας σε υδατάνθρακες. Συμπτώματα
μυοκυμίας στο πρόσωπο και στα άκρα ξεκινάνε από την ηλικία των 12 ετών. [2]
Οι Brunt και Van Weerden (1990) περιέγραψαν μια μεγάλη οικογένεια με
παροξυσμική αταξία και συνεχή συμπτώματα μυοκυμίας. Υπήρξε τουλάχιστον μια
περίπτωση μετάδοσης της νόσου από αρσενικό σε αρσενικό. Συχνά εμφανιζότανε
τρέμουλο στο κεφάλι και στα χέρια καθώς και σπασμοί στους μύες του προσώπου και
των χεριών. Τα επεισόδια, τα οποία διαρκούσαν λίγα λεπτά, ήταν δυνατό να
εμφανίζονται αρκετές φορές την ημέρα. Αρχικά εμφανιζότανε στην παιδική ηλικία
και έτειναν να μετριάζονται κατά την εφηβεία. Στους μισούς περίπου ενήλικες τα
συμπτώματα της μυοκυμίας μπορούσαν να διαγνωστούν με συνεχή και προσεκτική
παρατήρηση ενώ σε κάποιους ήταν εμφανή. Οι Brunt και van Weerden συμπέραναν
ότι η μυοκυμική δραστηριότητα οφείλονταν σε ερεθίσματα των περιφερειακών
νεύρων. [3]
Οι Vaamonde, Artienda και Obeso (1991) περιέγραψαν τη διαταραχή σε μια
26χρονη γυναίκα που εμφάνιζε επεισοδιακή αστάθεια για λίγα λεπτά έως την ηλικία
των 6 ετών. Η μητέρα της και μια αδερφή της εμφάνιζαν τα ίδια ακριβώς
συμπτώματα. Ηλεκτρομυογραφικές έρευνες αποκάλυψαν μυοκυμία (neuromyotonia).
H phenyrtoin είχε καλά αποτελέσματα στην καταστολή των συμπτωμάτων αλλά η
acetozolamide ήταν αναποτελεσματική. [4]
Σύμφωνα με τον οργανισμό National Ataxia Foundation, τα δεδομένα για την
επεισοδιακή αταξία κάθε τύπου χωριστά δεν είναι ακριβή, αλλά συνολικά (για όλους
μαζί τους τύπους) εμφανίζει μια συχνότητα τριών με πέντε ατόμων ανά 100.000
άτομα Η επεισοδιακή αταξία θεωρείται σπάνια, με την ΕΑ2 να αναφέρεται πιο συχνά
από την ΕΑ1.
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ
Σχήμα 3.2 Σχηματική αναπαράσταση της δομής του Kv1.1 (voltage-gated potassium
channel) όταν εμφανίζει το σύνδρομο ΕΑ1. Το Κν1.1 αποτελεί προϊόν του γονιδίου KCNA1.
Οι έντονες κουκίδες αποτελούν τη θέση της μετάλλαξης.
Σύνδρομο Dejerine-Sottas
To γονίδιο PMP22, κωδικοποιεί την πρωτεΐνη PERIPHERAL MYELIN
PROTEIN 22 των 22 κιλοβάσεων, η οποία αποτελεί το 2 με 5% της μυελίνης του
περιφερειακού νευρικού συστήματος. Αρχικά παράγεται από τα κύτταρα Schwann
και εκφράζεται, σε μικρή ποσότητα, σε όλες ουσιαστικά τις μυελινωμένες ίνες του
νευρικού περιφερειακού συστήματος. [5]
Η πρωτεΐνη PMP22 αποτελεί το κύριο συστατικό της μυελίνης και παράγεται
κυρίως στα κύτταρα Schwann. Οι μελέτες έχουν αποδείξει το σημαντικό ρόλο της
πρωτεΐνης αυτής στην ανάπτυξη και την διαφοροποίηση των κυττάρων Schwann. [5],
[6]
To 1992 (Martinoti et al., 1992), προτάθηκε ένας πιθανός ρόλος του γονιδίου
της PMP22 στην ανάπτυξη νεοπλασίας σε ασθενείς με νευρινωμάτωση
(neurofibromatosis) και σε μια εκφυλιστική για τη μυελίνη ασθένεια, το σύνδρομο
Charot-Marie-Tooth, που συνδέεται με το χρωμόσωμα 17p. [7]
H πρωτεΐνη PMP22 εκφράζεται σε κρανιακά νεύρα αλλά όχι στο ώριμο
κεντρικό νευρικό σύστημα. Η μεταλλάξεις στην PMP22 είναι δυνατό να
προκαλέσουν νευροαισθητήρια κώφωση από απομυελίνωση του όγδοου κρανιακού
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

νεύρου ή από κακή ανάπτυξη του εσωτερικού αυτιού ή από συνδυασμό και των δύο.
Η μικρή όμως συχνότητα σοβαρής κώφωσης ανάμεσα στις οικογένειες με
μεταλλάξεις στην PMP22 προτείνουν ότι οι περισσότερες μεταλλάξεις έχουν τις
ελάχιστες επιπτώσεις στην ανάπτυξη του εσωτερικού των αυτιών ή στη μυελίνη των
κρανιακών νεύρων. [8]
Το Dejerine-Sottas σύνδρομο χαρακτηρίζεται από υπερτροφική νευροπάθεια
και απομυελίνωση. Τα κλινικά συμπτώματα είναι παρόμοια αλλά πιο έντονα από
αυτά του συνδρόμου Charcot-Marie-Tooth τύπου 1A. Με γονιδιακή ανάλυση στην
περιοχή που κωδικοποιεί την PMP22 σε δυο ασθενείς με σύνδρομο Dejerine-Sottas,
οι οποίοι δεν είχαν καμία μεταξύ τους συγγένεια, αναγνωρίστηκαν οι ίδιες σημειακές
μεταλλάξεις (missense mutations) σε ετερόζυγη κατάσταση. [9]
Σε μια οικογένεια που και οι δύο γονείς ήταν αρνητικοί στην μετάλλαξη,
εμφανίστηκε ασθενής με το σύνδρομο. Έτσι προτάθηκε μια εκ νέου εμφάνιση μέσα
στον πληθυσμό. Σε έναν ασθενή συνέβη μεταστροφή θυμίνης σε αδενίνη (T-to-A
transversion) που οδήγησε σε υποκατάσταση της μεθειονίνης στη θέση 69 από
λυσίνη, ενώ στην άλλη περίπτωση είχαμε μετάπτωση (C-to-T transition) που οδήγησε
σε υποκατάσταση της σερίνης στη θέση 72 από λευκίνη. Ο ασθενής της πρώτης
περίπτωσης δεν εμφάνισε κάποιο ανιχνεύσιμο πρόβλημα κατά τη γέννα αλλά δεν
ξεκίνησε να περπατάει μέχρι τον 15ο μήνα και όταν περπάτησε είχε ένα παράξενο
τρόπο. Η αγωγή της νευρικής ώσης μετρήθηκε να υστερεί κυρίως στο αριστερό
ωλενικό άκρο στην ηλικία των 7 ετών. Έως την ηλικία των 18 παρουσίασε σοβαρή
εξασθένιση των κάτω άκρων του που τον ανάγκασε να κινείται σε αναπηρικό
καροτσάκι. Παρουσίασε επίσης απώλεια της ευαισθησίας των αισθητήριων νεύρων
του και στα τέσσερα άκρα. Κανένα άλλο μέλος της οικογένειας δεν εμφάνισε
παρόμοια πάθηση ή συμπτώματα. Ηλεκτρονική μικροσκοπία και βιοψία έδειξαν
υπερτροφισμό στο νευρικό σύστημα με απώλεια ή ανωμαλία στις ίνες μυελίνης. [9]
Ο ασθενής στον οποίο οι Roa, Dyck, Marks, Chance και Lupski παρατήρησαν
υποκατάσταση της σερίνης από λευκίνη στο 72ο αμινοξύ, ήταν ένα οκτάχρονο αγόρι
που είχε σοβαρή αδυναμία από τη γέννησή του, καθυστέρηση στην ανάπτυξη του
εριστικού συστήματος αλλά κανονική ανάπτυξη στην ικανότητα ομιλίας και
σταδιακή βελτίωση στις κινητικές του ικανότητες. Περπάτησε με μηχανική
υποστήριξη στην ηλικία των 7. Παρατηρήθηκε σε αυτό ατροφία στα κάτω άκρα και
απουσία αντανακλαστικών και στα τέσσερα άκρα. Η μητέρα του ασθενούς, που
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ
πέθανε τριάντα χρονών από ανεπάρκεια στο αναπνευστικό της σύστημα, είχε
ιστορικό παρόμοιων νευρομυικών παθήσεων. [9]
Σύνδρομο Li-Fraumeni
Σύμφωνα με τη βάση δεδομένων ΟΜΙΜ του NCBI, ο τύπος 1 του συνδρόμου
αυτού (LFS1) προκαλείται από μετάλλαξη στο p53 (TUMOR PROTEIN p53,
TP53) γονίδιο. Μια δεύτερη μορφή του συνδρόμου Li-Fraumeni (LFS2) προκαλείται
από μεταλλάξεις στο γονίδιο CHEK2 (CHECKPOINT KINASE 2). Μια ακόμη
μορφή (LFS3) έχει χαρτογραφηθεί στο χρωμόσωμα 1q23.
Το Li-Fraumeni Syndrome, (LFS), παρουσιάζει κλινική και γενετική
ετερογένεια και σχετίζεται με την καρκινογένεση. Χαρακτηρίζεται από αυτοσωμικό
επικρατή τύπο κληρονομικότητας, εμφάνιση όγκων σε πρώιμη ηλικία, πολλαπλούς
όγκους σε έναν ασθενή και εμφάνιση σε μέλη της ίδιας οικογένειας. Σε αντίθεση με
άλλα κληρονομήσιμα σύνδρομα, το LFS παρουσιάζει μια ποικιλία τους τύπους των
όγκων. [10]
Το Li-Fraumeni σύνδρομο διαγιγνώσκεται αν πληρούνται τα ακόλουθα τρία
κριτήρια (Birch et al., 1994):
1. the patient has been diagnosed with a sarcoma at a young age (below 45),
O ασθενής έχει διαγνωστεί με σάρκωμα σε νεαρή ηλικία (κάτω των 45).
2. a first-degree relative has been diagnosed with any cancer at a young age
(below 45), Ένας συγγενής πρώτου βαθμού έχει διαγνωστεί με καρκίνο σε
οποιαδήποτε νεαρή ηλικία (κάτω των 45).
3. and another first-degree or a second-degree relative has been diagnosed
with any cancer at a young age (below 45) or with a sarcoma at any age.
Ένας άλλος συγγενής πρώτου ή δευτέρου βαθμού έχει διαγνωστεί με
καρκίνο σε οποιαδήποτε νεαρή ηλικία (κάτω των 45) ή με σάρκωμα σε
οποιαδήποτε ηλικία. [11]
Ένας λιγότερο περιοριστικός ορισμός καθορίζει το σύνδρομο Li-Fraumeni-
like (LFL). Σε αυτό αρκούν δύο διαφορετικοί όγκοι που σχετίζονται με το LFS σε
συγγενείς πρώτου ή δευτέρου βαθμού σε οποιαδήποτε ηλικία. Περίπου το 70% των
περιπτώσεων LFS και το 40% των περιπτώσεων LFL περιλαμβάνουν βλαστικές
μεταλλάξεις στο γονίδιο p53 στο χρωμόσωμα 17p13.1. Σε περιπτώσεις μεταλλάξεων,
σε ασθενείς με LFS, που δε συνέβησαν στο p53 χρωμόσωμα, οι μεταλλάξεις
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

συνέβησαν στο γονίδιο CHEK2 στο χρωμόσωμα 22q11. Ένας τρίτος γονιδιακός
τόπος για το σύνδρομο έχει αναγνωριστεί στο χρωμόσωμα 1q23. [12]
Στα αρχεία των ιατρικών δεδομένων και των πιστοποιητικών θανάτου 648
παιδιών-ασθενών με ραβδομυοσάρκωμα, οι Li και Fraumeni αναγνώρισαν 4
οικογένειες στις οποίες αμφιθαλή αδέρφια ή ξαδέρφια εμφάνισαν σαρκώματα από
την παιδική ηλικία. Αυτές οι τέσσερις οικογένειες είχαν επίσης χαρακτηριστικό
ιστορικό καρκίνου του μαστού και άλλων νεοπλασμάτων, προτείνοντας ένα νέο
οικογενές σύνδρομο που οδηγεί σε διάφορους όγκους πολλών τύπων που
περιλαμβάνονται στο LFS [13].
Επόμενες έρευνες επιβεβαίωσαν την υψηλή συχνότητα εμφάνισης όγκων σε
οικογένειες που εμφανίζουν το LFS. [14]
Μελέτες σε ομάδες άλλων γεωγραφικών και εθνικών χαρακτηριστικών
επιβεβαίωσαν το σύνδρομο.Persons with LFS are at risk for a wide range of
malignancies, with particularly high occurrences of breast cancer, brain tumors, acute
leukemia, soft tissue , bone sarcomas, and adrenal cortical carcinoma. Τα άτομα με
FFS βρίσκονται σε κίνδυνο για ένα ευρύ φάσμα των κακοηθειών, με ιδιαίτερα υψηλά
περιστατικά καρκίνου του μαστού, όγκων του εγκεφάλου, οξείας λευχαιμίας,
σαρκώματα των οστών, και καρκινώματος του φλοιού των επινεφριδίων. [15]
Επειδή τα γονίδια των πρωτεϊνών καταπιεστών όγκων (tumor suppressor)
είχαν βρεθεί να σχετίζονται με νεοπλασίες, οι Malkin et al (1990) υποψιάστηκαν
μεταλλάξεις σε αυτού του τύπου τα γονίδια στο σύνδρομο Li-Fraumeni. Το γονίδιο
για το ρετινοβλάστωμα δεν ήταν υποψήφιο για μετάλλαξη επειδή το ρετινοβλάστωμα
δεν είχε παρατηρηθεί σε αυτές τις οικογένειες. Από την άλλη, το γονίδιο TP53 ήταν
πιο πιθανός υποψήφιος επειδή μεταλλάξεις που το αδρανοποιούσαν είχαν
συσχετισθεί με οστεαοσαρκώματα, σαρκώματα σε ιστούς, όγκους εγκεφάλου,
λευχαιμία και καρκινώματα του πνεύμονα και του στήθους. Επιπλέον ποντίκια που
φέρουν διαγονιδιακά μεταλλαγμένο το γονίδιο p53 είχαν αυξημένη εμφάνιση σε
οστεοσαρκώματα, σαρκώματα των ιστών και καρκινώματα στους πνεύμονες και στο
στήθος και εμφάνιση όγκων στα επινεφρίδια και στο λεμφικό τους σύστημα, όλοι οι
όγκοι που σχετίζονται με το LFS. Αυτή ήταν η βάση για την επιτυχή έρευνα των
μεταλλάξεων p53. [16]
Για τον προσδιορισμό της συχνότητας και της κατανομής των μεταλλάξεων
p53 σε οικογένειες με LFS, οι Frebourg et al. (1995) έκαναν σύγκριση των δέκα
εξωνίων της TP53 σε λεμφοκύτταρα και βλαστικά κύτταρα που παρέλαβαν από 15
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

οικογένειες με το σύνδρομο. Βλαστικές μεταλλάξεις παρατηρήθηκαν σε 8
οικογένειες, από αυτές στις έξι ήταν του τύπου missense μεταξύ των εξωνίων 5 και 8.
Μια μετάλλαξη ήταν του τύπου nonsense στο εξώνιο 6 και μια του τύπου splicing στο
εσώνιο 4. Κάθε μια από αυτές παρήγαγε μικρότερα τμήματα p53 RNA. Η μελέτες
έδειξαν ότι οι περισσότερες βλαστικές μεταλλάξεις ήταν τοποθετημένες μεταξύ των 5
και 8 εξωνίων και περίπου το 50% των ασθενών δεν παρουσίαζαν καμία βλαστική
μετάλλαξη στην κωδικοποιούσα περιοχή του γονιδίου p53. [17]
Παραπάνω από 50 οικογένειες έχουν πιστοποιηθεί με το LFS εξαιτίας
μεταλλάξεων του TP53. Το LFS ορίζεται με αυστηρά κριτήρια των κλινικών
συμπτωμάτων του. Ο ορισμός του LFS είναι δυνατό να διασταλεί ώστε να περιλάβει
και περιπτώσεις του συνδρόμου Li-Fraumeni-like. [18]
FIBROBLAST GROWTH FACTOR RECEPTOR 2; FGFR2
Ο παράγοντας ανάπτυξης υποδοχέα ινοβλαστών, Fibroblast Growth Factor
Receptor, περιλαμβάνει μια οικογένεια συσχετιζόμενων αλλά διακριτών μεταξύ τους
υποδοχέων. Αυτοί έχουν παρόμοια πρωτεϊνική δομή, με τρεις τομείς που μοιάζουν με
μόρια ανοσοσφαιρίνης (immunoglobulin-like domains) σε μια εξωκυτταρική περιοχή,
ένα τμήμα απλής μεμβράνης και τον τομέα κινάση της τυροσίνης στο κυτταρόπλασμα
(tyrosine kinase domain). Οι άλλοι παράγοντες που έχουν ανακαλυφθεί είναι οι
FGFR1, FGFR3 o οποίος είναι μεταλλαγμένος στην αχονδροπλασία και ο FGFR4. Η
ανάλυση της αλληλουχίας για την ανθρώπινη FGFR2 των 4,5 κιλοβάσεων, έδειξε ένα
ανοιχτό πλαίσιο ανάγνωσης που κωδικοποιεί την μεμβράνη και τον υποδοχέα
κινάσης της τυροσίνης. Δύο εναλλακτικά γονιδιακά προϊόντα έχουν χαρακτηριστεί:
τα KGFR και BEK. Αυτά τα δυο ισομερή είναι πανομοιότυπα εκτός από το 49ο
αμινοξύ μετρημένο από το δεύτερο μισό της τρίτης Ig θηλιάς της εξωκυττάριας
περιοχής. Αυτή η τοπική διαφοροποίηση οφείλεται στην παρουσία εναλλακτικών
εξωνίων εντός του FGFR2, του εξωνίου Β που εκφράζεται στο προϊόν ΒΕΚ και του
εξωνίου K26 στην KGFR. [19]
Pfeiffer σύνδρομο
Σε κάποιες λίγες περιπτώσεις του συνδρόμου Pfeiffer παρατηρήθηκε
μεταστροφή από αδενίνη σε κυτοσίνη (A-to-C transversion) στο νουκλεοτίδιο 1033,
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

αλλάζοντας τη θρεονίνη με προλίνη στη θέση 341, δίπλα στην κυστεΐνη της θέσης
342 στο υπολειμματικό τμήμα που βρέθηκε μεταλλαγμένο στις περιπτώσεις των
συνδρόμων Pfeiffer και Crouzon. [20] (Rutland et al. 1995)
Σύμφωνα με την εργασία του Cohen (1993) ορίζονται τρεις υποπεριπτώσεις
του συνδρόμου. Το σύνδρομο τύπου 1 είναι ο κοινός τύπος. Αποτελεί μια γενετική
διαταραχή που χαρακτηρίζεται από την πρόωρη συγχώνευση ορισμένων οστών του
κρανίου (craniosynostosis), η οποία αποτρέπει την περαιτέρω ανάπτυξη του κρανίου
και επηρεάζει το σχήμα του κεφαλιού και του προσώπου. Οι αντίχειρες και τα πρώτα
δάχτυλα του ποδιού εμφανίζονται με μεγάλο μέγεθος και οι ασθενείς έχουν κανονική
ή σχεδόν κανονική νοημοσύνη. H διαταραχή αυτή κληρονομείται με αυτοσωμικό
επικρατές πρότυπο, το οποίο σημαίνει ότι ένα αντίγραφο του τροποποιημένου
γονιδίου σε κάθε κύτταρο αρκεί για να την προκαλέσει. [21]
Το σύνδρομο του τύπου II παρουσιάζει πιο έντονα συμπτώματα από αυτά του
τύπου Ι που περιλαμβάνουν σοβαρή οφθαλμική πρόπτωση στην οποία εμπλέκεται το
κεντρικό νευρικό σύστημα και μια ποικιλία ανωμαλιών με μικρή συχνότητα όπως
γαστρεντερικές δυσπλασίες και στένωση της τραχείας. Αγκύλωση στους αγκώνες
εμφανίζεται σε μεγαλύτερη συχνότητα. Εξαιτίας των θανάτων τω ασθενών σε μικρή
ηλικία και την αδυναμία αναπαραγωγής, αυτός ο φαινότυπος παρατηρήθηκε μόνο σε
σποραδικές περιπτώσεις. Παρόμοιες σοβαρές περιπτώσεις χωρίς σχήμα κεφαλής
τριφυλιού καλούνται σύνδρομο Pfeiffer τύπου ΙΙΙ. [21]
Οι Grip et al. ανακάλυψαν μια μετάλλαξη της σερίνης στη θέση 351 σε
κυστείνη (ser351-to-cys, S351C) στο γονίδιο της FGFR2 σε έναν ασθενή που
θεωρήθηκε ότι έπασχε από σύνδρομο τύπου ΙΙΙ. Ο ασθενής εμφάνιζε υδροκεφαλισμό,
επιληπτικές κρίσεις, ακραία πρόπτωση, εξάρθρωση των ματιών και σοβαρή
καθυστέρηση στην ανάπτυξη. [22]
Ο ασθενής εμφάνιζε συμπτώματα που προσομοίαζαν με αυτά που
παρατήρησαν οι Kerr et al. (1996) σε μια περίπτωση ασθενούς με κανονικούς
αντίχειρες και οι Gripp et al. Σε μια περίπτωση υποκατάστασης της κυστεΐνης από
αργινίνη στη θέση 342 του γονιδίου FGFR2. Έτσι, το σύνδρομο Pfeiffer φαίνεται να
εμφανίζει ετερογένεια στα συμπτώματα. [23]
Ταξινόμηση
Συνοπτικά, το σύνδρομοPfeiffer syndrome is divided into three subtypes.
Pfeiffer χωρίζεται σε τρεις υποκατηγορίες.
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Type 1 or "classic" Pfeiffer syndrome has symptoms as described
above. Τύπου 1 ή "κλασικο" σύνδρομο Pfeiffer με συμπτώματα όπως
περιγράφονται παραπάνω. Most individuals with type 1 have normal
intelligence and a normal life span.Τα περισσότερα άτομα τύπου 1
έχουν φυσιολογική νοημοσύνη και η κανονική διάρκεια ζωής.
Types 2 and 3 are more severe forms of Pfeiffer syndrome, often
involving problems with the nervous system. Τύπου 2 και 3. Είναι πιο
σοβαρές μορφές του συνδρόμου Pfeiffer, συχνά συνεπάγονται
προβλήματα στο νευρικό σύστημα. ΤουType 2 is distinguished from
type 3 by more extensive fusion of bones in the skull, leading to a
"cloverleaf" shaped head. τύπου 2 διακρίνεται από του τύπου 3 με πιο
εκτεταμένη τήξη των οστών στο κρανίο, που οδηγεί σε ένα σε ένα
σχήμα κεφαλής σχήματος «τριφυλλιού».
Συχνότητα
Το Pfeiffer syndrome affects about 1 in 100,000 individuals. Pfeiffer
σύνδρομο επηρεάζει περίπου 1 στα 100.000 άτομα.
This condition is inherited in an pattern, which means one copy of the
altered gene in each cell is sufficient to cause the disorder.
Σύνδρομο and are used to confirm that somebody has this gene
mutation.Crouzon
Σε τρεις ασθενείς που δε σχετίζονταν μεταξύ τους, βρέθηκε μια μετάπτωση G
σε Α στο νουκλεοτίδιο 1037, στο Β εξώνιο του γονιδίου FGFR2. Αυτό οδήγησε σε
υποκατάσταση της κυστείνης της θέσης 342 σε τυροσίνη (cys342-to-tyr, C342Y)
εντός της τρίτου Ig τομέα. [24] Η ίδια μετάλλαξη βρέθηκε σε ασθενή με το σύνδρομο
Pfeifer [20].From Wikipedia, the free encyclopedia
Oι Steinberger et al. (1995) ανακάλυψαν μεταλλάξεις στο κωδικόνιο 342 σε
τρεις διαφορετικές περιπτώσεις σε ασθενείς με σύνδρομο Crouzon. Δύο από αυτές
αφορούσαν μετάπτωση (G-to-A transition) στη θέση 1037, μια μετάλλαξη που
περιγράφεται στην εργασία των Reardon et al. H τρίτη ήταν μια μεταστροφή (C-to-G
transversion) στη θέση 1038, αποτέλεσμα αντικατάστασης κυστείνης από
τρυπτοφάνη. Επισήμαναν ότι η μετάλλαξη στο κωδικόνιο 342 είχε παρατηρηθεί σε 8
από τις 17 περιπτώσεις και ότι στις 9 περιπτώσεις η μετάλλαξη συνέβη σε άλλες
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

πέντε θέσεις, προτείνοντας ότι το κωδικόνιο 342 του Β εξωνίου της FGFR2 οδηγεί
στο σύνδρομο. Η υποκατάσταση της κυστείνης που φαίνεται να οδηγεί στο σύνδρομο
εμφανίζεται στην τομέα immunoglobulin-like της FGFR2. [25]
Επίσης, σύμφωνα με τη βάση δεδομένων ΟΜΙΜ του NCBI, έχουν αναφερθεί
περιπτώσεις του συνδρόμου που οφείλονται στις εξής μεταλλάξεις:
cys342-to-arg (CYS342ARG) με T-to-C μετάπτωση στο 1036 νουκλεοτίδιο
cys342-to-ser (CYS342SER) με T-to-A μεταστροφή στο 1036 νουκλεοτίδιο
tyr342-to-his (TYR340HIS) με T-to-C μετάπτωση στο 1030 νουκλεοτίδιο
ser354-to-cys (SER354CYS) με C-to-G μεταστροφή στο 1030 νουκλεοτίδιο
ala344-to-ala (ALA344ALA) με G-to-A μετάπτωση στο 1044 νουκλεοτίδιο
Σύμφωνα με τη Wikipedia, The most notable characteristic of Crouzon
syndrome is cranial synostosis, as described above, but it usually presents as
brachycephaly, which results in the appearance of a short and broad head. (bulging
due to shallow eye sockets after early fusion of surrounding bones), (greater than
normal distance between the eyes), and (beak-like nose) are also symptoms.το πιο
αξιοσημείωτο χαρακτηριστικό του συνδρόμου Crouzon είναι κρανιακή synostosis,
όπως περιγράφεται παραπάνω.
Άλλα συμπτώματα είναι τα εξής:
Exophthalmos: διόγκωση των ματιών λόγω των ρηχών υποδοχών οφθαλμού
μετά της σύντηξης των οστών.
Hypertelorism: μεγαλύτερη από την κανονική απόσταση μεταξύ των ματιών.
For reasons that are not entirely clear, most Crouzon patients also have
noticeably shorter and bones, in proportion to the rest of their bodies, than members
of the general population.Για λόγους που δεν είναι απολύτως σαφής, οι περισσότεροι
ασθενείς έχουν σημαντικά μικρότερο βραχιόνιο και μηριαίο οστό, σε αναλογία προς
το υπόλοιπο του σώματός τους, από τα μέλη του γενικού πληθυσμού. A small
percentage of Crouzon patients also have what is called "Type II" Crouzon syndrome,
distinguished by partial .Ένα μικρό ποσοστό των ασθενών με σύνδρομο Crouzon
επίσης, αυτό που ονομάζεται Τύπου ΙΙ σύνδρομο Crouzon, διακρίνεται από μερική
συνδακτυλία.
Διάγνωση
Diagnosis of Crouzon syndrome usually can occur at birth by assessing the
signs and symptoms of the baby. Η διάγνωση του συνδρόμου Crouzon συνήθως
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

μπορεί να επιτευχθεί κατά τη γέννηση, με την αξιολόγηση των συμπτωμάτων του
μωρού. Further analysis, including radiographs, (MRI) scans, genetic testing, X-rays
and CT scans can be used to confirm the diagnosis of Crouzon syndrome. Περαιτέρω
ανάλυση που συμπεριλαμβάνει ακτινογραφίες, μαγνητική τομογραφία (MRI),
γενετικές εξετάσεις, ακτίνες Χ και αξονική τομογραφία μπορεί να χρησιμοποιηθεί για
να επιβεβαιώσει τη διάγνωση του συνδρόμου Crouzon.
Συχνότητα
Σύμφωνα με τον διαδικτυακό τόπο wikipedia, ηIncidence of Crouzon
syndrome is currently estimated to occur in 1 out of every 25,000 people out of the
general population. [ ] There is a greater frequency in families with a history of the
disorder, but that doesn't mean that everyone in the family is affected (as referred to
above). συχνότητα εμφάνισης του συνδρόμου Crouzon εκτιμάται επί του παρόντος σε
1 ανά 25.000 ανθρώπους του γενικού πληθυσμού. Υπάρχει μεγαλύτερη συχνότητα σε
οικογένειες με ιστορικό της διαταραχής, αλλά αυτό δεν σημαίνει ότι ο καθένας στην
οικογένεια πρέπει και να επηρεάζεται.
Denys-Drash σύνδρομο
From Wikipedia, the free encyclopedia ToDenys-Drash syndrome is a
characterized by the following conditions: Denys-Drash σύνδρομο είναι ένα
σύνδρομο που προκαλείται από μετάλλαξη στο γονίδιο WT1. Παρόμοια κλινικά
χαρακτηριστικά παρουσιάζουν τα σύνδρομα Meacham και Frasier.
Oι Drash, Sherman, Hartman Blizzard ανέφεραν 2 παιδιά χωρίς συγγενική
μεταξύ τους σχέση, με σύνδρομα που περιλαμβάνουν ψευδοερμαφροδιτισμό, όγκους
Wilms, υπέρταση και εκφυλιστικές νεφρικές ασθένειες. [26]
Οι Barakat et al., (1974) ανέφεραν 3 περιπτώσεις ψευδοερμαφροδιτισμού,
νεφρικής ανεπάρκειας και όγκων Wilms, κάνοντας επίσης αναφορά σε δυο
επιπρόσθετες μη καταγεγραμμένες περιπτώσεις. [27]
Σύμφωνα με την εργασία του 1985, 10 παιδιά αναφέρθηκαν με συμπτώματα
χαρακτηριστικά του συνδρόμου Denys-Drash. Όλα παρουσίασαν νεφροπάθειες. Τρεις
ασθενείς επίσης είχανε ψευδoερμαφροδιτισμό με φαινότυπο αρσενικού, δυο είχαν
όγκους Wilms και πέντε είχαν όλα τα παραπάνω συμπτώματα. Εννιά ασθενείς
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

κατέληξαν με χρόνια ή τελικού σταδίου νεφρική ανεπάρκεια μέσα σε δύο μήνες έως
δύο το πολύ χρόνια από την έναρξη των συμπτωμάτων. Το δέκατο παιδί εμφάνισε
νεφρική ανεπάρκεια από την ηλικία των 11. Σύμφωνα με την ίδια εργασία, ο
φαινότυπος όσων πάσχουν από το Denys-Drash σύνδρομο περιλαμβάνει είτε όγκους
wilms, είτε ψευδοερμαφροδιτισμό με κοινό παρανομαστή την πρώιμη εμφάνιση
νεφροπάθειας [28]
ΤοDenys-Drash Syndrome (DDS) is a very rare disorder. Denys-Drash
Σύνδρομο (DDS) είναι μια πολύ σπάνια διαταραχή.
Το 1991, σε μόνο έναν από τους δέκα ασθενείς με σύνδρομο Denys-Drash
βρέθηκε χρωμοσωμική ανωμαλία ελλείμματος (deletion) στην περιοχή 11p13-p12.
Αυτή ήταν η μόνη παρατήρηση αυτοσωμική χρωμοσωμική ανωμαλία ανάμεσα στους
ασθενείς που μελετήθηκαν. Οι συγγραφείς συμπέραναν ότι οι απαλειφές ή γενικά οι
μεγάλες αναδιατάξεις στο χρωμόσωμα δεν είναι συνηθισμένες σε ασθενείς με αυτό το
σύνδρομο. [29]
Βιβλιογραφία
1. Jen, J. C.; Graves, T. D.; Hess, E. J.; Hanna, M. G.; Griggs, R. C.; Baloh, R.
W.; CINCH Investigators :
Primary episodic ataxias: diagnosis, pathogenesis and treatment. Brain
130: 2484-2493, 2007.
PubMed ID : 17575281
2. Van Dyke, D. H.; Griggs, R. C.; Murphy, M. J.; Goldstein, M. N. :
Hereditary myokymia and periodic ataxia. J. Neurol. Sci. 25: 109-118, 1975.
PubMed ID : 1170284
3. Brunt, E. R. P.; van Weerden, T. W. : Familial paroxysmal kinesigenic
ataxia and continuous myokymia. Brain 113: 1361-1382, 1990.
PubMed ID : 2245301
4. Vaamonde, J.; Artieda, J.; Obeso, J. A. : Hereditary paroxysmal ataxia
with neuromyotonia. Movement Disorders 6: 180-182, 1991.
PubMed ID : 1647493
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

5. Snipes, G. J.; Suter, U.; Welcher, A. A.; Shooter, E. M. :
Characterization of a novel peripheral nervous system myelin protein
(PMP-22/SR13). J. Cell Biol. 117: 225-238, 1992.
PubMed ID : 1556154
6. Spreyer, P.; Kuhn, G.; Hanemann, C. O.; Gillen, C.; Schaal, H.; Kuhn, R.;
Lemke, G.; Muller, H. W. :
Axon-regulated expression of a Schwann cell transcript that is
homologous to a 'growth arrest-specific' gene. EMBO J. 10: 3661-3668, 1991.
PubMed ID : 1935894
7. Martinotti, A.; Cariani, C. T.; Melani, C.; Sozzi, G.; Spurr, N. K.; Pierotti,
M. A.; Colombo, M. P. :
Isolation and mapping to 17p12-13 of the human homologous of the
murine growth arrest specific Gas-3 gene. Hum. Molec. Genet. 1: 331-334, 1992.
PubMed ID : 1303210
8. Boerkoel, C. F.; Takashima, H.; Garcia, C. A.; Olney, R. K.; Johnson, J.;
Berry, K.; Russo, P.; Kennedy, S.; Teebi, A. S.; Scavina, M.; Williams, L. L.;
Mancias, P.; Butler, I. J.; Krajewski, K.; Shy, M.; Lupski, J. R. :
Charcot-Marie-Tooth disease and related neuropathies: mutation
distribution and genotype-phenotype correlation. Ann. Neurol. 51: 190-201, 2002.
PubMed ID : 11835375
9. Roa, B. B.; Dyck, P. J.; Marks, H. G.; Chance, P. F.; Lupski, J. R. :
Dejerine-Sottas syndrome associated with point mutation in the
peripheral myelin protein 22 (PMP22) gene. Nature Genet. 5: 269-273, 1993.
PubMed ID : 8275092
10. Li, F. P.; Fraumeni, J. R., Jr.; Mulvihill, J. J.; Blattner, W. A.; Dreyfus, M.
G.; Tucker, M. A.; Miller, R. W. :
A cancer family syndrome in twenty-four kindreds. Cancer Res. 48: 5358-
5362, 1988.
PubMed ID : 3409256
11. Birch, J. M.; Hartley, A. L.; Tricker, K. J.; Prosser, J.; Condie, A.; Kelsey,
A. M.; Harris, M.; Jones, P. H. M.; Binchy, A.; Crowther, D.; Craft, A. W.; Eden, O.
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ
B.; Evans, D. G. R.; Thompson, E.; Mann, J. R.; Martin, J.; Mitchell, E. L. D.;
Santibanez-Koref, M. F. :
Prevalence and diversity of constitutional mutations in the p53 gene
among 21 Li-Fraumeni families. Cancer Res. 54: 1298-1304, 1994.
PubMed ID : 8118819
12. Bachinski, L. L.; Olufemi, S.-E.; Zhou, X.; Wu, C.-C.; Yip, L.; Shete, S.;
Lozano, G.; Amos, C. I.; Strong, L. C.; Krahe, R. :
Genetic mapping of a third Li-Fraumeni syndrome predisposition locus to
human chromosome 1q23. Cancer Res. 65: 427-431, 2005.
PubMed ID : 15695383
13. Li, F. P.; Fraumeni, J. F. :
Rhabdomyosarcoma in children: an epidemiologic study and
identification of a familial cancer syndrome. J. Nat. Cancer Inst. 43: 1364-1373,
1969.
14. Li, F. P.; Fraumeni, J. F., Jr. :
Prospective study of a family cancer syndrome. JAMA 247: 2692-2694,
1982.
PubMed ID : 7077763
15. Birch, J. M.; Hartley, A. L.; Marsden, H. B.; Harris, M.; Swindell, R. :
Excess risk of breast cancer in the mothers of children with soft tissue
sarcomas. Brit. J. Cancer 49: 325-331, 1984.
PubMed ID : 6704308
16. Malkin, D.; Li, F. P.; Strong, L. C.; Fraumeni, J. F., Jr.; Nelson, C. E.;
Kim, D. H.; Kassel, J.; Gryka, M. A.; Bischoff, F. Z.; Tainsky, M. A.; Friend, S. H. :
Germ line p53 mutations in a familial syndrome of breast cancer,
sarcomas, and other neoplasms. Science 250: 1233-1238, 1990.
PubMed ID : 1978757
17. Frebourg, T.; Barbier, N.; Yan, Y.; Garber, J. E.; Dreyfus, M.; Fraumeni,
J., Jr.; Li, F. P.; Friend, S. H. :
Germ-line p53 mutations in 15 families with Li-Fraumeni syndrome. Am.
J. Hum. Genet. 56: 608-615, 1995.
PubMed ID : 7887414
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ
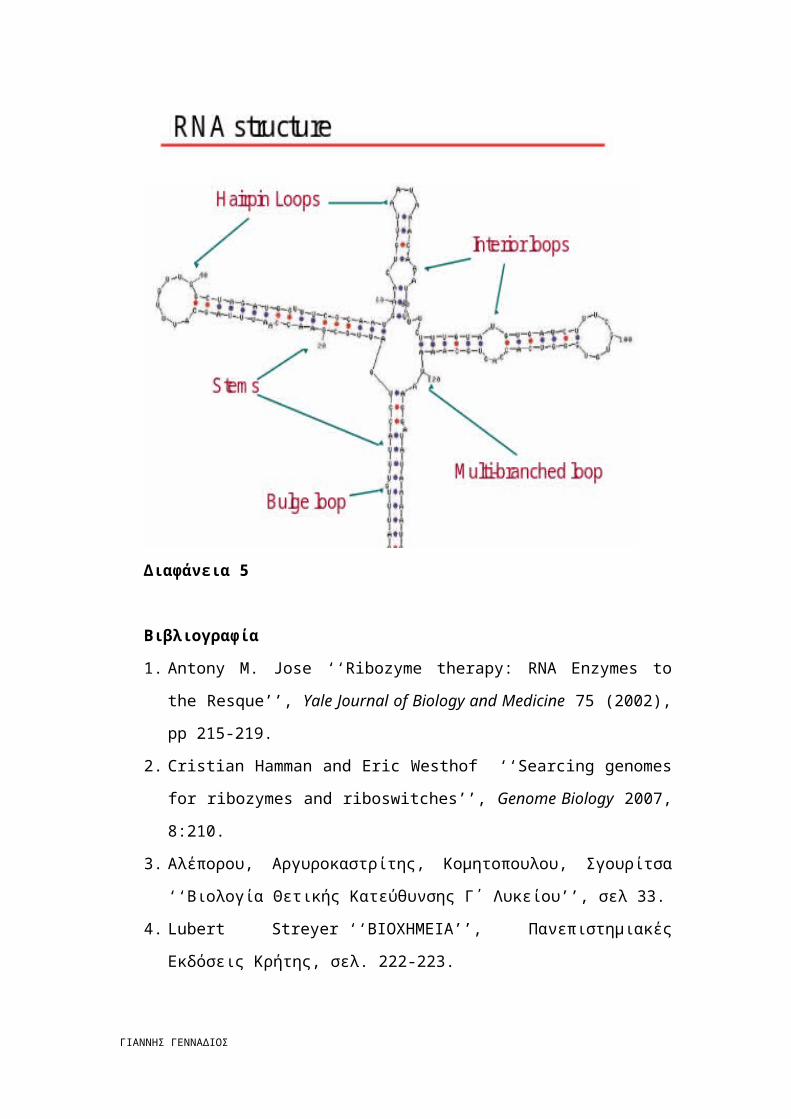
18. Varley, J. M.; Evans, D. G. R.; Birch, J. M. :
Li-Fraumeni syndrome: a molecular and clinical review. Brit. J. Cancer
76: 1-14, 1997.
PubMed ID : 9218725
19. Gilbert, E.; Del Gatto, F.; Champion-Arnaud, P.; Gesnel, M.-C.;
Breathnach, R. :
Control of BEK and K-SAM splice sites in alternative splicing of the
fibroblast growth factor receptor 2 pre-mRNA. Molec. Cell. Biol. 13: 5461-5468,
1993.
PubMed ID : 8355693
20. Rutland, P.; Pulleyn, L. J.; Reardon, W.; Baraitser, M.; Hayward, R.;
Jones, B.; Malcolm, S.; Winter, R. M.; Oldridge, M.; Slaney, S. F.; Poole, M. D.;
Wilkie, A. O. M. :
Identical mutations in the FGFR2 gene cause both Pfeiffer and Crouzon
syndrome phenotypes. Nature Genet. 9: 173-176, 1995.
PubMed ID : 7719345
21. Cohen, M. M., Jr. :
Pfeiffer syndrome update, clinical subtypes, and guidelines for
differential diagnosis. Am. J. Med. Genet. 45: 300-307, 1993.
PubMed ID : 8434615
22. Gripp, K. W.; Stolle, C. A.; McDonald-McGinn, D. M.; Markowitz, R. I.;
Bartlett, S. P.; Katowitz, J. A.; Muenke, M.; Zackai, E. H. :
Phenotype of the fibroblast growth factor receptor 2 ser351-to-cys
mutation: Pfeiffer syndrome type III. Am. J. Med. Genet. 78: 356-360, 1998.
PubMed ID : 9714439
23. Kerr, N. C.; Wilroy, R. S., Jr.; Kaufman, R. A. :
Type 3 Pfeiffer syndrome with normal thumbs. Am. J. Med. Genet. 66:
138-143, 1996.
PubMed ID : 8958319
24. Reardon, W.; Winter, R. M.; Rutland, P.; Pulleyn, L. J.; Jones, B. M.;
Malcolm, S. :
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Mutations in the fibroblast growth factor receptor 2 gene cause Crouzon
syndrome. Nature Genet. 8: 98-103, 1994.
PubMed ID : 7987400
25. Steinberger, D.; Mulliken, J. B.; Muller, U. :
Predisposition for cysteine substitutions in the immunoglobulin-like chain
of FGFR2 in Crouzon syndrome. Hum. Genet. 96: 113-115, 1995.
PubMed ID : 7607643
26. Drash, A.; Sherman, F.; Hartmann, W. H.; Blizzard, R. M. :
A syndrome of pseudohermaphroditism, Wilms' tumor, hypertension,
and degenerative renal disease. J. Pediat. 76: 585-593, 1970.
PubMed ID : 4316066
27. Barakat, A. Y.; Papadopoulou, Z. L.; Chandra, R. S.; Hollerman, C. E.;
Calcagno, P. L. :
Pseudohermaphroditism, nephron disorder and Wilms' tumor: a unifying
concept. Pediatrics 54: 366-369, 1974.
PubMed ID : 4153449
28. Habib, R.; Loirat, C.; Gubler, M. C.; Niaudet, P.; Bensman, A.; Levy, M.;
Broyer, M. :
The nephropathy associated with male pseudohermaphroditism and
Wilms' tumor (Drash syndrome): a distinctive glomerular lesion--report of 10
cases. Clin. Nephrol. 24: 269-278, 1985.
PubMed ID : 3000666
29. Jadresic, L.; Wadey, R. B.; Buckle, B.; Barratt, T. M.; Mitchell, C. D.;
Cowell, J. K. :
Molecular analysis of chromosome region 11p13 in patients with Drash
syndrome. Hum. Genet. 86: 497-501, 1991.
PubMed ID : 1849870
ΘΕΜΑ 4
Περίληψη
Τα miRNA είναι ενδογενώς προερχόμενα μικρά μόρια RNA ~22
νουκλεοτιδίων τα οποία μπορεί να παίξουν σημαντικό ρυθμιστικό ρόλο στα ζώα και
στα φυτά θέτοντας ως στόχο μόρια mRNA για να τα διασπάσουν ή να καταστείλουν
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

την έκφρασή τους. Παρότι ξέφυγαν της προσοχής μέχρι σχετικά πρόσφατα, τα
miRNAs αποτελούν μια από τις πιο άφθονες κατηγορίες μορίων της γονιδιακής
ρύθμισης στους πολυκύτταρους οργανισμούς επηρεάζοντας πιθανότατα την τελική
έκβαση πολλών περιοχών των γονιδίων. Σήμερα γνωρίζουμε ότι τα miRNAs
κατέχουν σημαντικό ρόλο στην ανάπτυξη, τη διαφοροποίηση, την απόπτωση και τον
καρκίνο, ενώ σημαντικοί είναι και οι ρόλοι τους στα φυτά.
H αρχή της ανακάλυψής τους έγινε στα εργαστήρια Ambros και Ruvkun όπου
παρατηρήθηκε ότι τα lin-4 RNAs είχαν αντινοηματική συμπληρωματικότητα με
πολλαπλές θέσεις στην 3΄-αμετάφραστη περιοχή (3΄-UTR) του γονιδίου του lin14.
Ακολούθησε η ανακάλυψη του let-7, ενός άλλου γονιδίου, επίσης στο C. elegans που
κωδικοποιεί ένα άλλο ρυθμιστικό μόριο RNA ~22 νουκλεοτιδίων και δρα κατά
παρόμοιο τρόπο με το lin-4. Ομόλογα με το let-7 γονίδια ταυτοποιήθηκαν σε
γονιδιώματα ανθρώπου και μύγας, καθιστώντας πλέον γνωστή την πολυάριθμη
παρουσία τους στα κύτταρα. Με όλα αυτά τα διαφορετικά γονίδια, υποθέτουμε ότι
κάθε τύπος κυττάρου σε πολυκύτταρους οργανισμούς, σε κάθε εξελικτικό στάδιο,
μπορεί να έχει ένα διαφορετικό εκφραστικό προφίλ.
Σήμερα έχουν αναπτυχθεί τεχνικές εύρεσης γονιδίων που δε βασίζονται μόνο
σε εύρεση ομόλογων ή παρόμοιων αλληλουχιών με ήδη γνωστά γονίδια. Δύο
χαρακτηριστικοί εκπρόσωποι των νέων μεθόδων είναι οι MiRscan και ΜiRseeker.
Όσον αφορά στο ρόλο τους, προσδιορίστηκε πολλές φορές με πειράματα in vivo σε
μεταλλαγμένα μόρια miRNA ή σε μεταλλαγμένες συμπληρωματικές αλληλουχίες που
ήταν δυνατό να διαταράξουν τη φυσιολογική τους λειτουργία. Με αυτό τον τρόπο
έχουν καταρτιστεί πίνακες με τον λειτουργικό ρόλο του κάθε μορίου. Για τη μεγάλη
όμως πλειοψηφία των miRNAs, οι φαινοτυπικές συνέπειες σε τροποποιημένα ή
μεταλλαγμένα μόρια δεν είναι ακόμη γνωστές.
Στη συνέχεια παρουσιάζονται οι αλληλεπιδράσεις μεταξύ ξενιστή-ιού κατά
την προσβολή του ξενιστή από έναν ιό. Η διαδραστικότητα που αναπτύσσεται έχει
σημαντικές επιπτώσεις, τόσο για τους ιούς όσο και για τον ξενιστή. Έχουν αναφερθεί
περιπτώσεις μορίων miRNA σε κύτταρα ξενιστών που καθορίζουν την έκφραση
διάφορων ιικών γονιδίων, παίζοντας έτσι σπουδαίο ρόλο για το ανοσοβιολογικό
σύστημα του ξενιστή. Τέλος, πρόσφατα έχουν αναγνωριστεί οι επιπτώσεις των
miRNAs σε παθολογικές διαδικασίες του καρδιαγγειακού συστήματος, γεγονός το
οποίο υπόσχεται ένα ταχύτητα αναπτυσσόμενα ερευνητικό πεδίο.
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Εισαγωγή
Μια ενδιαφέρουσα διαδικασία που σχετικά πρόσφατα έχει ανακαλυφθεί στα
βασίλεια των ευκαρυωτικών οργανισμών είναι η RNA σίγηση. Η RNA σίγηση
περιλαμβάνει ποικίλες διαδικασίες που εξαρτώνται από μόρια RNA, οι οποίες έχουν
ως κύριο αποτέλεσμα την αρνητική ρύθμιση της γονιδιακής έκφρασης. Κεντρικό
ρόλο παίζουν μικρά μόρια RNA της τάξης μεγέθους 20-26 νουκλεοτιδίων, που
ανιχνεύουν ομόλογες αλληλουχίες και επηρεάζουν τη μεταγραφή, τη σταθερότητα
του mRNA ή τη μετάφραση. Η RNA σίγηση συναντάται σε πρώτιστα, μύκητες, ζώα
και φυτά και πρόκειται για ένα καλά συντηρημένο εξελικτικά μηχανισμό που
μοιράζεται κοινά βασικά χαρακτηριστικά στα τέσσερα παραπάνω διαφορετικά
βασίλεια. Οι διαφοροποιήσεις και προσαρμογές όμως των μηχανισμών σε κάθε
βασίλειο έχουν καθιερώσει την ονομασία quelling στους μύκητες, RNA interference
(RNAi) και RNA silencing σε ζώα και φυτά αντιστοίχως.
Γενικά για το μηχανισμό της σίγησης
Τα μονοπάτια της σίγησης μοιράζονται τρία κοινά βιοχημικά χαρακτηριστικά:
τη δημιουργία δίκλωνου RNA (dsRΝΑ), την κοπή του σε μικρούς μονόκλωνους
ριβονουκλεϊκούς ρυθμιστές (μικρά RNAs) και τέλος την παρεμποδιστική δράση του
μικρού RNA – με τη βοήθεια κατάλληλων συμπλόκων – σε μερικώς ή πλήρως
συμπληρωματικό RNA ή DNA (Susi et al., 2004). [1]
Συγκεκριμένα, η πυροδότηση της RNA σίγησης γίνεται με την ανίχνευση
δίκλωνου μορίου RNA στο κύτταρο. Η δίκλωνη μορφή ενδέχεται να είναι
αποτέλεσμα δευτεροταγούς δομής ενδογενούς μεταγράφου ή διαγονιδίου, είτε
εξωγενής προερχόμενη RNA δομή. Η δίκλωνη μορφή του RNA αναγνωρίζεται από
πρωτεΐνες της οικογένειας Dicer και κόπτεται σε δίκλωνο RNA μικρού μεγέθους. Οι
πρωτεΐνες αυτές είναι ATP-εξαρτώμενες RNase III ενδονουκλεάσες που φέρουν
περιοχές πρόσδεσης δίκλωνου RNA και κατέχουν σημαντικότατο ρόλο στο
μηχανισμό (Vazquez, 2006). Τα φυτά διαθέτουν τέσσερις πρωτεΐνες της παραπάνω
οικογένειας (Dicer-like proteins, DCLs), με καθεμία να κατέχει διακριτό ρόλο, χωρίς
αυτό να συνεπάγεται μη αλληλεπικαλυπτόμενη δράση των DCLs. [2]
Στη συνέχεια, το μικρό μόριο RNA καθίσταται μονόκλωνο και
ενσωματώνεται σε σύμπλοκα σίγησης RISC (RNA induced silencing complex). Τα
σύμπλοκα αυτά παρουσιάζουν ετερογένεια, αλλά όλα ανεξαιρέτως περιέχουν
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

πρωτεΐνες της οικογένειας των Αργοναυτών (Argonaute) και είναι υπεύθυνα για την
παρεμποδιστική δράση που απορρέει από τη φύση των μικρών RNAs (Hammond,
2005). Οι πρωτεΐνες Αργοναύτες κατέχουν σημαντικό ρόλο, καθώς είναι αυτές που
με οδηγό το μικρό ριβονουκλεϊκό ρυθμιστή, οδηγούν σε καταστολή της έκφρασης,
είτε αποικοδομώντας το στόχο, είτε παρεμποδίζοντας τη μεταγραφή ή τη μετάφρασή
του. [3]
Ο ρόλος των miRNAs στη διαδικασία της σίγησης
Τα miRNAs είναι ενδογενώς προερχόμενα μικρά RNAs (Bartel, 2004). Στα
ζώα είναι γνωστό ότι κατέχουν σημαντικό ρόλο στην ανάπτυξη, τη διαφοροποίηση,
την απόπτωση και τον καρκίνο, ενώ στα φυτά εκτός της ρύθμισης αναπτυξιακών
προγραμμάτων, εμπλέκονται επίσης στη ρύθμιση διαδικασιών όπως η σηματοδότηση,
η απόκριση σε βιοτικές και αβιοτικές καταπονήσεις και η μετάβαση από βλαστική σε
αναπαραγωγική φάση (Zhang et al, 2007). [4]
Ta miRNAs προέρχονται από δύο διαδοχικές κοπές των πρώιμων μορίων
miRNAs (pri-miRNA) από μέλη της οικογένειας Dicer. Τα πρώιμα μετάγραφα είναι
προϊόντα της RNA πολυμεράσης ΙΙ και μεταγράφονται από τόπους διακριτούς από
άλλα χαρακτηρισμένα γονίδια (Lee et al, 2004). Ta miRNA γονίδια εντοπίζονται
ακόμη και σε ιντρόνια άλλων γονιδίων και μεταγράφονται είτε αυτόνομα, είτε ως
πολυκιστρονικά μετάγραφα. [5]
Συγκεκριμένα στα φυτά, τα πρώιμα μόρια miRNAs αναγνωρίζονται από την
πυρηνική DCL1 με τη βοήθεια της dsRNA προσδενόμενης πρωτεΐνης HYL1 και
παράγεται το πρόδρομο μετάγραφο με χαρακτηριστική δομή φουρκέτας, το οποίο
υπόκειται σε διαδοχική κοπή δίνοντας γένεση σε δίκλωνο miRNA/mRNA περίπου 21
νουκλεοτιδίων (Jones-Rhoades et al, 2006). [6] Το δίκλωνο αυτό μόριο εξέρχεται από
τον πυρήνα με τη βοήθεια της εξπορτίνης HASTY. Στα ζώα η πρώτη κοπή του
πρώιμου μεταγράφου γίνεται στον πυρήνα από την RNase III ενδονουκλεάση Drosha,
ενώ η δεύτερη στο κυτταρόπλασμα από τη Dicer. Η εξαγωγή του πρόδρομου μορίου
γίνεται από την εξπορτίνη 5. Οι συνεργάτες των ζωικών ενδονουκλεασών που
προσδένονται σε dsRNA είναι η Pasha και η Loquacious αντίστοιχα (Han et al, 2004;
Saito et al, 2005). [7]
Μία ακόμη διαφορά μεταξύ φυτικών και ζωικών ώριμων miRNAs είναι η
ύπαρξη της φυτικής μεθυλοτρανσφεράσης HEN1, η οποία προσδίδει μια μεθυλομάδα
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

στο 2΄-O της ριβόζης τους και θεωρείται ότι έχει προστατευτικό ρόλο (Yu et al,
2005). Και στις δύο περιπτώσεις, μετά το σχηματισμό του miRNA-mRNA επιλέγεται
η αλυσίδα με το λιγότερο 5΄ θερμοδυναμικά σταθερό άκρο (miRNA) για
ενσωμάτωση στο σύμπλοκο RISC και δρα σαν οδηγός για την ανίχνευση ομόλογων
αλληλουχιών. Η αλληλεπίδραση miRNA–μεταγράφου οδηγεί στην καταστολή της
έκφρασης του στόχου. [8]
Σχήμα 4.1 Αναπαράσταση βιογένεσης και δράσης φυτικού miRNA (Jones Rhoades
et al, 2006).
Η καταστολή επιτυγχάνεται με δυο τρόπους και βασίζεται στο βαθμό
συμπληρωματικότητας της αλληλεπίδρασης miRNA και μεταγράφου: ατελής
συμπληρωματικότητα οδηγεί σε παρεμπόδιση της μετάφρασης, γεγονός που
παρατηρείται κυρίως σε μεταζώα, ενώ σχεδόν πλήρης συμπληρωματικότητα οδηγεί
σε αποικοδόμηση του μεταγράφου, φαινόμενο που είναι σύνηθες στα φυτά. Μια
επιπλέον διαφορά μεταξύ φυτών και ζώων, όσον αφορά τη ρύθμιση από miRNAs,
εντοπίζεται στις περιοχές στόχευσης στα μετάγραφα. Στα ζώα η στόχευση γίνεται
στην 3΄ μη κωδική περιοχή του μεταγράφου, ενώ στα φυτά η πρόσδεση των miRNAs
έχει παρατηρηθεί στις κωδικές, όπως και στις μη κωδικές περιοχές. Ένα φαινόμενο
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

που έχει συχνά παρατηρηθεί στα ζώα, είναι ένα μετάγραφο να ρυθμίζεται από
περισσότερα του ενός miRNAs, ούτως ώστε να επιτευχθεί μεταφραστική καταστολή
(Lai, 2004), σε αντίθεση με τους φυτικούς οργανισμούς, όπου η αλληλεπίδραση
μεταγράφου και ενός miRNA είναι αρκετή για να οδηγήσει σε αποικοδόμηση. Τέλος,
συνηθίζεται στα φυτά ένα miRNA (ή μια οικογένεια miRNAs) να ρυθμίζει μέλη της
ίδιας οικογένειες γονιδίων. [9]
Ανακάλυψη
Σε μια έρευνα εμπνευσμένη τόσο από επιμονή όσο και από επιστημονικά
διεισδυτική ματιά, οι Victor Ambros και οι συνεργάτες του, Rosalind Lee και Rhonda
Feinbaum, ανακάλυψαν ότι το γονίδιο lin-4, γονίδιο γνωστό για τη ρύθμιση της
ανάπτυξης στο στάδιο της προνύμφης του νηματοσκώληκα C. elegans, δεν
κωδικοποιεί κάποια πρωτεΐνη αλλά παράγει ένα ζεύγος από μικρά μόρια RNA (Lee et
al., 1993). Το ένα RNA έχει μήκος περίπου 22 νουκλεοτιδίων και το άλλο περίπου 61
νουκλεοτιδίων. Το μεγαλύτερο αναδιπλώνεται σε μια χαρακτηριστική δομή (stem-
loop), που αποτελεί την πρόδρομη μορφή του μικρότερου μορίου στο οποίο και
μετατρέπεται αφού υποστεί διαδοχικές κοπές. Στα εργαστήρια Ambros και Ruvkun
παρατηρήθηκε ότι τα lin-4 RNAs είχαν αντινοηματική (antisense) συμπληρωμα-
τικότητα με πολλαπλές θέσεις στην 3΄-αμετάφραστη περιοχή (3΄-UTR) του γονιδίου
του lin14 (Lee et al., 1993; Wightman et al., 1993). H συμπληρωματικότητα στην
περιοχή αυτή είχε ως αποτέλεσμα την καταστολή της έκφρασης της πρωτεΐνης lin-14
μετα-μεταγραφικά, διότι η σύζευξη των δύο RNA οδηγεί σε αποικοδόμηση του lin-14
mRNA. Το εργαστήριο Ruvkun προχώρησε στη διαπίστωση της σημασίας αυτής της
συμπληρωματικότητας για τη ρύθμιση του lin-14 από το lin-4, δείχνοντας επίσης ότι
η ρύθμιση ουσιαστικά μειώνει την ποσότητα του προϊόντος του γονιδίου χωρίς
σημαντικές αλλαγές στην παραγωγή της ποσότητας του mRNA. Οι ανακαλύψεις
αυτές υποστηρίζουν το μοντέλο στο οποίο το RNA του lin-4 συζεύγνυται με την 3΄-
αμετάφραστη περιοχή του lin-14 για να οδηγήσει σε μεταφραστική καταστολή το
μήνυμα του lin-14 ως τμήμα μιας ρυθμιστικής οδού που οδηγεί τα κύτταρα σε
διαίρεση από το πρώτο στάδιο της προνύμφης σε αυτά του δευτέρου σταδίου (Lee et
al., 1993; Wightman et al., 1993). [10]
Το μικρότερο lin-4 RNA αναγνωρίζεται σήμερα ως το πρώτο μέλος μιας
τάξης άφθονων μικροσκοπικών ρυθμιστικών μορίων που καλούνται microRNAs ή
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

miRNAs (Lagos-Quintana et al., 2001; Lau et al., 2001; Lee and Ambros, 2001).
Πρόσφατα ανακαλύφθηκαν λειτουργίες των miRNA που περιλαμβάνουν τη ρύθμιση
του πολλαπλασιασμού των κυττάρων, το θάνατο των κυττάρων και τον μεταβολισμό
των λιπών στις μύγες (Brennecke et al., 2003; Xu et al., 2003). [10]
Για επτά χρόνια μετά την ανακάλυψη του lin-4 RNA, η γενετική αυτού του
τύπου των μικροσκοπικών μορίων φαινότανε απλή και δεν υπήρξε κάποιο παρόμοιο
εύρημα. Αυτό όμως άλλαξε με την ανακάλυψη του let-7, ενός άλλου γονιδίου στο C.
elegans που κωδικοποιεί ένα άλλο ρυθμιστικό μόριο RNA ~22 νουκλεοτιδίων. Το let-
7 RNA δρα με παρόμοιο τρόπο ρυθμίζοντας όμως τώρα τη μετάβαση των ώριμων
κυττάρων της προνύμφης στα κύτταρα της ενήλικης μορφής (Reinhart et al., 2000;
Slack et al., 2000). Επιπλέον, ομόλογα με το let-7 γονίδια ταυτοποιήθηκαν σε
γονιδιώματα ανθρώπου και μύγας και το ίδιο το let-7 ανιχνεύθηκε στον άνθρωπο, στη
Drosophila και σε άλλα έντεκα είδη ζώων (Pasquinelli et al., 2000). [10]
Εξαιτίας του κοινού ρόλου τους στον έλεγχο των σταδίων στην ανάπτυξη των
οργανισμών, τα lin-4 και let-7 RNAs είχαν αντιγραφεί σε μικρά προσωρινά μόρια
RNA (small temporal RNAs, stRNAs), με την προσδοκία ότι και άλλα ρυθμιστικά
μόρια αυτού του τύπου θα ανακαλυφθούν (Pasquinelli et al., 2000). Πράγματι, μέσα
σε λιγότερο από ένα χρόνο, τρία εργαστήρια κλωνοποίησαν μικρά RNAs από μύγες,
σκουλήκια και ανθρώπινα κύτταρα αναφέροντας συνολικά πάνω από 100 γονίδια
μικροσκοπικών μορίων RNA, περίπου 20 νέα γονίδια στη Drosophila, 30 στον
άνθρωπο και περίπου 60 σε σκώληκες (Lagos-Quintana et al., 2001; Lau et al., 2001;
Lee and Ambros, 2001). Τα προϊόντα RNA αυτών των γονιδίων προσομοίαζαν στα
lin-4 και let-7 stRNAs στο ότι ήταν ~22 νουκλεοτιδίων ενδογενώς εκφρασμένου
RNA. Πιθανότατα είχαν ληφθεί από τον ένα βραχίονα του πρόδρομου μορίου δομής
stem-loop (σχήμα 4.1) και παρέμεναν συντηρημένα κατά την εξέλιξη, άλλα σε
μεγάλο βαθμό και άλλα σε μικρές περιοχές όπως στα C. Elegans και C. Briggsae.
Όμως, σε αντίθεση με τα lin-4 και let-7 RΝAs, πολλά από τα 22 νουκλεοτιδίων RNAs
δεν εκφραζότανε σε συγκεκριμένα στάδια της ανάπτυξης αλλά φαινόταν να είναι
εκφρασμένα σε συγκεκριμένους τύπους κυττάρων. Έτσι, ο όρος miRNA
συνηθιζότανε να αναφέρεται στα stRNAs και σε όλα τα άλλα μικροσκοπικά RNAs με
παρόμοια χαρακτηριστικά αλλά άγνωστες λειτουργίες (Lagos-Quintana et al., 2001;
Lau et al., 2001; Lee and Ambros, 2001). Η ένταση των προσπαθειών για
κλωνοποίηση αποκάλυψε πολλά miRNA γονίδια σε θηλαστικά, ψάρια, σκουλήκια
και μύγες (Lagos-Quintana et al., 2002, 2003; Mourelatos et al., 2002; Ambros et al.,
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

2003b; Aravin et al., 2003; Dostie et al., 2003; Houbaviy et al., 2003; Kim et al.,
2003; Lim et al., 2003a, 2003b; Michael et al., 2003). Έτσι καταρτίστηκε κατάλογος
των mRNA και καθορίστηκε ο τρόπος ονομασίας των νεοανακαλυφθέντων γονιδίων
(Griffiths-Jones, 2004). [10]
Σχήμα 4.2 Παραδείγματα miRNAs σε πολυκύτταρους ζωικούς οργανισμούς. (Α) Οι
δομές stem-loop που ανακαλύφθηκαν στα miRNAs lin-4 και let-7 (Lee et al., 1993; Reinhart
et al., 2000). Η ακριβής αλληλουχία του ώριμου miRNA καθορίστηκε με κλωνοποίηση (Lau
et al., 2001). (Β) Παραδείγματα miRNAs από άλλα γονίδια πολυκύτταρων ευκαρυωτικών
οργανισμών, mir-1, mir-34 mir-124. (C) Παραδείγματα miRNAs σε γονίδια φυτών:
MIR165a, MIR172a2 και JAW. Φαίνονται οι δομές stem-loop από το Arabidopsis, έχουν
όμως βρεθεί ομόλογα στο ρύζι και σε άλλα φυτά (Park et al., 2002; Reinhart et al., 2002;
Palatnic et al., 2003).
Όπως στα lin-4 και let-7 γονίδια του C. elegans, τα περισσότερα γονίδια των
miRNAs βρίσκονται σε περιοχές του γονιδιώματος με αρκετή απόσταση από τα ήδη
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

γνωστά προαναφερόμενα γονίδια, αποδεικνύοντας ότι προέρχονται από ανεξάρτητες
μεταγραφικές μονάδες (Lagos-Quintana et al., 2001; Lau et al., 2001; Lee and
Ambros, 2001). Παρολαυτά, μια ευμεγέθη μειοψηφία (περίπου το 1/4 των
ανθρώπινων miRNA γονιδίων) βρίσκονται στα εσώνια των pre-mRNAs. Αυτά έχουν
κατά προτίμηση τον ίδιο προσανατολισμό με τα προβλεπόμενα mRNAs,
αποδεικνύοντας ότι τα περισσότερα δεν έχουν μεταγραφεί από τους δικούς τους
υποκινητές αλλά από τα εσώνια, όπως παρατηρήθηκε επίσης σε πολλά snoRNAs
(Αravin et al., 2003; Lagos-Quintana et al., 2003; Lai et al., 2003; Lim et al., 2003a).
Η διευθέτηση αυτή παρέχει ένα βολικό μηχανισμό για τη συντονισμένη έκφραση των
miRNAs και των πρωτεϊνών. Είναι εύκολο να φανταστούμε ρυθμιστικά σενάρια στα
οποία τέτοια συντονισμένη έκφραση θα μπορούσε να είναι χρήσιμη για να εξηγήσει
συντηρημένες σχέσεις μεταξύ miRNAs και mRNAs. Ένα χαρακτηριστικό
παράδειγμα τέτοιων συντηρημένων σχέσεων αφορά στο mir-7, το οποίο βρέθηκε στο
εσώνιο hnRNP K τόσο στα έντομα όσο και στα θηλαστικά (Aravin et al., 2003). [10]
Άλλα miRNAs γονίδια βρίσκονται σε συστοιχίες (clusters) στο γονιδίωμα με
μια διευθέτηση και ένα πρότυπο έκφρασης που υποδεικνύει μεταγραφή σε ένα
πολυκιστρονικό (multi-cistronic) πρωταρχικό μετάγραφο (Lagos-Quintana et al.,
2001; Lau et al., 2001). Παρόλο που η πλειοψηφία των γονιδίων στους σκώληκες και
στον άνθρωπο βρίσκεται απομονωμένη (Lim et al., 2003a, 2003b), περισσότερα από
τα μισά γνωστά γονίδια των miRNAs στη Δροσόφιλα βρίσκονται σε συστοιχίες
(Aravin et al., 2003). Τα miRNAs του γονιδιώματος που είναι ομαδοποιημένα, συχνά
αλλά όχι πάντοτε, σχετίζονται μεταξύ τους. Ομόλογες αλληλουχίες γονιδίων της C.
elegans με τα lin-4 και let-7 έχουν βρεθεί σε συστοιχίες στο γονιδίωμα της μύγας και
του ανθρώπου (orthologs) και συνεκφράζονται, μερικές φορές από το ίδιο πρώιμο
μετάγραφο, οδηγώντας στην ιδέα ότι ο γονιδιακος διαχωρισμός του lin-4 από το let-7
στους νηματοσκώληκες μπορεί να είναι μοναδικός στο γένος του σκώληκα (Aravin et
al., 2003; Bashirullah et al., 2003; Sempere et al., 2003). Το παράδειγμα αυτό τονίζει
ξεκάθαρα ότι ακόμη και σε περιπτώσεις που τα γονίδια σε συστοιχία δεν έχουν
προφανή ομολογία στην αλληλουχία τους, μπορεί να μοιράζονται λειτουργικές
σχέσεις. [10]
Η έκφραση των miRNAs
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Πολλά miRNAs παρουσιάζουν ένα ενδιαφέρον πρότυπο έκφρασης. Για
παράδειγμα, γονίδια paralogs και orthologs των lin-4 και let-7 του C.elegans
παρουσιάζουν εξειδίκευση κατά εξελικτικό στάδιο τόσο στην ανάπτυξη όσο και στη
λειτουργία τους παρόμοια με αυτή ορισμένων μορίων stRNAs (Pasquinelli et al.,
2000; Lau et al., 2001; Lagos-Quintana et al., 2002; Bashirullah et al., 2003; Lim et
al., 2003a). Άλλα ενδιαφέροντα παραδείγματα περιλαμβάνουν το miR-1, το οποίο
βρέθηκε κυρίως στην καρδιά θηλαστικών (Lee and Ambros, 2001; Lagos-Quintana et
al., 2002) το miR-122, το οποίο κυρίως βρέθηκε στο συκώτι (Lagos-Quintana et al.,
2002), το miR-223, το οποίο κυρίως εντοπίστηκε σε κοκιοκύτταρα και μακροφάγα
στο μυελό των οστών ποντικιών (Chen et al., 2004), miRNAs από mir-35-mir-42
cluster, το οποίο βρίσκεται κατά προτίμηση σε έμβρυο του c. elegans (Lau et al.,
2001) και αυτά της συστοιχίας mir-290-mir-295, τα οποία εκφράζονται σε βλαστικά
κύτταρα ποντικιών αλλά όχι σε ήδη διαφοροποιημένα κύτταρα (Houbaviy et al.,
2003) . [10]
Έχουν αποδειχτεί διαφορετικά τέτοια πρότυπα σε διαφορετικά εξελικτικά
στάδια ή περιοχές στον εγκέφαλο των θηλαστικών (Krichevsky et al., 2003). Με όλα
αυτά τα διαφορετικά γονίδια και εκφραστικά πρότυπα, είναι λογικό να υποθέσουμε
ότι κάθε τύπος κυττάρου σε πολυκύτταρους οργανισμούς, σε κάθε εξελικτικό στάδιο,
μπορεί να έχει ένα διαφορετικό εκφραστικό προφίλ. [10]
Μια αξιοπρόσεκτη παρατήρηση για την έκφραση του mRNA αποτελεί η
αφθονία των miRNAs στα κύτταρα. Για παράδειγμα, τα miR-2, miR-52 και miR-58
υπάρχουν σε - κατά μέσο όρο - πάνω από 50.000 μόρια ανά κύτταρο σκώληκα. Εάν
αυτή η υψηλή έκφραση αποδίδεται σε γρήγορο ρυθμό μετάφρασης ή σε αργή
αποδόμηση δεν είναι ακόμη γνωστό. Μερικά miRNAs εκφράζονται όμως σε πολύ
μικρότερα επίπεδα. Για παράδειγμα το miR-124 υπάρχει στα κύτταρα του ενήλικου
σκώληκα - κατά μέσο όρο - σε 800 μόρια ανά κύτταρο (Lim et al., 2003a). Αυτό
μπορεί να οφείλεται σε χαμηλή έκφραση πολλών κυττάρων ή σε υψηλή έκφραση για
λίγα κύτταρα. [10]
Η βιολογία των miRNAs
Τα miRNAs είχαν αρχικά περιγραφεί ως μακρά πρόδρομα μόρια RNA, που
καλούνται primary miRNAs. Αυτά απαιτούσαν το ένζυμο RNase III Drosha στον
πυρήνα για να δώσουν το πρώιμο miRNA. Το πρόδρομο αυτό μόριο χαρακτηρίζεται
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

από ένα τμήμα 33 νουκλεοτιδίων δομής stem-loop ή φουρκέτας σε ένα συνολικό
μόριο 60-70 νουκλεοτιδίων, το οποίο εξέρχεται προς το κυτταρόπλασμα όπου
μετατρέπεται στο ώριμο miRNA των 21-26 νουκλεοτιδίων από ένα άλλο ένζυμο
(RNase III enzyme Dicer). [11]
Το ώριμο miRNA μπορεί να αντιδράσει με πρωτεΐνες Αργοναύτες για να
σχηματίσει το σύμπλοκο RISC και έπειτα να οδηγήσει το RISC στον στόχο mRNA,
κυρίως στην 3-αμετάφραστη περιοχή (3΄ UTR). Για να σχηματίσει το miRNA
λειτουργικές αλληλουχίες, το 5΄-τελικό άκρο (7-8 νουκλεοτιδίων) πρέπει να έχει
ακριβή συμπληρωματικότητα με το στόχο mRNA (seed-region) και μερική
συμπληρωματικότητα με την υπόλοιπη αλληλουχία. Ένα miRNA μπορεί είτε να
παρεμποδίσει τη μετάφραση είτε να προκαλέσει την αποικοδόμηση του στόχου
mRNA. Αυτό εξαρτάται κυρίως από το συνολικό βαθμό συμπληρωματικότητας της
περιοχής πρόσδεσης, τον αριθμό των σημείων πρόσδεσης και την προσβασιμότητά
τους (καθορίζονται από την ενέργεια ελεύθερης κατάστασης). Όσο μεγαλύτερη είναι
η συμπληρωματικότητα των προσβάσιμων περιοχών πρόσδεσης, τόσο περισσότερο
πιθανή είναι η αποδόμηση του στοχευμένου mRNA. Τα miRNAs που παρουσιάζουν
ατελή συμπληρωματικότητα με τον mRNA στόχο καταλήγουν σε παρεμποδισμένη
μετάφρασή τους. Όσο περισσότερο προσβάσιμες είναι οι περιοχές πρόσδεσης για ένα
miRNA, τόσο μεγαλύτερη είναι η δράση αυτών των miRNAs που οφείλεται στο
δυναμικό συνέργειας των miRNA-mRNA. [11]
Έχει προταθεί ότι η ρύθμιση της μετάφρασης από τα miRNAs ταλαντώνεται
μεταξύ καταστολής και ενεργοποίησης κατά τη διάρκεια του κυτταρικού κύκλου.[11]
Ο υψηλός βαθμός συντήρησης των αλληλουχιών μεταξύ των πολυκύτταρων
ζωικών οργανισμών προτείνει μια δυνατή εξελικτική πίεση και συμμετοχή των
miRNAs σε ουσιώδης βιολογικές διεργασίες όπως στον κυτταρικό πολλαπλασιασμό,
στη διαφοροποίηση, στην απόπτωση και στον μεταβολισμό. Είχε προβλεφθεί ότι
κάθε μόριο miRNA μπορεί να έχει πάνω από 1000 γονίδια ως στόχους και κάθε απλό
γονίδιο μπορεί να ρυθμιστεί από πολλά miRNAs. Αυτό συμβαίνει απλά επειδή οι
δράσεις των miRNAs έχουν εξειδίκευση στην αλληλουχία ή στο πρότυπο πρόσδεσης
και όχι στο γονίδιο. Διαφορετικά γονίδια μπορεί να έχουν παρόμοιο πρότυπο
πρόσδεσης για δεδομένο miRNA και ένα δεδομένο γονίδιο μπορεί να έχει πρότυπα
πρόσδεσης για διακριτά μεταξύ τους miRNAs. [11]
Γονιδιωματική: Υπολογιστικές προσεγγίσεις και αριθμός γονιδίων
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Υπάρχει προβληματισμός σχετικά με το γιατί τα miRNAs δε ανακαλύφθηκαν
νωρίτερα. Η απάντηση είναι, εν μέρει, σαφής: πάντως όχι γιατί είναι σπάνια. Τα
miRNAs και οι συσχετιζόμενες πρωτεΐνες φαίνεται να είναι από τα πιο διαδεδομένα
ριβονουκλεοπρωτεϊνικά σύμπλοκα στα κύτταρα. Ωστόσο, τα miRNAs που η έκφρασή
τους είναι περιορισμένη σε λίγους τύπους κυττάρων ή σε ειδικές περιβαλλοντικές
συνθήκες μπορεί και να χαθούν κατά τις προσπάθειες κλωνοποίησής τους. Έτσι, οι
υπολογιστικές προσεγγίσεις έχουν αναπτυχθεί ώστε να συμπληρώσουν τις
πειραματικές προσεγγίσεις στην ταυτοποίηση των γονιδίων των miRNAs. Οι έρευνες
για την αναζήτηση ομόλογων σειρών έχουν αποκαλύψει γονίδια (orthologs και
paralogs) γνωστών γονιδίων miRNA (Pasquinelli et al.,2000; Lagos-Quintana et al.,
2001; Lau et al., 2001; Lee and Ambros, 2001). Μια ακόμη απλή προσέγγιση έχει
διερευνήσει τη γειτνίαση γνωστών miRNA γονιδίων για άλλες δομές του τύπου stem-
loop που μπορεί να αντιπροσωπεύουν επιπρόσθετα γονίδια σε γονιδιακές συστοιχίες
(Lau et al., 2001, Aravin et al., 2003; Seitz et al., 2003; Ohler et al., 2004). Η
στρατηγική αυτή είναι σημαντική επειδή μερικά από τα ταχύτερα αναπτυσσόμενα
γονίδια miRNA παρουσιάζονται ως διαδοχικές σειρές σε συστοιχίες τύπου οπερονίου
(operon-like clusters) και οι διαφοροποιημένες αλληλουχίες αυτών των γονιδίων τα
κάνουν σχετικά δύσκολα στην αναγνώριση με τη χρήση γενικών μεθόδων
προσέγγισης. [10]
Τεχνικές εύρεσης γονιδίων που δε βασίζονται σε εύρεση ομόλογων ή
παρόμοιων αλληλουχιών με ήδη γνωστά γονίδια έχουν ήδη αναπτυχθεί και
εφαρμόζονται σε ολόκληρα γονιδιώματα (Ambros et al., 2003b; Grad et al., 2003; Lai
et al., 2003; Lim et al., 2003a). Οι δύο πιο ευαίσθητες μέθοδοι είναι οι MiRscan, που
έχει συστηματικά εφαρμοστεί στους νηματοσκώληκες και στα σπονδυλωτά (Lim et
al., 2003a, 2003b), και η miRseeker που εφαρμόστηκε στα έντομα (Lai et al., 2003).
Και οι δύο μέθοδοι έχουν αναγνωρίσει δεκάδες γονίδια που έπειτα από μικρό χρονικό
διάστημα ή και ταυτόχρονα αναγνωρίστηκαν και πειραματικά. Εξαιτίας της σχετικά
μεγάλης ευαισθησίας τους, και οι δύο μέθοδοι έχουν προσδιορίσει 200-255 γονίδια
miRNA στο ανθρώπινο γονιδίωμα (Lim et al, 2003b), 103-120 στο γονιδίωμα της
C.elegans (Lim et al., 2003a; Ohler et al., 2004) και 96-124 στο γονιδίωμα της
Δροσόφιλα (Lai et al., 2003). Σε κάθε ένα από τα παραπάνω είδη, τα νούμερα αυτά
αντιπροσωπεύουν το 1% των αναμενόμενων γονιδίων του γονιδιώματος, ένα ποσοστό
εφάμιλλο με το ποσοστό και άλλων ρυθμιστικών γονιδίων. [10]
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Τα miRAs στα φυτά
Η κλωνοποίηση μικρών RNAs από τα φυτά αποκάλυψε επίσης miRNAs,
παρόλο που το πλήθος των άλλων 21 με 24 νουκλεοτιδίων RNAs που βρίσκονται στα
φυτά δυσκόλεψε την ταξινόμησή τους (Liave et al., 2002a; Mette et al., 2002; Park et
al., 2002; Reinhart et al., 2002). Όπως και στους πολυκύτταρους ζωικούς
οργανισμούς, τα φυτικά miRNAs (1) εκφράζονται ενδογενώς ως μόρια ~22
νουκλεοτιδίων, (2) είναι γενικά συντηρημένα κατά την εξέλιξη και (3) προέρχονται
από περιοχές του γονιδιώματος διακριτές σε σχέση με τα ήδη επισημασμένα γονίδια
(Reinhart et al., 2002). [10]
Η ανακάλυψη των miRNAs στα φυτά και στα ζώα απέδειξε ότι αυτή η τάξη
των μη μεταφραζόμενων μορίων RNA έχει διαφοροποιήσει τη γονιδιακή έκφραση
τουλάχιστον από τη στιγμή της ύπαρξης του τελευταίου κοινού γενεαλογικού
προγόνου (Reinhart et al., 2002). [10]
Ωστόσο, τα miRNAs στα φυτά και στα ζώα διαφέρουν σε κάποια σημεία που
φαίνεται να σχετίζονται με διαφορές στη βιογένεση. Η πλέον αξιοσημείωτη διαφορά
βρίσκεται στη δομή stem-loop. Οι αναδιπλώσεις της δομής αυτής στα φυτά
διαφέρουν στο μέγεθος και είναι τυπικά μεγαλύτερες από αυτές που υπάρχουν στα
ζώα (σχήμα 4.2). Άλλες πιο λεπτές διαφορές περιλαμβάνουν καλύτερο ζευγάρωμα
μεταξύ των miRNAs και του βραχίονα στα φυτά σε σχέση με τα ζώα και ίσως μια πιο
δυνατή προτίμηση για δομή U στο 5΄ τελικό άκρο στο φυτικό mRNA (Lau et al.,
2001, Reinhart et al., 2002; Bartel and Bartel, 2003). Οι διαφορές αυτές, σε
συνδυασμό με την απουσία αναφορών ότι συγκεκριμένα γονίδια miRNA
συντηρούνται μεταξύ φυτών και ζώων, αφήνουν ανοιχτή την προοπτική ότι τα
γονίδια miRNA εμφανίστηκαν ανεξάρτητα σε αυτούς τους πολυκυτταρικούς
γενεαλογικούς οργανισμούς έπειτα από τον τελευταίο τους κοινό πρόγονο, για τον
οποίο θεωρούμε ότι μάλλον ήταν μονοκυτταρικός. [10]
Ο ρυθμιστικός ρόλος των miRNAs
Το πιο πιεστικό ερώτημα που ανακύπτει από την ανακάλυψη εκατοντάδων
διαφορετικών miRNAs είναι το εξής: ποιος είναι ο ρόλος όλων των παραπάνω
μικροσκοπικών μορίων RNA; Για τα lin-4, let-7 και άλλα miRNAs που έχουν
αναγνωριστεί, κρίσιμες ενδείξεις για τη λειτουργία τους και τη ρυθμιστική τους
ικανότητα ανακαλύφθηκαν γρήγορα, πριν καν γίνει ακόμη γνωστό ότι αποτελούν μη
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

κωδικά μόρια (Meneelt and Herman, 1979; Chalfie et al., 1981; Ambros, 1989;
Weigel et al., 2000; Hipfner et al., 2002; Aukerman and Sakai, 2003; Brennecke et al.,
2003; Johnston and Hobert, 2003; Xu et al., 2003). Oι λειτουργίες αυτών καθώς και
άλλων γονιδίων που προσδιορίστηκαν με in vivo πειράματα, αναφέρονται στον
πίνακα 4.1. [10]
Πίνακας 4.1 Ορισμένα miRNAs και οι λειτουργίες τους.
Για κάποιες από αυτές τις περιπτώσεις, η λειτουργία προσδιορίστηκε με
πειράματα in vivo σε μεταλλαγμένα μόρια miRNA ή σε μεταλλαγμένες
συμπληρωματικές αλληλουχίες που ήταν δυνατό να διαταράξουν τη φυσιολογική
τους λειτουργία. Σε άλλες περιπτώσεις, η λειτουργία των miRNAs προέκυψε ως
συμπέρασμα από γεγονότα σε μεταλλαγμένες διαγονιδιακές περιοχές που οδήγησαν
σε εκτοπική έκφραση του miRNA. [10]
Για τη μεγάλη πλειοψηφία των miRNAs, οι φαινοτυπικές συνέπειες σε
τροποποιημένα ή μεταλλαγμένα μόρια δεν είναι ακόμη γνωστές. Ωστόσο έχουν
αναπτυχθεί υπολογιστικές προσεγγίσεις για να βρεθούν οι ρυθμιστικοί στόχοι των
miRNAs, που παρέχουν ενδείξεις για τη λειτουργία τους, βασισμένες στους γνωστούς
ρόλους αυτών των στόχων (Rhoades et al., 2002; Enright et al., 2003; Lewis et al.,
2003; Stark et al., 2003). Υπολογιστικά αναμενόμενοι στόχοι συνοδευμένοι από
πειράματα ή ανεξάρτητα φυλογενετικά δεδομένα δίνονται στον πίνακα 4.2. [10]
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Πίνακας 4.2 Ορισμένα μόρια miRNA και οι λειτουργίες τους.
Τα miRNAs των κυττάρων του ξενιστή και των ιών
Τα miRNAs αποτελούν ένα νέο παράδειγμα RNA-καθορισμένης γενετικής
έκφρασης και ρύθμισης ενός μεγάλου εύρους βιολογικών συστημάτων. Αυτά τα
μικρά μη-κωδικά μόρια RNA μπορούν να συνεισφέρουν σε ένα πλούσιο ρεπερτόριο
αλληλεπιδράσεων μεταξύ ξενιστή-ιού κατά την προσβολή του ξενιστή από έναν ιό. Η
διαδραστικότητα αυτή έχει σημαντικές επιπτώσεις, τόσο για τους ιούς όσο και για τον
ξενιστή. Έχουν αναφερθεί ενδείξεις μορίων miRNA σε κύτταρα ξενιστών που
καθορίζουν την έκφραση διάφορων ιικών γονιδίων, παίζοντας έτσι σπουδαίο ρόλο για
το ανοσοβιολογικό σύστημα του ξενιστή. Στο παιχνίδι μεταξύ παθογόνου ιού και
ξενιστή, οι ιοί ανέπτυξαν εξειδικευμένους μηχανισμούς σίγησης των γονιδίων ώστε
να αποφεύγουν τις συνέπειες της ανοσοβιολογικής απόκρισης. Πρόσφατες αναφορές
δείχνουν ότι οι ιοί κωδικοποιούν miRNAs που τους προστατεύουν από την κυτταρική
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

ανοσοβιολογική απόκριση. Επιπλέον, μπορούν να εκμεταλλευτούν κυτταρικές
μεταβολικές οδούς προς δικό τους όφελος. Παρόλα αυτά οι συνεχώς αυξανόμενες
γνώσεις μας για τις αλληλεπιδράσεις των ιών με τα κύτταρα του ξενιστή σε μοριακό
επίπεδο μας οδηγούν σε πιθανές εξηγήσεις για την ανάπτυξη αποτελεσματικής,
ανθεκτικής και μη τοξικής για τον ξενιστή αντιικής θεραπείας. [12]
Εφόσον τα miRNAs είναι ευρέως διαδεδομένα στη φύση, δεν αποτελεί
έκπληξη ότι οι ιοί κωδικοποιούν miRNAs. Κατά τη διάρκεια της εξέλιξης, οι ιοί
ανέπτυξαν μηχανισμούς ώστε να εκμεταλλεύονται τους βιοσυνθετικούς μηχανισμούς
του ξενιστή και να εξουδετερώνουν τους αμυντικούς μηχανισμούς. Η έρευνα σήμερα
αποκάλυψε αλληλεπιδράσεις που αφορούν την RNA σίγηση. [12]
Οι ιοί παρουσιάζουν ενδιαφέρουσες αλληλεπιδράσεις με τα κύτταρα των
ξενιστών, οι οποίες δημιουργούν προβλήματα στην αδρανοποίηση των ιών χωρίς να
προκληθεί ζημιά στο κύτταρο του ξενιστή. Η καταπολέμηση της ιικής μόλυνσης,
χωρίς να επηρεαστούν τα κύτταρα του ξενιστή, στοχεύοντας σε πρωτεΐνες και
μεταβολικές οδούς των ιών μοναδικούς για τους κύκλους της ζωής τους, κατέστη
δυνατή μόνο για λίγους. Έτσι, μόνο λίγα αντιικά φάρμακα υπάρχουν. Η αντίσταση
των ιών και η τοξικότητα των φαρμάκων αποτελούν σημαντικά προβλήματα στην
αντιική θεραπεία. Αυτό έχει οδηγήσει στην ανάγκη να τιθασεύσουμε τις δυνατότητες
της μεθόδου RNA interference (RNAi) ως έναν εσωτερικό αντιικό αμυντικό
μηχανισμό. [12]
Η μέθοδος RNAi αντιπροσωπεύει ένα ζωτικής σημασίας συστατικό της
εσωτερικής ανοσοβιολογικής απόκρισης στα φυτά και στα ασπόνδυλα ζώα. Επιπλέον
υπηρετεί ως ένας ρυθμιστικός για τα γονίδια μηχανισμός, ο οποίος ενεργοποιείται
από την έκφραση των μορίων miRNA. Ωστόσο, ο ρόλος των κυτταρικών miRNAs
στην άμυνα απέναντι στην ιική μόλυνση των θηλαστικών παραμένει ακόμη ασαφής.
Για αυτό είναι σημαντικό να κατανοήσουμε τις λεπτομέρειες σχετικά με την επιρροή
του πολλαπλασιασμού των ιών στην αφθονία και κατανομή των miRNAs μέσα στα
κύτταρα του ξενιστή. Έχει προταθεί ότι το κυτταρικό miRNA είναι δυνατό να έχει
ουσιώδες αποτέλεσμα στην εξάπλωση των ιών και να έχει το δυναμικό να ρυθμίζει
τον τροπισμό των ιών in vivo. Οι ιοί αξιοποιούν μηχανισμούς σίγησης γονιδίων που
επάγονται από μόρια miRNA, κωδικοποιώντας τα δικά τους miRNAs. Έτσι, η μελέτη
των αλλαγών στο τοπίο των miRNAs κατά την αναπαραγωγή των ιών μπορεί να μας
βοηθήσει να κατανοήσουμε τη ρύθμιση στην άμυνα των ξενιστών και την
προσπάθεια των ιών να ξεπεράσουν τους μηχανισμούς άμυνας. Ένα μεγάλο εύρος
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

αλληλεπιδράσεων είναι πιθανό διαμέσου miRNA-mRNA υβριδισμού κατά τη
διάρκεια των αλληλεπιδράσεων ξενιστή-ιού. [12]
Ο Thomas Tuschl και η ομάδα του στο πανεπιστήμιο Rockfeller ανέφεραν την
ύπαρξη ιικών miRNAs για πρώτη φορά στον ιό Epstein-Barr (EBV). Μέχρι σήμερα,
141 miRNA γονίδια έχουν ταυτοποιηθεί σε 15 ιούς από τρεις οικογένειες ιών
(herpesvirus, polyomavirus και retrovirus). [12]
Οι βιογενετικοί οδοί των miRNAs θέτουν ορισμένα σοβαρά προβλήματα για
τους RNA ιούς και ένα σύνολο DNA ιών στην κωδικοποίηση των miRNAs. Ωστόσο,
είναι πιθανό να ξεπεραστούνε ορισμένα προβλήματα υιοθετώντας μη συμβατικές
μεθόδους. Στην εργασία των Omoto et al., έχει αναφερθεί η παρουσία miRNAs σε
κύτταρα που έχουν προσβληθεί από τον ιό HIV-1 (o οποίος είναι RNA ιός), παρότι
εκτεταμένες έρευνες από τους Pfeffer et al. όπως και από τους Lin και Cullen έχουν
αποτύχει να επιβεβαιώσουν την παρουσία ιικών miRNAs στον ιό HIV. [12]
Για την κατανόηση του πιθανού ρόλου των miRNAs εξαιτίας των ιών και για
τις μελέτες ολόκληρου του φάσματος των αλληλεπιδράσεων μεταξύ ιών και ξενιστή
σε μοριακό επίπεδο, απαιτείται μελλοντικά ο ακριβής προσδιορισμός του χάρτη του
γονιδιώματος και της διαδικασίας της μεταγραφής για τα miRNAs. [12]
Ο ρόλος των miRNAs στις καρδιαγγειακές παθήσεις
Οι καρδιαγγειακές παθήσεις βρίσκονται ανάμεσα στα κυριότερες αιτίες
νοσηρότητας και θνησιμότητας στις ανεπτυγμένες χώρες. Οι παθολογικές
καταστάσεις της καρδιάς σχετίζονται με μεταβολές στο προφίλ των γονιδίων που
είναι σημαντικές για τις λειτουργίες της καρδιάς. Τα miRNAs έχουν αναδειχθεί ως
κεντρικός παράγοντας στη ρύθμιση της γονιδιακής έκφρασης. Οι επιπτώσεις των
miRNAs σε παθολογικές διαδικασίες του καρδιαγγειακού συστήματος έχουν
πρόσφατα αναγνωρισθεί και αντιπροσωπεύουν σήμερα ένα ταχύτητα αναπτυσσόμενα
ερευνητικό πεδίο. [11]
Οι εφαρμογές της RNAi τεχνολογίας στη ρύθμιση της έκφρασης θα
αποτελέσει σημαντικό παράγοντα στη θεραπεία σε μοριακό επίπεδο. [11]
Βιβλιογραφία
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

1. Susi P., Hohkuri M., Wahlroos T., Kilby N.J. ‘‘Characteristics of RNA
silencing in plants: similarities and differencies across kingdoms’’, Plant
Molecular Biology, 54: 157-174, 2004.
2. Mario Arteaga-Vazquez, Juan Caballero-Perez and Jean-Philippe Vielle-
Calzada ‘‘A Family of MicroRNAs Present in Plants and Animals’’, The Plant
Cell, Vol 18, 3355-3369, December 2006.
3. T. M. Hammond and N. P. Keller RNA ‘‘Silencing in Aspergillus nidulans Is
Independent of RNA-Dependent RNA Polymerases’’, Genetics Society of
America, 2005 February; 169(2): 607–617.
4. Zhang B., Pan X., Cobb G., Anderson T., ‘‘microRNAs as oncogenes and
tumor suppressors’’, Developmental Biology, 302 (2007) 1-12.
5. Lee P. Lim, Nelson C. Lau, Earl G. Weinstein, Aliaa Abdelhakim, Soraya
Yekta, Matthew W. Rhoades, Christopher B. Burge, and David P. Bartel ‘‘The
microRNAs of Caenorhabditis elegans’’ Genes Dev. 2003 April 15; 17(8):
991–1008.
6. Jones-Rhoades, Bartel DP, Bartel B. ‘‘MicroRNAs and their regulatory roles
in plants’’, Annu Rev Plant Biol. 2006; 57:19-53.
7. Kuniaki Saito, Akira Ishizuka, Haruhiko Siomi, and Mikiko C Siomi
‘‘Processing of Pre-microRNAs by the Dicer-1–Loquacious Complex in
Drosophila Cells’’ PLoS Biol. 2005 July; 3(7): e235. Published online 2005
May 24.
8. Junjie Li, Zhiyong Yang, Bin Yu, Jun Liu and Xuemei Chen Methylation
Protects miRNAs and siRNAs from a 3′-End Uridylation Activity in
Arabidopsis Current Biology vol 15, 1501-1507, 2005
9. Eric C Lai ‘‘Predicting and validating microRNA targets’’ Genome Biol.
2004; 5(9): 115 Published online 2004 August 3.
10. Bartel D. MicroRNAs: ‘‘Genomics, Mechanism, and Function’’, Cell, Vol
116, 281-297, January 23, 2004.
11. Wang Z., Luo X., Lu Y., Yang B., ‘‘miRNAs at the heart of the matter’’, J
Mol Med (2008) 86:771-783.
12. Ghosh Z., Mallick B., Chakrabarti J., ‘‘Cellular versus viral microRNAs in
host-virus interaction’’, Nucleic Acids Research, 2009, Vol. 37, No 4 1035-
1048.
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

ΘΕΜΑ 5
A) First, some clarification.Η διαφορά μεταξύ του RNA (ριβονουκλεϊκό οξύ)
και του DNA (δεοξυριβονουκλεϊκό οξύ) είναι η ύπαρξη της ομάδας του υδροξυλίου
στον 2΄ άνθρακα της ριβόζης.
Σχήμα 5.1 Η διαφορά μεταξύ ριβόζης και δεοξυριβόζης βρίσκεται στο 2΄-ΟΗ.
ΗThe removal of 2' hydroxyl groups from DNA does not occur after the DNA
has been synthesized, but rather the 2' hydroxyl groups are removed from the
nucleotides before they are incorporated into the DNA. αφαίρεση των ομάδων 2΄-
υδροξυλίου από το DNA δεν συμβαίνει αφού το DNA έχει συντεθεί, αλλά οι ομάδες
2΄-υδροξυλίου μάλλον απομακρύνονται από το νουκλεοτίδια πριν αυτά
ενσωματωθούν στο DNA.During , a portion of the nucleotide monophosphates
(NMP's) are dehydroxylated to 2'-deoxy-nucleotide monophosphates (dNMP's). Κατά
τη διάρκεια της νουκλεοτιδικής σύνθεσης, ένα μέρος των μονοφωσφορικών
νουκλεοτιδίων (NMP) είναι αποϋδροξυλιωμένα σε 2'-δεοξυ-μονοφωσφορικά
νουκλεοτιδία (dNMP). This means that GMP, AMP, CMP, and UMP are converted
into dGMP, dAMP, dCMP, and dUMP, respectively. However, before being
incorporated into the chromosomes, another modification, using as a catalyst,
methylates the uracil in dUMP to form a thymine making it dTMP.
Αυτό σημαίνει ότι και τα μονοφωσφορικά GMP, AMP, CMP και UMP
μετατρέπονται σε dGMP, dAMP, dCMP, και dUMP αντίστοιχα. Ωστόσο, πριν
ενσωματωθούν στα χρωμοσώματα, μια άλλη τροποποίηση, χρησιμοποιώντας φυλλικό
οξύ ως καταλύτη, μεθυλιώνει την ουρακίλη της DUMP για να σχηματίσει θυμίνη,
δημιουργώντας τη dTMP. After further phosphorylation, dGTP, dATP, dCTP, and
dTTP can be used as the building blocks to construct DNA. Μετά από περαιτέρω
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

φωσφορυλίωση, τα dGTP, dATP, dCTP, και dTTP μπορούν να χρησιμοποιηθούν ως
δομικά στοιχεία για την κατασκευή του DNA.
The important thing to notice is that while uracil exists as both uridine (U) and
deoxy-uridine (dU), thymine only exists as deoxy-thymidine (dT). So the question
becomes: Why do cells go to the trouble of methylating uracil to thymine before it can
be used in DNA?Το σημαντικό εδώ είναι να παρατηρήσουμε ότι, ενώ η ουρακίλη
υπάρχει με δύο μορφές, ουριδίνη (U) και δεοξυ-ουριδίνη (DU), η θυμίνη υπάρχει
μόνο ως δεοξυ-θυμίνη (dT). Προκύπτει λοιπόν το εξής ερώτημα: Γιατί τα κύτταρα
μπήκαν στον κόπο να μεθυλιώσουν την ουρακίλη δημιουργώντας θυμίνη, πριν αυτή
να μπορεί να χρησιμοποιηθεί στο DNA;
Με βάση τις παραπάνω σκέψεις, θα μπορούσαμε πλέον να
επαναδιατυπώσουμε το However, the real question is: Why does thymine replace
uracil in DNA?ερώτημα της εκφώνησης ως εξής: Γιατί η θυμίνη να αντικαταστήσει
την ουρακίλη στο DNA;
Σχήμα 5.2 Η θυμίνη διαφέρει κατά μια μεθυλομάδα σε σχέση με την ουρακίλη. Η
απαμίνωση της κυτοσίνης οδηγεί σε ουρακίλη.
The answer is: methylation protects the DNA.Η απάντηση θα μπορούσε να
είναι απλά η εξής: η μεθυλίωση προστατεύει το DNA.Beside using dT instead of dU,
most organisms also use various enzymes to modify DNA after it has been
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

synthesized. Εκτός από τη χρήση dT αντί του dU, οι περισσότεροι οργανισμοί
χρησιμοποιούν διάφορα ένζυμα για την τροποποίηση του DNA μετά την σύνθεσή
του. Two such enzymes, dam and dcm methylate adenines and cytosines, respectively,
along the entire DNA strand.Δύο τέτοια ένζυμα, τα dam και dcm, μεθυλιώνουν τις
αδενίνες και κυτοσίνες αντίστοιχα, κατά μήκος ολόκληρων των κλώνων του DNA.
This methylation makes the DNA unrecognizable to many Nucleases (enzymes which
break down DNA and RNA), so that it cannot be easily attacked by invaders, like
viruses or certain bacteria.Αυτή η μεθυλίωση καθιστά το DNA ως μη αναγνωρίσιμο
σε πολλές νουκλεάσες (ένζυμα που διασπούν DNA και RNA), έτσι ώστε να μην
μπορεί να προσβληθεί εύκολα από τους εισβολείς, όπως οι ιοί ή ορισμένα
βακτηρίδια.
Obviously, methylating the nucleotides before they are incorporated ensures
that the entire strand of DNA is protected.Προφανώς, η μεθυλίωση των
νουκλεοτιδίων πριν αυτά ενσωματωθούν στο DNA, διασφαλίζει ότι ολόκληρο το
DNA είναι προστατευμένο. ΗThymine also protects the DNA in another way. θυμίνη
προστατεύει επίσης το DNA με έναν ακόμη τρόπο. If you look at the components of
nucleic acids, phosphates, sugars, and bases, you see that they are all very hydrophilic
(water soluble).Αν δει κανείς τα συστατικά στοιχεία των νουκλεϊνικών οξέων,
δηλαδή τα φωσφορικά άλατα, τα σάκχαρα, καθώς και τις βάσεις, θα παρατηρήσει ότι
όλα είναι υδρόφιλα. Obviously, adding a hydrophobic (water insoluble) methyl group
to part of the DNA is going to change the characteristics of the molecule.Η προσθήκη
της υδρόφοβης (αδιάλυτης στο νερό) ομάδας του μεθυλίου σε τμήμα του DNA, είναι
ικανή να αλλάζει τα χαρακτηριστικά του μορίου. The major effect is that the methyl
group will be repelled by the rest of the DNA, moving it to a fixed position in the
major groove of the helix.Το σημαντικό αποτέλεσμα είναι ότι η ομάδα του μεθυλίου
απωθείται από το υπόλοιπο του DNA, και μεταφέρεται σε σταθερή θέση στις μεγάλες
αύλακες της δομής της διπλής έλικας. This solves an important problem with uracil -
though it prefers adenine, uracil can base-pair with almost any other base, including
itself, depending on how it situates itself in the helix.Αυτό λύνει ένα σημαντικό
πρόβλημα καθώς η ουρακίλη μπορεί να σχηματίσει ζεύγος με σχεδόν οποιαδήποτε
άλλη βάση, συμπεριλαμβανομένης της ίδιας, ανάλογα με το πώς τοποθετείται στην
έλικα. By tacking it down to a single conformation, the methyl group restricts uracil
(thymine) to pairing only with adenine.Η ομάδα του μεθυλίου περιορίζει την
ουρακίλη (θυμίνη πλέον) να συνδέεται μόνο με αδενίνη. This greatly improves the
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

efficiency of DNA replication, by reducing the rate of mismatches, and thus
mutations.Αυτό βελτιώνει σημαντικά την αποτελεσματικότητα της αντιγραφής του
DNA εφόσον μειώνει το ποσοστό των αναντιστοιχιών που θα προέκυπταν χωρίς τη
μεθυλίωση και συνεπώς των μεταλλάξεων.
To sum up: the replacement of thymine for uracil in DNA protects the DNA
from attack and maintains the fidelity of DNA replication. [ Moderator Note: In
addtion, the cytosine base can spontaneously deaminate to form a uracil base, which
would result in undetectable C -> U mutations if U were used routinely in
DNA.Επιπλέον, η κυτοσίνη μπορεί αυθόρμητα να υποστεί απαμίνωση και να
σχηματίσει ουρακίλη, η οποία θα οδηγήσει σε μη ανιχνεύσιμες C→U μεταλλάξεις
εάν η ουρακίλη βρισκότανε σε μεγάλη συχνότητα στο DNA.
Since Thymine is basically methyl-U, the cell's DNA repair mechanisms can
distinguish illegitimate U from legitimate methyl-U in DNA, and make the proper
repair (replacing any U with a C).Δεδομένου ότι η θυμίνη είναι μεθυλιωμένη σε
σχέση με την ουρακίλη, οι μηχανισμοί επιδιόρθωσης του κυτταρικού DNA μπορούν
να διακρίνουν τη νόθα U από τη νόμιμες μεθυλο-U στο DNA, και να προβούν στην
κατάλληλη ενέργεια (αντικατάσταση U με C). C -> U mutations in RNA do not
matter as much, because RNA is synthesized in large quantities and is rapidly
degraded in comparison to DNA. Οι C→U μεταλλάξεις στο RNA δεν έχουν σημασία
τόσο πολύ, διότι το RNA συντίθεται σε μεγάλες ποσότητες και η αποδόμησή του
είναι ραγδαία σε σύγκριση με αυτή το DNA.
-- Steve Mack, MadSci Moderator.]Β) Η ανακάλυψη της αντίστροφης
μεταγραφάσης ανέτρεψε το αρχικά θεωρούμενο κεντρικό δόγμα της βιολογίας,
εφόσον πρόσθεσε μια ακόμη παράμετρο: η γενετική πληροφορία μπορεί να ρέει από
το RNA στο DNA, από το DNA πίσω στο RNA και μετά στις πρωτεΐνες. Η μορφή
που έλαβε η ροή της γενετικής πληροφορίας με την προσθήκη της αντίστροφης
μεταγραφάσης έχει ως εξής:
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Ο ιός HIV περιλαμβάνει ένα γονιδίωμα που αποτελείται από δυο
πανομοιότυπα μόρια RNA ενωμένα μεταξύ τους με ένα μόριο tRNA. Καθένα από τα
δύο μόρια είναι συμπλοκοποιημένο με ένα μόριο αντίστροφης μεταγραφάσης. Θα
μπορούσαμε να φανταστούμε τι θα συμβεί σε ένα ευκαρυωτικό κύτταρο στο οποίο
υπάρχουν μόρια αντίστροφης μεταγραφάσης σε περίσσεια, αν παρακολουθήσουμε σε
αδρές γραμμές τη μολυσματική πορεία ενός ιού που περιέχει το ίδιο ένζυμο, σε
περιορισμένη βέβαια ποσότητα.
Εφόσον ο ιός εισέλθει στον ξενιστή, η αντίστροφη μεταγραφάση αντιγράφει
μέσα στο κυτταρόπλασμα τον κλώνο του RNA σε έναν κλώνο DNA, δημιουργώντας
έτσι ένα υβρίδιο RNA-DNA. Ακολούθως συντίθεται ένας συμπληρωματικός κλώνος
DNA από την ίδια μεταγραφάση με εκμαγείο τον πρώτο κλώνο DNA που συντέθηκε,
το δε RNA υδρολύεται από τη μεταγραφάση. Το δίκλωνο πια μόριο DNA, που στον
έναν κλώνο του περιέχει τα γονίδια του ιού, μεταφέρεται στον πυρήνα του ξενιστή,
όπου με την παρεμβολή του ιογενούς ενζύμου ιντεγράσης ενσωματώνεται στο
γονιδίωμα του κυττάρου ως προϊός. Ενώ είναι ενσωματωμένος ο προϊός στο
κυτταρικό γονιδίωμα, τα γονίδιά του μπορούν να μεταγράφονται από την κυτταρική
πολυμεράση του RNA, τα δε mRNA που προκύπτουν μεταφράζονται από τον
κυτταρικό μηχανισμό πρωτεϊνοσύνθεσης.
Αρχικά, για τα ευκαρυωτικά κύτταρα των ζωικών οργανισμών πρέπει να
υπολογίζουμε ότι το RNA βρίσκεται στον πυρήνα, στο κυτταρόπλασμα και στα
μιτοχόνδρια των ευκαρυωτικών κυττάρων. Το κυτταροπλασματικό RNA αποτελείται
από ριβοσωμικό RNA (rRNA), μεταφορικό RNA (tRNA), αγγελιοφόρο RNA
(mRNA) και άλλα μικρά τμήματα RNA. Στον πυρήνα βρίσκεται το ετεροπυρηνικό
RNA (hnRNA), πρόδρομος του RNA. Μόνο το 1-2% του ολικού RNA στα
ευκαρυωτικά κύτταρα είναι mRNA. Η πλειοψηφία του ολικού RNA είναι rRNA. Η
ποσότητα μάλιστα του απαντώμενου mRNA θεωρείται ότι αντανακλά το επίπεδο
μεταγραφής ενός γονιδίου.
Ας θεωρήσουμε για ένα κύτταρο ενεργή και σε μεγάλες ποσότητες
αντίστροφη μεταγραφάση. Εφόσον τα μόρια του ενζύμου υπάρχουν σε αφθονία,
συμπλέκονται με τα μόρια του mRNA, τα οποία και χρησιμοποιούνται ως καλούπι
για τη σύνθεση μονόκλωνου DNA. To μόριο που σχηματίζεται με τον τρόπο αυτό
είναι ένα δίκλωνο μόριο mRNA-DNA το οποίο πιθανότατα δεν είναι πολύ σταθερό.
Αυτό το υποθέτουμε επειδή γνωρίζουμε τις δυσκολίες που πειραματικά προκύπτουν
για την επιτυχή απομόνωση ακέραιου RNA από τα κύτταρα με τη μέθοδο που απαιτεί
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

την πραγματοποίηση τεσσάρων βημάτων (διάσπαση των κυττάρων - μετουσίωση των
συμπλόκων νουκλεοπρωτεϊνών - απενεργοποίηση της δράσης των ενδογενών
ριβονουκλεασών, RNase, και καθαρισμό του RNA από DNA και πρωτεΐνες).
Έτσι γνωρίζουμε ότι το RNA είναι επιρρεπές στην αποικοδόμηση και πρέπει
να λαμβάνονται πολλές προφυλάξεις κατά την απομόνωσή του.
To RNA λοιπόν θα υδρολυθεί και η ίδια μεταγραφάση με εκμαγείο τον πρώτο
κλώνο DNA που συντέθηκε αρχικά, θα συνθέσει ένα συμπληρωματικό κλώνο DNA.
Κατά αυτόν τον τρόπο οδηγούμαστε σε ένα δίκλωνο μόριο DNA. Για να
δημιουργηθεί βέβαια το υβριδικό μόριο mRNA-DNA απαιτείται ένα μόριο ως
εκκινητήρας (primer). Το ρόλο αυτό αναλαμβάνει το tRNA.
Ως συνέπεια της παραπάνω διαδικασίας, η ποσότητα του mRNA που
παράγεται κατά τη διαδικασία της μεταγραφής δεσμεύεται συνεχώς από την
αντίστροφη μεταγραφάση. Σε ένα κύτταρο που δεν υπάρχει πλέον mRNA για να
συνδεθεί με τα ριβοσώματα, δε λαμβάνει χώρα πρωτεϊνοσύνθεση. Το κύτταρο αυτό
δε θα έχει τις απαραίτητες πρωτεΐνες αλλά θα έχει DNA σε μια συνεχώς αυξανόμενη
ποσότητα.
Βέβαια, οι νεοσυντιθέμενες ποσότητες του DNA θα αφορούσαν ουσιαστικά
στα γονίδια που εκφράζονται σε κάθε κυτταρικό τύπο και όχι στις υπόλοιπες μη
κωδικοποιούσες περιοχές του γονιδιώματος. Και αυτό συμπεραίνουμε επειδή
εμπειρική παρατήρηση δείχνει ότι, μολονότι όλα τα εμπύρηνα κύτταρα του
οργανισμού έχουν το ίδιο ακριβώς DNA, δηλαδή τα ίδια ακριβώς γονίδια, εντούτοις
τα γονίδια τα οποία μεταγράφονται ανά πάσα στιγμή αποτελούν μικρό μόνο μέρος
του συνόλου του γονιδιώματος και το σημαντικότερο είναι ότι το είδος των γονιδίων
που μεταγράφεται εξαρτάται από το είδος του κυττάρου. Τότε, διαμέσου της
διαδικασίας της υπόθεσης (αντίστροφη μεταγραφάση σε περίσσεια) θα μειωνότανε η
ποσότητα των mRNAs και θα δημιουργούτανε αντίγραφα DNA των γονιδίων που η
ρύθμιση της έκφρασης επιτρέπει να εκφράζονται. Σε κάθε τύπο κύτταρου λοιπόν θα
υπήρχε αυξημένη ποσότητα DNA, πλέον της φυσιολογικής που συγκροτεί το
χαρακτηριστικό για το είδος γονιδίωμα, μόνο όμως των γονιδίων που εκφράζονται.
Μια συνέπεια που μπορούμε να φανταστούμε είναι ότι τα κύτταρα σε ένα
πολυκύτταρο ζωικό οργανισμό δε θα είχανε πλέον την ίδια ποσότητα DNA αλλά
σταδιακά θα κατέληγαν σε διαφορετική ποσότητα DNA ανά τύπο. Όσον αφορά στα
γονίδια που εκφράζονται σε όλα σχεδόν τα κύτταρα, θα παρήγαγαν DNA που θα ήταν
κοινό για όλους τους τύπους.
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ

Επίσης, εάν στον πυρήνα του υποθετικού ζωικού κυττάρου υπήρχε ένα ένζυμο
με δράση παρόμοια με αυτή της ιντεγράσης, το DNA θα ενσωματωνότανε στο
γονιδίωμα, οδηγώντας μακροπρόθεσμα σε αλλαγή της ποσότητας του γενετικού
υλικού των κυττάρων με απρόβλεπτες συνέπειες για το είδος αυτό. Εάν λάβουμε
υπόψη μας ότι το DNA είναι πιο σταθερό μόριο από το RNA, ίσως μακροπρόθεσμα
να αλλοιωνότανε ο αριθμός των χρωμοσωμάτων για το είδος που συζητάμε.
Γιάννης Γεννάδιος
ΓΙΑΝΝΗΣ ΓΕΝΝΑΔΙΟΣ
Recommended