
ISSN 2073-5529. Âåñòíèê ÀÃÒÓ. Ñåð.: Ðûáíîå õîçÿéñòâî. 2009. № 1
44
УДК 612.821/639.3
Н. Н. Федорова, В. В. Залепухин
ÈÑÏÎËÜÇÎÂÀÍÈÅ ÒÅÎÐÈÈ ÔÓÍÊÖÈÎÍÀËÜÍÛÕ ÑÈÑÒÅÌ Â ÐÛÁÎÕÎÇßÉÑÒÂÅÍÍÛÕ ÈÑÑËÅÄÎÂÀÍÈßÕ
Введение Начиная с 1950-х гг. академиком П. К. Анохиным разрабатывалась теория функциональных
систем, первоначально примененная к физиологии высшей нервной деятельности, а точнее – к системной организации нервных функций. В работах П. К. Анохина и его учеников [1–3] были заложены основы комплексного подхода к деятельности центральной нервной системы (ЦНС). Та-ким образом, среди разнообразных систем человеческого организма, включающего в себя крове-носную, дыхательную, двигательную и прочие постоянно взаимодействующие системы, в качестве объекта исследований была избрана именно ЦНС. В более поздних публикациях П. К. Анохин ана-лизировал функциональные системы кровообращения, питания, специфических форм передвижения, механизмы формирования и обеспечения функций, особенности их проявления и регуляции [4].
По П. К. Анохину, «… функциональная система – это замкнутая саморегулирующаяся структура, все компоненты которой взаимно содействуют достижению полезного для организма результата, с постоянной сигнализацией о достижении такого результата» [4]. Главными чертами каждой функциональной системы являются динамичность, гетерогенность компонентов, резуль-тативность, саморегуляция, наличие обратных связей: «…Функциональные системы складывают-ся из динамически мобилизуемых структур в масштабе целого организма, и на их деятельности не отражается исключительное влияние какого-либо анатомического типа участвующей структу-ры. Более того, компоненты той или иной анатомической принадлежности мобилизуются и во-влекаются в функциональную систему только в меру их содействия получению запрограммиро-ванного результата. Одним из наиболее важных свойств функциональной системы является дина-мическая изменчивость входящих в нее структурных компонентов, обеспечивающая получение полезного результата [1]. Частные физиологические процессы при выполнении какой-либо при-способительной функции образуют единое целое, обладающее спецификой связей, отношений и взаимных влияний составляющих компонентов именно в тот момент, когда все части организма мобилизованы на выполнение функции. П. К. Анохин выделял функциональную систему как строго очерченную группу процессов и структур, объединенных для выполнения какой-либо качественно своеобразной функции организма или акта его поведения [4].
При расширительном толковании такого определения и свойств функциональных систем
представления о них могут быть применены не только в физиологии высшей нервной деятельно-сти, но и в других отраслях естествознания. Примером использования концепции функциональ-ной системы в ихтиологии и рыбохозяйственной науке может служить исследование И. А. Баран-никовой [5], в котором детально проанализированы основы нейроэндокринной регуляции поведе-ния осетровых и лососевых рыб в период преднерестовых миграций. Кроме того, автором указано на роль функциональной активности желез внутренней секреции в развитии внутрипопуляцион-ной дифференциации рыб и формировании механизмов осморегуляции у молоди на различных этапах онтогенеза.
На наш взгляд, все вышеизложенные особенности вполне соответствуют описанию струк-тур и процессов, происходящих в репродуктивной системе рыб при реализации функции воспро-изводства (размножения). В пользу такой возможности говорит и подмеченное П. К. Анохиным свойство живого организма динамически изменять характер протекающих процессов до тех пор, пока организм как целое не получит приспособительного эффекта или конечного результата, ка-ковым применительно к функционирующей репродуктивной системе рыб являются созревание, овуляция, вымет половых продуктов и оплодотворение икры. Главным вопросом, возникающим при характеристике репродуктивной системы рыб как функционального комплекса, является ее соответствие критериям, выдвинутым П. К. Анохиным.
Известно, что процесс развития и созревания половых клеток требует значительных изме-нений метаболизма – белкового, углеводного, липидного, минерального. В связи с осуществлени-ем процесса размножения в организме происходят значительные физиолого-биохимические сдви-

Âîäíûå áèîëîãè÷åñêèå ðåñóðñû è èõ ðàöèîíàëüíîå èñïîëüçîâàíèå
45
ги, меняется поведение и т. д. Поэтому очевидно, что осуществление репродуктивного цикла тре-бует согласованного взаимодействия многих систем организма. Несомненно, что процесс созре-вания и овуляции ооцитов является весьма динамичным и сопровождается усиленным синтезом и перераспределением в органах и тканях разнообразных неорганических и органических соеди-нений, транспортом их в гонады. В процесс размножения, помимо репродуктивной, вовлечены многие другие системы организма:
− в печени и в самих ооцитах идет синтез веществ, необходимых для первых этапов эмбриогенеза и начала реализации генетической программы развивающегося зародыша [6];
− кровь обеспечивает доставку в гонады необходимых компонентов, причем рост плодо-витости связан с параллельным увеличением показателей системы эритрона – концентрациями гемоглобина и эритроцитов [7];
− ЦНС и эндокринная система обеспечивают регуляцию процессов созревания и овуляции [8]. Следовательно, действующей репродуктивной системе рыб присуща значительная гетероген-ность участвующих компонентов.
Примером разнообразных положительных и отрицательных обратных связей может быть схема нейрогуморального контроля размножения у рыб, приведенная И. А. Баранниковой [8].
Примером саморегуляции, на наш взгляд, может служить динамика содержания фосфогли-церидов в овулировавшей икре, описанная в [9]. Судя по полученным данным, накопление струк-турных соединений в ооцитах, к которым относятся фосфоглицериды, идет с опережением во времени по сравнению с другими биохимическим компонентами. Однако в период, непосред-ственно предшествующий овуляции, доля фосфоглицеридов в ооцитах сокращается – вполне ве-роятно, что это происходит при стабилизации и оптимизации состава биологических мембран созревающих ооцитов.
Наконец, главным результатом функционирования репродуктивной системы являются со-зревание и овуляция – два главных процесса в размножении рыб. Таким образом, можно говорить о соответствии признаков репродуктивного комплекса критериям функциональной системы.
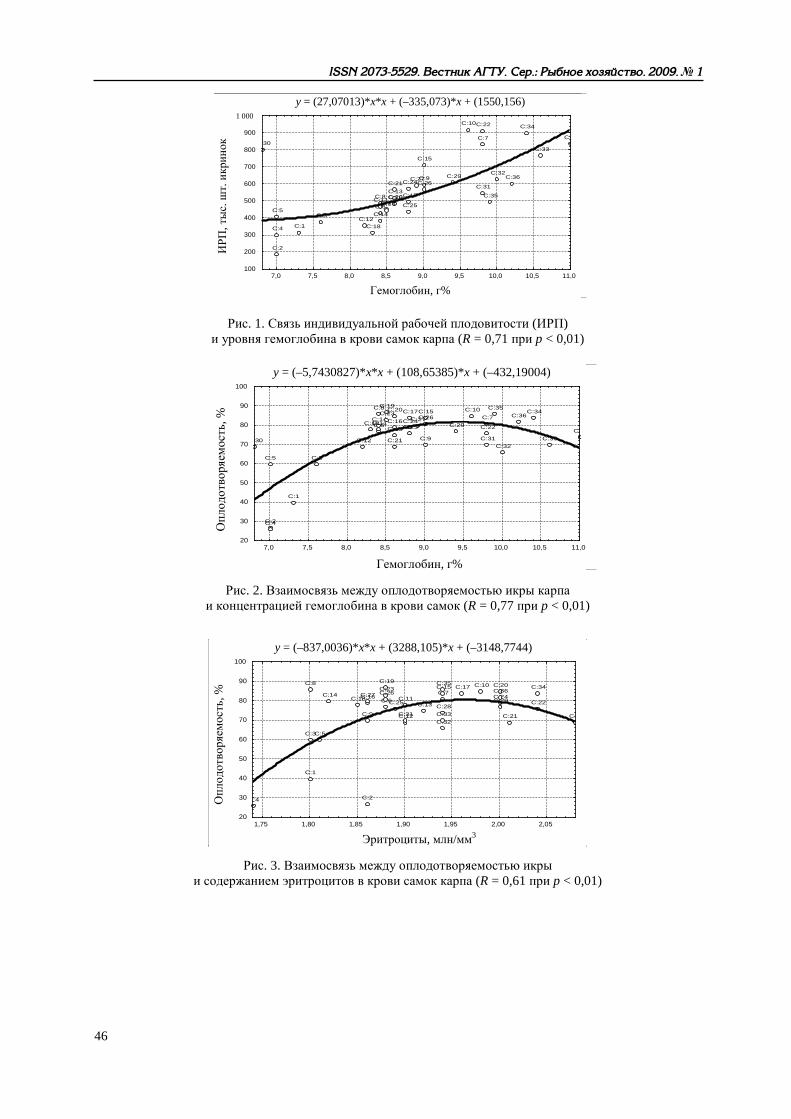
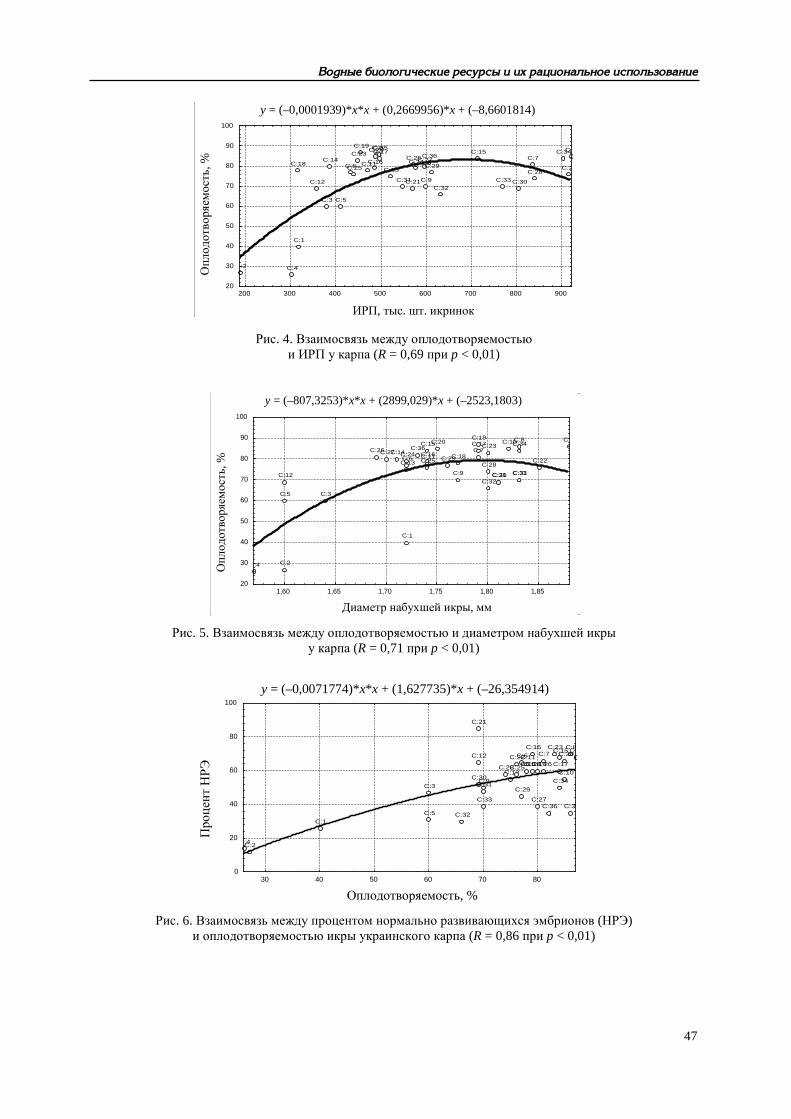
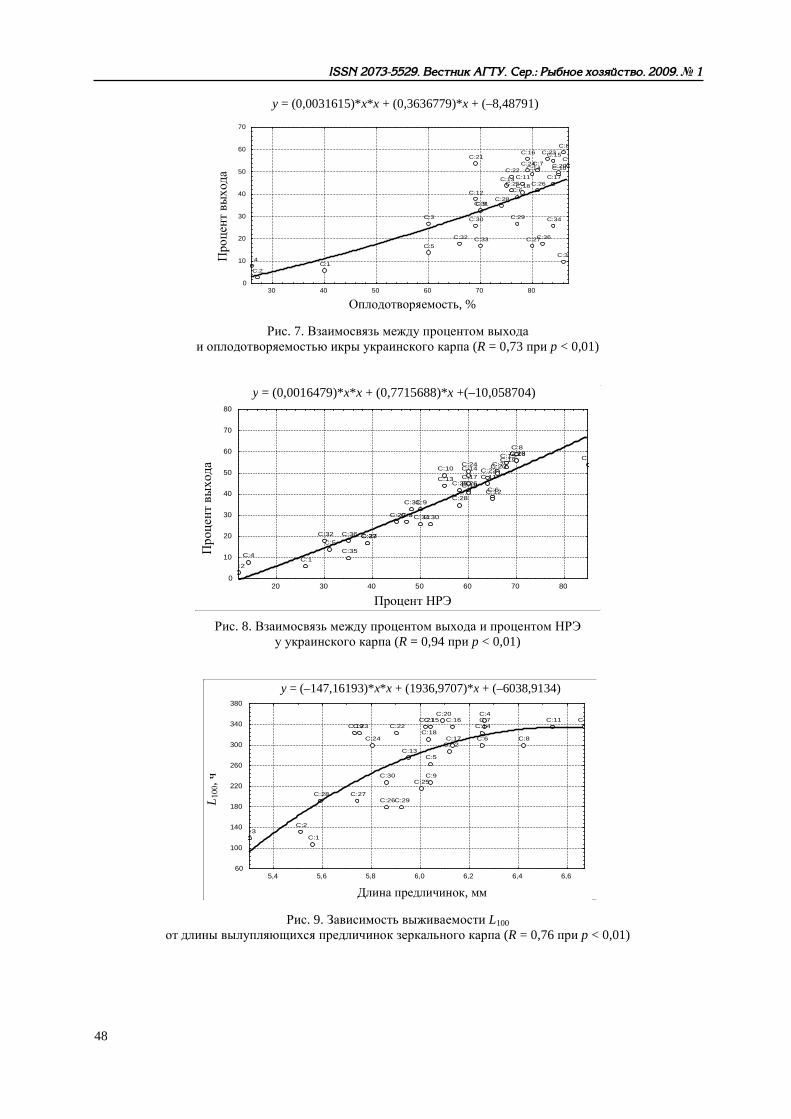
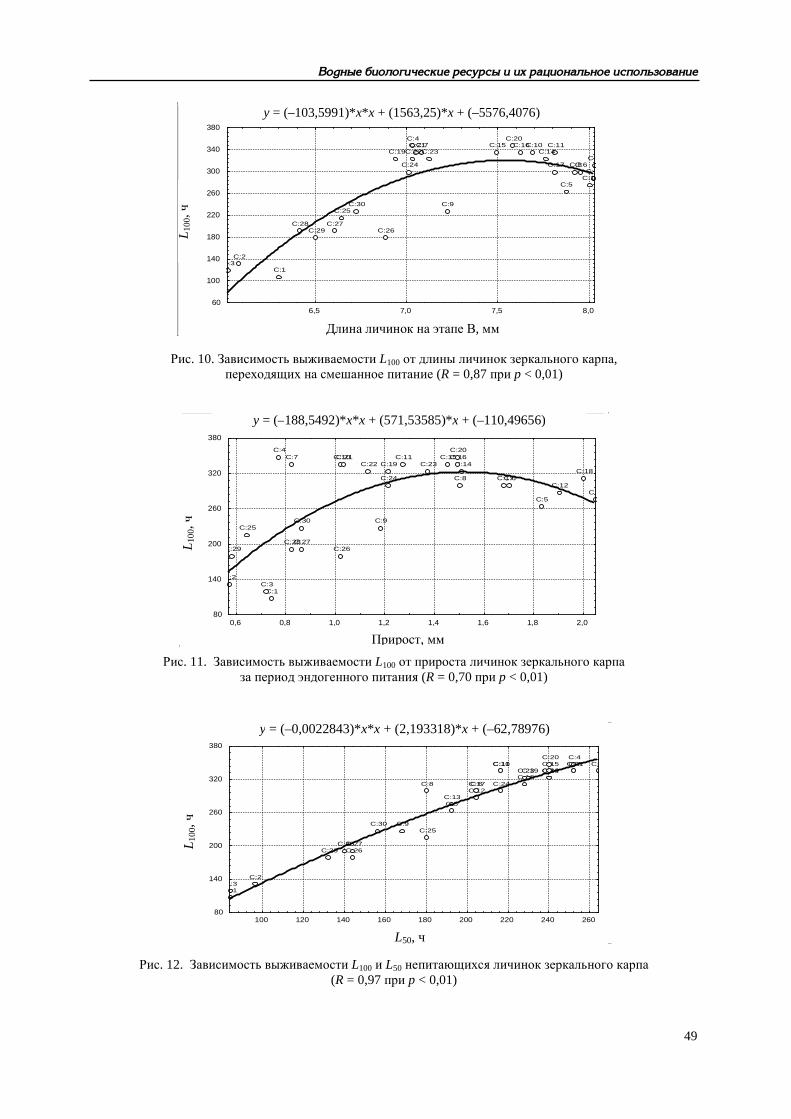
Другие аспекты функционирования репродуктивной системы рыб рассмотрены нами в [10], Показано, что разнообразные (прежде всего, физиолого-биохимические) характеристики произво-дителей карповых рыб в условиях искусственного стимулирования процессов созревания и ову-ляции существенно влияют на качество получаемых половых продуктов, жизнеспособность раз-вивающихся эмбрионов и личинок. Развивая представления В. И. Владимирова [11] о конститу-циональных и энергетических взаимосвязях между родителями и потомством у рыб при естест-венном размножении, мы пришли к выводу о том, что при искусственном разведении зависимо-сти гораздо сложнее – речь может идти о «функциональных связях». В это понятие должно быть включено представление об интенсивности обменных процессов в организме производи-телей и состоянии их нейрогуморальной системы. Параметры эритрона и концентрация гормо-нов в крови определяют, с одной стороны, фон, на котором происходит экзогенное стимулиро-вание созревания половых продуктов. Количественные характеристики гемоглобина и эритро-цитов, как отражение эндогенных обменных процессов, связаны определенными зависимостями с рабочей плодовитостью самок, биохимическим составом овулировавшей икры, оплодотворяе-мостью, ходом эмбрионального развития и даже с жизнестойкостью непитавшихся личинок. С другой стороны, созревание и овуляция у рыб считаются относительно независимыми про-цессами, но введенный извне комплекс гормонов, несомненно, сказывается и на интенсивности метаболизма, и на качестве половых продуктов. Таким образом, возникает цепь последователь-ных и взаимосвязанных параметров: система эритрона → рабочая плодовитость → биохимиче-ский состав овулировавшей икры → оплодотворяемость икры → диаметр набухшей икры → длина вылупляющихся предличинок → прирост личинок за период эндогенного питания → длина личинок, переходящих на смешанное питание → выживаемость L50 → выживаемость L100 непитавшихся личинок. Рост количественных величин одних параметров неизбежно ведет к со-ответствующему улучшению других показателей в раннем онтогенезе рыб, что можно проде-монстрировать на примере одного из изученных видов – украинского карпа (рис. 1–12).
ISSN 2073-5529. Âåñòíèê ÀÃÒÓ. Ñåð.: Ðûáíîå õîçÿéñòâî. 2009. № 1
46
y=(27,07013)*x*x+(-355,073)*x+(1550,156)
ãåìîãëîáèí, ã%
ÈÐÏ, òûñ. øòóê èêðèíîê
C:1
C:2
C:3
C:4
C:5C:6
C:7
C:8
C:9
C:10
C:11
C:12
C:13
C:14
C:15
C:16C:17
C:18
C:19C:20
C:21
C:22
C:23
C:24
C:25
C:26C:27
C:28
C:29
C:30
C:31
C:32
C:33
C:34
C:35
C:36
100
200
300
400
500
600
700
800
900
1000
7,0 7,5 8,0 8,5 9,0 9,5 10,0 10,5 11,0
Гемоглобин, г%
ИРП
, тыс.
шт.
икринок
y = (27,07013)*x*x + (–335,073)*x + (1550,156) 1 000
Рис. 1. Связь индивидуальной рабочей плодовитости (ИРП) и уровня гемоглобина в крови самок карпа (R = 0,71 при p < 0,01)
y=(-5,7430827)*x*x+(108,65385)*x+(-432,19004)
ãåìîãëîáèí, ã%
ïðîöåíò îïëîäîòâîðåíèÿ
C:1
C:2
C:3
C:4
C:5
C:6
C:7
C:8
C:9
C:10
C:11
C:12
C:13
C:14
C:15
C:16
C:17
C:18
C:19C:20
C:21
C:22
C:23
C:24C:25
C:26C:27
C:28C:29
C:30 C:31
C:32
C:33
C:34C:35
C:36
20
30
40
50
60
70
80
90
100
7,0 7,5 8,0 8,5 9,0 9,5 10,0 10,5 11,0
Оплодотворяемость, %
Гемоглобин, г%
y = (–5,7430827)*x*x + (108,65385)*x + (–432,19004)
Рис. 2. Взаимосвязь между оплодотворяемостью икры карпа и концентрацией гемоглобина в крови самок (R = 0,77 при p < 0,01)
y=(-837,0036)*x*x+(3288,105)*x+(-3148,7744)
ýðèòðîöèòû, ìëí / ìì 3
ïðîöåíò îïëîäîòâîðåíèÿ
C:1
C:2
C:3
C:4
C:5
C:6
C:7
C:8
C:9
C:10
C:11
C:12
C:13
C:14
C:15
C:16
C:17
C:18
C:19C:20
C:21
C:22
C:23
C:24C:25
C:26C:27
C:28C:29
C:30C:31
C:32
C:33
C:34C:35
C:36
20
30
40
50
60
70
80
90
100
1,75 1,80 1,85 1,90 1,95 2,00 2,05
Эритроциты, млн/мм3
Оплодотворяемость, %
y = (–837,0036)*x*x + (3288,105)*x + (–3148,7744)
Рис. 3. Взаимосвязь между оплодотворяемостью икры и содержанием эритроцитов в крови самок карпа (R = 0,61 при p < 0,01)
Âîäíûå áèîëîãè÷åñêèå ðåñóðñû è èõ ðàöèîíàëüíîå èñïîëüçîâàíèå
47
y=(-0,0001939)*x*x+(0,2669956)*x+(-8,6601814)
ÈÐÏ, òûñ. øòóê èêðèíîê
ïðîöåíò îïëîäîòâîðåíèÿ
C:1
C:2
C:3
C:4
C:5
C:6
C:7
C:8
C:9
C:10
C:11
C:12
C:13
C:14
C:15
C:16
C:17
C:18
C:19C:20
C:21
C:22
C:23
C:24C:25
C:26C:27
C:28C:29
C:30C:31
C:32
C:33
C:34C:35
C:36
20
30
40
50
60
70
80
90
100
200 300 400 500 600 700 800 900
ИРП, тыс. шт. икринок
Оплодотворяемость,
%
y = (–0,0001939)*x*x + (0,2669956)*x + (–8,6601814)
Рис. 4. Взаимосвязь между оплодотворяемостью и ИРП у карпа (R = 0,69 при p < 0,01)
y=(-807,3253)*x*x+(2899,029)*x+(-2523,1803)
äèàìåòð íàáóõøåé èêðû, ìì
ïðîöåíò îïëîäîòâîðåíèÿ
C:1
C:2
C:3
C:4
C:5
C:6
C:7
C:8
C:9
C:10
C:11
C:12
C:13
C:14
C:15
C:16
C:17
C:18
C:19C:20
C:21
C:22
C:23
C:24C:25
C:26C:27
C:28C:29
C:30 C:31
C:32
C:33
C:34C:35
C:36
20
30
40
50
60
70
80
90
100
1,60 1,65 1,70 1,75 1,80 1,85
Диаметр набухшей икры, мм
Оплодотворяемость, %
y = (–807,3253)*x*x + (2899,029)*x + (–2523,1803)
Рис. 5. Взаимосвязь между оплодотворяемостью и диаметром набухшей икры у карпа (R = 0,71 при p < 0,01)
y=(-0,0071774)*x*x+(1,627735)*x+(-26,354914)
ïðîöåíò îïëîäîòâîðåíèÿ
ïðîöåíò ÍÐÝ
C:1
C:2
C:3
C:4
C:5
C:6 C:7C:8
C:9
C:10
C:11C:12
C:13
C:14
C:15C:16
C:17C:18
C:19C:20
C:21
C:22
C:23
C:24C:25
C:26
C:27
C:28
C:29
C:30C:31
C:32
C:33
C:34
C:35C:36
0
20
40
60
80
100
30 40 50 60 70 80
Оплодотворяемость, %
Процент НРЭ
y = (–0,0071774)*x*x + (1,627735)*x + (–26,354914)
Рис. 6. Взаимосвязь между процентом нормально развивающихся эмбрионов (НРЭ) и оплодотворяемостью икры украинского карпа (R = 0,86 при p < 0,01)
ISSN 2073-5529. Âåñòíèê ÀÃÒÓ. Ñåð.: Ðûáíîå õîçÿéñòâî. 2009. № 1
48
y=(0,0031615)*x*x+(0,3636779)*x+(-8,48791)
ïðîöåíò îïëîäîòâîðåíèÿ
ïðîöåíò âûõîäà
C:1C:2
C:3
C:4
C:5
C:6
C:7
C:8
C:9
C:10
C:11
C:12
C:13
C:14
C:15C:16
C:17
C:18
C:19C:20
C:21
C:22
C:23
C:24
C:25 C:26
C:27
C:28
C:29C:30
C:31
C:32 C:33
C:34
C:35
C:36
0
10
20
30
40
50
60
70
30 40 50 60 70 80
Оплодотворяемость, %
Процент выхода
y = (0,0031615)*x*x + (0,3636779)*x + (–8,48791)
Рис. 7. Взаимосвязь между процентом выхода и оплодотворяемостью икры украинского карпа (R = 0,73 при p < 0,01)
y=(0,0016479)*x*x+(0,7715688)*x+(-10,058704)
ïðîöåíò ÍÐÝ
ïðîöåíò âûõîäà
C:1C:2
C:3
C:4
C:5
C:6
C:7
C:8
C:9
C:10
C:11
C:12
C:13
C:14
C:15C:16
C:17
C:18
C:19C:20
C:21
C:22
C:23
C:24
C:25C:26
C:27
C:28
C:29 C:30
C:31
C:32 C:33
C:34
C:35
C:36
0
10
20
30
40
50
60
70
80
20 30 40 50 60 70 80
Процент НРЭ
Процент выхода
y = (0,0016479)*x*x + (0,7715688)*x +(–10,058704)
Рис. 8. Взаимосвязь между процентом выхода и процентом НРЭ у украинского карпа (R = 0,94 при p < 0,01)
y=(-147,16193)*x*x+(1936,9707)*x+(-6038,9134)
äëèíà ïðåäëè÷èíîê, ìì
L 100, ÷àñû
C:1
C:2C:3
C:4
C:5
C:6
C:7
C:8
C:9
C:10C:11
C:12C:13
C:14C:15 C:16
C:17C:18
C:19
C:20C:21
C:22C:23
C:24
C:25
C:26C:27C:28
C:29
C:30
60
100
140
180
220
260
300
340
380
5,4 5,6 5,8 6,0 6,2 6,4 6,6
Длина предличинок, мм
L10
0, ч
y = (–147,16193)*x*x + (1936,9707)*x + (–6038,9134)
Рис. 9. Зависимость выживаемости L100 от длины вылупляющихся предличинок зеркального карпа (R = 0,76 при p < 0,01)
Âîäíûå áèîëîãè÷åñêèå ðåñóðñû è èõ ðàöèîíàëüíîå èñïîëüçîâàíèå
49
y=(-103,5991)*x*x+(1563,25)*x+(-5576,4076)
äëèíà ëè÷èíîê íà ýòàïå Â. ìì
L 100, ÷àñû
C:1
C:2C:3
C:4
C:5
C:6
C:7
C:8
C:9
C:10 C:11
C:12C:13
C:14C:15 C:16
C:17C:18
C:19
C:20C:21
C:22C:23
C:24
C:25
C:26C:27C:28
C:29
C:30
60
100
140
180
220
260
300
340
380
6,5 7,0 7,5 8,0
Длина личинок на этапе В, мм
L10
0, ч
y = (–103,5991)*x*x + (1563,25)*x + (–5576,4076)
Рис. 10. Зависимость выживаемости L100 от длины личинок зеркального карпа, переходящих на смешанное питание (R = 0,87 при p < 0,01)
y=(-188,5492)*x*x+(571,53585)*x+(-110,49656)
ïðèðîñò, ìì
L 100, ÷àñû
C:1
C:2C:3
C:4
C:5
C:6
C:7
C:8
C:9
C:10 C:11
C:12C:13
C:14C:15C:16
C:17C:18
C:19
C:20C:21
C:22 C:23
C:24
C:25
C:26C:27C:28
C:29
C:30
80
140
200
260
320
380
0,6 0,8 1,0 1,2 1,4 1,6 1,8 2,0
Прирост, мм
L10
0, ч
y = (–188,5492)*x*x + (571,53585)*x + (–110,49656)
Рис. 11. Зависимость выживаемости L100 от прироста личинок зеркального карпа за период эндогенного питания (R = 0,70 при p < 0,01)
y=(-0,0022843)*x*x+(2,193318)*x+(-62,78976)
L 50, ÷àñû
L 100, ÷àñû
C:1
C:2C:3
C:4
C:5
C:6
C:7
C:8
C:9
C:10C:11
C:12C:13
C:14C:15 C:16
C:17C:18C:19
C:20C:21
C:22C:23
C:24
C:25
C:26C:27C:28
C:29
C:30
80
140
200
260
320
380
100 120 140 160 180 200 220 240 260
L50, ч
L1
00, ч
y = (–0,0022843)*x*x + (2,193318)*x + (–62,78976)
Рис. 12. Зависимость выживаемости L100 и L50 непитающихся личинок зеркального карпа (R = 0,97 при p < 0,01)

ISSN 2073-5529. Âåñòíèê ÀÃÒÓ. Ñåð.: Ðûáíîå õîçÿéñòâî. 2009. № 1
50
Заключение Совокупность полученных экспериментальных данных и их математическое моделирова-
ние позволяют прийти к двум предварительным выводам. Во-первых, теория функциональных систем может быть использована при описании репродуктивной системы и начальных этапов онтогенеза рыб. Во-вторых, на существование функциональных связей в раннем онтогенезе ука-зывает рост количественных параметров по цепочке, охватывающей качественные характери-стики производителей, икры и личинок.
СПИСОК ЛИТЕРАТУРЫ
1. Анохин П. К. Очерки по физиологии функциональных систем. – М.: Медицина, 1975. – 448 с. 2. Судаков К. В. Теория функциональных систем. – М.: Наука, 1996. – 234 с. 3. Функциональные системы организма / под ред. К. В. Судакова. – М.: Медицина, 1987. – 431 с. 4. Анохин П. К. Избранные труды. Философские аспекты теории функциональной системы. – М.: Нау-
ка, 1978. – 400 с. 5. Баранникова И. А. Функциональные основы миграций рыб. – Л.: Наука, 1975. – 210 с. 6. Нейфах А. А., Тимофеева М. Я. Молекулярная биология процессов развития – М.: Наука, 1977. – 312 с. 7. Стрельников В. В., Залепухин В. В. Взаимосвязи рабочей плодовитости с физиолого-биохимическими
параметрами крови карповых рыб при искусственном разведении // Тр. Кубан. гос. аграр. ун-та. – Краснодар, 2007. – Вып. 2 (5). – С. 83–90.
8. Баранникова И. А. Гормональная регуляция репродуктивной функции у рыб с различной экологией // Биологические основы рыбоводства: актуальные проблемы экологической физиологии и биохимии рыб. – М.: Наука, 1984. – С. 178–218.
9. Федорова Н. Н., Залепухин В. В. Некоторые диагностические аспекты концепции эндогенной разно-качественности // Вестн. Астрахан. гос. техн. ун-та. – 2007. – № 3 (38). – С. 51–56.
10. Залепухин В. В. Концепция эндогенной разнокачественности в условиях искусственного воспроиз-водства карповых рыб: моногр. – Волгоград: Волгоград. науч. изд-во, 2006. – 320 с.
11. Владимиров В. И. Вариабельность размеров рыб на ранних этапах жизни и выживаемость // Разнока-чественность раннего онтогенеза у рыб. – Киев: Наук. думка, 1974. – С. 227–254.
Статья поступила в редакцию 28.03.2009
THE APPLICATION OF FUNCTIONAL SYSTEM THEORY
IN FISH RESEARCH
N. N. Fedorova, V. V. Zalepukhin
An attempt to prove the adaptability of P. K. Anokhin’s functional system theory to the description of fish spawning system has been made. Some examples of functional relations in the early ontogeny of fish are shown in the paper.
Key words: functional system theory, fish spawning system, early ontogeny.
Recommended

















